
A Novel Polymorphic AP-1 Binding Element of the GFAP Promoter is Associated with Different Allelic...
10
doi: 10.1111/j.1469-1809.2010.00614.x A Novel Polymorphic AP-1 Binding Element of the GFAP Promoter is Associated with Different Allelic Transcriptional Activities Tiziana Bachetti 1 , Eleonora Di Zanni 1 , Francesca Lantieri 1,2 , Francesco Caroli 1 , Stefano Regis 3 , Mirella Filocamo 3 , Innocenzo Rainero 4 , Salvatore Gallone 4 , Roberto Cilia 5 , Silvia Romano 6 , Mario Savoiardo 7 , Davide Pareyson 8 , Roberta Biancheri 9 , Roberto Ravazzolo 1,10 and Isabella Ceccherini 1∗ 1 Laboratorio di Genetica Molecolare, Istituto G. Gaslini, Genova, Italy 2 Dip. Scienze della Salute, Unit` a Biostatistica, Universit` a di Genova, Italy 3 Lab. Diagnosi Pre-Postnatale Malattie Metaboliche, Istituto G. Gaslini, Genova, Italy 4 Neurologia II, Dip Neuroscienze, Universit` a di Torino, Italy 5 Istituto per il Parkinson, Istituti Clinici di Perfezionamento, Milano, Italy 6 Dip Neurologia, Universit` a La Sapienza, Roma, Italy 7 Dip. Neuroradiologia, IRCCS Ist. Neurologico Carlo Besta, Milano, Italy 8 Neuroscienze Cliniche, IRCCS Ist. Neurologico Carlo Besta, Milano, Italy 9 Neuropsichiatria Infantile, Istituto G. Gaslini, Genova, Italy 10 Dipartimento di Pediatria e CEBR, Universit` a di Genova, Genova, Italy Summary The Glial Fibrillary Acidic Protein (GFAP) gene encodes a cytoskeletal protein belonging to the intermediate filament family whose expression is considered as a marker of astrocytes differentiation. GFAP expression, shown to be upregulated as a consequence of brain gliosis, depends on hormones, growth factors, cytokine, and transcription factors and, among these latters, activator protein 1 (AP-1) has been demonstrated to play a crucial role. In this study, we have focused on a 2.2 kb sequence of the regulatory region located upstream of the GFAP gene, searching in a panel of control individuals for single-nucleotide polymorphisms (SNPs) that could modulate GFAP transcription. Among four SNPs of the GFAP promoter whose alleles have been predicted by in silico analysis to induce differences in the pattern of binding transcription factors, we have identified a new AP-1 binding site lying at −250 bp upstream from the GFAP transcriptional start site. The two alleles of this polymorphic locus have shown to bind the AP-1 complex to different extents, thus promoting variable transcriptional activities of the GFAP promoter. Therefore, these SNP alleles may, among others, mediate the effects of GFAP mutations, thus explaining the phenotypic heterogeneity of Alexander disease. Keywords: GFAP promoter, single nucleotide polymorphisms, AP-1 Introduction The Glial Fibrillary Acidic Protein (GFAP) gene encodes for the GFAP that represents the main component of the astrocyte-specific intermediate filaments (Eliasson et al., ∗ Corresponding author: Isabella Ceccherini, Ph.D., Lab Genet- ica Molecolare, Istituto Giannina Gaslini, Largo Gerolamo Gaslini, 5-16148, Genova, Italy. Tel: +39-010-5636800; Fax: +39-010- 3779797; E-mail: [email protected] 1999). The GFAP protein is considered as a marker of as- trocyte differentiation, known to play also a role in astrocyte morphology and function. While levels of GFAP, and the state of its assembly into filaments, seem to be important in modulating astrocyte motility and shape (Laping et al., 1994), GFAP knock-out mice have been reported to show a nor- mal abundance of astrocytes, characterised by biochemical and morphological features of immature astrocytes, though they develop normally (Pekny et al., 1995; Menet et al., 2001). 506 Annals of Human Genetics (2010) 74,506–515 C 2010 The Authors Annals of Human Genetics C 2010 Blackwell Publishing Ltd/University College London
Transcript of A Novel Polymorphic AP-1 Binding Element of the GFAP Promoter is Associated with Different Allelic...

doi: 10.1111/j.1469-1809.2010.00614.x
A Novel Polymorphic AP-1 Binding Element of the GFAPPromoter is Associated with Different AllelicTranscriptional Activities
Tiziana Bachetti1, Eleonora Di Zanni1, Francesca Lantieri1,2, Francesco Caroli1, Stefano Regis3,Mirella Filocamo3, Innocenzo Rainero4, Salvatore Gallone4, Roberto Cilia5, Silvia Romano6,Mario Savoiardo7, Davide Pareyson8, Roberta Biancheri9, Roberto Ravazzolo1,10
and Isabella Ceccherini1∗1Laboratorio di Genetica Molecolare, Istituto G. Gaslini, Genova, Italy2Dip. Scienze della Salute, Unita Biostatistica, Universita di Genova, Italy3Lab. Diagnosi Pre-Postnatale Malattie Metaboliche, Istituto G. Gaslini, Genova, Italy4Neurologia II, Dip Neuroscienze, Universita di Torino, Italy5Istituto per il Parkinson, Istituti Clinici di Perfezionamento, Milano, Italy6Dip Neurologia, Universita La Sapienza, Roma, Italy7Dip. Neuroradiologia, IRCCS Ist. Neurologico Carlo Besta, Milano, Italy8Neuroscienze Cliniche, IRCCS Ist. Neurologico Carlo Besta, Milano, Italy9Neuropsichiatria Infantile, Istituto G. Gaslini, Genova, Italy10Dipartimento di Pediatria e CEBR, Universita di Genova, Genova, Italy
Summary
The Glial Fibrillary Acidic Protein (GFAP) gene encodes a cytoskeletal protein belonging to the intermediate filamentfamily whose expression is considered as a marker of astrocytes differentiation. GFAP expression, shown to be upregulatedas a consequence of brain gliosis, depends on hormones, growth factors, cytokine, and transcription factors and, amongthese latters, activator protein 1 (AP-1) has been demonstrated to play a crucial role. In this study, we have focused on a2.2 kb sequence of the regulatory region located upstream of the GFAP gene, searching in a panel of control individualsfor single-nucleotide polymorphisms (SNPs) that could modulate GFAP transcription. Among four SNPs of the GFAPpromoter whose alleles have been predicted by in silico analysis to induce differences in the pattern of binding transcriptionfactors, we have identified a new AP-1 binding site lying at −250 bp upstream from the GFAP transcriptional start site.The two alleles of this polymorphic locus have shown to bind the AP-1 complex to different extents, thus promotingvariable transcriptional activities of the GFAP promoter. Therefore, these SNP alleles may, among others, mediate theeffects of GFAP mutations, thus explaining the phenotypic heterogeneity of Alexander disease.
