
A Novel TaqI Polymorphism in the Coding Region of the Ovine TNXB Gene in the MHC Class III Region:...
14
A Novel TaqI Polymorphism in the Coding Region of the Ovine TNXB Gene in the MHC Class III Region: Morphostructural and Physiological Influences Oyeyemi O. Ajayi • Mufliat A. Adefenwa • Brilliant O. Agaviezor • Christian O. N. Ikeobi • Matthew Wheto • Moses Okpeku • Samuel A. Amusan • Abdulmojeed Yakubu • Marcos De Donato • Sunday O. Peters • Ikhide G. Imumorin Received: 13 August 2012 / Accepted: 14 March 2013 Ó Springer Science+Business Media New York 2013 Abstract The tenascin-XB (TNXB) gene has antiadhesive effects, functions in matrix maturation in connective tissues, and localizes to the major histocompatibility complex class III region. We hypothesized that it may influence adaptive physio- logical response through an effect on blood vessel function. We identified a novel g.1324 A?G polymorphism at a TaqI recognition site in a 454 bp fragment of ovine TNXB and genotyped it in 150 Nigerian sheep using PCR-RFLP. The missense Electronic supplementary material The online version of this article (doi:10.1007/s10528-013-9622-9) contains supplementary material, which is available to authorized users. O. O. Ajayi B. O. Agaviezor C. O. N. Ikeobi M. Wheto S. A. Amusan S. O. Peters Department of Animal Breeding and Genetics, University of Agriculture, Abeokuta, Nigeria O. O. Ajayi M. A. Adefenwa B. O. Agaviezor A. Yakubu M. De Donato S. O. Peters I. G. Imumorin (&) Department of Animal Science, Cornell University, 267 Morrison Hall, Ithaca, NY 14853, USA e-mail: [email protected] M. A. Adefenwa Department of Cell Biology and Genetics, University of Lagos, Lagos, Nigeria B. O. Agaviezor Department of Animal Science and Fisheries, University of Port Harcourt, Port-Harcourt, Nigeria M. Okpeku Department of Livestock Production, Niger Delta University, Amassoma, Nigeria A. Yakubu Department of Animal Science, Nasarawa State University, Lafia, Nigeria M. De Donato Department of Biomedicine, Universidad de Oriente, Cumana ´, Venezuela S. O. Peters Department of Animal Science, Berry College, Mount Berry, GA 30149, USA 123 Biochem Genet DOI 10.1007/s10528-013-9622-9
Transcript of A Novel TaqI Polymorphism in the Coding Region of the Ovine TNXB Gene in the MHC Class III Region:...

A Novel TaqI Polymorphism in the Coding Regionof the Ovine TNXB Gene in the MHC Class III Region:Morphostructural and Physiological Influences
Oyeyemi O. Ajayi • Mufliat A. Adefenwa • Brilliant O. Agaviezor •
Christian O. N. Ikeobi • Matthew Wheto • Moses Okpeku •
Samuel A. Amusan • Abdulmojeed Yakubu • Marcos De Donato •
Sunday O. Peters • Ikhide G. Imumorin
Received: 13 August 2012 / Accepted: 14 March 2013
� Springer Science+Business Media New York 2013
Abstract The tenascin-XB (TNXB) gene has antiadhesive effects, functions in
matrix maturation in connective tissues, and localizes to the major histocompatibility
complex class III region. We hypothesized that it may influence adaptive physio-
logical response through an effect on blood vessel function. We identified a novel
g.1324 A?G polymorphism at a TaqI recognition site in a 454 bp fragment of ovine
TNXB and genotyped it in 150 Nigerian sheep using PCR-RFLP. The missense
Electronic supplementary material The online version of this article (doi:10.1007/s10528-013-9622-9)
contains supplementary material, which is available to authorized users.
O. O. Ajayi � B. O. Agaviezor � C. O. N. Ikeobi � M. Wheto � S. A. Amusan � S. O. Peters
Department of Animal Breeding and Genetics, University of Agriculture, Abeokuta, Nigeria
O. O. Ajayi � M. A. Adefenwa � B. O. Agaviezor � A. Yakubu � M. De Donato �S. O. Peters � I. G. Imumorin (&)
Department of Animal Science, Cornell University, 267 Morrison Hall, Ithaca, NY 14853, USA
e-mail: [email protected]
M. A. Adefenwa
Department of Cell Biology and Genetics, University of Lagos, Lagos, Nigeria
B. O. Agaviezor
Department of Animal Science and Fisheries, University of Port Harcourt, Port-Harcourt, Nigeria
M. Okpeku
Department of Livestock Production, Niger Delta University, Amassoma, Nigeria
A. Yakubu
Department of Animal Science, Nasarawa State University, Lafia, Nigeria
M. De Donato
Department of Biomedicine, Universidad de Oriente, Cumana, Venezuela
S. O. Peters
Department of Animal Science, Berry College, Mount Berry, GA 30149, USA
123
Biochem Genet
DOI 10.1007/s10528-013-9622-9

mutation changes glutamic acid (GAA) to glycine (GGA). Among SNP genotypes,
significant differences (P \ 0.05) were observed in body weight and fore cannon bone
length. Interaction effects of breed, SNP genotype, and geographic location had a
significant effect (P \ 0.05) on chest girth. The SNP genotype was significantly
(P \ 0.05) associated with physiological traits of pulse rate and skin temperature. The
observed effect of this novel polymorphism may be mediated through its role in
connective tissue biology, requiring further association and functional studies.
Keywords Tenascin-XB gene � Sheep � TaqI � Nigeria � Morphology �Physiological status
Introduction
Tenascin-XB, encoded by the TNXB gene in the major histocompatibility complex
(MHC) class III region in sheep, is a large extracellular matrix protein that plays a
key role in the deposition of collagen by dermal fibroblasts, with its expression
observed in the dermis of the skin and in the connective tissues of heart and skeletal
muscles (Burch et al. 1997; Matsumoto et al. 1994). Similarly, extracellular matrix
molecules like tenascin-X (TNX) and fibrilin encoded by TNXB and FBN1 genes are
known to be expressed in connective tissues of muscles, tendons, and joint capsules
(Bosman and Stamenkovic 2003). Given these observations, mutations in genes
encoding these extracellular matrix molecules may alter structure and function of
muscle and tendon connective tissues and thus contribute to both muscle weakness
and hypermobility or contractures (Voermans et al. 2009).
Qin et al. (2008) reported that the class III region of the sheep and human MHC
are very similar with respect to length, location, and orientation between the class I
and class II regions and their gene composition. Loci within the class III region are
often highly conserved; in general terms they evolved prior to the class I and II loci
(Kumanovics et al. 2003). TNXB, an important gene in the gene-rich subregion of
the MHC class III of mammals, was discovered because of its 30 overlap with the
human CYP21A2 gene (Morel et al. 1989). Tenascin-X deficiency, in recessive or
dominant form, has been proposed as a cause of hypermobility type and tenascin-X
deficiency type Ehlers-Danlos Syndrome (EDS) in humans (Schalkwijk et al. 2001;
Zweers et al. 2004). Symptoms range from musculoskeletal pain to easy bruising
and velvety skin (Beighton et al. 1998) in hypermobility type and muscle weakness,
skin hyperextensibility, velvety skin, and easy bruising (Voermans et al. 2007) in
tenascin-X deficiency type EDS.
