
Six First Reports of Pin Nematodes from Portugal, with ... - MDPI
Upload
khangminh22Category
view
1download
0
STUDIES ON THE BEHAVIOUR OF NEMATODES
SUMM4RY
THESIS^UBMJTTED FOR THE DEGREE OF
©ottor of ^timtt IN
ZOOLOGY ALIGARH MUSLIM UNIVERSITY, ALIGARH
BW
ANWAR LUQMAN BILGRAMI Ph.D.
T'-^i^^ 'A ^j
DEPARTMENT OF ZOOLOGY ALIGARH MUSLIM UNIVERSITY
ALIGARH (INDIA) DECEMBER, 19 9 3

^A^^^
STUDIES ON THE BEHAVIOUR OF NEMATODES
THESIS SUBMJTTED FOR THE DEGREE OF
Bottor of ^titxiu IN
ZOOLOGY ALIGARH MUSLIM UNIVERSITY, ALIGARH
BY
KHV^NR LUQMAN BILGRAMI Ph. D.
DEPARTMENT OF ZOOLOGY ALIGARH MUSLIM UNIVERSITY
ALIGARH (INDIA) DECEMBER, 19 9 3

SUiM.MARY
The thesiE de^ls with the studies on certain
aspects of nematode behaviour v i z . , predatory
behaviour of nematodes, attrsction and feeding
behaviour of plant parasitic nematodes, effect of
cheroicais on the activity and survival of nematodes,
developmental biology, sex attraction etc.
Observations were made on different aspects of
predation such as attraction of predators towards
prey, pre- and post feeding attraction and aggregation
of predators at feeding sites, prey catching and
feeding mechanisms, prey selection., factors
influencing predation and attraction, food preference,
strike rate of the predators and resistance and
susceptibility of prey nematodes- to predation,
relationship between predation and . different prey
trophic categories, cannibalism, predator—predator
interaction etc. These studies were made on different
species of predators belonging to the Orders
Mononchida <Mvlonchulus dent atus, MyIonchulus
mi nor < MonanchvB a go ati cus) and Dipiogasterida
(Mononchoi des 1 onqi caudabtts , Mononchoi des f or t i dens ) ;
Sub orders Dorylaimina < Dor yl a i 'nits st-aqnaljs,
Aporre] ai me 1 1 u " n i v g lis, M p* s o d o r y 1 ^ i B t' •" biHsti ani ,

Pi scolai mus si 1vi col u s , Allodorylai mu s ameri canus)
and Nygolaimina (Aquat i des thornei > Super -family
Actinolai fljoidea (Neoact i nolai mus aqilis) and Family
Thaiassogeneridae (Thaiossoqenus shami mi ) .
Observations on the attraction o-f predators
towards prey rG\'eBl sd that the predators were
attracted towards excised and non excised prey in
response to pi^ey seer et i ons/at tract an t s-
Dipiogasterid predators also responded towards agar
containing una dent i-fi ed bacteria- The degree of
attraction o-f predators varied from prey to prey.
Some species of prey attracted predators more
(Hirschmanniel1 a oryz ae Tylenchorhynchus mashoodi >
than the others <HeIicotylenchus i n d i c u s) . Predators
were also attracted towards their own individuals.
Different factors' v i z . , prey number, temperature,
incubation period of prey, starvation of predators and
distance of predators from prey affected attraction of
predators towards prey.
Predators also e>;hibited pre- and post feeding
attraction and aggregation around prey at feeding
sites. Mc-e than one predictor was found attracted and
aggregated around the prey at feeding sites. This
phi?i»D,0enun of aggr t-g a t-i o,( is ;nore prominent m

3
m diplogasterid predators than the dorylaim or nygolai
predators. The phenomenon of aggregation of predators
at feeding sites has been attributed to
secretions/attractants released by the prey;
secretions from the predators at the time of feeding
or secretions of both the prey and predators. The
post feeding attraction or aggregation has been
attributed mainly to the lingering effects of the
prey/predator secretions/attractants.
The prey catching and feeding mechanisms of
majority of predators belonging to different groups of
nematodes may conveniently be divided into five more
or less distinct phases viz., encounter with prey,
a t t a c k r e s p o n s e , attack ,, sal i vati on/extra-corporeal
digestion and ingestion/feeding. All phases are quite
different in different species of predators belonging
to different orders and suborders. There is, however,
minute differences in these mechanisms as far as
closely related species of predators are concerned.
Many species of predators except mononchs showed
preference for different type of prey. Most predators
preferred the second stage juveniles of endo-parasitic
nematodes viz.. Aerobe!oi des 5p.^ Cephalobus sp.,
Meloidogyne incogi^i ta, Hel: fr orler a mothi Ancn<i nR

tritici » Tylenchulus sgmi pgnetrans etc. , but failed to
injure or kill sny af Hoplolai mus i ndi cus,
Hemi cr i congmoi des manqi ferse, Hemi cycliophora dhi ren —
deri Scutel 1 onema sp. Feeding by mononchs is
poiyvalant and al esttory as they -fed on all types of
prey.
The study on the gut content o-f over 1000
specimens of mononchs showed that they feed upon
tylenchs, dorylaims, mononchs and free living
saprophagus nematodes besides rotifers. Forty five
percent of the predators had prey in their intestine.
More <75X> predators had free living saprophagus
nematodes within the intestine rather than tylenchs
(457.) or dorylaims (42X) . Twenty percent predators
were cannibals while 27% dsvoured mononchs belonging
to other species/genera- Individuals of Mylonchulus
did not {zontai n any indi vi dual of any prey.
Various factors such as the type of prey, agar
concentration, agar thickness, temperature, starvation
C3f predato-5, activity of predators and prey, test
arena etc., governed the rate of predation by
different nematodes-
The nuHfgrical analysis of the predatory behaviour
of nematodes including stride rate of the predators.

encounters resulting in attacks, attacks resulting in
wounding- feeding a-fter wounding, prey left unconsumed
and duration of feeding upon an individual prey made
in relation to five prey trophic categories viz.,
bacterial feeders (saprophagus nematodes); fliigratory
juveniles (sedentary endoparasitic nematodes);
epidermal feeders (ecto-parasitic nematodes); cortical
feeders (ectoparasitic nematodes) and predatory
nematodes revealed that the resistance and
susceptibility of an individual prey and prey trophic
category varied as a whole. The endo-parsitic
nematodes were highly susceptible group of nematodes
to predation. Hoplolai mus i ndi cus, Hemi cri cnemoides
manqi ferae, Hemi cycl iophora dhi render! ., Scutel 1 onema
sp. were totally resistant to predation. Various
physical (thick body cuticle, cuticular annulation
over the body, double cuticle sheath, gelatinous
matrix); behavioural (active body response upon been
attacked; evasive actions; rapid undulatory movements)
and chemical (toxic/unattractive body secretions) were
attributed the cause of resistance of varying degrees
in nematodes.
Cannibalism is a phenomenon which occurs mostly
under non availability of prey- This phenomenon is
most common in mononchs- Observations on

cannibalistic tendency of the adults and juveniles of
Mononchus aquaticus revealed that the adults and
fourth stage juvenileB were stronger cannibals than
their younger stages. The second and first stages
were neither cannibalistic nor predatory but they were
carrion feeders. No cannibalistic tendency was
observed in any dorylairo, rfipiogasterid, nygolairo or
actinolaimid predator.
Feeding behaviour of plant parasitic nematodes
comprised of five distinct phases viz.^ wide spread
exploration, local exploration, stylet penetration
extra-corporeal digestion and feeding.
Studies on host finding abilities showed that the
adults and juveniles responded to attractants released
by the host roots, but the degree of attraction varied
with the age of nematodes. Responses of adults were
stronger than those of the juveniles.
Plant parasitic nematodes exhibited some degree
of preference towards different host. Cabbage and
carrot seedlings were preferred most than the other
plant roots. A study of factors influencing
attraction of plant parasitic nematodes
<Hi rschmanni el I a oryeHe and Tyl enchorhynchus.

7
mashhoodi ) towards host roots re\feai ed that all e>;cept
coloured lights and darkness a-ffected the degree o-f
attraction- The nematodes showed maxi/num attraction
towards 0-1 cm (root tip) and 2-3 (zone of maturation)
when the roots were incubated for 10—12 h in 3—4 mm
agar layer o-f 0-5X concentration at 20-25 "C- The
optimum size o-f the source of attraction varied with
adults and juvenile stages. The attraction decreased
with the increase in the distance of inoculation of
nematodes from the host roots- Nematodes responded
effectively for a ma>!imum distance of 4 cm- The
responses of these species of plant parasitic
nematodes declined with ageing roots and also with an
increase in the starvation period of nematodes. Live
roots attracted nematodes significantly but dead roots
were not attractive. The age of nematodes itself
affected their degree of attraction. Adults being
more attractive than ' their juvenile stages- The
responses of parasitic nematodes varied with different
concentrations of root homogenates.
Low concentrations of chemicals were favourable
for the activity and survival of predatory nematodes
(A Quat i des thornei and Dorylai mus staqnalis) and plant
parasitic nematode {Hir-gchmannieJ la or yj ae) . Ammonia

3
at all concentrations suppressed activity markedly.
Even 1 ppm concentrations o-f nematicides aldicarb and
furadan were lethal -for the nematodes-
Observations on the embryonic and post embryonic
developments of Dorylai mus staqnali s revealed a total
devel opiiiBnt period (from h a t c h i n g to sdult) of males
•from 112-131 days and that of females from 117-130
days at 25-30 'C, The first cleavage was
perpendicular to the long axis which appeared 15—24 h
after the egg was laid. The first stage juveniles had
two germinal and 4—6 somatic nuclei in genital
pri/Bordium, The second stage had 4—h germinal and 8 — 14
somatic nuclei- The third stage had 8—12 germinal and
20—45 somatic nuclei. The fourth stage female and
male had 20-60 and 13—61 germinal nuclei respectively-
The first stage juvenile required 17-22 days for
moulting; the second 24-23 days; the third 30-35 days;
the fourth stage male 46—52 days and the fourth stage
female 40-45 days for their development respectively.
During studies on the sex attraction behaviour of
H. or yi: af? it was observed that both the se.xes produced
attractants which resulted in a mutual response
towards each other. Male-^ of H_ aryxaf^ were more
re-?;n fiosive thH»n ff? tnalef - T'nere was no attraction of

9
males towards male attractsnts and females towards
female eKtractants, The distance of nematode
inoculation and the period of nematode exposure to
attractants influenced the degree of attraction.
Significant correlations were observed between the
attraction <Mean Score) and the period of nematode
exposure to attractants.
Some work' on the numerical ta>i onomy ^ hi st ochewi cal
localization of enzymes and u 11 r etst r uc ture of
predatory nematodes was also raade which has been
incorporated as a supplement in the thesis- Numerical
taxonomy mainly provides different methods, their
application, significance and utility in nematology.
Some of the important enzymes of the Embden Meyerhof
pathway, Glycolysis and Pentose phosphate shunt ytere
histochamical1y localized in a predatory nematode
Dor yl ai fiius st aqnal is. It .was found that enzymes
aldolase, phosphoglucomutase, fructose 1-6
diphosphatase, alcohol dehydrogenase, malate
dehydrogenase, succinate dehydrogenase and glucose 6-
phosphate dehydrogenase were present in the nematode.
The presence of these enzymes suggest that these
cycles are operative in predatory nematodes either
simultaneously qr £•! t er r>a I i vt'l y . The ul tr ast rue tr al

lO
(scanning electron mi cr oscopy) studies o-f the
predatory nematodes viz, , Ho none hoi des fori, i dens,
Dory? ai mus st aqnalis, and Thalossoqenus shami mi
revealed many new and interesting characters such as
the typical patterns on the longitudinal ridges o-f D.
staqnali R and H. forti dens.

STUDIES ON THE BEHAVIOUR OF NEMATODES
THESIS SUBMITTED FOR THE DEGREE OF
Bottor of ^timtt IN
ZOOLOGY ALIGARH MUSLIM UNIVERSITY. ALIGARH
BY
ANWAR LUQMAN BILGRAMI Fh. D.
DEPARTMENT OF ZOOLOGY ALIGARH MUSLIM UNIVERSITY
ALIGARH (INDIA) DECEMBER. 19 9 3

OTECkED-2C02
4
2 b J'::i "394
T4266

D E D I C A T E D TO
MY MOST BELOVED PARENTS
WHO HAVE ALWAYS BEEiM A SOURCE OF MY iNSPiRATSOSS!

rUniversity: 285 Phones [p^^J^•,^ ; 25646
D E P A R T M E N T O F Z O O L O G Y ALIGARH MUSLIM UNIVERSITY
ALIGARH-202002, U.P. (INDIA)
Sections : 1. ENTOMOLOGY 2 PARASITOLOGY 3 ICHTHYOLOGY a FISHERIES
4 AGRICULTURAL MEMATO^QGY
b. GENETICS
Ref. No.
Date J an . 25th, 1994
CERTIFICATE
This is to certify that the thesis entitled "Studies on
the behaviour of nematodes" embodies the research work done
by Dr. Anwar Luqman Bllgrami either singly or in collaboration
with other scientists. The entire work is original and a
significant contribution to the field of Nematology. The major
portion of the work is done by Dr. Bilgrami himself.
I have, therefore, allowed Dr. Anwar Luqman Bilgrami to
submit the present work in the fulfilment of the requirements
for the degree of DOCTOR OF SCIENCE in Zoology of the Aligarh
Muslim University, Aligarh. .
Man Mohan Agarwal ^"^
Professor & Chairman

ACKNOWLEDGEMENTS
I am deeply indebted to Professor M. Bhamim
Jairajpuri who trained me in Nematology and supervised
my research work for Ph.D Degree. It would have been
difficult for roe to prepare this thesis without his
constant help and encouragement.
I am grateful to the Chairman, Department of
Zoology, Professor M. M- Agarwal and the past Chairmen
Prof. B. M. Alam, Prof. N. H. Khan, Prof, Ather H.
Siddiqi and Prof. M. A. Khan for providing research
faci1i ti es.
I owe special gratitude to my teachers Prof. A,
K. Jafri , Prof. Jamil A. Ansari and Dr. Bhahid H. Khan
for encouragement Bnd help. My colleagues Drs. Irfan
Ahmad, Masim Ahmad, Samina Bhafqat and Dudisa Tahseen
deserves special thanks for their help and
cooperati on.
The help rendered by Dr. Zakaullah Khan, Mr-
Fazal Rehman, Ms. Tabassum Mirza and Noor-us-Sabha
Firdousi during compilation of the thesis is
acknowledged with thanks. I also wish to acknowledge
the help extended generously by my research students
Mr. Rajeev Kulshreshtha and Mr. Rashid Pervez.
Last but not least I shall remain very grateful
to all my co-authors for a fruitful collaboration and
excellent cooperation.

C O N T E N T S
Introduction................................. t — 31
Some -factors influencing predation by Hononchuu aquaticus ..................... 32 — 33
Observations on the predatory behaviour of Mononchus aquaticus ............ 34 — 38
Responses of Hononchus aquaticus to chemicals and pH. ..........«.<............... . 39 — 42
Cannibalism in Kononchus aquaticus........... 43 - 44
Factors influencing attraction of adult Hirschmanniel1 a oryzae towards cabbage seedlings 45 — 53
Responses of adult Hirschmanniel1 a oryzae towards different plant roots 54 - 60
Predatory behaviour of Aquatides thornei (Nygolaimina t Nematoda) ............ hi — 6h
Attraction and feeding behaviour of adult Hi r schmanni el 1 a oryzae &7 - 74
Effect of some chemicals on the activity of Hi r schmanni el 1 a oryzae 75 - 80
Responses of Hirschmanniel 1 a oryzae juveniles towards cabbage seedlings.......... 81 — 84
A study on the intestinal contents of some mononchs ............................ 85 — 88

2 1
Effect of different plants, singly and in combinations on the attraction of Xiphinema basiri 89 - 9
Evaluation of the predatory behaviour of DorylaimuB etagnalis CNematodas Dory! ai mi da> 94 - 1 DO
Attraction of Mononchoides longicaudatus and M. fortidens (Nematoda s Dipiogasterida) towards prey and factors influencing attraction 101 - 108
Interaction between mononchs in non—sterile agar piates................... , 109 — 110
Plant parasitic nematodes associated with paddy crops in Bihar, India........... Ill
Observations on the osmoregulation in an insect nematode Schewenchiel 1 a atheri (Thaiastomatidae s Oxyuroidea>...... . 112 — 114
Resistance of prey to predation and strike rate of the predators Mononchoides 1ongicaudatus and M. fortidens (Nematoda : Dipiogasterida) ...... . 115 - 119
Effects of concentrations and thickness of agar on the predatory behaviour of Mononchoides 1ongicaudatus and M. fortidens (Nematoda s Diplogasterida) and attraction towards prey 120 - 124
Aggregation of Mononchoides 1ongicaudatus and M. fortidens (Nematoda:Diplogasterida) at feeding sites 125 - 17

1 1 J
ReBponses o+ adult and juvenile Hirschroanniel 1 a oryzae towards different sizes of the source of attraction 129 - 132
Predatory nematodes. 133 —. 163
Dipiogasterid predators.............^...... 164 — 171
Predaceous Dorylaims..,- 172 — 177
Predatory abilities of MononchoideB 1ongicaudatus and M. fortidens (Neroatoda 5 Dipiogasterida> and factors influencing predation........ 17B - 191
Plant parasitic nematodes associated with paddy crops in Uttarpradesh, India. 192
Developmental biology of DorylaimuB stagnalis Dujardin 1S45 <Ne«»atoda t Dorylaimida) 193 - 203
Thaifissogenus shamimi n.sp. (Nematoda: Thaiassogeneridae>, a nematode predator from , India................ 204 — 208
Observations on the predation abilities of Aporcelaimel1 us nivalis (Nematoda; Dorylaimida) 209 - 218
Resistance and susceptibility of prey nematodes to predation and strike rate of the predators, Hononchus aquaticus, DorylaimuB stegnalis and Aquatides thornei 219 - 224
Biocontrol agents and their role in nematode management.................. 225 - 240

A nematode Feeding mitp, Tyrophagus putrescentiae (Sarcoptiformi5 s Acaridae) 241 - 242
Effect of chemicals on the activity and survival of Dorylaimus stganal i s> Dujardin, 1845 and their juvenile stages. 243 - 248
Analyses of relationships between predation by Aporeelaimel1 us nivalis and different prey trophic categories....... 249 - 25B
Predation abilities of nematodes and protozoan and their role in nematode management 259 — 313
Chemo~oriented sex attraction in a plant parasitic nematode. Hi r schmanni el 1 a oryzae 314 - 326
Predation abilities of Mylonchulus minor and factors influencing predation 327 - 332
A comparative study on the predation by Al1odorylaimuB americanus and Discolaimus silvicolus (Nematode: Dorylaimida) on different species of plant parasitic nematodes. . , 333 — 366
Predatory behaviour of a nematode feeding mite Tyrophagus putrescentiae (Sarcoptiformes t Acridae) 367 - 389
Evaluation of predation abilities of Mylonchulus dentatus 390 - 412
Observations on the predation abilities of Meoactinolaimu& agilis <Dorylaimida : Actinolaimoidea) 413 - 431

Feeding and attraction behaviour o-f Tyl encborhynchus mashhoodi 432 - 449
Factors influencing attraction o-f HirBchmanniel 1 a oryzae towards cabbage seedlings. 450 — 477
Attraction and food preference behaviour in predatory nematodes, Al1odorylaifflus americanus and Discolaimus silvicolus <Nematoda ; Dorylaimida) 478 - 497
Numerical analysis of the relationship between the predation by Mesodorylaimu5 bastiani (Nematoda : Dorylaimida) and different prey trophic categories.......................... 498 — 515
SUPPLEMENT
Study of variability in Xiphinema radicicola Goodey and X. brasiliense Lor del I D , 1951 ,
Hi«;torhemii«l }c>Cr»li;!ation of some p^»^yroe5 of the carbohydrate metabolism in a soil nematode Dorylaimus stagnalis 526 - 531
Specialized Techniques in Nematode Taxonomy. ,.....,.. 532 — 547
A scannirjQ electron microscop study Df» Dorylaimus stagnalis Dujardin, 1845 (Nematode : Dorylaimida) 548 - 552
SEM studies on Mononchoides fortidens <Nematodai Dipiogasterida>........ 553 - 560
LIST OF PUBLICATIONS 561 - 570

DECLARATION UNDER CHAPTER XXVI OF
THE ACADEMIC ORDINANCES
ALIGARH MUSLIM UNIVERSITY, ALIGARH
I. As required under Chapter XXVI o-f the Academic
Ordinances, Aiigarh Muslim University, I declare that
the major portion of the work presented in the thesis
entitled " Studies on the Behaviour of Nematodes"
submitted to the Aligarh Muslim University, for the
BWBrd of the degree of Doctor of Science, is an
original contribution made by the author. In all 50
papers have been incorporated in the thesis- These
Bre as followss
i> Papers published independently ......11
ii> Papers with senior authorship 23
iii> Papers with junior authorship Ih
The above papers have been classified hereunder.
A Complete list of publications by the author is
appended at the end of the thesis.
(i> PAPERS PUBLISHED INDEPENDENTLY
1. BILGRAMI, A, L. <1999). Specialized Techniques in
Nematode J aytonomy, In Nematode Pest
Identifir^ation. <Ed. Jairajpuri, M- S. >-,pp
155-168.

2
2. BILGRAMI, A. L. <1990>. Predatory Dorylaims. In
Nematode Bio-Contro} lAspects srtd Prospect g
<EdB. Jairajpuri, Alam ic Ahmad} CBS Publishers
and Distributors Pvt,, Ltd., New Delhi pp
143-148-
3. BILGRAMI, A. L- U 9 9 B ) . Di pi ogaster i d predators-
In Nematode Bio—control; Aspects and Prospects
<Ed5. Jairajpuri, Alam & Ahmad),CBS Publishers
and Distributors Pvt.,Ltd., New Delhi, pp 135—
142-
4 BILGRAHI, A. L. <1992). Biocontrol agents and
their role in nematode management. In Recent
Advances in Nematology (Ed- B. K". Dwivedi)
Pub- Bioved Research Society, Allahabad,
203-218-
5. BILGRAMI, A. L. U 9 9 2 ) . Resistance and
susceptibility o-f prey nematodes to predation
and strike rate of the predators, Mononchus
aquaticus, Dorylaimus stagnalis and Aquatides
thornei. Fundamental & Applied WematDlDgy- 15 :
265-270.

h. BILGRAMI, A- L. <2993>. Analyses of
relationships between predation by
Aporcel ai mel 1 us nivalis and di-f-ferent pf^ey
trophic categories, Nematologica. 39 : 356—365.
7. BILGRAMI, A. L. <1994>. r4uBjericsl analysis of the
relationship between the predation by
Mesodorylaimu5 bastiani (Nematoda s
Doryl ai fl»i da> and di-f-ferent prey trophic
categories. Nematol. medit. <in p r e s s ) .
8. BILGRAMI, A. L. <1994). Predation abilities of
nematodes and protoz pan and thei r role i n
nematode management. In Pesticide and
Environment Vol. II. (Eds. Gaur, H- C. &
Prasad, D . ) . (In p r e s s ) .
9. BILGRAMI, A- L. (1994). Predatory behaviour of a
nematode feeding mite Tyrophagus putrescentiae
. (Sarcoptiformes i Acridae). Fundamental and
Applied Nematology- (in p r e s s ) .
10. BILGRAMI, A. L. (199 ) . Factors influencing
attraction of Hirschmanniel 1 a oryzae towards
cabbage seedlings. Netsatol ogi ca . (Communicated)
11. BILGRAMI, A. L. (1994), Attraction and feeding
behaviour of Ty]enchorhynchus mashhood i. Annals
PI ant Pf nt ec t t on S c ? ^\^c.^'=,. (in p r e s s ) .

(ii) PAPERS WITH SENIOR AUTHORSHIP
1- bILGRAMI, A. L., AHHAD^ I. ?< JAIRAJPURI, H. S.
<}983), Some -factors influencing predation by
HonDnchus aquaticus. Revue Nematol . :, 6i 325-
326.
2. BILGRAMI, A. L., AHMAD, I. S< JAIRAJPURI,
MiS.<1984). Observations on the predatory
behaviour o-f Mononchus aquaticus. Nematol .
Medit-, 12 s 41-45.
3. BILGRAHI, A. L. ^ JAIRAJPURI, H. S. <1985). The
responses o-f Mononchus aquaticus to chemicals
and pH. Indian J. Nematol., 14 (1984): 171-174.
4. BILGRAMI, A. L. «c JAIRAJPURI, M. S. <1985).
Cannibalism in Mononchus aquaticus. Indian J.
Nematol. 14 (1984)j202-203.
5. BILGRAMI, A. L., AHMAD, I. ^ JAIRAJPURI, M. S.
(1985). Factors influencing attraction of adult
Hirschmanniel 2 a oryzae towards cabbage
seedlings- Revue Neraatol., 8:67-75-
h. BILGRAMI, A. L., AHMAD, I. ?< JAIRAJPURI, M- B.
<19S5>, ReBponses of laduit Hi r schmanni el 1 a
oryzae towards different plant roots- Revue
Nematol., S : 265-272.

5
7. BILGRAHI, A. L., AHMAD, I. S< JAIRAJPURI, M. B.
<1985)- Predatory behaviour o-f Aquatides
tbornei <Nygolaimina s Nematoda). Nematologica,
(1984)! 30 ! 457-462.
S. BILGRAMI, A. L., AHMAD, I.& JAIRAJPURI, M. S.
(1986). Attraction and feeding behaviour of
adult HiTBchroanniel 1 a oryzae. Indian J.
Nematol., 15 (1985)5 289-216.
9- BILGRAMI, A. L., AHMAD, I. ?* JAIRAJPURI, M. S.
(1986). Effect of some chemicals on the
activity of Hirschmanniel 1 a oryzae.
Helminthologia, 23 : 49-54.
ID. BILGRAMI, A. L., AHMAD, I. ?c JAIRAJPURI, M. S.
(1986). Responses of Hirschmanniel 1 a oryzae
juveniles towards cabbage seedlings.
HelminthDlogia, 23 s 119-122.
11. BILGRAMI, A. L., AHMAD, I. ^ JAIRAJPURI, M. S.
(1986). A study on the intestinal contents of
some mononchs. Revue Nematol., 9 : 191-194.
12. BILGRAMI, A. L., JAIRAJPURI, M. S. S< AHMAD,I.
(1986). Effect of different plants, singly and
in combinations on the attraction of Xiphineroa
basiri, Indian J. nematol., 16 ; 60-64.

D
13. BILGRAMI, A. L., ALAK, S. H. ?. AHMAD, I. (1987).
Hi stocberoi cal localization of BOBJe enzymes o-f
the carbohydrate roetaboliBm in a soil nematode
DorylaimuB stagnalis- Nematologica <1986)- 32 i
239-243.
14. BILGRAMI, A. L. & JAIRAJPURI, M- S. (1988).
Attraction o-f MononchoideB 1 ongi caudatus and
M. •fortidens (Nematoda : Di pi ogaster i da >
towards pi'ey and -factors in-fluencing
attraction. Revue Nematol., 11 : 195-202.
15. BILGRAMI, A. L., AHMAD, I. ?< JAIRAJPURI, M. S.
(1988). Interaction between mononcbs in non-
sterile agar plates. Indian J. Nematol., 17
<1987>- 339-340.
16- BILGRAMI, A. L- & JAIRAJPURI, M. S. <1989>.
Resistance of prey to predation and strike
rate of the predators Mononcboides
1 ongi caiidatus and M. fortidens (Nematoda:
Dipiogasterida)- Revue Nematol., 12 : 45-49.
17. BILGRAMI, A. L. ?< JAIRAJPURI, M. S. (1989).
Effects of concentrations and thickness of
agar on the predatory behaviour of
Nononchoides 1ongicaudatus and M- fortidens

7
(Nematoda:Dipi ogasterida> and on their
attraction towards prey. Indian J. Nematol.,
(19S8),18 : 222-227.
la. BILGRAMI, A. L. ^ JAIRAJPURI, «. S. (1989).
Aggregation o-f Mononchoides I ongi caudatus and
M- fortidens {Neroatcda s Dipiogasterida> at
deeding sites, Nematologica C19S8), 34 s 119-
121 .
19. BILGRAMI, A. L., AHMAD, I. & JAIRAJPURI, M, S.
(1989). Responses of adult and juvenile
Hir schmanni el 1 a oryzae towards dif-ferent sizes
D-f the source of attraction- Indian J.
Nematol. 18 (1988) s 357-360.
20. BILGRAMI, A. L. S< JAIRAJPURI , H. B. <1990>.
Predatory abilities o-f Mononchoides
1 ongi caudatus and M. -fortidens <Ne®atoda s
dipiogasterida> and factors influencing
predation. Neroatologica, 35 (1989)5 475-488.
21. BILGRAMI, A. L. S< AHMAD, W- (1990). Plant
parasitic nematodes associated with paddy crops
in Uttarpradesh, India. Int. jNematol . News
Letter. 7s pp 25.

8
22. BILGRAMI, A. L. ?< TAHSEEW, D, <1992)- A nematode
Feeding mite, Tyrophagus putrescentiae
(Sar copt i-f ormi Bs Acaridae). Fundamental &
Applied Nematology, 15: 477-47S,
23. BILGRAMI, A. L. ^ KULSHERESHTHA, R. (1994).
Evaluation o-f predation abilities of
Hylonchulus dentatus. Indian J. Nematol.
(in press).
(iii) PAPERS WITH JUNIOR AUTHORSHIP
1. RAHMAN, M. F., BILGRAMI, A. L. ^ JAIRAJPURI, M.
S. (1986). Study of variability in Xiphinema
radicicola Goodey and X. brasiliense Lordello,
1951- Helrointhologia, 23 : 179-188.
2. SHAFQAT, S. , BILGRAMI, A. L. S< J AIRAJPURI , M. S.
(1987), Evaluation of the predatory behaviour
of Dorylaimus stagnalis (Nematoda :
Dorylaimi da). Revue Nematol. , 10: 455-461.
3. AHMAD, W., KHAN, T. H, & BILGRAMI, A. L.
(3988). Plant parasitic nematodes associated
with paddy crops in Bihar, India. Inter.
Nematol. News Letter, 5 : 4 .

4. PARVEEN, R., BILGRAMI, A. L. S< JAIRAJPURI, D.
S, <19S9). DbBervations on the osmoregulation
in an insect nematode Schewc?nchi el i a atheri
(Thai astomati dae : D>:yuroi dea) - Indian J.
Nematol., <1988), 18 : 218-212.
5. JAIRAJPURI, M. S,, BILGRAHI, A. L- <199B>.
Predatory nematodeB. In Nematode Bi o—control i_
Aspect s and Prospects (Eds. Jairajpuri, Alam
?f Ahmad) CBS Publishers and Distributors Pvt.,
Ltd., New Delhi pp 95-125.
6. BHAFDAT, S.,JAIRAJPURI, M- S. & BILGRAMI, A.
L. (1991). Developmental biology of Dorylaioius
stagnalis Dujardin 1S45 (Neroatoda r
Dory1aimi da). Revue Nematol., 14 : 61-71.
7. SHAFQAT, S. , AHMAD, I., AHMAD, W- «< BILGRAMI,
A. L. <1991). A scanning electron microscop
study on Dorylaimus stagnalis Dujardin, 1845
(Nematoda : Dory1aimi da). Revue Nematol. 14 :
511-515.
S- AHMAD, W. , AHMAD, I., BILGRAMI, A.L. ?< KHAN, Z.
(1991).ThaiassogenuB shamimi n.sp.(Nematoda:
Thaiassogeneridae), a nematode predator from
India. Fundamental ?< Applied Nematology- 15:
127-131.

10
9. KHAN, Z., BILGRAMI, A. L. ?. JAIRAJPURI, M. S.
<1991), Some studies on the predation abilities
Df Aporcel ai mel 1 us nivalis ^Nefnatodai
Dorylaimida>. Nematologica. 37 i333-342.
ID. TAHSEEN, D. , AHMAD, I-, BIL(5RAMI, A. L. Sc
AHMAD, W- (1992>. SEM studies on Mononchoides
•fortidens <NematDda s Di pi cgasteri da> .
NematolDgica 38 : 296-303.
11. SHAFQAT, S., BILGRAMI, A.L. ^ JAIRAJPURI, H.
S.(1993)- Effect of chemicals on the activity
and survival of DorylaimuB stganalis Dujardin,
1845 and their juvenile stages. Indian J-
Nematol. 22 a992> s 19-24.
12. KULSHRESTHA, R- , BILSRAMI, A. L. ?f KHAN, Z.
<1994>. Predation abilities of Kyldnchulus
minor and factors influencing predation.
Annals Plant Protection Sciences. 1 : 79-84.
13. TAHSEEN, QUDSIA S< BILGRAMI, A. L. n994>. Chemo-
oriented sex attraction in a plant parasitic
nematode. Hirschmanniel 1 a oryzae- Indian. J.
Nematol. <in press).
14. KHAN, Z., BILGRAMI, A. L. 5< wlAIRAJPURI, M. S.
<1994>. A comparative study an the predation

11
by Al1odory1aimu5 americanuB n. sp- and
DiscolaimuB B I I V I C D I U B (Nematoda :
Dorylaimida) on different species of plant
parasitic: nematodes. Fundamental and Applied
Nematology, <in p r e s s ) .
15. KHAN, Z., BILGRAMI, A. L. ?f JAIRAJPURI, K. S.
<1994>. Attraction and -food pre-ference
behaviour in predatary nematodes.
All odorylai mus ameri canus and Pi scolai mus
silvicolns <Nematoda j Dor yl ai n»i da) .
Nemato3 OQJ ca. , (communicated) -
16. KHAf-J, Z., BILGRAMI, A. L. & JAIRAJPURI, M, S-
<1994). Observations on the predation
abilities o-f Neoac t i nol ai mus aqi lis
<Dory1aimidai Actinolaimoidea). Indi an J.
Nemetol. (in p r e s s ) .
I ^^urther declare that the following portion o-f
the work submitted herein was previously considered
•for the award o-f Ph.D degree of the Aligarh Muslim
University. The degree was awarded in the year 1983
on my thesis entitled "Observations on the behaviour
of nematodes".

••)
1. Some -f actors influencing predation by MononchuB
aquati cu£-»
2. Observations on the p'redatory behaviour of
MononchuB aquaticus-
3- The responseB of Mononchus aquaticus to chemicals
and pH,
A. Cannibalism in Mononchus aquaticus.
5. Factors influencing attraction of adult
Hirschmanniel 1 a oryzae towards cabbage seedlings-
6. Responses of adult Hirschraanniel 1 a oryzae towards
different plant roots-
7. Predatory behaviour of Aquatides thornei
(Nygolaimina : hJematoda).
S. Attraction and feeding behaviour" of adult
Hirschmanniel 1 a oryzae.
9. Effect of some chemicals on the activity of
Hirschmanniella oryzae.
ID. Responses of Hirschmanniella oryzae juveniles
towards cabbage seedlings.

1 •^
The foilowiPfg is the list of papers based on Ph.D
t hese.
1. BILGRAMI, A. L., AHHAD, 2. ?< JAIRAJPURI, H. S.
<1983)- Some -factors i n-f 1 uenci ng predation by
MononcbuB aquaticuB. Revue Neroatol., 6i 325—
326.
2. BILGRAMI, A. L., AMHAD, I. ?c JAIRAJPURI,
s1-S-<19S4>, Obser vat i OTf 5 on the predatory
behaviour o-f Hononchus aquaticus. Nematol .
Medit., 12 : 41-45.
3. BILGRAMI, A. L- ?c JAIRAJPURI, H. B. U 9 S 5 ) . The
responses o-f Mononchus aquaticus to cheffficals
and pH. Indian J. Nematol., 14 (1984): 171-
174.
4. BILGRAMI, A. L. ?c JAIRAJPURI, M. S. <1985).
Cannibalism in Mononchus aquaticus. Indian J.
Nematol. 14 U984>:2S2-283.
5. BILGRAMI, A. L., AHMAD, I. ^ JAIRAJPURI, M. S.
(1985). Factors influencing attraction of
adult Hirschmanniel 1 a oryzae towards cabbage
seedlings. Revue Nematol., 8:67-75.

14
h. BILGRAHI, A. L-, AHJiAD, I. ?( JAIRAJPURI, M.
S.<i9S5), Responses o-f adult Hirschmanniel 1 a
pr ys ae towards dif-ferent plant roots. Revue
Nematol. 8: 67-75.
7- BIL8RAHI, A. L., AKHAD, I. ?< JAIRAJPURI, H. S.
(1935>. Predatory behaviour o-f Aquatides
thornei (Nygolaimina s Nematoda).
Nematologica, <19a4>s 30 i 457-462.
8- BILGRAMI, A. L., AHMAD, I. S< JAIRAJPURI, M- S-
<1986>. Attraction and -feeding behaviour of
adult Hirschmanniel 1 a oryzae. Indian J.
Nematol., 15 <1985)i289-216.
9. BILGRAMI, A. L., AHMAD, I. & JAIRAJPURI, M. S.
<19S6>. E-ffect of some chemicals on the
activity o-f Hi r schmanni el 1 a orysae-
Helminthologia, 23 : 49-54.
ID- BILGRAMI, A. L., AHMAD, I. S. JAIRAJPURI, M. S.
(1986). Responses o-f Hi rschmanni el 1 a oryrae
juveniles towards cabbage seedlings.
Hel mintbologi a, 23 ; 119-122,.
\ 'k.\^~' Anwar L'. Bilgrami

15
INTRODUCTIOW
^4e^)atode5 sir& one o- the most important and
highly diversified group'S o-f multicellular animals-
Like other micro—organisms they are found in all
kinds o-f habitats. Dur knowledge of nematode
parasites of man and animals dates back to 4,500 B.C.
but the plant and soil nematodes were not discovered
until the 17th Century mainl y because of their small
size and the technical difficulties involved in their
isolation from the soil and plant tissues. Boreilus
(1656) recorded the first free living nematode, while
Needham (1743) was the first to discover the wheat
gall nematode now known as the Anqui na tritici .
Linnaeus (17 6 7 ) , Scopoli <1777>, Steinbuch (1799) and
others recorded the same species and noted it to be
a serious pest- Berkeley (1855) recorded the second
plant parasitic nematode H e l o i d o g y n e 5p., while
observing the galls produced on greenhouse grown
cucumber in England- Two years later in 1857 Kuhn
found yet another important nematode species which is
now known as Pi t ylenchus di psaci from the malformed
floral hieads of the fuller's teasel -
The economic importar»ce of plant parasitic
ne/tiatodes was first realised when the production of

u
commercially important Bugarbeet crop suddenly
declined in Germany in the second hair o-f the 19th
Century. In 1B59 Schacht discovered a serious
disease on sugar beet caused by a species of cyst
nematode whi cb v*as 1 ater named as Heteroder a schacht i
by Schmidt (1871). Due to serious losses in the
production of sogarbee.ts caused by H. schacht i
efforts were made by subsequent workers (Kuhn,
Liebcher, Koiz, Muller, Chatin and others) to study
its life history, habits, distribution and methods of
control- According to Thorne <196i>, the monograph
on Angui 1 1 ul i dae published by Bastian OS65) marked
the beginning of Nematology, It was only since the
early 20th century that true awareness of plant
parasitic nematodes and other important nematodes was
made.
The credit for the spread of awareness of plant
and soil nematodes goes to an American Neraatologist
Nathan August Cobb (1859-1933) who was employed by
the United States Department of Agriculture to spread
an awareness about the plant pests and other
beneficial nematodes. He was responsible for
initiating the formal study of these nematodes. His
great disservice was that he coined the term

17
Wematology. While -focuBing attention on the plant
pests he created a ZDol:^Qical artificial split by
distinguishing Mematology (the study of plant and
soil nematodes) -from the Hel mi nt hoi ogy (the study of
P'arasites of animals and man. Cobb published a
series of excellent papers on the taxoriDmy of fresh
water, soil Bnd plant nematodes. Detailed
morphological studies were made and new terminologies
were coined by him. The books by Chitwood (1937,
1950), Goodey (1961), Thorne (1961) and others
revolutionized the taxonomy of nematodes. These
contributions have laid a solid foundation of the
taxonomic and morphological research in Nematology-
Other aspects like physiology, biology, behaviour,
locomotion, feeding, predation etc., have remained
almost neglected even during the recent years when
Nematology has won recognition as an independent area
of biology. In spite of the spectacular progress
the research in Nematology is still by an large
confined to ba^ic aspects, like the taxonomy,
systematics and morphology certainly with many of the
advance techniques like SEM, TEM, Phase contrast
microscopy, Cryofacturing etc. It was only during
the last 15 years that attention has been paid to the
physiological, ecological and behavioural studies of

8
plant and soil nematodes- It is not unlikely that
the e>!ceedingly small size and the subterranean
habitat of the plant and soil nematodeE might have
largely been responsible -for this neglect-
During this &ra of research on nematodes when
extensive research was being pursued on their
taxonomy^ sjorphology, ecology and control, studies on
nematode behaviour were largely neglected- The
reason probably was their microscopic size* srnd their
habitats—marine, freshwater, soil and as parasites of
plants Bind animals. The behaviour of nematodes under
natural conditions has almost entirely escaped
observations. This is true because in the
laboratory investigations are essentially made under
artificial conditions and usually eliminate all other
factors e>!cept the one under investigation.
Behaviour in general is an observable activity
of a whole animal. The behaviour of nematodes is
assessed in terms of activity and locomotory patterns
which are the consequences of the sensory inputs- It
involves all the processes by which an animal senses
its external surroundings and responds to the change
it perceives iCarthy, 1958; Diether ?< Stellar, 1959?
C? ondnl ey-Tho<(»pson , ]9c>l) ^nd ihis in essence forms

19
the total behaviourel componerit in lower ani/nals-
Insects with their CDmp«le>; reactions and social
behaviour enjoy hierarchial dominance in terms o ^
behavioural patterns. Still higher sre the
vertebrates which display instinctive behaviour and
•finally the human beings who alone can look upon
themselves and become the object of their own
thoughts and can distinguish the world they know -from
themselves. Every organism living in an environment
has its own mechanical and physiological problems and
any environmental changes are bound to influence its
li-fe activities- The fluctuations in the
environmental conditions act as an internal or
external stimuli and the animals respond to them by
making adjustments. The analysis of these changes or
responses exhibited by the animals form the basis of
behavioural studies- The nematodes occupy a wide
range of ecological niches and show considerable
behavioural diversities in response to the
fluctuating environmental conditions. The behaviour
of these animals involves some form of movement,
either motion of certain parte of body or an outright
locomotion. The absence or cessation of activity is
itself an e>!preB5iDn of behaviour. Behaviour of an
cinima} is always depended upon its physiological

0
state and is divided into three basic types the
somatic behaviour, visceral behaviour and the genetic
behaviour- A22 differ only in their manifestation
but conform to the basic physiological processes.
Fraenkel ?< Gunn <1940) have done much to clarify and
standardize the various terminologies of animal
behaviour. They along with Gunn et al . (1937),
Kennedy (1945) and Ewer it Bryssel (1950) have
contributed richly to the study of behavioural
patterns in lower organisms including nematodes,
Most of the GBrlier information on the nematode
behaviour- are available in literature on the biology
and ecology which remained mostly confined to few .of
the important plant parasitic nematodes. These
studies have been summarized by Wallace, Rogers,
DrDp><in, Klingler, \fan Gundy, Doncaster, Crol 1 and
others.
Nematode behaviour, though, studied earlier was
centered on areas of biomedical and agricultural
importance. The study of nematode behaviour in a
broader perspective of analysing responses in
relation to other animals and the functional
morphology of nemato«Je itself is a relatively young
subject. Even then numerous responses analogous with

21
other invertebrates have been described by various
wor};ers. The ul tr astr uc t ur a i studies have brought
to lignt the comple>: nature o-f sense organs arid
provided clues for their possible mechanism of
•functioning (Wright, 1976).
The diversities o-f nematode behaviour are
apparent, however, its interpretations -from the point
o-f view o-f -functional organization ar& still in the
process of study. The nature of sense organs and
their practical -functional implications were
highlighted by Crol1 (1967) when he stated that
during movement the amphids, which are supposed to be
chemoreceptor5 did not lie side by side at right
angle to the plain of movement. It is therefore, the
complexity displayed in nematode behaviour seems to
be correlated with the degree of development of the
sense organs and the ability of perception since the
behaviour is measured in terms of activity and
locomotion (Croll, 1967). The sense organs are Unovtn
to respond to touch and chemical stimuli. Some
nematodes possess ocelli (Croll et al., 1972) while
others setae, bristle, papillae or free nerve endings
(Doncaster S< Seymour, 1973) and amphids which act as
chemoreceptnr5 (Ward, 1973; ZuKerman, 19S3).

22
Plant parasitic nematodes form a vast community
within the soi} mi cro--? auna. The phytophagous
species mainly belong to the order Tylenchida and
few to Dorylaimida- Many aspects of their feeding
behaviour, such as attraction, exploration, stylet
thrusting, salivation and ingestion have been
observed iWyss, 1 9 7 1 - 1 9 7 9 ) , Prior to 1970 these
observations were mostly restricted to a few
economically important phyto—parasihic nematodes.
Since then attraction end feeding behaviour of
numerous species have been studied with modern
methods and for a wide spectrum of behavioural
activities. During this era of behavioural studies
work carried out by Wyss on attraction and feeding
behaviour of plant parasitic nematodes drew
significant importance and laid a solid foundation of
this aspect of nematode behaviour. Later many
studies were mads on the attraction (Hayens ?< J o n e s ,
1976; Lee ic Evans, 1 973) j feeding types and
mechanisms (Doncaster ?< Seymour, 1973; Bridge ?<
Hague, 1 9 7 4 ) ; factors influencing attraction and
feeding of plant parasitic nematodes ^Boag, 1980;
Davies ?< Fisher, 1976; Moussa e_t aj_- •, 1 9 7 2 ) .
In t-QnirasI In thf-* plant tjarar-ifcic nematodes
vf r y 1 i t I 1 t-> wor k !ir«B !>'-H>I< dont^ on the predatory

22
behaviour of nematodes. Similar to plant parasitic
nematodes the predaceous nematodes also constitute
one of the largest group o-f Wematoda. They share the
soil environment with a multitude of soil microfauna.
The predatory nematodes fall under three categories
depending upon their mode of feeding and the type of
feeding apptaratus. The predaceous nematodes belong
to the order iMononchida, Di pi ogasteri da Bnti
Aphelenchida J suborder Dorylaimina and NygDlaimina,
superfamily Actinolaimoidea, family Ironidae,
Thaiassogeneridae etc. Predators are important from
the point of view of biological control of plant
parasitic nematodes. Though, the economic importance
of predatory nematodes was realixed since the early
part of this century but no serious effort was made
to evaluate their predatory potentials and study
predatory behaviour- Cobb's (1917) observations on
the biology and predatory behaviour of roononchs were
the first attempt which lead studies on predation by
few other worKers (Steiner ^ Heinley, 1922; Thorne,
1927^ Thorne ?f Swanger , 193o>. The nature and source
of food of majority of nematodes was not known till
Banage (19^1); E'=>-ier ^ Sobers (1964) and Veates
(]969^ made observations with different species of
p r- e f51=41 o r y n e m r» I o d e s , T h •> r '-? B c r c h xyr~i predatory

24
behaviour o-f nematodes which remairi&d by and large
neglected got. ar« ijnpetiis since the beginr'jing of
seventies. I n vest i cjat i ons were made on their 1 i-f e
cycle (Haertens, }*?7<S)| predatc-ry behaviour <WyBB &
Grootaert; 1977; Grootaert &r Msertens, 1976)5
ul t r est r uc tur e and function of feeding apF<aratu5
(Grootaert 2f Wyss, 1 9 7 9 ) ; predation abilities (Small
?f Grootaert, 1983) j factors influencing predation
<Nel»ne5, 1974) and other aspects- of predatory
behaviour. A significant and pioneering work on the
predatory behaviour of different types of predators
was carried out by Bilgrami et al. <1983-i993) which
has pa^ed the way to investigate other finer details
of this interesting aspect of nematode behaviour.
Pheromone behaviour has been described
extensively in insects (Shorey, 1976) but in
nematodes only the occurrence of 5e>j pberofnones is
known (Greet et aj_. , 1 9 6 8 ) , Nematode 5e>« pheramones
or sex attractants as they are more cojn.-nonly knovtrij
influence activity from only a very short distance.
Not mucji work Ui^'^ been done on the phy si co—cbemi cal
properties of nematode pheromones primarily because
of fche difficulty in obtaining a pure e>; tract.
F'rel 1 ;) i nar y work has showf^ that the sex plieromones

25
were volatile and labile which became i ne-f-f ec t i ve
a-fter 24 b <Green^ 1966) in contrast to the alarm
pheromone o-f the terniites which remained attractive
even a^^ter 4S h (Eisner et_ aj_. , 1 9 7 6 ) -
A l a r g e n u m b e r o^ internBl and e>iterrtal f a c t o r s
govern the behavioural activites o-f nematodes.
Ageing is one such factor which determines the out
come of response. Age affects activity of nematodes,
fecundity, the rate of oviposition and vulval
contractions sex attraction, rate of copulation,
osmotic fragility, specific gravity, feeding and many
other behavioural activities ICroll^ 1975j Duggal,
1978; Zukerman e ^ s2_. ^ 1 9 7 1 ) .
Very little information is available on the
migration of nematodes under the influence of pH
gradients, 1ight , temperature, gravity, electric and
meganatic fields, mineral salt concentrations " and
acids. Some of the work carried out by the earlier
workers (Wallace, 196? j Fraenkel ?< Guun, 1940;
Clapham, 1933; Bird, 1959> is the only information
available the effects of the above factors on
nematotle behaviour.
Dnrirjg ^^»e r^cpitt yeac'-; studies on the roif? of
neiiif t oiJf B as i ml j c iTi I or " r>+ en v i ronwon t" a 1 pollution

hav«=> gained Bomp mo/aentufn as initial results have
been encouraging as shown by the stuc'ies of Zu2 1ir<i
<1976^ and Shaw et al . 0 9B3>. The ne;na t ode/cc«pepod&
ratio is used in pollution fnonitoring as studied by
Crol1 et al. ^1981). The role of nefiiatodes along
with arthropods in decomposition o-f semi arid
ecosystem was studied by El kins ?< White-ford (1982)
eind further their abundance patterns in detection of
en vi r on»»ent al pertubrence was observed by PI al 1 et
al, <1984>.
The developmental biology o-f nematodes is an
aspect which has drainn considerable attention, be it
-for understanding the 1 i-fe cycle o-f economically
important species so as to device control programmes
or to elaborate the fundamental aspects of
development of an organism. It may be pertinent to
note that the early work on developmental biology of
nematodes started in the later part of the 19th
century. Some important work of that time (Butschli,
1875; Mailer , 188?.; Boveri^ 1893> later carried out
by the following works (Weuhaus, 19D3; Pai, 1928;
Hechler, 1963; Abdel-Rahman ^ Maggenti, 1 9 8 7 ) . The
work included different aspects of developmental
biology like the Bmbryonic and pest embryonic

27
developments, cleavage patterns, gonad development,
•factors in-fluencing development, hatching, ecdysis,
duration o-f development, devel opm€?nt of genital
primordia, multiplication ssnd di-f fer ent i at i on of
cells/nuclei -forming the gonad and the development o-f
secondary sexual characters. Still, it requires lot
to be done on developmental biology of nematodes
which could -fulfil the in-formation gaps in the
1i terature.
In the recent past behavioural activities
attracted some attention. Various aspects like the
movement o-f the juveniles in the egg and hatching,
molting, -feeding of plant parasitic and predators,
defecation, swarming, penetration, orientation,
copulation, sex attraction and chemoperception
behaviour of plant parasitic and predaceous nematodes
have been studied. Different workers have used a
wide variety of modern methods like tracking (Croll,
1976), micro-cinematography ^Doncaster, 1971) and
micro surgery (Hung et al.^ 1982) to study nematode
behaviour and recently scientists have started
interpreting behaviour at molecular level.
In India research in Nematology started rather
late, buV the ecnnnmir. importance of the nematodes

28
as pests and predators quickly led to the development
D-f numerous nemat oi ogi cal research centers all over
the country. Wo doubt nefiiatode problems have existed
in our cultivated crops ^or a long time but the real
recognition o-f plant and soil nematodes came in the
late -fifties when several important developments led
to the escpansion of Hematology in India. Inspite of
the extensive development of Nematology in India
specially aspects like the taxonomy, morphology and
control of nematodes research on nemaiiode behaviour
totally remai n&d n&qlected and there was not a
single centre where this kind of work was going on.
Even today no Hematology Centre in India is engaged
in carrying out behavioural studies of nematodes
e>{cept the one in ^he Department of Zoology, Aligarh
Muslim University.
The Department of Zoology of the Aligarh Muslim
University where the present research work involving
studies on the behaviour of plant and soil nematodes
was carried out happens to be the pioneer centre of
nematological research in India. The work in
Hematology was initiated here in 1955 under the
leadership of Prof. M. A, Basir which blossomed into
a fullfledged Section. In this short span of about

29
40 yfc?ars extensive research work sjostly on the basic
aspects like th? ta>!onoff»y and morphology has been
carried out which brought international recognition
and light to this Centre of Newatology mainly because
of the devoted and high esteemed research carried out
here. Presently this centre o-f Nematology is not
known only -for its fundamentBl reBearch but also for
the signi-ficant behavioural research which was
carr i ed out -for a little over 15 years. Much o-f the
work carried out on behavioural aspects is pioneer in
nature which has achieved recognition world over.
Literature lacked i n-f ormat i one on various
aspects o^ nematode behaviour in general and
predatory behaviour of nematodes in particular due to
which research could not be pursued. It was
therefore, indepth studies were made on different
aspects of predatory behaviour of nematodes- Most of
these studies were carried out for the first time and
was pioneering which include detailed
characterization of the mechanisms and principals
involved in P«"ey catching and feeding mechanisms,
attraction of predators towarrSs prey, pre- and post-
feeding aggr t Or {-i on and attraction of predators at
f ee<l i ng - i h f*s , f i r i nr s i nf 1 nt^nr i f»g , 11 r . c t i on anci

0 n
predatioTf, pr-edation abilities, p?^ey pre^^erence,
canni bal i 5fr« , interactions, r el at i onshi p-s between
predation and dir-ferent P^ey trophic categories,
nature snd type o-f -food and resistance and
susceptibility o-f prey neroatodes to predation and
strike rate of the predators. Models were proposed
to measure the strike rate of the predators and
resistance sind susceptibility of prey nematodes to
predation quantitatively. Besides, the effects of
chemicals were also observed on the activity of
predatory nematodes. The work on predatory behaviour
of nematodes has aroused interest which received
world wide acclaim and recognition. This is
indicative frooj the fact that out of the total 50
publications 32 papers were published on behaviour of
predatory nematodes alone. Two papers published on
the predatory behaviour of a nematode feeding m i t e ,
Tyi-ophanus put r escen t i ae are also included in the
thesis as they depict prey behaviour of nematodes
Bnd maintsiin a c o n t i n u i t y .
Observations were also made on the attraction
e^nd feeding behaviour of plant parasitic nematodes,
factors influencing abtrachion and feeding, responses
of par<^=ijtic nematodes to plant roots and effect of

31
dif-ferent hosts on the attraction of phytophagous
nematodes. In all eight papers were published on these
aspects of nemettode behaviour. The rest o-f the papers
deal with the 5e>; attraction behaviour, ef-fect of
chemicals on the activity, devej opinent ai biology and
distribution o-f nematodes. A -few o-f them deals with
the numerical taA-onomy, ul t r astruc t ur e , hi stochemi cal
localization of the enzymes and variability which are
supplemented at the end o-f the thesis.
The present research work is an e-f-fort to
consolidate studies on some o-f the behavioural aspects
of plant and soil nematodes which have remained
neglected altogether. Whatever have been accomplished
in this thesis cover wide range experiments with the
limited facilities- available in the laboratory.
Nevertheless, foundation has been laid for better
future research on nematode behaviour in general and
prGdatory behaviour of nematodes in particular.

Notes breves
SOMK FACTORS r \ F r j ; [ ' : \ ( ; r . \ G J'RKDATfO.X lA' \rOXO.\CHUS AOi: ATICUS
Anwar L. Bi lgrami*, Irfan Ahmad* and M. S h a m i m Jairajpuri" *
"2
Ft'w .-ludies ha \ e be(>n inadr on llie firedatory abilily of inoiionchs. X('lnu'> .1974) studied tlic predalory behaviour of 7'rionc/!w/((.s punc/a/((.s- ^Dujar-din. IHlfr & \Vu Hoeppli, 1929, while thai of Monun-cliiis (Kjuaticu^ CoeLzee, 19()S wa- ^tudied b \ Oroo-taerL and \ \ ' \M^ (1979). Jairajpuri and A/.rni '1978 observed tlie predatory potenliai of Miilonchulun (Icnlalus and liie praclicabibty of usinp; Inonon^h^ to con! rot nematodes \va^ as^es>ed by Cohn and Mordechai il974) and Small 11979'i.
The rate of predation on different species was determined by plaein<j five Monoiichiis aqualicus with 2r) specimens of prey in cavity blocfcs containinir 1 % water ajjar. Five species of [irey, viz., (^hiloplacus syniiii('lricu)i [Thorne. 1925i Thorne, 19o7. (Jephalobus sp., Mesorliabdilis sp., Prisiiialoiaimus >p.. and Aglcnchus parvus Siddi([i, 19615 were used separately. To observe the effect of tempera!ure on predation, >[)ccimens of .1 / . aqualicus wei-e kept at different temperatures ranj^ing from 10 to 40" together with 23 s|iecimcns of (J. symmetricns in ca \ i ly blocks containing 1 % water au'ar. Predation was recorded after 24 iir. Similarly, the effect of agar concentration was obser\-ed by placing iise predators with 23 C. si/nunetric(is in 1, 2, '5. 4. 3 and 6 % water agar. The number of prev killed or dcsoured was counted after 24 hr.
Acliveness (activity) was measured in terms of two jiaraioelers. In the firsi experiment whei-e different pre\ were used, it was expressed as di.s])lacemenl. i.e., distance i mmj moved [)er minute while in the lemjierature and agar concentralion experiments it was expressed as waves per minute. A swing of the head to (me side, then back over the initial point to the oilier extreme and then back to the initial point was considered as one wave.
The frequency of predation of different nematode species by M. aquaiicus was found to \ a ry , Cepha-lobus sp. and Aglenchus parvus were preyed on more
Displacement
^ AgLenchus
parvus Cepha-lobussp
C symm-etncus
Prey
Mesorhab-i'iis sp
Pnsrnato-laimus sp
20 .
15 i
10 ^
5 a
Fig. f. FlTect of type of prey and prey acliveness on predation rate.
than the others. This diiferential predation rale was inversely related to the acliveness of the prey. The most active prey, Prismatolaimus sp.. was killed the least while the least active, Cephalobus sp., was devoured (he most [P'ig. 11. The number of C^epha-Inhus sp. killed was significantly higher [p < 0.05) than those of C. si/mmelricus or Mesorhabdilis sp. It was also higher than Prismalolainius sp. [p < O.OFi, but from Aglenchus parvus it showed no significant dilference (p > 0.03).
The rale of predation was also found to be influenced by differences in temperature. The optimum temperature appeared to be 23° at which maximum predation occurred (Fig. 2), and was significantly reduced at lower or higher temperatures ^p < 0.011. The increased activity of prey and predator increased the rate of predation while decreased activity reduced it. The a(4iveness of both the predator and the prey was maximum at 23°. Predation also depended on the conceniration of the agar medium used. One or 2 % water agar was more suitable for the predators
Section of Xematoloyy, Depiiriment of Zoology, Aligarh Muslim University, Aligarh - 202001, India. Comniowealth Institute of J'arasiloloyy, 395 A JIatfield Road, St. Albans, Herts. AL4 OXU, England.
Revue Mmalol. 6 (2) : 325-326 (1983) 325

33 .1 . /,. Bilijnimi, 1. Ahmad ct M. S. Jdira/puri
20
15
10
o—o Predator activeness •—• Prey activeness X—xf
CO
"30^
^ £ in '20 o a, >
in n)
"10
10 15 20 25 30 35 iO
Temp °C
MfZ. 2. KITccf of diflcrcnl tempcriilurcs on [)rc(lation rale.
more active, and il may be for Ihis reason that sitrni-ficantly more Cephalobus sp. were killed than Pria-matolaimwi ^p. Maertens (197r)) usino; J'rionchulus pundalus observed an averafi;e of ten Panagrelliis rediviviis killed per day while Nelmes (1974) using the same predator but Aphelenchus avenae as prey recorded (en kills [)er hour. It seems possible that tliis ditference could be due lo difTei-ences in prey behaviour, and one of the most likely causes may be the diilerenlial [)rey actisencss, as frec-livin": nematodes jiosscss hifi'tier end(i<i;enous ac t i \ i ty (Croll. 1972'. Besides, the experimental techniques miprht have also caused (iilTerences in predation.
Like [)rey activeness, predator activeness is also an important factor ii:overnin<i; predation. Those factors, such as temjierature and agar concentration, which influenced predator activity also influenced predation rate.
and not much dilTerence in the rate of predation was observed between them ;Fif>;. oj. Hii):her concentrations reduced predation si<>;nificanlly fp < 0.011 and in () % ap:ar no prey was killed. In this case also [)redati(m a[)])eared lo be related lo the acliveness of the predalors and I he prey as was the case for I.emfierature. (liffher affar concenirations reduced ac l i \ i ty and also firedation. Maximum predation occurred in 1 % agar.
From the data [iresenled, i t i s e \ i den t that .1 / . aqua-liciis showed no consistent predatory pattern with dilTer(>nt prey species and that some were devoured more than others. Such differences could {)robably be related to the acli\-eness of llie prey, the less active bein"- more vulnerable to attack than the
1 2 3 ^ 5 6
Agar concentration (%)
Fig. 3. I'lfTecl of different agar concentrations on predation rale.
.\cKxowi.i':i)(;t;Mh\Ts
We thank the Chairman, Dejiartment of Zoology for laboratory facilities and the l,'(JC and CSIR for financial assistance to the first and second aulhor.
Hi'.FERKxi;i:s
CoiiN, i;. & MoRDiocuAi, M, (1974). Experiments in suppressing citrus nematode populations by use of marigold and a predacious nematode, .\eniatol. medit., -1 : 43-53.
(JROI.I., N . A . I'197'2I. Mehavioural activities of nematodes. Ildminihol. Absir.. 41 iH) : 87-103.
(iROOTAKHT, P, & W'YSS, L'. :1979). L4trastru(4ure and function of the anterior feeding apparatus in Monon-chus aqualicus. .\ematologica, 2:") : 163-173.
J.\iHA,)Pi HI, M. S. & .\7,Mi, M. 1. (1978). Some studies on the predatory lK4iaviour of Mylonchuliia den-tatus. yemalol. medil., 6 : 20r)-21-i.
MAERTKXS, U . ,197.") .. Observations on the life cycle of Prioruimlus punctaius iCobb, 1917) (.Nematoda; and some culture conditions. Jiiol. Jb. Dodonaeu., 43 : 197-218.
NELMES, A. J. '1971). Evaluation of the feeding behaviour of Prionchulus punclatus, a nematode predator. J. anini. EroL, 43 : .^BS-OGJ.
SiMAr.!., R. W. (1979). 4"he effects of predatory nematodes on [)0[)ulation of plant-f)arasitic nematodes in pots. Xemaiologicfi, "io : 94-103.
Acrepte pour publication Ic 29 juillel 19H2.
32r) Revue M'malol. 6 (2) : 325-326 (1983)

Scmatid medil. l iyS4' . 12; 41-45.
4 Section of Nematology, Department of Zoology, Aligarh Muslim University, Aligarh-202001, India
OBSERVATIONS ON THE PREDATORY BEHAVIOUR OF MONONCHUS AQUATICUS
by A. L. B[LGRAMt, I. AHMAD and M.S . JAIRAJPURI
There have been few studies on the predatory potential of mon-onchs since the work of Steiner and Heinly (1922). Thorne (1927) suggested that Clarkus papillatus (Bastian) might be used to control sugar-beet nematodes. More recently, Nelmes (1974) described the feeding behaviour of Prionchtdus punctatus (Cobb). The life cycle of Mononchus aquaticiis Coetzee has been studied by Grootaert and Maertens (1976). Jairajpuri and Azmi (1978) studied the predatory behaviour of Mylonchuliis dentatiis.
This paper describes the predatory behaviour of adults and juveniles of Mononchus aqiiaticus Coetzee.
Material and Methods
Mononchus aqiiaticus were collected from the campus of the Aligarh Muslim University. Soil samples were processed by sieving and decantation and Baermann's funnel method. This method was used for all stages except the first stage juveniles, which were obtained by placing gravid females in 1% water agar at 25° C; the eggs laid by these females then hatched and the juveniles were collected.
The rate of predation was observed over a period of 24 h with five specimens each of adults and juvenile stages of M. aqiiaticus which
41

were kept separately with 25 individuals of Chiloplacus symmetncus (Thorne) or Cephalohus sp. as prey, in separate cavity blocks containing a small quantity of f/o water agar. Observations were made after 24 h.
To determine whether M. aquaticiis is attracted towards its prey, a plastic tube 5 cm long and 5 mm in diameter with a hole in the middle was used. At one end of the tube another 1 cm piece of tubiiig ol the same diameter was attached and a piece of filter paper was placed between the joint, thus forming a 6 cm long tube, divided into a 5 cm part and a 1 cm part. The tube was filled with agar from both ends and pieces of filter paper applied to prevent drying. 50 specimens of nematodes to serve as prey were put in the smaller part of the tube and the predator, M. aquaticiis, in the larger tube through the central hole. The distribution of predators was observed after 3 h by cutting the tube into 1/2 cm blocks. Attraction was tested by using C. symmetricus alone as prey or a combination of the following nematode species: C. symmetricus, Cephalohus sp. and Aglenchus parvus Siddiqi.
To detect whether the prey numbers affect the rate of predation, separate cavity blocks containing 25, 50, 100 and 150 specimens of C. symmetricus and Cephalohus sp. in l 'O water agar were introduced with five M. aquaticiis each. After 24 h the number of prey killed or ingested was counted for each block.
To find out whether M. aquaticiis showed any preference for prey, various combinations of prey were subjected to predation. The prey combinations used were: 20 C. symmetricus + 5 Cephalohus sp.; 20 C. symmetricus + 5 Mesorhahditis sp.; 20 Mesorhabditis sp. + 5 Cephalohus sp.; 20 Cephalohus sp. + 5 Mesorhahditis sp. The prey and predators were placed together in cavity blocks containing 1% water agar. The number of prey killed or ingested was counted after 24 h.
To determine the rate of feeding over a period of ten days, 5 M. aquaticiis were placed together with 25 prey specimens in cavity blocks containing 1% water agar. Observations were made at 24 h and the predators were transferred to fresh media containing 25 prey specimens of the same type. C. symmetricus and Cephalohus sp. served as prey.
The rate of predation of M. aquaticiis which were starved for 0, 2, 4, 6, 8, 10, 12 and 14 days was determined by using 25 specimens each of C. symmetricus and Cephalohus sp. as prey, separately.
— 42 —

Results
Mononchus aquaticus moves in a typical undulatory wave pattern. During locomotion contacts were made with the prey. After one of many such contacts the predator moved its head vigorously and attacked the prey. The attacks did not always result in killing as very often the prey moved out of the predators grip. The prey was usually attacked when full contact was made with the lip region of the predator. The predators showed no preference in attacking any particular part of the body of the prey. The smaller prey nematodes were swallowed whole but the cuticle of the larger ones was ruptured and the internal parts then ingested. The prey species apparently seemed unable to detect the presence of the predator and took no evasive action. Often the predator and the prey were seen moving in close proximity.
While observing the feeding behaviour of different stages of M. aquaticus, it was found that adults, fourth stage and third stage juveniles were predacious in behaviour. Adults and fourth stage juveniles killed the maximum number of prey during 24 h. The number of prey removed by these two was not significantly different (P > 0.05). Third stage juveniles also attacked the prey but in lesser numbers compared with adults and fourth stage juveniles. This low rate of predation among third stage juveniles was significantly different (P < 0.02) from the adults and fourth stage juveniles. Second and first stage juveniles were not predacious as they were not observed to remove any prey. They did not attempt to feed on prey even when lip contact was made with them. They appeared to ingest agar by rapid side ways movements of the head. Fourth and third stages preyed in the same manner as adults. Adults, fourth stage and third stage juveniles killed significantly (P < 0.01) more individuals of Cephalobus sp. than of C. symmetricus.
M. aquaticus was not attracted towards its prey either alone or in mixed populations. Movements appeared to be somewhat inhibited and the majority of the specimens remained in the zones adjoining the point of inoculation. The number of M. aquaticus on the two sides of the point of inoculation showed no significant difference (P > 0.05).
The abundance of prey did not influence predation as there was no significant difference (P > 0.05) in the rate of predation when 25,
43 —

%1 50, 100 or l.SO specimens cf Cephalobtts sp. or C. symmetricus wei'e used as prey.
M. aquaticHS when placed in \ar ious combinations of prey viz., Cephalohiis sp., Mesorhabdilis sp. and C. symmetricus, showed some degree of selection. Ccphalohns sp. in combination with Mesorhabditis sp. or C. symmetiiciis was always preyed on more than the other two nematodes. In the combination of Mesorhabditis sp. and C. symme-Iriciis the former was killed more frequently than the latter, but always less than Cephalobiis sp.; C. symmetricus was always devoured the least.
M. aquaticiis did not show any set pat tern of predation and in observations ovei' a period of ten days the number of prey killed by five predators was only slightly ditierent each day.
The starvation of predators did not alter the rate of predation. The difference in predation after two and fourteen days was not significant (P > 0.05).
Discussion
Steiner and Heinly (1922) and Esser (1963) suggested that predacious nematodes were unable to detect their prey even from a very short distance. Their opinion was supported by Grootaert and Maertens (1976) in their study of M. aquaticus. Our observations on .'W. aqnalicus also reveal that it was not at tracted towards potential prey species or their secretions. Predation depends purely on chance encounters, as was also suggested by Nelmes (1974) for P. punctatus. Theoretically, the increase in the number of prey should also inciease the probability of contacts and consequently also the rate of predation. Our observations contradict this as an increase in the number of prey did not significantly increase the rate of predation.
The prey catching mechanism, however, seems to vary slightly from that reported by Nelmes (1974) and Grootaert and Wyss (1979). While lip contact was essential, and contact with any other parts of the body did not provoke an attack, not all lip contacts initiated feeding or killing. Prey was attacked more frequently when the head of the predator made full contact with the prey, than when there were glancing contacts. Probing before attack has been described by Grootaert and Wyss (1979) but the same could not be detected in M. aquaticus. In P. punctatus short term deprivation of food tended
— 44

3B to enhance predation rate, while prolonged starvation tended to reduce it (Nelmes, 1974). Jairajpuri and Azmi (1978) also observed enhanced rate of predation in unfed Mylonchulus dentatus compared to well fed ones. With M. aquaticus however, starvation did not produce any significant effect on predation.
We thank the Chairman, Department of Zoology, for providing necessary laboratory facilities and Professor Ather H. Siddiqi for his suggestions. The first author is grateful to U.G.C. and the second to C.S.I.R. for financial assistance.
S U M M A R Y
The predatory behaviour of aduh and juvenile Mononchus aquaticus was studied in agar plates. The resuks indicate that M. aquaticus is not attracted towards its prey and contact is necessary to initiate an attack. Smaller prey nematodes were usually swallowed whole, but with larger ones only the internal parts were ingested. Adults, fourth stage and third stage juveniles were predacious, while second and first stage juveniles were assumed to feed on the agar. Prey number and starvation did not significantly influence the rate of predation.
L I T E R A T U R E C I T E D
Essi R R. P., 1963 - Nematode interactions in plates of non-sterile water agar. Proc. Soil Crop Sci. Soc. Fla., 23: 121-138.
(jROorAERT p. and M.\ERrE\s D., 1976 - Cultivation and life cycle of Mononchus aquaticus. Nematologica, 22: 173-181.
GROOrAERT P. and WYSS U., 1979 - Ultrastructure and function of the anterior feeding apparatus in Mononchus aquaiicus. Memalologica, 25: 163-173.
JAIRAJPI'RI M.S. and Az\ti M. I., 1978 - Some studies on the predatory behaviour of Mylonchulus dentatus. Neniaiol. inedit., 6: 205-212.
\'I:EMES A. J., 1974 - Evaluation of the feeding behaviour of Prionchulus punctatus, a nematode predator. J. Anim. EcoL, 43: 553-565.
SrEiNiiR G. and HEINLY H. , 1922 - The possibility of control of Heierodera radi-cicola and other plant injurious nemas by means of predatory nemas specially by Mononchus papillalus Bastian. J. Wash. Acad. Sci., 12: 367-385.
THOR\E G., 1927 - The life history, habits and economic importance of some mononchs. / . Agric. Res., 34: 265-286.
Accepted for publication on 3 September 1983.
— 45 —

irtdian J. Nematol l4(2), (1984) : 171-lf4
THE RESPONSES OF MONONCHUS AQUATICVS TO CHEMICALS AND pH
ANWAR L BILGRAMI and M SHAMIM JAIRAJPURl
Section of Nematology, Department of Zoology, Aligarh Muslim University, Aligarh-202001, India
Abstract Different concentrations of salts and furadan 3 G and pH grad ents were used to determine their effects on the activity of the predatory nematode, Mononchus aquaticus Except urea, the higher coneentrations (0 06M) of all the other salts proved highly toxic and except for cupric sulfate and ammonia, the lower concentrations (OOOIM) had stimulating effect Furadan 3-0 proved very toxic, even at low concentration, suppressing activity much belnw control pH 5 8 proved optimal for nematode activity The effectiveness of salts, with some exceptions, was directly proportional to their molecular weight.
Key words Mononchus aquaticus, responses, effect of salt concentration.
The toxicity of chemicals on different species of nematodes has been demons trated to vary (Pitcher & McNamara, 1972, Stephenson, 1945, Johnston, 1959 and Banage & Visser, 1965). Jairajpuri et al. (1974) determined the effect of pH and mineral salts on the survival of Hoplolaimus mdicus, Helicotylenchus indicus, Xiphmema basin and Mylonc hulus minor
In this work, the effects of some chemicals including a nematicide and hydrogen ion concentration on the predatory nematode, Mononchus aquaticus Coetzee, 1968 are reported.
Four concentrations (0 001, 0 01, 0 02 and 0 06 M) of cupnc sulfate potassium chloride, potassium nitrate, calcium chloride, urea and ammonia, and six concentrations (5, 25, 50, 100, 200 and 400 ppm) of the ne-raaticide, Furadan 3-G, were prepared Boric acid borate buffer of pH gradient 4, 5 4, 5 8,74 and 8 were used Distilled water served as control treatment.
The activity of M. aquaticus was measured in terms of its head movements One dorsal and one ventral head movement was considered as one wave. The observations were made, 1, 2, 4, 6, 8, 10
and 12 h after transferring the nematodes into a medium. In Figs 1 & 2, nematode activity IS plotted in terms of percentage increase and decrease compared with control. The concentrations, in which activity ceased withm the first hour, have not been plotted.
The nematodes were regarded as dead when repeated p-oddmgs with a needle 15mts. after immobilization, failed to produce any sign of activity Each treatment was replicated ten times and their mean values were calculated
M aquaticus were collected from soil samples of the Aligarh Muslim University Campus. The extracted nematodes were rinsed repeatedly in distilled water and kept in It till use Only adult M aquaticus were tested to chemicals and pH.
RESULTS
Activity in salts : No noticeable differences were observed in the activity of M. aquaticus at low concentrations, but at higher (concentrations the differences became significant.
All salts at 0.06 M pro.ved toxic. The activity declined sharply from 1 h in all

AO
172 ANWAk L. BILGRAMl AND M. §HAMIM JAIRAJPURI
80'
> • • ^
> <
c o o
> o m
60
^0
2U
Control
:s 20"
J. o 60
CuSO/i KN03
» _ KCL
0—a CaCl2
»—• Urea
. ^ N H 3
— I H
Hours
-+- •4 10 12
80"
— C > o
I 20"
Control
t^ 20
2 40-c
5 6ff o
80-
B
80" 5=
0 60" 2^ ^ . - c > 0 - " 40 0
> 0 -S 20
Control
:» 20"
^1 40-E c
*> 0
< J 60' 0 CJ
't _ 1 ^
-
- \
K
X-\
•— D
• • '
^ ^ x —
c , • - •
' • \ .
"**-A
80 4 - 4-
4 6 8
Hours
^ - 1 -10 12 10 12
Fig. 1 . Activity pattern m : A—0.06 M concentration, B—0.02 M concentiafion, C—0 01 M concentration, D—0.001 M concentration of salts.

41
RESPONSE OF MONONCHUS AQLJATICUS 173
concentrations and ceased in 4 h (Fig. 1, A) except in the case of urea where significant reduction in activity was recorded after 12 h (p<0.001).
At 0.02 M, activity was maximum during ] h but subsequently decreased below control level in potassium chloride, potassium nitrate and calcium chloride. In cupric sulfate, the nematodes did not survive after 1 h, while in urea they showed a significant (p<0.02) increase in activity from the 1 h to 12 h (Fig. 1,B).
Calcium chloride and urea stimulated activity at 0.01 M (Fig. 1, C) but in other salts it declined after 1 h.
The most favourable concentration for the activity of A4 aquutious proved to be 0.001 M. Enhanced activity above control was evident in all sahs except cupric sulfate
and ammonia (Fig. 1, D). In all salts, activity increased after 1 h.
Activity in Furaclan 3 G : All concentrations proved toxic and greatly suppressed activity much below control. Even at 5 ppm, a very low rate of activity was evident although the nematodes did survive for more than 12 h (Fig. 2, A).
Activity in hydrogen ion concentrations : The maximum activity occurred at pH 5.8 (Fig. 2, B) and increased significantly (p< 0.001) from 1 to 12 h.
DISCUSSION
The effect of salts on activity was directly proportional to their molecular weight which supports Stephenson (1945) and Johnston (1959). The effectiveness of salts
80-
- 60' o :t: i
(J
< 2" iff 01
I 20' to
Control
:= 2ff
•4-1 O
5 o
X I
6or
8ff i
• — • 5 ppm ^
0—o 25 ppm
• — , 50 ppm
A—4 100 ppm
Fif. ; : Activity pattern in A—Furadan 3-G, B—pH gradients

42 174 ANWAR L. BILGRAMI AND M. SHAMIM JAIRAJPURI
on activity is given below and it corresponds with their molecular weight : cupric sulfate (mol. wt. 249)>potassium nitrate (mol. wt. 101.11 )>potassium chloride (mol. wt 74.56)>calcium chloride (mol. wt. 110 99) >urea (mol. wt. 60.00). Calcium chloride was an exception because it proved less toxic than potassium nitrate or potassium chloride despite its greater molecular weight. Ammonia, which has the lowest molecular weight of all the salts, proved highly toxic even at 0.001 M. The high toxicity of ammonia could be due to its nematicidal power, which results from its capacity to generate high osmotic pressure, even in very low concentration (Vassallo, 1967).
The observations suggest that M. aquati-cus is greatly influenced by salts, particularly cupric sulfate which is highly toxic. It is also vulnerable to slight changes in hydrogen ion concentrations. Urea, which is frequently used as a fertilizer, enhanced activity of M. aquations at different concentrations compared to the other salts. The high toxicity, at all concentrations, of Furadan 3-G suggests that the continuous use of these pesticides may result in lowering the population of mononchs by disturbing the natural ecosystem, which may also result in an increase in the population of plant parasitic nematodes to noxious level.
We thank Prof. Nawab H. Khan, Chairman, Department of Zoology for providing necessary laboratory facilities, to Prof. Ather H. Siddiqi for suggestions and Dr. Irfan Ahmad for the help. The first author is thankfull to the U.G.C., New Delhi for financial assistance.
REFERENCES
BANAGE, W.B. & VissER, S.A. (1965). The effects of some tatty acids and pH on soil nematodes. Nematologica 11 : 255-262.
JAIRAJPURI, M.S., AzMi, M I. & BAJAJ, H . K . (1974) Effect of pH and sail concentrations on the Survival of Hoplolaimus indicus, Helicotylenchus indicus, Xiphinema basiri and Mylonchuliis minor, indian J- Nemalol. 4: 171-181.
.JOHNSTON. T .M. (1959). Effect of fatty acid mixtures on the rice siylet nematode i.Tylciichorli}nclws martini Fielding, 1959). Nature, London 183 : 1392.
PITCHER R .S . & MCNAMARA, D . C . (1972). The toxicity of low concentrations of silver and cupric ions to three species of plant parasitic nematodes. Nematologica 18 : 385-390.
STEPHENSON, W (1945). The effect of aci s on soil nematode. Parasitology 36 : 158-164.
VASSALLO, M A. (1967) The nematicidal power of ammonia (Abstract). Nematologica 13: pp 155.
Accepted for publicftion : March, 1984

43
202 SHORT COMMUNICATIONS
Cannibalism in Mononcfaus aquatlcus
ANWAR L Bl lGRAMI nmlM SH \ M I M J AlRAJPURl
StciiOFi of Ncmaloloey, IXp i r lm tn l of 7ooloRy, AliRjrl i Muslim IJnnirMly, AliiMiii-2n2O0l, India
Cannibalism in difTercnt stages of A/ aquaiuus was studied using 25 juveniles of each stage (first, second, third and fourth) alongwith 5 adults with each stage separately Further, juvenile juvenile combinations were used also in the following manner 5 fourth stage f25 third stage, H- 25 second stage, +25 first stage, 5 third stage jvwcmles (-25 second stage, \ 25 fust stage, 5 second stage +25 first stage juveniles All experiments were carried out at con .tant temperature (25°C) and replicated five times Individuals killed or injured were counted at 6 h intervals during a period of 24 h When adults were in \arious combinations with the juveniles it w.is found that tirst and second stages were killed more than the others (fig 1, a & b) and significantly higher (p<0 01) than the number of third and fourth stage juveniles kilkd by the same number of adults (Fig I, c & d ) The dillcrcnce between lirst and second s(,igc juveniles killed was insigmlicant (p'>U 0^) When third, second and first stage ju\eniles were placed with 5 fourth staee juveniles separately, again the first stage juveniles were preyed upon more in comparision with the others (Fig 2 a, b & c) The rate of pre dation on first stage juveniles was also significantly higher (p<0 01) than the second or third stage juveniles The number of first stage juveniles killed by 5 third stage juveniles was significantly more (p<0 HI) than the number of second stage juveniles, where no
killing was recorded (Fig 3) During observations It was noted that young stages were more prone to attack by their older stagis Only on vi.iy few occasion it w,is found th.it M iiqtiaiKUs was attacked by fourth st.iizi Juveniles
The insignificant difference between IIK rale of cannibalism of adults and fouriji stage |n\cnilcs suggest that they are m par in their predator) ability The U.wer rate ol cannibalism of third stage luvenilcs than the adults and Uurtrt stage juveniles gives an idea ol then low predatory potential ( aiini-balism in \l)lon<hulii^ cicntalti'; was reported by A7mi & Jairajpuri (1979) who found it to resort to cannibalism when placed exclu sivcly with their own juveniles in absence of prey In M nt/uar/CHv, besides adults, foiiitli and third stage juveniles also showed c.inni-balistic behaviour when placed with their own juveniles in absence of prey Adults iiid finirth stage juveniles show ,i more slroiu' cannibalistic tendency than their joiiiiger stages I lie third stage juveniles are neither predatory nor cannibalistic in behavioui
We thank Prof Nawab H Khan, Chairman, Department of Zoology, for providing ncccssaiv laboratory facilities and to Prof Athcr H Siddiqui for his valuable suggestions The first author is also thank-full to U G C lor financial assistance
nrrrRiN( i AzMi M I A. JAIRAJPURI M S (1979) Ncmaiiil
Midi! 7 201 207

4
SHORT COMMUNICATIONS
2Ch
203
20,
J5 *-
:2a~
Fig.i
12 18 24 Time(h)
2(h
12 IB —I
24
Time (h)
Fig. 3
Fig. 1. Ujte of cannibalism in adult /M oquaiirui (d^5A I-2M,; b^5A+25J,; c = 5A+25Jj; d=5A I-25J4 )-
Fig. 2. Rate of cannibalism in fourth stage juveniles of M. aqiiali'us. (a=3J|+25Ji; b = 5J|f25J,;c=5J| + 25J3)
Fig 3. Rale of cannib.Tlism in third stage juveniles of W oquaiiLUs (5Ji + 25J)) Kl Y-A--Adults; Jj—First stage juveniles, J] -Second stage juven.les, Jj— Third stage juveniles; Jj—Fourih stage juveoiles.
Key—A—Adults, ),, Jj J, & Jj-Firsl, second, third & fourth stage juveniles.

^^
Factors influencing attraction of adult Hirschmanniella oryzae towards cabbage seedlings
Anwar L. BiLGRAMi, Irfan AHMAD and M. Shamim JAIRAJPURI
Section of Nematology, Department of Zoology, Aligarh Muslim University, Aligarh-202001, India.
SUMMARY
Observations on various factors influencing attraction of Hirschmanniella oryzae towards cabbage roots revealed that all factors except coloured lights and darkness affected attraction. Responses of H. oryzae declined as the age of the roots increased and also with an increase in the period of food deprivation of nematodes. Maximum attraction was recorded towards 4-6 day-old cabbage roots with fed nematodes as well as 2, 4 and 6 day unfed ones. Live roots attracted nematodes but roots killed by heating lost attractiveness. Maximum number of nematodes were attracted when roots were incubated for 10-12 h in 3-4 mm thick agar layer of 0.5 % concentration at 20-25°. The nematodes showed preference to the region of root tip and aggregated most around it. Attraction decreased with increase in the distance of inoculation of nematodes from the roots.
RESUME
Facteurs influengant I'attraction des adultes d'Hirschmarmiella oryzae par les planlules de chou
L'etude de I'influence de differents facteurs sur I'attraction des adultes d'H. oryzae par le chou revele que tous ces facteurs, sauf les lumieres colorees et I'obscurite, ont un effet. Les reponses d'H. oryzae diminuent en intensite lorsqu'augmentent soit I'age des racines soit la duree du jeune chez les nematodes. L'attraction maximale est observee avec des plantules de chou de 4-6 jours et des nematodes fraichement nourris ou soumis au jeune depuis seulement 2, 4 et 6 jours. Les racines vivantes sont attractives, mais perdent ce caractere lorsqu'elles sont tuees par la chaleur. Un nombre maximum de nematodes sont attires lorsque les racines sont incubees pendant 10-12 heures dans une couche de 3-4 mm d'agar a 0,5 % maintenue a 20-25°. Les nematodes montrent une preference pour rextremite apicale de la racine autour de laquelle la plupart d'entre eux se rassemblent. L'attraction decroit lorsqu'augmente la distance entre la racine et le lieu d'inoculation des nematodes.
Several workers have studied the attraction and feeding behaviour of plant-parasitic nematodes (Luc, Lespi-nat & Souchaud, 1969; Azmi & Jairajpuri, 1977a; Prot & Netscher, 1978; Prot, 1980; Wyss, 1981) but few have dealt with the factors influencing attraction of nematodes towards plant roots. Attractiveness was proportional to the rate of root growth (Weiser, 1955; Lavallee & Rohde, 1962) but if the roots were killed by heating they lost their attractiveness (Linford, 1939; Bird, 1962). Lownsbery & Viglierchio (1961) suggested that roots became unattractive when their growth ceased. Moussa, De Grisse & Gillard (1972) did not observe any significant differences in the attraction of Globodera rosto-c/ziemw juveniles when coloured Ughts were used. Davies & Fisher (1976) found temperature, initial inoculum density, duration of exposure and the distance at which the juveniles were placed from the root to be limiting factors in the invasion of barley seedUngs by Heterodera avenae juveniles. Prot and Van Gundy (1981a; 1981b) concluded that the soil texture, clay components, photo-period and temperature acted as influencing factors on the migration of the juveniles of Meloidogyne incognita and M. hapla towards tomato roots. Recently, Prot and
Komprobst (1983) observed decreased rate of penetration of tomato roots by M. javanica juveniles when an increased concentration of seed extract was used.
The present work was undertaken to find out the effect of various biotic and abiotic factors on the attraction of adult Hirschmanniella oryzae (Van Breda de Haan, 1920) Luc & Goodey, 1963 to cabbage seedlings (Brassica oleracea L. var. capitata) in agar plates. Bilgrami (1983) has shown that cabbage roots attract adult and juveniles H. oryzae though it is not as good a host as rice (Oryza saliva). The present observations were made on cabbage seedlings to study in detail the mode of attraction of adult H. oryzae.
Materials and methods
The soil samples containing H. oryzae were collected from paddy fields near the village Jamalpur, Aligarh and the nematodes were extracted by decantation and sieving and modified Baermann's funnel techniques.
Most experiments were carried out by Petri-dish techniques (cf. Ahmad & Jairajpuri, 1980). Excised root tips of 4-6 day old cabbage seedlings were placed in the
Revue Nematol. 8 (I): 67-75 (1985) 67

u A. L. Bilgrami, I. Ahmad &. M. S Jairajpun
inner circle of the Petri-dishes containing 0.5 % water-agar and incubated for 12 h. Five adult H. oryzae were then introduced at different places on the reference circle. Observations on the distribution of nematodes were made after 4 h and were converted into scores and log scores. All experiments were carried out at 25 ± 1° and replicated fifteen times. Controls were run without host roots.
The above conditions were similar in all experiments unless mentioned otherwise.
EFFECT OF DIFFERENT AGE GROUPS OF ROOTS ON THE ATTRACTION OF FED AND UNFED NEMATODES
To determine the effect of different age groups of cabbage roots on the attraction of fed (kept on roots in agar for one day) and unfed (kept without roots in tap water) nematodes, six age groups of roots viz., 2, 4, 6, 8, 10 and 12 day old were subjected to attraction of 0, 2, 4, 6, 8, 10 and 12 day unfed nematodes. The water in which they were placed was changed each day. Each group of unfed nematodes was tested towards each age group of the roots separately. The 0 day refers to fed nematodes.
EFFECT OF LIVE AND DEAD ROOTS
The influence of live and dead cabbage roots on the attraction of H. oryzae was observed by placmg in the inner circle of separate Petri-dishes and incubating prior to the introduction of nematodes. Cabbage roots were killed by boiling in water for 5 min.
EFFECT OF LENGTH OF INCUBATION PERIODS OF ROOTS
To observe the mfluence of different periods of incubation of roots on the attraction of nematodes, roots were incubated separately for 2,4,6,8,10 and 12 h prior to the introduction of nematodes.
EFFECT OF REGIONS OF ROOT
To observe the effects of different regions of the root on the attraction two experiments were performed. In the first experiment attraction was tested towards intact non-excised roots. Water-agar (0.5 %) was poured on a glass slide and was allowed to cool. The root was placed in the agar parallel to the length of the slide and divided into three 1 cm regions by markings lines at the bottom of the slide (Fig. 1). The shoot remained outside the agar. After incubating roots for 12 h, 25 adult H. oryzae were introduced at the edges of the slide opposite the middle region of the root. Distribution of nematodes was observed after 4 h.
In the second experiment the Petri-dish technique was used. A 3 cm long excised root was cut into three equal parts viz., root tip (0-1 cm); zone of elongation (1-2 cm) and zone of maturation (2-3 cm). These were incubated in separate Petri-dishes prior to nematode introduction.
0-1 1—2 2—3 cm cm cm
Glass slide"
Agar Point of inoculation of nematodes
Fig. 1. Expenmental design for testing the anraction of H. oryzae towards different regions of cabbage roots 0-1 cm : root tip; 1-2 cm : zone of elongation; 2-3 cm : zone of maturation.
EFFECT OF AGAR THICKNESS
To determine the effect of different thicknesses of agar on the attraction, nematodes were placed with roots in Petri-dishes containing 1, 2, 3, 4, 5 and 6 mm thick agar layers.
EFFECT OF AGAR CONCENTRATION
To observe whether agar concentrations influence attraction, roots were incubated in separate Petri-dishes containing 0.5, 1, 2, 3, 4, 5 and 6 % water-agar prior to the introduction of nematodes.
EFFECT OF LIGHT AND DARKNESS
Light from 60 Watt lamp kept at a distance of 65 cm from the Petri-dish was considered as normal light. Coloured effects were obtained by covering the lamp with red, yellow, green and blue cellophane papers. Roots were incubated in Petri-dishes under the different light conditions after which the nematodes were introduced and the dishes were replaced under similar lights.
EFFECT OF TEMPERATURE
To find the effect of temperature on the attraction of H. oryzae the roots were incubated in agar at 5, 10, 15, 20,25,30 and 35°. The nematodes were then introduced and Petri-dishes were again kept at the same temperature at which they Vi ere stored previously.
EFFECT OF DISTANCE OF ROOT ON ATTRACTION
The distance from which nematodes can be attracted towards roots was .studied in 7 cm Petri-dishes. These were marked into 7 equal zones at the bottom similar to Azmi and Jairajpun (1977a). A 1 cm long part of root from cabbage seedlings was placed in zone 1 and incubated for 12 h. After incubation 25 H. oryzae where introduced in zone 2. Similarly, the nematodes were introduced in zone 3,4, 5,6 and 7 of separate Petri-dish and their distribution was recorded after 4 h.
68 Revue Nematol. 8 (1): 67-75 (1985J

Attraction o/Hirschmanniella oryzae towards cabbage roots
Results
The attraction of H. oryzae was influenced by all factors except coloured lights and darkness.
EFFECT OF DIFFERENT AGE GROUPS OF ROOTS ON THE A T T R A C T I O N OF FED AND UNFED NEMATODES (Fig. 2 and 3)
All groups of roots attracted fed nematodes except 12 day old roots (mean log score 1.35; p > 0.05). Maximum attraction was recorded towards 4 and 6 day old cabbage roots (mean log score 2.59 and 2.58 respectively) (Fig. 2). This was higher than the attraction towards 2 day (mean log score 2.07) and 8 day old roots (mean log score 2.24) or 10 day old roots (mean log score 1.98) (p <0.05). The attraction towards 2, 8 and 10 day old roots was not different from each other (p > 0.05).
The two day unfed nematodes were attracted most by 4 and 6 day old roots (mean log score 2.59 and 2.55 respectively) (Fig. 3, A). The roots of 2, 8 and 10 days attracted lesser number of nematodes compared to 4 and 6 day old roots (p < 0.01). No attraction was observed towards 12 day old roots (mean log score 1.45, p > 0.05).
Similarly, when 4, 6 and 8 day unfed nematodes showed maximum attraction towards 2, 4 and 6 day old roots (Fig. 3, B, C, D) which declined significantly in the presence of higher are groups and became unresponsive towards 12 day old roots (p > 0.05).
H. oryzae which were deprived of food for 10 days showed positive response towards 2, 4, 6 and 8 day old cabbage roots (Fig. 3, E) and there was no difference in the attraction towards individual age groups (p > 0.05). The nematodes were unresponsive towards 10 and 12 day old roots (mean log score 1.45 and 1.42 respectively, p > 0.05).
Nematodes kept away from food for 12 days showed maximum attraction towards 4 day old roots (mean log score 1.76). This attraction was not different either from 2 day old (mean log score 1.75) or 6 day old roots (mean log score 1.70) (p > 0.05). However, roots 8, 10 and 12 day old did not attract nematodes significantly (p > 0.05) (Fig. 3, F).
EFFECT OF LIVE AND DEAD ROOTS (Fig. 4)
Live roots attracted nematodes (mean log score 2.55, p < 0.05) while dead roots failed to attract them (mean log score 1.38, p > 0.05).
EFFECT OF LENGTH OF INCUBATION PERIODS OF ROOTS
(Fig. 5)
The period of incubation of roots influenced the degree of attraction. While 2 and 4 h incubation did not attract the nematodes (mean log score 1.18, p > 0.05), the attraction became positive only after 6 h (mean log score 1.63, p < 0.05). Further, it increased significantly up to 10 and 12 h incubation (p < 0.05). There was no difference in the attraction after 10 or 12 h incubation (mean log score 2.40 and 2.55 respectively, p > 0.05).
2£5i 2.5a
2-2CLJ o
^ 1-9CLI o
c 1.601 U
1.301
1.00 I "I 10 12
Agt of rooig (days)
Fig. 2. Effect of different age groups of cabbage roots on the attraction of fed H. oryzae (Black circles : control; open circles : with roots).
EFFECT OF REGIONS OF ROOT (Fig. 6 & 7)
In Petri-dish experiments when excised roots were used maximum attraction (mean log score 2.58) was observed towards 0-1 cm region of roots, while least (mean log score 1.80) towards 1-2 cm region (Fig. 6). Similar results were obtained in glass slide experiments also when whole roots were used (Fig. 7). However, in Petri-dish experiments the attraction towards 0-1 cm was more than towards the 2-3 cm region (mean log score 2.20, p < 0.05) while in glass slide experiment there was no difference (p > 0.05).
EFFECT OF AGAR THICKNESS (Fig. 8)
In various thicknesses of agar, the nematodes responded positively but their attraction was influenced by different agar layers. Attractiion was maximum in 3 and 4 mm (mean log score 2.51 and 2.38 respectively) but declined in other layers, thicker or thinner than these two
Revue Ne'matol. 8 (1): 67-75 (1985) 69

4 8 A L Bilgrami, I Ahmad & M S Jairajpun
ZGSn 2.5a
Z
V3a
1.00.
o
2.65-1 2.5a
2.2a
1.90.
| i . 6 a j
z
1.30.
1.00
• — • — • -
1 I I I I ' I 2 A 6 8 10 12
1 I 1 1 I I 2 I. 6 8 10 12
zsa
2.2a V 0
g1.60. « Z
t30.
too
• — * ^
•—•—
' 1 1
E
\ - .
-•—,—••—• — 1 — 1 1 1
2 A 6 8 10 12 Agt of roots (day^
2.6§-| 2.50. • — • — •
2.20.
1.9a
1.60.
UO.
1.00
2.65-, 2.5a
2.2a
1.9U.
1.60.
1.30.
1.00
2.65 2.5a
2.20.
1.90.
1.60.
1.3a
1.00
B
• — — • •
1 I 1 I I 1 2 4 6 8 10 12
' \ . • \ .
T r - — I 1 1 1 2 4 6 8 10 12
•—•Control
0—0 With roots
F
\
. « — • — ^ • -• — - •
— •
2 k 6 8 10 12 Ago of roots (days)
Fig 3; Effect of different age groups of cabbage roots on the attraction of unfed H oryzae A : 2 day unfed adults, B : 4 day unfed adults; C 6 day unfed adults; D 8 day unfed adults, E : 10 day unfed adults, F 12 day unfed adults
70 Revue Ne'matol. 8 (1) 67-75 (1985)

Attraction o/Hirschmanniella oryzae towards cabbage roots 49
s
2.5a
1.90.
2 1.60J X
1.30.
Control Live roots Dead roots
• ^
% ZA Roots
Fig. 4. Effect of live and dead cabbage roots on the attraction of H. oryzae.
2.65^ 2.5a
2.2a «
§1.904 ot o cl.BQj 10 V
1.30,
1.00
Control 0-4 cm
1—2 cm
2-3cm
/ / / /
y///
7''/' Region of roots
Fig. 6. Attraction of H. oryzae towards different regions of excised cabbage roots (Petri-dish experiments). 0-1 cm : root tip; 1-2 cm : zone of elongation; 2-3 cm : zone of maturation.
2.651 2.50-
2204
I .
§ 1.90.
o c 1.60J n «>
1.30.
1.00
•—•Control e—oWith roots
T-I, 6 8 10 12
Incubation period of roots(h9
Fig. 5. Effect of length of incubation periods of cabbage roots on the attraction of H. oryzae.
11 10,
o
E 6 «» c
6 Z
2.
77;
i 0-1 cm 1—2cm 2—3cm
Region of roots
Fig. 7. Attraction of H. oryzae towards different regions of intact cabbage roots (Glass slide experiments). 0-1 cm : root tip; 1-2 cm : zone of elongation; 2-3 cm : zone of maturation.
Revue Ne'matol. 8 (1): 67-75 (1985J 71

50 A. L. Bilgrami, I. Ahmad & M. S. Jairajpun
o
2.651 2.5a
2.2a
1.90.
= 1.6QJ n
1! 1.3a
1.00
• — • Control 0—e w i th roots
— I I I I 1 I 1 2 3 ^ 5 6
Agar thickness (mm)
Fig. 8. Effect of agar thicknesses on the attraction of H. oryzae towards cabbage roots.
(p < 0.05). The attraction was least in 6 mm (mean log score 2.01).
EFFECT OF AGAR CONCENTRATION (Fig. 9)
The attraction was greatly influenced by all concentrations of water-agar, except 0.5 and 1 % in which it was
2.65-1 2.50L
« 2.20. o o M » 1-90.
S1-50L| «
i.3aJ
, _ , Control
o—oWith roots
— 1 — f 1 1 1 r-06 1 2 3 4 5
Agar concentration (Ve)
- I 6
Fig. 9. Effect of agar concentrauons on the attraction of H. oryzae towards cabbage roots.
maximum. Attraction in 2 % water-agar (mean log score 1.83) declined significantly (p < 0.01) and there was no attraction in 3, 4, 5 and 6 % agar dishes (p > 0.05).
EFFECT OF LIGHTS AND DARKNESS (Fig. 10)
Red, yellow, fn"een, blue, normal lights and darkness did not significantly influence attraction of nematodes (p > 0.05).
2.65-2.saJ
o
CD
o
2 ^
tsa
c I6QJ « X
\ 3a
1.00
1—I 1;^ Control
i 1 1 Nr. Rd. Gr. yw. 81 . Ok.
Fig. 10. Effect of lights and darkness on the attraction of H. oryzae towards cabbage roots (Nr : Normal; Rd : red; Gr : green; Yw ; yellow; Bl: blue; Dk : darkness).
EFFECT OF TEMPERATURE (Fig. 11)
Attraction was influenced by different temperatures. Maximum attraction (mean log score 2.55) was at 20 and 25° and was higher than the attraction at 15 or 30° (p < 0.01). Nematodes were attracted least at a temperature of 35°. Lower temperatures of 5 and 10° proved
2.55-1 •—•Control 2.5CL
2.20. «i u O
•" 1.90. at o
c 1.604
z
1.30.
1.00
With roots
« = -
To 15 20 25 30 35
Temperature fC)
Fig. 11. Effect of temperatures on the attraction of H. oryzae towards cabbage roots.
72 Revue Nematol. 8 (1): 67-75 (1985)