Keywords: GFAP promoter, single nucleotide polymorphisms, AP-1
Introduction
The Glial Fibrillary Acidic Protein (GFAP) gene encodesfor the GFAP that represents the main component ofthe astrocyte-specific intermediate filaments (Eliasson et al.,
∗Corresponding author: Isabella Ceccherini, Ph.D., Lab Genet-ica Molecolare, Istituto Giannina Gaslini, Largo Gerolamo Gaslini,5-16148, Genova, Italy. Tel: +39-010-5636800; Fax: +39-010-3779797; E-mail: [email protected]
1999). The GFAP protein is considered as a marker of as-trocyte differentiation, known to play also a role in astrocytemorphology and function. While levels of GFAP, and thestate of its assembly into filaments, seem to be important inmodulating astrocyte motility and shape (Laping et al., 1994),GFAP knock-out mice have been reported to show a nor-mal abundance of astrocytes, characterised by biochemicaland morphological features of immature astrocytes, thoughthey develop normally (Pekny et al., 1995; Menet et al.,2001).
506 Annals of Human Genetics (2010) 74,506–515 C© 2010 The AuthorsAnnals of Human Genetics C© 2010 Blackwell Publishing Ltd/University College London

New polymorphic AP-1 binding site in GFAP promoter
The regulation of GFAP expression has been investigatedfor several years (Laping et al., 1994; Gomes et al., 1999; Suet al., 2004; de Leeuw et al., 2006; Lee et al., 2008) andhas been shown to depend on hormones, growth factors, cy-tokine, and transcription factors. In particular, transcriptionof the GFAP gene has been investigated to disclose mecha-nisms of astrocyte differentiation and response to brain injury(Kahn et al., 1997). Moreover, the identification of regulatoryregions on its promoter has proved to be useful to create pro-moter constructs able to direct the expression of genes intoastrocytes (Brenner et al., 1994).
An important role of the activator protein 1 (AP-1) com-plex, containing proteins belonging to the Jun and Fos families(c-Jun, JunB, JunD and c-Fos, FosB, Fra-1, Fra-2, respectively,Chiu et al., 1988; Raucher et al., 1988), in regulating GFAPexpression has recently been reported after observing that in-hibition of the c-Jun component of AP-1 induced a drasticdecrease in GFAP expression (Gopalan et al., 2006). Simi-larly, GFAP transcriptional regulation has also been shown todepend on the nuclear factor-kappa B (NF-κB), as aspirin-induced decrease of the NF-κB activation effectively loweredGFAP protein levels in human astroglial cells (Bae et al., 2006).
Mutations in the GFAP coding region have been detectedin individuals affected with Alexander disease (Brenner et al.,2001), a rare genetic leukoencephalopathy characterised bythe presence in astrocytes of deposits called Rosenthal fi-bres, containing GFAP, small heat shock proteins (sHSPs),and ubiquitin (Quinlan et al., 2007). Although the molecu-lar pathogenesis of Alexander disease seems to rely on toxicoligomeric forms of mutant GFAP (Tang et al., 2010), it hasbeen shown that also overexpression of the wild-type GFAPprotein can induce pathogenetic cell events (Messing et al.,1998; Koyama & Goldman, 1999; Tanaka et al., 2007; Choet al., 2009), suggesting that either disease occurrence in pa-tients lacking mutations of the GFAP coding region or severeexpressivity in early onset infantile patients could depend onincreased protein levels.
In this light, we have hypothesized that the nucleotidesequence variability, commonly present within the GFAPpromoter of normal individuals, may account for differentexpression levels shown by this neurofilament.
To this end, we have first genotyped a number of healthyindividuals for apparently neutral SNPs of the GFAP pro-moter and, secondly, we have checked whether binding oftranscription factors usually interacting with the GFAP pro-moter could be altered by such nondeleteriuos substitutions.Finally, we have verified the effect of one specific SNP onmodulating the GFAP transcriptional activity.
Materials and Methods
DNA Extraction from Healthy Subjects
DNA was extracted from 50 anonymous blood samples obtainedfrom blood donors of the Istituto Gaslini, according to standardprocedures. Informed consent was obtained from these individ-uals to use their DNA as control samples for research purposes.
DNA Analysis of the SNPs in the GFAP Promoter
The 2123 bp segment upstream of the GFAP transcriptionalstarting site (GenBank accession no. NT_010783) has been sub-divided into four regions that have been analysed by polymerasechain reaction (PCR) amplification (GC-RICH PCR system,Roche, Indianapolis, IN) using the four couples of oligonu-cleotides (GP1F/1R, GP2F/2R, GP3F/3R, and GP4F/4R) de-scribed in Table 1.
Direct sequencing of each amplicon was performed on an ABI3100 DNA automated Sequencer (Big Dye terminator, AppliedBiosystem, Carlsbad, CA, USA), by using the above mentionedspecific primers; in addition, to overcome technical problemscaused by the large poly-A stretch of amplimer 1 and the poly-T stretch of amplimer 4 of the GFAP promoter, further DNAsequencing was carried out with the forward GP1bis and GP4bisprimers, respectively (Table 1).
Table 1 Sequences of the primers usedto amplify the four promoter segments
Amplicon Primer 5′->3′ sequence1 Amplimer size (bp)
1 GP-1F GCACTTTGGGAGGCTGAT 589GP-1R AAGATGCCAGGCTGTCAGGGP1bis CCTGGTGTGGAGTAGGGG
2 GP-2F CGAGAAGCCCATTGAGCAG 696GP-2R CATGGCCTCTATGTGGCAG
3 GP-3F GATCCAGGGGCTAAAGTCC 459GP-3R AGCTGTGTTTGCGCCACTC
4 GP-4F CAGGTCAGAGGTCATCTGG 538GP-4R GGGATGCGAGGGCTTTATGGP4bis GACAAGGTCTCTCTCTG
1Primers used for direct sequencing only are shown in italics.