Although tenascin-X protein is abundantly expressed in almost all the connective
tissues (Zweers et al. 2003), it is not expressed in lymphoid organs. Therefore, the
apparently incongruous location of TNXB within the MHC class III region prompted us
to hypothesize its association with morphostructural and physiological indices because
any of the MHC class III genes could contain sequences that affect productivity either
alone or in linkage disequilibrium with other disadvantageous alleles in other genes
(Walsh et al. 2003). In this study we identified a polymorphism defined by a TaqI
restriction site in the coding region represented by a 454 bp fragment of the ovine TNXB
Biochem Genet
123
gene in Nigerian sheep. This was genotyped in 150 animals and analyzed for association
of genotypes at this locus with morphostructural and physiological indices.
Materials and Methods
Animals and DNA Samples
Samples were collected from 150 randomly selected, sexually mature sheep, of
approximately the same age ([15.5 months), representing the four major breeds of
sheep in Nigeria: 40 Uda sheep, 40 Yankasa, 40 Balami, and 30 West African
Dwarf. The sheep were sampled from farms and markets across Nigeria, in the
states of Bauchi and Borno in the northeast, Kano and Sokoto in the northwest,
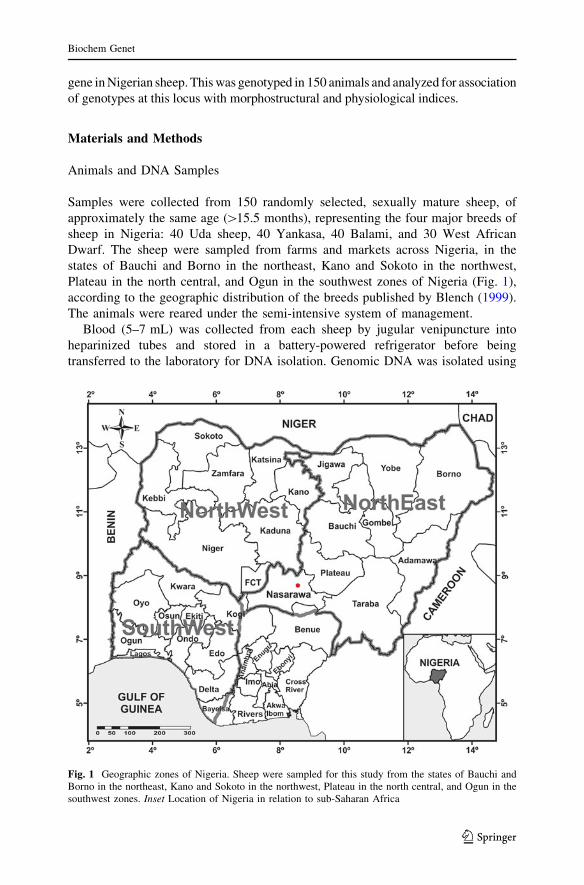
Plateau in the north central, and Ogun in the southwest zones of Nigeria (Fig. 1),
according to the geographic distribution of the breeds published by Blench (1999).
The animals were reared under the semi-intensive system of management.
Blood (5–7 mL) was collected from each sheep by jugular venipuncture into
heparinized tubes and stored in a battery-powered refrigerator before being
transferred to the laboratory for DNA isolation. Genomic DNA was isolated using
Fig. 1 Geographic zones of Nigeria. Sheep were sampled for this study from the states of Bauchi andBorno in the northeast, Kano and Sokoto in the northwest, Plateau in the north central, and Ogun in thesouthwest zones. Inset Location of Nigeria in relation to sub-Saharan Africa
Biochem Genet
123

the ZymoBead Genomic DNA kit (Zymo Research Corp., Irvine, CA, USA). DNA
yield was quantified and its quality assessed using a Nanodrop ND-100 UV/Vis
Spectrophotometer (Nanodrop Technologies, DE, USA).
PCR Amplification, Sequencing, and Multiple Sequence Alignment
The ovine TNXB gene was amplified using forward 50-TCACCACAACAAAGAT-30
and reverse 50-GGACCTTGAAGGAGTCAAAT-30 primers (Qin et al. 2008) to
generate a fragment of 454 bp. PCR amplifications were carried out in a Techne
thermal cycler in a total reaction volume of 20 lL, containing an average of 20 ng
DNA and 10 pmol each primer, in AccuPower PCR Premix (Bioneer Corp., Alameda,
CA, USA). The PCR cycling conditions were denaturation at 95�C for 4 min; followed
by 35 amplification cycles of denaturation at 94�C for 30 s, annealing at 60�C for
1 min, and extension at 72�C for 3 min; and a final extension at 72�C for 10 min. PCR
products were detected on 1.5% agarose gel including ethidium bromide and
photographed under UV light. The amplified fragments were sequenced using the
same PCR primers with the Applied Biosystems Automated 3730 DNA Analyzer, Big
Dye Terminator chemistry, and AmpliTaq-FS DNA polymerase.
The sequences obtained were verified by comparing them to published sequences
of sheep and other species in Genbank. Multiple sequence alignment of deduced
protein sequences from humans, mice, and cows was accomplished using multiple
sequence comparison by log-expectation (Muscle) software (http://www.ebi.ac.uk/
Tools/msa/muscle/), which generally achieves better average accuracy and better
speed than ClustalW2 or T-Coffee, depending on the chosen options.
PCR-RFLP Genotyping of Sheep TNXB
To identify the SNP in the TNXB gene, we aligned the DNA sequences derived from
nine animals using the ClustalW program (http://www.ebi.ac.uk/Tools/clustalw/).
The nonsynonymous SNP in codon 106 was detected. This SNP was analyzed by PCR-
RFLP in 150 sheep. TaqI (Fermentas Life Sciences, Glen Burnie, MD, USA), which
recognizes the palindromic tetranucleotide sequence ;TCGA, was used for restriction
enzyme digestion according to NEB cutter version 2.0 (Vincze et al. 2003). About 10
lL of each PCR product, 2 lL 109 fast digest green buffer, 1 lL TaqI fast digest
enzyme, and 17 lL nuclease-free water were added in this order in a PCR tube and
incubated inside a heat block for 10 min. The restriction fragments were separated by
electrophoresis in 2.5% agarose gel dissolved in 19 TBE and stained with 0.5 lg/mL
ethidium bromide. Genotypes were scored from restriction fragments.
