
Monitoring the effects of different conservation treatments on paper-infecting fungi
Upload
independentCategory
view
0download
0
©20
12 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
A novel method for infecting Drosophila adult flieswith insect pathogenic nematodes
Julio Cesar Castillo, Upasana Shokal and Ioannis Eleftherianos*Insect Infection and Immunity Lab; Department of Biological Sciences; The George Washington University; Washington, DC USA
Keywords: Drosophila, Heterorhabditis,Photorhabdus, infection model,nematode parasitism, model host
Abbreviations: IJ, infective juveniles;Imd, immune deficiency;PBS, phosphate-buffered saline;WT, wild-type
Submitted: 01/25/12
Revised: 03/30/12
Accepted: 04/02/12
http://dx.doi.org/10.4161/viru.20244*Correspondence to: Ioannis Eleftherianos;Email: [email protected]
Drosophila has been established asan excellent genetic and genomic
model to investigate host-pathogen inter-actions and innate immune defensemechanisms. To date, most informationon the Drosophila immune responsederives from studies that involvebacterial, fungal or viral pathogens.However, immune reactions to insectparasitic nematodes are still not wellcharacterized. The nematodes Hetero-rhabditis bacteriophora live in symbiosiswith the entomopathogenic bacteriaPhotorhabdus luminescens, and they areable to invade and kill insects. Interest-ingly, Heterorhabditis nematodes areviable in the absence of Photorhabdus.Techniques for infecting Drosophilalarvae with these nematodes have beenpreviously reported. Here, we have deve-loped a method for infecting Drosophilaadult flies with Heterorhabditis nema-todes carrying (symbiotic worms) orlacking (axenic worms) their associatedbacteria. The protocol we present can bereadily adapted for studying parasiticstrategies of other insect nematodes usingDrosophila as the host infection model.
Introduction
The use of insect models to investigatepathogenic infection processes and hostinnate immune mechanisms offers certainadvantages over the use of mammalianmodels. Insects are easy to maintain inlarge quantities and their small sizefacilitates artificial infections and extrac-tions of tissues. Insects also share manyphysiological processes with mammalian
hosts, and they can be used as efficientmodels to discover toxins and virulencefactors some of which are also required tosubvert mammalian defenses.1-6
Drosophila melanogaster has been estab-lished as an ideal model for studyinghost-pathogen interactions, as it benefitsfrom the development of a wide range ofmolecular and cellular tools, impressiveadvances in the application of highthroughput forward and reverse genetic/genomic screens and the body ofknowledge accumulated by thousands ofinvestigators.7 In addition, innate immu-nity in Drosophila can be studied as anintegrated system at the level of thewhole organism.8 In recent years, geneticstudies in Drosophila have resulted in thediscovery of distinct immune signalingpathways in response to microbial infec-tions.9 These advances have enjoyed ahigh profile because they have producedthe major spin-off of rekindling interestin the innate immune system ofmammals. Largely thanks to work donefirst on Drosophila, it is now recognizedthat the nuclear localization of nuclearfactor kappa B (NFkB) transcriptionfactors is a universal feature of innateimmune reactions,10,11 and that Toll-likereceptors are of outstanding importancein transducing innate immune responsesin vertebrates.12-15
Drosophila has a multilayered immuneresponse consisting of humoral and cellu-lar mechanisms.16 The hallmark of theDrosophila host defense is the definitionof two main signaling pathways, Toll andimmune deficiency (Imd), which lead tothe activation of distinct members of theNFkB family of transcription factors, andresult in the expression of hundreds of
PROTOCOL
Virulence 3:3, 339–347; May/June 2012; G 2012 Landes Bioscience
www.landesbioscience.com Virulence 339

©20
12 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
target genes, including those encodingantimicrobial peptides (AMPs).17-19
Activation of these pathways depends onrecognition of certain microbial elicitorssuch as bacterial peptidoglycans and fungalglucans.20,21 Two other signaling path-ways, the c-Jun N-terminal kinase (JNK)and Janus kinase-signal transducer andactivator of transcription (JAK-STAT),also participate in regulating immunityeffector genes in Drosophila.22,23 Thebody cavity of Drosophila, like that ofall arthropods, is filled with a circulat-ing hemolymph that contains bothfree-floating and sessile blood cells (hemo-cytes). These are responsible for a numberof cellular defenses, while they can takepart in humoral reactions.24,25 In addition,Drosophila can activate complex proteo-lytic cascades that regulate coagulation andmelanization of hemolymph,26,27 defensesassociated with the production of reactiveoxygen and nitrogen species,28,29 andepithelial responses in the gut that alsoplay important roles in fighting microbialinfections.8,30
Drosophila has previously been usedas a model host to investigate immuneresponses to diverse pathogenic organisms,including bacteria, fungi, viruses andparasitoid wasps.9,20 Recent work has alsobegun to use the power of Drosophila todissect the molecular basis of the insectimmune response to the combined insultof insect parasitic nematodes and theirmutualistic bacterial pathogens.31-34 Unlikemany animals associated with bacterialsymbionts, entomopathogenic nematodesare viable in the absence of their mutualisticbacteria.35 Consequently, each partnerof the mutualistic relationship can beseparated and studied in isolation or incombination, thus enabling pathogenesisto be studied individually or together.36
In addition, to being highly virulentparasites of insects, Heterorhabditis bacterio-phora nematodes maintain a mutualisticrelationship with the entomopathogenicbacteria Photorhabdus luminescens.37 Inparticular, the bacteria are found in thegut of infective juvenile (IJ) worms thatare able to attack and invade insects.38
Once inside the insect, the IJ regurgitatesPhotorhabdus into the hemolymph wherethe bacteria begin to divide exponentiallyproducing a wide range of toxins and
hydrolytic enzymes that result in insectdeath.39 At the same time, the IJ exitsdiapause and develops into an adulthermaphrodite nematode in a processcalled IJ recovery. The adult herma-phrodite lays eggs that hatch and developthrough four juvenile stages into adultnematodes. Remarkably a single IJ enter-ing an individual insect will result in theproduction of . 100,000 IJs over a time-scale of 2–3 weeks. This extremelyefficient relationship provides a fascin-ating model system for studying bacterialpathogen, nematode-vector and insecthost interactions.40-44
Protocols for infecting Drosophilalarvae with Heterorhabditis nematodeshave recently been published;31-33 how-ever, it is not currently known howDrosophila adults respond to nematode
infection. Interestingly, differences havebeen found between the adult fly andlarval immune systems that are probablydue to physiological differences betweenlarvae and adults, and differences in theirlifestyles.45-48 Here we report the develop-ment of a new assay for infecting adultflies with axenic and symbiotic worms(Fig. 1). We have used this method tomonitor fly survival and nematode loadand show that Heterorhabditis nematodesare able to develop in Drosophila adults.In future studies, we will employ thisinfection protocol to assess immune geneexpression in nematode infected wild-type(WT) and mutant adult flies in orderto elucidate how parasitic infections aresensed, and how their presence is commu-nicated both within and among cells andtissues of the host.
