
A review and tests of hypotheses about causes of the KwaZulu-Natal sardine run
33
PLEASE SCROLL DOWN FOR ARTICLE This article was downloaded by: On: 16 November 2010 Access details: Access Details: Free Access Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37- 41 Mortimer Street, London W1T 3JH, UK African Journal of Marine Science Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t911470580 A review and tests of hypotheses about causes of the KwaZulu-Natal sardine run P. Fréon a ; J. C. Coetzee b ; C. D. van der Lingen bc ; A. D. Connell d ; S. H. O'Donoghue e ; M. J. Roberts f ; H. Demarcq a ; C. G. Attwood c ; S. J. Lamberth bd ; L. Hutchings cf a IRD, Sète, France b Branch Fisheries, Department of Agriculture, Forestry and Fisheries, Rogge Bay, South Africa c Marine Research Institute, University of Cape Town, Rondebosch, South Africa d South African Institute for Aquatic Biodiversity, Grahamstown, South Africa e School of Biological and Conservation Sciences, Westville Campus, University of KwaZulu-Natal, Durban, South Africa f Oceans and Coasts, Department of Environmental Affairs, Rogge Bay, South Africa Online publication date: 08 November 2010 To cite this Article Fréon, P. , Coetzee, J. C. , van der Lingen, C. D. , Connell, A. D. , O'Donoghue, S. H. , Roberts, M. J. , Demarcq, H. , Attwood, C. G. , Lamberth, S. J. and Hutchings, L.(2010) 'A review and tests of hypotheses about causes of the KwaZulu-Natal sardine run', African Journal of Marine Science, 32: 2, 449 — 479 To link to this Article: DOI: 10.2989/1814232X.2010.519451 URL: http://dx.doi.org/10.2989/1814232X.2010.519451 Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of A review and tests of hypotheses about causes of the KwaZulu-Natal sardine run

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by:On: 16 November 2010Access details: Access Details: Free AccessPublisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
African Journal of Marine SciencePublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t911470580
A review and tests of hypotheses about causes of the KwaZulu-Natalsardine runP. Fréona; J. C. Coetzeeb; C. D. van der Lingenbc; A. D. Connelld; S. H. O'Donoghuee; M. J. Robertsf; H.Demarcqa; C. G. Attwoodc; S. J. Lamberthbd; L. Hutchingscf
a IRD, Sète, France b Branch Fisheries, Department of Agriculture, Forestry and Fisheries, Rogge Bay,South Africa c Marine Research Institute, University of Cape Town, Rondebosch, South Africa d SouthAfrican Institute for Aquatic Biodiversity, Grahamstown, South Africa e School of Biological andConservation Sciences, Westville Campus, University of KwaZulu-Natal, Durban, South Africa f
Oceans and Coasts, Department of Environmental Affairs, Rogge Bay, South Africa
Online publication date: 08 November 2010
To cite this Article Fréon, P. , Coetzee, J. C. , van der Lingen, C. D. , Connell, A. D. , O'Donoghue, S. H. , Roberts, M. J. ,Demarcq, H. , Attwood, C. G. , Lamberth, S. J. and Hutchings, L.(2010) 'A review and tests of hypotheses about causes ofthe KwaZulu-Natal sardine run', African Journal of Marine Science, 32: 2, 449 — 479To link to this Article: DOI: 10.2989/1814232X.2010.519451URL: http://dx.doi.org/10.2989/1814232X.2010.519451
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

African Journal of Marine Science 2010, 32(2): 449–479
Printed in South Africa — All rights reserved
Copyright © NISC (Pty) Ltd
AFRICAN JOURNAL OFMARINE SCIENCE
ISSN 1814–232X EISSN 1814–2338
doi: 10.2989/1814232X.2010.519451
African Journal of Marine Science is co-published by NISC (Pty) Ltd and Taylor & Francis
A review and tests of hypotheses about causes of the KwaZulu-Natal sardine run
P Fréon1*, JC Coetzee2, CD van der Lingen2,3, AD Connell4, SH O’Donoghue5, MJ Roberts6, H Demarcq1, CG Attwood3,
SJ Lamberth2,4 and L Hutchings3,6
1 IRD, UMR 212 EME, CRHMT, avenue Jean Monnet, 34209, Sète, France2 Branch Fisheries, Department of Agriculture, Forestry and Fisheries, Private Bag X2, Rogge Bay 8012, South Africa3 Marine Research Institute, University of Cape Town, Private Bag X3, Rondebosch 7701, South Africa4 South African Institute for Aquatic Biodiversity, Private Bag 1015, Grahamstown 6140, South Africa5 School of Biological and Conservation Sciences, Westville Campus, University of KwaZulu-Natal, Private Bag X54001,
Durban 4000, South Africa6 Oceans and Coasts, Department of Environmental Affairs, Private Bag X2, Rogge Bay 8012, South Africa
* Corresponding author, e-mail: [email protected]
Manuscript received June 2010; accepted August 2010
The term ‘sardine run’ is part of the cultural heritage of the South African nation and refers to a natural
phenomenon that is well known to the general public but still poorly understood from an ecologi-
cal perspective. This lack of understanding has stimulated numerous hypotheses, often contradic-
tory, that try to explain why (ultimate factors) and how (proximate factors) the run occurs. Here, we
provide a new definition of the term sardine run, review the various hypotheses about the run, and
propose ways to test those hypotheses. Where possible, the results of tests that have been conducted
thus far are presented and discussed. Our interpretation of the causes is that the sardine run most
likely corresponds to a seasonal (early austral winter) reproductive migration of a genetically distinct
subpopulation of sardine that moves along the coast from the eastern Agulhas Bank to the coast of
KwaZulu-Natal (KZN) as far as Durban and sometimes beyond, in most years if not in every year. This
eastward migration is constrained close to the coast by the thermal preference of sardine and the
strong and warm offshore Agulhas Current. The run is facilitated by the presence of a band of cooler
coastal water and by the occurrence of Natal Pulses and break-away eddies that enable sardine shoals
to overcome their habitat restrictions. These enabling mechanisms are most important in the area
where the shelf is at its narrowest and feature most prominently off Waterfall Bluff, which has led to the
coining of the ‘Waterfall Bluff gateway hypothesis’. Based on the collection of eggs off the KZN coast,
sardine remain there for several months and their westward, return migration during late winter to
spring is nearly always unnoticeable because it likely occurs at depth as the fish avoid warmer surface
waters. Years in which the sardine run is not detected by coastal observers could reflect either its real
absence due to high water temperatures and/or other hydrographic barriers, or an eastward migration
that is farther offshore and possibly deeper and is enabled by hydrographical anomalies.
Keywords: migration, proximate factors, Sardinops sagax, South Africa, ultimate factors
The term ‘sardine run’ (or ‘KwaZulu-Natal sardine run’)
is part of the cultural heritage of the South African nation
and refers to a well-known natural phenomenon off the
east coast of South Africa. The sardine run has long been
known by the coastal population in that region because of
its economic importance (van der Lingen et al. 2010a), and
over the last few decades has been a topic of much media
attention and become the focus of a growing tourism industry
(Dicken 2010, Myeza et al. 2010). Despite this heightened
interest, the ecological aspects of this phenomenon remain
poorly understood, as evidenced by numerous hypotheses,
often contradictory, that try to explain why and how the
sardine run occurs. In this paper, we first attempt to provide
a neutral and consensual meaning of the term sardine run
as perceived by the general public and scientists alike.
Because of the commercial/recreational/public nature of
the sardine run, which requires some kind of management
response that will depend on how accurately the timing
and intensity of the ‘run’ can be predicted, our description
of what constitutes the run focuses on its visible aspects
Introduction
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings450
when it reaches the coast and elicits a human reaction. The
ecological meaning of the sardine run and its management
implications are separate issues that are also addressed in
this paper.
We define the term sardine run as the visible effects of the
coastal, alongshore movement during early austral winter of
a small and variable fraction of the South African population
of sardine Sardinops sagax from the eastern Agulhas Bank
to the KwaZulu-Natal (KZN) coast, as far as Durban and the
north coast of KZN. The sardine run is usually associated
with foraging top predators including seabirds, mammals
(O’Donoghue et al. 2010a, 2010b), and sharks and gamefish
(Dudley and Cliff 2010, Fennessy et al. 2010) that facilitate
its visual detection. The north-east displacement is usually
initiated from late May to early June and is mostly visually
detected and commercially exploited (via beach-seine
netting and tourism operations) when it occurs within a few
hundred metres of the coast. The duration of this coastal
spectacle is highly variable, lasting until the end of June or
even September at times (O’Donoghue et al. 2010c). Little is
known about farther offshore movements of sardine and even
less about the likely return of surviving fish to the Agulhas
Bank thereafter, although eggs in plankton samples suggest
that sardine remain in KZN waters until as late as December
(Connell 2010). Consequently, it is unknown whether or not
the sardine run corresponds to a conventional seasonal
migration that occurs every year but is only partly visible
in some years. Similarly, it is uncertain whether or not the
individuals that perform the sardine run belong to a different
population or subpopulation of sardine from those that inhabit
the west and south coasts of South Africa. Furthermore,
questions regarding the reasons for the variable occurrence,
intensity and duration of the sardine run at different spatial
and temporal scales are still unanswered.
Several hypotheses have been proposed to explain why
and how the sardine run occurs. In this paper, we describe
these hypotheses and examine them in the light of evidence
in the literature, including papers presented in this suite, and
either reject the hypothesis or suggest future research that
might yield further evidence to support a particular hypoth-
esis. General concepts of spatial behaviour that make
a distinction between ‘proximate’ and ‘ultimate’ factors
motivating this behaviour are first presented, followed
by the methods specific to this study and, where relevant,
summaries of those employed in preceding papers of the
suite (van der Lingen et al. 2010b). Hypotheses related to
ultimate and proximate factors of the sardine run and their
corresponding tests, some of which are relevant to more
than one hypothesis, are then detailed. Results of the tests
performed are presented and summarised, followed by a
general discussion that provides insight gathered from these
analyses, and contributes to our present understanding of
the sardine run.
General concepts of spatial behaviour
Proximate versus ultimate factors
Our investigation into the causes of the sardine run is
set against a conceptual framework of animal movement
behaviour and its ultimate and proximate causes. Following
Tinbergen (1963), Noakes (1992) applied the general
terms ‘ultimate’ and ‘proximate’ to refer to different factors
influencing behaviour. Ultimate refers to the final, long-term,
evolutionary consequences of behaviour (function), whereas
proximate refers to the immediate, short-term, physiological
mechanisms of behaviour (causation). Instantaneous habitat
selection is related to proximate cues and is expected to
occur on a small spatial scale (microhabitat) in response
to changes in the environment. A common example of a
proximate factor, likely relevant to the sardine run, is the
avoidance of warm waters and associated oxygen depletion.
This instantaneous selection and its relationship with the
environment are usually easy to observe and quantify. In
contrast, ultimate relationships may be much more difficult to
resolve. Additionally, once a selective pressure has led to the
evolution of a particular behaviour, that behaviour may last
for epochs even when major change occurs in the environ-
ment, providing that the behaviour (termed relic behaviour)
has at most a minimal cost regarding survival and reproduc-
tive success. Examples of ultimate factors are availability of
appropriate food, low potential predation (not instantaneous
predator avoidance) or favourable conditions for reproduc-
tive success.
An understanding of ultimate causes should also provide
insight into the long-term effects of fishing on movement
behaviour. Modern fishing practices have massive eco logical
influences that can impose strong selection differentials (Law
2007). Of particular relevance to this review are studies of fish
that display inter- and intra-population variability in movement
behaviour. The contributions of conditional (environmental)
and unconditional (inherited) factors to this variation is
important. These contributions will be enhanced if the
conditional variation is density-dependant or if the uncondi-
tional variants are exposed to differential fishing mortality
(Attwood and Cowley 2005, Dawson et al. 2006), as is likely
the case for the sardine run.
Types of movement
Although mechanisms of movement may differ, the selection
pressures experienced by aquatic and terrestrial species
are not dissimilar. Classification schemes and definitions
of movement are numerous, and scientists studying fish
movement frequently refer to, among others, residency, natal
homing, nomadism, dispersal, migration and emigration,
although some of these terms are applied loosely and often
not consistently. When considering movement rules for
individual-based models of fish, Tyler and Rose (1994) drew
a distinction between pattern-matching rules and process-
matching rules. Apart from the patterns, the theoretical and
evolutionary models that underpin each type of behaviour
need to be considered.
Some classifications have been based on physical descrip-
tions of the movement (e.g. distance, consistency in
direction, repetition, etc.), whereas others have been based
on biological events (e.g. feeding, spawning, home-range
relocation, etc.). The classification scheme presented by
Dingle (1996) draws on studies across the animal kingdom,
and provides a useful reference (Table 1). Station-keeping
is a widely adopted strategy, even among fish (e.g. Gerkin
1959), but is not likely to apply to sardine. It is the distinc-
tion between ranging and migration that is of relevance to the
current investigation. Ranging (Dingle 1996), or nomadism
Downloaded At: 21:50 16 November 2010

African Journal of Marine Science 2010, 32(2): 449–479 451
(Sinclair 1984), is a movement pattern that results from
a search for suitable resources and perhaps for a new
home-range. Animals searching for new resources cease
moving once these resources have been located. Definitions
of the home-range are not always practical and include
the following: the area in which an animal normally lives
(Smith 1980); the area that an animal occupies exclusive of
long-range migrations and erratic wanderings (Mace et al.
1984); a specific area that is repeatedly used in the course
of an animal’s activities; and a relatively circumscribed
area over which an organism travels to acquire resources it
needs for survival and reproduction (Dingle 1996). Under the
umbrella of the optimal foraging theory, the marginal value
theorem states that an animal will remain in a particular
location (patch) until the rate of energy gain falls below the
rate achievable elsewhere (Charnov 1976).
Sinclair (1984) uses three terms to describe one-way
movements: emigration, by which the animal moves along
a predetermined direction; dispersal, by which the animal
moves in an unpredictable direction; and nomadism, by
which the animal moves with no consistent direction. Harden
Jones (1968) draws a similarity between wanderings and
dispersal, both of which describe widespread movements of
fish away from the breeding area in search of food. Shields
(1984) defines dispersal as the movement of an animal away
from their site of origin to a new area, or succession of areas.
Dingle’s (1996) ranging is perhaps similar to Sinclair’s (1984)
nomadism and Shields’ (1984) dispersal, but Dingle (1996)
notes that there will remain confusion between foraging and
ranging and between ranging and migration, largely because
specific distinctions have not been looked for.
Migration is typically viewed as a regular round-trip within
the lifespan of an animal (Sinclair 1984), with the basic
plan of a fish’s migration usually being a triangle (Harden
Jones 1968). The free-floating egg is spawned at point A,
whereupon it drifts with the current and develops into a larval
stage. The larval stage reaches a nursery ground, B, where
it develops into a juvenile fish that ultimately recruits to the
feeding ground, C. The adult fish migrates from C to A to
spawn, an act that it may do once only, as is the case for
eels, or repeatedly, over several years, as is the case for
plaice. The most favoured explanation for such a pattern is
that the spawning ground must be upstream of a favourable
nursery ground, hence adults must undertake a compen-
satory upstream migration. The definition of migration may
be broadened and defined as a syndrome that involves
behavioural, anatomical and physiological adaptations.
Harden Jones (1968), for example, defines migration as
‘the class of movement which impels the migrants to return
to the region from where they have migrated’. According to
Dingle (1996), migration should not be defined in terms of
the movement, but rather in terms of the mover. He adopts
a modification of Kennedy’s (1985) definition: ‘Migratory
behaviour is persistent and straightened-out movement
effected by the animal’s own locomotory exertions or by
its active embarkation on a vehicle. It depends on some
temporary inhibition of station-keeping responses, but
promotes their eventual disinhibition and recurrence’. Accord-
ing to this definition, migration involves five attributes that
distinguish it from other movement types (Dingle 1996):
the animal displays persistent motion that takes it beyond 1.
the home-range;
the movement is direct, and not erratic in direction;2.
the animal passes suitable resources that would otherwise 3.
not be overlooked;
the animal engages in specific departure and arrival 4.
behaviour; and
physiological adjustments are made to reserve energy for 5.
a long journey.
Migrants commonly depart before existing resources are
depleted, and, ultimately, migration places the animal (or its
offspring) among favourable resources, thus avoiding the
possibility of it being marooned in a degrading environment
with insufficient resources. Distinctions have been made
between alimental, climatic and gametic migration (Harden
Jones 1968). Often associated with gametic migration is the
phenomenon of natal homing (Carr 1967), made famous by
salmon, the eel Anguilla and turtles, but known to occur also
among truly pelagic fish (Cury 1994, Rooker et al. 2008).
These definitions generate testable hypotheses that we
apply to the sardine run.
Mixed strategies
A difference in behavioural patterns among individuals
within a population is not an anomalous situation, but rather
it might be an evolutionary stable strategy that maintains
a balance between station-keepers and colonisers. At the
root of the split is density dependence. Crowded popula-
tions near or over carrying capacity may respond with
ranging behaviour gaining ascendancy over station-keeping,
implying also that movements at the extreme of the range
are linked to resource limitation within the population. An
example of this is detailed below under the basin theory
hypothesis (MacCall 1990).
Mixed behavioural strategies are common within species
and populations (Swingland 1984) and can easily confuse
the study of fish movement. Movement behaviour is rarely
consistent throughout a species, because most species
Movement type Characteristics
Station-keeping
Kinesis
Foraging
Commuting
Territoriality
Ranging (= nomadism, or = dispersal)
Migration (= emigration + immigration, or = one-way emigration)
Movements that serve to keep an animal stationary
Feeding movements within a home-range
Diel movements between day and night locations
Territorial defence and aggression, non-overlapping home-ranges
Exploratory movements over wide areas in search of resources
Persistent, directed, non-exploratory, predictable, physiological adaptation
Table 1: Dingle’s (1996) classification of animal movement behaviour, with synonyms inserted in brackets
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings452
show qualitative (i.e. type of movement) and quantitative
(extent, routes, timing, etc.) variation (Dingle 1996). Nowhere
is this variation more apparent than among salmonids,
where migrants (e.g. sea trout) and residents (e.g. lake trout)
show a behavioural distinction that is linked to phenomenal
life-history differences. The migration option involves trade-
offs: salmonid migrants have access to a marine food supply
and grow faster, and as a result are more fecund. However,
they also run a higher mortality risk on their seaward journey,
and they start breeding one or two years later than resident
salmonids (Jonsson and Jonsson 1993). The various forms
among salmonids actually look different (in size, colour
and shape), but among other species external differences
between migrants and residents are not generally obvious.
Japanese sardine Sardinops sagax melanosticus can be
divided into two groups (Hiramoto 1991): a coastal group
that lives in bays and has a limited migration range, and an
oceanic group that displays extended seasonal migrations.
This species shows a correlation between growth and
migration range. If the South African sardine that engage in
the run form a separate race, they should display discern-
ible physical or physiological characters, and their behaviour
should be explained against current and perhaps past
environments. For the sardine run to be a successful and
repeatable strategy, as opposed to maladaptive behaviour, it
will surely be contingent on an overall advantage regarding
reproductive success, given the high mortality likely experi-
enced by fish engaged in the run. What are the biotic and
environmental conditions that might favour such a trade-off
of life-expectancy for reproductive success? If such a
trade-off exists, it must surely leave a discernible mark on
population structure and individual resource allocation.