Attraction o/Hirschmanniella oryzae towards cabbage roots 51
unfavourable and no positive response was observed (p > 0.05).
EFFECT OF DISTANCE OF ROOT ON ATTRACTION (Fig. 12)
The number of nematodes reaching zone 1 increased as the distance of their introduction from the root decreased. Maximum numbers were recorded in zone 1 when the nematodes were introduced in zones 2 and 3 i.e., 1 cm and 2 cm from roots. They also showed positive
response when placed 3 cm and 4 cm away from the roots i.e., zone 4 and 5 but not when introduced in zones 6 and 7 i.e., 5 and 6 cm from the roots.
Discussion
From the above observations it is evident that as the cabbage roots grew older, their attractiveness decreased and failed to elicit a response from fed nematodes,
16.
12.
M
? 12.
I. c
i 1&1
M
1 12J
S ^
o t..
' ' • - - ' 1 2 3 4 S 6 7
Dh I I I 1 1 2 3 ^ 5 6 7
i hAJ 1 2 3 4 5 6 7
Zones
20-1
16.
12
8.
4,
16-j
12.
8.
i.
16-1
12
8.
i.
I i ••>
1 2 3 4 5 6 7
a=3 1 2 3 4 5 6 7
h m 1 2 3 4 5 6 7
Zones
Fig. 12. Effect of distance of cabbage roots on the attraction of H. oryzae.
Kevue Nematol. 8 (1).- 67-75 (1985) 73

^'? A L Bilgrami, I Ahmad & M S Jaimjpun
suggesting that they were no longer attractive Similarly, as the period of food depnvation of nematodes increased their response towards increasing age groups of roots also decreased indicating that besides the unattractive-ness of roots some other factor(s) within the nematodes was (were) also governing their responses Doncaster and Seymour (1973) concluded that in starved nematodes the mimmum response threshold decreased and unfed nematodes could percieve weaker sdmuh These observations show that in H oryzae, perhaps, depnvation of food caused an increase in minimum response threshold as 12 day unfed nematodes failed to respond to some of those age groups of roots which attracted lesser unfed nematodes or perhaps, a loss in the degree of attraction with an increase in the penod of food depnvauon may have resulted from loss of energy, as starving nematodes utihze their reserve food matenal (Cooper & Van Gundy, 1970, Reversat, 1981a, 1981b) Hence, both factors, age of roots and penod of food depnvation appear to independently influence attraction of H oryzae towards host roots
Weiser (1955), and Lavallee and Rohde (1962) found attracuveness of roots proportional to their growth and Viglierchio (1961) observed attraction to be independent of the age of the roots The present observations are contrary to both This could be due to different hosts and nematode species involved but it may also be possible that all nematodes are not equally sensitive to the same stimulus or all plants do not produce similar attractants (Prot, 1980) The possible inhibition of growth of roots in Petn-dishes contaimng only water-agar may have also influenced attraction of H oryzae as was suggested by Viglierchio (1961) Moreover, roots killed by heating lost their attractiveness (Linford, 1939, Bird, 1962) which confirms the results on the responses of adult H oryzae towards live and dead cabbage roots
The observations on the effect of vanous coloured lights support the findings of Moussa, De Gnsse and GiUard (1972) who also didn't find any difference m attraction of )uvemles of Globodera rostochiensis
The influence of different incubation penods of roots possibly suggest that a minimum penod is necessary for the development of perceptible attractant gradients by the roots The effect of agar concentrations and temperatures on the attraction may be related to the activity of nematodes Wallace (1969), and Azmi and Jairajpun (1977b) suggested that the activity of nematodes declined in increasing agar concentrations Perhaps this reduced activity inhibited attraction of H oryzae adults in more than 2 "o water-agar Similarly, it has been shown (Wallace, 1963) that nematodes became inactive at temperatures 5-10° and 30-40° and that activity of Meloidogyne incognita and M hapla juveniles was inhibited by low temperatures (Prot & Van Gundy, 1981b) Thus no response m H oryzae at 5-10° may be attn-buted to inhibited activity Besides, the diffusion of root attractants in the agar may also have been influenced by
different temperatures and agar thicknesses which resulted m different nematode responses
The maximum attraction of H oryzae towards root tip, both excised as well as intact, suggests that perhaps this IS the most attractive region as was also proposed by other workers (Pitcher & MacNamara, 1970, Schilt & Cohn, 1975, Stirling, 1976) However, excised roots may introduce into the medium additional unknown substances due to cell breakdown But these substances apparently did not seem to influence behaviour as the attraction towards different regions of roots, excised and intact, showed a similar pattern
As compared to Globodera rostochiensis juveniles which migrated upto 45 cm in soil (Rohde, 1962) and Anguina tntici juveniles which travelled 30 cm upwards to reach their host (Leukel, 1962), H oryzae showed a comparatively weak response as it could reach its host only from a distance of 4 cm which is similar to Dity-lenchus dipsaci (Blake, 1962) Prot (1977) has shown a 50 cm honzontal and vertical migration in soil of Meloidogyne javanica in 9 days towards a susceptible tomato vanety Similarly, the juveniles of M incognita migrated upto 20 cm in soil to reach tomato roots (Prot & Van Gundy, 1981a)
ACKNOWLEDGEMENTS
We thank Prof Nawab H Khan, Chairman, Department of Zoology for providing laboratory facilities and Prof Ather H Siddiqi for his valuable suggestions The first author is thankful! to U G C , New Delhi for financial assistance
REFERENCES
AHMAD, I & JAIRAJPURI, M S (1980) Sex attraction and copulation in Chiloplacus symmetncus (Nematoda Acro-belinae) Nematologica, 26 139-148
AZMI, M I & JAIRAJPURI, M S (1977a) Attracuon of plant-parasitic nematodes to host roots Nematologica, 23 119-121
AZMI, M I & JAIRAJPLRI, M S (1977b) Studies on nematode behaviour III, LocomoUon in juveniles of Hoplolaimus indtcus and Hehcotylenchus indicus in different concentrations of agar Indian J Nematol, 5 240-242
BILGRAMI, A L (1983) Attraction of Hirschmanniella oryzae towards cabbage seedlings Proc Third Nematol Symp, Solan, India (Abstr) 15
BIRD, A F (1962) Onentation of the larvae of Meloidogyne javanica relative to roots Nematologica, 1 275-287
BLAKE, C D (1962) Some observauons on the onentation of Dnylenchus dipsaci and invasion of oat seedlings Nematologica, 7 177-192
COOPER, A F & VAN GUNDY (1970) Metabolism of glycogen and neutral lipids by Aphelenchus avenae and Caenorhab-ditis sp m aerobic, micro-aerobic and anaerobic environments J Nematol, 2 305-315
DAVIES, K A & FISHER, J M (1976) Factors influencing the
74 Revue Nematol 8 (I) 67-75 (1985)

Attraction o/Hirschmanmella oryzae towards cabbage roots
number of larvae of Heterodera avenae invading barley seedlings tn vitro Nematologtca, 22 153-162
DoNCASTER,C C & SEYMOUR, M K (1973) Exploration and selection of penetration site by tylenchids Nematologica, 19 137-145
LAVALLEE, W H & RoHDE, R A (1962) Attractiveness of plant roots to Pratylenchus penetrans (Cobb) Nematologica, 8 252-260
LEUKEL, R W (1962) Nematode disease of wheat and rye Farmer's Bull, 1607 1-16
LiNFORD, M B (1939) Attractiveness of roots and excised shoot tissue to certain nematodes Proc helminth Soc Wash, 6 11-18
LowNSBERY, B F & ViGLiERCHio, D R (1961) Importance of response of Meloidogyne hapla to an agent from germinating tomato seeds Phytopathology, 51 219-221
Luc, M , LESPINAT, P & SOLCHAUD, B (1969) Marquage direct de Hemicycliophora paradoxa par le phosphore ra-dioactif Utilisation pour I'etude des deplacements des nematodes phytoparasites dans le sol Nematologica, 15 35-43
MoussA, F F , De GRISSE, A T & GILLARD, A (1972) The effect of hght on the attractiveness of tomato seedlings to larvae of the golden cyst nematode, Globodera rostochiensis Med Fac Landbouww Rijksuniv Gent, 37 315-321
PITCHER, R , S & MCNAMARA, D C (1970) The effect of nutntion and season of year on the reproduction of Tn-chodorus viruliferus Nematologica, 16 99-106
PROT, J -C (1977) Amplitude et cinetique des migrations du nematode Meloidogyne javanica sous I'lnfluence d'un plant detomate Cah ORSTOM, ser Biol, 11 157-166
PROT, J -C (1980) Migration of plant-parasitic nematodes towards plant roots Revue Nematol, 3 305-318
PROT, J C & KORNPROBST, J M (1983) Effects of Azadi-racta tndica, Hannoa undulata and Hannoa klameana seed extracts on the ability of Meloidogyne javanica juveniles to penetrate tomato roots Revue Nematol, 6 330-332
PROT, J-C &NETSCHER, C (1978) Improved detection of low population densities of Meloidogyne Nematologica, 24 129-132
PROT, J-C & VAN GUNDY, S D (1981a) Effect of soil texture and the clay components on migration of Meloidogyne incognita second stage juveniles J Nematol, \3 213-217
PROT, J C & VAN GUNDY, S D (1981b) Influence of photopenod and temperature on migranons of Meloidogyne juveniles J Nematol, 13 217-220
REVERSAT, G (1981a) Effects of ageing and starvauon on respiration and food reserve contents in adult Hirschman-niella spinicaudata Revue Nematol, i 125-130
REVERSAT, G (1981b) Consumption of food reserves by starved second stage juvemles of Meloidogyne javanica under conditions inducing osmobiosis Nematologica, 27 207-214
ROHDE, H (1962) Untersuchungen uber das Wandervermo-gen von Larven des Kartoffelnematoden (Globodera rostochiensis) in Modell Versuchen mit verschiedenen Bodener-ten Nematologica, 7 74-82
SCHILT, H G & CoHN, E (1975) Pathogenecity and population increase of Paratnchodorus minor as influenced by some environmental factors Nematologica, 21 71-80
STIRLING, G R (1976) Paratnchodorus lobatus associated with citrus, peach and apncct trees m South Australia Nematologica, 22 138-144
VIGLIERCHIO, D R (1961) Attraction of parasinc nematodes by plant root emanations Phytopathology, 51 136-142
WALLACE, H R (1963) 77ie biology of plant parasitic nematodes Edward Arnold, London, 280 pp
WALLACE, H R (1969) Wave foi-mation of infective larvae of the plant parasitic nematode Meloidogyne javanica Nematologica, 15 65-75
WEISER, W (1955) The attractiveness of plants to larvae of rootknot nematode I The effect of tomato seedhngs and excised roots on Meloidogyne hapla Chitwood Proc helminth soc Wash, 22 106-112
WYSS, U (1981) Ecto-parasitic root nematodes feeding behaviour and plant cell responses In Zuckerman, B M and Rohde, R A (Eds), Plant Parasitic Nematodes Vol III, New York, Academic Press 325-351
Accepte pour publication le 3 decembre 1984
Revue Nematol 8 (1) 67-75 (1985) 75

Responses of adult Hirschmanniella oryzae towards different plant roots
Anwar L. BILGRAMI, Irfan AHMAD and M. Shamim JAIRAJPURI
Section of Nemaiology, Department of Zoology, Ahgarh Muslim University, Ahgarh-202001, India
SLMMARY
Expenments were performed to study the host preference behaviour of Hirschmanniella oryzae and to observe the effects of plant root homogenate concentrations on the activity of H. oryzae Attraction was tested in 0.5 "o water-agar in Petri-dishes towards plant roots of cabbage, carrot, radish and onion either alone or in different combinations. The effect of 20, 40, 60, 80 and 100 "o concentrations of the above plant root homogenates was observed on the activity (head movements minute) of nematodes. Cabbage and carrot attracted most H oryzae while onion the least. Similarly, homogenate concentrations of cabbage and carrot roots increased activity of nematodes but those of onion and radish suppressed it markedly. The results suggest that H oryzae shows some degree of preference and attraction depends upon the host root, attractants and activity of nematodes.
RESLME
Reactions des adultes de Hirschmanniella oryzae envers differentes plantes hdtes
Une sene d'expenences a ete reaiisee pour etudier le comportement de Hirschmanniella oryzae envers differentes plantes hotes et pour observer les effets d'homogenats de racines sur I'activite du nematode. L'attraction par les racines de chou, de carotte, de radis et d'oignon a ete testee sur agar a 0,5 "o dans i'eau, en boite de Petn. Les effets de concentrations a 20, 40, 60, 80 et 100 "o d'homogenats de racines des plantes precitees sur I'activite du nematode (nombre de mouvements de la parti e anterieure par minute) ont ete egalement observes. Le chou et la carotte attirent le plus H oryzae et I'oignon le moms. De meme, les homogenats de racines de chou et de carotte accroissent I'activite du nematode tandis que ceux d'oignon et de radis la suppnment de fa^on nette. Ces resultats suggerent que H oryzae montre un certain degre de preference envers les plantes hotes et que l'attraction depend des racines de I'hote, des substances attractives et de I'activite du nematode.
There is ample evidence to suggest that plant-parasitic nematodes accumulate around roots of host plants (Lownsbery & Viglierchio, 1960; 1961; Azmi & Jairajpuri, 1977; Prot & Van Gundy, 1981a; 1981b). Viglierchio (1961) found a correlation between plant attractiveness and its efficiency as a host in case of Heterodera schachtii. He (I. cj also observed a strong attraction of Meloidogyne hapla juveniles towards a non-host (Kanota oat) and strong repulsion or weak attraction towards Merced Rye grass. There was no significant difference in the attraction of Meloidogyne hapla towards resistant and susceptible cultivars of alfalfa when tested separately but when compared simultaneously susceptible cultivars attracted more nematodes (Griffin, 1969). Lee and Evans (1973) found a strong correlation between the degree of attraction of rice seedling extract and its susceptibility to Aphelenchoides besseyi. Mathur and Prasad (1973), and Mohandas, Pattanik and Prasad (1979) observed the host range of rice root nematode, Hirschmanniella oryzae, on some commonly cultivated crops and weeds while Park, Han and Lee (1970) studied its behaviour towards different varieties of rice. The attraction and factors influencing the migration of
H. oryzae towards cabbage roots was studied by Bilgrami (1983) and Bilgrami, /Vhmad and Jairajpuri (1985).
In the present work, behaviour of adult Hirschmanniella oryzae (van Breda de Haan) was observed towards roots of cabbage (Brassica oleracea L. var capitataj; carrot (Daucus carota L. var safivus); radish (Raphanus sativus L.) and onion (Allium sepa L.) singly and in combinations of two plants. The effects of different root homogenate concentrations of the above mentioned plants on the activity of H. oryzae have also been observed.
Materials and methods
H O S T SELECTION BEHAVIOUR
The responses of adult H. oryzae were observed towards roots of cabbage, carrot, radish and onion singly and towards different combinations of two hosts. All experiments were carried out in 0.5 cm thick layer of 0.5 "o water-agar in the presence of 1 cm long excised
Revue Nematol, 8 (3) : 265-272 (1985) 265

55 A I Bilgranu, I Ahmad &. M S Jaimjpun
roots of 4-6 day old seedlings (Bilgrami, Ahmad & Jaira)pun, 1985) The roots before being excised were surface sterilized with 0.1 . mercuric chloride and the nematodes with 0 01 mercuric chloride solution All experiments were carried out at 25 ± I" and replicated five times unless mentioned otherwise. Controls were run without roots.
AriK.\cno\ IOW^RDS s i \ ( ,n HOSI ROOI
Two experiments were performed to test host preference behaviour of / / oryzae In the first 7 cm diameter Petn-dishes were marked at the bottom into seven equal zones of 1 cm width These zones were numbered as 1, 2 and 7 (Fig. 1). 'Ihe roots were placed in zone 1 of the Petn-dishes and left fe)r 12 h after which twenty-five
^/^""^sT^ /
/ ' •
\ \
/
/ 1 •
\ \
• Point of inocu of nematoc
Sec
^ \ \ 1
\ \
/ y Lation es
tors
~1
2
3 i
5 6
7
2 3 /,
5 6
7
A
B
Fig 1 Diagram showing different zones of the Petn-dish which was employed in the expenment to study the attraction of H oryzae towards single host and combination of two hosts
H oryzae adults were introduced in zone 4. The number of nematodes present m each zone was counted after 4 h In the second attraction was tested by the Petn-dish method (Ahmad & Jairajpun, 1980; Bilgrami, Ahmad & Jaira)puri, 1985) A 1 cm part of excised root from a 4-6 day old seedlings was placed in the inner circle of Petri-dishes containing a 0.5 cm thick layer of 0.5 "o watcr-agar After 12 h five specimens of adult H oryzae were placed at different points of the reference circle. The distribution of H oryzae was recorded after 1, 2, 3 and 4 h and was converted into scores and log scores. Experiments were replicated fifteen times.
A n R . \ C I I O N lOWARDS DlhFhRtM HOST Rt)01 COMBI WIIONS
As above, attraction towards different root combinations was studied m Petri-dishes marked at the bottom into seven zones (Fig 1). An excised root of one host was placed in zone 1 and that of the other in zone 7 and left for 12 h before fifty adult nematodes were introduced in zone 4 The distribution of nematodes was recorded after 4 h The following combinations of host roots were used • cabbage and carrot, cabbage and radish; cabbage and onion, carrot and radish; carrot and onion and onion and radish.
Ac IIVTIT I \ ROOr HOMOGEN'^rE CONCENTRAIIONS
Twenty gram of roots of 4-6 day old seedlings were surface sterilized with 0 1 °o mercunc chloride and homogenized in 20 ml of sterile distilled water in a Warring blender to obtain the stock (100 "u) root homo-genate soluuons. This solunon was then filtered to remove the sohd materials. From the stock solution 20, 40, 60 and 80 '' o dilutions were obtained. Stock solutions of homogenates of cabbage, carrot, radish and onion roots were prepared and tested separately on the activity of H oryzae adults. Two ml solution from each concentration of various root homogenate taken in separate cavityblocks and five adult H oryzae were placed m each. The activity of nematodes was measured in terms of head movements per minute (Bilgrami, Ahmad & Jairajpun, 1983). The observations were made at 2, 4, 6, 8, 10 and 12 h after transferring nematodes into medium Activity in tapwater served as control. Data of those solutions in which the activity ceased within 2 h have not been plotted.
Results
ATTRACTION TOWARDS SINGLE HOST ROOT (Fig. 2,3 & 4)
Most of the nematodes remained near point of introduction (zone 4) in the absence of roots (Fig. 2, A) Onion and radish seedlings attracted fewer nematodes (Fig. 2, B & C) than carrot and cabbage (p < 0 05) (Fig 2, D & E)
In the second experiment the distribution of adult H oryzae corresponded to ranks 4, 5 and 6 in absence of roots (Fig. 3). However, in the presence of onion, radish, carrot and cabbage roots the distribution of nematodes corresponded to ranks 5, 6, 7, 9 and 10; 6, 7, 8 and 9; 10, 11, 12, 13, 14, 15, 16 and 17 and, 18, 20 and 21 respectively (Fig. 3, A, B, C & D). The nematodes showed weak responses towards onion and radish seedlings (mean log score 1.56 and 1.97 respectively, p < 0.05) while they responded strongly towards cabbage seedhngs (mean log score 2.58, p < 0.05; Fig. 4).
266 Revue NematoL, 8 (3) 265-272 '1985J

Responses o/Hirschmanniella oryzae towards plant roots
ii ~o o m £ c 'q o
« 1
^4
•a o ra E c
o 6 z
16-1
12,
i_
1 2 3 ^ 5 6 7 Zones
B
1 2 3 ^ 5 6 7 • lb
1 2 3 ^ 5 6 7
in ii •U o ra
e u c
o
16-1
12-.
T 1 2 3 ^ 5 6 7
Zones I L.
15-
12.
H] 1 2 3 -i 5 6 7
Zones
1 2 3 ^ 5 6 7
Fig 2 Attraction of adul t / / yrvzae towards different plant roots A without roots, B onion, C radish, D carrot, E cabbage
A n R . \ ( I I O \ lOW^RDS niFH-RPM HOST ROOT tOMBI \A I IONS (Fig 5)
In Lombinations of cabbage and onion and cabbage and radish, H oryzae was attracted more towards cabbage than other plant roots (p < 0 05) (Fig 5, C & D) but in the combination of cabbage and carrot there was no significant difference m the attraction (p > 0 05j (Fig 5, B) The roots of carrot when placed in the combination of radish (Fig 5, E) and onion (Fig 5, F) attracted significantly more H oryzae than the latter two plants (p < 0 05) The differences in the attraction of nematodes m combmation of radish and onion were insignificant (p > 0 05) (Fig 5, G) In the absence of roots most of the nematodes remained near point of introduction (Fig 5, A)
ACTIVITY I \ ROOT HOMOCIENAIF CONCEN'TRATIONS (Fig 6)
All concentrations of cabbage root homogenate appeared favourable for H oryzae The homogenate of 60 and 80 ''o stimulated activity most and the increase in activity from 2 to 12 h was significant (p < 0 05) (Fig 6, A) 20, 40 and 60 "o concentrations of carrot appeared to be more suitable for nematodes (p < 0.05) The 80 and 100 "o concentrations seemed unfavourable (Fig 6, B) All concentrations of radish and onion suppressed the activity of nematodes (Fig 6, C & D). No activity was observed in 100" o solution of radish and onion root homogenate The nematodes became immobile after 6 h m 80 " 1 solution of radish and after 4 and 10 h in 80 and 60 " 0 solutions of onion root homogenate
Revue Nematol, 8 (3l 265-272 f 19851 267

B7 A. L. Bilgramt, I. Ahmad & M. S. Jairajpun
>>
2.
With roots
\l
I I I I I I I I I I I I I I I I I I I I I 5 10 15 20
6n
u '•-I c (U D
LL
B
1 I I I I I I I I I I I I I I I I I I I I 1
5 0 15 20
o L c
^2.
I I I I I I I I I I I I I I I I I I I I I 5 10 15 20
o c
81 i
6
D
I I I I I I I I I I I I I I I I M I I 5 10 15 20
Ranks
L.
Control
I I I I I I I I I I I I I I I I I I I I I 5 10 15 20
L.
2.
A
I I I I I I I I I I I I I I I I I I I I 1 5 10 15 20
7
4
2J
n
I I I I I I I I I I I I I I I I I r I I I 5 10 15 20
6.
2.
I I I I I I I I I I I I I I I I I I I 5 10 15 20
Ranks
Fig. 3. Distribution of adult H. oryzaein presence and absence of different plant roots, A : onion; B : radish; C : carrot; D : cabbage.
268 Revue NematoL, 8 (3): 265-272 a 985,

Responses o/Hirschmanniella oryzae towards plant wots
z:/tr 2.5CL
a, 2.00, L. o o (/) a,1-50J o
<u Z
0.50-
/ >
^ C o n tro L
1 I Ca. Ct. Rd. Roots
On.
Fig. 4. Attraction of adult H. oryzae towards different plant roots (Petri-dish expenments). Ca '. cabbage; Ct : carrot, Rd : radish; On : onion.
Discuss ion
Prot (1980) considered the attraction of plant-parasitic neinatodes towards host roots to be non-specific. Khera and Zuckerman (1963) found no significant difference in the attraction of Hemicycliophom simibs towards roots of host and non-host plants. However, juveniles of Meloidogyne hapla were attracted equally by the resistant and susceptible cuitivars of alfalfa when tested separately, but when compared simultaneously susceptible cuitivars attracted more nematodes (Griffin, 1969). Our observations showed that all plant roots were attractive to H. oryzae but these nematodes showed some degree of preference for cabbage and carrot as indicated by their distribution pattern. Cabbage was preferred most when tested separately or in different combinations of plant roots. These differences in the attraction of H. oryzae towards different plant roots may be attributed to the sensitivity of the nematodes to stimuli of the roots or due to the attractants themselves as all plants do not produce similar attractants (Prot, 1980).
The differential activeness of H. oryzae in different concentrations of various root homogenates may be attributed to the effects produced on the nematodes by the attractants and possibly by other substances present in the host roots. Luc, Lespinat and Souchaud (1969) and Prot (1975) believed that root diffusates simultaneously stimulate the rate of movement and attract the nematodes. Perhaps the same phenomenon seems to
occur in H. oryzae adults. The maximum attraction towards cabbage and least towards onion roots as well as enhanced activity in different concentrations of cabbage and reduced activity in onion root homogenate suggests that perhaps attraction may be directly related to the activity of nematodes, the more active being attracted more.
Ac KNOWLEDCFMEN 1 S
We thank Prof. Nawab H. Khan, Chairman, Department of Zoology for providing laboratory facilities and Prof. Ather H. Siddiqi for suggestions. The first author is grateful to U.G.C., New Delhi for the financial assistance.
RfchHRENCLS
AH.MAD, I. & JMFLMPLRI, M . S. (1980). Sex attraction and copulation m Chiloplacus symmetncus (Nematoda : Acro-belinae). Nematologica, 26 : 139-148.
AzMi, M. I. & JAIRAJPLRI, M . S. (1977). Attraction of plant-parasitic nematodes to host roots. Nematologica, 23 : 119-121.
B1LGRA.MI, A. L. (1983). Attraction of Hirschmanniella oryzae towards cabbage seedlings. Proc. 3rdNematol Symp., Solan, India : 15 (Abstr.).
BiLGRAMi, A. L., AHMAD, 1 & J.' IR.'VJPLRI, M . S. (1983). Some factors influencing predation by Mononchus aquaticus. Revue Nematol, 6 : 325-326.
BiLGRAMi, A. L., AH,MAD, I & Jj,iRAjPLRi, M. S. (1985). Factors influencing attraction of adult Hirschmanniella oryzae towards cabbage seedlings. Revue Nematol, 8 : 67-75.
GRIFFI\, G. D . (1969). Attractiveness of resistant and susceptible alfalfa to stem and root-knot nematodes. J. Nematol, 1 : 9 (Abstr.).
KHERA, S. & ZOCKERVIAN, B. M . (1963). In vitro studies of host-parasite relationships of some plant-parasitic nematodes. Nematologica, 9 : 1-6.
LEE, Y. B. & EVANS, A. A. F. (1973). Correlation between attraction and susceptibilities of nce varieties to Aphelen-choides besseyi Christie, 1942, Kor. J. PI Protect., 12 : 147-151.
LovcNSBERY, B. F. & Vu GLiERCHio, D. R. (1960). Mechanism of accumulation of Meloidogyne incognita acnta around tomato seedlings. Phytopathology, 50 : 178-179.
LovcNSBERY, B. F. & ViGLiERrHio, D. R. (1961). Importance of response of Meloidogyne hapla to an agent from germinating tomato seeds. Phytopathology, 51 : 219-221.
LLC, M. , LESPINAT, P. & SOLTHAID, B. (1969). Marquage direct de Hemicychophora paradoxa par le phosphore ra-dioactif. Utilisation pour I'etude des deplacements des nematodes phjtoparasites dans le sol. Nematologica, 15 : 35-43.
M M H L R , V. K. & PR.' S.AD, S. K. (1973). Survival and host range of the nce root nematode Hirschmanniella oryzae. Indian J Nematol, 3 : 88-93.
Revue Nematol, 8 (3j: 265-272 (1985) 269

59 A L Bilgramt, I Ahmad & M. S Jairajpun
30'
25 J V)
•S 20. O
° 15
^ 10.
1 2 3 A 5 6 7 Zones
en
- o o
-•-•
ra E c
o z
in 01
•a o
E (i> c
o z
in
• D O
•4-*
m £ c
o Z
20-jCa
15
10.
5
B ct
20'
15.
10-
5.
1 2 3 /. 5 6 7
Ca D
Rd
1 2 3 /, 5 6 7
25
20
15
10
5,
-:t F
i
1
On
1 2 3 / 1 5 6 7
Zones
20-
15.
10.
5.
Ca. !
II
c
On
20-1,
15
10
5J
1 2 3 / 1 5 6 7
CL E
Rd
1 2 3 4 5 6 7
25-
20.
15.
10.
5.
Rd
—
^ G
1
On
1 2 3 4 5 6 7
Zones
Fig. 5. Attraction of adult H. oryzae towards plant roots in combinations. A ; without roots, Ca : cabbage; Ct : carrot; Rd : radish; On : onion 'Arrotvs indicate point of introduction of nematodes/.
270 Revue NematoL, 8 (3j: 265-272 (1985j

Responses oj Hirschmanniella oryzae towards plant roots
•"•inne ( h )
Fig. 6. Activity of adult H oryzae in different concentrations of plant root homogenates. A : cabbage, B : carrot; C ; radish; D onion.
MoHANDAi, C , PAI-IAMK, N . K C. & PR.AS\D, S K (1979). Host range of the rice root nematode Hirschmanmella oryzae Indian J Xernatol, 9 . 177-178
PARK, J S., H\\, S. C. & Li-f-, Y B. (1970). Studies on the ecology and feeding preference of Hirschmanmella oryzae Res Rep Office rural Develop, Korea PI Environ, 13 • 93-98.
PROI, J. C. (1975). Research on movement of Meloidogyne spp., juveniles towards roots. Cah ORSTOM, ser Biol, 10 251-262
PROI, J. C. (1980). Migration of plant-parasitic nematodes towards plant roots. Revue Nematol, 3 : 305-318.
PROI, J. C. & VAN GLNDY, S. D (1981a). Effect of soil texture and the clay components on migration of Meloidogyne incognita second stage juveniles J. Nematol, 13 : 213-217.
PROF, J. C. & Vw GLNDY, S. D . (1981b). Influence of photopenod and temperature on migrations of Meloidogyne juveniles. J Nematol, 13 : 217-220.
VioLiERCHio, D. R. (1961). Attraction of parasitic nematodes by plant root emanations. Phytopathology, 51 : 136-142.
Accepte pour publication le 29 avril 1985
Revue Nematol, 8 '3j: 265-272 1985) 271

Nematologica 30 (\984) 457-462, E J Brill, Leiden U J
PREDATORY BEHAVIOUR OF AQUATIDES THORNEI (NYGOLAIMINA: NEMATODA)
BY
ANWAR L BILGRAMI, IRFAN AHMAD and M SHAMIM JAIRAJPURI
Section of Nematology, Department of Zoology, Aligarh Muslim University, Aligarh-202001, India
Studies on its predatory behaviour revealed that Aquatides thornei was not attracted towards live nor bisected prey nematodes, but made contact with the prey by chance labial contact was necessary to initiate an attack Several predators aggregated and fed on prey injured by another predator No preference was shown for any part of the body for attack, Rhabditis sp was selected in preference to Chiloplacus symmetncus A thornei seemed to paralyse the prey alter its mural tooth was inserted deep into the body Ingestion was intermittent with short periods of sucking activity, the duration of feeding on a single prey varied from 3-5 mm to 4-6 h Little variation was found in the daily rate of predation during 12 days The rate of predation increased as the number of prey increased Maximum predation occurred with 150 prey and least with 25 prey nematodes
Keywords Behaviour, predator, prey, biological control, feeding
Mononchs and dorylaim nematodes are known to be predacious and Thorne (1930, 1939) had suggested that nygolaims including Sectonema were predatory in habit Since then several species of Dorylaimus, Discolaimus and Actinolaimus have been described as predatory (Linford & Oliveira, 1937). Esser (1963) had also observed that species of Dorylaimus, Labronema, Discolaimus and Car-charolaimus were predatory. The dorylaim predators are omnivorous and derive their lood from other sources also such as algae and fungi (Hollis, 1957; Ferris, 1968; Wood, 1973) Wyss & Grootaert (1977) studied the feeding mechanism of Labronema vulvapapillatum Loof & Grootaert, 1981, a predatory dorylaim, while the ultrastructure and function of its anterior feeding apparatus was studied by Grootaert & Wyss (1978). Small & Grootaert (1983) observed the predatory abilities of nematodes belongmg to different orders on a range of prey species and found Panagrellus redivivus (Linn, 1767) Goodey, 1945 to be most susceptible to attack of the free-living or plant-parasitic species used as prey.
The following observations were made on a nygolaim nematode, Aquatides thornei (Schneider, 1937) Ahmad & Jairajpuri, 1979 to study its predatory behaviour
MATERIALS AND METHODS
Soil samples containing Aquatides thornei were collected from paddy fields near Jamalpur, Aligarh. These samples were processed by sieving and decan-

6 4 5 8 BIICRAMI AHMAD & JMRAJPLRI
tation and a modified Baermann funnel method The nematodes were rmsed repeatedly m distilled water and kept m tapwater before use Only male and female predators and adult prey were selected for study All experiments were earned out at 25 + 1°C m an incubator
The feeding of A thornei at 66 x magnification was observed by inverting the Petri-dishes containing A thornei and prey nematodes in 1 % water-agar under the binocular microscope To study the feeding mechanism at 450x oi higher magnifications simple observation chambers similar to those used by Wyss & Grootaert (1977) were made These consist of glass slides with a small cjuantity of 1 % water-agar The agar when cool, was cut into 2 x 1 5 x 0 5 cm blocks Ten specimens of A thornei, both males and females, were placed in agar blocks together with 100-150 specimens of Rhabditis sp and Chiloplacus syrnmetncus (Thorne, 1925) Thorne, 1937 which served as prey To facilitate entry of nematodes, the agar surface was incised with a needle The agar block containing predators and prey was gently covered by a cover-slip and kept at 25 + 1°C for 15 mm before starting the observations
To determine whether A thornei were attracted towards prey, petri-dishes 5 5 cm m diameter were divided into three zones viz , inner, middle and outer by drawing two concentric circles 0 5 and 2 5 cm m diameter A plastic straw 5 mm high, 5 mm in diameter, with a small piece of filter paper glued at one end was placed vertically in the inner zone of the petn-dish, with the filter paper m contact with the petri-dish The petri-dish and straw was then filled with a 0 5 cm thick layer of 1 % water-agar The response of yl thornei was tested towards live and bisected (cut into two pieces) prey To observe its attraction towards live prey, 25 Rhabditis sp were placed in the straw pipe and left for 4 h Five predators were then released on the periphery of the middle zone The distribution of^ thornei was recorded after 1, 2, 3 and 4 h Scores were obtained by summing up the products of the number of worms in each zone with their corresponding weighting factors The weighting factors were obtained by dividing the area of outer zone by that of each of the three zones These scores were then converted into log scores and plotted against time (Ahmad & Jaira-jpuri, 1980) The second experiment was identical to the above except that the prey were bisected Each experiment was replicated fifteen times
To determine the pattern of feeding during a period of 12 days five A thornei were placed with 25 specimens of C syrnmetncus in cavity-blocks containing a 0 5 cm thick layer of 1 % water-agar Observations were made every 24 h and the predators then transferred to fresh cavity-blocks with 25 fresh prey A similar experiment was carried out with A thornei and Rhabditis sp Each experiment was replicated five times
To observe the effect of prey numbers on the rate of feeding, 25, 50, 75, 100, 125 or 150 specimens of C syrnmetncus were placed in separate cavity-blocks containing a 0 5 cm thick layer of 1 % water-agar Five A thornei were placed m each cavity-block The same experiment was earned out with Rhabditis sp and

AQl'ATIDES THOKXEI BEHAVIOUR 4 5 9
A. thornei. Observations were made after 24 h. Each experiment was replicated five times.
RESULTS
A. thornei encountered prey purely by chance. To attack a prey contact with the lips was necessary; the predator did not start an attack after contact with any other part of the body. When contact was established, the predator moved its lip region over the body surface of the prey and made a few thrusts with its mural tooth. Penetration of the cuticle occurred quickly with 8-10 thrusts of the tooth. Once the tooth penetrated deep into the body of prey the latter became immobile and only a few weak oesophageal contractions in the predator were observed. To ingest the body contents the predator inserted its tooth deep into the body of the prey and then commenced side-ways movements and thrusting of the tooth. The lip region of the predator remained attached to the body of the prey throughout the entire period of feeding. Ingestion appeared intermittent, with short periods of sucking activity lasting a few seconds. All through the ingestion period the mural tooth remained inserted into the prey. Feeding usually ended when the entire body contents had been ingested but the cuticle of the prey remained intact. Frequently, 4-5 A. thornei were seen feeding on a single nematode, the duration of feeding on which varied from 3-5 min to 4-6 h depending on size and type of the prey and the number of predators. Rhahditis sp. were killed faster than C. symmetricus.
Female A. thornei laid eggs in the agar and showed affinity to its own eggs: males behaved identically. When a randomly moving predator (male or female) came in contact with its own eggs it rubbed its lips around the eggs a few times without thrusting its mural tooth and then moved away without any attempt to feed. By contrast with this, the predator did not respond to contact with the eggs of prey nematodes.
A. thornei was not attracted towards live or bisected prey. In each case the distribution after each hour was only slightly different from control (p>0.05) . Distribution towards live prey after 1 h (mean log score 1.46) was not different from that after 4 h (mean log score 1.49, p>0 .05) (Fig. 1, A). Similarly, towards bisected prey it also did not differ significantly after 1 h (mean log score 1.53) and 4 h (mean log score 1.54, p>0 .05) (Fig. 1, B).
The rate of feeding of 4. thornei on C. symmetricus and Rhabditis sp. varied little during the period of observation lasting 12 days. The number of prey killed differed slightly from day to day but the differences were insignificant (p>0.05) . The predators, however, seemed to prefer Rhabditis sp. and preyed more on them than on C. symmetricus (p<0.05) .
The number of prey influenced the rate of predation by A. thornei. The feeding rate increased with increase in prey number (Fig. 1, D). The number of individuals killed in the population of 150 prey specimens of C. symmetricus or
L/ J

64 460 BILGRAMI, AHMAD & JAIRAJPURI
1.55
fe 1-50. u
3 US. c ra
I 1. 0 J
1.35
o_oWi th prey A
.— •Cont ro l
1 2
Time
• D
*• 2
/ \ / \ / \ ^ D O o — O O—o
° - ° R h a b d i t i s sp. • — • C.svmmetricus
• —•. • • — • — • a / v v \ / ~\—1—I—I—I—I—I—I—I—I—I—I
1 2 3 4 5 6 7 8 9 10 11 12
Days
1.601
1.55.
° 1-50.
o 1.45,
c n)
^ 1.40.
1.35
o—oWith prey B .— •Cont ro l
3
Time (h )
14-1
^ 10.
^ 6. o d
2_
0—0 Rhabdit is sp.
•— • C -symmetricus
. / /
— I — 25 50
—r— 75
No. of prey
D.
100 125 150
Fig 1 A, Attraction of ^ i/!orn« towards intact prey B, Attraction of ^ //iorria towards bisected (cut into half) prey C, Predatory pattern of A thornei o\er a period of 12 days D, Effect of prey
numbers on the rate of predation by A thornei
Rhabditis sp. was significantly more than when 25 specimens of the same prey were used (p<0.05) . A thornei agam preyed more on Rhabditis sp. than on C. symmetricus
DISCUSSION
These observations show that A thornei can be predacious in nature. The rate of predation by Diplenteron potohikus Yeates, 1969 and Pnonchulus punctatus (Dujardin, 1845) Wu & Hoeppli, 1929 increased with increase in the number of prey (Yeates, 1969; Nelmes, 1974). A thornei also behaved similarly when placed with different populations of prey. The higher rate of predation with in-

A(ILA I IDES rHUR\EI BEHAMOLR 4 6 1
creased prey number may be due to improved chances of contact betv/een predators and prey However, Mononchus aquaticus Coetzee, 1968 did not show any significant difference in predation when placed with different numbei s of prey nematodes (Bilgrami et al , 1984) This disparity between different predators reflects their predatory capabilities
A thornei were not attracted towards potential prey either live or bisected Wyss & Grootaert (1977) found the same for Labrontma vulvapapillatum A live prey cut m half attracted several dorylaim predators from a short distance (Esser, 1963), a reaction not found in this study However, the aggregation of several predators about an injured prey suggests some kind of attraction This may perhaps be due to oesophageal secretions of the predators themselves released during feeding instead of chemical attractants released by the prey (Esser, 1963) However, if the predators were attracted to any attractants from the prey (Esser, 1963) then their effect may be excercised over too short a distance to be discerned by our experimental technique The affinity of A thornei for their eggs in the presence of eggs of free-living nematodes suggests that these nematodes are capable of recognizing their own chemical attractants (Esser, 1963)
The feeding behaviour of ^ thornei does not differ much from that reported by Wyss & Grootaert (1977) for L vulvapapillatum except for the duration of feeding on single prey, where A thornei fed longer than L vulvapapillatum and killed more prey per day These differences could be due to differences in ihe nematode species used as prey Seinura spp of Aphelenchoidea paralysed their prey by injecting salivary or oesophageal secretions (Linford & Ohveira, 1937, Hechler, 1963) while dorylaim predators did not paralyse or pre-digest their prey (Hollis, 1957, Wyss & Grootaert, 1977) A thornei, which is a nygolaim predator probably paralysed the prey by its mural tooth
Small & Grootaert (1983) noted that several predators preferred some prey to others and found Panagrellus redivivus more susceptible to attack than other free-livmg or plant-parasitic nematodes used as prey A thornei also exhibited preference for some prey as they killed significantly more Rhabditis sp than C symmetncus This differential predation may be due to differences in behaviour or size of prey, or even differences in texture of the prey cuticle
We thank Professor Nawab H Khan, Chairman, Department of Zoology for providing laboratory facilities and Professor Ather H Siddiqi for his suggestions The first author is thankful to U G C , New Delhi for financial assistance
ZL S AMMENFASSL NG Das rauberuche Verhalten von Aquatides thornei (Nygolaimina Nematoda)
Untersuchungen uber das rauberiscfie Verhalten ergaben, dafi Aquatides thornei weder von le benden noch von durchschnittenen Beutenematoden angelockt wurde Der Beutekontakt erfolg-te zufalhg, und zur Auslosung eines Angriffes war Lippenkontakt notwendig Mehrere Raubcr
65

6 4 6 2 BILGRAMI AHMAD & JAIRAJPLRI
sammelten sith und frafien an der von einem anderen Raubcr verietzten Beute Btim Angnff konnte keine Bcvorzugun^ irgendeines Korptrabschnittes festgestellt werden Bei gleichzeitiger Anwes>eheit von Rhabdiln sp und Chiloplacus symmetncus wurde Rhabditu bevorzugt angefallen A Ihurnei schien seme Beute zu lahmen, nachdem der Zahn tief m den Korper emgedrungen war Die Nahrungsaufnahme wurde von kurzen Saugpenoden unterbrochen An einem Beutenema-toden blieb em Rauber von 3-5 Mmutcn bis zu 4-6 Stunden Uber einen Beobachtungszeitraum von 12 l a g e n blieb der tagliche Beuteverzehr ziemlich gleich Die An/ahl der Beutefange nahrn mit hohercr Beutedichte zu, wobei die meisten fange bei einer Dichte von 150 und die wenigsten bei 25 Beutenematoden auftraten A thornei war in dei I age, eigent Eier von Eiern der Beute nematoden zu unterscheiden
REFERENCES
AHMAD, I & JAIRAJPLRI, M S (1980) Sex attraction and copulation in Chiloplacus symmetncus (Nematoda Acrobelinae) Nematologtca 26, 139-148
BIIGRAMI, A L , AHMAD, I & JAIRAJPLRI, M S (1984) Some observations on the predatory
hehaMioya oi Mononchus aquaticus Nematologia Meditenanea 12, 41 45 issER, R P (1963) Nematode interactions in plates of non-stenle water agar Proceedings of the
Soil & Crop Science Society of Florida 23, 121-138 FERRIS, V R (1968) Biometric analysis in the genus Laftronsma (Nematoda Dorylaimida) with
a description of L thornei n sp hlematologica 14, 276-284 GROOTAERT, P & WYSS, U (1978) Ultrastructure and function of the anterior feeding apparatus
in Labronema sp Nematologtca 24, 243-250 HozLis,J P (1957) CnXtura^ iinAvts, \N\l\i Dorylaimui eltersbergensii Phytopathology 47, 4:6Q-i73 LiNFORD, M B & OLIVEIRA, J M (1937) The feeding of hollow spear nematodes on other
nematodes Science 85, 295-297 NEIMES, A J (1974) Evaluation of the feedmg behaviour of Pnonchulus punctatus (Cobb), a
nematode predator Journal of Animal Ecology 43, 553-565 SM-VLL, R W & GROOTAERT, P (1983) Observations on the predation abilities of some soil
dwelling predatory nematodes Nematologtca 29, 109-118 THORNE, G (1930) Predacious nemas of the genus Nygolaimus and new genus Sectonema Journal
of Agricultural Research 41 445-446 THORNE, G (1939) A monograph of the nematodes of the superfamily Dorylaimoidea Capita
Zoologica 8, 1-261 WOOD, F H (1973) Biologv of Aporcelaimellus sp (Nematoda Aporcelaimidae) Nematologtca
19, 528-537 WYSS, U & GROOTAERT, P (1977) Feeding mechanism of Labronema sp Mededelmgen van de
Fakulteit Landbouwwetenschappen Rijksuniversiteit Gent 42, 1521-1527 YEAFES, G W (1969) Predation by Mononchoides potohikus (Nematoda Diplogastendae) m
laboratory culture Nematologica 15, 1-9
Accepted for publication January 7, 1985

4i-(2l ^'^s '^^a^fiA^
Indian J. Nematol. 15 (2), (1985) : 209-216
ATTRACTION A N D FEEDING BEHAVIOUR O F ADULT HIRSCHMANNIELLA ORYZAE
ANWAR L. BILGRAMI, IRFAN AHMAD and M. SHAMIM JATRAJPURI Section of Nematology, Department of Zoology, Aligarh Muslim University,
Aligarh-202001, India
Abstract: Feeding behaviour of adult/f('«cAwa««(V//a oryzae comprised of four phases viz., exploration of feeding site, stylet penetration, ingestion and departure from the feeding site. Ingestion of cell contents was intermittent and lasted to 20-110 min but often stretched to 3-4 h. Salivation was not observed. H. oryzae was attracted towards cabbage seedlings and its homogenates. The attraction was greatest in higher concentrations (80, 100 per cent) while least in the lower concentrations (20, 40 per cent) of cabbage root homogenate.
Key words : H. oryzae, behaviour; feeding, attraction, cabbage seedlings.
Several studies have been conducted on the feeding and attraction of plant-parasitic nematodes (Khera & Zuckerman, 1963; Azmi & Jairajpuri, 1977; Prot, 1980; Wyss, 1981; Prot & Van Gundy, 1981a; 1981b; Prot &Kornprobst, 1983). Feeding of Tylenchorhytichus daytoni was studied by Krusberg (1959). Wyss (1971; 1973) described different phases in the feeding of Trichodorus similis and Tylen-chorhynchus dubius respectively. While observing the exploration and selection of feeding sites, Doncaster & Seymour (1973) described probable succession of stimuli and the responses leading a nematode to feed on its host. Passive ingestion during feeding was reported by Doncaster & Seymour (1974) in Hexatylus viviparus. Barbatola & Bridge (1980) observed feeding behaviour of Hirschmanniella oryzae, H. spinicaudata and H. immamuri adults and juveniles on rice while Bilgrami (1983) found cabbage seedlings attractive to the adults and juveniles of H. oryzae. Bilgrami et al. (1985) studied several factors which influenced the attraction of H. oryzae towards cabbage seedlings.
In the present work, the mechanism of feeding and attraction of rice root nematode, Hirschmanniella oryzae (van Breda de Haan, 1902) Luc & Goodey, 1963 on
cabbage {Brassica oleracea L. var capitata seedlings is described.
MATERIALS AND METHODS
Adult H. oryzae were extracted from soil from paddy fields. Cabbage roots, grown in 0.5 per cent water agar were surface sterilized with 0.1 per cent and the nematodes with 0.01 per cent mercuric chloride prior to the experiments. Water agar was prepared by autoclaving at 15 lb pressure for 30 min. Experiment was carried out at 25 ± r c .
Feeding mechanism : Feeding of H. oryzae was observed in chambers, similar to those used by Wyss & Grootaert (1977) which consisted of glass slides with a 0.5 cm thick layer of 0.5 per cent water agar. When cool, the agar was cut into blocks of 2 X 1.5 cm. A 3 cm long excised root of a 4-6 day-old cabbage seedlings was placed in the middle of agar and 25 adult H. oryzae were placed along the edges of the agar. A coverslip was gently placed over the agar and it was sealed with vaseline to prevent drying.
Attraction behaviour of adult H. oryzae : It was tested towards cabbage seedlings by the following methods:

68
210 BILGRAMI, AHMAD AND JAI RAJPURI
Petridish experiments : Petridishes 5.5 cm in diameter, weie marked at bottom into three zones by drawing two concentric circles of 0.5 and 2.5 cm diameter. The latter was termed as reference circle. One cm excised root, from a 4-6 day-old cabbage seedling, was placed in the inner circle of petridishes containing 0.5 cm thick layer of 0.5 per cent water agar. After 12 h five specimens of adult H. oryzae were placed at different points on the reference circle. The distribution of nematodes was recorded afler J, 2, 3 and 4 h and was converted into scores and log scores (Ahmad & Jairajpuri, 1980). Experiments were replicated fifteen times. Controls were run without host roots.
Polythene tube experiments : Polythene tubes, similar to those used by Bilgrami et al. (1984), were designed and filled with 0.5 per cent water agar. One cm excised root from 4-6 day-old cabbage seedling was placed in the smaller tube. After 12 h, 25 adult H. oryzae were introduced in the larger tube through the central hole. The distribution of nematodes was recorded after 1,2,3 and 4 h by cutting the tubes into 1 2 cm blocks. Experiments were replicated five times and tubes without roots serveed as control.
Attraction towards cabbage root homo-genates : Twenty g roots of 4-6 day-old cabbage seedlings were surface steri'ized and homogenized in 20 ml sterile distilled water in a warring blender to obtain stock (100 per cent) root homogenate solution. This was then filtered to remove the solid materials. From the stock solution, 20, 40, 60 and 80 per cent dilutions were made. To observe the attraction of nematodes, 7 cm diameter petridishes, marked at bottom into 7 equal zones (Azmi & Jairajpuri. 1977), filled with 0.5 cm thick layer of 0.5 per cent water agar were taken. 0.5 ml of homogenate solution was placed in a cavity made in zone 1 of the petridishes and after 12 h, 25 adult H oryzae were introduced in zone 4. Nematodes were tested against
each concentration separately. The number of nematodes present in each zone was recorded after 4 h. Petridishes without root homogenate solutions servad as control. Each experiment was replicated five times and fresh stock solution was prepared for each experiment.
RESULTS
Feeding mechanism : Nematodes exhibited random movement upon introduction into the agar and few oF them became quiescent. After about 3-4 h, almost all had started feeding or were in the vicinity of the toots. The following phases were observed :
Widespread exploration : Nematodes moved a'ong the root and made several contacts with the roots by their lips both at right angles and with the sides of the head. A right angle contact of the lips with the root resulted in a few quick stylet thrusts (10-15) otherwise occasional probings with (he stylet was observed during this phase which lasted about 24 (15-29) min.
Local exploration: Regular stylet thrusts were made by nematodes in order to perforate the cell wall. These thrusts often resulted in the lips to lift away from the root surface, reverting nematodes to widespread exploration. However, cell wall was pushed in with each stylet thrust when nematode had located a favourable feeding site and the contact between the lips and cell surface was never broken.
Stylet penetration : While penetrating, the nematode remained immobile. At the onset, there were 80-94 stylet thrusts per minute which increased later to 170-198/rain and about l/4th of the stylet was protruded Thereafter, stylet thrusts declined to 118-133/min. becoming more deeper (about 1/2 of the stylet was penetrated). Cell wall perforation was achieved in about 15 (10-18) min. Nematodes

ATTRACTION OF H. ORYZAE 211
reverted to widespread exploration they failed to perforate the cell wall.
if
Ingestion : The ingestion was intermittent (short pauses of sucking aclivity). During ingestion (20-110 min, some times stretched to 3-4 h), the nematodes did not remain completely motionless, bending of tail was quite frequent. The lumen of metacorporeal pump opened with the contraction of the muscles of the median bulb. The rate of pulsation of the pump ranged from 55-68/min. After Ingestion, some nematodes reverted to widespread exploration and fed again, while others became quiescent. No salivary secretions were observed.
Retraction of the stylet and departure from feeding site: After ingestion the pump stopped pulsating, stylet was retracted and the lips detached from the cell surface. While moving away from the roots the nematodes made 8-10 stylet thrusts in an exploratory fashion. After withdrawl, the nematodes moved back and forth, a few times, and left the feeding site.
Attraction behaviour of adult H. oryzae
Petridish experiments (Figs. 1 & 2) : Maximum attraction (mean log score 2.6) was observed after 4 h and least (mean log score 1.7) after 2 h (Fig. 1). No attraction was evident after 1 h (mean log score 1.2, p >0.05).
In the presence of cabbage seedlings the distribution of K o/-j'zaecorresponded to ranks 4, 5 and 6 after 1 h; 4, 5, 7, 8 and 9 after 2 h; 13, 14, 16, 17 and 18 after 3 h; and 18, 20 and 21 after 4 h (Fig. 2). In absence of roots, the distribution corresponded to ranks 4, 5 and 6 after each successive hour (Fig. 2).
Polythene tube experiment: (Fig. 3): The majority of nematodes remained near the point of introduction in the absence of roots. In the presence of roots, attrac-
2.65
2.5a
2.20.
1.90.
1.60.
1.30.
1.00
•—e Control
0—0 With root
Fig.
1 2 3 i Time (h)
Attraction of H. oryzae towards cabbage seedlings.
t:on was evident after 1 h. (p < 0.05, Fig. 3, A). More H. oryzae reached + 5 , +4 , 4-3 and + 2 regions after 2 h (Fig. 3, B) and about half the number was found in + 5 region after 3 h (p < 0.05, Fig. 3, C). The maximum number of nematodes reached the -f5 region only after 4 h (Fig. 3, D).
Attraction towards cabbage root homo-genates : Nematodes were attracted least by 20 per cent homogenate of cabbage roots (Fig. 4, A). The attraction increased in the presence of 40 and 60 per cent solutions (p < 0.05, Fig. 4, B & C) and was maximum towards 80 and 100 per cent root homogenates of cabbage (p < 0.05, Fig. 4, D & E).
DISCUSSION
The feeding phases described in Tricho-dorus similis (Wyss, 1971), Tylenchor-hynchus dubius (Wyss, 1973), Naccobus

70
212 BILGRAMI, AHMAD AND JAI RA3PUR1
itn
C
I -
2.
With roots
A
I I I I I I I I I I I I I 1 I 10 15 20
3-
8.
r 6. ^ L
2. TL
L 1
5 1 t
10 1 1 1 1
15
B
20
10-
8L
o 6.
2.
I I I I 11 • I I I I 10 15
10-
a.
3
f> i. u
L. 21
TT'. 20
D
I I I '. I I I . I I I 1 I I 1 I 5 10 15
Ranks 20
Control
6.
L
2.
10-
8.
6.
i .
2.
I I- I I I I I I I I I I I I 10 15 20
I I I ' I I I M I I 1 M I 10 15 20
10-
8.
6.
4.
a 6
i-
2.
n
i J I I I I I I I I M I I I 10 15 20
J
1 I i I I I I I I I I I I I I I I "I 5 10 15 20
Ranks
Fig. 2. Distribution of H. oryzae in the presence and absence of cabbage seedlings. A—1 h; B—2]i; C—3h; D-4h .
batatifoimis and Heterodera cruciferae ingestion of cell contents by H. oryzae juveniles (Doncaster & Seymour, 1973) was intermittent as was also the case in were also observed in H. oryzae. The Xiphinema index (Gotten, 1973; Weischer

ATTRACTION OF H. ORYZAE 213
1\
12;
iti
e c A.
o Z
• With roots 12-
8.
i .
Control
+5+4 +3+2 +1 -1 -2 -3 - 4 - 5 +5+4 +3+2+1 - 1 - 2 - 3 - 4 - 5
•o o
• • - '
n E c
»> o 6 2
12-1
8J
4.
-
1 "H .
B
1 . 1
12i
+ 5+4 +3+2 + 1 -1 - 2 - 3 - 4 -5 - d m ,.
+5+4 +3+2+1 -1 - 2 - 3 -4 - 5
14i
.n 12J V
•o o 10 8J
e <u -c
>^ 4J o o Z
\
-c
c
i
1 . . . 1 I . I
14 C 12
8.
4.
+ 5+4+3+2+1 -1 - 2 - 3 - 4 - 5 H L +5 +4 +3+2+1 - 1 - 2 - 3 - 4 - 5
I 12-1 n
I ' o
r. ""Si D 16.
12.
8.
+ 5+4 +3+2+1 -1 - 2 - 3 - 4 - 5
Regions
OIL. +5+4 + 3+2 +1 - 1 - 2 - 3 - 4 - 5
Regions
Fig. 3, Distribution of H. oryzae in different regions of polytiiene tube in the presence and absence of cabbage seedlings. Arrows indicate point of nematode introduction. A—1 h; B—2 h; C—3h; D - ^ h .
& Wyss, 1976). However, it differs from lack of the median bulb in the oesophagus Hexatylus viviparus where the ingestion (Doncaster& Seymour, 1974). of cell contents is passive because of the Salivation in H. oryzae could not be

72
214 BILGRAMI, AHMAD AND JAI RAJPURI
in « o m E v c
q 6 Z
16
12.
8.
/ i .
16' t/i
to
e c
o z
>—•' -T 1 r-
2 3 i,
•--•Control With homogenate
— 1 — I — I — I — I — I — t 1 2 3 4 5 6 7
Zones
B
*><*—•
16"
12.
8.
./K 1 2 3 4
—1 1 1
5 6 7
U1 V •o o to E <u c
o
z
16i
12.
i..
D
1 2 4 5 Zones
16
12,
i..
•i
I I
•—•-' ' • -T 1 1 1 1 1
2 3 4 5 6 7 Zones
Fig. 4. Attraction of H. oryzae towards different concentrations of cabbage root homogenate. A—20% homogenate concentration; B— 0% homogenate concentration; C—60% homogenate concentration; D—80% homogenate concentration; E—100% homogenate concentration.
observed though it has been reported in quite possible that salivation might have other plant parasitic nematodes (Wyss, taken place between the termination of 1971; Doncaster& Seymour, 1973). It is cell wall perforation and ingestion of the

ATTRACTION OF H. ORYZAE 215
cell contents (Wyss, 1971) but could not be detected perhaps due to its non granular nature (Doncaster, 1971).
The accumulation of H. oryzae near tissue paper appears similar to Meloi-dogync sp. (Lownsbery & Viglierchio, 1960; 1961) and Meloidogyne incognita (Peacock, 1959). It is most likely that root attractantis were responsible for the attraction and accumulation of nematodes near the tissue paper, since factors like the presence of micro-organisms on the root surface (Edmunds & Mai, 1967) and the formation of water film around the roots (Sandstedt & Schuster, 1962) which could cause similar responses, were absent.
Attraction towards various concentrations of cabbage root homogenate suggest that nematodes responded more strongly as the concentration of root homogenate increased. The diiferences in the attraction towards various homogenate concentration may be attributed to differences in the attractant gradients. Perhaps the lower concentrations failed to produce optimum perceptible gradients as compared to the higher concentrations and this may have resulted in former attracting fewer nematodes than the latter.
ACKNOWLEDGEMENTS
We thank Prof. Nawab H. Khan, Chairman, Department of Zoology for providing laboratory facilities and Prof Ather H. Siddiqi for suggestions and U.G.C. for financial assistance to one of us (ALB).
REFERENCES
AHMAD, I. & JAIRAJPURI, M.S. (1980). Sex attraction and copulation in Chiloplacus symme-tricus (Nematoda : Acrobelinae). NeinatO' logica 26: 139-148.
AzMi, M.I. & JAIRAJK'RI, M.I. (1977). Attraction of plant-parasitic nematodes to host roots. Nematologica 23 :119-121.
BARBATOLA, J .O. & BRIDGE, J. (1980). Feeding behaviour and histopathology of Hirschman-
niella oryzae, H. immamuri and //. spinicau-data on rice. / . Nematol. 12 : 48-53.
BrLGRAMr, A.L. (1983). Attraction of Hirschman-niella oryzae towards cabbage seedlings. Proceedings of Third Nematology Symposium, Solan, India. (Abstr) pp. 15.
BtLGRAMi, A.L., AHMAD, I. & JAIRAJPURI, M.S. (1984). Observations on the predatory behaviour of Mononchus aquaticus. Nematol Medit. 12 : 41-45.
BiLGRAMt, A.L., AHMAD, I. & JAIRAJPURI, M.S. (1985). Factors influencing attraction of Hirschmannieha oryzae towards cabbage seedlings. Revue Nematol. (In press).
COHEN, J. (1973). Feeding behaviour and reproduction of Xiphinema index on some herbaceous test plants. Nematologica 19 • 516-520.
DONCASTER, C.C. (1971). Feeding in Plant parasit'C nematodes : Mechanism and behaviour. In Zuclcerman, B.M., Mai, W.P. & Rohde, R.A. (cds) Plam-parasitic Nematodes. Vol. II, Academic press, New York 137-157.
DONCASTER, C.C. & SEYMOUR, M . K . (1973). Exploration and selection of penetration site by tylenchids. Nematologica 19 : 137-145.
DONCASTER, C.C. & SSYMOUR, M.K. (1974). Passive ingestion in a plant nematode, Hexatylus viviparus (Neotyljnchidae : Tylen-chida)'. Nematologica 20 : 297-307.
EDMUNDS, J .E . & MAJ, W . F . (1967). Effect of Fusarium oxysporum on movement ot Pratylenchuspenetrans towards alfalfa roots. Phytopathology 57 : 468-471.
KHERA, S. AZLXKERM^N, B.M. (1963). In vitro studies of host parasite relationships of some plant parasitic nematodes. Nematologica 20 277-282.
KRUSBERG, L .R. (1959). Investigations on the life cycle, reproduction, feeding and host range of Tylenchorhynchus claytoni Steiner. Nematologica 4 : 187-197.
LOWNSBERY, B .F . & VjGUtRCHro, D.R. (1960) Mechanism of accumulation of Meloidogyne incognita acrita around tomato seedlings. Phytopathology 50 . 178-17 *.
LOWNSBERY, B .F . & VIGLIERCHIO, C .R. (1961). Iinportance of response of Meloidogyne hapla to an agent from germinating tomato seeds. Phytopathology 5\ 210-221.
PEACOCK, F . C . (1959). The development of a teclitiique for studying the host parasite relationship of the root-knot nematode.

74
216 BILGRAMI, AHMAD AND JAI RAJPURI
Mehidogyne incognita under controlled conditions. Nematologica 4 : 43-55.
PROT, J .C . (1980). Migration of plant parasitic nematodes towards plant roots. Revue Nematol. 3 : 305-318.
PROT, J.C. & KoRNPROBST, J.M. (1983). Effects of Azadiracta indica, Hannoa undulata and Hannoa klaineana seed extract on the ability of Mehidogyne j'avanica juveniles to penetrate tomato roots. Revue Nematol. 6 : 330-332.
PROT, J.C. & VAN GUNDY, (1981a). Effect of soil texture and the clay components on migration of Mehidogyne incognita second stage juveniles. / . Nematol. 13 : lii-in.
PROT, J.C. & VAN GuNoy (1981b). Influence of photoperiod and temperature on migrations of Mehidogyne]men[\es. J. Nematol. 13 : 217-220.
SANDSTEDT, R. & SCHUSTER, M . L . (1962). Liquid trapping of Meloidogyne incognita around roots in agar medium. Phytopathology 52 : 174-175.
WEISCHER, & WYSS, U. (1976). Feeding behaviour and pathogenecity of Xiphinema index on grapevine roots, Nematologica 22 : 319-335.
WYSS, U . (1971). Der mechanismus der nahrung-saufnahme bei Trichodorus similis. Nematologica 17 : 508-518.
WYSS, U . (1973). Feeding of Tylenchorhynchus dubius. Nematologica 19 : 125-136.
WYSS, U . (1981). Ecto-parasitic root nematodes : Feeding behaviour and plant cell responses. In Plant Parasitic Namatodes. Vol. lit (Editors Zuckerman, B.M. & Rohde, R.A.) New York, U.S.A. Acodemic press pp. 325-351.
Accepted for publication : December 1985

HELMINTHOLOGIA 23 49—'i4, 1986
Effect of some chemicals on the activity of Hirschmanniella oryzae (Nematoda)
A L BILORAMI ! AHMAD M S JAIRAJPURI
Section of Nematologv, Department of Zoology, AUgarh Muslim University, AJigarh — 202 001, India
Received Jun> 20, 1984
Summary
Low contentrdtions of 0 001 M and 0 01 M of sodium chloride and 0 001 M of potassium chloride and ferric chloride were favourable for the activity of H oryzae Ammonia solution at all concentrattons suppressed activity markedly though it had least effects at 0 001 M Except 1 ppm all other concentrations of neraaticide (Furadan 3—G) proved unfavourable for the nematodes At 1 ppm tne activity regime was almost similar to the control
Keywords chemicals,activity HirsctimwniellaorvzaeC^ematoda)
Introduction
Chemicals affect different types of nematodes differently According to Soroczan (1969) low concentrations of mineral salts favoured Rbdbditis sp but low concentrations of silver and cupric ions were not favourable to Pratylenchus penetrans and Xiphinema d/versicaudatum (Pitcher and McNamara, 1972) The effect of pH and mineral salt concentrations on the survival of Hoplo-laimus mdicus. Xiphinema basin, Hehcotylenchus indicus and Mylonchulus minor was observed by Jairajpuri et a/ (1974) Ahmad and Jairajpun (1982) observed the toxicity of pH, mineral salts and fatty acids on a predatory nematode Parahadronchus shakih Very recently, Babatola (1983) while observing the activity and survival of Hirschmanniella oryzae, H immamuri and H spmicaudata in graded solutions of sodium chloride, urea, sucrose and artificial sea water found these chemicals adversely affected the nematodes
In this work the activity of Hirschmann/ei/a oryzae (van Breda de Haan, 1920) Luc et Goodey, 1963 was tested in different concentrations of mineral salts and a nema'icide
Material and methods
To study the effects of mineral salts on the activity of H oryzae, four concentrations, viz , 0 001 M, 0 01 M, 0 02 M and 0 06 M of ferric chloride, sodium chloride, poiassium iodide and ammonia (30 %
49

70 solution) were prepared Similarly for the netndticide Furadan 1—G five concentrations of 1, 5, 10 15 and 20 ppm were prepared Nematodes were treated in each concentratior of chemicals in cavity blocks containing 2 ml solution at a constant temperature of 25 °C The activitv of nematodes was measured in terms of head movements per minute A swing of the head to one side and then back to initial point, and again to the other extreme and back to the initial point constituted one wave Observations were made 0, " ' 4 6 8 10 and 12 h after transferring nematodes in a solution The period immediately preceding immersion of nematodes in solution was regarded as 0 h All experiments were replicated ten times and activity in tap-water served as control
Results
No noticdble differences were observed m the activity of ff or/zaeat low concentrations, but at higher concentrations the nematodes responded differently
Activity in sodium chloride (Fig 1) Activity of H orvzae was not influenced much by sodium chloride solutions Concentration of 0 06 M suppressed ad ivity below control after 8, 10 and 12 h (p < 0 05) However, there was no difference in the activity of nematodes during 2 4 and 6 h (p > 0 05) In 0 02 M solution activity increased significantly at 2, 4 and 6 h (p < 0 05) but declined thereafter The activity remained above control at 0 01 M while 0 001 M inhibited activity upto 4 h, after which this concentration had little effect and the activity became more than the activity observed in tap-water
Activity in feme chloride (Fig 2) High concentrations of ferric chloride suppressed activity At 0 06 M and 0 02 M it remained much below the control throughout the entire period of observation (p < 0 05) Concentrations 0 01 M and 0 001 M appeared more suitable and the activity regime was similar to tap-water except that during 0 and 6 h in 0 01 M where it was less than the control
Activity m potassium iodide (Fig 1) All except 0 001 M concentrations of potassium iodide were unfavourable for the nematodes The activity at 0 06 and 0 02 M declined sharply and remained below control throughout entire period of observation (p < 0 05) 0 01 M
20
15
I 10 Q < LU
I ^ 5
^
© 4 6 TIME(h)
10 12
Fig 1 Activity of H oryzae in different concentrations of sodium chloride (Legend of Fig 1 see p 511
'iO

20
^''
I 10h Q < LU
I
>
©
t X 77
4 6 TIME (h)
10 12
Ftg 2 Activtty of H oryzae in different concentrations of ferric chloride
23r
15 z LLI
z LU
I 10 Q < IXJ
I > 5 H > I -
© + + Control • • 0 06 M (Legend of Figs 1—3)
4 6 TIME(h)
10 12
0—
3—
— O 0 02M — • 0 01 M —L] 0 001 M
Fig 3 Activity of H oryzae m different concentrations of potassium iodide.
stimulated activity upto 8 h (p <{).05) After 8 h activity declined below control (p < 0.01). TTie most favourable concentration for activity was 0.001 M where nematodes showed enhanced activity upto (10 h (p < 0.05).
Activjfy in dmmonia (30 % so/ufion)(Fig. 4): The activity of H. oryzae at 0.06 M declined after 2 h and remained below control (p < 0.05). 0.02 M enhanced activity only upto 4 h but reduced it thereaftertoabouthalf of control at the end of 12 h(p < 0.01). The activity pattern
5]

20
H15
LU
g lO Q <
> U <
© 4 6 TIME(h)
10 12
Fig. 4. Activity of H orv/at- in different concentrations of ammonia (30 % volution)
-+ Control - • 0 06 M
O O 0.02 M B H 0.01 M c_—r, 0.001 M
20
15
Z ui
> o r Q < LU
I 5 > -
10
^
©
• • Control O—O 1 ppnfi O—O S ppm
••• • •10 ppm CV"Ol5 ppm O—O20ppm
4 6 TIME(h)
10 12
Fig. S. Activity of H. oryzae in different concentrations of Furadan 3—G
52

at 0 01 M was almost similar to 0 02 M but slightly higher 0 001 M had kast effects and the activity remained equal to the control except during 0 and 2 h
Acfivify in Furadan 3—G(Fig 5) The activity of H oryzae was greatly suppressed by all concentrations of Furadan 3—G except 1 ppm in which it was almost similar to control (p < 0 01) At 5, 10, 1 "i and 20 ppm activity decreased and was least at 20 ppm
Discussion
The effectiveness of salts on the activity of H oryzde corresponded to their molecular weights and the molar concentrations Salts having higher molecular weights, viz , potassium iodide (mol wt 166 01) and ferric chloride ( mol wt 162 21) inhibited activity of adult H oryzae to a greater extent, while sodium chloride having low molecular weight (mol wt 58 45) had least effects The same conclusion was drawn by S t e p h e n so n (1945) and J o h ns t o n (1959) who also found effectiveness of salts to be proportional lO their molecular v/tights in Rhabditis ferrestris and Tylencborhyncbus martini, respectivelv The ineffectiveness of low concentrations of almost all salts on the activity of nematodes is in agreement with Soroczy n (1969) and Ja i ra jpur i ef ai (1974) Both the authors found that low concentrations of salts were most suitable for the survival of Rhabditis sp , and Mylonchulus mmoi Helicotylenchus indicus, Hoplolaimus mdicus and Hiphinema basin, respectively Ahmad and J a i r a j p u n (1982) also reached the same conclusion in their study of Parahadronchus shakuli
Although ammonia had the least molecular weight (mol wt 17 03 Sp Gr 0 89) it proved most unvafourable for nematodes This was perphaps due to its capacitv to generate higher osmotic pressure even at ow concentrations (cf Vassa l lo , 1967) Furadan 3—G which contains 3 % active ingredients of Carbofuran reduced activity at all concentrations The nematodes seemed to have narrow range for the nematicide Puiadan 3—G also effectively controlled juvenile populations of H oryzae in soil (S a m a n t a r a i and Das 1971)
Acknowledgements
We thank Professor Ndwab H Khan Chairman Department of Zoology for providing laboratory facilities and Professor Ather H Siddiqui for his suggestions The first author is thankfull to L G C, New Delhi for financial assistance
References
AHMAD, N , JAIRAJPURl, M S (1982) Fffect of pH, mineral sails and fatty acids on the predatorv nematode Parahadronchus shaki/i (Jairajpun 1969) Indian J Sematol 12 22—30
BABATOLA, J O (1983) In vifro studies on the activity and survival of HirschmannieWa spp in some chemical solutions Anna/s of App/iedBio/ogy, 102 355—363
JAIRAJPURl, M S , AZMl, M I, BAJAJ, H K (1974) Effect of pH and salt concentrations on the survival of Hoplolaimus indicus, Helicotylenchus mdicus, Mylonchulus minor and Xiphmema basin Indian J Nematol,4 171—181
JOHNSTON, T M (1959) Fffect of fatty acid mixtures on the rice stylet nematode (Tylenchorhyncbus martini Fielding, 1956) Nature (London), 183 1392
PITCHER R S,McNAMARA, D G (1972) The toxiaty of low concentrations of silver and cupric ions to three species of plant parasitic nematodes Nematologica, 18 385—390
53

80 SAMANTARAl, K G DAS S N (1971) Control of plant parasitic nematodes in rice soils Proc
Jndun Acad Sc,, 37 B 372—376 SOROCZAN. W (1969) The effect of pH and the concentrations of mineral salts of the medium on
survival rate of larvae and adults of Rhabditis spp , under laboratory conditions Annals Univ \4ana Curw Sklodowska 24 153—164
STEPHENSON, W (1945) The effects of acids on a soil nematode Parasifo/ogy, 36 158—164 VASALLO M A (1967) The nematicidal power of ammonia Nematologica, \3 155(Abst)
B.iiiHHHe HeKOTopbix XHMmecKHX BeuiecTB Ha HeMaTonu Hirschmaniella oryzae
BblBOflhl
HH3KHe KOHUeHTpauHH - 0 001 M M 0,01 M xjiopHfla HaxpHH H 0,001 M xjiopHfla KajiHJi HJIH TpHXJiopHfla )Ke;ie3a OKasa.iM SjiaronpHsi Hoe BJiHHHHe Ha aKTMBHOcTh H orvzae PacTBopbi nama-TbipHoro CHHpra so ectx KOHueHrpauHHx apKO noaaajlsijiM aKTMBHOCTb, XOTH caMot cjiaSoe BJIHUHHC HMe.i.' KOHueHTpauHJi 0 001 M 3a HCKjiioteHHeM I ppm ocTajibHbie KOHueHxpaiiMH HeMJTHUHfla (Furadan 3—G) aejia.iHCb J I H 3TOM HeMaroflbi BpcAHbiMH HpH KOHueHxpaitHH 1 ppm npoasjieHHa
aKTHBHOCTH HBJlHJIHCb OMCHb nOfloSHblMH KOHXpOJIK)
A M BHJjrpaMH M AxMea
M C ^upaiinypH
Der Einfluss einiger chemischer Stoffe auf Nematoden Hirschmaniella oryzae
Zusammenfdssung
Niednge Konzentrationen — 0 001 M und 0,01 M von Natrium-chlorid und 0,001 M von Kaliumchlorid oder Eisentrichlorid beeinflussten gunstig die Aktivitat von H oryzae Die Losungen von Ammoniak in alien Konzentrationen unterdruckten markant die Aktivitat, auch wenn die schwachste Wirkung die Konzentration 0,001 M hatte Mit Ausnahme von 1 ppm zeigten sich die ubrigen Konzentrationen des Nematizide (Furadan 3—G) fur die Nematoden schadlich zu sein Bei der Konzentration 1 ppm waren die Aktivitatsausserungen sehr ahnlich der Kontrolle
A L Btlgrami J Ahmad
M S Jairajpun
Vplyv niektorych chemickych latok na nematody Hirschmaniella oryzae
Subrn
Nizke koncentracie — 0,001 M a 0,01 M chloridu sodneho a 0,001 M chloridu draselneho alebo chioridu zeleziteho pnaznivo ovplyvnovali aktivitu H oryzae Roztoky cpavku vo vsetkych koncentra-ciach vyrazne potiacovali aktivitu, i ked najslabsie posobenie mala koncentracia 0,001 M S vynimkou 1 ppm ostatne koncentracie nematidicu Furadan 3—G sa ukazali byt pre nematody skodhve Pri koncentracii 1 ppm prejavy aktivity boh vefmi podobne kontrole
A L Bilgrami I Ahmad
M S Jairajpun
54

Ill I NHNillf)! ()(,IA, n 119 \)}. V)'U
KespOBBSiL* oJ MiiischiiiiiiuneHh} o/fjzae jisvu'oiJos toward' raW)iiige seedfjiigs
I BILGRAMI.A L , AHMAD 1 , JAlf^AJPURI, M S
Section of NenuKolury, DcpjiiiiiLiil p[Zoology, Alig.iili Mihliin
Uiuvcibiiy, Aligdih —202 001, Indu
I
Rcccncd C)Llubcr26, 198^ I
Siiii'uiinry
'] lie .lulhof'; sliidicd llic pliLiuiiiiLiiDm (if .illrjclioiiof ////si/;i(ia;i/c//j or>7.ic (Van Bicda dc llj.vii, 1920) Luc cl Goudcy, 1963 JUVLIU'C sin;;! S tow.irds cnblijyc bcedliiiys (Or.uiic.i O/CT.T(CJ L var cipnol^) oi aqar pl.ilcs 'Hic juvenile H ory/ic jppciicd lo be posKivciy inraclcd bv cibbigc scodlin;; Hic aUr.iLlion liLpeiu). on the .ij'e of ';i,c(llirij' loot'i TIKI on (lie a[;c of ni.niatodcb pioptr I oiirili dcvelopniciiljl sla{;c was found lo be attracted more than third or second stages
Key words" /{if;>chmi-'nnie//a or>?ac (Ncmatodaj, Bla^•;lC5 olnncca L , alfracfun (jt(ra<(Tii(3J
//ifjor/ijt/jon
'11 e pi ml pal isiiu ucniaiinKs n ^poud lo am ul ml piodui.( d li) llit mois (l!i i d , l"^'i, I'KiO
L.owcnsbcry and Viy l i c rc l i io , IVOO, 1961 , La va I Ice and Kol ide , 196^ , A/in i i nd Ja ir i j()i)J i
1977) Glol'ndcni rosmi/i/c/iMS jnv inks migrated towards [loinlo and sujjii bti t roots \/\n n in rwpi 'ct
uilh tlicir rui'I diffiisatts (Wci sc l i e i , I9')5) Al l r ic lmb persisted in the soil .\i\{.\ influLiitcd ihr
iitiiuitodes e\eii litter the pliint. were removed (Wal l aec , IVit!, Luc , l'>(>2. 1 ro t , 19^5) Uic
Httr n.tuiuof the juveniles of Afcfcu/o^vne ineo;;ni( i, M li [i( i and At jav inic i tov ards ti m iio loni. .</1'
obsersed bv I'rot and Van G u n d y (1981 a, ii) l i i ly r i in i L; a/ fin prcss^ ol ser\ed some faciors
ip'Uiein-iiig the allraction of iidult /iirsc/i/i).innit//a ury^ac toi/ards cabbafc sccd!.m;s Ihc present v.in'
was carried out to ob'crvc the ultifiction of juvenile stages of Ihrschm.iunitll> oi)/ac (^'in Hri d i ih
H lan, 1920) Luc el Goodey, 1963 towaids cabbage seedlings ([ir.issica olcracc i L \ ir c ipiiai i) m .w ir
pi lies.
A/.itcTKii nnd mcilioc/s
'Ilij soil samples containing juveniles of / / oij/ac were collected from ti j p idd^ field; I'c ir I'l
villajjc Jaiiialfxir, Ali^arli and (lie (icrii.Ho<ics ucrc isoliicd by dcciniadoii and ,c\u'[' ,ii.d i.iodifi !
Oacrnnnn's funnel tcclinu|Ucs '1 he (abbTgeseedlin{',s \,'eie gciinuiatcd in 1 % water a" ir and stcfi'i/c(l
111 0 1 7u iiiercurie chloride solution as \/rrc also the nciii ilodes Allrat tionof juv( tiib s lo / iidsi'iffr n nt
a;;e [roups of cabba(;c roots sva, levied b) I'eiii disli i"X])eninenis (Aliinad Mui J a i r . i j p u i , , I'lo'i)
U9

82
Pi;iri-iJislic5 5.5 cm in uiaaitjtcr were diviilcil INIO ilucL-Z()ncsbydi.iwing(v/()cuin:cnlric,Liiclc5i)f 'j ';,,,(( 2.5 cm in diaineicr, the latter also known as reference circle. E.xcised roots from 2,4, 6. 8. 10 and 1/ (J,-,Y old roots were placed in the inner circle of se|)aralc I'etri-dislies containing 0.5 % vvater-aj;,i( ,|,|j incubated for 12 h. After incubation, 5 specimcs of each juvetiili- stage (secojid, third and foiirlli) f/^f^ introduced in separate Pclri-dishes, at different points on the reference circle. Tlius juvenile stat;(:i, ^„,.f^. tested separately towards each ago group of root. The disirihulion of juveniles v/as rccoided afi,., ,j i, converted to score; and log scores. Each experiment v/as replicated 15 times and controls v/i;//; ju^ without host roots, l l ie experiments v/crc carried out at 25 °± 1 °.
Results
Ma,\ifnum atlraclion ofsccond singe juveniles was ob.served towards 4 and ^dayo ld /oo ts
(moan loy score 1.77 and 1.84, respectively, p < O.O.'i), v/liicli was, liov/ever, not differciK / / „ , . ,
tiie aitraclion towards 2 and 8 day old roots ( p > 0.(J5). Second stage juveniles did tioi rci; [ ,( ,„ j
o
Ai
C fO
MS-] 2-5a
2J0.
1.90-
UO.
130-
l.on
\
A
2 4 5 8 1Q 11
Ago o} roots (dnyc)
2.65-| 2.5a
720.
o ^ 1.9a o
c 1-60.
1-30]
1.00
-oWlth rootu -oControl B
\ . \
:o
2 i 0 0 10 12
Ago of tiwta (dayc)
2.2a
2 L G
Aoa of roots (days)
-1 ; 1
0 10 12
Fig. l.Effcctofdiffcrcntagegroupsofcahbagcrootsonihcaitractionof//.oryz.-iejuveniles. A — .'v.-(:f,r,d stage juveniles; D — 'Ili ird stage juveniles; C — Fourth stage juveniles
120

83
towards lOand 12 Jay nkl looir, (iiiuaii loy score 1.37aiid J.25, res(,'L-ciivcly) (l-'iij. l,A).'lbc'', and 6 day old cabbage seedlings attracted maxiniuni niinibcr of third stage juveniles (mean lop, score 2.00 and 2.02, respectively, p < 0 . 0 5 ) . In the presence of 2 and 8 day old roots juveniles showed lesser degiec of attraction (mean log score 1.71) and 1..57, respectively). Third siace juveniles were uniesponsive towards U) <\\u\ 12 day old roots (Fij;. 1, B). The fourth si.r';-juveniles were attracted by all age groups of roots e;(Cepl 12 day old toots (mean log score I.4(S, p>().05) The 2, 4 and 6 d ly old roots were most attractive to them (mean log score 2.29, 2,37 and 2.37, respectively) Attraction of these juveniles declined .significantly lov/ards 8 and 10 day old roots (mean log score 2.02 and 1.6.5, respectively, p <0.05) (f-'ig. 1, C).
The observations on the attrtiction of 11. (Hyz.'ic juveniles indicate that they v<ere ,:!ractcd positively towards the seedlings of cabbage. The attractatits produced by the roots v^erc most likely rcspvmsibic for ;\ccumnlalio i of the juveniles ariound llieni (Ijird, 1959 ; La va I l ace and R o h d e , 1962). Factors suci as miero-organisni on ilic root surface ( l i d m u n d s and Mai, 1967) and the [ormaiion of water film around roots (Sands tsd I and S c h u s t e r , 1962) which were avoided, supports that the nematodes were attracted due to attractants produced by the roots, llie attraction of H.oryzac juveniles depended upon the age of roots as v/cllas on their own age, fotuth stage being most attractive than third or second stage juveniles. This differential attraction may be attributed to the differences in their inherent locomotory speed (Azmi and J a i r a j p u r i , 1977). Bi !gram i ct;il. (in press) concluded that increase in the age of roots, decreased their attractiveness and 12 day old r()ots failed to attract fed U. oryzuc. hi case of H. oryzae juveniles also a similar phenomenon was c)bscrved, some older age groups of roots (10 and 12 day old) failed to attract diffcient juvenile stages.
References
AHMAD, 1., JAIRAJI'URI, M. S. (1980); Sex utlraclion UIKI coiiulalioii in Cliihpheur, synimelricns (Ncinaluda: Acioheliu[ic). Ncn\iiioloi;iv:i, 26: 1.19—MK
AZMI, M. 1., JAIRAJl'URI, M. S. (1977): Attraction of phml para.silic iicniniodcn to host rools. /Ve;ii.i(ol()^;ic;i,23; 119—121
HlLCiRAMI, .A. L., AIIMAO, I.. JAIRAJl'URI, M. .S. (in press): I'anois inllucriciiigaitracliiin of aduli //iVsc/ini.'i/iii)e//a oryzLtc towards cabbage scddliiig. Revue Ncnuiol.
BIRD, A. F. (lyi'J): The altractivciicsu of roots to llic p|ynt-p:irasiti(; nematodes Mcloidogyiic j:n:u>icii and Af. hnph. Nemato/og/ca, 4: 322—335
GIRD. A. 1". (ly(il)): Additional notes on the nllraciivcncs.s of roots to plam-parii-.ilic ne; laJodc;. Neina(o/oi,'ic.T, 5:217 1
EDMUNDS, J. E., MAI, W. F. (1967): Effecls of Fu.s.inuni (),v)S7)orum on niovemcru (if /Va/y/e.-ic/ju'; peiictra/is tov.'ards alfalfa roots. F/iylo/^Jt/id/oi:)', 57 : 4{i8—471
LAVAl-LEF. \V. H., ROllDE, R. S. (19()2): Atlraelivcne.'.s of plani roots to PrMytcnehw; pcueli:iin (Cool)li), Kcniaio(uj:ic'.H;252—2<)U ;
LOWNSBERY, B. F., VIGLIERCHIO, D. R. (1960): Mechanism of accumubtion of Meloidogync incoi;iii(;! acrua around tomato seedlinc.s. F/iytopat/io/ogy, 50; 17K- 179
LOWNSBERY, li. F., VIGLIERCHIO, D. R. (1961): Inipoitanjcof rcsponso ul ,Mc/oidogync /ia/)la to an aitcnt ftoui [lerminiuin!; tomato seeds. I'hytopjiholoi'.y, 51; 219—221
LUC, W. (I9i'2); Jiar roricmalioii dcplacenienl ile //ennVyc;/()/)/ior,'i piinuloxj (lain li; MJI (,.'. '• IICIHI. Seuuc. 5Vi. I'.iri.s-.. 251: 3124—3126 • I
PROT, J. C. (1975); Research on movement of Afe.'oulogync spp., juveniles towards roots. Scric. iJio;oi.';e/Vejii.iiologie, 10: 251—262- » '
PROT, J. C, Van GUNDY (19Kla); Effect of soil tcMure and the clay conipoiicnis on migration of Afe/oKfoi;\iie iiicogiiiia second ^tiigc juvenilei;. J. Ne/nato/., 13; .''13—217
1 2 J

I 'ROI , J C , V.iii GUNDY (l'>hll)) IiifluLiiLC of plKJlopLnod .uiil inn,it, iiiic on IMIJ-I iiioi of MLloidoi'yncJUVLIUILS J Nmuiiol, 11 217—2?0
S A N D S l ^ D I , R , SCHUSrCK, M L (1962) I iqiml trapping of Aft/o/c/i;; yn. //iccy/jj J jrouiul roo.s 111 ,ii',ar iiKiliiini I'hjlopr'lwki^'y, 52 174—175
W A l L A C r . I l R (19S8) OIISLIV uioiis on the i-!iu rfi iici, froni cysts iml tin. oru iit iii ri of lar i of llun: spULiL'i of till, genus //LfcnH/cni m llic p ic tnc i ; of lio.t pi nit tool. N( iii iioloyn i 1 : 3 ( . - 2 4 3 I
WEISCIILR B (1959) Ep(.nniL'ntLllo linlcrMicliiingci) ubcr d<c WjiidL-rtiiij; von Nuni iod^n Nc-iii.i(o/oi;/c,i.4 172—186 '
llpiniiinu.iiiiio loiiciiiuit.iii.ix tnijiint iitM.nn'i llir<(.liiii,uiiiii.ll:i vry/tic II piH(. j , i iM KiiPiyf. 11,1
B LniTi)L" auTopbi oiiiicijiunK)T)ii"icHiif[ipin^rMii.iiimi ioii^muii,iii,ix r r j iiifi hir',chn]jiutlUt orv/.iL (V.iii BrLd.i dc Haaii, 1970) I tit. ct Goodev, 1963 K p<iccj/i;i" K.iiiycri.i (/Jriss/c.i o/tr.icci f '..ir c jpint j ) IIJ i\r.ipoiu,ixtiJiJcriiiiKax OKai.iJuici.,'!ro loiicmuu.iii.ic / / ory/jct'Oiiiriii'iio niiiir.ii ,t( uoti -i K pacca;iaM KaiiycTiji ripninriiuaiiiic jauiicuT oi DOJpacTa iie'iaTonui 'icrrcin isi 'iii'iiiiio'iHaH crra i,')i HBjmcTCS Go;iee npnTsirnuaiitiori MCM ipcTi.ii luiii uio|)aM
! A // /:>iij7f7jaMi(
/ / Ax". '7 M C ''\iip i.uftpii
I Die Aiizichi-ii|; jmciiiJL'r Slndien voii Ncniulodcii limchiPaunitUti
ory7'Jomi Kroulpiluiuhn;;e I
Itli Ucilrag wird die rrbcliciiuing tier Allraklioii jiivciiilcr SladiLn lIu'^Lhiiuinnitlli oiy/n (Von Bred 1 dc Ha,in, 1920) Liit ct Cioodi y, 1963 ,iii Kniupfl in/lini'i (/J; nwi i oft f m 11 \ n ( I/I/M; i ) mf Agaipl itlcn l)(.ol),ulitcl I . /ci/'lc bii li, d is, juvc mlc II i>i)nt .in Kr,iiil|)fl III/IIII|'L posiliv nij'i /o r n wcrdcii Uic Atliakiion h mgt voiii Mtcr ikr VVIU/LIM dcr i'fl iii/linrc sciv ic aui.' '.ciiii c'ljct'cn Alitr J( s Nciiiatodcii ab das vitrtc LntwickUiiigsstadiuiii isl mclir angc/ogcn als das tli itic odcr das Z\/LIIL
! A L Hih'idtm
I \lw,id ,M S J.uroijjiin
I
Pritiilioviiiiic juvciiihijih sljdii iieinuludov H:nil!!n.!ii.uLh] (Uy,aiciv pricsudi'Di kmiustir
Suhrn
V prisp(.vkii s 1 slcdiijc jav atrakcic juvi nihntli sladii IhrsduuMMiLll i orr/ it (\'dii BrnI i dt 11 i MI 1920) I IK CI CjDudcy, 1963 k puts ul iiii k.ipiisty (/(/ i sn i o/t/ iti / I s ir ( ipii it i| n i .ir 'lo ,i li platii idi Uk.i/alo sn, 2c jnvcnilnc / / o i ) / i c su |)o/iliviic iirilahovanc k iiri-s id iiii k.'pujiv Alrat ti i /avisi lia VLkii korcnov pu^s.'d ako .ij n.i \lisnu)in VLIIU iitmalodi sl\nL \ ) , i no \ c sladunn jt pri(ahu\anc viae ako trciic alcbo driiiit.
A L HI T.IIIII
I /Mini III \l S Jtr.iipun
122

85
A study of the intestinal contents of some mononchs Anwar L. BILGRAMI, Irfan AHMAD and M. Shamin JAIRAJPURI
Section of Nematology, Department of Zoology, Aligarh Mushm University, Aligarh-202001, India.
SLMMARY
The intestinal contents of over one thousand specimens of thirty three species belonging to nine genera of the order Mononchida were analysed to determine their range and preference for prey. The study showed that mononchs feed upon tylenchs, dorylaims, mononchs and frce-livmg saprophagous nematodes besides rotifers. Cuticular remains of all types including trophi of rotifers and other unidentifiable objects of prey were of frequent occurrence. Forty five percent of the predators had prey m their mtesune. Feeding of different mononchs is polyvalent and predation aleatory. However, more predators (75 "n) had free-living saprophagous nematodes within the intestine rather than tylenchs (45 "n) or dorylaims (42 'H). Pratylenchus, Hoplolaimus, Tylenchorhynchus, Tylencholaimus, Aporcelaimus, Thomenema, lotonchus, Mylonchulus, Rhabditis, Acrobeloides and Chiloplacus were found most frequently in the intestine of different species of mononchs. Parahadronchus shakih appeared to be most active predator as 68 "n of us specimens had prey m their intestine while Coomansus indicus was least active with only 21 "o having prey. Twenty percent predators were cannibals while 27 'o devoured mononchs belonging to other species genera Mylonchulus spp., and lotonchus haqni did not contain any prey or its remains m their intestine. None to eight prey could be present in the intestine of a single predator.
RESUME
Etude du contenu intestinal de quelques Mononchides
Le contenu intestinal de plus de mille specimens appartenant a 33 especes et neuf genres de 1 ordre des Mononchides a ete analyse en vue de connaitre I'eventail des proies et les preferences dans la predation. La presente etude a montre qu'en sus des rotiferes, les proies des Mononchides appartiennent aux Tylenchides, Dorylaimides, Mononchides et aux nematodes libres saprophages. II a ete frequemment observe des restes cuticulaires de nature vanee, y compris des trophi de rotiferes et d'autres portions de proie non identifiables. L'lntestin de 45 "o des nematodes predateurs contenait une ou des proies. La nournture des Mononchides est polyvalente et la predation aleatoire. Cependant, un plus grand nombre de predateurs (75 "o) contenait des nematodes libres saprophages plutot que des Tylenchides (45 "o) ou des Dorylaimides (42 "o). Les representants des genres Pratylenchus, Hoplolaimus, Tylenchorhynchus, Tylencholaimus, Aporcelaimus, Thomenema, lotonchus, Mylonchulus, Rhabditis, Acrobeloides et Chiloplacus sont les proies les plus frequemment observees. Parahadronchus shakih parait etre le predateur le plus actif car 68 "o de ses specimens ont une proie dans leur intestin, tandis que ce pourcentage n'est que de 21 "o chez Coomansus irdicus, espece la moms active Vingt pour cent des predateurs sont « cannibales », tandis que 27 "o devorent des Mononchides appartenant a une espece ou a un genre differents. Mylonchulus spp et lotonchus baqni nc contiennent aucune proie ni aucun reste de celles-ci Les proies peuvent etre au nombre de zero a huit dans I'lntestin du meme predateur.
Observations on the predatory behaviour of nematodes of the order Mononchida Jairajpun, 1969 have been made (Cobb, 1917; Steiner & Hemly, 1922; Thome, 1927; Nelmes, 1974; Jairajpuri & Azmi, 1978; Bilgrami, Ahmad & Jairajpuri, 1984) and the possibility of using them in controlling population levels of some species of plant-parasitic nematodes has been assessed (Cohn & Mordechai, 1974; Small, 1979). Mohandas and Prabhoo (1980) found Rotylenchulus remformis Linford & Oliveira, Meloidogyne incognita (Kofoid & White), Rhabditis sp. and Monohysterella sp. to be most common in the gut of mononchs. Arpin and Kilbertus (1981) made observations on the intestinal contents and epithelium of these nematodes. While observing the predatory abilities of nematodes belonging to different orders on a range of prey species. Small and Grootaert (1983) found Panagrellus redwwus (L.) to be most
susceptible to attack than other free-living or plant-parasitic species. In the present work the intestinal contents of mononchs belonging to 33 species of nine different genera were studied. This study was carried out to determine and enumerate different kinds of ingested prey and to find out whether these mononchs have any choice for a particular type of prey.
Materials and methods
Observations were made on the intestinal contents of mononchs that were collected during past several years from all parts of India. Over one thousand specimens of mononchs belonging to nine genera viz., Parahadronchus {ont species), Mononchus {pfio species), Miconchus (three species), Clarkus (two species),
Revue Nematol, 9 12) : 191-194 '1986/ 191

u A. L Bilgrami, I. Ahmad & M S Jatrajpun
Pnonchulus (one species), Sporonchulus (one species), Coomansus (one species), lotonchus (thirteen species) and Mylonchulus (nine species) were studied.
Results
All species had vanable number of prey in their intestine. The nematodes identified as prey are given in Table 1. In lotonchus, I. tnchuns and /. monohystera were most active predators while /. basidontus was least active. Other species are of intermediate grouping
except /. baqni which contained no prey or its remains in the intestine. /. trichuris preyed upon three genera each of Tylenchida and Rhabditida and two each of Dorylaimida and Mononchida. Three genera each of Tylenchida and Mononchida, one of Dorylaimida and three of Rhabditida were identified as prey in the nematode genera were most commonly found in had preyed only upon one genus of Tylenchida and two of Rhabditida. In Miconchus, more specimens of M. aquaticus contained prey in their intestine than M. dalhousiensis. M. aquaticus preyed upon two genera each of Tylenchida, Mononchida and Rhabditida and
Table 1
Predator-prey list
Predators
Parahadronchus shakth
Mononchus aquaticus M. tunbndgensis Miconchus aquaticus M. Clin M. dalhousiensis Clarkus papillatus C. shen Pnonchulus muscorum Sporonchulus vagabundus Coomansus indicus lotonchus antedontus I. baqnt I basidontus I tndicus I. jam I. longtcaudatus I monohystera I. parabasidontus I. prabhooi I. nsoceiae I shafii I. tnchuns lotonchulus sp. Mylonchulus spp.
Tylenchs
1, 2, 3, 4, 5, 6 ,7 ,8 2 ,3 ,8 2 ,3 ,7 4,6 1,3 9
7,9 3,8
2, 3, 7, 8 1,6,9 1,3,7
3 — 3
3,4 2
2,5 1,2,5
5 — 1 2
1,2,3 5 —
Dorylaims
10, 12, 17, 18, 19,21
10, 13, 14, 19 10, 17
10, 15, 18 11
10,20 11, 13
11 11, 12, 15 11, 12, 19
18 14 — —
10, 12 16 12 10 16 11 11 12
12, 18 — —
Prey
Mononchs
22, 23, 24, 27, 29
23,24 24
22,28 23,28
23 25,27 25,28
22, 23, 28 28, 30
28 23 — —
22,23 25
24,26 22, 24, 27
22,23 —
22,23 —
22,26 — —
Free-Living
31, 32, 33, 36, 37
32, 33, 34 31, 34 31, 32
33 34
31, 32 34
31, 34 33, 34 31, 33 34, 35
— 31, 32 33, 34
33 33, 34
31, 34, 35 32, 33 32, 35
32 31, 32
31, 34, 35 — —
KEY :
1. Pratylenchus 2. Hoplolaimus 3. Tylenchorhynchus 4. Helicotylenchus 5. Hirschmanniella 6. Hemicychophora 7. Hemicnconemoides 8. Aphelenchus 9. Aphelenchoides
192
10. Tylencholaimus 11 Aporcelaimus 12. Thomenema 13. Discolaimus 14. Belondira 15. Axonchium 16. Dorylaimellus 17. Nygolaimus 18. Xiphinema
19. Tnchodorus 20. Diphtherophora 21. Campydora 22. lotonchus 23. Mylonchulus 24. Mononchus 25. Clarkus 26. Coomansus 27. Pnonchulus
28. Miconchus 29. Parahadronchus 30. Sporonchulus 31. Rhabditis 32. Acrobeles 33. Acrobeloides 34. Chiloplacus 35. Mesorhabdiiis 36. diplogastend 37. monohystend
Revue Nematol, 9 12): 191-194 11986)

87 Intestinal contents of mononchs
Parahadronchus Mononchus Miconchus Clarkus Prionchulus Sporonchulus Coomansus lotonchus Mylonchulus
Total
Number of specimens observed
164 198 34 62
105 59 24
173 190**
816
Table 2 Analysis of intestinal contents of mononchs
Number of specimens containing
prey
112(68%) 87 (44 "o) 15 (44 °o) 26 (42 »o) 32 (30 "o) 16 (27 %) 5 (21 "o)
75 (43 %) 0
368 (45 %)
Dory-laims
42 22 10 4
18 4 4
50 0
154
Number
Tylenchs
48 24
8 6
20 6 5
49 0
166
of sped
Fre^ living
68 55 15 17 22 16 4
70 0
277
mens containing diffi
Unidentified prey
48 33 6 8
24 8 4
52 0
184
Cuti-cular parts
21 19 3 4
16 7 2
24 0
96
3rent preys
Mononchs Mononchs of other genera
38 10 4 8
12 3 1
24 0
100
of same genus
14 23
3 8 0 6 0
20 0
74
Including monohysterid, diplogasterid and rhabditid nematodes. Not included in total as no specimen of this genus had prey or its remains in the intestine.
three of Dorylaimida while M. dalhousiensis on one genus each of Tylenchida, Mononchida and Rhabditida and two of Dorylaimida. Mononchus aquations preyed upon more varied type of nematodes (three genera each of Tylenchida and Rhabditida, four of Dorylaimida and two of Mononchida) than M. tunbridgensis (thrtt genera of Tylenchida, two each of Dorylaimida and Rhabditida and one of Mononchida). Similarly, Clarkus papillatus preyed upon two genera each of Tylenchida, Dorylaimida, Mononchida and Rhabditida while C. sheri predated only upon two genera each of Tylenchida and Mononchida and one each of Dorylaimida and Rhabditida.
Table 2 shows the distribution of prey in the intestine of different species of mononchs. The following nematode genera were most commonly found in the gut of mononchs : Pratylenchus, Hoplolaimus, Tylenchorhynchus, Tylencholaimus, Aporcelaimus, Thomenema, lotonchus, Mylonchulus, Rhabditis, Acrobeloides and Chiloplacus. Other prey not too frequently encountered belonged to the genera Helicotylenchus, Hirschmanniella, Hemicycliophora, Hemicriconemoides, Aphelenchus, Aphelenchoides, Discolaimus, Belondira, Axonchium, Dorylaimellus, Nygolaimus, Trichodorus, Xiphinema, Diphtherophora, Campydora, Mononchus, Clarkus, Coomansus, Prionchulus, Miconchus, Acrobeles and Mesorhabditis besides some diplogasterids and monohysterids. Tylenchorhynchus, Tylencholaimus, Mylonchulus and Chiloplacus were found within the intestine of maximum number of species of mononchs. Tylenchorhynchus was observed in the gut of eleven species, Tylencholaimus in seven, Mylonchulus in nine and Chiloplacus in eleven species belonging to different genera (Tab. 1).
None of the species of Mylonchulus (M. brachyuris, M. contractus, M. dentatus, M. lacustris, M. hawaiiensis, M. amurus, M. muradi, M. sigmaturus, M. nainitalensis) contained prey or its remains in the intestine. Forty five percent of the remaining predators had prey in their intestine (Tab. 2). The preys were observed either whole or in semi-digested conditions or their cuticular remains were found in the intestine of the predators. An analysis of intestinal contents showed that mononchs feed upon tylenchs, dorylaims, mononchs and free-living nematodes. Some specimens of lotonchus trichuris, I. longicaudatus, I. indicus, Clarkus papillatus, Coomansus indicus, Sporonchulus vagabundus and Parahadronchus shakili also preyed upon rotifers besides soil and plant-parasitic nematodes. Various types of cuticular remains such as spear, spicules, buccal cavity, cuticle, trophi of rotifers and other unidentifiable structures of prey were found frequently in the intestine of the predators.
Discussion
Under natural conditions mononchs probably feed upon all types of nematodes besides rotifers and some soil microorganisms. Arpin (1979) and Mahapatra and Rao (1981) found signicant correlation between the populations of mononchs and free-living nematodes but Nelmes and Mc CuUoch (1975) did not find such a correlation. Since the present observations were made only on mounted specimens and not on actual populations, the relation of mononchs with other nematodes present along with them in the soil could not be determined. It cannot, therefore, be suggested with certainty that the widespread presence of free-living nematodes in the intestine of different mononchs is due
Revue Nematol, 9 (2) : 191-194 (1986) 193

A L Bilgrami, I Ahmad & M S Jairajpun
to any preference for them as prey or it has resulted as a consequence of a correlation between the populations of prey and predators or due to their widespread occurrence in the soil besides their susceptibility to predation A few species of tylenchs and dorylaims are reported to be resistant to some extent to predation perhaps because of having thick cuticles (Esser, 1963, Small & Grootaert, 1983) The heterogeneity of prey withm the intestine of mononchs suggests that these predators have no choice of prey, their feeding being polyvalent and predation aleatory A similar conclusion was also drawn by Mohandas and Prabhoo (1980) and Small and Grootaert (1983) However, we have found that more predators (75 ' o) had free-living saprophagous nematodes in their intestine rather than tylenchs (45 "o) or dorylaims (42 "o) (Tab 2)
Cannibalism appears to be more of a natural phenomenon than occurnng only under conditions of prey non-availability (Azmi & Jairajpun, 1979, Bilgrami & Jairajpun, 1985) as in most instances the mononchs (prey) occurred together with other types of prey in the intestine of mononchs (predator) All species except Pnonchulus muscorum, Coomansus indicus, lotonchus antedontus, I baqni, I basidontus, I jain, I longicaudatus, I prabhooi, I shafii and all species of Mylonchulus showed cannibalistic tendencies Twenty percent predators having prey in their intestine were cannibals while 27 o devoured mononchs belonging to other species genera also (Tab 2) Most predators devoured a greater number of mononchs of other genera than their own except species of Mononchus and Sporonchulus which showed a reverse trend
Of all the mononchs studied Parahadronchus shakili was the most active predator as 68 "o of us specimens had prey in their intestine while Coomansus indicus was least active with only 21 ' o having prey Eight genera of Tylenchida, six of Dorylaimida, five of Mononchida, three of Rhabditida and one each of Diplogasterida and Monohystenda were identified as prey of Parahadronchus shakili, while Coomansus indicus fed upon three genera of Tylenchida, one Dorylaimida and two each of Mononchida and Rhabditida (Tab 1) Mohandas and Prabhoo (1980) did not find any prey within the intestine of predators belonging to the genus Mylonchulus The same holds true in the present work also The number of prey present in the intestine of a single predator vaned from none to eight
ACKNOVt 1 EDGEMbMS
We thank Prof Athtr H Siddiqi, Chairman, Department of Zoology for providing laboratory facilities
RFFhRPNCtS
ARI'IN, P (1979) Ecologie et systematique des nematodes Mononchidcs dcs zones forestieres et herbdcees sous climat
Accepte pour publication le 23 fevner 1986
tempere humide I Types de sol et groupements specifi ques Revue Nematol, 2 211221
ARI'I\ , P &K1LBERILS, G (1981) Ultrastructure du contenu digestif et de Tepithelium intestinal chez quelques nemato des predateurs (Mononchides) tt bactenophages Revue Nematol, 4 131-143
AzMl, M I & jAiRAjPbRi, M S (1979) Effect of mechanical sDmulanon and crowding on Mylonchulus deniatus \ematol medit, 7 203-207
BILGRAMI, A L , AHMAD, I & JAIRAJPLRI, M S (1984)
Observations on the predatory behaviour of Mononchus aquaticus Nematol medi , 12 41-45
BILGRAMI, A L & JAIRAJPI RI, M S (1985) Cannibahsm in Mononchus aquaticus Indian J Nematol, 14 (1984) 202-203
COBB, N A (1917) The mononchs {Mononchus, Bastian) a genus of free-living predatory nematodes Soil Sci, 3 431-486
CoH\, E & MoRDECHAi, M (1974) Expenments in suppressing citrus nematode populations by use of mangold and a predacious nematode Nematol medit, 2 43-53
EssER, R P (1963) Nematode interactions in plates of non-stenle water-agar Proc Soil Crop Sci Soc Fla, 23 121-138
JAIRAJPLRI, M S & AZMI M I (1978) Some studies on the predatory behaviour of Mylonchulus dentatus Nematol medit,6 205-212
MAHAPATRA, N K & RAO, Y S (1981) Bionomics of lotonchus monohystera, a predacious nematode in rice soils Proc Indian Acad Parasitol, 2 85-87
MOHANDAS, C & PRABHOO, N R (1980) The feeding behaviour and food preference of predatory nematodes (Mononchida) from the soil of Kerala (India) Revue Ecol Biol Sol, 17 53-60
NELMES, A J & MCCLLLOCH, J S (1975) Number of Pnonchulus punctatus (Cobb), a nematode predator J amm Ecol, 43 553-565
NELMES, A J & M C CLLLOCH, J S (1975) Number of Mononchid nematodes in soil sown to cereals and grasses Annls appl Bwl, 79 231-242
SM^LL, R W (1979) The effects of predatory nematodes on population of plant-parasitic nematodes in pots Nematologica, 25 94 103
SMAIL, R W & GROOTAERT, P (1983) Observations on the predation abilities of some soil dwelling predatory nematodes Nematologica, 29 109-118
STEINER, G & HI INLY, H (1922) The possibility of control of Heterodera radicicola and other plant injunous nemas by means of predator/ nemas specially by Mononchus papillatus Basuan J Wash Acad Sci, 12 367-385
THORNE, G (1927) The life history, habits and economic importance of some mononchs J agnc Res, 34 265-286
194 Revue Nematol, 9 (2) 191-194 (1986)

89
Indian J. Nematol. 16(1), (1986) : 60-64
EFFECT OF DIFFERENT PLANTS, SINGLY AND IN COMBINATIONS, ON THE ATTRACTION OF ADULT XIPHINEMA BASIRI
ANWAR L. BILGRAMI, M. SHAMIM JAIRAJPURI and IRFAN AHMAD
Section of Nematology, Department of Zoology, Aligarh Muslim University, Aligarh-202 001, India
Abstract : Observations on the effect of different plants, singly and in combinations of two plants, were made on the attraction of adult Xiphinema basiri in agar plates. Cabbage and tomato were most attractive, carrot and brinjal moderately attractive, chilli and radish least attractive while onion and mustard failed to attract nematodes significantly when tested alone, OT together in one combination. The differential attraction behaviour of X. basiri is attributed to the plant root attractants and the intensity of the attractants.
Key words : Xiphinema basiri, behaviour, attraction
The attraction and feeding of plant-parasitic nematodes has been studied by many workers (Wyss, 1971a, 1971b; Doncaster & Seymour, 1973; Azmi & Jairajpuri, 1977; Prot, 1980). Biigrami era/. (1985b) observed the influence of single plant and combinations of two plant roots on the attraction of rice root nematode, Hirschmanniella oryzae. The feeding of Xiphinema basiri on tomato roots was studied by Bajaj & Jairajpuri (1977) while Malik (1985) reported its attraction towards several plant roots. X. basiri is widely distributed (Yadav & Verma, 1967; Roy, 1973) and is suspected to bs the vector of Cowpea Mosaic Virus (cf. Caveness e/ al., 1975). The present investigations were made on its attraction towards different plant roots, singly and in combinations of two plants.
MATERIALS AND METHODS
The nematodes were extracted afresh for each experiment from soil around the roots of mulberry (Moras indica) by modified Baermann's funnel method and kept in tap-water before use. Experiments were carried out in 7 cm diameter Petri dishes, containing a 5 mm thick layer of 0.5 per cent autoclaved (15 lb for 30 min)
water-agar. The attraction was tested towards 4-6 day old seedlings at a temperature of 28''C. The plants were grown in 1 per cent water agar at the same temperature and sterilized with 0.1 per cent mercuric chloride solution prior to the experiment. Experiments were replicated 5 times. Only adult X. basiri were used. The attraction was tested towards eight plants viz., cabbage {Brassica oleracea), radish {Raphanus sativus); carrot {Daucus carota); onion {Allium sepa); tomato {Lycopersicon esculentum); brinjal {Solarium melongena); mustard (Brassica com-pestn's) and chilli (Capsicum annuum)
Attraction of X. basiri towards single plant : Petri dishes were marked at bottom into 7 zones, 1 cm wide, and numbered serialy 1-7 (Biigrami et al, 1985b). Plants (one plant per dish) were embedded in zone 1 (root parallel to the long axis of zone while shoot remained outside) and left for 12 h. Water, if present around roots, was soaked out with a piece of filter paper to avoid formation of water film; 25 nematodes were then placed in zone 4 and their distribution was recorded after 12 h. Controls were run without seedlings and nematodes were tested separately towards each plant.

so
61 EFFECT OF DIFFERENT PLANTS SINGLY AND IN COMBINATIONS
Attraction of X. basiri towards combinations of two plants placed at a distance from each other: All conditions were same as above except that the nematodes were tested towards combinations of two plants, placed at opposite ends, one in zone I and other in zone 7.
All 28 possible combinations were studied. In figures, the total number of nematodes present in zones 1, 2 and 3, and 5, 6 and 7 have been plotted.
A ttraction of X. basiri towards combinations of two plants placed together : In this experiment also, all conditions were same as in previous experiments except that two plants were placed together in zone I and not separately. All 28 possible combinations were tested separately.
RESULTS
Attraction of X. basiri towards single plant: X. basiri showed maximum response towards cabbage and tomato (p < 0.05, Fig. 1, B & F). Carrot and brinjal attracted moderate number of nematodes (Fig. I, C & I ) . Radish and chilli plants were least attractive to the nematodes (Fig. 1, D & H) while onion
and mustard seedlings were not attractive to X basiri (p > 0 05, Fig. 1, E & G).
Attraction of X- basiri towards combinations of two plants placed at a distance fram each other : In the presence of cabbage, the attraction towards carrot, tomato and brinjal was similar (p > 0.05) but declined towards radish (p < 0.05, Fig. 2, A). No attraction vt/as observed towards onion, mustard and chilli (p > 0.05) when tested either with cabbage or carrot (Fig. 2, A & B). In presence of carrot, tomato and brinjal were more attractive (p < 0.05) but radish attracted lesser number of nematodes (Fig. 2, B). Onion in all combinations failed to attract nematodes significantly (p > 0.05, Fig. 2, C). The maximum attraction was towards tomato while lesser towards radish, chilli and brinjal when tested with onion (Fig. 2, C). Nematodes responded more towards tomato and brinjal in presence of radish (p < 0.05, Fig. 2, D) but the latter was more attractive when tested with chilli (p < 0.05, Fig. 2, D). Mustard also failed to attract nematodes (p > 0.05) in presence of onion, radish, tomato and brinjal (Fig. 2, C, D, E & F). Tomato attracted more nematodes when tested either with chilli or brinjal (Fig. 2, E).
A B C D E F G H l
Plant roots - '
Fig. 1. Attraction of X. basiri towards single plant. Striped bars = total number of nematodes in zones 1, 2 and 3; Plain bars = total number of nematodes in zones 5, 6 and 7. A —without roots; B —cabbage; C —carrot; D —radish; E —onion; F —tomato; G - - mustard; H — Chilli; I — brinjal.