Figure 1. Outline of the method for infecting Drosophila adult flies with Heterorhabditisnematodes. We use 4–6 d old adult flies in the infection experiments. Nematode infective juveniles(IJ) are kept in sterile water in tissue culture flasks. Filter paper discs are transferred to the bottom ofsmall plastic cups. Approximately 1,000 nematodes are pipetted onto the filter papers and 10 fliesare transferred to each test cup. A second identical plastic cup is inserted into the test cup torestrict movement of flies between the filter papers and the bottom of the inserted cup.
340 Virulence Volume 3 Issue 3

©20
12 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
Experimental Design
Fly strain.We used 4–6 d old adult flies ofthe Drosophila strain OregonR. Both maleand female flies were used in the assays.Flies were reared at relatively low densities(20 individuals per vial) to prevent over-crowding conditions. All experiments wereperformed using flies from the same batch,whenever possible, to reduce variation.
Bacterial/nematode strains. Heterorha-bditis bacteriophora nematodes and Photo-rhabdus luminescens subsp laumondiistrain TT01 bacteria were used in allexperiments. We also used 2–4 weeks oldHeterorhabditis IJ, as newly hatched andold nematodes show reduced infectivityand host seeking behavior (unpublisheddata). To generate axenic nematodes, weused bacteria of the Photorhabdus tem-perata mutant strain RET16 that supportthe growth of Heterorhabditis withoutcolonizing the nematodes.31 Nematodeswere cultured on lipid agar plates supple-mented with carbenicillin and gentamicin.This resulted in selection of Photorhabdusluminescens cells that were obtained bythe worms. Consequently, nematodesfed on RET16 bacteria turn to axenic(Photorhabdus-free) once they completetheir life cycle.
Materials
Organisms. Insects. OregonR adult flies(kindly provided by Prof. J.M. Reichhart,University of Strasbourg and CNRS,Strasbourg, France); Galleria mellonellasixth instar larvae (PetCo).
Nematode species. Heterorhabditis bac-teriophora strain TT01.
Bacterial strains. (a) Photorhabdusluminescens subsp laumondii strain TT01(kindly provided by Dr David Clarke,University College Cork, Cork, Ireland)
(b) RET-16 mutant, a GFP-labeledvariant of Photorhabdus temperata strainNC1 (gentamicin resistant) (kindly pro-vided by Dr Todd Ciche, Michigan StateUniversity)
(c) Escherichia coli (streptomycin resistant)(ATCC #25254), kindly provided by Prof.Michael R. Strand, University of Georgia).
Reagents. Luria-Bertani (LB) broth(Miller) (1% tryptone, 0.5% yeast extract,1% NaCl) (Amresco, J106)
Bacteriological agar (Amresco, J637)Corn Syrup (MP Biochemicals, 101413)Cod liver oil (MP Biochemicals, 901405)0.5M magnesium chloride solution
(Acros organics, 7791-18-6)Gentamicin (stock solution 10 mg/mL)
(VWR, 97061-370)Carbenicillin (stock solution 100 mg/mL)
(VWR, 97063-146)Phosphate-buffered saline (PBS) (Fisher
Scientific, BP399-500)Sterile water: autoclaved distilled water1% Bleach solution for nematode
sterilization10% Bleach solution for cleaning surfacesEthanol (Absolute), 70% solution for
surface sterilization (Decon-Labs, 2716)Active dry yeast (Fisher Scientific,
S802452)Instant Drosophila media, Equation
4-24 (Carolina Biol. Supply Co., 173202)0.85% Sodium chloride solution
(Fisher Scientific, BP358-1)Glycerol (Fisher Scientific, 633-500)Mineral oil (Alpha Aesar, 8020-83-5)Equipment. Fly incubators (25°C tem-
perature, 60–75% humidity, 12-h light/12-h dark photoperiod.
Paintbrush (size 0) (Fisher Scientific)General-purpose microbiological incu-
bator (VWR)Petri dishes (150 � 15 mm) (VWR,
25384-326)Petri dishes (100 � 15 mm) (VWR,
25384-342)Compartmentalized Petri dishes (Bi-
plates) (100 � 15 mm, 2 sections)(Fisher Scientific, 0875150)
Whatman filter paper, Grade 1 (90 mmdiameter) (Whatman, 1001090)
Whatman filter paper, Grade 1(150 mm diameter) (Whatman, 1001150)
Graduated disposable pipettes (25 mL)(Corning)
One liter bottles (Corning)96 well plates (Corning)Microscopy glass slides (Fisher
Scientific, 12-550-15)Glass coverslips (60 � 24 mm) (Fisher
Scientific, 12-545-M)Cell culture flasks, 80 cm2 culture area
(Thermo, 153732)Cell culture flasks, 175 cm2 culture area
(Thermo, 156502)Plastic cups 0.5 Oz (Solo Cup Co.,
P050-0100)
15 mL conical polypropylene tubes(VWR, 21008-103)
50 mL conical polypropylene tubes(Falcon, 352098)
Kimwipes (4 � 8 in) (Fisher Scientific,S47299)
Orbital shaker (New Brunswick)Needle puller P-1000 (Sutter
Instruments)Borosilicate capillary needles, Sutter
Instruments (OD: 1.0 mm, ID: 0.50 mm,B100-50-10)
Centrifuge with interchangeable rotorsfor 1.5 mL microcentrifuge tubes and 15,50 mL conical tubes (Eppendorf)
Stereomicroscope (Tri-Tech)Nanoject II (Drummond)10 mL disposable syringe (BD Medical,
14-823-2A)25 G syringe needle (Fisher Scientific,
BDC-5124)0.22 mm syringe filter sterile (Fisher
Scientific, 09-719C)Color tape (Fisher Scientific, 15-901-
15A)1.5 mL microcentrifuge tubes (US
Scientific, 1615-5510)Nanodrop spectrophotometer (Thermo
Scientific)Disposable spectrophotometer cuvettes
(VWR, Cat. No. 89005-758)Micropipettes (10, 100, 200, 1000 mL)
(Rainin)Drosophila vials (VWR, 734-2255)Disposable bacterial spreaders (VWR,
89042-018)Drummond #5 Forceps (Fine science
tools, 11252-40)Pneumatic pump for pipettes (VWR,
53502-211)Water bath, Isotemp 215 (Fisher
Scientific)Graduated 100 mL cylinder (Fisher
Scientific)Carbon dioxide (CO2) anesthetizing
system for Drosophila (Genesee Scientific)Rubbermaid container, 6.5 qt.