Morphostructural and Physiological Measurements
All measurements were made on each animal within a 2-week sampling period when
weather conditions were similar across the geographic regions (variation in environ-
mental conditions was ± 5%). Each animal was measured for body weight (kg) and
eight linear body measurements (cm), following standard procedures and anatomical
reference points (Yakubu et al. 2010). Height at withers was taken as the distance from
Biochem Genet
123

the surface of the ground to the withers; rump height was measured vertically from the
ground to the top of the pelvic girdle; body length was from the tip of the mouth to the
pin bone; ear length was from the base of the ear to its tip; fore cannon bone length
measured the lower part of the leg extending from the hock to the fetlock; chest girth
was the body circumference just behind the forelegs; chest depth was the dorsal–ventral
distance between the most dorsal point of the withers and the ventral surface of the
sternum; and rump width was the distance between the two tuber coxae. Rectal
temperature was taken on each animal using a digital thermometer inserted into the
rectum at the display of the lowest readable temperature (�C) and removed at the sound
of the alarm signal. Respiratory rate was determined by counting the number of
movements of the abdomen per minute using a stop watch. Pulse rate was determined
by placing a stethoscope just below the chest region and counting the number of pulses
per minute using a stop watch. The heat stress index was derived from the relationship
between pulse rate and respiratory rate together with their normal average values using
the formula of Oladimeji et al. (1993), H = (AR/AP) 9 (NP/NR), where H is the heat
stress index, AR is the average respiratory rate value, AP is average pulse rate value,
NP is normal pulse rate value, and NR is normal respiratory rate.
Statistical Analysis
Morphological and physiological trait data were analyzed using the generalized
linear model in the SAS (2009) software package. In the linear model,
Yijkl ¼ lþ Si þ Aj þ Bk þ Ll þ ðSAÞij þ ðSLÞil þ ðABÞjk þ ðALÞjl þ ðBLÞkl
þ ðSABÞijk þ ðALSÞjli þ ðBLSÞkli þ eijkl;
Yijkl is the trait of interest, l is the overall mean for the trait of interest, Si is SNP genotype
(i = 1–3), Aj is fixed effect of jth breed (j = 1–4), Bk is fixed effect of kth sex (k = 1–2),
Ll is fixed effect of lth geographic location (l = 1–4), (SA)ij is interaction effect of SNP
genotype and jth breed, (SL)il is interaction effect of SNP genotype and lth geographic
location, (AB)jk is interaction effect of jth breed and kth sex, (AL)jl is interaction effect of
jth breed and lth geographic location, (BL)kl is interaction effect of kth sex and lth
geographic location, (SAB)ijk is interaction effect of SNP genotype with breed and sex,
(ALS)jli is interaction effect of breed with geographic location and SNP genotype,
(BLS)kli is interaction effect of sex with geographic location and SNP genotype, and eijkl
is the random error associated with each record (normally, independently, and identically
distributed with zero mean and constant variance). Significant means were separated
using Duncan’s multiple range test procedure from the same statistical software. Inter-
actions that were not significant were removed from the model. Correlations were also
computed to ascertain relationships among measurable traits.
Results
The TNXB gene fragment was successfully amplified with primers designed by Qin
et al. (2008) to produce a DNA fragment of 456 bp after amplicon sequencing. The
Biochem Genet
123

sequences have been submitted to Genbank (awaiting accession numbers). The
alignment of sequences led to the discovery of the g.1324 A?G mutation (Genbank
accession no. EF 197845.1) in the coding region. The mutation was a transition at a
TaqI restriction enzyme site. Blastn analysis of the TNXB sequence in Nigerian
sheep revealed 99% nucleotide identity with Ovis aries (EF 197845.1). Prediction of
the amino acid sequence from the TNXB sequence in Nigerian sheep showed 90%
similarity with that of Bos taurus (acc. no. NP_777128.1), and this led to the
discovery of a p.E2000G missense mutation resulting in a change from glutamic
acid (GAA) to glycine (GGA). Enzymatic digestion of PCR products at the TaqI
polymorphic site showed AA homozygotes with two digested fragments of 316 and
140 bp, GG homozygotes with an undigested fragment of 456 bp, and AG
heterozygotes with all three bands.
The observed polymorphic site corresponded to lysine from the complete
alignment by Muscle of the human, mouse, and bovine protein sequences. Tracing
this position, it was observed that in humans and sheep, glutamic acid (E) was
present, and that position corresponded to amino acids K in cows and W in mice,
proving that the amino acid position is not conserved.
Effects of SNP Genotypes on Morphology
The highest frequency of the AA genotype (0.5) was found in Uda sheep, West
African Dwarf sheep had the highest AG frequency (0.6), and Uda sheep also had
the highest GG frequency (0.15). The highest frequency of the A allele was found in
Uda sheep, and the lowest was found in West African Dwarf sheep. The frequencies
of the A allele in the Yankasa and Balami breeds were intermediate between these
two extremes. AG was the most frequent genotype in the breeds studied, except for
the Uda breed. Chi-square values observed to be significant among the four breeds
were suggestive of Hardy–Weinberg equilibrium (Table 1).
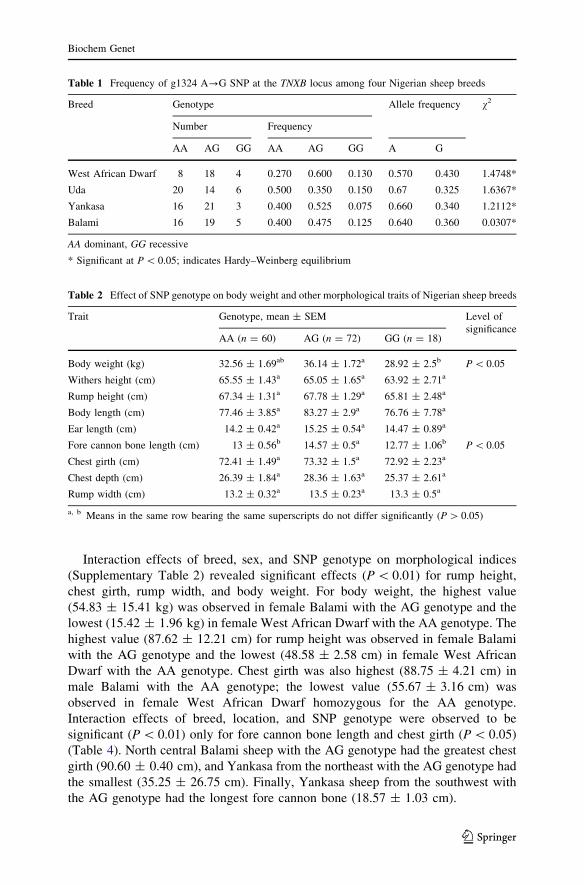
The SNP genotype had significant effects (P \ 0.05) on body weight and fore
cannon bone length, with the highest values observed in the AG genotype
(36.14 ± 1.72 kg and 14.57 ± 0.54 cm, respectively). No significant differences
(P [ 0.05) were observed among the SNP genotypes for the remaining morpho-
logical traits examined (Table 2). For all the morphological indices, the highest
values were observed in the AG genotype. The lowest values were observed in the
GG genotype, except for rump width and ear length. The differences observed
among the AA, AG, and GG genotypes were not significant, however, except for
body weight and fore cannon.