Annals of Human Genetics (2010) 74,506–515 507C© 2010 The AuthorsAnnals of Human Genetics C© 2010 Blackwell Publishing Ltd/University College London

T. Bachetti et al.
Haplotype and Linkage Disequilibrium (LD)
Haplotypes for the six SNPs were reconstructed by the use ofthe software PHASE (Stephens et al., 2001), and LD was esti-mated with Haploview (Barret et al., 2005). The SNP −1278was excluded from the LD analysis since its minor allele fre-quency (MAF) was lower than 10%, and SNP −504 was ex-cluded because it was genotyped in only 18 of 50 controls. Only46 individuals that had genotype calls for at least four SNPs orthree SNPs plus the SNP −250 polymorphism were used for theanalysis.
In Silico Analysis of Transcription FactorBinding Sites
To detect possible differences in transcription factors predictedto bind these SNPs and surrounding regions, the two allelevariants of SNPs thus identified were subjected to the Geno-matix Matinspector software, containing the MatBase database(http://www.genomatix.de), and to the Promo3.0 software, con-taining information from the TRANSFAC database (Farre et al.,2003; Messeguer et al., 2002). The rate of dissimilarity consid-ered for each analysis was 20%.
Plasmids Construction
pGL3basic-GFAP promoter(A) – A sequence of about 2.2 kb,upstream of the GFAP transcription start site, was ampli-fied from DNA extracted from a healthy individual homozy-gous for the −250 A allele with the following primers:GFAPprom[F] 5′-taccctgggctgggcgca-3′ and GFAPprom[R]5′-aattaagcttggcgagcagcggaggtga-3′, containing the underlinedHindIII restriction site. The PCR product was inserted into thepCR2.1 vector (TOPO TA Cloning, Invitrogen, Paisley, UK),downstream of a HindIII site, completely sequenced, HindIIIreleased and transferred into the pGL3basic vector (Promega,Madison, WI, USA) previously digested with the same enzyme.
pGL3basic-GFAP promoter(C) – Amplimer 4 of the GFAP pro-moter was amplified starting from DNA of a −250C homozy-gous healthy individual, inserted into the pCR2.1 vector (TOPOTA Cloning, Invitrogen) and completely sequenced. The 230bp region obtained from a NsiI-Bsu36I double digestion, andcontaining the C allele, was inserted into the pGL3basic-GFAPpromoter(A) construct, previously NsiI-Bsu36I digested.
pGL3basic-GFAPprom(MIN) – A region of about 600 bp wasamplified starting from the full length A clone with the GP4Fand GFAPprom[R] primers; the PCR product was inserted intopCR2.1 vector downstream of a HindIII site, completely se-quenced, HindIII released and transferred into the pGL3basicvector (Promega) previously digested with the same enzyme.
Cell Culture and Luciferase ReporterActivity Assay
The U251-MG human astrocytoma cell line was grownin RPMI medium (Euroclone, CELBIO, Pero, MI, Italy)supplemented with 10% fetal brain serum (FBS) (New
Zealand), 1% L-glutamine (100×), 100 U/ml penicillin, and100 mg/ml streptomycin in a humidified atmosphere with 5%CO2. Transient transfections were performed plating U251-MGcells directly with the transfection mix containing 1 μg of theGFAP reporter construct and 3 μl of Lipofectamine 2000 (In-vitrogen) in 35 mm diameter dishes.
The plasmid pRL-CMV, expressing the Renilla Luciferase gene,was used as an internal control for each sample. Twenty-fourhours after transfection, cells were assayed for Luciferase activity(Dual-Luciferase Reporter Assay System, Promega) using a TD-20/20 Luminometer following manufacturer’s instructions.
Nuclear Extracts and ElectromobilityShift Assay (EMSA)
Nuclear extracts from U251-MG and HeLa cells were preparedfollowing the manufacturer’s instructions (Nuclear and Cytoplas-mic Extraction Reagents, CELBIO, Pero, MI, Italy) and, alter-natively, a protocol already published (Schreiber et al., 1989).The C probe (5′-agagagggtcctcttgcttcagcggtcagg-3′) and the Aprobe (5′-agagagggtcctcttgattcagcggtcagg-3′), both biotinylatedat the 5′ end, were incubated for 20 min with an equal amountof nuclear extracts in the presence of 10% glycerol, 1 μg dIdC,binding buffer 10×. In competition assays, a 200-fold excess ofC or A cold probes and the probe containing the underlined AP-1 consensus sequence (5′-cgcttgatgactcagccggaa-3′, Santa CruzBiotechnology, Santa Cruz, CA, USA) were added during incu-bation with the mix. The c-Jun antibody (Santa Cruz Biotech-nolgy) was preincubated with nuclear extracts, glycerol and bind-ing buffer on ice for 20 min. The DNA-protein complexes wererun on a 6% acrylamide gel, transferred to a N+ nylon mem-brane, cross-linked and revealed by chemiluminescence (LightShift Chemiluminescent EMSA kit, Pierce).
Analysis of Correlation Between GFAP −250C/AGenotype and Gene Expression
To investigate differential levels of expression in the presenceof either A or C alleles of SNP rs2070935, we checkedthe publicly available expression databases of the laboratoryof functional neurogenomics of the University of Miami(http://labs.med.miami.edu/myers/LFuN/LFuN.html) that in-cludes gene expression and genotypic information from thebrain cortex of 193 individuals of European descent withoutcerebrovascular or neurological diseases (Myers et al., 2007).LD and haplotypes were estimated on the CEU sample fromHapMap (http://hapmap.ncbi.nlm.nih.gov/, release 27) investi-gating SNPs in the 200 kb around rs2070935 using haploview(Barret et al., 2005).
Quantitative association analysis of the GFAP level of ex-pression with the SNP rs17629022 was performed with plink(Purcell et al., 2007; http://pngu.mgh.harvard.edu/purcell/plink/). Comparisons of expression levels between the genotypegroups were performed by the one-way ANalysis Of Variance(ANOVA) and by the unpaired t-test implemented in SPSS, ver-sion 17.0 (SPSS, Chicago, IL, USA).