(Rubbermaid, 3Q31)UV Transilluminator, Spectrolinker
XL-1000 (Spectronics corporation)Setup of reagents. Drosophila food
preparation. Prepare Drosophila instantmedia according to the manufacturer’sinstructions. Briefly, take one small scoopof media and add it to a Drosophila vial.Add 10 mL of sterile water and let the mix
www.landesbioscience.com Virulence 341

©20
12 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
sit for 2 min; add 5–10 yeast granules andcover the tubes with cotton balls. Labelaccordingly. Always use fresh vials whenpreparing fly cultures.
Preparation of antibiotic stocks. Preparestock solutions of gentamicin (10 mg/mL)and carbenicillin (100 mg/mL) by dilutingthe antibiotics in sterile water and filteringthe solution through a 0.2 mm syringefilter. Prepare rifampicin stock solution(75 mg/mL) by dissolving the antibioticin DMSO and vigorous vortexing (nosterilization is required). Keep antibioticstocks at -20°C in the dark.
Preparation of lipid agar plates. They areused for growing axenic nematodes. Add25 g of LB broth, 17 g of bacteriologicalagar, 10 mL of corn syrup in 1 L of doubledistilled water and autoclave the solution.After autoclaving, keep the solution at60°C in a water bath. Add 5 mL of codliver oil (autoclaved), 1 mL of 0.5 MMgCl2, and carbenicillin, gentamicinantibiotics (final concentration in water:100 mg/mL and 0.75 mg/mL, respectively).Mix well and pour into one side of acompartmentalized-plate. The plates arewrapped in aluminum foil and stored at4°C until needed.
Preparation of LB agar plates. These areused for culturing bacteria. Plain LB agarplates are prepared by mixing 25 g of LB,17 g of agar in 1 L of double distilled waterand the solution is autoclaved. A separatebatch of LB agar plates supplemented withrifampicin (75 mg/mL) is prepared to testfor the presence of Photorhabdus bacteriain axenic worms. Rifampicin plates shouldbe kept in the dark. All plates are storedat 4°C.
Counting nematodes. Agitate a flaskcontaining nematodes in suspension andpipette ten individual drops (aliquots) of50 mL each of water-containing nematodeson a Petri dish; count the number ofworms in each drop using a stereo-microscope. Calculate the total numberof worms by adding all ten counts andmultiply by 20 to obtain X number ofnematodes per mL. Use the number ofnematodes/mL to calculate doses.
Precautions. (1) All surfaces shouldbe sterilized with 10% bleach to avoidcontaminations.
(2) Forceps and brushes that are usedto manipulate flies should be sterilized for
6 min using an UV trans-illuminator.Alternatively, forceps should be cleanedwith 70% ethanol.
(3) Add water to bi-plates under sterileconditions.
(4) All contaminated materials shouldbe discarded in a biohazard container forproper disposal.
(5) All solutions should be autoclavedunless otherwise indicated and antibioticsshould always be filter sterilized.
(6) Wear gloves at all times.
Procedure
Culturing symbiotic nematodes: Timing14–20 d. To grow and amplify Hetero-rhabditis symbiotic nematodes, we usecaterpillars of the greater wax mothGalleria mellonella. The larvae are infectedwith nematodes carrying P. luminescensstrain TT01 bacteria.
(1) Plate preparation: 1 h. Cover thebottom of a large Petri dish (150 mmdiameter) with a single filter paper, add1 mL of sterile water and transfer 6–10Galleria larvae. Pipette a drop of watercontaining 50–100 IJ on each insect.
(2) Incubation: 11 d. Galleria larvaesuccumb to nematodes 2 d after theinfection. Heterorhabditis/Photorhabdusinfected Galleria larvae exhibit a charac-teristic “red brick” color (Fig. 2A). Insectsturning black or gray are contaminatedby other pathogens and they should beimmediately discarded. Monitor thesurvival of larvae daily and add a fewdrops of water, if needed, to keep theinsect cadavers moist but not soaked.
(3) Water trap preparation: 1 h. On day11 after infection, transfer the Gallerialarvae to a water trap. This consists ofa large Petri dish (150 � 20 mm), inside ofwhich a smaller standard size Petri dish(100 � 15 mm) is placed upside down.We place a piece of filter paper (cut asshown on Fig. 2B) on top of the smallPetri dish and fill the large Petri dish withwater until the edges of the filter paperare submerged.
(4) Insect cadavers as prey for nematodes:10 min. Using a pair of plastic forceps,gently transfer and arrange the infectedGalleria larvae on the filter paper of thewater trap. Place the traps in a Rubbermaidcontainer to prevent contamination of the
Figure 2. Water trap setup. (A) Galleria mellonella cadavers are arranged on top of the filter paper(note the characteristic “red brick” color of nematode infected larvae). Aerial (Top) and lateral(bottom) views showing the insect larvae arranged on top of the cut filter paper centered inthe Petri dish. Note the arrangement of the small Petri dish, centered in the large Petri dish,the position of the filter paper and the edge of the paper making direct contact with the water.(B) Filter papers are folded and cut in the corners to create “projections” that will be in contact withthe water. This facilitates the transfer of moisture and the migration of Heterorhabditis IJ fromthe insect cadavers to the water trap.
342 Virulence Volume 3 Issue 3

©20
12 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
cadavers by other pathogens. Store in thedark until the IJ emerge from the insectcadavers.
(5) Nematode harvesting: 30 min.Nematodes usually emerge from the deadinsects 14–20 d after infection. Emergingnematodes directly fall into the waterallowing for easy collection. They can bereadily collected from the water trap usinga 100 mL graduated cylinder. Fill thecylinder to the top (100 mL mark) withwater containing nematodes and let theworms settle at the bottom (this takesapproximately 30 min). Remove 85 mL ofwater (down to the 15 mL mark) usinga volumetric pipette. Fill back up withfresh water to the 100 mL mark, let thenematodes settle, and remove the excesswater as indicated above. Repeat threetimes until the water in the cylinder isclear. Transfer 20 mL aliquots in smallculture flasks, and store in the dark.Nematodes can be collected daily for atleast 2 weeks or longer depending on thenumber of worms required for experiments.