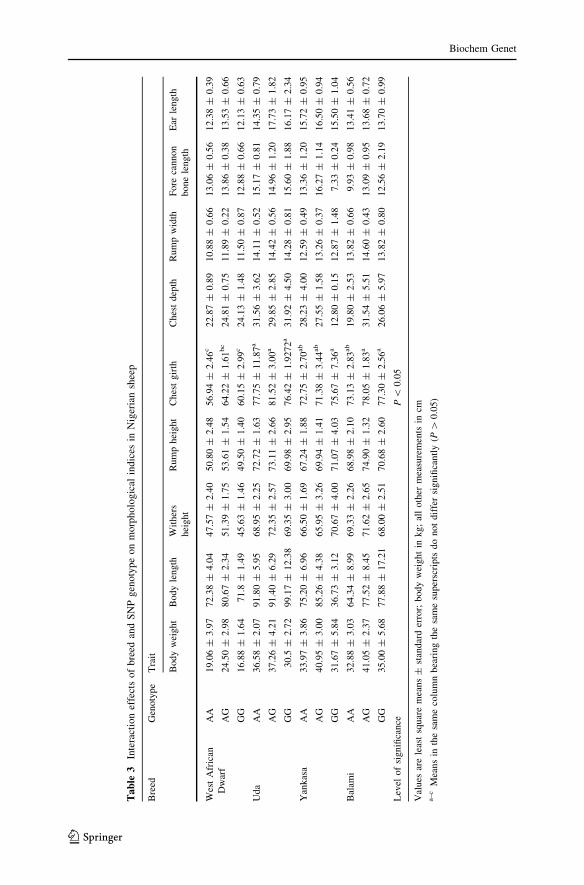
The interaction effects of breed and SNP genotype (Table 3) did not significantly
affect morphological indices, except for chest girth. The highest value for chest
girth (81.52 ± 3.00 cm) was observed in Uda with the genotype AG, and the lowest
value (56.94 ± 2.46 cm) was seen in West African sheep homozygous for A. A
significant effect (P \ 0.01) on chest girth was also observed in the interaction
between location and SNP genotype (Supplementary Table 1), with the highest
value (81.80 ± 2.75 cm) observed in sheep from the north central region with the
AG genotype; the lowest value (60.15 ± 2.99 cm) was observed in sheep from the
southwest with the GG genotype.
Biochem Genet
123
Interaction effects of breed, sex, and SNP genotype on morphological indices
(Supplementary Table 2) revealed significant effects (P \ 0.01) for rump height,
chest girth, rump width, and body weight. For body weight, the highest value
(54.83 ± 15.41 kg) was observed in female Balami with the AG genotype and the
lowest (15.42 ± 1.96 kg) in female West African Dwarf with the AA genotype. The
highest value (87.62 ± 12.21 cm) for rump height was observed in female Balami
with the AG genotype and the lowest (48.58 ± 2.58 cm) in female West African
Dwarf with the AA genotype. Chest girth was also highest (88.75 ± 4.21 cm) in
male Balami with the AA genotype; the lowest value (55.67 ± 3.16 cm) was
observed in female West African Dwarf homozygous for the AA genotype.
Interaction effects of breed, location, and SNP genotype were observed to be
significant (P \ 0.01) only for fore cannon bone length and chest girth (P \ 0.05)
(Table 4). North central Balami sheep with the AG genotype had the greatest chest
girth (90.60 ± 0.40 cm), and Yankasa from the northeast with the AG genotype had
the smallest (35.25 ± 26.75 cm). Finally, Yankasa sheep from the southwest with
the AG genotype had the longest fore cannon bone (18.57 ± 1.03 cm).
Table 1 Frequency of g1324 A?G SNP at the TNXB locus among four Nigerian sheep breeds
Breed Genotype Allele frequency v2
Number Frequency
AA AG GG AA AG GG A G
West African Dwarf 8 18 4 0.270 0.600 0.130 0.570 0.430 1.4748*
Uda 20 14 6 0.500 0.350 0.150 0.67 0.325 1.6367*
Yankasa 16 21 3 0.400 0.525 0.075 0.660 0.340 1.2112*
Balami 16 19 5 0.400 0.475 0.125 0.640 0.360 0.0307*
AA dominant, GG recessive
* Significant at P \ 0.05; indicates Hardy–Weinberg equilibrium
Table 2 Effect of SNP genotype on body weight and other morphological traits of Nigerian sheep breeds
Trait Genotype, mean ± SEM Level of
significanceAA (n = 60) AG (n = 72) GG (n = 18)
Body weight (kg) 32.56 ± 1.69ab 36.14 ± 1.72a 28.92 ± 2.5b P \ 0.05
Withers height (cm) 65.55 ± 1.43a 65.05 ± 1.65a 63.92 ± 2.71a
Rump height (cm) 67.34 ± 1.31a 67.78 ± 1.29a 65.81 ± 2.48a
Body length (cm) 77.46 ± 3.85a 83.27 ± 2.9a 76.76 ± 7.78a
Ear length (cm) 14.2 ± 0.42a 15.25 ± 0.54a 14.47 ± 0.89a
Fore cannon bone length (cm) 13 ± 0.56b 14.57 ± 0.5a 12.77 ± 1.06b P \ 0.05
Chest girth (cm) 72.41 ± 1.49a 73.32 ± 1.5a 72.92 ± 2.23a
Chest depth (cm) 26.39 ± 1.84a 28.36 ± 1.63a 25.37 ± 2.61a
Rump width (cm) 13.2 ± 0.32a 13.5 ± 0.23a 13.3 ± 0.5a
a, b Means in the same row bearing the same superscripts do not differ significantly (P [ 0.05)
Biochem Genet
123
Ta
ble
3In
tera
ctio
nef
fect
so
fbre
edan
dS
NP
gen
oty
pe
on
morp
holo
gic
alin
dic
esin
Nig
eria
nsh
eep
Bre
edG
enoty
pe
Tra
it
Body
wei
ght
Body
length
Wit
her
s
hei
ght
Rum
phei
ght
Ches
tgir
thC
hes
tdep
thR
um
pw
idth
Fore
cannon
bone
length
Ear
length
Wes
tA
fric