Material and methods
Most of the evidence relating to the sardine run and the tests
proposed for the various hypotheses are based on material
and methods already used and described in this suite (van
der Lingen et al. 2010b) or in historical papers, and are
briefly described below.
Van der Lingen et al. (2010a) provide information on
catches of sardine by beach-seine nets along the KZN coast
during some years between 1965 and 2009. The total number
of catches sampled by month is not always available, but
data collected recently (2003–2009) suggest values ranging
from 1 to 98 hauls (SD 31.1). Van der Lingen et al. (2010c)
also supply information on some biological characteristics of
sardine sampled from KZN beach-seine catches, including
data on caudal length and age distributions, and meristic
and morphometric data, and compare these with similar
data collected during research surveys and from commercial
purse-seine catches taken elsewhere along the South African
coast.
Connell (2010) presents a monthly time-series of sardine
egg abundance data derived from weekly, surface ichthyo-
plankton samples collected from 1987 to the present off
Park Rynie (about 60 km south of Durban). Initially, samples
were collected from a site 4–5 km offshore in a water depth
of 40–50 m, but from early 1994 a second sample was also
collected from 500–800 m offshore (15 m water depth). This
sampling strategy is in contrast to sampling during large-scale
biomass surveys conducted from 1984 to 2007, during which
eggs were collected using a CalVET net (van der Lingen and
Huggett 2003) and, more recently, a continuous underway
fish egg sampler (CUFES; van der Lingen et al. 1998) along
cross-shelf survey transects.
O’Donoghue (2009) and O’Donoghue et al. (2010a, 2010c)
report on visual surveys of sardine and their predators during
the run, made from the shore, small planes and small boats
along the KZN coast from 1988 to 2005. Dudley and Cliff
(2010) report on the number of sharks captured by shark
nets deployed along the KZN coast and on the incidence of
sardine in their stomach contents.
Coetzee et al. (2010) describe the various acoustic and
biological sampling methods performed during the 2005
sardine-run survey conducted onboard the RS Africana in
association with a small inflatable boat that investigated
shallow waters. Amongst other data, hydro acoustic estimates
of sardine density, and length frequency measurements and
age data, were collected and processed. Oceanographic
data such as sea surface temperature (SST), current speed
and direction as well as chlorophyll a (Chl a) concentrations
were also analysed. Additional oceanographic data collected
during the 2005 survey are also presented by Roberts et
al. (2010), together with continuous current data recordings
from ADCP moorings deployed off Port Edward and Port
Alfred since 2005.
Satellite data were also used to examine spatial and
temporal patterns in SST and Chl a. SST monthly daytime
averages of the NOAA Pathfinder version 5 (V5; available
at http://data.nodc.noaa.gov/pathfinder) were processed to
provide a compilation of the data from the AVHRR sensor
at 4 km resolution (Kilpatrick et al. 2001). In order to avoid
coastal contamination and lack of data, the coastal SST
data were extracted for distances of 5–10 km from the coast.
Ocean colour data used to estimate Chl a concentration
were downloaded from the NASA Ocean colour group web
portal using Seadas software (http://oceancolor.gsfc.nasa.
gov/seadas/). Altimetry data were also downloaded from
the CCAR near-real time altimetry site (http://argo.colorado.
edu/~realtime/welcome/). In some Hovmöller diagrams the
monthly data were smoothed temporally and spatially.
Hypotheses about ultimate factors (Hu) and potential tests
Hypotheses related to ultimate factors that attempt to answer
the question why the sardine run occurs, and the tests of
these hypotheses, are listed in Table 2 and described below.
Hu1: The sardine run represents a subpopulation spawning
migration
Recent advances in fishery genetics dismiss the conventional
view that fish, especially those with pelagic early stages
and a highly mobile adult phase, tend to form geographi-
cally extended and genetically homogeneous populations
with limited local adaptation and speciation (see reviews in
Hauser and Carvalho 2008, Fromentin et al. 2009). Evidence
for some degree of subpopulation structuring in the southern
Benguela sardine population is accumulating, with studies of
distribution patterns at varying biomass levels (Coetzee et al.
2008) and observations of consistent spatial differences in
some biological characteristics (van der Lingen et al. 2009)
suggesting the presence of at least two subpopulations.
Downloaded At: 21:50 16 November 2010
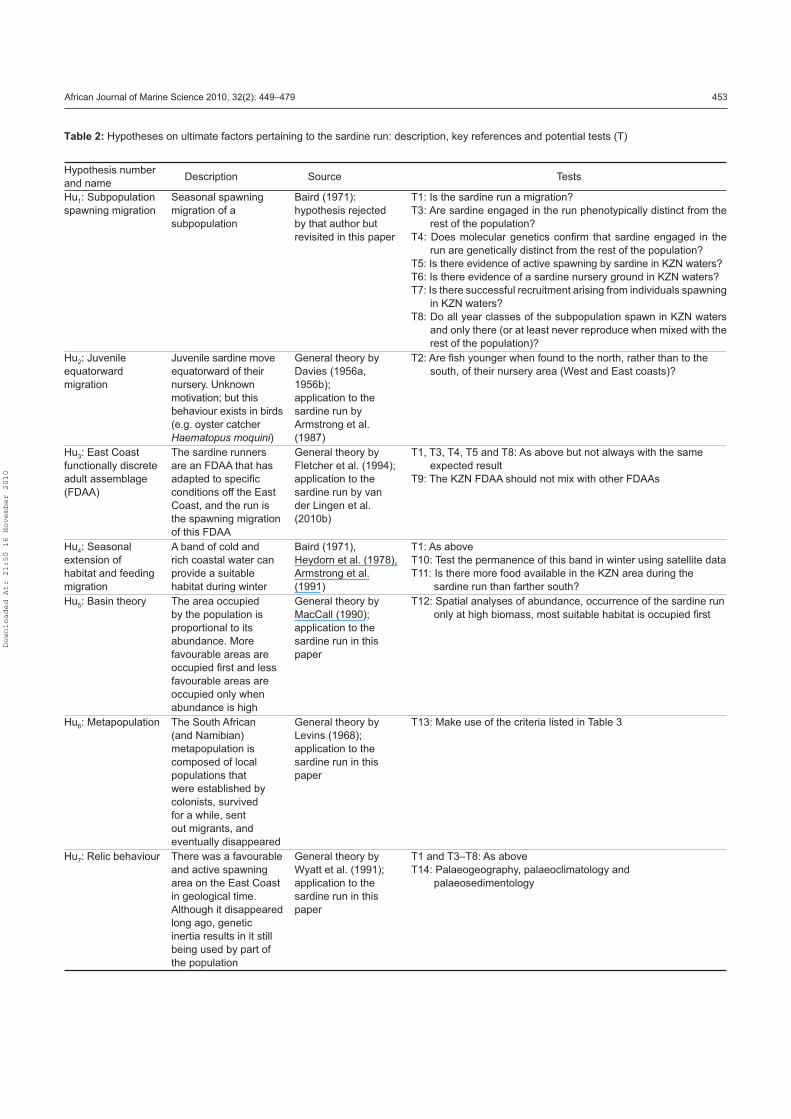
African Journal of Marine Science 2010, 32(2): 449–479 453
Hypothesis number
and name Description Source Tests
Hu1: Subpopulation
spawning migration
Seasonal spawning
migration of a
subpopulation
Baird (1971):
hypothesis rejected
by that author but
revisited in this paper
T1: Is the sardine run a migration?
T3: Are sardine engaged in the run phenotypically distinct from the
rest of the population?
T4: Does molecular genetics conÞ rm that sardine engaged in the
run are genetically distinct from the rest of the population?
T5: Is there evidence of active spawning by sardine in KZN waters?
T6: Is there evidence of a sardine nursery ground in KZN waters?
T7: Is there successful recruitment arising from individuals spawning
in KZN waters?
T8: Do all year classes of the subpopulation spawn in KZN waters
and only there (or at least never reproduce when mixed with the
rest of the population)?
Hu2: Juvenile
equatorward
migration
Juvenile sardine move
equatorward of their
nursery. Unknown
motivation; but this
behaviour exists in birds
(e.g. oyster catcher
Haematopus moquini)
General theory by
Davies (1956a,
1956b);
application to the
sardine run by
Armstrong et al.
(1987)
T2: Are Þ sh younger when found to the north, rather than to the
south, of their nursery area (West and East coasts)?
Hu3: East Coast
functionally discrete
adult assemblage
(FDAA)
The sardine runners
are an FDAA that has
adapted to speciÞ c
conditions off the East
Coast, and the run is
the spawning migration
of this FDAA
General theory by
Fletcher et al. (1994);
application to the
sardine run by van
der Lingen et al.
(2010b)
T1, T3, T4, T5 and T8: As above but not always with the same
expected result
T9: The KZN FDAA should not mix with other FDAAs
Hu4: Seasonal
extension of
habitat and feeding
migration
A band of cold and
rich coastal water can
provide a suitable
habitat during winter
Baird (1971),
Heydorn et al. (1978),
Armstrong et al.
(1991)
T1: As above
T10: Test the permanence of this band in winter using satellite data
T11: Is there more food available in the KZN area during the
sardine run than farther south?
Hu5: Basin theory The area occupied
by the population is
proportional to its
abundance. More
favourable areas are
occupied Þ rst and less
favourable areas are
occupied only when
abundance is high
General theory by
MacCall (1990);
application to the
sardine run in this
paper
T12: Spatial analyses of abundance, occurrence of the sardine run
only at high biomass, most suitable habitat is occupied Þ rst
Hu6: Metapopulation The South African
(and Namibian)
metapopulation is
composed of local
populations that
were established by
colonists, survived
for a while, sent
out migrants, and
eventually disappeared
General theory by
Levins (1968);
application to the
sardine run in this
paper
T13: Make use of the criteria listed in Table 3
Hu7: Relic behaviour There was a favourable
and active spawning
area on the East Coast
in geological time.
Although it disappeared
long ago, genetic
inertia results in it still
being used by part of
the population
General theory by
Wyatt et al. (1991);
application to the
sardine run in this
paper
T1 and T3–T8: As above
T14: Palaeogeography, palaeoclimatology and
palaeosedimentology
Table 2: Hypotheses on ultimate factors pertaining to the sardine run: description, key references and potential tests (T)
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings454
A ‘western’ subpopulation that is distributed to the west of
Cape Agulhas and an ‘eastern’ subpopulation distributed to
the east have been postulated (de Moor and Butterworth
2009a). The existence of these two putative subpopulations
is supported by results from separate assessment models
(de Moor and Butterworth 2009b), and from coupling an
individual-based model of larval drift to a 3D-hydrodynamic
model of the southern Benguela (Miller et al. 2006), which
indicated that sardine life-history strategy could be divided
between two main systems, separated at Cape Agulhas.
Regardless of the existence of the two abovementioned
subpopulations, sardine participating in the KZN sardine run
could represent the seasonal spawning (gametic) migration of
another discrete sardine subpopulation. This hypothesis was
assessed in earlier research (e.g. Baird 1971) by comparing
some biological characteristics of KZN sardine with those
sampled from commercial catches made off the Western
Cape (see T3 in Table 2). Variability in phenotypic charac-
teristics may be due to genetic factors or environmental
influences, and is usually associated with the geographical
region occupied by a subpopulation (Begg and Waldmann
1999). Baird (1971) rejected the hypothesis of an East Coast
subpopulation after comparing length–mass relationships,
mean length at age, vertebral counts and otolith morphology,
finding no difference between sardine from KZN and those
from the Western Cape. He concluded that sardine partici-
pating in the KZN sardine run were migrants from the Cape
subpopulation, and were ‘...probably spawned in areas
with a similar environment and during the same season’
as sardine from the Cape. That conclusion was supported
by subsequent workers (Heydorn et al. 1978, Armstrong
et al. 1987, 1991). Grant (1985) also rejected this hypoth-
esis following results from electrophoresis analysis (see T4,
including all genetics approaches).
To further test the hypothesis of a subpopulation gametic
migration, it will be necessary (i) to ensure that the sardine
run is a migration, according to the definition and attributes
provided in the Introduction (see T1), and (ii) test if sardine
engaged in the run are genetically distinct from the rest of
the population, using modern molecular approaches (T4).
Assessing whether or not the sardine run is a spawning or a
feeding migration can be tested by looking for the presence
of sexually mature individuals in the area and, even better,
by sampling ichthyoplankton for eggs and larvae in the KZN
area (T5, and see reviews in Connell 2001, 2010). To test
the hypothesis of a substock it will be necessary to obtain
evidence of a nursery area in, and successful recruitment
arising from spawning in, KZN (T6) and to assess whether
cross-reproduction between subpopulations does not occur
or is limited enough to secure genetic differentiation. This
implies that all year classes of the putative subpopulation
spawn in KZN, and only there (allopatric spawning), or at
least never reproduce when mixed with the rest of the popula-
tion (sympatric spawning with seasonal segregation; T7).
Hu2: Juvenile sardine move equatorwards of their nursery
worldwide
Early research (Davies 1956a, 1956b) suggested an equator-
ward movement of juvenile pilchards (= sardine) from the
southern Benguela nursery grounds and a poleward move-
ment of adult fish when sexually mature, based on the slightly
smaller modal size classes in Namibian catches compared
to Cape catches in the 1950s. Davies postulated that the
sardine run on the East Coast was also an expression of this
equatorward movement. However, Newman (1970) rejected
this theory based on extensive tagging exercises in Namibia
(141 000 fish) and South Africa (74 344 fish), which suggested
largely separate populations in the northern and southern
Benguela. Fish from the Lüderitz area mingled with fish from
the Walvis Bay area, and only a very small proportion of
these fish actually moved to the southern Benguela. Sardine
in St Helena Bay remained in the southern Benguela and did
not move northward to Namibia at all, according to Barange
et al. (1999).
To further test Hu2, one can determine whether fish to the
north of their nursery areas are younger than those found on,
or to the south of these areas, both on the west and east
coasts of South Africa (T2 in Table 2).
Hu3: The sardine run represents an East Coast functionally
discrete adult assemblage
The concept of functionally discrete adult assemblages
(FDAAs) was initially proposed by Fletcher et al. (1994) who
hypothesised that two centres of spawning by Australian
sardine Sardinops sagax neopilchardus observed off Albany
and Bremer Bay on the south coast of Western Australia
(WA) could represent separately functioning adult populations
that were not reproductively isolated. A subsequent analysis
by Gaughan et al. (2001) provided further evidence for the
existence of three distinct centres of sardine spawning off the
southern Western Australian (WA) coast (off Albany, Bremer
Bay and Esperance) that were joined by areas of lower
spawning activity along a continuous distribution. Because
the spawning centres were distinct rather than strictly
separate, Gaughan et al. (2001) termed these groups FDAAs.
Regional differences in age compositions and gonadosomatic
indices of sardine from WA indicated that there was little or
no wide-scale mixing of mature age classes between the
three spawning centres (Gaughan et al. 2002). Those authors
further hypothesised that the persistence of these FDAAs
arose from phenotypic (as opposed to genetic) fitness-
related ties to localised areas of higher habitat suitability. In
contrast to the subpopulation hypothesis, the different FDAAs
share the same genetic pool because their individuals come
from a common pool of earlier stages, as for instance the
‘juvenile pool’ (Gaughan et al. 2001). Based on egg distribu-
tion patterns, the Transkei coast and KZN south coast could
represent a centre of sardine spawning that is distinct from
spawning centres off the West Coast and South Coast (van
der Lingen and Huggett 2003). Following Gaughan et al.
(2002), van der Lingen et al. (2010b) suggest that this could
indicate that fish participating in the KZN sardine run may be
an FDAA that has shown a phenotypic adaptation to specific
conditions off the East Coast. The run could represent the
spawning migration of an East Coast FDAA, with sardine
displaying behavioural patterns (in this case an eastward
migration) and possibly phenotypic adaptation that are
different to those displayed by FDAA(s) that remain on the
Agulhas Bank and spawn there in winter, although all these
FDAAs would come from a common pool of earlier stages.
Most of the tests that can be used for the FDAA hypothesis
are the same as those usable for the subpopulation (Hu1)
Downloaded At: 21:50 16 November 2010

African Journal of Marine Science 2010, 32(2): 449–479 455
hypothesis, with expected results being opposite in some
cases but in others similar. As for Hu1, sardine from the run
are expected to be phenotypically distinct from the rest of
the population (T3), but genetically indistinct from it (T4).
Both hypotheses imply evidence of active spawning in KZN
waters (T5), but in contrast to Hu1, there is no need for a
nursery area there (T6), nor the need for successful recruit-
ment related to spawning off the KZN coast (T7) to support
the FDAA hypothesis. The need for all year classes of the
putative KZN FDAA to spawn in KZN waters (T8) is shared
between Hu1 and Hu3. There is a specific test for the FDAA
hypothesis, directly related to its definition, which is that the
KZN FDAA should not mix with other FDAAs (T9).
Hu4: The sardine run arises from a seasonal extension of
habitat and is a feeding migration
Southern African sardine are known to prefer temperatures
between 14 and 20 °C (Barange et al. 1999), with those
along the East Coast previously observed in temperatures
below 20 °C (Baird 1971, Heydorn et al. 1978). The seasonal
occurrence of sardine along the East Coast during the winter
led Armstrong et al. (1991) to conclude that the sardine
run was a seasonal extension of sardine habitat due to the
favourable cooling of coastal and continental shelf waters
(i.e. a climatic migration). This hypothesis was tested (T10) by
comparing evidence of sardine run events with a time-series
of remotely sensed SSTs.
Sardine dietary carbon intake along the South African east
coast appears to be derived predominantly from zooplankton
(Mketsu 2008), chiefly calanoid copepods, with phytoplankton
rarely important (van der Lingen et al. 2010c). A paucity of
data pertaining to zooplankton abundance during the sardine
run precludes an investigation into whether sardine move up
the East Coast to occupy more favourable feeding conditions.
Any attempt to use remotely sensed Chl a concentration as
a proxy for sardine diet would need to be based upon an
established relationship between Chl a concentration and
zooplankton abundance. To our knowledge, this relation-
ship has not yet been established along the South African
east coast. A further caveat is the as yet untested accuracy
of remotely sensed Chl a concentrations within East Coast
waters. Nonetheless, despite these restrictions, sardine distri-
bution was compared with a remotely sensed Chl a concen-
tration time-series along the South African east coast (see
T11). Testing whether there is more food available along
the KZN coastline than farther south during the sardine run
would allow a decision regarding the possibility of the sardine
run being part of a seasonal feeding (alimental) migration.
Obviously, to be supported, both the climatic and alimental
migration hypotheses developed here must display all the
attributes of a migration (T1).
Hu5: The sardine run can be explained by the basin model
theory
According to basin model theory, the density and distributional
area occupied by marine fish are related to their biomass,
and fish movement can be implicitly considered as ranging
rather than migrating. This is an extension of the concept of
‘density-dependent-habitat-selection’ (DDHS) and the ‘ideal
free distribution’ (IFD) of Fretwell and Lucas (1970), in which
population size and density influence the choice of habitat,
and therefore also the distribution of the population among all
potential habitats, so as to optimise individual fitness (MacCall
1990). Specifically, MacCall (1990) portrayed this model of
fish density and distribution as an irregular basin, the shape
of which could potentially vary over time, filled with a liquid
under the influence of gravity. In this basin, habitat suitability
increases downward in each of the habitats and the depth
of the liquid at any location is proportional to the density-
dependent reduction in realised suitability at that location and
is also proportional to local density. Because densities tend to
be highest toward the centre of a species’ range, the deepest
area of the basin (i.e. the highest realised basic suitability)
will be central, and the topography will become progres-
sively shallower towards the periphery. The total volume of
liquid in the basin is related to total population size. Dynamics
leading to realisation of spatial distribution equilibrium arise
from movement orientated to gradients in habitat suitability
(viscosity) and rate of movement orientated to gradients in
population density (diffusivity). Per capita growth rate (fitness)
influences both the rate of population growth and migratory
flow, i.e. individuals are rewarded by increased reproductive
value for responding to a habitat-suitability gradient. It is worth
noting that habitat suitability/fitness is not well defined (see
review in MacCall 1990) and could possibly be used in the
context of maximisation of reproductive output, which could
translate into MacCall’s per capita growth rate.