91
ANWAR L. BILGRAMI et ol. 62
In mustard-chilJ] combination, both plants failed to attract nematodes (Fig. 2, F) but brinjal was attractive in presence of mustard (p < 0.05, Fig. 2, F). In combination of chilli and brinjal. only the latter attracted nematodes (p < 0.05) while the former failed to attract them (Fig. 2, G).
Attraction of X. basin towards combinations of two plants placed together : Cabbage and tomato together were most attractive (p < 0 05, Fig. 3, E). Carrot, radish, brinjal and chilli combinations with cabbage were attractive to lesser degree (p <0.05, Fig. 3. A, B, F & G). The attraction declined further towards combinations containing either onion or mustard plants (p < 0.05, Fig. 3, C& D). X. basni showed lesser degree of attraction towards all combinations with onion (p < 0.05, Fig. 3, H, O, P, R & S) except with mustard where they did not respond
(p 0.05, Fig. 3, Q). Carrot and tomato together attracted more nematodes (Fig. 3, J) the carrot combinations with radish, mustard, chilli and brinjal (Fig. 3,1, K, L & M). Tomato and brinjal when tested together attracted more nematodes (Fig. 3, X) than tomato-mustard and tomato-chilli (p < 0.05, Fig. 3, V & W). Attraction of X. basiri vi'as more towards radish-tomato and radish-brinjal (Fig. 3, Y & Z:) but declined towards radish-mustard and radish-chilli combinations (Fig. 3, Z & Zi). The nematodes did not respond towards mustard-chilli combination (p > 0.05, Fig. 3, T) but they were attracted by the (mustard-brinjal Fig. 3, U) and chilli-brinjal combinations (Fig. 3, N).
DISCUSSION
Malik (1985) found chilli most attractive, tomato and brinjal moderately
, i 6 Z [ I
IlliU Mustard
1.1 Tomoto
i
E •
Comblnotlons of plant roots
Fig. 2. Attraction of X. basiri towards combinations of two plants placed at opposite ends/.e., zone 1 and 7. A — combinations of cabbage; B — carrot; C — onion; D —radish; E — tomato; F — mustard; G — chilli.

Q2
63 EFFECT OF DIFl EREXT PLANTS SINGLY AND IN COMBINATIONS
B C 0 E F G H I M N
0 P a - R 5 T U V W X V Z Z , 22
Combinations of plant roots
Fig. 3. Attraction of X. basiri towards combinations of two plants placed together (zone 1). Stripped bars = total number of nematodes in zones 1, 2 and 3; Plain bars = total number of nematodes in zones 5, 6 and 7. Combinations of cabbage with, A — carrot, B — radish, C ~ onion, D — mustard, E — tomasto, F —chilli, G — brinjal; combinations of carrot with, H ~ onion, I — radish, J — tomato, K — mustard, L — chilli, M—brinjal; combinations of chilli with N — brinjal; combinations of onion with, O — radish, P — tomato, Q — mustard, R — chilli, S — brinjal; combitutions of mustard with, T — chilH, U — brinjal; combinations of tomato with, V — mustard, W — chilli, X —brinjal; combinations of radish with, Y —tomato, Z - mustard, Zi —chilli, Zj —brinjal.
attractive and radish not attractive to X. basiri. Our observations show that tomato was most attractive, brinjal moderately and chilli and radish least. These differences may have resulted from different experimental techniques as some oi the experimental factors are known to influence attraction of plant-parasitic nematodes (Bilgrami et at., 1985a).
Khera & Zuckerman (1963) and Grifiin (1969) respectively found Hemi-cycliophora similis and Meloidogyne hapla juveniles equally responsive towards a host and non-host seedlings and towards
resistant and susceptible cuitivars of alfalfa (tested separately). Prot (1980) also considered attraction in plant parasitic nematodes to be non-specific. Our observations also indicate that attraction was non-specific but X. basiri showed some degree of preference similar to H. oryzae (Bilgrami et al., 1985b) and M. hapla juveniles for susceptible cultivar when tested simultaneously with a resistant one (Griffin, 1969). The preference of cabbage and tomato in presence of chilli, radish, carrot and brinjal and towards the letter two plants in presence of chilli and radish in different combinations

93
AMWAR L BILGRAMI et ul 64
reflects an idea of dissimilar attractants emitted by different plant roots (Prot, 1980)
The rndximum response towards cabbage and tomato, when tested either alone, separately or together in one combination, may be related to the intensity of the stimuli, as strong stimuli produce a vigrous response (Doncaster & Seymour, 1973) The decrease in the attraction towards carrot-brinjal and radish-chiUi combinations also suggests that their attractants were of moderate and week intensities respectively The attractiveness of chilli alone and Us un-attractiveness in different combinations except with those plants which were either equally attractive oi non-attractive also supports that its attractants were of week intensities and thus failed to attract nematodes in the presence of plants having attractants of higher intensities
The unresponsiveness towards onion and mustard, tested alone, separately or together in one combination may either be due to the failure of attractants in producing a perceptible gradient oi that the exudates were not attractive at ail However, a decrease in attraction towards some combinations containing these two plants and total inhibition of response towards mustard-chilli combination may perhaps be due to a neutralizing effect of root exudates of onion or mustard exerted on exudates of other plants
ACKNOWLEDGEMENT
We thank the Chairman, Department of Zoology for providing necessary laboratory facilities. The first author also acknowledges the financial assistance given by ICAR, New Delhi under All India Coordinate Research Project on nematode pests of crops and their contiol
REFERENCES
AzMi, M I & JAIRAJPUM, M. S (1977) Attraction of plant parasitic nematodes to host roots Nematohgtca 23 119-121
BAJAJ, H K &JAiRiijpuRi MS (1977; Studies on nematode behaviour X Observations on ktdin% of Xiphinema basin on tomato Indian J Nematol 7 01-93
BiLGRAMi, A L , AHMAD, I & JAIRAJPLRI, M S (1985a). Factors influencing attraction of adult Hirschmanmella oryzae towards cabbage seedlings Revue Nematol (in press)
BaoRAMi A L , AHMAD, 1 & JAIRAJPURI, M S (1985b) Responses of adult Hirschmanmella oryzae towards different plant roots Revue Nematol (in press)
CAVENESS, F F , GIEVIER, R M & WILLIAMS, R J (1975) Transmission of cowpea mosaic by Xtphinema basin m Western Nigeria, In nematode vector of plant virus Eds F Laraberti, C E Taylor & J W Semhorst Plenum Press, London 289-290
DONCASTER, CC & SEYMOUR, M K (1973) Exploration and selection of penetration site by tylenchids Nematologica 19 137-145
GRIFFIN, G D (1969) Attractiveness of resistant and susceptible alfalfa to stem and root knot nematodes J Nematol 1 9 (Abstr)
KHERA S & ZLCKERMAN, B M (1963) In vitro studies of host-parasite relationships of some plant parasitic nematodes Nematologica 9 1-6
MALIK Z (1985) Attraction of Xiphmema basin towards different hosts Indian J Nematol. 14 . 199-200.
PROT, J C (1980) Migration of plant parasitic nematodes towards plant roots Revue Nematol 3 305-318
ROY, TK (1973) Notes on the pathogenecity of Xiphmema basin and its host records Indian J Nematol 3 161-163
WYSS, U (1971a) Saugverhalten and pathogeni-tat von Tnchodorus spp in steriler agarkul-ture Nematologica 17 501-507.
WYSS, U (1971b) Der mechanismus der nahrungsaufnahme bei Tnchodorus similis Nematologica 17 : 508-518.
YADAV, BS &VERM\, M K (1067). New host plants of A'i/)/ii/iema 6a«n and X indicum. Nematologica 13 469
Accepted for publication July 1985

94
Evaluation of the predatory behaviour of Dorylaimus stagnalis T>u]dix6m, 1845 (Nematoda : Dorylaimida)
Samina SHAFQAT, Anwar L. BILGRAMI and M. Shamim JAIRAJPURI
Section of Nematology, Department of Zoology, Aligarh Muslim University, Aligarh-202001, India
SUMMARY
Observations on the predatory behaviour of Dorylaimus stagnahs Dujardin, 1845 were made in agar-plates Predation was on chance encounters Predators were not attracted towards hve (non-excised) or excised prty D stagnahs probed over a short distance along any part of the body of prey and paralysed it by disorganizing its internal body organs with the help of their odontostyle Ingestion of prey contents was intermittent and duration of feeding lasted for 85 mn (25-180 mn) on Rhabditis sp , and 70 mn (15-150 mn) on Hirschmanniella oryzae Rhabditis sp was prefered most while Acrobeloides sp , Tylenchorhynchus mashhoodi and Hehcotylenchus indicus the least No specimen of Hoplolaimus indicus was either injured or killed by the predators There was no difference in the rate of predation over a period which lasted for ten days Adults (male and fema e) preyed more than their younger stages All factors viz , prey number, temperature and agar concentration influenced predation by D stagnahs except the starvation of predators Maximum predation took plate when predators were placed in a population of 125 individuals of prey Agar concentrations 1, 2 and 3 " and temperatures 25 and 30 were most suitable for predation by D stagnahs
RESLME
Evaluation du uomportement predateur de Dorylaimus stagnahs Dujardin, 1845 (Nematoda Dorylaimida)
Des observations sur le comportement predateur de Dorylaimus stagnahs Dujardm, 1845 ont ete faites en utilisant des boites de Petn contenant de I'agar-agar La predation est en relation avec des contacts dus au hasard, le predateur n'est en effet pas attire par les proies, que celles-ci soient intactes ou incisees D stagnahs se nourrit sur une longueur resiremte de n'lmporte quelle partie du corps de la proie et paralyse celle-ci en desorganisant ses organes internes par action du stylet L'lngestion du contenu de la proie est intermittente et la prise de nournture dure 85 mn (20-180) avec Rhabditis et 70 mn (15-150) avec Hirschmanniella oryzae Rhabditis est une proie preferee tandis que Tylenchorhynchus mashhoodi et Hehcotylenchus mdicus sont les proies les moins attaquees Aucun individu de Hoplolaimus indicus n'a ete blesse ou tue par le predateur II n'est apparu, au cours d'une penode de dix jours, aucune difference dans le taux de predation Les adultes, males ou femelles, ont un comportement predateur plus actif que les juveniles Tous les facteurs etudies (nombre de proie, temperature, concentration en agar-agar) influencent la predation de D stagnahs, a I'exception du jeiine du predateur La predation maximale a lieu lorsque les predateurs sont places parmi une population de 125 proies Les concentrations de 1, 2 et 3 "o en agar-agar et les temperatures de 25 et 30 sont les plus favorablcs a la predation par D stagnahs
Thome (1930; 1939) had suggested that species of many dorylaims and nygolaims may be predatory in habit. Since then species of Dorylaimus, Labronema, Discolaimus, Carcharolaimus and Actinolaimus have been reported to be predacious (Lmford & Oliviera, 1937; Esser, 1963) Detailed account on the feeding mechanism and predation abihty of a dorylaim predator, Labronema vulvapapiUatum Loof & Grootaert, 1981 was given by Wyss and Grootaert (1977) and Small and Grootaert (1983). Very recently, Bilgrami, Ahmad and Jairajpuri (1985a) for the first time made detailed observations on the nygolaim predator, Aquatides thor-nei (Schneider, 1937) Ahmad & Jairajpun, 1979 The dorylaim predators derive their food also from algae and fungi besides feeding on nematodes and hence are
Revue Nematol 10 I4j 455-461 I987i
considered omnivorous (HoUis, 1957; Ferris, 1968, Wood, 1973).
In the present work a comprehensive study on the predatory behaviour of Dorylaimus stagnahs Dujardin, 1845 was made to assess it^ predatory abihties and to determine the factors affecting predation.
Materials and methods
Soil samples containing Dorylaimus stagnahs, collected from paddy fields near Jamalpur, Aligarh, were processed by sieving, decantation and Baermann's funnel methods D stagnahs was cultured in 1 "o water-agar at 25 ± 1 using Rhabditis sp. as prey Five milligram of infant milk powder (Lactogen) was spread over the
455

s S Shafqat, A L Btlgrami & M S Jairajpun
surface of the agar to allow bacteria to grow This served as the food of prey nematodes Predators were transferred to fresh culture dishes containing prey after every 10-15 days Rhabditis sp , Acrobeloides sp and Cephalo-bus sp were cultured in agar plates by spreading the same amount of Lactogen over the surface of agar Plant-parasitic prey species Hoplolaimus indicus, Helico-tylenchus indicus, Hirschmanniella oryzae and Tylen-chorhynchus mashhoodi were isolated fresh from the soil
FEEDING MECHANISM
Feeding in D stagnahs was studied by observing inverted Petri-dishes containing predators and prey under microscope at 66 x magnification For higher magnifications 450 x or more, simple observation chambers consisting of a glass slide with a block of 1 " o water-agar measuring 2 x 1 5 x 0 5 cm was used (Wyss & Grootaert, 1977, Bilgrami, Ahmad & Jairajpun, 1985 a) Ten aduh (male and female) specimens of D stagnahs were released together with 100 to 150 prey nematodes m the agar block which was gently covered by a coverslip and kept at 25 ± 1 Observations were started after one hour Feeding of D stagnahs was observed on Rhabditis sp , and H oryzae
pieces) and excised H oryzae Control was run without prey nematodes
RATF OF PREDATION OF ADLLTS AND JLVENILES
Rate of predation of male, female and the four juvenile stages (differentiated on the basis of their odonto-style length and body length) of D stagnahs was determined by releasing five specimens of each stage with twentyfive prey nematodes in cavity-blocks containing 1 "o water-agar Predation by each stage was tested separately upon Rhabditis sp and H oryzae Number of prey killed or ingested was counted after 24 h Experiments were replicated five times and carried out at 25 ± 1
P R E D A I O R Y P A n t R N
To observe predatory pattern over a period of ten days, five female predators were released in cavity blocks containing 1 "o water-agar with twentyfive prey nematodes at 25 ± 1 Observations were made at 24 h intervals The experiment was replicated five times Each day after observations the predators were transfered to fresh cavity blocks containing prey of the same type Rhabditis sp and H oryzae was used separately as prey
P R F Y I ' R F F F R E N ( I
To determine whether D stagnahs has any preference for prey Rhabditis sp , Acrobeloides sp, Cephalobus sp , Hoplolaimus mdicus, Helicotylenchus indicus, Hirschmanniella oryzae and Tylenchorhynchus mashhoodi were subjected to predation Five predators were placed with 25 prey nematodes in cavity-blocks containing 1 °o water-agar The number of prey killed or ingested was recorded after 24 h Each type of prey was tested separately at 25 ± 1 Expenments were replicated five times
A I I R. C I ION I im \RDS P R E \
Attraction of D stagnahs towards prey was studied by the methods of Azmi and Jairajpun (1977) and Bilgrami, Ahmad and Jairajpun (19856) Seven zones, each of km width, were marked at bottom of 7 cm diameter Petri-dish A plastic straw pipe 5 mm high and 5 mm in diameter, one end sealed with filter paper, was placed vertically m zone 1 (sealed end remained in contact with the surface of the Petn-dish) Straw pipe and Petn-dish were then filled with 1 water-agar (0 5 cm thick layer) Twentyfive prey nematodes were placed in straw pipe and whole set was left at 25 ± 1 for 24 h Twentyfive female predators were then released at different places in zone 4 Number of predators present in each zone was counted after four hours Attraction was tested separately towards Rhabditis sp , live and excised (cut in two
FACTORS INFLLENC ING PRE DA HON BY D srAC,\AnS
Observations on the effect of different factors viz , prey number, temperature, agar concentration and starvation of predators (3n the rate of predation were made Five female D stagnahs were placed with twentyfive specimens of prey nematodes in small cavity blocks containing 1 'o water-agar at 25 ± 1 Number of prey killed or ingested was counted after 24 h In all the experiments Rhabditis sp and H oryzae were used separately as prty Each experiment was repeated five times The above conditions remained same during all experiments unless mentioned otherwise Effect of prey number on predation was observed by placing predators with 25, 50, 75, 100, 125, 150, 175 and 200 prey nematodes in separate cavity blocks To observe the effect of different temperatures, prey nematodes were subjected to predation at 5,10,15,20,25, 30,35 and 40 To determine the effect of starvation, the predators were kept without prey for 0, 2, 4, 6, 8, 10, 12 and 14 days Predators of each group were tested in separate cavity blocks Six concentrations of water-agar viz , 1, 2, 3, 4, 5 and 6 "o were made to study their effect on the predation of D stagnahs
Results and discussion
F E F D I N G MECHANISM
Observations show that D stagnahs feeds by predation Predation depended on chance encounters with the
456 Revue Nematol 10 (4, 455-461 11987

Predatory behaviour o/Dorylaimus stagnalis
c
prey (Nelmes, 1974, Grootaert & Maertens, 1976, Wyss & Grootaert, 1977; Bilgrami, Ahmad & Jairajpun, 1984, 1985 a). D stagnalis attacked prey only if it made lip contact and there was no preference shown for attacking any particular part of the body of prey. Probing before attack was reported by Wyss and Grootaert (1977) and Bilgrami, Ahmad and Jaira)puri (1985a) in Labronema vulvapapillatum and Aquatides thomei respectively. D stagnalis also probed any part of the body of prey over a short distance before thrusting its stylet so as to puncture the cuticle of the prey. Dunng probing a few weak oesophageal pulsations were also observed in predators. Probing by D. stagnalis was, however, not as vigorous as that of L vulvapapillatum (Wyss & Grootaert, 1977) Stylet thrusting was also not of the same intensity with which L vulvapapillatum aimed to perforate the cuncle of its prey It was more like that of A thomei where gradual and intermittent stylet thrusting was observed (Bilgrami, Ahmad & Jairajpun, 1985 a). Normally six to eight stylet thrusts were enough to puncture the cuticle but some times the prey escaped from the grip of the predator if the later failed to penetrate its odontostyle into the body of prey.
Aphelenchid predators (Seinura spp.) are known to paralyse their prey by injecting salivary and eosophageal secretions into the prey (Linford & Ohviera, 1937; Hechler, 1963). D stagnalis also paralysed its prey probably by disorganizing internal body parts of the prey with Its odontostyle similar to other dorylaim predators (Holhs, 1957; Wyss & Grootaert, 1977; Bilgrami, Ahmad & Jairajpun, 1985 a) However, if predators injected secretions into the prey, these might have been used for extra-corporeal digestion After penetration, thrusting and side ways movements of the odontostyle began and during ingestion odontostyle remained within the body of the prey The ingestion was intermittent involving short periods of sucking activity. The body contents of prey could be seen passing through the lumen of odontostyle and oesophagus of the predators D stagnalis explored and fed at different spots of the same prey to ingest all the body contents Only the cuticle was not consumed when the feeding was completed The rest of the sequenses of prey catching and feeding mechanisms did not differ much from those reported by Wyss and Grootaert (1977) and Bilgrami, Ahmad and Jairajpun (1985a) for Labronema vulvapapillatum and A thomei The rate of predation by D stagnalis was, however^ more than the above two predators This differential rate of predation could be due to different nematodes used as prey for the above predators The duration of feeding on Rhabditis sp (85 mn . 25-180 mn) was slightly more than that on H oryzae (70 mn : 15-150 mn)
Often two to four predators were seen feeding together on a single prey which was injured or killed earlier by another predator D stagnalis readily fed on the eggs of Rhabditis sp it had contacted but not on its own eggs
They rubbed them with lips few times before moving away. In the presence of eggs of free-living nematodes, affinity of D stagnalis (both male and female) for their own eggs suggests some form of recognition of their own chemical attractants (Esser, 1963; Bilgrami, Ahmad & Jairajpun, 1985 a)
P k t Y PREFERLNCE (Fig. 1)
D stagnalis showed some degree of preference. Rhabditis sp. were killed in greater numbers (p < 0.05), while Acrobeloides sp., T mashhoodi and Hehcotylenchus indi-
U
10_
o Z
5_,
2_
A B
prey
Fig 1 Prey preference in D stagnalis A Rhabditis sp , B Hirschmanmella oryzae, C Cephalohui sp , D Hehcotylenchus indicus, E Acrobeloides sp , F Tylenchorhynchus mashhoodi, G Hoplolaimus mdicus
cus were prefered least. Other prey nematodes viz , Hirschmanmella oryzae and Cephalobus sp. were killed in moderate numbers. No specimen of Hoplolaimus indicus was either injured or killed by the predators. This disparity in the rate of predation by D stagnalis upon different nematodes that were used as prey may be attributed to the size and behaviour of prey and the texture of their cuticle. Hoplolaimus indicus and Hehcotylenchus tndtcus seemed to resist predation probably due to the toughness and thickness of their cuticle and secretion of toxic chemicals as was suggested by Esser (1963) for other species of Hoplolaimus and Hehcotylenchus respectively
A riR. \C I ION TOWARDS PREY (Fig 2)
The predacious nematodes were unable to detect their prey even from short distances (Esser, 1963). This
Revue Nematol 10 14 45'i-461 11987) 457

97 S. Shafqat, A. L. Bilgrami & M. S. Jairajpun
o r) h 01 c_
'o o Z
8 -
6 .
;_
2_
A
2 3 4 5 6 7
Zones
in at x> o o E c
o z
8 "
6 -
4,
2_
B
1 2 3 ; 5 6 7
Zones
01
O
O E 01
o Z
8 "
6 _
?
c
3
V)
8 "
6 -
2_
D
1 3 4
Zones
Fig. 2. Attraction of D. si oryzae (excised).
Zones
w towards A : without prey; B : Rhabditissp. (whole); C : Rhabditissp. (excised); D : Htrschmanmella
opinion was supported by Grootaert and Maertens (1976); Wyss and Grootaert (1977) and Bilgrami, Ahmad and Jairajpuri (1984; 1985a) on their studies on Mononchus aquaticus, L. vulvapapillatum and A. thor-nei. Esser (1963) was also of the opinion that live prey cut in two halves attracted several dorylaim predators. During present observations on D. stagnalis such a phenomenon was not observed either towards excised Rhabditis sp. or H. oryzae as the number of predators present in zones 1, 2 and 3 in the presence and absence
of prey were not significantly different (p > 0.05). However, aggregation of D. stagnalis around an injured prey suggests some kind of attraction (Wyss & Grootaert, 1977; Bilgrami, Ahmad & Jairajpuri, 1985 a).
RATE OF PREUAT ION OF ADULT S AND JUVENILES
(Fig. 3)
All except the first stage juveniles preyed upon Rhabditis sp. and H. oryzae. First stage juveniles did not
458 Revue Nematol. 10 (4/ : 455-461 '1987,

Predatory behaviour o/Dorylaimus stagnalis
9 8
10-
6_
2_
• R hobditis sp
H orvzQg
i I- g _l I A B C D E F
Fig 3 Rate of predation of adult and juvenile stages A Female, B Male, C Fourth stage, D Third stage, E Second stage, F First stage juveniles
kill or injure any prey but fed on prey killed by elder stages Adults (male and female), fourth and third stage juveniles killed more Rhabditis sp than H oryzae Second stage juveniles preyed equally upon both the pre> There was no significant difference in the number of prey killed by male and female D stagnalis (p > 0 05) Fourth and third stage juveniles killed lesser number of Rhabditis sp and H oryzae than their adults Differences in the rate of predation by various stages of D stagnalis reflect their predatory potentials Adults being stronger predators than their younger stages
PRED'XIORV PATTERN (Fig 4)
D stagnalis showed little variation in its predation on Rhabditis sp and H oryzae over a period lasting ten days Number of prey killed by predators after 2nd and 10th day was not significantly different (p > 0 05) Mononchus aquaticus aho showed a consistent predatory pattern over the same period (Bilgrami, Ahmad & Jairajpuri, 1984)
FAC lORS I \ H L h \ ( I\C, PRED\ iK) \ B\ D SMC,Mils
(Fig 5)
The increase in prey number increased the predation by Diplenteron potohikus, Pnonchulus punctatus and A thornei (Yeates, 1969, Nelmes, 1974, Bilgrami, Ahmad & Jairajpun, 1985a) but predation by Mononchus aquaticus remained unchanged in varying prey populations (Bilgrami, Ahmad & Jairajpun, 1984) Obser-vanons on D stagnalis revealed that predauon increased significantly upto a population of 125 prey nematodes (p < 0 05, Fig 5, A) Prey number more than this did not affect predation and the rate of killing per day was same (p > 0 05) This suggests a certain ration between predators and prey for maximum predation at which / )
U
•a ^ 10.
k-a. o 6.
2 .
• • •
^ O O
0 0 Rhabditis sp.
O — O H.orvzge
10
Days
Fig 4 Predatory pattern in D stagnalis
stagnalis might have been satiai ed and did not attack or kill any prey thereafter The increase in predation was perhaps the outcome of improved chances of encounters between predators and prey vi'hich, however, did not alter predation beyond an optimum level of prey population
Predation was also influenced by different temperatures Maximum number of Rhabditis sp and H oryzae were killed at 25 and 30 (Fig 5,B) Temperatures lower (5-15 ) and higher (40 ) declined predation markedly (p < 0 05) At 20 and 35 moderate number of Rhabditis sp and H oryzae were killed At different temperatures the rate of predation by D stagnalis may be different because of the activity of predators and prey which might have been influenced b> the change in temperature (Bilgrami, Ahmad & Jairajpun, 1983)
The starvation of predators was the only factor which did not alter the number of prey killed per day by D stagnalis (Fig 5, C) Differences in predation after 0 and 14 days were insignificant (p > 0 05) Bilgrami, Ahmad and Jairajpun (1984) also found similar results in M aquaticus
The rate of predation b> D stagnalis was also altered by the change in agar concentrations as was also observed by Bilgrami, Ahmad and Jairajpun (1983) in M aquaticus Maximum killing was recorded in 1, 2 and 3 "() water-agar (p < 0 05, Fig 5, D) Concentrations higher than these declined the rate of feeding significantly (p < 0 05) In 6 ' 0 no specimen of H oryzae was cither killed or ingested but predators succeeded in kilhng d few Rhabditis sp at th s concentration It seems that similar to temperature, agar concentrations also governed the activity of prey and predators which affected the rate of predation by D stagnalis
Revue i\ematol 10 4 455-461 1987/ 459

99 S Shafqal, A L Bilgrami & M S Jairajpun
Ik
20_
T3 Of
= 16.
°. 12. o
8 .
18"
-1 1 1 1 1 1 1 1 25 50 75 100 125 150 175 200
Prey number
•? 12.
• — • Rhabditis sp.
O—O H.oryzae B
1 1 1 \ 1 1—I r
5 10 15 20 25 30 35 AG
Temperature (°C)
18'
12_
o 5. d
. - . / — ^ . / • ^ . - .
O O -°\„/ o^\
n — I — I 10 12 U
Starvation period (days) Agar concentration(V.)
Fig. 5. Factors influL-ncing predation b> D stagnalis A : Prey number, B Temperature, C . Starvation of predators; D : Agar concentration.
A( KNOW. I tD(,L.Mr\ I S
We thank the Chairman, Department of Zoology, Aligarh Mushm University, Ahgarh for providing necessary laboratory facilities.
RtFhRLNCFS
A/Ml, M. I. & JAIKAJPLRI, M . S. (1977). Attraction of plant-parasitic nematodes to host roots. Nematologica, 23 : 119-121.
460 Revue NematoL 10 (4j . 455-461 '19871

Predatory behaviour o/Dorylaimus stagnalis
BiLGR^^Mi, A. L., AHMAD, 1. & JAIRAJPLRI, M . S. (198^). Some factors influencing prcdation by Mononchus aquaticus. Revue Ne'matoL, 6 : 525-326.
BiLc,R.\Mi, A. L., AHMAD, I. & JAIR/\JFI RI, M . S. (1984).
Observations on the predatory behaviour of Mononchus aquaticus. Nematol medtt, 12 • 41-45.
B11GR.AMI, A L., AHMAD, I & J.\IR.AJPI KI, M S. (1985a)
Predatory behaviour of Aqualides thornei (Nygolaimina Xematoda). Nematologita, 30 (1984) : 457-462.
BiiGR^wii, A. L., A H V U D , I & JAIR,A]PIRI, M . S. (19856).
Responses of adult Hmchmanniella oryzae towards different plant roots Revue \ematoL, 8 ; 265-272.
Eisi-R, R. P. (1963). Nematode interactions in plates of non-sterile water-agar. Proc. sod Crop Sci. Sac. Ha., 23 : 121-nH.
FbRRis, V. R. (1968). Biometric analysis in the genus Labro-nema (Nematode : Dor>'laimida) with a description of /,. thornet n. sp. Nematologica, 14 • 276-284.
G R O O I \ F R I , P . & M A I R U N S , D . (1976). Cultivation and hfe
cycle of Mononchus aquatKus Nematologica, 22 : 173-181.
H K H U R , H C (1963\ Description, developmental biology and feeding habits of Setnura tenmcaudam (de Man) J. B. Goodey, 1960 (Nematoda : Aphelenchoididac), a nematode predator. Proc helminth. Soc. Wash., 30 : 182-195.
Hot u s , J. P. (1957). Cultural studies with Dorylaimus etters-bergensis Phytopathology, 47 : 468-473.
LiNFORD, M. B. & OiiVEiRA, J. M. (1937). The feeding of hollow spear nematodes on other nematodes. Science, 85 : 295-297.
N L I Ml s, A. J. (1974). Evaluation of the feeding behaviour of Pnonchulus punctalus{Cobh) a nematode predator. J anim. Ecol, 43 : 553-565.
SMAI 1, R. W. & G R O O I \ I R I , P. (1983). Observations on the
predation abilities of some soil dwelling predatory nematodes. Nematologica, 29 : 109-118.
T H O R M , G. (1930). Predacious nemas of the genus Nygolai-mus and a new genus Seaonema. J agnc. Res., 41 : 445-446.
T H O R M - , G . (1939). A monograph of the nematodes of the superfamily Dorylaimoidea. Capita ZooL, 8 : 1-261.
W O O D , F . H . (1973). Feeding relationships of soil-dwelling nematodes. Sod Biol. Biochem., 5 : 593-601.
WYSS, U . & G R O O I A I R I , P. (1977). Feeding mechanism of Labronema vulvapapillatum. Med. Fac. Landbouw Rijks-umv Gent., 42 : 1521-1527.
Yl-AiPS, G. W. (1969). Predation by Mononchoides potohikus (Nematoda : Diplogastendae) in laboratory culture. Nematologica, 15 • 1-9.
Accepte pour publication le l> decembre 1986.
Revue Nematol. JO 14/: 455-4t>l "1987/ 461

CI
Attraction of Mononchoides longicaudatus and M. fortidens (Nematoda : Diplogasterida) towards prey and factors
influencing attraction Anwar L. BILGRAMI and M. Shamim JAIRAJPURI
Section of Nematology, Department of Zoology, Aligarh Muslim University, Aligarh-202001, India
SUMMARY
The attraction oi Mononchoides longicaudatus and M fortidens tov/ards prey and factors influencing their attraction were studied on agar plates. These predators were attracted towards exased and non-excised (live) prey in response to prey secretions, attractants. Predators also responded positively towards agar containing unidentified bactena. M longicaudatus and M fortidens showed preference for the prey nematodes which were tested either alive or by cutting into two pieces (excised). Hirschmanniella oryzae, Tylenchorhynchus mashhoodi and Hoplolaimus indicus attracted predators most while Longidorus sp , and Hehcotylenchus mdicus elicited least response from the predators. The rest of the prey nematodes attracted predators moderately. Both species of predators were also attracted towards their own individuals Different factors viz., prey number, temperature, incubation penod of prey, starvation of predators and distance of predators from prey affected attraction of the predators towards Hirschmanniella oryzae and the second stage juveniles of Meloidogyne incognita Both predators were attracted m maximum numbers when 50 prey nematodes were incubated for 12 h in Petn-dishes, at 25-30 . Starvation of predators increased their attraction towards prey; increase in prey distance decreased attraction
RESLME
L'attraction de Mononchoides longicaudatus et de M. fortidens 'Nematoda et les facteurs qui I'lnfluencent
Diplogastenda) par les proies,
L'attraction de Mononchoides longicaudatus et de M fortidens par les proies et les facteurs qui I'lnfiuencent ont ete etudies en boites de Petn Ces predateurs sont attires par les proies, dissequees et non-dissequees (vivantes), en reponse a des substances attractives secretees par ces proies. Les predateurs repondent egalement positivement a la presence sur I'agar d'une bactene non identifiee. M longicaudatus et M fortidens montrent une preference parmi les proies, que celles-ci aient ete testees vivantes ou coupees en deux : Hirschmanniella oryzae, Tylenchorhynchus mashhoodi et Hoplolaimus indicus ont la meiUeure attraction tandis que Longidorus sp. et Hehcotylenchus indicus provoquent la reponse la plus faible. Les autres especes testees n'attirent les predateurs que moderement L'une et I'autre especes sont egalement attirees par des individus appartenant a leur propre espece. L'attraction par Hirschmanniella oryzae et par les juveniles de 2' stade de Meloidogyne incognita est mfluencee par differents facteurs : nombre des proies, temperature, penode d'lncubation des proies, jeune des proies, distance entre proie et predateur Les deux especes predatnces sont attires en nombre maximal lorsque 50 proies ont sejourne 12 heures sur I'agar de la boite de Petn, a 25-30°. Le jcune des predateurs accroit l'attraction; enfin celle-ci est inversement proportionnelle a la distance entre proie et predateur.
Predatory nematodes are unable to detect their prey and their predation depends on chance encounters (Yeates, 1969; Grootaert & Maertens, 1976; Small & Grootaert, 1983; Bilgrami, Ahmad & Jairajpuri, 1984). Esser (1963) speculated that dorylaim predators were attracted towards excised prey from short distances. Bilgrami, Ahmad and Jairajpuri (1985 a), and Shafqat, Bilgrami and Jairajpun (1987), however, did not find any positive attraction in Aquatides thomei (Schneider) and Dorylaimus stagnalis (Du)ardm) respectively towards excised prey, but observed aggregation of more than one predator taking place around a prey injured by another
Revue Nematol 1] d) 195-202 (1988)
predator. Yeates (1969) and Wyss and Grootaert (1977) also reported similar aggregation in Diplenteron potohi-kus (Yeates) and Lahronema vulvapapillatum (Loof & Grootaert). Bilgrami and Jairajpuri (1988) observed the pre- and post-feeding aggregation of Mononchoides longicaudatus (Khera) and M. fortidens (Schuurmans Steckhoven) at feeding sites and attributed this phenomenon to lingenng prey attractants in that area. The predatory mononchs, however, did not show such a response towards their prey (Bilgrami, Ahmad & Jairajpuri, 1984).
In the present work observations on Mononchoides
195

IC A. L. Bilgrami & M. S. Jairajpuri
longicaudatus (Khera) and M. fonidens (Schuurmans Stekhoven) were made to study their attraction towards prey and factors influencing their attraction.
Materials and methods
G E N E R . - \ L CONDI'lIONb
Mononchoides longicaudatus and M. fonidens were cultured separately in 1 "i water-agar using Rhabditis sp., as prey. Five mg of infant milk powder (" Lactogen ") was spread on the surface of the agar to encourage bacterial growth which served as food for the prey nematodes. The predators, as and when their population declined, were transferred to fresh culture dishes containing prey.
In all experiments, attraction of M. longicaudatus and M. fonidens was tested in 0.5 cm thick layer of 1 "o water-agar using ten prey nematodes at 28 ± V. The most suitable concentrations and layer of agar was 1 or 2 ''o and 0.5 cm respectively. It was therefore, these characteristics used for all experiments. The nematodes were incubated for eight hours in Petri-dishes containing autoclaved water-agar (without bacteria) prior to the inoculation of predators. The distribution of predators was recorded after four hours. Each experiment was replicated 20 times. All nematodes used as prey were extracted fresh from the soil and sterilized with 0.01 "n mercuric chloride. Unless mentioned otherwise the conditions remained identical in all experiments.
All experiments were carried out in 5.5 cm diameter Petri-dishes, divided into three zones viz., inner, middle and outer by drawing two concentric circles 0.5 and 2.5 cm in diameter on the bottom. A plastic straw pipe 5 mm high, 5 mm in diameter, with a small piece of filter paper glued at one end was placed vertically in the inner zone of the Petri-dishes, with the filter paper in contact with the Petri-dish. Water-agar was then poured into the Petri-dishes and straw- pipe to make a 0.5 cm thick agar layer. Prey nematodes were released in the straw pipe and left for incubation. Five predators were then released at the periphery' of the middle zone. The distribution of predators was recorded and scores were obtained by summing up the product of the number of worms in each zone with their corresponding weighting factors. The weighting factors were obtained by dividing the area of outer zone by that of each of the three zones (Ahmad & Jairajpuri, 1980; Bilgrami, Ahmad & Jairajpuri, 1985 a). In the text MLS stands for " Mean Log Score ".
ATrR.-\(rnoN IN M. I.OXGICAVDAIVS .\SV) M. I-ORIIDEXS
Attraction was tested separately towards excised (cut in two pieces) and non-excised (live) Rhabditis sp., and agar containing unidentified bacteria (cultivated on agar
by adding Lactogen milk powder). Petri-dishes without prey or bacteria served as control. Observations were made after 1, 2 , 3 and 4 h intervals.
ArrRiVCTiON TOWARL:)S 15II-TF.RENT PREY NEMATODES
Attraction of M. fonidens and M. longicaudatus was tested separately by Petri-dish experiments with excised and non-excised prey nematodes. Adult Hirschmanniella oryzae, Hoplolaimus indicus, Helicotylenchus indicus, Hemicriconemoides mangifeme, Paralongidonis sp., Lon-gidorus sp., Xiphinema americanum, Tylenchorhynchus mashhoodi, Mononchoides fonidens, Mononchoides longicaudatus and the second stage juveniles of Meloido-gyne incognita and Anguina tritici were used as prey.
FACTORS INFLUENCINC} ArrR^MrnoN
To determine the effect of the following factors on the attraction, live adult H. oryzae and the second stage juveniles of M. incognita were used. M. fonidens and M. longicaudatus were tested separately towards individual prey nematodes.
Prey number .-To observe the effect of prey numbers on the attraction of M. fortidens and M. longicaudatus 5, 10, 15, 20, 25, 30, 35, 40, 45 and 50 prey nematodes were placed in the straw pipe. They were incubated prior to the inoculation of predators.
Incubation of prey nematodes : Prey nematodes were incubated for 2, 4, 6, 8, 10 and 12 h by placing in the straw pipe of the separate Petri-dishes. After incubation M. longicaudatus and M. fortidens were released and their distribution recorded after the desired time.
Temperature : Prey nematodes were incubated at 5, 10, 15, 20, 25, 30, 35 and 40" to determine the effect of temperatures on the attraction of M. longicaudatus and M. fonidens. After incubation the predators were released and Petri-dishes containing prey and predators were again placed at the same temperatures at which they were kept earlier. Observations on the distribution of predators were made when required.
Starvation of predators : M. longicaudatus and M. fortidens were starved for 0, 2, 4, 6, 8, 10 and 12 days by placing them in cavity-blocks containing water (without prey). Predators were transferred to fresh water each day. Day 0 refers to fresh predators (well fed). Each group of starving predators was tested separately towards previously incubated prey nematodes.
Distance of predators from prey : The effect of the distance of predators from the prey on their attraction was tested in 7 cm diameter Petri-dishes containing water-agar. The dishes were marked at the bottom by straight lines into seven zones and numbered serially as 1, 2, 3... 7 (Bilgrami, Ahmad & Jairajpuri, 19856). A plastic straw pipe with one end sealed with a piece of
196 Revue Nemalol. II '2i : 195-202 '1988,

Attraction o/Mononchoides spp. towards prey
filter paper was placed vertically in zone 1 so that the sealed end remained inside the agar. Prey nematodes (H. oryzae I were released in the straw pipe and the whole set was left for incubation. Twenty-five predators were then released at different points in zone 2. Similarly, predators were released in the zones 3, 4, 5, 6 and 7 of the separate Petri-dishes and observations were made after 4 h. Attraction of M. longicaudatus and M. fortidens was tested in separate Petri-dishes.
Results
AnR^U'lION' I \ M. tOR 1 IDh\S .Wll M. lOSGKAVDAlUS
(Fig. 1)
Both the predators responded positively towards live and excised Rhabduis sp., and agar containing bacteria. Attraction increased significantly from 1 h to 4 h (p < 0.05). The attraction of M. longicaudatus (Fig. 1 A) and M. fortidens (Fig. 1 B) (MLS 2.43 and 2.44 respectively) was more towards Rhabduis sp., than the live (MLS 2.26 and 2.28 respectively) or agar containing
L^yyi Rhq&d t 5 sp non exc sed )
I I Rhobditis sp (excised J
n
Agar containing bacterit
B
bacteria (MLS 1.90 and 2.13 respectively) after 4 h (p < 0.05). Minimum number of both the predators were attracted in the agar containing bacteria.
AriRACTION TOWARDS DII-hERE\T PRFY NEMAIODFS
(Fig. 2)
M. longicaudatus and A l fortidens responded differently towards live and excised prey. When tested towards live prey, H. oryzae was most attractive to M. longicaudatus (Fig. 2 A). Prey nematodes viz., Helicoty-lenchus indicus, H. mangiferae and Longidorus sp., were least attractive. The attraction towards H. oryzae (MLS 2.06) was higher than towards Helicotylenchus indicus, H. mangiferae and Longidorus sp., (MLS 1.58, 1.58 and 1.59 respectively, p < 0.05). Excised prey attracted more M. longicaudatus than the non-excised ones (Fig. 2 B). Excised T. mashhoodi attracted most (MLS 2.34) while Helicotylenchus indicus the least (MLS 1.46) M. longicaudatus. There was no significant difference in the
" 3D0^
:c^
I ^
^ I i
n
^ ^ ^
M
Fig. 1. Attraction of M. longicaudatus and M fortidens. A : M. longicaudatus; B : M. fortidens
Fig. 2. Attraction of AI. longicaudatus and M. fortidens towards non-exciscd and excised prey nematodes. Above : Non-excised. Below : Excised prey. A : Hirschmanmella oryzae, B : Hoplolainius mdicus; C : Helicotylenchus indicus, D : Hemicriconemoides mangiferae; E : Paralongidorus sp.; F : Longidorus sp.; G : Xiphinema amencanum; H : Tylenchor-hynchus mashhoodi; I : Anguina tntici juvemies; J : Meloido-gyne incognita juveniles; K • Mononchoides longicaudatus;h : Mononthoides fortidens
Revue Nematol. 11 2 : 195-202 1988 197

1C4 A L Bilgrami & M S Jairajpun
at
o
u»
c o 0/
?-3n
2.10.
1.411
1.70.
l.SO
• ^ y
/ — A •
/ •
T 1 1 1 1 1 1 1 1 1 5 10 15 20 25 30 35 40 i 5 50
Prey number
2-45
" 2 00.
% 170.
1.40
2.20'
2.0CL
1.80.
-1 1 1 i 1 1 2 4 6 8 10 12
Incubation periods of prey( h)
2 150.
1.40 -\ 1 1 1 1 1 1 1 5 10 15 20 25 30 35 40
Temp CC)
2-301
2.10-
1.90.
1-70.
1.50
245'
20a
1.70.
UO
/ .o-o-
/ O - o ^ o ^ o - — o
. /
/
. / B
/
/ /
- 1 — I — I — I — \ — I — I — I — I — I 5 10 15 20 25 30 35 40 45 50
Prey number
O O H-oryzge
0 0 M.incoqnito juv.
D
Jy - f — I 1 1 1 1
2 4 6 8 10 12
Incubation periods of prey(h)
2-2 tf
2.00.
O 180_
• 1.60_
Temp (°C)
Fig. 3. Factors influencing attraction of M fortidens and M. longicaudatus A : Effect of prey numbers on the attraction of M fortidens; B : Effect of prey numbers on the attraction of M longicaudatus, C : Effect of incubation penod of prey on the attraction of M fortidens, D Effect of incubation period of prey on the attraction of M. longicaudatus; E : Effect of temperature on the attraction of M fortidens; F : Effect of temperature on the attraction of M. longicaudatus.
198 Revue Nematol. 11 Q) • 195-202 (1988)

Attraction o/Mononchoides spp towards prey
attraction of M longicaudatus towards excised H ory-zae, Hoplolaimus indicus and A tntici juveniles (MLS 2 20, 2 19 and 2 16 respectively), and M incognita )uvc-mles and H mangiferae (MLS 2 04 and 2 02 respectively, p > 0 05) M longicaudatus was also attracted towards its own individuals
Among all the prey nematodes tested alive, H oryzae attracted most M fonidem (MLS 2 15) while Hebcoty-lenchus indicus did the least (MLS 1 53, Fig 2 B) There was little vanation in the attraction towards non-excised T mashhoodi {MLS 2 08) and Hoplolaimus indicus (MLS 2 02), and A tntici )uveniles (MLS 1 81) and Paralongidorus sp , (MLS 1 84) Excised H oryzae attracted maximum predators and the attraction (MLS 2 38) was more than that towards other nematodes (p < 0 05) Secretions of excised Hebcotylenchus indicus
were least attractive to M fortidens (MLS 1 65) These predators also responded positively towards their own individuals when tested alive (MLS 1 72) or excised (MLS 2 12)
FAC rORS I \FILE\( I \C, ArTRACno\ Prey numbers (Fig 3 A & B) Attraction of M
fortidens increased with the increase in the number of prey (Fig 3 A) The attraction towards five M incognita juveniles and H oryzae (MLS 1 74 and 2 14 respectively) increased significantly and was maximum when 50 individuals of the same prey nematodes were present (MLS 2 17 and 2 22 respectively, p < 0 05) M longicaudatus also responded indentically (Fig 3 B)
Period of incubation of prey (Fig 3 C & D) Attraction of both type of predators increased with the increase in the period of incubation of prey The predators responded more towards iVf incognita )U'veni\th than H oryzae which were incubated for the maximum duration (12 h) and least when incubated for only 2 h (p < 0 05)
Temperature (Fig 3 E & F) M fortidens and M longicaudatus were attracted more towards M incognita juveniles and H oryzae at 30 and 35 Higher and lower temperatures showed dechning attraction of both the predators (p < 0 05) Least attraction was recorded at 5
Starvation of predators (Fig 4 A & B) Twelve day starved M fortidens (Fig 4 A) and eight day starved M longicaudatus (Fig 4 B) exhibited maximum attraction towards H oryzae and the second stage juveniles of M incognita Their attraction was more than the fresh predators (p < 0 05) M longicaudatus, starved for more than eight days showed considerable decline in the attraction (Fig 4 B)
Distance of predators from prey (Fig 5) Increase in the distance of predators from the prey affected their movement towards prey M fortidens and M longicaudatus were attracted more towards H oryzae (zone 1) when tested from the zones 2 and 3 The attraction was moderate when predators were tested from other zones of the Petn-dishes
Discussion
Predatory nematodes are unable to detect their prey even from short distance (Nelmes, 1974) However, Esser (1963) and Yeates (1969) observed aggregation of predators around previously injured prey and attributed this phenomenon to prey secretions Bilgrami and Jai-rajpun (1988) in their studies on Mononchoides longicaudatus and M fortidens have attnbuted the pre- and post-feeding aggregation of these predators at feeding sites to the lingering eff( ct of prey secretions attractants, they have also speculated on the role of predators
2 40'
2 20.
° 2 00
o
160
. /
A
. / ' . • ^ , ,
X 180. y^
"T 1 1 1 1 1 1 0 2 ; 6 8 10 12
240'
2 20.
2 00.
180.
160
O — o y oryzae
• — • M incoqnitd jiiv.
B
°^o
• - - .
n 1 1 1 i 1 1 0 2 4 6 a 10 12
Starvation of predators (days) Starvation of predators(days)
Fig 4 Effect of starvation of predators on their attraction towards prey, A M fortidens, B M longicaudatus
Revue Nematol 11 2) 195-202 '1988/ 199

Qcf A. L. Bilgrami & M. S Jairajpun
15
10.
5.
i
—m ,
A
( I
12
U1
2 ^-•u
i _
1 2 3 A 5 6 7
c
1
1 2 3 4 5 6 7
• 10
^
E
—
1 2 3 A 5 6 7
9-r-] G
1 2 3 4 5 6 7
«, 9-1
ii 6. o •o
2 3. O.
O
^ I
o z
8"
1 2 3 A 5 6 7
1 2 3 4 5 5 7
Z ones
^
B
1 1 1
1 2 3 4 5 6 7
12'
6 . i
D
1 1 2 3 4 5 6 7
10 F
m^
1 2 3 A 5 6 7
9
'J H
1 2 3 4 5 6 7
^ J
1 2 3 4 5 6 7
8 '
4
L
1 2 3 4 5 6 7
Zones
Fig. 5. Effect of distance of predators from prey on the attraction. A, C, E, G, I & K : M. fomdens; B, D, F, H, J & L • M longicaudatus. Arrows indicate point of inoculation of predators.
200 Revue Nematol. 11 12) : 195-202 (1988)

Attraction o/Mononchoides spp towards prey
secretions (released during extra-corporeal feeding and digestion) in the aggregation of other predators
The present observations on the attraction of M longicaudatus and M fortidens reveal that these predators respond positively towards different prey m response to prey secretions/attractants Nematode reach bactena by chance or wilful movements (Poinar & Hansen, 1986), their orientation is determined by probing movements of the head (Ward, 1973, Seymour, Wnght & Doncaster, 1983) and location of bactena depends upon papillae and amphids (McLaren, 1976, Wnght, 1983). The present study also indicates attraction of M longicaudatus and M fortidens towards bactena
The differential response of predators towards different prey suggests their preferential behaviour It could also be possible that some prey nematodes secrete repellents which may be undesirable for the predators Esser (1963) has suggested that species of Hehcotylen-chus secrete some toxic substances which make them resistant to predation The fact that the two predators, M longicaudatus and M fortidens were less attracted towards Helicotylenchus indicus, Longidorus sp , and X amencanum indicates that these prey nematodes may secrete substances which are either toxic or unfavorable, leading to meager or moderate response of the predators Besides, the composition, concentration, quantity and quality of prey secretions may also play a role in differential responses of M longicaudatus and M fortidens towards different prey since these charactenstics may vary from species to species and individual to individuals
Wallace (1963) found lower (5-10 ) and higher (40 or more) unfavorable for the nematodes and Bilgrami, Ahmad and Jairajpun (1983) found temperatures influencing activity of the predatory nematodes, Monon-chus aquaticus The decrease in the attraction of M longicaudatus and M fortidens at temperatures lower than 25 and higher than 35 may be attnbuted to their activity which might be inhibited by the lower and higher temperatures Besides, temperatures may also influence the dispersion of prey attractants in the surrounding resulting in a differential predator responses
The differential response of M longicaudatus and M fortidens towards prey nematodes, which were incubated for vanable durations suggest that a minimum penod is necessary for the development of the perceptible gradients of prey attractants Doncaster and Seymour (1973) concluded that the minimum response threshold decreases in starving nematodes which could sense even the weaker stimuli. However, Bilgrami, Ahmad and Jairajpun (1985^) had suggested an increase in the minimum response threshold level in starving H oryzae Dunng present observations it seems that the minimum response threshold of M longicaudatus and M fortidens decreased with the increase in the penod of starvation resulting in increased attraction from two to twelve day
starved M fortidens and two to eight day starved M longicaudatus In the latter predator the threshold level increased after eight of starvauon and resulted in their lesser attraction towards prey
Attraction of nematodes is influenced by the distance of nematodes from the source of attraction (Bilgrami, Ahmad & Jairajpun, 19856) The effect of distance of M longicaudatus and M fortidens from the prey on the attraction may be attnbuted to the time taken by the prey secretions/attractants to form an optimum threshold level in agar as well as the time taken by the attractants to disperse in the surroundings and to reach the predators Stimuh of stronger intensities may require lesser time to develop a minimum response threshold and could easily be spread m a particular area faster than those of the weaker intensities
Cannibalism seems to be a natural phenomenon occumng m most predators (Bilgrami, Ahmad & Jairajpun, 1986) under conditions of non-availability of prey nematodes (Bilgrami & Jairajpun, 1985) The ability of predators to recogmze their own chemical secretions (Esser, 1963, Grootaert & Small, 1982, Bilgrami, Ahmad & Jairajpun, 1985 a, Shafqat, Bilgrami & Jairajpun , 1987) may induce cannibalism D u n n g present observations the attraction of M longicaudatus and M fortidens towards their own individuals suggests their ability to recognize their own chemical secretions which may also lead them to prey upon their own individuals (cannibalism)
ACKNOWLEDGEMENTS
The authors are thankful to the Chairman, Department of Zoology, Aligarh Muslim University, Aligarh for providing neccessary laboratory facilities and Dr Irfan Ahmad for his suggestions
REFtRENC ES
AHMAD, I & JAIRAJPLRI, M S (1980) Sex attraction and copulation in Chiloplacus symmetncus (Nematoda Acro-belinae) Nematologica, 26 139-148
BILGRAMI, A L , AHMAD, I & JAIRAJPLRI, M S (1983) Factors influencing predation by Mononchus aquaticus Revue Nematol, 6 325-326
BILGRAMI, A L , AHMAD, I & JAIRAJPLRI, M S (1984)
Observations on the predatoryf behaviour of Mononchus aquaticus Nematol medit, 12 41-45
BILGRAMI, A L , AHMAD, I & JAIRAJPLRI, M S (1985a)
Predatory behaviour of Aquatides thomei (Nygolaimina Dorylaimida) Nematologica, 30 (1984) 457-462
BILGRAMI, A L , AHMAD, 1 & JMRAJPLRI, M S (19856)
Factors influencing attraction of adult Hirschmanniella oryzae towards cabbage seedlings Revue Nematol, 8 67-75
BILGRAMI, A L , AHMAD, I & JAIRAJPLRI, M S (1986) Study on the intestinal contents of mononchs Revue Nematol, 9 191-194
Revue Nematol 11 <2) 195-202 1988) 201

A L. Bilgrami & M S jfatrajpun
BiLGRAMi, A. L. & JMR.'VJPLRI, M . S. (1985) Cannibalism in Mononchus aquaticus. Indian J. Nematol, 14 (1984) • 202-203.
BliGRAMi, A. L. & J\iR.^jPLRl, M. S. (1988). Aggregation of Mononchoides longicaudatus and M fortidens (Nematoda . Diplogastenda) at feeding sites. Nematologica (in press).
D o \ c \sii-R, C C. & Sf\Moi R, M. K. (1973) Exploration and selection of penetration site by tylenchids. Nematologica, 20 : 297-307
EssFR, R. p . (1963). Nematode interactions in plates of non-stenle water agar. Proc Soil Crop Sa Soc Fla, 23 121-138.
G R O O I \ E R I , P & MAPRIJ -NS , D . (1976). Cultivation and life cycle of Mononchus aquaticus. Nematologica, 22 : 173-181.
G R O O I \i RT, P. & S M \ I I , R . W . (1982). Aspects of the biology of Labronema vulvapapillatum (Meyl) (Nematoda, Dory-laimidae) in laboratory culture. Biol. Jb. Dodonaea, 50 : 135-148.
M v i.\Rl \ , D J. (1976). Nematode sense organs. In Dawes, B. (Ed.) Adv Parasttol, 14, Academic Press, London 195-265.
N F L M L S , A . J. (1974). Evaluation of the feeding behaviour of Pnonchulus punctatus (Cobb), a nematode predator. J anim Ecol, 43 553-565.
PoiNAR, G. O Jr & H \ N s t \ , E . L.( 1986). Association between
nematodes and bacteria. Helminthol. Abstr Ser. B, 55 : 61-81.
StvMOLR, M. K . , W R I G H I , K A . & D O \ C \ S I L R , C C . (1983). The action of antenor feeding apparatus of Caenorhabdms elegans (Nematoda : Rhabditidea). J ZooL, 201 . 527-539.
S M \ I I , R. W. & G R O O I \LRi, P (1983). Observations on the predation abilities of some soil dwelling predatory nematodes. Nematologua, 29 . 109-118.
S H A F Q M , S., BIU.RAMI, A . L . & JMRAJPIRI , M . S. (1987).
Evaluation of the predatory behaviour of Dorylaimus stagna-/wDujardin, 1845 (Nematoda : Dorylaimida). Revue Nematol., 10 : 455-461.
WAI 1 \( p, H. R. (1963). The biology of plant-parasitic nematodes. London, Edward Arnold, 280 p.
W\RD, S. (1973). Chemotaxis by the nematode Caenorhabdms elegans . Identification of attractants and analysis of the response by use of mutants. Proc natl Acad. Sci USA, 70 : 817-821.
W R R . H I , K . A . (1983). Nematode chemoscnsiUa • form and function. J. Nematol, 15 : 151-158.
WYSS, U . & G R O O I AiRi, P. (1977). Feeding mechanism of Labronema vulvapapillatum Medd. Fac. Landbouww. Rijk-sumv Gent, 42 : 1521-1527.
Yi \1LS, G. W. (1969). Predation by Mononchoides potohikus (Nematoda Diplogastendac) in laboratory culture. Nematologica, 15 . 1-9
Accepie pour publication le 12 juin 1987
202 Revue Nematol. 11 (21 : 195-202 '1988,

A T. 09
Indian J. Neinawl. 17(2) : 339-340 (1987)
Interaction between mononchs in non-sterile agar plates
ANWAR L. BILGRAMI, IRFAN AHMAD and M. SHAMIM JAIRAJPURI
Section of Nematology, Department of Zoology, Aligarh Muslim University, Aligarh-202 001, India
Several studies have been made on the predatory behaviour of mononchs (Nelmes, 1974, Cohn & Mordechai, 1974; Jiirajpuri & Azmi, 1978; Small, 1979; Bilgrami Ci-a/., J983; 1984) and few on their cannibalistic behaviour (Jenson & Mulvey, 1968; Azmi & Jairajpuri, 1979; Bilgrami & Jairajpuri, 1985). However, interspecific and intraspecific interactions between mononchs have not been studied much. Esser (1963) was the first to study interactions between different nematodes including mononch and dorylaim predators in non-sterile agar plates. In the present work experiments were performed to observe interactions between three species of mononchs in agar plates.
The interaction between the predators was studied by placing varying numhers of lotoncfuis indicus Jairajpuri, 1969; Mononchus aquaticus Coetzee, 1968 and Mylonchulus fife/7?fl?wj'Jairajpuri, 1970 together in Petri-dishes containing 1% water-agar. The four combinations studied were 25 M. dentatus ~ 15 M. aquaticus - 1 0 7. indicus; 25 /. indicus 4-25 M. aquaticus; 25 /. indicus -f 25 M. dentatus; 25 M. aquaticus ~i- 25 M. dentatus. The number of individuals of different species killed or injured was counted after 24 h. only adults were used for all experiment. Experiments were replicated five times.
When 25 M. dentatus, 15 M. aquaticus and 10 /. indicus were placed together, M. dentatus was killed by both /. indicus and M. aquaticus. The number of M. dentatus killed by /. indicus in ai! combinations was significantly higher than those killed by M. aquaticus (p < 0.05). When /. indicus and
aquaticus
M. aquaticus were kept together, the latter species was attacked and devoured by the former. No specimen of /. indicus was either killed or injured, though attacked frequently by M-(Fig. 1).
From the evidences presented it may be inferred that M. dentatus was the v/eakest predator being attacked and devoured by both M- aquaticus and /. indicus without itself being able to injure the latter two species. M. aquaticus was a stronger predator than M. dentatus. while /. indicus appeared to be strongest
^M.aquaticus
20 riM.dentatus
0)
in ro
>
c
15.
10.
5-J
i FJG. ]. Interaction between diiTerent species of
mononchs
a = 25 M. dentatus '- 15 M. aquaticus -\ 10 /. indiCus.
b = 25 /. indicus -t- M. aquaticus. c • - 25 /. indicus + 25 M. dentatus.
d '- 25 M. aquaticus ',- 25 M. dematiis.

110
340 SHORT COMMUNICATIONS
being able tokillanddevoure M- dentatus as well as M. aquaticus. The predatory capabilities of these species may be dependent on size of the nematodes as well as their activity characteristics {Bi\gv&m\ et ai, 1983).
The authors are grateful to the Chairman, Department of Zoology Aligarh Muslim University, Aligarh for providing neccessary laboratory facilities.
REFERENCES
AzMi, M.I. & JAIRAJPURI, M.S. (1979). Nematol. Medit. 7 : 203-207.
BiLGRAMi, A.L. , AHMAD, I. & JAIRAJPURI, M.S. (1983). Revue Nematol. 6 : 325-326.
BiLGRAMi, A.L., AHMAD, [. & JAIRAJPURJ, M . S (1984). Nematol. Medit. 12 : 41-45.
BiLGRAMi, A.L. & JAIRAJPURI, M.S. (1984). Indian J. Nematol. 14 : 202-203.
CoHN, E. & MoRDECHAi, M. (1974). Nematol. medit. 2 : 43-53.
EssER, R.P. (1963). Proc. Soil. Crop Sci. Soc. Fla. 23 : 121-138.
JAIRAJPURI, M.S. & AZMI, M.I. Nematol. Medit. 6 : 205-212.
(1978).
JENSON, H.J. & MULVEY, R.H. (1968). Ore. St. Monogr. Stud. Zool. No 12 : 1-57.
NELMES, A . J . (1974). / . Anim. Ecol. 43 : 553-565.
SMALL, R . W . (1979). Nematologica IS : 94-103.

Int. Neraatol. Netv/ork Newsl. 5(4) :4. 1983
PLANT-PARASITIC NEMATODES ASSOCIATED V ITH PADDY CROP IN BIHAR, INDIA
Wasiin Ahmad, Tarique H. Khan, and Anwar L. Bilgrami Post Doctoral Research Fellow, Research Scholar and
Post Doctoral Research Fellow, respectively Section of Nematology Department of Zoology
Aligarh Muslim University Aligarh 202 002
INDIA
A random survey was conducted to identify and assess the frequency of occurrence of economically important plant-parasitic nematodes of paddy Oryza sativa L. in three rice-growing districts of Bihar, India. A total of 36, 26, and 11 soil samples were collected at random from paddy fields in the districts of Hadhubani, Sitamarhi, and West Champaran, respectively. The soil samples were processed by Cobb's decantation and Baermann's funnel techniques. The nematodes per 200g of soil from each sample were counted and identified.
The rice root nematode Hirschmanniella oryzae (van Breda da Haan, 1902) Luc & Goodey, 1964, was found in 90% of samples from Madhubani, 61% from Sitamarhi, and 73% from West Champaran. Average populations per 200g soil ranged from 50 to 226. Tylen-chorhynchus mashhoodi Siddiqi & Basir, 1959, was found in 90% of samples from Madhubani, 47% from Sitamarhi, and 100% from West Champaran, Average populations per 200g soil ranged from 16 to 104. Therefore, in addition to H. oryzae, a predominant pest of paddy in these areas, T. mashhoodi may also be a potential threat.
Most samples contained lower populations of Helicotylenchus indicus Siddiqi, 1963 [<.40/200g]; H. dihystera (Cobb, .1893) Sher, 1961 [<54/200g]; Pratylenchus coffeae (Zimmerman, 1898) Filipjev & Schuurmans Stekhoven, 1941 [<60/200g]; Basiria tunida (Colbran, 1960) Geraert, 1968 [<53/200g]; and Ditylenchus sp. [£40/200g]. Other important plant-parasitic nematodes encountered were Aphelenchus avenae Bastian, 1865; Aphelenchoides besseyi Christie, 1942; Coslenchus costatus (De Man, 1921) Siddiqi, 1978; Tylenchorhynchus divittatus (Siddiqi, 1961) Jairojpuri, 1984; Plenicriconemoides cocophillus (Loos, 1949) Chitwood & Birchfield, 1957; Hoplolaimus indicus Sher, 1963; Neo-psllenchus maqnidens (Thorne, 1949) Thorne & Malek, 1968; Pratylenchus thornei Sher & Allen, 1953; Paratrichodorus porosus (Allen, 1957) Siddiqi, 1974; Xiphinema insiqnis Loos, 1949; and Malenchus sp.

Indian J. Neimtol. 18(1) : 110-112 (.1988)
Observations on the osmoregulation in an insect nematode, Schwenkiella at her i (Thelastomatidae : Oxyuroidea)
RAFIA PARVEEN, ANWAR L. BILGRAMI and DURDANA S. JAIRAJPURI
Department of Zoology, Aligarh Muslim University, Aligarh-202001, India
112
Observations on the eifects of different dilutions of salt solution on the osmoregulation in Schwenkiella atheri Parveen & Jairajpuri, 1982, a nematode parasite of the cockroach {Periplaneta americana), are reported here.
Specimens (S. atheri) were collected from the intestine of P. americana, washed with distilled water, kept in normal saline and used within 30 mts. For each experiment fresh nematodes were collected from the cockroach gut. 1.0 M solutions of potassium chloride, calcium chloride, potassium iodide and sodium chloride were prepared and then dilutions of 0.01, 0.05,0.10,0.15, 0.20 and 0.25 M were made for evaluation of their effects on the body length of nematodes. The small size and fewer number of S. atheri have precluded us from using weight changes as the parame ter. Adult nematodes were placed in cavity blocks, containing small quantity of experimental solution. To prevent evaporation and to keep osmotic pressure of the solution constant, cavity blocks were covered with glass lids and edges were sealed with vaseline, and then placed in an incubator at 25 ± 1°C. These were removed at hourly intervals for observations. Fach experiment was replicated five times. The results are p resented in figure (1, A-D) as per cent increase or decrease in length from the Original (control) which was taken soon after dissecting out nematodes from intestine of the host. The nematodes were measured from camera lucida drawings (Stephenson, 1942).
Concentrations of 0.01, 0.05 and 0.10 M potassium chloride were hypotonic and the length of the body increased significantly (p < 0.05) (Fig. 1, A). The
body length was more in 0.15 M but decreased below control after 5 h. Reduction in the body length was also observed in 0.20 and 0.25 M (hypertonic solutions).
The 0.1 M of calcium chloride was isotonic. No difference from the original length was observed except at 2 h when the length was slightly more. Concentrations 0.01 and 0.05 M were hypotonic and 28 and 22 per cent increase in the length was observed at the end of 8 and 1 h, respectively (Fig. 1, B). The nematodes showed a maximum of 12 per cent decrease in length in 0.15 and 27 per cent in 0.20 and 0.25 M dilutions of calcium chloride which were hypertonic to the parasite.
Body length increased upto 14 and 7 per cent, respectively, when nematodes were treated with 0.01 and 0.05 M potassium iodide (Fig. 1, C). In 0.10 M the nematodes behaved somewhat similarly as in the above dilutions but after 6 h showed signs of shrinking. The 0.15,0.20 and 0.25 M were hypertonic to nematodes.
All concentrations, of sodium chloride were hypotonic except 0.25 M in which initially (upto 3h) the nematodes lost their length but gained afterwards. The maximum (13 per cent) increase in the length was in 0.01 M and least (3 per cent) in 0.25 M solution (Fig. 1, D).
Lee (1960) found shrunken Hammer-schmidtiella diesingi from the hind gut of Blatta orientalis which he attributed to the increase in the osmotic pressure of the gut contents resulting in the f uid of the gut becoming hypertonic during the period of moulting. The change in the osmotic pressure during moulting in

n 111 SHORT COMMUNICATIONS
•—•—•-
Hours Hours
c ^
15
10- . Q> w s: a • " 4) C7I 1- C ^ ^ "•
c
\ d) '
0) D» *- c u (b 0, — Q
b.
0
5
10.
15.
20.
Mours
Fig. 1. Observations on ihe osmoregulation in
A—Potassium chloride, B—Calcium chloride, C-
.01 M — 0 .05M
.10 M ®-® .15M .—. .20M , _ . .?5M
5
loj
15.
20
1 2 3 'i 5 6 '" 8 Hours
an insect nematode, Schwenkiella atheri -Potassium iodide, D—Sodium chloride
P. americana may also affect the respon- the external medium. Pannikar & sesofS. atheri. The present observa- Sporston (1941) observed the body fluid tions on S. atheri show that adults can of Angusticaecum sp. hypertonic even in survive changes in osmotic pressure of distilled water. Stephenson (1942) showed

1 14
SHORT COMMUNICATIONS 112
increase and decrease in length due to flow of water in and out of Rhabditis terrestris under the influence of osmotic forces. He (1. c) also observed a reduction in the body swelling after a prolonged immersion of nematodes in distilled water and suggested that this recovery was due to the existance of an active method of osmotic regulation. The hypertonic solutions decrease the length due to flow of water from the body while hypotonic increase it because of water absorption by the body (Stephenson, 1942; Lee, 1960; Viglierchio et al., 1969). During present observation on S. atheri adults the change in body length may be attributed to the hypo-and hypertonicity of salt concentrations and also to different osmotic pressures exerted upon nematodes. Some of those concentrations (0.15M of potassium chloride, 0.10 M of calcium chloride and
potassium iodide and 0.20 M and 0.25 M of sodium chloride) in which the body length was either equivalent to the original body length or increased upto a certain time but decreased below control thereafter or vice-versa suggest that the period of exposure to different dilutions of salts also affects the responses of S. atheri. Thus it seems that water regulation also takes place in S. atheri under the influence of osmotic forces.
REFERENCES
LEE, D. L. (1960). Parasitology 50 : 241-246. PANNIKAR, N. K. & SPORSTON, N. G.
(1941). Parasitology 33 : 214-223.
STEPHENSON, W. (1942). 253-265.
Parasitology 34 :
VIGLIERCHIO, D. R. CROLI, N. A. & GORTZ, J. H. (1969). Nematologica 15 : 15-21.

r
Resistance of prey to predation and strike rate of the predators, Mononchoides longicaudatus and M. fortidens
(Nematoda : Diplogasterida) Anwar L. BILGRAMI and M. Shamim JAIRAJPURI
Section of hematology, Department of Zoology, Ahgarh Muslim University, Aligarh-202002, India.
S i MVIAR"!
Resistance of prey ncmauxies to predanon, stnke rate of the predators, Mononchoides longicaudatus and M. fortidens and their ability to wound prey and feed upon them was determined on agar plates. M. longicaudatus and M. fortidens attacked all kinds of prey nematodes. The second stage juveniles of Meloidogyne incognita and Anguina tntici and the adult Acrobeloides sp., Cephalobus sp., and Panagrellus redivivus were most susceptible to predation while Hemicnconemoides mangiferae and Hoplolaimus mdicus were totally resistant. Hirschmanniella oryzae, Helicotylenchus indicus, Tylenchorhynchus mashhoodi, Xiphinema amencanum, Rhabditis sp., Tnchodorus sp., and Longidorus sp., were attacked with intermediate frequencies. M. longicaudatus took a minimum of 15-25 mm. to finish a luvenile of M. incognita and maximum 80-110 min. for the Longidorus sp., white M. fortidens consistently consumed prey at a faster rate.
RFSI MI
Resistance des praxes a. la predation el taux d'attaque des pre'dateurs Mononchoides longicaudatus et M. fortidens (Nematoda : Diplogastenda)
La resistance des nematodes-proies a la predation, le taux d'attaque des predateurs, Mononchoides longicaudatus et M. fortidens, amsi que leur habilete a blesser les proies et s'en nournr ont ete determines sur plaques de gelose. M. longicaudatus et M. fortidens attaquent tous les types dc proies. Les juveniles de deuxieme stade de Meloigyne incognita et A'Anguina tntici et les adultes d'Acrobeloides sp. Cephalobus sp. et Panagrellus redivivus sont les plus exposes a la predation tandis que Hemicnconemoides mangiferae et Hoplolaimus mdicus sont totalement resistants. Hirschmanniella oryzae, Helicotylenchus indicus, Tylenchorhynchus mashhoodi, Xiphmema amencanum, Rhabditis sp., Tnchodorus sp. et Longidorus sp. sont attaques avec des frequences moyennes. II faut a M. longicaudatus un minimum de 15 a 25 minutes pour achever un juvenile de M incognita, et un maximum de 80-110 minutes pour Longidorus sp., tandis que M fortidens ingere ses proies plus rapidement.
The predatory nematodes use a variety of mechanisms to obtain prey. Similarly, prey nematodes possess certain qualities such as the thick cuticle, annulations, chemical secretions, etc., which may protect them from predators (Esser, 1963). Esser (1963) provided a list of nematodes that he found to be susceptible or resistant to predation by different mononchs and dorylaims. Grootaert, Jaques and Small (1977) observed prey selection in Butlenus sp., and measured susceptibility and resistance to predation of tylenchid, aerolaimid, enoplid and dorylaimid nematodes. Small and Grootaert (1983) described anti-predation adaptations in prey nematodes. Very recently, Shafqat, Bilgrami and Jairajpuri (1987) observed prey selection in Dorylaimus stagnabs Dujardin, 1845 and suggested that the thick cuticle of Hoplolaimus indicus Sher, 1963 and chemical secretions of Helicotylenchus indicus Siddiqi, 1963 may represent anti-predation devices.
The present work describes experiments conducted to determine the degree of resistance/susceptibility of different plant-parasitic and free-living nematodes to predation by Mononchoides longicaudatus (Khera, 1965) Andrassy, 1984 and M. fortidens (Schuurmans Stekho-ven, 1951) Taylor & Hechler, 1966.
Materials and methods
M. fortidens and M. longicaudatus were cultured by the methods of Bilgrami and Jairajpuri (1988). Adult Acrobeloides sp., Cephalobus sp., Rhabditis sp., Panagrellus redivivus, Trichodorus sp., Longidorus sp., Xiphinema amencanum, Hirschmanniella oryzae, Tylenchorhynchus mashhoodi, Hoplolaimus indicus, Helicotylenchus mdicus, Hemicnconemoides mangiferae and the second stage juveniles of Meloidogyne incognita and Anguina tritici were used as prey.
Revue Ne'matol. 12 'L • 45-49 '1989i 45

u A I Bilgraini & M S Jairajpun
The free-livmg nematodes were obtained from the cultures mamtamcd m the laboratory These prey nematodes were cultured m Petn dishes containing 1 "o water-agar 5 mg of infant milk powder (Lactogen®) was spread over the surface of agar to grow bactena which served as food for the prey nematodes The second stage juveniles of M incogmia and A tntici were obtained from population maintained on tomato and the wheat galls respectively Other prey nematodes were isolated fresh from soil for each experiment 500 g of soil containing nematodes was processed by decantation and Baermann's funnel techniques
The encounters (lip contact of predator with the prey at right angle) between predators and prey were observed in Petn dishes containing 1 "u water-agar using a stereoscopic binocular microscope For each encounter a new predator and a prey individual was used (irrespective of whether the predator failed or succeeded m attacking the prey) A prey nematode was placed in front of the head of an active predator with the help of a fine needle without touching (disturbing) the predator in any manner Onl^ those observations were recorded where the predators behaved as normally as could be ensured To reduce the effects of satiation and prey habituation, 4-6 day starved adult predators were tested Fifty encounters were observed between predator and prey
The strike rate of predator ma> be defined as the percentage of success with which it attacks prey (attack represents the rapid movement of head and the application of suction created by oesophagous of the predator) and may reflect its predatory potential The strike rate SRi of M longicaudatus and M fortidens could be determined by the followiing formula
FA
tance susceptibility since it is an important factor in predation (Small & Grootaert, 1983) Esser (1987) also suggested that resistance of a prey is lost if it is wounded or IS m a weakened condition A wound may result in the loss of hydrostatic pressure of the body affecting locomotion and thereby making the prey more vulnerable to predation Wounding may also permit invasion of pathogenic micro-organisms resulting into the death of prey The mere encounters between the predator and the prey may not necessarily result into wounding of the prey
The percentage of piedators which started feeding after wounding the prey and the percentage of prey which were left unfinished by the predators were also determined Both species of predators were tested separately The untransformed results were analysed to obtain correlation coefficients '" r "), standard error 'SE) and level of significanc(" '" P "J with the help of the following statistical tests
IXY Correlation coefficient '" r "j =
V IX lY
Standard error (SE ±) 1 - r
SR X lOO
Where X and Y are the deviations measured from their respective means and A-'is the number of replicates Significance of " r " values was obtained directly from the Table for the significance of the Correlation coefficient
Results
Where EA = total number of encounters resulting into attack, E = total number of encounters made by the predator
1 he resistance of a prey may be defined as the degree of their ability to a\ old predation (wounding) by physical, chemical or behavioural means The percentage degree of resistance of a prey may be calculated as follows
PR Cu = EA-AW
EA X 100
Where PR = pre> resistance, AW = total number of attacks resulting into prey wounding
The susceptibility of a prey may be defined as their degree of succumbing to predation (wounding) The percent susceptibility of prey to predation may thus be obtained by the following formula
I PS 1%) = 100 — PR PS -= prey suscepubilityy
During the present study the wounding was considered as the primary factor in measuring prey resis-
The results show a high degree of correlation between the two species of predators for encounters resulting into attack (r = 0 89, S£ ± 0 05, p < 0 05), attacks resulting into wounding (r = 0 98, S£ ± 0 01, p < 0 05), feeding after wounding (r = 0 98, S i ± 0 01, p < 0 05) and number of prey left unfinished (r = 0 83, SL ± 0 05, p <005)
R E S I S 1 -WCP Oh PKt V 1 () I'RbD^ 1 I O \ ^ ND S I RIkE R A 1 E OE M Li)\OI(. HDATI. S
M longicaudatus attacked all types of prey nematodes with lowest strike rate o n / / mangiferae {ib'•\)) Encounters with P redivivus, Acrobeloides sp , Cephalobus sp , and the second stage juveniles of M incognita and A tntici resulted m maximum strike rate (92-96 " 0 and prey wounding (95-100"' ) byM longicaudatus (Tab 1) The success of M longicaudatus in attacking other prey species ranged between 68-90 " i This predator did not succeed m wounding any individual belonging to Hoplo-laimus indicus or H mangiferae but attacked 74 ' i and
46 R'vue Nematol 12(11 45-49119891

Predation fcy Mononchoidts longicaudat us and M fortidens 117
Tabic 1 Resistance of prey to predation and strike rate of Mononchoides longicaudatus
Prey nematodes
Panagrellus rediiiius Acwbeloides sp Cephalobus sp Rhahditis sp Anguina intici juv Meloidogyne incognita juv Hirschmanmella oryzae 1 ylenchorhynchus
mashhoodi Hehcotylenchus indicus Longidorus sp Xiphinema aniencanum Hoplolaimus indicus Hemicnconemoides mangiferae Tnchodorus sp
\uinber
of encounters
E
50 50 50 50 50 50 50
50 50 50 50 50
50 50
Number
of encounters resulting into attack
PA
47 i8 48 45 46 48 43
42 34 35 34 37
23 39
Strike rate
of predator
SR n
94 96 96 90 92 96 86
84 68 72 68 74
46 78
Sumber
of attacks resulting in pray i^oundmg
AW
46 48 48 29 43 46 35
33 25 22 22
0
0 22
Degree of resistance susceptibility
PR ,
4 0 0
36 7 4
19
21 26 39 35 0
0 44
PS
96 100 100 64 93 96 81
79 74 61 65
0
0 56
Feeding after wounding prey
/
100 96
100 86 98 98 94
73 48 •56
45 0
0 77
Prey left
Duration
of unfinished feeding
>
0 0 6
16 10 7
30
25 58 50 50
0
0 47
on single prey
'mn
30-35 20-25 20-25 30-45 25-30 15-25 40-55
35-50 60-70
80-110 70-85
0
0 45-60
\\\ figures are nearest to whole numbers
46 ' of the individuals respectively The second stages of M incognita and A tntzcz along with the adult/jcrafce-loides sp , P redivivus and Cephalobus sp , were highly susceptible to predation having only 0-6 i resistance against M longicaudatus (Tab 1) Other prey nematodes VIZ, H oryzae, I mashhoodi, Helicotylenchus indtcus, X amencanum, Rhabditis sp , Tnchodorus sp , and I ongidorus sp , had low to moderate degree of resistance against predation The percentage of predators which continued feeding on the prey after wounding them was also highest for the most susceptible prey nematodes as mentioned above The percentage of predators feeding on Rhabditis sp , was also high (86 "o) although this species was not wounded as frequently as the most susceptible prey species In contrast, fewer M longicaudatus (27 " ) continued feeding after wounding 74 'o Helicotylenchus indicus When feeding occurred no individual of P redtvivus and Acwbeloides sp , was left unfinished by M longicaudatus while 58 "i of Helicotylenchus indicus, 50 X amencanum and 50 "o Longidorus sp, were not completely consumed (p < 0 05) M longicaudatus took 15-25 min to finish a M incognita juvenile but 80-110 min for Longidorus sp
RiSISI A\Ct Oh PRI Y rOPRlD^vlION A\L:iSlRlKt R \ l l - OV M hORIIDtW
Though M fortidens attacked Hoplolaimus indicus and H mangiferae at success rates of 66 "o and 56 "( respectively, but none of them were ever wounded Maximum strike rates occurred against P redivivus, Acrobeloides sp , Cephalobus sp , and the second stage juveniles of TVf incognita and A tntici (96-100 "o) These nematodes were highly susceptible to predation (PR = 0-2 "() Consumption of these prey was also high (96-100 V) (Tab 2) Least number of attacks by M fortidens resulted into wounding X amencanum and Longidorus sp , as with AI longicaudatus Few attacks by M fortidens resulted in wounding Rhabditis sp , but wounded animals were usually consumed In contrast, only 51 "u of the predators fed on 78 "o of Helicotylenchus indicus which were wounded and 56 "o of these individuals were not completely consumed No individuals of P redivivus and Acrobeloides sp , were left unfinished by the predators M fortidens required 15-20 mm to consume a A/ incognita juvenile Acrobeloides sp , and Cephalobus sp , and 60-80 mm to consume a Longidorus sp, (Tab 2)
Rezue Nematol 12 1/ 45-49 1989 Al

8 A. L Bilgrami & M. S Jairajpun
Table 2
Resistance of prey to predation and stnke rate of Mononchoides fortidens
Prey nematodes
Panagrellus redivwus Acrobeloides sp. Cephalobus sp. Rhabditis sp. Anguina tnticx juv. Meloxdogyne incognita juv. Hirschmanmella oryzae Tylenchorhynchus mashhoodi Hehcotylenchus indicus Longidorus sp. Xiphmema amencanum Hoplolaimus indicus Hemicnconemoides mangiferae Tnchodorus sp.
Number
of encounters
<E
50 50 50 50 50 50 50
50 50 50 50 50
50 50
Number
of encounters resulting into attack
(EA,
49 48 50 48 48 50 44
44 45 39 40 33
28 42
Stnke rate of predator
SR (%)
98 96
100 96 96
100 88
88 90 78 80 66
56 84
Number
of attacks resulting in prey wounding
AW
49 47 50 34 47 49 37
39 35 30 33
0
0 25
Degree of resistance susceptibility
PR '%,
0 2 0
29 2 2
16
11 22 23 18 0
0 40
PS (%J
100 98
100 71 98 98 84
89 78 77 82
0
0 60
Feeding after wounding prey
l%i
99 100 100 88 96
100 89
77 51 67 61
0
0 80
Prey left
Duration
of unfinished feeding
'%l
0 0 4
13 4 2
18
20 56 50 45
0
0 40
on single prey
(mni
20-25 15-20 15-20 25-35 15-25 15-20 30-40
35-40 50-55 60-70 40-55
0
0 35-45
All figures arc nearest to whole numbers
Discussion
The high degree of correlation between M. fortidens and M. longicaudatus for encounters resulting into attack, attacks resulting m prey wounding, feeding after wounding and number of prey left unconsumed indicates that both species of predators are very similar in their behaviour and requirements and also that the predator-prey relationship measured for one species of predator strongly supports the observations made with other predator.
The prey nematodes can resist predation by physical, chemical and behavioural characteristics (Esser, 1963, 1987; Esser & Sobers, 1964) such as thick cuticles (Esser, 1963; Small & Grootaert, 1983); annulations (Esser, 1963); speed (Esser, 1963; Bilgrami, Ahmed & Jairajpuri, 1983); vigorous escape response (Grootaert, Jaques & Small, 1977; Small & Grootaert, 1983); toxic, unfavourable chemical secretions (Esser, 1963). Lack of predation, during the present experiments, on Hoplolaimus indicus and H. mangiferae might have resulted from their thick cuticles and annulations respectively, that provided resistance to prey from wounding. Mylon-chulus dentatus and Dorylaimus stagnabs also failed to kill or injure Hoplolaimus indicus (Jairajpuri & Azmi, 1978; Shafqat, Bilgrami & Jairajpuri, 1987). These authors also attributed thick cuticle as an anti-predation
characteristic. Small and Grootaert (1983) attributed active body undulations and vigorous escape response of Rhabditis oxycerca, Pelodera sp., and Plectus sp., as characteristics providing resistance against predation. During present study a combination of high attack rate against Rhabditis sp., comparatively lower rate of wounding and higher rate of feeding upon wounded individuals suggests that this prey nematode also relies primarily on active body undulations and vigorous escape response movements to resist predation by M. longicaudatus and M. fortidens.
The high degree of susceptibility to predation of Cephalobus sp., Acrobeloides sp., P. redivivus along with the second stage juveniles of M. incognita and A. tritici may be due to small body size, slow rate of movement and lack of protective cuticle adaptations (Bilgrami, Ahmad & Jairajpuri, 1983). These characteristics may be common among endoparasitic, phytophagous nematodes (Esser, 1963).
Esser (1963) observed that the dorylaim predators rarely attacked or succeeded in devouring Hehcotylenchus spp., and suggested some sort of chemical resistance in species of this genus. The present study supports these observations since wounded Hehcotylenchus indicus were incompletely consumed more than any other prey species. Bilgrami and Jairajpuri (1988) also suggested the possibility of unfavourable secretions in Helicoty-
48 Revue Nematol. 12 '!) : 45-49 (1989)

Predalwn by Mononchoides longicaudatus and M fortidens 1 19
lenchus indicus which ehcited meager responses of M fortidens and M longicaudatus besides X amenca-num and Longidorus sp The number of Helicotylenchus indicus, X amencanum and Longidorus sp , left uncon-sumed by the predators also suggests the presence of unfavourable repellent substance(s) in the body of these species of prey The texture of the cuticle of the dory-laims, X amencanum, Longidorus s p , may conform resistance to wounding However, the stnke rate against this group was also generally lower suggesting other defensive mechanisms may operate It may be inferred from the present observations that Hoplolaimus indicus, Tnchodorus sp , and H mangiferae acquire physical resistance in the form of thick cuticle and annulations respectively, Rhabditis sp behavioural resistance in the form of active body movements and vigorous escape response, Hebcotylenchus indicus chemical resistance in the form of toxic/unfavourable secretions, and X amencanum and Longidorus sp , resisting predation partly by physical and chemical means
Ectoparasitic nematodes living in close proximities to predacious nematodes may have developed more anti-predation devices These mechanisms as mentioned above may be more effective against dorylaim, nygolaim actinolaim and diplogastend predators since most of these predators possess piercing type of feeding apparatus and feed by punctunng the cuticle of prey However, mononchs overcome the resistance of prey of different kinds to a larger extent as these do not puncture the cuticle of prey but engulf them whole The ectoparasitic nematodes are therefore more vulnerable to predation by mononchs This is evident as many species of mononchs have been found containing species of Hoplolaimus, Hebcotylenchus, Xiphinema, Hemicnco-nemoides, Rhabditis s p , e t c , besides other tylenchs, dorylaims, rhabditis and mononchs entire in their intestine (Bilgrami, Ahmad & Jairajpun, 1986) Mohandas and Prabhoo (1980) also found intact Tylenchorhynchus nudus and Xiphinema elongatum in the intestine of lotonchus kherai and / monohystera
ACKNOVt LEDGEMENTS
The authors are thankful to the Chairman, Department of Zoology, Aligarh Muslim L Diversity, Aligarh for providing neccessary laboratory facilities
RFF RFNCFS
Bii c RAMI, A L , AHMAD, I & JAIRAJPLRI, M S (1983) Some factors influencing predation by Mononchus aquaticus Revue Nematol, 6 325-326
BiioRAMi, A L , AHMM:), [ & JAIRAJPLRI, M S (1984) Observations on the predatory behaviour of Mononchus aquations Nematol medit, 12 41-45
BILGRAMI, A L , AHMAD, I & JAIRAJPLRI, M S (1986) A
study of the intestinal contents of some mononchs Revue \'ematol,9 191-194
BiicRAMi, A L & JMR-MI'LRI, M S (1988) Attracuon of Mononchoides longicaudatus and M fortidens (Nematoda Diplogastenda) towards prey and factors influencing attraction Revue Nematol 11 195-202
EsshR, R P (1963) Nematode interactions m plates of non-sterile water agar Proc Soil Crop Sci Soc Fla, 23 121-138
EssER, R P (1987) BiolojTical control of nematodes by nematodes I Dorylaims (Nematoda Dorylaimina) Fla Dept Agnc & ConsumerServ, Nematol Circ, No 144 4p
EssLR, R P & SoBi RS, E K (1964) Natural enemies of nematodes Proc Soil Crop Sci Soc Fla, 24 326-353
GROOI\ERI , P , J^QLES, A & SMALL, R W (1977) Prey selection m Butlenus sp (Rhabditida Diplogastendae) Med Fac Landbouww Ryksumv Gent, 42/2 1559-1563
JAIRAJPLRI, M S & AZMI, M I (1978) Some studies on the predatory behaviour of Mylonchulus dentatus Nematol medit, 6 205-212
MOHANDAS, C & PRABHOO, N R (1980) The feeding behaviour and food preference of predatory nematodes (Mononchida) from the soil of Kerala (India) Revue Ecol Biol Sol, 17 53-60
SHAFQAF, S , BILGRAMI, A L & JAIRAJPLRI, M S (1987)
Evaluation of the predatory behaviour of Dorylaimus stagna-/isDujardin, 1845 (Nematoda Dorylaimida) Revue Nematol, 10 455-461
SMAI L, R W &GROOIAERI P (1983) Observations on the predation abilities of some soil dwelling predatory nematodes Nematologica, 29 109-118
Accepte pour publication le 22 janvier 1988
Revue Nematol 12 (1) 45-49 11989) 49

Inndian J. Nematol. 18(2) : 223-227 (1 f 88)
EFFECTS OF CONCENTRATION AND THICKNESS OF AGAR ON THE PREDATORY BEHAVIOUR OF MONONCHOIDES LONGICAUDATUS
AND M. FORTIDENS (NEMATODA : DIPLOGASTERIDA)
ANWAR L. BILGRAMI and M. SHAMIM JAIRAJPURI
Section of Nematology, Department of Zoology, Aligarh Muslim University, Aligarh-202001, India
20
Abstract : The attraction of Mononchoides longicaudatus and M. fortidens was affected by different concentrations and thickness of agar layers. The maximum attraction took place at 1 and 2 per cent concentration and in agar layers of 4-6 mm in thickness. Agar concentrations of 1 and 2 per cent were also suitable for predation. M. longicaudatus and M. fortidens killed maximum number of Hirschmanniella oryzae and Meloidogyne incognita juveniles at these concentrations. However, the rate of predation remained unaltered in all agar layers. The activity of predators and prey is attributed as the cause of differential response of predators.
Key words : Predator, Prey.
Mononchoides longicaudatus, M. fortidens, Predation, Attraction,
Several studies have been made on the predatory behaviour of mononchs (Nelmes, 1974; Cohn & Mordechai, 1974; Small, 1979; Small & Grootaert, 1983, Bilgrami et al., 1984); dorylaim (Wyss & Grootaert, 1977; Shafqat et al., 1987); nygolaim (Bilgrami et al., 1985) and diplogasterld predators (Yeates, 1969; Grootaert et al., 1977; Small & Grootaert, 1983; Bilgrami & Jairajpuri, 1988). The pre- and post-feeding aggregation of Mononchoides longicaudatus and M. fortidens around a previously injured prey at feeding sites and their attraction towards diiferent prey has been observed by Bilgrami & Jairajpuri (1987a; 1987b). The concentrations and thicknesses of agar govern the activity of nematodes (Wallace, 1969; Azmi & Jairajpuri, 1977) and their attraction (Bilgrami et al., 1985b). Bilgrami e?o/. (1983) attributed decreased activity of predators and prey in increasing agar concentrations as the cause of the decline in predation by Mononclt'js aqiiaticus. Later. Shafqat et al. (1987) also found similar results
while evaluating predation abilities of Dorylaimus stagnalis. In the present work the effect of different agar concentrations and thicknesses was observed on the predation by M. longicaudatus (Khera, 1965: Andrassy, 1984). and M. fortidens (Schuurmans Stekhoven, 1961; Taylor & Hechler), 1966, and on their attraction towards the prey nematodes.
MATERIALS AND METHODS
M. longicaudatus and M. fortidens were cultured by the techniques used by Bilgrami & Jairajpuri (1987b). Hirschmanniella oryzae and the second stage juveniles of Meloidogyne incognita were used as prey. The former prey was isolated fresh from the soil while the latter was obtained from the culture maintained on tomato. Predatory behaviour was observed in water agar of 1, 2, 3, 4, 5 and 6 per cent concentration and 2, 4, 6, 8 and 10 mm thickness. All experiments were carried out at 28 ± 1 'C.

