
A paleoscience approach to estimating the effects of climatic warming on salmonid fisheries of the...
8
489 A paleoscience approach to estimating the effects of climatic warming on salmonid fisheries of the Columbia River basin James C. Chatters1 and Virginia L. Butler2, Michael J. Scott, David M.Anderson, and Duane A. Neitzel Chatters. J.C..V.L. Butler, MJ. Scott,D.M. Anderson, and DA Neitzel. 1995. A plaeoscience approach to estim~_~g the effectsof climatic sarming on salmonidfisheriesof the ColumbiaRiver basin,p. 48~96. In R.J. Beamish [ed] Climate change andnorthernfish populations. Can. Spec. Publ. Fish. Aquat. Sci. 121. Abstract: Efforts to estimate the effect of climatechange on fisheries are hampered by the lack of models that ~ject realistic JMluatic habitatconditions at the regional scale. Data from the palcosciences area suitable alternative both for environmental scenario development and model validation. We are using a paleoscience approach to calculate the potentialeffect of global warming on anadromous salmonid stocks of the ColumbiaRiver basin,western North America. First, archaeologically dated fluvial sediments and bivalveswere used with terrestrialpaleoecological datato reconstruct the flow, flow patterns, temperatures, and bedconditions6000-7000 yr ago,whenpaleoclimatic indicatorsand atmospheric modelssuggest regionaltemperatures were up to 2°C warmer.Next, these conditionswere imposed on Columbiasystem subbasins andtheir effectson salmon stocks were modeled. Results thus far indicatea 3~OO/o declinein salmon stocks relative to currentconditions. Finally, fish remains from archaeological sites were analyzed for evidence of actualsalmon productionunderthe reconstructed s1ream conditions to assess the validity of model projections. Preliminaryfindings are comparable to model predictions. Resume: Les efforts visant a estimer les effetS du changement climatiquesur les~ches sootentraves par Ie manque de modeles exprimantdesconditions realistes pour les habitats aquatiques a l' echelle regionale. Les donnees despal60sciencesconstituent uneoption valable autant pOur l'elaborationde scenarios environne- mentaux que pour 1a validation desmodeles. Nous utilisons la demarche pal60scientifiq~ pour calculer I'effet eventuel du rechauffement global sur les stocks de salmonides anadromes du bassin du tleuve Colum- bia, dans l'ouest de I' Amerique du Nord. En premierlieu, nous noussommes servis de sediments tluviaux arcMologiquement dates etde bivalvesen plus desdonnees paleoecologiques terrestres, pour reconstituer Ie debit, les configurations d'6coulement, les temperatures et les conditionsdu lit du tleuve it y a 6 ou 7 000 ans, epoque ou les indicateurs paleoclimatiques et les modeles atmosph6rlques permettent de croire que lestemperatures regionales etaientjusqu'a2° C plus chaudes. Par 1a suite,nousavonsapplique ces conditionssur les sous-bassins du bassin du Columbiaet avonsmodelis6 leurs effets sur les stocks de sau- mons.Les resultatsjusqu'a,maintenant foot 6tat d'un d6clin de 30 a 60 % desstocks de saumons par rapport aux conditions actuelles. Entin, nousavons analyse les fossiles de poissons de sites arch60logiques pour trouver des preuves de 1a productionreelle de saumons selonles conditions reconstituees du tleuve en vue d'6valuer la validite desprojectionsfaites avecIe modele. Les constatations preliminaires sontcomparables a cellesqui resultent dumedele. WA 99352, USA. I Praent address: North American PaleoScience, 648 SaiIIt Street, Richland, W A 99352, USA. 2 Present addreSS:University Museum. University of Colorado, Boulder, CO 80309, USA. Scott and D.M. Anderson. Technology Policy Analysis WA, 99352, USA. D.A. Neitzel. Environmental Sciences DeparbDeut. Pacific Northwest Laboratory, P.O. Box 999, Richland, WA 99352, USA.
Transcript of A paleoscience approach to estimating the effects of climatic warming on salmonid fisheries of the...

489
A paleoscience approach to estimating theeffects of climatic warming on salmonidfisheries of the Columbia River basin
James C. Chatters1 and Virginia L. Butler2, Michael J. Scott, David M. Anderson,and Duane A. Neitzel
Chatters. J.C.. V.L. Butler, MJ. Scott, D.M. Anderson, and DA Neitzel. 1995. A plaeoscience approach toestim~_~g the effects of climatic sarming on salmonid fisheries of the Columbia River basin, p. 48~96. InR.J. Beamish [ed] Climate change and northern fish populations. Can. Spec. Publ. Fish. Aquat. Sci. 121.