A feature of the sardine run is that it seems to involve
only a small fraction of the sardine population at its fringe.
The following aspects of the basin theory are tested in
terms of the sardine run (T12): changes in population size
result in contraction or expansion of the population range,
so that at low biomass individuals occupy habitats with the
highest suitability, whereas at high biomass other, previously
less suitable, habitats become equally attractive and are
colonised, providing for population range expansion.
Hu6: The sardine run represents the behaviour of a meta-
population
The concept of metapopulations was first proposed by
Levins (1968) as ‘a population of local populations which
were established by colonists, survive for a while, send out
migrants, and eventually disappear. The persistence of
a species in a region depends on the rate of colonisation
successfully balancing the local extinction rate’. Levins (1968)
defined the metapopulation as:
there are a large number of sites, each supporting a 1.
single local population;
each local population has a probability of going extinct 2.
that may depend on its genetic composition;
the allele frequencies are governed by the classical 3.
genetic equations; and
vacant sites are recolonised by migrants from within the 4.
metapopulation.
The concept was then enriched with new ideas and
summarised by McQuinn (1997) when he applied it to
populations of Atlantic herring Clupea harengus. He synthe-
sised Levins’s ideas as ‘The population structure of many
species can be considered as an array of local populations
linked by variable degrees of gene flow’.
Another point is that continuous populations cannot be
considered as metapopulations. Kritzer and Sale (2004)
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings456
define a metapopulation as ‘a system of discrete local
populations, each of which determines its own internal
dynamics to a large extent, but with a degree of identifi-
able and nontrivial demographic influence from other local
populations through dispersal of individuals’ (Figure 1).
Hanski (1999) provides a list of 12 conditions necessary to
fulfil the status of metapopulation, and F Gerlotto (IRD, pers.
comm.) completed this list with four additional conditions
expressed by other authors (Table 3). We assessed
these 16 conditions with regard to present knowledge of a
putative sardine metapopulation off southern Africa (T13)
with two local populations potentially located off the coast
of Namibia, at least in historical periods of high abundance
(Bakun 2005), and three local populations off the coast of
South Africa (van der Lingen et al. 2010c). The three South
African local populations considered were: West Coast
and Western Agulhas Bank; Eastern Agulhas Bank; and
KwaZulu-Natal.
Hu7: The sardine run is relic behaviour
Carscadden et al. (1989) found that the only stock of
capelin Mallotus villosus that was not an intertidal spawner
was selecting areas of gravel deposition corresponding
to ancestral beaches of the Wisconsin glaciation period.
Wyatt et al. (1986) noted that the unusually deep location
of spawning grounds for sardine Sardina pilchardus off
North-West Europe is in an area that was coastal during the
post-Pleistocene transgression. These two examples show
that habitat selection during spawning can be the result of
remote ultimate factors that have resulted in what could be
termed relic behaviour, i.e. behaviour that was the norm in
some previous climate regime (e.g. a glacial period) but,
while still occurring, is no longer the norm in the present
regime (e.g. Romero 1985). A subpopulation of sardine on
the east coast of South Africa could potentially have been
brought about through a combination of bet-hedging and/
or relic behaviour (Coetzee et al. 2010). The relic behaviour
could have arisen with a northwards shift of polar conditions
and population separation during the last ice age and glacial
maximum some 16 000 years ago, when sea level was
approximately 130 m lower than at present (Miller 1990). At
that time, the southern extremity of the continental landmass
was 200 km farther south than the present day, subtropical
SSTs were 4 °C or more lower, the Subtropical Convergence
(STC) was farther north, the Agulhas Current retroflection
was farther eastwards, and there was little or no exchange
between the Indian and Atlantic oceans (Hutchings 1992,
Peeters et al. 2004, Parkington 2006). Each of these or a
combination thereof, together with the northward shift in cool
polar conditions, may have enforced separation of sardine
off the West and South coasts, with each having their own
independent spawning patterns and/or migrations. The end
of the ice age would have seen the west and east subpopu-
lations being reconnected with South/West Coast spawning
becoming dominant, possibly as a result of higher levels
of productivity in that region. A similar sequence of events,
borne out by molecular genetic studies, may have led to
the differentiation of the Atlantic and Indian populations
of albacore Thunnus alalunga (Chow and Ushiama 1995,
Viñas et al. 2004, AJ Penney, National Institute of Water &
Atmospheric Research, New Zealand, pers. comm.).
Relic behaviour has been demonstrated elsewhere. The
diadromous life histories of most anguillid eels and salmonids
are thought to have arisen from ‘migration loops’ expanding
away from low-productivity, food-poor origins in the tropical
marine and cold temperate riverine environments respec-
tively (Tsukamoto et al. 2002, Inoue et al. 2010). In both
families, the ‘ancestral loops’, including spawning migrations,
have been retained. The East Coast sardine ‘spawning
run’ differs from anguillid and salmonid relic behaviour by
not being dominant but, similarly, may have been partially
driven by higher productivity in this region during the glacial
periods. Relic behaviour may not necessarily arise over a
single glacial period but over successive ones, becoming
reinforced and more stable within each cycle (Kawanabe
1977). It follows that relic behaviour such as the East Coast
migration of sardine could be retained as an alternative stable
state to counter recurring glacial periods and by default be
a bet-hedge against biological and environmental pertur-
bations that befall the population on a shorter time-scale.
Nonetheless, to be retained in the genome of sardine, this
behaviour must be viable during the protracted interglacial
periods. This means that reproduction off the KZN coast must
be successful enough to produce recruitment to this subpop-
ulation, despite heavy predation during the sardine run.
Beyond the fact that the relic behaviour hypothesis is a
gametic migration of a subpopulation that must fulfil the five
attributes of a migration (T1) and also the other tests related
to the hypothesis of a subpopulation spawning migration
(Hu1; T3 to T8), this hypothesis is difficult to falsify. The
only specific tests that could allow differentiating it from Hu1
would be related to palaeogeography, palaeoclimatology
and palaeosedimentology (T14).
A. Network of closed populations
B. Metapopulation
C. Patchy population
Pro
babili
ty
Dispersal distance
Pro
babili
ty
Dispersal distance
Pro
babili
ty
Dispersal distance
Figure 1: Three types of spatially structured populations with general-
ised dispersion curves for each local population. Case A: closed local
population with no exchange between them; case B: metapopulation;
and case C: patchy population inside a single ‘super-population’.
The thicker lines represent the overall geographical limit of the
population(s). From Kritzer and Sale (2004)
Downloaded At: 21:50 16 November 2010

African Journal of Marine Science 2010, 32(2): 449–479 457
Hypotheses about proximate factors (Hp) and potential
tests
During the sardine run, sardine shoals need to swim along
the eastward-narrowing shelf of the Pondoland and Transkei
coasts against the fast, warm Agulhas Current to reach
KZN waters. In some years, the sardine have not reached
southern KZN or have arrived there late in the year. Here,
we list the hypotheses related to proximate factors (enabling
conditions) of the sardine run in an attempt to answer the
question relating to how the sardine run occurs (Table 4).
Hp1: Natal homing and imprinting
Natal homing is defined as the return of spawning adults
to their region of origin (Carr 1967), which in most cases is
allowed by imprinting of very early stages (larvae or even
eggs) by environmental cues (Stabell 1984). It differs from
spawning site fidelity in which fish may spawn for the first
time at a site different from where it was born. Natal homing
is a strategy that ensures the best chance of successful
reproduction. Adults return to natal sites to spawn irrespec-
tive of environmental change (the effects of which are likely
to be unpredictable to an animal), because that strategy is
likely to be most beneficial (in fitness terms) in the face of
environmental uncertainty. Natal homing is a strong driver
of genetic differentiation and it secures genetic isolation, by
preventing or limiting interbreeding. Although natal homing
could also be considered as an ultimate factor and environ-
mental cues associated to imprinting as proximate factors,
we classified it as a proximate factor because it is a process
that can be associated with two ultimate factor hypoth-
eses: the spawning migration of a subpopulation and relic
behaviour.
There are several examples of natal homing. Thorrold et
al. (2001) showed that between 61% and 80% of weakfish
Cynoscion regalis, an estuarine-spawning marine fish,
returned to their natal estuaries on the east coast of North
America after reaching sexual maturity offshore. Mature
Icelandic plaice Pleuronectes platessa were tagged on
spawning and feeding grounds off Iceland (Solmundsson
et al. 2005), and after weighting the number of recaptures
from standardised fishing effort, a minimum level of fidelity
to the spawning ground was estimated at 94% for the
spawning season one year after tagging. Fidelity of 72%
for the second and third spawning seasons was observed
after migrating from distant feeding grounds. The North
Atlantic bluefin tuna Thunnus thynnus has been shown to
consist of two fairly discrete populations, one based in the
Mediterranean and the other on the east coast of Canada
and the United States. Although they mix in feeding
aggregations as sexually immature fish in the North Atlantic
Bight (Rooker et al. 2008), the return of spawning adults
to their region of origin is high and remarkably similar for
both eastern and western spawning regions: 95.8% for the
Mediterranean, 99.3% for the Gulf of Mexico, 94.8% in the
Gulf of Maine and 100% in the Gulf of St Lawrence. Among
open-water coastal pelagic shoaling fish species, significant
isolation of stocks in the North Atlantic herring population
implies natal return to maintain such integrity (Stephenson
et al. 2009). Mullon et al. (2002) showed that observed
Metapopulation characteristics Relevance to the case of southern African sardine
1 Population size or density is significantly affected by migrations
Not obvious. At least this is not the case for migration between Namibia and South Africa due to the cold thermal barrier created by the Lüderitz upwelling cell (Lett et al. 2007)
2 Population density is affected by patch areas and isolation Yes, but mainly at a relatively small scale (clusters of schools)
3 Existence of asynchronous local dynamics Yes between Namibia and South Africa, but not between the three putative South African local populations
4 Population turnover, local extinctions and establishment of new populations
No, to the best of our knowledge
5 Presence of empty habitats Not permanent, except, to a certain extent, in the vicinity of the Lüderitz upwelling cell
6 Metapopulations persist despite population turnover Yes
7 Extinction risk depends on patch area No, except for Namibia.
8 Colonisation rate depends on patch isolation ? This condition seems irrelevant for large pelagic populations
9 Patch occupancy depends on patch area and isolation ? This condition seems irrelevant for large pelagic populations
10 Spatially realistic metapopulation models can be used to make prediction about metapopulation dynamics in particular fragmented landscape
? No specific modelling exercise done so far
11 Metapopulation coexistence of competitors ? Except possibly for the Namibian versus South African anchovy putative metapopulation
12 Metapopulation coexistence of prey and its predator ? This condition seems irrelevant for large pelagic populations
13 Evidence of genetic linkage ? Only electrophoresis analyses performed so far
14 Genetic or morphometric, meristic or biological/behavioural differences
Yes for morphometrics and meristics (similar to T3)
15 Existence of source/sink populations ?
16 Discrete local populations No
Table 3: List of characteristics describing a metapopulation (from Hanski 1999 for the first 12 conditions and F Gerlotto, IRD, pers. comm.,
for the last four) and assessment of their application to southern African sardine
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings458
anchovy Engraulis encrasicolus spawning patterns in the
southern Benguela Current system off South Africa could
be accurately reproduced by simulating a natal homing
reproductive strategy, i.e. individuals spawning at their natal
date and place.
Natal return would explain both the very small percentage
of the southern African sardine population that migrates
annually up the East Coast and into KZN waters, generally
considered to be about 10–15% (e.g. Armstrong et al.
1991), but as low as 1% when abundance is high (Coetzee
et al. 2010), and the fact that the intensity and occurrence
of the annual sardine run appears to bear no relationship
to the state of the overall population (see above). Whereas
it would seem reasonable to assume that fish arising from
successful spawning in KZN waters would tend to remain
at the eastern extreme of the distributional range of the
South African sardine population (Port Elizabeth to Cape
St Francis), recent focus on the adventure diving and tele-
vision documentary potential of the sardine run has seen
the development of early-season tracking of the eastward
movement of sardine shoals, with reports coming from as
far west as Mossel Bay (M Addison, Blue Wilderness Dive
Expeditions, pers. comm.).
The recent advance in knowledge of natal returns in
marine fish is largely as a result of the development of new
genetic (T4) and chemical techniques (described in T7). The
latter, in particular, has enabled detailed chemical tracking
of larval history, through the microchemistry of the growth
stages within the sagittal otoliths (ear bones). During their
migration along the Eastern Cape coast and into KZN
waters sardine spawn prolifically, but nothing is known of
the success or otherwise of this spawning in contributing to
recruitment to the southern African sardine population. The
natal return hypothesis relies on survival of recruits from the
spawning that occurs during the migration.
The most promising technique currently available to test
this hypothesis is to analyse the chemical constituents
of the core of the otoliths (T15) of juveniles from the four
major regions, namely the West Coast, the Agulhas Bank,
Algoa Bay and the KZN coast, in order to establish whether
juveniles can be separated by unique elemental constitu-
ents. If this proves to be the case, then analysis of the same
core area of adult fish participating in the annual sardine run
will establish what proportion of these adult fish are natal
returns, and provide other valuable insights into the structure
of the migrating shoals. If the run consists mainly of natal
returns, it will add a new dimension to management of the
resource, because preservation of within-species diversity is
an important element of an ecosystem approach to manage-
ment. Another test, although less powerful, could be based
on vertebral count data because it has been demonstrated
that this meristic characteristic depends on the temperature
experienced by larvae during the first few days following
hatching, with higher temperatures resulting in fewer
vertebrae (Ben Tuvia 1964, Andreu 1969). For instance,
young Sardina pilchardus found at the extreme equatorward
range of their distribution (14°43! N) had lower vertebral
counts than those from any other area in the Atlantic Ocean
(Fréon and Stéquert 1982).
Hp2: Are sardine pushed northward or shoreward by predators
Armstrong et al. (1991) proposed that the KZN component of
the sardine run is motivated by repeated attacks of predators
that would result in sardine being pushed northward and/or
Hypothesis number
and name Description Source Tests
Hp1: Natal homing and
imprinting
Fish return to the site of their
birth to reproduce because they
are imprinted at early stages by
environmental cues
Carr (1967), Stabell
(1984)
T15: Sardine engaged in the run are mostly born
in KZN waters
Hp2: Predator-driven
movement
Predators push sardine to the
north and/or to the shore once
arrived in Natal waters
Armstrong et al. (1991) T16: Fine-scale observations of predation and
response of sardine
Hp3: Environmental drive:
(a) favourable inshore
seasonal countercurrent
A seasonal onshore coastal
countercurrent (of the Agulhas
Current) would favour the
sardine run during all winter
Baird (1971) T17: Continuous coastal current measurement
(e.g. ADCP mooring line)
T18: Embedded, very high resolution numeric
model
Hp4: Environmental drive:
(b) favourable cooling of the
coastal area
Sporadic northward movement
of quanta of cooler waters
inshore along the East Coast
Heydorn et al. (1978) T18: As above
T19: Compare remotely sensed SST and
sardine run data (visual observations,
catches, eggs, acoustics) at high resolution
Hp5: Environmental drive:
(c) Waterfall Bluff gateway
During their north-east
displacement, sardine are
blocked by warm water flowing to
the south-west, north of Waterfall
Bluff, due to the topographic
configuration of this area. During
Natal Pulses the ‘gateway’
opens and the sardine run can
extend to the north-east
Armstrong et al. (1991),
Roberts et al. (2010)
T18: As above
T19: Look for the coincidence of Natal Pulses or
break-away eddies in remote-sensing data
with sardine run events
Table 4: Hypotheses on proximate factors pertaining to the sardine run: description, key references and potential tests (T)
Downloaded At: 21:50 16 November 2010

African Journal of Marine Science 2010, 32(2): 449–479 459
shoreward. Their evidence for this generalised statement was
the presence of sardine in warm water along this coastline. It
is difficult to test this hypothesis, because although there is a
very close association between sardine and their predators
(O’Donoghue et al. 2010a, 2010b, 2010c), particularly Cape
gannets Morus capensis and common dolphins Delphinus
capensis, it is uncertain whether the predators are following
or chasing their prey. Nonetheless, the application of
fine-scale observation techniques (T16), general knowledge
of fish behaviour and common sense could assist in reaching
a conclusion.
Hp3: Environmental drive: (a) favourable inshore seasonal
countercurrent
With little oceanographic information available, Baird (1971)
speculated that a well-established countercurrent inshore of
the Agulhas Current along the entire coast during winter could
be the prime cause of the sardine run. However, as pointed
out by Schumann (1987), the National Research Institute
for Oceanography (NRIO) measurements undertaken off
East London between April and December 1984, during
a year of substantial sardine catch on the Transkei/KZN
border, showed no indication of a defined seasonal counter-
current. From current meter data, Schumann (1987) sug-
gested that the mechanism by which sardine could be
assisted in their migration northwards occurred sporadi-
cally. This supported the views of Heydorn et al. (1978) who
were sceptical of the association of the sardine run with a
cool water countercurrent that originated off the southern
Cape and flowed up the East Coast in winter. Hp3 can be
tested by analysing data from ADCP moorings positioned
in shallow waters off the KZN coast (T17). Hydrodynamic
models in 3D with fine-scale resolution and one or two
levels of embedding (T18) could also aid in evaluation of this
hypothesis.
Hp4: Environmental drive: (b) favourable cooling of the
coastal area
Rather, Heydorn et al. (1978) thought it more likely that water
movement in the form of northward advection of pockets
(quanta) of cooler water inshore of the Agulhas Current
provided the physical mechanism for the run to occur. This
would be coupled with seasonal cooling of the inshore water
that is also conducive to the appearance of this temperate
species in KZN waters. Heydorn et al. (1978) also noted
that it is possible that a distinct nearshore regime may exist
within a kilometre from the shore. This region, which would
include the breaker zone, would probably be governed
by swell-driven mass transport and would reflect changes
in the wind field with a south-westerly swell generating
a northward nearshore current. To test this hypothesis,
fine-scale measurement of temperature, using, for instance,
high-resolution MODIS or aerial survey data, and relating
them to sardine run evidence, would be useful (T19). As
for Hp3, numerical models could help in demonstrating the
existence of quanta of cooler water (T18).
Hp5: Environmental drive: (c) Waterfall Bluff gateway hypothesis
The first dedicated attempt to assess and understand the
sardine run was undertaken by Armstrong et al. (1991)
using data from three ship-borne surveys along the East
Coast in August 1986, June 1987 and June 1990. They
observed sardine along the narrow Pondoland shelf as far
east as Port St Johns, but to the north of this the sardine
apparently descended into deeper, cooler water. Armstrong
et al. (1991) proposed that the sardine run is a phenomenon
arising from the expansion of the suitable environment for
this temperate species, during the cooler water conditions
that prevail in winter (Hp4) and, furthermore, that sporadic
current reversals and upwelling of cooler water onto the
narrow shelf results in the leakage of sardine schools along
the shelf between Port St Johns and Durban. Armstrong et
al. (1991) initially speculated that the Natal Pulse was the
key mechanism, but its intermittent occurrence (only found
20% of the year according to Lutjeharms and Roberts 1988,
or at intervals varying between 50 and 150 days according
to de Ruijter et al. 1999) meant that it could not be primarily
depended on for the sardine run.