121
224 ANWAR h. BILGRAMI AND M. SHAMIM JAIRAJPURI
Twenty-five preynematodes alongwitH five adult specimens of predators were released in cavity blocks containing water-agar with five replications. The number of prey killed or ingested was recorded after 24 h.
To observe attraction 5.5 cm diameter Petri-dishes were marked at the bottom into three zones viz., inner, middle and outer by drawing two concentric circles of 0.5 and 2.5 cm in diameter. A plastic straw pipe 5 mm high, 5 mm in diameter, with a small piece of filter paper placed at one end (sealed end remained in contact with the Petri-dish) was placed vertically in the inner zone of the Petri-dish. Petri-dish and straw pipe was filled with water agar. Twenty-five prey nema-tode,s were released in the straw pipe and left for incubation for eight hours. After incubation five adult predators were released at the periphery of the middle zone. Twenty replications were maintained. Petri-dishes without prey served as control. The distribution of predators was recorded after 4h. The scores were obtained by summing up the product of the number of worms in each zone with their corresponding weighting factors obtained by dividing the area of the outer zone by that of each of the three zones (Ahmad & Jairajpuri, 1980; Bilgrami etal., 1985b).
RESULTS
Effect of concentration and thickness on predation (Fig. 1) : There was a negative correlation between the number of prey killed and concentration of agar (p < 0.05). Maximum number of H. oryzae and M. incognita juveniles were killed by M-fortidcns (Fig. 1, A) and M. longicaudatus (Fig. 1, B) at 1 and 2 per cent agar concentrations (p < 0.05). The rate of predation by M. fortidens on M. /«cog/7z7fl juveniles remained unaltered at 3 per cent (Fig. 1, A) but on H. oryzae it declined significantly (p < 0.05). Concentrations higher than 2 per cent reduced predation markedly. At 6 per
cent minimum number of H. oryzae and the second stage juveniles of M. incognita were killed by M. fortidens and M. longicaudatus.
The predation by M. fortidens (Fig. 1, C) and M. longicaudatus (Fig. 1, D) was independent of the thickness of agar layer. There was no difference in the number of H. oryzae and M. incognita juveniles killed by both species oi diplo-gasterid predators in all agar layers (p > 0.05). The second stage juveniles of M. incognita were preyed upon more by both predators than ff. oryzae in all agar layers (p < 0.05).
Effect of agar concentration and thickness on attraction (Fig. 2) : M. longicaudatus and M. fortidens were attracted maximum towards H. oryzae and the second stage juveniles of M. incognita at agar concentrations of 1 and 2 per cent. Concentrations higher than these declined attraction of M- fortidens (Fig. 2, A) and M. longicaudatus (Fig. 2, B). Predators were not attracted either towards H. oryzae or M. incognita juveniles at 6 per cent as there was no significant difference in the attraction of predators in the absence or presence of prey (p > 0'.05).
Agar thickness of 4 and 6 mm were most favourable for the predators M. fortidens (Fig. 2, C) and M. longicaudatus (Fig. 2, D) and were attracted most towards H. oryzae and M. incognita juveniles at these agar layers (P < 0.05). Agar thickness of 2 and 8 mm decreased attraction of both species of predators. Minimum number of M. fortidens and M. longicaudatus were attracted towards both species of prey in 10 mm thick agar layers (P < 0.05).
DISCUSSION
Agar concentrations govern the activity (Wallace, 1969; Azmi & Jairajpuri, 1977) while agar thickness affects attraction of nematode (Bilgrami et al, 1985b).

122
PREDATORY BEHAVIOUR OF MONONCHOIDES SPP 225
, r.--a95 y>- ^Wl • ^5 •16 ' ^ incoqnilo )v)
I ' I I I I I
1 2 3 4 5 6
Agor concfnlrolions(Vi)
15.
S. 10.
1 2 3 4 5 6
Agar concentrotions (%)
20.
18.
^ 16.
X - Observed vaiure(t) iftfoonitn |v)
«-Observed values (H oryzae) C
J X X X *
'••-0-i2
V = - 0-03x • 19.82 (M. incognita jvj
r= 0.27 '"v.- 0.06x-18 64 (U-SIYZflS)
4 6 8
Agar Ihichness (mm)
—I 10
20
16
l . " - 0 - 2 3 y=-o.05«-i3.3i'Min£29i!i!a i»)
_ r : - a 2 2 "v---005x-1530(Horvzcel
- I 1 1 1 '-t 2 4 6 8 10
Agor thickness {mmj
Fig. 1. A — Effect of agar concentration on predation by Moiwnchoides fortidens. B — Effect of agar concentration on predation by Mononchoides longicaudatus. C — Effect ef agar thickness on predation by Mononchoides fortidens. D — Effect of agar thickness on predation by Mononchoides longicaudatus.
Bilgraml etal. (1983) attributed lower rate of predation by M. aquatkus in increasing agar concentrations to inhibited activity of predators and prey. The predation by M. longicai datus and M. fortidens and their attraction towards prey is influenced by different biotic and abiotic factors (Bilgrami & Jairajpuri, 1987b; 1988). During present observations different concentrations and thicknesses of agar layers affected predation by M. fortidens and M. longicaudatus and their attraction towards H. oryzae and M. incognita juveniles. The activity of M. fortidens and M. longicaudatus along with the prey nematodes might have
been inhibited by higher concentrations resulting in a fewer predator-prey encounters and hence, could be the cause of lower rate of predation (Bilgrami et at., 1983) and less attraction (Bilgrami et al., 1985b) towards prey. Besides, the dispersion of prey attractants may have also affected by different agar concentrations resulting into diffential predator response. Shafqat et al. (1987) also found higher concentrations of agar unfavourable for predation by Dorylaimus stagnalis.
Bilgrami et al. (1985b) found that the adult Hirschmanniella oryzae were attract-

123
226 ANWAR L. BILGRAMI AND M. SHAMIM JAIRAJPURI
2-35
w 1.80.
I 1-50.
1.20
2-20
200.
\ \
—1 1 1 1 1 1 ) 2 3 « 5 6
Agar concentration (%)
2.351
-
1.80.
1.50.
1.20
B \ •—•M.incoanit
V o—0 H oryzae
• — K \
\ \
' 1 1 1 1 T 1
Agar concentration (•/.)
Z 1.60.
\.L0
\
/ ~ " \
- I 1 8 10
2-20"
200.
1.80-
1.60.
1.40
o/°\.
•-v. 1 1 1-
D
\ o
- r • 1
Agor thickness (mm)
2 4 6 8
Agar thickness (mm)
Fig. 2. A — Effect of agar concentrations on the attraction of Mononchoides fortidens towards prey.
B — Effect of agar concentrations on the attraction of Mononchoides longicaudatus towards prey.
C — Effect of agar thickness on the attraction of Mononchoides fortidens towards prey. D — Effect of agar thickness on the attraction of Mononchoides longicaudatus
towards prey.
ed towards cabbage seedlings in 3 and 4 mm thick agar layers. The attraction of M. fortidens and M. longicaudatus was also influenced likewise by various thicknesses of agar layers. This differential attraction could have resulted due to prey attractants requiring more time to form an optimum perceptible attraction gradient in thick agar layers in compari-sion to thinner ones. The present observations suggest that there exists a correlation between the nematode activity and its surrounding environment.
ACKNOWLEDGEMENTS
The authors are gratefull to the Chairman, Department of Zoology, Aligarh Muslim University, Aligarh for providing necessary laboratory facilities.
REFERENCES
AZMT, M.T. & JAIRAJPURI, M.S. (1977). Studies on nematode behaviour IJf. Locomotion in juveniles of Hoplolaimus indicus and Helicotvlenchus indicus in different concentrations of agar. Indian J. Nematol. 5 : 240-242.

124
PREDATORY BEHAVIOUR OF MONONCHOIDES SPP. 227
BILGRAMI, A.L , AHMED, I & JAIRAJPURI, M S (1983) Some factors influencing predation by Mononchus aquaticus. Revue Nematol 6 : 325-326.
BILGRAMI, A L , AHMAD, I. & JAIRAJPURI, M S. (1984). Observations on the predatory behaviour of Mononchus atqua-ticus Nematol medit 12 : 41-45.
BILGRAMI, A.L., AHMED, I. & JAIRAJPURI, M.S (1985a) Predatory behaviour of Aquaiides thoinei (Nygolaimina : Nema-toda). Nematohgica, 30 : 457-462
BILGRAMI, A L., AHMAD, I. & JAIRAJPURI, M S. {1985b). Factors influencing attraction of adult Hirschmanniella oryzae towards cabbage-seedhngs. Revue Nematol. 8 : 67-75.
BILGRAMI, A L. & JAIRAJPURI, M S (1987a) Aggregation of Mononchoides loijgicaudatus and A:/ fortidens (Nematoda : Diplogasterida) at feeding site. Nemato-iogica (in press)
BILGRAML A.L.& JAIRAJPURI, M S. (1987b) Attraction of Mononchoides fortidens and M longicoudatus (Nematoda: Diplogasterida) towards prey and factors influencing attraction. Revue Nematol. (in press)
BILGRAMI, A.L. & JAIRAJPURI, MS. (1988). Predation by Mononchoides longi-caudatiis and M. fortidens (Diplogasterida • Nematoda). Revue Nematol. (m press).
COHN, E. & MORDECHAI, M (1974). Experiments in suppressing citrus nematode populations by use of mangold and a predacious nematode. Nematol. medit. 2 :
43 53.
GROOTAERT, P , JAQUES, A & SMALL, R.W. (1977). Prey selection in Butlermi sp. (Rhabditida : Diplogasteridae). Med. Fac Landbouww. Rijksuniv. Gent. 4212 : 1559 1563.
NELMES, A J (1974) Evaluation of the feeding behaviour of Pnonchulus punctatus (Cobb.) a nematode predator / . Amm. Ecol 43 : 553-565.
SHAFQAT, S , BILGRAMI, A.L & JAIRAJPURI, MS. (1987). Evaluation of the predatory behaviour of Dorylaimus stag-nalis Dujardin, 1845 (Nematoda : Dorylai-mida) Revue Nematol. (in press).
SMALL, R W. (1979). The effects of predatory nematodes on population of plant-parasi-tic nematodes in pots Nematohgica 25 : 94 103.
SMALL, R.W & GROOTAERT, P. (1983). Observations on the predation abilities of some soil dwelling predatory nematodes. Nematohgica 29 : 109-118.
WALLACE, HR. (1969) Wave formation of infective larvae of the plant parasitic nematode Meloidogyne javamca Nematohgica 15 : 65-75.
WYSS, U. & GROOTAERT, P. (1977). Feeding mechanism of Lahronema tuha^ papillatum. Med Fac Landbouww. Rijk-sumv Cent 42: 1521-1727.
YEATES, G.W (1969) Predation by Mononchoides potohikus (Nematoda : Diplogasteridae) m laboratory culture. Nematohgica 15 . 1-9.
(Accepted for publication : February, 1988)

12S SHORl ( < ) \ I \R \ l ( A.[K)\S 1 1 9
L I T h R A T U R
ABOII -PJ ID , H Z SI COOM^NS, A (1966) Intersexuality in Lotigidorus macrosoma Nematolo^ica 12, 343-344
ANDR^SSY, I (1952) Frcilebendi" Nt-rndtoden dus dem Bukk-CJebiige Annates Hutonco Saturaki Musei nationah\ huni^anci 2, 13-65
ANDRASSY, I. (1985) The genus Plectm Baitian, 1865 and its neaiest lelativcs (Nematoda Plec tidae) Ada Zoolosica Hunganca 31 , I 52
BR/tbKi, M (1963) Materialy do poznania krajowych nicieni z lodzjow Plectm Bastian i Ana-plectui De Cloninck et Sch Stekh (Nematoda, Plectidae) Iragmenta Jaunntica 11, 21-30
CoHN, E & MoRDECHAi, Vf (1968) A case of intcrsexualuy and occurrence of males in Z.on zo'o-••ui ajncanus hemalulugua 14, 591-592
DADAY, E V (1905) Untersuchungen uber die Suliwassei-Mikroiauna Para^uays III Nematoda. Zoolngica 18, 48-71
J A I R \ ] F I R I , M S & SiDDiQi, A H (1964) Intersexuality m Tyleptui ilnatui \emaiolo^ica 10, 182-183
RuHM, W (1956) Dit Nematoden der Ipiden Paraatologische Schnjtenmhe 6, 1-437, SiFiNER, G (1923) Intersexes in nematodes Journal vj Heredity 14, 14/-158
A. L. BiLGRAMi & M. S. JAIRAJPI'RI ') : Aggregation 0/Mononchoides longi-caudatus and M fortidens {Nematoda Diplogasterida) at feeding sites
Studies on predatory nematodes reveal that predation depends on chance encounters with the prey (Yeates, 1969; Grootaert & Maertens, 1976; Small & Grootaert, 1983; Bilgrami et al , 1984). Esser (1963) was of the opinion that excised (cut) prey attracted several dorylaim predators. However, Aquatides thornei (Schneider, 1937) Ahmad & Jairajpuri, 1979 and Dorylaimus stagnalis Dujardin, 1845 did not respond positively towards excised prey but aggregated around prey injured by another predator suggesting some kind of attraction (Bilgrami et al., 1985; Shafqat et al , 1987). A similar aggregation of predators viz., Diplenteron colobocercus Andrassy and Labronema vulvapapillatum Loof & (jrootaert, 1981 around prey was also observed by Yeates (1969) and Wyss & Grootaert (1977) respectively. No aggregation of mononchs around wounded prey has been recorded (Bilgrami et al , 1984). In the present work observations on Mononchoides longicaudatus (Khera, 1965) Andrassy, 1984 and Mononchoides fortidens (Schuurmans Stekhoven, 1961) Taylor & Hechler, 1966 were made to observe their attraction and aggregation at feeding sites.
Mononchoides longicaudatus and M fortidens were cultured in 1 % water-agar using Rhabditu sp. as prey. Five mg of infant milk powder {Lactogen) was spread over the surface of agar to provide for growth of bacteria which served as food tor the prey nematodes. The predators were transferred to fresh culture dishes containing prey as and when the population of the predators started declining. Both the predators were cultured separately. The culture dishes were kept in the dark when not under observation. Observations on the aggregation of M. longicaudatus and M. fortidens were made in 5.5 cm diameter Petri-dishes con-
') Stction of Nematology, Department of Zoology, Aligarfi Muslim University, Aligarh-202002, India

1 i M
1 2 0 MIUR I C ( ) \1 \U \ I ( \ I 1 ( ) \ S
taming a 2 mm thick layer of I'^c water-agar Fifty specimens of prey nematodes {Rhabditu sp ) were released at different places on the surface of agar Iwtn ty six adult predators (thirteen each of male and female) were then leleased m the Petri-dishes Fi\e dishes were examined every half hour by inverting Petri dishes o\er the stage of the stereoscopu microsco[ie The point when a [)rey was killed was marked at the bottom of the dish by a circle 2 mm m diametc r The whole area of the ciicle is referrc d to as a leeding site During observations the sites formed pre\iously were looked at first and then the new feeding sites were recorded The experiment was terminated after five hours The mean \alues were obtained from ten observations which were replicated five times I he time taken b\ the predators to form a feeding site, the total duration of the feeding site (i e , until all the predators dispersed from the feeding site), the time of actual feeding (i e , until the prey wa' completely finished) and the time of their aggregation after finishing off a prey (post-feeding aggregation) were calculated from the records Similarly, the number of predators aggregated before and after finishing the prev at the feeding site and those actually feeding lAere also recorded The experiments vvJere earned out at 2^1+1° Both predators were tested separately
Both species of predators viz , ,W lom^itaudatm and M fortidens aggregated around the injured or killed prey but there was no obvious change m their pattern of aggiegation with sequences of feeding sites Up to 16 feeding sites of M lons,icaudatus and 15 of M jortidens were seen on a dish with an average 4, range t-b predators at any one site However, all predators present at the feeding site were usually not feeciing, the average number actually feeding (three) was less than the total number of predators present there (tour) Post-feeding aggregation of predators at the feeding site was common to both species and on an average three predators per feeding site were observed The duration of post-feeding aggregation was almost the same for both species of predatois i e , about 50 mm but time at the feeding site (mean 1 lb min, range 84-148 mm. SF + 10 09 for M lom^icaudatus and mean 104 mm, range 72-156 mm, SF ± 9 ^8 for M Jortidem) and actual leeding time (mean 56 n i n , range 35-77 min, SK + 5 76 for M lon^icaudatus and mean 48 mm, range 24-66 mm, bF> ± 4 07 foi V/ fortidens) differed ( p < 0 05), being longer tor M lonsJtcaudatus However, the number of feeding sites formed and the average tirn • taken to form (approximately 20 min) each feeding site was almost the same for both predators
A thornei and D sta^nalis did not respond positively towards excised (cut) prey bu' aggregated around an injured prey during teeding which led to the speculation that there was some kind ot attraction towards prey (Bilgrami et al , 1985, Shatqat et al , 1987) During the present study M loni^icaiidaths and M fortidens showed aggregation (i e during actual leeding) around prey injured or captured bv another picdator 1 his phenomenon ot aggregatiC/D by M loni>icaudalui and \i fortidens during leeding is difficult to explain at present but

SlIORI ( ( ) \ 1 \ U M ( \ I 1 ( ) \ S 1 2 1
may bf duf to (i) a response ot predators to seeretions (attra( tants) released by the prey (ii) secretions from the predators at the time of feeding or (iii) secretions of both prey and predators. Sueh aggregation has not previously been observed in any of the predatory nematodes belonging to the Order Mononchida viz., Pnonchulus pundatus, Mylonchulus dmtatus and Mononchus aquatuus (Nelmes, 1974; Jairajpuri & Azmi, 1978; Bilgrami et a' , 1984). The pheiiomt non of post-ieeding aggregation has not been reported in any predatorv nematodes where aggregation has been reported during actual feeding [Diplenteron colobocercm, Labronema vulvapapillatum, Aquatides thornei and Dorylaimus stagnabs) (Yeates, 1969; Wyss & Grootaert, 1977; Bilgrami et al , 1985, Shafqat et al , 1987). During our observations the post-feeding aggregation of A/ longicaudatus and M Jortidem seemed to be due to a lingering effect of the prey/predator secretions (attractants). M. jortidens appeared a stronger predator, devouring its prey faster than M longuaudatus, and, consequently, feeding sites were maintained for a shorter period. Further observations are needed to ascertain the mechanism involved.
REFERENClvS
BiK.R^Mi, \ 1- , AHMAD, 1 & JAIRAJPIRI, M S (1984-) Observationb on the predatory bi.-hd\iour ot Mononchus aquatuus \ematologia Mediterrama 12, 41-45
BiiGR\M',A 1. , \HVIAD, t & JMRAJPI Ri, M S (1*^85) ?TvddUm hi^hdwoux oi Aifuahde', thornei (.\\t;oldimina- Dorylaimida) Sematoloi^ica 30 (l'J84), 457-462
EiSER, R P (!963) Nematode interactions in plates of non-btenle watcr-agar Pro, eedinqs of Sod and Crop Science Society of Florida 23, 121-138
(jRooFAtRT, P (4 M' ERTENs, D (1976) Cultivation and iik cycle of Mononchus aquaticus Xemuloluiiua 22, 173-181
JMRAJPCRI M S siA^Mi.M I (1978) Some studies on the predatory behavioui o) .V/y/ont/iii/Mj dentatu) \irnatolos,ia \iediterranea 6, 205-212
.NFLMFS, \ J (1974) Evaluation of the Itedin^ behaviour oi Prwmhulw, pumtatus (Cobb), A nemaiode predator Journal of Animal Ecology 43 , 553-565
SHAK^AI , S , BILCRAMI, A L & JAIRAIPLRI, M S (1987) Evaluation ot the predatory
behavioui ol Dorylaimus sta^nahs Dujardin, 1845 (."viematoda Dorylamudai Revue de .\'emat(iloi>u 10, 455-466
.SMALI , R W s GROOI'VERT, P (1983) Observations on the predation abilities ol some soil dwelling predatory nematodes \ematologica 29, 109-118
WYSS, U & GRtiDPAERi, P (1977) Feedmg mechanism of Labronema bulvapapillatum Mededel-tn^en Fatultnl Landbouuwetenschappen Rijksunwersiteit Gent 42, 1521-1527
\ ' tMES, (j W (1969) Predaiion b \ Mononchoides polohikus (Nematoda Diplogasleridae) m laboiatorv culture Sematoloi^ua 15, 1-9
R. A. Flow RIGHT'): Rice callus for the monoxemc culture of root ectoparasitic
nematodei
Successes m the monoxenic culture of root ectoparasitic nematodes on plant callus tissue (Khera & Zuckerman, 1962; Krusberg, 1961; Townshend. 1974; Khera et al , 1969 and Wood, 1979), prompted studies which showed that
') CAB International Institute ol Parasitology. 395a, Hatfield Road, St .Albans, Herts, England

128
1 2 2 SHORl C OMMUNK ^IIONS
Tylemhorhynchui annulatus and Helicotylenchus crenacauda feed on rice callus tissue. The present work clarifies these studies and examines the influence of callus explant derivative and the concentration of 2,4-dichlorophenoxyacetic acid (2,4-1)) on callus growth and nematode reproduction.
Hulled rice seed cv IR 36 was surface sterilised (sodium hypochlorite, 1% available chlorine for 20 minutes), rinsed in sterile distilled water and placed on water agar. After 18 hours embryos were excised from half of the seed and the remainder were allowed to germinate. Embryos and 1 cm excised root tips were transferred to 1 % agar slopes of Gamborgs B5 medium (Gamborg et al., 1968) plus CaNOs, 440 mg/L; NH4NO3, 1650 mg/1 and glycine, 2 mg/1 with 2% glucose, supplemented with 2-100 mg/1 2,4-D. and incubated in the dark at 2 5 ^
Nematodes, washed from around rice roots, were extracted by wet sieving and concentrated in sterile distilled water. Batches of 120 nematodes were surface sterilised (0 .1% malachite green for 15 minutes) in sterile watch glasses and rinsed 5 times in sterile distilled water. Nematodes were clumped in the base of the watch glass by gentle swirling and liquid removed using a sterile Pasteur pipette.
Forty eight days after initiation each callus was inoculated with 30 nematodes, which were placed one at a time directly onto the callus. Of 12 calluses for each treatment, 4 were inoculated with T. annulatus, 4 with H. crenacauda and 4 were left uninoculated. After a further 105 days nematodes were extracted from the calluses and counted.
Root tip and embryo callus was first observed after 7 days. Callus development occurred in the range of 2-15 mg/1 2,4-D and was significantly faster on root tips: however, after 153 days callus weights from both jurces were the same (Table 1). At 10 and 15 mg/1 2,4-D, calluses had a watery, smooth appearance while at 2 and 5 mg/1, calluses were hard and friable containing roots and root hair like structures.
Tylenchorhynchus annulatus reproduced on callus from both sources and nematodes from high and low concentrations of 2,4-D remained infective when re-inoculated onto rice roots. The multiplication of T. annulatus was variable within treatments and was not related to callus fresh weight. On embryo callus multiplication occurred at 5, 10 and 15 mg/1 2,4-D and numbers were low at other concentrations (Fig. 1). The highest number recovered from a single embryo callus was 9750 at 15 mg/1 2,4-D. On root tip callus, there were no clear differences in numbers between treatments but the highest number developed at 10 mg/1 2,4-D. There were no significant differences between nematode numbers on embryo and root tip callus.
Helicotylenchus crenacauda did not multiply on any callus though a few, apparently healthy nematodes were recovered from each treatment.
I thank Mr. A. Macarthur for his technical assistance and the U.K. Overseas Development Administration who funded this research.

•C
Indian J. Nemalol. 18 (2) : 357-360 (1988)
Response of Hirschmanniella oryzae towards different sources of attraction
ANWAR L. BILGRAMI, IRFAN AHMAD and M. SHAMIM JAIRAJPURI
Section of Nematology, Department of Zoology, Aligarh Muslim University, Aiigarh-202001, India
Several studies have been made on the attraction and feeding behaviour of plant-parasitic nematodes (Wyss, 1971; Doncaster & Seymour, 1973; Azmi & Jairajpuri, 1977; Prot, 1980; Bilgrami et al., 1986a\ While observing the attraction of juvenile stages of Hirschmanniella oryzae Bilgrami et al.
(1986bj found fourth stage juveniles most responsive towards cabbage seedlings. Weiser (1955) and Lavallee & Rohde (1962) found that the attraction by the roots was proportional to their rate of growth but roots killed by healing loose their attractiveness (Linford. 1939; Bird, 1962; Bilgrami ct ah, J 985a). Adult

130
358 SHORT COMMUNICATIONS
H. oryzae were attracted mostly towards cabbage seedlings as compared to other plant roots that were tested (Bilgrami et ai, 1985b) and factors i.e., agar concentration, agar thickness, age of roots, incubation period of roots, starvation of nematodes etc., affected attraction (Bilgrami et al., 1985a). Recently, Bilgrami et al. (1986c) found cabbage and tomato roots most attractive while chilli and radish least attractive to adult Xiphinema basiri. In the present work the responses of adult and juveniles of H. oryzae were tested towards different sizes of the source of attraction.
Adult and juvenile stages of Hirschmanniella oryzae were isolated from soil collected from paddy fields from Village Jamalpur, near Muslim University, Aiigarh by sieving and decantation and modified Baermann's funnel techniques. The intensity of attraction of adults and their juveniles towards different sources of attraction was tested in 5.5 cm diameter Petri-dishes containing 0.5 percent water agar. The source of attraction of 2, 4, 6, 8 and 10 mm of diameter was obtained from previously incubated agar with 4-6 day old excised cabbage roots for 12h. The diflferent source of attraction was placed at the centre of separate Petri-dishes by replacing the same amount of agar. A portion equal to that of the source to be tested was drawn at the bottom of the Petri-dish (i.e., with 2mm source the diameter of the inner circle was 2mm and with 4mm it was 4mm ). This circle containing attraction source is referred to as inner circle. Another circle, referred as middle, was drawn concentric to the centre of the Petri-dish at a distance of 1 cm from the periphery of the source of attraction to be tested. Thus, the diameter of the middle circle varied with different sizes of the source of attraction keeping the distance of nematodes inoculated from the source identical in
each case (i.e., with 2mm source the diameter of the middle circle was 2.2 cm and with 4mm it was 2.4 mm ). Five nematodes were inoculated at different places at the periphery of the middle circle. The observations on the distribution of nematodes were made after 4hrs. Each experiment was replicated ten times and carried out at 28 ± 1°C. Control experiments were conducted without using previously incubated agar with roots. The area within the inner circle is referred as inner zone while that between the inner and middle circle as middle zone. The circumferal area of the Petri-dish comprised the outer zone.
The attraction of the juvenile stages as well as adults depended on the size of the source of attraction as it increased with the increase in the size of source. The optimal size varied for each stage including the adults. All juvenile stages and adults of H. oryzae were unresponsive towards 2 and 4 mm sources (p > 0.05). The attraction of the second stage juveniles was maximum towards the source size of 10 mm (p < 0.05). Least attraction of this stage was observed towards 6 mm attraction source (Fig. 1, A). Like the second stage the 10 mm attraction source was also optimal for attraction of the third stage juveniles of H. oryzae (Fig. 1, B). Minimum number of third stage juveniles were attracted by 6 mm diameter agar disc (p < 0.05). Fourth stage juveniles were attracted most by 8 and 10 mm sources (Fig. 1, C). The differences between them were insignificant (p > 0.05). The 6 mm source attracted least number of nematodes. The adult H. oryzae when placed with different source sizes were attracted most by 6 mm or larger agar pieces (Fig. 1, D). The number of nematodes reaching the inner zone was found significantly more than that the number reaching the same region containing 2 or 4 mm sources (p < 0.05).

131
SHORT COMMUNICATIONS 359
•° c
o " e 0, i. °
o
-o c o o
li 2 0,
a* i: •o c 0 o
01 O
•D o a £ Z
•51 • 4-
•3-
•2-
.1
-1--2-
-3--4. -5-
o—Dinner zone / •—•middlezone / A ,—xouterzone/o '^
\ .
•5 •A
•2
• 1 .
-1-1
-2
-3^
-A-
^ .
/
8 10
"X.
y
\ .
•5' •4-
•3.
f2-
• 1 .
-1--2 -J
-5-
•5-•A-•3.
•2-
• 1.
- 1 --2.
-3-1 -4
-5,
1 I 6 5~15 Source of attraction
(mm diam)
1 I 1 r— 2 A 6 8 -^
^yh
y~.
"na 1 I 1
2 \ 6 Source ofattraction
(mm diam) Fig. 1.' Effect of size of attraction source on the attraction of H. oryzae. A — Second stage
juveniles; B — third stage juveniles; C —fourth stage juveniles; D — Aduhs.
The attration of adults and juveniles of/T. oryzae was affected by different sizes of the source of attraction. No response of adult and juveniles towards 2 and 4 mm agar pieces suggests a correlation between the size of the source and dispersion of attractants. It could be possible that small sources of attraction took more time (more than the period of observation) to form an optimum attraction gradient in the surroundings in comparision to the larger ones or have failed at all to
develope an optimum attraction threshold level. Besides, the differential responses of H. oryzae adults and juveniles may have also resulted due to their sensitivity to percieve stimuli as all nematodes may not have the same degree of perception which can vary from individual to individual (Prot, 1980).
We are thankful! to the Chairman, Department of Zoology, Aligarh Muslim University. Aligarh for providing necessary laboratory facilities.

^2
360 SHORT COMMUNICATIONS
REFERENCES
AZMI, M.I. & JAIRAJPURI, M.S. (1987). Nematologica 23: 119-121.
BILGRAMI, A.L., AHMAD, I. & JAIRAJPURI, M.S. (1985a). Revue Nematol. 8 : 67-75.
BILGRAMI, A.L., AHMAD, I. & JAIRAJPURI, M.S. (1985b). Revue Nematol. 8 : 265-272.
BILGRAI, A.L., AHMAD, I. & JAIRAJPURI, M.S. (1986a). / . Nematol. 15 : 209-216.
BILGRAMI, A.L. AHMAD, L & JAIRAJPURI, M.S. (1986b). Helmintho-logiali : n9A22.
BILGRAML A.L., JAIRAJPURI, M.S, & AHMAD, I. (1986c). Indian J. Nematol. 16 : 60-64.
BIRD, A.F. (1962). Nematologica 7 : 275-287.
DONCASTER, C.C. & SEYMOUR, M.K. (1973). Nematologica 19 : 137-145.
LAVALLEE, W.H. & ROHDE, R.A. (1962), Nematologica 8 :252-260.
LINFORD, M.B. (1939). Proc. Helminth. Soc. Wash. 6: 11-18.
PROT, J.C. (1980). Revue Nematol. 3 : 305-318.
WIESER, W. (1955). Proc. Helminth. Soc. Wash. 22-.106-112.
WYSS, U. (1971). Nematologica 17 : 501-507.

0 ^
PREDATORY NEMATODES
M. SHAMIM JAIRAJPURI & ANWAR L. BILGRAMI
DcparLmcm of Zoology,
Aligarii Muslim Umversity, Aligarh-202002, India.
Introduction
The soil-inhabiting nematodes dominate over all other animals, botli in numbers and species. Tliey mainly fall under four groups based on their feeding t)'pcs-Llic micro-phagous which feed on microorganisms, the saprophagous obtaining food from ilie dead and decaying organic matter, the phytophagous feeding on plants and the prc-daccous which icill otlier organisms for food. The predatory nematodes fall under tlu-ec categories depending upon their mode of feeding and the type of feeding apparatus. The first type are lliose which feed by cutting the body of prey and tlien sucking its body contents as they arc unable to engulf intact preys. These prcdatois possess' small but well developed buccal cavities and belong mainly to the order Diplogasterida (Fig. 1, F). The second type of predators are tiiosc wi,.ch feed by a combined action of cutting and sucking as well as at times, engulfing a prey whole. The latter group of nematode predators which belong to die Older Mononchida possesses a strong and comparalJvely larger buccal cavity provided widi teeth,
NlIMARJDi-. BIO-CON'nU)!, ASPIiCl'S &PROSPEaS(EJs.-M S Jjir.ijpuii. M"f/I Alain .t: 1 AhnuJ) CUS Publiihcrs & Dismbuiois, Delhi India. 1990

0. a I 3^
A- Mononchus.B-Dorvlaimus, C-Aquatides. D-Actmolalmus E-5^inurg^ F-Mononchoides
FIG. 1.
DIFFERENT TYPES OF PREDATORY MEMATODES

1 ^ t f:
A/ S Jairajpun & A L Bd^ranu 97
tooiJi and denticles (Fig 1,A) In fact ihc entire group of mononchs is prc laceou^ The tluid group is of those which feed only by puncturing tlic cuticle of prey and then suck their body contents The feeding apparatus of this letter gioup i e belonging to the Suborders Dorylanmna, Aphelenchina and Nygolaimina is of piercing and sucking type (stylet/s})ear/mural tooth) (Fig 1, B,C & E)
The possibility of using predatory nematodes Si ccially the mononchs for checking populations of plant parasitic nematodes in soil was speculated by Cobb (1917) while making cbscrvalions on their biology and predatory ability Later, Cobb (1920) and Stciner & Homlcy (1922) suggested the use of Clarkus papillaius in controlling populations of nematodes in sugar beet fields However, Thome (1927) considered that the specie, of mononchs present in sugar-beet iulds are usually insufficient to bring about encouraging results as far as control of phytoparasiuc nematodes was concemc l Cassidy (1931) concluded that under suimble condiuons lotonchus brachylaimus might partially control populations of pest nematodes Christie (1960) suggested lliat the use of predatory nematodes should be assessed for detenmning tfie piacticability of their use in the control of plant parasitic nematodes Further studies in this direction were made by Mulvey (1961), Esscr (1963), Esser & Sobers (1964) and Rntcr & Laumond (1975) The nature and source of food of a majority of mononchs has not yet been determined Of those known, lliey feed on a variety of soil micro organisms including nematodes (Banage, 1961, Arpm &. Kilbcrtus, 1981). Accoiding to Webster (1972) and Jones (1974) non-specific predators like mononchs exert only partial conuoland tlie possibility of tlicsc being successful agents of biological conlrol aie remote Nclincs (19 74) compared iJie predation abilities of/'riOAic/ii./uj punctatus with those of prcdaiory arthropods (Solomon, 1949 , Holling, 1966) AccordingtoCohn&Mordcchai(1974)A/>/onc/i«/uj sigma^rw; predates upon the citrus nematode, 7y/enc/!u/i« semipeneirans and when the predator density IS high the population of r penefra^j IS low Consequent, upon these observations research o" predatory mononchs gained some impetus and investigations were made on tlicir life cvcle (Macrtcns, 1976), prcditory behaviour (GitxiUcrt&Maertens, 1976, Jairajpuri&A/mi, 1978, Bilgramier al, 1984), ultrastruc-ture and function of feeding apparatus (Grootacrl & Wyss, 1979), prcdauon abilities (Small, 1979,Small AGrwucit, 1^83), effect of mechanical stimulation and crowding on prcdauon (Azmi & Jairajpun, 1979), factors innucncing predjtion (Bilgrami er a/, 1983 & 1984, Nclmes, 1971), cannibalistic tendency (Azmi & Jairajpun, 1978, Bilgrami & Jairajpun, 1985a) and iiiu a jxxific interactions (Bilgrami et al, 19^8)
The predatory poicniial o) ncinatodi.s belonging to suborders Dorylaimina, Nvgolaimma and Aphclciiclma has not >ci been fully explored Cobb (1929), Thome (19^0 c*L 1939) and I iniord ^ Oliviira (1937) reported se\jral species of AponelaimiLs Nygolaunus, Sec onLina Labronenia Dorylaimus and Aciinolaimus

0 0
98 Predatory Nematodes
as predaceous Esscr (1963) ai' o observed siKcies of diesc genera besides (iiosc of Carcharolaimus to be predaceous Many species ol Seinura arc prrdatoty in nalurc feeding upon different t)pes of prey (Linford &01iv'era, 1937, Hechlcr, 1963) Wyss &Groolaert(1977) and Bilgramier al, (1985a) were the first to giv: detailed accounts of the predation abiliticsof dorylaim and nygoiaim predators, vi? , Lafcronerai vul-vapapillatum and Aquatides thornet respectively Recently, Shafqatera/ (1987) have studied the prey catching and feeding mechanisms inDorylaim'^s stagnali^ and factors influencing its predation It seems possible tJiat these groups may possess more species which could be predaceous in nature particularly tliose belonging to Cudorylaimus, Mcsodorylaimw;, Aporrclaimus, AporcelaimeUiis, Laimydorus, NygolatmiiS and Stcioncnui 1 lie dorylaim predators aie omnivoioiis asllicy jl>o feed on algae and fungi besides feeding on nematodes (Hollis, 1957, Ferris, 1968, Wood, 1Q73)
The d.plogasterid predatois arc also iniixirtant and may prove to be promismg agents of biological control, but only furtlicr studies can confirm tiieir utilit} The studies which have been carried out so far on Diplenieron colobocercus,Builerius sp , B degressei, Mononclwides fortidens and A/ longicaudatus C eatcs, 1969, Grootaert et al, 1977, Small & Grootaert, 1983, Bilgiami & Jairajpun, in press) suggest tliat they are quite active animals possessing great predatory abilities as compared to other group of predators The nematodes belonging to Monohysterida, Bathyodon-tina and Actmolaimoidca arc also perhaps predaceous as is evident from the struc turc of their feeding apparatus In the following a review of (he work that has already been done on tlic predatory ncmatodi-s is given as to assess tlieir predator) potential and to explore the possibility of thtir use in th' bio control of plant paiasitic nematodes
FEEDING APPARATUS
7/it' mononclis (fig 2, A & B) Though, only some species of other nematode groups are predaceous, all s[)ecics of tlic Oidcr Mononthida feed exclusiNcK by picdalion and hence may be of considerable importance to agriculture The mononchs gcneially have large stout bodies, a strongly sclcroti/cd usually barrel shaped Iceding apparatus (TKO known as sioma or buccal cavih) consisiing of two cls of three plaies (one dorsal and two subvenuals) each The alienor set is more dc\clopcd, lan'c and \criicall) pliLcd asco.npirnl lo the [xisierior ct which is small, oblique and cmtvdJcd in the ankrior p III ol iK'sophai'us The CIOIMI pi ilc ul the VLrticil ct i l ,vaysKusa l.irgc s'harpK poinkd U oth while ihe sulnuuril walN usuall) hear either similar or smaller tooth, teeth or dentii.les whose number and arrangemenl is greatly \an.ble

.1 o rf i 61
MS Jairajpuri <i. AL Ddgraru 99
The muscles in llie cci'lialic region iJial help in catching llic prey arc of two types The labial muscles arc a set of six muscles attached postcnorly to the oesophagus Towards the anicnor region thc^e are bifurcated and each band is connected to a diffeient lip In this way each labial muscle can control two adjacent lips and each hp IS conuoUed by two diffcreni muscles The outer set is of stomal muscles which are attached to the vertical walls posterior to the denticles. Two of these TIUSCICS are subdorsal, two subventral and four sublatcral. The buccal cavity with its associated muscles forms the trophicoscnsory organ of the mononchs As soon as contact wit]i iJic prey is established, tlic labial muscles coulract and pull llic lips outwards and backwards so tliat tlic mouth and the vesubule arc wide open (Coomans &. Lima, 1965). The buccal armature helps ip oitching the prey, puncltunng its body and engulfing It eitlier intact or by cutting it into pieces. The food consisting of either intact prey or Its internal body organs or body fluids is sucked into tfie oesophagus by repeated actions of Uic stoma. The oesophageal muscles and the hydrostatic pressure of the body help to open and close the lumen of tlie oesophagus and push the food through llic oesophagus into the intestine (Roggen, 1973)
The dorylairns (Fig 2, E) Dorylaini predators can not engulf or devour the entire prey or cut its body into pieces bcxause of their fccd.ng apparatus which is merely of tlic piercing and sucking tjpc Widi the help of ihcu- dagger-shaped odonlostyle (stylct/si)car) they puncture the body wall of the prey and suck its body contents The feeding apparatus, consisting of a vesabule, guiding ring, guiding sheath, odonloslyie and odonlophore p<-rform this act The odoniostyle bcs at the base of the guiding apparatus o\crlying tlie odonlophore The odonlophore is in conti-nuily with llie ocsopluj'eal lumen and is usually surrounded by an ellipsoidal swelling.
In typical dorylairns the odontost)le is axial in position but in n>golaims it is non axial and is called mural tootli The former has a dorsal appcrtuie and groove but tJie later docs not have an opening or groove. The prey contaiis (semi solid, paniculate) pass through iJic odoniostyle of dorylairns while it is suckixl up by t)ie combined action of tlie guiding shcaih (pharynagcal cavity) and the bas il part of '* e stoma (tclostonc or odonlophore) in nygolaims (Fig 2, F)
The aciinolatms Inactinolaiins the vestibule is reinforced with pLue-like or ribbed basket like stnicturcs which nia^ li:qucntly bo accompanied bylaige onchia with or wiilunii duilkks 1 his addition.il inn Uiirc iiu) 1-c licl(ifiil in killing the pn ) and cuUing o[\:n Its txjdy wall etc llucc SLIS of specialized muscles dialaior butcac, protractor and retractor muscles aic atUiJicd to the feeding apparatus of aLiinobini nematodes and bring about ils lorwcU J and backwaid movcmcnis during feeding

lOO Predalory Nematodes
1c8
{Mononch )
Prwnchulus ( Mononch )
Lab ronema (Doryl aim ) ( NijCjolaim )
FIG.:
FEEDING APPARATUS OF PREDATORY NEMATODES

MS Jairajpun <SLAL Dilgrami 101
139
Thalassoqenus ( Pelagonematid )
Paractinolaimus (Actinolaim )
I ronus {EnopUd)
Mononcho ide 5 iDiplogastend)
RG 2. (Contd)
FEEDING APPARATUS OF PREDATORY NEMATODES

a
102 Predatory Nematodes
The aphdenchs The predatory feeding is known lo occur in the species of Seinura These nematodes possess a small narrow spear, with a lumen but witliout basal knobs, supported by muscles for it5 forward movement dunng feeding. The spear is piercmg aiid suckmg type. Once the body of the pre> is punctured, the secretions of the oesophageal glands of the prfdator are poured into the prey with llic acuon of the powerful median oesophageal bulb, i" o prey is paralysed almost instantaneously and its body contents are sucked by the predators. The median oesophageal bulb helps the ingesaon of prey contcnLs by creating a suction force
Ihedtplogasterids (Fig.2, H): Species o( Mononchoides( =7ridonius) arc excellent example of predators in tins group of nematodes Though, they possess a comparatively small buccal cavity but it is well armed with teeth of different sizes which are located at different positions (Jairajpuri et at, 1973) The feeding apparatus of diplogaslcrid predators is of cutting and sucking type and called stoma or buccal cavity The mouth of tlicse predators is circular with 6 pairs of 'r-ugue' and 10 labial papillae, lips arc inconspicuous and may be fused The buccal cavity is made up of components similar to otlier rhalxlitid nematodes It may be divisible into a wide, strongly sclcroti/xd anterior part and a comparatively narrower, longer and lightly sclerolizcd hinder part. The chcilorhabdions, which arc antenad in position and fonn cheilostome of tlie stoma/buccal cavity, arc undivided (unlike otJier rhabdiuds) and appear to protrude outside tlie lip region. Pro, meso- and inctii-rhabdions form tlie middle part of the buccal cavity. Tlie pro- and mesorhabdions are strongly sclerouzed. The dorsal meta rhabdions arc heavily scleroUzcd and provided with a claw-like tooth, having its apex directed anteriorly This claw-like tooth situated on tlie dorsal meta-rhabdion is called dorsal tootli which is movable and hollow from inside. It receives a duct from the dorsal oesophageal gland which opens in the region of the buccal cavity The large dorsal tooth seems to be tlie main killing weapon It is also resixinsible for the ejccuon of secretions from the oesophageal glands for tlie purpose of cxtra-corporcol digestion The subvcntral walls opposite to the dorsal tootli bear small, rose-thorn shaped icctti while the narrower hinder part of tlic buccal cavity bears a small tooth at ils base Some species also possess denticles which help in grinding ilie food particles Tclo rhjtxlions arc weakly sclerotized, situated at llie posterior end fonning tlie telostoine of tlie buccal cavity
Oilier predators (Fig 2, C,D, i G ) Tlie prcdalors like r/ia/a550^?;iuj (Pclagneinatoi-de.i) possess a buccal eavii) .uid arniatiirc which su[)erhcially icscmblcs that of a mononch (Ortoii WiUi.ims A. Jairajpuri, 1984) Ihcy may ptrhaps have similar feeding habits (convergence) The enoplid iicmatcxlc,/ronii,f and other rcLucd genera (van dor HciJcn, 1974) have three sharplv pointed icetli .vhich act as npi'ini; organ

41
MS Jairajpun i' AL Bilgranu 103
for teanng open Uic Ussucs of the prey The large cylindrical pharyngeal (oesophageal) cavity acl5 as sucking organ Well developed muscles are associated wiLh the feeding apparatus m members of Ironidae
PREY CAT CHING AND FEEDING MECHANISMS
The prey catching and feeding mechanisms of the majonty of predators belonging to different groups of nematodes may conveniently be divided into five more or less distinct phases (Fig 3) viz , encounter with prey, attack response, attack, sahvatwnlextra-corporeal digestion and mgestionlf^edmg (Bilgrami & Jairajpun, in press)
Predator-prey encounter This is an important factor which miuates prcdauon and most hkely governs the rate of prcdauon These encounters largely depend upon chance. Increase in encounters also increases rate of predaLion The dorylaim predators, Labronema vulvapapillatum, Aquatides ihornei and Dorylaimus stagnalis arc attracted towards excised prey (Esscr, 1963) and aggregated at fecdmg sites around the prey which has been injured or captured by another predator (Wyss & Grootacrt, 1977, Bilgrami et a! , 1985, Shafqat^r a!, 1987) These observations have led to the speculation that some kind of chemo-attraction was involved m predator-prey encounters The diplogastend predators viz , M longicaudatiis and M fortidens are able to perceive prey ailraclants and respond positively and significantly towards live and excised frce-living, ecto and endo-parasitic nematodes and bactena (Bilgrami & Jairajpun, 1988) The pre- (dunng feeding) and post-feeding aggregauon (after the prey is completely consumed) of these predators and that ol D colobocercus (YoJiLS, 1969) h.is also been observed at feeding sites This suggcsLs more iJia/i a casual role of chemo attractants m establishing predator-prey encounters, besides merely the chance contacts The attraction and aggregation, of all these predators seems to be in response to prey secretions/atiractants.
The chemo attracuon m dorylaim and nygolaim predators is rather poor as the nematodes take quite a long time to reach their feeding sites and also aggregate only m fewer numbers 2-4 indi\idua!s) Though positive attracUon is evident in these predators, it occurs only v. hen the prey nematode is artificialK excised (Esscr, 196'i) It, therefore, shall have liitle impact on tlic actual predator prey encounters as tlic response, if it has to be signillcancc, shoula be towards live ard intact prey like the diplogasicrui predators whith arc aitncicd towards live prc\ besides excised ones I hey also aggregate ralJicr quickly, in large iiuinl>crs (up lii 10 ndividu iK) The ability ol diplogastend predators to rcs[X)nd towards tJie aiLracLinLs of live prey increases signifiCcUitly tlie predator prey cneountcrs In Inononehs the predaior-prc) encounters depend onl> on chance Tlie absence of chi mo altraeiion in

104 Predatory Ncnialodfi
>'
F
t
Lip contect with prey
^ I .
1 Probing of 1 prey
\
Is I
t
Acceptable contour and texture of prey cuticle
Random nnovemen of predator
1 \ \ 1 1
1
Pos^ feeding aqgregotion "* -
I I \
\ \ \ x
Aggregation of other predators
""-1
Feeding terminated |
^'^A
<:[[__
Lf..,., ^
Entire preyl engjlfpd \
k
\
Feeding/ ingestion by cutting or sucking
A 1
1
1 '
1 1
\ I
Escape of prey
+
Salivary/ oesopf-aqeol secretions
A
«'« _ ; 1 ] ' 3 1
Attack by pr eckitor —
si-
1
stylet thrusting/ punr t i innn/ Slit ting prey cuti :le
""--
Cuticle perforation
/ 1,
FIG 3.
SCHEMATIC nEPRESENTATION OF PnF-Y CATCHING AND FEEDING MECHANISM

4 -s
MS Jairajpuii &AL Dilgranu 105
mononclis is compensated to a great exlcnl by Ihc activity of tJicsc predators and their efficient prey catching mechanism.
Tlie dorylaim predators perceive prey attraclanls only from sliorl distances (Ess<.T, 1963) and their perception level varies from species to species arid widun a species from individual to individual (Doncasicr & Seymour, 1973). Tlie attraction of diplogastcrid predators, M. longicaudatus and M.fonidens is governed by different biotic and abiouc factors including their distance from the source of prey altractanis. Factors, like temperature and period of starvation of predators, could also be responsible for varying degree of aitracuon in these predators. Studies on dipiogasicnd predators indicate tlicir attraction also towards bactcna. Yeatcs (1969) observed attraction oiDiplenteron colobocercu5\owai6sahac\cn\imBaciUuscereuj \zxmycoides. M. longicaudatus and M. fortidens responded positively towaids unlcnown bacteria cultjvated on agar by adding infant milk powder (Bilgrami & Jairajpuri, 1988). These predators could reach prey nematodes, bacteria and other source of food by chance or wilful movements (Poinar & Hansen, 1986), their orientation is determined by probing movements of tlic head (\Vard, 1976; Seymour el. al., 1983) and location of prey or bacteria depends upon papillae and amphids (McLaren, 1976; Wright, 1983).
Attack response Attack response of predators is indicated by llieir head probing and oesophageal pulsations. The probing is an important phenomenon helping the predators not only to idcnufy tlicir prey but also to locate a suitable spjt on the prey body for initiating an attack and thereby attaining a suitable posture for launching an attack. The most appropriate posture is attained when the predator e<.lablishes lip contact with the prey usually at right angles and a vacuum is crcvilcd by iJie oesophagus. The body of the prey is then slit opened or punctured. TTie attacks are usually successful when right angle contacts with the prey are established. Glancing contacts or contacts other dien those at right angles do not ncccessarily initiate an attack resulting into prey wounding (Bilgrami el. al., 1984). Such unsucccsful contacts lead to tlie escape of prey from attack. .
The probing may be quite aggressive in P. punciatus, M. deniaius fNelmes, 1974; Jairajpuri & A/mi, 1978); vigorous and confined only to ^liort distarKcs in L. vulvapapillaiuin (Wyss & Grcwuat, 1977); or gradual and rcstncu;d to short distances in A. tlwrnei and D. stagnalis (Bilgrami et al., 1985r; Shafq.it a. at., 1987). The piobing in A/, aquaiicus is rapid, side to side lip rubbing of the prey cuucle lasting for very short durations (Grootaert & W)ss, 1979). M. longicaudatu.^ probed its prey for slightly longer duration but probing by A fortidens was more vigorous (Bilgrami & Jairajpuri, in picss). The probing in Butle,'ius sp., occurred in the form of quick head shaking and lip rubbing in order ic establish i full contact widi llie prey (Grootaert er al, 19771.

(1 / H ^
106 Predatory Nemaiodes
Attack When predator comes in contact witli tlic prey, either by chance or wilful movement, it establishes a lip contact Contact with any other region Cnon-labial contacts) does not sian an attack as full lip contact is essential for tins act While alla:lQng, the predatory nematodes do not show preference for any particular part of the body of prey (Grootaert & Wyss, 1979) as they attack rather mdiscnminatly involving all regions of prey body
The mechanism of attack differs from predator to predatoi depending upon the type of feeding apparatus M aquaticus attacks prey by moving its lips in a backward direction ri'sulting in its mouth being wide open and the tooth coming in close contact withlheprey (Grootaert & Wyss, 1978) M deniatus iniUdtcs attack by widely o[x;ning lis vesubule with the help of labial muscles and exposing the doisal looih and denudes to bite the prey (Jairajpun & Aznii, 1978) L vuhapapiUaium begins an attack'by thrusung its stylet vigorously (Wyss & Grootaert, 1977) and A ihornei and D stagnalis attack prey by gradual and inicrmiitcnt tlirusting of their mural tooth and odoniostyle respccuvely (Bilgrami ei al, 1985a, Shafqat et al, 1987) D coiobocercus, Butlenus sp , M longicaudatus and A/ foriidens attack witli the help of their movable dorsal toodi and the oesophageal suction (Yeatcs, 1969, Grootaert et al, P77, Bilgrami & Jau-ajpun, in press)
CutiCle perforation may be achieved rather quickly with tlie help of odonto-si) le (dorylaims), mural tooth (nygolaims), spear (aphclenchs), dorsal tooth (mononchs), onchia (acunolaims), teeth (cnoplids) and die combined action of high suction force of the oesophagus and mo% able dorsal tooth etc (diplogastends) L vulvapapillaium required not more than five odontostyle tlirust to puncture tlic prey cuticle but A ihornei and D stagnahs needed 8-10 mural/stylet tlinists for achieving this The mononchs also attain cuticle perforation rather quickly Tlic aphclcnchid predators, chiefly the species of Semwa, paralyse tlic prey by injecting Lhcm with salivary secrcuons (Linford & Olniera, 1937, Hcchler, 1963) The dorylaim and nygoLiim predators paralyse their prey by disorganizing tiieir internal body organs with the help of their odontostylc and mural tooili respectively Mononchid and diplo-gastcrid predators inactivate their prey by tlie high suction of liicir oesophagus
Sah\aiwnlexira corporeal digestion Tlie ingcsUon and feeding upon prey contents staits soon after the prey cuticle is slit opened or punctured Extra corporeal digestion, smular to plant parasitic nematodes (NVyss, 1971) also occurs in piedatory nematodes (Lee & Atkinson, 1976) In stylet bearing predators (dorylaims, nygolaims nphe-lenchs) the ( esophageal sccanions contaming cn/ymes are inja ted inio the prc> body widi die help ol (ceding apparatus and diese cause the pre digestion of die food before it is sucked up by ilie preditors Tlirough tJie action of these enzymes (histolytic) the complex food globules arc broken down into simpler and smaller paiticlcs which can enter die hollow stylet Tlic extra corpoical digestion

1 4 5
MS Jairajpuri & AL^L'dgranu 107
has been observed in D. colobocercus (Yealcs, 1969), Builerius sp. (Grootacrt et al., 1977); L. vulvapapillatum (Wyss & Groolacrl, 1977); M. longkaudatus and M.fortidens (Bilgrami & Jairajpuri, in press). In case of mononchs no such observation has been made which could indicate the occurrence of extra-corporeal digestion. It could be possible that tJiese predators do not require salivation because of tlie fact lliat they possess a well developed, wide and strong buccal cavity with which they could engulf even an intact prey.
Ingestion/feeding The feeding starts as soon as tiie cuticle of liic prey has been punctured. The ingesuon of prey contents is intermiitcnt (.short periods of sucking activity) in almost ail tlie predators. The mononchs swallow llieir prey intact or puncture its cuticle with the help of suction arid buccal armature. However, species of Mylonchulus do not engulf whole prey (Bilgrami et. al., 1986) and feed by cutting it into pieces (Jairajpuri & Azmi, 1978). The swallowing of whole prey is aided by the contraction of oesophageal muscles which pull the prey into tlie oesophageal lumen and tlie posterior oblique pbies assuming vertical position (Grootaert & Wyss, 1979).
The stomatal muscles along with the suction probably provide a gcxxi hold on the su-uggling prey. Further, after the feeding has been accomplished these muscles restore the head region to its original position thereby acting as retractors. The marguial muscles of the oesophagus assist in the closure of lumen. The act of devouring an entire prey may lead the mononchs to move randomly once again in search of yet another prey but sometimes tiiey also become inactive for short durations (Nclmes, 1974).
The doryiaim, nygolaim and apheienchid predators cannot swallow or enguif their i rcy whole or cut them into pieces because of the piercing and sucking t}7)e of ilicir feeding apparatus. These predators feed by puncturing llie cuticle of the prey and then sucking its body contents. After penetrating the body of prey the predators move tiieu" feeding apparatus 'ji'lcways. During these movements each odontost>le protrusion is accompanied by a suction which is caused by the conu-action of oesophageal niu.scles. As soon as the ingestion begins the slylel/tooth which is nearly protrutcd fully With tlie anterior fold of tlie guiding shcaiti close to llie oral aperture. The ingestion is typically intcniiiticnt < .id tlie suction is achieved by die contracuon of radial muscles of oesophagus. Whenever, tlic radial muscles withni the oesophageal bulb contract, the triradial oesophageal luinen is diaiatcd and the bulb is suctched (Wyss <i Grootaert, 1977) Upon relaxation and simuluincous shortening of bulb, the oesophageal lumen narrows (roni Iroiit to its back. This.il!o\\% the sucking iii ol the body con tents and lore in g them into tJic intestine tlirough llic cx'iophago-mtestinai junction. The depletion of ilic prey's conlcnLs at the feeding spot is usually followed by scaicliing ino\cnicnis of the feeding apparatus and finally by the separation of the lips from the jjrcy and subsequent

1 4i
108 Predatory Nimalodei
ictracuon of the slylct/looth The same prey is then attacked at some otiicr place and feeding starts all over again These predators also feed upon the eggs of pre/ ncrnairxJcs but not on their own eggs When in contact with their own eggs they rub theii with their hps several times but do not make an attempt to feed upon them (Esser, 1963 &. 1987) The rubbing seems to be for the purposes of recognition cf eggs
The diplogasterid predators viz , D colobocercus Builenus sp , M lonqicaudatus and M foriidens also feed by punctunng the cuticle and llien sucking in tiie contents of the body of prey However, M fortidens were observed to devour intact firj' and second stage juveniles of small sized prey nematodes, Aiwbeloides sp , znd Cei^halobus sp , and also cut larger prey into pieces for feeding purposes The ingestion and feeding in these predators is helped by the movable dorsal tooth and the suction force generated by the oesophagus witli the help of oesophageal muscles Nclmes (1974) observed 'shunung' behaviour in P punctatus as in the free hving nematodes (Croll, 1974), a phenomenon which was also observed in M hngicaudatus and M fortidens The predators moved forward or backward only up to a distance equal to about theu' OV>TI
Ixxly lengtli and ilitn reverting back to tlicir onginal positon for fccdmg once apjin on the prey The type of feeding apparatus possessed by the aciinoiaims and enoplid predators suggests that they can feed by cutting the body of the prey
The prey nematodes may, sometimes, escape from the grip of predator cither due to an unsuccessful attack or the inability of prcdaioi to pucture the prey cuticle or due to evasive actions uiken by tne prey nematode The evasive action could be the abilit) to retreat instantly when attacked or the acuve undulatory movenienis However, an injured prey while escaping may attract other predators which would then con lerge upon it This phenomenon is seen commonly in dorylairn, nygolaun and diplogasterid predators but not in the mononchs Though, predators may feed togcthci but a defmite struggle among them to feed first upon the pre) could be noticed Th^ is more signilicant m diplogasterid predators Group feeding is an interesting phenomenon as it allows predators to nnish their prey rather quickly and then continue hunting 1 his results ui increased predation as a group of predators may taie far less Uine to consume tlicir pre\ than a single predator During aggregauon at feeding sues tlie average number of predators actually feeding is less than the total numtx,r of predators present (Bilgrimi A. Jjirajpuri, 1989a) 1 he duuiion of aLjrcgaiioii t feeding sites and the actual fccdingj tune may vary from prcdaior to predator and dc[xnds on the t)]x; of prcN tciure and contour of prey cuucle, composition, conccnurauon, quality and quanta, ol the pre> contLnus I
Feeding in picdilors is completed soon alter tJic prey is complLi lv tunsum.d (diplogastcndb, mononchs), devouring an mtacl picy (mononchs) or suckiiii; m u\c whole lxxJ\ contents of nicy except tlic cuticle (dorylaims, nygolnms, iphclcnchs)

'•7 H
MS Jairajpuri &.AL Btlgrami 109
Prey rreference/ Range of I'rey
The knowledge of predalor-prcy rclauonship and range of prey has great significance in formulating biological control programmes and in proper utilization of different kinds of predators. The obser/ations on the predation abilities and preference of predators for different prey species were mainly made on live nematodes in aqueous suspensions (Thome, 1924; 1927); intestinalcontentsofpreserved predators (Banage, 1%3; Szczgiel, 1971; Bilgrami era/., 1986);in wrro (Jairajpuri & Azmi, 1978; Grootaerte( a/., 1977) and in pot experiments (Cohn & Mordechai, 1974). Though, no predator is really prey specific but tlic range of prey is rather limited and differs from species to species. Esscr (1963) observed lliat the predatory dorylaims and mononchs prefer Meloidodera floriden-sis, Pratylenchus penetrans, P. vulnus, Paratylenchus curvitatus and Meloidogyne spp. while Pnonchulus punctalus preferred Anguina iriiici and Apheknchoides avenue but not liehcotylenchus (Nelmes, 1974). Mylonchulus dentatus, Dorylaimus stagnalis, Mononchoides longicaudatus and M.fonidens also killed fewer Helicotylemhus indicus (Jairajpuri & Azmi, 1978; Shafqat el al. 1987; Bilgrami & Jairajpuri, m press) Tht dorylaim predator, Lahronema vulvapapillalum preferred Aphelenchus avenae, Panagrellusredivivus and Angiana triuci in place of P. penetrans and Xiphinema index (Wyss & Grootaert, 1977). Prey selection/preference has also been observed in dipiogastcrid predators. Builenus sp. preferred soil stages of endoparasitic nematodes in place of ectoparasitic ones (Grootaert et al, 1977). The prey nematodes viz., Aphelenchoides frcigariae,Mesodiplogasterlhentieri,Cephalobus sp., Almmiu sp.,Rhab-diiis sp. (larvae); the juvenile stages of/'rarj/enc/iuvsp ,Plectus sp ,Monocnhus aquaticus and the adults and juveniles oi lihalxhiisoxycerca wdPclodrra sp. were also attacked and consumed by Bullenussp Roiylenchus rohustus, 1 ylcnchus macrurus, Meloidogyne naasi, Pratylenchus sp , Rlkibdiiis sp., Plectus sp. and Mesodorylaimus sp. were attacked rarely but served as food for Builenus sp., when aitificially wounded. Tylenchorhynchus sp., Merlinius sp , Henucydiophora sp , and Trichodorus sparsus were neither attacked nor killed by these predators. Builenus degressei killed a large number of P.redivivus (Small & Grootaert, 1983). While observing the predauon abilities of A/, longicaudatus and M. fortidcns, Bilgrami & Jairajpuri (in press) found that these prcdjtois prefer Acrobeloides sp., Ceplialobus sp.,P rediMvus, Uirschmanniella orvzae and the sceond stage juveniles oi Meloidogyne inrognua and A tritici, but did not feed u[X)n lloplolaimus indicus and llctmcnconcnioidcs inangrerac Oilier prey nematodes, viz., Rrabdiw; ^p, Longidonis sp., Xiphinema amcncanum, Tylenrhorhynrhw; mashhoodi and Heliint\U'nt hits indirus were inoderaieK preferred lloplolaimw; indirw; ucic prckrrccl neither by Myloinludus dentatus (Jairaiputi cV, A/nii, 1978) nor b_, DoryUwuK sta'^miU'^ (Slialqat et al, 1%1). Diplenteron colopoLcrcus, however, did not sliow prLlercntc lor

UiS
n o Predatory Nematodes
any particular t>pe of prey and killed Mesorhabditis httoralis, Panagrolaimus austrahs, Acrobeloides syrtisus, Zeldia punua in equal numbers. Cohn & Mordechai (1974) reported that the feeding by Mylonchulus sigmatums was more on Tylenchulus semi-penetrans or Meloidogyne javanica juveniles than on Hslicotylenckus multianctus or Longidorus qfricanus.
The selection of prey depends on the activity, size and the behaviour of prey nematodes. M. aquaticus preferred less active nematodes like Cephilobus sp., Aglcnchus parvus as compared to more acuve Pnsmatolaimus sp. (Bilgrami etal., 1983). The prey selection may differ from species to species of predators. Thome (1932) found remnants in lotonchus acutus of those prey {Rotylenchus robustus, Trichodorus sparsus) which were not killed by Butlenus sp. (Grootaert etal., 1977).
Bilgrami etal. (1986) have analysed the intestinal contents of over 1(XX) specimens of 33 species of nine genera of Mononchida to assess their choice of prey. It was found that the choice differed widely and there was no clear-cut pattern; Sporonchulus (one species); Pnonchulus (one species); Coomansus (one species); Mononchus (two species); Parahadronchus (one species), Clarkus (two species); Miconchus (three species); Mylonchulus (nine species) and lotonchus (thirteen species) were analysed. The species of lotonchus preyed upon five genera each of Tylenchida and Rhabdmda and seven of Dorylaimida and those of Miconchus on five genera of Tylenchida, four of Dorylaimida and three of Rhabdiuda. The prey nematodes belonging to three genera each of Tylenchida and Rhabdiuda and four of Dorylaimida were found in the intesune oiMononchus^i four of Tylenchida, two of Dorylaimida and three of Rhabdiuda in Clarkus were identified as prey. The most diversified type of prey nematodes belonging to eight genera of Tylenchida, six of Dorylaimida and five of Rhabditida were found in the intestine of Parahadronchus shakdi Predators belonging to Coomansus were found containing three genera of T)lenchida, one of Dorylaimida and two of Rhabditida in their intestine. The nematodes belonging to four, three and two genera of Tylenchida, Dorylaimida and Rh,^bduida respeciiMely were identified as prey of Pnonchulus. Sporonchulus pre>ed upon three genera each of Tylenchida and Dorylaimida and two of Rhabdiuda. None of the species of .Mylonchulus (M. brachyuris. M. contractus. M. dcniaius.M.lacustns, M ha%>,aiensis, M amurus, M. muradi, M. sigmaiurus. Si. namtalen-sis) contained prey or thcu' remains in the intesune. This confomis with the obscrvauons made by Mohandas & Prabhoo (1980).
The species of Prat\lenchus, Hoplolaimus. Tylenchorhynchus, T\icncholaimus. Aporcclaunus. Thomenema. MylonchuUs. Rhabduis, Acrobeloides and Chiloplacus wcr: tound most frcqucnil) in the mtcsime of different mononchs. Other prc\ ncmoicd;s not 100 frcqucntl) encountered belonged [O the gcncrd. Uclicotylcnrhw;, llirscrjruinnicll.2. !le'".ii.\cUopnora. llciniLni.oncmoidcs. Aphclcnchus. Apkelcnchoides Discolaiin i:. Bciondira, Axonci.ium, Dor.lainicU.iS. Nygolawus. Tnchodorus. Xipt.incma. Dipn-

149
MS Jairajpwi & AL Bilgr^rra 111
tkerophora, Campydora, Mononrkus, Clarkus, Miconchus, Acrobeles and Mesorkabdias besides few diplogasterids and monohysterids. Tylencholaimus, Tyler.chorhynchus. Mylonckulus and Chdloplacus were found within the intestine of maximum number of species of mononchs. Tylenchorhynchus was observed in the gut of eleven species, Tylencholaimwi in seven, Mylonchulus in nine and Chiloplacus in eleNen species belonging to different genera. Kind and number of prey nematodes v,aried and these were found either intact or in semi-digested condition in the intestine. The remains of the cuticle, spear/stylet, spicules, buccal cavity etc. were also observed frequently in the intestine. loionchus trichuris, I. longicaudatus, I. indicus, Clarkus papUlatus, Coomansus indicus, S. vagabundus and P. shakiii also preyed upon rotifers besides soil and plant parasitic nematodes.
Arpin (1979) and Mahapatra & Rao (1981) found a significant correlauon between the populations of mononchs and free-living nematodes but Nelmes & McCulIoch (1975) did not find such a correlation. Bilgrami el al , (1986) observed more free-living saprophagous (75%) nematodes than either the tylenchs (45%), dor\laims (42%) or the mononchs (27%)in the gut of mononchs. The heterogeneity of prey in the intestine of mononchs suggests that they are polyvalent and their prcdation aleatory. Similar conclusions were also arrived at by Mohandas & Prabhoo (1980).
Prey Resistance and Susceptibility to Predation
Every organism has one or the other kind of strategy, hereditary or aquired, for its survival or existence and to protect itself from the adverse climatic ronditions including the enemies. Like predatory nematodes which possess different kinds of mechanisms to overpower and then feed upon their prey or the plant parasitic nematodes which have developed mechanisms to locate their hosts, the prey nematodes have also evolved certain devices for defending themselves from the onslaught of the predators. This mechanism is termed here as prey resistance and may be defined as the ability to avoid predation (wounding). The susceptibility of prey represents the sus':epubility to predation (Bilgrami & Jairajpuri, 1989b). The resistance is of varied type and it may be physical, chemical or behavioural in nature. Physical resistance is due to physical charactenstics, such as, thick cuticle, body annulaaons, double cuticle, gelaunous mairu, etc. (Esser, 19t)3; Small & Grootaert, 1983; Bilgrami & Jairajpuri, 1989b); the chemical resistance IS because of toxic/unfavourable/rcpcilent secretions (Esscr, 1963; Bilgrami & Jairajpuri. 1989b;, while behavioural charactcnstic represents certain mechiinisms of the prey nematodes to resist or to avoid predation by active body undulations (Bilgrami & lau-ajpuri, 19S9b), ability to retreat instantly when attacked (Esser, I%3); vigorous escape response (Grootaert etal.. '.977; Small & Grootacn, 1983) and Lhc inability of 3rcdators to attack and injure them inert mechanisms). The wounding is an important

1 5 0
112 Predatory Nematodes
detciT" nant factor in predaticn A wound may result m loss of hyclrostauc pressure of bod> affecting locomotion and thereby making prey more vulnerable to predation Wouiding also permits invasion of pathogenic micro organism, resulting into death of prey Besides, the resistance of prey is lost if it is A-ounded or is in a weakened condition. The predaaon is measured m terms of number of prey kiJIed, mere encounters between predator-prey may not necessanly result m wounding or death Besides, wounding is the only mtermediate factor between the mortality and survivahty of prey organisms and hence could be taken into account for determming the degree of resistance of prey (ability to avoid wounding by pnysical, chemical or behavioural charactenstics) and suscepubility (to succumb to wounding) to predation (Bilgrami & Jau^jpun, 198Qb).
The ability of different prey nematodes to defend themselves from predators may vary from species to species and from individual to individual and determmes the degree of their resistance/susceptibiity to predation. Esser (1963) was the first to give a list of prey nematodes that were resistant or susceptible to predation by different dorylaims and mononchs Taking into the consideration the abihty of prey to resist repeated attacks of predators successfully he considered Hoplolaimus cylenchformis, Belonolaimus lon-gicaudatus, Hemicychophora sinuhs, Dohchodorus heterocephdus, Cnconemoides spp. Scutellonema spp ,and //e/icor>/^/ic/!U^spp.as resistantpreyspecies.On the other hand Meloidoderafloridensis, Praiyknchuspenetrans, P vulnus, Pararylenchuscur\itaius and Meloidagyne spp, were extremely susceptible to predauon by different mononchs and dorylaim predators Cohn & Mordechai (1974) found Hehcorylenchus muhicmctus and Longidorus africanus as resistant prey species while Tylenchulus senupenetrans as the most susceptible Groctaert « a/ (1977) while studying predation by fluf/enui sp., concluded that soil (juveniles) stages of endoparasitic nematodes arc extremely susceptible to predaceous nematodes but the ectoparasuic nematodes resist predauon fairly well Tylenchorhynchus sp , Tylenchas macrurus, Meloidogyne naasi, Menminus sp. Pratylenchus sp (adults), Tnchodorus sparsus and Hemicyclwphora sp were totally resistant to predation by Budenussp Small & Grootaert (1983), *hilc observing the predauon abibties of Pnonchulus punctatus, Mononchus aquancus, Butlerius degrissei, Labronema vuhapapillaium, Neoactinolaimus duphcidentatus, recorded Rhabdnis oxycercafeloderasp andRorylenchusfallorobustus^STCSisVimptcy species Panagrellus redivnus was the most susceptible prey while other free-living and ectoparasuic species were rclauvely less resistant to predation Recently, Bilgrami & Jairajpun (1980b) reached almost similar conclusion and found P redivivus, Cept^lobus sp, Acrobe'oides sp, and the second stage juveniles of M mcogmia and A iri.,^ias most suscccuble to predation by M longicaudaius and V/ forttdens Hoplolaimus ir.aicus and HenuLriconemoides mangiferae were totalh resistant to predauon Tlic other trec-li\inj and plant paraiiuc nematodes [Rhabdms sp TylenLWrnvnchus tTiaMiooai \,Dninuiki

I 51
MS Jairajpiu-i S:.AL Bdgrcirru 113
amencandm Longidjrus sp Hehcotylenchus mdicus, Hirschmanniella oryzae) w.rt
moderately suscepub'e/resi^tant to predauon by M longicaudatus and M fomdens The thick or double (sheath) cuticles are common amongst ectoparasitic
nem.atDdes {Hoplola.Tus spp, cnconematids) and may represent an anti-predauon device in otherwise exposed and vulnerable prey species Weil developed annulauons possibly provide further protection to prey nematodes from attacks by predators. Esser (1963) sp&ubted that the resistance in Hoplolaimus sp, was due to their tough and thick cuticles Small & Grooiaert (1983) also suggested that the thick cuticle is a better defence against rapid attacks of mononchs and diplogastend predators. Esser (1963) suggested that species of Hehcotylenchus have a chemical resistaiKC against predation as the predators rarely contacted or if contacted did not feed on them. He (1 c) records two observations m which dorylaim predators withdrew suddenly after attacking a Hehcotylenchus sp and appeared to regurgitate some substance which oould be noxious. The same holds true for the predators, Af fonidens and M longicaudatus which were attracted least towards Hehcotylenchus indicus (Bilgrami & Jairajpun, 1988) and did not prefer to feed upon these nematodes (Bilgrami & Jairajpun, in press) Nelmes (1974) also observed Pnonchulus punctatus refusmg to feed upon Hehcotylenchus dihysiera Besides, some prey nematodes (R oxycerccr, Plectus sp, and other rhabdiud nematodes) may avoid predation by thcu" thick '"uucles and rapid escape responses (Small & Grooiaert, 1983, Grooiaen et al, 1977). The escape response may be more successful against a comparatively slow predator like L vulvapapillatum, D stagnalis, A thomei and probably against all dorylaim and nygolaim predators which require several stylet thrusts to achieve penetration However, aphelenchid predatcrs {cf Semura spp.) may overcome these responses of their prey with the help of their toxic sccreuons which paralyse the prey quickly (Linford & Ohviera, 1937, Hechler, 1963). The high degree of sucepubility of Cephalobus sp , Acrobiloides sp , P redivivus along mlh the second stage juveniles of M uicognita and A trmci to predation by M lon^caudatus and M. fortukns happened to be due to their small body size, slow rate of movement and lack of protecuve cuncle adaptauons (Bilgrami and Jairajpun 1989b) Such characteristics are common among endoparasmc nematodes (Esser, 1963, 1987).
The ectoparasitic nematodes hvmg m close proximities to the predaceous nematodes may have developed more ana-predauon devices. These chairactensucs such as thick body cuucle, annulauons, toxic unfavourable/repellent chemical secreuons, escape response, active body undulations etc , may be more ef]'"ective against diplogastend, dorylaim and nygolaim predators since these predators possess cutDng or piercjig type of feeding apparatus and feed by sucking the contents of prey Mcnonchs may, however, overcome resistance oi prev of different kinds to a larger extent as these predators often do not cut the cuticle o*" prcN but engulf them whole The ec-loparasiuc nematodes are, therefore, more vulnerable to predauon by mononchs This

152
114 Predatory Nerr^odes
IS evident as many spec es of mononchs ha\B been found containing species of Hoplolaimus.Hehcotylenckus.Xiphinema.Hemicnconemoides.Rhabditis, etc .besides other tylenchs, dorylaims, rhabdiuds and nionochs entire in their intestine (Bilgrami et al., 19S6). Mohandas & Prabhoo (1980) also found intact Tylenchorhynchus nudus and Xiphiner^M elongavjm in the intestine oilotonckus kherai and / monckystera.
A predator may be considered as an efficient agent of bio- control if it possesses high strike rate (ability to attack and wound prey) against a panicular type of prey depending upon the degree of resistance/suscepubihty of prey against predauon. The most effective and suitable combinauon of predator and prey could be the most efficient predator (in terms of highest stnke rate) and most suscepable prey (i e., most vulnerable to wounding) Bilgrami & Jairajpun (1989 b) have proposed certain formulae to deter-mme the stnke rate of predators and resistance/suscepubility of different prey nematodes against predauon. The stnke rate of the predator is defined as llie percentage of success with which It atncks prey (attack rep-esents the rapid movement of head and the apphcation of suction created by oesophagus of the predators) The stnke rate of the predator may reflect its predatory potenual and could be detennined as follows-
SR(%)= ^ ;tlOO E
The resistance of a prey may be calculated as percentage degree of resistance by the following formula:
PR(%) = ^ ^ ^ X 100
whereas, the suscepubihty of prey nematode may be determined by the followmg formula:
PS(%) = 100-PR
Where SA=stnke rate of the predators; PR=prey resistance; PS = prey susceptibility; EA=total number of encounters resulting mto attack; E=total number of encounters made by the predators; AW=total number of encounters resuitmg into prey wounding.
Factors Influencing Predation
DilTeient biotic and abiotic factors influence predation as observed i/j VUTO and the same is likely to De true m natural habitats. The following factors may either mcrease or decrease the rate of preaauon.

^>3
MS Jairsjpuri ISLAL B.lg'-cmi 115
Prey activity. Predation depends on the type of prey. Bilgrami el al. (19S3) found prcdation by M aquaiicus inversely proportional to the actmty of the pre>. The most acuve prey, Pnsmatolaimus sp, was killed least, while the least active Cephzlobus sp., was devoured most . Maertens (1976) usingP.punctatus obser/ed that ten? redivivus were killed per day while Nelmes (1974) using the ssnne predator but Apheienchus aveiae as prey recorded ten killing per hour. This could be due to the differences In pre acuvnty as P. redivivus is far more active than A avenae. In general, the free-living nematodes possess higher endogenous acuvity (CroU, 1972) than the parasiuc species. Besides activity, other factors such as body cuticle, annulations, unfavourable body secretions of prey, inability of predators to attack or puncture prey cuucle etc., also govern directly or indaecdy the rate of predauon as already menuoned earlier under prey suscepubiliiy and resistance.
Sex and age of predators: The male and female D. stagnalis, M.fortidens and M. lon-gicaudatus did not differ m their rate of predauon (Shafqat et al., 1987; Bilgrami & Jairajpun, in press). However, males of D. colobocercus preytd more than iheu- females (Yeates, 1969) The age of predators usually governs prcdation. Aduk and fourth stage juveniles being stronger, kill a larger number of prey than their yoang stages (Bilgrami & Jau-ajpuri, 1985; 1989b). The first and second stages of some may neither be predaceous nor cannibalistic in nature, but feed as free-living nematodes e.g., M. aquaticus (Bdgrami et al., 1985) or canion feeders e.g., P. punctatus (Nelmes, 1974). Prey number. The prey density affects the rate of predation by increasing chances of encounters. Yeates (1969) found that rate of predaaon by D. colobocercus was proportional to prey density. Simdar observauons were recorded by Bilgrami et al. (1985) and Bilgrami & Jairajpuri (1989b) with A. thomei and, M. longicaudatus and M.fomdens. However, the abundance of prey did not mfluence predation by M. aquaticus (Bilgrami etal. 1984). In case of D. stagnalis predation increased significantly up to a populauon level of 125 prey nematodes per five predators, but prey numbers higher than this did not affect predation or the rate of killing prey per day. The increase m predauon was perhaps the outcome of improved chances of encounter between the predators and prey which, however, did not alter prcdation beyond an optimum level of prey pcpulanon.
Temperature: In nematodes, temperature produces certain interesung changes, more specially with respect to their activity and movement. There are usually two types of effects that can be produced by temperature: one is on the sensory behavioural responses mediated through the sensory receptors and the other is thermodynamic effect resulting from changes m the rate of metabolic processes of the nematodes.
Wallace (1963) concluded that majority of nematodes become inacuve at 5-15^ and 30-JCPC ana the optimum temperature lies bctwen 2C-30''C. These may of course vary for different nematode species under different environmental conditions. .Azmi i Janaj-pun (1977) also found temperatures between 25-30°C being oDtimum tor the nematodes

154
116 Predatory Nematodes
they had tested. Bilgrami et ai, (1983) concluded that the temperature influnced the activity of predators and prey and caused decline in the rate of predation by M. aquaticus. Grootaert & Maertens (1976) also found temperatures affecting activity of M. aquaticus. D.stagnalis preyed maximum at temperatures ranging between 25-30°C (Shafqat etal, 1987). In case of Af. longicaudatus and M.fortidens, a significant correlation was found between the prey killed and temperature (Bilgrami & Jairajpuri, in press). The optimum temperature for these diplogasterid predators was found to be between 25-30°C. Since predation is a physical phenomenon, the affected sensory behavioural response may be one of the factor responsible for this differential rate of predation at different temperatures. Starvation of predatory. Starvation is a physiological phenomenon occurring under conditions of non-availability of food. The change in the appet;itive condition may also bring about changes in the response of predators with respect to iJieir activity, movement, attraction, feeding, mating etc. The short-term deprivation of food enhances predation by P. punctatus. but declines when the predators are starved for a long duration (Nelmes, 1974). According to the latter author the predator; loose their appetative behaviour if star\ed for more than three weeks. This could have happened due to loss of energy as starving nematodes utilize their reserve food materials (Cooper & van Gundy, 1970; Reversal, 1981a 1981b), thereby affecting their movements and other behavioural responses or due to their becoming habitual of prey non-availability which may occur due to acclimatizing effects of starvation on predators. Jairajpuri & Azmi (1978) found the rate of predation by hungry M. dentatus distinctly higher than the firesh predators. Similarly, Bilgrami & Jairajpuri (in press) also found significant conelation between the number of prey killed by M.fortidens and M. longicaudatus, and the number of days they were starved. In contrast D. colobocercus.M- aquaticus dnd D.stagnalis were not influenced by starvation and the rate of predation by fresh and starved predators remained same (Yeates, 1969; Bilgrami et al., 1984; Shafqat et ai, 1987). Agar concentration: The movement and locomotion are necessary for feeding and mating activites. In experimental studies agar concentrations govern the activity of nematodes O'allace, 1969; Azmi & Jairajpuri, 1977). Bilgrami « a/., (1983) attributed lower rate of predation by M. aquaticus to increasing agar concentrations which inhibit activities of both the predators and prey. The activity was maximum at lower agar concentrations and consequently the rate of predation was higher. The increased prey and predator activities may also inrrcase chances of conuci between predators and prey. Shafqat etal. (i987) while evaluating the predation abilities of I'. 5Mgnato also reached similar conclusions and found lower concentrations most favourable for prcdauon. A high significant correlation was found by Bilgrami & Jairajpuri (1989c) for M. longicaudatus and M.fortidens.

55
MS Ja.rajpur. i.AL Bdgrami 117
Cannibalism
Little is knov-Ti about the cannibalistic behaviour of predators and only a few of them have been reported as cannibals. ATini & Jairajpuri (1979) found M. dentatus to be cannibalisuc in the absence of prey nematodes. Mulvey & Jenson (1968) also reported that some species of mononchs resort to cannibalism. In M aquaticus besides adults, the fourth and third stage juveniles also show cannibalistic behaviour when placed with then own juvenile stages in absence of prey nematodes (Bilgrami & Jairajpun, 1985a). Nelmes (1974), however, did not observe this phenomenon in P. punctatus. The cannibalistic tendency is known only in mononchs and not m other predators except M. longicaudatus and M fortidens (Bilgrami & Jau^ajpun, m press). Af./om^e/iioccasionally resorts to cannibalism in the absence of prey instead of feeding on bacteiia. The ability of predators (dorylaims, nygolauns, diplogastends) to recognize their own chemical secretion (Esser, 1963, Grootaert & Small, 1982; Bilgrami etai, 1985a) and to attract their own members (Bilgrami & Jau^ijpun. 1988) possibly induces cannibalism .It may be possible that dorylaim and nygolaim predators also resort to cannibalism a fact that is unknown so far.
Cannibalism appears to be more of natural phenomenon rather than occumng only due to prey non-availabilicy as in most iristances the mononchs (prey) occurred together with other types of prey in the intestine of mononchs (predators) (Bilgrami et al., 1986). All species except? muscorum, Coomansus indicus, lotonchus basidontus, 1. jain, I baqru, I. shafii, I. antedontus, I. longicaudatus, I. prabhooi and Mylonchulus spp. show cannibalistic tendency. Twenty percent predators having prey m iheu intestine were cannibals. The cannibalistic tendency may vary from predator to predator as species of Mononchus and Sporonchuius kiJled their own species more than the other mononchs (predators).
Biology and Culture of the Predators
A huge populauon of predators is required to execute biological control programmes successfully against a plant parasitic nematode species m the field. The better understanding and advanced knowledge to elevate the populauon of predators both under in virroand in vivo conditions is thererore required. This could perhaps be the main hurdle, so far, which has restricted the use of predatory nematodes in combatmg pbnt parasitic nematodes. Some species of prcdaionj. such is mononchs, have a very res-Uricicl distribution but others lUe the dorjlaims and the nygolaims arc of cos-mopohtan m nature. Stil!, \ery liitle is known about thcu" biology, reproductne potential and culture techniques. The various culture lechinqucs arc a fascinating area of study

I b*^
l i s Preda orj \erralocLs
and have been reviewed by several workers Following Esser (1963) who cultured predator} dcr;lajms ard mononchs, Yeates (1969) cjitivated diplogastend predator D coloboce'-cus on agUT plates using a batenum. Bacilluscereusvzimycoides Nelmes (19"4) reared P punctatus on A a\enae and P redivivus in soil extract agar medium at pH 6 8 similar to Spnngets (1964) who reared enchytreads Maertcns (1976), Grootaert & Maertens(1976) and Sr^all (1977) also used the same techiii.,ue) to elevate populauons ot M aauancjs and P punctatus using P redv.ivus as prey Shafqat er a/. (1987) and Bilj-ami &. Jau-ajpun (1988) cultivated the predators,D stagnalis andM longicaudatus and M fortidc is respectively using Rhaocitis sp , in agar plates supplemented with mfant milk powder (Lactogen) under in vitro conditions Addition of milk powder boosted bactenal growth in maintaining theu- populauons for long durations M fortidens and Af longicaudatus are not obligate predators, as they also feed on bacteria besides prey nematodes These nematodes can also be cultured easily using bacteria and their populations may be elevated to desu-ed level by supplementmg organic compounds which support bactenal growth Studies on the biology of predatory nematodes need more attenuon as only few observations ha^e been made on their life cycle and reproduc-uve potenual (Maertcns, 1976, Gnxitaen & Maertens, 1976, Grootaca & Small 1982).
Conclusions
In view of the success achieved by entomologists m controlling some insect pests by mtroducing ih"ir natural insect encm'es, it is logical that the nematologists should also consider the possibilities of controlling the plant parasiuc nematodes with the help of predatory nematodes Though, the use of chemicals and nematicides in controlling populauons of plant parasiuc nematodes have yeilded some posiuve results but thetr use is now being discouraged because of theu- exorbitantly high costs, theu- toxicity to other beneficial soil fauna, theu- residual effects on human bemgs and live-stock and because of the fear that the conunuous use of these chemicals may result in the nematodes becommg resistant to chemicals The studies which have oeen earned out on different aspects of predators, like the identificauon of preuaceous species, ecology of predators, range of prey, reproducuon and their rclaucnships with prev nematodes has led manv nematologists to test them in agncultural fields for cono-olling bpccics of plant parasiuc nematodes However, infcrmauon on Lhe predauon ibiliues oi predatory nematodes iS •sparse and their assessment as potenual biological control agerts is largclv spcculauve Linford ».v Oliviera (193") observed increase in the popuiaiion of predators wnen chipped leaves are added to the llcld Lai ^.tal (1983) also recorded a two-fold increase in the populauon of predators nemaiodci). Dorylaimus Discolaimus and Monvncnus when gavn manure is used in soil Thev have concluded diat the decrease in the population of plan parasitic nematod'"- occurred because ol increased popubLon

157
MS Jatrajpuri & AL Bdgrami 119
levels of predators Christie (1960) was of the view that further studies on biological control by nematodes \w3uld be of interest and it could possibly be of some practical use According to Jones (1974) non-speciCc predators excert only a partial control, if any, and are therefore unlikely to be of much value as good agents of biological control Similarly, the results obtained by Small (1979) were not conclusive of a bio- control potentul fcT predatory nematodes but he suggested further studies on the biology of predator and on those predator-prey combinauons which could result in partial control. Small & Grootaert (1983) also suggested further soidies on predatory nematodes as biological control agents. They emphasised that these studies should concentrate on the migratory stages of endoparasitc nematodes as these are more susceptible to predation. Ahmad & Jau-ajpuri (1982) found an m%«rse relationship between the populations of predator, Parahadronchus shakih and the prey species belonging to Tnchodorus, Henucnconemouks sp and Thomenema sp. under natural conditons. This could have been due to large popuLiuons of P shahh in that area. The mononchs, however seem to make Lttle impact on plant parasiuc nematode populauons although, they are efficient predators capable of preying on a wide vorietyof nematodes. The mam drav/back m usuig mononchs for biological control purposes lies m their isolated and restricted populations, non-availabihty of naturally occumng large populaDons in agricultural fields, longer life cycles, low reproducnvc potcnual, susceptibility to chjinging environmental conditons and a change m soil texture, soil type, etc. The mononchs arc very sensmve to chemicals and pH (Bilgrami & Jairajpun, 1985b). Esscr (1963), however, found dorylaim predators most efficient and highly potent as biological control agents In vitro studies which have been carried out so far on different diplogasterid predators also uidicate theu' ability as promising bio-control agents. These predators require attention of nematologists forexploringpossibilitiesofusingihemasagentsofbiological control The most advantageous and encouraging aspect of the dorylaim, nygolaim, and diplogasterid predators is the easy mauilenance of theu populations which can be achieved by adding orgamc substances in the fields (Webster, 1972) and also theu-pohphagous nature due to which they may be maintained without prey nematodes. Above all.bemg polyphagous they do not generally resort to cannibalism which helps m avoiding setback \n intraspecific mteracuons which is a natural phenomenon amongst mononchs The mononchs requu'e prev nematodes exclusively for theu- survival More studies on the biology, ecology, behaviour, rcproducuve potenual and longivit> of predatcTS are required in relation to prey and on their predatory behaviour, factors uilluencing predation, range of prev and predator-prey relationships before making pracucal use of these predators.

158
120 Predniory Nematode
REFERENCES
AHMAD, N. & JAIRAJPURI, MS. (1982). Populadon Huctuatio'ns of the predatory nematode, Parahadronchus shakili (Jairajpuri, 1969) Mulvey, 1978 (Mononchida).Froc. Symp. Ecol. Anim. Popul. India. Pt. 3,1-12.
ARPIN, P. (1979). Ecologie et systematique des nematodes Mononchidae des zones foresteries et haipaeces zones climaie, Tempere humide I. Types de sol ei groupements specifiques. Revue Nematol. 2,211-221.
ARPIN, P. & KILBERTUS, G. (1981). Ultrastructure de contenu digestif et de I'epithelium intestinal chez quelques nematodes predateurs (Mononchids) et bacteriophages. Ravue Nematol. 4,131-143.
AZMI, M.I. & JAIRAJPURI, M.S. (1977). Studies on nematode behaviour HI. Locomotion in juveniles of Hoplolaimus indicus and HeUcotylenchus indicus in different concentration of agar. Indian J. Nematol. 5,240-242.
AZMI, M.I. & JAIRAJPURI, M.S. (1978). Studies on nematode behaviour IV. Comparative xtivity of Tylenchorhynchus mashhoodi and Rhabdias sp. Indian J. Nematol. 6,96-98.
AZMI, M.I. & JAIRAJPURI, M.S. (1979). Effect of mechanical stimulation and crowding on Mylonchulus dentatus. Nematol. medit. 7,203-207.
BANAGE, W.B. (1961). The ecological importance of free living soil ncmtodes with special reference to those of Moorland soils. J. Anim. Ecol. 32, 133-140.
BILGRAMI, AJ,.; AHMAD, I. & JAIR.^JPURI, M.S. (1983). Some factors influencing predation by Mononchus aquadcus. Revue Nematol. 6, 325-326.
BILGRAIvn, AL.; AHMAD, I.& JATRAJPL'RI, M.S. (1984). Observaoons on the predatory behaviour of Mononchus aquaticus. Nematol. medit. 12, 41-45.
BILGR.A.MI, A.L.; AHMAD, I.& JAIRAJPURI, M.S. (1985a). Predatory behaviour of Aquoiides thornei (Nygobimina: Nematoda). Nematolo^ca (1984) 30. 457-461
BILGRAMI, A.L.; AHMAD, I. & JAIR.AJPL'RI, M.S. (1985b). Factors influencing attraction of adult Hirschinanniella oryzae towards cabbage seedlings. Re\iie Nematol. 8, 67-75.
BILGR-AVn, AL.; AHMAD, I. & JAIR.JJPL'RI. M.S. (1986). A study of ihe intestinal conLnts of some mononchs. Revue Nematol. 9, 191-194.

59
MS Ju'.rajpur: ii.AL Biigrojni 12!
BILGRAMI. AL.; AHMAD, I. & JAIRAJPURI, M.S. (1988). Intcracuon between rr.ononchs in non-stcnle agar plates. Indian J. Nematol. ''1987) 17, 339-340.
BILGRAMI, A.L. & JAIRAJPURI, M.S. (1985 a). Cannibalism in Mononchus aqacacus. IrJian J. Nemawl. 14 (1984) 202-203.
BILGRAMI, A.L. & JAIRAJPURI, M.S. (1985b). The responses of Mononchus fl^uaficor to chemicals and pH. Indian J. Nematol. 14(1984) 171-174.
BILGRAMI, A.L. & JAIRAJPURI, M.S. (1988). Attraction of Monochoides lon-gicaudatus and \f. foriidens (Nematoda: Diplogasterida) towards prey and factors influencing itazcnon. Revue Nematol. 11,195-202.
BILGRAMI, AJL. & JAIRAJPL'RI, M.S. (1989a) Aggregauon of Monochoides lon-gicaudatus and M.fortidens (Nematoda: Diplogastenda) at feeding sites. Nematologica ((1988) 34, 119-121.
BILGR.AMI, A.L. & JAIRAJP' 'RI, M.S. (19S9b). Resistance of prey to predauon and stnke rate of the predators, Mononchoides longicaudaius md M.fortidens (Nematoda: Diplogastenda). Revue Nematol. 12, 4549.
BILGRAMI, AX. & JAIRAJPURI, M.S. (1989c). Effects of concentrauons and thickness of agar on the predatory behaviour of Mononchoides longicaudatus and M.fortidens (Nematoda: Diplogasterida) and on their attraction towards prey. Indian J. Nematol. (1988) 18,222-227.
BILGRAMI, A.L. &. JAIRAJPURI, M.S. Predat:on by Monochoides fomdens and M. longicaudatus (Nematoda: Diplogastenda). Nematologica (In press).
CASSIDY,G. (1931). Some mononchs of Hawaii. WaH-aijf to. ^ec. 35, 305-339.
CHRISTEI, J.R. (1960). Biological control-predaceous nematodes. In Sasser, J. M. & Jenkings, W.R. (eds). Nematology: fundamentals and recent advances wii/i emphasis on plant parasitic and soil forms. Univ. of. N. Carolina Press Chapel Hill. pp. 480.
COBB, N.A. (1917). Tlie mononchs (Mononchus, baslian) a genus of free living nematodes. Soil Sci. 3,431486.
COBB, N.A. (1920). Transfer of nematodes (mononchs) from place to phce for economic purposes. Science 51,640-641.
COBB. N.A. (1929). Ncmas of the genus Dorylaunus attacking the eggs of mites. J ParasitoL 15, 284.
COHN, E., & MORDECH AI, M. vl9T4). Experiments in suppressing :iirus populations by use of mangold and a predacious nematode. Nematol. medxt 2:-i3-53.

GO
122 Predatory Nematodes
COOMANS, A. & LIMA; M.B (1964). Dcscripuon of Anatonchus ameciae n. sp. (Nematoda : Monochidae) with observations on its ju\cnile stages and anatomy. Nemaiologica 11,413-431.
COOPER, A.F. & VAN GUNDY (1970). Metabolism of glycogen and neutral lipids by Aphelenchus avenae and Caerwrhabditis sp. in aerobic, micro-aerobic and anaerobic environments. / . NemaioL 2, 305-315.
CROLL.N.A. (1972). Behavioural activities of nematodes. Helminlhol. Abstr .41(B), :87-105.
CROLL, N.A. (1974). Locomotory behaviour, energeuc, and disposal in nematodes. Adv. Parasitology 12.
DONCASTER, C.C. & SEYMOUR, M.K. (1973). Explorauon and selecuon of penetration site by tylenchids. Nematologica 19, 137-145.
ESSER, RP. (1963). Nematode mtemcuons in plates of non- sterile water agar. Proc. Soil Crop Sa.Soc.Fla. 23, 121-138.
ESSER, RP. (1987). Biological control of nematodes by nematodes. I. Dor>'iamis (Nematoda: Dorylaimida). Nemutology Circular No. 144. pp. 4.
ESSER, R.P. & SOBERS, E.K. (1964). Natural enemies of nematodes. Proc. Soil Crop Sci. Soc. Fla. 24, 326-353.
FERRIS, V.R. (1968). Biometnc analysis in the genus Labronema (Nematoda: Dorylamiida) with a description of Z,. r/io/Tiei n. sp. Nemaiologica 14, 276-284.
GROOTAERT, P.; JAQUES, A. & SMALL, R.W. (1977). Prey Selection in Butlerius sp. (Rhabditida: Diplogastcndae). Med. Fac. Landbouww Rijksumv. Gent 42, 1559-1562.
GROOTAERT, P. & MAERTENS, D. (1976). Culuvation and life cycle of Mononchus aquaticus. Nematologica 22, 173-181.
GROOTAERT, P. & SMALL, R.W. (1982) Aspects of the biology of Labronema vulvapapillaium (Mcyl) (Nematoda: Dorjiaimida). in bboratory culture. Biol. Jb. Dodonaea 50. 135-1.48.
GROOTAERT, P. & WYSS. U. (1978). The ultrasiructure and function of the anterior feeding apparatus in Labronema sp Nematologica 24, 243-250.
GROOTAERT, P. & WYSS, U. (1970). Ultrastnicture and function of the antenor feeding apparatus m Mononchus aquaticus. Nematologica 25.: 163-173.