Abstract: Efforts to estimate the effect of climate change on fisheries are hampered by the lack of modelsthat ~ject realistic JMluatic habitat conditions at the regional scale. Data from the palcosciences are asuitable alternative both for environmental scenario development and model validation. We are using apaleoscience approach to calculate the potential effect of global warming on anadromous salmonid stocksof the Columbia River basin, western North America. First, archaeologically dated fluvial sediments andbivalves were used with terrestrial paleoecological data to reconstruct the flow, flow patterns, temperatures,and bed conditions 6000-7000 yr ago, when paleoclimatic indicators and atmospheric models suggestregional temperatures were up to 2°C warmer. Next, these conditions were imposed on Columbia systemsubbasins and their effects on salmon stocks were modeled. Results thus far indicate a 3~OO/o decline insalmon stocks relative to current conditions. Finally, fish remains from archaeological sites were analyzedfor evidence of actual salmon production under the reconstructed s1ream conditions to assess the validity ofmodel projections. Preliminary findings are comparable to model predictions.
Resume: Les efforts visant a estimer les effetS du changement climatique sur les ~ches soot entraves par Iemanque de modeles exprimant des conditions realistes pour les habitats aquatiques a l' echelle regionale. Lesdonnees des pal60sciencesconstituent une option valable autant pOur l'elaboration de scenarios environne-mentaux que pour 1a validation des modeles. Nous utilisons la demarche pal60scientifiq~ pour calculerI'effet eventuel du rechauffement global sur les stocks de salmonides anadromes du bassin du tleuve Colum-bia, dans l'ouest de I' Amerique du Nord. En premier lieu, nous nous sommes servis de sediments tluviauxarcMologiquement dates etde bivalves en plus des donnees paleoecologiques terrestres, pour reconstituer Iedebit, les configurations d'6coulement, les temperatures et les conditions du lit du tleuve it y a 6 ou7 000 ans, epoque ou les indicateurs paleoclimatiques et les modeles atmosph6rlques permettent de croireque les temperatures regionales etaientjusqu'a 2° C plus chaudes. Par 1a suite, nous avons applique cesconditions sur les sous-bassins du bassin du Columbia et avons modelis6 leurs effets sur les stocks de sau-mons. Les resultatsjusqu'a,maintenant foot 6tat d'un d6clin de 30 a 60 % des stocks de saumons par rapportaux conditions actuelles. Entin, nous avons analyse les fossiles de poissons de sites arch60logiques pourtrouver des preuves de 1a production reelle de saumons selon les conditions reconstituees du tleuve en vued'6valuer la validite des projections faites avec Ie modele. Les constatations preliminaires sont comparablesa celles qui resultent dumedele.
WA 99352,USA.
I Praent address: North American PaleoScience, 648 SaiIIt Street,
Richland, W A 99352, USA.2 Present addreSS: University Museum. University of Colorado, Boulder,
CO 80309, USA.
Scott and D.M. Anderson. Technology Policy Analysis
WA, 99352, USA.
D.A. Neitzel. Environmental Sciences DeparbDeut. Pacific Northwest
Laboratory, P.O. Box 999, Richland, WA 99352, USA.
~
490
Introduction extrapolation from recent climatic variation is uncertain.Currently available GCM-generated climatic scenariosaddress the globe at grid scales that often mask regional-scalevariability; regional-scale models derived from GCMs are stillin the development stage. Therefore, to predict the effects oflarge-scale climate changes on fISheries, it is necessary toassume that great rises in air and sea-surface temperatures, forexample, will produce regional climate changes of the sametype, but simply of greater magnitude than those observedhistorically. This assumption may be invalid; historical recordscontain evidence of climatic fluctuations within one equilibriumstate, but air temperature increases of2-5°Cmay generate newequilibria, with regional effects that are not mere extrapola-tions of what we currently observe. In their present forms, notwo GCMs agree on the regional-scale characteristics offutureclimates (O_Jshm~n and Spring 1989; Kalkstein 1991).
At Pacific Northwest LabOratory, we have been exploringthe use of data from the paleosciences as a solution to thisdilemma. Paleosciences are the disciplines, including geology,paleoecology, paleontology, paleohydrology, paleooceano-graphy, and archaeology, that use the physical traces andremnants of processes and organisms to reconstruct and under-
The standard approach to predicting the effect of climaticchanges on fish populations is a three-part process. Theresearcher begins either by conducting a careful analysis of thebehaviour, habitat needs, and reproductive biology of thetarget organism and the influence of temperature on thesevariables, or by making a series of observations on populationresponse to small-scale climatic variations occurring withinthe present-day equilibrium climate. Observations are thenanalyzed to generate an empirical model of a species' responseto environmental conditions. Finally, the potential effects offuture climates are inferred by driving the empirical modelwith scenarios generated by general circulation models (GCMs)and extrapolating from the effects of observed climaticextremes (see, for example, DeAngeles and Cushman 1990).