Whereas the timing of current reversals between Port
Alfred and East London will slightly quicken or retard
progress of the sardine shoals moving up the East Coast, the
encroachment of the Agulhas Current onto the Pondoland
shelf is considered to be the first serious barrier for the
sardine shoals to overcome during their northward migration
to the KZN Bight. However, according to satellite imagery
(Roberts et al. 2010), this high-velocity warm temperature
barrier is quite variable and probably only impedes passage
for periods lasting several days. Anecdotal sightings
(O’Donoghue 2009) of sardine shoals near Waterfall Bluff
for extended periods, however, supports the earlier proposal
by both Schumann (1987) and Armstrong et al. (1991) of this
foremost barrier. According to Roberts et al. (2010), Natal
Pulses and break-away eddies provide the means for sardine
shoals to overcome this barrier and to move northward
along the narrow shelf, although their arrival on the KZN
south coast will depend on the timing of these features. The
process is schematically illustrated in their Figure 14, which
shows the ‘open gateway’ and ‘closed gateway’ situations,
with the latter allowing the shoals to reach the KZN south
coast and even Durban. They refer to these mechanisms
and their potential influence on the timing and appearance
of sardine in KZN waters as the ‘Waterfall Bluff gateway’
hypothesis.
Testing the existence and quantification of Natal Pulses
and break-away eddies can be partly addressed using
numeric models (T18), but a real test of Hp5 itself requires
the investigation of coincidental occurrences of Natal Pulses
or break-away eddies with sardine run events by combining
T17 and T19 approaches.
Results
Testing hypotheses
In this section, the tests listed in Tables 2 and 4 are reviewed.
They are dealt with separately from the hypotheses because
a given test can be applied to several hypotheses. Tests
already performed and documented in the literature are
recapitulated and updated when appropriate. Wherever
possible, new tests are performed and suggestions given
for further tests that could shed light on yet unanswered
questions, as methods are developed or data become
available.
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings460
T1: Is the sardine run a migration?
The five conditions listed in the Introduction that have to
be met before movement can be attributed to migration are
evaluated below.
T1a: Does the animal display persistent motion that takes
it beyond the home-range? The answer is definitely
yes. The sardine run is a seasonal displacement over
hundreds of kilometres.
T1b: Is the movement direct, and not erratic in direction?
Although the sardine run can be stopped (potentially
due to unfavourable environmental conditions farther
to the north-east; see below), the movement is always
direct and not erratic.
T1c: Does the animal pass suitable resources that would
otherwise not be overlooked? The answer to this
question is not obvious, except maybe at the onset of
the migration where sardine leave the relatively food-
rich eastern Agulhas Bank and enter into the poorer
food environment of the KZN coastal area (Hutchings
et al. 2010). Farther north, as sardine move past the
KZN Bight (limited observations by Pearce 1977),
they would encounter and pass suitable resources
originating from persistent upwelling in the region
between Richards Bay and St Lucia which spreads
southwards over the KZN Bight (Lutjeharms et al. 1989,
Hutchings et al. 2010).
T1d: Does the animal engage in specific departure and arrival
behaviour? Both the departure point and departure date
are variable but thought to be on the eastern Agulhas
Bank and generally sometime in May. Arrival in the area
north of Port Edward, as deduced from aerial surveys,
beach-seine catches, increased predator fish avail-
ability and rapid increase in the occurrence of sardine
eggs (O’Donoghue et al. 2010c, van der Lingen et al.
2010a, Fennessy et al. 2010 and Connell 2010 respec-
tively), occurs predominantly in June or July. There
is, however, little evidence of where sardine eventu-
ally go once they pass Durban (to the KZN Bight or
farther north?) and how long they remain off the KZN
coast, although sardine eggs are regularly collected
in samples taken off Park Rynie up until December
(Connell 2010). Whether sardine engage in any form
of display to signal departure or arrival, as is common
among migratory birds, is unknown. One may expect
changes in school structure, orientation, or feeding. The
test will require structured submarine observations that
have not yet been attempted, and indeed may not be
practical.
T1e: Are there physiological adjustments made to reserve
energy for a long journey? According to van der Lingen
et al. (2006), fish from the South Coast (20–27° E)
have a higher mean condition factor (CF) than those
from both the West (north of 33.5° S) and South-West
coasts. In contrast, sardine caught during the run
display a lower CF than in any other area (Coetzee
et al. 2010). Although CF is a complex indicator
reflecting not only condition but also morphology and
sexual maturation stage, these observations could be
interpreted as fish building up reserves on the South
Coast prior to departure and then quickly losing these
reserves as they migrate northwards along the KZN
coast.
From the above, it can be concluded that despite a few
uncertainties regarding mainly departure time and location,
as well as the ultimate arrival location, tests for confirming
the migration theory (T1) are largely positive.
T2: Are fish younger when found to the north (rather than
the south) of their nursery area?
Van der Lingen et al. (2010c; their Figures 6–8) show
that the age structure of sardine caught off the KZN coast
since 1955, although discontinuous, is dominated by
age classes 2–4. Age classes 0 and 1, although possibly
underestimated in beach-seine net catches, are seldom
abundant. Sardine caught in midwater trawls between
Port Alfred and Port Edward during the 2005 sardine run
survey were smaller in the south than farther northward
with no clear pattern in age structure (Coetzee et al. 2010).
Furthermore, van der Lingen et al. (2010c, Figure 2) show
that sardine caught off KZN typically ranged from 13 to
21 cm caudal length (CL) with an overall mode at 18 cm,
although bimodal structures are observed during some
years. The annual mean CL of KZN sardine caught by
beach-seines in recent years (1997–2005) appears to be
different from that of sardine caught by the purse-seine
fishery off Port Elizabeth during June and July of the same
years, with the KZN data showing less individuals of size
classes 16.0–18.0 cm and more of classes 18.5–19.5 cm
than the South Coast data (!2 test, p < 0.02; Figure 2).
Disaggregated commercial catches (beach seines) in
KZN in 2004 and 2005 compared to experimental catches
(midwater trawl) performed during pelagic spawner
biomass surveys during the same years show different
caudal length distributions (Figure 3), especially for young
fish, which is due to the different areas sampled (pelagic
spawner biomass surveys extend from Hondeklip Bay off
the West Coast to Port Alfred off the South Coast) and to
FR
EQ
UE
NC
Y (
%)
20
20 22 23 2421
15
10
5
12 13 14 15 16 17 18 19CAUDAL LENGTH (cm)
Port Elizabeth (n = 1 274)
Sardine run (n = 4 127)
Figure 2: Comparison between frequency distributions of caudal
length of sardine sampled from commercial purse-seine catches
taken off Port Elizabeth during June and July, and those sampled
from beach-seine catches taken off KZN during the sardine run, for
the period 1997–2005
Downloaded At: 21:50 16 November 2010
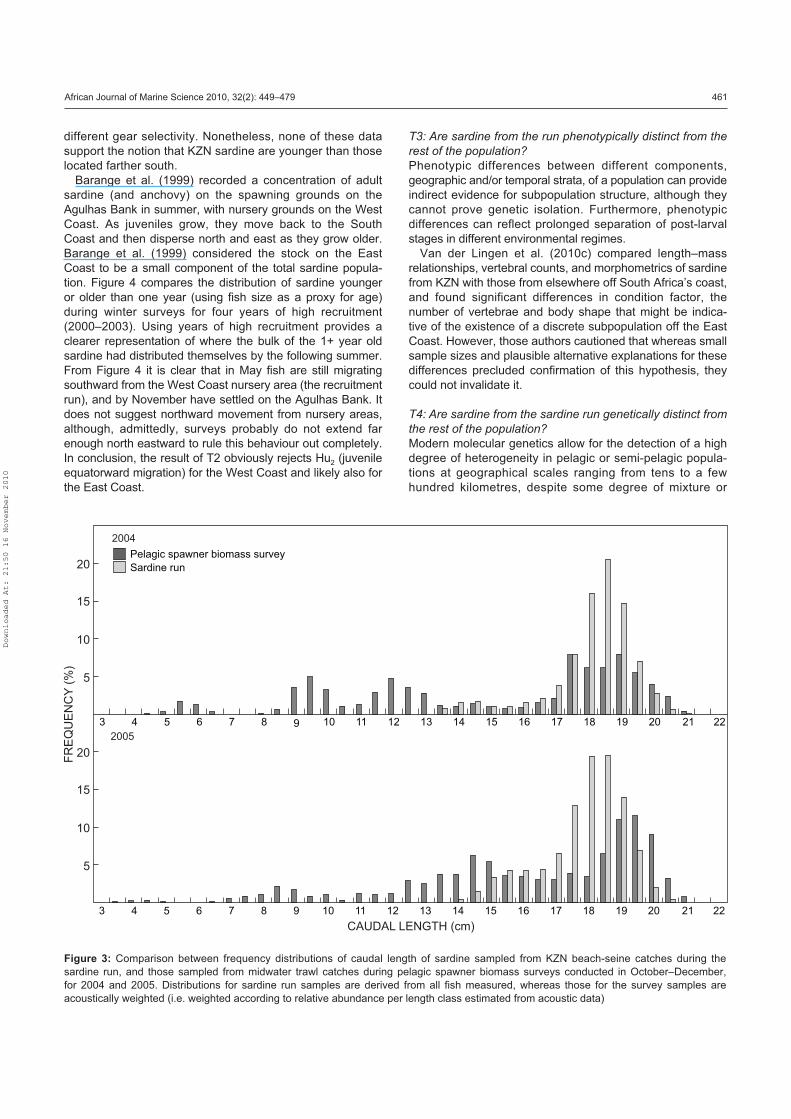
African Journal of Marine Science 2010, 32(2): 449–479 461
different gear selectivity. Nonetheless, none of these data
support the notion that KZN sardine are younger than those
located farther south.
Barange et al. (1999) recorded a concentration of adult
sardine (and anchovy) on the spawning grounds on the
Agulhas Bank in summer, with nursery grounds on the West
Coast. As juveniles grow, they move back to the South
Coast and then disperse north and east as they grow older.
Barange et al. (1999) considered the stock on the East
Coast to be a small component of the total sardine popula-
tion. Figure 4 compares the distribution of sardine younger
or older than one year (using fish size as a proxy for age)
during winter surveys for four years of high recruitment
(2000–2003). Using years of high recruitment provides a
clearer representation of where the bulk of the 1+ year old
sardine had distributed themselves by the following summer.
From Figure 4 it is clear that in May fish are still migrating
southward from the West Coast nursery area (the recruitment
run), and by November have settled on the Agulhas Bank. It
does not suggest northward movement from nursery areas,
although, admittedly, surveys probably do not extend far
enough north eastward to rule this behaviour out completely.
In conclusion, the result of T2 obviously rejects Hu2 (juvenile
equatorward migration) for the West Coast and likely also for
the East Coast.
T3: Are sardine from the run phenotypically distinct from the
rest of the population?
Phenotypic differences between different components,
geographic and/or temporal strata, of a population can provide
indirect evidence for subpopulation structure, although they
cannot prove genetic isolation. Furthermore, phenotypic
differences can reflect prolonged separation of post-larval
stages in different environmental regimes.
Van der Lingen et al. (2010c) compared length–mass
relationships, vertebral counts, and morphometrics of sardine
from KZN with those from elsewhere off South Africa’s coast,
and found significant differences in condition factor, the
number of vertebrae and body shape that might be indica-
tive of the existence of a discrete subpopulation off the East
Coast. However, those authors cautioned that whereas small
sample sizes and plausible alternative explanations for these
differences precluded confirmation of this hypothesis, they
could not invalidate it.
T4: Are sardine from the sardine run genetically distinct from
the rest of the population?
Modern molecular genetics allow for the detection of a high
degree of heterogeneity in pelagic or semi-pelagic popula-
tions at geographical scales ranging from tens to a few
hundred kilometres, despite some degree of mixture or
2004
2005
Pelagic spawner biomass survey
Sardine run
3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22
FR
EQ
UE
NC
Y (
%)
20
15
10
5
20
15
10
5
CAUDAL LENGTH (cm)
Figure 3: Comparison between frequency distributions of caudal length of sardine sampled from KZN beach-seine catches during the
sardine run, and those sampled from midwater trawl catches during pelagic spawner biomass surveys conducted in October–December,
for 2004 and 2005. Distributions for sardine run samples are derived from all fish measured, whereas those for the survey samples are
acoustically weighted (i.e. weighted according to relative abundance per length class estimated from acoustic data)
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings462
close proximity of the spawning populations to each other
(see review by Hauser and Carvalho 2008). Estimates of
FST, the fraction of total genetic variation attributable to
differences among populations, is of particular interest in
this context. Indeed, FST estimates the relative decrease
in heterozygosity due to population substructure relative
to the expected heterozygosity in an equivalent panmictic
population. This kind of approach, not yet conducted in
South Africa at a spatial scale relevant for the sardine run,
is certainly one that could allow the best possible insight
for understanding the structure of the sardine population,
despite some limitations related to large populations. Ideally,
it would require simultaneous collection of samples at the
time of the sardine run from various locations along the
5
10
15
20
1 2 3 4 5 6 7 8 9AGE (years)
CA
UD
AL L
EN
GT
H (
cm
)
For each year, fish <1 year old in May were assumed to be
<12.5 cm and fish older than 1 but younger than 2 years in
November were assumed to be !12.5 and <16 cm
2000 2001
2002 2003
<1-year-olds in May
1- to 2-year-olds in November
<1-year-olds in May
1- to 2-year-olds in November
<1-year-olds in May
1- to 2-year-olds in November
<1-year-olds in May
1- to 2-year-olds in November
Sardine density (g m–2)
1 5 25 50 100
Figure 4: Density of sardine <12.5 cm CL in May and "12.5 and <16.0 cm CL in the following November for four years (2000–2003) during
which sardine recruitment was high (upper panel); and cut-off lengths for minimum and maximum caudal length of 1-year-old fish from the
sardine age–length curve (lower panel; courtesy D Durholtz, Department of Agriculture, Forestry and Fisheries)
Downloaded At: 21:50 16 November 2010

African Journal of Marine Science 2010, 32(2): 449–479 463
entire southern African coast, including Namibia. The ideal
distance between samples should be in the order of 100 km.
Coupling molecular genetics with otolith microchemistry (see
below) would increase the power of this approach.
T5: Is there evidence of active spawning in KZN?
The 21-year time-series of ichthyoplankton data collected
weekly from surface shelf waters at Park Rynie on the KZN
south coast since 1987 (Connell 2010) clearly demonstrates
that sardine spawn regularly in this area from June to
December. As expected with monospecific plankton data,
the dataset is dominated by zero values (78%) and extremely
skewed to the right (skewness = 370), to the point that
a lognormal transformation still provides an extremely
skewed distribution. Nonetheless, the dataset does not fit a
Pareto law. The most intense spawning occurred from June
to October (2.2–5.8 eggs m#3), despite some degree of
inter annual variability. Sardine egg concentrations off Park
Rynie, and those from near-surface samples collected along
the East Coast using a CUFES during the 2005 sardine
run survey (which ranged from 0 to 43.2 eggs m#3, and had
an average concentration of 0.6 eggs m#3 [SD 2.9] for both
survey phases combined; see Coetzee et al. 2010), are
substantially lower than those observed elsewhere off the
South African coast. Concentrations of sardine eggs collected
using a CUFES on the western Agulhas Bank ranged from
0 to 106.8 eggs m#3 and had an average of 5.0 ± eggs m#3
(SD 1.9) (van der Lingen et al. 1998), and similar average
and maximum concentrations have been observed in CUFES
samples collected off the West Coast (van der Lingen
and van der Westhuizen 2000) and South Coast (CDvdL
unpublished data).
The distance travelled between egg sampling sites and
the site of spawning can be roughly estimated from current
data and an estimation of the age of eggs collected. A
sardine egg takes about 24–36 h to hatch in warm waters
(King 1977), akin to the KZN south coast area during the
period June–December (average monthly AVHRR satellite
SST ranging from 19.5 to 25.8 °C). Egg stages vary from
freshly spawned to well developed, and on average eggs
can be estimated to be 12 h old (ADC pers. obs.). Current
direction in the sampling area off Park Rynie from June to
December is dominated by alongshore flow (to either the
north or south) and this pattern is consistent throughout the
year (results not shown). Semi-quantitative data on current
speed, backed-up by a few current meter measurements
from June to December 2006, indicate that the alongshore
current speed varied between 0.15 and 0.88 m s#1, with
an estimated average of 0.31 m s#1 regardless of their
estimated direction. According to these estimates, which
are in agreement with ship-borne (S–ADCP) measurements
(Roberts et al. 2010), the average distance travelled by an
egg in the alongshore direction can be estimated at 13 km,
varying from 6 to 38 km. Cross-shelf currents are seldom
observed in this area (1.8% inshore and 0.4% offshore) and
are much weaker, with speeds varying from 0 to 0.15 m
s–1. Therefore, the maximum distance travelled by an egg
in the cross-shelf direction can be estimated to be 6 km.
Intermediate current direction is dominated by south-inshore
currents (7.1%) whereas zero or undetectable currents
($0.1 m s–1) represent 6.1% of the dataset. These results
clearly confirm that the KZN area is an active spawning
area, with the possible exception of 2006 when no sardine
eggs were found in samples (Connell 2010).
Historical egg surveys conducted in KZN waters confirm the
spatial extent of spawning in this region (see Connell 2010)
as did the 2005 sardine run survey (see Figure 9 of Coetzee
et al. 2010). In conclusion, T5 is clearly positive, upholding
the hypothesis of active spawning in KZN and thereby
supporting Hu1 of a subpopulation spawning migration.
T6: Is there evidence of a nursery area off the KZN coast?
Connell (2010) gathered several lines of evidence from the
literature that supported the existence of a nursery area
in the KZN region, including the presence of early sardine
larvae (Beckley and Hewitson 1994, Beckley and Naidoo
2003), early juveniles (Heydorn et al. 1978, van der Byl
1980) and late juveniles (van der Byl 1978). Connell (2010)
concluded that locally spawned larvae may tolerate higher
temperatures on the KZN coast than elsewhere in South
Africa. Juvenile sardine were also found south of the KZN
region, from Port Edward to Port Elizabeth, as reported by
Miller et al. (2006) who linked the presence of these recruits
to local spawning. Nonetheless, one cannot exclude the
possibility that these individuals result from spawning in
KZN and farther northward, their transport or migration being
subsequently favoured by the dominant Agulhas Current.
Hence, the existence of a nursery area off the KZN coast is
likely.
T7: Is there successful recruitment attributed to individuals
spawned in KZN?
Whether or not all these early stages and juveniles contribute
significantly to recruitment of the South African sardine
population remains an unanswered question to date. Coetzee
et al. (2008) show that high sardine recruitment as a whole
is mainly associated with successful West Coast recruit-
ment, although the importance of recruitment emanating
from successful KZN spawning must be of significance in the
maintenance of a local spawning migration, particularly if it
results from a genetically distinct subpopulation. It is likely
that the use of otolith microchemistry, coupled with molecular
genetics, would provide a definitive answer to this question.