NOTE: It is important to achieveoptimum numbers of nematodes in eachflask: high density will result in nematodeaggregations and eventually nematodedeath; low density will not produce theappropriate number of worms requiredfor the experiments. We keep around1,500–2,000 IJ/mL of water. Shake flasksregularly to aid aeration. Nematodes can bestored at room temperature (RT) (~25°C)for months. We always use less than1-mo-old nematodes in our experiments.
Culturing axenic nematodes: timing14–20 d. Axenic nematodes (nematodeslacking Photorhabdus bacteria) are culturedin bi-plates on a bacterial lawn producedby P. temperata mutant strain RET16.
(6) Bacterial inoculation and plating:18 h. Grow P. temperata RET16 bacteriain LB broth at 30°C with vigorous agita-tion for 18 h; transfer 500 mL of pureculture to LB agar bi-plates supplementedwith 100 mg/mL of carbenicillin and0.75 mg/mL of gentamicin. Incubate platesfor 48 h at 30°C to form a bacterial lawn.
(7) Incubation of nematodes mixed withbacterial lawn: 30 min–12 d. Transfer500–1,000 surface-sterilized IJ to thecenter of the bacterial lawn; gentlyagitate plates to distribute the wormsevenly and incubate at 30°C for 12 d.
NOTE: A container filled with sterilewater is placed in the incubator to preventdesiccation. Sterile water (approximately400 mL) is also added to the platesafter 1 week (or whenever necessary), tomoisten the agar.
(8) Water trap setup: 10 min. After 12 dof incubation, fill the other half of thebi-plate with sterile water and incubate atRT to allow emerging IJ to fall into thewater trap as they migrate from the agar tothe water.
(9) Nematode collection: 1 h. After 14–20 d, IJ nematodes are collected from thewater traps. Aspirate the water using a10 mL disposable pipette and replace withfresh water. Transfer the water containingnematodes to a culture flask. Washnematodes several times with sterile waterand store flasks in the dark. Nematodesshould always be surface sterilized beforeuse, as described above.
(10) Testing axenicity: 2–3 d. To deter-mine whether axenic Heterorhabditis con-tain Photorhabdus cells, pellet a sampleof newly generated axenic nematodes(200 mL of a nematode suspension in a1.5 mL tube) for 5 min at 8,000 rpm.Remove the supernatant and homogenizethe worms using a plastic pestle. Add100 mL of LB broth, mix and plate out thesolution on LB agar plates supplementedwith rifampicin (see above); incubate for2 d at 30°C. The absence of bacterialcolonies indicates that no P. luminescenscells are present in the worms.
Surface sterilization of nematodes:timing 1 h. (11) Pelleting nematodes: 10–15 min. Centrifuge 20 mL of a nematodesuspension in a 50 mL conical tube at1,500 rpm (standard speed used in allsteps) for 5 min; discard the supernatant.
(12) Bleach treatment: 10–15 min.Resuspend the nematode pellet in 20 mLof 0.1% bleach solution in water, gentlyinvert the tube several times, incubate for7 min at RT and centrifuge for 5 min at1,500 rpm. Remove the bleach solutionleaving the pellet in approximately 1 mLof solution left, add fresh water (40 mL),mix gently and repeat centrifugationstep. Repeat four times. Nematodes areresistant to low concentrations of bleach.We did not observe any negative effectsof bleach on the fitness or survival ofthe nematodes.
(13) Washes: 25 min. Add 20 mL of 1xPBS, invert the tube three times andcentrifuge for 5 min. Repeat this stepthree times. Finally, the nematodes areresuspended in sterile water or 0.85% ofNaCl and stored in sterile flasks untilneeded.
Presence of bacteria in infected flies(survival experiment controls): timing20 h. Positive and negative controls inthe survival experiments include fliesinjected with pathogenic P. luminescensand non-pathogenic E. coli bacteria,respectively.
(14) Bacterial growth: 18 h. Inoculatebacteria from frozen glycerol stocks in10 mL of LB broth and incubate withconstant agitation (250 rpm) for 18 hat 30°C for P. luminescens and 37°C forE. coli.
(15) Wash bacteria: 45 min. Centrifugebacterial cultures for 5 min at 5,000 rpmto pellet the bacterial cells; discardthe supernatant. Wash the pellet twiceusing sterile 1x PBS and centrifuge for5 min at 5,000 rpm; resuspend thepellet in 7 mL of 1x PBS. Measure theoptical density (OD) using a spectro-photometer. Adjust the concentrationat an OD of approximately 0.1 to obtainadequate cell density.
Bacterial injection (survival experimentcontrols): 1–2 h. We use a nanoinjectorand glass capillary needles for deliveringprecise doses of bacteria into flies.
(16) Preparation of capillary needles:10 min. Secure a borosilicate needle tothe pipette puller and press the “Pull”button. Pipette puller settings are: ramp:561, velocity: 20, heat: 560, delay: 1, pull:100, pressure: 500. The needle tip ismanually broken under the stereoscopeusing a pair of clean forceps. The capillaryis then filled with mineral oil using aplastic syringe and 25G gauge needle.
(17) Injector assembly: 3 min. Attach theneedle to the injector by connectingall o-rings and securing the cap tightly.Pipette 50 mL of washed bacterial cells ofthe appropriate concentration on a pieceof parafilm. Fill the needle by pressing the“Fill” button.(18) Injections: 1.5–2 h. Place the flies
on a fly pad and anesthetize them withCO2; flies are arranged with a paintbrushand injected with 18.6 nl of a bacterial
www.landesbioscience.com Virulence 343

©20
12 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
suspension corresponding to an OD of0.1 (approximately 400–700 CFUs forP. luminescens and 1,000–4,000 CFUs forE. coli). Transfer infected flies to freshvials and monitor their survival daily andup to 7 d.
Nematode infection assays: 5–7 d. (19)Preparation of infection units: 45 min–1 h.Cover the bottom of the cups with 4–5pieces of filter paper; use the bottom of thecup as measurement.
(20) Mixing nematodes and flies: 15 min.Surface sterilized nematodes in solution(100 IJ/fly) are pipetted to each infectionunit. Flatten the filter paper at the bottomof the cup using a micropipette tip. Addmore water to the filter paper as needed(usually in aliquots of 100 mL) until it isattached to the bottom of the cup; thisstep is important because extra water keepsthe nematode parasites active.
Tip: For four layers of filter paper, themaximum total volume of added water isapproximately 600 mL; for five filter papersit is about 800 mL. Once nematodes andflies are added to the cup, a second emptycup is placed on top and secured usingtape (Fig. 1). Keep the infection units atRT in a humid box (e.g., a pipette tip boxlined with wet filter paper).