This approach produces thought-provoking results, as manyof the papers in this symposium demonstrate. Certainly, thecareful collection of empirical data and the derivation ofempirical models therefrom is indispensable to our under-standing of climatic effe:Cts on an important resource base.However, the plausibility of predictions based on GCMs and
-SON. Polen.
.: .'.. EXI8nt of foI8It ~2700 B.P .
("" ') Extent of for.- 7000-«)00 B.P.
_Mcxi8mfol88;~
\
~ --- ~~.=8
n"
\~ -4rN
'. ~.".Archaeologica1 :
.~.~,.~
-~
~.' Grand
RondeSubbasin
."."':"~ ~-. " ."". ." ,
:",. "
118"W124'W
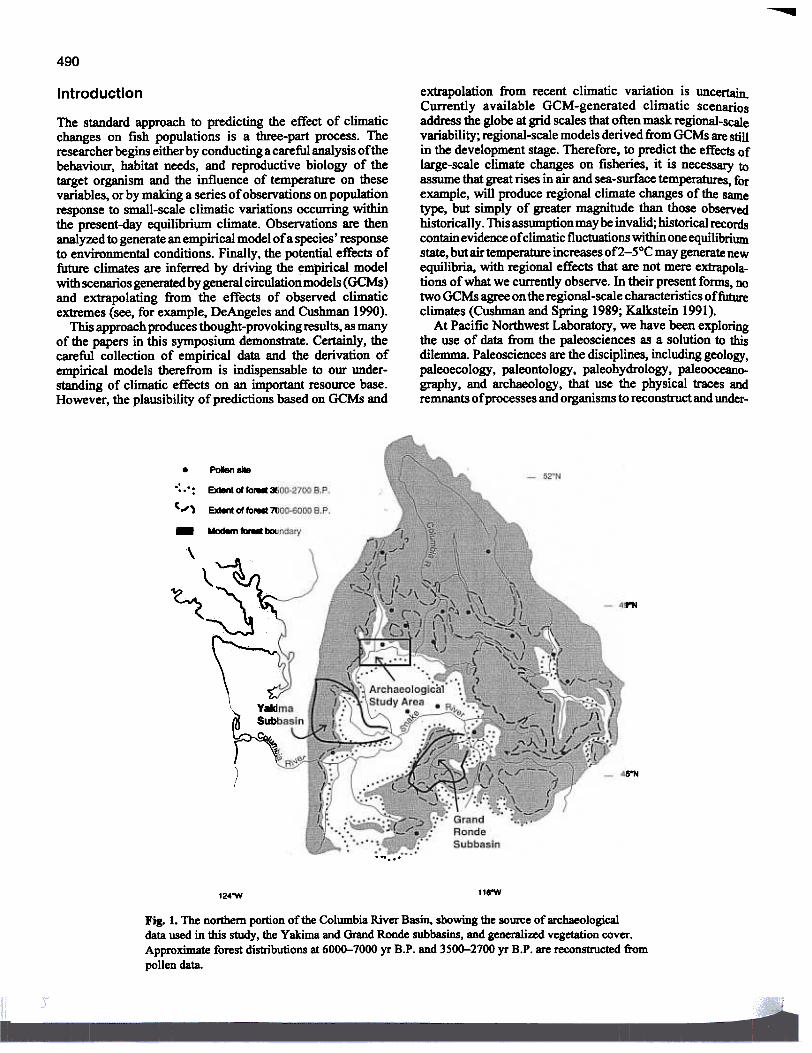
Fig. 1. The northern portion of the Columbia River Basin. showing the source of archaeologicaldata used in this study, the Yakima and Grand Ronde subbasins, and generalized vegetation cover.Approximate forest distnoutions at 6000-7000 yr B.P. and 3500-2700 yr B.P. are reconstructed frompollen data.

491
stand ecosystems and cultures of the past. Such data can beused to generate plausibl, even probable, environmental analogsfor possible future climate states and to validate dynamicenvironmental models (e.g., COHMAP members 1988).
We are applying the paleoscience approach to estimate thepotential effect of climates 2°C warmer than today on theanadromous salmonids of the Columbia River basin, westernNorth America (Chatters et al. 1991; Neitzel et at. 1991) andto validate the model used in ~ analysis. The results areintended to assist the Northwest Power Planning Council(NPPC), Bonneville Power Administration, and otherresource-management agencies that are developing initiativesto enhance salmonid productivity in this basin. The study isalso designed to help the U. S. Department of Energy andthe U. S. Environmental Protection Agency identify theconsequences global wanning might have for water resourcesand national energy policy. This work is still in progress, so theresults reported here are prelin1inary.