an
Dw
arf
AA
19.0
6±
3.9
772.3
8±
4.0
447.5
7±
2.4
050.8
0±
2.4
856.9
4±
2.4
6c
22.8
7±
0.8
910.8
8±
0.6
613.0
6±
0.5
612.3
8±
0.3
9
AG
24.5
0±
2.9
880.6
7±
2.3
451.3
9±
1.7
553.6
1±
1.5
464.2
2±
1.6
1bc
24.8
1±
0.7
511.8
9±
0.2
213.8
6±
0.3
813.5
3±
0.6
6
GG
16.8
8±
1.6
471.8
±1.4
945.6
3±
1.4
649.5
0±
1.4
060.1
5±
2.9
9c
24.1
3±
1.4
811.5
0±
0.8
712.8
8±
0.6
612.1
3±
0.6
3
Uda
AA
36.5
8±
2.0
791.8
0±
5.9
568.9
5±
2.2
572.7
2±
1.6
377.7
5±
11.8
7a
31.5
6±
3.6
214.1
1±
0.5
215.1
7±
0.8
114.3
5±
0.7
9
AG
37.2
6±
4.2
191.4
0±
6.2
972.3
5±
2.5
773.1
1±
2.6
681.5
2±
3.0
0a
29.8
5±
2.8
514.4
2±
0.5
614.9
6±
1.2
017.7
3±
1.8
2
GG
30.5
±2.7
299.1
7±
12.3
869.3
5±
3.0
069.9
8±
2.9
576.4
2±
1.9
272
a31.9
2±
4.5
014.2
8±
0.8
115.6
0±
1.8
816.1
7±
2.3
4
Yan
kas
aA
A33.9
7±
3.8
675.2
0±
6.9
666.5
0±
1.6
967.2
4±
1.8
872.7
5±
2.7
0ab
28.2
3±
4.0
012.5
9±
0.4
913.3
6±
1.2
015.7
2±
0.9
5
AG
40.9
5±
3.0
085.2
6±
4.3
865.9
5±
3.2
669.9
4±
1.4
171.3
8±
3.4
4ab
27.5
5±
1.5
813.2
6±
0.3
716.2
7±
1.1
416.5
0±
0.9
4
GG
31.6
7±
5.8
436.7
3±
3.1
270.6
7±
4.0
071.0
7±
4.0
375.6
7±
7.3
6a
12.8
0±
0.1
512.8
7±
1.4
87.3
3±
0.2
415.5
0±
1.0
4
Bal
ami
AA
32.8
8±
3.0
364.3
4±
8.9
969.3
3±
2.2
668.9
8±
2.1
073.1
3±
2.8
3ab
19.8
0±
2.5
313.8
2±
0.6
69.9
3±
0.9
813.4
1±
0.5
6
AG
41.0
5±
2.3
777.5
2±
8.4
571.6
2±
2.6
574.9
0±
1.3
278.0
5±
1.8
3a
31.5
4±
5.5
114.6
0±
0.4
313.0
9±
0.9
513.6
8±
0.7
2
GG
35.0
0±
5.6
877.8
8±
17.2
168.0
0±
2.5
170.6
8±
2.6
077.3
0±
2.5
6a
26.0
6±
5.9
713.8
2±
0.8
012.5
6±
2.1
913.7
0±
0.9
9
Lev
elof
signifi
cance
P\
0.0
5
Val
ues
are
leas
tsq
uar
em
eans
±st
andar
der
ror;
body
wei
ght
inkg;
all
oth
erm
easu
rem
ents
incm
a–c
Mea
ns
inth
esa
me
colu
mn
bea
ring
the
sam
esu
per
scri
pts
do
not
dif
fer
signifi
cantl
y(P
[0.0
5)
Biochem Genet
123

Ta
ble
4In
tera
ctio
nef
fect
so
fb
reed
,lo
cati
on
,an
dS
NP
gen
oty
pe
on
mo
rph
olo
gic
alin
dic
esin
Nig
eria
nsh
eep
Bre
edL
oca
-
tion
Gen
o-
type
Tra
it
Body
wei
ght
Body
length
Wit
her
s
hei
ght
Rum
p
hei
ght
Ches
t
gir
th
Ches
tdep
thR
um
pw
idth
Fore
cannon
bone
length
Ear
length
Yan
kas
aN
CA
A30.2
8±
4.2
766.8
0±
9.9
765.2
2±
2.3
164.7
6±
2.7
370.1
6±
4.1
1ab
cd
e21.0
7±
2.9
311.9
9±
0.6
610.4
4±
1.4
6d
efg
h15.3
8±
1.4
7
AG
32.4
0±
3.5
368.8
0±
12.9
868.7
8±
2.7
570.6
6±
3.2
977.8
0±
1.9
1ab
cd
e21.8
0±
4.0
014.2
6±
0.4
012.5
6±
2.1
2b
cd
efg
h17.1
0±
2.1
1
GG
31.6
7±
5.8
436.7
3±
3.1
270.6
7±
4.0
071.0
7±
4.0
375.6
7±
7.3
6ab
cd
e12.8
0±
0.1
512.8
7±
1.4
87.3
3±
0.2
4g
h15.5
0±
1.0
4
SW
AA
44.7
5±
11.4
392.0
0±
2.6
870.7
5±
3.0
969.5
0±
3.1
074.6
3±
4.8
6ab
cd
e31.0
0±
0.9
113.5
0±
1.1
917.6
3±
0.8
0ab
c18.0
0±
1.2
9
AG
45.0
0±
3.6
794.4
3±
1.6
265.7
5±
4.8
070.1
8±
1.6
774.2
5±
1.7
2ab
cd
e30.8
9±
0.9
313.0
0±
0.4
318.5
7±
1.0
3a
17.2
1±
0.8
8
Bal
ami
NE
AA
32.8
0±
3.6
834.3
0±
3.3
865.9
2±
4.8
269.6
2±
2.5
373.7
0±
3.8
5ab
cd
e11.5
6±
0.9
013.9
8±
0.8
56.8
0±
0.3
7h
14.7
±0.5
8
AG
33.6
4±
2.9
956.3
5±
8.8
066.3
3±
3.9
172.5
4±
1.6
974.9
3±
2.4
7ab
cd
e31.0
6±
9.6
113.8
5±
0.4
811.7
3±
1.3
2cd
efg
h14.0
5±
0.9
0
GG
26.5
0±
6.5
037.7
5±
4.7
565.5
0±
2.5
067.2
0±
3.2
073.5
0±
2.5
0ab
cd
e11.7
5±
0.7
512.4
5±
1.7
57.2
5±
0.2
5g
h15.7
5±
0.7
5
NC
AA
29.6
7±
7.9
654.4
0±
11.3
670.9
0±
8.2
564.8
7±
5.3
969.0
0±
9.0
7b
cd
e16.2
3±
3.3
013.8
7±
1.8
39.5
3±
2.3
8efg
h14.3
3±
2.0
3
AG
40.5
0±
1.5
042.7
5±
0.2
576.6
0±
0.6
078.6
0±
3.6
090.6
0±
0.4
0a
14.3
0±
0.8
015.1
0±
1.7
07.3
0±
0.1
0g
h12.2
0±
0.2
0
NW
AA
36.4
3±
5.6
394.3
6±
12.3
272.0
0±
2.1
971.2
9±
3.8
376.0
7±
4.6
1ab
cd
e28.7
0±
3.1
914.2
0±
1.1
512.8
9±
1.2
7b
cd
efg
11.8
6±
0.5
5
AG
77.1
7±
