
A single origin of gall association in a group of parasitic wasps with disparate morphologies
12
A single origin of gall association in a group of parasitic wasps with disparate morphologies Alejandro Zaldivar-Rivero ´n a, * , Sergey A. Belokobylskij b,c , Virginia Leo ´ n-Regagnon a , Juan Jose ´ Martı ´nez d , Rosa Bricen ˜o e , Donald L.J. Quicke f,g a Departamento de Zoologı ´a, Instituto de Biologı ´a, Universidad Nacional Auto ´ noma de Me ´xico, 3er. Circuito Exterior, Cd. Universitaria, Ap. Postal 70-153, C. P. 04510, Me ´xico D. F., Mexico b Zoological Institute, Russian Academy of Sciences, Universitetskaya nab. 1, St. Petersburg 199034, Russia c Museum and Institute of Zoology, PAN, Wilcza 64, Warsaw 00-679, Poland d Divisio ´ n de Entomologı ´a, Museo Argentino de Ciencias Naturales ‘‘Bernardino Rivadavia’’, Av. Angel Gallardo 470, C1405DJR, Buenos Aires, Argentina e Universidad Centroccidental ‘‘Lisandro Alvarado’’, Decanato de Agronomı ´a, Depto. de Ciencias Biolo ´ gicas, Seccio ´ n de Entomologı ´a, Cabudare, Estado Lara, Venezuela f Division of Biology and Centre for Population Biology, Imperial College London, Silwood Park Campus, Ascot, Berkshire, SL5 7PY, UK g Department of Entomology, Natural History Museum, London SW7 5BD, UK Received 17 August 2006; revised 10 May 2007; accepted 17 May 2007 Available online 9 June 2007 Abstract The braconid wasp subfamily Doryctinae mainly comprises idiobiont ectoparasitoids of other insect larvae. In recent years, however, members of a few genera have been discovered to be associated with galls from various unrelated host plant families, with some of these being gall inducers whereas others are suspected as being predators of gallers. Because of their considerable morphological differences, these gall-associated taxa traditionally have been placed in separate tribes or even in other subfamilies. In this study, we investigate the phylogenetic relationships among representatives of a number of different doryctine genera, including five of its seven gall-associated genera using two genetic markers. Here we analyzed the length-variable 28S sequence data based on secondary structure both excising the unalignable regions and recoding them according to indel length. In addition, multiple alignments were carried out with a range of gap-opening and extension parameters. The combined (28S + CO1) phylogenetic hypotheses obtained, both excluding and recoding the unalignable regions, recover a clade comprising the five gall-associated genera, and most of the analyses using multiple alignments also support this relationship. These results support a scenario in which secondary phytophagy evolves from initially attacking primary gall- forming hosts. The relationships recovered are also more congruent with a model that explains the macroevolution of insect plant asso- ciation in the Doryctinae as reflecting geographic proximity rather than host plant relationships. Further, our phylogenetic hypotheses consistently show that one of the main morphological features employed in the higher level classification of the Doryctinae is actually highly homoplastic. Ó 2007 Elsevier Inc. All rights reserved. Keywords: Braconidae; Doryctinae; Cyclostomes; Character evolution, Molecular phylogeny; Secondary structure alignment; Multiple alignment 1. Introduction In the Hymenoptera, gall formation, predation of gall- inducers and inquilinism are related phenomena that occur in several lineages, principally in the Chalcidoidea and Cynipoidea, but which have also been discovered recently in a few members of the cyclostome group of the otherwise almost entirely parasitic family Braconidae (see Wharton 1055-7903/$ - see front matter Ó 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.ympev.2007.05.016 * Corresponding author. Address: Museo Nacional de Ciencias Naut- rales, Departamento de Biodiversidad y Biologı ´a Evolutiva, C/Jose ´ Gutie ´rrez Abascal 2, 28006 Madrid, Spain. E-mail address: [email protected] (A. Zaldivar-Rivero ´ n). www.elsevier.com/locate/ympev Molecular Phylogenetics and Evolution 44 (2007) 981–992
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of A single origin of gall association in a group of parasitic wasps with disparate morphologies

www.elsevier.com/locate/ympev
Molecular Phylogenetics and Evolution 44 (2007) 981–992
A single origin of gall association in a group of parasiticwasps with disparate morphologies
Alejandro Zaldivar-Riveron a,*, Sergey A. Belokobylskij b,c, Virginia Leon-Regagnon a,Juan Jose Martınez d, Rosa Briceno e, Donald L.J. Quicke f,g
a Departamento de Zoologıa, Instituto de Biologıa, Universidad Nacional Autonoma de Mexico, 3er. Circuito Exterior, Cd. Universitaria,
Ap. Postal 70-153, C. P. 04510, Mexico D. F., Mexicob Zoological Institute, Russian Academy of Sciences, Universitetskaya nab. 1, St. Petersburg 199034, Russia
c Museum and Institute of Zoology, PAN, Wilcza 64, Warsaw 00-679, Polandd Division de Entomologıa, Museo Argentino de Ciencias Naturales ‘‘Bernardino Rivadavia’’, Av. Angel Gallardo 470, C1405DJR, Buenos Aires, Argentina
e Universidad Centroccidental ‘‘Lisandro Alvarado’’, Decanato de Agronomıa, Depto. de Ciencias Biologicas, Seccion de Entomologıa, Cabudare,
Estado Lara, Venezuelaf Division of Biology and Centre for Population Biology, Imperial College London, Silwood Park Campus, Ascot, Berkshire, SL5 7PY, UK
g Department of Entomology, Natural History Museum, London SW7 5BD, UK
Received 17 August 2006; revised 10 May 2007; accepted 17 May 2007Available online 9 June 2007
Abstract
The braconid wasp subfamily Doryctinae mainly comprises idiobiont ectoparasitoids of other insect larvae. In recent years, however,members of a few genera have been discovered to be associated with galls from various unrelated host plant families, with some of thesebeing gall inducers whereas others are suspected as being predators of gallers. Because of their considerable morphological differences,these gall-associated taxa traditionally have been placed in separate tribes or even in other subfamilies. In this study, we investigate thephylogenetic relationships among representatives of a number of different doryctine genera, including five of its seven gall-associatedgenera using two genetic markers. Here we analyzed the length-variable 28S sequence data based on secondary structure both excisingthe unalignable regions and recoding them according to indel length. In addition, multiple alignments were carried out with a range ofgap-opening and extension parameters. The combined (28S + CO1) phylogenetic hypotheses obtained, both excluding and recoding theunalignable regions, recover a clade comprising the five gall-associated genera, and most of the analyses using multiple alignments alsosupport this relationship. These results support a scenario in which secondary phytophagy evolves from initially attacking primary gall-forming hosts. The relationships recovered are also more congruent with a model that explains the macroevolution of insect plant asso-ciation in the Doryctinae as reflecting geographic proximity rather than host plant relationships. Further, our phylogenetic hypothesesconsistently show that one of the main morphological features employed in the higher level classification of the Doryctinae is actuallyhighly homoplastic.� 2007 Elsevier Inc. All rights reserved.
Keywords: Braconidae; Doryctinae; Cyclostomes; Character evolution, Molecular phylogeny; Secondary structure alignment; Multiple alignment
1055-7903/$ - see front matter � 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.ympev.2007.05.016
* Corresponding author. Address: Museo Nacional de Ciencias Naut-rales, Departamento de Biodiversidad y Biologıa Evolutiva, C/JoseGutierrez Abascal 2, 28006 Madrid, Spain.
E-mail address: [email protected] (A. Zaldivar-Riveron).