NOTE: It is important to preventdesiccation. The addition of extra waterto the cups depends on nematode density:the higher the density, the more waterneeds to be added, and vice versa. Excesswater will result in fly drowning; lackof water will result in desiccation andnematode death. It is also important toflatten the filter papers at the bottom ofthe cup to avoid trapping of flies inbetween the filter papers. Cups should beprepared 15–30 min before use.
Survival experiments: 7 d. (21) Set up:30 min. Infect 4–6 d old OregonR adultflies with 100 IJ/fly as described above.
(22) Monitoring survival: 7 d. Countand record the number of dead flies daily.
Dose-response survival experiments: 7 d.(23) Set up: 30–45 min. Infect 4–6 d old
OregonR adult flies with 10, 25, 50 and 100nematode IJ/fly. Estimate nematodenumbers as described above.
(24) Monitoring survival: 7 d. Incubateinfected and uninfected control flies for7 d at RT and count the number of deadflies daily.
Persistence experiments: 2 d. (25) Setup: 30 min. Infect 4–6 d old OregonR
adult flies with 100 IJ/fly as describedabove. Incubate flies at RT.
(26) Nematode counting: 2 d. At 24 and48 h post-infection, wash infected (dead oralive) flies in a drop of 1x PBS to removeany nematodes attached to the exterior ofthe flies (use the microscope). Dissect fliesin 1x PBS using a pair of clean forcepsto determine the number of nematodespresent in each fly.
Nematode development: 5 d. Todetermine whether Drosophila adult fliesare suitable hosts for supporting thedevelopment of Heterorhabditis nema-todes, we infect flies with symbioticnematodes (50 IJ/fly), as described above.We have observed that infection with ahigh number of nematodes slows downnematode development, probably due tocrowding conditions in the fly.
(27) Set up: 15 min. Infect Drosophilaadult flies as previously described andincubate insects at RT.
(28) Dissection and fixation: 1–2 h. Fivedays after infection, dissect flies in sterile1x PBS. Incubate dissected cadavers incold 4% paraformaldehyde diluted in 1xPBS, in a 96-well plate for 1 h. Thistreatment eliminates any remaining livenematodes. Gently remove the fixativesolution and wash samples three times in
1x PBS. Carefully mount the dissectedflies on a glass slide using 50% glyceroldiluted in 1x PBS solution, and coverthe samples with a coverslip. Specimensare ready for viewing under the micro-scope. Slides can be stored at 4°C for afew days.
(29) Imaging: flexible. Time neededfor viewing depends on the number ofprepared samples and experience of theresearcher with microscopy techniques.Images are assembled and illustrationsprepared using Adobe Illustrator andPhotoshop software packages (AdobeCS5 suite).
Results and Further Studies
We first infected Drosophila WT adultflies with Heterorhabditis nematodescarrying or lacking Photorhabdus bacteriaand estimated the ability of flies to surviveinfection over time (Fig. 3). We foundthat in both cases flies succumbed tonematode infection after 5–6 d. Survival offlies infected with axenic or symbioticnematodes was highly significant (p ,0.05; Chi-square test) compared withuninfected controls. High fly mortalitycaused by axenic nematodes merits furtherinvestigation and it will be the focus of ourfuture studies. Injection of Drosophilawith Photorhabdus (positive control) also
Figure 3. Survival results after infection of Drosophila wild-type (WT) adult flies with axenicHeterorhabditis (nematodes lacking Photorhabdus), symbiotic Heterorhabditis (nematodes carryingPhotorhabdus) or Photorhabdus bacteria alone. Each replicate consisted of 10 Oregon 4–6-d-oldflies infected with approximately 1,000 nematodes (100 IJ per fly). Flies were injected with 18.4 nlof PBS buffer containing approximately 500 Photorhabdus cells. Infections with E. coli bacteria(500 cells) and injections with PBS buffer alone served as controls. Results showed that both axenicand symbiotic nematodes were able to kill adult flies at a similar rate. Survival of flies wasmonitored daily and up to day six following infection. Data represent the percent survival ofinfected and control flies. The averages from at least three experiments are shown.
344 Virulence Volume 3 Issue 3

©20
12 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
killed all flies within the same length oftime. Injection of flies with E. coli bacteriaor uninfected flies (negative controls)produced minimum insect mortality.
We then infected Drosophila adultflies with different numbers of axenicor symbiotic Heterorhabditis IJ. Interest-ingly, we found that flies infected with25 or 100 axenic Heterorhabditis diedslightly faster (3 d after infection) com-pared with those infected with 10 or50 axenic worms (4 d after infection)(Fig. 4A). Different number of axenicnematodes caused mortality that was signi-ficantly higher compared with uninfectedcontrols (p , 0.05; Dunnett’s multiplecomparison test). One explanation forthis discrepancy could be variation innematode seeking behavior or increasedfly activity. We have found that hyperac-tive flies significantly lower Heterorhabditisinfectivity (unpublished data). However,we observed that all flies infected withdifferent numbers of symbiotic Heter-orhabditis succumbed within 4 d ofinfec>tion (Fig. 4B). Again, mortality ofuninfected control flies was significantlylower (p , 0.05; Dunnett’s multiple com-parison test) compared with mortality offlies infected with symbiotic nematodes.