The paleoscience approachTo understand how climatic Wamling might affect salmonidpopulations. we are following a four-step procedure. incorpo-rating paleoenvironmental data at the beginning and endingpoints. First. we used geomorphic. paleobotanical. and paleo-malacological data to reconstroct s1ream conditions during thelast 8000 yr. Second. we estimated the effect on salmon(Oncorhynchus) of conditions extant approximately 6000-7000radiocarbon years before present (B.P.). when temperatureswere as much as 2°C warmer than at present (Kutzbach 1983).This became an analog of future warmer climate and its effectson spawning. incubation. and rearing parameters of the NPPC' sTributary Parameter Model (TPM) for estimating salmonidproduction. Third. we ran the TPM in conjunction with theNPPC System Planning Model (SPM) to calculate the effect ofthese analog conditions on the population of returning adultfish in selected stream systems. Ultimately. we will run themodels for all salmon-accessible subbasins of the ColumbiaRiver system. Finally. we are identifying fish remains obtainedfrom archaeological sites along the Columbia River tocompare variations in the taxonomic composition of ancientfish assemblages with model predictions.
proportional to that available for plant growth. Estimatedflows ranged from 30 to 400/0 below modem levels underwarm. arid climates prior to 6000 yr B.P. to as much as 30%above modern levels in the coolest, wettest times (Chatters andHoover 1992).
Composition of the bed and suspended sediment loads ofancient streams were inferred partially from the texture,bedding structure, and rate of build-up of fluvial sediments,but primarily from the taxonomic composition of musselassemblages (Chatters et al. 1991). There are two species oflarge mussels occurring in the flowing water of the U. S. Pacificnorthwest. Margaritiferafalcataprefers swift-flowing, clear,gravel-bedded, stable streams, while Gonidea angulatafavours slower water, higher suspended sediment loads, sandto silt bottoms, and unstable beds (Lyman 1980; Vannote andMinshall 1982). The percent Gonidea in assemblages from atime interval was taken as a crude indicator of the percent sandand silt in the river's bed. Gonidea ranged from zero between300 and 700 yr B.P. to nearly 40% around 6700 yr B.P. (Fig. 2;Chatters 1986).
The annual distribution of flow, or hydrograph, was partiallyreconstructed from the season at death of river mussels fromarchaeological middens. Assuming, as one Native Americanelder stated, that people consumed mussels at any time of year,as long as they were available, and knowing that high streamflows make mussels unavailable as a result of water velocityand turbidity, the period when people were not collectingmussels indicates the period of peak flow. The pre-damColumbia River, which was snowmelt fed, rose in May,peaked in mid-June during the freshet, and fell to near baseflow by mid-July. Season at death for mussels was determinedby measuring annual growth increments along the resilialtuberosity, where the hinge ligament attaches to the shell(Chatters 1987). A growth index was calculated by dividingthe final growth increment by the expected growth for thedeath year. The growth indices were compared to the annualgrowth curve for modern M falcata to estimate the time ofdeath for each sample of mussel shells, or collection event.Finally, collection events were graphed by time period todetermine the timing of the freshet. Results showed thatfreshets typically ended by late June between 7900 and5500 yr B.P ., and ended as late as early August between 3300and 2200 yr B.P. (Fig. 2; Chatters 1986).
Stream temperatures are also derived from increments ofmussel shell growth. Unpublished research by Chatters onmodem M falcata has shown that growth rate is closelycorrelated with water temperatures. Mean growth in theninth and tenth years has a linear relationship with the numberof days above 6, 8, and 10°C, with the strongest correlation(R2= 0.89) at 10°C. Growth rates in mussels from the WellsReservoir area of the Columbia River indicate remarkablechanges in water temperature over the past 8000 yr. Priorto 3900 yr B.P., the water was above 10°C for approximately200 d, but since that time this has dropped to an average oflessthan 130 d (Fig. 2).
As an analog of possible future climate, we chose the periodbetween 6000 and 7000 yr B.P., a time during which bothpaleoscientific indicators (Chatters 1989) and climate models(Kutzbach 1983) indicate temperatures in western NorthAmerica were approximately 2°C warmer than at present.
Reconstructing ancient stream environmentsEstimates of stream flows, annual flow patterns, bed character-istics, and temperatures have been obtained by analyzing pastvegetation patterns, geologic stratigraphy of archaeologicalsites, and archaeological collections of freshwater mussels(Chatters 1989; Chatters and Hoover 1992). Data for thisreconstruction come primarily from the Wells ReservoirArchaeological Project in Okanogan and Douglas Counties,Washington (Chatters 1986). Associated climate scenarios arederived from a synthesis of paleoclimatic indicators, in~ludingpollen sequences, cave and lake sediments, sub fossil wood,and geomorphic evidence of glacial activity (see Chatters andHoover 1992).
The distribution of vegetation. as indicated by studies offossil pollen sequences throughout the river basin (Fig. 1; seeMehringer 1985), was used to estimate mean annual flows;the amO\Dlt of water available for runoff is assumed to be

~492
u00.-
.>0.a..>-.
'a-
.~~-.~GD-
EtI.-
~...~
Aug.
Jul
Jun
Radiocarbon Years B.P.