1. Introduction
In the Hymenoptera, gall formation, predation of gall-inducers and inquilinism are related phenomena that occurin several lineages, principally in the Chalcidoidea andCynipoidea, but which have also been discovered recentlyin a few members of the cyclostome group of the otherwisealmost entirely parasitic family Braconidae (see Wharton

982 A. Zaldivar-Riveron et al. / Molecular Phylogenetics and Evolution 44 (2007) 981–992
and Hanson, 2005 for a review). Starting from the firstreport of phytophagy in Allorhogas Gahan almost 20 yearsago (Macedo and Monteiro, 1989), cecidogenesis (i.e., galldevelopment) in the Braconidae has subsequently beenconfirmed only in other two cyclostome genera, Monitoriel-
la Hedqvist (Infante et al., 1995) and Mesostoa van Achter-berg (Austin and Dangerfield, 1998). Some other braconidwasps have only ever been reared from galls, though someof these appear to display various forms of gall associationrather than being the primary gall-formers themselves.Most of these gall-associated species belong to the largeand cosmopolitan subfamily Doryctinae, though this fea-ture is also displayed by species of three genera of the Mes-ostoinae (sensu Zaldivar-Riveron et al., 2006) (Mesostoa,Aspilodemon Fischer, and Hydrangeocola Brethes; Brethes,1927; Austin and Dangerfield, 1998; Oda et al., 2001).
Doryctine wasps are mainly idiobiont (i.e., they arrestthe host development at the time of parasitization) ectopar-asitoids of xylophagous Coleoptera larvae (Belokobylskijet al., 2004a,b). However, gall association is exhibited byspecies of seven Neotropical doryctine genera, most ofwhich have been reared from different unrelated host plantfamilies. In Allorhogas, several species are known to inducegalls in seeds of species of Melastomataceae, Bignoniaceaeand at least four legume (Fabaceae) genera, though someother species are thought to be parasitoids of gall inhabit-ants (Macedo and Monteiro, 1989; Macedo et al., 1998;Marsh, 2002; Marsh et al., 2000). Moreover, the type spe-cies of Percnobracon Kieffer, Pe. stenopterus Kieffer, wasreared from cecidomyiid (Diptera) galls on the legumeProsopis strombulifera (Lam.) Benth (Kieffer, 1910), andrecently this and two other species of the genus have beenreared from the Argentinian endemic Pr. caldenia Burkart(Martınez, 2006). Species of Monitoriella, on the otherhand, induce leaf galls on Philodendron Schott (Araceae)(Infante et al., 1995), whereas some members of Labania
Hedqvist and Psenobolus Reinhard have been observed todevelop in galls located on aerial roots and leaves, andon the syconia of Ficus L. (Moraceae), respectively (Marsh,2002; Ramirez and Marsh, 1996). Finally, the onlydescribed species of Donquickeia Marsh and Mononeuron
Fischer were reported to be reared from cecidomyiid gallsof Mikania sp. (Asteraceae) and Eugenia rotundifolia Casar(Myrtaceae) (Penteado-Dias, 2000) and from a species ofDuguetia A.St.-Hil (Annonaceae) (Fischer, 1981),respectively.
Since the process of host shift in phytophagous insectsinvolves a number of genetic and ecological factors (e.g.,secondary metabolites, competitors, predators; Thompson,1999), this is normally expected to occur more between clo-sely related host plant species rather than in distant, unre-lated families (e.g., Becerra, 1997; Kopf et al., 1998). Thedifferent host plant families inhabited by the above doryc-tine genera would therefore suggest that at least the con-firmed cecidogenic Allorhogas and Monitoriella aredistantly related. This idea has been generally supportedby the considerable external morphological differences
shown by the different gall-associated doryctine genera.For this reason, these taxa have been placed in separateand in some cases exclusive tribes (e.g., Labanini, Perc-nobraconini) or even in other subfamilies (e.g., Monitoriel-
la: Wharton, 1993; but see Zaldivar-Riveron et al., 2006). Ithas been argued, however, that the external morphology ofdoryctine wasps is quite homoplastic (Belokobylskij et al.,2004b), and thus several features employed to differentiateits higher taxa could have actually evolved separately inrepeated occasions.
Recently, a simultaneous molecular and morphologicalphylogenetic analysis among representatives of the cyclo-stome braconid subfamilies strongly supported the gall-associated Labania and Monitoriella as being sister taxa(Zaldivar-Riveron et al., 2006). That study, however, didnot sample other gall-associated doryctine genera and onlyincluded a scarce number of members of this group. Thus,relationships both within the subfamily and among thegall-associated genera were far from conclusive.
Different strategies have been proposed for aligninglength-variable rDNA sequence data, though unalignableregions present in rDNA sequence alignments still are com-monly excluded and therefore the results obtained arebased exclusively on regions that appear to be length-con-served. In the case of secondary structure alignment(Gillespie, 2004), strict criteria of matched base-pairingare used to delimit putatively homologiseable stem regionsfrom ambiguously alignable loops and expansion zones.However, coding approaches also exist that align length-variable regions conditionally based on indel length (e.g.,Simmons and Ochoterena, 2000; Lutzoni et al., 2000;Zaldivar-Riveron et al., 2006). Alternatively, computeralgorithms are often employed to find best matchesbetween bases in regions of ambiguous alignment, andthe incorporated variation may contribute to the phyloge-netic analysis performed. These include multiple alignment(e.g., Clustal X, Thompson et al., 1997; MALIGN,Wheeler and Gladstein, 1994) and dynamic-based align-ments (e.g., POY, Wheeler, 1996; Gladstein and Wheeler,1996), and when using them it is increasingly common touse a ‘sensitivity’ approach, where the stability of theresults is assessed across a range of user-defined parame-ters. Here we utilize three different alignment strategies,secondary structure alignment excluding unalignableregions, secondary structure alignment with conditionalalignment of indel regions, and multiple alignment usinga range of parameters to assess the sensitivity of our resultsto alignment procedure.
In this study, we report a close relationship among mostof the currently known gall-associated doryctine waspsbased on a molecular phylogenetic analysis using two genefragments. We discuss the implications of this finding onthe origin of cecidogeny and on the evolution of host plantshifts displayed among these taxa. Moreover, we assess theevolution of one of the key features employed to distin-guish among supraspecific taxa within the group, the lengthof acrosternite (strongly sclerotised basal sternal plate) of

A. Zaldivar-Riveron et al. / Molecular Phylogenetics and Evolution 44 (2007) 981–992 983
the first metasomal segment, and consider the conse-quences that our results will have in the higher level classi-fication of the Doryctinae.
2. Materials and methods
2.1. Taxon sampling
DNA sequence data were generated for a total of 51taxa belonging to 50 different doryctine genera. The taxonsampling comprised representative genera from the Afro-tropical, Australian, Neotropical, Oriental, and Palaearcticregions, which belong to most of the tribes recognized byBelokobylskij (1992). Monophyly of the Doryctinae, previ-ously supported by relatively little convincing morpholog-ical evidence (see Quicke et al., 1992), was tested byincluding members of seven cyclostome subfamilies, withthe rhyssaline Rhyssalus Haliday used for rooting all trees:previous studies have consistently recovered the Rhyssali-nae as the sister group of the remaining cyclostome sub-families except the Aphidiinae + Mesostoinae s.l. clade,which appeared as the sister group of all of them (Belshawet al., 1998, 2000; Dowton et al., 2002; Zaldivar-Riveronet al., 2006). Details of the included material, voucherinformation, and GenBank/EMBL Accession Numbersare given in Table 1.
2.2. DNA sequence data and alignment
The genetic markers examined included �650 bp of thesecond and third domains of the nuclear 28S rDNA geneand 603 bp of the COI mtDNA gene. Genomic DNAwas extracted from ethanol preserved and up to 15 yearsold dry pinned specimens. The DNA extraction and ampli-fication procedures employed were those described inZaldivar-Riveron et al. (2006). 28S sequences wereobtained using the primers designed by Belshaw andQuicke (1997) (fwd: 50-GCG AAC AAG TAC CGTGAG GG-30) and Mardulyn and Whitfield (1999) (rev:50-TAG TTC ACC ATC TTT CGG GTC CC0-30). COIsequences were amplified using the primers designed byFolmer et al. (1994) (LCO 50-GGT CAA CAA ATCATA AAG ATA TTG G-30 HCO 50-0TAA ACT TCAGGG TGA CCA AAA AAT CA-30).
COI sequences were unambiguously aligned based ontheir translated amino acids; however, alignment of thelength-variable 28S gene fragment was more problematicand here we employ three different strategies and comparethe results to assess the sensitivity of our findings to choiceof alignment method.