28.9
117.8
3±
3.4
084.5
0±
5.8
8101.0
±24.1
778.2
5±
2.8
6ab
cd
e35.0
0±
1.1
815.5
0±
1.0
515.3
3±
0.7
1ab
cd
ef
12.6
7±
1.1
5
GG
47.5
±4.5
0113.5
±4.5
073.5
0±
1.5
076.5
0±
0.5
083.2
5±
0.2
5ab
c36.5
0±
3.5
015.0
0±
0.0
016.0
0±
1.0
0ab
cd
e11.5
±0.5
0
Lev
elof
signifi
cance
P\
0.0
5P
\0.0
1
Val
ues
are
leas
tsq
uar
em
eans
±st
andar
der
ror;
body
wei
ght
inkg;
all
oth
erm
easu
rem
ents
incm
a–
hM
eans
inth
esa
me
colu
mn
bea
ring
the
sam
esu
per
scri
pts
do
not
dif
fer
signifi
cantl
y(P
[0.0
5)
Biochem Genet
123
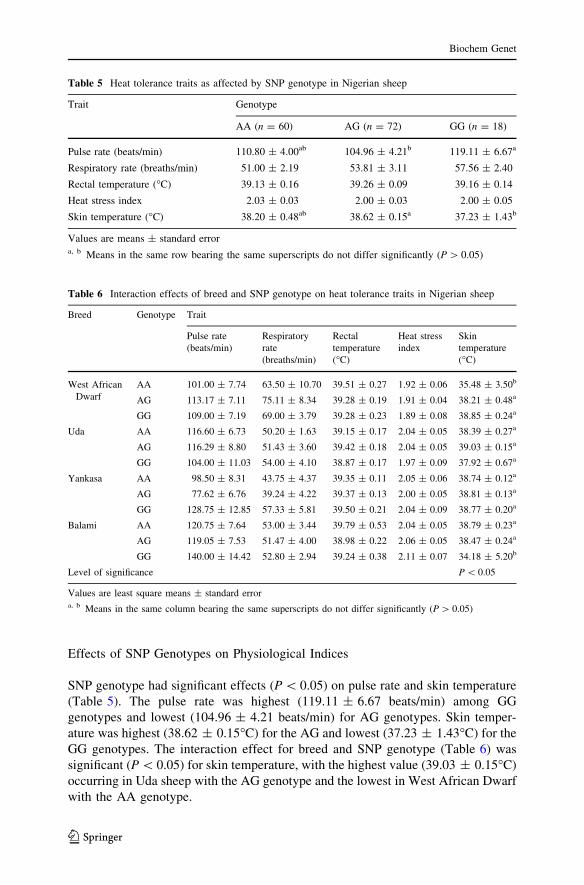
Effects of SNP Genotypes on Physiological Indices
SNP genotype had significant effects (P \ 0.05) on pulse rate and skin temperature
(Table 5). The pulse rate was highest (119.11 ± 6.67 beats/min) among GG
genotypes and lowest (104.96 ± 4.21 beats/min) for AG genotypes. Skin temper-
ature was highest (38.62 ± 0.15�C) for the AG and lowest (37.23 ± 1.43�C) for the
GG genotypes. The interaction effect for breed and SNP genotype (Table 6) was
significant (P \ 0.05) for skin temperature, with the highest value (39.03 ± 0.15�C)
occurring in Uda sheep with the AG genotype and the lowest in West African Dwarf
with the AA genotype.
Table 5 Heat tolerance traits as affected by SNP genotype in Nigerian sheep
Trait Genotype
AA (n = 60) AG (n = 72) GG (n = 18)
Pulse rate (beats/min) 110.80 ± 4.00ab 104.96 ± 4.21b 119.11 ± 6.67a
Respiratory rate (breaths/min) 51.00 ± 2.19 53.81 ± 3.11 57.56 ± 2.40
Rectal temperature (�C) 39.13 ± 0.16 39.26 ± 0.09 39.16 ± 0.14
Heat stress index 2.03 ± 0.03 2.00 ± 0.03 2.00 ± 0.05
Skin temperature (�C) 38.20 ± 0.48ab 38.62 ± 0.15a 37.23 ± 1.43b
Values are means ± standard errora, b Means in the same row bearing the same superscripts do not differ significantly (P [ 0.05)
Table 6 Interaction effects of breed and SNP genotype on heat tolerance traits in Nigerian sheep
Breed Genotype Trait
Pulse rate
(beats/min)
Respiratory
rate
(breaths/min)
Rectal
temperature
(�C)
Heat stress
index
Skin
temperature
(�C)
West African
Dwarf
AA 101.00 ± 7.74 63.50 ± 10.70 39.51 ± 0.27 1.92 ± 0.06 35.48 ± 3.50b
AG 113.17 ± 7.11 75.11 ± 8.34 39.28 ± 0.19 1.91 ± 0.04 38.21 ± 0.48a
GG 109.00 ± 7.19 69.00 ± 3.79 39.28 ± 0.23 1.89 ± 0.08 38.85 ± 0.24a
Uda AA 116.60 ± 6.73 50.20 ± 1.63 39.15 ± 0.17 2.04 ± 0.05 38.39 ± 0.27a
AG 116.29 ± 8.80 51.43 ± 3.60 39.42 ± 0.18 2.04 ± 0.05 39.03 ± 0.15a
GG 104.00 ± 11.03 54.00 ± 4.10 38.87 ± 0.17 1.97 ± 0.09 37.92 ± 0.67a
Yankasa AA 98.50 ± 8.31 43.75 ± 4.37 39.35 ± 0.11 2.05 ± 0.06 38.74 ± 0.12a
AG 77.62 ± 6.76 39.24 ± 4.22 39.37 ± 0.13 2.00 ± 0.05 38.81 ± 0.13a
GG 128.75 ± 12.85 57.33 ± 5.81 39.50 ± 0.21 2.04 ± 0.09 38.77 ± 0.20a
Balami AA 120.75 ± 7.64 53.00 ± 3.44 39.79 ± 0.53 2.04 ± 0.05 38.79 ± 0.23a
AG 119.05 ± 7.53 51.47 ± 4.00 38.98 ± 0.22 2.06 ± 0.05 38.47 ± 0.24a
GG 140.00 ± 14.42 52.80 ± 2.94 39.24 ± 0.38 2.11 ± 0.07 34.18 ± 5.20b
Level of significance P \ 0.05
Values are least square means ± standard errora, b Means in the same column bearing the same superscripts do not differ significantly (P [ 0.05)
Biochem Genet
123

Discussion
Genetic variability within the MHC region has been discovered to influence
resistance or susceptibility to diseases and productivity of sheep (Larruskain et al.
2012; Hui et al. 2012; Hickford et al. 2011; Castillo et al. 2011; Keane et al. 2007;
Geldermann et al. 2006). In the present study, the g.1324G?A mutation resulted in
an amino acid change from hydrophilic glutamic acid to hydrophobic glycine,
which may plausibly alter the structure of the expressed tenascin-XB protein.