1 o l
MS.JairajpuT-i&AL.Bilgraml 123
H E C H L E R , H.C. (1963). Description, developmental biology and feeding habits of Siinura lenuicaudata (de Man). J. B. Goodey, i960 (Nematoda : Aphelenchida), a nematode predator. Proc. helmruh. Soc. Wash. 30, 182-195.
HOLLINGS, C.S. (1966). The functional response of invertebrate predators lo prey density. Mem. Ent. Soc. Can. 48,1-86.
HOlxIS, I.P. (1957). Cukuiul studies with Trkhodorus cknsici on tomato roots. / . Sirmtol. 5, 22^-219.
JAIRAJPURI. M.S. &AZMI,M.I. (1978). Some studies on tlie predatory behaviour oi Mylonchulus dentalus. Nematol. medit.. 6, 205-212.
JAIRAJPURI, M.S.; BAQRI, Q.H. & AHMAD, S. (1973), Nematodes of hi^. altitudes in India I. Four new species of Tylencyhida. Nematologka 19,19-30.
JENSON, HJ. & MULVEY, R.H. (1968). Predacious nematode (Mononchidae) of Oregon. Oregon. State Monognph; Oregon State University press. 12,1-57
JONES, F.G.W, (1974). Control of nematode pest, background and outlook for biological control. In Price-Jones, D &. Solomon. MJE. (eds.). Biology in Pest and disease control. 13th Symposium British Ecological Society Oxford.
LAL, A.; SANWAL. K.C. & MATHUR, V.K. (1983). Changes in the nematode populations of undisturbed land with the introduction of land development practices and cropping sequences. Indian .f. Nematol. 13, 133-140.
LEE, D.L. & ATKINSON, HJ. {1916). Physiology of Nematodes. The Macmillian Press, London. Ltd pp. 215.
LINFORD, M.B. & OLIVIERA, J.M. (1937). The feeding of hollow spear nematodes on other nematodes. Science 85, 295-297.
MAERTENS, D. (1976). Observations on the life cycle of Prionchuluspunaatus (Cobb, 1917) and cultural conditions, flio/. i* . Diidond^fl. 43, 197-218.
MAH.-srATRA, N.K. & RAO, Y.S. (1981) . Bionomics of lotonchus monohystera, a predxeous nematode in rice soil. Proc.Indian Acad.Parasitol. 2, 85-87.
MCLAREN, D J . (1976). Nematode sense organs. In Dawes, B. (ed.) Adv. Parc^sitoloty, 14, Academic Press, London, pp. 195-265.
MOHANDAS. C. &. PRABHOO, NJl. (1980). The feeding behaviour and food preference of predatory nematodes (Mononchida) from the soil of Kerala, India. i?evu« <i'EcologieetdeBialogie,du.Sol. 17,153-160.
MLXVBY. R.H. (1961). The Monochidae : A family of prcdaceoiis nematodes I. Genus: ^ /^ylonckulus (EnocHda r Mooochidae), Can. J. Zool. 39,666-6%.

w
124 Predator, Sematodi.s
KELMES, AJ (1974) Evaluaiion of Ihe feeding behaviour of Pnonchulus punctaius (Cobb), A nematode predator. J Aran Ecol. 43, 553-565.
NELMES, AJ. & MCCULLOCH, J S. (1975). Number of mononchid nematodes in soil sown \a cereals and grasses Annls. app Bwl. 79, 231-242.
ORTON WILLIAMS, KJ. & JAIRAJPURI, M.S. (1984). Observation on the morphology and systemaacs of Thalassogenus Andrassy, 1973 (Nematoda : Thalassogenendae) n.[2Lm.RecAus:rahanMus 36, 1-6.
POINER, G.O. Jr. & HANSEN, EL. (1986). Association between nematodes and bactena. Helrmnihol Absir 5erB-55,61-81.
HITTER, M. & LAUMOUND, C. (1975) Review of the use of nematodes in biological control programmes against parasites and pests of cultivated plants. Bull. des. Resch. Agrvno.des Gem Sem d'EludeAgncet Htjej desPlants pp. 331-344,
RIVERS AT, G (1981a) Effects of ageing and starvauon on respirauon and food reserve contents in adult Ilirschmanruelia spinicaudata Revue Nematol 4,125-130.
RIVERSAT, G. (1981b). Consupmuon of food reserves by starved second stage juveniles of Meloidogynejavanica under condiuons inducing osmosis. Nematologica 11, 207-214.
ROGGEN, D.R. (1973). Functional morphology of the nematode pharynx I. theory of the soft wailed cylindrical phar>nx. Nematologica 19,349-365.
SEYMOL'R. M K; WRIGHT, K.A. & DONCASTER, C.C. (1983). The action of antenor feeding apparatus of Caenorhabdiuselegans (Nematoda : Rhabditida) / Zool 201, 507-539.
SHAFQAT, S.; BILGRAMI, A.L. & JAIRAJPURI, MS (1987). Evaluauon of the prcdation ability ol Dorylaimus siagnalis Dujardin, 1845 (Nematoda : Dorylaimida). Revue Nematol 10, 455-461.
SMALL, R W (1977). The culture and ecology of predatory nematode populauons. spcciailv o\ Pnonchulus punaatus. Ph.D. thesis. University of Lonodon ,451 pp.
SMALL, R W (1979; The effects of predatory nematodes on populauon of plant parasitic nematodes in pots Nematologica 25, 94-103
SMALL. R W. & GROOTAERT, P (1983). Observations on the prcdation abiiiucs of some iOii uweiling predatorv nematodes. Nematologica 29, 109-118
SOLOMON, M F .(1949) The natural control ol animal population J A mm Ecol 18. 1-35
SPRINGETT J A (1«64) A m ihod of cultunng Enchytrcids. (9iioT 15, 175-177.

u3
MS Jairajpiu-i <i. AL Bdgrami 123
STEINER, G. & HEiNLEY, H. (1922). The possiDility of control of Hetewdera radic'.cola and other plant mjunc s remas by means of predatory ncmas specially by 'for.ortchiispapiUatus Basaan./.'.. jsh. Acad, Sci 12, 267-385.
SZCYGIEL, A. (1971). Wystepcwine drapienych nicieni 2 rodziny Monochidae w. glekuch uprawynch >v./'oL i:/:e.2<;52. Probl.Postep Nauk.roln. 121,145-1^8.
TKORNE, G. (1924). Utah nematodes of the gtnusMononchusTrans Am. Microscope 5oc. 43,157-171.
THORNE, G. (1927). The life hiitory, habits and economic importance of some mononchs. J.Agric.Res. 34, 265-286.
THORNE, G. (1930). Predaceous nemas of the genus Nygolaimus and new genus Sectonema.J.Agric.Res. 44, 445-4^.
ThORNE, G. (1932). Apparant successful transfer of Mononchus papdlatus, M. mus-corum and M. amphigonicus from Utah to Cahfornia. /. Parasitol. 19,90.
THORNE, G. (1939). A monograph of the nematodes of the super-family Dorylaimoidea. Capita Zool. 8,1-261.
VANDER HEIDEN (1974). The strjcture of the anterior feeding apparatus in members of the Ironidae. (Nematode: Enopiida). Nema'.ologica 20,419-436.
W.ALLACE, H.R. (1963) The biology of plant parasitic nematodes. Edward Arnold. London. 280 pp.
WALLACE, H.R. (1969). Wave formation of infective larvae of the plant parasitic nematodes, Meloidogynejavanica. Nematologica 15, 65-75.
W.ARD, S. (1976). Chemotaxis by the nematode Caenorhabditiselegans. Identification of attractants and analysis of the response by use of mutants./"roc.jVa/..Acad. 5c;. USA 70,817-821.
WEBSTER, J.M. (1972). Nematode cJ biological control. In Websi£r, JM. (ed.). Econonuc Nematology: Academic Pre^s. Ne-w York. 563 pp.
WOOD, F.H. (1973). Feeding rebtionship of soil duelling nematodes. Soil Biol. Biocnem. 5,593-601.
WRIGHT, K.A. (1983). Nematode sonsi' a: form ana function./. Nemaim , 15.151-158.
WYSS, U. & GROOTAERT. P iI97"l Feeding mechanism o't Labronema \-ul\apapii-lj:.jn Med Fac. LandhouHw RrLuniv Gent 42. 1521-1527.
YEATES, G.W. (1969). PrcdJtion by Mononcr.ouics potohiL.is (Xcmatodj-Diplo^jstcndjc) in lalx rator)' cjlture. .Vt7mi.ViV'i,';i,a 15, 1-9.

j 4
11
DIPLOGASTERID PREDATORS
ANWAR L BILGRAMI
Department of Zoology,
Aligarh Muslim University, Aligarh 202002, India.
Introduction
The diplogasterid predators which feed by cutting and sucking their prey remained largely neglected until Yeates (1969) evaluated the predatory abilides of Diplemeron colobocercus {=Mononchoides potokikus). Subsequent studies on these predators have brought to light more informadon on their biology, behaviour, predator-prey relationships, ecology, predation abilities, etc. Grootaert (1976) cultured Mesodiplogaster Iherineri and studied its population growth, reproduction , egg production, embrj-ohic and post-embryonic developments and effect of temperatures on its development. Grootaert e: al. (1977) studied prey selection by Butlehus sp., and elucidated some aspects of its feeding habit and possible application of the same in the biological control of plant parasitic nematodes. While observing the predation abilities of Buderiusdegrissei Small &. Grootaert (1983) [oundPana§re!IusrediMviis and the first and second stages of Rhabdius oxycerca as most suscepnble prey species. Very recendy, a comprehensive study on the predatory abihties of two species of diplogasterid predators belonging to die genus Mononchoides viz.,.V/. longicaudaius and M.fortidens was made and the role of these predators as agents of biological control was assessed. These studies include pre- and posi-fccding aitmcuon and aggreganon around the prey at feeding sites
.NE.MATODG BiO CONTROL. ASPECTS A PROiFECTS (Eds — M S Jairajpun, S1..M. AUm i I AhmaJ) CBS PubLshcrs &. Dssmbuton, Ddhi, India 1990

136 Diplogasterid Predators
(Bilgrami & Jairajpuri, 1989a); attraction of predators towards excised and non-excised Ciive) prey and factors influencing at*j3c:icr. (Bi!^'~.; & Jairajpuri, 1988); strike rate of tie predators and resistance and susce-dr'Iir/ of "srr/ letpatodes to predation (Bilgrami &. Jairajpuri, 1989b); prey c^tchiag ' . ' feeding mechanisms, prey preference etc., (Bilgrami & Jairajpijri. in rrecs) :„x-':ic:ors influencing predation and attraction (Bilgrami & Jairajpuri, ]9S9a; 1989c).
In the following the potential of diplogasteridi as predators of nematodes is assessed and their possible role in the biological control of plant parasitic nematodes is evaluated.
Feeding Apparatus
The nematodes of the Order Diplogasterida possess a comparatively small buccal cavity armed with teeth of variable sizes which are located at different positions. There is a strong well developed movable dorsal tooth (Jairajpuri et al., 1973). The large tooth is the main killing weapon and is responsible for injecting the oesophageal secretions into the body of the prey for the purpose of exira-corporeal digestion. The entire feeding apparatus is of cutting and sucking type.
Aggregation and Attraction
The ability of diplogasterid predators to perceive prey attractants (Bilgrami & Jaiijapuri, 1988), to aggregate around an injured prey at feeding sites (Yeates, 1969; Bilgrami & Jairajpuri, 19S9a) and to feed in groups on a single prey at feading sites (Yeates, 1969; Bilgrami &. Jairajpuri, in press) seems to put forth some interesting aspects which could lead to establish a strong predator-prey relationship. It may not be out of place to discuss these mechanisms here, since th^se have been observed in diplopsterid predators only and could lead to a better understanding of the mechanisms involved and their possible impact on the responses of predators in relation to their utility in the biological control programmes. Mononchoides longicaudatus and Af./om'Jer.s respond positively towards Ii\e and excised bacteriophagous nematodes {Acrobcbidcs sp., Cephalobus sp., Rhabaiiis sp., Panagreilus redivivus); ecioparasitic nematodes [Tnchodoriis sp., Xipnmena amcncanum, Lon^dorus sp., TyienchorhynchiiS mash-hoodi, Paralon'^doms sp., Hirsohmanr.i'jtla oryzae, Hemicnconemoidcs mzr.zfcrac, Hoplolaimi's ind:cus, Heiicccyienchus •.r,di"ds); endoparasitic nematodes (second stage juveniles of Meloido{;}ne'.ncosni[a and An^:i:r.a :riitci) (Bilgrami & J;iirjjpun, N8S) as well as bactena (Yeates, 1969; Bilgnimi & Jairajpuri, 1988). Tlie attraction and aggregation has also been ob.sen'ed in dop.ljim predators viz., Labronema \'ulvap.ir:'.lMiun, AqiLiiides thornci. and Doni^urius s:j'>:r.i2::s I WN-SS & Grootaeri, 1977; Bilgrani c'r J/..

u
AL. Bilgrami 137
1985; Shafqat et al, 1987). Though, this suggests some kind of chemoattraction in dorylaims but it appears mild and insignificant as only a few prediitors (2-4) aggregate around a prey requiring more time to reach the feeding sites. However, in case of diplogasterids it appears more evident and positive as more predatc>rs aggregate around the prey (up to ten) taking lesser time to reach the feeding site (tlie aggregation stans within minutes after the prey is punctured by another predator).
The degree of attraction, duration of post-feeding aggregation, duration of feeding sites and actual feeding time m.ay vary from individual to individual and species to species depe.-iding upon the type of prey, composition, concenlration, quality and quantity of prey secretions/attractants. These may be important determinant factors, besides those studied by Bilgrami & Jairajpuri (1988; 1989c). The lower and higher temperatures other than the optimum also adversely affect both the attraction and predation. These factors could thus be manipulated to modify the activities of predators to maximum extent.
Prey Catching and Feeding Mechanisms
The diplogasterid predators feed by puncturing the cuticle of prey and sucking its body contents. However, M.fonidens can devour whole the first and second stage juveniles of small prey nematodes like ,4 cTcie/o/kfej sp., and Cephalobus sp., and cut larger prey into pieces to feed upon them. The feeding/ingestion is helped by the suction force created by the oesophagous and the movable dorsal tooth. The duration of feeding varies in different indiuduals and depends upon the size and t>pe of prey, volume of prey contents and texture of the prey cuticle.
The positive attraction of diplogasterids towards prey, instantaneous prey catching and attack, faster rate of feeding, high rate of predation, bacteriophagous nature, besides being predaceous. more tolerent to ad\erse conditions as evident from-the formation of daurve larvae, shon life cycles, easy Culture conditions, wide range of prey and probably higher longivit>, are some of the characteristics which suggest these predators as efficient agents of biological control ofplant-parasitic nematodes. In diplogasterids chemoattraction as well as chance encounters bring about predator-prey contacts. The predator-prey encounter is an important factor which initiates predation and most likely governs the rate of predation. The improvement and increase in the number of such encounters should also increase the rate of predation. Tne ability of diplogasicrid predators to poreei\e prey aitractanis could lead predators to move directly towards prey and enhance chances of more frequent prcdaior-prcy contacts rcsulung in higher rate of killing. This could be one of the rca.sons that these predators possess a high rate of predation.

1u7
13S Diplogasterid Predators
All diplogajterid ps'edators viz., Mononchoides longicaudatus, M. fortidens, But-leriussp.,Buderi'usdegrissei and Diplenteron cclobocercis get hold of their prey with the help of high suction force created by their oesophagus. The prey catching and attack is instantaneous and only one or two attacks are enough to pucture the cuticle of prey. These predators attack and puncture the cuticle of their prey with Lhe help of their movable donal tooth (Yeates, 1969; Grootaert et al., 1977; Small & Grootaert, 1983; Bilgrami & Jairajpuri, in press). The oesophageal suction enables predators to take a firm grip on their prey and prevent its escape.
Probing in dipiogasterid predators lasts only for a v:;ry short duration. The diplogasterid predators also feed upon bacteria in the absence of prey nematodes (Bilgrami &Jairajpuri; in press). The bacteriophagous nature ;«ems to be an important phenomenon and this may avoid intra-specific interxtions among the fffcdators themselves {cannibalism).
Prey Range and Preference
The predator-prey relationship and range of pa-ey are unportant aspects to evaluate predaceous activities of predators and in assessing their utility in the biological control programmes. The diplogasterid predators which have a wide range of prey nematodes (Table-I) also show diiTerent degrees of preference for diJTerent species of prey. Cbser\-ations on Budenus sp., carried out by Grootaert sf a/. (1977) and their results on prey selection conform the general conclusions drawn by Esser (1963) for the predatory dorylaims and mononchs. The soO stages of endo-parasitic nematodes are preferred most by the predators. Buderius sp., attacked, wounded and fed upon a number of free-living, ecto- and endo-parasitic nematodes. Buderius degressei killed more PanagreUus redivivus 'than Rhabditis oxycerca (Small & Grootaert. 1983). Bilgrami & Jairaipuri (in press) observed M.-for.idens and M. longicaudatus preferring Acrobeloides sp., Cepkalobus sp., P. redivivus, Hirschmanniella oryzae and the second stage ju%eniies of M. incognita, and Anguina triad. Diplenteron colobocercus, however, did not prefer any particular type of prey (Yeates, 1969).
Tne prey selectioa'prefcrence depends on the activity, size and behaviour of prey nematodes. Soft-bodied and less active nematodes being preferred most as compared to !ajge or more active prey nematodc-s (Bilgrami Hal., 1983). It may also depend on the predators as some are stronger v.hiie others hase low predatory potentials.
Theoretically, the most suitable (attractive) prey should also be killed to the maximum e.xicnL This, however, does not seem to be the case. H. oryzae, Tyier.chor-hynchus mas'n'iwodi and lloplolaimus indicus attracted maximum number of M. lon-^^iciudatus and .M. fonidens. The fonner two prey were killed in moderate numbers whereas indi\iduals oilloplolaimus indicus ox Ilemicriconemoides mang.fcrze was

I b
AL.Bilgraml 139
neither killed nor injured by these predators. In contrast, Meloidogyne incognita and Anguina triiici juveniles which are preysd upon more by the predators attracted moderate number of M. longicaudatus and M.fortidens. It seems that the prey preference of these predators and probably that of others is not solely dependent upon the prey secretions/attractants. Tne ability of predators to perceive chemical attractanls may, however, certainly help in locating prey and in increasing predation on a particular type of prey by increasing predator-prey encounters. The differential conditions such as confinment of prey during attraction and allowing prey to move freely during predation experiments may have caused the differences. Besides, the predation is a physical phenomenon, which essentially requires a lip con'act of the predators with the prey, depends upon the activity, escape response, texture and thickness of the cuticle of prey nematodes, number of prey, temperature and the inert behaviour of the predators themselves (ability of predators to attack prey). In contrast, attraction being a chemical phenomenon may not neccessarily be affected by these factors but will depend on the composition, concentration, quality and quantity of the prey secretions, besides how fast the attractants are dispersed in the surrounding area.
Biology and Culture
To execute any biological control programme successfully against plant parasitic nematodes in field conditions a huge population of predators, advance knowledge of the populations of predators and beuer understanding of their biology, behaviour, prey range, preference, ecology etc., is required. The inability so far, to raise . iss cultures of the predators seems to have been the real hurdle in combatmg populations of plant parasitic nematodes. Yeates (1969) cultivated D. colobocercus in agar plates using a bacterium Bacilluscerusvar mycoides. Bilgrami & Jairajpuri (1988) uscdRhabdids sp., m agar plates supplemented with infant milk powder (Lactogen) for rearing M.fonidens and M. longicaudatus in vitro. The milk powder boosted bacteria! growth and ser-'ed as the primary source of food for the prey nematodes. Grootaert (1976) cultured Mesodiplogastcr ihenten and studied its population groMh, reproduction, egg production, embryonic ana post embn.onic developments and effects of temperatures on its development. The total duration of embryonic and post-embryonic de\tlopments in M.fortidens has been found :o rang: between 5-7 days (Q. Tahseen, personal communication). Since diplogastcnd predates also feed upon bactcna and thr.ve equally v.ei! in absence of nematodes ihcy can easily be cultured using bactcna as the source of food. Their populations can also be elevated to desired levels by supplementing organic amendments which support bactenal growth under natural conditions. The studies on the biology and culture techniques of these predators are fc.'. and rcquue more studies.

1^9
140 Diplogasterid Predators
Conclusions
The studies en dipiogastend predators carried out so far indicate their abilitv as promising agents of biological control. Tne mam advantages of using these predators are their ability to perceive prey attractants (predation may increiise), high rate of predation, shon life cycle and high fecundity (build up of population), formation of dauer larvae and longivity and bactenophagous nature. Besides, iheyare easy to culture. The population of diplogastend predators can be maintained by adding organic amendments (Webster, 1972). The prospects of using diplogastend predators in the biological control programmes seem to be bnght. However, further studies on their biology, ecology, behaviour, reprodocuve potentials, fecundity, longivity, range of prey and predator prey relationships are requu-ed. These studies will further help m determining theu- role as predators of plant parasiDc nematodes and the possibilities of using them m fields.
TABLE I
PREY NEMATODES OF DIPLOGASTERID PREDATORS
Predators Prey Reference
Mononchoides AngmnatTitici]\., fortidens Mebidogyneincogrutajv
Hirsckmanmella oryzae, Tylenchorhynchus maskhoodi, Hehcocylenchusindicus, Longtdorous sp , Xiphinema amencanum^ Tnchodrussp. Fanagrellus rediMvic: Acwbebidessp , Cepr^lobus sp , Hoplolaimus indic'^,* Hermcriconemoides mangife'ae *
Bilgrami & Jau-ajpun, (m press)
Mononchoides longicaiidatus
•do — Bilgrami & Jairajpun (in press^
Mononchoides Mesorkabduis htioralis, coLobocercus Panji^rolaimusaustrjlis,
ALrcbeloidessynisus, Zc'L.J punna. Ercpionema injlj!X'^i T,i\.iLa \',aipiikea. Emnolatmus sp , E^r^stonuna sp ,Nygo!aimusdirectus, DiscolainL^m sabuh. MyloncnJus
Ycaics i!%Q)

1 7 0
psammophilus.
Butlerius PanagreHusredivivus, Rhabditis degressei oxycerca, Meloidogyne naasi*
Ciobcdera rostochiensis, Rotylenchus fallorobustus* Labronema vulvapapillatum*
Butlerius sp. Tylenckorkynchus sp.,* Rotylenchus robustus* Tylenchus macrums, M. naasi, Merlinius sp.,* Aphelenchoides fragrariaeJ'ratylenchussp*.,iy.Hemi-cycliophora sp.* R. oxycerca*,Pelcdera sp.* Cephalobus sp.,* Rhabdontolaimus stigmatus, Mesodiplogaster Iherideri, Plectus sp*. Tripyla sp., Alaimus sp. Mononchus aqundcus jv.Mesodorylaimus sp.,* Eudarylaimus sp., Trichodorus sparsus*
A.L.Bilgrami 141
Small & Grootaert (1983)
Grootaert, Jaaues &SmaU(1977)
*Prey attacked by predators but not wounded. Predators feed only when the prey is
artificially wounded.
REFERENCES
BILGR AMI, AI-.; AHMAD, I. &JAIRAJPURI,M.S. (1983). Some factors influencing
predation by Mononchus aquaticus. Revue Nematol. 6, 325-326.
BILGRAMI, A.L. & JAIR.-UPURI, M.S. (1988). Attraction of Mononchoides lon-gicaudatus and M. fortidens (Nematoda : Diplogasterida) towards prey and factors influencing a.im.z^Qn. Revue Nematol. 11,195-202.
BILGRAMI, A.L. & JAIR.AJPURI, M.S. (19S9a). Aggregation df Mononchoides lon-
gicaudatus and .\!. fortidens (Nematoda: Diplogastcrida) at feeding sii£S. Nemaiologica
(1988), 34, 119-121.
BILGRAMI. A.L. & JAIR.AJPLT^I, M.S. (1989b). Resistance of prey to predaticn and
strike rate of predators, Mononchoides longicaudatus and M. fortidens (Nematoda:
Diplogastcrida!. Revue h'eirjitol. 12,45-49.
BILGRAMI, A.L. &JAIR.^JPURI.NLS. (1989c). Effect of concentration and thickness
of agar on the predator)' behaviour of Mononchoides longicaudatus and M. fortidens
(NomatCKla: Diploga-Stcrida) and oi; their attraction towards prey. Indian J. Ncn^tol.
18, 222-227.

17
142 Diplogasterid Predators
BILGRAMI, A.L. & JAIRAJPURI, M.S. Predation by Mononchoides fortidens and M. longicaiuiatus (Nematcda : Diplcgasterida). Nematologica (In press).
ESSER, RJ*. (1963). Nematode interactions in plates of non-sterile water agar. Proc. Soil. Crop Sci. Fla. 23, 121-138.
GROOTAERT, P. & 3MALL, R.W. (1982). Aspects of the biology of Labronema vulvapapillatum (Me>i) (Nematoda, Doryiaimidae) in Laboratory culture. Biol. Fb. Dodonaea SO, 135-1-8.
GROOTAERT, P.; JAQUES, A. & SMALL, R.W. (1977). Prey selection in Butlerius sp. (Rhabditida: Diplogasteridae). Med. Fac. Londbouww Rijksuniv. Geru. 42, 1559-1562.
SMALL.R.W. & GRGOTAERT.P. (1983). Observations on the predation ability of some soil dwelling predatory nematodes. Nematologica 29,109-118.
SHAFQAT, S.; BE.GRAMI, A.L. & JAIRAJPURL M.S. (1987). Evaluation of the predation ability of Dorylaimus stagnalis Dujardin, 1845 (Nematoda; Dorylaimida). Revue Nematol 10, 455-461.
WEBSTER, J. M. (1972). Nematodes and Biological control. In Webster, J. M. (ed). Economic Nematology, Academic Press, New York, 563 pp.
WYSS, U. & GROCAERT, P. (1977). Feeding mechanism of Labronema vulvapapillatum. Med. Fac. Laruibouww. Rijksuniv. Gent 42, 1521-1527.
YEATES, G.W. (1969). Predation by Mononchoides potohikus (Nematoda: Diplogasteridae) in laboratory culture. Nematologica 15,1-9.

7 7
12 PREDACEOUS DORYLAIMS
.^\WAR L BILGRAMI
Department of Zoology,
Aligarh Muslim University, Aligarh, 202002, India.
Introduction
Dorylaims are perhaps one of the most ubiquitous group of nematodes occurring in all types of climates and habitats. The presence of two, three or more genera of dorylaims at one place/ field is quite usual and their abundance has been estimated to be 200-500 million/acre (Thorne, 1930). Thome (1930; 1939) had suggested that dorylaims and nygolaims may be predaceous. Since then many species belonging to these groups have been reported as predators of phytophagous nematodes. It is quite possible that many more species of dorylaims and nygolaims may be predatory. Further investigations are needed to identify such species and to establish their predaceous nature, besides carrying out studies for assessing their role as agents of biological control. The dorylaim predators are usually larger than their prey and possess a large, hollow protrusible stylet (odontostyle) or a mural tooth.
Feeding Mechanism
The dorylaim predators cannot engulf their prey wjiole or cut ihem into pieces as their feeding apparatus is of piercing and sucking t>pe. They thus, puncture the cuticle of prey with the stylot/tooih and suck its body contents. Predauon depends largely upon chance encounters with tiie prey but there is some evidence of chcmoatu^action (Esscr,
N'E.MATODE BIO-CONTROL: ASPECTS i PROSPECTS i,Eas,—M.S. iairaipun, M..M. AUm Ji. I. Ahmad). TBS Publishers &. Disinbutors, Delhi, India. 1990.

173
144 Predaceous Doryla'jns
1965) and then- aggregation around an injured prey at feeding sites ("Esser, 1963, Biigrami etai, 1985 Shafqat etai, 1987), though it may not be very significant. The attraction occursonly when the prey nematodes are either excised artificially or wounded/killed hj ancLher predator. Tiiis -^tms to have lifJe impact en the possibiliues of increased predator-prey encounters.
Most dorylaim predators usually have a low rate of predation (Wyss & Grootaert, 1977; Biigrami e/c / , 1985) as ccnpared to mononchs (Nelmes, 197-; Jairajpun &. Azmi, 1978; or dipiogastends (Biigrami i Jairajpun, m press) as is evident from the experimental studies. Whether the same occ TS in nature also remains yet to be determined. Like other predators the dorylamis fir: probe their prey upon contact pnor to attack. This may also be reierred to as pre-penetration exploration or probing (Esser, 1987). Tne dorylairr predators show a strong aiTia,:y for theu- own eggs as they probe and explore thera with iheir lips for quite some ume without making any attempt to feed upon them. They, however, attack and suck the contents of eggs of other nematodes (Esser, 1963; Biigrami et al., 1985; Shafqat et al., 1987). This phenomenon is not reported in other grotips of predators.
Prey Nematodes of Dorylaim Predators
The dorylaims attack and feed on many species of nematodes, oligochaetes, roufers and other mvenebrates in addition to a wide vanety of invertebrate eggs (Table-I). However, regarGing the food of Donlaimus spp.. Theme & Swanger (1936) commented "The intestinal contents of hundreds of doryiaims have been observed. Generally they have been filled with matenal from the higher plants, indicating that the;se are their preferred soiirce of food. Frequently, the intesunes are filled with highly coloured matter that can be identified as probably coming from bnght coloured roots among which they are ccDected. ?vlatter apparently onginaung from algae fill the intestine of aquaDc forms. Frequently, specimens are obserNed in such situations which suggest them to be predaceous. The author has repeatedly found larval specimens of Dorylaimus orscurus in the cysts Oi" Sic sugar-beet nematode, Heieroaera schachtu and on two occasions they have been observed with eggs impaled on spe^ " Cobb (1929) and others (Thome, 1939; Sieiner, 1924), howeser, had provided evidences which indicated the preoiceous namre of some dorylaim species. Cobo (1 c) observed two species of dor}'laims feeaing en t'-.e eggs of mitcs m deca>ing n.:TCissus buibs. The eggs of H schachni were found !~:a.,ea en Lhe styict of acu "-ddu^^iids obtus.csiidaius (= Dorylaimus obfdsica.^jrjs) (T::cr"e,1939). Stcincr (1924) obsc.".ed setae of c'lgochactes in the intestine of D '•esius. Thome (1930,1939) found species o\ Aporcelaimus. Nygolaimus and Seccnema and at least one species of Labrone-^i (L fc-ox) feeding mostly on small oligo:.hactcs. Theme also observed Lhat almost all species ol Ducolainv.LS could be prcJjceoj:> and

AL. Bilgrami 145
some possibily be nemativorous. Linford & Oliviera (1937) determined ten species of Dorylaimus, two ofDiscolaimusand. one ofActinolaimus to be predators of nematodes. It seems that manyspeciesof dor>1aims and those of related genera are omnivorous and derive their food from various organisms like algae, fungi, rotifers etc. (7able-I). Hollis (1957) found the latter to be true for Thonus etiersbergensis (= Dorylaimus ettersbergensis) as it fed on various micro-organisms including the globose vegetative cells of blue-green algae (Chroococcus sp.), two species of green algae {Chlorellavulgariscmd Teraedron sp.), the globose cysts of a ciliated infusorium {Drepanomonas sp.), besides feeding on nematodes. Phytophagous nematodes viz., Hoplolaimus tylenchiformis, H. indicus, Belonolaimus longicaudatus, Hemicycliophora similis, DoUchodoms heterocephalus, Criconemoides spp., Scutellonema spp. and Helicotylenchus indicus have shown resistance to predation by different predators.
Biological Control Potential
The widespread and abundant presence of predaceous dorylaims in soils could possibly indicate some biological control activity by these predators. However, it is not known to what extent and under what conditions the populations of phytophagous nematodes are reduced since, such an evaluation has never been made to provide evidence to this effect. The dorylaims, in large numbers, may certainly ha%« an advantage over other predators which have localized populations. These predators need only slight modifications in the field conditions to maintain and establish themselves in high populations. Being omnivorous, i.e., feeding on algae, fungi, nematodes and small invertebrates, it may be conceived that they could be raised on algal or fungal cultures for utilization in field conditions for biological control. The detection of Eudorylaimus obtusicaudatus feeding on the eggs inside the H. schackai cysts or the increase in the population of predators {Thornia sp.) in presence of citrus nematode and decrease in absence of them in pot trails (Boosalis &. Mankau, 1965) indicates a potential rather than a casual role of these nematodes in the bio-control of plant-parasitic nematodes.
The biological control oi plant parasitic nematodes is an attractive and unexplored field of study awaiung further research on different aspects of the biology of predators specially under the natural conditions. These Lnvestigations will provide much needed informauons which could lead to the practical utility of doPi'laims as predators of plant parasitic nematodes.

i 75
146 Predaceous Dorjla.ir.s
TABLE I
PREDACEOUS DORYLAIMS A ND THEIR PREY
Predators Prev Refsrence
Aporcalaimus sp.
Actinoiaimus sp. Aquandes tkornei
Aporcelaimus sp.*
Aporcelaimellus sp.* Carcharolaimus sp.
Carcharolaimus sp. Dorylaimus obtusi-caudatus (= Aporcelaimellus obtusicaudaius)
D.carteri D. obscurus
D. stagnalis
Dorylaimus sp.
D. ettcrsborgcnsis (= T'tionus er.crsber^cn'iis sp),
D'.scoLiunus .-.p.
Discc-lc:in\L^ ^\t.
Bacteriophagous nematodes
Meloidogyne sp. Chiloplacus symmetricus Rhabditis sp,. Ac^obeloides sp., Meloidogyne sp.
Rotylenchulus reruforrms Meloidogyne sp. Acrobeles sp. Eses oiHeterodera schachtii
Eggs of mites Eggs of//, schachni
Rhabdius sp., Cepnalobus sp.. Acrobeloiaes sp., Tylenchorhynchus mashhoodi. Hoplolaimus indicus.* Heiicocylenckus indicus.* Hirsch-manniella oryzae. Meloidogyns. sp.
3Iue green aigae (Ch''oococcus sp green algae {C'niorcila \ul-^.:r:sjeraca'-cn sp.l, globose CN' ti of eiiuiej intusonum
ci lungus yC.r''.Jic'.'':(:r;x7! so.) ^"".ciicnc.^ sr..
\?".C'.L~.C".'.ii> ^?
Linford & Oliviera, (1937). SmaU (1979) Bilgrami era/. (1985)
Bilgrami (Unpublishec
Linford & Oliviera (1937) Esser (1963) Cobb (1929)
Cobb (1929) Thome & Swanger, (1936) Shafqateffl/. (1987)
Linford & Oliviera, (1937) Hoili: ^557)
Linfcra i OIi\iera (1937) Ebser i,'i^t>3)

i / M
Discolaimiis sp.*
Discolaimoides sp. Eudorylaimus sp.
Eudorylciim-ds sp.* LabronervM. vulvapapiliatum
L. vulvapapiliatum
Labronema sp.* Laimydorus sp.* Mesodorylaimus sp.* Mesodorylaimus goodeyi* Neoactinolaim^ duplicidentatus Nygolaimus sp. Thornia sp.
Acrobeloides sp., Meloidogyne incognita — do — Algae, f\mgi, Bursapkelenchus zylophclus Acrobeloides sp. iW. incognita Aphslenchus avenae, Anguina tritici, Pratylenchus penetrans;"* Xipkinema index. Clobodera rostochiensis, Meloidogyne naasi, Rhabditis oxycerca,*" Panagrellus redivlvus Rorylenchus fallorobustus.** Acrobeloides sp.,A/. incognita —do—
Pjedivivus,R. oxycerca;** R. fallorobustus.** Acrobeloides sp.,M. incognita Amneba. sp. A. avenae. Tylenchulus semipenetrans
A.L. Bilgrarr.i 147
Bilgrami (Unpublished)
— do — Thome (1930)
Bilgrami (Unpublished) Wyss & Grootaert (1977)
Small &. Grootaert, (1983)
Bilgrami (Unpublished)
—do—
Small & Grootaert (1983) Bilgrami (Unpublished) Boosaiis & Mankau, (1965)
*Unidentir~ied species, obser\-ations based on feeding in cultures.
**Prey nematodes were nerJier wounded nor killed by the predators except species of Helicotylenchus where wounding took place but predators were repelled.
REFERELNCES
B1LGR.\MI, A.L.;AHM.AD,I. &JAIR.\JPURI,M.S. (1985). Predatory behav.our of Aquav.des thomei (Dorrjimida: N)Xohimiv,a)..'»'en:atoiogica (1984), 30,457-46-.
BILGR-ANn. A.L. & JAIR-^JPURI, M.S. (19SS . Attracuon of Mononckoidcs Icn-
§icaujj.'u.s :ind \{. fortidcrs (Ncnutoda: Diplogasionda) towards prey and fcctcrs influencing attraction. Revuv SerrutcL II. 195-202.
BILGRAMI, A.L. &. JAIRAJPLT:!, M.S. Prcdator>- abihnjs of Mononcr.oiJcs Icr.-
gicau.ia::^<: and Mononchoist's fcry.der.s i NcnutOvia: DipiogJstenda) and factors in
fluencing prcd.uion. Nc!rvj::loi^i:.z (In press).

I / /
148 Predaceous Dorylaims
BOOSALIS, M-G. & MANKAU, R. (1965). Parasitism and predation of soO microorganisms. In Baker, ifCF. & Synder, W.C. (eds). Ecology of soil borne pathogens. Univ. Calif. Press., Berkeley, pp 374- 391.
C0B3,N.A.(I929). Nemas of the genus Dorylaimus attacking the eggs of mites. Science 51,640-641.
ESSER, R.P. (1963). Nen:ato(fe interactic-ns in plates of non-sterile water agar. Soil & CropSci. Soc. Flo, 23,121-138.
ESSER, R.P. (19S7). Biological control of nematodes by nematodes!. Doryiaims (Nematoda: Yycry\3lmii2L).Nema:ology circular'Ho 144, pp 1-4.
MOLLIS, JP.(195Tj. Cultural studies with Dorylaimus ettersbergensis. Phyiopathology 47, 468-473.
JAIRAJPURI, M.S. &. AZMI, M.I. (1978). Some studies on the predatory behaviour of Mjionchulus deniatus.Nemaiol. Medit. 6, 205-212.
LL-TORD, M.B. & OLIVIERA, J.M. (1937). The feeding of hoUow spear nematodes. Science 85,295-297.
NELMES, AJ. (1974). Evaluation of the feeding behaviour of Prionchulus punctatus (Cobb), A nematode predator, J.Anim.Ecol. 43, 553-565.
SHAF3AT. S^ BILGRAMI, A.L. & JAIRAJPURL M.S. (1987). Evaluation of the predatory behaviour of Dorylaimusstagnaiis Dujardin, 1845. (Nematoda: Dcrylaimida). Revue Nerrialol. 10, A55^\.
SMALL, R.W. (1979). The effects of predatory nematodes on populations of plant parasitic nematodes in pots. Nematologica IS, 94-103.
SMALL. R.W. & GROOTAERT, P. (1983). Observations on the predator ' abilities of some soil dwelling predator)' nematodes. Nematologica 29, 109-118.
STEINER,G. (1924). On some plant parasitic nemas and related forms. /. agric.Res. 28,1062-1064
THORNE. G. (1930). Predacious nemas of the genus Nygolaimus and a new genus Sectonema.J.AgricRes. 41, 445-446.
THORNE. G. (1939). A monograph of the nematodes of the Saperfamily Dorylaimoideo. Cjpi:a ZocL 8.1-261.
rriORNE. G. & SVr'.- vNGER, H.H. (1936). A monograph of the ncmaiode genera Dorylaimus. Dujardm, Aporauimus n.gen., Dorylaimoidesvi. gen. and Pun^nius n. gen. Capita Zed. 6 , 1-223.
WYSS, U. Sc GRCOTAERT, P. (1977). Feeding mechanisms of Lji?ro/ic'~.J sp. Med. Fac. Lindlwuww. Ri/Ksumv. Gcr.t. 42, 15:i-!527.

178 Nematologica 35 (1989): 475-488, E. J . Brill, Leiden
PREDATORY ABILITIES OF MONONCHOIDES LOA'GICAUDATUS AND M FORTIDENS {NFMAT01X\: DIPLOGASTERIDA)
AND FACTORS INFLUENCING PREDATION*
BY
.'\. I.. B I L C ; R A M I and M. S JAIRAjPL'RI
SeiMin ol'Xemdtolof^v, Department of Zoology. Aligarh Mu.slirii rnixersity, .Migarh-2()2(K)2, India
The predatory abilities of all stages of Mononchoides lon^icaudatus and M. for-tiderii were studied in vitro ineluding prey catching and feeding mechanisms, rate oi predation by adult and juvenile predators, prey preference, and effect of prey numbers, starvation of predators and temperature on predation. Prey catching and feeding mechanisms were divided into five phases, i.e., encounter with prey, attack response, attack, salivation!extra-corporeal digestion and ingestion! feedini^. Most phases were comparable in both predators. M. fortidens preyed more frequently and took less time to consume its prev than AI. longicaudatus. The latter species probed its prey gently but for longer, while probing by the former was more vigorous but for a shorter time. Lhe rate of predation by male and female predators remained unchanged over twelve days and prey numbers, starvation of predators and temperatures governed predation. Maximum predation took place in a population of 200 prey indixiduals and when predators starved for tweh'e days were used at 25-30°C. Acrobeloides sp. Cephalobus sp., Panagrellui redivivm and second stage juveniles ni' Me/oidogjme incognita and Anguina tritici were most preferred by both predators. No predation on Hoptolaimus indicus or Heniicriconemoides rnangiferae occurred.
Ever since Cobb (1917) first speculated on the possible economic importance of mononchs in agriculture, studies have been made of the predatory abilities of mtmonchs (Nelmes, 1974; Cohn & Mordechai, 1974; Small, 1979; Bilgrami et al., 1983, 1984; Small, 1987); dorylaims (Wyss & Grootaert, 1977; Shafqat el al., 1987) and nygolaims (Bilgrami et al., 1985). However, few attempts have been made to assess the role of diplogasterid predators in the biological control of plant parasitic nematodes. Yeates (1969) first studied in vitro lhe predatory beha\-iour of Diplenteron colobocercus (Andrassy). All stages of diplogasterid predators may not be obligate predators as they also feed upon bacteria in the
* Pre.scrited at the XIX International Xematolog) S\inpo>>iuni ot the Kiiropean Society ol XeinaioloL;i->th held at Uppsala, .Sweden, .August 7 13. 1988.

I 7 9
4 7 b Bll C RAMI \ )AIR\)Pl Rl
dbsenct oi pre) nemattjcles (Yeates, 1969) Grootaert et al (1977) elucidated the feeding habits of Butlenus sp , and assessed its possible use in the biological contiol of nematodes Later, Small & Grootaert (1983) evaluated the predation abilities ol Butlenus degnssei (Grootaert & Small) Recently, obstrvations ha\e been made of the attraction of Mononchoidei lon^icaudatus (Khera) and M for tiden\ (Sihuurmans Stekhoven) towards different prey nematodes (Bilgiami & Jaiiaipuii 1988) and pre- and post-fetding aggregation of these predators around a previously injured prty at feeding sites (Bilgrami & Jairajpuri, 198915) 1 he strikt rate of W longicaudatui and M fortidens and resistance and susceptibility of different prey nematodes to these predators was also dtter-mined (Bilgrami & Jaira|puri, 1989a)
III th( present woik Mononthoidei lon^icaudatus and if joHidens ha\c been studied in iitro to evaluate their predatory abilities and to determine the effect of bK)tic and abiotic factors on predation
XUlfRI^LS WD \U 1 HODS
Monoruhoides longicaudatus and \d fortidens were cultured by using the tech niques of Bilgrami & Jairajpun (1988) All experiments were made in "i cm diametei Petn-dishes containing a 5 mm thick layer of 1 % water-agar at 2 8 + 1 ' G The predatoi species were tested separately Twenty-fi\e prey nematodes were subjected tei predation by five female predators The prey and predators were inoculated simultaneously Observations on the number of pie) killed or ingested were made alter 24 h pAcry experiment was replicated five times The conditions in all experiments lemaincd the same unless stated otherwise lo maintain eonsistenc), the same speeies of prey were used for all the experiments with the two predators, i e , attraetion (Bilgrami & Jairajpun, 1988), resistance and suseeptibilit) oi prey (Bilgrami & Jairajpun, 1989a) and predation (present observations)
t o r eompanson Iree-living eeto- and endo-parasitic nematodes were used as [)rev Seeond stage |uveniles ol Meloidogyni incognita and An^uina tntici were eolleetcd fiom cultures maintained on tomato and wheat respectively, while iree-living nematodes (viz , Acrobeloides sp , Cephalobus sp , Rhabditis sp , Panas^rellus rediinus) were isolated irom tulture dishes maintained in the laboratorv The lest of the speties of plant-parasitic nematodes (viz , lykn-chorhynchus rnashhoodi, Xiphinema amencanum, Hehcotylenchus indicui, Hoplolaimus induus, Hir;(hmanmeUa oryzae, Hemicnconemoidei mam^firae anei 1 on^idorus sp ) were obtained fresh irom soil
Prey catthins^ and ffdini> meihanisms Observations on pre\ catthing were made bv stereoscopic mieroscope at 60 x magnification on inverted culture dishes eontaining M longicaudatus and M fortidens To study the feeding mechanism in detail, observation chambeis were made of a glass slide with a 2 x 1 5 X 0 5 em block of 1% water-agar (W)ss & Grootaert, 1977, Bilgrami et

180 \i()\()\( !i(i/ii/ \ i'Rin\ii()\ 477
al , 1985) and studied at more than 100 x mat^mification as required Ten specimens of M fortidem and \1 lom^icaudatui, (male and female) wcie placed m blocks of separate obsei\ation chambeis togethei with 50-75 / / oryzm The agar blocks contaming piedaiois and pre\ wcit genth co\ered with a cc5\er-slip and observations were made alter 15 min
Prey preferena To determme pieferente by W foHidms cind M loni^icaudatui tv\() sets of experiments were performed In the first, all the abo\e mentioned pu") nematodes were subjected to predation singK In the second set of experiments, all paired comf)inations of plant-paiasitic nematodes weie made using twent\-ri\e specimens of t a th pie\ species
Rate of predation b} ]uitniU \taiifs The rate of predation b\ diffeient juxemle stages of \i lom^uaudatus and 1/ fortidtm on H oryzae oi T/ uuos^nita was determined
Rate of predation bj adults To observe the late of predation bv adult predatois over a penod of twelve davs 1/ forlidens and M Itmi^uaudatm weie plated with H oryzae ox \i incognita \\x\(\\\\ii% The male and female piedators weie tested separatelx and each dav the piedatois weic tiansfcircd to fresh media contain mg pie\ of the same tvpe and original number
Effect oj tfmperature To find out whether temperatuie affects piedation, both predatois were tested w i t h / / oryzae dnd M imugmta ]u\cn\]cs dl 5, 10, 15, 2.0, 25, «), '35 and 40°C
Effct oJ prey numbers Cavit\ blocks containing 25. 50, 75, 100, 125 150, 175, or 200 / / oiyzat or \l incognita ]u\vm\cs were subjected to piedation bv \1 fortuhm or "V/ longuaudatus
Lffid of starvation of prtdators Al fortidem and Al longicaudatus weic staivcd bv keeping them m water without prev lor 0, 2, 4, b, 8, 10 oi 12 davs Predators were changed each dav to fiesh water to avoid bacterial contamination f.ach starved group of predators was tested separatelv against / / oryzai and the second stage juveniles ol \l incognita
Kl SL1 IS
Prey catching and fcfding muhani^ms [Iur \) The prev catching and feeding mechanisms of both predators could be divided into five moie or less distinct phases, i c , encouritir with prey (lip contact of predatois with pic) ), attack rapon'c (^probing and oesophageal suction), attack (attempts to puncture the cuticle ol piev), salivation'extra corporeal digestion (release of oesophageal secietions) and ingatwn/Jeedtng (intake of prev contents)
Piedatorprev contact was establisht^d bv tandom movement or bv attraction ol predators towaids prev (Fig 1, \ ) Pull lip contact of piedatoi with the ()iev was netessaiv to establish a contact between them {encounter uith puy) (Pig 1,B) Non-ldbial contacts failed to elicit responses from the predatois Aftei establishing contact, predators probed the prev's bodv with then lips and

IB^ 478 l!l ( R Wll I \1K \ I U<l
Probing of Prey (attack response)
Acceptable contourand texture of prey cuticle
Encounter and lip contact with prey
(B)
(0
:D)
(E)
Rand of
onn nnovennent predators
(A)
Attack by predators
(attack)
T" \
5 \ L)
1\
Post feeding attract on and aggregation of predators
m \ Feeding
completed
I ' ^ i
%\
\\
(J) Aggregation of other predators and feeding
^^.
(H)
o
Feeding/ ingestion by sucking
Escape of prey (G)
Salivary/ oesophageal secretions (extra-corporeal digestion)
\
Puncturinc slitting pr? cuticle
I , u j)u M 1 It III! I I In p n s I lu l i in i , iiul l v i u u \ ^ i n n h iiiiMiis ol \l m ii hi i h
ii^uaiidalu iiul \l 'i i' liii
tiisuicd a lull hp contact with the help ot oesophageal suction {attack teiponse ^l^ 1 ( I 1 In duiation ot probmt; dillercd m both predators il lon^uaudatui [)iobcd si uhtu lont(ci but less vit^oiously than \l foitidim It the contoui and tcvtuic ot du p u \ cuticle suited the j)udatois the pre\ was attacked (Fii^ 1, 1)) otliciwisi pudatois piobid sonic othci ict^ion in cjuick succession (iinnicdia'cK altti findini^ the tiist contact unsuitable) Pre) was ininiobihsed and attached onh when the predator attained an attacking posture (i t , hp contact \\ 'h the pie\ <it ^){)°) and cicated hit^h oe sophai^eal suction [at'ack F ^ 1 F) 1 he (Utulc ot picv nematodes was punctured b\ the combined action ol the dorsal 'ooth and oesophat^eal suction (I itj; 1, F) \ltei the cuticle was puiKiuieei oe soi)haneal secretions were obseived passing; into the buieal capsule ol tlu [)teclaiois thiouL;li a poic at the IMSC ol the doisal tooth [saliia

182 \l(l\(>\( IIOIDI \ P R I 1 ) \ 1 I ( ) \ IT-)
tion/extra corporeal digestion) (Fie; 1, Ci) feeding/iniieiiwn (Fie, 1, Pi) starttd after the prey cuticle was punctured by a combination oi the hit^h suction of the oesophagus and the movable dorsal tooth These predators feed by ingesting the body contents of their prey M fortidens also engulfed occasionally the entire fiist stage juveniles oiAcrobeloides sp , and Cephalobus sp (if the pre y and predator were end to end) During ingestion the metacorporeal pump pulsates at legular intervals usulting m intermittent flow of picv body contents mlo the predators About 20-25 feeding bouts/min (bout = single pulsation) were recorded in \1 fortidens and 15-20 in M longicaudatus During feeding, both predators exhibited ' shunting behaviour" m which \4 jortidem and M longicaudatus teleased their pre}, moved forward/backwaid up to a distance equal to their body lengths before stopping and reverting back to reach the prev to feed again Predatory activity (shunting bcha\ loui) was punctuated by feeding bouts copulation and egg laving Some times prev escaped fiom a predator's grip bv evasive actions taken by the prev or because of inability of the predator to wound or injure the prey (Fig 1,1) I his led predators to move again in search of prev Howeyer, the escape of an injured or wounded pre) attracted other piedators (Fig 1, J ) Generally 6-8 predators, sometimes more than ten, fed together on a single prev Although predators fed together, they stiuggled among 'hemselves to be first to feed on the prev feeding finished soon after the pre> was completely eomsumed (fig 1, K) bul preditors remained aggregated at feeding sites for some time (post-feeding aggregation. Fig 1, F)
Pre) prejereme ( I able s i & If) When prey nematodes were subjected to predation singlv , V/ fortidens and Vf /on icawc/a wf showed maximum pre dation on Acrobeloide-i sp . Cephalobus sp , P redwivus and the second stage juyeniL'S of M imoi^mta and A tritici ( p < 0 05, Table 1) f he number of different pre) killed by \4 fortidens and M longicaudatus was positively correlated (r = 0 97) M incognita and A tritici were most preferred by both predators in all the combinations in which they were tested ( p < 0 05, Table II) When tested together in one combination M incognita and A tritici juyeniles were killed in equal numbers by both predators ( p > 0 05) Helicotylenchus indicus were killed when tested alone (Table I) or in combinations (1 able II) but were only partially consumed by the predators on many occasions fewest Longidorus sp , and X amencanum were killed m all combinations Predators, however, prefeired X americanum when tested with Longidorus sp , in one combination ( p < 0 05. Table II) No specimen of Hoplolaimus indicus or H rnagniferae was killed or injured by the predators under an) conditions
Rate ojpredation by juvenile stages (fig 2) All stages of both predators k lied both types of prev 1 he fourth stage of each predator killed most of each prey and the first stage, least ( p < 0 05) There was no dilference m the number of prey killed bv the same stage of each predator ( p > 0 05)
Rate of predation by adults (Fig 3) The number of prey killed bv males and

a3 w o lill ( R Wll I \Ik \ | l ' l RI
1 ABI E I
Rate nf predation by Mononchoides longicdudatus and M iortidens in a single prey population
l ' ie\ siHcies Mean + Standaid dtvnt ioi i p \1 furtidetn \1 Ionian i udatw,
Pariiu^uhu lediinus
Ainilxu idt\ sp
( (/ Hall I u ^p
Rhuhdil •,]!
Mill ld(ijrn iruon^mta ]\
hnutnii I ilui
Hii ilnnutinulla ory^at
I)lituh(>h\7uhH\ maihhiiidi
\iph nnnii amtruanum
HI In I lni(hui induus
I (iriiiidoru sp
Hupli laim I induu\ Him riiuturni tdis numi^ifiiai
C ( i t t luunt ol coiichtion r - 0 " l? ( p < 0 ()))
2i + 1 01 [U 25)
22 f 1 41 (20 24)
2 2 - 1 41 (20 24)
12-r2 28 (10 If))
22+ 1 85 (\^ 24)
12 1 1 60 (20 24)
14 t 1 r i (17 21)
10 + 2 78 (14 22)
If)+ 2 b5 (14 19)
lb r 2 Oi (12 19) 0 ^ 2 0^
00 00
22 + 2 OJ (20 2.)
2) + lbl (20 24)
18+1 n (lb 20)
10 + 2 )7 (8 14)
2 0 + 1 41 (18 22l
21 + 1 0 i (19 24)
17 M 2) (1) 18)
1b + 1 b> (14 18)
n + 1 49 (12 18)
15 + 2 r (12 18) 4+ 1 01
00 00
Iis^uits IP paunthLses induatt rant^t oi p i t \ killed b\ piedators p \alu s a u cakulatcd b t l w i m du numbtr oi i puv killtd b\ \4 fortidin and \i lona^uaudatus + \ ( ) i Mt^nilicant + + Sit,nih( ml It 0 05
ftindles of M Jortidens (Fig ^, A & C) and \I loni>icaudatus ( t ig ^, B & D) prey pel ddv tor 12 days did not differ ( p > 0 OT) More individuals of A/ incognita juveniles than H oryzae were killed by males and females of both predators ( p < 0 OT)
tfffct of temperature (Fig 4, A & B) There was a significant increase in numbers of both species of prey killed by M fortidens (Fig 4, A) and M loni^icaudatus (Pig 4,B) i the temperature rose to 30° Maximum predation occurled between 25 and 30° and declined sigmf'icandy ( p < 0 05) at higher temperatures
£ffeit of prey numbers (Fig 4, C & D) There was a positive correlation between the number of prey killed by M fortidem (Fig 4, C) and \4 hn^icaudaiu^ (Fig 4, D) and numbers of prey increased ( p < 0 05) Both predators showed maximum predation when 200 prey individuals of// oryzae and Ivl inLos,nita ]u\cn\\e?, were present Minimum predation was recorded in

\IO\0\l HDini s I'KI l ) \ l l ( ) \
i
K
__
i ^ .
I I
H o r y Z Q e
V n e o g n i '
M
I I I F^
Juveni e stoges Juvenile stages
ti^ 1 Rat<. of |)ie(icili(»ri b\ |ine)iilc statjcs. A, Mononchoidf. jorlidin\, B, 1/ lonnuauilatu\ / / /•"/M/ slaiie jmnnlis }> Simnd tluni iuii>iili\ ] ^ I hud vlii!>i jiiiniiln / / I'oiiilh \liii;i imniilit
22
2a -oi7.p>o.ns
xy---003x*20.67
'^-G.2L. P>Om
- I 1 1 1 1 1 ' 1 1 '
1 2 3 4 5 6 7 8 9 10
No. of days
22
20.
18
l i
U
X
5 "
X
""•~
X
X
r = -O.Oq.P>0OR X X
X
•
" > ' - o o 5 7 ^ i : i 2 ^ •
B
X
X
X
•
• •
No. of days
22
2(1
-E=0.i9.p>o.ns. ^ = -004x«19.25
- | 1 1 1 1 1 1 1 1 1 1 2 3 i 5 6 7 8 9 10
No of days
2) '
19.
17
1b_
13
x
•
X X
X
r=-007, P>005 y = -0.0lx-15 49
•
—1 r- - I - r-
X
•
— 1 —
X
• l
•
—I 1
D
X
•
•
1 1 1 2 3 i; 5 6 7 8 9 10
No. of days
Fit? 3 Rale .of prcdation b\ adult predators A, Malt- Moriuiiihoidt^ fortidin\, B, .Male .\t loni^icaudatui C, hernalc M fortidins. 1), Female W lonpcaudatus • = Huschmanrurlla iiryzae,
X = Meluidneyne inioi^mla |u\eniles

IB':)
1-82 Bll I l< W l l ^ I \ll< \ | l ' l Kl
1 \BI V II
Rfilf of pr datii n b\ Monoiu hoidc s tor t id ins and \\ l()n^K ducldtus in diffinnt
comhi>iation\ of pri\
( m i l '
111 | i i (
H O
I K )
H O
H O
H O
H O
H O
H O
1 O
1 O
1 O
1 O
1 O
1 O
1 O
HI
111
HI
HI
HI
) i i i
\
f
»
-
'
4
4
•
-
*
-f
1 i t i i t n s
1'
1 O
HI
M l
\ l
1 \ !
W
H O ! '
I I \ I
H I
\ l l
\ l
1 \ I
W
H O I '
l l \ l
M l
\ l
1 M
\ \
H O I '
i'l
1 _' i )
IH) 1 1 "H
1 > Ml|
11 1<)) 1 ' III!
1 ' - ! i m
" 1 8't
, 181
1 IH
" , 1 (M
'(I
' (I'l
1 ) I'M
' 1 41 ) 'M
1 * 1 ()H
1 ()1
1 -!()
1 1 ~
1) i 1
. r 1 " t
1 M t . ' ( I < l
1 1 > 1 ) 1
IJ » (1 7-I-
1 1 1 )
1 ' t 1 7J
'• 1 1-
1 8 )
8 1 ii
t ~ 1 m
l l l l 1 ( > )
') vl '11
{\1 I'M
'(I ^ 1 t l
18 l.l\
M ( in si i n d
1'
K I T 1 H
('1 12,
\1 r I (.1
(8 KM
J l T J (1 1
(18 i l l
11 4 1 78
( i l l 21-1 .'II 1 ()2
(17 11\
n . 1 «'i
(11 11.)
1 8 + 2 K)
(11 2(1)
2 . 1 1)7
tli> 2 1
2 . - 1 ,i .
( I ' l 211
1! 1 >1
(1 • 21 )
1 ) t 1 I'l
(11 18)
2 . t 1 71
[211 2 1i
2 ' * 1 <)"
20 21
18 t 2 )2
(1 > 21) 1 2 - 1 H
(HI 1 t |
i i d (1( \ 1 i l i o n
I'l
2(1 T 1 72
(18 2 ! )
18 r 2 DO
(1 • 21 1
11 2 11
(11 18) 1 i f 0 8')
(12 11)
1 ) t 1 i l
(11 un 11) • 2 OJ
(12 I'l)
1 1 f 1 72
(1 > 18) 17 t 1 41
(1 ) I 8 | ) t 1 'M
I l 1 1 )
2 4 1 11
(0 H i 1 ()7
lO ))
1 > 1 11
(2 0) 2 - 0 18
U' i) (-) t 1 07
( I ' l )
. 1 41
( H i )
10 4 1 8 )
(8 1 i)
8 4-2 40
(0 10)
1 2 + 1 f)7
(10 14)
14 I 2 00
(10 16)
1 8 - 1 72
( I ' l 20)
\>
(. - 1 72
( M l )
10 r 1 11
(•1 I2l 20 t 1 '1 .
(17 21)
21 t 1 ()7
(18 2 i ) 10 4 1 11
(11 18) 1 0 - 1 07
(8 12)
1- 4 2 28
( 11) 1()|
22 4 1 41
(20 21)
2 > 1 07
(20 2 )) ]' > 1 .2
(11 20) 1 .4 -2 ')()
(10 17 |
2 1 4 2 0(1
(211 21)
2 i ( 1 74
(20 21)
1 0 ^ 1 10
i l 4 18)
11 4 0 74
^1(1 12)
R c ^ K ssion
( ( X I I U R I U 1
P, F
(1 41
0 00
(1 81
(I 0 1
0 1()
0 <)1
(1 O'l
0 8 1
0 8 )
0 <)2
( 1 - )<)
0 '!_!
0 "(i
(1 "1
0 <)1
0 1>
0 1')
0 )•••.
0 7b
0 <)()
0 08
0 <)4
1) '14
0 '12
(1 '10
0 7-)
—
0 11
0 <)7
0 8
1 (10
0 ) i
—
—
0 07
0 80
(1 Oh
0 81
—
\ l l p i i \ I nib ilioiis ((iiiipiisid (iMitn | )u \ incln idudls (tv\( nt\ li\( uf ( a( h (\|)(.) wc rt siib|ct(ed Ui pi idi t idi . t)\ l \ ( Ml and Ml MpaiatfK Ris^ussion (o t t fuunl i was taUulatid b a w a n (ht luuiibti (il I p a r I iildi p u \ kill( d In Mh and MI p mdii att s Mi(nih( ami of i I i ^ U R s i n p a i i n llH M s iiulii iK I mm i) l | ) i ( \ Lilkd l)\ picditoi * Sm;nilK inl al 0 0 ) — \(i t Ms^iiilnant
Kl \ \'I tu^( [)ii\ P . s((oiid pi( \ 1)1 (onibmatuin MP Mojioruhaidis jvitidi i\ Ml Morion iliiiilc liDi^ii iurlali,\ HO fhrulmunoiiiltu on ai l O 11 nt^idoru^ sp li\ - Hilh(i!\tenihu\ uiditus MI Mihndi ^\ri miiiinila \u\ \ 1 - Arii^uina tntlii ]u\ I \ 1 I dtnchor/nnihu\ tnashliuodt W-\ifiliiinvia an run nil HOP Hi pliilaimu\ indiiii^ \\\{ ~ Himiiruunniiouhs maniiifirai

1 8 6
M()\(>\( H()II>1:\ I'RI'DAIION 4 8 3
TABLE; II (continued)
Rate of predation bj Mononchoides fortidens and M. longicaudatus in different combinations of prey
( ; ( i m binaticms ol prcv
I'l
HI
MI
Mi
MI
MI
MI
\r AT
A T
A'l
I'M
TM
T M
XA
XA
H O P
!'_,
+ H M
+ A T
+ TM
4 XA
^ H O P
+ HM
4 I M
+ XA
* H O P
r H M
^ XA
* H O P
4- HM
4 H O P
4 HM
-, H M
i'l
18 t 2.3'2 ():)-2!i
20 T 1. H) (lH-22)
(20-21) 2(! 1 1 07 (;r-2o
122-2,-!) 2:; *• ;.o9 ,22-21)
24 Wi 74 l2:i 27)
2 i t 1 07 1 21-2:1)
2'i : r ; . n ,22-27))
11 ± 1 00 ,19-2'))
18 4 1 41 (10-20)
18 J 1.72 (17-20)
17:, 1.20 (1(1-19)
19 , 1.78 (11.-20)
i;i , 2.31 (10-15)
Mean + star
P ;
C C
22+ 1.72 (20-2,5)
14 ± 1 54 ( I : M 7 )
9 i 1.41
(8-11) 0 .-
r> •-__ ^
l b ± \M (14-18)
10±0.74 (9-11)
' ,-
, . • , ,
9 ± 2 00 (0-12)
idard deviation
1^
17 4-2.09 (15-20)
22+1 .16 (21-24)
21 I 2.00 (20-24)
22 ±2.41 (19-25)
2:1 t 1.49 (20-24)
21 3:2.19 (18-24)
2:!-r 1.09 (21-24)
20 ± 2 00 (17-2;5)
21 4 2.41 (18-24)
22 + 2,00 (19-25)
14 + 1.41 (12-16)
12 t 1 78 (10-14)
14± 1.41 (12-16)
14+ 1.1b ( 1 ; M 6 )
10 +2.00 (10-10)
^1
c c
20 4 1.41 (18-22)
10* 1.41 (8-12) 6 4 1.78 (4-8)
-
C
124 1 41 (10-U) 64 1 01 (5-81
^
0+ 1.41 (^8 )
--
Regi 'fssitin tocflicient ' - '
f'l
0.60
0.91
0 80
0.70
0.84
0 68
0 70
0.7;;
0.87
1.00
1 00
0.78
0.84
0.66
0.80
—
P_.
- •
0 98
0.68
0.95
—
—
0.94
0.70
—
—
0.99
—
—
—
—
--
'P '
1^ Pi
— —
* *
*
— —
.\11 prey combma'iuns. tciinprised ol'fift)- pir \ indixiduals (twcntv live til caoh type) were siibjcited tu predation by li\e MF and ML separateh' Regression eoel'lieient ' r ' was laieulated Ix'tween the number ol a particular prey killed b\ MF and ML 'p indicates significance o l ' r " . Figures in paren-(hr,scs indivutt ;.jngc lA prey "k'lWvd hv prcddtor. * = .SjijniiJc am at 0.05; - - ^ .Xot sigiiijiiant.
Kev: Pi - Fiisl pte\'; I\i -second pie\ of combination; \\V = M(,?i(inJi(ndi-\ fcilidrriw ML= \fori(in-ikoide\ 'iniiiuaudntu\. \{0-'Hirsclimannnilu oryzae: \X) ~ Ijini'idnrw. sp.. HI ^ Heluoliliruliu^ niduuy. \U ^ Mf'laidagyrir tm ot^tiila j u \ . , AT - An^uina Irttki j u \ . , '\'\l = I'yli'ruiiurhvnihui mmhhondi; X.-^ ^ Xipliinema ameruanhm, HO[-'= Hoplolaunu\ induu\; hWl =- Hemunconniwides mane^iferar

484
22
2(51
o m y /
BUC RWl l & J \ 1 R \ I K Rl
A
• — • H oryzoe
X—X Nl incognito juv
1 1 1 ! 1 1 \ 1 5 10 15 20 25 30 35 W
Temperature Cc)
15H
• — • \\_ oryzoe
X'—X M incognito juv
T 1 [ 1 1 1 1 1 5 10 15 20 25 30 35 40
Temperature Cc)
T 1 1 r . . 25 50 75 100 125 150 175 'CO
No. of prey
—1 1 1 1 1 1 1 1 25 50 75 100 125 150 175 200
No Of prey
' I
z 4.
r 0% P«0^ y 0 33x*1972
r 7 " j 90P t005
1 ' I I I 1 I 0 2 t 6 8 10 12
Period of starvation (days) Period of starvotion(days)
I-ii 4 I a a o i i mflueniin^ prcddtion bT, Mononchoidn Jortidim and M hm^icahdatm \ tffi tt of umpt-idturL on prLdatioii b\ W fortidem B Hftct of t t inperatuit on picdation b \ M lim^uauauius ( Efitn of pit v numbers on prtddtion bv M fortidern D t fftcl of pic\ numbers on puddtKin b\ \1 lon^uaudahn \ Effttt of stdrvation of prtdators on pifddtion 1)\ \1 fortidtns F tttect uf starvation on pudat ion bv W ton^uaudatui • - m d i c a t t s tht attual number ot Hmchmmniiella ory^ae killed x - mdicdtts the actual number of Mdoidogyne ivcosnita ]u\cnilcs
killed b\ tht pitddtors

I B S
\U)\()\( HOIDi \ TRIDMION 48^^
a population ol 25 prey individuals belonging to both types ot prev ( p < 0 05) EJJect of itariatiun oj predators (Fig 4, PL & F) The starvation of .V/ Jortidens
and M lunqicaudalui influenced their predation More H oryzae and M incognita juveniles were killed by M Jortidens (P'lg 4, FL) and M lone^icaudatus (Fig 4, F) starved for 12 days than by freshly isolated predators ( p < 0 05) The rate ol piedatK)n b\ all starving groups was higher on M mcoi^nita |uveniles than H oryzae
DISC LSSlOX
The great similarity of piey catching and feeding mechanisms and ot the influence ot various factors on predation bv M fortidens and M lon^icaudatus indicates that V)oth species oi predators are very smiilar m then behaviour and lequirements Thus, the predator prey relationship measuied tor cine species ot predator ( W fortidens) strongly supports the observations made with the other predator [M toni^icaudaiui)
Predation bv Pnonchulus punctatus, Monorichus aquatKus. Mylonchulas dentatus and Aquatides thornei follows chance encounters with the prev (Nelmes, 1974, (irootaeit & Matrtens, 197b, Jairajpuri & Azmi, 1978, Bilgranu et a< , 1985) However lecent studies on the attraction of M Jortidens and \1 lonoicaudatus towards ditterent prey (Bilgrami & Jairajpuri, 1988) and their aggregation at teedmg sues around a previously injured prey (Bilgrami & Jairajpuri, 1989b) suggest chemoieception in these predators to establish contact with prev as well as chance contacts which occur during random menement ot predators and pre) 1 he chcmosensory ability ot these predators should enhance predation on a particular type ot prey by increasing the likelihood of contacts between piedator and prey VVyss & Grootaert (1977), Bilgrami et al (1985) and Shatqat et al (1987) reported probing in Labronema vukapapillatum, A thornei and Dorylaimus stai^nalis respectively The present predators also probed their prey belore attacking possiblv to establish a suitable spot to puncture the cuticle ol prey
1 heoretic allv. more ot the most attractive piey should be killed by the predators However, Bilgrami & Jairajpuri (1988) showed that H oryzae, I mashhoodi and Hoplolaimus indicus attracted more \1 lonoicaudatus and Al Jortidens than other species, but the former two pre) species were killed only m moderate numbers and no individual of Hoplolaimus indicus or H mangijerae was either killed or injured bv the two diplogasterid predators By contrast, M imos^nita and A tritici |uvemles weie preyed upon more by M lons^icaudatus and \1 fortidens but attracted only moderate numbers ot these predators (Bilgrami & Jairajpuri, 1988) Difterences in experimental conditions (i e , confinement ot prey at one place during attraction experiments (Bilgrami & Jairajpuri, 1988) and allowing prey to move treely in the agar during the present s'udy) mav have led to these differences Predation is a physical phenomenon requir-

;33 4 H b liUi.KAMl .. J MRAJl'lKI
iri^ essentially, lif) contact of predatoi with prey and depends upon the activity, escape response, texture and thickness of the cuticle and undulation movements of prev. CMiemoreception is not influenced by the same physical factors bu! depends rather on the compositicm, concentration, equality and quantity of prev secretions/metabolites, as well as the rate of dispersal of such secretions in the surrounding medium (Bilgrami & Jairajpuri, 1988). It seems that prey preference in these predators does not depend solely on prey secretions but also on the factors mentioned above.
Siinura s{)p.. paralyse their prev bv injecting paralysing agents (Linford & ()li\iera, 1937; Hechler. 1963). The nygolaitn and dorylaim predators immobilize their prey bv disorganizing the internal organs with their mural tootlr'odontostyle (Bilgrami et al.. 1985; Shafqat et al., 1987; VVyss & CJrootaert, 1977). M. furtidens and M. lungicaudatin seemed to hold their prey by suction from the oesophagus. I'he release of oesophageal secretions into the buccal capsule of Al. fortidens and M. loni^icaudatus through a pore a' the base of the ciorsal tooth after the prev cuticle was punctured suggests the occurrence ol e.xtra-corporeal digestion, a pfienomenon that occurs in plant-parasitic nematodes (Wyss, 1971). (jrootaert el al. (1977) also ()bser\-ed oesophageal secretions released into the buccal capsule of Butlerius sp. The shunting beha\ iour of .\/. furtidem and M. lon((icaudatu.s\ a phenomenon which commonlv occurs m free-li\ing nematodes (C'roll, 1974), seems independent of prey densitv (.Xelmes, 1974) but dependeci on the number of predators feeding together upoti a prey at a time. The greater the number of predators the more evideni was their shimtiiig bcha\-iour. (Cannibalism which occurs under nalurai conditions (Bilgrami et al., 1986), has also been observed in vitro (Xelmes. 1974; Azmi & Jairajpuri, 1979; Bilgrami & Jairajpuri, 198,5). During the present observations A/, loni^icaudatu.s exhibited a stronger tendency to cannibalism in the al)sence ol prey as compared to M. fortidens under similar conditions. This disparitv may be due to the bacteriophagous nature of .1/. fortidens whicli fei'ds on bacteria in the absence of prey nematodes, hwX M. lons^icaudatus did not do well on bacteria alone.
Neinatodes may resist predation by physical, chemical and behavioural characteristics (Ksser, 1963, 1987; Ksser & .Sobers. 1964; Small & Grootaert, 1983; Bilgrami & Jairajpuri, 1989a). Lack of predation oi Hoplolaimus indicus and // . manf^iferae might have been due to their thick cuticles and annulations respecti\-ely, pro\-iding resistance against predation. M. dentatu.i and D. stai^naln also lalled to kill or injure Hoplolaimu.'i indicus (Jairajpuri & .'\zmi, 1978; Shafqai et ai. 1987). Ksser (1963) suggested chemic:al resistance in Helicotylen-chus spp. Bilgrami & Jairajpuri (1989a) also suggested the possibility of unlavouralile secrcti(^)ns in Heltcotylenchus indicus. X. amencanum and Longidurus sp. Tins could be a reason lor less predation of these species.
The rate of predation by M. longicaudatus and M. fortidens increased when the numljcr of |)rey increased as it did with Diplenteron colobocercus and A. thornei

• 1 9 t J
uu\(n<//(!////• \ I'K] })\iH>\ 4 8 7
(\'eates, 1969; Bilgraiiii i-t al., 1985) but for .W. aquaticus remained unaltered in different ntuabers of prey (Bilgrarni et al., 1984). Shafqat et al. (1987), however, found an optimum ratio between predators and prey numbers for maximum predation. The increase in the rate of predation by the two diplogasterid predators during the present study was prt)bably due to improved predator prey eneounters. Starvation of .W. longicaudatui and M. Jortidens also influenced their rate oi predation with predation increasing with increase in the period of starvation. The starvation of predators. M. aquaticus and I), stagnalis, did not alter the rate of predation (Bilgrami et al., 1984; Shafqat et al., 1987), while (jnl) short term deprivation of food enhanced predation by P. punctatus (Nclnies, 1974). However, D. putuhikus and M. dentatm killed more prey when starved for a longer period (Yeates, 1969; Jairajpuri & Azmi, 1978). The difference in predation ')y M. fvrttdens and Af. longicaudatui at different temperatures may be attributed to the change in activity of both predators and prey as a result of changed temperature (Bilgrami et al., 1983).
ZL SAMMKNF.ASSSUNC;
[)i( iauheri\fhen hahiiilittleii 'on \l(inon( hoult-s lont^Kduddtus «?irf Mononchoutc.s Ibrtidcns l.\cnia-loda. Oijjjdtjd.s fndd) und Faktuini, dif da> Beulemachi'ri bieinjluuen
In IT, idiu Studien wuHlen die idubeusclien Fahigkciten aller Lntwii klungbstddit-n von Monnn-ihoide. tani^icaudaiut und Ai ihiiidms unlcrsucht, finschliffiiich Beutcfang, Mcchanibnius der Ndhrjngsaufiiahmt*, Lnitarig des Beuteniachens Lei Aduhcn und Juvenilrn, Beutcbt-vorzusrung bijvvic der Eintlufi der AiuAil Beutetiere, des Hungers der Rauber und der 1 emperaiur auf das Beuteuiachen. Die MethaniMuen deb Beutetan^s und der \ahrunt ;sduinahme werden in fiinf Fhascn euigeteilt; Begcgnui."; mil der Beute. Angrift'sreaktion. Angiitt. Speithelabgabe/exlra-korporale W r d a u u n g und .Wihrungsaufnalirne.
Die nieisten Phasen Wditn Ijei beiden Rdubern \ergleiclibdr. M iuriidens ting haufigei und brauchte weniger Zeit zum V eizehr dei Beute dl.s I/, [ungiiaudalut Lelzterei priifte seine Beute sdntier iind tin Idngere Zeit. vviihrend die Priitung von \i. furtidem lieftiger und kiirzer v\ar. Die Rdit dcs Beuteiiiachens bli( b bei niannlichen und vveiblithen Raubern iiber zwfilt l age hinweg glen hi. Sie vvuide dureh die .\nzahl tier \'oriiandenen Beutetiere. den Hungerzustand der Rauber and die reinperatui gcsteuert. Der hocliste Beutelang land in einer Population von '. 00 Reulet:tren iiei 25-!'iO''(. n^.t Raulx'rn statt. die 12 Page geiiungert hatten. Acrubetoide\ sp., CV-phaliibu^ sp , Panagrellu- redi.uu\ und die J2-Stadien von Meloidug)ne incuiJmta und Ani^uina tiitui wurden von den beidt n Raubern am siarksteii bevorzugl Hoplolaimus induu^ und Hemuncune-nuiiue\ man^iitiai vvuideii nu 'n getresseu.
RKPKREM.KS
.AzMi, M. 1. & jAiRAjin Ri, \ 1 S (19791. hftect ol meehaniLal stimulation and i rowding on .Myloiuhutus denlahii .\iiniit(iloi^ia meditrnanea 7. 2()!5-2()7.
Bin,K,\.vu, A. 1. . . - \ H \ I \ I ; 1 8i jMR.a,jPt'Ri, \ 1 . S. (1983). Some taitors iniluencing piedatioii bv Miinonclm.s aquntnw Reiae de Seriiatolugit 6, .•i2>.'i26.
BiK.R.A.vii, .\. I ,, .Aii\i.\i>. 1 & jviRvjPtRi. \ 1 . S. (1984). Observations on the piedatorv behaviour of Munuilia^ aquataia StynuUdoii^ia meditmavea 12, 41-4,T
Bn <;K.-V.\II, A L , AHVIVU, 1 & IVIRAJPCRI, M . S (1985). Piedatorv behav lour of .-l /uaZ/rfn thornn (Nygolaimina: Dorv .ainiida). Semaluioi^La 30 (1984). 457-462
Bii.GR.vMi, A. L., A H . I V D . 1, i )viR.3ijPURr M. S. (19861. A studv on the intestinal contents C'f some mononchs Rnut de Semawtogu 9, 191-194.

191 t 8 8 Bll ( RWIl ^ l^ilRAIPl Rl
KiMKWii \ I &J\iR\jpiRi \ I S ^198T) C annibaliMn in \4ononchus aqualuus Indian Journal „ \ema/ologi 14 (1484) 202 20^
BiKKWii \ L SijMR^jPi Ri \ I S (1488) ^itraition of \iononchoide\ lani^tiaudalus <md \I foi l'din\ (Nenidtoda Diplot^astcnda) towards j )u \ and tactois inlluenciiiiJ <ittraction Reiue de \i'malulume 11 I'J') 202
BiK.KWii \ L &J\iR\jpiRi M S (1980d) Resistanu of prev to pu ( ition and stiikc rale ol tht pitdatois Mimanthoidei luni;uaudaiu\ and V/ fortidtns ( \ ( .matoda J)iplot;asttiida) lieiue de \emaWloi;ie 12 45 49
Bn(.R\\u \ 1 SiJ\iR\jPiRi \ 1 S (1989b) Xa^nvt t^Miun of Manunclwidis lon^uaudatus dnd A-l rarlideni {\emdtodd Diploctastinda) at sitis \(m«('«/oi;ua 34 (1988) 119 121
( oBB \ \ (1917) 1 hi rnononchs (\4ononthus Bastian) a genus of Irte livnic; ncmatodis S,d Siiem, 3 4J1 48b
C nH\ 1 6. MoRDfCUxi \1 (1974) txp t i in t tn t s in suppressing cuius populations lj> use of mangold and a prtdatious ntmatodt \ematoloi^ia meditenanea 2 4i 53
C KOI 1 \ \ (1975) 1 otoinotorv beha\ lour energetics and dispersal in iitinatodts Advamis in Parasitoloiy 13 71 115
EsshR R P (196-i) Xeniatode intcidctions in plates of non sterile water aeai Proceeding'^ of the Soil and Crop Scienie Society of Florida 23 121 H8
1 ssi-R R P (1987) Biological eontiol ot nematodes I Dor\laims (Ntiiiatoda Dorvlaimma) \cmalolo^j Circular \° 144 pp 4
I ssi R R P & SoBfRs F R (1964) Natural enemies ol nematodes Procecdini^i of the Soil and Crop 'science Socut) of Honda 24 32b 355
(iRoe)! \ tRr P & M^hRihss 1) (1976) Cultivation and life e\cle (jl \lononctius aquatiem \ematolo!;ica 22 17 5181
(rROe)i \i-Kr P J \Qi Fs \ &S\ i \ ] i R W (1977) Pre) selt e tion in Bu^/c««\ sp (Rhabditida Oiplogastendae) Mededelin^en faculleit I andhounivetenuhappen Rr/ksunn eruleit (nnt 42/2 r n 9 1563
HicHLiR H C (1963) Description de\ tlopnie ntal biologv and feeding habits of Seinura
enuuaudala (de Man) } B Croodev 1960 (Nematoda Aphelcnchoidea) a nematode picdatoi Proceediniii of the Hilmintholo^ical Socict\ of U a^hington 30 182 19T
J\ iR\ |PiKi \1 S & Ai/Ml \ I I (1978) Some studie s on the predatorv behaviour ol \4]loncliulus dentatus \emataloi;ia mediterranea 6 205 212
1 iNhoRD \ I B SiOiiviER\ J \ 1 (1937) The feeding ot hollow spcai nematodes on other nematodes Science 85 295 297
XpLMhS \ J (1974) Evaluation of the feeding behaviour oi Pncmchulus piactalus (( obb) a nematode predator Journal oj Animal hcolog) 43 553 565
SHAFCJAI S BHCRAMI \ \ Ik JMRAJPIRI M S (1987) hvaluation of the pitdatory
behavioui of Dorylaimw, ^ta^nah\ Du|ardin 1845 (Nematoda Dorvlaimida) Reiue de \'-malcloiiu 10 4 T 5 461
SM^LI R V\ (1979) 1 he efltets of predatorv nematodes on population of plant paiasitic nematodes in pots \emaloloi;ica 25 94 103
S M \ I I R V\ (1987) A rev ltv^ of the pre) of pre datoiv soil nematodes Pedobiology 30 179 206 S M \ I I R VS & GROOI \S-RI P (1983) Observations on the predation abilities of some soil
dwelling predatorv ntniatodis \ematologtca 29 109 118 \ \ ' i s s L (1971) Drr Mechamsmus der \ahrungsaufnahme bei Inchodorus suniti'< \ematologua
17 508 5l8 W-iss L &(rRooia.fKi P (1977) P e c ding mec hanisni of / oAroKf mfl sp Mcdid'lingen I aculttit
I andhoua iietcnnhappen Riikwnutrutcil Cent 42 1321 1)27 \ f \ r E b Cr \\ (1969) Predation bv \4onoclwides polohikm ( \ ema toda Diplogasteridae) in
laboratoi\ cultures \ernalolo^ua 15 1 9 YEVTFS ( I V\ (1987) Sigmficanie of developmental stages in the CO existence 01 three species
of Mononchoidea ( \enia toda) in pasture soils Biolog) & Pertilit} of Soil 5 ^25 229
AcccfHed for publication \Iav 15 1989

102
Inll Ncmatol. Network Ncwsl. 7(4): 25. 1990
Planl-p.irasific Nematodes Associated with Paddy Crops in Utlar I'radcsli, India
Anwar L Bilgrami and Wasim Ahmad Lecturers, Section of Nematology
Department of Zoology Aligarh Muslim University
AJigarh-202002 ''iNDlA
A randvom s ar\'ey was conducted to identify and assess the frequency of occurrence of economically important plam-parasiiic ncm itodes of paddy {Or,za sataa L.) m the districts Deona, Basti, Gorakhpur and BareiUy of Uttar Pradesh, India A total of 36, 24, 38 and 42 samples were collected from different localities of the above districts, respectively Tnc soil samples were processed by Cobb's decantation and Baerrnann's funnel techniques. Tlie nematodes per 200 gm of soil from each sample were counted and identified.
Thence root ntmalode, Htrschmanniella oryzae (Van Breda da Haan) Luc& Goodeywas found to dominate all other parasitic species of nematodes associated with paddy in Deona and Basti districts, as also was reported by Ahmad et al (]9(SS) during their survey of paddy in Bihar State of India. Data on Gorakhpur and Bareilly districts indicated dominance of Tylenchorhynchus mashhoodi (Siddiqi & Basir, 1959). In the present study, H oryzae was found .n 80% (£204'200 gm), Hehcotylenchns dih}slera (Cobb) Shcr m 60% (j<84/200 gm); Hoplolaimus indicus Sher in SO't (^4I)/2(K) gm), J mashhoodi in 50% (j<32/200 gm) m&Xiphinema insigne Loss was present in 20% (;< 10/200 gm) ofsoil samples which were collected from Deona district. Resultsof the survey of paddy crops from Basti district indicate 87.5% occurrence of T. mashhoodi (j<252/200 gm); 86% of// oryzae (j<56/200 gm); 50%o of// dihystera (_<52/2(K) gm), 62 5% oiAphelenchus avenue Basiian (j<16/2(X) gm); and 37.5% of Basirui tumula (Colbran) Gcraert (j<24y2(K) gm) // oryzae was found to be the most dominant and prevalent pest of paddy in Gorakhpur district. / / oiyzae occurred m 67.6% of samples with a population of (_< 112/200 gm). Hehcotylenchns dihystera and T mashhoodi each occurred in 45% of samples (<7,9flQO gm and 60/200 gm, respectively). The population of / / indicus, Fiatylenclvis ihomei, A avenae, and X insigne were comparatively low (_<24/200 gm; 8/200 gm; 8*200 gm and 16,'2fX] gm, respectively). These nematodes occurred in 36%, 18%, 36% and 36% of soil samples, respectively.
It appears that H oryzae is the key pest of paddy, in addition to T. mashhoodi (Ahmad et a!., 1988; Darekar et al, 1990), in many pans of India.
References
Ahmad, W , T. H Khan, and A. L. Bilgrami 1988. Plant-parasitic nematodes associated with paddy crops in Bihar, India. Int! Ncmatol. Network Newsl. 5.4.
Darekar, K S , N L. Mhase, and S S. Shelke. 1990. Plant nematodes of cereal crops from Klaharashtra, India. Intl Ncmatol. Network Newsl. 7:15-16.

Developmental biology of Dorylaimus stagnalls Dujardin, 1845 (Nematoda : Dorylaimida)
Samina SHAFQAT, M . Shamim JAIRAJPURI and Anwar L. BILGRAMI
Section of Netnaiolosy, Depanment of Zoology, Aligarh Aluslim Umventty, Ahgarh, 202002, India.
SUMMARY
Tlie obsenations are made on the embr>'onic and post-embryonic developments of Dorylaimus stcgnaiis. The eggs, measunng 71-95 X 33-45 (82 x 39) jim, are smooth, elongate o'.al, densely granulated and dark brown in colour. The first cleavage is perpendicular to the long axis of die egg and appears 15-2 i h after tne egg uas laid. The blastula and gastrula stages are formed 4C-50 and 75-90 min after the first cleavage. The tadpole stage is formed 215 min later and the juvemle hatched out of the egg 6 h ?fte- lae formation of their internal organs. Maumum hatching is recorded at temperatures 30-35 "C. The first stage )u\eniJes have v.>a germinal and 4-6 somatic nuclei in the genital primordium; the second stage have 4-6 germinal and 8-14 somatic nuclei. Tlic third stage have 8-12 germinal and 20-45 somatic nuclei. The fourth stage female and male have 20-60 and 18-61 germinal nuclei rcspccuve.y. The first stage juveniles require 17-22 da>s for moulting; the second 24-28 days; the third 30-35 days The founh stace female and male require 46-52 and 40-45 dajs for their development, respectively In the founh and final moult, the long filiform tail of fourth stage male juveniles transforms into a shon conoid tail. The total ume (from hatching to adult) of males vanes from 111-131 da>s and that of females from 117-138 days at 25-30'C.
RESUME
Bwlogie dit diveloppement de Dorylaimus stagnalis Dujardin, 1845 (Nematcda : Dorylmmida)
Dcs observadons ont ete faites sur les developpements embryonnaire et postembryonnaire de Dorylaimus stagnalis. Les osufs, lisses, allonges-ovales, a granulauon dense et dc couleur brun-noir, mesurcnt 71-95 x 45 (S2 x 39) [im. La premiere division, perpendiculaire au grand axe de I'ceuf, a lieu 15-24 h apres que I'oeuf ait ete pondu. Les stades blastula et gastrula sont realises 40-00 et 75-90 mm, respocuvcment, apres la premiere division; le stade « en tetard » est realise 215 min plus tard et les juvemles eclcsent 6 h apres la formation de leurs organes internes. L'eclosion maximale est observee a 30-35 °C. Le pnmordium gemtal des juveniles de 1 ' stade comporte deux noyaux germmaux et quatre a cmq noyaux somatiques; chez les juveniles de 2= stade ces chiffres sont de 4-6 et 8-14, et de 8-12 et 20-45 pour les juveniles de 3"= stade; chez les juveniles de 4= stade, le nombre de noyaax germinaux est de 20-60 pour les males et 18-61 pour les femelles. La durec du \" stade juvenile est de 17-22 jours, celle du 2" 24-28 jours, cellc du 3' 30-35 jours, celle des 4" stades femelle et m Jle de 46-52 et 40-45 jours, respcctivemcnt. Lor5 de la quatnemc et dcmicre muc, la qujuc dcs juveniles de 4' stade, allongee, filiforme se transforme en une queue courte, conoidc La durec cntre 6closion et appanuon des adukes, i 25-30 ""C, vane de 111 a 131 jours pour les males et de 117 i 13S jours pour les femelles.
Studies on die developmental biology of dor-j'laims are few, and most of die informations available relates tc the phncphagous species. The life cycle oiXiphinema index and A', mediterrancum was studied by Radevvald and Raski (l9o2), and Dalmasso and Younes (1969, 1070) respecavcly. F]egg(1966; lQ68a,-19686), and Colin and Mo. lechai (196Q) observed die yearly reproduction in the species o(Xiphinema and Longxdonts. Jairajpun and B.ij.m, P'^S). and Malik and Jairajpuri (1983) studied the ciubnonic development of A', basin, X. insipit and X amer.canum. Of the members of Dor\-lainioidoa. the bioloi;y of only two species have been studied in det.ul : Apacclaiiiicllus by Wood (1073) and Labroneiiia vurca-papu'.atum by Grootaert and Small (1982). Piilai and
Taylor (1967) and Flegg (1969) determined the effect of different temperatures on the embryonic development and hatching of some phytophagous nematodes. TTie formation and development of odontostyle in some dorj'laim nematodes was studied by various authors (Coomans, 1963; Coomans & De Coninck, 1963; Coo-mans & Van der Heiden, 1971; Carter & Wnght , 1979; Grootaert .Sc Coomans, 1980).
In the present work the cmbrv-onic and postembrvo-nic development patterns have been studied m DoryL:-;;i!;5ifa^;ui/w Dujardin, IS 15. Besides, obsep.'ations were also made on the dc\ elopment of odontost^, le, gonad and male tail. 1 he effect of tempo-atures on the embryonic development was also observed.
Rciuc .Wmaiol. N d) : 61-71 [19911 61

S Sha/jal, M S Jpi-tvpun iC A L. Bxlgraim I 94
Materi;U a n d m e t h o d s
EMBRYONIC DEVELOI'MENT
7 o obtain fresh eggs, graviJ females were transferred to smnll caviC)' blocks containing 1 "o. water agar and allowed to lay eggs. These eggs were then placed in the cavit)' slide containing 1 % water agar. A cover-slip was placed gendy over it to prevent desiccation of eggs.
ErrECTO.-T EMPER(\TURE ON EMBRYO.N'IC DE\'ELOPMENT
To determine the effect of different temperatures on die embiyonic development, the eggs were kept in small cavit} blocks containing 1 % water agar. Five eggs were placed in each cavity block and ten eggs were used at each temperature. The blocks containing eg;;"? v.crc then placed at different temperatures (5, 10, 15, 20, 25, 30, 35 and 40 "C) maintained in an incubator. Observations were started after 24 h, diree times a day, to observe the
hatching and time required for cmbr}onic devclopmert at each temperature.
POSI-E.MBRYCNIC DC'»LLOr.MFN-T
The moukmg and inter-moulting juvenile stages v.ere collected direcdy from die soil, fixed and dehydrated by slow method. Observadons were made on these stages for tracing the development of odontcstyle and the gonads. For the latter live star%'ed nematodes were also stained in 1 % lact'-accto-orcein. T o observe the moulting time, gravid females wer; cultured in 1 % water-agar.
Desc r ip t ion of DoTylaitmis stagjialis Duja rd in , 1845
DIMENSIONS
See Table 1.
Table 1
Dimensions of adults and juvenile stages of Dorylaimus stagnabs (means are between brackets, n = 50)
Charaaen
Lengtli (L) m m
a
b
c
c'
\
T
Gi
G:
FunctiomI odontosuie d im) Replacement odontO'.n'c (nm) Od, 3nlophore ((im)
0 ^oplngus (nm)
Pre
An
.-•c-tum dim)
il boJ\ di muter (,mn) T i l 1 ((im^
Aduh 9
2 76-166 (3 66)
27 3S-41 86 (36 65)
3 97-6 18 (5 0!)
12 3-32 76 (1941)
2 47-6 !5 (4 1=5)
3 1 8 8 x 1 2 l (40 53)
—
11 41-22 67 (1745)
12 8-20 77 (20 041
37 5-52 5 (42 SO)
—
30-12 (46b'>^
631(1 <;,-l (728 -^1
201 1-U8 (285 01)
3S-5-(17 - >
io:(>-vi (193 7..'
Adult d
2 63-4 41 (3 30)
30 69-42 02 (35 04)
3 78-5 63 (4 73)
68 32-145 0 (86 84) 0 5-0 84 (0 70)
—
57 95-75 59 (67 16)
—
—
37 5-46 5 (40 61)
—
39-16 5 (43 67) 608-811 (695 69)
277 4-770
(465 87) 49 !-6S 4
("31 52) 30 4-11 S
(38 10)
U
0 37-0 87 (0 18)
10 47-3! I (24 37) 2 6-3 7 (2 02)
6 3-102 (7 82) 3 5-6 3 (4 72)
—
—
—
—
7 5 - 1 5 0 (0 61
10 5-20 0 (12451
10^-2! (14 221
125 4-250 {!6l 72 '
30-5-
{-- 21
11 1-19 ( i : o o '
15 p-05
(61 101
L:
0 93-1 49 (124)
26 4-38 0 (32 08) 3 1-40 (3 60) 8 8-119 (1041) 3 8-8 0 (6 05)
—
—
—
—
18 0-25 5 {22 70)
10 5-30 (20 19) 2.-30
(2- 581 292r . -3618
(V.SOI)
c-0-100 5 ( f ; T 7 ) K-22 8
( ' - T 9 ) 91 2-152
( i : o I)
LiO
I 27-2 53 (2 07)
20 5-41 01 (35 07) 3 4-4 7 (4 10)
10 33-19 74 (13 53) 2 8-6 62 (5 07)
—
—
—
—
27-30 (28 71)
37 5-12 (30 01)
31 5-37 5 ( 3 ^ 6 - )
327 1-551 (501 00)
76 5-US6
(15.N54)
26 6-58 (3147)
lOo 1-201 4 (15070)
L> 6
1 OS-2 92 (2 33)
26 0-12 7? (33 60) 3 4-5 1 (4 29)
10 0-34 81 (15 11) 3 0-5 81
(4 5)
—
—
—
—
27-30 (29 35)
31 5-42 (39 94) 27-30
(3107) 311 6 600 1
(519)
115 5-235 6
(200 20) 10-!5 6
(35 53) 6 1 6 - 1 9 7 6 (159 72)
UO
2 10-2 97 (2 40)
23 5-44 66 (34 49)
4 06-5 32 ( 1 4 3 )
11 12-17 47 (15 46)
2 28-6 4 (4 93)
—
—
—
—
31 5-40 5 ( 3 ( 3 2 )
37 5-49 5 (42 87) 33-13 5
O7 05) 505 1 630 8
(519 15)
1 13-201 4
(173 73) 26 6-''S 0 3 5 1 ) 76-212 8
(165 77)
u d
2 17-3 10 (2 56)
33 5-40 46 C36 63)
4 03-5 65 (4 68)
12 64-10 37 (14 78)
3 25-6 54 (193)
—
—
" —
—
3 . 5-35 0 (32 25) 30 45
f43 13) •1 5 - ( 0 5 ,1175) 51 •-'/)(. 6 ^ .< "2)
: 0 ^ 27 5 &
.22'^ 89) '4 2-15 6 . " 36)
; ' . 212 s i s : 55)
62 Rnue i\o-uiu-'. 14 (I) '! il'yQ!'