2.2.1. Secondary structure alignment
Alignment of the 28S sequences followed the 28S braco-nid secondary structure alignment proposed by Gillespieet al. (2005). Length variable regions were identified andinitially characterized according to the three categories pro-posed by Gillespie (2004) [regions of ambiguous alignment
(RAAs), regions of slipped-strand compensation (RSCs),and regions of expansion and contraction (RECs)]. RAAsand their adjacent RECs were then combined into singleunalignable regions for subsequent analyses using condi-tional alignment (see below). The secondary structurealignment excluding these ambiguous regions is termed28SN.
2.2.2. Indel coding and conditional alignment
Regions of the 28S data set whose positional homologycould not be confidently assigned were recoded followingthe approach used by Zaldivar-Riveron et al. (2006). Thisapproach offers an objective criterion for preserving allthe nucleotide variation that is not in conflict due tosequence length variation and additionally includes infor-mation on indel length by attributing a ‘morphological’character state to indels of identical length regardless totheir base composition. This data set, comprising the sec-ondary structure alignable regions (28SN) together withthe unalignable regions of identical length and the ‘mor-phological’ states referring to indel lengths is referred toas 28SA.
2.2.3. Multiple alignment
Multiple alignments of the 28S data were created withClustal X (Thompson et al., 1997) with each of the follow-ing range of gap opening-gap extension penalties: 1:1, 2:1,2:2, 4:1, 4:2, 4:4, 6:1, 6:2, 6:4, 6:6, 8:1, 8:2, 8:4, 8:6, 8:8, 10:1,10:2, 10:4, 10:6, 10:8, 10:10. All of these used a DNA tran-sition: transversion weight ratio of 1.0: 1.0, and the defaultpairwise alignment parameters.
2.3. Phylogenetic analysis
Bayesian MCMC analyses were performed for the sepa-rate and combined data sets (with 28SN and 28SA matri-ces) using MrBayes version 3.1.2 (Ronquist andHuelsenbeck, 2003). Two independent analyses were runsimultaneously for each data set, each consisting of 2 mil-lion generations, sampling trees every 1000 generations,and using four chains and uniform priors on three topolo-gies, with the latter implying non-uniform prior probabili-ties of clades due to these are dependent on the number oftaxa in a clade as well as the number of taxa in the analysis(Pickett and Randle, 2005). The GTR + I + C model ofsequence evolution was the model selected for the two genemarkers examined according to the likelihood ratio testimplemented with MrModeltest version 2.2 (Nylander,2004). There is no reason to assume a priori that the statesderived from the 28SA indel characters, whose states arecoded as ‘morphological’ (i.e., with numerical coding),could have different frequencies. The Mk + C (Markov k;Lewis, 2001) model of evolution was therefore used forthe above characters. This model was originally designedto deal with morphological characters and considers equalbase frequencies, equal mutation rates and makes the like-lihood conditional on characters being variable (Lewis,

Table 1Provenances and voucher and EMBL/GenBank Accession Numbers of the taxa included
Voucher No. 28S COI
TaxonAcanthorhogas sp., Costa Ricaa Jo-713 DQ498942 DQ498975Acrophasmus exilis Marsh, Costa Ricad IB-02 DQ498943 DQ498976Aivalykus arawak, Costa Ricaa Jo-725 AY935471* AY935398*
Allorhogas sp., La Pampa, Argentinad� IB-18 DQ498927 —Allorhogas sp., Cartago, Turrialba, Costa Ricad� IB-20 — DQ498959Aphelopsia striata Braet & Barbalho, French Guyana (holotype)b Jo-838 DQ498941 DQ498974Barbalhoa contophleba Marsh, Costa Ricaa� Jo-723 DQ498935 —Barbalhoa licina Marsh, Costa Ricaa� Jo-739 — DQ498968Caenopachys hartigii (Ratzeburg), Corsica, Francea Jo-793 AY935474* AY935401*
Caenophanes sp., Africaa Jo-781 AY935465* DQ498961Curtisella hansoni, Costa Ricad IB-03 DQ498944 DQ498977Doryctes heydenii Reinhard, Palaearcticd — DQ498913 DQ498945Doryctes leucogaster (Nees), Israela Jo-876 AY935472* DQ498946Ecphylus sp., South Americae — AJ302894* —Euscelinus sarawacus Westwood, Thailanda Jo-871 DQ498920 DQ498952Hansonorum sp. nov. aff. pauli, Costa Ricaa Jo-728 DQ498934 DQ498967Hecabolus sulcatus Curtis, Ascot, UKa Jo-650 AY935473* AY935400*
Hemidoryctes carbonarius (Ashmead), Diego Suarez Province, Madagascara Jo-592 DQ498921 DQ498953Heterospilus prosopidis Viereck, Silwood culture, UKe — AY935469* AY935396*
Heterospilus sp., Costa Ricaa IB-01 DQ498926 DQ498958Histeromeroides sp., Rio Napo, Ecuadora Jo-866 DQ498940 DQ498973Hypodoryctes sibiricus Kokujev, no datae� — AJ302895* —Hypodoryctes sibiricus kokujev, Finlandd� Jo-981 — DQ498965Ipodoryctes sp., St. Phillippe, Reunion Islanda Jo-929 DQ498916 DQ498949Jarra maculipennis Marsh & Austin, Australiaa — DQ498925 DQ498957Johnsonius sp., Costa Ricaa Jo-719 DQ498939 DQ498972Labania sp., Costa Ricae — DQ498928 AY935397*
Lamquetia marshi Braet & Barbalho, French Guyana (holotype)b Jo-833 DQ498938 DQ498971Leptorhaconotus sp. (1), Madagascara Jo-655 AY935479* AY935406*
Liobracon sp., Costa Ricae Jo-714 AY935467* AY935394*
Masonius sp., Cuyagua, Venezuelab Jo-578 DQ498933 DQ498966Megaloproctus sp., Colombiaa Jo-654 AY935466* AY935393*
Monitoriella sp., Costa Ricaa Jo-733 AY935457* AY935387*
Notiospathius sp., Caucagua, Venezuelab Jo-596 AY935477* AY935404*
Odontobracon sp., Costa Ricaa Jo-712 AY935468* AY935395*
Ondigus bicolor Braet, Barbalho & van Achterberg, French Guyana (holotype)b Jo-834 DQ498937 DQ498970Ontsira imperator (Haliday), Honshu Ibaraki, Japana Jo-971 DQ498919 DQ498951Pedinotus sp., Costa Ricaa Jo-742 DQ498936 DQ498969Percnobracon cf. Stenopterus Kieffer, Santa Rosa, La Pampa, Argentinad IB-19 DQ498930 DQ498960Platyspathius sp., Benina AL-183 DQ498915 DQ498948Psenobolus sp., Camboto, Venezuelad Jo-666 DQ498929 —Pseudodoryctes sp., Malawia Jo-679 DQ498922 DQ498954Rhaconotus emarginatus Marsh, Costa Ricaa Jo-720 DQ498914 DQ498947Rhacontsira sp., Hoa Binh Province, Vietnama Jo-923 DQ498917 DQ498950Rinamba opacicollis Cameron, Madagascara Jo-588 DQ498923 DQ498955Schlettereriella sp., Kibale, Ugandaa Jo-673 AY935478* AY935405*
Semirhytus sp., Munipality of Lara, Yacambu, Venezuelac� Jo-628 DQ498931 —Semirhytus sp., no datad� Jo-983 — DQ498962Spathiomorpha sp. aff.longipalpis, Japana Jo-711 DQ498924 DQ498956Spathius agrili Yang, China — AY920299* AY920285*
Spathius sp., Tollara Province, Madagascarb Jo-591 AY935476* AY935403*
Spathius (Antespathius) sp., Madagascara Jo-910 DQ498918 —Stenocorse sp., Belizea Jo-750 AY935475* AY935402*
Syngaster lepidus Brulle, Australiae — AJ245698* DQ498963Tarasco sp., Camboto, Venezuelab Jo-926 DQ498932 —Tarasco sp., Camboto, Venezuelab Jo-587 — DQ498964
OutgroupsCystomastax sp., Caucagua, Venezuelab Jo-574 AY935445* AY935369*
Hormius sp., Mahajanga Province, Madagascarb Jo-582 AY935455* AY935385*
Oncophanes sp., Ascot, UKe — AY935481* AY935407*
Pambolus sp., Choroni, Venezuelab Jo-597 AY935458* AY935388*
(continued on next page)
984 A. Zaldivar-Riveron et al. / Molecular Phylogenetics and Evolution 44 (2007) 981–992

Table 1 (continued)
Voucher No. 28S COI
Pseudorhysipolis sp., Costa Ricab Jo-758 AY935450* AY935377*
Rhyssalus clavator Haliday, Kazimierz, Polanda Jo-890 AY935482* AY935409*
Stiropius sp., Costa Ricaa Jo-730 AJ784961* AY935373*
*Sequences obtained from previous works. Voucher specimens: aZoological Institute, St. Petersburg, Russia; bNationaal Natuurhistorisch Museum,Leiden, Netherlands; cUniversidad Centrooccidental, Tabudare, Venezuela; dInstituto de Biologıa, Universidad Nacional Autonoma de Mexico, Mexico;enot retained/none remaining. �Specimens whose sequences were used to form a single terminal taxon.