508 Annals of Human Genetics (2010) 74,506–515 C© 2010 The AuthorsAnnals of Human Genetics C© 2010 Blackwell Publishing Ltd/University College London

New polymorphic AP-1 binding site in GFAP promoter
Results
SNPs at the GFAP Locus in a Set of ItalianIndividuals
Fifty healthy subjects were analysed for the presence of SNPsin 2.2 kb of the GFAP promoter. Direct nucleotide sequenc-ing of this region, subdivided into four amplimers named 1–4,led to the identification of six SNPs, defined through theirposition with respect to the transcriptional start point.
As shown in Table 2, the SNPs were identified with dif-ferent allele frequencies in the above subject panel. In par-ticular, one SNP was detected in amplimer 1 (−1926G/A),three in amplimer 2 (−1312G/C, −1278C/T, −986T/C)and two in amplimer 4 (−504T/A and −250C/A), respec-tively. Among these SNPs, variations at the −986, −504, and−250 loci have already been reported, while the others arereported here for the first time. Haplotype and LD analysesshowed that −250C/A is in fairly high LD with −986T/Cand −1312G/C and, in particular, the A allele is generally onthe same haplotype with the −986 T allele, while the C alleleis mostly on the same haplotype with the −1312 G allele (datanot shown).
In silico analysis was then performed by using the Geno-matix Matinspector software, to identify possible differences,between the two alleles of each SNP, in the transcription fac-tors pattern predicted to bind each SNP locus. In two of thesix SNPs, the less frequent variant allele did not result in alter-ation of the binding sites predicted for the most frequent allele,while alleles of the remaining four polymorphisms turned outto be associated with variations in binding proteins, as listedin Table 3. In particular, SNPs lying in potentially regula-tory loci have been subdivided into two groups: one showingdifferent transcription factors predicted to bind each of the
Table 2 SNPs alleles and their frequencies
SNPs Allele Allele frequency
−1926 G 0.88A 0.12
−1312 G 0.86C 0.14
−1278 C 0.97T 0.03
−986 C 0.64T 0.36
−504 T 0.71A 0.29
−2501 C 0.55A 0.45
1NCBI dbSNP (rs2070935).
two alleles (qualitative differences between alleles), the otherone whose variant allele is predicted to gain a binding site(quantitative differences between alleles).
Because of the crucial role played by the AP-1 complexin the regulation of GFAP expression (Gopalan et al., 2006;Masood et al., 1993), we focused on the −250C/A locus,where the A allele was expected to modify the sequence ofthe site by inducing interaction with the AP-1 complex. Firstof all, to confirm the Matinspector prediction, the −250C/Aallele was analysed also with the Promo software. Despite afew differences, the two prediction software packages pro-vided similar results. In particular, while Matinspector hadrecognised the AP-1 binding only for the A allele, the Promosoftware was able to identify a putative binding site for c-Junon the C allele and binding sites for c-Jun, c-Fos, and AP-1on the A allele.
Interaction Between the −250C/A Siteand AP-1
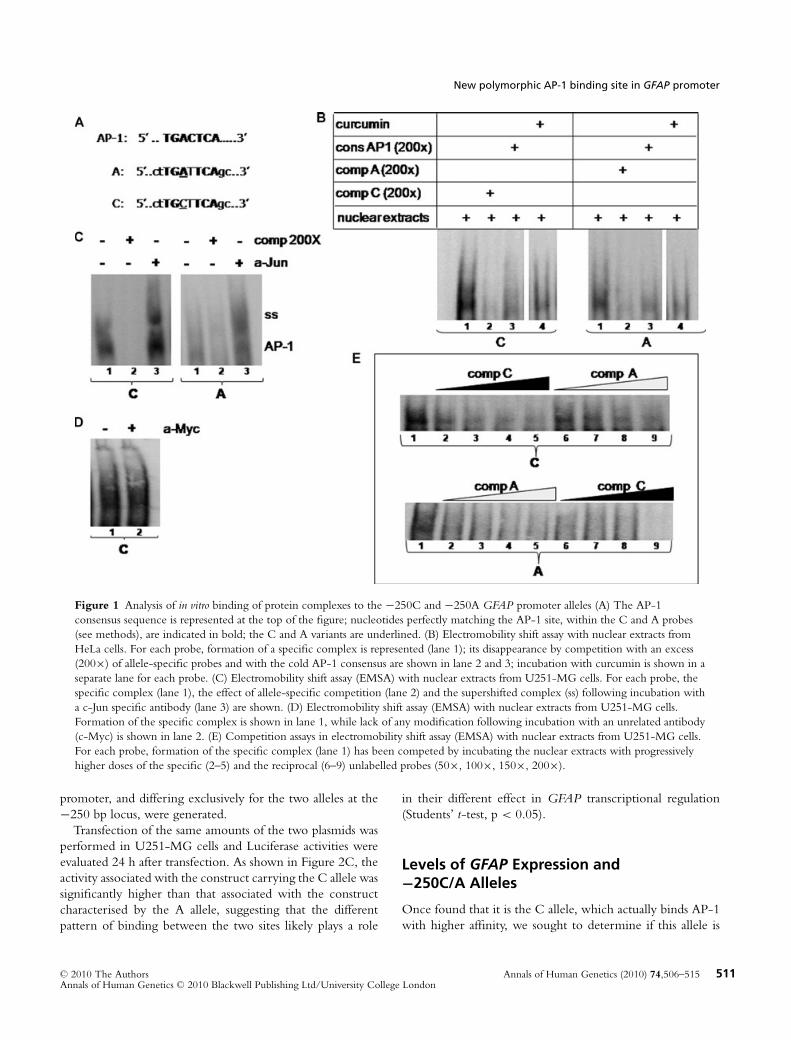
To verify the hypothesis of the insertion of a novel binding sitefor the AP-1 transcription factor into the GFAP promoter,EMSA were performed. First, nuclear extracts obtained fromHeLa cells, characterised by high expression of AP-1, wereincubated with two probes, each containing one of the two al-leles (Fig. 1A). Incubation was also performed in the presenceof a cold probe containing a nonbiotinylated AP-1 consensussequence or in the presence of 10 mM curcumin, a moleculeable to specifically inhibit formation of the AP-1/DNA com-plex in vitro (Juhyung et al., 2006). As a result, formation ofa specific complex, disappearing in the presence of the coldprobe, was demonstrated for both alleles. Moreover, the samecomplex was shown to decrease in the presence of eitheran excess of the cold AP-1 consensus or of curcumin, thussuggesting it likely contained the AP-1 factor. In particular,although the in silico analysis predicted that only the A allelecould induce AP-1 binding, the complex bound both probes,curiously with a higher intensity in the presence of the Callele than in the presence of the A allele (Fig. 1B).