FiIo 2. A summary of evidence for stream conditions in the Wells Reach of the Columbia River since8000 yr B.P .and their inferred effects on salmon production if they were manifested basinwide.
Based on the four lines of evidence described above, theColumbia River during this period had 30% lower flow,freshets ending nearly a month earlier, temperature above10°C for nearly twice as long, and a bed with nearly 200/0 moresand and silt than found under predam conditions.
may experience ~ predation during a prolonged mi-gration. Differences between present and past flow rates wereused to directly modify smolt capacity, which was asswned tobe proportional to ~ volume (VanHyning 1973; Smith1985). For the details of parameter value changes made in onesimulation, see Chatters et aI. (1991).
Simulating the effects 016000-7000 yr B.P. streamconditionsThus far, we have conducted simulations for two tributaries ofthe Columbia River system: the Yakima subbasin, on the eastflank of the Cascade Mountains of Washington. and the GrandRonde subbasin in mountainous northeastern Oregon (Fig. 1).Simulations estimated the productivity of the spring chinooksalmon (0. 1sbawytscha) under a continuation of currentenvironmental conditions and with the 2°C climate warmingscenario represented by the 6000-7000 yr B.P. analog. Changesto the four TPM parameters differed slightly between basinslargely because of differences in stream gradient and expectedresidence time of smolts. Relative to current conditions,productivity est1~at~ ODder climate change were 600/0 lowerfor the Yakima subbasin (Chatters et aI. 1991) and 300/0 lowerfor the Grand Ronde subbasin (Fig. 3). Although we cannotassert that these two subbasins sample the full range ofvarla-bility in the Columbia River system, they indicate an averagedecline in productivity of 45%.
Alterinl parameters of the TPMThe NPPC system for simulating salmon production uses boththe TPM. which calculates the effects of environmental chang-es on life stages critical to salmon survival in subbasins, andthe SPM, which calculates cumulative salmon swvival overmultiple generations from TPM outputs, plus data on oceanand main-stem river survival (NPPC 1989, 1992). Althoughneither is biologically elegant, both models are readily under-standable and, more importantly, guide the NPPC's salmonenhancement program.
The TPM contains four key parameters: adult prespawningsurvival, egg-to-smolt survival, smolt-to-smolt survival, andsmolt capacity, which are computed for each reach of atributary basin. We used reconstnK:tions of 6000-7000 B.P.conditions in the Wells Reach of the Columbia River to modifyeach of these parameters (Chatters etal. 1991). Water temper-ature was used to estimate changes in adult prespawningsurvival because elevated temperatures increase the incidenceand severity of disease at this life stage (Becker .and Fujihara1987). Texture of the stream bed was used toalteregg-to-smoltsurvival because increased sedimentation can suffocate eggsand displace fry (McNeil and Anhel 1964; Hall and Lantz1969). Freshet timing affects smolt-to-smolt survival byaccelerating or retarding oubnigration to salt water. If thefreshet is too early, smolt may reach salt water before they arephysiologically prepared, or if left behind by the freshet, they
Evaluating the model1ritb &h remains fromarchaeololical sitesHuman inhabitants of the Columbia River system have beenexploiting the fishery for at least 10 000 yr (Cressman 1960;Butler 1993). Presumably. within the limits of available

49310000
rn~ Current
~fJ~~ii~~~~~~~.:.:-:.:.:-:.:.:.:
~:::::::::::::::::i
.:.:.:.:.:.:.:.:.:',.;:1~1~1~1~1~1~1~~1~1~~~1~1~1~~1~1~~1~
':::::::::::::::::::::::::::::::::::::::::-~;::::~:::::::::::::~::::::::::::::~
;!j1111~~~1111~111~11111111111~11111~;
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
:.:.:.:.:.:.:.:.:.:.:.:.:.:.:.:.:.:.:.:.:.,
~:::::::::::::::::::::::::::::::::::::::::~;:::::::::::::::::::::::::::::::::::::::::~
:~1~1~111~1~1~1~1~1~j~1~1~1~1~j~j~j~j~1~~~
~ Climate Change
7500
ID~~
'C<QC'E~~u:~
0~G)
.0E~z
5000
2500
:r;I!!!~:
I!~ltiilti0Yakima Grand Ronde
Subbasin
Fig. 3. Results of model nms under current and climate change analog stream conditions for twoColumbia River system subbasins.
archaeofaunas, in an attempt to generate a history of the fisherythat is chronologically comparable with the reconstroctions ofstream conditions.