A. Zaldivar-Riveron et al. / Molecular Phylogenetics and Evolution 44 (2007) 981–992 985
2001). The 28S nucleotide and indel characters and thethree COI codon positions were each treated as unlinkedpartitions. Likelihoods reached their plateau ranges before100,000 generations in all the analyses; therefore, their first100 sampled trees were discarded as burn-in. Relationshipsderived from the postburn-in samples were similar in thetwo analyses run for each data set and thus they werepooled and used to produce a 50% majority rule consensustree with posterior probabilities of clades.
The analyses for the combined data sets derived fromthe 28S multiple alignments used the same search strategiesand parameters as above, except for the number of rungenerations, which was of 1 million. Burn-in in all the anal-yses was determined to be completed by 100,000 genera-tions. We also carried out maximum parsimony (MP)analyses for the combined and separate data sets thatresulted from the 28S multiple alignments with PAUP* ver-sion 4.0b10 (Swofford, 1998). Alignments from eachparameter combination were analyzed using 10,000 ran-dom additions followed by TBR branch-swapping andholding no more than one most parsimonious tree (MPT)for swapping at any one time. The resulting MPTs fromeach search were then used as starting trees for furtherTBR searches with unlimited maxtrees. Application ofreweighting on these using the maximum value of the reten-tion index as the reweighting function, followed byreweighting characters back to unity (Quicke et al., 2001)failed to recover any more parsimonious trees.
The matrices of the separate and combined data setswith 28SN and 28SA and their topologies reconstructedcan be downloaded from the TreeBase web page (accessionno. S1800). The matrices with the multiple alignments ana-lyzed and their resulting topologies can be obtained uponrequest to AZR.
2.4. Test of alternative hypotheses
A Bayesian approach (Buckley et al., 2002; Reeder,2003; Brandley et al., 2004) was implemented for hypothe-sis testing of alternative topologies not present in ourmajority consensus trees derived from the two simulta-neous analyses. In particular, we examined the monophylyof each of the large tribes Doryctini, Hecabolini and Spath-iini (sensu Belokobylskij, 1992). Of these, the gall-associ-ated Allorhogas and Psenobolus were placed in theHecabolini and Spathiini, respectively. In this approach,a 95% credible set of trees sampled after burn-in wasassembled for each simultaneous analysis, considering a
given alternative phylogenetic hypothesis as statisticallyrejected when this was absent in all of the credible set oftrees. The presence of the alternative hypotheses withinthe 95% posterior intervals was detected by creating theselected topological constraint with MacClade 4.06 (Madd-ison and Maddison, 2003) and then using this to filter theset of credible trees using the option ‘compatible with con-straints’ included in PAUP* version 4.0b10 (Swofford,1998).
The Shimodaira-Hasegawa (SH; Shimodaira and Hase-gawa, 1999) test was also performed with PAUP* version4.0b10 (Swofford, 1998) to test for significant differencesbetween the likelihood of the 28SN + COI Bayesian phy-logeny and the likelihood of alternative Bayesian topolo-gies that enforced the above tribes each as monophyletic.The SH tests were carried out using the full optimizationsampling with 1000 replicates. The alternative Bayesiantopologies were obtained with the constraint option avail-able in MrBayes version 3.1.2, using the same strategiesand parameters employed in the unconstrained analyses.Burn-in in these analyses was determined occur after300,000 generations.
2.5. Analysis of character evolution
The evolution of gall-association and of one morpholog-ical feature used to distinguish higher-level taxa in the Dor-yctinae, the length of acrosternite of the first metasomaltergite, were investigated mapping their selected characterstates onto the 2SN + COI Bayesian phylogeny employingmaximum likelihood character optimizations using Mes-quite 1.6 (Maddison and Maddison, 2004). The morpho-logical feature was defined to have three states [(0) 0.2–0.25 times as long as tergite, petiole absent; (1) 0.3–0.5times as long as tergite, petiole present but short; (2) 0.6–0.85 times as long as tergite, petiole present and long].The gall association trait was defined as binary [(0)absent/unknown; (1) present]. Ecological information wasscored from literature and personal observations, and mor-phological data was obtained from the sequenced speci-mens and from literature. The character states scored forthe two features examined can be retrieved from the elec-tronic supplementary material. The Markov k-state oneparameter model (Mk1; Lewis, 2001) was used for the like-lihood character optimizations, which assumes a single ratefor transitions between character states. A likelihood ratiotest was carried out to discriminate for the best estimate ofancestral state at each node following Pagel (1994). In this

986 A. Zaldivar-Riveron et al. / Molecular Phylogenetics and Evolution 44 (2007) 981–992
test, a state is considered as the best estimate for a givenbranch if it differs by P2.0 from one or more states withhigher negative log-likelihood values, and considers theancestral reconstruction as ambiguous when the differencebetween states is 62.0.
The full hierarchical Bayesian method for inferringancestral states (Huelsenbeck and Bollback, 2001) was alsoperformed for the gall association character with MrBayesversion 3.1.2 (Ronquist and Huelsenbeck, 2003) using the28SN + COI data set. This method incorporates uncer-tainty in the tree, branch lengths, and substitution modelparameters, approximating the posterior probability ofancestral states employing Markov Chain Monte Carlo(Huelsenbeck and Bollback, 2001). The Bayesian analysisperformed to infer the ancestral state for the above charac-ter was run using the same parameters and settings as theoriginal phylogenetic analyses, except by including the gallassociation character in the matrix analyzed and constrain-ing the node with the five gall associated genera examined(see Section 3).
3. Results
3.1. Phylogenetic reconstructions
Despite several attempts were made to obtain both 28Sand COI sequences for all the ethanol preserved and drypinned specimens examined, COI sequences could not beobtained for three taxa [Psenobolus, Spathius Nees (Antes-
pathius Belokobylskij), and Ecphylus Foerster]. Moreover,in five genera (Tarasco Marsh, Hypodoryctes Kokujev,Barbalhoa Marsh, Semirhytus Szepligeti, and Allorhogas)sequences of the two genetic fragments were obtained fromdifferent specimens, but in the case of Tarasco sp. and Hyp-
odoryctes sibiricus Kokujev, the sequences were from differ-ent individuals of the same species. The 28S data setconsisted of 538 unambiguously aligned positions and 15unalignable regions. Inclusion of the recoded unalignableregions added 263 characters to the data set, of which249 were from the nucleotide clusters created and 14 fromthe ‘morphological’ indel characters.
The majority rule consensus trees that resulted from theseparate 28SN and COI Bayesian analyses performed areshown in Figs. 1 and 2, respectively. The COI Bayesianphylogeny recovered the highest number of significantlysupported clades in comparison to the analyses with the28SN and 28SA data sets (COI = 21; 28SN = 18;28SA = 19). The topologies reconstructed from the 28SNand 28SA data sets were mainly similar, with no conflictingsignificantly supported clades among them. The three sep-arate analyses performed recovered a clade with gall-asso-ciated genera. A clade with Heterospilus Haliday and thegall-associated genera included except Labania was recov-ered in the 28SN topology (BPP = 0.66), whereas in the28SA topology the five gall-associated genera were recov-ered in a single clade together with the above species ofHeterospilus (BPP = 0.89). Moreover, of the four gall-asso-
ciated genera examined in the COI Bayesian analysis, threeof them (Allorhogas, Labania, and Monitoriella) were sig-nificantly recovered as monophyletic (BPP = 1.0). Noneof the separate analyses recovered the taxa with elongatedfirst metasomal tergite to be monophyletic.