We further looked at the persistence ofaxenic and symbiotic Heterorhabditis inDrosophila WT flies at 24 and 48 h afterinfection (Fig. 5). We found that bothgroups of nematodes were present in fliesat both time-points, which suggests thatDrosophila adults can serve as valuablemodel for studying host-parasite interac-tions and nematode parasitism. We didnot observe any significant difference inthe number of axenic or symbiotic wormsbetween the two time-points (p , 0.5,df = 3; one-way ANOVA)
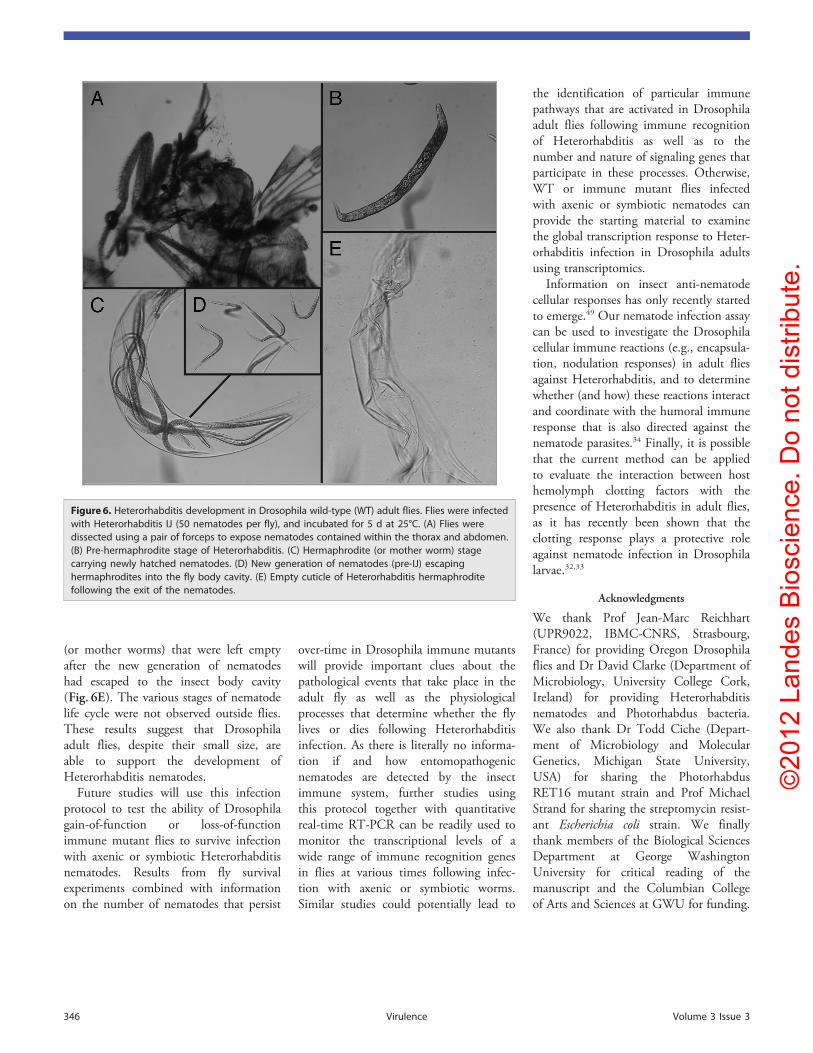
Finally, we asked whether Drosophilaadult flies permit nematode development.For this, we used Heterorhabditis IJ toinfect WT flies that were dissected 5 dlater (Fig. 6A). We found that infectedflies contained nematodes at differentstages of development; these includedpre-hermaphrodite nematodes (Fig. 6B),hermaphrodites harboring newly hatchedL1 nematodes (Fig. 6C), and pre-IJ wormsthat disrupt the nematode cuticle andmove to the fly hemocoel (Fig. 6D). Wealso observed cuticles of hermaphrodites
Figure 4. Dose-response survival results for Drosophila wild-type (WT) adult flies infected with(A) axenic Heterorhabditis (nematodes lacking Photorhabdus) and (B) symbiotic Heterorhabditis(nematodes carrying Photorhabdus). We used 10, 25, 50 and 100 IJ per fly. Ten Oregon 4–6-d-oldflies were used for each treatment. Uninfected flies served as controls. Survival of flies wasestimated daily and up to day seven following infection. Data represent the percent survival ofinfected and control flies. The averages from at least three experiments are shown.
Figure 5. Persistence of axenic and symbiotic Heterorhabditis in Drosophila adult flies. Ten Oregon4–6-d-old flies were used in each replicate. Flies were infected with approximately 1,000 nematodes(100 nematodes per fly), and dissected at 24 and 48 h after infection. Nematode numbers werecounted under a dissecting microscope. Colored horizontal lines indicate the arithmetic means ofnematode numbers and error bars indicate standard errors. Data represent the averages from atleast three independent experiments.
www.landesbioscience.com Virulence 345
©20
12 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
(or mother worms) that were left emptyafter the new generation of nematodeshad escaped to the insect body cavity(Fig. 6E). The various stages of nematodelife cycle were not observed outside flies.These results suggest that Drosophilaadult flies, despite their small size, areable to support the development ofHeterorhabditis nematodes.
Future studies will use this infectionprotocol to test the ability of Drosophilagain-of-function or loss-of-functionimmune mutant flies to survive infectionwith axenic or symbiotic Heterorhabditisnematodes. Results from fly survivalexperiments combined with informationon the number of nematodes that persist
over-time in Drosophila immune mutantswill provide important clues about thepathological events that take place in theadult fly as well as the physiologicalprocesses that determine whether the flylives or dies following Heterorhabditisinfection. As there is literally no informa-tion if and how entomopathogenicnematodes are detected by the insectimmune system, further studies usingthis protocol together with quantitativereal-time RT-PCR can be readily used tomonitor the transcriptional levels of awide range of immune recognition genesin flies at various times following infec-tion with axenic or symbiotic worms.Similar studies could potentially lead to
the identification of particular immunepathways that are activated in Drosophilaadult flies following immune recognitionof Heterorhabditis as well as to thenumber and nature of signaling genes thatparticipate in these processes. Otherwise,WT or immune mutant flies infectedwith axenic or symbiotic nematodes canprovide the starting material to examinethe global transcription response to Heter-orhabditis infection in Drosophila adultsusing transcriptomics.
Information on insect anti-nematodecellular responses has only recently startedto emerge.49 Our nematode infection assaycan be used to investigate the Drosophilacellular immune reactions (e.g., encapsula-tion, nodulation responses) in adult fliesagainst Heterorhabditis, and to determinewhether (and how) these reactions interactand coordinate with the humoral immuneresponse that is also directed against thenematode parasites.34 Finally, it is possiblethat the current method can be appliedto evaluate the interaction between hosthemolymph clotting factors with thepresence of Heterorhabditis in adult flies,as it has recently been shown that theclotting response plays a protective roleagainst nematode infection in Drosophilalarvae.32,33
Acknowledgments
We thank Prof Jean-Marc Reichhart(UPR9022, IBMC-CNRS, Strasbourg,France) for providing Oregon Drosophilaflies and Dr David Clarke (Department ofMicrobiology, University College Cork,Ireland) for providing Heterorhabditisnematodes and Photorhabdus bacteria.We also thank Dr Todd Ciche (Depart-ment of Microbiology and MolecularGenetics, Michigan State University,USA) for sharing the PhotorhabdusRET16 mutant strain and Prof MichaelStrand for sharing the streptomycin resist-ant Escherichia coli strain. We finallythank members of the Biological SciencesDepartment at George WashingtonUniversity for critical reading of themanuscript and the Columbian Collegeof Arts and Sciences at GWU for funding.
Figure 6. Heterorhabditis development in Drosophila wild-type (WT) adult flies. Flies were infectedwith Heterorhabditis IJ (50 nematodes per fly), and incubated for 5 d at 25°C. (A) Flies weredissected using a pair of forceps to expose nematodes contained within the thorax and abdomen.(B) Pre-hermaphrodite stage of Heterorhabditis. (C) Hermaphrodite (or mother worm) stagecarrying newly hatched nematodes. (D) New generation of nematodes (pre-IJ) escapinghermaphrodites into the fly body cavity. (E) Empty cuticle of Heterorhabditis hermaphroditefollowing the exit of the nematodes.