To confirm this pattern of binding also in a cell line ex-pressing endogenous GFAP protein, we performed EMSAwith nuclear extracts obtained from the astrocytoma U251-MG cell line. As shown in Figure 1C, we observed a specificcomplex for both alleles, and the binding to the C probe wasstronger than to the A probe. Moreover, the presence of c-Junin the complex triggered by both alleles has been suggestedby formation of a supershifted band, following incubation ofnuclear extracts with a c-Jun specific antibody. Thus, AP-1is likely to be part of the complex, as previously suggestedby in silico analysis and by results obtained with HeLa nuclearproteins. This effect has been shown to be specific, as the
Annals of Human Genetics (2010) 74,506–515 509C© 2010 The AuthorsAnnals of Human Genetics C© 2010 Blackwell Publishing Ltd/University College London

T. Bachetti et al.
Table 3 Pattern of transcription factorsbinding each allele of the SNPs
SNPs Allele Qualitative differences1
−1926 G ARNT: Aryl hydrocarbon receptor nuclear translocatorHESF: Hey-like-bHLH transcriptional repressorSREBP: Sterol regulatory element-binding protein 1 and 2
A SRF: Serum response element-binding factorPARAXIS/TCF15: Transcription factor 15MIT: Microphthalmia transcriptional factorIRX5: Iroquois homeobox gene 5
SNPs Allele Quantitative differences2
−1278 C –T RTR: Germ cell nuclear receptor family
−504 T –A ABDB: Abdominal-B type homeodomain transcription factor
−250 C c-JunA c-Jun, c-Fos and AP-1 activator protein 1
MEIS/HOXA9: Meis1a and Hoxa9 complex
1Different pattern of transcription factors binding the two alleles.2Aquisition/lack of binding sites by one of the two alleles.
incubation with an antibody specific for a tag absent from ournuclear extracts did not show any significant shift variation(Fig. 1D).
Finally, to further confirm that the complex binds the Callele more effectively than the A allele, we performed com-petition assays by using increasing doses of the A and C coldprobes in the mix containing each allele. As shown in Fig-ure 1E, when labelled C allele was incubated with increasingdoses of cold A allele, a pattern more intense than that re-sulting from incubation with an equal dose of cold C allelewas obtained, confirming that the C allele is characterisedby a stronger binding ability than the A allele. Conversely,because of the small amount of the complex binding the Aallele, every dose of the cold C probe tested could efficientlycompete with the A allele and enable the complex to disap-pear. Consistently, the lowest amount of cold A allele (50×)could only partially compete with the labelled A allele forcomplex binding, as shown by the weak complex that is left,thus confirming that the A allele does bind the −250A/CSNP much more weakly than does the C allele.
Functional Validation of the Novel AP-1 Site
We then sought to assess whether the binding of the AP-1factor to the −250 bp site of the GFAP promoter was alsotranscriptionally functional. To this end, we used a reporterplasmid, the pGL3basic-GFAPprom(MIN), which containedonly the GFAP promoter region proximal to the transcrip-tion start site and including the −250 locus. We avoided theinclusion, in the reporter plasmid, of the distal portion of theGFAP promoter that contains an additional already charac-
terised AP-1 site (Gopalan et al., 2006; Brenner et al., 1994)and that could mask the effects of the −250 bp site underanalysis.
First, we could demonstrate that this region is transcrip-tionally active by transfecting the reporter plasmid in U251-MG cells and comparing the Luciferase activity with thatinduced by the empty vector pGL3basic (Student’s t-test, p <
0.01). Indeed, as shown in Figure 2A, the proximal region ofthe GFAP promoter does maintain a residual transcriptionalactivity.
To confirm AP-1 binding, the plasmid was then transfected,either in the presence or absence of the specific inhibitorof the AP-1/DNA interaction curcumin in the cell culturemedium. Twenty-four hours after transfection, a 50% reduc-tion of transcriptional function (Students’ t-test, p < 0.01)was detected following the curcumin treatment, thus suggest-ing that the proximal portion of the GFAP promoter clonedinto the reporter construct does contain a still unreported andfunctionally active AP-1 site (Fig. 2B).
Functional Effect of the C/A Variationat the −250 bp Locus
After the identification of the AP-1 complex induced by thepresence of both the C and A alleles, though characterisedby different extent of interaction, and the confirmation ofthe functional activity of the novel AP-1 site, we investigatedwhether a different GFAP promoter transcriptional activitycould be ascribed to each of the two alleles.
To study the effect of the −250 bp SNP, two versionsof a Luciferase reporter plasmid containing the entire GFAP
510 Annals of Human Genetics (2010) 74,506–515 C© 2010 The AuthorsAnnals of Human Genetics C© 2010 Blackwell Publishing Ltd/University College London
New polymorphic AP-1 binding site in GFAP promoter
Figure 1 Analysis of in vitro binding of protein complexes to the −250C and −250A GFAP promoter alleles (A) The AP-1consensus sequence is represented at the top of the figure; nucleotides perfectly matching the AP-1 site, within the C and A probes(see methods), are indicated in bold; the C and A variants are underlined. (B) Electromobility shift assay with nuclear extracts fromHeLa cells. For each probe, formation of a specific complex is represented (lane 1); its disappearance by competition with an excess(200×) of allele-specific probes and with the cold AP-1 consensus are shown in lane 2 and 3; incubation with curcumin is shown in aseparate lane for each probe. (C) Electromobility shift assay (EMSA) with nuclear extracts from U251-MG cells. For each probe, thespecific complex (lane 1), the effect of allele-specific competition (lane 2) and the supershifted complex (ss) following incubation witha c-Jun specific antibody (lane 3) are shown. (D) Electromobility shift assay (EMSA) with nuclear extracts from U251-MG cells.Formation of the specific complex is shown in lane 1, while lack of any modification following incubation with an unrelated antibody(c-Myc) is shown in lane 2. (E) Competition assays in electromobility shift assay (EMSA) with nuclear extracts from U251-MG cells.For each probe, formation of the specific complex (lane 1) has been competed by incubating the nuclear extracts with progressivelyhigher doses of the specific (2–5) and the reciprocal (6–9) unlabelled probes (50×, 100×, 150×, 200×).
promoter, and differing exclusively for the two alleles at the−250 bp locus, were generated.