The 28SN + COI Bayesian topology with its posteriorprobabilities and the posterior probabilities of similarclades recovered in the 28SA + COI Bayesian topologyare shown in Fig. 3. The number of significant posteriorprobabilities of clades considerably increased in the twocombined analyses with respect to the separate ones. Atotal of 22 clades were similar and significantly supportedin the two combined analyses, but two clades were only sig-nificantly supported in the analysis with 28SN and fourclades increased their BPP to significant values in the anal-ysis with 28SA (Fig. 3).
The two simultaneous analyses both show a non-mono-phyletic Doryctinae, though the clades implicated are notsignificantly supported and involve the morphologicallyless derived genera Doryctes Haliday and Ontsira Cam-eron. In both topologies, a clade with the species of the lat-ter two genera (BPP: 28SN, 28SA = 1.0) appears as thesister group of a clade with two divisions, one containingall the non-doryctine subfamilies except rhyssalines, andthe other one with the remaining doryctine genera. Thismajor doryctine clade (BPP: 28SN, 28SA = 1.0) shows anapparent geographic structure that is composed of twomain clades. One of these contains genera mainly distrib-uted in the Palaearctic, Oriental, and Afrotropical regions(BPP: 28SN = 0.62; 28SA = 0.56). The second clade hasthe Neotropical Megaloproctus Schulz at the base (onlywith 28SN; BPP = 0.58), followed by a clade with two spe-cies of Spathius + Caenophanes Foerster (BPP: 28SN,28SA = 1.0), then by a clade with the two genera of thecosmopolitan Holcobraconini, Liobracon Szepligeti andOdontobracon Cameron (BPP: 28SN, 28SA = 1.0), a cladewith two endemic Australian genera, Jarra Marsh & Aus-tin and Syngaster Brulle (BPP: 28SN, 28SA = 1.0), andfinally by a large, significantly supported clade representedby doryctine genera that are mainly distributed in the Neo-tropics (BPP: 28SN = 0.69; 28SA = 0.98).
The five gall-associated doryctine genera were recoveredtogether in the two simultaneous analyses in a single cladewithin the large ‘Neotropical’ clade, and recoding of theunalignable regions increased its BPP, though not to a sig-nificant value (BPP: 28SN = 0.49; 28SA = 0.7). Moreover,a clade with all the gall associated genera excluding Perc-
nobracon appeared significantly supported in the28SA + COI topology (BPP = 0.95). The clade with thefive gall-associated genera was recovered as the sister groupof a clade with the two included species of the widely dis-tributed and polyphagous Heterospilus, and this relation-ship increased its BPP to a significant value when theunalignable regions were recoded (BPP: 28SN + COI =0.69; 28SA + COI = 0.99).
Most of the topologies derived from the MP and Bayes-ian analyses using the multiple alignments for the 28S with

RhaconotusPlatyspathius
LeptorhaconotusIpodoryctes
Spathius (Antespathius)Rhacontsira
Hemidoryctes
CaenopachysEuscelinus
LiobraconOdontobracon
SchlettereriellaPseudodoryctes
RinambaCystomastax
PseudorhysipolisPambolus
Megaloproctus
Caenophanes
Spathius sp.Spathius agrili
SyngasterJarra
Heterospilus prosopidisHeterospilus sp.
AllorhogasPsenobolus
MonitoriellaPercnobraconSemirhytus
JohnsoniusPedinotus
LamquetiaAcanthorogas
Curtisella
HecabolusStenocorse
AphelopsiaBarbalhoa
Histeromeroides
EcphylusAivalykus
OndigusAcrophasmus
Notiospathius
MasoniusHansonorum
TarascoStiropius
SpathiomorphaHypodoryctes
HormiusDoryctes leucogaster
Doryctes heydeniOntsira 0.01 substitutions/site
OncophanesRhyssalus
**
Labania
Fig. 1. 28SN Bayesian phylogram. Black circles above branches indicate Bayesian posterior probabilities P0.95 recovered in both the 28SN and 28SABayesian analyses. Asterisks indicate Bayesian posterior probabilities P0.95 only recovered in the separate 28SN Bayesian analysis. Hollow circlesindicate P0.95 only recovered in the 28SA Bayesian analysis. Names of the gall-associated genera are in bold.
A. Zaldivar-Riveron et al. / Molecular Phylogenetics and Evolution 44 (2007) 981–992 987
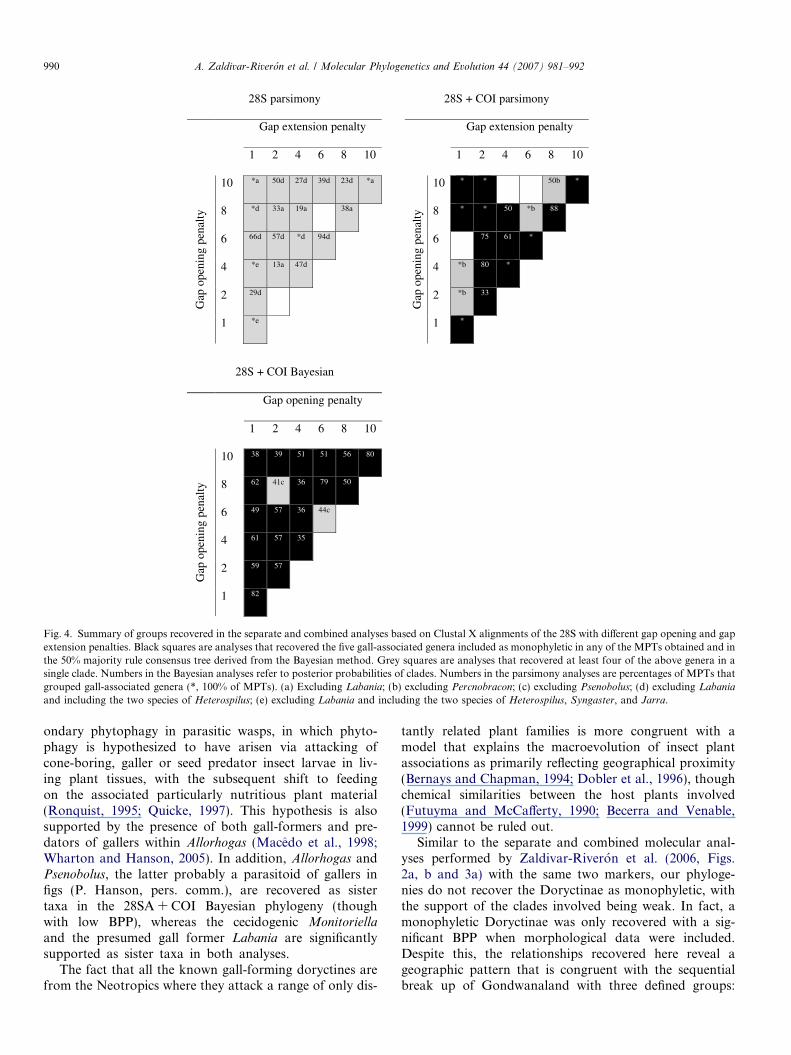
various gap opening-gap extension penalties recovered aclade with at least four of the gall-associated generaincluded (Fig. 4). Moreover, all these analyses placed thetaxa with elongated first metasomal tergite in separateclades along the tree. Six of the 21 MP analyses with the28S alone recovered MPTs containing a group with thegall-associated genera except Labania, and 13 analysesreconstructed MPTs that placed them together with thetwo species of Heterospilus or with the latter and the Aus-tralian Jarra and Syngaster. Fourteen of the combinedanalyses using MP recovered the monophyly of the five
gall-associated genera in at least some of their MPTsobtained, whereas 19 of the 50% majority rule consensustrees derived from the simultaneous Bayesian analyses alsorecovered this relationship.