This part of our research program is the least developed.Thus far, we have analyzed samples from the Chief JosephDam Archaeological Project (CillAP) (Campbell 1985) andWells Reservoir Archaeological Project (WRAP) (Chatters1986), two of the largest and most detailed archaeological datarecovery efforts yet conducted in the region. Chatters (1986)has identified the fish remains from the WRAP; Butler hasre-analyzed selected archaeofaunas from the CillAP and, tomake the data as comparable as possible, is reanalyzing WRAPassemblages. Comparability is somewhat affected by thecollection procedures used on the two projects. The CillAPused 3.2-mm mesh screen for data recovery and sifted thematrix dry; the WRAP used I.O-mm mesh for 2.5% of thematrix and 6.4-mm mesh for the remainder, all screened usingwater. Quantification of the CillAP data is preliminary; eachsite contained material from multiple habitation events, oftenspanning several thousand years. We have not yet separatedthe archaeofaunas by ~e period, but in each case, over 75%
technology, fish exploitation patterns were consistent withthe expectations of optimal foraging theory (MacArthur andPianka 1966; Winterhalder and Smith 1981). Fish would havebeen taken in numbers proportional to their size and relativeease of capture. Salmon, among the largest and at times mostnumerous fishes in the ecosystem, would have been among thefirst exploited, followed by smaller and/or less numerousor accessible species. Assuming discard of fish remains inhabitation sites or kitchen middens and equivalence in thedurability of the remains of various species, taxonomic abun-dances among fish remains in archaeological bone assemb-lages (archaeofaunas) should reflect availability of variousspecies, particularly the more desirable salmonids. Therefore,archaeofaunas offer data with which to evaluate models of theclimate's effect on salmon productivity.
Archaeological research has been conducted in the ColumbiaRiver Basin for four decades, stimulated largely by hydro-electric projects along the Columbia and Snake rivers. Archae-ofaunas have been collected systematically since the 1960s,providing an extensive data base for model evaluation. Weare assembling and systematically identifying these fish

~
494
of th fish remains come from one time interval. A generalpicbJre of fish use through time can be obtained by assumingthat all fish in a site come from the dominant time interval.
To date, we have results on 13 archaeofaunas from 10 sites,ranging in age from 300 to nearly 8000 yr B.P. Although theoldest archaeofaunas date within the time period of om analogstream environment, we are uncertain how to use them. Before4500 yr B.P., occupants of the region moved frequentlythroughout the year, exploiting foods that were in season ateach locality (Campbell 1985; Chatters 1986). F or this reason,the three archaeofaunas we have from this period may not fullyrepresent the exploitation of Columbia River fish. Analyzedsites that postdate 4500 yr B.P., however, consist almostentirely ofpithouse-containing base camps, which were eitheroccupied year round or were die winter setting for consumptionof foods gathered during all times of year (Lohse 19848, b;Chatters 1986). The remainder appear to have been fishingcamps (Miss 19848, b).
As a preliminary evaluation of the model, we can comparethe fish archaeofaunas from a period of time with streamconditions similar to those of 6000-7000 yr B.P. with thosetimes when stream conditions existed under an essentiallymodem climate. Stream conditions comparable to thoseexisting under the warmer climate persisted until approxi-mately 3900 yr B.P. (Fig. 2). Although the freshet ended atapproximately the same time as it does now, water tempera-tures continued to be warm and the stream bed contained asignifi~t amount of sediment. Conditions for salmonidproduction before 3900 yr B.P. can be characterized as poor,equivalent to the results of climate change simulations for theYakima and Grand Ronde subbasins. The period after 1200 yrB.P., with modem freshet timing, much cooler waters, andvariable amounts of bed sedimentation, is equivalent to thecurrent climate scenario. We characterize these conditionsas good. Between 3400 and 2300 yr B.P., conditions wereoptimum for salmon production, with cool waters, a belatedfreshet, and virtually no bed sedimentation. Data are largely
lacking for the periods between 1200 and yr 2300 yr B.P. andbetween 3400 and 3900 yr B.P., and we make no suggestionsabout their suitability for salmon.
Representatives of six genera have been identified fromthe fish arcbaeofaunas: Oncorhynchus (steelhead. trout, andsalmon), Prosopium (whitefish), Caloslomus (suckers),Ptychocheilus (squawfish), Mylocheilus (peamouth), andAcipenser (sturgeon). For this analysis, we present only thepercentage of Oncorhynchus in each assemblage (fable 1).
Fish archaeofaunas dating between 300 and 4500 yr B.P.closely follow the expected pattern (Fig. 4). Results fromthe CffiAP and WRAP are remarkably similar, despite thedifferences in analysts and data-recovery techniq~. Thepacentage of Oncorhynchus is lowest before 3700 yr B.P.(mean = 23.2%), highest from 3300 to 2200 yr B.P.(mean = 95.5%) and moderate thereafter (mean = 69.8%). Thedifference in Oncorhynchus frequencies between our analogfor current conditions and the warmer climate scenario is66.88/0, which is remarkably close to the 600/0 differencebetween the current and climate change simulations for theYakima subbasin.