Di:telop>ncnt o/Doryhimu', jtagnnli";
DESCRIPTION Resul t s
Female : Body slightly vcntrally curved upon fLxation, rapcnng gradi'ally towards extremities but more posteriorly. Cuticle with fine transverse striations, 6-9 [im tliick at mid-body and marked wiih 32-34 longitudinal ridges which gradually fade out towards extremeties. Lateral chords 1/4-1,'3 of corrcspci.' ':ng body width wide at mid-body. Dorsal and ventral body pores 7-10 and 25-30 respective!; '' 'pregionsHghtly narrower than adjoming body, marked h\ a slight depression, 15-23 ^im wide and 6-8 |im higii. En face view shows six amalgamated lips arranged as foilows : two ventrosubdorsal, two lateral and two ventrosublateral; each lip with a p?pil!a on the inner circlet and a papilla on the outer circlet; subdorsal, and subventral lips with an additional papilla: this making a total of 16 papillae.
Ampliids sdrrup-shapcd with slit-like apertures, 8-11 [im wide, occupying about 1/2 of corresponding body v."iddi, 8-9 um from anterior end. Odontostyie cylindroid, 5-6 lip region '.vidths long, forked at its junction with cdontophore. ape.ture 13-15 |im wide or 29-35 "o of odontostyie length. Odontophore simple, rod-like, slightly longer than odontostyie, embedded in the oesophageal tissues. Gl iding ring " double " ; fixed nng at 17-32 |im from antenor end. Length of guiding sheath variable, depending upon the posiuon of odontostyie. Oesophagus doryiaimoid. Nerve ring encircling antenor slender part of oesophagus at 165-247 [im from antenor end. Expanded pan of oesophagus 327-460 (j.m long or 51-52 % of oesophageal length. Location of oesophageal gland nuclei and their orifices are as follows :
DO = S,N, = S,N, == S,N, -
50-54 "o (52 %), 68-83 % (76 °o), 71-86 To (79 °o), 84-92 "o (88 °o\
D N = 53 -56% (55%) S,0 , = 6 6 - 7 8 % (72%) S,02 = 70-83 % (77 °'o) SjO, = 83-90 % (87 %)
04-93 ° J (89 % ), 5 , 0 , = 85 -89% (87%) Cardia well developed, 23-57 jxm long, elongate-cy-
lindroid wiih a disc and surrounded by intestinal tissues. Vulva transverse, slit-Iikc, pre-equatorial or equatorial. Reproductive system ampludelphic, each sexual branch consisting of a reflcxcd o\ary, oviduct and uterus. Prcrectum 201-418 jim long, 5-7 anal body widths. Rectum 45-68 um longer 1.1-1.2 anal body widths. Tail elongate, tapering gradually. 2.7-5.3 anal body widths; tlaree caudal pores on each side.
Ma'.c . Similar to females m general body shape and morphology except for tlie ventrally cun.'ed posterior exiremip,-. Reproductive system diorchic. Spicules dory-b i m o i ' , ventrally cun-cd, 75-112 )j.m long, lateral guiding pieces 9-17 (.im long. S^ipplements consisting of an adanal pair and a contiguous series of 41-47 vcntromc-di.in. Prcrectum 5-1! anal body widths long, extending beyond i.iiige of supplcme:i:s. Tail short, conoid witii blunt'}- lOL'iidcd terminus, c.iudal pores varying fiom 3-5 on C.KII .';idc.
E.MBRVGN;C DE-.-ELOV'.VXNT (Fig. 1)
Each sexual branch of the gravid female possessed 10-19 oocytes at one time. Generally not more than four mature eggs were present in the uterus The c?g shell is made up of three distinct layers (Fig. 1, A) and measures 71-95 X 33-45 (82 x 39) |_im (n = 20). The eggs are smooth, elongate, densely granulated and dark brown in colour. The single-celled egg divided perpendicularly to the long axis into two equal blastomeres 15-24 h after
J / — v:\ F.
I
Fig. I. Embn-cmc development. A • Singlc-ccilcd ^•.-'ec; B : Two-celled stage; C : Four-celled st.ici'; D : Six-cc.loj st.ngc; E : Eighi-ccllod stage; F : Tuclvc celled stage; G 5.xtcen-cellcd st.igc; H : Thirty si.x-celled st.nge; I • Blastuli ^•,.ii;c; ] • G.istruia stage; R • " Lima bean " static; L • " CcrrniM ' stage; M : " Tadepole " stane; N : One fold embryo; O : T.vo fold cnibrj'o; P : Firs; stage )uvcnilc ready to hatch iB^'- -- :0 fun'.
R^vue W'tmlol. !•) {V : 61-71 :'^9l, 63

S. Shafiiat, M. S. Jairajpuri & A. L. Bilgrami Q
cyg laying (Fig. IB). TIic anterior cell divided after 16 mill forming a three celled stage wliereas the posterior one divided 10 min later resulung in four equal blasto-ir.crcs with conspicuous nviclei (Fig. IC). Tlie p^an of division was oblique to die long axis in botli cases. After another 25 min the nvo anterior cells divided perpendicularly fonning a six-celled egg with four small anterior ai.d iv.o la.ge posterior cells (Fig. ID). 40 min later the r.vo posterior cells diviaea leacmg to an eiglit-cfll'-d stage v/ith equal blastomcres (Fig. IE) . The 12-celled stage appeared after 45 inin (Fig. IF) and the 16-celied stage vritltin about 60 min (Fig. IG). In tlie next 2 h the 16 cells divided to form die 36-celled stage (Fig. IH). r'ifter'vvaids the cells multiplied rather rapidly and it became difficult to follow the sequence and plan of cell divisions. 40-90 min later the blastula was formed (Fig. '. I). TTie gastrula stage was formed 75-90 min after the foimation of blastula stage. The gastrxila forming cells were differentiated into two r\'pes : the outer layer of cuboidal cells forming die hyaline portions and the anterior and posterior cells of die inner mass forming the granular portions (Fig. IJ). The anterior region was broad having large ceils formed the future stomodeum (feeding apparatus, ^oesophagus) while the posterior region with smaller cells formed the intestine and tail. Tne gastjula yansformcd into the " iima-bean " stage after 105 min by a slight invagination in the middle of the developing embryo (Fig. IK) and to " comma " stage after anodier 75 min (Fig. IL). Tne " tadpole " stage appeared 35 min later as a result of the formadon of deep invagination (Fig. IM). The mouth depression was formed at the broader (anterior) end of the developing embiyo in the late " tadpole " stage. 160 min later gradual and intermittent movements began in the anterior region of the embn'o. The movements were very slow during the first 15 min but became vigorous 45 min later and spread over the entire body. The developing crnbno increased in length, became two-fold within 25 min (Fig. 10) and three-fold in another 160 min (Fig. 1P). The movemcnis witJiin the egg shell continued during enure process of elongation of embr,'o. The different regions like tlte oesophagus and intestine made a faint appearance after die diree-fold stage was reached. About 150 min later the tip of the functional odontostj'le appeared. The odontost)!e secreting cell was not \'isible at "iiis stage due to heavy granulation and continuous movement. First the tip of the functional odontostylc was formed and dicn it grew furtlier posteriorly. Near the middle of the odontosiyle two refractive dots appeared on the two sides of cdontost^ie and formed the 1 iturc guiding ring. The complete guiding ring along-wit''- the intestine appealed about 4 h after the odontostylc ti[i was formed. The formation of a functional odontostylc took 5 h, followed by the formation of the rep'.accnient odontostylc and the walls of the odonto-p!iorc. The movement oi' the cinbiTO became vciy slow aiicr the foiuuuion of tlic tiigcstivc svstem. This de
crease in the movement was an indication that the juvenile is ready to hatch. Some secretions on the tip of odontostylc of the new fully formed juvenile were seen.
HATCHING (Fig. IP)
The juveniles hatched out of the eggs arpro;dma:e!y 6 h after the fonnation of their internal organs. The egg shell became thin, flexible, increased in v.idtli, and form a blister-like structure at the anterior region. After the formation of the blister, continuous activity of juvenile ruptured the egg membrane. The total duration of embn.'onic development i.e., from single cell stage to hatching varied from 30 to 59 h.
EFFECT OF TE.MPERJ\TURE OK T H E E\!B:',VO:;!C DEVEL-OPAiENT (Fig. 2)
Maximum hatching (70-80 %) was recorded at 25, 30 and 35 ^C. However, the duration of embryonic development was more at 25 °C (86 h) than at 30 'C (54 h) or at 35 '"C (44 h). Least number of eggs hatched at 15 ""C (40 °a) whereas all the eggs remained single celled at 5 and 10 "C and no hatcliing was recorded up to a period of 10 days. Maximum time required for hatching was 144 h a t ' l5 "C and minimum 24 h at 40 'C.
POST-E.MBRYONTC DEVELOPMENT
The post-embr^'onic development of D. siagnalis was studied with particular emphasis on the development of odontost},'le, oesophagus, cardia, gonads and tail.
Development of odoniastyle (Fig. 3)
The odontostyle develops from the cell present in the submedian wall of the anterior slender part of oesophagus. The position of the cell varies in different juvenile
5 10 15 ;0 25 30 35
Icmpfrcjlurp (°c)
Fig. 2. Effect of tc.'iiiK'raturc on cmbn,of;cn?j;s (a) hatching x) of Doiyiaimus stagnaiis.
64 Rciiic h'niuilo!. I-I II' : ol-7I :l''''l!

i S 7 DcveAopnir.nl o/Oonyiaimui "ta^nali-*
Fig. 3. Development of odontostyle in second and tJiird stage juveniles. A-D : Second stage juveniles; E-H ; Tliird stage juveniles.
Revue Kernaiol. 14 (1) : 61-71 :1991) 65

S Shafqat, M S Jairajpun li A I Btlgrami C8
stapes The tip of the replacement odonto<:t> Ic appears fii-st close to the posterior region of odontophore After tip formation, the basal forked part of the odontost>le is formed G^'ieraOy two odontosD,les (functional and replacement) are present in first, second and th^rd juveniles dunng interrnoulting stages As the moulting piocess IS initiated, the functional odontostjle migrates antenorlj and die replacement odontostyle turns on its axis shifting to occup} the position adjacent to the funcuonal odontostvlc The ecd>sis starts with the separation of old cuticle fiom the new one which is secreted b> the h>podennal cells All the cuticular stniciures like lining of the labial papillae, amphids, guiding nng, odontostjle, lumen of odontophore and rectum, e t c , remain attached to the old cuticle The fourth and final moult differs from, all the other moult-mg stages due to absence of a new replacement odon-tostj'le
Development of oesophagus
Along with the shcddingof odontostyle dunng moulting, die lumen of odontophore is also cast off The shed odontophore hning can be seen along with that of the limng of stoma (odontostyle) in the moulted cuticle At tJie time of moulung the structure of oesophagus is pardy obscured due to accurrulation of granules
Development of cardia
The oesophago-intestinal luncuon or cardia is an elongated cone-like suucture The cone shaped structure IS made up of tissues m idults but of cardiac cells in juveniles At each moulting stages, with the formation of odontostyle and other bod\ parts, a new cardia is formed b> the multiplication of cardiac cells These cells appear dark upon staining The number of cardiac cells vanes in different juveniles stages The first and second stage juveniles have 5-8 and 9-12 cardiac cells respective!}, while the third and fou-th stages have 11-14 and 13-16 cells respectivel}
DeiJopmem of gonads
Fust stage juzeniles, I able 1 (Fig 4A) The pn-mordium(9-21 x 6-12 fim) consists of two large coarse, granular germinal nuclei and four to six small somatic nuclei The ventnl choid nuclei vanes from 14-35 from the base of oesophagus to the middle of the pnniordium The Dme taken from hatching to the completion of the first moulting vanes from 17 22 days
Second stage j!izini!,s,Tah]c I (Tig 4B) The gcrmi-nil primoidmm is 15-30 yvm lonc, consisting of four to SL\ gcimiml and 8-1 1 somatic nuclei There arc 40-75 ventnl choid nuclei between base of ocsoplngus and middle of pcimiml pnniordium The second stage juveniles transloim into thud stigc in 21-28 davs
llurd stage ju emles. Table 1 (Fig 4C-E ct 5A-Uj The sexes can be differenuated at this stage The length of male and femn'e p n n c r d i a measured 21-143 ^im and 42-135 |im respceuvelj and each possess 8-12 germinal and 20-45 somatic nuclei The number of ventral chord nuclei vanes from 17Q-190
The juveniles t in t develop into females havf two-four dark oval shaped speciahzed ventral chord nuclei (Fig 4C) In the late third stage the middle part of the p m i o r d i u m bulges out slightly to form a "cone " — like structure (Fig 4E) lepresentmg the future vagina and vulva
The juveniles destined to become males have a small compact mass of daik stained nuclei of spicular pn-mordium in the anal region below thb rectum (Fig 5D; In the third stage male dunng moulung a beickwardlv directed tube (gonoduct) appears in the middle of the genital p n m o r J . „ m which forms the future vas deferens and ejaculatory duct in the adults (Fig 5B) The time interval between the completion of the second and third moulting vanes from 30-35 days
Fourth stage juveniles (Females), Table 1 (Fig 4F) TTie number of germinal nuclei in the pnmordium vanes from 20-60 The somatic nuclei can not be counted because of their large numbers and overlapping arrangement Few somatic nuclei are also arranged on both the sides of the antenor and postenor anns along the wall of the developing gonad which form the epithelial covenng of the gonad The number of specialized ventral chord nuclei increases to 14-20 and arrange themselves around the " cone " With the completion of moulting, the cuticular linings of the vagina and vulva are formed (Fig 4H-J) T h e final moulung took 46-52 days after the third moulung
Fourth stage juvenile': 'Alales), Table 1 (Fig 5C, F, G) The development of gonads in the fourth stage male juv enilcs IS faster than females 'I he number of germinal nuclei vanes from 18-61 The backwardl) directed tube (gonoduct) mcreases in size as the antenor and postenor arms of the developing S)'stem elongate The somatic nuclei are arranged alternately along the walls of the gonoduct D u n n g early stages of moulting, the spicuhr pnmordium becomes more compact and the spicuLs appear as faint hvaline refractor> lines which get shortened and become hard at later stage assuming their normal shape and size (Fig 6) Alanv rows of darklv Stained nuclei arc arranged on the ventral side of the bodv between the cloaca and mtestmc-rrercctum junction (Fic 5E) and giv e nse to copulaton muscles md the vcntromedian supplements The male fourth stage juveniles moult fmall) into the adults 40-15 d i>s after the thud moulting
65 Rc-ue h'cnmul HI 61 ~1 I!" I

5. Shnfqat, M S. Jairajpuri & A. L Dxlsmmx
A ; r s
':::[
t
9
e c 0 0 0
: -/V^. • • : i'pr.'l '
l i / ' l" ' . 0 i : ' « ' »
• • , ; o > ^ = '.' ''n .1 •
_22um_ A,D
SOum
Fig. 5. Development of male genital system. A. : Primordium of the third stage juvenile; B : Primordium of the moulting tliird stage juvenile; C : Primordium of tlic foartii stage juvenile; D ; Posterior region of the third stage juvenile; E : Posterior region of the fourth stage juvenile; F : Fully de^•eloped testes of tlic fourth stage moulung juvenile; G : Moulung fourth stage juvenile.
6S Rcvuc Kemaiol 1' 61-71 (199V

2 C 0 D.ve!o"'-irnt o/ Diyrjl.iitnj'^ -.(-•'i
A
/ .
%'
r • I
s i
Ci ol
1 f . :i /
I? * . l :
' > < • <
r-v
D,/.
. e„
fr"
jaanL.
' : • ; • • • ' I '
..:y «•,
m
F ' . •
r - •* • •
, ' • • . '
K
H;.
, ^
^
^
F'.g 4 Development of Tcr,! no genua! system .\ .'U'.enile; C • Pnmordr.'in f the t'.ird stage )Lvcr Ic,
. Pr.niordium of the fiist <:tage juvenile; B : Pninordlum of the second --lagc D Abnormal pnmordu of third stage ju.cnile, E : Pnmordiuni .M the moulting
third stage )U\e tile, F Pi.inordiuni of fc^''tii stage juvenile; G : Pnmord'um ot the moulting foiuth stag; ]u\cm!e fornniion of viai.a); H : Early fourth. SMITC juvenile; I : Mid-fourth stage juvenile; J • Late fourth stage juvenile
Rjue Ni,r.a!0l 14 (V : ''I- 7 1991) 67

2iA Dcvetcpmcni o/Dor>ljimU3 stagnalis
Fig 6. Development of spicule and tail. A ; Postciior region of ijte third stage nivcnile; B : Early fourth stage juvenile: C, D • Mid fourth stage juvenile; E : Late fourth stage juvenile; F, G : Moulung fourdi stage juvcrule; H : Fourdi stage juvcrule just before final moult.
Rivue Kematol. 14 (J) ; 61-71 (1991) 69

S. Shafqat, M. S. ]a,irajpun & A. L Bilgrami
O .'•: '•) •• I • - '
Development of tail in males (Fig. 6)
The adult males have a shon; conoid tail as compared to female"; which possess a long filiform tail. The male juveniles of the third and fourtti stages also have filiform tails much like that of adult females. The development of male tail starts rather late during the third stage of male juvenile. At the time of fourth or the last moulting, when the old cuticle of tail is getting separated, a new short and conoid tail is formed. The tail of the old elongated juvenile is detached from the newly formed conoid tail of adult male.
Discussion
T~he embryonic development of Dorylaimus stagnalis is on the same pattern as those of Xiphinema spp. (Fiegg, 1968a; 19686); Aporcelaimellus sp. (Wood, 1973); Xiphinema baziri, X. nrsi^Me (jairajpuri & Bajaj, 1978); X. americanum (Malik & Jairajpuri, 1983) except for the differences in the arrangement of blastomeres after the second cleavage. Aporcelaimellus sp., took 230-250h (Wood, 1973); Xiphinema sp. 19-30 days (Flegg, 1968a; 1968W;^. basiri. seven days Qairajpuri & Bajaj, 1978) and X. americanum 100-120h (Malik & Jairajpuri, 1983) to hatch. However, D. stagnalis took only 30 to 59 h to hatch out of the egg. Pillai and Taylor (1967) and Fiegg (1969) obsen'ed the effect of temperatures on the mycophagous nematodes and X. diversi-caudatum and found decrease in the time of hatching with die increase in temperature to a certain limit. During present observations the development and hatching of D. stagnalis is also affected by various temperatures in a similar fashion.
The position and length of odontostyle, replacement odontostyle, odontophore, size of primordia as well as the number of primordial cells were used for differentiating juvenile stages. The development of odontostyle which originate from an elongated flask-shaped cell is similar to that of other dor>'laim nematodes (Coomans & De Coninck, 1963; Grootacrt & Coomans, 19S0; Coomans &,Van Der Hcidcn, 1971). The cardia cells were obser\-ed in juveniles only. The primordium of the first stage juvenile is oval witli two germinal and four to sLx somatic nuclei. With the increase in body and primordial length, the number of germinal, somatic nuclei and ventral chord nuclei also increases. The sexes could be differcniiatcd at the third stage due to the presence of speciali;:cd ventral chord nuclei in females and spicular primordia in males (Hirschmann, 1962). The third and fourth juvenile stages could not be differentiated on the basis of primordial lengths as these overlap.
The fourtit stage juveniles mature sexually earlier than the females as was also observed by Cooninns and Limn (1965) in Anatonchus anuciac, and Ahmad and Jairajpuri (19S2) in Parahadronchus shakili. L^brvnema
ferox and L. tliomci took 3-4 months and 5-6 month") respectively to complete their life cycles (I-crris, 1968; whereas L. vulvapapillatum required 27-126 days for its development (Grootaert & Small, 1982). Aporcelaimellus sp. completed its cycle in 75-114 days as was obser^-ed by Wood (1973). During present observations, D. stagnalis took 17-22 days for the completion of first moulting, 24-28 days for the second and 30-36 dayj for the third moulting. The fourth stage male juveniles required 40-45 days whereas female juveniles took '.5-52 days to complete the final moult. The total time in D. stagnalis from the first stage (hatching) to the adult varied from 111-131 days in males and 117-138 days in case of females.
ACKNOWLEDGE.MENT
The authors arc grateful to the Chairman, Dcpanmcnt of Zoolog>', Aligarh Muslim University, for providing ncccssan,-laboratory facilities.
REFERE.N'CES
AH.SiAD, N. & JAIR-Mi't.-R!, M. S. (1982). Observations of the development of juveniles and adults of Parahadronchus j/iafei/i (Jairajpuri, 1969). Revue Nematol, 5 : 79-91.
CARTER, R. F. & NS 'RIGHT, K. A. (1979). Odontostyle formation in Xiphinema americanum. J. Nematol, 11 : 295.
CoHN, E. & MoRDECHA!, M. (1969). Investigations of the life cycles and host preference of some species of Xiphinema and Longidorus under controlled conditions. Nematologica, 16 : 423-428.
CocviANS, A. (1965). Xiphinema basilgoodeyi n. sp. vWth observations on its larval stages (Nematoda : Dorylaimida). Nemaiologjca, 10 : 581-593.
CooNWNs, A. & DE CONINCK, L. (1963). Observations on spear fomiation in Xiphinema. Nemaiologica, 9 : 85-96.
COOMANS, A. & LIMA, M . B. (1965). Description of Anatonchus amiciae n. sp. (Nematoda : Mononchidac) with observations on its juvenile stages and anatomy. Nemaiologica, II : 413-431.
CooM.'\NS, A. & VAN DER HEIDEN, A. (1971). Structure and formation of the feeding apparatus in Aporcelaimus and Aporcelaimellus(Nemaxoda : Dor\laimida). Z. Aiorph. Tiere, 70 : 103-118.
DALAWSSO, A. & YOUNES, T . (1969). Ovogcnese et em'bn'oge-gencse chez Xiphinema index (Nematoda : Dorylaimidae). Annls Zool. Ecol anim., I :. 265-272.
FLEGG, J. J. M. (1966). Once j-carly reproduction in Xiphinema vuittenezi. Naiure (London', 212 : 741.
FLEGG, J. J. M. (lQ68a). Embrs-ogcnic studies of some Xiphinema :\nd Longidorus species. Nemaiologica, 14 : 137-145.
FLEGG, J. J. M. (1968fc\ Life cycle studies of some Xiphinema and Longidorus species in South eastern England-. Nemaiologica, 14 : 197-210.
70 Revue Nar..iiol. LI II' : 61-71 ilOQlj

Development o/Dorylaimus stagnalis
FLEGG, J. J. M. (IQd''). The effect of temperature on the embrjogcnc'is of Xiphmema dizersicaudalum. Nemaiolo-gica, 15 : 235-286.
FERRIS, V. R. (1968). Biometnc analysis in the genus Labro-ncma (Nemaioda : Dorv'laimid.)) with descnption of L. chomei n. sp. Nemawlogica, 14 : 276-281.
GROOFAERT, P & Coo\i\N-s, A. (1980) T h e formation of the anterior feeding apparatus in dorylaims. Netriatologica, 26 : 406-431.
G R O O T A E R I , P & S ' . L \ L L , R \ V . (1982). Aspects of the biology-of Labronema vuhapapillaium (Meyl) (Nematoda : Dory-laimida) m laboratory culture. Biol Jaarb. Dodonaea, 50 : 135-148.
HiRSCHMANN, H. (1962). The hfe cycle of Duylenchus tnformxs
with emphasis on post embryonic dc%elopment Proc helminth Soc. X^'ash, 29 : 30-49
JAIRAJPURI, M . S & BAJAJ, H . K . (1978) Obser.ations on the biology of Xiphmema basin and X. inngne Rezue Nematol, I : 227-239.
M.%L1K, Z. & jAii^\jPLRi, M S (1983) Population d>namics and life cycle of Xiphmema aniencanum Cobh, 1913. Indian J NemaioL, 7 : 21-28
PiLt^M, J K. ^^TA'TLOR, D P (1967) Effect of temperature on the time requ'red for hatclimg and duration of life cycle of five mycophagous nematodes. Nematologica, 13 : 512-516
R\DFa-ALD, J. D. & RASKI, D . J. (1962). A study on the hfe cycle of Xiphmema index Phytopatholcgy, 52 : 748
W O O D , F H . (1973). Biology of Aporcelaimellus sp (Nematoda : Aporcelaimidae). Nematologica, 19 : 528-5 37.
Accepie'pour pubhcaiton le 16 janvn'r 1990.
Rcvui AVmnto/. 14 11} .- 61-71 11091)

1^ \^ 7
Fundam appl Nematol, 1992, IS (2), 127-131
Thalassogenus shamimi n. sp. (Nematoda : Thalassogeneridae) a nematode predator from India
Wasim AHMAD, Irfan AHMAD, Anwar L. BILGRAMI and Zakaullah KHAN
Section of Nematology, Department of Zoology, Aligarh Muslim Unweistty, Altgarh 202002, India
Accepted for publication 5 December 1990
Summary — Thalassogenus shamimi n sp is descnbed and illustrated It has 1 5-2 3 mm long body, a = 22-32, b = 3 9-5 4, c = 19-31, V = 56-67, buccal cavity 39-58 nm long and 18-27 |j.m wide and is closely related to the two previously descnbed species However, it differs from T paradoxus knAra%'iy, 1973 in having slit-like amphidial aperture, smaller buccal cavity, presence of four cardiac glands and a longer tail It differs from T archaeops Orton Williams & Jairajpun, 1984 in the shape of hp region and longer tail Observations made on the gut contents suggest that it possesses a high predatory potential
Resume — Thalassogenus shamttnt n. sp. (Nematoda : Thalassogeneridae), un nematode predateur provenant de I'Inde — Thalassogenus shamimi n sp est decnt et illustre II presente les caractenstiques suivantes L = 1,5-2,3 mm, a = 22-32, b = 3,9-5,4, c = 19-31, V = 56-67, cavite buccale longue de 39-58 |im et large de 18-27 \xm Cette espece est tres proche des deux autres especes du genre Toutefois, elle differe de T paradoxus Andrassy, 1973 par I'ouverture amphidienne en feme, une cavite buccale plus petite, la presence de quatre glandes associees au cardia et une queue plus longue Elle se separe de T archaeops Orton Williams & Jairajpun, 1984 par la forme de la region labiale et une queue plus longue Les observations faites sur le contenu du tube digestif suggerent que cette nouvelle espece possede une capacite predatnce elevee
Key-words : TTialassogenus, predator
Andrassy (1973) descnbed an interesting nematode species Thalassogenus paradoxus from Papua New Guinea Because of the presence of an eye spot, generally found only in marine nematodes, Andrassy considered It to be a marine residual form Orton Williams and Jairajpun (1984) descnbed a closely related species, T archaeops, from Western Samoa They made detailed observations on the morphology of both the species, discussed their systematic position and proposed a new family Thalassogenendae under Pelagonematoidea (Enoplida) for this genus Regarding the systematic position of this genus we concur with Orton Williams and Jairajpun (1984) Its placement in the family Thalassogenendae (Pelagonematoidea Oncholaimina Enoplida) seems more appropnate than in Mononchina as done by Lorenzen (1981) or m Tryploidina
Dunng 1989-90 we collected two populations of this genus from tropical rain forests in India A detailed light microscopic and SEM study of these specimens revealed that they represent a new species closely related to the previous two known species Observations were also made on the gut contents of these specimens m a bid to understand their predatory ability
Materials and methods
Specimens for light microscopy were killed, and fixed in hot 4 % formalin, dehydrated by the slow method, mounted in glycenne and measurements were made with an ocular micrometer For SEM, specimens were fixed in 3 % glutaraldehyde, washed m buffer, dehy
drated in alcohol senes and cntical point dned using CO2 Dned specimens were mounted on aluminium stubs, coated with 30 nm gold and observed m a Hitachi S 2300 SEM
Thalassogenus shatnimi n. sp. (Figs 1, 2)
MEASUREMENTS
See Table I
DESCRIPTION
Female Body ventrally curved upon fixation, tapering slightly antenorly and more towards postenor extremity Cuticle very finely transversely stnated appearing smooth under light microscope, 3-4 p.m thick at midbody and 6-7 |j.m at tail Lip region continuous with body 32-42 (xm wide, truncated, narrowing shghtly antenorly near oral aperture to become crater-shaped Lips marked with six to eight longitudinal ndges (furrows) running from tip to the base which may help in widening the oral aperture Labial and cephalic papillae distinctly setose, projecting outwards and are arranged in charactenstic pattern of 6 -I- 6 -I- 4 Each papilla projecting from a circular cavity with a thickened nm The SIX outer labial papillae are always larger than the inner and cephalic papillae Inner and outer labial papillae 4 p,m apart while outer and cephalic papillae 6 ptm apart Amphidial apertures small, elliptical slits about 2 5 |j,m wide Three lateral cervical papillae
ISSN 1164-5571/92/02/127 05 $250/© Gauthier-ViUars - ORSTOM 127

20-5 W Ahmad et al
Fig. 1. Thalassogenus shamimi n sp A Entire body, B Antenor region, C Nerve nng and eye spot, D Oesophago-intestinal junction, E Gonad, F Tail, G Tail tip
128 Fundam appl Nematol

iCo Thalassogenus shamimi n sp
Fig. 2. Thalassogenus shamimin sp A En face view, B Lips showing ndges, C Antenorend, D Labial and cephalic papillae, E Lips (laterals view), F Amphids and cervical papillae, G Transverse stnations, H Vulva, I Tail, J Tail tip (Bar = 10^m in A, C, 4 /im in B, D, E, G, J, 5 fim in F, 20 //m in H, I)
Vol 15, n" 2 1992 129

C7 W Ahmad et al
Table 1. Measurements and diagnostic
n L (mm) Diameter a b c c' V G 1 G 2 Bucc cav length Bucc cav width Dorsal tooth from base of bucc cav Cardia Rectum Tail ABD
features of females of Thalassogenus species
Thalassogenus shamimi
Paratypes
1 68-2 3 55 5 84 0 25 5-29 3
4 3-5 4 19-31
16-2 3 56 67
4 4-6 5 2 5-5 7
45 0-58 5 21 27
9-13 5 30-42
210-40 5 72 102
37 5-51
14 (1 94 ± 0 28) (67 7 ± 10 9) (28 8 ± 3 10)
(4 7 ± 0 37) (24 2 ± 3 0)
( 1 9 ± 021) (59 8 ± 3 01)
(5 4 ± 0 85) (4 1 ± 0 78)
(5128 ± 4 01) (24 3 ± 2 46)
(12 0 ± 15) (38 2 ± 5 45) (32 7 ± 7 03)
(83 04 ± 11 13) (416 ± 3 95)
Holot>pe
1 2 1
75 0 28 0
4 8 25 0
181 59 71
6 14 5 7
52 5 27 0
10 5 37 5 40 5 84 0 46 5
n sp T
Itanagar population
1 51-2 1 54-75
22 5 32 0 3 9 4 8
23 9-30 4 1 64-1 80 56 1-62 0
3 5-7 4 3 1-5 6 39-43 5 18-24
9 0 10 5 21-49 30-42 57 72
37 5-42 0
9 (1 79 ± 0 19) (61 3 ± 5 68) (29 2 ± 2 37)
(4 5 ± 0 27) (27 3 ± 2 1) (1 69 ± O i l ) (59 8 ± 1 95)
( 4 8 + 1 34) (4 1 ± 0 89)
(41 5 ± 344) (19 1 ± 1 95)
(9 5 ± 1 06) (30 0 ± 10 25) (30 0 ± 4 45) (65 5 ± 4 4) (38 8 ± 1 84)
paradoxus'
2 2 3 2 5
— 28-31
4 5-4 6 30-36 1 3-1 7 57 60
— —
61-63 25
— — — — —
T archaeops"
4 1 66 1 9
— 29-34
4 3-4 7 34-37
0 9-1 3 59-62
— —
41-47 22-24
9 11 38-48 30-36 39 53 38 45
All measurements m \im cxecpt L * from Andrassy (1973) '* from Orton Williams and Jairajpun (1984)
present just below the amphidial aperture in a characteristic pattern of 2 -I- 1 Stoma barrel-shaped, 18-27 jim wide and 39-58 )xm long, composed of a set of three large vertical and a set of three small oblique plates The vertical plates are 32-48 |j.m Ipng, unarmed and sloping antenorly to form the stomal aperture 7 5-10 5 [im across below the oral aperture The basal oblique plates bear a tooth on each plate at about 9 0-13 5 |j,m from base of buccal cavity Oesophagus 326-520 |j.m long, cylindncal, antenorly surrounding obhque plates and postenorly terminating in a muscular cardia Oesophageal glands opening through dorsal tooth Neither glands nor onfices visible in the oesophagus Oesophageal lumen strongly sclerotized Nerve nng at 142-147 (im from antenor end connected by means of a short commissure to an eye spot Eye spot 7 5-10 5 [im wide and 9 0-13 5 .m long, heart shaped, located mid-ventrally at about 25-34 m from antenor edge of nerve nng and 160-218 .m from antenor end of body The structure of eye spot similar to that descnbed by Orton Williams and Jairajpun (1984) Cardia muscular with four large, almost spheJical glands arranged sub-laterally around the oesophago-intestinal junction Anal opening a large slit almost coV enng entire ventral side of anal region Vulva a transverse sht, 22 |j.m wide, vagina short, 9-14 jim across Gonad amphidelphic, both branches ventrally situated, genital branches small, ovanes reflexed with few oocytes, usually extend upto or beyond vulva Tail conoid, ventrally curved, 57-102 [im long, tip cap-like with a slit-like opening (spinneret) Caudal glands large, prominent arranged in tandem in two sets
130
of three each These glands open in a sac which leads to the extenor through the spinneret
Male Not found
TYPE MATERIAL
Holotype Deposited in the nematode collection of Zoology Department, Aligarh Mushm University, Ali-garh
Paratypes 14 females Two females deposited in the following collections Museum national d'histoire na-turelle, Laboratoire des Vers, Pans, France, National Nematode Collection, lARI, New Delhi and National Zoological Collection, Zoological Survey of India, Calcutta, India and rest in the nematode collection of Zoology Department, Aligarh Muslim University, India
TYPE HABITAT AND LOCALITY
Soil around roots of wild tree from Silent valley (altitude 1500 m), Malaparum, Kerala State, India Collected in December, 1989
Other habitat and locality Soil around roots of banana, Musa paradiasica (L) from near Ganga Lake, Itanagar (altitude 500 m) Arunanchal Pradesh, India Collected in March, 1990
DIAGNOSIS AND RELATIONSHIP
Thalassogenus shamimi n sp comes very close to T paradoxus Andrassy, 1973 because of the shape of lip region and tail However, it differs from it in having a shorter body, slit-like amphidial apertures, smaller buc-
Fundam appl Nematol

Thalassogenus shamimi n sp
20 8
cal cavity, in the presence of four cardiac glands and a longer tail (amphidial aperture pore-like and three cardiac glands in T paradoxus) It differs from T archaeops Orton Williams & Jaira)pun, 1984 in the shape of lip region and a longer and differently shaped tail (labial region almost flat and inner nng of labial papillae inwardly directed, tail not ventrally curved in T archaeops)
O B S E - R \ ^ I I O \ S O \ G L T ( o \ i t - \ i s
The analysis of intestinal contents of T shamimi suggests that it is predaceous in nature and possesses a high predatory potential A total of 26 specimens were analysed and 22 of them had prey either intact or in semidigested condition in their intestine A large vanety of prey nematodes viz species of Hehcotylenchus, Tylen-chorhynchus, Hemicnconemmdes, Pratylenchus, Dory-laimellus, Mesodorylaimus, Aporcelaimellus, Nygellus, Onverutus, Thomenema, Xiphinema, Tnchodorus and Mononchus besides the trophi of rotifers were encountered in the intestine of these predators More dorylaims were present than tylenchs Predators consumed species of Tylenchorhynchus, Xiphinema and Aporcelaimellus m maximum numbers and species of Dorylaimellus, Onverutus, Mononchus and Pratylenchus m least numbers (Table 2) The presence of the trophi of rotifers in fourteen specimens indicate that they were also preferred by the predators The cuticular parts of the prey nematodes such as the stylet, spear, spicules, e tc , were frequently encountered in the gut of T shamimi n sp besides many unidentified (semidigested) species of prey nematodes
Table 2. Analysis of the gut contents of Thalassogenus shatmmi Total number of specimens analysed = 26 Total number of specimens containing prej = 22
Prey
Dorjlaims Tylenchs I ongidorids Mononchs Rotifers
Total number of specimens
containing intact prej
14 11 2 4 0
Total number Total number of specimens
containing unidentified
prey
9 7 1 1 0
of specimens containing cuticular
parts
13 11 0 0
14
Acknowledgments
We thank Prof M S Jairajpun for examining the specimens and for his suggestions This work was supported by the research grants from the ICAR, New Delhi and the DRS programme of the LGC, New Dtlhi
References
A\DR.\ssY, I (1973) Ein Meeresrelikt und eimge andere bemerkenswertc Nematodenarton aus Neuguinea Opusc zoo!, Bpest, 12 3-19
LoREN^FN, S (1981) Entwurf eines phylogenedschen Systems der freilebenden Nematode n Veroff Inst Meeresforsch Bremerhaven Suppl, 7, 472 p
ORION Will lAMS, K J & ]MRAJPL m, M S (1984'> Observations on the morphology and systematics of Thalassogenus Andrassy, 1973 (Nematoda Thalassogenendae n fam) Rec Aust Mus, 36 1-6
Vol 15, n" 2 1992 131

2 G 9 Si'malaluiiua J7 (\9'^l) n 'J-J42. t j Brill, Lcide
OBSKRX'ATIONS OX T H E FRKDATION ABILITY OF APORCKI.AIMELLIJS MVALIS {\\:VU?.RK, 1932) HFA'NS, 1966
(NKMAFODA: DORYLAIMIDA)
/ KHAN. \ L. BILC,RAMI and M S j \ IRAjPL RI
SittiMii 111 Xi'mdtolot^v Dip jnmcn t ol /(niiu!J\ \li^a h Muslim Lni\ciMt\ \lis(drh-J()2(Hli. India
Prt'dation i)\ AtKiriflaimellus ruiah\ dipt'iuk'd lai!{fl\ on ihan t i ciKounters with the pu-\ but the pre-fccdin<^ atiiaction and ai(^rcgation ol up to 7 ptcdatois aioiind an alicach in|uictl pre\ at a Iccding si(<- is an induatioii ol positi\( pcucption ol pic\ bodv lluids Pic\ catching and kcding mechanisms comprised ll\e distinct phases \ i / , eiicounlei with p ie \ . att<ick lesponsc. attack. saliN a'iciii • \ti a-coipoic al clmestion and mm'stion teedini; Rate ol piedation un iamed unchani^ed o\ c i a |)c nod ol eight da\ s but pie\ luiiiibei tempeiatuH , agai concc nliatioiis. stai-vat ionol thc picdators, t\pc ol the pie\ etc , dc lemiined piedation A nwa/n pielei led second stage juvenile s ol Miloidvi^mi imiis,nita (Koloid & V\hite, I'M')) C.hitwood, 1')!-') and Helerodera miithi Khan & Husain, 19b5 to other spec le.s ol piev nenidtocles \c) piedation took place when Hiiplolaimu\ uidu'i-' Shei. I'lb'i, I!ihiut%lenchu\ induiis SiddiC|i, I'ldi and Himu\(tiopliiita sp weie used as pie\ thick cuticle, bocK annulations and toxit or unlaxouraljlc pre\ secieticins weie consideied till like 1\ cause .-Xdult (male and fc male) .4 nna/n killing maximuin nuinbei ol pre\ nematodes w.is the most active picdator stage
Kiuvords ipdiii 'aimillu^ nivalh, piedatoi. | ) ic\ , piedation
Many s]K'ci('s of dorylairris belonging to the genera Dorylaimus, Labronema, Discolaimus. C.'anhaiolaimu^. Aporcelaimus, Aponelainiellw, ha\-e been reportecd to be pretJatciry in nature ( Iliorne, 1930, 1939; Thorne & Swanger, 1935; Lin-ford & OliMera, 1937; FJsser, 1963). Apart Irom these easual observations, few detailed studies of predatoiy behaviour oi dorylaim nematodes are available. V\Vss & Cjiootaert (1977) and Small & (irootaert (1983) were the first to gi\e a detailed account oi predation by a dorylaim nematode, Labronema vulvapapillatum Loof & Grootaert, 1981 iollowcd by Bilgrami et al (198.T) on a nygolaim nematode, Aquatides thornei (Schneider, 1937) Ahmad & Jairajpuri, 1979 and Shdlcjat et al (1987) on another dorylaim, Dorylaimm btagnahs Dujar-din, 1845, I'he dorylaim predators are omnivorous feeders which also derive their food from algae and fungi as well as feeding on nematodes (Hollis, 1957; Ferris, 1968; Wood, 1973). Fsser (1987) and Bilgrami (1990b) have summarized various aspects of predation by dorylaim predators but more information is recjuired to establish a true picture ol their predatory proiile
A detailed study on the predation abilities of Apunelavnellm nivalis (Altherr, 1952) Heyiis, 1966 was made to assess its predatory potential and to determine the effects of various factors governing predation.

210
MM PRIM S WD \lh IHODS
Ditli i ini stdt^ts of Apurnlaimellus nualis wi rt obtained irorn tht soil collected Irorn pddd\ fields ncdi Jairidlpur Alit^arh India All txpcnmtnts were carried cnu in siiiall cavity blocks contdinint^ a 10 mm deep layer ot 1% water atjar Fi\c 1( malt prcdatois were tested together at a time using 25 individuals of Htr<;cfimannuUa oryzai (van Rieda de Haan 1902) Luc & Goodev 1964 and the sKond stage juveniles ol \i(loidoi>yn( incognita (Koioid & White, 1919) Chit wood 1949 in scpdraie cdvitv blocks Fach experiment was replicated five times md caiucd out at 25 + 1 °C 1 he number of pre) killed or ingested was counted after 24 h \\\ conditions vsere similar during each experiment unless othenvise slated
Pre} (atdiiniJ and fadinti rmcharmmi Prev catching and feeding mechanisms of 4 nualis were studied bv inverting the culture dishes containing predators and prev uiidei a steicoscopie microscope l o study the feeding mechanism in detail ol)sc IV atioiis vveie made at x 400 oi higher magnifications using obser V ition eliambeis similar to thcjsc used bv Ahmad & Jairajpuri (1979)
lllraition tuiiards pn\ '\ttiaction o{ A nivalis towards prey nematodes was testcil using 7 em diameter Petri dishes which were marked at the bottom into s( ve n see tors e)f 1 em width each as in \/m\ & Jaira|pun (1977) and Bilgrami et al {198 i) I he sectors were marked as 1 2 ^ 7 A. straw pipe 1 cm long and I mil m di iinctci se tied at one end with a small piece of filter paper was placed \eiticallv in the middle ol seetoi 1 I he sealed end of the straw pipe lemamed it the bottom of the dish Fiftv prey individuals were incubated within the stiavv pipe tor 24 h bclore releasing 25 predators at various places in secioi 1 1 he distribution of predators was recorded after 24 h Separate e \pe rimen s we re conducted to test the attiac tion of predators towards live and dissccKd ((Ui into two parts) piev nematode^ Pctii dishes without prey neniilodes weie tieated as contiol The lesponscs of 4 nualis were tested tow aids H orx^ai
Ral( (i pudatwn 1 he late of predation b\ th( male female and various juvenile st ii e s of 1 nualis was detcimined usiiif both the pre\ separatelv
hidin^ paUtrn I he feeding tiend was obseived over an eight dav period 1 he numbi t ol piev individuals killed was counted after cveiv 24 h The preciatois we rt tiansftircd to another cavit^, block containing the same number and tvpe of prev each da\ after observation
Pre] pufmiKt To determine the choice of piedator M incos^nita Heterodera mothi H or] ai 1 ilemhorhimhus inashhoodi Siddicp & Basir 1959 Hoplolaimm induus HiliK tihruhus induus and Himuycliuphora sp were subjected to preda tion bv 1 III alls m scpatatc cavitv blocks
Pnt SI lit turn in combination Diile le nt prev combniitions (v iz 25 M incognita + l)H ut\^a< 25 \i incoi;nita + 2.1 I mashkoodi 25 W incognita + 2.5 H mothi, 25 / / or\..ai + 25 I mashlwodi) were used to determine the preftienet by A nualis

r> 1 1 PRl l ) \ l U ) \ (.)! U(>K< H ^l\IH I I \ \/l It [\ 3 3 5
Effect uj prt} dnisity The effect ot prey denbity on the rate of preddtion bv 4 niialis was observed using 21, 50, 75, 100, 125, 150 175, and 200 mdi\ iduals ot pre\
Effect oj itanatwn 1 he effect of starvation on the rate of predation was oljservcd using starved predators (kept without piev m water) tor 0 (control), 2, 4, 6, 8 and 10 davs
Effect oJ Ifmpfratiire To studv the influence ot tempeialuu on tlu laK of picdation bv A niialu, the prev nematodes vvcre subjected to predation at 5, 10, 15, 20, 25, iO, 35 and 40°C
Effect of as,ar concentrations The predators were released in cavitv blocks containing 0 5 , 1 0 1 5, 2 0, 2 5, 3 0, 3 5, 4 0, 4 5, 5 0, 5 5 and 0 0% water agar together with prev to observe the effect of agar concentrations on their rate of predation
R l S L f I ' s
Prey catctiimi and feeding mechanisms The prev atclung and leeding mechanisms of .4 niialis mav be divided into five more or less distinct phases VIZ , encounters with prev, attack response, attack, salivation with extracorporeal digestion and mgestion/ieeding (Jairajpuri a Bilgrami, 1990)
Predator piev encounters occurred mostly during the apparently random movements and hence were a matter of chance The pre\ nematodes were attacked onlv when predators made lip contact with the prev at 90•" 1 he predator pre)bed the prey's body with its lip betore mitiating an attack A nivalis did not show a preference for anv particular par of the bodv of prev to attae k During probing a few oesophageal pulsations were obse rv ed Alter probing a suitable spot, the predator attacked the prev with its stvlet Generally 7-12 stylet thrusts were enough to achieve cuticle penetration However, not all attacks resulted in prev wounding and otten the prev took evasive action I he stvlet was thiust deep into the bodv oi prev and rendered it imniobile bv disorganizing the bodv organs During ingestion the stvlet remained within the body of the prey and oesophageal secretions were released bv the predators tor extra-corporeal digestion Ingestion was intermittent i e , with short periods between sucking activities A niialis punctured the prev cuticle at more than one place m order to ingest all the body contents Only the distorted cuticle of the prey remained when ieedmg was completed A nivalis took ^0-^5 mm to completel) consume an individual ot H oryzae Up to seven predators were observed to aggregate around an injured prev and teed together at one site A nivalis was also observed lapping (side to side lip rubbing) their eggs but attacked and fed on the eggs of prey nematodes in culture
Attraction touards prey {Fie, 1) A ;2«'a/n was not attracted either towards live or dissected prev There was no significant diflerence in the distribution pattern oi predators in diflerent sectors ot the Petri-dishes ( p > 0 05)

9
i3b K H W B I I C R W I I V ) \ I R A ) l ' l Rl
6 \^ o 'a A •o a/
Live prey ^ E x c i s e d prey
^
L 1
I I,
^
L ^
I 1 2 3 A 5 6
Zones
Rate of predation (Fig 2) All stages except the first stage ju\enile ot A niialn art predatoi\ Males and females killed equal number^ of prev ( p > 0 01) The number of M imognita ]uveniles and H oryzae adults killed bv fourth, third and second stage ju\ eniies was less than that consumed b\ then adults ( p < 0 05) However, all stages including the adults consumed sigmfkanth nioie \l ;«co rHta ju\enik s than / / oryzae adults
teedini^ pattern (fig 3) 1 he rate of predation bv A rmahs o\ er a period of eight days remained unchanged ( p > 0 05) There was no difference in the number of prev killed bv the predators on the first and eighth day ( p > 0 05)
Pny preffnriK (Table I) More second stage juveniles (jf M incosiuta and H mothi were killed ihan any other species used as prev ( p < 0 05) H oryzae was killed in nroderate numbers while F rnaMoodi the least iNone of Hoplolaimus indicm, Heluolilemhui induui and Hemuychophora sp was injured or killed bv the predators
Prey selediori in combination (Table II) When placed in various combinations of prey T ^aa/n killed and consumed more V/ incognita dr\A H mo/A; juveniles than other prev nematodes ( p < 0 05) 7 maihhoodi was the least preferred species of { rev Predators preferred// orjzae lo i / inro 'Wito in this experiment also
Effect of prf] density (Pig 4^.) \ Iaximum picdation occurred wrth 150 oi more prev and the mrnimum in a population of 25 piev individuals ( p < 0 05) More M incoi>nita juveniles were killed than H oryzae ( p < 0 05)
Effect of stariation (Fig 4B) Starvation of A niialn for varvrng perrods drd not affect Its rate of predation on M tncoi;ntta juveniles and H oryzae adults The number of prev killed bv fresh predators and those starved for 10 d remamcd unchanged ( p < 0 05)

P R F I ) \ 1 \0\ Ol APOlU H AIMH H S \ / l Al IS
213 337
8. T3
S 6,
o d
2.
I
1 1
M.incognita juv. ^ H.oryzae
i i I B D
'liT 2 Ratt ot pii'ddtion In adult and ]uvcnile stancs ol ,1 niialu A Mak B Fcniak (' hdurth stage juvenilf D Third stage juvtnik h Snond stagi juvenik
12
1QJ
TJ 8 01
S.Z,. o
i 2 X—X M.incoanitg juv.
• — • H . o r y z a e
3 ^ 5
No. of days
Fig 3 Predatory pattern ot A nivalis
Effect of temperature (Fig. 4C): Temperatures between 25 and 30°C were most suitable for A nivalis predation. Temperatures below 25 and above 30°C markedly decreased predation (p<0.05) .
Effect of agai concentrations (Fig. 4D): Predation by A nivalis decreased with increase in agar concentration. Maximum predation was recorded in 1 to 2% water agar. Concentrations above these caused a significant (p<().05) declme in predation No prey was killed by the predator at 5.5% or higher concentration of water agar.

s - .
« 8 k H \ \ KU (,R \ M I s ) \ I K \ | 1H RI
1 Mil t I
Prey i elect ion
\'lt\
\lelnidoi!\tH iiuvi^nila | u \ Hetaoder(s viatni ]\\\
HiTschmu'inul''! or\^ai
1 \limhoih]niiiii\ ma^lihuudi
Helnolyliiuhin in(luu\
Hupliilainiu\ J tutu i\
Hnnuyaiiiphiiiu sp
\ i . ot inoc ul;
r i ' i
1') 2') li
!-> i )
pif\ i t cd
\ 1) o) pr<r\ k l l k d
11 l(t
~ .
(1
0
0
R ins(t
') l i
*l 12
'i 8 > ) — — —
S 1) +
1 M 1 iO 1 14 1 14 — — —
S ?
I
0 70 0 =)8 0 =)() (1 11 — — —
r \ B i F I I
Prij seldlum h] \ [ii\ ali-i in difftunt t_ombinatuin\ ol pu\
F K V
\irlindu^ ni rnitiinita ] u \
Husihrnanrii'l'a (ir\zai
\liloidn^]n( uHounita )u \
/ \leruhi rlnmhin mashluxidi
\i(loul(i^)n( nioiinita Ju \
Hitiroaaa nmlhi fu\
Hnsihrnami iiui ur^zai
11 /(IKn irh] III nil ma hhuorii
\ o ol p n \
inoculdlfcl
Ii
>'-> li
J i
li
li J)
r-)
\ ( ) ol pr t V
killed
11
) HI
) 10
4
b
i
R an m
<) H
K) ci 1 i
J T
4 12
8 11 ) 8
2 )
S D +
1 )8
0 84 2 07
1 14 1 50
1 10 1 iO
1 U
S L ±
0 70
0 40 0 42
0 ")1 0 58
0 58 0 58
0 50
DISC LSSK)\
Oui ohsi r\ dtioiis inditdtt that \ nnahs is prcdaceous in naturi Prt dation dt'pt iided on thanct cntounttrs (Xtlmes, 1974, (Jrooiaeit k Maeitens 1976, Wvss &. Cirootaeit, 1977, Small & Grootaeit, 198J, Bilt(iarni et al 1984, Bilt^iami et al , 198"). Shatqat et al , 1987) with diiferent nciiiatodc species I h e i e was no attiattion ol A niialu towards pie\ Howcxer, the ag"i r(. g'atton ol more than one predator around an injured piev at a leeding site (pie-lecdins^ atrgretijanon, Bilgrami & Jairajpun, 1989b) and thtir subsequent let ding togethei on that pre\ is simpl) an induation oi a positi\c iespouse of pudators towards piev secietions 'I his phenomenon is ol common occurrence in most of the dorvlaim and nvgolaim predators (Essei 196J, V\ \ss & Grootaeit, 1977, Bilgiami el al , 198'i, Shalqat et al , 1987) Stiong chemoperception has been

? 1 5 PKl / ) \ I l<)\ Ol iPOR( H mil-11 I \ Ml U /s rw
25
2Ql •o
15.
°- 1Q o
5.
X X M.i'ncoqnitg juv.
12'
IQJ
0) -
>- ^ 0;
Q.
O
I
A 1A
ia
i-hiA
— 1 — 1 — I — I — I — I — I — I 25 50 75 100 125 150 175 200
Prey Number
/?xj/-'~^n\i/i a
\ /
rl\M'H^A-\
TTTTTTTTTTiB Starvation (days)
~i r 1 1 1 1 r 1 5 10 15 20 25 30 35 ^0
Temperature (°C)
1 1 1 1—I 1 1 1 — r 1-0 2-0 3D A.0 5.0
Agar concentration (•/,)
Fig 4 Rati (il prcddtion b) A mvain A Etiect of p u v nunibiis B Fllt-ci of s tauat ion i', Effett of temperature D Effect of agai tomenliat ion
observed in cliplogasterid predators (Yeates, 1969; Grootaer t et al , 1977;
Bilgranii & Ja i ra jpur i , 1988, 1989b; Bilgrami, 1990a)
T h e prev catching and feeding mechan isms oi A nivalis are similar to o ther
doryjaim and nygolaim predators , e .g. , Labronema vulvapapillatum (Wyss &
Groo tae r t , 1977); Dorylaimus stagnalis (Shafqat et al., 1987) and Aquatides thornei
(Bilgrami et al , 1985), but was a more active predator and kills more prey per
day.
The rate of predation by A nivalis remained unaltered over a period of eight
days but some of the prey nematodes viz., M incognita and H mo/Az juveniles

3 4 0 KUAN BII (.R \\4I \ I \ IR \ | I 'L Rl
were killed and devoured most. These differences could be attributed to the size and type of prey and the activity of predators and prey (Bilgrami et al , 1983) and to the physical, chemical and behavioural characteristics that provide resistance and susceptibility of prey nematodes to predation (Crrootaert et al , 1977, Small & Grootaert, 1983; Esser, 1987; Bilgrami & Jairajpuri, 1989a; Jairajpuri & Bilgrami, 1990) The lack of predation on Hoplolaimus indicus, Helicot\lenchus indkui and Hemtcyclwphvra sp. may be attributed to these factors-jjhysical characteristics (thick cuticle and body annulations) in Hoplolaimw, indicui and Hemicycliophora, respectively, and chemical (unfavourable body secretions) in Helicotylenchm indicus As in Oiplentewn colobocercus (Yeates, 1969), Prwnchulus punctatui (Nelmes, 1974), Aquatidei thornei (Bilgrami et al , 1985) and Mononihoides longicaudatus and JW Jortidens (Bilgrami & Jairajpuri, 1990a) predation b\ A mvalh also depended upon the density oi prey. The increase in predation per day with the increase in the number of prey may be attributed to increased chances of contacts between predat(jrs and prey The activity of predators and the prey is affected by the temperature and agar concentration and thus governs predation (Bilgrami et al , 1983). This could be the reason for differential responses of predators at different temperatures and agar concentrations The differences in the number of prey killed by the adult (male and iemale) and various juvenile stages reflects their predatory potential, adults having greater predatory abilities than their juveniles.
Like A Ihurnei and D stagnahs, A nivalis exhibited alfinitv towards its own eggs (Bilgrami et al , 1985, Shalqat et al , 1987). The predators attacked and consumed the eggs of prey nematodes when in contact with them in culture dishes.
/ U S A M M F N I - A S S b N G
B(obaihtUTii^en ulitT die raubenschen Fa/n^kulen ion ApoueLumelius nivalis (Althin I'liJ) Hnn\ / % 6 (\imaloda Donlaimidaj
Das Bt utenidchcn \ on A port elaimillus niialis beruhte vvcitt; hcnd aui zulalhs^cn Bft;c^nuni^t'n nut Biutfticri II, doih sind die voi deni Fresscn bcobaLhtat Ankxkun^ und die Ansanimlunii \(in bis /_\j 7 Raubern an emei schon \ei let / ten Beute Hmweist aut eine positive Wahinchniunsr del Korpdflussiijlveit von BeutetRren Die Mechanismen \on Beutetani> und Nahrungsaut-nahme uniiassien luni deutlieh untersihiedene Phasen Bet^egnung; mil der Beule. Angrittsreak-tion, Ant;ritl SpeKhelabgabe'e\iiakt)ipuiale W i d a u u n g Nahiungsauinahnu
Die Rate d( s Beutemaihens blieb ubcr einen Zeitiauni \ )n acht 1 agen un\ ei<iiid( it. doth wurde SK bestimmt duuli dii ' \n/ahl dei Beutenue. Umpt ia tu i Agaikon/cntiatioii, Cirad des Hungers di r Rauber, Art der Beute usw A nnaln /og Ji-Statlien von Mdoidnt^ini incni;nila (Kofoid (t VMiite. 1919) (;hitw()()d 1949 und HeUrodera molhi Khan & Husam 19bT anderen Arten von Beutenematoden vor Hoplolaimus induus Sher, 19bk Heluotyleruhus indicus Siddiqi, 1963 und Herniiyiltophora sp vvurden nieht getangen Duke Cutuula , staike Korpenii igt lung und unangenehme Ausseheidungen vvurden als wahrst heinluhe I'rsachen hiertui bttrachtet Erwachiene (Mannehtn und Weibchen) von/I nH'a/n totetin die mtisten Beutetieu undwaren somit dds aktivste lauberisthe LntvvKklungsstadium

- > ' 7 PRl DA I l ( ) \ OF iPOKC Fl \IMI n I S Ml il IS 3 4 1
R P P t R l \ ( 1 S
\ H M \ I ) 1 (t IxiRxjfi Ki \ 1 S (1979) l)i vtlopinint il hiolot^v oi ihiloplaius s\mnielruus Indian journal )f \emaliiloii) (1977) 7 125 H9
^/Mi \ I I S J A I R \ ) P L R I M S (1977) \ t t ra( tion of plant parasitu m m itodes to hiibt roots \ematolotica 23 119 U l
BiiGK\Mi \ L (199(ld) Diplogdsti rid puda to i s In \emalode Bwcontrol Aspati and Pruspeits ( t d s \ I S Jaiiajpun M M Mam & I Ahmad) ( BS publishers and Distributors Pvt 1 td \ e w Dtlhi pp 13-1 142
BiLCRWii \ [ (1990b) Pifddtcous IJoi)laiins In \nnalodi Buxonlrol lipiili and Pwipali ( t d s \ I S )diraipuii Vl \1 Mam & I Ahm ul) C BS pubhshus and Distiibutors Pvt 1 td \ o v D.lhi pp 143 148
Bii(R\Mi \ I A H M \ D I & J\iR\jPiRi M S (1983l Sonit lattois inllutncint; pudat ion b \ \Uirionihus aquaticm Reiue de \ematolo^ie 6 i25 26
BiifRAMi \ I AHMAD I & J A I R \ | P L R I M S (1984) Obstixations on tht bihaviour of Mononihui aqualicus \ematulo^ia mediterranea 12 41 45
BiKRAVii \ I AHMAU I (tjAiRxjFiRi M S (198i) Prtddtorv behaviour of/li^ua^irffi Moraf! (\vtjolaimina \(ma(ocld) \ematolo^ica 30 (1984) J57 4(i2
BiKRAMi A I STJAIRAJPI Ri \ I S (1988) Attraction of \'luniin(holder tunt;uaudatui dmi \1 for tidtns ( \ n n itoda I)iplot;dstt rida) tow irds prt v and lac tors in fluent int) dttidction Reiue de \irnali/(,i>ii 11 195 202
BIIGRAVII -X I n. JAIRVJPI Ri \1 S (1989a) Rts i s tmt t ol piev to pitdation dnd stiiki latt ot tht predators Munonchoidei longicaudatus and \4 firtidtm ( \ emdloda Diplos^asttiida) Riiue de \em.at<ilogu 12 43 49
Bii&R'Wii A I SI JAIRVJPI Ri \ I S (1989b) \ge^n^M]on of Vlummihoidei lon^icaudalus dnd \^ fortiden ( \ ematoda DiploEfdstiridd) at feeding sitts \ematolo^ica 3'i (1988) 119 121
hssFR R P (1963) \ t m a t o d t interactions in platts ot non-stt rik watti agdr Proieediniii of Soil & ( rop Science Society Florida 23 121 138
EssER R P I 1987) Biological control ol plant parasitK m matodcs bv nematodes I Dorvlaims (Ncrnatoda Dorvlaimida) \ernaiolog} Circular \i) 144 pp I 4
FERRIS \ R (1968) Bionu trie analysis in the genus/airon^ma ( \ cmdtoda Doiv laiimda) with a description of L tliornei n sp \ematoloi;iia 14 276 284
GROOIAITRT P J^yLLS A SISMAII R W (1977) Pie> sc lection in/Ju^feni/j sp (Rhabditida Diplogastcndac) Xiededeltni^en van de Facutteit I andbauixaelinschappen Ri^ksunwer^iteit Gent 42 1559 1563
(jRooiAFRT P & MAFRTLNS [) (1976) Cultivation and life cycle of Vlanomkus aquatuus \ematolo(;ica 22 1 7 ^ 8 1
HoLi IS J P ( 1 9 D 7 ) Cultural studies with Dor))/a!mui c«frr/ier?cnjH Phylopatholui^) 47 468 47 5 JAIRAJPI Ri \1 S it BiicRWii A 1 (1990) Predatory nematodes In \imatodi Bwcontrol
A ipeiti and Prospects (hd',\l S Jaira|puri M M Alan &I \ h m a d ) C BS Publishers and Distributors Pvt l t d New Delhi pp 95 12)
1 iNfoRD M B & OiiviERA J M (1957) The feeding of hollow spear nematodes on other nematodes Science 85 295 297
\ t iMFb \ I (1974) tvaluation of the feeding behavioui ol Pnonchulus punclatus (Cobb) a nematode predatoi Journal of Animal Ecology 43 i55 565
SHAFyvi S BiiGRAMi A L & JMRvjPLRi M S (1987) Evaluation of the predatory bchavioui ot Durylaimu^ sla^nahs Du|aidin 1841 (Ntmatoda Dorylamuda) Reiui de \ematoloi;ie 10 445 461
SMAI I R W & GROOIAERI P (1983) Observations on the prcdation abilities ol some soil dweliing predatory nematodes \ematolo!{ua 29 109 113
1 HORNE () ( 930) Predactous nemas oi the genus \yi>okimus and a new genus Seclonema Journal oj Ai^ricultural Research 41 445 446
1 HORMF ( I (1959) / monograph ot the nematodes of the superfamily Dorylaimoidea Capita /ooloi;ica 8 1 261
1 HORNE (j Si S\v \NGER, H H (19 56) A monograph of the n matode genera Z^ory/afmns Du|ar din \pcircelairnus n g Dorylaimuidei n g and Pungcntus n g Capita Zooloi;tca 6 1 22 5

2'iS 342 KUW BII CRWII •. IMRAIPl Rl
Wuul) 1 H (l')7"i) hmiiiiE; ulatioiiships ol soil-clvvtllint; ntmaluclts Sail Biolai^, unit Bwiluynisir} 5, )4 i ()()]
V\ •> ss L it ( J R O O M P R I P ( 1 ' ) 7 7 ) PttdiiiE; medianism ol Labmnema sp Mrdidilim^iri lan di tacultitl I.andbouuuetenuhappen Ri/k\uniiirMteil Cnnl 42, 1 J 2 1 - 1 " ) 2 7
\tATF!> CJ V\ (1964) Prtcldtion b\ Monomhoidis polohikw, (Neniatodd Diplo^aslti ida) in laboratoiv culturt' \emalnlniiua 15, 1 ')
Xcitpted for publuatiun Apii l lH 1991

bundam appl Nematol 1992 15 (3) 265 270
Resistance and susceptibility of prey nematodes to predation and strike rate of the predators,
Mononchus aquaticus, Dorylaimus stagnalis and Aquatides thomei
Anwar L BiLGRAMi
\(chon i)f N(rnahd()ify Department of/oology Aligarh Muslim Umiersity Alii^arh 202002 India
Accepted for publication 10 June 1991
Summary — The strike rate of the predators, Mononchus aquaticus, Dorylaimus stagnalis and Aquatides thomei and the degree of resistanee and susccptibihty of/4cro6e/o!rfe5 sp Cephalobussp Rhabditt sp , Panagrellus redwivus, Tylenchorhynchus mashhoodi, Hoplolaimus indtcus, Heticotylenchus indicus, Scutellonema sp , Hemicycliophora sp , Hemicnconemoides mangiferae, Hirschman niella oryzae, Longidorus bp , Paralongidorus am, Paratnchodorus sp , Xiphmema amencanum and the second stage juveniles of Meloidogyne incognita Heterodera mothi and Anguina tntici to predation was measured and quantified using methods of Bilgrami and Jairajpuri (1989ay M aquaticus was the most successful predator with maximum strike rate (SR = upto 100 ) on various species of nematodes \11 saprophagous nematodes were highly susceptible to predation (PS > 90 ) except Rhabditu sp which showed some degree of behavioural resistance in the form of active body undulations Hehcotylenchus indicus resisted predation by chemical means, i e toxic unfavourable secretions X amencanum, P can, Longidorus ^p , and Paratnchodorus sp arc provided with physical characteristics (cf cuticle) which provided partial resistance against predation Hoplolaimus indicus, Scutellonema sp , Hemicycliophora sp and H mangiferae were totally resistant to predation by D stagnalis and A thomei, their individuals were neither injured nor killed by the two predators
Resume — Resistance et sensibtlite a la predation de nematodes-protes, taux d'agressivite des predateurs Mononchus aquaticus, Dorylaimus stagnalis e< Aquatides thornei — La methode de Bilgrami et Jairajpuri (1989a) a permis de mcsurer et de quantifier le taux d agressivite des predateurs Mononchus aquaticus, Djrylaimus stagnaltstt Aquatides thomei ainsi que le degre de resistance ou de sensibilite des nematodes Acrobeloides spp , Cephalobus spp , Rhabditis spp , Panagrellus redivivus, Tylenchor hynchus mashhoodi, Hoplolaimus mdicus, Hehcotylenchus indtcus, Scutellonema sp , Hemicycliophora sp , Hemicnconemoides mangiferae Hirschmanniella oryzae, I ongidorus sp , Paralongidorus citri, Paratnchodorus sp , Xiphinema amencanum et des juveniles de deuxieme stade de Meloidogyne incognita, Heterodera mothi et Anguina tntici M aquaticus s est montre le predateur Ic plus performant avec un taux d agressivite (SR) maximum (jusqu a 100 ) envers differentes especes de nematodes lous Ics nematodes saprophages sont trcs sensibles a la predation (PS > 90 " ) sauf Rhabduis sp qui a un comportement de resistance grace a des ondulations rapides du corps Hehcotylenchus indicus resiste a la pn dation au moyen de substances chimiques (substances toxiques ou non favorables) X amencanum, P citn, I ongidorus sp , et Paratnchodorus sp ont des caractenstiques physiques liees a la cuticule qui leur conferent une resistance partielle contre la predation Hoplolaimus indicus, Scutellonema sp , Hemicycliophora sp et H mangiferae sont totalement resistants a la predation par D stagnalis et A thomei, leurs individus ne sont en effet jamais blesses ou tues par ces deux predateurs
Key-words : Mononchus, Dorylaimus, Aquatides, prey nematodes, resistance, predation
The predatory nematodes possess different mechanisms to overpower their prey and to feed upon them Similarly, the prey nematodes also have charactenstics, hereditory or acquired, to defend themselves from predation (Bilgrami, 1990 a, b, Jairajpuri & Bilgrami, 1990) The anti-predation adaptations are in the form of physical, chemical and behavioural characteristics (Esser, 1963, 1987, Esser & Sobers, 1964, Bilgrami & Jairajpuri, 1989a) Esser (1963) provided a list of resistant or susceptible prey nematodes to predation by different mononchs and dorylaims Grootaert et al {\911) measured susceptibility and resistance of tylen-chid, areolaimid, enoplid and dorylaimid nematodes to predation while Small and Grootaert (1983) descnbed their anti-predation adaptations Very recently Bilgrami and Jairajpuri (1989a) have defined resistance and
susceptibility of prey nematodes and stnke rate of the predators and proposed methods to determine and quantify them using Mononchoides longicaudatus and M fortidens as predators against a vanety of free-living, ecto- and endo-parasitic nematodes The present ex-penments were conducted to determine the degree of resistance and susceptibility of different trophic groups viz , saprophagous, ecto- and endoparasitic and pre-dacecus nematodes to predation and the strike rate of the piedators, Mononchus aquaticus, Dorylaimus stagnalis and Aquatides thomei
Materials and inethodes
The predaceous nematodes viz , Mononchus aquaticus (Mononchida), Dorylaimus stagnalis (Dorylaimina)
ISS\ 1164 SS71 92 0^ 2f>-^ 06 ' 60 <D Gauthier Vtllars - ORSTOM 265

• ^ 20 A L Bilgrami
and Aquatides thornei (^Nygoldimina) were cultured by the methods of Bilgrami and Jairajpun (1988) The adult individuals oi Acwbeloides i,p , Cephalobussp , Rhabditis sp , Panagrellus redii ivus, Tylenchorhynchus mashhoodi, Hoplolaimus indicus, Hehcotylenchus indicus, Scutello-nema sp , Hemicycliophora sp , Hemicnconemoides nian-giferae, Hirschmanniella oryzae, Paralongidorus citn, Longidorus sp , Paratnchodorus sp , Xiphinema amen-canum and the second stage juveniles of Meloidogyne incognita, Anguma tritici and Heterodera mothi were used as prey The three predators were also used as prey against each other
The ectoparasitic nematodes were isolated fresh from the soil by decantation and Baermann's funnel techniques for each experiment The second stage juveniles of M incognita, H mothi and A tntici were obtained from their populations maintained on tomato, moth grass and wheat galls respectively The free living nematodes were obtained from the culture maintained m Petn-dishes containing 1 water-agar supplemented with infant milk powder (Lactogen)
Fifty encounters were observed between predators and prey The encounters (i e , lip contact of predators with the prey at right angles) between predators and prey were observed in Pctri-dishes containing 1 ' i water agar using a stereoscopic bionocular microscope A prey nematode was placed m front of the head of an active predator with the help of a fine needle without touching (disturbing) the predators in any manner Only those observations were recorded where the predators behaved as normally as could be ensured For each encounter a new predator and prey individuals were used (irrespective whether the predator failed or succeeded in attacking prey) To reduce the effect of satiation and prey habituation, 4-6 day starved predators were tested The strike rate (SR) of the predators M aquaticus, D stagnahs and A thornei and the degree of prey resistance (PR) and susceptibility (PS) to predation were determined using methods of Bilgrami and Jairajpuri (1989a) The percentage of predators which started feeding after wounding the prey and the prey which were left unconsumed by the predators were also determined (calculated respectively out of the total number of prey individuals wounded and individuals upon which feeding actually initiated) All species of predators were tested separately
Results (Tables 1 and 2)
RiSISl ANCl AN ) SLS( 1 PIIRII [IY OH PRhY lO
PREDATION AND S I RIKE- R.\ 11- OP Af 4QI t IK I S
Among saprophagous nematodes, encounters with P redivivus, Cephalobus sp , and Acrobeloides sp , resulted in maximum strike rate (SR = 98 — 100 'o) and prey wounding (AW = 98 — 100 "() while amongst endo parasitic nematodes M aquaticus had maximum stnke rate against A tntici juveniles (SR = 98 "o) with 98 "o prey wounding M aquaticus was most successful
against T mashhoodi and H oryzae (SR = 92 — 94 'o) and wounded maximum individuals (AW = 93 — 94 S ) in comparison to other ecto-parasitic nematodes M aquaticus exhibited high degree of success upon their own individuals (SR = 88 ' ) but not on other predators viz , D stagnahs and A thornei Hoplolaimus indicus ( P R = 87 ' 0 , Scutellonema sp (PR = 91 ' 0 , D stagnalis(PR= 83 ^ ) and A thornei (PR = 88 ^ ) were most resistant prey species while T mashhoodi (PS = 93 ')), H oryzae (PS = 94 ' ^), A tritici )uv (PS = 98" i ) ,A l incognna)uv (PS = 1 0 0 " . ) , / / mothi luv (PS = 96 ' ), P redivivus (PS = 98 ), Cephalobus sp (PS = 98 " , ) , Acrobeloides sp , (PS = 100 "o) and Longidorus sp (PS = 91 "o) were highly susceptible species of prey nematodes to predation Predators which started feeding upon prey after wounding was maximum (FW = 1 0 0 '()) on P redivivus, Cephalobus sp , Acrobeloides s p , Rhabditis sp , A tntici, and M incognita luveniles Feeding after wounding (FW) H oryzae, T mashhoodi and H oryzae was 91 "o and 95 'o respectively, while that on other prey nematodes ranged between 50-89 "() M aquaticus though wounded, 80 "( of Hehcotylenchus indicushm fed upon 25 "o of individuals leaving 60 "( of them either unconsumed or partially eaten 36 "i individuals of H mangiferae and 23 ' ( of Scutellonema sp , were also left unconsumed by the predators Other prey species were left unconsumed in lesser numbers (0-16 'o) M a^wancMS took a minimum of 7-11 (9) mm and 8-15 (10) mm to consume completely an individual of M incognita and Acrobeloides 5p , respectively and maximum of 86-113 (96) min to finish D stagnahs
Ri SIS I WCh AND SLSCFPTIBILITY OF PREY TO PREDATION \NDS1RIKE RAlbOE D SM(j\AIIS
Encounters with M incognita, H mothi and A tntici juveniles yielded maximum success (SR = 82 " i , 80 "o and 82 " ) and wounding (AW = 93 "o, 80 "o and 90 "o) respectively Among ectoparasitic nematodes D stagnahs attacked T mashhoodi with maximum success iSR = 84 " ) and wounding (AW = 86 "o) Encounters with saprophagous nematodes yielded maximum success with Cephalobus sp , (SR = 80 ' o) but more P redivivus were wounded by the predators (AW = 95 " o) With others it resulted into a moderate to high rate of wounding (AW = 5-90 " o) D stagnahs failed to wound any individuals of Hoplolaimus indicus, Scutellonema 5p , H mangiferae, Hemicycliophora sp , and D stagnahs (PR = 100 "()) P redivivus, Cephalobus sp, A tntici and M incognita juveniles were highly susceptible to predation by D stagnahs (PS = 90-95 ' o) Acrobeloides sp , r mashoodi sp , H oryzae, H mothi, X america-num, Longidorus sp , Paratnchodorus sp , possessed low 10 moderate degree of resistance (PR = 10-24 'o) Rhabditis s p , and Hehcotylenchus indicus exhibited moderate to high degree of resistance (PR = 30-53 ' o) against predation D stagnahs initiated feeding upon
266 bundam appl Nematol

T a b l e 1. Strike rate of Mononchus aquaticus ( M A ) , Dorylaimiis siagnahs ( D S ) , Aquatides thomei (AT) and resistance and susceptibility of prc> to prcdation
Prt\
SMK()IH«(lls\t\ni()l)IS
P reJl I II
(tphal till' ^p
-tink kh ^p
RhibJiii sp
••( i()i'\MMii \ b \ n I as
H n.ut
H "/ .; ««(> nJh I
SiU!elhneiiij \p
H mciiki/iidt
Hemi \ h tpli WQ sp
\ jmmiihiiin
L 'ng<J ni sp
P ir
Pjralrkn J • n
PriJdtor^
\\\ DS
\l
VU
DS
AT
\[\
DS
AT
\[\
DS
AT
\ U
DS
AT
\1A
DS
AI
\!A
DS
AT
\1A
DS
AT
\!A
DS
\I
\1A
DS
Al
MA
DS
Al
\1A
DS
AT
MA
DS
AI
MA
DS
AI
MA
DS
AI
tA
iO
is
40
t^
40
42
49
h
]b
44
iQ
42
40
42
41
4' r
iQ
4b
4 r
b
40
42
44
40
42
3Q
32
34
12
]]
% li
34
32
43
i4
it)
4'
i4
i>
b
3h
30
SR
100
"6
80
100
HO
84
98
'0 "T
*i8 'H
•i4
92
M
8X
94
"4
"H
92
^2
"4
90
SO
'<4
88
80
84
'8
64
68
84
66 "2
86
68
o4
S6
68 ~i
% 68
66
90
"6 - 1
% oundins
A% PR
98
95
95
98
90
9i
100
89
94
86
0"
"6
93
S6 9
94
86
90
13
0
0
X9
4'
i2
9
0
0
11
0
0
33
0
0
84
"6
91
91
'6
ii
84
>i-
'6
91
42
44
2
5
5 1
,0
" 0
11
0
14
)3
24
-14
11
6
14
10
8"
00
100
u 5i
48
91
100
00
i9
00
100
67
00
00
16
24
19
9
24
)i
16
1
24
9
58
56
PS
98
95
95
98
90
9i
100 89
94
86
6' '6
9)
86
89
94
86
90
n [
0
89
r 52
9
0
0
41
0
0
3i
0
0
84
76
91
91
"6
0'
8
Si
'6
91
42
14
K
n
100 "S
& 100
()" 82
100
"1
S2
100
"" '\
4-1
"" oM
m 26
% — 0
0
25
1
i6
6"
0
0
-)[)
0
0
10
0
0
- 1
69
H3
'9
09
"5
s9
i2
M
XK
^ 1
M
iJing
prL\ li.*l un
I msumid
0
" 6
0
S
) 0
9
1
1
15 s
I
" " 0
12
i
h
0
1
40
i i
Sh
2i
I
0
i6
0
0
1)
0
0
i
~ 13
12 n
T»
16
26
29
9
! i
Duratiin i*
lading jpoT
MIUJIL pra
ram
10-16 M
9 15 14
8 12 10
n-19 10
14-22 19
12 10 15
x 16 h
10 r 15
10 22 14
19 20 2)
26 i4 i(i
21 24 22
H 2 1 19
22 iH 2s
s i2 it
22-ii M
i l -4 ' io
iO .D 31)
i0 51 11
11
0
35 42 3-
i'i-i6 46
it-i2 i9
20-19 i9
0
0
2129 20
0
0
20 i i 28
0
t)
28-4i i '
12 oi 12
18 19 52
*1 50 18
5016 i'
11 M 6S
-tO i9 V
* - ' 2 60
01-94 ' 2
2i i4 i l
4 40 i2
)1 49 4'
\U figures are in nearest «hok numbers Figures in paranthcsis indicate mean feeding time EA = Encounters resulted into attack, AW = Attacks resulted into pre} wounding, PS = Pre> susceptibilit> to prcdation, SR = Strike rate of predators, PR = Pre> resistance M prcdation, FVt = Feeding after wounding pre>
Vol l";, n" 3 1992 267

^ ^_ u«
A L Bilgrami
T a b l e 2 . St r ike ra te of Mononchus aquaticus ( M A ) , Dorylaimus stagnaks ( D S ) , Aquatides thomei ( A T ) a n d res is tance a n d
suscept ibi l i ty of p rey t o p r e d a t i o n
Pa\
b\D(lUMMII( \L\HIODti
•i nil
\l n null
H mil
IRhD IOR"! \h\U 0)ti
D lif;ndl
•i ill niei
1/ Jijuan
Predator LA
W lundini!
SR \Vi PR PS
\u DS
\! \ U
DS
AT
MA
DS
AT
\1A
DS
AI
MA
DS
AI
\1A
DS
AI
49
4(1
4.
-t«
41
41
48
40
42
)0
1
14
V.
8
44
11
H
% % Ql
% «2
« % SO
H4
60
10
28
04
14
16
88
i8
)6
98
90
8
10„
9
95
85
so 86
1"
0
) n
I
0
68
1
0
T
10
" 0
1
4
20
14
Si
lOO
lOi
8
100
100
C
95
100
98
90
9'
100 9i
95
96
80
86
1
k
0
12
0
0
68
"\ 0
pLidin? Durjti T t
't dmt jp P F\\ pro kft un ,,nek pa\
. nsumid mm
100
81
85
100
"4
96
90
i]
6
0
) "1
0
I
6
100
0
1)
10
6 T
-) 6
18
6
>) 0
0
0
0
0
0 1
i
8 14 1
12^8 2i
10 '6 ><
14 9
16 i2 24
P iO 21
8 15 10
l5 )0 1
P M 18
86 1 ' 96
0
0
59 "8 64
0
4 62 5^
M h 1
All figures art in nearest whole numbers Figures in partntheseis indicate mean feeding time EA = Lneeiunters resulted into attaek AW = Attaeks resulted into prey wounding PS = Prc> susceptibility to predation SR predators PR = Prey resistanee to predation FW = Feeding after wounding prey
Strike rate of
31 "u of individuals belonging to Helicotylenchus indicus but 33 "i of them were left unconsumed Maximum feeding occurred on H mothi juveniles (FW = 90 "o) and Tnchodorus sp (FW = 81 'o) D stagnahs took a minimum time of 9-15 (14) min to consume completely an individual of P redivivus and maximum of 58-72 (60) mm to finish P citn
RESISTANCE AND SLSCFFI IBII I IT OF PREY ro PRFDAT ION ^ N D ST RIKE R A T E OF A IHOK \bl
A thomei attacked all species of prey nematodes with highest strike rate (SR = 84 "o) on Rhabdms sp , among saprophagous nematodes, 82-84 "o on the second stage juveniles of endo-parasitic nematodes, M incognita, H mothi and A tntict and 88 "u on ectoparasitic nematodes, 7 mashhoodi A thomei was least successful! on D stagnahs, M aquaticus and upon its own individuals (SR = 16-36 "(i) Hoplolaimus indicus, Scutellonema sp Hemicychophora sp , D stagnahs, M aquaticus and A thomei were totally resistant to predation by A thomei (PR = 100 'o P redivivus, Cephalobus sp ,
Acrobeloides sp , H oryzae and the second stages of M incognita, H mothi and A tritici were most vulnerable species of prey (PS = 86-95 ' i) Rhabditis sp, 1 mashhoodi, Hehcotylenchus indicus, X americanum, P citn, L ongidorus sp , and Paratnchodonis sp , possessed low to moderate degree of resistance (PR = 10-50 ' o) Maximum feeding occurred on Cephalobus sp (FW == 82 '()), A tntici (FW = 77 %) and H oryzae (FW = 86 "()) among different trophic groups of prey A thomei initiated feeding only upon 36 "o individuals of Hehcotylenchus indicus but left maximum of them unconsumed (88 'n) The number of individuals of P redivivus, Cephalobus sp , Acrobeloides sp , Rhabditis sp , T mashhoodi, H oryzae, H mothi and M incognita juveniles left unconsumed were significantly low (3-8 "(, p < 0 05) However, comparatively, greater number of individuals of X americanum (13"o), Longidonis sp (22 "o) P citn and Paratnchodorus sp (13 "o), were left partially consumed A thomei required a maximum of 61-94 (72) mm to consume an individual of Paralongi-doms (itn and minimum of 8-12 (10) mm to finish P redivivus
268 hundam appl Sematol

223 Resistance and susceptibility of prey nematodes
Discussion
Wounding is an important factor in predation (Bil-grami & Jaira)pun, 1989 a) Small and Grootaert (1983) and Esser (1987) also suggested that resistance of a prey is lost if wounded or in a weakened condition A wound may also result in the loss of hydrostatic pressure of the body affecting locomotion and thereby making the prey more vulnerable to predation Wounding may also permit invasion of pathogenic micro-organisms resulting into the death of prey Thus, it is the wounding which determines the degree of resistance and susceptibility of a prey and ability of predators more precisely and hence, was considered as the primary determinant
The prey nematodes can resist predation by physical, chemical and behavioural characteristics (Esser, 1963, 1987, Esser & Sobers, 1964, Bilgrami & Jairajpun, 1989a) such as thick cuticle (Esser, 1963, Small & Grootaert, 1983), annulations in criconematids (Esser, 1963, Bilgrami & Jairajpun, 1989a), gelatinous matnx (Small & Grootaert, 1983), speed (Esser, 1963, Bilgrami & Jaira)puri, 1989a), vigorous escape response (Grootaert et al, \()11, Small & Grootaert, 1983, Bilgrami & Jairajpun, 1989a) and toxic unfavourable secretions (Esser, 1963, Bilgrami & Jairajpun, 1989a) The environmental soil factors e g , temperature, pH, moisture, soil type, chemical composition of soil, etc, also play a significant role m governing activities of predators as is evident from experiments on temperature (Wallace, 1969, Bilgrami et al, 1983), crowding (Azmi & Jairajpun, 1979), chemicals (Bilgrami & Jairajpun, 1985a), pH (Bilgrami & Jairajpun, 198"^a) and soil types (Small, 1979)
Dunng present stud> the high susceptibility of P redivivus sp , Cephalobus sp , Al incognita, A tntici and H mothi juveniles to predation by M aquaticus, D stagnalis and A thornei may be attributed to their small body size, slow rate of movement (Bilgrami et al, 1983) and lack of anti-predation devices (Small & Grootaert, 1983) The ecto-parasitic nematodes seem to be more vulnerable to predation specially by mononchs This is evident as many species of mononchs have been found containing resistant species of nematodes viz , Hoplo-laimus, Hebcotylenchus, Scutellonema, Hemicnconemoi-des, Hemicychophora, Rhabditis, Xiphinema, etc , entire in their intestine besides many other tylenchs, dorylaims, mononchs and rhabditids (Bilgrami et al, 1986) No predation on Hoplolaimus indicus, Scutellonema, H mangiferae and Hemicychophora sp , by D stagnalis and A thomei suggests resistance due to thick cuticle and annulations (physical resistance) (Bilgrami & Jairajpun, 1989 a) Moderate to high degree of resistance (PR = 24-58 "()) in Paratnchodorus sp , Longidoms sp , P citri and X americanum to predation by D stagnalis and A thomei appeared to be of physical nature D stagnalis also possessed physical resistance in the form of thick and tough cuticle
Esser (1963) suggested chemical resistance in the species of Hehcotylenchus as he observed dorylaim predators rarely attacking or succeeding in devounng the indiv duals of Hehcotylenchus spp The meagre responses of Mononchoides longicaudatus and M forti-dens towards Hehcotylenchus mdicus — live or excised — (Bilgrami & Jairajpun, 1988), their inhibited feeding and leaving greater number of wounded prey unconsumed (Bilgrami & Jairajpun, 1989a, 1990) support chemical resistance in Hehcotylenchus indicus The same has been observed dunng present investigations except in M aquaticus which wounded and killed a large population of Hehcotylenchus indicus and fed equally upon them, leaving least number of prey unconsumed This disparity is possible specially with mononchs as 1) they are capable to overcome resistance of varying types (Bilgrami & Jairajpun, 1989a), 2) their predation feeding IS aleatory (Bilgrami et al, 1986) depending exclusively on chance encounters (governed mostly by me-chanoreceptors) and 3) no evidence of chemical perception (prey secretions) is available in mononchs (Bilgrami et al, 1984) In contrast, the dorylaim (Esser, 1963, Shafqat et al, 1987), nygolaim (Bilgrami et al, 1985) and diplogastend predators (Bilgrami & Jairajpun, 1988, able to perceive prey secretions, attracted towards them and aggregated around a previously injured prey at feeding sites (Bilgrami & Jairajpun, 1989fc)
Rhabditis oxycerca, Pelodera and Plectus sp , possess characteristics such as active body undulations and vigorous escape response which provide resistance against predation (Small & Grootaert, 1983) Rhabditis sp , seems to rely pnmanly on active body undulations and vigorous escape response to resist predation by M aqua ticus, D stagnalis and A thomei as is evident by the combination of high strike rate of the three species of predators against Rhabditis sp , and comparatively low er rate of wounding and higher rate of feeding upon wounded prey individuals The abilities of predators themselves govern their efficiency as predators and also the degree to overcome prey resistance as some predators possess strong predatory potentials This is evident from the present observations on M aquaticus which showed maximum strike rate upon all the prey nematodes and almost all prey nematodes were most susceptible to M aquaticus m comparison to D stagnalis and A thomei
M aquaticus avoided predation by other predators VIZ, D stagnalis and A thomei mainly with its active body undulations (behavioural resistance) However, their high degree of susceptibility to predation by their own members conforms cannibalistic tendency in mononchs as has been reported earlier (Azmi & Jairajpun, 1979, Bilgrami & Jairajpun, 19856, Bilgrami et al, 1986) The phenomenon of cannibalism is not evident either in D stagnalis or A thomei (Shafqat et al, 1987, Bilgrarni et al, 1985)
Vol \S, n" 3 J 99 : 269

A L. Rilgrann
References
A/MI, M. I. & J\iR\jPLRi, M. S (1979) Effect of mechanical stimulation and crowding on Mylonchulus dentatus. Nema-tol medit., 7 . 203-207
BlL(,K\\ii, A L. (1990a). Diplogastend predatorb In ; Jairaj-puri, M. S., Alam, M. M & Ahmad, I. (Eds). Nematode hio-coniwl. Aspects and Prospects. Delhi, India CBS Publishers • 135-142.
Bu (..HLWii, A. L. (1990b). Predacious dorylaims. In Jairajpuri, M. S., Alam, M. M. & Ahmad, I. (Eds). Nematode bio-control, Aspects and Prospects. Delhi, India CBS Publishers : 14 3-148.
B u c k U i i , A. L., A H \ I \ 1 ) , I & J\iR.MPLRi, M. S. (1983). Some factors influencing predation by Mononchus aquaticus. Revue Ne'matol., 6 : 325-526.
Biu,R\\ i i , A. L., AiiNUD, 1. & JAIR-MPLRI, M . S . (1984). Observations on the prcdalor>' behaviour of Mononchus aquaticus Nematol. medit, 12 : 41-45.
Bnc,R.\Mi, A. L., A H \ I \ I ) , I. & J\IR. \JI ' I RI, M S. (1985).
Predator> behaviour of Aquatides thornex Nygolaimina: Nematoda). Nematohgica (1984), 30 ; 457-462.
Bn<,K.uii, A. L., A H \ I U ) , I. & ]MR.\JPL'RI, M . S . (1986). A
study of the intestinal contents of some mononchs. Revue Ne'matol., 9 : 191-194.
Bii t,R.\\ii, A. 1.. & J.»iiR\jPi RI, M. S. (1985a). Cannibalism in Mononchus aquaticus Indian J. Nematol., 14 (1984) : 202-203.
BiK.RWii, A. L. & J \iR.\li'i RI, M. S. (1985fc). The responses of Mononchus aquaticus to chemicals and pH. Indian J. Nematol, 14 (1984) : 171-174.
Biic,R.\Ml, A. L. & J\iR\|l ' lRi, M . S . (1988). Attraction of Mononchoides longicaudatus and M. fortidens (Nematoda : Diplogasterida) towards prey and factors influencing attraction. Revue Nematol., 11 : 195-202.
BucRwi i , A. L. & JuR\i. ' i Ri, M. S. (1989(j). Resistance of prey to predation and strike rate of the predators, Mononchoides longicaudatus and M fortidens (Nematoda : Diplogasterida). Revue Nematol, 12 • 45-49.
Bu c,R wii, A. L. & JMR,\JI>I- RI, M S. (1989fc). Aggregation of Mononchoides longicaudatus and M. fortidens (Nematoda : Diplogasterida) at feeding sites. Nematologica (1988), 34 : 119-121.
BiiuR-^vii, A. L. & J^iRAji'iRi, M . S . (1990). Predation by Mononchoides fortidens and M. longicaudatus (Nemadota : Diplogasterida). Nematologica, 35 : 475-488.
Esii-R, R. P. (1963). Nematode interactions in plates of non-sterile water-agar. Proc Soil Crop Sci. Fla, 23 : 121-138.
EssER, R. P. (1987). Biological control of nematodes by nematodes I. Dorylaims (Nematoda : Dorylaimida). Nematol Cir No. 4, 4 p.
EssiR. R. P. & SoBiRS, E. K. (1964). Natural enemies of nemalodiis. Proc. Soil Crop Sci. Soc. Fla., 24 : 326-353.
G R O O I A L R I , P . , J A Q L I S , A. & S M M I , R . W . (1977). Prey
selection in Butlenus sp. (Rhabditida : Diplogastendae). Meded tac. Landbouww. Rijksuniv. Gent. 42: 1559-1563.
J \ IR. \JPLRI, M . S. & BII t,R,\\ii, A. L. (1990). Predatory nematodes. In : Jairajpuri, M. S., Alam, M. D. & Alam, I. (Eds) Nematode Bio-control . Aspects and Prospects New-Delhi, CBS Publishers & Distributers Pvt. Ltd. : 95-125.
S H . M Q M , S., BILOR.\MI, A . L . & JMRAJRLRI, M . S. (1987).
Evaluation of the predation ability of Dorylaimus stagnalis Dujardin, 1845 (Nematoda : Dorylaimida). Revue Nematol, 10 .455-461.
S\i^i 1, R W . (1979). The effect of predator}' nematodes on population of plant parasitic nematodes in pots. Nematologica., 25 • 94-103.
S.vniL, R. W. (1987). A review of the prey of predatory soil nematodes. Pedobiologia, 30 : 179-206.
S U M I , R . W . & G R O O I M-RI, P. (1983). Observations on the predation abilities of some soil dwelling predatory nematodes. Nematologica, 29 : 109-118.
WAI 1 \( 1, H . R . (1969). Wave formation of infective larvae of tne plant parasitic nematode, Meloidogyne javanica Nematologica, 15 : 65-75.
270 Fundam. appl. Nematol

r: ^ ^
A Z 0
Recent Advances in Nematology, 1992:203-218
BIO-CONTROL AGENTS AND THEIR ROLE IN NEMATODE MANAGEMENT
Anwar L. BUgrami
Section of Nematology, Department of Zoology, Aligarh Muslim University, Aligarh-202002, India.
Nematode control measures may either be corrective (eg. chemical) or preventive (cultural) but the biological control has both these possibilities. It may be preventive in the sense that it can help in evading the disease but if the disease has already set in the malady can be corrected by reducing the population density of pests. Biological control is a means of promoting organisms that are harmless to plants but attack and destroy organisms antagonistic to them. The biological control is an ecological and biological problem which calls for thorough understanding of the ecosystem. It is determined by predator-prey relationships and other such associations. Such relationships or association in the ecosystem which are deterimental to pest population can be helpful in nematode management programmes. These associations are important part of energy flow of the biological world. If somehow the energy flow in the ecosystem is manipulated without disturbing the ecological balance the aim of biological control can be achieved. Though, the advocacy of biological control in nematology dates back to six to seven decades but awareness about it and its potentials were brought into sharp focus only in recent times. The biological control has achieved some success in entomology, plant pathology and insect nematology but it has remained by and large a theoretical approach in plant nematology at least for the time being. This should not sound discouraging because the biological control programme unlike chemical and physical control methods is a long time strategy. Nematologists are lucky in the sense that they have numerous kinds of organisms antagonistic to plant parasitic nematodes at their disposal which can be explored for their use in nematode bio-control.
Many possibilities are there and some more will be discovered but how well these could be exploited would largely depend on the anthusiasm and the patience of the future nematolgists and the amount and time that they devote on this aspect of control. The antagonistic organisms may be predaceous or parasitic in nature. Some of them have exhibited greater predatory potential, efficient characteristics and possible future role in nematode management while others still await exploration and experimentation.
Predaceous nematodes (Bilgrami, 1990a,b; Bilgrami and Jairajpuri, 1990); parasitic and predaceous fungi (Jatala, 1986;) predaceous and parasitic protozoans (Weber etal., 1952;) viruses (Zuckermanefa/., 1973); rickettsias (Walsh, 1979, Kerry, 1986); bacteria (Sayre, 1971, Mankau, 1981); tardigrades (Sayre and Powers, 1966); •earthworms (Yeates, 1969); collembolane (Murphy and Doncaster, 1957) enchytraeids (Sayre, 1971, Boosalis and Mankau, 1985); mites (Rodriguezesa/., 1962) Muraoka and Ishibishi, 1970, enemy and trap crops (Brown, 1978;Gommers, 1981)

\.j
204 Recent Advances in Nematology
microbial decomposition products (Sayre, 1971); neem (Siddiqi and Alam, 1988) are the possible nematode bio-control agents. The effectiveness and success of these organisms depend upon some desired specific characteristics viz., (1) easy culture conditions; (2) high reproductive potential; (3) longevity; (4) easy applicability; (5) safety to non target organisms; (6)searchingcapabilities; (7) ecological compatability; (8) temporal compatability; (9) environmental adaptability; (10) persistance capability; (11) dispersal capacity; (12) control potential; (13) biologically competitive and (14) broad spectrum efficiency. The aim of this chapter is to focus attention on various agents of nematode bio-control, their potentials, constraints and future role in nematode management. It is therefore the discussion is mostly confined to these aspects ony rather than to discuss various other aspects which are very well documented in recent reviews by (Alam etal., 1990a .) (Ahmad, 1990a, 1990b); Ahmad (1990); Bilgrami (1990a;1990b) and Jairajpuri and Bilgrami, (1990).
Biological control potentials of difTerent agents
The biological control potential of different agents vary from species to species and individuals to individuals. However, the one most deterimental to the population of plant parasitic nematodes can be a good agent of nematode bio-control.
1. Nematode destroying fungi
Fungi is one on the most studied group. Since the first inception of a nematode trapping fungi nematode (Zopf, 1888) more than 200 such species have been reported (Jatala, 1986; Rodriguez Kabana, et. al., 1987; Mankau, 1980a,b). The nematode destroying fungi (predaceous, parasitic or opportunistic) are widespread in soil and associated with nematode bio-control. A few namely Dactylella oviparasitica, Nematopthera gynophiia, Verticillium chlamydosporium, Arthrodtrys robustus (Stirling and Mankau, 1977, Kerry, 1982, Cayrol et al., 1978, Cayrol and Frankowaski, 1979) have been reported suitable for practical utilization. Handling, isolation, culturingand inoculation besides their obligate nature may at times make their utilization a little cautious organic soil amendments; soil treated with a particular fungi, both treatment given together or fungi concomittantly with other agent have significantly elevated theirbiological control potential of fungi and overcome greatly some of theconstraints.
The predaceous fungi belong mostly to Hypomycelidae; Deuteromycetes; Zygomycetes and few other groups, sticky traps (adhesive branches, adhesive network, adhesive knobs); and mechanical ring traps (constricting and non constricting ring) are basic mechanisms involved in their predation. The adhesive branches are few cells of the fungal mycelium (e. g., Monacrosporiunt cionopagum while adhesive knobs are produced on a single or double celled lateral branches (e. g. Dactylaria Candida). The constricting rings (e.g. Arthrobotrys Dactyloides, Dactylella bembicodes) caught nematode in the ring by bulging the inner walls of the lumen. The non-constricting ring (e.g. D. Candida) traps nematodes only when the later tries to escape. Their ability to produce endigenous nemin make them potential bio-control agents of plant parasitic nematodes. Commercial preparations {A. robustus, marked as strain 300 Royal and Royal 350) antagonised Ditylenchusdipsaci on mushroom and reduced greatly the root galling on tomato.
Endozoic fungi : It was suggested the role of Harposporium anguillulae in

'^?7 Anwar L. Bilgrami 205
regulating nematode populations. Since then more than 50 species of endozoic fungi have been described from Blastocladiales, Chytridiales, Legenidiales, Entomopthorales, Zoopagales and Monilials. However, only few have shown considered potentials. Acrostalqmus. H. nemathoptera and Haptoalossa are some common genera which can be cultured and remain dormant in soil for long periods (upto five years) without nematodes (Kerry, 1984). These endozoic fungi produce adhesive spores which upon contacting a nematode quickly and firmly attached to its cuticle. It then germinates and enter the nematode body through germ tube. Fungi forming adhesive spores appear better than those whose spores to be ingested by nematodes because it is self limiting phenomenon and depends upon the size of the spear lumen which in most plant parasitic nematodes is too narrow to ingest these spores. The infection of endozoic fungi has been observed on H. Schachtii (Khun, 1877); H. avenue and G. rostochiensis (Kerry, 1975; Kerry et al., 1976); H. carolae, H. cruciferae, H. gottengiana, H. schachtii, H. trifoli (Kerry and Crump, 1977) besides the natural control of H. avenae by Nematopthera qvnophilus (Kerry, 1974; Kerry and Crump, 1977) suggested more than a causal role of these organisms in reducing nematode populations.
Opportunistic fungi : These fungi rapidly colonize nematode reproductive structures and destroy females, cysts and eggs belonging to Globoder a. Heterodera and Meloidogyne. The opportunistic fungi which can survive host free periods and can be grown axenically belong mostly to the gener a Cylidrocopon. Exophiala, Fusarium, Galiocladium, Paceilomyces, Phoma and Verticillium etc. Paceilomyces lilacinus, The subject of many investigations, has been produced on commercial scale under trade name of Biocon (Jatala, 1986). V. chlamydosopnim is another excellent example parasitizing Meloidogyne arenaria (Morgan et al., 1981) and K glycines (Gintis et ai, 1983).
Constraints: The future role of predaceous fungi in nematode bio- control appears bright but to achieve this and other significant success hurdles and constraints in their handling, culturing, inoculation etc., are to be removed. The lack of culture techniques preventing mass inoculation, their slow colonizing abilities and non specific predation are other limitations. Moreover their limited predaceous activity msy also pose great problems in their exploitation as successful bio-control agent. There appears some of the obvious reasons that produced irraiic results on nematodes. Trap forming fungi depends upon nutritive values of the soil making nematodes as an alternate food, carbohydrate, nitrogenous compounds and microbial decomposition products are always required to produce and increase the production of traps (Linford, 1937). Means have to develop to manipulate soil and environmental factors such as pH, moisture, temperature, soil type, soil constituents, soil composition etc. which greatly influence fungi and their trap formation. (Johnson, 1962, Mankau, 1968). Lastly, it is the isolation and identification that often pose serious hurdles and offer their restricted utilization.
Future prospects : No doubt all types of fungi posses greater potentials of nematode bio-control agents but those species which rapidly colonize eggs and cysts and those which parasitize the reproductive organs provide effective role in future control strategies if not alone then concomitantly with other bio- control agent under integrated pest management.