3.2. Tests of alternative hypotheses
The Bayesian test of alternative phylogenetic hypothesesfor the two combined analyses showed that the topologiesrecovering the tribes Doryctini, Hecabolini and Spathiini(sensu Belokobylskij, 1992) each as monophyletic are statis-

RhaconotusPlatyspathius
RhacontsiraLeptorhaconotus
IpodoryctesCaenopachysEuscelinus
HemidoryctesRinamba
SchlettereriellaPseudodoryctes
HypodoryctesSpathiomorpha
MegaloproctusSpathius sp.
Spathius agriliCaenophanes
SyngasterJarra
Heterospilus sp.Pedinotus
LamquetiaCurtisella
HecabolusNotiospathius
MasoniusHansonorium
HisteromeroidesTarasco
AphelopsiaStenocorse
AcrophasmusBarbalhoa
Ondigus
SemirhytusJohnosonius
AivalykusAcantorhogas
LiobraconOdontobracon
DoryctesCystomastax
HormiusPseudorhysipolis
PambolusStiropius
Doryctes heydoniOntsira
OncophanesRhyssalus
MonitoriellaLabania
Allorhogas
Percnobracon
Heterospilus prosopidis
0.01 substitutions/site
Fig. 2. COI Bayesian phylogram. Black circles above branches indicate Bayesian posterior probabilities P0.95. Names of the gall-associated genera are inbold.
988 A. Zaldivar-Riveron et al. / Molecular Phylogenetics and Evolution 44 (2007) 981–992
tically rejected (P < 0.05). The SH tests also show that thethree Bayesian topologies derived from the 28SN + COIdata set with the above tribes forced to be monophyleticare significantly worse explanations of the data than theunconstrained Bayesian topology (P < 0.0001). The Bayes-ian alternative topologies used for the latter test can beretrieved from the electronic supplementary material.
3.3. Character evolution
Fig. 3 shows the ancestral states of the two examinedfeatures that were mapped onto the 28SN + COI Bayesianphylogeny based on maximum likelihood optimizations.
Origin of gall association appears significantly supportedonce in the Percnobracon + Psenobolus + Allorho-
gas + Labania + Monitoriella clade. On the other hand,at least eight origins are shown for a moderately to consid-erably elongated acrosternite in the first tergite. This fea-ture is shown to have evolved three times within theRhaconotini clade [Platyspathius Viereck, Leptorhaconotus
Granger and Spathius (Antespathius)], twice within theremaining members of the ‘Palaearctic-Oriental-Afrotropi-cal’ clade with Schlettereriella Szepligeti and Spathiomor-
pha Tobias, once with the species of Spathius (Spathius
Belokoblyskij), and two times within the ‘Neotropical’clade, one with the gall-associated Percnobracon and Pse-

RhaconotusPlatyspathius
LeptorhaconotusIpodoryctes
Spathius (Antespathius)Rhacontsira
CaenopachysEuscelinus
HemidoryctesRinamba
SchlettereriellaPseudodoryctes
SpathiomorphaHypodoryctes
MegaloproctusSpathius sp.
Spathius agriliCaenophanes
LiobraconOdontobracon
SyngasterJarra
Heterospilus prosopidisHeterospilus sp.Allorhogas
LabaniaMonitoriella Gall associated
generaPsenobolusPercnobracon
SemirhytusJohnsoniusPedinotus
LamquetiaAcanthorogas
HecabolusCurtisella
Stenocorse
AphelopsiaHisteromeroides
NotiospathiusMasonius
HansonorumTarasco
EcphylusAivalykus
Barbalhoa
OndigusAcrophasmus
CystomastaxPseudorhysipolis
PambolusStiropius
HormiusDoryctes leucogaster
Doryctes heydeniOntsira
OncophanesRhyssalus
0.01 sustitutions/site
Uncertain length of acrosternite
Non gall associatedAcrosternite 0.2-0.25 x as long as tergite
0.3-0.5 x as long as tergite
0.6-0.85 x as long as tergiteGall associated
*
Fig. 3. Character evolution of gall association and length of acrosternite of first metasomal tergite on the Bayesian 28SN + COI phylogram for doryctinegenera using maximum likelihood optimizations. Black circles indicate clades supported by BPP P 0.95 in both the 28SN + COI and 28SA + COIBayesian phylogenies. Open circles indicate clades supported by BPP P 0.95 only in the 28SA + COI Bayesian phylogeny. The asterisk indicates thesignificantly supported clade (BPP = 0.95) recovered in the 28SA + COI Bayesian analysis, but containing Psenobolus instead Percnobracon.
A. Zaldivar-Riveron et al. / Molecular Phylogenetics and Evolution 44 (2007) 981–992 989
nobolus, and the remaining one in the Notiospathius Mat-thews & Marsh + Masonius Marsh + Tarasco + Hansono-
rum Marsh clade.The full hierarchical Bayesian method shows that the
ancestral state being gall associated has a probability of0.93 for the clade involved.
4. Discussion
Our study shows phylogenetic evidence that suggests aclose evolutionary relationship among a group of gall-asso-
ciated wasps within the mainly parasitic wasp subfamilyDoryctinae. This relationship was recovered in our twosimultaneous phylogenetic analyses, though the cladeinvolved was only significantly supported in the 28SA +COI Bayesian topology. The result obtained was surprisingbecause the taxa involved display disparate external mor-phologies and are associated with distantly related plantfamilies.
The exclusive presence of cecidogenic and presumedgaller-predating taxa within a same clade agrees withthe generally accepted scenario about the origin of sec-
28S parsimony 28S + COI parsimony
Gap extension penalty Gap extension penalty
1 2 4 6 8 10 1 2 4 6 8 10
10 *a 50d 27d 39d 23d *a 10 * * 50b *
8 *d 33a 19a 38a 8 * * 50 *b 88
6 66d 57d *d 94d 6 75 61 *
4 *e 13a 47d 4 *b 80 *
2 29d 2 *b 33
Gap
open
ing
pena
lty
1 *e
Gap
open
ing
pena
lty
1 *
28S + COI Bayesian
Gap opening penalty
1 2 4 6 8 10
10 38 39 51 51 56 80
8 62 41c 36 79 50
6 49 57 36 44c
4 61 57 35
2 59 57
Gap
open
ing
pena
lty
1 82
Fig. 4. Summary of groups recovered in the separate and combined analyses based on Clustal X alignments of the 28S with different gap opening and gapextension penalties. Black squares are analyses that recovered the five gall-associated genera included as monophyletic in any of the MPTs obtained and inthe 50% majority rule consensus tree derived from the Bayesian method. Grey squares are analyses that recovered at least four of the above genera in asingle clade. Numbers in the Bayesian analyses refer to posterior probabilities of clades. Numbers in the parsimony analyses are percentages of MPTs thatgrouped gall-associated genera (*, 100% of MPTs). (a) Excluding Labania; (b) excluding Percnobracon; (c) excluding Psenobolus; (d) excluding Labania
and including the two species of Heterospilus; (e) excluding Labania and including the two species of Heterospilus, Syngaster, and Jarra.
990 A. Zaldivar-Riveron et al. / Molecular Phylogenetics and Evolution 44 (2007) 981–992
ondary phytophagy in parasitic wasps, in which phyto-phagy is hypothesized to have arisen via attacking ofcone-boring, galler or seed predator insect larvae in liv-ing plant tissues, with the subsequent shift to feedingon the associated particularly nutritious plant material(Ronquist, 1995; Quicke, 1997). This hypothesis is alsosupported by the presence of both gall-formers and pre-dators of gallers within Allorhogas (Macedo et al., 1998;Wharton and Hanson, 2005). In addition, Allorhogas andPsenobolus, the latter probably a parasitoid of gallers infigs (P. Hanson, pers. comm.), are recovered as sistertaxa in the 28SA + COI Bayesian phylogeny (thoughwith low BPP), whereas the cecidogenic Monitoriella
and the presumed gall former Labania are significantlysupported as sister taxa in both analyses.
The fact that all the known gall-forming doryctines arefrom the Neotropics where they attack a range of only dis-
tantly related plant families is more congruent with amodel that explains the macroevolution of insect plantassociations as primarily reflecting geographical proximity(Bernays and Chapman, 1994; Dobler et al., 1996), thoughchemical similarities between the host plants involved(Futuyma and McCafferty, 1990; Becerra and Venable,1999) cannot be ruled out.