Indeed, otoliths take up several chemical substances and
their stable isotopes (e.g. strontium, strontium/calcium ratio,
barium, oxygen ["18O] or carbon ["13C] isotopes) in propor-
tion to their concentration in the ambient environment (review
in Fromentin et al. 2009). It is likely that there is a high
contrast in chemical tracers found in the Indian and Atlantic
oceans. Therefore, one can expect to trace the history of fish
movement from early stages to age at catch and to infer the
contribution of fish born in the Indian Ocean to the overall
population. Similarly, chemical tagging with tetra cycline or
isotopes, injected into mature females, has allowed identifi-
cation of returning recruits, by detecting the injected chemical
in the core of the returning juvenile otolith (Jones et al. 2005,
Almany et al. 2007). Nonetheless, the application of such
approaches are limited by the monetary expense of the
method per fish sampled, which would probably constrain
sampling to fewer individuals than required to obtain a precise
estimate of the contribution to recruitment of fish spawned in
KZN.
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings464
T8: Are all year classes of the subpopulation spawning in
KZN and only there (or at least never reproduce when mixed
with the rest of the population)?
Van der Lingen et al. (2010c; their Figures 6–8) show that
sardine caught by beach-seine off KZN since 1955 are
dominated by mature age classes (2- to 4-year-old fish) with
large interannual variability. Year classes 4, 5 and 6 are
often absent from samples, which can be attributed to a low
sampling rate, or to higher mortality than in the rest of the
population owing to repeated migrations through an area
with heavy predation pressure (see also general discussion
on this point).
200 400 600 80020 1 000
Onboard counts; n = 1 461
Eggs m!2
Hondeklip Bay
Cape TownMossel Bay
Port Elizabeth
Port Alfred
East London
Port St Johns
Port Edward
Durban
(a)
16° E 18° E 20° E 22° E 24° E 26° E 28° E 30° E
30° S
32° S
34° S
36° S
32° E
Hondeklip Bay
Cape TownMossel Bay
Port Elizabeth
Port Alfred
East London
Port St Johns
Port Edward
Durban
16° E 18° E 20° E 22° E 24° E 26° E 28° E 30° E
30° S
32° S
34° S
36° S
32° E
Eggs m!3
(b)
Min. = 0; Max. = 163.5
100 m
200 m
100 m200 m
Figure 5: (a) Composite distribution of sardine eggs (areal abundance expressed as eggs m–2) around the South African coast derived from 8 016
CalVET net samples collected during 24 annual pelagic spawner biomass surveys from 1984 to 2007 (updated from van der Lingen and Huggett
2003); (b) distribution of sardine eggs (volumetric abundance expressed as eggs m–3 at 6 m depth; symbol size proportional to concentration)
derived from CUFES samples collected during the 2008 spawner biomass survey (CDvdL unpublished data; note that the figure is generated
from onboard counts which require laboratory validation). The solid line in (a) indicates the maximum survey coverage (note that not all surveys, in
particular those during the early years, attained this coverage), and dashed ellipses on both maps indicate the location of spawning centres possibly
indicative of functionally discrete adult assemblages or subpopulations; 100 and 200 m isobaths are shown
Downloaded At: 21:50 16 November 2010
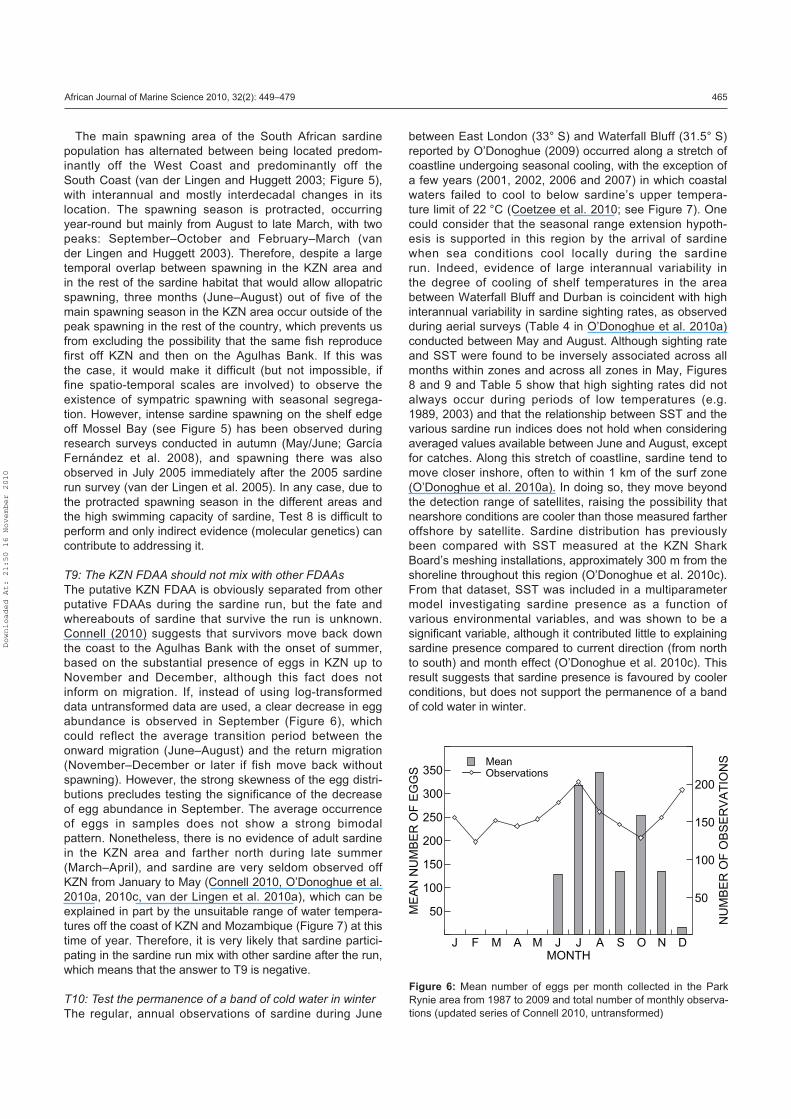
African Journal of Marine Science 2010, 32(2): 449–479 465
The main spawning area of the South African sardine
population has alternated between being located predom-
inantly off the West Coast and predominantly off the
South Coast (van der Lingen and Huggett 2003; Figure 5),
with interannual and mostly interdecadal changes in its
location. The spawning season is protracted, occurring
year-round but mainly from August to late March, with two
peaks: September–October and February–March (van
der Lingen and Huggett 2003). Therefore, despite a large
temporal overlap between spawning in the KZN area and
in the rest of the sardine habitat that would allow allopatric
spawning, three months (June–August) out of five of the
main spawning season in the KZN area occur outside of the
peak spawning in the rest of the country, which prevents us
from excluding the possibility that the same fish reproduce
first off KZN and then on the Agulhas Bank. If this was
the case, it would make it difficult (but not impossible, if
fine spatio-temporal scales are involved) to observe the
existence of sympatric spawning with seasonal segrega-
tion. However, intense sardine spawning on the shelf edge
off Mossel Bay (see Figure 5) has been observed during
research surveys conducted in autumn (May/June; García
Fernández et al. 2008), and spawning there was also
observed in July 2005 immediately after the 2005 sardine
run survey (van der Lingen et al. 2005). In any case, due to
the protracted spawning season in the different areas and
the high swimming capacity of sardine, Test 8 is difficult to
perform and only indirect evidence (molecular genetics) can
contribute to addressing it.
T9: The KZN FDAA should not mix with other FDAAs
The putative KZN FDAA is obviously separated from other
putative FDAAs during the sardine run, but the fate and
whereabouts of sardine that survive the run is unknown.
Connell (2010) suggests that survivors move back down
the coast to the Agulhas Bank with the onset of summer,
based on the substantial presence of eggs in KZN up to
November and December, although this fact does not
inform on migration. If, instead of using log-transformed
data untransformed data are used, a clear decrease in egg
abundance is observed in September (Figure 6), which
could reflect the average transition period between the
onward migration (June–August) and the return migration
(November–December or later if fish move back without
spawning). However, the strong skewness of the egg distri-
butions precludes testing the significance of the decrease
of egg abundance in September. The average occurrence
of eggs in samples does not show a strong bimodal
pattern. Nonetheless, there is no evidence of adult sardine
in the KZN area and farther north during late summer
(March–April), and sardine are very seldom observed off
KZN from January to May (Connell 2010, O’Donoghue et al.
2010a, 2010c, van der Lingen et al. 2010a), which can be
explained in part by the unsuitable range of water tempera-
tures off the coast of KZN and Mozambique (Figure 7) at this
time of year. Therefore, it is very likely that sardine partici-
pating in the sardine run mix with other sardine after the run,
which means that the answer to T9 is negative.
T10: Test the permanence of a band of cold water in winter
The regular, annual observations of sardine during June
between East London (33° S) and Waterfall Bluff (31.5° S)
reported by O’Donoghue (2009) occurred along a stretch of
coastline undergoing seasonal cooling, with the exception of
a few years (2001, 2002, 2006 and 2007) in which coastal
waters failed to cool to below sardine’s upper tempera-
ture limit of 22 °C (Coetzee et al. 2010; see Figure 7). One
could consider that the seasonal range extension hypoth-
esis is supported in this region by the arrival of sardine
when sea conditions cool locally during the sardine
run. Indeed, evidence of large interannual variability in
the degree of cooling of shelf temperatures in the area
between Waterfall Bluff and Durban is coincident with high
inter annual variability in sardine sighting rates, as observed
during aerial surveys (Table 4 in O’Donoghue et al. 2010a)
conducted between May and August. Although sighting rate
and SST were found to be inversely associated across all
months within zones and across all zones in May, Figures
8 and 9 and Table 5 show that high sighting rates did not
always occur during periods of low temperatures (e.g.
1989, 2003) and that the relationship between SST and the
various sardine run indices does not hold when considering
averaged values available between June and August, except
for catches. Along this stretch of coastline, sardine tend to
move closer inshore, often to within 1 km of the surf zone
(O’Donoghue et al. 2010a). In doing so, they move beyond
the detection range of satellites, raising the possibility that
nearshore conditions are cooler than those measured farther
offshore by satellite. Sardine distribution has previously
been compared with SST measured at the KZN Shark
Board’s meshing installations, approximately 300 m from the
shoreline throughout this region (O’Donoghue et al. 2010c).
From that dataset, SST was included in a multi parameter
model investigating sardine presence as a function of
various environmental variables, and was shown to be a
significant variable, although it contributed little to explaining
sardine presence compared to current direction (from north
to south) and month effect (O’Donoghue et al. 2010c). This
result suggests that sardine presence is favoured by cooler
conditions, but does not support the permanence of a band
of cold water in winter.
50
100
150
200
250
300
350
J F M A M J J A S O N DMONTH
ME
AN
NU
MB
ER
OF
EG
GS
50
100
150
200
NU
MB
ER
OF
OB
SE
RV
AT
ION
SMeanObservations
Figure 6: Mean number of eggs per month collected in the Park
Rynie area from 1987 to 2009 and total number of monthly observa-
tions (updated series of Connell 2010, untransformed)
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings466
Additionally, the 4 km resolution used to create the satellite
image SST time-series possibly masks smaller-scale
features of the sardine distribution. For instance, in 2006, few
sardine were observed north of the Mbhashe River (32.2° S)
whereas dense aggregations of feeding predators were
observed off Morgan Bay (32.5° S, O’Donoghue 2009). In
this case, the predator-feeding aggregations coincided both
temporally and spatially with the confluence of cool upwelled
water and warm Agulhas Current water, as evidenced from
satellite imagery that clearly depicted a thermal front in this
region. But again this suggests that the presence of cooler
water is ephemeral, allowing us to conclude that T10 is
negative.
T11: Is there more food available in the KZN area during the
sardine run than farther south?
The remotely sensed Chl a concentration time-series (Figure
10) clearly shows that the cool conditions to the south of
SST (°C)
1980
1990
2000
YE
AR
SST (5–10 km) Shark catches Sardine catches Sightings of sardine Sardine eggs
MONTH
1971
2009
1980
1990
2000
1971
2009
Catches (number) Catches (kg0.3) Sightings (%) 0
Log (eggs+1)
F A J A O D F A J A O D F A J A O D F A J A O D F A J A O D
20 22 24 >26 0 200 400 600 0 20 40 0 2 4 6 8 0.5 1.0 >2.0
Figure 8: Hovmöller diagrams of the mean monthly SST values (AVHRR data, including second pixel from the coast) in the coastal band
30°–32° S and approximately 5–10 km from the coast, compared to different monthly indices of the occurrence and intensity of the sardine
run. Diagonal lines represent missing data. The vertical line is located on 1 June. Method and original data (although displayed differently)
are available in Dudley and Cliff (2010) for shark catches; van der Lingen et al. (2010a) for sardine catches; O’Donoghue et al. (2010a) for
sardine sightings; and Connell (2010) for sardine eggs
16
18
20
24
26
28
22
29° S
30° S
31° S
32° S
33° S
Thukela River
Durban
Mdoni
Waterfall Bluff
Mbhashe River
East London
Port Alfred
Port St Johns
SST (°C)1982 1984 1986 1988 1992 1994 1996 19981990 2002 2004 2006 20082000
YEAR
Figure 7: Hovmöller plot of SST data from Pathfinder, V5 (4.5 km resolution) from the AVHRR sensor, from 28° to 34° S along the East
Coast, 1985–2009. Contours indicate the 21 and 22 °C limits; the vertical bars indicate 1 August of each year. To increase the number of
available data the second pixel from the coast was retained
Downloaded At: 21:50 16 November 2010
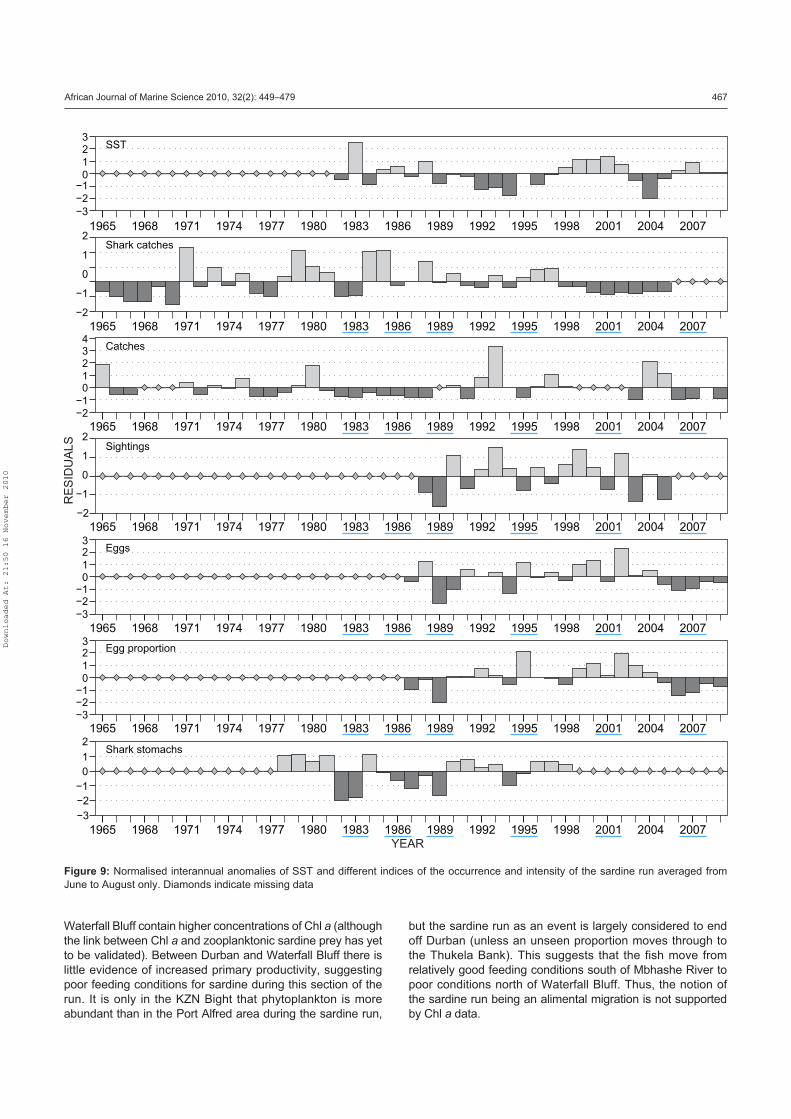
African Journal of Marine Science 2010, 32(2): 449–479 467
Waterfall Bluff contain higher concentrations of Chl a (although
the link between Chl a and zooplanktonic sardine prey has yet
to be validated). Between Durban and Waterfall Bluff there is
little evidence of increased primary productivity, suggesting
poor feeding conditions for sardine during this section of the
run. It is only in the KZN Bight that phytoplankton is more
abundant than in the Port Alfred area during the sardine run,
but the sardine run as an event is largely considered to end
off Durban (unless an unseen proportion moves through to
the Thukela Bank). This suggests that the fish move from
relatively good feeding conditions south of Mbhashe River to
poor conditions north of Waterfall Bluff. Thus, the notion of
the sardine run being an alimental migration is not supported
by Chl a data.
1965 1968 1971 1974 1977 1980 1983 1986 1989 1992 1995 1998 2001 2004 2007
"2
"3
"10
1
23
"2
"1
0
1
2
"2"1
01
234
"2
"1
0
1
2
"3
"2"1
01
23
"3"2"1
0
1
32
"2
"3
"1
0
1
2
SST
1965 1968 1971 1974 1977 1980 1983 1986 1989 1992 1995 1998 2001 2004 2007
Shark catches
1965 1968 1971 1974 1977 1980 1983 1986 1989 1992 1995 1998 2001 2004 2007
Catches
1965 1968 1971 1974 1977 1980 1983 1986 1989 1992 1995 1998 2001 2004 2007
Sightings
1965 1968 1971 1974 1977 1980 1983 1986 1989 1992 1995 1998 2001 2004 2007
Eggs
1965 1968 1971 1974 1977 1980 1983 1986 1989 1992 1995 1998 2001 2004 2007
Egg proportion
1965 1968 1971 1974 1977 1980 1983 1986 1989 1992 1995 1998 2001 2004 2007
Shark stomachs
YEAR
RE
SID
UA
LS
Figure 9: Normalised interannual anomalies of SST and different indices of the occurrence and intensity of the sardine run averaged from
June to August only. Diamonds indicate missing data
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings468
From the analyses of plankton data collected during the
2005 sardine run survey, Coetzee et al. (2010) concluded
that primary and secondary productivity levels in the KZN
area are much lower than elsewhere on the western and
eastern Agulhas Bank. Furthermore, the lower condition
factor of sardine on the East Coast, compared to the
long-term average for fish caught on the West and South
coasts (van der Lingen et al. 2006), further supports the
idea that feeding conditions on the East Coast are not ideal,
although this decrease could be attributed to the cost of
migration (van der Lingen et al. 2010c).
T12: Spatial analysis of abundance
Before testing whether or not the basin theory can be used
to explain the occurrence of the sardine run, it is necessary
to first conceptualise the design of the basin for the South
African sardine population. A hypothetical model that depicts
the available habitat of sardine in terms of the concepts
provided by MacCall 1990 (Figure 11), and which incorpo-
rates information on changing productivity (Hutchings et al.
2010) and observed sardine distributional patterns (Coetzee
et al. 2008), suggests that the basin topography should
allow for two basins. There are two reasons why: levels of
primary and secondary productivity tend to be higher on the
West Coast and western Agulhas Bank than farther east,
and data from biomass surveys conducted since 1984 show
that some sardine are located on the less productive South
Coast even at low biomass levels. If range expansion of the
sardine population was based entirely on density-dependent
habitat selection (DDHS), it would be expected under a
single-basin scenario that sardine would be restricted to
the most productive West Coast system at low biomass.