Although this analysis is preliminary, it does indicate thatoW" parameter manipulations and the TPM may accuratelysimulate effects of climate change. If anything, our resultsare conservative estimates of climate's effects. However,several nonenvironmental factors could also affect patterns ofOncorhynchus representation. First, differences in bonedensity between Oncorhynchus and other genera may causerelatively accelerated deterioration of salmon bone. Two ofus(V .L. Butler and ] .C. Chatters) are investigating this issue BOOits relationship to taxonomic composition of archaeofaunas.Second, human beings interact selectively with their environ-ment, and may not assemble a representative sample of theavailable biota (Grayson 1981). Optimal-foraging theory andthe general edibility of salmon argue against the importance ofselectivity in this instance, however. Third, technologicaladvances can improve the accessibility of fish species. For
..f
~
'~ij~ I~~ ;
.-~ 1~~: i- !
I.ii
II
!I
Table 1. Data on fish an:baeofaunas from Columbia River sites after 4S00 yr B.P.
Screen siu(mm)
Site Assemblage Age range inradiocarbon yr B.P.
Analyst Percent Number ofQrrcorhyrachw fish bones
Reference forradiocarbon ages
45-00-28545-00-37245-00-2144S-00-3724S-00-37245-00-21145-00-37245-OK-3834S-OK-ll4S-OK-382
290%90 - 340%80
630*701035%65 - 1205%1602220:t:60 - 2400:1:802760%70 - 3000%90
2712%80 - 3117%1192960%170 - 3340:1:803770%80 - 4010%90
3720%80 - 4490:1:110
4O4O:t: II 0
3.26.4,13.2
6.4, 16.4,13.2
6.4,16.4,13.2
6.4,1
V.L. ButlerJ.C. ChattersV.L. ButlerJ .C. Chatters
J . C . Chatters
V.L. ButlerJ.C. ChattersJ . C . Chatters
V.L. Butler
J . C . Chatters
83.661.064.797.19S.093.496.437.0.6.S
26.0.
116 '2510
173 '14131227394 '5679
1537'41
Miss 1984a:112Chatters 1986:89Miss 1984b:l12Chatten 1986: 89Chatters 1986:89Lohse 1984a:26Cbatten 1986:89Chatters 1986:89Lohse 1984b:31SChatters 1986:89
ZoDel~OccupationllZcxa 2, 3 ...
OccupationmOccupation IVZone 4Occupation VOccupation IIKartarOccupation"
8111Cae sites ce oot pidIOUIe occu.-tioDS. M) depositional coIIditiOIIImId peIerV8tion maydiffer from die rest of die 8IDp1es.~ Counts ~ from d1e site IS a whole, which containa material from odIer time periods, but more than 12% of fish elements are from this mile.. Butler bas also identified these collections, obtaining Oncoril}lltC1III.J pen:eDt8geI of 28.0% for 4S-OK-383 aDd 14.3~ for 4S-OK-382.~ This Dumber excl\.Jes vertebrae u.~. spines, n"bs, aDd gill raters.


496
AcknowledgementsWe thank Adeline Fredin of the Colville Confederated Tribes,Charles Robins of the P.U.D. No.1 of Douglas County,Washington, and Lawr Salo of the Seattle District, Corps' ofEngineers, for the opportunity to analyze fish and molluskremains from archaeological collections in their care. Thiswork was supported by the U.S. Department of Energy(Contract DE-AC06- 76RLO 1830) and the Northwest Collegeand University Association for Science (Washington StateUniversity) under contract FGO6-89ER-7SS22 with the U.S.Department of Energy.
I
ReferencesBecker, C.D., and M.P. Fujihara. 1987. The bacterial
pathogen F/exibacter co/umnaris and its epizoologyamong Columbia River fish. Am. Fish. Soc. Monogr. 2.American Fisheries Society, Bethesda, MD.
Butler, V.L. 1993. Natural versus cultural salmonidremains: origin of the Dalles Roadcut Bones, ColumbiaRiver, Oregon, U.S.A. I. Archaeol. Sci. (In press)
Campbell, S.K. 1985. Summary of results, Chiefloseph DamArchaeological Project, Washington. Office of PublicArchaeology, University of Washington, Seattle, WA.
Chatters, I.C. 1986. The Wells Reservoir ArchaeologicalProject. Vol. I. summary of findings. Cent Wash.Archaeol. Surv. Archaeol. Rep. 86-6. CentralWashington University, Ellensburg, WA. '
Chatters I.C. 1987. Shell of Margaritifera margaritifer,afa/cola as a source of paleoenvironmental and culturaldata. Appendix F. In M.S. Kelly, E. Nilsson, andI. Cleland [ed.] Archaeological Investigations at LakeBritton, California. Pacific Gas and Electric Company,San Francisco, CA.
Chatters, I.C. 1989. Hypsithermal paleohydrology as ananalog of greenhouse effects on the Columbia RiverBasin. Washington. Geol. Soc. Am. Abstr. Prog. 21(6).