Similar to the separate and combined molecular anal-yses performed by Zaldivar-Riveron et al. (2006, Figs.2a, b and 3a) with the same two markers, our phyloge-nies do not recover the Doryctinae as monophyletic, withthe support of the clades involved being weak. In fact, amonophyletic Doryctinae was only recovered with a sig-nificant BPP when morphological data were included.Despite this, the relationships recovered here reveal ageographic pattern that is congruent with the sequentialbreak up of Gondwanaland with three defined groups:

A. Zaldivar-Riveron et al. / Molecular Phylogenetics and Evolution 44 (2007) 981–992 991
one mainly composed of taxa from the Palaearctic,Oriental, and Afrotropical regions, a second one of Aus-tralian, and the remaining one of Neotropical taxa, withthe latter two being significantly supported as sistergroups.
The presence of suites of characters associated withadaptations of parasitism has commonly led to distantlyrelated wasp taxa being classified together (Achterberg,1988; Quicke, 1997; Quicke and Belshaw, 1999). Ourphylogenetic analyses have consistently shown that oneof the main morphological features employed to distin-guish the Spathiini, one of the largest tribes of the sub-family, has evolved on numerous separate occasions. Wesuggest that an extended first metasomal tergite in dis-tantly related lineages probably reflects convergence asa result of a shared life history strategy: the attack ofconcealed host larvae, as in the case of wood- andbark-boring coleopterans through enabling use of alonger ovipositor while maintaining a near vertical ‘dril-ling’ posture. Finally, the results of this study also sup-port a previous work that suggests the existence ofhigh levels of morphological homoplasy among the dif-ferent doryctine genera (Belokobylskij et al., 2004b),which can be inferred by the high disparity betweenthe morphological classification of the group and ourresulting hypotheses of phylogenies. This high level ofmorphological homoplasy is also observed by the signif-icant differences found between the likelihood of the28SN + COI phylogeny and the likelihood of alternativetopologies that enforced the larger doryctine tribes Hec-abolini, Spathiini, and Doryctini each as monophyletic.The significantly supported relationships presented inour study should therefore be used to construct an alter-native higher classification in the Doryctinae.
Acknowledgments
We thank Kees van Achterberg, Yves Braet, Jose Clav-ijo, Brian Fisher, Georg Georogen, Carolina Godoy, PaulHanson, Nina Laurenne, John Noyes, and Mark Shaw forproviding material for this study. This work was in partsupported by a Natural Environment Research CouncilGrant NE/C519538/1 to DLJQ (and Andy Purvis), a post-doctoral fellowship given by the Universidad NacionalAutonoma de Mexico (UNAM-DGAPA) to AZR, a Grantgiven by the Russian Foundation for Basic Research (No.07-04-00454) to S.A.B., a PhD scholarship given by theConsejo Nacional de Investigaciones Cientıficas y Tec-nologicas (CONICET) to J.J.M., and by two Grants givenby FONACIT-Venezuela (S1- 2000000479, LAB-2000001593) to R.B.
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at doi:10.1016/j.ympev.2007.05.016.
References
Achterberg, C. van., 1988. Parallelisms in the Braconidae (Hymenoptera)with special reference to the biology. In: Gupta, V.K. (Ed.), Advancesin Parasitic Hymenoptera Research. E.J. Brill, Leiden, pp. 85–115.
Austin, A.D., Dangerfield, P., 1998. Biology of Mesostoa kerri (Insecta:Hymenoptera: Braconidae: Mesostoinae), an endemic Australian waspthat causes stem galls on Banskia marginata. Austr. J. Bot. 46, 559–569.
Becerra, J.X., 1997. Insects on plants: macroevolutionary chemical trendsin host use. Science 276, 253–256.
Becerra, J.X., Venable, D.L., 1999. Macroevolution of insect-plantassociations: the relevance of host biogeography to host affiliation.Proc. Natl. Acad. Sci. 96, 12626–12631.
Belokobylskij, S.A., 1992. On the classification and phylogeny of thebraconid wasps subfamilies Doryctinae and Exothecinae (Hymenop-tera, Braconidae). Part I. On the classification, 1. Ent. Obozr. (InRussian) 71, 900–928, English translation 1993 Ent. Rev. 72: 109–137.
Belokobylskij, S.A., Iqbal, M., Austin, A., 2004a. Systematics, distribu-tion and diversity of the Australian doryctine wasps (Hymenoptera,Braconidae, Doryctinae). Rec. S. Austr. Mus. 8, 1–150.
Belokobylskij, S.A., Zaldivar-Riveron, A., Quicke, D.L.J., 2004b. Phy-logeny of the genera of the parasitic wasps subfamily Doryctinae(Hymenoptera: Braconidae) based on morphological evidence. Zool. J.Linn. Soc. 142, 369–404.
Belshaw, R., Fitton, M.G., Herniou, E., Gimeno, C., Quicke, D.L.J.,1998. A phylogenetic reconstruction of the Ichneumonoidea (Hyme-noptera) based on the D2 variable region of 28S ribosomal RNA. Syst.Ent. 23, 109–123.
Belshaw, R., Dowton, M., Quicke, D.L.J., Austin, A.D., 2000. Estimatingancestral geographic distributions: a Gondwanan origin for aphidparasitoids. Proc. R. Soc. (Lond.) B. 267, 491–496.
Belshaw, R., Quicke, D.L.J., 1997. A molecular phylogeny of theAphidiinae (Hymenoptera: Braconidae). Mol. Phylogenet. Evol. 7,281–293.
Bernays, E.A., Chapman, R.F., 1994. Host-plant Selection by Phytoph-agous Insects. Chapman & Hall, London.
Brandley, M.C., Schmitz, A., Reeder, T.W., 2004. Partitioned Bayesiananalyses, partition choice, and the phylogenetic relationships ofScincid Lizards. Syst. Biol. 54, 373–390.
Brethes, J., 1927. Nouveau hymenopteres parasites du Chili. Rev. Chil.Hist. Nat. 31, 194–200.
Buckley, T.R., Arensburger, P., Simon, C., Chambers, G., 2002.Combined data, Bayesian phylogenetics and the origin of the NewZealand cicada genera. Syst. Biol. 51, 4–18.
Dobler, S., Mardulyn, P., Pasteels, J.M., Rowell-Rahier, M., 1996. Host-plant switches and the evolution of chemical defence and life history inthe leaf beetle genus Oreina. Evolution 50, 2373–2386.
Dowton, M., Belshaw, R., Austin, A.D., Quicke, D.L.J., 2002. Simulta-neous molecular and morphological analysis of braconid relationships(Insecta: Hymenoptera: Braconidae) indicates independent mt-tRNAgene inversions within a single wasp family. J. Mol. Evol. 54, 210–226.
Fischer, M., 1981. Mononeuron, ein neues Doryctinen-Genus aus Brasilien(Hymenoptera, Braconidae). Z. Arb. Gem. Ost. Ent. 33, 47–51.
Folmer, O., Black, M., Hoeh, W., Lutz, R., Vrijenhoek, R., 1994. DNAprimers for amplification of mitochondrial cytochrome c oxidasesubunit I from diverse metazoan invertebrates. Mol. Mar. Biol.Biotechnol. 3, 294–299.
Futuyma, D.J., McCafferty, S.S., 1990. Phylogeny and the evolution ofhost plant associations in the leaf beetle genus Ophraella (Coleoptera,Chrysomelidae). Evolution 44, 1885–1913.
Gillespie, J.J., 2004. Characterizing regions of ambiguous alignmentcaused by the expansion and contraction of hairpin-stem loops inribosomal RNA molecules. Mol. Phylogenet. Evol. 33, 936–943.
Gillespie, J.J., Yoder, M.J., Wharton, R.A., 2005. Predicted secondarystructure for 28S and 18S rRNA from Ichneumonoidea (Insecta:Hymenoptera: Apocrita): impact on sequence alignment and phylog-eny estimation. J. Mol. Evol. 6, 114–137.