Furthermore, Coetzee et al. (2008) showed that the area
occupied by sardine in the area west of Cape Agulhas was
relatively smaller at the same level of biomass than the area
to the east of Cape Agulhas. In terms of the above theory,
SSTShark
catches
Sardine
catches
Sighting
rate
Log
(eggs+1)
Egg
proportion
Shark
stomachs
SST 1 0.546 0.008** 0.883 0.132 0.579 0.320
Shark catches #0.130 1 0.572 0.876 0.969 0.387 0.005**
Sardine catches #0.538 0.102 1 0.052 0.578 0.494 0.124
Sighting rate 0.038 0.040 0.548 1 0.218 0.221 0.095
Log (eggs+1) 0.323 0.010 0.141 0.306 1 0.000** 0.084
Egg proportion 0.122 #0.211 0.172 0.303 0.827 1 0.079
Shark stomachs #0.257 0.594 0.356 0.529 0.519 0.526 1
Table 5: Correlation matrix of annual normalised mean values of SST and different indices of sardine run occurrence and intensity (same as
in Figures 8 and 9). Note that the correlations were computed using all available data for a given couple of variables, regardless of missing
values in other variables in the same year (variable degrees of freedom). Due to their highly skewed distribution, egg data are analysed
according to two modalities: their mean number and their mean proportion of presence in samples. Correlation coefficients appear below the
diagonal and p-values above; ** indicates statistical significance at the 1% level
53
210.5
0.20.1
CHL a(mg m!3)
60
20
10
Thukela River
1998 1999 2000 2001 2002 2003 2004
Durban
Mdoni
Waterfall BluffPort St Johns
Mbhashe River
East London
Port Alfred
YEAR
Figure 10: Chlorophyll a concentration between Port Elizabeth and Richards Bay, 1998–2005. The solid and dashed vertical lines denote the
1 June and 1 September respectively for that year (adapted from O’Donoghue et al. 2010a)
Downloaded At: 21:50 16 November 2010

African Journal of Marine Science 2010, 32(2): 449–479 469
this would suggest that the area west of Cape Agulhas can
accommodate higher densities and is thus more suitable
than the area east of Cape Agulhas. Additionally, Coetzee
et al. (2008) reported that the central Agulhas Bank area
off Cape Agulhas is only inhabited at very high levels of
biomass, hence the shallow topography of the basin model
in that area. The hypothetical model also considers the
thermal limits to sardine expansion farther northwards,
both on the West Coast, where the Lüderitz upwelling cell
is a permanent barrier to further expansion north westward
(Lett et al. 2007) and on the East Coast, where increasing
temperatures in the vicinity of Port St Johns during summer
(Armstrong et al. 1991, Roberts et al. 2010) prohibit further
expansion of sardine into the coastal region of KZN. The
position of both these temperature barriers is variable,
dependent on the intensity of upwelling off Lüderitz on the
West Coast, and on seasonal changes in the dynamics
of the Agulhas Current, which strongly influences coastal
temperatures on the East Coast.
Having established a likely hypothetical model, it is possible
to evaluate some of the basin theory attributes, originally
put forward by MacCall 1990, in the context of the South
African sardine population, which enable evaluation of the
proposed tests.
The total area occupied by sardine during November
during the period 1984–2007 was calculated from distribution
maps of sardine obtained from acoustic biomass surveys
and related to changes in sardine biomass during the same
period for both the entire population (Barange et al. 2009)
and separately for sardine in the area west and east of Cape
Agulhas (Coetzee et al. 2008). These studies both showed
that contraction and expansion of the sardine distributional
range were related to sardine abundance, consistent with
MacCall’s basin theory. Nonetheless, despite this consist-
ency with basin theory, this expansion and contraction
cannot explain the occurrence of the sardine run. From the
hypothetical model (Figure 11), expansion into the area east
of Port St Johns would only be possible in times of very high
sardine biomass, because the suitability of this area is low
relative to the rest of the shelf area. It is therefore reasonable
to conclude that the basin theory does not account for the
occurrence of the run because sardine have been present in
KZN waters even at times when the biomass of the sardine
population was very low.
Additionally, it is unlikely that DDHS alone influences the
distribution of sardine. Sardine distribution patterns from
biomass surveys reveal that even at very high biomass
levels, large productive areas of the West Coast are void of
sardine, whereas densities in less productive areas of the
South Coast are high (Figure 12). This is contrary to the
DDHS theory because fish increasing their fitness through
selection of the most suitable available habitat will do better
KZN coast(low suitability)
Maximumcarryingcapacity
Cold thermal barrier
shifts slightly north or
south throughout the
year, depending on
upwelling intensity
Hig
h s
uitabili
ty
Modera
te s
uitabili
ty
Warm thermal barrier
shifts north-east in
winter and south-west
in summer
Low sardine biomass
(1984–1997 and 2005–2009):
no sardine run expected
Medium sardine biomass
(1998–2004):
no sardine run expected
High sardine biomass
(2002–2003):
sardine run expected
Port St JohnsCape AgulhasLüderitz
Figure 11: Hypothetical basin model of the habitat available to the South African sardine population (top panel) at various levels of biomass
(fullness; bottom panel)
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings470
by migrating north-westwards up the West Coast instead
of north-eastwards up the East Coast. This argument also
holds in the context of improved reproductive value (better
transport of early stages to the West Coast nursery grounds
than compared to farther east; Miller et al. 2006) or in the
context of competition with anchovy (predation pressure on
sardine eggs spawned on the South Coast is likely to be
higher there compared to farther west).
November 2003
November 1986
200 m
200 m
Hondeklip Bay
Doringbaai
Cape Town
Mossel Bay
Port Elizabeth
Port Alfred
Port St Johns
30° S
31° S
32° S
33° S
34° S
35° S
36° S
37° S17° E 18° E 19° E 20° E 21° E 22° E 23° E 24° E 25° E 26° E 27° E 28° E 29° E
Hondeklip Bay
Doringbaai
Cape Town
Mossel Bay
Port Elizabeth
Port Alfred
Port St Johns
30° S
31° S
32° S
33° S
34° S
35° S
36° S
37° S17° E 18° E 19° E 20° E 21° E 22° E 23° E 24° E 25° E 26° E 27° E 28° E 29° E
Sardine density (g m"2)
1 5 25 50 100 500
(a)
(b)
Figure 12: Relative density and distribution of sardine during (a) the pelagic spawner biomass surveys of 1986 (low biomass) and (b) 2003
(high biomass)
Downloaded At: 21:50 16 November 2010
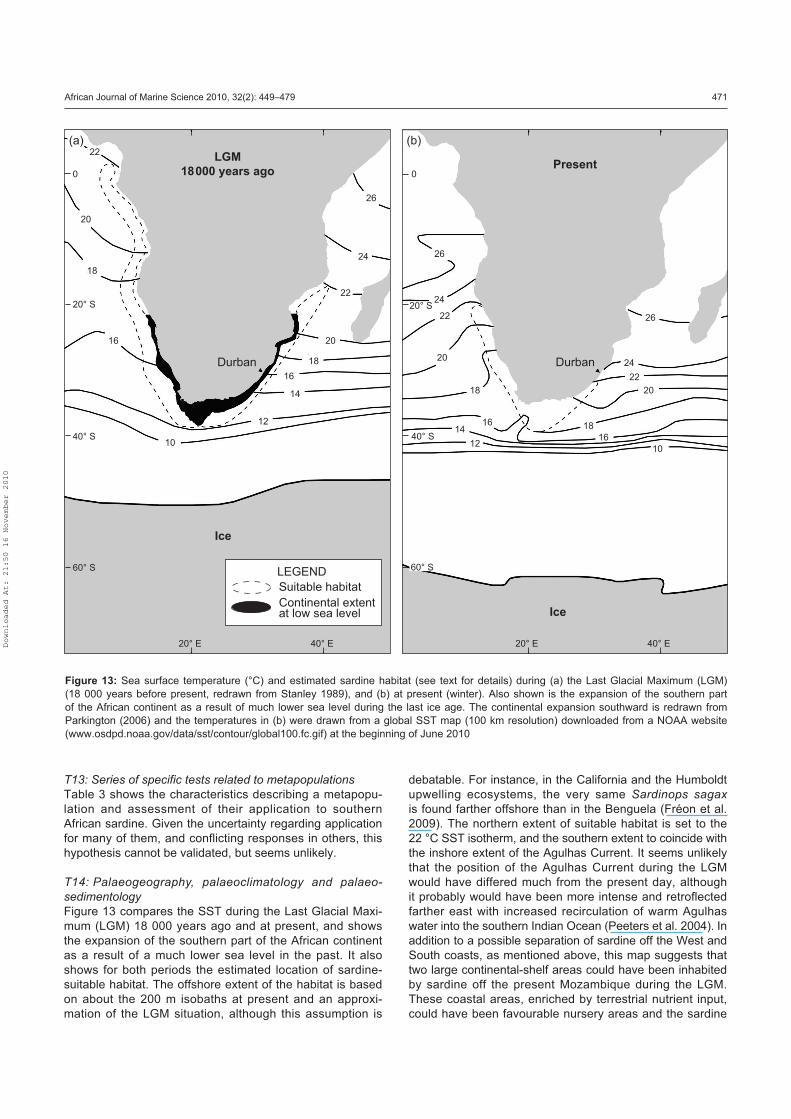
African Journal of Marine Science 2010, 32(2): 449–479 471
T13: Series of specific tests related to metapopulations
Table 3 shows the characteristics describing a metapopu-
lation and assessment of their application to southern
African sardine. Given the uncertainty regarding application
for many of them, and conflicting responses in others, this
hypothesis cannot be validated, but seems unlikely.
T14: Palaeogeography, palaeoclimatology and palaeo-
sedimentology
Figure 13 compares the SST during the Last Glacial Maxi-
mum (LGM) 18 000 years ago and at present, and shows
the expansion of the southern part of the African continent
as a result of a much lower sea level in the past. It also
shows for both periods the estimated location of sardine-
suitable habitat. The offshore extent of the habitat is based
on about the 200 m isobaths at present and an approxi-
mation of the LGM situation, although this assumption is
debatable. For instance, in the California and the Humboldt
upwelling ecosystems, the very same Sardinops sagax
is found farther offshore than in the Benguela (Fréon et al.
2009). The northern extent of suitable habitat is set to the
22 °C SST isotherm, and the southern extent to coincide with
the inshore extent of the Agulhas Current. It seems unlikely
that the position of the Agulhas Current during the LGM
would have differed much from the present day, although
it probably would have been more intense and retroflected
farther east with increased recirculation of warm Agulhas
water into the southern Indian Ocean (Peeters et al. 2004). In
addition to a possible separation of sardine off the West and
South coasts, as mentioned above, this map suggests that
two large continental-shelf areas could have been inhabited
by sardine off the present Mozambique during the LGM.
These coastal areas, enriched by terrestrial nutrient input,
could have been favourable nursery areas and the sardine
LGM
18000 years agoPresent
Ice
Ice
Suitable habitat
Continental extentat low sea level
22
20
18
16
16
16
22
20
18
16
26
24
22
20
18
26
24
22
20
18
26
24
12
10
14
1210
14
0
20° S
40° S
60° S
0
20° S
40° S
60° S
20° E 40° E 20° E 40° E
Durban Durban
LEGEND
(a) (b)
Figure 13: Sea surface temperature (°C) and estimated sardine habitat (see text for details) during (a) the Last Glacial Maximum (LGM)
(18 000 years before present, redrawn from Stanley 1989), and (b) at present (winter). Also shown is the expansion of the southern part
of the African continent as a result of much lower sea level during the last ice age. The continental expansion southward is redrawn from
Parkington (2006) and the temperatures in (b) were drawn from a global SST map (100 km resolution) downloaded from a NOAA website
(www.osdpd.noaa.gov/data/sst/contour/global100.fc.gif) at the beginning of June 2010
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings472
run could be a relic spawning migration to the small remains
of this spawning ground: the KZN Bight and adjacent areas.
This scenario is speculative, and further tests should include
at least searching for evidence of sardine presence off the
present Mozambique during the LGM, which could be investi-
gated in core sediments providing that conditions preserved
fish remains (scales, vertebrae, etc.).
T15: Are sardine run fish mostly born in KZN?
Otolith microchemistry should also provide an answer to the
question of natal homing. Under this hypothesis, the chemical
composition of the core of the otoliths should be repeated
during every reproductive season, but possibly excluding the
first one (see Discussion).
It is noteworthy that vertebral counts of sardine run fish
performed recently (van der Lingen et al. 2010c) were lower
than those from the rest of the population. Notwithstanding
the limited number of samples, the finding that sardine
collected from the KZN coast had lower vertebral counts
than elsewhere along the South African coast supports the
idea that sardine run individuals were hatched in warmer
waters than the rest of the sardine population, most probably
in the KZN area, supporting the natal return hypothesis.
Nonetheless, because Baird (1971) found no significant differ-
ences in his vertebral count, one cannot reach a firm conclu-
sion for this test.
T16: Fine-scale observations of predation
There is some circumstantial evidence to suggest that
sardine are not pushed northward by predators. Observations
of feeding predators off the East Coast indicate that only
common dolphins have the ability to herd their prey into a
‘baitball’ from which they can feed (SHO’D pers. obs.). A
comparison of sardine and predator sighting rates along
the South African east coast (see Figure 3 in O’Donoghue
et al. 2010a) indicates that common dolphin abundance
decreases markedly northwards of the Eastern Cape/
KZN border. Sardine movement towards shore peaks
along the stretch of coastline between Mdoni and Durban.
From that figure it is clear that predator sighting rates are
lower there than those farther south along the coast. The
peak in sardine shoreward movement does, however,
coincide with the shoreward intrusion of warm water from
the Durban Eddy, which can be measured as an increase
in temperature at the KZN Sharks Board meshing installa-
tions (O’Donoghue et al. 2010c), making this the more likely
cause for the shoreward movement observed in this region.
Furthermore, although it is possible that predators control
the movement of their prey during a few hours and over
a limited distance, it is unlikely that this process can work
around the clock during several days and over hundreds of
kilometres. Underwater footage of the sardine run released
in the media (e.g. the film Ocean) show that successful
attacks by predators mostly arise in the vertical plane, from
above in the case of gannets and from below in the case
of sharks and dolphins. Successful attacks in the horizontal
plane are performed in open water by predators circling
sardine. Straight attacks by sharks in shallow waters, as
illustrated in Figure 13 in Dudley and Cliff (2010), are likely
to be successful as well. However, because of the ‘fountain
pattern’, a circular movement around the detected danger
displayed by prey to maintain visual contact with their
predator (Hall et al. 1986), it is unlikely that predators will
be able to push sardine horizontally over long distances. In
conclusion, despite a lack of firm evidence against it, it is
unlikely that Hp2 is valid.
T17: Continuous coastal current measurement (e.g. ADCP
mooring line)
The ADCP mooring data collected near Port Edward in 2005
(Roberts et al. 2010) confirm high south-westward velocities
of 50–100 cm s–1 in the inshore region (30 m) of the shelf,
with bottom temperatures commonly exceeding 20 °C. Both
factors are likely to constitute a barrier to the sardine run,
and consequently allow rejection of the seasonal inshore
countercurrent hypothesis (Hp3). Most importantly, Roberts
et al. (2010) show that current reversals on the shelf are
associated with both the Natal Pulse and the more common
Durban break-away eddies, which are necessary conditions
for supporting Hp5.
T18: Embedded very high resolution numeric model
Embedded numerical hydrodynamics models in 3D (with
two or more levels) allow fine-scale studies of key areas of
ecological interest (e.g. Penven et al. 2006). Providing that
small-scale local atmospheric forcing and detailed bathym-
etry are available, such models can accurately reproduce
current structure, although in the case of the KZN area
the steep slope of the shelf and the proximity of the strong
Agulhas Current could generate some technical challenges.
A frame model and some medium-scale embedded
models already exist for the southern African region (Veitch
et al. 2009), from which such a small-scale model could be
constructed. This model would, at least, allow testing of the
permanence of a seasonal coastal countercurrent (Hp3),
the sporadic northward movement of quanta of cool water
inshore (Hp4), and the existence of Natal Pulses (Hp5).
However, validation of Hp4 and Hp5 will remain difficult at
present given limitations on the temporal resolution of data
related to past sardine run events.
T19: Compare remotely sensed SST and sardine run data
at high resolution
The results from Roberts et al. (2010) verified that, whereas
the Agulhas Current is the single-most dominant feature on
the east coast of southern Africa and strongly influences shelf
circulation, there are both stationary and transient eddies
that exist between Algoa Bay and Durban (e.g. between
Cape Padrone and Keiskamma River, East London and
Haga Haga, Rame Head and Waterfall Bluff, and Park Rynie
and Balito; see figures in Roberts et al. 2010) and result in
coastal countercurrents. This dismisses the hypothesis by
Heydorn et al. (1978) who proposed that water movement in
the form of northward advection of pockets (quanta) of cooler
water inshore of the Agulhas Current provided the physical
mechanism for the run to occur (i.e. Hp4).
The hydroacoustic survey data collected during the 2005
sardine run survey showed that between 18 and 22 June
(Leg 1) sardine were distributed between Algoa Bay and the
Mbhashe River, with the main shoals north of Haga Haga
(see Figure 10 in Coetzee et al. 2010). Satellite images
shown in Figure 11f of Roberts et al. (2010) indicate that the
Downloaded At: 21:50 16 November 2010

African Journal of Marine Science 2010, 32(2): 449–479 473
Port St Johns–Waterfall Bluff cyclonic eddy was in existence
on 16 June, with the Agulhas Current encroaching close
to the coast near the Mbhashe River. Expanded views of
the SST depict warm water (~23 °C) drawn into the Port St
Johns eddy as a northward filament that formed a distinct
thermal front. The inshore boundary of the Agulhas Current
moved offshore near Haga Haga, where the highest fish
density was measured. Figure 11g in Roberts et al. (2010)
shows that on 25 June, the inshore boundary of the Agulhas
Current had moved slightly farther offshore resulting in
water of 21–22 °C near the coast. The close association
of sardine distribution with the inshore boundary of the
Agulhas Current off the Haga Haga coast suggested that
the northward movement of the sardine shoals (and other
small pelagic species) was controlled by the dynamics of the
Agulhas Current, i.e. the Pondoland encroachment barrier.
The oceanographic situation appeared to have remained
similar for the next 4–5 days with the Port St Johns–
Waterfall Bluff eddy re-established on the 28 June and
the cooler (21–22 °C) band of coastal water terminating
north of Waterfall Bluff at Mkhambathi. To the north of
this, 24 °C water was close to the coast, extending to
Durban, i.e. the gateway was closed. The hydroacoustic
survey data collected on the southward-bound Leg 2 by
Coetzee et al. (2010) showed that sardine distribution had
spread northwards past Waterfall Bluff but no farther than
Mkhambathi (Figure 11 in Coetzee et al. 2010). The close
association between the northern extent of the sardine distri-
bution and the inshore boundary of the Agulhas Current,
and, moreover, that no sardine were found north of the
Agulhas Current coastal disjuncture, suggested again that
the dynamics of the Agulhas Current control the northward
movement of the sardine shoals. The lateral movement of
the inshore boundary of the Agulhas Current (opening and
closing of the gateway) off Waterfall Bluff for the three-month
period May–July 2005 is schematically shown in Figure 14c
in Roberts et al. (2010), and clearly illustrates that the ‘gate’
was closed during the last days of June, when the survey
was conducted. However, a transient break-away eddy,
observed in the satellite data, soon followed and the gate
opened on 13 July.