2 2 8
206 Recent Advances in Nematology
PREDACEOUS NEMATODES
The predaceous nematodes are of different kinds. They may be classified into three different types depending upon their feeding apparatus and mode of feeding. One with strong and well developed buccal cavity, with Jooth teeth and denticles and efficient prey catching and feeding mechanisms belong to the order Manonchida. Their feeding apparatus is cutting, sucking and engulfing type {t.g.,Mononchus. Mylonchulus lotonchus ) Bilgrami et ai, 1984, Jairajpuri and Bilgrami, 1990). The second group of predators are those which are commonly known as stylet bearing predators and belong to suborders Dorylaimina, Nygolaimina and Aphelenchina. The feeding apparatus of dorylaim predators, known as stylet or odontostyle, is always axial in position and provided with a lumen. It is called as mural tooth in Nygolaim predators where it is solid and without any lumen. Ingestion occurs throught the oral aperture. Aphelenchid predators C. seinura are provided with spear a pointed needle like feeding apparatus with definit lumen for ingestion. Members belonging to the order Diplogsterida also posess a well develope and strong bucal cavity with dorsal movable tooth, teeth etc. It is cutting and sucking type.
BIOLOGICAL CONTROL POTENTIAL OF MONONCHS
The interest in using mononchs for controlling plant parasitic nematodes was initiated since Cobb (1917; 1920) and Steiner and Heinley (1922) regarded C.papillatus as a promising species to work. Until 1974 not much work was done on predatroy nematodes when Cohn and Mordechai (1974)) found correlation between high population levels of Mylonchulus sigmaturus and low populations of T. semipenetrans in pot experiments. Small (1979) found a significant reductioon in G. rostochiensis and M incognita in presence of Prionchulus punctatus in pots. Ahmad and Jairajpuri, (1982) from a field data found a significant correlation between populations of Parahadronchus shakili and Trichodorus sp., Hemkriconemocides sp. Azmi (1983) indicated an increase in the population of lotonchus monohystera and reduction in Helicotylenchus dihystera. Factors influencing predation (Bilgrami et al., (1983), predation (Small and Grootaert, 1983; Bilgrami etai, 1984), food choice (Bilgrami et al., 1986), strike rate of predators and resistance and susceptibility of prey to predation cannibalism (Bilgrami and Jairajpuri, 1985a), effect of chemicals on their survival (Bilgrami and Jairajpuri, 1985b), and prey range etc (Small, 1979) are other recent efforts which have been made to establish predator prey relationships of mononchs.
Biological control potentials of Dorylaim and Nygolaim predators
Though, Dorylaim and Nygolaim predators possess predatory potentials unequal to Mononchus but their role in biological control of plant parasitic nematodes seems to be more promising. Their wide spread and abundant pre-sence could indicate their biological control activites. However, it is not known upto what extent and under what conditions phytoparasitic nematodes are reduced since such an evaluation has never been made. The Dorylaims and Nygolaims are one of the most ubiquitous group of nematodes occuring in all types of soil, climates and habitats. The presence of 2,3 or more genera of the^e predators at one place/fleld is quite common. Their abundant and naturally occuring populations have advantages over other predators. They need only slight modifications under field conditions to establish their populations' at desired levels. The predators can also be maintained on algae and fungi (HoUis, 1957),

229 Anwar L. Bilgrami 207
Ferris, 1968; Wood, 1973, Bilgrami, 1990b) besides nematodes (Bilgrami et al., 1985; Shafqat et al., 1987). The detection ooi Eudorylaimus obtusicaudatus feeding on the eggs inside the//, schachti cysts or the increase in the population of predators (Thomia) in the presence of citrus nematodes and decease in absence of them in pot trails (Boosalis and Mankau, 1965) indicates potential rather than their casual role in nematode bio-control. The recent observations on A. nivalis and D. stagnalis also favours their possible future role in namatode management.
Biological control potential of deplogasterld predators
Studies made on diplogasterid predators indicate that they possess useful characteristics of and efficient bio-control agents. Observations on their ability to percieve prey attractants (Bilgrami and Jairajpuri, 1988), pre- and post- feeding aggregation and attraction (Bilgrami and Jairajpuri, 1989a) strike rate of the predators and resistance and suceptibility of prey to predation small and Grootaert, 1983; Bilgrami and Jairajpuri, 1989b), prey catching and feedings mechanisms, prey preference etc., (Bilgrami and Jairjapuri, 1990) and factors governing their predation (Yeates, 1969; Bilgrami and Jairjapuri, 1989c) have revealed many interesting and promising features for the first time. Their short life cycle, high fecundity and bacteriophagus nature (Bilgrami, 1990a) besides easy culture conditions are some of the other advantages with these predators. Organic substances (leaves, cow dung, manure and other nutrients) enhance their population and helps in culturing them Webster, 1972; Bilgrami, 1990a). Being polyphagus they can be maintained using bacteria without nematodes (Bilgrami and Jairajpuri, 1990) and thus interspecific competition may be avoided.
Constrainst
The biological control of plant parasitic nematodes using predaceous nematodes may by more challenging than of any other pest as they inhibit soil and attack mostly underground parts of the plant. This could be one of the reason that our eyes go unnoticed about what these minute organisms are doing under soil conditions. Still means are lacking for better culture conditions for large inoculums. The limited knowledge on their taxonomy, classification, indentification, behaviour, predation, biology, ecology, longevity, predator-prey relationships etc., are some of the constrainsts preventing their better understanding and practical utilization.
Future prospects
With very little informations available on various aspects of nematode predation, their role in nematode management can be thought only in positive and hopeful perspective. The future prospects of predaceous nematodes depends greatly on what advances have been made on other control methods, their effectiveness and the resources provided to establish research programmes. Naturally occuring huge populations of Dorylaim and Nygolaim predators and significant role played by them in decressing population of parasitic nematodes under natural conditions exhibit their tremendous potentials and indicate their possible future role in nematode bio-control.
Microbial parasites and predators
The various kjnds of microbial parasites of plant parasitic nematodes are mainly the viriKes, rickettsias, bacteria, protozoans etc.

2 30 208 Recent Advances in Nematology
Biological control potentials of viruses
The abnormal and putative changes in the cuticle of nematodes were attributed to various surface of the cuticle, hypodermis, muscles and the digestive and reproductive systems have been observed to be affected by the viruses. Lowenberg et al. (1959) have reported a viral disease of M incognita, which resulted in the reduced activity and the loss of their gall forming ability, subsequent workers Zuckerman et al, 1973; Ibrahim and HoUis, 1973; Ibrahim et al., 1973) also reported virus infections in plant parasitic nematodes. Dolichodorus meterocephalus and T. martini
Biological control potential of r/c/retts/a
The potential of rickettsia like organisms lie in their ability to parasitize nematodes and to be transmitted from one generation to the next via the oocytes (Walsh, 1979). The first rickettsia infection was observed in two species of cyst nematode G. rostochiensis and H. gottengiana (Shepherd et al., 1973) and later in the second stage juvenile of H. glycines (Endo, 1979; Nfoel and Wilson 1981). These observations gave credence to the idea of their being parasites of plant parasitic nematodes rather than the symbionts.
Bacteria
Pasturia penetrans an obligate parasite of M incognita, M. arenaria, M.javanica, Pratylenchus sp., P. scribneri, P. penetrans. Helicotylenchus sp., Scutellonema sp., Heterodera sp., H. elabista, Tylenchorhynchus martini has emerged as a possible bio-controi agent (Rodriguez ef A/., 1965; Nishizawa, 1987; Mankau and Prasad, 1977; Stirling, 1985; Bird and Brisbane, 1988; Page and Bridge, 1985). Sayre (1971) and Mankau (1981), however, expressed reservations against their parasitic nature. P. penetrans parasitzes by developing germ tubes which give rise to vegetative hyphae. Proliferating colonies of these undergo periodic lysis of the interaciliary cells allowing daughter colonies to establish at points further away from the site of infection (Sayre and Wergin, 1977). Terminal cells of the fragmented colonies then differentiate to form ovate structures and giving rise to spores which are highly resistant to dissication (Sayre, 1980). The number of spores produced in an infected female is estimated to be over two million (Mankau, 1975). The specificity of spores in adhering to different nematodes (SpauU, 1984); possible exsistance of new bacterial species (Sayre and Golden, 1987), antagonism and lysing nematodes (Katznelson et al., 1964); effectiveness of producing toxic metabolites (Johnston^ 1959) and effectiveness in controlling species of Meloidogyne sp. (Page and Bridge, 1985) are some of characteristics of P. penetrans whicii establishes their predatory potentials at much higher levels than other organisms.
Protozoans
The protozoans may be parasitic or predaceous. The earlier observations or predaceous amoeba {Theratromixa webri) feeding on G. rostochiensis was of Weber et al., (1952). T. webri feed by forming digestive cysts and entering into resting stages under adverse conditions. The parasitic protozoans parasitizes either by forming resistant oocytes (Agerella helminthorum) or by aggregating around the excretpry pore and vulval opening and entering into the body of the nematodes (Flagellates). '
Flagellates specially Bodo caudatum which produces resistant oocytes and infect

o Anwar L. Bilgrami 209
nematode through the body openings particularly the excretory pore and vulva and T. webri which forms digestive cysts to feed upon nematode and undergo resting stages under adverse conditions do hold some promise of an efficient agent of nematode bio-control.
CONSTRAINTS
Not enough effort has been expanded to search microorganisms to be used in biological control programmes. The paucity of knowledge about their occurrence, taxonomy, biology, ecology, behaviour, predator-prey relationship, range of prey etc., particularly of microorganisms has so far restricted the expansion of efforts in establishing the true nature of these organisms. One of the major problems with virus, bacteria, protozoans is our inability to culture them in vitro and in vivo for large inoculums. Besides some of their self limiting behavioural patterns, susceptibility to changing environmental conditions are to be studied in depth for their better understanding and utilization. Means and ways are to be developed to study various factors governing their behaviour, population build up, population stabilization etc., for their effective utilization in nematode bio control.
Future prospects
Some of the microbial agents do posses future prospects to be used in nematode management while many others appear to exhibit potentials and prove credentials as and when their abilities are explored. However, at present P. penetrans is one such microorganism which posses greater potentials. Their effectiveness lines in their antagonism in lysing nematodes by producing toxic metabolites, occurrence under different climatic conditions, forming resistant spores under adverse climatic conditions, having broad range of prey and degradation as bacterial products. The protozoans specially T. webri and B. caudatum does hold some promise and can be considered for future studies.
Biological control potentials of predaceous soil in vertebrates
In the complex food chains that exist in the soil environment nematodes may serve as prey to predaceous nematodes, protozoans or act as host to a variety of prokaryotic parasites. A number of soil invertebrates are known to antagonistic plant parasitic nematodes which could, be useful in reducing nematode populations. Though there is a dirth of knowledge about various aspects of predation by the invertebrate organisms, nevertheless they posses many attributes of an efficient bio-control agent.
Tardigrades
Tardigrades commonly known as the water bear always made distinct tear in cuticle and remained firmly attached to prey during feeding. Macrobiotus and Hypsibius have been observed feeding on Trichodorus sp., Tyknchus sp., M. incognita and P. rediviyus.
Turbeliarians
No ellucidation of the effects of turbeliarians on the population of plant parasitic nematodes has been made which could lead to speculate their role as nematode bio-control agents. However, Adenoplia sp. Feeding on M incognita and P. redivivus is perhaps one such observtion (Sayre and Wergin, 1979) which have indicated reduction

2 3 2 210 Recent Advances in Nematology
of galling on tomato roots.
Enchytraeids
A very few species viz., Fridericia and Enchytraeus have been observed feeding on H. schachti and controlled their populations (Schaerffenberg and Tendl, 1951).
Collemboians
Collembolans are active predators, found in large numbers on organic matters around roots (Sayre, 1971). Brown (1954) and Murphy and Doncaster (1957) found Onychiurus armatus to be the most active Collembolan on H. cruciferae. They found 30% cysts perforated and body contents consumed by O. armatus
Mites
The feeding of mites on nematodes was first observed by Linford and Oliviera (1938). Rodriguez et al., (1962), cultured Macrocheles sp., on Rhabditis sp., and found more than 30 species of miter devouring nematodes whole out of the 41 species which were identified as predators (Muraoka and Ishibishi, 1976). Lasioseius sculpatus (Imbriani and Mankau, 1983) and Tyrophaqus putrescentiae fed voracioulsy on plant and soil nematodes.
Earthworms
With a constant passage of soil through the gut of earthworms, it may be expected that a sizeable number of plant and soil nematodes will! be consumed. Though too few studies on earthworms permit for any sound conclusion on their role in suppressing nematode populations but 53.7% and 37-67% reduction in different types of soil (Dash etal., 1980; Yeates, 1981) suggest their potential as predators of namatode.
Constraints
It is difficult to pinpoint constrains! in practical utilization of these organisms as the knowledge about various aspects of their predation is still limited. However, the inability of turbellarians to move in smaller pore spaces where the nematode move may limit their role as biological control agents. Similarly, the saprophagus nature of enchytraeids and devouring endoparasitic nematodes only under particular conditions may exert little impact on nematode populations. Mites and earthworms are such organisms which posses potential, need to be explored for better culture conditions, understanding of various aspects of predation and more importantly predator-prey relationships.
Future prospects
The future use of the soil invertebrates vi2., mites and earthworms appears to be eminent because of their behavioural characteristics and potentials they posses. To utilize these organisms in nematode control efforts are required to be expanded in all directions to identify various aspects possesing potential, better culturing methods and effects of physical, chemical and environmental factors on predation.
Biological control potentials of enemy plants
Antagonistic plants also serve as useful tool for nematode bio control. Many of these plants have good nematode control potential. There is an age old practice to

Anwar L. Bilgrami 211 •^ 'J
interplant mustard, marigold, neem etc. with wheat and barley. A large number of plants are known to suppress nematode populations (Gommers, 1981).
Tagetus
Tagetus is one such crop which has effectively reduced P. penetrans on narcissus and reduced root rot disease. Different varieties of T. patula and T. erecta reduced population of leason nematode upto 90% besides Tylenchorhynchus sp. and P. erenatus. The antagonistic action of these plants is due to their root diffusates containing high concentrations of alpha terthenyle together with biogenetically related bithenyle.
Mustard
Mustard (Brassica hirta; B. nigra and B. compestris) has inhibiting effects on G. rostochiensis; T. brassicae; Hoplolaimus indicus, M. incognita, R. rkniformis, Helicotylenchus indicus, T. filiformis and T. mirzii (Alam et. ai, 1977). The toxicity of mustard species is considered to be due to the presence of isithiocynate in their root system. Besides having antagonistic values mustard is also an important oil crop.
Asparagus
The members of the family Liliacae specially i. e., Asparagus {A. officinaliis) which contain nematicidal glycosides with systemic nematicidal action to reduce activity of Paratrichodorus minor (Rohde and Jenkins, 1958); Trichodorus christei (Rohde, 1960) completely inhibited emergence of G. rostochiensis and H. glycines juveniles form the cyst (Takagusi et al, 1975) and produced toxic effect on M hapla. P. penetrans aa6 P. curvitatus (Takagusi er A/., 1975).
Grasses
Studies justify antagonism of meadow grasses {Poa pratensis and P. trivalis) (Family : Graminae) which have resulted in the reduction of cyst contents of G. rostochiensis.
Crotalaria
Crotalaria spectabilis and C. junces are known to suppress plant parasitic nematodes, M incognita, Hoplolaimus indicus, T. mirzii, R. reniformis, T. brassicae, Helicotylenchus indicus, T. filiformis etc., (Mcbeth and Taylor, 1944; Alam etai, 1979) Monerotatum a pyrrolizidine ester is believed to be the chief toxic factor (Fassuliotis and Skucas, 1%9).
In rural India neem trees are allowed to grow in fields near residential areas mainly because it purifies air and possess medicinal and insecticidal properties. Various parts of neem and its seed cake have suppressed the multiplication of M. incognita. R. reniformis and T. brassicae (Alam et ai, 1977) Siddiqui and Saxena, 1987a,b). The commonly occurring chemicals in neem eg., azardiractin, kaemfor, nimbin, nimbirdin, nimbidic acid, querctin and thionemone are believed to be the toxic factors.
Cor)straints
Non-acceptability of many of the antagonistic plants by the formers on account of farm economics is one the the major constraint in their utilization as nematode bio-control agent.

^ 1 d ^ 0 " 212 Recent Advances in Nemalology
Future prospects
Recent studies have high lighted the antagonistic nature of many plants e. g., mustard, neem and tagetes. The future use of these plants appear to be more bright mainly because of their nematicidal properties. Some of them are important oil crops and posses economic values. So the suspections of the farmer can also be removed and their farm economics improved by intercropping and using by products of these plants for nematode management.
Conc/us/ons
Biological control always enjoys advantages over other conventional methods like physical and chemical control because of its specific, non pollutant and inexpensive nature. The use of chemicals and nematicides in controlling plant parasitic nematodes have yielded positive results, but their use is now been discouraged because of their exhorbitantly high costs, toxicity to other beneficial soil fauna, residual effects on human beings and live stocks and because of the fear that the continuous use of these chemicals may result in the nematodes to become resistant.
In view of the success achieved by entomologists, plant pathologists and insect nematologists in controlling pest population through their natural enemies, the plant nematologists must also consider possibilities of using organisms to control plant parasitic nematodes in fields. It may sound discouraging that at present nematode management by natural enemies is largely theoretical but some classical exceptions indicate their tremendous potentials and bright future in nematology too. The success of biological control depends upon exploration of various aspects viz., knowledge about their prey predation, reproduction, life cycle, culture conditions, ecology, behaviour, longevity etc. The correct indentification and classification of these organisms as well as sets the term of biological control. It is therefore required to carry out future studies on these organisms extensively with strong interaction, coordination determination and by involving more people to work out their practical feasibiliy in controlling plant parasitic nematodes.
The author is greatful to Ms Tabassum Mirza without whose help this work would have never completed.
REFERENCES
Ahmad, I., 1990a. Microbial parasites and predators. Nematode Bio-control: Aspects and Prospects, (eds. Jairajpuri elal,}'. CBS Pub. and dist. Pvt. Ltd. New Delhi pp. 83-94.
Ahmad, i., 1990b. Predaceous soil invertebrates. In Nematode Bio-control: Aspects and Prospects. (Eds. Jairajpuri et al,) CBS Pub. and Dist. Pvt. Ltd. New Delhi pp. 149-152.
Ahmad, N. and Jairajpuri, M.S., 1982. Population fluotuationsof predatory nematode, Parahadronchux shakili (Jairajpuri; 1969). (Mononchida). Proc. Symp. Ecol. Anim. Popul. India Pt.,3: 1-12.
Ahmad, W, 1990. Predatory mononchida. in Nematode Bio- control:Aspects and Prospects. (Eds. Jairajpuri ei al.,). CBS Pub. and Dist. Pvt. Ltd. New Delhi pp.

Anwar L. Bilgrami 2S'5 " - ' - ' - '
127-134.
Alam, M. M., 1990. Nematode destroying fungi, In Nematode Bio-control:Aspccts and Prospects. (Eds.Jairajpuri et al.,). CBS Pub. and Dist. Pvt. Ltd. New Delhi pp. 57-69.
\lam, M. M. and Jairajpuri, M. S., 1990. Natural enemis of nematodes. In Nematode Bio-control: Aspects and Prospects. (Eds. Jairajpuri et al.,). CBS Pub. and Dist. Pvt. Ltd. New Delhi pp. 17-40.
^jam,M. M.; Saxena.S.K. and Khan, A. M., 1977. Influence of interculture of marigold and margsa with some vegetable crops on plant growth and nematode population. Acta Bot. Indica, 5 : 33-39.
.Mam, M. M.; Saxena, S. K. and Khan, A. M., 1979. Suitability of crops to the reniform, stunt and root-knot nematodes. In Recent Researches in Plant Science (Ed. S. S. Bir), Kalyani Pub. New Delhi pp. 630-634.
Alam, M. M.; Siddiqui, M.A. and Ahmad, A., 1990. Antagonistic plants. In NematodeBio-control: Aspects and Prospects. (Jairajpuri et al.,) CBS Pub. and Dist. Pvt. Ltd. New Delhi pp. 41-50.
Azmi, M. I., 1983. Predatory behaviour of nematodes. Biological control of Helicotylenchus dihystera through the predaceous nematode, lotonchus monogystera. Indian, J. Nemalol., 13 : 1-8.
Azmi, M. I. and Jairajpuri, M. S., 1979. Effect of mechanical stimulation and crowding on Mylonochulus dentaus. Nematol. Medit., 7 : 203-207.
Bilgrami, A. L., 1990a. Diplogasterid predators: In NematodeBio-control: Aspects and Prospects. (Eds. Jairajpuri et al.,) CBS Pub. and Dist. Pvt. Ltd. New Delhi pp. 135-142.
Bilgrami, A. L, 1990b. Predaceous Dorylaims. In nematode Bio-control; Aspects and Prospects. (Eds. Jairajpuri et al.,). CBS Pub. and Dist. Pvt. Ltd. New Delhi pp. 143-148.
Bilgrami, A. L; Ahmad, I. and Jairajpuri, M. S., 1983. Some factors influencing predation by Mononchus aquatkus. Revue Nematol., 6 325-326.
Bilgrami, A.L.; Ahmad, I. and Jairajpuri M. S., 1984. Observations on the predatory bc\i&\\o\xr oiMononchus aquaticus. Nematol. Medit., 12:41-45.
Bilgrami, A. L.; Ahmad, I. and Jairajpuri M. S., 1985. Predatory behaviour of Aquatides thornel (Nygolaimina:Nematoda). Neniatologica, 30:457-462.
Bilgrami, A. L; Ahmad, I. and Jairajpuri, M. S., 1986. A study of the intestinal contents of some mononchs. Revue Nematol., 9:191-194.
Bilgrami, A. L; Ahmad, I. and Jairajpuri M. S., 1988. Interaction between Mononchus in non-sterile agar plates. Indian J. Nematol., 17: 339-340
Bilgrami, A. L. and Jairajpuri, M. S., 1985a. Cannibalism in Mononchus aquaticus Indian/ Nematol., 14 : 1984,202- 203.

214 Recent Advances in Nematology
Bilgrami, A. L. and Jairajpuri M. S., 1985b. The responses of Mononchus aquaticus to chemicals and pH. Indian]. NematoL, 14 : 1984,171-174.
Bilgrami, A. L. and Jairajpuri, M. S., 1988. Attraction of Mononchoides, iongicaudatus and M. fortidens (Nematoda : Dipligasterida) towards prey and factors
influencing attraction. i?evwe A emflto/., 11: 195-202.
Bilgrami A. L. and Jairajpuri, M. S., 1989a. Aggregation of Mononchoiam iongicaudatus and M. fortidens (Nematoda: Diplogasterida) at feeding s.i^B Nematologica, 1988,34 :119-121.
Bilgrami, A. L. and Jairajpuri, M. S., 1989b. Resistance of prey to predation and strike rate of the predators, Mononchoides Iongicaudatus and M. fortidens (Nematoda:Diplogasterida). Revue Nematol, 12 : 45-49.
Bird, A. F. and Brisbane, P. G., 1988. The influence of Pasteuriapenetrans in field soils on the reproduction of root-icnot nematodes. Revue NematoL, 11: 75-81.
Boosalis, M. G. Mankau, R., 1965. Parasitism and predation of soil microorganisms. In Baker, K. F. and Synder, W. C. (Eds.). Ecology of soil borne pathogens. Univ. Calif. Press. Berkeley, pp. 374-391.
Brown, E. B., 1978. Cultural and biological control methods. In "Plant Nematology". (Ed. J. F. Southey), GD, Min. Agr. ^ish. Food, HMSO, London, 269-282.
Brown, W. L., 1954. CoUembola feeding on nematodes. Ecology, 35:421.
Cayrol, J. C. and Frankowski, J. P., 1979. Une methode de lutte biologique contre les nematodes a galles des racines appartenant au genre Meloidogyne. Pepinieristes, Horticulteurs, Maraichers,/?evue//ort/co/e, 193:15-23.
Cayrol, J. C; Frankowski, J. P.; Laniece, A.; D'Hardemare, G. and Talon, J. P., 1978. Contre les nematodes en champignoniere. Mise au point d'une methode de lutte biologique a I'aided'un Hyphomycete predateura; Arthrobotrys Mraichers./?ev«e Horticole, 184: 23-30.
Cobb, N. A, 1917. The Mononchus {Mononchus, Bastian) a genus of free-living nematodes. Soil ScL, 3: 431- 486.
Cobb, N. A, 1920. Transfer of nematodes (ipononchs) from place to place for economic purposes. Science, 51: 640-641.
Dash, M. G.; Senapati, B. K. and Mishra, C. C, 1980. Nematode feeding by tropical earthworms. Oikos, 34:322-325.
Endo. B. Y., 1979. The ultrastructure and distribution of an intracellular bacterium-like microorganism in tissues of larvae of the soybean cyst nematode Heterodera glycines. J. Ultrastruc. Res., 67 :1-14.
Fa.ssulliotis, G. and Skucas, G. P., 1969. The effect of pyrralizidine alkaloid ester and plants containing pyrrolizidine on Meloidogyne incognita acrita. J. Nematol., 1 : 287-288.
Ferris, V. R., 1968. Biometric analysis in the genus Labroneam (Nematoda:Dorylaimida) with a description of L. thornei n. sp. Nematologica, 14

Anwar L. Bilgrami 215
: 276-284. "^ '"'
Gintis, B. O.; Morgan-Jones, G. and Rodriguez-Kabana, R., 1983. Fungi associated with several developmental stages o{Heteroderaglycines form an Albania sobean field soil. Nematropica, 13:181-200.
Gomers, F. J., 1981. Biochemical interactions between nematodes and plants and their relevance to control. Helm. Abstr., 50 : 9-24.
Mollis, J. P., 1957. Cultural studies with Trichodorus christei on tomato roots / NematoL, 5 : 228- 289.
Ibrahim, I. K. A. and Hollis, J. P., 1973. Electron microscopic studies on the cuticle of swarming and non swarming Tylenchorhynchus martini. J. NematoL, 5:275-281.
Ibrahim, I. K. A.; Joshi, M. M. and Hollis, J. P., 1973. The swarming virus disease of Ty/enc/ior/iync/iu.ymflrfin/.Int. Cong. P/flnf/'fl/Zio/. Minneapolis, Minn. Abstr. 555.
Imbriani,J. L. and Mankau, R., 1983. Studies on Lasioseius sculpatussi neostigmated mite predaceous on nematodes. J. NematoL, 15 : 523-528.
Imbriani, J. L. and Bilgrami, A. L., 1990. Predatory nematodes. In NematodeBio-control; Aspects and Prospects. (Eds. Jairajpuri etal.,). CBS Pub. andDist. Pvt. Ltd. New Delhi pp. 95-125.
Jatala, P., 1986. Biological control of plant-parasitic nematodes. Ann. Rev. PhytopathoL, 24:43-489.
Johnson, L. F., 1962. Effect of addition of organic amendments to soil on root-knot of tomatoes. II. Relation of soil temperature, moisture and pH. Phytopathology, 52 : 410-413.
Johnston, T. M., 1959. Effect of fatty acid mixtures on the rice stylett nematode {Tylenchorhynchus martini Fielding,; 1950). Nature, 183:
Katznelson, H.; Gillespie, D. C. and Cook, F.D., 1964. Studies on the relationship between nematodes and other soil microorganisms III. Lytic action of soil myobacters on certain species of nematodes. Can./. MicrobioL, 10:699-704.
Kerry, B. R., 1974. A fungus associated with young females of the cereal cyst-nematode, Heterodera avenae. Nematologica, 20:259-260.
Kerry, B. R., 1975. Fungi and the decrease of the cereal cyst-nematode population in cereal monoculture. f/'POJSu//., 5:353-361.
Kerry, B. R., 1982. The decline of Heterodera avenae populations. EPPO Bull., 12 : 491-496.
Kerry, B. R., 1984. Nematophagus fungi and the regulation of namatode populations in soil. Helm. Abstr., 53 :B: 1-14.
Kerry, B. R., 1986. An assessment of the role of parasites in the regulation of cyst-nematode populations. In cyst nematodes, NATO ASl series. (Eds. F. Lamberti. and C. E. Taylor.) Plenum Pub. Co. New York. pp. 433-450.
Kerry, n M\d Ciump, D. H., 1977. Observation on fungal parasites of females and

216 Recent Advances in Nematology
2' '; 8 "" eggs of the cereal cyst-nematode Heterodera avenae. and other cyst nematodes.
Nematologica, 23: 193-201. Kerry, B. R. ; Jenkinson, S. C. and Crump, D. H., 1976. Fungal parasites of cyst
nematodes. Rothamasted Exp. Sta. Rep. Part 1:201-202.
Kuhn, J., 1877. Vorlaufiger Bericht uber die bisherigen Ergebnisse der seit dem Jahre; 1875. In Auftrage des Vereins fur Rubenzucker Industrie ausgefuhrten Versuche zur Ermittenlung der Revue Revue Ursuche Ermittenlung der Ursuche der Rubenmudigkeit des Bodens and zur Eforschung der Natur der Nematoden. Z. Ver. Rubenzucker Ind. Dtsch. Reiches pp. 452-457.
Linford, M. B., 1937. Stimulated activity of natural enemies of nematodes. Science, 85 : 123-124.
Linford, M. B. and Oiviera, J. M., 1938. Potential agents of biological control of plant parasitic nematodes. Phytopathology, 28:14.
Lowenberg, J. R.; Sullivan, T. and Schuster, M. L., 1959. A virus disease of Meloidogyne incognita the southern root-knot nematode. Nature, 184:1986.
Mankau, R., 1%8. Reduction of root-knot disease with organic amendments under semifield conditions. Plant. Dis. Rep., 52:315-319.
Mankau, R., 1975. Bacillus penetrans n.comb causing a virulent disease of plant parasitic nematodes. /. Invert. Pathol., 26:333-339.
Mankau, R., 1980a. Biological control of nematode pests by natural enemies.v4««. Rev. Phytopathol., 18: 415-440.
Mankau, R., 1980b. Fungi as nematode control agents. /. Nematol., 12:244-252.
Mankau, R., 1981. Microbial control of nematodes. In Plant Parasitic Nematodes Vol. III. (Eds. B. M. Zuckerman and R. A. Rohde.) Academic Press, New York pp. 475-495.
Mankau, R. and Prasad, N., 1977. Infectivity oi Bacillus penetrans IntPlant parasitic nematodes. /. Nematol. 9:40-45.
Mcbeth, C. W. and Taylor, A. L, 1944. Immune and resistant cover crops valuable in root-knot infested peach orchard. Proc. Am. Soc. Hon Sci., 45:158-166.
Morgan-Jones, G.; Godoy, G. and Rodriguez-Kabana, R., 1981. Phytonemalodr pathology : Ultrastructural studies. 2. Parasitism of Meloidogyne arenaria eggs and larvae by Paecilomyces lilacinus. Nematropica, 14: 57-71.
Muraoka, M. and Ishibisi, N., 1976. Nematode feeding mites and their feeding behsviom. App. Entomol. andZooL, 11:1-7.
Murphy P. W. and Doncaster, C. C, 1957. A culture method for soil meiofauna and its application to the study of nematode predators. Nematologica, 2 :202-214.
Nishizawa, T., 1987. A decline phenomenon in a population of upland rice cyst nematode Heterodera elachista, caused by a bacterial parasite, Pasteuria penetrans. J. Nematol., 19:546.

Anwar L. Bilgrami 217
^ O 'J Noel, G. R. and Wilson H. J., 1981. Association of rickettsia-like organism with
reduced Heterodera populations. J. Nematol, 30 :
Page, S. L. J. and Bridge, J., 1985. Observations on Bacillus penetrans as a parasite of Meloidogyne acronea. Nematologica, 31: 238-240.
Rodriguez-Kabana, R.; Jordan, J. W. and HoUis J. P., 1965. Nematodes:Biological control in rice fields: Role of hydrogen sulfide. Science, 148: 524-526.
Rodriguez-Kabana, R.; Morgan-Jones, G. and Chet, I., 1987. Biological control of nematodes. Soil amendments and microbial antagonists. Plant and Soil, 100 : 237-247.
Rodriguez-Kabana, T. G.; Wade, C. F. and Wells, C.N., 1962. Nematodes as natural food for Macrocheles muscaedomesticae. (Acarina : Macrochelidee) a predator of the house fly egg. Ann. Entomol. Soc. Amer., 55:507-515.
Rohde, R.A. and Jenkins, W.R., 1958. The chemical basis of resistance oiAsparagus to the nematode Trichodorus christei. Phytopathology, 48 :463.
Shyre, R.M., 1971. Biotic influences in soil environment. ImPlant Parasitic Nematodes. Vol., I. (Eds. B.M. Zuckerman, W.F. mai and R.A, Rohde.). Academic Press, New York., pp. 235-256.
Sayre, R.M., 1980. Bio-control: Bacillus penetrans and related parasites of nematodes. J. Nematol., 12 : 260-270.
Sayre, R.M. and Golden, M.P., 1987MorphologicaI differences of endospores among the members of Pasteura penetrans group parasitizing nematodes. 7. Nematol., 19:555.
Sayre, R.M. and Powers, E.M., 1966. A predaceous soil turbellarian that feed on free-living and plant parasitic nematodes. Nematologica, 12 :619-629.
Sayre, R.M. and Wergin, W.P., 1977. Bacterial parasite of a plant nematode : Morphology and Ultrastructura./. Bacteriol., 129:1090-1101.
Sayre, R.M. and Wergin, W.P., 1979. The use of SEM to classify and evaluate the parasites and predators of pest nematodes. SEM, 11:89-96.
Schaerffenberg, B. and Tendl, H., 1951. Untersuchung uber das Enchytraeiden gegenuber dem Zuckerrube nematoden. H. schachtii Schm. Z. A. Angew. Entomol., 32:476-488.
Shafqat, S.; Bilgrami, A.L. and Jairajpuri,M.S., 1987. Evaluation of the predation ability oiDorylaimus stagnalis Dujardin, (Nematoda: Dory\iiiva.\Aai). Revue Nematol., 10 : 455-461.
Shepherd, A.M.; Clari, S.A. and Kempton, A., 1973. An intracellular microorganism associated with tissues of Heterodera spp. Nematologica, 19:31-34
Siddiqi, M./. and Alam, M.M., 1988. Effect of seed treatment with azadiractin on root-knot development on growth of some vegetables. Tests of Agrochemicals and Cultivars 9 (Ann. appl. Boil. 112 supplement): 20-21.

218 Recent Advances in Nematology
24Q Siddiqi, M.A. Saxena, S.K., 1987a. Effect of interculture of margosa and Persien lilac
with tomato and egg plant on root-knot and reniform nematodes. Int. Nemotal, Network News!., 4(2): 5-8
Siddiqi, M.A. Saxena, S.K., 1987b. Studies on control of the nematode Tylenchorhynchus brassicae by interculture of margoss and Persian lilac. Int. Nematol. Network Newsl., 4(4): 27-29.
Small, R. W., 1979. The effects of predatory nematodes on population of plant parasitic nematodes in pots. Nematologica, 25 : 94-103.
Spaull, V.W., 1984. Observations on Bacillus penetrans infecting Meloidogyne in sugarcane fields in South Africa. Revue Nematol., 7 :277-282.
Steiner, G. and Heinley, H., 1922. The possibility of control of Heterodera radicicola and other plant injurious nemas by means of predatory nemas specially by Mononchus papillatus Bastian J. Wash. Acad. Sci., 12 : 267-385.
Stirling, G. R.,1985. Host specificity of Pasteuria penetrans within the genus Meloidogyne. Nematologica, 31:203-209.
Stirling, G. W. and Mankau, R. 1977. New fungal parasites of Meloidogyne spp. J. Nematol., 9:285-286.
Takagusi, M.; Yachida, Y.; Anetai, M.; Masamuni, T. and Kegasawa, K., 1975. Identification of asparagusic acid as a nematicide occuring naturally in the roots of Asparagus, Chemical Letters, 43-44.
Walsh, J. A., 1979. An intracellular microorganism in tissues of the potato cyst nematode Globodera rostochiensis and the pea cyst nematode Heterodera goettingiana. J. Nematol., 11:317.
Weber, A. P.; Zwillenberg, L. O. and Van Ddr Laan, P. A., 1952. A predaceous amoeboid organism destroying larvae of the potato root eelworm and other nematicides. Nature, 169:384-385.
Webster, J. M., 1972. Nematode and biological control. In Webster J. M. Economic Nematology: Academic Press New York. pp. 563.
Wood, F.H., 1973. Feeding relationship of soil dwelling nematodes. Soil Biol. Biochem., 5:593-601.
Yeates, G. W., 1969. Predation by Mononchoides potohikus (Nematoda : Diplogasteridae.) in laborator>' c\i\lm&.Nematologica, 15 :1-9.
Yeates, G. W. 1981. Soil nematode populations depressed in the presence of earthworms. Pedobiologia,22:191-195.
Zopf, W., 1988. Zur Kenntnis der Infektions- Krankheiten Niederer Thiere and Pflanzen Nova Acta Leopold Carol., 52 : 314-376.
Zuckerman, B. M.; Himmelhocn, S. and Kisiel, M. J., 1973. Virus-like particles in Dolichodorus heterocephalus. Nematologica, 19:117-124.

Acknowledgments
The author thanks Dr. Pinochet for suggesting this study and Dr Inserra for useful procedure suggestions. Thanks are also given to Mrs. Casanovas for typing this manuscript.
References
B\iMS, R C , C\MPRo\, J. W. & Soosi, R. K. (1974). Four biotypes of Tylenchulus ieniipenetrans m California identified, and their importance in the development of resistant citrus root-stocks. J Nematol, 6 : 63-66.
B r n o , A., N^ws, A., BLI \ R I , C. & ALMR.\, P. (1986). Los nematodos de los citncos. Monografia Excelcntisimo Ayun-tamiento de Castellon dc la Plana, 222 p.
Go n I ILK, Y., Q)H\, E. & Si'ii c.i i -ROY, P (1986). Biotypes of the citrus nematode (Tylenchulus ssmipeneirans Cobb) m Israel. Phytoparasitica, 14 : 193-198.
I\srRR\, R. N., Vovi \s, N. & 0 ' B \ \ \ o \ , J H. (1980). A
classification of Tylenchulus semipenetrans biotypes. J Nematol, 12 : 283-287.
LAMBIRU, F., VOVLAS, N . & TIRRO, A. (1976). An Italian biotype of the citrus nematode. Nematol. medit., 4 : 117-120.
MARiiNb/ BERIN(,OL\, M . L. , C\RcriFS, A. & GLTILRRF/, M. P. (1987). Ensayos de nematicidas contra el nematodo de los agnos, Tylenchulus semipenetrans. Boln Samd. Veget. Plagas, 13 : 261-271.
McSoRLt\, R., P\RR.ux), J. L. & DAXKHRS, W . H . (1984). A quantitative comparison of some methods for the extraction of nematodes from roots. Nematropica, 14 : 72-84.
O R I L N O MARII.Nt/, A., GOMI-Z GoMh/, J. & CANOV4S C\\
DFi, F. (1969). Poblaciones nematologicas fitoparasitas en los suelos de la huerta de Murcia. An. Edafol. AgrobioL, 28 ; 389-398.
TARJAN, A. C. (1972). Observations on extracting citrus nematodes, Tylenchulus semipenetrans, from citrus roots, PI Dis. Reptr, 56 : 186-188.
A N E M A T O D E FEEDING MITE, TYROPHAGUS PUTRESCENTIAE ( S A R C O P T I F O R M I S : ACARIDAE)
Anwar L. BILGRAMI * and Qudsia T A H S E E N *
* Section of Nematology, Department of Zoology, Aligarh Muslim University, Aligarh-202002, India.
Accepted for publication 14 October 1991.
Key-words : Tyrophagus, Acandae, nematodes.
There are few reports confirming nematodes as the diet of soil mites. Linford and Oliviera (1938) observed mites feeding on root knot nematodes. Murphy and Doncaster (1957) reported mjury of Heterodera cysts by a mite. The most definite association came from the work of Rodriguez et al. (1962) who observed that when given equal choice, the adult mite Macrocheles muscae-domesticae preferred house fly eggs over nematodes while proto- and deutonymph under same conditions preferred nematodes. An onbatid mite Pergalumna sp., fed upon Pelodera lambdiense and Tylenchorhynchus martini in large numbers (Rockett & Woodring, 1965). Muraoka and Ishibishi (1976) described feeding by many species of mites which were identified as nematode predators. Recently, Imbriani and Mankau (1983) observed feeding of a neostigmatid mite, Lasiosetus scapulatus on Aphelenchus avenae and Cephalobus sp.
In the present work observations were made on the predatory behaviour of Tyrophagus putrescentiae (Sar-coptiformis : Acridae) using nematodes as prey.
Materials and methods
Prey catching and feeding mechanisms were studied in culture dishes and special observation chambers. A
plastic ring (1 cm high; 2 cm diam.) glued to a coverslip at one end, was fixed in the middle of a metallic slide. The chamber, thus formed, was filled with 1 "o water agar. The mites and prey nematodes were then inoculated and the ring was sealed with another coverslip to prevent air drying and escape of mites. Predation was observed on Rhabditis sp., Cephalobus sp., Hirschman-niella oryzae and Tylenchorhynchus mashhoodi. The rate of predation by T. putrescentiae via% determined by using five adult mites against 50 individuals of prey. The effect of prey density on the rate of predation by T. putrescentiae was observed by placing 25, 50, 75, 100, 125, 150, 175 and 200 individuals of H. oryzae separately with five predators. The number of individuals killed or consumed by the mites was recorded after 24 h. All experiments were carried out at 28 ± 2 "C and replicated five times.
Predatory behaviour
T. putrescentiae feed on nematodes and other microorganisms in culture dishes. During routine observations the cultures of saprophagous species of nematodes viz., Acrobeloides, Cephalobus, Rhabditis, Pana-grellus and predaceous nematodes viz., Mononchus
Vol 15, n" 5 - 1992 477

2 4 2
aquaticus, Mononchoides fortidens, M. longicaudatus, Dorylaimus stagnalis, Aquatides thomei were all found to be contaminated and consumed by T. putrescentiae. It restricted multiplication and reproduction of nematodes in culture dishes and as a result of predation brought down their population at a low level.
T. putrescentiae could hold its prey using its palp (used to identify prey organisms and other objects) and legs as soon as the prey came in contact. The legs are generally used for holding and wounding the prey. Chelicerae are the main killing and feeding organ, also used for grasping and crushing the prey. The ingestion of prey contents was intermittent (two or three feeding bouts) with short periods of resting activity at regular intervals. Mites also consumed its own faecal matter while moving randomly generally in absence of prey nematodes. No cannibalistic tendency was observed in T. putrescentiae. The nymphal stages of T. putrescentiae fed mostly on dead nematodes, their remains or upon the agar containing bacterial and fungal growth. T. putrescentiae killed Rhabditis sp. and H. oryzae most while Cephalobus sp. the least (Table 1). Maximum predation occurred in the population of 200 prey individuals which was double the number of prey killed when 25 individuals of H. oryzae were used as prey (Table 2).
Conclusions
Observations on T. putrescentiae suggest its pre-daceous nature. These predatory mites appeared to possess greater predatory potentials mainly because of their ability to kill variety of nematodes in large numbers. T. putrescentiae fed most upon Rhabditis sp. and H. oryzae. Besides, mites also consumed bacterial and fungal populations grown over the agar. Increased predation by T. putrescentiae with increasing number of H. oryzae suggests density dependant predation as is the case with many predaceous nematodes (Bilgrami et ah, 1984; 1985; Bilgrami & Jairajpuri, 1990). The copro-phagous behaviour of T. putrescentiae may be an added advantage as it could avoid intraspecific interactions (cannibalism).
References
BILGRAMI, A. L. & J.'\IRAJPURI, M . S. (1990). Predation by
Mononchoides fortidens and M. longicaudatus (Nematoda : Diplogasterida). Netnatologica, 35 : 475-488.
BILGRAMI, A. L., AHMAD, I. & J.\IR.MPLR!, M . S. (1984).
Observations on the predatory behaviour of Mononchus aquaticus. Nematol. medit., 12 : 41-45.
BILGRAMI, A. L., AHMAD, I. & JAIRAJPURI, M . S. (1985). Predatory behaviour of Aquatides thomei Nygolaimina : Nematoda). Nematologica, 30 (1984) : 457-462.
IMBRIANI, J. I. & MANKAU, R. (1983). Studies on Lasioseius
Table 1. Rate of predation by Tyrophagus putrescentiae upon different prey nematodes.
Prey species
Rhabditis sp.
(Cephalobus sp.
Hirschmanniella i
Tylenchorhynchus
:>ryzae
mashhoodi
No. of prey killed
29-33 (34 ± 1.6)
21-27 (24 ± 2.4)
28-34 (30 ± 2.6)
24-31 (27 ± 2.5)
Table 2. Effect of prey density (Hirschmanniella oryzaejon the rate of predation by Tyrophagus putrescentiae.
Prey number
25
50
75
100
125
150
175
200
No. of prey killed
6-14 (10 ± 3.0)
23-29 (26 ± 2.0)
39-43 (41 ± 1.9)
58-62 (61 ± 3.0)
82-88 (85 ± 2.8) 108-113
(109 ± 2.3) 131-136
(133 ± 2.6) 158-164
(161 ± 2.3)
"o
40
52
55
61
68
73
76
81
All figures are nearest to whole numbers.
scapulatus, a neostigmatid mite predaceous on nematodes. J. Nematol, 15 : 523-528.
LiNFORD, M. B. & OLIVIF.RA, J. M. (1938). Potential agent of biological control of plant parasitic nematodes. Phytopathology, 28 : 14.
MuRAOKA, M. & IsHiBiSHi, N. (1976). Nematode feeding mites and their feeding behaviour. Appl. Ent. Zool, 11 : 1-7.
MURPHY, P. W. & DONCASTER, C. C. (1957). A culture method
for soil microfauna and its application to the study of nematode predation. Nematologica, 2 : 202-214.
RoCKEiT, C. L. & WOODRING, J. P. (1966). Biological investigation on a new species of ceratozetes and of Pergalumna. Acarologia, 8 : 14-18.
RODRIGUEZ, T . G. , WADE, C. F . & WELLS, C. N . (1962). Nematodes as natural food for Macrockeles muscaedomesti-cae (Acarina : Macrochelidae) a predator of the house fly egg. Ann. Ent. Soc. Am., 55 : 507-511.
478 Fundam. appl. Nematol.

Indian J. Nematol 22 W •.0992]
-i -4 0
EFFECT OF CHEMICAI^ AND NEMATICIDES ON THE ACTIVITY AND SURVIVAL OF DORYIAIMUS STAGNAUS,
A PREDACEOUS NEMATODE
SAMINA SHAFQAT, ANWAR L. BILGRAMl AND M. SHAMIM JAIRAJPURI
Section of Nematology, Department of Zoology, Aligarh Muslim University, Aligarh-202002
Abstract :Ter concentrations of potassium chloride, sodium chloride, potassium iodide, urea, ammonia and five concentrations of aldicarb and carbofuran were used to determine their efiects on the activity and sua'ival of adult and juvenile stages of a predatory nematode, Doryloimus stagnalis. It v-as found that high concentrations (0.6-O.SM) of allchemicals were very toxic. In low concentrations tf-,e activity decreased with an Increase in the period of exposure but the nematodes survived for ccmparolivtiy longer durations (2880 mini. The effectiveness of salts on the activity and survival of nematodes corresponded vvllh their molecular weights and salts with higher molecular weights (e. g., potassium iodide; Mol. Wt - 166.01) were more toxic as compared to the others with lesser molecular weight (Sodium chloride; Mol Wt - 58.44).
Key words: Activity, chemkaJs,Doryfa(musstagna/ts,ncniatic Ides, survival, toxicity.
Effect of Inorganic and organic chemicals on the activity and survival of nematodes have been studied by many workers. It is evider.t that acid and mineral salts with higher molecular weights are generally very toxic to nematodes (Fenwick, 1957; Moje & Thompson, 1963; Bird & McGuire, 1966; Vassailo, 1967; Esser, 1972; Prot, 1979; Bilgrami& Jairajpuri, 1985; Bilgramietal., 1986 Stephenson(1945), Johnston (1959), Banage & Visser (1965) and Pitcher & McNamara (1972), however, observed that low concentrations of silver and cuprlc ions were toxic to Pratylenchus penetrans, Xiphinema diversicaudaturr,, Rhabdltis terresus, T^^lencborh^nchus martini and Dnr\;!Lnmus sp. The hydrogen Ion concentrations (iKo affected the behaviour of nema-fofitis (l.owenberg et al, 1960). Husaln & MdSooJ (1974) studied the effects of some organic chemicals on Helicot\,'lenchus sp. and Jairajpuri et al. (1974) observed the effect cf chemicals on/fopfo/alrnuslndicus, Xifhir.cwa basin, llelicqti'lenchus indlcus and M-,]onchuhis minor. Soroczon (1969) (ie! 'nr.i.-ied the suivival rate of Rhabdltis sp. undci d.'ferent concentrations of pH and mir.o.i:;! <alts and suggested that lower con
centrations of mineral salts and pH range 7. 5-8.8 were most suitable for nematodes. Effect of pH, mineral salts and fatty acids was observed by Ahmad & Jairajpuri (1982) on ParahacfronchusshalciliandMalik& Jairajpuri (1984) onXiphinemaamericanum.
Dory/o(miisstagna//s has great predatory potentials (Shafqat eta/., 1987; Bilgrami, 1990). This predatory species is found in cultivated fields and is most likely to be subjected to chemical treatments including nematicides. Therefore, the effect of different concentrations of various salts and nematicides was observed on the survival and activity of its adult and juvenile stages.
MATERIALS AND METHODS
Dor\;lalmus stagnalis were collected from soil samples from paddy fields from the Jamalpur village close to the Aligarh Muslim University campus. The nematodes were washed repeatedly with tap water before they were used In experiments. Ten concentrations of 0.1, 0.2 0.4, 0.6, 0.8, 0,01, 0.02, 0.04,0.06 and O.OS M of potassium chlodde, sodium chloride, potassium iodide,

- '1
20 SAMINASHAFQATe/o/
ammonia, urea and fiv€'concentrations viz., 1,10,100, 1000, 10,000 ppm of aldlcarb and carbofuran were 'prepared In distilled water by using the following fonnula.
V.N,
Where
N, = Standard
V.N,
Nj == Required solution; molar concentration
V, = Required volume of standard solution
Vj = Volume of H^O in which molar
concentration is to be prepared
Twentyfive adults and juveniles were tested in 2 ml solution of each concentration separately In small Cavity blocks. The activity of nematodes was measured in termsof head movements / min. Swing of head to one side and then back to the initial point, and again to the other extreme and back to the initial point constituted one urave / movement (Bilgrami&Jairajpuri, 1985). Observations were terminated if the nematodes died (nematodes were considered dead if they did not react when touched with a needle) within an hour after being inoculated in the solution. Each experiment was replicated five times. Experiments were carried out at room temperature (25 ± 2=0.
RESULTS
Table-1 gives maximum activity (head movements) and survival o each stage of D stJi^na/is in different chemical solutions The coiicentiations which piovsd least and most favourable ate given in paientheses.
At higher conccntiations of salts and nematicidesD stagno.wsacultsandjuveniles showed movoments bofoic death The dead spoclniens generally had i)rotrudcd spear
and spicules. The body posture of adults a'so changed and become spiral, C-shaped, closed C-shaped and figures of '8 ' and 'S' at higher concentrations. Body shapes of juveniles, however did not show any change At lowerc6ncentrations(0.01M) the activity of different stages decreased gradually.
Higher concentrations of potassium chloride were veiyi toxic and no activity was observed. Minimum survival of juvenile and adults was recorded at 0.02 and 0 04 M concentrations Concentration of 0 01 of potassium chloride was least toxic to juveniles and adults (p>0 05) Their survival rate was also maximum at this concentration
Sodium chloride was less toxic than potassium chloride. The nematodes exhibited maximum activity at 0 01 and 0 02 M concentrations. No activity of fiist, second and third stage juveniles was observed at 0 4 M concentration of sodium chloride which was toxic to all stages. The maximum survival time of adults and juveniles was recorded at 0.01 M.
Potassium iodide was most toxic. The activity of male, female and all tlie four juvenile stages was inhibited greately at all concentrations. The adult survived for 12 hr, fourtli and third stages for 8-9 brand second and first stages for 5 hr at 0 01 M The nematodes survived for minimum duration at 0 2 M of potassium iodide The first and second stage juveniles did not exhibit any actlwty at 0.1 M concentration of potassium iodide
All concentralions of urea suppiessed the activity of adult and juvenile stages The survival of nematodes v*.as, houover. net affected Thenematodessui\ii.odmaMiiHim atO 01 MandminimuniatO 6Mconccntiations of uica

? 4
DOHYLAI'-iUS STAGNALIS A5 I fFLUENCED BY CHEMICALS 21
Like urea, the ammonia was also unfavourable for the activity of all stages of D stagnalis. At 0.01 M concentration of ammonia the survival of nematodes was maximum and at O.OIM it was minimum. AJdicarb at 1 ppm concentration was least toxic and the nematodes survived for more than 7 hr while minimum survival for all stciges was recorded at 1000 ppm. Carbofuran proved comparatively less toxic than aidlcarb. At 1 ppm it was least toxic and nematodes survived for maxinrium duration. 1000 ppm was proved highly toxic and all stages of D. stagnalis survived for least duration
DISCUSSION
The adults of D. stagnalis were generally more active and survived for longer periods than juvenile stages at all concentrations. Ttie effectiveness of salts on the activity and survival corresponded to their molecular weights and molar concentrations. Sodium chloride, (mol. wt. 58.44) was least effeciive. The effectiveness of chemicals . (salts) can be arranged in the folllwing order depending upon their toxicity - potassium iodide > potassium chloride > urea > ammonia > sodium chloride. Urea and ammonia seem to be exceptions because these have low molecular weights (mol. wt. 60.06 and
17.03 respectively) but still inhibit activity of nematodes significantly at all concentrations. The toxicity of ammonia concentrations including lower (O.OIM) may be attributed to its nematicidal power '[Vassallo, 1967). The latter may be attributed to its capacity to generate high osmotic pressures even at very low concentrations. Stephenson (1945), Johnston (1959), Bilgrami&Jairajpuri (1985) and Bilgrami et al. (1986) were also of the same opinion that effectiveness of salts is directly proportional to their molecular weights. The favourable effects of low concentrations (0.01, 0.02, 0.04 to 0.08M) of all salts on the activity of D. stagnalis adults are in conformity with Soroczon (1969), Jairajpuri et al. (1974); Ahmad & Jairajpuri (1982); Bilgrami et al. (1986). Aldicarb, because it contained 10 % active ingradient might have influenced activity of nematodes at all concentrations more than the carbofuran (Samantarai & Das, 1971; Bilgrami et ai, 1986).
From the above observations, It Is evident that D. stagnalis and its juvenile stages are very sensitive to various salts and nematicides. It may be concluded that limits of tolerance become narrower with the increase in the molar concentration of the salts and pH.

n O
22 SAMINA SHAFQAT e( a!

0 '1 17
DOKYLAIMUS STAGNAUS AS INFLUENCED BY CHEMICALS 23
S 6 •-' o a. o CM a o ^ ci
O CO
6 "
e a a
o „ a
o " cl
= • 0 E ^ « a o -a- d d
E D. a
~ o 6 V o a
d
E S [^ a o "^ d
S CO g. — 00 a O (N „
c d
E a a
c r o E ^ CO a •- 00 a O CM --
O ° •£?
" 2 a
iS o a < ^ a.
o c
o o e CM o a
§ 1 2 &
^ g E r g a
d CI.
° o a .-I a
o t
°2i];
o o a r-< a
2 6 o ~ o a " o a
2 2
§ E r a Lo a
o „ ^ a
E a a
o '' E "^ a. m LI
ON • CL
a
SB
E a a
'^ E
E a a
E a a
o o „ CO ° C
2 2 a r-i ^ O.
o o -—
•c o E a
o o „ 00 o g
o E a
•— a
'= S — '-D e E in o 2 n-d a
2 &
=?g e o "r a
o o
•-1 O
o o <;- O t-H O
O 0 8
O o CO o O o
32 <
I u
5

2^,8
24 SAiMINA SHAFQAT el a!
REFERENCES
Ahmad, N. Jairajpurf, M . S. (1982). Elect ol pH, mineral salts and fatty acids on the predatory nematode Parahadronchus shakill (Jairajpuri, \969].IndianJ Nemato). 12:22-30.
Banage, W. B. & Vlsscr, S. A. (1965) The effects of some fatty acids and pf-f on soil nematodes. Nematohgica 11:255-262
Bil>jrami, A . L. (1991) Predacious dorylaims In Nematode Biocontrol, Aspects and Prospects (Eds Jairajpun, Alam, & Ahmad CBS Pubhshers & Distributors, New Deihu pp 143-148.
Bilgrami, A. L. & Jairajpuri, M.S. (1985) The responses cf Mononchus aquaticus to chemicals and pH Indian J Nematol 14:171-174
Bilgrami, A.L.Ahmad,I .&Jairajpuri ,M.S.(1986) Effect of some chemicals on the activity of Hirschmanmellaorvzae Helmmthologia23-A9-54
Bird ,A.F.&McGuire ,R.J . (1966) Theeffectofanti metabolites on the growth of Meloidogyne puanica Nema/o/ogicQ 12:637-640.
Esser. R . P . (1972) . Effect of sodium hydrochloride concentrations on selected genera of nematodes Proc he'.minlhsoc Wash 39:108-114.
Feiiwiclt,D.W. (1957) PI elimintry studies on the effect of ethylene dibromlde fumigation on the hatch-ability of Hcterodera rostocbiensis (Woli), Nematologica 2 :241 -249.
Husain, S . I. & Masood, A. (1974) Effect of certain organic chemicals on the mortality of Hel/co(ylenc/ius sp Indian J Nematol 3 : 134-137.
Jairajpuri, M . S . , Azml, M. I. & Bajaj, H.K. (1974) Effect of pH and salt concent'-ations on the suvivalof/-foplolalmus/ndicKS Helicotvlenchus indicus. Xiphinema tiisiri and Mi'loncfiulus nii nor Indian J Nematol 4 :171-18]
Johnston, T . M .(1959) fffccl of f.il'v ^CId inixti •> ^ on the rice stylet ncmTlodc (TVILVC /ior/i^r r/i J'; marlm/Fielding 1956 Natu •i. London 183 1392
Loewenberg, J . R., Sullivan, T. ft Schuster , M . L. (1960) The effect of pH and minerals on the hatching and survival of Meloidogyne incognlt't larvae. Phyiopalfio/ogy 50:215-217
Malik, Z. & Jairajpuri, M . S . (198^) E"ect of pH some mineral salts and fatty acids on survival of Xiphimmaamencanum Nematol Medit 12 : 73-79.
Moje, W. & Thompson, I . J . (1963) Toxicity of mlx'turesof 1, 2-dibromo-3-ch'orcpropane and 1, 2-dibromomethane to root knot nematode, Meloidogynejavanica Phytopathology 53:428 431 .
Pitcher, R . S . & McNamara, D . G . (1972) The toxicity of low concentrations of silver and cupnc ions to three species of plant poras.tic nemD todes Nematologica 1 8 : 385 39U
Prot, J . C . (1 979) Infli^ence of concentration qndients of salts on the behaviour of four p'ant parasitic
• nematodes RevueNeniatol 2 :1 ) 16
Samantaral , K . G . & Das, S . N. (1971) Control of plant parasitic nematodes in nee soils Proc IndionAcad Sa 37:372-376
Shafqat, S., Bilgrami, A . L . & Jairajpuri, M . S . (1987) Evaluation of the predatcv beht viour of DoryfoimusstagnalisDujardin IS l5(Nern:)toda :Dorylaimida) RevueNematol 10 :455 461
Soroczon, W . (1969) Tf)e effect of pH and t ie concentrations of nm>ciol salts c! the iiKdi'imni survival rate of larvae and adulf: "' Hhabditis sp under laboratory conditiorB Anr s Unii; Manat: Curia Sktodewsk 24:153-164
Stephenson, W . (1945) The effect c' acids on a soil nematode Parasitofoqy 36:1 5*^ 1 f<4
Vassallo, M . A . (1967) The ncm-'"L'dal povi.cr of ammonia (Abstrai t) Nematolc^ :i\3: 155

3 \emalologua 39 I' I^J 3i6-3b5, E J Brill, Leiden
ANALYSIS OF THE PREDAl'ION BY APORCEIAIMELLUS MVALIS ON PREY NEMATODES FROM DIFFERENT PREY TROPHIC
CATEGORIES
BY
A \ \ \ A R L BILGRAMI
Stction of Neiridtolot^N, D-pdrtment of Zoologv, Aligarh Muslim I'niversity, Aligarh-202002, India
\umerKdl dnaKsis ol thi predator) behaviour o( Apurcelaimellui rmahs, includins; strike rate, rncounters risuiting in attack, attacks resultina; in v\ounding, feeding after wounding, prey left •mronsumed and duration of feeding upon an individual, was made in i elation to five different pie\ trophic categories 1 Bacterial feeders .Saprophagous nematodes 2 Migratory juveniles Sedentor\ endoparasitic nenatodes , 3 Epider-nal feeders lEctoparasitic nematodesl, 4i Cortical
tetders Etloparasitit nematodes and 5 Predatorv nematodes The degree of susceptibility of iiidnidual pre\ and pre) tiophir categories as a whole was found to varv The endoparasitic r tmatodts were highK susceptible iprev sus( eptibilit) = 11" a but predators resisted predation well prt \ resistance PR =78"u Charartenstit s such as thick body cuticle, body annulations and j-'ick longitudinal lutiiular folds provided physical resistance to Hoplolaimus indicus, Scutellonema sp Hemicrteunemoides mangijirae, Hemicycbophora dhirmdn and Mononchoides Jortidens iPF=100%^ \limomhus aqualicus and Rhabditis sp exhibited behavioural resistance in the form of rapid undula-uirv movements and took evasive action when attacked No individuals oi Hehcotylenchus indicus wtri consumed bv the predators This was attributed to unfavourable/toxic pre) contents ^cheml-(al resislani e Duration of feeding on an individual prey depended on the size of the prey and other phvsteal and chemical factors
ht\ui)rd^ Aporcelaimellus mialu. Predator, Prey, Predation, Resistance, Susceptibility
There are various characteristics which may reduce the susceptibihty of prey nematodes to predation these may be physical, chemical or behavioural. Esser lM63i provided a list of nematodes susceptible to predation by different mono-
nchs and dorylaims. Quialitative estimations of the degree of susceptibility of t)lenchids, araeolaimids, enoplids and dorylaim nematodes to predation were made by Groolaert et al. < 1977). Small & Grootaert 'J983) measured the degree of susceptibility of various prey nematodes using five species of predatory nematodes. Bilgrami & Jairajpuri 1989a) modelled the strike rate oi Mononchoides longicaudatus and Al. Jortidens and the susceptibility of free living, endo-and ecto-parasitic nematodes. Such quantitative characterizations are gener-all\ lacking but become significant if the biological control potential of preda-ceous nematodes is to be understood and effective combinations of predators and prey are to be determined (Small, 1987).
In this study the strike rate of the predator, Aporcelaimellm nivalis (Altherr, 19')2, Heyns, 1966 and the susceptibility of various plant and soil nematodes to

: ,0 PREDATION BY APOKC ELAIUtLU S V/I 4LIS 357
predation was measured. The relationships between strike rate of predators, prey susceptibihty, wounding, feeding etc., and different prey trophic categories were studied.
MATERIALS A \ D METHODS
Aporcelaimellus nivalis was collected from the Victoria Gate, Aligarh Muslim University, and cultured in Petri dishes containing 1% water-agar (Bilgrami & Jairajpuri, 1988). Nematodes belonging to five different prey trophic categories (PTC were used as prey (Table I).
TABLE I
Characterization of prey by trophic categories and sources of prey nematodes
Prey trophic categories)
Prey species Source
Bdcteridl feeders isaprophagous
Migratory juveniles isfdentory endo-parasites
Cortica. feeders 'e( to-parasites
Epidermal feeders ^ectoparasites
Panagreilus redwivus Chiloplacus symmetncus Cephaloous, Aaobela Rhabditis, Acrobeloides spp
Second stage juvenileb of Aieioidogyne incognita Het'rodera moth Anguina tritici
Xiphinema ammcanum X insigne Paralongidorus cttn Longidorui
Tytencfiorhynchus mashhuudi Htrschmanmella oryzae Hophlmmus indicus
Predators
Heli''otylenchui indtcus Hemicnconemoides mangijerae Hemicycliophora dhirendn Scutdlonema, Aphelenchoides Ban'ia spp
Mononchus aquatum Mylonchutus minor Dory'aimus stagnalu Apori elaimellus mvalf, Mesodorylatmus bastiani Aqualides thornei Moncnchotdes Jortiden i M longicaudatui
Prom laboratory cultures
Prom culture on tomato motha grass wheat galls
Isolated fres from soil samples
Isolated fresh from soil samples
I rom laboratory cultures

O " 1
3 5 8 BI .GRAMl
Each prey species was encountered 50 times; each encounter involved new predator and prey individuals. The encounters (i.e., lip contact of predators with the pre} at right angles) between predators and prey were observed in Petri dishes containing unused 1% water agar (i.e., track and pheromone free) using a stereoscopic microscope. All encounters were observed at room temperature 28 ± 2° C during the day time. A potential prey nematode (previously rinsed in tap water; was placed in front of the head of an active predator with the help of a fine needle without to aching (disturbing) the predator. Abnormal predator behaviour led to the obstruction being rejected. To reduce any possible effect of satiation and prey habituation, adult predators were starved for 4-6 days. The attack represents the rapid movement of head and the application of suction created by the oesophagus of the predator .^Bilgrami &
Jairajpuri, 1989a), The strike rate (SR) of T nivalis and the degree of prey resistance PR) and susceptibility (PS) were determined by the methods of Bilgrami & Jairajpuri (1989a).
Strike rale (7ol = £ ^ x 100
„ , . . EA - AW , „„ Relative resistance = x 100 of prey i%) EA
where, E-Total number of encounters made by the predators, EA=total number of encounters resulting in attack, A\V=total number of attacks resulting in wounding.
The proportion of predators which started feeding after wounding prey (FW) and the prey individuals left unconsumed iPL) by the predators were also determined.
RESULTS
A significant correlation between EA and AW was observed for all the P T C s • Table IIA\ Similar correlations were obtained between AW and FW for PTCs bacterial feeders, endoparasites and epidermal feeders but for cortical feeders and predators the relationships were insignificant Table IIB'i.
PT(J - I: Bacterial feeders (saprophagous nematodes) (Table IIIA^
Encounters with Panagrellus redivivus and Cephalobus sp. yielded most success iSR=88% and 84%); most wounding ;AW=77% and 74'''o! and maximum feeding TW=97"/o and 100%). Among bacterial feeders Rhabditis sp. was most resistant :PR=65%' while P. redivivus (PR=23%) and Cephalobus sp. (PR=26%i

fcfc w w
PREDATION EY APORCELAIMEUl S MIAIIS 359
A. Correlation between
PTCs
1 Bacterial feeders 2 Mie;idturyjuveniles 3 Epidermal feeders 4 Cortica, feeders 5 Predators
encounters
B. Correlation between
PTCs
1 Bacterial feeders 2 Migratory juveniles 3 Epidermal feeders 4 Cortical feeders 5 Predators
attacks
TABLE II
resulting in attacks
Correlation coefficient
r
+ 0 76 + 1 00 + 0 76 + 0 83 + 0 92
resulting in
Correlation coefficient
+ 0 78 + 0 8 1 + 0 73 + 0 32 + 0 48
and attacks resulting
Standard error ,± SE1
0 07 0
0 08 0 06 0 02
in wounding
P
< 0 05 < 0 05 < 0 05 < 0 05 < 0 05
wounding and feeding after wounding
Standard error ± SE
0 06 0 11 0 09 0 18 0 09
P
< 0 05 < 0 05 < 0 05 > 0 05 > 0 05
Si8;nificant at p < 0 05
the least Aporcelaimellus nivalis fed readily after wounding them (FW= 77-100"ol Few prey were left unconsumed (PL=0-14%), the maximum being for Acrobelus and Chiloplacus symmetncus (PL= 13-14%). Aporcelaimellus nivalis took least time to consume an individual of P. redwwus iFT= 19-30 min.; Mean=23 min.i and longest to finish Acwbeloides sp. (FT=38-51 min.; Mean=44 min.).
PTC-II: Migratory juveniles (sedentary endoparasites) (Table IIIB^
Heterodera moth was predated to the greatest extent (FW=87%^ and Anguina tntici least (FW=83%). Aporcelaimellus nivalis attacked Meloidogyne incognita, H. mothi and A. tritici with SR= 74-78% and K\M-1^-11% All three species avoided predation to the same degree (PR=23-24%, with 8-13% mdividuals unconsumed). Minimum feeding time was recorded on H. mothi (FT= 16-29 min., Mean=25 min.^ and maximum on xi tntici [FT=23-37 min., Mean=32 min. .
PTC - III: Cortical feeders (ectoparasitic nematodes) ^Table I I IC
Aporcelaimellus nivalis attacked Xiphinema amencanum and X. insigne with greater success SR=78-84%) than Paialongidorus citri i^SR=58%) However, more individuals of P. citri AW=86%, PS= 14%) and fewer Paratrichodorus sp. (AW=677o,

n ^^ O s'o. - ^ ^
360
TABLE III
Strike rate of the predator Aporce.aimellus nivalis and susceptibility of different prey Iroph categories
Prfy sprcifs
Prf>
\ttacks
Encounters "^Strilie resulted in attack
EA
BACTERIAL FEEDERS P rediiuus Cep/iatobui sp Arrobeloide': sp Rhahditis sp Acrobele\ sp C symmetniu^
MIGRATORY M incognita H motlii A Iritui
44 42 38 40 36 39
rate of predators SR o/o
Wounding
Attacks resulted wounding
AW %
SAPROPHAGOL'S^
JUVENILES 37 39 39
UORTICAl, FEEDERS j X amencanum X insigne Lmigidorus sp P citri Paratncfiodoru >
EPIDERMAL 1 r maifi/ioudi H orvzae Hoplolaimus indicus Helicolyiencliw indicui Scutellonema ^p H mangiferai H d/iirendri Aphelemholder sp Basina sp.
PREDATORS M aquaticus M minur D stagnah) A nnahs M bastiam A IhuniH M lunguaudalus M fortiden\
39 42 33 29 36
[•ELDERS 37 41 39
33
37 31 33
•. 42 40
17 13 22 23 24 29
9 11
88 84 7b 80 72 78
77 74 66 35 44 79
ENDOPARASITES' 74 78 78
76 77 77
ECTOPARASITES) 78 84 bb 58 72
77 76 79 86 67
ECTOPARASITES 74 82 78
bb
74 62 bb 84 80
34 26 44 46 48 58 18 22
86 95
0
70
0 0 0
76 75
24 31 23 35 25 41 11 0
Relative resistance of prev PR'%
23 26 34 65 56 21
24 23 23
23 24 21 14 33
14 5
100
30
100 100 100 24 25
76 69 77 65 75 59 89
100
Feeding after wounding
FW ,"'0
97 100 84 86 88 77
86 87 83
83 88 85 80 75
78 92
0
26
0 0 0
91 83
100 75 80
100 83
100 100
0
Feeding
Prey left uncon-sumed PL X
9 0 0 8
14 13
13 8
12
24 18 36 -lO 17
12 11 0
100
0 0 0
21 12
0 0
75 50 20
8 0 0
Duration of feeding
FT \vcnn. Range:Medi
19-30,23 18-28;25 38-51;44 21-33;25 27-44;39 21-36;29
21-33;26 16-29;25 23-37;32
48-59;53 42-55;50 68-96;77 71-108;85 31-43,36
29-42;34 38-53;4b
-
5-10;7*
---
26-39;32 I9-34;27
39-57;44 20-28;25
143** 53-71,62 31-36;33 45-69;57 17*
-
+ 50 encuiiUters made lor each species of prey ** Predatoi fed upon only one individual of prey * - Preddto^^ discontinued feeding All \alues ail nearest to v\hole numbers.

o f- r
362 BILGR^MI
TABLE IV
Summary and comparison of different prey trophic categories
Prrv trophic categories
Bd( teridl fet'deri (sap-rophas;ousi
Migratory juveniles ^sedentary endoparasiteb
C:ortical feeders 'ectoparasites
Epidermal leeders ectopara
sites
Predators
EA M+SD
40±3.0 i7'
38±1.0 3
36±5.0 il4^
37±4.0 i l l i
19+7.0 38^
SR M+SD
80± b.O 7
76± 2.0 3
72±10.0 vl4
74+ 8.0 .11
38± 4.0 i3a>
AW M±SD
63+16.0
a9
11± 1.0 i>
77± 7.0 (91
45±43.0 ;95-
18±13.0 i70'
PR M±SD
371180 49^
23± 1.0 4
23± 7.0 i30i
5,i±43.0 ;78^
78±13.0 (17^
FW M±SD
89± 9.0 ^lOi
85± 2.0 .21
82± 5.0 •61
41±41.0 (99]
80±34.0 (42)
PL M±SL)
7± 6.0 87]
11+ 3.0 24)
27110.0 (39
17 + 32.0
19128.0
FT M+SD
311 9.0 (281
281 4 0 (14;
60+20.0 (34)
351 8.0 (231
54142.0 (781
All figures are nearest to whole numbers. Figures in parentheses indicate Coefficients of Variability jper cent). For the meaning of EA, SR, AW, PR, FW, PL, FT, see the text. MlSD=Mean Standard Deviation
Attacks resulting in wounding: Attacks on endoparasitic nematodes and cortical feeders resulted in maximum wounding with 1 and 9% variation respectively, but with predators fewer attacks terminated in wounding. More bacterial feeders were wounded, with considerable variation (CV=29%). Maximum variation (7%) was recorded for epidermal feeders (CV=95%). Prey resistance: The predaceous group was the most resistant prey trophic category. Epidermal feeders exhibited least resistance to predation by A. nivalis (PR=MSD 55% ± 42.9) with CV=787o. Feeding after wounding: The endoparasitic nematodes were preferred as A. nivalis fed most readily on them after wounding. The maximum variation has been observed in epidermal feeders (CV=99%) with a low rate of feeding. Prey left unconsumed: The maximum number left unconsumed was recorded for cortical feeders and the least for bacterial feeders but these varied most (CV=87%).

I 'RtD\IIO\B\ iPORlhUnihlnS MULIS 3 6 1
PR=33%'; were wounded after being attacked. The species of cortical feeders were equally susceptible to predation ^PS = 76-79%). A. nivalis fed most on X. amencanum, X. insigne and Longidorus sp. iFW=83-88%' and least on Para-truhodorus sp. ^F\V=75''/o), but left more individuals o f f citn and Longidorus sp. unconsumed PL=36 and 40% respectively). A. nivalis required least time to consume completely an individual of Paratrichodorui sp. ^Fl'=31-43 mm ; Mean=36 min.) and most to finish P citn ^FT=71-108 min , Mean=85 min. .
PTC - IV Epidermal feeders (ectoparasittc nematodes) ^Table HID'
Aporcelaimellus nivalis attacked Basiria sp., Aphelenchoides sp. and Hirschmanniella oryzae with most success (SR=80-84%\ Most individuals of T. mashhoodi and H. oryzae were wounded ^AW=86-95%^ but fewer Basina sp. ^AW=75%). No individuals of Hoplolaimu. indicus, Helicotylenchus indicus, Scutellonema sp., Hemi-cruonemoides mangiferae anc Hemicycliophora dhirendn were either killed or injured PR=100%). Tjlenchorhynchus mashhoodi and H. oryzae were highly susceptible
epidermal feeders (PS = 86-95%). A. nivalis fed most frequently on Aphelenchoides sp. and H. oryzae (FW=91-92%). Generally fewer individuals of this PTC were left unconsumed i PL= 11-12%) with the maximum of Aphelenchoides sp. No individual of Helicotylenchus mdicus was completely consumed ^PL=100%) as predators fed upon them For a very short period and appeared to have tasted some unfavourable substance, but 70% were wounded. Individuals oiBasiria sp. were consumed more quickly (FT= 19-34 min., mean=27 min.) than H. oryzae •FT=38-53 min.; Mean=46 min.).
PTC - V: Predatory nematodes (prey) (Table HIE'
Aporcelaimellus nivalis achieved maximum success (SR=58% when predating Aquatides thornei; for other species SR remained below 507o. Most species of predators were highly resistant to predation (PR=75-100%) but A. thornei, Mylonchulus minor and A. nivalis were most susceptible (PR=59-69%). A. nivalis (predator) fed upon all wounded individuals (FW= 100%) oiMononchus aquaticus, A nivalis, A. thornei and Monochoides longicaudatus but failed to injure or kill any M. jortidens i^PR=100%). Most individuals oi Dorylaimus stagnalis were left unconsumed (PL=75%) but none of M. aquaticus, Mylonchulus minor and M. longicaudatus was left A. nivalis took 143 minutes to consume the only individual of D. stagnalis that it wounded in comparison to M. minor where the FT was least (20-28 min.; Mean=25 min ).
Numerical analysis of prey trophic categories (Table IV)
Strike rate of the predators: A. nivalis was most successful as a predator on bacterial feeders. When predators were used as prey fewer encounters resulted in attacks but there was a relatively high degree of variation (CV=38%).

u
]'Rhl)\[\0\ RY ll'()R(H-U.Ml:I.Ll.'< \/l'IZ,/S 3 6 3
DLSCL'SMON
X'arious factors, are known to provide prey nematodes with resistance against predation lEsser, 1963; JairaJDuri & Bilgrami, 1990; Small & Grootaert, 1983'. Wounding is one factor which determines the degree of prey susceptibility as a wound may result in the loss of hydrostatic pressure of the body and affect locomotion, thus making the prey individual more vulnerable to predation, as well as allowing secondary infections (Small & (jrootaert, 1983). Esser 'J987^ also suggested that a wounded or weakened prey loses its resistance.
In this study physical characteristics such as thick body cuticle (e.g. Hop-lulairnm indtcui; Scutellonema sp.i; body annulations [H. mangiferae; H. dkirandn; and thick longitudinal cuticular folds [M. fortidens] might have provided resistance against A. nivalis, as has been observed for other predators ;Esser, 1963; Small & Grootaert, 1983, Bilgrami &Jairajpuri, 1989a'i. Acrobelm sp. and all predaceous species except A/, aquaticus resisted predation well, possibly because of their thick body cuticle. The smallest number of encounters resulting in attacks, the attacks resulting ir. wounding, and the minimal success of .4. nivalis against predators iprey'i may be an indication of less predator-predator interaction.
Predators generally requirea more attacks to injure/wound active prey. The high SR of A. nivalis against AI. aquaticus and Rhabditis sp. but low rate of wounding suggests that the two prey species avoided predation primarily by such characteristics as active body undulations and vigorous escape response which restricted wounding but not feeding. Such an active prey when wounded was consumed more easily.
Esser 1963) suggested that resistance to predation in Helicotylenchus indicus may be chemical, which was later supported by observations on M. fortidens and M. lungicaudatus which were little attracted to Helicotylenchus indicus (Bilgrami & Jairajpuri, 1988'i. Also feeding was inhibited and fewer wounded individuals were consumed (Bilgrami & Jairajpuri, 1989a; 1990). In this study A. nivalis exhibited similar behaviour towards Helicotylenchus indicus. Mononchs may be the only exception as they feed indiscriminately on their prey and many species oi mononchs were found containing intact Hoplolaimus, Helicotylenchus, Hemi-criconemoides, Scutellonema, Hemicydiaphora, Xiphinema, Dorylaimus in their intestine and these have been reported to be partially or completely resistant to other species of predators (Bilgrami et ai, 1986). Mononchs may overcome resistance of differing kinds (Bilgrami &Ja.irajpuri, 1989a; Jairajpuri & Bilgrami, 1990) as their feeding is aleatory (Bilgrami et ai, 1986; Small, 1987'i and depends exclusivelv on physical encounters (Bilgrami et ai, 19841 unlike dorylaims (Shafqat el ai, 1987; Khan el. ai., 1991); nygolaims (Bilgrami et ai, 1985) and diplogasterid predators (Bilgrami & Jairajpuri, 1988) which perceive prey secretions and aggregate around an injured prey at a feeding site and continue feeding i Bilgrami & Jairajpuri, 1989b).

2 5 7 ;^64 BU.GRAMI
I'hf high degree of susceptibihty of P. redivivus, C. symmetricus, P. citri, T. mashkoodi, H. oryzae and A. thornei may be attributed partly to their relatively soft and smooth cuticle iBilgrami et ai, 1983; Jairajpuri & Bilgrami, 1990) and partly to their activity. Less active prey are attacked and wounded more than active nematodes (Bilgrami et ai, 1983;. Environmental factors e.g., temperature, pH, moisture, soil type, chemical composition of soil, predator density t i c , also play a significant role in governing predaceous activity (Wallace, 1969; Small, 1979; Bilgrami et ai, 1983; Azmi & Jairajpuri, 1979; Bilgrami &
Jairajpuri, 1985:. There appears to be a correlation between the time spent feeding and size as predators took more time to consume large prey.
The characterization of PTC's has revealed that migratory juveniles of sedentary endoparasites were most vulnerable to predation as they appeared to lack anti-predation characteristics (Esser, 1963; Grootaert et al, \911; Small & Cirootaert, 1983; Bilgrami & Jairajpuri, 1989). Bacterial feeders which have a soft body and smooth cuticle are always wounded with a high degree of suscejitibility despite their activity. The ectoparasitic nematodes (cortical and epidermal feeders; and predators possess physical and behavioural characteristics which resisted predation better than other PTC's. The reason why cortical feeders were more susceptible to predation could be their inactivity, large body size and inabihty to take evasive action (body undulations, escape response etc.) when attacked.
RESUME
Anah :i' de la predation par .f\porrelaimcllus nivellus envers des nematodes appartenant a differents types trophiques
11 a eit" procede a une analyse numerique du comportement predateur d'Aporcelaimeltus nivalis, analyse portant sur le taux d'aggressivite, les contacts suivis d'attaque, les attaques provoquant des blt•^surt^, I'ingestion apres blessure, les proies non consommees et la duree de 1,'ingestion d'une pniie. ( etle analyse comprenait des proies appartenant a cinq types trophiques: 1) nematodes ba( teriophages saprophages,; 2\ juveniles mobiles de nematodes endoparasites sedentaires; 3) nematodes ectoparasites se nourrissant sur I'epiderme; 4) nematodes ectoparasites se nourrissant dans les tissus du cortex racinaire; 5;, nematodes predateurs. Le degre de sensibilite a la predation de chaque proie et celui de chaque type trophique considere comme un tout sont I'un et I'autre variable-. Les nematodes endoparasites sont hautement sensibles (sensibilite a la predation=77%), tandis que les predateurs resistent bien (resistance a la predation ou PR=78"/o). Certaines carac-teristiques tels une cuticule epaisse, des annelations cuticulaires prononcees ou des replis cuticulaires longitudinaux bien marques, confercnt une resistance physique a Hoplolaimus indicus, Sculellonerna sp., Hemicriconemoides rtangiferae, Hemicycliophora dhirendri et Mononchoides fortidens !PR~10ll"o . Mononchus aquaticus et Rhabditis sp. montrent un comportement de resistance consis-tanl en de rapides mouvements ondulatoires el peuvent ainsi esquiver les attaques. Aucun individu d'Heluolylenchus indicus n'est consomme par le predateur. Ce phenomene est attribue a un contenu defavorable ou toxique resistance chimique). La duree de I'ingestion d'une proie depend de la taille de cellc-ci et de certains facteurs physico-chimiques.
REFERENCES
AZMI. M . I. & J.MRAJPIRI, M , S. ; .979;. EfTect of mechanical stimulation and crowding on Myliinchulus denlatus. Nematulogia mediterranea 7, 203-207.

PREDATION BY APORCELAIMELLUi NlVAlh 3 6 5
BiLGRAMi, A L SCJAIRAJPURI, M S 1985) The responses of Mo«onc/(«j aiywaficui to chemicals and pH Indian Journal of Mematologf W [I'iM], 171-174
BiLGRAMi, A L &JAIF AjpuRi, M S (1988) l^nr^iCUonof Mononchotdes longicaudatui diViA M fortidens • (Nematoda Diplogastcrida) towards prey and factors influencing attraction Revue de Nema
iologie 11, 195-202 BiLGRAMi, A L &JAIFAJPURI, M S (1989a) Resistance of prey to predation and strike rate of the
predators, Mononrhoides longtcaudatus and M forttdens (Nematoda Diplogasterida) Revue de Mmatologte 12, 45 49
BiLGRAMi, A L & JA RAJPURI, M S ^ 1989b) Aggregation o( Motionchoides longtcaudatus and M forlidens (Ncmatoca Diplogasterida) at feeding sites J^ematologica 34 (1988), 119-121
BILGRAMI, A L &JAIRAJPURI, M S 09901 Predatwnhy Mononchoidesforttdens and M longtcaudatus (Nematoda Diplcgasterida) Nematobgica 35, 475-488
BILGRAMI, A L , AHM\D, I &JAIRAJPURI, M S (1983) Some factors influencing predation by Mononchus aquattcus Revue de JVematologte 6, 325-326
BILGRAMI, A L , AHMAD, I &JAIRAJPURI, M S (1984) Observations on the predatory behaviour of Mononchus aquattcus Nematologia Medilerranea 12, 41-45
BILGRAMI, A L , AHMAD, I &JAIRAJPURI, M S (1985) VrcdAloxy hchAwouv oi Aquatides Ihornei (Nygolaimina NeTiatoda) jVcma/o%(:a 30 (1984), 457-462
"?!ILORAMI, A L , AHMAD, I &JAIRAJPURI, M S (1986) A study of the intestinal contents of some mononchs Revue de Nematologie 9, 191-194
EsSER, R P 1 1963) nematode interactions in plates of non-sterilc water-agar Proceedings Soil &. Crop Science Society of Florida 23, 121-138
EssER, R P (1987) Biological control of nematodes by nematodes I Dorylaims (Nematoda Dorylaimida) Nemalologf Circular 4, 4 pp
EssER, R P & SOBERS, E K (1964) Natuial enemies of nematodes Proceedings Soil &i Crop Science Society of Florida 24, 326-353
GROOTAERT, P , JA^UES, A & SMALL, R VV (1977) Prey selection in Bw/Zenui sp (Rhabditida Diplogastendae) Mededdelingen Faculteit Landbouwwetenschappen Rijksuniversiteil Gent 42, 1559-1563
JAIRAJPURI, M S &B11GRAMI, A L (1990) Predatory nematodes In Metnalode Bw control Aspects and Prospects,-p^) 9^-125 (Eds M S Jairajpun, M M Alam & I Ahmad) CBS Publishers Pvt Ltd , New Delhi
KHAN, Z , BILGRAMI, A L & JAIRAJPURI, M S (1991) Some studies on the predation abilities of Aporcelaimellus nival ( (Nematoda Dorylaimida) Mematologica 37, 333-342
SHAFCJAT, S , BILGRAMI, A L & JAIRAJPURI, M S ^1987) Evaluation of the predation ability of Dorylainius stagnalis Dujardin, 1845 Nematoda Dorylaimida 1 Revue de JVematologte 10, 455-461
SMALL, R VV (1979) The effect of predatory nematodes on population of plant parasitic nematodes in pots JVematologica 25, 94-103
^ALL, R VV (1987) A review of the prey of predatory soil nematodes Pedohiologia 30, 179-206 SMALL, R W & GROorAFRX, G (1983) Observations on the piedation abilities of some soil
dwelling predatory nematodes Nematoiogica 29, 109-1 18 WALLACE, H R (1969 Wave formation of infective larvae of the plant parasitic nematode,
Meloidogynejavanica Nematoiogica 15, 65-75
25
Accepted for publication 17 August 1992

259
PREDATION ABILITIES OF NEMATODES AND PROTOZOAN AND THEIR ROLE IN NEMATODE MANAGEMENT
By
ANWAR L. BILGRAMI
Section o-f Nematology, Department of Zoology, Aligarb Muslim Uni ver si ty , Al i garh-202t802 , Indi a .
The nematodes share their environment with a
multitude o-f soil micro-fauna. Interaction of one
organism with other or amongst themselves not only
becomes unavoidable but also neccessary. Of-f the
various types of associations, which may occurr or
exist between these groups, the ones that can be
detrimental to nematode populations may be helpful in
the biological control programmes. Although, the
advocacy for the biological control of nematodes may
date back to six to seven decades, but awareness in
this field and potential of biological control was
brought into sharp focus only in recent times. As
scientists looked for cheaper, non-toxic and pollution
free control methods, the prospects of evaluating the
role of soil micro—organisms recieved greater
importance. The biological control is determined by
predator-prey relationships which is an important part
of energy flow of the biological world. If the energy
flow in the ecosystems can be regulated by governing

I 'O U
the population of pests either by inducing the bio-
control agents or manipulating naturally occurring
populations of predators positive results could be
achi eved-
Control may either be corrective (chemical)
or preventive (Cultural) but the biological control
has both these possibilities. It may he preventive in
the sense that it can help in evading the disease but
i-f the disease is already set in the malady can be
corrected by reducing the population densities of the
pest populations.
The protozoans and predaceous nematodes are such
organisms which can play significant role in nematode
management if given equal attention and opportunity.
However, before making practical use of any bio
control agent it becomes neccessary to study their
predatory behaviour, range of prey, prey preference,
rate of predation, fecundity, life cycle, population
dynamics, ecology, predator-prey relationships, strike
rate of predators, degree of resistance and
susceptibility of parasites to predation etc.,
PROTOZOANS
BASIC MECHANISMS AND PRINCIPALES INVOLVED
Several types of protozoans are known to

parasitize nematodes but results are still lacking to
justi-fy passimism in using them as bio-control agents-
The protozoan may be parasitic or predaceous
(Bilgrami, 1992a). Some species of protozoans
parasitize nematodes by forming resistant oocysts
<DDcyst5> (Coccidians) while others by aggregating
around the excretory pore and vulval opening and
entering into the body o-f the nematode (Flagellates).
Some species of predaceous protozoans kill nematodes
by forming spores while still others predate upon by
•forming digestive cysts <Theratromv>i a webri ) . The
CDCcdian parasite Lagerel1 a helmi nthorum parasitizes
Coomansus composti col a and forms resistant oocysts
under adverse conditions which are then discharged
into the lumen of the gut. These oocysts infect other
nematodes if ingested,.a self limiting mechanism in
the sense that nematodes with broad stoma may be
infected while the parasitic nematodes (tylenchs)
remain unaffected as they possess spear lumen too
narrow to ingest the cysts- The promastigote,
Leptomonus infections are known only in Tri1 obi us
qraci1i 5 (Butschali, lS78)j Pi pioqaster 1onqi rauda
(Goodey & Triffit, 1927) and Poroi qrolai mel1 us
<Bovien, 1937) but has never been reported from any
parasitic nematode. A flagellated protozoan, Bodo

*L U .C
caudatuia was observed in tbionasin treated as well as
untreated dead Di tylencbus di psaci (Canning, 1973).
Bodo caudatua does bold some promise as an agent o^
biological control-
Many species of Amoeba ars predaceous. Tbe
earliest observations o-f an amoeboid organism feeding
on Globodera rostochi ensi s 1arvae was by Weber et al. ,
(1952) . Tbe specaes was later identified as Tj^ webri .
As examined ender the Electron Microscope (SEM> the
trophic stage of T. webri covered the entire surface
of the nematode. After development, a digestive cyst
is formed. This cyst may be characterized by a rigid
wall which persisted in cultures even after
excystment. Amoeba failing to feed on nematode entered
into resting stages which are spherical and smaller
than the digestive cysts. The digestion is completed
after 24 h at 21•C. In Urostyl a the nematodes that
were engulfed either whole or partially were generally
able to rupture protozoans, themselves being remained
unaffected <Doncaster & Hooper, 19A1>. Esser's (1983)
observations on a species of Thecamoeba indicates some
weakness in prey catching mechanism. The nematodes
that were ingested lengthwise were killed by
compression of the body but others escape.

J- o3
ADVANTAGES OF USING PROTOZOANS
The protozoans may have certain advantages in
the sense that their rate o-F multiplication is very
high and the population buildup is rapid. Dif-ferent
ecological -factors are to be identi-fied which retard
and stimulate the rate of multiplication and
population o-f these organisms. The populations D-f
species forming ress-istant cyst, spores and oocysts can
be taken through the adverse climatic conditions to
favourable conditions-
DISADVANTAGES OF USING PROTOZOANS
The main disadvantage of using protoroans as
agents of nematode bio-control lies in their
sensitiveness to changing environmental conditions-
The large size of many parasitic protozoans do not
permit them to enter the body of nematode specially
the parasitic nematodes. The life span of protozoans
is not as per the requirement of bio-control agent and
sufficient amount of water is required for their
movement in the soil.
CONSTRAINTS IN USING PROTOZOANS IN NEMATODE MANAGEMENT
No doubt the above observations indicate limited
and partial role of protozoans in influencing nematode
populations. With these little informations it is
difficult to consider protozoans a doubtful starter as

agents o-f nematode bio-control- We lack informations
on various aspects o-f parasitic and predaceous
protozoans. The identification and classification of
antagonistic species ars one such factor. The
relationships between protozoans and parasitic
nematodes determining predacity, range of prey, pf^y
specificity and factors governing predation,
multiplication, behaviour, longivity, ecology, etc.,
still not clearly known are other factors which
require more and more attention. Besides, techniques
to culture and lab to land programmes are also to be
strengthend to obtain better results.
FUTURE PROSPECTS OF PROTOZOANS IN NEMATODE MANAGEMENT
Few observations have indicated possible future
role of protozoans in nematode management; e.g., those
which enter through the pore (parasites) and those
which feed by forming digestive cysts and resistant
oocysts <predators>. More experiments and efforts are
required to improve culture conditions and to improve
means to transfer them into fields infested with
parasitic nematodes. It is therefore, with these
little informations, that effort are required on all
fronts and different aspects besides the availability
of initial finances which are needed to explore the

possibilities o-f using these mi cro-organi sms -for
practical purposes i-f not alone than concoroi tt ant 1 y
with other bio-control agents at one or the other
stage o-f nematode management programme.
PREDATORY WEHATDDES
Types of predators? The predatory nematodes, depending
upon their mode o-f feeding and type of -feeding
apparatus, may -fall under three catagories. The first,
which feed by combined action of cutting and sucking
and at times engulf a prey whole, belong to the Order
Mononchida- The entire group of mononchs is
predaceous. The second type of predators feed by
cutting the body of prey and sucking its body contents
<Drder Dipiogasterida>. The third group, belongs to
the Sub Orders Dorylaimina, Nygolaimina and
Aphelenchina, is piercing and sucking type
(Stylet/spear/mural tooth).
FEEDING APPARATUS
The Mononchs s The mononchs generally have large
stout bodies, a strongly scierotired usually barrel
shaped feeding apparatus (also known as stoma or
buccal cavity) consisting of two sets of three plates
each <one dorsal and two subventrals)„ The anterior

set is more developed, large and vertically placed-
The posterior Bet is Bmail, oblique and smbeddBd in
the anterior part o-f oesophagus- The dorsal plate o-f
the vertical set always bear a large sharply pointed
tooth, while the subventral walls usually bear either
similar or smaller tooth, teeth or denticles whose
number and arrangement is greately variable. The
buccal srmsiture helps in catching the prey, puncturing
its body and engulfing it either intact or by cutting
it into pieces. The oesophageal muscles and the
hydrostatic pressure of the body help to open and
close the lumen of the oesophagus and push the food
through the oesophagus into the intestine.
The Dor yl ai ms : The -feeding apparatus of dorylaim
predators is consisted of a vestibule, guiding ring,
guiding sheath, odontostyle and odontophore. The
odontostyle lies at the base of the guiding apparatus
overlying the odontophore. The odontophore is in
continuity with the oesophageal lumen and is usually
surrounded by an ellipsoidal swelling.
The Nyqol ai .-as. In typical dorylaim the? odonstotyle is
axial in position but in nygolaims the feeding
apparatus is non-axial and called mural tooth. The
former has a dorsal apperture and groove and the later

2o7
does not have an opening or groove. The prey contents
<semi—solid or particulates) passes through the
odontostyle of dorylaims while it is sucked up by the
comnbined action o-f the guiding sheath <pharangeal
cavity) and the basal part o-f the stoma (telostome or
odontophore) in nygolaims.
The Acti nolai ros; In actinolaims the vestibule is
rein-forced with plate like or ribbed basket like
structures which may -frequently be accompanied by
large onchia with or without denticles. This
additional armature may be help-ful in killing the prey
and cutting open its body wall etc.
The Aphelenchs: These predators possess a small
narrow spear, with a lumen but without basal knobs,
supported by muscles -for its -forward movement during
feeding. The spear is piercing and sucking type.
The Di piooasteri ds; These predators possess a
comparatively small buccal cavity, well armed with
teeth D-f di-f-ferent sizes located at di-f-ferent
positions- The claw like tooth situated on the dorsal
meta-rhabdion is called dorsal tooth which is moveable
and hollow -from inside- It recieves a duct -from the
dor'sal oesophageal gland which opens in the region o-f

Or
the buccal cavity. The large dorsal tooth is the main
killing weapon. It is also responsible -for the
ejection o-f oesophageal secretions -for extra-corporeal
digestion. The sub-ventral walls bear small, rose
thorne shaped tooth while the hinder part o-f the
buccal cavity bears a small tooth at its base.
Other Predators s The predators like Thaiassoqenus
<Pelagnematoidea> possess a buccal cavity and armature
which super-f i ci al 1 y resembles to that o-f mononchs.
They have similar feeding habits (convergence). The
enoplid nematode Ironus and other related genera have
three sharply pointed teeth which act as ripping organ
to tear open the tissues o-f the prey. The large
cylindrical pharyngeal cavity act as sucking organ-
Well developed muscles are associated with the feeding
apparatus in members of Ironidae.
BASIC MECHANISMS AND PRINCIPLES INVOLVED
Prey catchi nq and feedi nq roechani sros; The prey
catching and feeding mechanisms of majority of
predators may conveniently be divided into five more
or less distinct phases <Fig. 3> viz.. Encounter with
prey Attack response. Attack , Salivation/
e>t tracer poreal di qest i on and Ingest i on/f eedi nq
<Bilgrami ?< Jairajpuri, 1990).

2u9
Predator—prey encounter ? The encounters between
predators and pi"ey depend up-on chance as well as
chemo-attraction. The dorylaim predators are
attracted towards excised prey and aggregated around
an injured prey at -feeding sites (Wyss & Grootaert,
1977; Bilgraroi et^ si_. , 1985? Sha-fqat et_ aL* » 1987).
The dipiogasterid predators percieve prey attractants
and respond positively towards live and excised -free-
living, ecto and endo-parasitic nematodes as well as
bacteria <Bilgra«ni S< Jairajpuri, 19SS). The pre-
(during -feeding) and post-feeding aggregation (after
the prey is completely consumed) o-f these predators
(Yeates, 1969? Bilgrami & Jairajpuri, 1989a) has also
been observed at feeding sites. The recent
observations on them suggest more than a causal role
of chemo-attraction in predator-prey encounters. In
mononchs encounters depend purely on chance encounters
with the prey. The absence o-f chemo-at tract i on in
mononchs is compensated to a greater extent by their
activity and efficient prey catching mechanisms- The
perception of prey seer eti ons/at tract ant s vary -from
species to species and within the species from
individual to individual (Jairajpuri 8f Bilgrami,
1990). Dip1ogasterid predators viz., D. colobocercus,
M. fort i dens and M. Ionqi caudatus are also attracted

270
towards bacteria and fed upon them, a phenomenon which
could pi ay a sigrsificant. role in maintaining cultures
of these prfsdators iYeates, 19-&95 Bilgrami &
Jairajpuri, 1 9 8 S ) ,
Attack Response; The probing is Bn important
phenomenon helping predators not only to identify
their prey, to locate a suitable spot to initiate an
attack and to attain a suitable posture to launch an
attack. The most appropriate posture is at right
angle <98">. The body o-f the prey is then slit open
or punctured- Glancing contacts or contacts other
than at right angles do not neccessarily result into
prey wounding <Bilgrami et. ai_-,19S4). Such
unsuccesful contacts lead prey to escape from attack.
The probing by H. aquat i cus is rapid, side to side lip
rubbing lasting -for very short durations (Grootaert Sc
W y s s , 1979>; quite aggressive in P. punctatus and M.
dent atus (Nelmes, 19745 Jairajpuri 5!< Azmi, 197S>;
vigorous and confined only to short distances in L.
vulvapapi 1 latum (Wyss St Grootaert, 1977>j gradual and
restricted to short distances in A. thornei and D.
st aqnal i s (Bilgrami et. al . , 1985; £3hafqat et. al . ,
1 9 9 7 ) . Mj^ 1 onqi caudatus probed its prc»y for slightly
longer duration but probing by M. fort i dens was more

271
vigorous (Bilgrami & Jairajpuri, 1993). In But Ieri us
5p. probing is in the form of quick he?sd shaking Sind
lip rubbing (Grootaert et. al.< 1977).
Attack i When predator comes in contact with th^
prey, either by chance or will-ful movement
(attraction) it establishes a lip contact. The
roechani 53J o-f attack di^-fers from predator to predator.
H.aguati C U B attacks prey by moving its lips in
backward direction resulting in its mouth being wide
open and the tooth coming in close contact with the
prey. M. dentatus attacks by widely opening its
vestibule with the help of labial muscles and exposing
dorsal tooth snd denticles to bite the preyfJairajpuri
St Azmi , 1978). L^ vulvapapillaturo attacks by
thrusting its stylet vigorously (Wyss it Grootaert,
1977) and A ^ thornei and D,^ staqnal i s by gradual and
intermittent thrusting o-f their mural tooth and
odontostyle respectively (Bilgraroi et. al., 1985a;
Shafqat et. al., 1987) whereas the dipiogasterid
predators with the help o-f their movable dorsal tooth
(Bilgrami, 1990j Bilgrami & Jairajpuri, 1990).
Cuticle perforation may be achieved rather quickly
with the help of odontostyle (dor/laims), mural tooth
(nygolaims), spe-sr (aphel enchs) , dorsal tooth

211
<«»DnDncb5), onchia < ac t i nol ai ms> , teeth (enoplids) and
the combined action of high suction force of the
oesophagus and f?)ovable dorsal tooth < di pi ogaster j ds) ,
Salivati on/e>i tr a-corporeal di qest i on ; In stylet
bearing predators <dorylaims, nygolaifss, ephelenchs)
thfcf oesophageal secretionB containing enzymes are
injected into the prey for partial digestion of the
food be-fore it is sucked up by \LY\G predators (Bilgrami
?< Jairajpuri, 1993). The extra corporeal digestion
has been observed in D. colobocercus (Yeates, 1969);
But1eri us sp. (Grootaert et. al. , 1977) L.
vulvapapillatu/n (Wyss 8t Grootaert , 1977) j M.
1onqi caudatuB and H. forti dens <Bilgrami & Jairajpuri,
1990).
Ingesti on/feedi nq? The dorylaim, nygolaim and
aphelenchid predators cannot swallow or engulf their
prey whole or cut them into pieces becuase of the
piercing and sucking types of their feeding apparatus.
Each odontostyle protrusion is accompanied by a
suction v^hich is caused by the contraction of the
DHSophageal muscles. The ingestion is typically
intermittent in almost all types of predators. The
mononcbs swallow their prsy whole or puncture its
cuticle and suck its body contents. However, species

O 7 73
D-f Hyl onchul us do not ingest a prey whole and intact
(Bilgrami et . al - , 1986) instead -feed by cutting and
Bucking <Jairsipiiri S< Asmi, 1978). The di pi ogesteri d
predators -feed by puncturing the cuticle and sucking
the body contents D^ prey-
An injured prey while escaping may attract other
predators which would then converge upon it leading to
group -feeding. This phenomenon is seen comrooniy in
dorylaims, nygolaims and dipiogasterids but not in
mononchs- The group feeding is an interesting
phenomenon as it allows predators to finish prey
quickly and then continue bunting. This results into
increased predation as a group o-f predators take less
time to consume their prey than a single predator.
The duration of aggregation at -feeding sites and the
actual -feeding time may vary -from predator to predator
and depjends on the type o-f prey, tewture and contour
o-f prey cuticle, composition, concentration, quality
and quantity of the prey contents. The feeding is
completed soon after the prey is completely consumed
(dipiogasterids, mononcbs), devouring an intact prey
<mononcbs) or sucking in the whole body contents of
prey e>*cept the cuticle (dorylaims, nygolaims,
aphei enchs).