992 A. Zaldivar-Riveron et al. / Molecular Phylogenetics and Evolution 44 (2007) 981–992
Gladstein, D.S., Wheeler, W., 1996. Optimization alignment: the end ofmultiple sequence alignment in phylogenetics? Cladistics 12, 1–9.
Huelsenbeck, J.P., Bollback, J.P., 2001. Empirical and hierarchicalBayesian estimation of ancestral states. Syst. Biol. 50, 351–366.
Infante, F., Hanson, P., Wharton, R., 1995. Phytophagy in the genusMonitoriella (Hymenoptera: Braconidae) with description of newspecies. Ann. Ent. Soc. Am. 88, 406–415.
Kopf, A., Rank, N.E., Roininen, H., Julkunen-Tiitto, R., Pasteels, J.M.,Tahvanainen, J., 1998. The evolution of host-plant use and sequestra-tion in the leaf beetle genus Phratora (Coleoptera: Chrysomelidae).Evolution 52, 517–528.
Kieffer, J.J., 1910. In: Kieffer, J.J., Jorgensen, P. Gallen und Gallentier ausArgentinien. Centr. F. Bakt. Parasit. Infekt. 27: 362–442 (in German).
Lewis, P.O., 2001. A likelihood approach to estimating phylogeny fromdiscrete morphological character data. Syst. Biol. 50, 913–925.
Lutzoni, F., Wagner, P., Reeb, V., Zoller, S., 2000. Integrating ambig-uously aligned regions of DNA sequences in phylogenetic analysiswithout violating positional homology. Syst. Biol. 49, 628–651.
Maddison, D.R., Maddison, W.P., 2003. MacClade version 4.06. Sinauer,Sunderland, MA.
Maddison, W.P., Maddison, D.R., 2004. Mesquite: a modular system forevolutionary analysis, v. 1.05. Available from: http://www.mesquiteproject.org.
Macedo, M.V., Monteiro, R.T., 1989. Seed predation by a braconid wasp,Allorhogas sp. (Hymenoptera). J. New York Ent. Soc. 97, 358–362.
Macedo, M.V., Pimentel, M.C.P., Vieira, R.C., 1998. Response ofPithecellobium tortum Martius (Leguminosae) seeds to the attack ofthe phytophagous braconid wasp, Allorhogas sp. (Hymenoptera). J.New York Ent. Soc. 97, 358–362.
Mardulyn, P., Whitfield, J.B., 1999. Phylogenetic signal in the COI,16S, and 28S genes for inferring relationships among genera ofMicrograstrinae (Hymenoptera; Braconidae): evidence of a highdiversification rate in this group of parasitoids. Mol. Phylogenet.Evol. 12, 282–294.
Marsh, P.M., 2002. The Doryctinae of Costa Rica (excluding the genusHeterospilus). Mem. Am. Ent. Inst. 70, 1–319.
Marsh, P.M., Macedo, M.V., Pimentel, M.C.P., 2000. Descriptions andbiological notes on two new phytophagous species of the genusAllorhogas from Brasil (Hymenoptera: Braconidae: Doryctinae). J.Hymenopt. Res. 9, 292–297.
Martınez, J.J., 2006. Three new species of Percnobracon Kieffer &Jorgensen (Hymenoptera: Braconidae) from Argentina, reared fromcecidomyiid (Diptera) and eurytomid (Hymenoptera) galls. Zootaxa1282, 49–58.
Nylander, J.A.A., 2004. MrModeltest, v2. Evolutionary Biology Centre,Uppsala University. Available from: http://www.ebc.uu.se/systzoo/staff/nylander.html.
Oda, R.A.M., de Macedo, M.V., Quicke, D.L.J., 2001. First biologicaldata for Aspilodemon Fischer (Hymenoptera: Braconidae: Hydran-geocolinae): parasitoids of cecidomyiid fly galls on Asteraceae inBrazil. J. Hymenopt. Res. 10, 126–130.
Pagel, M., 1994. Detecting correlated evolution on phylogenies: a generalmethod for the comparative analysis of discrete characters. Proc. R.Soc. (Lond.) B 255, 37–45.
Penteado-Dias, A.M., 2000. Biological notes and larval morphology ofDonquickea (Hymenoptera: Braconidae: Doryctinae). In: Austin,A.D., Dowton, M. (Eds.), Hymenoptera Evolution, Biodiversity and
Biological Control. CSIRO Publishing, Collingwood, Australia, pp.296–299.
Pickett, K.M., Randle, C.P., 2005. Strange Bayes indeed: uniformtopological priors imply non-uniform clade priors. Mol. Phylogenet.Evol. 34, 203–211.
Quicke, D.L.J., 1997. Parasitic Wasps. Chapman & Hall, London.Quicke, D.L.J., Belshaw, R., 1999. Incongruence between morphological
data sets: an example from the evolution of endoparasitism amongparasitic wasps (Hymenoptera: Braconidae). Syst. Biol. 48, 436–454.
Quicke, D.L.J., Ficken, L., Fitton, M.G., 1992. New diagnostic ovipositorcharacters for doryctine wasps (Hymenoptera, Braconidae). J. Nat.Hist. 26, 1035–1046.
Quicke, D.L.J., Taylor, J., Purvis, A., 2001. Changing the landscape: anew strategy for estimating large phylogenies. Syst. Biol. 50, 60–66.
Ramirez, W.B., Marsh, P.M., 1996. A review of the genus Psenobolus
(Hymenoptera: Braconidae) from Costa Rica, an inquiline fig waspwith brachypterous males, with description of two new species. J.Hymenopt. Res. 5, 64–72.
Reeder, T.W., 2003. A phylogeny of the Australian Sphenomorphus group(Scincidae: Squamata) and the phylogenetic placement of the crocodileskinks (Tribolonotus): Bayesian approaches to assessing congruenceand obtaining confidence in maximum likelihood inferred relation-ships. Mol. Phylogenet. Evol. 27, 384–397.
Ronquist, F., 1995. Phylogeny and early evolution of the Cynipoidea.Syst. Ent. 20, 309–335.
Ronquist, F., Huelsenbeck, J.P., 2003. MRBAYES 3: Bayesian phyloge-netic inference under mixed models. Bioinformatics 19, 1572–1574.
Shimodaira, H., Hasegawa, M., 1999. Multiple comparisons of log-likelihoods with applications to phylogenetic inference. Mol. Biol.Evol. 16, 1114–1116.
Simmons, M.P., Ochoterena, H., 2000. Gaps and characters in sequence-based phylogenetic analyses. Syst. Biol. 49, 369–381.
Swofford, D.L., 1998. PAUP*: Phylogenetic Analysis Using Parsimony (*and other methods), version 4.0b10. Sinauer Associates, Sunderland,MA.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins,D.G., 1997. The ClustalX windows interface: flexible strategies formultiple sequence alignment aided by quality analysis tools. Nucl.Acids Res. 25, 4876–4882.
Thompson, J.N., 1999. What we know and do not know aboutcoevolution: insect herbivores and plants in a test case. In: Olff, H.,Brown, V.K., Drent, R.H. (Eds.), Herbivores: Between Plants andPredators. Blackwell Science, Oxford, pp. 7–30.
Wharton, R.A., 1993. Review of the Hormiini (Hymenoptera: Braconi-dae) with a description of new taxa. J. Nat. Hist. 27, 107–171.
Wharton, R.A., Hanson, P.E., 2005. Gall wasps in the family Braconidae(Hymenoptera). In: Raman, A., Schaefer, W.C., Withers, T.M. (Eds.),Biology, Ecology, and Evolution of Gall-inducing Arthropods. SciencePublishers, Enfield, New Hampshire, pp. 495–505.
Wheeler, W.C., 1996. Optimization alignment: the end of multiplesequence alignment in phylogenetics? Cladistics 12, 1–9.
Wheeler, W.C., Gladstein, D.S., 1994. MALIGN: a multiple sequencealignment program. J. Hered. 85, 417–418.
Zaldivar-Riveron, A., Mori, M., Quicke, D.L.J., 2006. Systematics of thecyclostome subfamilies of braconid parasitic wasps (Hymenoptera:Ichenumonoidea): a simultaneous molecular and morphologicalBayesian approach. Mol. Phylogenet. Evol. 38, 130–145.




