Transfection of the same amounts of the two plasmids wasperformed in U251-MG cells and Luciferase activities wereevaluated 24 h after transfection. As shown in Figure 2C, theactivity associated with the construct carrying the C allele wassignificantly higher than that associated with the constructcharacterised by the A allele, suggesting that the differentpattern of binding between the two sites likely plays a role
in their different effect in GFAP transcriptional regulation(Students’ t-test, p < 0.05).
Levels of GFAP Expression and−250C/A Alleles
Once found that it is the C allele, which actually binds AP-1with higher affinity, we sought to determine if this allele is
Annals of Human Genetics (2010) 74,506–515 511C© 2010 The AuthorsAnnals of Human Genetics C© 2010 Blackwell Publishing Ltd/University College London

T. Bachetti et al.
Figure 2 Transcriptional activity of the −250C and −250A GFAP promoter alleles (A)The diagram represents the Luciferase activity induced by a fragment of (−250A) GFAPpromoter containing the proximal 400 bp and lacking the distal AP-1 site. Values arerepresented as fold induction with respect to the empty vector pGL3basic (arbitraryvalue = 1). (B) Values of Luciferase activity in the presence of curcumin are represented aspercentage of values obtained without any treatment (untr, value = 100). In bothdiagrams, values are the mean ± SD of three independent experiments performed induplicate. Significant differences are indicated by asterisks (Students’ t-test ∗p < 0.05). (C)The diagram represents the Luciferase activity induced by the two GFAP promoter allelestransiently transfected in U251-MG astrocytoma cell line. Values of Luciferase activity arerepresented as percentage with respect to the activity of the C allele (normalised to“100”) and are the mean ± SD of four independent experiments performed in duplicate.The significant difference between the two alleles is indicated by an asterisk (Students’t-test ∗p < 0.05).
512 Annals of Human Genetics (2010) 74,506–515 C© 2010 The AuthorsAnnals of Human Genetics C© 2010 Blackwell Publishing Ltd/University College London

New polymorphic AP-1 binding site in GFAP promoter
also correlated with higher GFAP expression by checkingpublicly available expression databases. Unfortunately, neitherSNP rs2070935 nor any other tagSNPs in complete LDwith it were reported in the University of Miami database(http://labs.med.miami.edu/myers/LFuN/LFuN.html), asite where both gene expression and genotypes are providedfor 193 neuropathologically normal individuals. Nonetheless,we have examined rs17629022, which in the CEU-Caucasiansample is in LD with rs2070935 (r2 = 0.399 and D′ = 1),and found that the C allele is always in phase with the Aallele of rs2070935 (20.1% of the possible haplotypes), whilethe alternative T allele is mainly in phase with the C alleleof rs2070935 (61.3%) but also, more rarely, with the A allele(18.3%). The mean expression value of the GFAP gene forindividuals homozygous for the C allele at rs17629022 inthis database is 11,708, for individuals CT is 13,296 and forindividuals TT is 13,675. However, given the small numberof CC genotypes (11 individuals), the present analysis did notyield any statistically significant quantitative association tothe SNP, or any difference among genotype groups, neitherby performing a univariate ANOVA test (data not shown),nor with a t-test that compared the CC group to the groupwith CT and TT genotypes pulled together (p = 0.371).Nonetheless, since CEU individuals heterozygous for theC allele at rs17629022 are mostly heterozygous for the Aallele at rs2070935 (79%), but could also be AA (21%), andindividuals TT are not only CC (57%), but also CA (39%)and AA (4%) at rs2070935, we are tempted to speculate thatthe C allele at rs2070935 is associated with a higher GFAPexpression level than the A allele, presenting more extremerates than rs17629022, where the possible presence of the Aallele could lead to underestimation of the GFAP expressionlevel.
Discussion
In this study, we report the genetic analysis of a 2.2 kb se-quence of the GFAP promoter in a set of Italian healthyindividuals, to identify common variants that could take partin the transcriptional regulation of GFAP expression and, po-tentially, play a role in the phenotypic variability of Alexanderdisease.
In particular, we have detected SNPs at six loci, three ofwhich are reported here for the first time. These are char-acterised by distinct allelic frequencies and, in four cases, anin silico pattern of binding has predicted the involvement ofdifferent factors with the two allelic variants.
Among these SNPs, we have focused on the −250C/Apolymorphism, whose A variant was supposed to create anovel AP-1 binding site.
EMSA assays showed that, differently from what was ex-pected, both alleles could bind the AP-1 complex, and thenucleo-protein complex was more effective when induced bythe presence of the C allele. The presence of AP-1 withinthe complex was assessed through the observation of differentcircumstances of partial or total disappearance of the com-plex, namely (1) in the presence of the AP-1 cold consen-sus probe, (2) following treatment with curcumin, a naturalmolecule widely used as an inhibitor of the formation of theAP1/DNA complex, (3) following incubation with the c-Junspecific antibody.
The GFAP promoter has already been demonstrated tocontain a functional AP-1 specific site, lying in a more distalportion of the regulatory region (Brenner, 1994). To assessthe possible functional role of the AP-1 binding site lyingon the −250 bp SNP, we first generated a construct lackingthe already described, distal AP-1 consensus sequence of theGFAP promoter, to avoid the possibility that the effect of theproximal AP-1 sequence under investigation could be masked.Luciferase activity of this construct showed about 30-fold in-duction with respect to the empty vector, thus demonstratingthat this region of the promoter is characterised by a de-tectable transcriptional activity; moreover, in the presence ofthe inhibitor of AP-1/DNA binding curcumin, the activitydrastically decreased, suggesting that AP-1 likely takes part inthe regulation of this region.