Qin et al. (2008) suggested that despite structural similarity between the human and
sheep MHC, it is likely that further studies would show species-specific differences
between the class III members of the sheep MHC and those of other species. Since this
is the first study of a mutation in this gene associated with morphostructural and
physiological indices, it is difficult to compare our results with similar studies. It is
interesting to note that significant differences in morphological traits for the AG
genotype indicated heterozygous superiority similar to the overdominant effect
inferred from studies involving MHC genes in mice coinfected with Salmonella and
Theiler’s murine encephalomyelitis virus (McClelland et al. 2003).
The prominent expression of tenascin-X protein as revealed by intense staining in
the muscle (Matsumoto et al. 1994) lends credence to its ubiquitous expression in
collagen, which is the most abundant protein in vertebrates. Muscle cells not only
play a great role in the body’s internal life, helping other systems to function, but
also may likely affect body weight, possibly through collagen deposits. Mutations in
the TNXB gene have been associated with reduced deposition of collagen (Egging
et al. 2006) and muscle weakness (Voermans et al. 2009). The mutant genotype
(GG) observed in Nigerian sheep may plausibly contribute to the inability of
individuals of that genotype to cover large distances in search of good-quality
forage, possibly due to musculoskeletal pain, a situation decidedly exacerbated by
the extensive or semi-intensive system of management practiced by most local
farmers in sub-Saharan Africa. This may be supported by observations that tenascin-
X interacts with types I, III, and V fibrillar collagen molecules when in native
conformation, suggesting that tenascin-X, via trimerization and multiple interac-
tions with components of collagenous fibrils, plays a crucial role in the organization
of extracellular matrices (Lethias et al. 2006). Although the exact mechanism of
how TNXB contributes to variations in body weight remains obscure, its effect in
regulating the structure and mechanical properties of connective tissues has been
well documented (Elefteriou et al. 1997, 1999; Lethias et al. 2006; Egging et al.
2007; Jollymore et al. 2009). For example, results from using single-molecule
atomic force microscopy to investigate the mechanical properties of bovine
tenascin-X indicated that it is an elastic protein, in that the fibronectin type III
domains can unfold under a stretching force and refold to regain their mechanical
stability when the force is removed. Mechanical stability, mechanical unfolding
kinetics, and contour length increment upon domain unfolding may be disrupted by
mutations yet uncharacterized (Jollymore et al. 2009). This is further buttressed by
the presence of distinct tenascin-X proteins composed of distinct molecular species
that retain functional activity (Egging et al. 2007), although none of these proteins
found in humans have been reported in sheep.
Biochem Genet
123

The SNP genotype had significant effects on pulse rate and skin temperature.
This could be attributed to a reduction in the collagen deposition, which greatly
affects the tensile strength of the blood vessels, thereby leading to stiffness and
hardening of the arteries, arterioles, capillaries, and veins (Lethias et al. 2006;
Egging et al. 2007; Jollymore et al. 2009). Reduction in the amount of blood
circulating around the body due to the stiffness of the blood vessels was
compensated for by an increase in the pulse rate. Our hypothesis is supported by the
ubiquitous and prominent expression of tenascin-X protein in blood vessels and
heart muscles, validated by intense staining in these regions (Matsumoto et al.
1994). Similarly, loss of tenascin-X from the vessel wall could affect collagen fibril
and elastic fiber properties, thereby causing weakness of the aortic wall (Zweers
et al. 2004), consistent with recent findings that a TNX-knockout mouse model
showed muscle weakness, mild myopathic features on histology, and functional
upregulation of genes encoding proteins involved in extracellular matrix degrada-
tion and synthesis (Voermans et al. 2011).
Tenascin-X expression in the heart during embryogenesis and adulthood suggests
its vital role in the development of the heart (Matsumoto et al. 1994, 2012; Burch
et al. 1995; Jing et al. 2011). Absence of TNXB in gene-targeted mice, however, did
not result in an apparent cardiac phenotype (Mao et al. 2002), but skin biopsies of
tenascin-X deficient humans showed severe abnormalities in elastic fibers and
collagen, which are also principal components of blood vessels (Zweers et al. 2004).
There is currently no published work on TNXB gene mutations on physiological
indices in ovine species. Therefore, its possible use as a candidate gene that affects
the rhythmic expansion of blood vessels in sheep awaits further investigation. The
reason for the obvious effect of SNP genotypes and interaction effect of breed and
SNP genotype on skin temperature is largely unknown. Significant results obtained
for interactions of breed with SNP genotype and location with SNP genotype on
chest girth suggest the sensitivity of the expressed forms of TNXB genotypes to
environmental components and breed differences. This may reflect a role in skeletal
muscle, heart muscle, esophagus, and gut, given the large amounts of deposited
collagen in these organs (Matsumoto et al. 1994; Burch et al. 1997). The observed
effect of this polymorphism may be mediated through its role in connective tissue
biology, and it may be a potential molecular genetic marker for morphological
growth and physiological indices when validated in a larger population supported by
functional studies. In addition, further studies are required to test the biochemical
effects of TNXB isoforms on quantitative traits.
Acknowledgments Financial support from the College of Agriculture and Life Sciences, Cornell
University, Ithaca, New York, is gratefully acknowledged.
References
Beighton P, De Paepe A, Steinmann B, Tsipouras P, Wenstrup RJ (1998) Ehlers-Danlos syndromes:
revised nosology, Villefranche. Ehlers-Danlos National Foundation (USA) and Ehlers-Danlos
Support Group (UK). Am J Med Genet 77:31–37
Biochem Genet
123

Blench R (1999) Traditional livestock breeds: geographical distribution and dynamics in relation to the
ecology of West Africa. Working Paper 122. Overseas Development Institute, London
Bosman FT, Stamenkovic I (2003) Functional structure and composition of the extracellular matrix.
J Pathol 200:423–428
Burch GH, Bedolli MA, McDonough S, Rosenthal SM, Bristow J (1995) Embryonic expression of
tenascin-X suggests a role in limb, muscle, and heart development. Dev Dyn 203(4):491–504
Burch GH, Gong Y, Liu W, Dettman RW, Curry CJ, Smith L, Miller WL, Bristow J (1997) Tenascin-X
deficiency is associated with Ehlers-Danlos syndrome. Nat Genet 17:104–108
Castillo JA, Medina RD, Villalobos JM, Gayosso-Vazquez A, Ulloa-Arvızu R, Rodrıguez RA, Ramırez
HP, Morales RA (2011) Association between major histocompatibility complex microsatellites,
fecal egg count, blood packed cell volume and blood eosinophilia in Pelibuey sheep infected with
Haemonchus contortus. Vet Parasitol 177(3–4):339–344
Egging DF, Van Vlijmen I, Starcher B, Gijsen Y, Zweers MC, Blankevoort L, Bristow J, Schalkwijk J
(2006) Dermal connective tissue development in mice: an essential role for tenascin-X. Cell Tissue
Res 323:465–474
Egging DF, Peeters AC, Grebenchtchikov N, Geurts-Moespot A, Sweep CG, den Heijer M, Schalkwijk J
(2007) Identification and characterization of multiple species of tenascin-X in human serum. FEBS J
274(5):1280–1289
Elefteriou F, Exposito JY, Garrone R, Lethias C (1997) Characterization of the bovine tenascin-X. J Biol
Chem 272(36):22866–22874
Elefteriou F, Exposito JY, Garrone R, Lethias C (1999) Cell adhesion to tenascin-X mapping of cell
adhesion sites and identification of integrin receptors. Eur J Biochem 263(3):840–848
Geldermann H, Mir MR, Kuss AW, Bartenschlager H (2006) OLA-DRB1 microsatellite variants are
associated with ovine growth and reproduction traits. Genet Sel Evol 38(4):431–444
Hickford JG, Forrest RH, Zhou H, Fang Q, Frampton CM (2011) Association between variation in faecal
egg count for a mixed field-challenge of nematode parasites and ovine MHC-DQA2 polymorphism.