In conclusion, Armstrong et al. (1991) proposed that
the Natal Pulse was the main mechanism for the sardine
shoals to move north of Waterfall Bluff, but they dismissed
the notion because of the infrequency of the pulses. They
were, however, unaware of break-away eddies, identified in
Roberts et al. (2010), but otherwise their theory (Hp5) was
correct.
Discussion
The sardine run has stimulated the imagination of scientists
over several decades, leading to the proposal of several,
often contradictory, hypotheses to explain its existence
(Table 6). In this study, these have in the first instance been
classified according to their cause (ultimate factors that may
explain why the sardine run occurs) and enabling conditions
(proximate factors that detail how this run can take place).
Tests of these hypotheses were also proposed (Tables
2 and 4) and, where possible, applied to existing data.
Interestingly, all hypotheses with the possible exception
of one (Hu7) were found potentially falsifiable or refutable,
which means that there is a logical possibility that they
can potentially be shown false by an observation or by an
experiment, which is of particular value in science (Popper
1962). Results from these tests are summarised in Tables
7 and 8 and suggest that none of the proposed hypoth-
eses can be confirmed beyond doubt. Nonetheless, our
results are coherent since Table 6 shows (a) that when any
given hypothesis is supported, that automatically means
rejection of any other hypotheses that are contradictory, and
(b) virtually all hypotheses that are supported (or at least
inconclusive) do not contradict each other. Most hypoth-
eses related to ultimate factors can be rejected and the
two hypotheses that are supported (Hu1 and Hu7) are not in
conflict (Tables 6 and 7).
Strong support emerged for one ultimate factor hypothesis
(Hu1) and one proximate factor hypothesis (Hp5). It is likely
Hypotheses Hu1 Hu2 Hu3 Hu4 Hu5 Hu6 Hu7 Hp1 Hp2 Hp3 Hp4 Hp5
Hu1: Subpopulation spawning migration – – – – – # # # # # #
Hu2: Juvenile equatorward migration – – # – # – – # # # #
Hu3: East Coast functionally discrete adult assemblage – – # – – – – # # # #
Hu4: Seasonal extension of habitat and feeding migration – # # # # – – # # # #
Hu5: Basin theory – – – # – – – – – – –
Hu6: Metapopulation – # – # – – – # # # #
Hu7: Relic behaviour # – – – – – # # # # #
Hp1: Natal homing and imprinting # – – – – – # # # # #
Hp2: Predator-driven movement # # # # – # # # – – –
Hp3: Environmental drive: (a) favourable inshore currents # # # # – # # # – – –
Hp4: Environmental drive: (b) favourable cooling of the coastal area # # # # – # # # – – –
Hp5: Environmental drive: (c) Waterfall Bluff gateway # # # # – # # # – – –
# Compatible
– Incompatible
Table 6: Compatibility of hypotheses
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings474
that the sardine run is the seasonal spawning migration of
a genetically distinct subpopulation of sardine that is found
mixed with other subpopulations during the rest of the year.
Although genetic and otolith microchemistry tests (T4, T7
and T15) are still needed to fully validate Hu1, this hypoth-
esis is supported by positive tests based on migratory
behaviour, phenotypic differences and evidence of different
spawning sites and nursery areas (T1, T3, T5 and T6). The
existence of several subpopulations with different migratory
behaviour is common in the animal kingdom and well known,
particu larly for some bird species where only a fraction of the
population performs a seasonal migration (Berthold 2001).
Such disparity could arise from a mixed behavioural strategy
of the species which results in an evolutionary stable
strategy.
Table 8 shows that only one hypothesis (Hp5) was
supported whereas another (Hp1) remains inconclusive,
but both hypotheses are not in conflict (Table 6). Indeed,
two complementary processes can explain how the sardine
run occurs despite the presence of several strong barriers
(ecological and environmental) to its success. The first one
is the likely (but as yet not fully tested) existence of a natal
homing strategy by sardine (Hp1) which, possibly due to
imprinting of environmental cues at an early stage of egg
and/or larval development, ensures that maturing individuals
migrate back to the place where they were spawned, guided
by physical or chemical signals. Fréon and Misund (1999)
believe that natal homing is more generalised than expected
in pelagic fish and suggested that permanent chemical
stimuli, related to the local marine ecosystem and/or
continental inputs (e.g. from rivers) present in the spawning
grounds, could provide the imprinting cues. The second
likely enabling process relates to the hydrography and its
influence on functioning of the so-called Waterfall Bluff
gateway (Hp5). SST satellite data and current ADCP data
show how hydrographic processes enable sardine passage
through a normally highly unfavourable thermal and current-
related barrier, aided by the sporadic presence of Natal
Pulses and break-away eddies. The Waterfall Bluff gateway
was unlikely to have been a constraint to the northward
migration of sardine shoals during the last glacial maximum,
and is probably only a current-enabling process that was
not present in the recent past. Hp5 is incompatible with Hp4
(cooling of the coastal zone) and the two hypotheses are
Hypotheses on ultimate factors T1 T2 T3 T4 T5 T6 T7 T8 T9 T10 T11 T12 T13 T14 Conclusion
Hu1: Subpopulation spawning migration##
*
#
**
…
***
##
**
#
**
…
***
0
***
Supported
**
Hu2: Juvenile equatorward migration––
***
Rejected
***
Hu3: East Coast functionally discrete adult
assemblage
##
*
#
*
…
*
##
**
0
**
–
***
Rejected
*
Hu4: Seasonal extension/feeding migration##
*
–
**
––
**
Rejected
**
Hu5: Basin theory–
**
Rejected
**
Hu6: Metapopulation–
**
Rejected
*
Hu7: Relic behaviour##
*
#
**
…
***
##
**
#
**
…
***
0
***
0
**
Supported
*
Result of the test: … to be performed; – negative; –– strongly negative; 0 inconclusive; # positive; ## strongly positive
Strength of the test and conclusion: * low; ** medium; *** high
Table 7: Different tests (Tn) applied to different hypotheses about ultimate factors (Hun) and corresponding results and final conclusion.
Note that the strength of a test may also differ according to the tested hypothesis. Blank cells mean that the test is not relevant for the
corresponding hypothesis
Hypotheses on proximate factors T15 T16 T17 T18 T19 Conclusion
Hp1: Natal homing and imprinting
…/0
*** Inconclusive
Hp2: Predators-driven movement–
**
Rejected
**
Hp3: Favourable inshore currents—
***
…
*
Rejected
**
Hp4: Favourable cooling of coastal area…
*
–
**
Rejected
*
Hp5: Waterfall Bluff gateway#
**
Supported
**
Table 8: Different tests (Tn) applied to different hypotheses on proximate factors (Hpn) and corresponding results and final conclusion (see
Table 7 for definition of symbols). Note that T15 has been performed twice for vertebral count data, and is still to be performed for otolith
microchemistry
Downloaded At: 21:50 16 November 2010

African Journal of Marine Science 2010, 32(2): 449–479 475
tested differently (T10 and T11 versus T8 and T9 respec-
tively) due to differences in the processes involved and their
duration (short term for Hp5), although there are similarities
between the end result of each process.
The choice to migrate does not need to be a conscious
one and in the case of the sardine run, in which several
impediments and challenges to migration are encountered,
it probably is not. During the 2005 sardine run survey, one of
the authors (LH) jotted some wisdom down in his notebook
about conditions in KZN: ‘Line 20 Scottburgh, 25/06/2005,
3am. Very similar to line 16. Warm water, deep weak thermo-
clines, low concentrations of cells, flat particle spectra. Why
anything wants to live in this godforsaken place, let alone
migrate hundreds of km to get here in the face of incredible
predation, is quite beyond me. Life must be tough at home!’.
This aptly sums up the obstacles faced by migrating sardine
and suggests that only a strong instinctive behaviour, geneti-
cally hard-wired and generating automated responses to
sensory cues, can explain a counter-intuitive behaviour such
as the sardine run.
Analogies can be drawn with other spectacular natural
migrations such as the obstinate upstream migration of
salmon challenged by natural or artificial obstacles or the
‘trek of the Wildebeest’ (a migration of 1.5 million ungulates)
across the vast plains of Africa, relentlessly tracked by their
predators, in search of rain-ripened grazing. Such behaviour
has to be rewarding in terms of an individual’s reproduct ive
success. In the case of the sardine run the reward is
obviously not food-related, nor is it related to predator
avoidance. It is more likely that in depositing eggs upstream
of a recruitment ground, as do many other species in the
region (Hutchings et al. 2002), sardine will gain an advantage
for the survival of their offspring.
The hypothesis that the sardine run exists due to relic
behaviour (Hu7) inherited during the Last Glacial Maximum,
when one or several large sardine nursery areas were
likely located off the coast of the present Mozambique, was
also supported, mostly based on the same tests as those
performed for Hu1 because the hypotheses are closely
related. Indeed they both imply a seasonal migration of a
subpopulation, the only difference being that Hu1 does not
imply the existence of prehistoric distant nursery areas and,
conversely, Hu7 does not imply that present day spawning
on the KZN coast is highly rewarding in terms of repro ductive
success. Even so, for a subpopulation to have persisted for
the long periods of time implied by Hu7, it must have a viable
reproductive strategy. There is, however, at least one factor
that could counter the validity of Hu7 and to a large extent
Hu1 as well: if the same cohorts, after reaching maturity,
repeatedly take part in the sardine run up until the end of
their life (which is a prerequisite of Hu1 and Hu7), and if
mortality is much higher for the sardine run subpopulation
than for the rest of the population, then the age structure
of the former should be biased toward younger individuals.
Analyses of the few available data (van der Lingen et al.
2010c) do not tend to support this, although accurate ageing
of sardine is difficult (Durholtz 2005). Another explanation
could be that young adult sardine do not migrate during their
first or second season after attaining sexual maturity in order
to build up their resources and size in the south, such that
they are more likely to make a greater and more successful
contribution in the following year. This assumption is linked
to the fact that larger fish migrate farther, as discussed in
Coetzee et al. (2010). Further analyses of length and
age data would assist in either supporting or refuting this
hypothesis.
The similar length distributions for sardine >15 cm CL
obtained from commercial purse-seine catches and midwater
trawl catches when compared with beach-seine catches off
KZN (Figures 2 and 3) is interesting and could possibly result
from an interaction between the higher natural mortality rate
experienced during the sardine run being offset by a low
exploitation rate. Indeed, the average commercial catch of
sardine taken in KZN since the 1970s is <150 t year#1 (van
der Lingen et al. 2010a). If one assumes a sardine run
biomass of 30 000 t (Coetzee et al. 2010), this amounts to
an exploitation rate of only 2.5%, which is far lower than that
for the rest of the population (11% for the period 1987–2007,
Coetzee et al. 2008).
Some tests are so powerful (indicated by stars in Tables
7 and 8) that, had they been performed, they would have
allowed conclusive validation or rejection of some hypoth-
eses. This applies particularly for the molecular genetics
(T4) and otolith microchemistry (T7, T15) methods, and even
more so if the two approaches are applied concurrently on
the same individuals. Doing this would have allowed better
support of Hu1, Hu7 and their related proximate hypothesis
Hp1. Some other powerful tests were performed, but their
results were not always conclusive owing to data paucity,
the number of repetitions and technical issues related to
data collection. Improved datasets are advised. A difficulty
inherent in some potential tests (e.g. T19) is that the
different indices of the sardine run occurrence and intensity
are seldom in agreement with each other (Figure 8),
because they are inaccurate or were not computed at similar
temporal and spatial scales. For instance, in contrast to all
other time-series, egg data collected off KZN throughout
the year (Connell 2010) provide evidence not only of the
sardine run, but also of the fact that sardine remain off the
KZN coast, and reproduce there for several months until
their return migration in spring.
The return migration remains poorly understood and is
only evidenced by the presence of eggs in the vicinity of Park
Rynie long after the sardine run has occurred (Figure 6). It is
likely that sardine move back to the South Coast using the
fringe of the powerful Agulhas Current to aid migration at a
depth that is suitable in term of temperature. This strategy
should be energy saving and would strongly limit predation,
at least by birds.
Recommendations for future work that could improve
understanding of the sardine run include: (1) a combi na tion of
molecular genetics (T4) and otolith microchemistry (T7, T15);
(2) collection of further evidence of a sardine nursery ground
in the KwaZulu-Natal Bight area in summer; (3) employment of
high resolution (<1 km) satellite data to study oceanographic
conditions present in the inshore area; (4) repetition of ADCP
moorings and determination of oxygen content at strategic
sites along the KZN coast (T18), including shallow waters;
and contrast of ADCP records with evidence of sardine run
events. Further suggestions relate to improved knowledge on
palaeogeog raphy, palaeoclimatology and palaeosediment-
ology (T14); improved monitoring of commercial catches
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings476
during the sardine run to derive an index of abundance;
monthly acoustic surveys along the KZN coast to test for the
possible existence of a deeper northward migration of sardine
shoals to the KZN spawning area during June–July and to
gain knowledge about the return migration; or alternatively
perform stomach content analyses of predators in the KZN
area from mid-September to mid-December.
In conclusion, the efforts of the scientific community over
the past five decades, as summarised in van der Lingen
et al. (2010a), have substantially improved understanding
of ecological processes associated with this extraordinary
wildlife phenomenon. The basis of this paper was to evaluate
the validity of existing hypotheses, several of which were
generated at a time when the ecological processes affecting
the sardine run were poorly understood. Several of these
have now been rejected, and others can only be tested after
more data become available, or advanced techniques are
applied. These limitations notwithstanding, our approach
suggests that the sardine run is most likely a seasonal
spawning migration of a genetically distinct subpopulation
of sardine responding to a strong instinct of natal homing.
The mechanism enabling passage through unfavourable
thermal and current-related barriers is the sporadic presence
of Natal Pulses and break-away eddies that allow sardine to
migrate into a hostile environment to complete their life cycle
following successful spawning in KZN waters. In summary,
this subpopulation is likely to follow the conventional
triangular plan of migration (Harden Jones 1968) described
in our Introduction.
Acknowledgements — Laurent Drapeau (IRD, US 140 ESPACE,
France) is sincerely acknowledged for his contributions to satellite
data analysis and time-series analysis. Mike Armstrong (CEFAS)
is thanked for his comments on an early draft of the manuscript,
and George Branch (University of Cape Town) and an anonymous
referee are warmly acknowledged for their detailed and rapid reviews
of this paper.
References
Almany GR, Berumen ML, Thorrold SR, Planes S, Jones P. 2007.
Local replenishment of coral reef fish populations in a marine
reserve. Science 316: 742–744.
Andreu B. 1969. Las branquispinas en la caracterizacion de las
poblaciones de Sardina pilchardus (Walb.). Investigaciones
Pesquera 33: 427–523.
Armstrong MJ, Berruti A, Colclough J. 1987. Pilchard distribution
in South African waters, 1983–1985. In: Payne AIL, Gulland JA,
Brink KH (eds), The Benguela and comparable ecosystems.
South African Journal of Marine Science 5: 871–886.
Armstrong MJ, Chapman P, Dudley SFJ, Hampton I, Malan PE.
1991. Occurrence and population structure of pilchard Sardinops
ocellatus, round herring Etrumeus whiteheadi and anchovy
Engraulis capensis off the east coast of southern Africa. South
African Journal of Marine Science 11: 227–249.
Attwood CG, Cowley PD. 2005. Alternate explanations of the
dispersal pattern of galjoen (Dichistius capensis). South African
Journal of Marine Science 27: 141–156.
Baird D. 1971. Seasonal occurrence of the pilchard Sardinops
ocellata on the east coast of South Africa. Investigational Report,
Division of Sea Fisheries, South Africa 96: 1–19.
Bakun A. 2005. Seeking an expanded suite of management tools:
implication of rapidly-evolving adaptive response mechanisms (e.g.
“school-mix feedback”). Bulletin of Marine Science 76: 463–483.
Barange M, Coetzee J, Takasuka A, Hill K, Gutierrez M, Oozeki
Y, van der Lingen C, Agostini V. 2009. Habitat expansion and
contraction in anchovy and sardine populations. Progress in
Oceanography 83: 251–260.
Barange M, Hampton I, Roel BA. 1999. Trends in the abundance
and distribution of anchovy and sardine on the South African
continental shelf in the 1990s, deduced from acoustic surveys.
South African Journal of Marine Science 21: 367–391.
Beckley LE, Hewitson JD. 1994. Distribution and abundance of
clupeoid larvae along the east coast of South Africa in 1990/91.
South African Journal of Marine Science 14: 205–212.
Beckley LE, Naidoo AK. 2003. Exploratory trials with light-traps to
investigate settlement stage fishes in subtropical, coastal waters
off South Africa. African Zoology 38: 333–342.
Begg GA, Waldman JR. 1999. An holistic approach to fish stock
identification. Fisheries Research 43: 35–44.
Ben Tuvia A. 1964. Influence of temperature on vertebral number
of Sardinella aurita from Eastern Mediterranean. Israel Journal of
Zoology 12: 59–66.
Berthold P (ed.). 2001. Bird migration: a general survey (2nd edn).
Oxford: Oxford University Press.
Carr A. 1967 (ed.). So excellent a fish: a natural history of sea
turtles. New York: Schribner.
Carscadden JE, Frank KT, Miller DS. 1989. Capelin (Mallotus
villosus) spawning on the Southeast shoal: influence of physical
factors past and present. Canadian Journal of Fisheries and
Aquatic Sciences 46: 1743–1754.
Charnov EL. 1976. Optimal foraging, the marginal value theorem.
Theoretical Population Biology 9: 129–136.
Chow S, Ushiama H. 1995. Global population structure of albacore
(Thunnus alalunga) inferred by RFLP analysis of the mitochondrial
ATPase gene. Marine Biology 123: 39–45.
Coetzee JC, Merkle D, Hutchings L, van der Lingen CD, van den
Berg M, Durholtz MD. 2010. The 2005 KwaZulu-Natal sardine
run survey sheds some light on the ecology of small pelagic
fish off the east coast of South Africa. African Journal of Marine
Science 32: 337–360.
Coetzee JC, van der Lingen CD, Fairweather T, Hutchings L. 2008.
Has the fishery contributed to a major shift in the distribution
of South African sardine? ICES Journal of Marine Science 65:
1676–1688.
Connell AD. 2001. Pelagic eggs of marine fishes from Park Rynie,
KwaZulu-Natal, South Africa: seasonal spawning patterns of the
three most common species. African Zoology 36: 197–204.
Connell AD. 2010. A 21-year ichthyoplankton collection confirms
sardine spawning in KwaZulu-Natal waters. African Journal of
Marine Science 32: 331–336.
Cury P. 1994. Obstinate nature: an ecology of individuals. Thoughts
on reproductive behaviour and biodiversity. Canadian Journal of
Fisheries Aquatic Sciences 57: 1664–1673.
Davies DH. 1956a. The South African pilchard (Sardinops ocellata).
Sexual maturity and reproduction 1950–1954. Investigational
Report, Division of Sea Fisheries, South Africa 22: 1–155.
Davies DH. 1956b. The South African pilchard (Sardinops ocellata).
Migration 1950–55. Investigational Report, Division of Sea
Fisheries, South Africa 24: 1–52.