Finally, we observed that the different extent of bindingdue to the C and A alleles was associated with a transcrip-tional activity of the C variant significantly higher than thatof the A allele. We could confirm such a conclusion throughthe analysis of data, in the public domain, concerning GFAPexpression and genome-wide genotypes in a set of healthysubjects, which showed higher GFAP expression in the pres-ence of the C allele than with the A allele.
These results might have important implications under par-ticular conditions like brain injury or inflammation. As anexample, astrogliosis is characterized by proliferation of astro-cytes and their processes, and associates with enhanced ex-pression of GFAP, thus representing a remarkable response ofastrocytes to several types of CNS injuries. Noteworthy, AP-1is one of the downstream targets of the Toll-like receptors,whose activation has recently been observed in associationwith neurodegeneration (Okun et al., 2009); we thereforesuggest that the −250 bp SNP might modulate the extent ofGFAP expression increase under stress conditions.
Moreover, these findings might help to understand Alexan-der disease pathogenesis. In particular, several mutations havebeen associated with different disease phenotypes, suggestingthat molecular changes in the GFAP protein are likely notsufficient to determine a given degree of severity of clin-ical symptoms. Molecular events and pathogenetic mecha-nisms able to explain such a phenotype heterogeneity are still
Annals of Human Genetics (2010) 74,506–515 513C© 2010 The AuthorsAnnals of Human Genetics C© 2010 Blackwell Publishing Ltd/University College London

T. Bachetti et al.
uncharacterised. Based on present results, we are tempted topostulate that, in addition to environmental factors or to theeffects of other genetic loci, the presence of the C or the Aallele of the −250C/A SNP in cis with a GFAP mutationcould determine a different level of transcription of the mu-tant allele, thus making the mutation differently penetrant andvariably expressed in the phenotype. Indeed, because of thedominant negative effect of mutant GFAP proteins on fila-ment organization (Perng et al., 2006; Bachetti et al., 2008),accounting for a GFAP dominant gain of function, relativelevels of incorrect and wild-type GFAP proteins within a cellare important for the expression and function of the wild-typeallele and can influence the cellular outcome.
Moreover, levels of GFAP protein are crucial for aggre-gate formation, as it has been observed that a threshold foraggregate induction is lowered in cases where mutant GFAPproteins are expressed (Tanaka et al., 2007).
In conclusion, we suggest that the −250 bp SNP mayact as a genetic modifying factor in Alexander disease, bymodulating the relative expression of the wild type and mutantGFAP alleles, thus accounting for the clinical heterogeneityof the disease.
Acknowledgements
T.B. has partially been supported by the ELA (EuropeanLeukodystrophy Association) Research Foundation (Laxou,France). EDZ is a PhD student partially sustained by the gener-ous financial support of “Ristorante San Marco” in Sestri Levante(Genova, Italy).
ReferencesBachetti, T., Caroli, F., Bocca, P., Prigione I., Balbi, P., Biancheri,
R., Filocamo, M., Mariotti, C., Pareyson, D., Ravazzolo, R. &Ceccherini, I. (2008) Mild functional effects of a novel GFAPmutant allele identified in a familial case of adult-onset alexanderdisease. Eur J Hum Genet 16, 462–470.
Bae, M. K., Kim, S. R., Lee, H. J., Wee, H. J., Yoo, M. A., Ock,O. S., Baek, S. Y., Kim, B. S., Kim, J. B., Sik-Yoon & Bae,S. K. (2006) Aspirin-induced blockade of NF-κB activity restrainsup-regulation of glial fibrillary acidic protein in human astroglialcells. Biochim Biophys Acta 1763, 282–289.
Barret, J. C., Fry, B., Maller, J. & Daly, M. J. (2005) Haploview:analysis and visualization of LD and haplotype maps. Bioinformatics21, 263–265.
Brenner, M. (1994) Structure and transcriptional regulation of theGFAP gene. Brain Pathol 4, 245–257.
Brenner, M., Johnson, A. B., Boespflug-Tanguy, O., Rodriguez, D.,Goldman, J. & Messing, A. (2001) Mutations in GFAP, encodingglial fibrillary acidic protein, are associated with Alexander disease.Nat Genet 27, 117–120.
Brenner, M., Kissebert, W. C., Su, Y., Besnard, F. & Messing, A.(1994) GFAP promoter directs astrocytes-specific expression intransgenic mice. J Neurosci 14, 1030–1037.
Chiu, R., Boyle, W. J., Meek., Smeal, T., Hunter, T. & Karin, M.(1988) The c-Fos protein interacts with c-Jun/AP-1 to stimulatetranscription of AP-1 responsive genes. Cell 54, 541–552.
Cho, W., Hagemann, T. L., Johnson, D. A., Johnson, J. A. &Messing, A. (2009) Dual transgenic reporter mice as a toll formonitoring expression of glial fibrillary acidic protein. J Neurochem110, 343–351.
de Leeuw, B., Su, M., ter Horst, M., Iwata, S., Rodijk, M., Hoeben,R. C., Messing, A., Smitt, P. S. & Brenner, M. (2006) Increasedglia-specific transgene expression with glial fibrillary acidic proteinpromoters containing multiple enhancer elements. J Neurosci Res83, 744–753.
Eliasson, C., Sahlgren, C., Berthold, C-H., Stakenberg, J., Celis,J. E., Betsholtz, C., Eriksson, J. E. & Pekny, M. (1999) Interme-diate filaments protein partnership in astrocytes. J Biol Chem 34,23996–24006.
Farre, D., Roset, R., Huerta, M., Adsuara, J. E., Rosello, L.,Mar Alba, M. & Messeguer, X. (2003) Identification of patternsin biological sequences at the ALGGEN server: PROMO andMALGEN. Nucleic Acids Res 31, 3651–3653.
Gomes, F. C. A., Paulin, D. & Moura Neto, V. (1999) Glial acidicfibrillary protein (GFAP): modulation by growth factors and itsimplication in astrocytes differentiation. Braz J Med Biol Res 32,619–631.
Gopalan, S. M., Wilczynska, K. M., Konic, B. S., Bryan, L.& Kordula, T. (2006) Astrocyte-specific expression of the α1-antichymotrypsin and glial fibrillary acidic protein genes requiresactivator protein-1. J Biol Chem 281, 1956–1963.