Vet Immunol Immunopathol 144(3–4):312–320
Hui WQ, Jia B, Zhao ZS, Du YC, Shen H (2012) Differential expressions of MHC-DQB1 mRNA in
Chinese merino sheep infected with Echinococosus granuclosus. Parasitol Res 110(5):2075–2079
Jing L, Zhou LJ, Zhang FM, Li WM, Sang Y (2011) Tenascin-x facilitates myocardial fibrosis and cardiac
remodeling through transforming growth factor-b1 and peroxisome proliferator-activated receptor cin alcoholic cardiomyopathy. Chin Med J (Engl) 124(3):390–395
Jollymore A, Lethias C, Peng Q, Cao Y, Li H (2009) Nanomechanical properties of tenascin-X revealed
by single-molecule force spectroscopy. J Mol Biol 385(4):1277–1286
Keane OM, Dodds KG, Crawford AM, McEwan JC (2007) Transcriptional profiling of Ovis aries
identifies Ovar-DQA1 allele frequency differences between nematode-resistant and susceptible
selection lines. Physiol Genomics 30(3):253–261
Kumanovics A, Takada T, Lindahl KF (2003) Genomic organization of the mammalian MHC. Annu Rev
Immunol 21:629–657
Larruskain A, Minguijon E, Arostegui I, Moreno B, Juste RA, Jugo BM (2012) Microsatellites in
immune-relevant regions and their associations with Maedi–Visna and ovine pulmonary adenocar-
cinoma viral diseases. Vet Immunol Immunopathol 145(1–2):438–446
Lethias C, Carisey A, Comte J, Cluzel C, Exposito JY (2006) A model of tenascin-X integration within
the collagenous network. FEBS Lett 580(26):6281–6285
Mao JR, Taylor G, Dean WB, Wagner DR, Afzal V, Lotz JC, Rubin EM, Bristow J (2002) Tenascin-X
deficiency mimics Ehlers-Danlos syndrome in mice through alteration of collagen deposition. Nat
Genet 30:421–425
Matsumoto K, Saga Y, Ikemura T, Sakakura T, Chiquet-Ehrismann R (1994) The distribution of tenascin-
X is distinct and often reciprocal to that of tenascin-C. J Cell Biol 125(2):483–493
Matsumoto KI, Satoh K, Maniwa T, Araki A, Maruyama R, Oda T (2012) Noticeable decreased
expression of tenascin-X in calcific aortic valves. Connect Tissue Res 53(6):460–468
McClelland EE, Penn DJ, Potts WK (2003) Major histocompatibility complex heterozygote superiority
during coinfection. Infect Immun 71:2079–2086
Morel Y, Bristow J, Gitelman SE, Miller WL (1989) Transcript encoded on the opposite strand of the
human steroid 21-hydroxylase/complement component C4 gene locus. Proc Natl Acad Sci USA
86(17):6582–6589
Biochem Genet
123

Oladimeji BS, Osinowo OA, Alawa JP, Hambolu JO (1993) Estimation of average values for pulse rate,
respiratory rate and rectal temperature and development of heat stress index for adult Yankassa
sheep. Bull Anim Prod Hlth Africa 45:105–107
Qin J, Mamotte C, Cockett NE, Wetherall JD, Groth DM (2008) A map of the class III region of the sheep
major histocompatibility complex. BMC Genomics 9:409
SAS (2009) Statistical analysis system user’s guide. SAS Institute Inc., Cary, NC, USA
Schalkwijk J, Zweers MC, Steijlen PM, Dean WB, Taylor G, Van Vlijmen IM, Van Haren B, Miller WL,
Bristow J (2001) A recessive form of the Ehlers-Danlos syndrome caused by tenascin-X deficiency.
N Engl J Med 345(16):1167–1175
Vincze T, Posfai J, Roberts RJ (2003) NEB cutter: a program to cleave DNA with restriction enzymes.
Nucleic Acids Res 31:3688–3691
Voermans NC, Altenburg TM, Hamel BC, De HA, Van Engelen BG (2007) Reduced quantitative muscle
function in tenascin-X deficient Ehlers-Danlos patients. Neuromuscul Disord 17:597–602
Voermans NC, Altenburg TM, Hamel BC, De HA, Van Engelen BG (2009) Joint hypermobility as a
distinctive feature in the differential diagnosis of myopathies. J Neurol 256:13–27
Voermans NC, Verrijp K, Eshuis L, Balemans MC, Egging D, Sterrenburg E, van Rooij IA, van der Laak
JA, Schalkwijk J, van der Maarel SM, Lammens M, van Engelen BG (2011) Mild muscular features
in tenascin-X knockout mice, a model of Ehlers-Danlos syndrome. Connect Tissue Res
52(5):422–432
Walsh EC, Mather KA, Schaffner SF, Farwell L, Daly MJ, Patterson N, Cullen M, Carrington M,
Bugawan TL, Erlich H (2003) An integrated haplotype map of the human major histocompatibility
complex. Am J Hum Genet 73:580–590
Yakubu A, Salako AE, Imumorin IG, Ige AO, Akinyemi MO (2010) Discriminant analysis of
morphometric differentiation in the West African Dwarf and Red Sokoto goats. S Afr J Anim Sci
40(4):381–387
Zweers MC, Bristow J, Steijlen PM, Dean WB, Hamel BC, Otero M, Kucharekova M, Boezeman JB,
Schalkwijk J (2003) Haploinsufficiency of TNXB is associated with hypermobility type of Ehlers-
Danlos syndrome. Am J Hum Genet 73(1):214–217
Zweers MC, Vlijmen-Willems IM, Van Kuppevelt TH, Mecham RP, Steijlen PM, Bristow J, Schalkwijk J
(2004) Deficiency of tenascin-X causes abnormalities in dermal elastic fiber morphology. J Investig
Dermatol 122:885–891
Biochem Genet
123




