346 Virulence Volume 3 Issue 3

©20
12 L
ande
s B
iosc
ienc
e. D
o no
t dis
tribu
te.
References1. Vallet-Gely I, Lemaitre B, Boccard F. Bacterial
strategies to overcome insect defences. Nat RevMicrobiol 2008; 6:302-13; PMID:18327270; http://dx.doi.org/10.1038/nrmicro1870
2. O’Callaghan D, Vergunst A. Non-mammalian animalmodels to study infectious disease: worms or fly fishing?Curr OpinMicrobiol 2010; 13:79-85; PMID:20045373;http://dx.doi.org/10.1016/j.mib.2009.12.005
3. Limmer S, Quintin J, Hetru C, Ferrandon D. Virulenceon the fly: Drosophila melanogaster as a model geneticorganism to decipher host-pathogen interactions. CurrDrug Targets 2011; 12:978-99; PMID:21366519;http://dx.doi.org/10.2174/138945011795677818
4. Apidianakis Y, Rahme LG. Drosophila melanogaster as amodel for human intestinal infection and pathology.Dis Model Mech 2011; 4:21-30; PMID:21183483;http://dx.doi.org/10.1242/dmm.003970
5. Glavis-Bloom J, Muhammed M, Mylonakis E. Ofmodel hosts and man: using Caenorhabditis elegans,Drosophila melanogaster and Galleria mellonella asmodel hosts for infectious disease research. Adv ExpMed Biol 2012; 710:11-7; PMID:22127881; http://dx.doi.org/10.1007/978-1-4419-5638-5_2
6. Vlisidou I, Waterfield N, Wood W. Elucidating the invivo targets of photorhabdus toxins in real-time usingDrosophila embryos. Adv Exp Med Biol 2012; 710:49-57; PMID:22127885; http://dx.doi.org/10.1007/978-1-4419-5638-5_6
7. Schneider D. Using Drosophila as a model insect. NatRev Genet 2000; 1:218-26; PMID:11252751; http://dx.doi.org/10.1038/35042080
8. Charroux B, Royet J. Drosophila immune response:From systemic antimicrobial peptide production in fatbody cells to local defense in the intestinal tract. Fly(Austin) 2010; 4:40-7; PMID:20383054
9. Lemaitre B, Hoffmann J. The host defense ofDrosophila melanogaster. Annu Rev Immunol 2007;25:697-743; PMID:17201680; http://dx.doi.org/10.1146/annurev.immunol.25.022106.141615
10. Hetru C, Hoffmann JA. NF-kappaB in the immuneresponse of Drosophila. Cold Spring Harb Perspect Biol2009; 1:a000232; PMID:20457557; http://dx.doi.org/10.1101/cshperspect.a000232
11. Ganesan S, Aggarwal K, Paquette N, Silverman N. NF-kB/Rel proteins and the humoral immune responsesof Drosophila melanogaster. Curr Top MicrobiolImmunol 2011; 349:25-60; PMID:20852987; http://dx.doi.org/10.1007/82_2010_107
12. Leulier F, Lemaitre B. Toll-like receptors–taking anevolutionary approach. Nat Rev Genet 2008; 9:165-78;PMID:18227810; http://dx.doi.org/10.1038/nrg2303
13. Kawai T, Akira S. The role of pattern-recognitionreceptors in innate immunity: update on Toll-likereceptors. Nat Immunol 2010; 11:373-84; PMID:20404851; http://dx.doi.org/10.1038/ni.1863
14. Valanne S, Wang JH, Rämet M. The Drosophila Tollsignaling pathway. J Immunol 2011; 186:649-56;PMID:21209287; http://dx.doi.org/10.4049/jimmunol.1002302
15. Chtarbanova S, Imler JL. Microbial sensing by Tollreceptors: a historical perspective. Arterioscler ThrombVasc Biol 2011; 31:1734-8; PMID:21775770; http://dx.doi.org/10.1161/ATVBAHA.108.179523
16. Uvell H, Engström Y. A multilayered defense againstinfection: combinatorial control of insect immune genes.Trends Genet 2007; 23:342-9; PMID:17532525; http://dx.doi.org/10.1016/j.tig.2007.05.003
17. Lemaitre B, Nicolas E, Michaut L, Reichhart JM,Hoffmann JA. The dorsoventral regulatory genecassette spätzle/Toll/cactus controls the potent anti-fungal response in Drosophila adults. Cell 1996;86:973-83; PMID:8808632; http://dx.doi.org/10.1016/S0092-8674(00)80172-5
18. Georgel P, Naitza S, Kappler C, Ferrandon D, ZacharyD, Swimmer C, et al. Drosophila immune deficiency(IMD) is a death domain protein that activatesantibacterial defense and can promote apoptosis. DevCell 2001; 1:503-14; PMID:11703941; http://dx.doi.org/10.1016/S1534-5807(01)00059-4
19. Imler JL, Bulet P. Antimicrobial peptides in Drosophila:structures, activities and gene regulation. ChemImmunol Allergy 2005; 86:1-21; PMID:15976485;http://dx.doi.org/10.1159/000086648
20. Ferrandon D, Imler JL, Hetru C, Hoffmann JA. TheDrosophila systemic immune response: sensing andsignalling during bacterial and fungal infections. NatRev Immunol 2007; 7:862-74; PMID:17948019;http://dx.doi.org/10.1038/nri2194
21. De Gregorio E, Spellman PT, Tzou P, Rubin GM,Lemaitre B. The Toll and Imd pathways are the majorregulators of the immune response in Drosophila.EMBO J 2002; 21:2568-79; PMID:12032070;http://dx.doi.org/10.1093/emboj/21.11.2568
22. Agaisse H, Perrimon N. The roles of JAK/STATsignaling in Drosophila immune responses. ImmunolRev 2004; 198:72-82; PMID:15199955; http://dx.doi.org/10.1111/j.0105-2896.2004.0133.x
23. Delaney JR, Stöven S, Uvell H, Anderson KV,Engström Y, Mlodzik M. Cooperative control ofDrosophila immune responses by the JNK and NF-kappaB signaling pathways. EMBO J 2006; 25:3068-77; PMID:16763552; http://dx.doi.org/10.1038/sj.emboj.7601182
24. Meister M. Blood cells of Drosophila: cell lineages androle in host defence. Curr Opin Immunol 2004; 16:10-5; PMID:14734104; http://dx.doi.org/10.1016/j.coi.2003.11.002