Chatters, I.C., and K. A. Hoover. 1992. Response of theColumbia River fluvial system to Holocene climaticchange. Quat Res. 37: 42-49.
Chatters, I.C., D.A. Neitzel, MJ. Scott, and S.A. Schankle.1991. The effect of climate change on s1reamenvironments: the salmonid resource of the ColumbiaRiver Basin. Northwest Environ. I. 7: 271-293.
COHMAP Members. 1988. Climatic changes of the last18,000 years: observations and model simulations.Science (Wash., D.C.) 241: 1043-1052.
Cressman, L.S. 1960. Cultural sequences at The Danes,Oregon. Trans. Am. Philos. Soc. 50(10).
Cushman, RM., and P.N. Spring. 1989. Regionaldifferences among GCM climate projections for resourceanalysis. Environ. Manage. 13: 739-795.
DeAngeles, D.L., and RM. CnqhmSin Potential applicationof models in forecasting the effects of climate change onfisheries. Trans. Am. Fish. Soc. 119: 224-239.
Grayson, D.K. 1981. A critical review of the use ofarchaeological vertebrates in paleoenvironmentalreconstruction. I. Ethnobiol. 1: 28-38.
Hall, I. C., and RL. Lantz. 1969. Effects of logging on thehabitat of coho salmon and cutthroat trout in coastalstreams. H. R MacMillan Lectures in Fisheries,University of British Columbia, Vancouver, B.C.
Kalkstein, L.S. [ed.]. 1991. Global comparisons of selectedGCM control nms and observed climate data. Publ. 21P-2202 U. S. Environmental Protection Agency.Washington. DC.
Kutzbach, J.E. 1983. Modeling of Holocene climates.p. 271-277. In H.E. Wright. Jr. [ed.] Late Quaternaryenvironments of the U.S. Vol. 2. The Holocene.University of Minnesota Press. Minneapolis. MN.
Lohse. E.S. 1984a. Archaeological investigations at site45-DO-211. Chief Joseph Dam Project. Washington.Office of Public Archaeology. University of Washington.Seattle. W A.
Lohse. E.S. 1984b. Archaeological investigations at site 45-OK-II. Chief Joseph Dam Project. Washington. Office ofPublic Archaeology. University of Washington. Seattle.WA.
Lyman, R.L. 1980. Freshwater bivalve mollusks in southernPlateau prehistory. Northwest Sci. 54: 121-135.
MacArthur. R.H.. and E.H. Pianka. 1966. On optimal use ofa patchy environment. Am. Nat. 100: 603-609.
McNeil, W.H.. and W.H. Anhel. 1964. Success of pinksalmon spawning relative to size of spawning bedmaterials. Spec. Sci. Rep. Fish. 469. U.S. Fish andWildlife Service. Corvallis. OR.
Mehringer. P.J. 1985. Late Quaternary Pollen Records fromthe Interior Pacific Northwest and Northern Great Basinof the United States. p. 167-189. In V. Bryant andR. Holloway [ed.] Pollen records of late QuaternaryNorth American sediments. American Association ofStratigraphic Palynologists. Dallas. TX.
Miss. C.J. 1984a. Archaeological investigations at site 45-DO-285. Chief Joseph Dam Project. Washington. Officeof Public Archaeology. University of Washington.Seattle. W A.
Miss. C.J. 1984b. Archaeological investigations at site 45-DO-214. Chief Joseph Dam Project. Washington. Officeof Public Archaeology. University of Washington.Seattle. W A.
Neitzel, D.A.. M.J. Scott. SA. Schankle. and J.C. Chatters.1991. The effect of climate change on streamenvironments: the salmonid resource of the ColumbiaRiver Basin. Northwest Environ. J. 7: 271-293.
NorthWest Power Planning Council (NPPC). 1989. Salmonand steelhead system planning documentation. userinformation for the System Planning Model. Monitoringand Evaluation GroUP. Portland. OR.
Northwe:~ Power Planning Council (NPPC). 1992. Salmonand steelhead system planning documentation. 1992update of user information for the System PlanningModel. Monitoring and Evaluation GroUP. Portland. OR.
Smith, R.I.F. 1985. The control offish migration. Springer-Verlag, New York, NY.
VanHyning. J.M.. 1973. Factors affecting the abundance offall chinook salmon in the Columbia River. FishCommission of Oregon. Portland. OR.
Vannote. R.L.. and G. W. Minshall. 1982. Fluvial processesand local lithology controlling abundance. structure. andcomposition of mussel beds. Proc. Natl. Acad. Sci.U.S.A. 79: 4103-4107.
Winterhalder. B.. and EA. slitith. 1981. Hunter-gatherer ;,'foraging strategies. University of Chicago Press. ;jChicago. IL. ~:




