Dawson MN, Grosberg RK, Botsford LW. 2006. Connectivity in
Marine Protected Areas. Science 313: 43–44.
de Moor CL, Butterworth DS. 2009a. A 2-stock hypothesis for
South African sardine: two discrete stocks. Unpublished report,
MCM/2009/SWG-PEL/23. Cape Town: Marine and Coastal
Management, Department of Environmental Affairs and Tourism.
de Moor CL, Butterworth DS. 2009b. A two discrete stock hypothesis
for South African sardine resource. Unpublished report, MCM/2009/
SWG-PEL/47. Cape Town: Marine and Coastal Management,
Department of Environmental Affairs and Tourism.
de Ruijter, WPM, van Leeuwen PJ, Lutjeharms JRE. 1999.
Downloaded At: 21:50 16 November 2010

African Journal of Marine Science 2010, 32(2): 449–479 477
Generation and evolution of Natal pulses: solitary meanders
in the Agulhas Current. Journal of Physical Oceanography 29:
3043–3055.
Dicken ML. 2010. Socio-economic aspects of boat-based ecotourism
during the sardine run within the Pondoland Marine Protected Area,
South Africa. African Journal of Marine Science 32: 405–411.
Dingle H (ed.). 1996. Migration. The biology of life on the move.
New York: Oxford University Press.
Dudley SJF, Cliff G. 2010. Influence of the annual sardine run on
catches of large sharks in the protective gillnets off KwaZulu-
Natal, South Africa, and the occurrence of sardine in shark diet.
African Journal of Marine Science 32: 383–397.
Durholtz D. 2005. Report on sardine age determination. Unpublished
report, SWG/AUG2005/PEL/06. Cape Town: Marine and Coastal
Management, Department of Environmental Affairs and Tourism.
Fennessy ST, Pradervand P, de Bruyn PA. 2010. Influence of the
sardine run on selected nearshore predatory teleosts in KwaZulu-
Natal. African Journal of Marine Science 32: 375–382.
Fletcher WJ, Tregonning RJ, Sant GJ. 1994. Interseasonal variation
in the transport of pilchard eggs and larvae off southern Western
Australia. Marine Ecology Progress Series 111: 209–224.
Fréon P, Arístegui J, Bertrand A, Crawford RJM, Field JC, Gibbons
MJ, Tam J, Hutchings L, Masski H, Mullon C, Ramdani M, Seret
B, Simier M. 2009. Functional group biodiversity in eastern
boundary upwelling ecosystems questions the wasp-waist trophic
structure. Progress in Oceanography 83: 97–106.
Fréon P, Misund OA (eds). 1999. Dynamics of pelagic fish distrib-
ution and behaviour: effects on fisheries and stock assessment.
Cambridge: University Press.
Fréon P, Stéquert B. 1982. Note sur la présence de Sardina
pilchardus (Walb) au Sénégal : étude de la biométrie et interpré-
tation. Rapports et Procès-verbaux des Réunions du Conseil
international pour l’Exploration de la Mer 180: 345–349.
Fretwell SD, Lucas HL. 1970. On territorial behaviour and other
factors influencing habitat distribution in birds. I. Theoretical
development. Acta Biotheoretica 19: 16–36.
Fromentin J-M, Ernande B, Fablet R, de Pontual H. 2009.
Importance and future of individual markers for the ecosystem
approach to fisheries. Aquatic Living Resources 22: 395–408.
García Fernández I, van der Lingen CD, Moloney C. 2008.
Characterization and comparison of autumn and spring
spawning habitat of sardine (Sardinops sagax) and round herring
(Etrumeus whiteheadi) on the Eastern Agulhas Bank, South
Africa. Symposium guide and book of abstracts, South African
Marine Science Symposium 2008, Cape Town, South Africa.
Cape Town: South African Network for Coastal and Oceanic
Research. pp 37–38.
Gaughan DJ, Fletcher WJ, McKinlay JP. 2002. Functionally distinct
adult assemblages within a single breeding stock of the sardine,
Sardinops sagax: management units within a management unit.
Fisheries Research 59: 217–231.
Gaughan DJ, White KV, Fletcher WJ. 2001. The links between
functionally distinct adult assemblages of Sardinops sagax: larval
advection across management boundaries. ICES Journal of
Marine Science 58: 597–606.
Gerkin SD. 1959. The restricted movements of fish populations.
Biological Reviews of the Cambridge Philosophical Society 34:
221–242.
Grant WS. 1985. Population genetics of the southern African
pilchard, Sardinops ocellata, in the Benguela upwelling system.
In: Bas C, Margalef R, Rubies P (eds), International symposium
on the most important upwelling areas off western Africa (Cape
Blanco and Benguela), Barcelona, 1983. Barcelona: Instituto de
Investigaciones Pesqueras. pp 551–562.
Hall SJ, Wardle CS, MacLennan DN. 1986. Predator evasion in a
fish school : test of a model for the fountain effect. Marine Biology
91: 143–148.
Hanski I. 1999. Metapopulation ecology. New York: Oxford University
Press.
Harden Jones FR. 1968. Fish migration. London: Edward Arnold
Publishers Ltd.
Hauser L, Carvalho GR. 2008. Paradigm shifts in marine fisheries
genetics: ugly hypotheses slain by beautiful facts. Fish and
Fisheries 9: 333–362.
Heydorn AEF, Bang ND, Pearce AF, Flemming BW, Carter RA,
Schleyer MH, Berry PF, Hughes GR, Bass AJ, Wallace JH, van
der Elst RP, Crawford RJM, Shelton PA. 1978. Ecology of the
Agulhas Current region: an assessment of biological responses
to environmental parameters in the south-west Indian Ocean.
Transactions of the Royal Society of South Africa 43: 151–190.
Hiramoto K. 1991. The sardine fishery and ecology in the Joban and
Boso waters of Central Japan. In: Kawasaki T, Tanaka S, Toba Y,
Taniguchi A (eds), Long-term variability of pelagic fish populations
and their environment. Oxford: Pergamon Press. pp 117–128.
Hutchings L. 1992. Fish harvesting in a variable, productive
environment – searching for rules or searching for exceptions?
South African Journal of Marine Science 12: 297–318.
Hutchings L, Beckley LE, Griffiths MH, Roberts MJ, Sundby S, van
der Lingen C. 2002. Spawning on the edge: spawning grounds
and nursery areas around the southern African coastline. Marine
and Freshwater Research 53: 307–318.
Hutchings L, Morris T, van der Lingen CD, Lamberth SJ, Connell
AD, Taljaard S, van Niekerk L. 2010. Ecosystem considerations
of the KwaZulu-Natal sardine run. African Journal of Marine
Science 32: 413–421.
Inoue JG, Miya M, Miller MJ, Sado T, Hanel R, Hatooka K, Aoyama
J, Minegishi Y, Nishida M, Tsukamoto K. 2010. Deep-ocean
origin of the freshwater eels. Biology Letters 6: 363–366.
Jones GP, Planes S, Thorrold SR. 2005. Coral reef fish larvae
settle close to home. Current Biology 15: 1314–1318.
Jonsson B, Jonsson N. 1993. Partial migration: niche shift versus
sexual maturation in fishes. Reviews in Fish Biology and Fisheries
3: 348–365.
Kawanabe H. 1977. Relic social structure hypothesis on the terri-
toriality of ayu, Plecoglossus altivelis (Pisces: Osmeridae).
Proceedings of the Japan Academy, Series B 53: 74–77.
Kennedy JS. 1985. Migration, behavioural and ecological. In: Rankin
MA (ed.), Migration: mechanisms and adaptive significance.
Contributions to the Marine Sciences 27: 5–26.
Kilpatrick KA, Podesta GP, Evans R. 2001. Overview of the NOAA/
NASA Advanced Very High Resolution Radiometer Pathfinder
algorithm for sea surface temperature and associated matchup
database. Journal of Geophysical Research 106(C5): 9179–9197,
doi:10.1029/1999JC000065.
King DPF. 1977. Influence of temperature, dissolved oxygen and
salinity on incubation and early larval development of the South
West African pilchard Sardinops ocellata. Investigational Report,
Division of Sea Fisheries, South Africa 114: 1–35.
Kritzer JP, Sale PF. 2004. Metapopulation ecology in the sea: from
Levins’ model to marine ecology and fisheries science. Fish and
Fisheries 5: 131–140.
Law RM. 2007. Fisheries-induced evolution: present status and
future directions. Marine Ecology Progress Series 335: 271–277.
Lett C, Veitch J, van der Lingen CD, Hutchings L. 2007. Assessment
of an environmental barrier to transport of ichthyoplankton from the
southern to the northern Benguela ecosystems. Marine Ecology
Progress Series 347: 247–259.
Levins R (ed.). 1968. Evolution in changing environments. Princeton,
New Jersey: Princeton University Press.
Lutjeharms JRE, Grundlingh ML, Carter RA. 1989. Topographically
induced upwelling in the Natal Bight. South African Journal of
Science 85: 310–316.
Lutjeharms JRE, Roberts HR 1988. The Natal Pulse: an extreme
transient on the Agulhas Current. Journal of Geophysical Research
Downloaded At: 21:50 16 November 2010

Fréon, Coetzee, van der Lingen, Connell, O’Donoghue, Roberts, Demarcq, Attwood, Lamberth and Hutchings478
93(C1): 631–646, doi:10.1029/JC093iC01p00631.
MacCall AD (ed.). 1990. Dynamic geography of marine fish popula-
tions. Seattle: University of Washington Press.
Mace GM, Harvey PH, Chutton-Brock TH. 1984. Vertebrate
home-range size and energetic requirements. In: Swingland IR,
Greenwood PJ (eds), The ecology of animal movement. Oxford:
Clarendon Press. pp 32–53.
McQuinn IH. 1997. Metapopulations and the Atlantic herring. Reviews
in Fish Biology and Fisheries 7: 297–329.
Miller DCM, Moloney CL, van der Lingen CD, Lett C, Mullon C,
Field JG. 2006. Modelling the effects of physical-biological
interactions and spatial variability in spawning and nursery areas
on transport and retention of sardine Sardinops sagax eggs and
larvae in the Southern Benguela ecosystem. Journal of Marine
Systems 61: 212–229.
Miller DE. 1990. A southern African Late Quaternary sea-level curve.
South African Journal of Science 86: 456–458.
Mketsu QK. 2008. Comparative dietary analysis of sardine and
other clupeids from presumed mixed shoals off South Africa’s
east coast. MSc thesis, University of Cape Town, South Africa.
Mullon C, Cury P, Penven P. 2002. Evolutionary individual-based
model for the recruitment of anchovy (Engraulis capensis) in the
southern Benguela. Canadian Journal of Fisheries and Aquatic
Sciences 59: 910–922.
Myeza J, Mason RB, Peddemors VM. 2010. Socio-economic
implications of the KwaZulu-Natal sardine run for local indigenous
communities. African Journal of Marine Science 32: 399–404.
Newman GG. 1970. Migration of the pilchard (Sardinops ocellata) in
southern Africa. Investigational Report, Division of Sea Fisheries,
South Africa 86: 1–6.
Noakes DLG. 1992. Behaviour and rhythms in fishes. In: Ali MA
(ed.), Rhythms in fishes. New York: Plenum Press. pp 39–50.
O’Donoghue SH. 2009. The sardine run: investigating sardine,
Sardinops sagax, and predator distributions in relation to
environmental conditions using GIS and remotely sensed products.
PhD thesis, University of KwaZulu-Natal, Durban, South Africa.
O’Donoghue SH, Drapeau L, Dudley SFJ, Peddemors VM. 2010c.
The KwaZulu-Natal sardine run: shoal distribution in relation to
nearshore environmental conditions, 1997–2007. African Journal
of Marine Science 32: 293–307.
O’Donoghue SH, Drapeau L, Peddemors VM. 2010a. Broad-scale
distribution patterns of sardine and their predators in relation to
remotely sensed environmental conditions during the KwaZulu-
Natal sardine run. African Journal of Marine Science 32:
279–291.
O’Donoghue SH, Whittington PA, Peddemors VM, Dyer BM 2010b.
Abundance and distribution of avian and marine mammal
predators of sardine observed during the 2005 KwaZulu-Natal
sardine run survey. African Journal of Marine Science 32:
361–374.
Parkington J (ed.). 2006. Shorelines, strandlopers and shell middens.
Cape Town: Krakadouw Trust.
Pearce AF. 1977. The shelf circulation off the east coast of South
Africa. CSIR Research Report No. 361. Stellenbosch: Council for
Scientific and Industrial Research.
Peeters FJC, Acheson R, Brummer GJA, de Ruijter WPM,
Schneider RR, Ganssen GM, Ufkes E, Kroon D. 2004. Vigorous
exchange between the Indian and Atlantic oceans at the end of
the past five glacial periods. Nature 430: 661–665.
Penven P, Debreu L, Marchesiello P, McWilliams JC. 2006.
Application of the ROMS embedding procedure for the Central
California Upwelling System. Ocean Model 12: 157–187.
Popper K (ed.). 1962. Conjectures and refutations: the growth of
scientific knowledge. New York: Basic Books.
Roberts MJ, van der Lingen CD, Whittle C, van den Berg M. 2010.
Shelf currents, lee-trapped and transient eddies on the inshore
boundary of the Agulhas Current, South Africa: their relevance to
the KwaZulu-Natal sardine run. African Journal of Marine Science
32: 423–447.
Romero A. 1985. Ontogenetic change in phototactic responses
of surface and cave populations of Astyanax fasciatus, Pisces.
Copeia 4: 1004–1011.
Rooker JR, Secor DH, De Metrio G, Schlosser R, Block BA, Neilson
JD. 2008. Natal homing and connectivity in Atlantic bluefin tuna
populations. Science 322: 742–744.
Schumann EH. 1987. The coastal ocean off the east coast of South
Africa. Transactions of the Royal Society of South Africa 46:
215–229.
Shields WM. 1984. Optimal inbreeding and the evolution of
philopatry. In: Swingland IR, Greenwood PJ (eds), The ecology
of animal movement. Oxford: Clarendon Press. pp 132–159.
Sinclair ARE. 1984. The function of distance movements in vertebrates.
In: Swingland IR, Greenwood PJ (eds), The ecology of animal
movement. Oxford: Clarendon Press. pp 240–258.
Smith RL (ed.). 1980. Ecology and field biology (3rd edn). New
York: Harper & Row Publishers.
Solmundsson J, Palsson J, Karlsson H. 2005. Fidelity of mature
Icelandic plaice (Pleuronectes platessa) to spawning and feeding
grounds. ICES Journal of Marine Science 62: 189–200.
Stabell OB. 1984. Homing and olfaction in salmonids: a critical
review with special reference to the Atlantic salmon. Biological
Review 59: 333–388.
Stanley SM. 1989. Exploring earth and life through time. New York:
Freeman WH and Company Ltd.
Stephenson RL, Melvin GD, Power MJ. 2009. Population integrity
and connectivity in Northwest Atlantic herring: a review of
assumptions and evidence. ICES Journal of Marine Science 66:
1733–1739.
Swingland IR. 1984. Intraspecific differences in movement. In:
Swingland IR, Greenwood PJ (eds), The ecology of animal
movement. Oxford: Clarendon Press. pp 102–115.
Thorrold SR, Latkoczy C, Swart PK, Jones CM. 2001. Natal homing
in a marine fish metapopulation. Science 291: 297–299.
Tinbergen N. 1963. On aims and methods of ethology. Zeitschrift
für Tierpsychologie 20: 410–433.
Tsukamoto K, Aoyama J, Miller MJ. 2002. Migration, speciation,
and the evolution of diadromy in anguillid eels. Canadian Journal
of Fisheries and Aquatic Sciences 59: 1989–1998.
Tyler JA, Rose KA. 1994. Individual variability and spatial
heterogeneity in fish population models. Review of Fish Biology
and Fisheries 4: 91–123.
van der Byl PCN. 1978. The annual Natal sardine run 1978.
Internal Report. Cape Town: Sea Fisheries Branch, Department
of Industries.
van der Byl PCN. 1980. The Natal sardine run. Annual Report No.
3. Cape Town: Sea Fisheries Institute, Department of Agriculture
and Fisheries..
van der Lingen CD, Checkley D, Barange M, Hutchings L, Osgood K.
1998. Assessing the abundance and distribution of eggs of sardine,
Sardinops sagax, and round herring, Etrumeus whiteheadi, on the
western Agulhas Bank, South Africa, using a continuous, underway
fish egg sampler. Fisheries Oceanography 7: 35–47.
van der Lingen CD, Coetzee JC, Hutchings L. 2010a. Overview of
the Kwazulu-Natal sardine run. African Journal of Marine Science
32: 271–277.
van der Lingen CD, Coetzee JC, Hutchings L (eds). 2010b. The
KwaZulu-Natal sardine run. African Journal of Marine Science
32: 269–479.
van der Lingen C[D], Coetzee J, Hutchings L, Peddemors V, Merkle
D, van den Berg M, van der Westhuizen J, Dyer B. 2005. Report
on the 2005 sardine run survey. Unpublished report, SWG/
AUG2005/PEL/04. Cape Town: Marine and Coastal Management,
Department of Environmental Affairs and Tourism.
van der Lingen CD, Durholtz MD, Fairweather TP, Melo Y. 2009.
Downloaded At: 21:50 16 November 2010

African Journal of Marine Science 2010, 32(2): 449–479 479
Spatial variability in biological characteristics of southern Benguela
sardine and the possible existence of two stocks. Unpublished
report, MCM/2009/SWG-PEL/39. Cape Town: Marine and Coastal
Management, Department of Environmental Affairs and Tourism.
van der Lingen CD, Fréon P, Fairweather TP, van der Westhuizen
JJ. 2006. Density-dependent changes in reproductive parameters
and condition of southern Benguela sardine Sardinops sagax.
African Journal of Marine Science 28: 625–636.
van der Lingen CD, Hendricks M, Durholtz MD, Wessels G,
Mtengwane C. 2010c. Biological characteristics of sardine caught
by the beach-seine fishery during the KwaZulu-Natal sardine run.
African Journal of Marine Science 32: 309–330.
van der Lingen CD, Huggett JA. 2003. The role of ichthyoplankton
surveys in recruitment research and management of South African
anchovy and sardine. In: Browman HI, Skiftesvik AB (eds), The big
fish bang: proceedings of the 26th annual larval fish conference.
Bergen: Institute of Marine Research. pp 303–343.
van der Lingen CD, van der Westhuizen JJ. 2000. Use of a
continuous, underway fish egg sampler in South Africa. In
Checkley DM Jr, Hunter JR, Motos L, van der Lingen CD (eds),
Report of a workshop on the use of the continuous underway fish
egg sampler (CUFES) for mapping spawning habitats of pelagic
fish. GLOBEC Report 14: 27–30.
Veitch J, Penven P, Shillington F. 2009. The Benguela: a laboratory
for comparative modelling studies. Progress in Oceanography
83: 296–302.
Viñas J, Alvarado Bremer JR, Pla C. 2004. Inter-oceanic genetic
differentiation among albacore (Thunnus alalunga) populations.
Marine Biology 145: 225–232.
Wyatt T, Cushing DH, Junquera S. 1991. Stock distinctions and
evolution of European sardine. In: Kawasaki T, Tanaka S,
Toba Y, Taniguchi A (eds), Long term variability of pelagic fish
populations and their environment. Oxford: Pergamon Press. pp
229–238.
Downloaded At: 21:50 16 November 2010

Downloaded At: 21:50 16 November 2010




