25. Williams MJ.Drosophila hemopoiesis and cellular immu-nity. J Immunol 2007; 178:4711-6; PMID:17404248
26. Tang H. Regulation and function of the melanizationreaction in Drosophila. Fly (Austin) 2009; 3:105-11;PMID:19164947
27. Eleftherianos I, Revenis C. Role and importance ofphenoloxidase in insect hemostasis. J Innate Immun2011; 3:28-33; PMID:21051882; http://dx.doi.org/10.1159/000321931
28. Davies SA, Dow JA. Modulation of epithelial innateimmunity by autocrine production of nitric oxide. GenComp Endocrinol 2009; 162:113-21; PMID:18952086;http://dx.doi.org/10.1016/j.ygcen.2008.09.012
29. Bae YS, Choi MK, Lee WJ. Dual oxidase in mucosalimmunity and host-microbe homeostasis. TrendsImmunol 2010; 31:278-87; PMID:20579935; http://dx.doi.org/10.1016/j.it.2010.05.003
30. Ryu JH, Ha EM, Lee WJ. Innate immunity and gut-microbe mutualism in Drosophila. Dev CompImmunol 2010; 34:369-76; PMID:19958789; http://dx.doi.org/10.1016/j.dci.2009.11.010
31. Hallem EA, Rengarajan M, Ciche TA, Sternberg PW.Nematodes, bacteria, and flies: a tripartite model fornematode parasitism. Curr Biol 2007; 17:898-904;PMID:17475494; http://dx.doi.org/10.1016/j.cub.2007.04.027
32. Wang Z, Wilhelmsson C, Hyrsl P, Loof TG, Dobes P,Klupp M, et al. Pathogen entrapment by transgluta-minase–a conserved early innate immune mechanism.PLoS Pathog 2010; 6:e1000763; PMID:20169185;http://dx.doi.org/10.1371/journal.ppat.1000763
33. Hyrsl P, Dobes P, Wang Z, Hauling T, WilhelmssonC, Theopold U. Clotting factors and eicosanoidsprotect against nematode infections. J Innate Immun2011; 3:65-70; PMID:20948189; http://dx.doi.org/10.1159/000320634
34. Castillo JC, Reynolds SE, Eleftherianos I. Insectimmune responses to nematode parasites. TrendsParasitol 2011; 27:537-47; PMID:21982477; http://dx.doi.org/10.1016/j.pt.2011.09.001
35. Ciche T. The biology and genome of Heterorhabditisbacteriophora. WormBook 2007; 1-9; PMID:18050499
36. ffrench-Constant RH, Eleftherianos I, Reynolds SE.Nematode symbionts shed light on invertebrate immu-nity. Trends Parasitol 2007; 23:514-7; PMID:17964855;http://dx.doi.org/10.1016/j.pt.2007.08.021
37. Waterfield NR, Ciche T, Clarke D. Photorhabdus and ahost of hosts. Annu Rev Microbiol 2009; 63:557-74;PMID:19575559; http://dx.doi.org/10.1146/annurev.micro.091208.073507
38. Ciche TA, Ensign JC. For the insect pathogenPhotorhabdus luminescens, which end of a nematode isout? Appl Environ Microbiol 2003; 69:1890-7; PMID:12676661; http://dx.doi.org/10.1128/AEM.69.4.1890-1897.2003
39. ffrench-Constant RH, Dowling A, Waterfield NR.Insecticidal toxins from Photorhabdus bacteria andtheir potential use in agriculture. Toxicon 2007; 49:436-51; PMID:17207509; http://dx.doi.org/10.1016/j.toxicon.2006.11.019
40. ffrench-Constant R, Waterfield N, Daborn P, Joyce S,Bennett H, Au C, et al. Photorhabdus: towards afunctional genomic analysis of a symbiont andpathogen. FEMS Microbiol Rev 2003; 26:433-56;PMID:12586390; http://dx.doi.org/10.1111/j.1574-6976.2003.tb00625.x
41. Joyce SA, Watson RJ, Clarke DJ. The regulation ofpathogenicity and mutualism in Photorhabdus. CurrOpin Microbiol 2006; 9:127-32; PMID:16480919;http://dx.doi.org/10.1016/j.mib.2006.01.004
42. Goodrich-Blair H, Clarke DJ. Mutualism and patho-genesis in Xenorhabdus and Photorhabdus: two roads tothe same destination. Mol Microbiol 2007; 64:260-8;PMID:17493120; http://dx.doi.org/10.1111/j.1365-2958.2007.05671.x
43. Clarke DJ. Photorhabdus: a model for the analysis ofpathogenicity and mutualism. Cell Microbiol 2008;10:2159-67; PMID:18647173; http://dx.doi.org/10.1111/j.1462-5822.2008.01209.x
44. Eleftherianos I, ffrench-Constant RH, Clarke DJ,Dowling AJ, Reynolds SE. Dissecting the immuneresponse to the entomopathogen Photorhabdus. TrendsMicrobiol 2010; 18:552-60; PMID:21035345; http://dx.doi.org/10.1016/j.tim.2010.09.006
45. Tian C, Gao B, Rodriguez MdelC, Lanz-Mendoza H,Ma B, Zhu S. Gene expression, antiparasitic activity,and functional evolution of the drosomycin family.Mol Immunol 2008; 45:3909-16; PMID:18657321;http://dx.doi.org/10.1016/j.molimm.2008.06.025
46. Fellous S, Lazzaro BP. Larval food quality affects adult(but not larval) immune gene expression independentof effects on general condition. Mol Ecol 2010;19:1462-8; PMID:20196811; http://dx.doi.org/10.1111/j.1365-294X.2010.04567.x
47. Fellous S, Lazzaro BP. Potential for evolutionarycoupling and decoupling of larval and adult immunegene expression. Mol Ecol 2011; 20:1558-67; PMID:21299661; http://dx.doi.org/10.1111/j.1365-294X.2011.05006.x
48. Shia AK, Glittenberg M, Thompson G, Weber AN,Reichhart JM, Ligoxygakis P. Toll-dependent anti-microbial responses in Drosophila larval fat body requireSpätzle secreted by haemocytes. J Cell Sci 2009; 122:4505-15; PMID:19934223; http://dx.doi.org/10.1242/jcs.049155
49. Eleftherianos I, Joyce S. ffrench-Constant RH, ClarkeDJ, Reynolds SE. Probing the tri-trophic interactionbetween insects, nematodes and Photorhabdus.Parasitology 2010; 26:1-12.
www.landesbioscience.com Virulence 347
Copyright © 2022 FDOKUMEN




