Juhyung, L., Chi-Hoon, P., Wook-Hwan, K., Yung Ha, H.,Kyungchae, J. & Chul-Hak, Y. (2006) Inhibitory effects of Mo-mordin I derivatives on the formation of Fos-Jun-AP-1 DNAcomplex. Bull Korean Chem Soc 27, 535–538.
Kahn, M. A., Huang, C. J., Caruso, A., Barresi, V., Nazarian, R.,Condorelli, D. F. & de Vellis J. (1997) Ciliary neurotrophic factoractivates JAK/Stat signal transduction cascade abd induces tran-scriptional expression of glial fibrillary acidic protein in glial cells.J Neurochem 68, 1413–1423.
Koyama, Y. & Goldman, J. (1999) Formation of GFAP cytoplasmicinclusions in astrocytes and their disaggregation by αB-crystallin.Am J Pathol 154, 1563–1572.
Laping, N. J., Teter, B., Nichols, N. R., Rozovsky, I. & Finch,C. E. (1994) Glial fibrilary acidic protein: regulation by hormones,cytokines and growth factors. Brain Pathol 4, 259–275.
Lee, Y., Messing, A., Su, M. & Brenner, M. (2008) GFAP pro-moter elements required for region-specific and astrocyte-specificexpression. Glia 56, 481–493.
Masood, K., Besnard, F. & Brenner, M. (1993) Analysis of a seg-ment of the human glial fibrillary acidic protein gene that directastrocyte-specific transcription. J Neurochem 61, 160–166.
Menet, V., Gimenez, y., Ribotta, M., Chauvet, N., Drian, M. J.,Lannoy, J., Colucci-Guyon, E. & Privat, A. (2001) Inactivation ofthe glial fibrillary acidic protein gene, but not that of vimentin,improves neuronal survival and neurite growth by modifying ad-hesion molecule expression. J Neurosci 21, 6147–6158.
Messeguer, X., Escudero, R., Farre, D, Nunez, O., Martınez, J. &Mar Alba, M. (2002) PROMO: detection of known transcriptionregulatory elements using species-tailored searches. Bioinformatics18, 2, 333–334.
Messing, A, Head, M. W., Galles, K., Galbreath, E. J., Goldman,J. E. & Brenner, M. (1998) Fatal encephalopathy with astro-cyte inclusions in GFAP transgenic mice. Am J Pathol 152, 391–398.
514 Annals of Human Genetics (2010) 74,506–515 C© 2010 The AuthorsAnnals of Human Genetics C© 2010 Blackwell Publishing Ltd/University College London

New polymorphic AP-1 binding site in GFAP promoter
Myers, A. J., Gibbs, J. R., Webster, J. A., Rohrer, K., Zhao, A.,Marlowe, L., Kaleem, M., Leung, D., Bryden, L., Nath, P.,Zismann, V. L., Joshipura, K., Huentelman, M. J., Hu-Lince, D.,Coon, K. D., Craig, D. W., Pearson, J. V., Holmans, P., Heward,C. B., Reiman, E. M., Stephan, D. & Hardy, J. (2007) A sur-vey of genetic human cortical gene expression. Nat Genet 39,1494–1499.
Okun, E., Griffioen, K. J., Lathia, J. D., Tang, S. C., Mattson, M. P. &Arumugam, T. V. (2009) Toll-like receptors in neurodegeneration.Brain Res Rev 59, 278–292.
Pekny, M., Leveen, P., Pekna, M., Eliasson, C., Berthold, C. H.,Westermark, B. & Betsholtz, C. (1995) Mice lacking glial fibrillaryacidic protein display astrocytes devoid of intermediate filamentsbut develop and reproduce normally. EMBO J 14, 1590–1598.
Perng, M. D., Su, M., Wen, S. F, Gibbon, T., Prescott, A. R.,Brenner, M. & Quinlan, R. A. (2006) The Alexander disease-causing Glial Fibrillary Acidic Protein mutant, R416W, accumu-lates into Rosenthal fibers by a pathway that involves filamentaggregation and the association of αB-Crystallin and HSP27. AmJ Hum Genet 79, 197–213.
Purcell, S., Neale, B., Todd-Brown, K., Thomas, L., Ferreira,M. A., Bender, D., Maller, J., Sklar, P., de Bakker, P. I., Daly,M. J. & Sham, P. C. (2007) PLINK: a tool set for whole-genomeassociation and population-based linkage analyses. Am J HumGenet 81, 559–575.
Quinlan, R. A., Brenner, M., Goldman, J. E. & Messing, A. (2007)GFAP and its role in Alexander disease. Exp Cell Res 313, 2077–2087.
Raucher, F. J., III, Voulalas, P. J., Franza, B. R., Jr & Curran, T. (1988)Fos and Jun bind cooperatively to the AP-1 site: reconstitution invitro. Genes Develop 2, 1687–1699.
Schreiber, E., Matthias, P., Muller, M. M. & Schaffner, W. (1989)Rapid detection of octamer binding proteins with ‘mini extracts’,prepared from a small number of cells. Nucleic Acids Res 17,6419.
Stephens, M., Smith, N. & Donnelly, P. (2001) A new statisticalmethod for haplotype reconstruction from population data. Am JHum Genet 68, 978–989.
Su, M., Hu, H., Lee, Y., d’Azzo, A., Messing, A. & Brenner, M.(2004) Expression specificity of GFAP transgenes. Neurochem Res29, 2075–2093.
Tanaka, R., Takebayashi, H., Yamakazi, Y., Ono, K., Naruse, M.,Iwasato, T., Itohara, S., Kato, H. & Ikenaka, K. (2007) Murinemodel of Alexander disease: analysis of GFAP aggregate formationand its pathological significance. Glia 55, 617–631.
Tang, G., Perng, M. D., Wilk, S., Quinlan, R. & Goldman,J. E. (2010) Oligomers of mutant glial fibrillary acidic pro-tein (GFAP) Inhibit the proteasome system in alexander dis-ease astrocytes, and the small heat shock protein alphaB-crystallin reverses the inhibition. J Biol Chem 285, 10527–10537.
Received: 29 December 2009Accepted: 12 August 2010
Annals of Human Genetics (2010) 74,506–515 515C© 2010 The AuthorsAnnals of Human Genetics C© 2010 Blackwell Publishing Ltd/University College London




