2 7 4
Prey preference/range; The knowledge o-f predator-prey
relationships and range o-f F<rey has grteat
significance in -fDrmulafcing biological control
prografljme end in proper utilization of different kinds
of predators. The observations on the predation
abilities and preference of predators for different
prey species were mainly made on live nematodes in
aqueous suspensions; intestinal contents of preserved
predators <Bilgrami et. al . , 1 936) j i_ri_ vi tro (Bi 1 gra/ni
?< Jairajpuri, 1998) and pot experiments (Cohn St
Mordechai , 1974j Bilgrami, unpublished). Though, no
predator is really prey specific but the range of prey
is rather limited and differs from species to species
<Table-I, II Sr III). The dorylaims and mononchs prefer
Meloi dodera f1ori densi B , Pratylenchus penetrans, P.
Vu 1 nus, P. curvi tatus and Meloi dogyne sp. , whereas P.
punctatus preferred A. tri tici and A.
avenae but not Heli cotylenchus sp. (Esser, 1963; 1987;
Nelmes, 1974). M. si qmaturus fed more on T.
semi penetrans and M. javaniCB juv, than on
Heli cotylenchus roultici netus or Lonqi dorus afri canus.
M. dent atus, D. staqnalis, M. forti dens and M.
1onqi caudatuB also killed fewer Heli cotylenchus
i ndi cus (Jai r iri jpur i 8< Ai mi , 1978; Shafqat et. al . ,
1987; Bilgrami ?< Jairajpuri, 1990). The di pi ogaster i d

275
predator, But 1 er i us sp-, pre-ferred soil stages of
endo-parasitic nematodes (Grootaert et. al., 1977),
The prey nematodes Aphelenchoi des fraqariae^and
Pratylenchus juv. were attacked and consumed by
But 1eri us sp besides other ecto— and endo-parasitic
nematodeB-
RotylenchuB robustus, Tylenchus macrurus, H.
naasi, Pratylenchus sp-, were attacked and served as
•food for But 1 er i us sp. when wounded artificially.
Species of Tyl enchor hynchus, Heiaicycl i ophor a and
Tri chodorus were neither attacked nor killed by the
predators. The second stage juveniles of H.
i ncpqni t a Heterodera mothi and A. tr i ti ci favoured
predation by M. 1onqi caudatus and H. forti dens
<Bilgrami & Jairajpuri, 1990) but not Hoplolaimus
indieus, Helicotylenchus i ndi cus and Hemycyli ophora
sp. Hoplolai mus i ndi cus was preferred neither by W.
dentatus nor by D. staqnal i s . D. colobodercus did
not show preference for any particular typeof prey and
killed equal number of prey- L. vulvapapillatum
preferred A. avenae and A. tr i ti c i in place of P.
penetrans and Xi phi nema i ndex (Wyss 8e Grootaert,
1977). Bilgrami et. al . <19BA) have analysed the
intestinal contents of over 1C520 specimens of 33

Z7 i. ij
species o-f nine genera of Mononchida to B S S B B B their
choice of prey. Tylenchorhynchus, Tylencholaimus and
Chi Icpl acus were -found in the gut of maximum number of
predators- More free-living saprophagouB klS'/.'i
nematorieB were observed than either the tylenchs
(4S%>, dory!aims (42%} or the mononchs <27%). The
heterogenity of pr-ey in the intestine of moncnchs
suggests that they Are pelyvalant and their ^redatian
aleatory, Arpin (1979) and Mahapatra S< Rao <19S1)
found a significant correlation between the
populations of mononchs and free—living nematodes but
Nelmes %< McCulioch (1975) did not find such a
correlati on.
Resi stance and susceptibi1i ty of prey to predati ons
The prey nematodes have certain devices for defending
themselves from the onslaught of the predators. This
mecbanisro is termed as prey resistance (PR) and
defined as the ability to avoid predation <wounding).
The Prey susceptibility (PS) of nematodes represents
the degree of an individual being vulnerable to
predation (Bilgrami & Jairajpuri, 1989b). Prey
resistance may be physical, chemical, behavioural or
environmental in nature (Bilgrami, 1992b5 1993).
Physical resistance is due to physical

O r-; '7 11
charsc t er i st i C5 , such as thict< cuticle, body
annulations, double? cuticle, gelatinous matrix e't^. ^
(Bilgreroi & Jairajpuri, 1 98*?b > , the chemical
resistance is due to t ox i c/un-f a vour abl e/repel 1 en t
secretions <E5ser, 1963; Bilgrarai Sc Jairajpuri, 1989b>
while behavioural characteristics represent certain
mechanisms of nematodes v i z , , active body undulations
(Bilgrami Sf Jsirajpuri, 1 9 3 9 b ) , ability to retreat
instantly when attacked (Esser, 1963)5 vigorous escape
response (Small & Grootaert, 19S3) and the inability
of predators to attack etnd injure (inert mechanism)
which avoid predation. The environmental resistance
may appear in the form of temperature, humidy,
moisture, pM etc. which govern the activity of
predators and prey. The wounding is an important
determinant factor in predation. It results in the
loss of hydrostatic pressure of body affecting
iDcomotion thus making prey more vulnerable to
predation. Wounding also permits invasion of
pathogenic micro-organisms resulting into death of
prey. Besides, the resistance of prey is lost if it
is wounded or is in a weakenod condition- The
predation is measured in terms of number of prey
killed and not number of Bncounters as these
encounters may not neccessarily result always in

78
wounding or death of prey. Hence wounding could only
be considered to determine the degree o-f prey
resistance and their susceptibility to predation
<Bilgrami t< Jairajpuri, 19S9b>, Th€? ect o-par asi t i c
nematodes living an close proximities of predaceous
nematodes develope wore anti-predation devices- These
characteristics such as thick cuticle, body
annulations, toxic/unfavourable body secretions,
escape response, active body undulations etc., may be
more e-f-fective against di pi ogaster i d, dorylaim and
nygolaim predators, Mononchs may, however, overcome
resistance o f dif-ferent kinds to a larger extent as
these predators often do not require to puncture the
cuticle o-f pi 'ey but er<gulf them as a whole. These prey
arB there-fore more vulnerable to predation by
<Bononchs. This is evident from the observations of
Bilgrami et. al. <1986> who found maximum number of
mononchs containing species of Hoplolai mus,
Helicotylenchus, Hemi cycli ophora, Hemi cri conemoi des,
Xi phi nema, Rhabdi ti s etc., besides other tylenchs,
dorylaims and rhabditids entire in their intestine.
Hohandas 8< Prabhoo (1980> also found intact
TyIenchorhynchus nudus and X^ elongatum in the
intestine of I . kher i and jL^ monohyst er a. Bilgrami ?f

? 7 q
Jairajpuri <19S9b> have proposed following forroulae to
determine the strihe rate of the predators and
resistance and susceptible of prey nematodes \.o
predation. The stril<e rate (SR) of predators is
defined as the percent success with which it stttackB
prey. The strike rate of predators reflects their
predatory potential stnd could be determined as
foilDW5J
EA Strike rate (SRV. > = _ _ _ >; 100
E
The resistance may be calculated as percent degree
of resistance by the follwing formulas
EA - AW Prey resistance (PR %) = _ _ >« iS0
EA
whereas, the susceptibility of prey is determined by
Prey susceptibility <PS y.) = 100 - PR
Where, EA = Total niimber of encounters resulting into
attacJj; E - Total number of encounters made by the
predators; AW = Total number of encounters resulting
into prey wounding.
These formulae may be helpful in determining the
best possible combinations of predators and prey. The

80
Riost e - f f e c t i v e a n d s u i t a b l e c o m b i n a t i o n of p r e d a t o r
a n d p r e y c o u l d b e t h e m o s t e f - f i c i e ' n t p r e d a t o r ( i n
t e r m s nf h i g h e s t s t r i k e r s t e ^ and mo&t s u s c e p t i b l e
p r e y { i „ e . , m o s t v u l n e r a b l e t o w o u n d i n g ) .
TABLE - I
LIST DF PREY NEr ATCDES OF MONONCHS (ONLY PLANT PARASITIC)
Predf'Hi;or!3 Ps rasv i t i c n'?/-f(atodes<prey) Re-ferernvce
AnatonchuB emici ae 31 Coorf<ar(B ?c L i ma, 1965
A.q inqlymodontus 4 , 42 Szc'i-ygi e l ,1 966 j 1967.
A . t r i d e n t a t u s 38 Mulvey ,1961
CIar><us /Rulveyi 5,, 5 5 , 2 4 , 40 Cobb ,1917; Men2el ,1920
Clart;u5 p a p i l l a t u s 2 , 8 , 9 , 19 , 37 S t e i n e r $< H e i n l e y , 1 9 2 2 B i l g r a f f l i , e t . a l . ,1 986 Thor ne ,1927 ;Over gaar d -N e i l B o n , 1 9 4 8 , 1 9 7 7 .
C. s h e r i I S , 22 B i l g r a m i , e t . a l . , 1 9 S 6
Coomansus i n d i c u s 2 2 , 2 7 , 3 1 , 37 E<i 1 gr ami , e t . a 1. ,1 986
l o t one b u s an tedon tus - 22 B i l g r a m i , e t . al . , 1 986
I . b a s i d o n t u s 22 B i l g r a m i , e t , al.,1986
I . i n d i c u s 12 ,22 B i l g r a m i , e t . a l . , 1 9 8 6
I . j a j i r i ID B i l g r a m i , &k. al . ,1986
I . l o n q i c a u d a t u s 10 , 20 B i l g r a m i , et_,_ a l . ,1986
I . roonohystera 5 , I P , 1 5 , 1 6 , 2 0 , 2 4 , 3 7 , 4 ( 3 A?jmi , 1983
I . p a r a b a s i d o n t u s 20 Bilgraf^i i , e h . al . ,1986
I . t r i c u r i s 10, 22,, 3 1 , 37 B.i 1 gr ami , et.._ a l . , 1 986

?81
Table-I contd.
I. acutus 33, 4 J,. 45 Cobb, } '1 7^ Ti,or ne, 1 931
I> Bgiphiqonicus 6 Tl-(orne, 1924
I. Uhf=r-ai 5, .t.5,21 ,24,32,33,4fe Mc<harfdas 5c PrDbbDo,19aS
I . p'"r t,<hpoi 5, 43 Hohandas ?t PrDbhoD,19S0
I . s/ttl vapapi } 1 rituB 22 Aridra55y,19cj4
Mrjpr.nchi.'? --ir;urji-i CI'.F 1 , 3 , 5 , 6 , 7 , IB, 13,11 , 17 Bilgrami, e t . al . ,1983; 21,23,25,28,29,33, 19805 &T(all & Grootaert
34,35,44 1983jBilgra«ji, 1992
Mj_ tunbridqeneJ!? 13, 18, 39 Bilgrami, e t . a2_-,19a6
Hi cor-fJ-turi. riqut.>ticus 12, 25, 31 Bilgrarai, e t . el . ,1984
M c i t r i 22, 37 Bilgrarai, e t . a l . ,1986
N. d».l housic riBJ B 19 Biigrami, e t . a l . ,1936
Hyl onrhu.l us dentrttu'S 2 , 13, IS, 23 Jairejpuri & Azmi,1978
M. si qmaturns 42 Cohn ?< Mordechai ,1974;
Mankau,1982
M. aqi 1 j s 14, 36, 39 Dc3Ucet,19SB
H. hawaiensJF. 5 , 21, 24, 40 Mohandas ?< PrabhDD,1980
!!L fflinSill 1, 7, 11, 33, 42 Kulshrestha et. al.. 1993
Prionc-hulus t^Mnutiiiiii. 1 ,3 ,5 ,6 ,16 ,18 ,26 ,39 Nelmes, 1974;Sflia 11 ,1979; Small S< Grootaert, 1983
P_ muscoriKf, 13, 18, 22, 27 Bilgraroi, e t . al . ,1936 P.^rahHdronchus nU^iiiil 10,12,20,22,31,37,44 Ahmad & Jairajpuri ,1982
Bi 1 grami , e t . a l . , 1986
illOL-9iL'll"'L'L!-ii'i •/Htjiil'O.iidu'-e 1 9 , 2 5 , 3 7 , 44 Bilgraroi, e t . a l . ,1936

2 82
TABLE - II
PREY NEMATODES OF DIPLDGASTERID AND APHELENCHID PREDATORS (ONLY PARASITIC NEMATODES)
Predators Parasitic nematodes (prey) Re-ference
Butleriur. sp. 3,23,25,37,41 ,4tt,48,49,50 LVooteert, ei:.al.l977
But! pr-i UB dpqre' .gpj 3, 6, 39 Email ?f Grootaert ,1983
Pi pi pqastpr sp- 5B Lin-f Drd,1937;Lin^Drd ?<
Olivier a, 1938
Da pi ente>rnn potoh-i kus'. free living nematodes Yeates, 1969
Mononcho? des -fori idens 1,11,13,21,23,23,33, Bilgrami & Jairajpuri ,
35,39,44 1990 M. 1 onq i CRUfiaf t<? 1 , 1 1 , 1 3 , 2 1 , 2 3 , 2 8 , 3 3 , B i lg rami i< J a i r a j p u r i ,
3 5 , 3 9 , 4 4 1998
M. chanqj IS Goodr ich , e t . a l . , 1 9 6 8
S e i n u r a CP) er-i s 18 Hechler Sf T a y l o r , 1966
S.defflani 18 Wood, 1974
S - o H v t H r a e 18 Hechler & T a y l o r , 1966
S. O i u r a 18,5B Hechler & T a y l o r , 1966
5 . s t e i PF-r i 18 Hechler ^ T a y l o r , 1966 S. teoHJ caudr- ' i-^ 4 , 1 8 , 5 1 , 5 2 , 5 3 , 5 4 , 5 5 L in - fo rd , 1937?Lin-ford ?<
0 1 i v i e r a , 1 9 3 7 ; H e c h l e r , 1963

O Q O 83
TABLE - I I I
P r e y nematDdes o f d o r y l a i m and n y g o l a i m p r e d a t o r s <only p a r a s i t i c n e m a t o d e s )
P r e d a t D i - s P s s r a s i t i c n e m a t o u e s Rf£--ference
<p rey )
A p c r c e l a i m e l I u s s p - So U o c d , 1973
A. o b s c u r u s 57 T i i o r n p s Swang&r ,1 V30
A. obtufe i t -audatuB r<7 T}-.Df?.fc', 1^23
A. n i v a l i s ! , 5 , 7 , 3 1 » 1 3 , 1 <?,21 , 2 3 K h a n , e t . a l . . , 1 9 9 1 2 S , 2 9 , 3 3 , 3 4 , 3 5 , E11 g r aff.i , 1 9 9 3
5 9 , < ^ 0 , 6 1 , 6 2 C a r c h a r o l e i m u s e p - 1 , 4 0 L i r . f o r d ?' O l i v i e r d i ,
1 037
D o r y l a i m u s o b t u s i c a u d a t u s 5 7 C o b b , 1 9 2 9
D. o b s c u r u s 57 Tl-iorne S< Swange^ , i 936
D. s t a q n a l i s 1 , 2 , 5 , 7 , 1 1 , 1 3 , 2 1 , 2 3 , 2 8 , S h a f g c ^ l , e t . a l . , 1 987 2 9 , 3 3 , 3 4 , 3 5 , 4 4 , 5 9 B t l g r d j r . i , 1 9 9 2
D o r y l a i m u B s p - 2 L ^ x f o r d S< O i l v i e ) - a , 1937
Pi s c o l a i mus s p . 1 7 , 5 1 Lmford ?<• O l i v i e r a , 1937
Di s c o l a i mus sf-. 17 E s s e r , 1 9i)3
Di s c o l a i mus £>p. 5 , 2 1 , 1 8 , 2 3 Bi 1 graflti ( u n p u b i i shed^ Wood, 1 9 7 3
Labroneffla vul vapapi 11 atum 3 , t t , 18 ,21 Wyss Si Grooiae'^ 1 , i 977 5<i>,57 S...ail & G r o o L a e r i ,
19B3
Mesodoryla i f f tus q o o d e y i 5^21 Di 1 y r uimi ( u n p u b i i'.aheci)
N e o a c t i n o l ai mus 39 S-i.ai 1 ?< G r o o L a e r c ,
d u p l i c i d e n t a t u B 1 9 9 3
T h o r n i a s p . 2 3 , 3 9 , 4 2 E-ioosalas S, flcirikau , 1 9^5
A q u a t i d e s t h o r n e i 1 , 5 , " , 1 1 , 13 ,1 B , 21 , ' , 33, , 34 , 3 5 , 4 4 )Ri 1 g r am , 1 9 9 2
->~r r-\r

2 8 4
KEY TO PREY SPECIES Anquina tritici (1> H. hapla <A) j^etBradera mot h i (7) H o p l o l a i m u s sp, ( tH) H. i n d i c u s (13) H. d i h y s t g r a (:5 A> Aphel e n c h o i d g s sp. <!*?) Ty 1 e n c h o r h y n c h u s <27?) T . H e m i c y c l i o p h o r a s p ( 2 5 > H. fflanqi-ferae X i p h i n e m a Gp„ P a r a l o n Q i d o r u s c i t r i Pratylenchus sp.
Roty1enchulus reni-formJB Tylenchulus sp. T. macrurus T. sparsus P. paratensis Hi. tri-foli P, penetrans Hemi c ycli ophor a dbirender!
(78)
(34)
(37)
<40)
<43) (4A> <4!;')
( v b l >
M e l o i d o q y n e n p , (2> H. i n c o g n i t a <5> Hi. s c h a c h t i (8^ H. i n d i c u s <11> HJL v u l Q a r i s (14> A p h e l e n c h u s s p . <17 > Hi r s c h m a n n i e l 1 a (20^ T . m a s h h o o d i (23> M. t y p i c a (2o^ S c u t e l 1 onefpe 5p-. (29) X. e l o n qa t uR* <3?> L o n q i d o r u s . &p. <35>
P a r a t y l e n c h u s (38) m a c r o p h a l u s R. r o b u s t u s (41)
T r i c h o d o r u s sp^ (44) H e r l i n i u s s p . (47) D. ffiyci I i c3 i3haqu5(5g) A. p a r i e t i n u s (53 i X. inde>i (56> P a r a t r i c h o d o r u s (59> B a s i r i a 5 p . ( i2>
M- naas i (3) G. r p s t o c h i e n s i s (6) H^ r a d i c i c o l a (9) H e l i c o t y 1 e n c h u s s p . (12) H. fflulticinctus (15) A. a v e n a e (18) \L. o r y z a e (21) T, n u d u s (24) Hemi c r i conentoi d e s (27 > S . c u r v a t a (30) X„ a m e r i c a n u m (33) L , c a e s p i t i c o l a (36)
R o t y l e n c h u s (39) •f a l l o r o b u s t u s T y l e n c h u l u s (42) semi penetrans T. o b t o s u s (45) A. • f r ag rae i (48) H. g>er ion i (51) D. d i p s a c i (54) eggs o-f H. schachti (57) Xiphinema insiqne (hD)
FACTORS INFLUENCING PREDATION
The e-f-ficacy of biological control agents is
more likely to be affected by the environment. The
organisms which grow in soil ars more affected by soil
conditions. Each species has specific conditions and
requirements for various environmental factors such as
temperature, moisture, pH, soil type, constituents
etc., for their growth and so it is tedious job to
generalize soil conditions favouring biological
control. Thoi.jgb different biotic and abiotic factors

2 8 5
are hnown to in-fluence activity of predators i n vitro
but similar experiments under natural cDnditionB are
very -few. Nevertheless. the same -factcrs may also
in-fluence e-f-feciency of predators under natural
conditions (Table- IV>. Predation depends upon the
type and activity of prey (Bilgrami et.al.« 1983),
type of body cuticle, annuiations, body secretions,
and other behavioural patterns (Bilgrami Sc Jairajpuri,
1989}. The age of predators (Veates, 1969; Bilgrami,
1990bj Jairajpuri 8< Bilgrami, 1990), prey density/prey
number (Yeates, 1969; Bilgrami et.al . , 19S4; Bilgrarai
St Jairajpuri, 1990; Bhafqat et. al . ,19B7i Khan et_ al . ,
1991; Kulshrestha et_ al>, 1993), temperature (Bilgrami
et. aj_' 5 1934; 1985), starvation of predators
(Bilgrami et . al . , 1984; Bilgrami ?< Jairajpuri, 19S9c;
1990; Khan et_ al_. , 1991} Kulshrestha Bjt §!_'» 1993),
environmental conditions, seasonal fluctuations, soil
conditions, soil pH and the chemical composition of
the soil are other factors which govern predation.
Besides, the major restriction in the development
of and effective biological control of nematodes is
the large bulk of soil that is to be treated to ensure
contact between agent and prey and bull; population of
predatory nematodps that is requi rrjd for the purpose.
The soil environment is to be manipulated to encourage

^ 8u
TABLE - 2V
FACTORS INFLUENCING PRErATORS AND THEIR A C T I V I T I E S
F a c t o r s P r e d a - M o b i l - F e c u n d - L i f e - P o p u l a t i o n i t y i t y c y c l e t i o n
1 . Type o f p r e y +•+
2 - P / c t i v i t y o f p r e y ++
3 . A c t i v i t y o f p r e d a t o r s ++ ++
4 . S t a r v a t i o n o f ++ ++ +4-p r e d a t o r s -
5 . Physical s t a t e o f +•*- ++ p r e d n t o r s and p r e y
6. Te fT i pe ra tu re ++ +•-<- ++ +4- ++
7 . M o i s t u r e ++ ++
S . S o i l T y p e ++ ++
9 . S o i l pH ++ +4 +•+•
1 0 . Cl-)emical c o n s t i - ++ ++ t u e n t s o f B o i 1
i l . T r a n s p l a n t a t i o n s ++
12- P l a u g h i n g ++ ++
1 3 . F a l l o w i r . g ++ ++ ++ ++ ++
1 4 . O t f i e r far iT i p r a c t i c e s ++ ++ ++
1 5 . A g r o - c h e m i c a l B ++ ++ +•+•
b u i 1 d u p o f t h e p o p u l a t i o n o f p r e d a t o r s . A d d i t i o n o f
o r g a n i c m a t e r i a l s may i n c r e a s e t h e p o p u l a t i o n o f
p r e d a c e o u s n e m a t o d e s b u t s u c h a d d i t i o n s b e s i d e s o t h e r
s t i m u l a n t s may n o t b e w i t h o u t s i d e e - f - f e c t s . T h e e n d o -
p a r a s i t i c n e m a t o d e s ars p r o t e c t e d w i t h i n t h e r o o t s a n d

287
are attacked only during their soil phase o-f
development- Nematodes such as the species o-f
Pi tylenchus that multiply rapidly within the host may
be most di'fficult to control biologically.. A number
D-f endo- and semi endo-parasi t i c nematodes {e.g., T.
semi penetrans, Weioi dogyne spp,> develope saccate
•females that srs sedentary on roots and lay their eggs
in a gelatinous matrix may be vulnerable to parasitism
and predation. Little is known about the agronomic
practices that a-f-fect the expression of biological
control.
Canni bali sm and its si qni fi cances The cannibalism
appears to be more o-f natural phenomenon rather than
occurring only due to prey non availability as in most
instances the mononchs occurred together with other
prey nematodes in the intestine o-f other mononchs
(Bilgrami et. al., 1986). Still very little is known
about cannibalism in predatory nematodes but is
certainly a negative aspect -for biological control
programmes. Cannibalism is known to occurr only in
mononchs <Azmi S< Jairajpuri, 1979$ Bilgrami ?<
Jairajpuri, 19S5a| Jairajpuri & Bilgrami, 1990).
Bi oloqy and culture of predators: Mononchs have very
restricted distribution but others like the dorylaim

288
and nygolaim predators are o+ cosmopolitan nature and
ubiquitous group of nematodes. they occur in all
types D-f climates and habitats. The presence of two,
three or more genera o-f dorylaim or nygolaim predators
at one place/field is quite usual and their abundance
has been estimated to 200-508 million/acre (Tborne,
1930). Still very little is known about their
biology, reproductive potentials and culture
techniques. Following Esser (1963), who cultured
predatory dorylaims and mononchs Yeates (1969)
cultivated dipiogasterid predator ^ D. colobocercus in
agar using bacterium Baci11 us cercus var wycoi dus.
Nelmes (1974) reared P^_ punctatus on A^, avenae and P.
redi vi vus in soil extract agar medium at pH 6.8
similar to Bpringets (19A4) who reared enchytreads.
Same techniques were used to elevate populations of W.
aquati cus and P. punctatus using P. redi vi vus as prey,
Bilgrami & Jairajpuri (1990); Bilgrami et. al. (1985)
and Sha-fqat et. aj_. (1987) cultivated M. f orti dens and
W. lonqicaudatus., A. thornei and D. st aqnal i s using
prey nematodes in agar plates supplemented with
nutrients. M_j_ f orti dens, H. 1 onqi caudatus, D.
colobocercus and other dipiogasterid predators also
•feed upon bacteria besides prey nematodes and thus can
be cultured using bacteria whose population can be
raised to desired level by adding organic compounds in
the culture medium (Bilgrami 1990a).

289
At tr i butes o-f an e-f-f i ci ent predatory nematode; The
characteristics of an ef-ficient predaceous nematode
Bre similar to those -for other soil borne predatorB.
These depend generally upon the type o-f control
required and as such no predator has all the
attributes listed. Basically, there are three
practical approaches to Biological control which may
be applied to predaceous nematodes also i.e., i)
Jnundai i on i where the predators are introduced in
large numbers to control pests rapidly. Frequent
application o-f predators may be required. Using
predaceous nematodes this technique could be use-ful in
glasshouses, transplantation crops and other such
short term treatments. ii> Introduction or mass
re^leasei When the predators are absent normally from
the soil but upon introduction they can spread and
established in the soil- iii) HaturaJ Controls Where
the predators are increased fortuitously in soil and
their manipulation is confined to preserving or
enhancing conditions that favour their activity. Such
control is more likely to develop with monoculture and
perenniel crops- Safety to non-target organisms.
Ecological corapatabi1ity, Temporal compatabi1ity,
Control potentials. Environmental adaptability.
Persistence capablity, Dispersal capacity. Biological

230
compatabi 1 i ty, High •fecundity. Short li-fe cycle,
Longivity, Searching capabilities. Easy culture
conditions, Compatable to standard farm practices.
Easy to handle and use -for practical utilization are
some D-f the characteristics which can be attributed to
an e-ffecient predaceous nematodes.
Constrai nts; The biological control (using predaceous
nematodes) o-f plant parasitic nematodes may be more
challenging than o-f any other pest in the sense that
they usually inhabit soil and attack underground parts
of the plants. This could be one of the main reason
that our eyes go unnoticed about what these tiny
predators Bre doing inside the soil with their prey.
Secondly, a large population of predaceous nematodes
is required for a successful bio-control. Means are
lacking to culture them and elevate their populations
en mass for large inoculums. Thirdly, the defficient
knowledge about their taxonomy, classification,
identification and behaviour is another setback which
has prevented sofar their better understanding as
agents of bio-control. These aspects need to be
strengthend. At times the inundation of predaceous
nematodes at a very high dose may become impractical
for treating large field/area. This may not be

291
achieved unless and untill a resistant stage oi
predacecus nemBtode is -found si mi 1 ar to the insect
nematode predstor Neoplectana carpocapcae strain DD
BIOLOGICAL CONTROL POTENTIALS OF DIFFERENT
PREDATORY NEMATODES
The biological control potentials c-f predatory
nematodes vary •from group to group, genus to genus and
species to species. However, the one roost deterimental
to the population of pest nematodes can be a good
agent o-f Bio-control.
Bi ol oqi cal control potent i al s o-f mononchs; The
interest in using mononchs for controlling plant
parasitic nematodes was initiated since Cobb (1917j
1920); Steiner ?< Heinley (1922) regarded C. papillatus
as a promising species to work. Not much work was
done on these aspects until 1970's when observations
on predatory abilities of mononchs were made. Cohn &
Mordechai (197-4> -found correlation between high
population levels of M. si qmaturus and low population
levels of T. semi penetrans in pot e>! per i ment s. Small
(1979> found a significant reduction in the population
of G. rostochiensi5 and M. i ncoqni ta in the presence

0 O 0
of P. punct atus in pot experiments. Ahmad S<
Jairajpuri <1982) -from a field data found a
significant correiation between populations of P.
shaki1i < Tr i chodorus sp. Hemi cri conemoi des sp. Azmi
<1983> indicated that with the increase in the
population of I. fl»onohystera a gradual reduction took
place in the population of Heli coty1enchus di hystera
but after some time an equillibriuro is maintained
between them. The mononcbs msy be proved effecient
predators because of their stranger predatory-
potentials, high rate of predacity and high strike
rate. The main difficulties with mononchs are their
e>;treme sensitiveness to temperature, moisture, soil
types, soil pH, soil constituents etc., restricted
occurrence, longer life cycles, low rate of
reproduction and lesser degree of population
estability. The possibilities of using mononchs in
biological control may not be ruled out merely because
of the above mentioned limitations as many of these
conditions can be manipulated and efforts are still
required to e>}plore further their predatory behaviour,
ecology, biology etc., which may prove vital for
future course of action.
Bi oloqi cal control potenti als of dorylaim and nyqolai m
predators; Though, dorylaim and nygolairo predators

233
possess predatory potentials unsqual to mononchs but
their role? in biological control of plant parasitic
nematodes seems to be more promising. The wide spread
and abundant presence o-f these predators could
indicate their biological control activities.
However, it is not known to what e>:tent and under what
conditions the population o-f phyt o-par asi t i c nematodes
are reduced since, such stn evaluation has never been
made to provide evidence. The dorylaims and nygolaims
are one o-f the 'nost ubiquitous group o-f nematodes
occurring in all types o-f soil, climates, and
habitats. The presence of 2, 3 or more genera of
these predators at one field/place is quite common.
The abundant and naturally occurring populations of
dorylaim and nygolaim predators have advantages over
other predators. These predators need only slight
modifications in the field conditions to maintain and
establish their populations. Being omnivorous i.e.,
feeding on algae, fungi /Hollis, 1957; Ferris, 1968;
Wood, 1973) and nematodes their populations could be
raised more easily using all these hinds of prey. The
detection of Eudor yl ai )>>U5 obt usi caudatus feeding on
the eggs inside the H ^ scharht i i cysts or the increase
in the population of predators (Thorni a) in the

7 0 4
presence o-f citrus nematode and decrease in absence o-f
them in pot trisi 1 5 <BoD5alis ?< ManJ;att, 1 9< 5> indicates
potential rather than their casual role in nematode
bic-CDntrcl. Similar to other predatory groups same
attention is required on various aspects of these
predstors.
Bi ol oqi cal control potent i sil B^ o-f diploqasterid
predators t The studies c arr i ed out so -far on
dipiogasterid predators indicate their promising
nature- The main advantage o-f di pi ogast eri d predators
lies in their ability to percieve prey attractants
{predation may increase), high strike rate (Bilgrami 8c
Jairajpuri, 19995 Small ?< Grootaert, 1983), high rate
o-f predation, short life cycle (minimum being around h
days e.g., M. < -f ort i dens) , High fecundity <build up of
population), and bacteriophagous nature. Besides they
are easy to culture. A mere application of organic
substances (leaves, cow dung, manuare sr.d other
nutrients) enhance their population and helps in
culturing them <Webster,1^72;Bi1grami,1990a>-Being
polyphagous they can be maintained without nematodes
using bacteria. They seldom resort to cannibalism thus
the setback of intera-specific competetion may be
avoi ded.

:o5
Future prospects? The •future prospects o-f
predaceous nematodes and protorcans are difficult to
asses without making elaborate studies and obtaining
evidences in favour or against their use in nematode
management. The assessment also depends greatly on
what advances have been fT:ade on other control methods,
their effectiveness and on the resources provided to
establish research prograoimes. With very little
informations and knowledge currently in hand and with
a very limited research and efforts and insufficient
e>!pertiBe it can only be thought in the positive and
hopeful perspectives- Very little research effort
(9X> on predaceous nematodes and (2%) on protosoans
<Table - V> itself attracts attention on various
aspects of their predation and parasitism. It may
sound discouraging that at present nematode management
by predaceous nematodes and porotosoans is largely
theoretical barring few studies, at least for the time
being, unlike its sister disciplines of Entomology,
Plant Pathology and Insect Nematology where biological
control is feasible practically. However, significant
roles of predators and tremendous potentials of many
of them indicate a possisble bright future of
biological control in Nematology too. The success in
fvlematology also depends upon how much interaction is

*L ^' iV
made between various aspects, the mere the interaction
the more and earlier the chances of getting sound
results. Biological control always require e>!pert
handling of the situation, previous knowledge about
the predator and prey, their ecology, biology,
behaviour, environmental -factor^; governing their
activities wnd more importantly their correct
i dent i-f i c at i on and c 1 assi-f i cat i on . The development and
advancements in Genetics Bnd HoSecular Biology greatly
improved the prospects o-f biological control- The
mutations, genetic engineering and manipulations can
produce more «a-ffective nf^matode predato'^s. Lastly the
success and utilization o-f predaceous nematodes and
protozoans depend upon the funding agencies and
interested researchers.
TABLE - V
RESEARCH EFFORTS ON DIFFERENT ANTAGONESTIC ORGANISMS
Crgani&ms ReuPtirct-i r-f + crtn Orgi n i r?-fl)C Re'3f.-firch e-f-forts
Predaceous -fiu'igj 5Q Col emhol an's 4
P a r a s i t i c •fungi \9 R i c i t p t t ^ . i c1(r
Predaceous nofriatod^-'O- ^ Virur=.ps 1
B a c t e r i a 4 Tard i grridf^F 1
P r o i DiL'C'ans • 2 Tn rbe l 1 H.ri MCts 1
Mitf?'.- 1 Enr.hyi-rr ipi d'=. 1
nt'tt.-f im nt^d -i-rnni an f?M ^iiu ttr^i t on (j-? mnr*^ M-t,-in --"PP \-<r^\'<i^r'-•.

O n 7
Future studi es requi red on predaceous neynatodes sti\6
protoz oans; The -future studies on pr pdaceous nep)atcdf?B
and protozoans are de-?inp1-?y required as these agents
specially the predaceouB nematodes in most cases have
proved their credentials. Means have fco be developed
to get more BVI^ more people engaged on predaceous
nematodes. Trul •/ Bpertking -future studies require a
strong interaction ^vi'di coordinatjon between fche
nematode tavonomists, behaviourists, ecologists and
pathoiogistB,Future studies are needed on all
predatory nematodes dorylaim, nygolaim and
dipiogasterid predators. The whole of the Family
Dory 1 ai mi dae and a large number o-f species of the
Order Dip1ogasterida appears to be predaceous. Almost
all the ^nembers o-f the Sub-Order Nygolaimina Stre
suspected predators and thus need proper
i dent i f i art i on, It may not be wrong i-f said that
taxonomy and behaviour s^r e the integral parts o-f
biological control. It is throtjgh an e>ipert
ta>!onon»i5t and behaviourist that correct
i dent i-f i cat i on of the species involved and predator
prey rel r h i onshi ps &r^ established. More studies are
required on culture techniques both i n vitro and i n
vi vr> for their large inoculum ir> fields.

2£3
Concl u si on Si Irj view o-f the success achieved by the
Entomologists, Plant Pathologists and Insect
Nematol ogist5 in controlling pests through their
natural enemies, it becomes logical that the
nematologist5 should also consider the possibilities
of controlling populations of plant parasitic
nematodes through their natural enemies. Though the
use of chemicals and nematicides in controlling plant
parasitic nematod&B have yielded Bome positive
results, but their use is now been discouraged because
of their exorbitantly high costs, their tonicity to
other beneficial soil fauna including predatory
nematodes <Bilgrami 8t Jairajpuri, 39S5a), their
residual effects on human beings and live stocks and
because of the fear that the continuous use of these
chemicals may result in the nematodes becoming
resistant to chemicals. The studies carried out so
far on different aspects of predatory nematodes like
identification, ecology, predation, range of prey,
reproduction and their relationships with plant
parasitic nematodes has led nematol ogists to test them
in agricultural fields for controlling populations of
plant parasitic nematodes. Some of these effects have
resulted in understanding some of the mechanisms of
predatory nematodes hwich have remained unexplored

233
e.g., attraction <BilQrami ?< Jairajpuri, 198S)5
aggregation (Bilnrami S< Jairajp^urij 1989; Pilgrami et
al . , 1 9B5) ; gut content anal yse?. <Bilgremi et. al . ,
1986j Ahmad et_ al., 1991)| quantitative determination
end estimation o-f the strike rate o-f the predators end
resistance end susceptibility of plant parasitic
nematodes to predation ^Bilgran-^ ?< Jairajpuri, 1939),
characterization of the prey catching and feeding
mechanisms of predaceous nematodes (Bilgrami ?f
Jairajpuri, 1990), besides, experiments in pots or
field conditions <Linford & Oliviera, 1922; Boosalis S<
Mankau, 1965; Cohn & Mordecbai, 1974; Small, 1979; Lai
et. al., 1983; Armi, 1983) also indicate encouraging
results. The efforts of predaceous nematodes have
also yielded several reviews during the recent times
(Esser, 1963; 1987; Webster, 1972; Brown & Kerry,
1987; Small, 1987; Bilgrami, 199Ba; 1990b; Jairajpuri
St Bilgrami, 1990). Still more informations are
required on various aspects of predaceous nematodes.
At last it becomes pertinent to quot Jatala
<1986)..... the proper and efficient utilization of
bio-control agents in suppressing nematode populations
is a great challenge. Removal of the obstacles in
achieving "tir goals can only be attained by
dedication, per si-stance and hard wor J< .

0 r 00
REFERENCES
AHMAD, N. S< JAIRAJPURI, M. S. (1982). Population
fluctuations of the predatory nefnatodes,
Parahadronchus shaki1i <Jairajpuri, 1969)
< Hon one hi da > , Prpc . Symp. Ani >.n. Popul . Indi a Pt.
3 s 1-12.
AHMAD, W. AHHAD, I., BILGRAiil, A. L. Sf KHAr4, Z.
O 991 ) . Thai assoqenus shami oii n . 5p .
< Wemat oda ! Thai asBogBDer i dae) , a nejnahode predator
•fro(B India- Fund Bment al and Appl i ed Nemaiol oqy»
15 : 127-131.
ANDRASSY, I. <1964>. Subwasserneroetoden au5 groben
gebi rgsgegenden Osta-frika, Acta. Zool . Acad.
Sclent. Hung.. 10: 1-59.
ARPIN, P. n 9 7 9 } . Ecologie et systemati que des
neinatodeB Hononchidae des zones -forBsteries et
harpaeces zones climate, teropere humid
I.Types de sol et groupments specifiques. Revue
Nematol. 7 J211-221.
A2MI, M. I. n 9 a 3 ) . Predatory behaviour of nematodes.
Biological controL of Heli cotylenchus di hystera
through the predaceous nematode,Iotonchus
monohyst &r s. Indi an J. Neroatol . 13 » 1-S.

On
AZMI,H.I. ^ JAIRAJPURI,M,S- (.1979), Effect of
mechanical st i mi-tl at i on and crowding on
Wylonchulus dentatus. Hematpi medi t. 7 j 203-
207.
BILGRAMI, A. L. <1990a>. Pi pionasterid predators: In
nematode bio—control s Aspects snd Prosptects
(eds Jairajpuri et.al.) CBS publishers St
Distributors, New Delhi pp. 143-148-
BILGRAMI, A. L. (199Pb), Predaceous dorylaims; In
Nematode Bio- control} Aspects and Prospects
(eds Jairajpuri et. al . ) CPS publishers ?<
Distributors Pvt. Ltd. New Delhi pp. 143-148.
BILGRAMI, A. L. <1992a>. Biological Control agents
and thei r rol e i n nematode wanagement. In
Recent Advances in Nematology <Ed. B. K.
Dwivedi) Publ. Bioved Research Society,
Allahabad, 203-218 pp.
BILGRAMI, A. L. (1992b). Resistance and
susceptibility of prey nematodes to predation
and strike rate of the predators Mononchus
aquati ens, Dnrylai mus stagnali s and Aguati des
thornei . Fundament al and Appli ed Nematoloqy.
15 : 265-270.

»-> - ' w ' v_. t«.
BILGRAMI, A. L- (3 993). Relationship between
predation by Aporcelaimellus ni vali s and
di-f-ferent prey trophic categories. b^ema tol oqi ca
< i n press) ,
BILGRAJil, A„ L. Sc vTAIRAJPURI, «, S. (1985a),
Cannibalism in Mononchus aquati cus, Indi an J.
Nematol, 14 (1984) ; 202-203-
BILGRAMT ,A.L. ?< JA.TRAJPURI, H, S. (1995b). The
responses of Hononchus agnat i cus to chemicals
and pH. Indian J. Nematol. 14 (19S4) sl71-174.
BILGRAMI, A. L. ?. JAIRAJPURI, M. S- (1988). Attraction
of Mononchoi des lonqicaudatus and M, forti dens
(Nematodas Di pi ogaster i da) towards P'"ey and
•factors in-fluencing attraction. Revue Nematol . ,
11:195-202.
BILGRAKl, A. L . S< JAIRAJPURI, M, S.(1989a>. Aggregation
D-f Mononchoi des 1 onqi caud at LIS and M. f ort i dens
(NematodasDipingastprida) at feeding sites.
Nematoloqica, (19RS) 34s 119-121.
BILGRAMI, A. L. ?< JAIRAJPURI, M. S. (1989b).
Resistance of prey to predation and striJre rate
o-f the predators^ Mononchoi des 1 onqi caudatus and
M. f ort i dens ( ne«r<at oda : Di p I ogasrter i da ) . Revue
Nematol.. 12 J 4 5-4 9.

3 C 3
BILGRAMI, A. L. ^ JAIRAJPURI, H. S. <19B9c>. Effects
o-P con cent r at. i ons and tbicknes"^ o-f agar on the
predatory behaviour of Mononchoi des
lonqicaudat«.<5 and M. f orti dens {Nematoda
I Di pi oga^iteri da > and on their attraction
tD«*r»rd5 prgy, Indi an J.Nematol 0 933) «iai222-227.
BILGRAM3, A. L. ?< JAIRAJPURI, K. S. U 99iS> . Predation
abilitip?5 of Hononchoi des lonqicaudatus and M.
f or t i dens < Nemat oda ; Di pi ogar^t er i da > and factors
ir»fluencing predation, Nematol oqi ca, 35 i 4 75-
433,
BILGRAMI, A. L., AHMAD, I, & JAIRAJPURI, M. S, <1983>.
Some factors influencing predation by Mononchus
aquat i cus. Revue Nematol , 6 : 325- 326.
B.ILGRAMI, A, L., AHMAD, I. & JAIRAJPURI, M. S- <19B4>.
DbservationB on the predatory behaviour of
Mononchus aquati cus. Nematol. Medi t. 12 s41-45,
PILGRAMI, A. t-, AHMAD, I. It JAIRAJPURI, M. S.
<1985a}- Predatory behaviour of Aquati des
thornei { Nygolaimina: Nematoda). Nematoloqi ca
<1984V 30 : 457-4o2. *
BILGRAMI, A. L-, AHMAD, I. ?< JAIRAJPLfRI, M. S-
(1935b). Factors influencing attraction of

0;l
adult Hi rgchmanni el 1 a oryz ae towards cabbage
BPedlings, Pevue Nematol.^ 8: 67-75.
BILGRAMI, A. L., AHHAD, I- 9< JAIRAJPURI , «. S. <1986}.
A study oi the inttjstinal contents o-f some
mononchrB. Revue Nemaiol . , 9 s 191—194.
BILGRAHI, A, L-, AHHAD, I. ?< JAIRAJPURI, «. S. <1988>.
Interaction between mononchs in non-sterile agar
plates- Indian J. Nematol . , U987> 17 : 339-340.
BOVIEN, P. ?1937). Some types o-f associations between
nematodes and insects. Vi denskabeli qg
Neddelelser Fra Dansk Naturhi stori sk Foreni nq
Copenhagen 101 s 1-144.
BODSALIS, M- G. & MANKAU, R, <1965). Parasitism and
predation of soil micro-organisms. In K- F.
Baker 8f W- C. Snyder <eds). Ecology o-f
soil borne plant pathogens. Ber kl ey ,Uni v . o-f
Cali-fornia P r e s s pp 3 7 4 - 3 9 1 .
B R O W N , R, H. S< K E R R Y , B. R- < 1 9 S 7 ) . P r i n c i p l e s and
practices o-f nematode control in crops.
Academic Press, New York, 447 pp.
BUT8CHALT, 0. (1878). Beitrage zur Kenntanias der
Plagellal-en und einigpr verwandten organisman,
2. Wi'^s. 7ool . 30 J 205-281.

-•> 's r w.' iy' '^
CAhSNING, E. U. (1973). Protoroan parasites as agents
for biological control of plant parasitic
nefoatodeB. Nematol oqi ca , 19 : 342-34So
COBB, N. A. <J9!7). The mononchE (Mononchus^ Bastian)
a genus of free living nematodes, Soi I Sci . 3 ;
431-4SA.
COBB, N. A. n 9 2 8 } . Transfer of nematodes (mononchs
from place to 'place for ecorjomic purposes.
Science^ 51 s 640-641,
COHN, E. ?. MORDECHAI, M- <1974>. Experiments in
suppressing citrus nematode populations by use
of merigold and a predaceous nematode.
Nematol. Medit, 2 : 43-53.
COOMAMB, A, ?< LIHA, B, (1965). Description of
Anatonchus ami ci ae n, sp. <Nen)atoda j
Mononchidae) with observations on its juvenile
stages and anatomy. Nematoloqi ca, 11 s 413—431.
DDNCASTER, C. C. ?* HOOPER, D, J. <1961). Nematodes
attaclted by protozoa and tardigrades.
Nematoloqi ra, 6 i 333—335.
DOUCET, M. E. <1980). Description d'une nouvelle
t»spe{:p de genre Myl onchul us < Nema tod a s
Doryl ai mi da> . Nematol . Hedi t , 8 s 37--42.

w V, '••'
ESSER, R. P. <1963), Nematode interactions in plates
o-f non-sterile water-agar. Proc. Soi 1 Crop Sci ,
Fla. 23 ; 121-138.
ESSER, R. P. <19S3>. Amoebic predation upon
nefnatodes. Nematol oqy Ci rcul ar , 98 i 2pp.
ESSER, R. P. <19S7). BiolDgicel control of nematodes
by ne^setodes I. Dorylaims <Nematoda s
Dorylairi)ide> . Memat ol oqy Ci ret.'} ar 144 jpp4.
FERRIS, V. R. n968>- Biometeric analyses in the
genuB Labronema (Nematoda : Dory1aimi da> with a
discription of L. thornei n. sp. Nematoloqica ,
14 J276-284.
GOODEV, T. ?. TRIFFIT, M. J. <1927>. On the presence
o-f •flagellates in the intestine o-f the nematode
Di pioqaster 1onqi cauda. Protorooloy, i : 47—58
GROOTAERT, P., JADUES, A. S< SMALL, R. W. (1977). Prey
selection in But Ieri us sp. (Rhabditida s
Dipiogasteridae>. Med. Fac . Landbouww.
Ri jksuniV. Gent. 42 5 1559.
GROOTAERT, P. St SMALL, R. W. (1982). Aspects of the
biology of Labronema vulvapapi11atum (Meyl)
(Nematoda s Dorylaimida) in laboratory culture.
Bi ol. Jb. Dodonaea, 135-148.

1 r 7
GRDOTAERT, P. 3, WVSS, U. <197a). The ul t r astruc ture
and -function o-f the anterior feeding apparatus
in Labronema 5p. Nematoloqica, 24 : 243—25B.
HECHLER, H. C. <19.^3>. Description, developmental
biology and -feedinrj habits of Sei nur a
tenui caudata (De Man) {J. B, Goodey, 1960
<Nematoda j Aphelenchoididae>, a nematode
pr&dator, Proc. Helmi nth. Soc. Wash. 30 J 183-
195,
HECHLER, H. C. ?< TAYLOR, D. P. <1966>. The life
h i s tnr i eB of Sei nura eeleri s, 5. oli vei rae, S.
o>iura and S. stei neri (Nematoda :
Af>hBlr?r.ch{3ididae> . Proc ._ Hel tni nth . Soc . Mash . ,
33 : 71-B3.
HOLLIS, J. P. <1957). Cttlture studies with
Tri chodoruB chri stei on tomato roots. J.
Nematol., 5 s 228-229.
JAIRAJ^URI, M. S. Sc AZM3, M. I. <1979>- Some studies
on the predatory behaviour of Mylonchulus
denhatus. Nematol Medit., A s 205-212.
JAIRAJPURT, M. S, ^ 8TLBRAMT, A. L. (1990). Predatory
nematodes. In Nematode Pio-cDntrol s Aspects
and Prospects (Eds Jairajpuri et.al.> CBS

3 C 8
P u b l i s h e r s & D i s t r i b u t e r s P v t , L t d . , riB,>3 3':?1 h i
pp 9 5 - 1 2 5 .
JATALA5 P- U 9 8 ^ } . B i o l o g i c a l c o n t r o l o f p l a n t
p a r a ' i i t i c n e . n a t o d s s - A n n . R e v . P h y t o p a t h o l . 24 :
4 5 3 - 4 8 9 .
KHAN, Z., BIL6RAMI, A. L. ^ JAIRAJPURI, K. S. <1991).
Sime '=:tudies on the predation abilities o-f
Aporcel ai mgl 1 us ni vel i s <Ne<natoda :
Dor "/I a J/ni da > . Nematol ogi ca i 37 s 333—342.
KULSHRESTHA, R., BILGRA«I, A. L. ^ KHAN, 2. <1993).
Predation abilities of Mylonchulus mi nor and
f^sictors influencing predation. Annal 5 of
PI ant Protect i on Sci ences (in press).
LINFDRD, M. B. (1937). The feeding of some hollow
B.tylf>t ne/R.=^todeB. Proc. Hel mi nth Soc. Wash. , 4 ;
41-4A.
LIWFORD, M- B. ?< OLIVIERA, J. M. (1937). The feeding
of hollow spear nematodes on other nematodes.
Science, 85 : 295-297.
MAERTENS. D. U97<?^>. Observations on the life cycle of
Pr i onehulus pnncf atus (Cobb, 1917) and cultural
conditions, Biol. Jb. Dodonaea, 43 s 197-19SQ.

3C9
HAHAPATRA, N. K. & RAO, Y. S. (1981>- Bionomics of
lotonchus monohystera a predacectus nematode in
rice 5Di1, Proc. Indi an ftcad. Parasi tol- , 2
: B5-87.
HENZEL, R. <i92S>. Ub&r die Nehrung der freilebenden
Nematoden und die art i hr er Au-f nahme. Ein
Eeitrag zur Kenr<tnis der Ernabrung der
Wuc msF - Verhandl . Natur•f . Ges. Basel . 31 s
153-J8S.
MOHANDAS, C. & PRABHOO, H. R, <198(3). The -feeding
behc^viour eir^d food pre-ference o-f predatory
nematodes (Hononchida) -from the soil o-f
Ker fil a , I rtdi a . Revuf? d ' Ecol oqi p et de Bi ol oqi e
du Sol . , 17 i 153-160.
HULVEV, R- H. il9al). The Hononchidae a family of
predi=»ceous nematodes I, Genus Myl onchul us
(Enoplida ; Nematoda), Canadi an J. Zool. ,
39 J 665-696.
NELMES, A. a. <1974>. Evaluation of the feeding
beiiaviour of Pr i onchul u5 punctatus (Cobb), A
neinr^hode predator. J. Ani m. Ecol . < 43 : 553-565-
NELMES- A. J. ?< M C C U L L O C H , J . S . < 1 9 7 5 ) . Number of
tfjononchid nematodeB in soil sown to cereals

2 i 0
and grasseB. AnnIs. App. Bi ol. <79 r 231-242.
DVERGAARD-NELSEN, C U 9 4 3 ) . Studies on the soil
iTti c cf aiipB I. The nrtosB inhabiting nematcdeB and
rotifers, pL'bl ._ Soc. 5c. Lettr. d ' . Aarhus.
ser.5c. Nat. 1 : 1-9S-
OVERGAARD-MIELSEW, C. <1949). Studies on soil
micro-fa ?jn^ TI. The soil inhabiting nsinatodes.
N^tura jHtl andir<=^ 2 t 1-131-
BHAFQAT, S., BILGRAMI, A. L. ?. JAIRAJPURI, ». S.
(19S7). Evaluation of the predation ability of
Dorylai mus st aqnal is Dujarding, 1345 (Nematoda j
Dory1 axroida> . Revue Nereatol, , IB i 455-461.
SMALL, R. W. <\987>. A review of the prey of
predatory soil nematodes. Pedobi ologi a , 30 :
179-206.
SPRINGETT, J. A. U964>- A method of the culturing
Enchytreids. 0i kos, i5s 175-177.
STEI.MER, G. ?< HEINLEY, H. <1922>. The possibility of
control of Heterodera radi c i col a and other
plant injurious nemas by means of
prefiatory nemt^s specially by Hononchus
papi11 atus Bastian. 3, Mash. Acad. 5ci. , 12 :
267-385.

31 1
SZCZYGIEL, A. U 9 < ^ 6 ) . studies on the fauna and
population dynamics o-f nematodes occurring on
strawberry plantations. Ekol . Pol. , 14 : 651-
709.
BZC7VG1E'»_, A. (19"?' \. Wystep'owani & dr apieznycb
nicieni z rod^iny Mononchidae w glebach
uprawnych w polsce. Z * ? ^ ?- o b 1 . Post ec.
Nf>ui - -o2n, , t21 t t45-t58.
SAYFE^ R. ^5. <1973>. 7r- e"^ ^f'- o<n v •; P webr i , an amoeba
prs'dahory on plant parasitic nematode. J.
U^J^:^±S:J_. ^ 5 I 258-264.
THORNE, S- <I924>. Utah nematodes of the genus
f-'ir.'-i'jir.r He . Jrercr:^, A-n, H^ cr e' cr-r- . ?r'C . , 43 i 157-
171.
THDRNE, P. 192S>. W6>^atode5 inhabiting the cysts of
the sugar beet nematode (i-!et f r oder g ? ch achl i ) .
J , A<Tf i r . Research . , 37 : 571-575.
THDRNE, G. (1932). Apparaot successful transfer of
Hrcrtf'pr b<." ppp^llatus, H . ff' u •= c c i u m and M.
Amn li 1 noH < .r.'iJS- from Utah to California. J .

o I 12
THHRME, G. ?< SWAfJER, H, H. il93<b>. A m o n o g r a p h o-f t h e
n err; a t o d e g e n e r a r> --> r v 1 ^ i -•;> M =5 , A ri* f j r c ? • ^ •. f>tu 3 ,
D""" y " ir- ^ ff>o": df^-e a n d p j i r i g e n t >.',-3 ,- CFIp i t fj- 2 _ o o j _ . , 6 :
W E B E R , A . P . , Z y i I L L E r 4 B E R G , L . 0 . S< VAhS DER L A A r 4 , P. A .
t3 9 r > ' < ^ , A p r e d a c e o u B afnoeboid o r g a n i s f n
d e ^ r r c j y i n g I p i r v a s or t h e p o t a t o r o o t
WEBSTER, J . M. ( 1 9 7 2 ) » f'te-rt^atade i<riCj B i o J o g i c a l
conf r-ol . In W e b s t e r J , M. , Eccjno<?jic Nesnat D1 o g y :
Academic P r e s s New Y o r k . 563 p p .
WOOD, P , H. ( 1 9 7 3 ) . F e e d i n g r e l a t i o n s h i p of s o i l
d w e l l i n g ne-Tta todes , Fc-i 1 B i o 3 . Bi ochem. 5 ;
5 9 3 - 6 8 1 .
WYSS, U. 3. GROOTAERT, P , ( 1 9 7 7 ^ F e e d i n g m e c h a n i s m o-f
Lfubr anf^-n.tt s p - Hed •• Fac , L aodboM w»<? R i j »- s u n i v .
Geo; . , 42 : 1 5 2 1 - 1 5 2 7 .
YEATES, G. W. (19,5.9). P r e d a t i o n by Hcmo -r_ h p j d e s
P o t - i> " t; " '-- (^4em.^tcdrt : D i p i o g a s t e r i d a e > i n
labo<'af<Trv c u l t u r e . ^•i(T>:-.-< r T 1 <-»-i ' f - , 15 : 1-*^.

3 1 3
VEATES, G. W- (1971). Feeding types and feeding
groups in plant and soil nematodeB-
Pedobj Q } oqji. e .. 11 s 173-179.
VEATES, G. W- <19e7>- Nematode feeding and activity:
the importance of devel np^nent al stageB. Biol.
Pert. S o i I .

3 1 4
CHEMD-ORIENTED SEX ATTRACTION IN A PLANT PARASITIC NEMATODE, HIRSCHHANNIELLA ORVZAE
By
OUDSIA TAHSEEN and ANWAR L- BILGRAMI
Section of Nefnatology, Department of Zoology, Aligarh Muslim University, Aligarb-202002, India
Abstract: During the present observations on the 5B>! attraction behaviour of H. or ys ae it was observed that both the se^es produced attractants which resulted in a mutual response towards each other-Males of H. orvz ae were more responsive than females. There was no attraction of maleEi towards male attractants and females towards female attractants-The distance of nematode inoculation and the period of nematode e>;posure to attractants influenced the degree of attraction. Significant correlations were observed between the attraction <Mean Score) and the period of nematode exposure to attractants.
Key word: Sex attraction, behaviour, H. oryz ae.
Many studies were made on reproductive and .sex
attraction behaviour of free living nematodes- Greet
(1964) studied sex attraction in a free living
nematode, Panaqrellus ri qi dus. Later, Jones (1966)
observed attraction of male: Pel oder a t er es .towards
females. Ahmad tt Jairajpuri (19S0a, 1931a) observed
sex attraction in Chi 1oplacus symmetr i cus and
Cruz nema 1anbdi ense respectively. They have also
studied the effects of various factors on the sex
attraction of C. symmetr i cus (Ahmad ?< Jairajpuri,
1980b) and copulatory behaviour of C. 1anbdi ense
(Ahmad ?< Jairajpuri, 19Slb>. Fewer observations are
available on the reproductive flr^d sex attraction

behaviour o^ plant parasitic nematodes. Green <1966,
19£>7> and Green S< Plumb (1970) reported the presence
D-f male attractants in Gl oboder a rostochiensi s and
HeterOder a schacht i. Green and Greet (1972) showed
that attr-actirta substances were secreted -from the
entire body DT G. rostochiensis and H, schacht i.
Greet et al . {1968) observed that males o-f B.
rostochi ensi s were attracted more strongly towards
•females- Armi & Jairajpuri <1977> reported that male
attracting substances also ocurr in active -females of
HoplP1ai mus i ndi cus.
In the present work observations were made on the
sex attraction behaviour of Hi rschmanni elI a oryz se.
MATERIALS AND METHODS
Adult Hirschmanniella oryz ae were isolated -from
soil collected around tbe roots D-f paddy (Prys a sat i va
L-) by seiving, decantation and Baermann's funnel
methods- T!»e nematodes isolated were washed with tap
water repeatedly. They were sterilised with 0.1%
solution of mercuric chloride prior to experiment.
For each experiment fresh specimens of H. oryz ae were
i BoJ ated.
Dbser va h i on"? on the se:.- attraction of Hj; oryz ae
were made in 6 cm diameter Petri dishes with a 5 mm

u I (. D
thick layer of 0.5% water agar. The Petri-dishes tnere
marked into three zones (outer, fniddle and inner
circles) at the bottom by drawing two concentric
circles o^ 1 and 2 cm radii., fires \ r" > o-f each zone
was calculated Bnd weighting -factors were obtained by
dividing the sr&a of outer zone with each o-f the three
zones (Table—I),
Table - I i Area and weighting factors o-f the
three zones marked on the Petri—dishes-
Inner zone (Sq, cm)
Middle zone (Sq. cm)
Outer zone < Bq, cm)
Area ;. 34
Weighting 5.0 Factors
9.42
2.0
16.0
1 .0
Weighting factors are nearest to whole number
Scores were than obtained by summing up the
products of the number of vjorms present in each zone
with their corresponding weighting factors. Mean
scores <MS> were calculated for each e>;pBriQent and
represented in Figures I A-F. Each e>:periment was
carf ied out at rooin temperature 23 + 2 C and
replicated five times.
Attraction of males was tested towards fnale and
female secretions and of f t?fsia 1 es towards f en>a 1 e and

o ^ 7 o
male secretionE- Sex attractants were obtairsed by
placing 50 individuals o-f each in 2 ml o-f distilled
water for 24 h in a cavity block. One drop of
attractant was placed in the centre of the inner
circle and ten individuals to be tested were released
at various points of the periphery of the outer circle
o-f the Petri—dish- The distribution o-f nematodes in
each zone was noted after 4, B and 16 h,
Similarly the attraction of male and female H. oryae
was tested by releasing thero in at the periphery o-f
the middle and inner circles- In control e>!periment5 a
drop of water without male and female attractants was
used-
RESULT
The present observations on the 5e>! attraction
behaviour of Hi rschmar^niel 1 a or yz ae show that there is
a positive response of male individuals to female
attractants and female individuals to male
attractants. The males were more responsive than
females. Ma>;imum attraction v*as observed after 16 h
and there was a significant influence of the period of
exposure of nematodes on their responses. Males
towards male and females towards female attractants
were rarely attracted. The distance of the
inoculation of male and females also influenced the
ilt t rt * nf attraction.

8
Attraction o-f males towards -female attractants (Fig.
1, B>. t*ihen male i ndi vi dtjai EJ O ^ U . c- •" y i. a e were
released at the periphery of the OLiter c^rcl^ they
showed a gradual increase in the attraction from ^ h
<h5 - 3.66) to 16 h (M5 = 10.46, p < 0.05). ivhen
tested -fro/j) the mi ddJ e i::iri:.'sEf n&mBtadeE e>;hibited
stronger response towards i^emBle attractants (p <
D.D5>. Attraction was minimuro after 4 h fMS = 7.46)
while it was may, i mum after 3 6 h ^HS = ID-43) (p <
D-D5>. The attraction of male individuals was more
than the control ( p < 0.05) whert tested from the
inner circle. Ma>i a mura attraction was recorded after
16 h <H3 = 15.40; p v 0.05> while winiinum after 8 h
<M3 = 1 5 . 2 6 ) . There was a significant correlation
between the attraction and period of e;'.posure when the
nematodes were inoculated in the outer ^r = + 0.65, p
^' 0 . 0 5 ) , middle (r = + 0-^0, p < 0.05^ and inner
circles <r = + 0.93, p < 0 . 0 5 ) .
Attraction of males towards male attractants (Fig. 1,
C ) : Male H. o r y •; a e when tested towards male
attrertants there was no significant difference in
th^^ir response <p > 0 . 0 5 ) . Inoculation of males at
th)e periphery of either outer, middle or inner circle
f i a rJ not evrd:e any po'^iLive r e^^poriE^e. Hov;e\ pr , males

3 1 9
showed a significant negative correlation with the
period of ei:poBure (r = - 0-93, p < 0,05) when placed
at the inner circle. In other cases the coe-F-f i ci ent
o-f correlations were insignificant ( p > D. 05> ,
Attraction of females towards male attractants (Fig.
1, E>: Attraction of ferijales towards mstl e attractants
declined with the period of exposure < r = — 0.62, p <
0,05) wljeff tt?sted from the inner zone. Maximum
attraction was recorded •r 4 h iMS = 15.2) while
the least after Ih h (MS = 1 3 - 9 ) . The inoculation of
females in the middle circle ^'^oked more response than
the control ( p < 0.05) but the period of exposure did
not have any sionificnt effect on their response to
attractants (r = + D.4S, p > 0. 0 5 ) . Though, there was
no positive response of females to male secretions ( p
> 0.05) wh:en ttiey were tested from the outer circle
but a positive correlation was evident with the period
of exposure (r = + 0-79, p < O.D5>.
Attraction of females towards female attractants (Fig.
1, F>: Female H ^ oryz ae did not show any positive
resfjonse towards female hormones in either of the
conditions ( p > D . D 5 ) . Mean Score declined with the
increase in thf period of exposure (r = ~ 0.74, p <
0,05) wh{?n tested from the inner zone. It

u i. U
increased with the increase in the period of exposure
ir = + 0.95, p < 0-05> when -feroaleB v*ere released at
the periphery of the outer circle. Inoculation of
-females at the middle circle did not set any pattern
ir = -i- 0.44, p > O.G5> .
DISCUSS.T0r4
During present observations on tbe* se,-; attraction
behaviour both the se>:es of H. or yz ae produced sew
attractants r*hich resulted in a !T»utuai response
towards each other. A similar behavioiir has also been
observed in Panaqrol aimus r i o j d t< s <Greet, 19i4) ,
Panaqrel1 us si 1usi ae <Cheng ^ Samoiloff, 1 9 7 1 ) , P.
redi vi vus (Duggal, 1978) and Cruzrfems 1 ambdi ense
(Ahmad ?< Jairajpuri, 1 9 8 1 a ) . Males of H. oryz ae
appeared more responsive than their females as was
also observed by Ah/nad Sr Jairajpuri • (1980a) in
Chi 1oplacus symmetr i cus and Greet et al. (1968) in G.
rostonchi ensi s. No response of males and female to
their own attractants suggests that these individuals
perceive attractants of opposite sex n;cre than their
own body secretions and that they 3r t^ specific in
their attraction responses. Theoretically, the
attraction should be more when the nernatodes were
released at the periphery of the inner circle.

0 9 1 hr/ Ilk, I
However, in all such the e>i per i ment s except Fig. I B
^attraction of fnales to -female attractants) the dBore-B
of attraction {Mean Score) decreasecj after every 4 h-
The reason could either be the attract ante. had a
little effect on the nesj-atodes to change their course
frofn 5uch a E-hort distamze or neifiatodes were repelled
due to high attraction gradients. Due to these
factors the jTtovement of nematodes from the inner
circle aj:«peared more random similar to control where
t^tean Score declined after every 4 h. In all these
e>! per i Rient B negative correlations were obtained
between the attraction and the period of nematode
exposure to attractants. • In rest of the cases where
the nematodes were inoculated either at the periphery
of the middle or outer circle the relationships were
positive and there was an increasing trend in the
attraction of nerrtatodes.

w X. >_
REFERENCES
AHMAD, I. ^ JAIRAJPURI, ^5. S. <1980a>. Se>: attraction
and copulation in Chi 1 opl ecus symmet r i CUE-
(Mematoda J Acrobelinae>. NematoloQica 26 ; 139-
148,
AHMAD, I. , JAIRAJPURI, H, S- (19SDb). Factors
in+luencing sex attraction in Chi 1oplacus
s y <n me t r i c u 5 . I n d i art Q^. Nemat ol . t 0 j 107 — 117.
AHHAD, I. S JAIRAJFMJRI , H, S- n 9 S l a } . Sex attraction
in Cru^nema 1 anbdi ense <MeRjatoda ; Rhabdi t i dae) .
Revue Nematol . A_ ; 145-150,
AHHAD, I. ?,- JAIRAJPURI, H. S. (193}b>. The copulatory
behaviour of Cruz nema 3 ambd i ense (Newatoda ;
Rfjabdi ti dae) - Revue Nematol . 4 ; 151—156.
A Z H I , M, I. & JAIRAJPURI, Vt. S. <1977>. Chemo-
oriented 5e>; attraction in Hopl ol ai mus i ndi cus,
Indi sn J . Par asi tol . i_ : 63-64.
CHENG, R. & SAMOILOFF, M. R. <1971>, Sexual
attraction in free living nematode Panaqer alius
5i 1 t4s i ae < Ceph a 1 ob i dae > . Can . J . Zool . 49 :
1443-1440.

•? 0 o v/ i . >J
DUGGAL^ C. L. (397S} , Sex a t t r a c t i o n i n r r e e l i v i n g
netTiatode P^r^ p,c\r tal 1 u a r -e H i v :t v t.< s . NematoloQi ca 24
GREEN, C, D- < i966>. O r i e n t a t i o n o^ male He tg rodera
r o s t onch i pns i s. Wol 1 and H e t e r o d e r a 5chacht i i
Schfij t o t h e i r f e m a l e s . Ann. Apr?l . Bi o l , 53 ;
327 -339 .
GREEra, C. D. M967> . The a t t r a c t i o n of male c y s t
nematodes to t h e i r f e i na ies . ^4e<i^atol og i c a 13 ;
172-173.
GREE?4, C. D. ^ GREET, D. N. < i972>. The l o c a t i o n s of
s e c r e t i o n s t h a t a t t r a c t male Ne te rode ra
schacht i i and H. r o s t o c h i en s i s t o t h e i r fesBales.
Newato loq i ca IS : 347—352.
GREEN, C. D. ?< PLUMB, S. C. <1970) . The
i n t er r e l at i onsh i ps o f 5D/ne H e t e r o d e r a spp . as
i n d i c a t e d by t h e s p e c i f i c i t y o f t h e male
a t t r a r t a n t s e m i t t e d hy t t i e i i - f e tna les .
N >? m a t o ] c) q i c: a 1 A i 3 9 - 4 o .
GREET, D. N- < i964>. O b s e r v a t i o n s on sexua l
a t t r a c t i o n and c o p u l a t i o n i n nematode
p^t)r>uer o l a i mu'? r i g i riu s ( S c h n e a d e r ) . Na t ur e
<l l.ndu^^ 204 : 9 6 - 9 7 .

24
G R E E T , D . N- , GREEN^ C, D . ?f P O U L T O N , M. E , U 9 6 S > .
E x t r a c t i o n , s t a n d a r d i z a t i o n a n d a B S S B B F i e n t D t
t h e v a l i d i t y o-f 5e>i a t t r e r t a n t £ o f H B t e r o d e r a
r o-E-t o c h i e n g - i 5 Viol I a n d H . s c h a c h t i t S c h m . A n n .
Ar . r« . B i o l i . fcl s 5 1 1 - 5 1 9 .
J O N E S , T , P . < I 9 6 6 > . Se>: a t t r a c t i o n snd c o p u l a t i o n i n
P<B1 O d e r a t e r e s . N e w a t o l o q i c a 1 2 : 5 1 S - 5 2 2 .

2 5
LEGENDS
Fig. 1-
A - Attraction or males in absence o-f females <CDntro3 >
B — Attraction o-f males towards T^emale attractant;
C — Attraction of males towards msls attractents
D — Attraction o+ females in absence of males <Contr ol )
E — Atraction o-f -females towards^ male attractants
F — Attraction of males towards ffiale attractants

15 1 \ ,
y^y%t<A 12 x—x Inner circle--^ x
o—oMiddle circle Outer ci(
81
8 -12 16
TimG(h)
i5'
8. r=~058
^=-18.5+130.25 o
o •if
or=0.84 X=6.5y-21.07
12
16"
r = 0.9 S x^7.0r2:-.G,P<0.05
A ~ T ; p-
8 12 Tinne(h)
16
16

3 2 /
Predation Abilities of Mylonchuhis minor and Factors Influencing Predation
R. Kulshrestha, A.L. Bilgrami and Zakaullah Khan
Section ofNemalolo^; Department of Zoology, Aligarh Muslim University, Aligarh 202002, India
ABSTRACT
Observa.ions on hlyionchiihis minor revealed Ihal Ihe prey talchjng and feeding mechanisms are comparaDle to olhcr mononchs. P/tdiiion by M. minor depended on ctwnce cncounlcra wild Ihe prey, P/cdaiors fed by culling the cuticle and shredding )hc prey in pieces. There was no set predatory profile and adult females were more active predators than Ihe young juveniles. M. minor preferred second stage juveniles of Meloidogym incognita and Tylcnchulus semipeixctrans when tested singly or in combinations with olhcr prey nematodes. Number of prey, letnperalure, agar concentration and thickness and starvation of predators affected predation by Af. minor.
Key Words i Mylonchulus minor, Nematoda, Predation, Predator, Prey.
The possibility of using r )nonchs against plant parasitic nematodes was speculated by Cobb (1917). Cohn and Mordechai (1974) studied the effect of Mylonclmlus sigmatunis on citrus nematode, Tylendmltts semipenelrans. Further investigations on predatory behaviour of mononchids (Grootaert and iMaertens, 1976); ultra structure and function of feeding apparatus (Grootaert and Wyss, 1979); predation ability (Small and Grootaert, 19S3); effect of mechanical stimulation and crowding on predation (Azmi and Jairajpuri, 1979); factors influencing predation (Bilgrami et al., 19S3; 1984); cannibalistic behaviour (Bilgrami and Jairajpuri, 1985); inlraspccitlc interactions (Bilgrami et al, 1988) and strike rate (Bilgrami, 1992) have been reported.
In the present investigations, the prey catching and feeding mechanisms of Mylonchulus minor and factors influencing predation have been studied.
Materials and Methods
Mylonchulus minor was cultured in Petri-dishes containing \% water agar using Rluibditis sp. as prey. A small amount of milk powder was spread over the surface of agar to encourage the growth of bacteria to serve as food for the prey nematodes [Rhabdiiis sp.).
All experiments were performed in small cavity blocks containing one per cent water agar at 28 + 2°C. Twenty-five individuals of Meloidogyne incognita were subjected to predation by five .^f. minor. The number of prey killed or injured was counted after 24 h. Each experiment was replicated five times. Conditions remained same for all experiments unless mentioned othervvise.
Prey catching and feeding mechanisms of j \ / . minor were studied by inverting the Pctri-dishcs containing M. minor and prey nematodes under stereoscopic binocdlar microscope. To studv
the feeding mechanism at 100 X or higher
Armiils of Plant Protection Sciences 1(2) : 79-S-l {1993) 70

a Kidshrctha A! lUi^rami and 7akaul nn Khun
328
nng iific^' ons, tcii dd JILS of A/ rr^rt^r were phccd separately in observnlioii chambers wilh 1(X) specimens of Rliabditis sp. ab prey. Tlie observalioii chambers consisted of a glass shde with a 2 X 1 X 0 5 cm block of one per cent water agar (Bi'granu et a!, 1985) Predator and nrey were left together and agar block was then covered gciill) ivilb a cover slip The ncnintodcs were allowed to accli-ni.iti7e for 20 nunutcb bclore crscrvalio.is were recorded To find out the rale of pred?lion by adult and ju\ciiilcs of M minor, eacn stage was p'ace separately in cavity blocks containing water agir with second stage juNcniles o[ M. incognna as prey.
To determine prey preference of M. minor, second stage juveniles of A/, mcoqniui, T. senipene-trans, Angiava trtiici and adults of Hoplolaimtis Indian, Xipluncma basin and Rhubditis sp were used as prey Each prey was subjected to preda-tion by M minor in separate ca\il_\ blocks.
Prey selection in combination : To study the prey selection by Af, minor 15 two species combinations (T.ible l)ofA/ incognita, A. tntici, H. mdiciis, X. americaniim and Rnchditis sp. were subjected to prcdation. In each combination 25 prey of each species, total 50 individuals, were used. The predatory profile of Af. minor o\er 10 days was stuuicd by Irnnsfcrnng the predators to new ca\ity block with the same number of prey on fresh agar. The effect of starvation up to 10 days and relations between the number of prey and temperature from 5 to 40°C with rate of prcdation were also studied.
The cflcct ol agiir concentration Irom 1 to 6% and thickness from 2 to 14 mm on the prcdation of A/, minor wcTC also observed.
Prcdation depended purely on chance cmouiUcrs with the prey with no evidence of chcmo'.aclic activity. During locomotion predator contacted prey, established lip contact and moved its lip vigorously over prey Alter probing the prcditor attacked prey with its buccal armature to acI/Kvo cuticle perforation. All a'lacks did not result in killing the prey as \er)' r i tm the prey moved out of the grip of predator The conlnciion of labial muscles pulled the I'ps outwards and backwards so that the mouth and the vestibule became wide open (Coomans and Linn, 1965). As a result the dorsal tooth and the denticles became fully exposed and ready to attack and puncture the cuticle of prey. The prey became miinobile as soon as it was punctured, shredded into pieces with the help of buccal annafure and consumed by the predator. During feeding the prey was held by the tooth, denticles and lips of the predators. The food which consisted of body fluids and internal organs of prey was ingested by repeated muscular activity of oesophagus and feeding apparatus. Once the prey was completely consumed the predator left the feeding site cither in search of another prey or moved randomly in agar.
RATE OF PREDATION BY ADULT AND JUVENILE STAGES
All Stages of Af minor vvcre predaceous in nature. Females killed the maximuiii number of prey (p <0 05) and the first stage juveniles the least (Fig. 1).
PRT/SELECTION
Results
PREY CATCHING AND FEEDING M E C - L A M S M S
M. minor moved in a topical urdulator} wave pilScrn m Tg.ir m the presence cf prey nematodes.
When phccd with ditfcrcnt species of prey iicmntodes A/, minor killed the niaximuni numbers of second stage juveniles of A/, incognita ard T. scmipcnetrans in comparison to other species of prey (p < 0 05). X anicricaniun was predated upon least by the predators (p < 0 05) Other
so Awwh rfrijnl Pro'^i I on 5. •enca .'(-"I 79 i I (lOOJj

329
rri^iat cii chilili^oj ^f) oncl tt is r-ii or
90
?70-
;5o-
0-30
20
10
JA J3 J2
Fig 1 R;ilcofprcaaiionbyadullandju\cnilesiagcsof \A/om:A"/ minor
species of prey viz^/ / . indicns, A iriiici and Rliabditis sp were killed in moderate numbers (Fig 2 and Table 1)
Pr V com
P>
M! M P 1 M' A" RH PI lU HI HI TS TS TS
•s ^
+ + +•
+ + + + + + f
+ 4-
+ + +
Tabic Prey stleciion by \f
binalioir;
P.
AT RH AT XA XA X.\ XA MI AT RH HI AT MI XA RH
1 minor in \ a rc —
conibmaiions of ptcy
Per cent kjlUng
P,
84 SO 64 84 72 60 40 20 40 2S 84 80 52 76 72
P:
40 36 44 32 24 32 52 80 64 72 20 44 72 28 40
S i p J
d c v p
P>
34 32 28 35 3 I 26 1 4 1 1 1 6 19 2 9 2 4 2 0 2 3 30
i r d
en
P
1 9 18 2 ' 1 5 1 9 2 2 2 3 19 2 1 2 6 2 0 1 6 1 6 19 2 I
P| = first prey, P, = Second prev MI = Melo Jo!:;\ne irco^i iia AT = Angmna tniici RH = RhitMns sp, XA = Xiphmana amencanum HI = Hoplolaimus mdicus, TS - Tylciich ilus scmipt:nclrans
100
90-
80-
70-
c60
;55o-
"?iO
^ 3 0 a. 20
10
r.g 2
I i i. A
v j
C D Type of prey
i
R\li.ofi icJjuca by Af>/o/ic/»/iij/iii/ici-ondiffi-rcnl species
of prey
In various Iwo species combinations of prey ncnnlodcs M minor showed diffcrcnlnl degree of preference The second stage ju\cnilcs of M incoi^ntta and T semipenetrans were killed most b^ the predators in all combimtioiis (p <0 05) M miror least preferred X amencanum and H indiats m ail combinations of prey A tritici and Rhabdiiis sp were preferred inodenlei) by the predators
PRLDATION BEHAVIOUR
There was no consistent predatory profile o\er 10 di\s Predators killed minimum number of prey after third (76'^) and maximum aficr fifth (9lQ) and tight diys (91'^) of prcdilion Tlitrc was no sicuificant dillcrcnce (p < 0 05) in t c ntc of daily killmg t>et\vecn the first and tenth <^^^ (Fig 3 \-C)
Amnis of Plan! r euioii Su<:nc<rs 1(2) 79i^(JP9?j Si

R. Kiilshrvllm, A L. Hilgram and ZakaiiUnli Khan
lOCi
en
£ 70
fl6.
c U-j
c .o82-
£ 80-
8 5-,
75
.S 65'
ix55' •*->
£ A 5 -o a3 35-
Q.
25-1
15
A
-1 1 1 r 1 1 1 1 i 1
2 4 6 8 10
Number of days
"so ' 1^0 ' 1^0 ' 260 Number of prey
E
— I 1 1 1 1 ,
1 2 3 ^ 5 5 Agar concentration ('/o)
1C0-
90-
80-1
70
t.
100
90 -
80-
70
50
50
iO
30-1
T 1 1 T-2 A T^
starvation period (days)
1 — I 10
15 20 25 30 35
"Temperature °c AO
2 ^ 6 8 10 12 14
Agar thickness (mm) Fii;.-V Tri'd.ilory bch.niourcfA/>'/o/icV»/iijm/ncr:Prcdnlor>'P-itloni over 10d.iys(A); Effects ofst3n'allon(n\ Prey pcpul.ilionJcnsily
(.C), Tcnipcniurc (D), Agar conccnlration (H) anJ Agar tlm-' ncss (1").
6 .: U
82 Annals ofPhml Protccliun Sc/f.uvj IQ) : 70.<;j j.'OpJj

PrcAoticn ahiliuc^of M^lonch-jlus minor
331
The rate of prcdation increased (p <0.05) with an increase in the period of starv'alion. Prey density influenced prcdation by M. minor. Predators killed more individuals of prey in a population of 200 individuals than 25 (p <0.05).
ETECl' OF TEMPERATURE
Temperatures between 25 and SS 'C were better for prcdation (p <0.05) than lower and higher ones iu observations after 12 and 24 h (Fig. 3 D).
EFFECT OF AGAR CONCENTRATION ANT) THICKNESS
Meloidogyne incognita juveniles were killed more in 1-2% of water agar (p <0.05) than at higher concentrations (Fig. 3 E, F).
Thickness of agar layers also influenced the rate of prcdation by M. minor. M. incognita juveniles were killed maximum on 2-6 mm thick agar layers (p <0.05) than in thicker layers.
Discussion
The prey catching and feeding mechanisms of M. minor are comparable to other mononchs except some minor variations. Prcdation by A/. minor depended upon chance encounters with the prey as there did not appear any chemotactic response of predator towards prey. The chance prcdation appears to be a general phenomenon in mononchs. (Nclincs, IS'74; Cohn and Mordcchai, 1974; Grootacrt and Macrtens, 1976; Bilgrami et al., 1983, 1984). Many mononchs such as Mononclins, loiouclius, Cobbonchus, Sporonchuhts', Clarkus etc., arc capable of engulfing their prey whole but species of Mylonclmlus have not been found to contain any species of prey nematode intact in their inicstine (Bilgrami, ct al, 19S6).
Mylonclmlus minor exhibited preference for M. incognita and 7. semipcnctrans when tested cither singly or in combination. Tin's preferential behaviour of M. minor may be attributed to lack of auti-predation adaptation in these nematodes.
Endopsrasitic nematodes lack characteristics which provide resistance against predalion (Bilgrami and Jairajpuri, 1989a; Bilgrami. 1990a, 1990b; Jairajpuri and Bilgrami, 1990). The physical, chemical and behavioural characteristics of ectoparasites probably protect them from predalion (Small and Grootacrt, 1983). Preferential preying has also been reported earlier (Shafqal et al., 1987; Bilgrami and Jairajpuri, 1988; Bilgrami and Jairajpuri, 19S9a; Bilgrami, 1992). Less predation on Rliabdicis sp. and least prcdation on H. indicus may be attributed to their activity and thick body cuticle (Esser, 1963; Bilgrami et al., 1983; Bilgrami and Jairajpuri, 1989a).
The increased rate of predation by M. minor in increasing population of prey indicates that prcdation depended on the number of prey (Bilgrami et al, 1985; Yeates, 1969; Bilgnmi and Jairajpuri, 1990). The increased predalion by M. minor may be attributed to increased chance of contacts between predator and prey. Bilgrami et al. (1984) did not observe any effect of prey density on the rate of predation of Mononclins aquaticiis. The differential rate of predation by M. minor at different temperature? and agar concentrations may be the result of inhibited activity of predators and prey as was also observed by Bilgrami et al. (1983); Bilgrami et al. (1985); Shafqat et al. (1987); Bilgrami and Jainjpuri (19S9b), Khan et al. (1991) with M. cquaticus, Mononchoides longicaudalus, M. furtidens, Aqitatides thornei, Dorylaimus stagnalis and Aporcclaimelliis nivalis. Less predation in thick agar layers may be because the predators and ' prey got more area to move resulting in fewer contacts between them.
During the present observations adult predators killed more prey than their younger stages reflecting their differential predatory potcn;ial. Bilgranti ct al. (1984) also recorded higher rate of prcdation by adult i\f. iiqii.r.iciis than juveniles.
Amuih of Flam Vrouxlicn Sciences 1(2) : 79-S I (/'-'O.'l Sj

(Cc^'OlhOi (J/ ' -"Ji '^ '^i " ^i JI i"'l lI'H""'V IS
6 I ST 't):)'foioiDuat/ 3Jn||nD Xioiiioqi-i ui ( ip i j j bi.8o]ilig iponujisj)
siyjoiod sr^i ),ji .3I/D;V Xq u. p^ij (69til) A \0"^ - ' l " \ .
fcl 611 6c r j 3O/O;PIJ£IJY sopoiLiu^u
XjoiBpsjd 3Qi]|J\\p [los aiuob jo soi|i|iqL uoriLpjid 3 1 uo SDOiiLvio-cqo (eS6l) -JMOOiQ J puc / , \ a •iicras
t9t-fjf 01 '•/o;r>u;PA/?n.Jj/ (LpinirE|UOQ i:--oiEni3Kj) fj-g]'uipjcfriQ SIJVUSDIS snu.}ri^oQ jo ^limi U0i|ip3jd oiji Jo noi; Eilt^^a Csol) undfuitf s 1\ P"E iuiEj3|ig i y ' S li-bjcijs
/• JOiEpaid spoiciusa B '(qqo3) srivpuiid rnirrqovouj
JO jnoiAEqsq Sjip^Dj o ji jo no En|5Ag (t'Z.6l) fV 'ssmisfsj : r t tee i£ t>:i iJioinuoi^ (ipiujiEiuoQ
'EpoiEuisN) 9961 'su(3H (_;j6I 'JsqUV) f ' / cuu iniiiwiDpoJody jo XiipqE uoi tpojd sqi uo suoriEuss -qo 3UJ0S (U'5l) urdfLjiLf g j«j pjE imEiSpg ^ V "^ '"^TO
"fzi jeti' iijisa-^^'N'sJoi"q"»'a puB SJsqsqqn^ gg^ '( spj) jr p undfLJiEf siosdsojj
pun siDsd^ lOJiiioo oig ap^ivui^i^ xs\ sopojEtDja
X;oiEpojj (066l) >uiEj3|ig i y P"8 SW 'undfeJiBf
mJDrniiD smj:>ucruo)^ UI sniEJpJJu Suipsij jopsitre sqi jo
uoipunj pUB sjruDniisEJiifi (fi.cl) sS' /rt. fl P°^ d 'porjoojo 181 ILl Zt VDiSoioivuo\ i7i:iiipnbv snijDuouoji, ]0
sp^o ijq puE LoiiEMiin^ (9/.6I)sneiJ3Bi'ij Q P"^ d UJEJOOJO
K l - l t l CZ ' vij :>o£ loy doj2) I'o^ XJJ JESB J31E/I\ siuais-Doa
JO s3iE|d ui snoipBJsiui spo 6UI3M (C96l) dli 'J 'ssg
XuioieUB puE saSpis 3iia3\'-r sji uo suoiiEAJssqo qi"rt (3BpiqDL0uoj<j 'EpoiE_j3[vj) ds u svioiiuv S7ii/3uo;ouv JO coi|dijoS3Q (5^,51) EOJII g p'lE-y "suBtuoo^
ii VV Z I'l^}'^ loiDi—"^ :poiEui3u sno5orp3jd B puB ppSiJEiu JO 3m Xq suopB ndod 3po)BUi3u stuia 3ai tsjjdJns ui siuamijsdx] (pLt'i itq^spJoj'^ y^ pue -g aqo^
Sot' I£f C ' " ? ! • 5' l.-»p0|ElU3U AJOlCpijd 3ui\ij 33JJ JO sr-oS y st,Duc_{j[i] 3qj_ (i,T6l) 'V'^ 11°D
S31E|d J E 3 E 3,Ii31S liOU UI SqOLO^OLU U3DYi|3q UOriOEJJiaj
(S86l) undfEJiEf S N P"^ pEiuqy I ' 1 y imejSjig
f6I 161 6 'lo)vw3f]3n\3\j -sqouououi auioi jo s)J3iuoo |EJIIS3I0I sqi
JO Xpms V i'jhtl)'^^^^^"'^i S )> P'JE PEuiMy I " l y itLBi3|ig
c9t-ifr lt-S6l)0C'»^'^oi'"»'-^Y (EpoiEui3[\J 'EuiuJicioSXjSj) 3uj^^i Sip )Drby 10 inoi\zi\jC\ <JOI>T)3JJ (^^aij undfiJiBf s,^pLEpEiuqy j -]-y itHEjSiig
fV IV Zl 'vpy{ joivuaj^ siiDiwnbv sii^Duouofy^o JHOKE oq OoiEpsjd 3qi no saoi) EMs^Q (t-;:^!) undfiJiEf t j ; , pUL peuiqy J "["y laiEiSng
9:E JZ£ 9';''"''^^A'""^Jf r; :)j;''nl'« suij:>utiiO}i{ Xq uoi)Lp3jd Suiousnijui SJOpij ='"'-''5 (£S6I ) unifuiif S N P"c pBU'MV 1 " I V 'iu'u3|ig
uoiiipsjd Saionsnijui sjopLj pus (Epu3ist3o]diQ
'i-poicui3[xj) S7j;r rp^iSiwj ;^ pus STopi;vo/r)p)oipi/ouo;y
jos-ii|i|iqE Xjoiipxij (oftoi) undfuiBf g j - ^ puE-] y'miEiS]ig
IZZZZZ (8861)81';°'""'^///""'"'"'/ ^^"^ SpJEWO] UOIlDEillE JI3qi uo put ( rp iJ3 |SE3o |d ig Ep01EtU3{vj)
sii^pujoj i\' puE stiivpiwDiSuoj ^pioi(ouoiio)ij JO jnoUEqiq XiojEpjjd sqi DO JVSB JO s«tt!(orqi puB SUOIIBJ] -U3DU00 JO s)03jJ3 (O686T) pndfEJiBf g JAJ pus -[ y 'iiuiiSjig
(rpiJ3iSE3oidiQ 'EpoiEm3[,j) sioptuoj }•, puB ^iiDpnoDiStioj npioi{oiiouo}i< 'sioiEp3jd sqi jo 3iEi 3)(uis puE uonipsjd 01 XsJd JO 30uqsis3y (B686I) l-indfEJiE/ g }< pus -[ y 'luiuSpg
ZQZ-ib\ I I tOIUUJ3{^3nA3)J UOMOBJIH. 8ui3U3n|jui sJO.acj puB <3Jd spJEwoi (Bpi]3)si3o|diQ 'LpoiEUJ3;vj) sxtipiuo] JYPUB m]upiwoi2uo\ npwijyiiouoj^i JO uoiioEJuy (SS6T) wndfEjiBf g JAJ pne -^ y 'iuiLj8[ig
£0Z-Z0Z
, (t>86l) {"I ' lOjDuoff f uvtpuj snoiiDiibB snqououoyj ui uisijcqiuuB^ ( J 8 6 I ) I-indfEiiEf g y^ pus -q y ')uiBj3iig
QLZ -J9J gi'/Soioivu'7>ff p3iiddypiiv ]vp/)WDpinj aujotji soptivrby^it siimSos snwiDiKuOQ 'snouvribn smfjuoiiojy SJOlBpajd 31)1 JO 91Bi 33|U1S pUB UOjIEfWjd O] £3p01EUU3U
XsJd JO Xiqiqiidjosns puB ODUEisissy (Z66l) 1 V '[WuSpg
et-i et't 'id -'iqioQ -AO^ 'wcnquisiQ 2? sjsqsiiqnj g g ^ ( spg) IV i> iJndfBJiBf 'uo^dsoJj pun si33dsy 'jOJiu03 oig
3po)vw3{^ n] 'sxuiEiXjop sno303p3jj (q066l) T V ''uiEj3|ig Stl £ -1 dd 'njisa A\3N 'SJOinqiJisia y SJsqsriqnj g g 3
'( SP3) ;" r? und EJIE; 's7D3dsojj piw si^x/yy IOJJUOJ oig
3poiDu.~}f^ n[ "sjoiEpsjd pu3isi,3o]diQ (1.O66T) T V 'imajSpg
LOZ £0Z L ' I'P'fV \oiuui3f^ sn}viu}p snin^ouoi^jij uo SuipMOja put uoiiEinuiijs
lEOiutq33Ui jo I03J3 (6i6l) undfuiBf g }\ puE J J\ 'iiuzy
•saippjs assqi loj ODUHSISSB JEIDUEUIJ
3iiipr\ojd joj rqpQ A\0{VJ 'Eipuj jo )U3UIUJOAO9
'ASojouqoax P " B aouoiog 30 luauiiJEdsQ aqi
0) [nj3)Ei9 3JB sjoqjne puooas pus jsjg aqjL
i N i K J o a n A\o t rsD V
uvify{ ijoiini'^02ptiD luivj^iiq fV'vtf^J'f^'i^i i{

" 0 ' Ci J 0
A COMPARATIVE STUDY ON THE PREDATION BY
ALLODORYLAIMUS AHERICANUS AND DISCOLAIHUS SILVICOLUS
(NEHATODA .- DORVLAIMIDA) ON DIFFERENT SPECIES OF
PLANT PARASITIC NEMATODES
By
Z- KHAN, Anwar L- BILGRAMI and M- Sharoim JAIRAJPURI
Institute o-f Agriculture, Sha-fi House, Qi 1 a Road, Aligarb Muslim University, Aligarh-202002, India

3 3 4
SUMMARY
Studies on Allodorylai mus ameri canus and Di 5col ai mus si 1 vi col *»s revealed that these predators are similar in their predatory behaviour and requirements- The predator-prey relationship measured -for one species strongly supports the observations made -for the other predator. The prey catching and -feeding mechanaisms are comprised o-f -five phases viz., encounter wi th prey , att ac k response att ack , e>! tr a — corporeal di qest i on/Sal ivation, end i nqest i on/-f eedi nq. Predators were not attracted towards live and intect f«rey but responded positively towards excised prey. Both species o^ predators preferred second stage juveniles o-f H, i ncoqni t a, A. tritici H. roothi and T. semi penetrans. No individual of Hoplolai mus i ndi cus, Helicotylenchus i ndi cus and H. dhi renders was either injured or killed. Temperature, agar concentration, age of predators, prey density etc., affected predation but starvation of predators did not show any effect. Both predators achieved maximum success on endo-parasitic nematodes (SR = 90/t> - Cortical and epidermal feeders exhibited maximum resistance <PR=55—65%>. Maximum feeding after wounding occurred on endo—parasitic nematodes. Hoplolai mus i ndi cus, Helicotylenchus i ndi cus and H. dherenderi were 100% resistant to predation- A significant correlation was observed between encounters resulting in attack and attack resulting in wounding and feeding after wounding except for endoparasites for A. ameri canus and cortical feeders for D. si 1vi col us. Various physical, chemical and behavioural characteristics are attributed to
different degrees of resistance in prey nematodes. Female predators probed their own eggs when came in contact but attacked, punctured and fed egges of prey nematodes (Rhabdi ti s sp.) Predators took maximum time to consume an individual of Lonqi dorus sp. (FT = 61-68m> and minimum to T. semipeneterans <FT = 22-25m>-
Resume

r> , r
Linford ?t Oliviera (1937) reported Aporcel ai mus,
Nyqolai mus, Sectonema, Labronema Dorylai mus and
Actinolaimus 5pp, as predaceous. Esse <19&3; 1987)
also observed species of these genera besides
Carcharol ai <nu5 as predaceous. Wo detailed studies
v*ere mside until Wyss & Grootaert (1977) and Bilgrami,
Ahmad tc Jairajpuri (1985} gave a descriptive account
o-f the predation abilities o^ a dorylaim and nygolaim
predator, Labronema vulvapapillatum and Aquati des
t hornei respectively. Small St Grootaert (19S3>
observed predation by L. vulvapapi11atum and Shafqat,
Bilgrami ^ Jairajpuri (1987) studied prey catching
and feeding mechanisms of Dorylai mus staqnalis. Khan,
Bilgrami ?< Jairajpuri (1991) used dif-ferent species of
P'lant parasitic nematodes to study predation by
Aporcel ai mel 1 us nival is. Jairajpuri €< Bilgrami (1990)
and Bilgrami (1990) speculated about more species of
Eudorylaimus^ Mesodorylai mus, Aporcelaimus,
Aporcelaimellus, Lai mydorus, Nyqolai mus and Sectonema
as predaceous- Bilgrami (1.993) analysed the
relationships between Aporcelai mel1 us ni vali s and
different prey trophic categories.
In the present work experiments were carried out
to study predation by two species of dorylaim
predators v i z . , AIlodorylai mus amer i canus (Tjepkema,

3 36
Ferris and Ferris, 1971) Andrassy, 19S6 and
Di scpl ai mus si Ivicolus., Sauer and Annells, 1985 using
plant parasitic.nematodes.
MATERIALS AND METHODS
All odor yl ai mus stmer i canus and Di scol ai mus
si 1vi col us were cultured in seperate Petri—dishes
containing IX water—agar using Rhabdi t i s sp. as prey.
Small amount (5-10 mg) of infant milk powder
(Lactogen) was spread over the sur-face of agar to grow
bacteria which served as food for the prey nematodes.
Plant parasitic nematodes <prey) belonging to -four
trophic categories were collected -fresh from different
hosts and localities of Aligarh (Table-I). All
experiments were made in small cavity blocks <2.0 x
2.0 y, 1 cm) containing 5. mm thick layer of 17. water-
agar at 25 + 1 "C. Typical e>!periments were cerr led
out using twenty five individuals of Hirschmanniella
oryz ae which were subjected to predation by five adult
individuals of A. amer i canus and D- si 1vi col us i n
separate cavity blocks. The number of prey killed or
injured was counted 24 h after inoculation of
predators and prey. Similarly, predation was tested
on second stage juveniles of M. i ncoqni ta. The
experiments v<ere replicated five times. All the above

3 3 7
mentioned conditionB remained BBuie -for all experiments
unless mentioned otherwise.
Prey catchi nq and -f eedi nq mechani sms; Observations on
the prey catching and -feeding mechanisms of the two
predators were made at 60 x magnifications by
inverting petri-dishes containing predators and prey
over the stage of stereoscopic microscope. To study
feeding mechanism in detail observation chambers
consisted of a glass slide with 2 x 1-5 x 0,5 cm block
of IX water-agar were used at more than 100 x
magnification <Bilgrami, Ahmad St Jairajpuri, 1985).
Twenty specimens of A. ameri canus and D, si 1vi col us
were released in observation chamabers with 50—75
individuals of H. oryz ae. The agar blocks were
gentely covered with a cover-slip. Observations were
made after 15 m and continued upto about one hour.
Attracti on of predators towards live and exci sed prey;
Attraction of A, amer i canus and D, silvicolus was
tested towards 1ive . (intact) and excised (cut in two
pieces) individuals of H, oryz ae and H. i ncoqni t a
using petri-dish method (Ahmad 8c Jairajpuri, 1980;
Bilgrami & Jairajpuri, 198Sa>, Petri-dishes 5.5 cm in
diameter, filled with agar, marked at bottom in three
zones viz., inner, middle and outer by drawing two

3:3
concentric circles of 0-5 and 2.5 cm in diam. A
plastic straw pipe 5 mm high and 5 mm in diam., sealed
at one end with a piece o-f filter paper was placed
vertically at the centre of the inner circle in such a
way that the sealed end remained in touch with the
bottom D-f petri-dish. The straw pipe -filled with agar
was inoculated with 50 living or excised indviduals of
H. or yz ae and H. i ncoqni t a and incubated -for 12 h.
Five adult individuals of A. ameri c anus and D.
siIvicolus were released at various points of the
periphery of the middle circle. The distribution of
predators was recorded after 2, 4, 6 and 8 h. Scores
were obtained by summing up the products of the number
of worms in each zone with their corresponding
weighting factors. The weighting factors were
obtained by dividing the Br&& of outer zone by that of
each of the three zones. Scores were then converted
into log scores. Experiments were replicated 20
times. Petri-dishes without prey served as control.
Ef feet bf type of prey: Effect of different species
of prey on predation by A. ameri canus or D. si 1vi col us
was determined by subjecting each species of prey
<Table-I) to predation.
Pray selacti on i n combi nat i ons; To determine prey
selection all possible paired combinations of plant

o •J 0
parasitic ne/riatodeB were subjected to predation by A,
amer i canw^L and D ~ si Ivicolus, Twenty five individuals
of each prey was used in each CDmbination. Thus each
combination bad 50 individuals o-f prey.
Ef -feet of prey densi ty s The effect of prey density en
the rate of predation was determined fay releasing five
predators in cavity blocks containing 25, 5D, 75, 100,
125, 150, 175 and 200 individuals of prey.
Effect of starvati on of predators; To study the
effect of starvation of predators on their rate of
feeding prey nematodes were subjected to predation by
O <fresh>, 1, 2, 3, 4, 5, 6, 7, 8, 9 and ID day
starved (keeping in water without prey) predators.
Ef feet of temperature? To determine the influence of
temperatures, predators were placed in cavity blocks
containing prey at verious temperatures ranging from 5
to 40 'C (with intervals of 5 " O .
Ef feet of agar concentr at i ons r To study the effect of
agar concentrations predators were released with prey
in cavity blocks containing 1, 2, 3, 4, 5 and 6% water
agar .
Ef fert of age of predat or s: To find out the rate of
predation by adults and juvenile stages, each stage

o •;• u
of the two predators was released in cavity—blocks
containing prey nematodes.
Feedi nq patterni The number of prey killed by the
predB\.or5 each day was studied over a period of ten
days. Predators and prey were placed together in
cavity blocks. Number of prey killed or injured was
counted after every 24 h- Each day after observations
the predators were ^:r&nB^erred to another cavity block
containing fresh agar and prey.
Resi stance and suscepti bi1i tv of prey nematodes to
predati on snd stri ke rate of the predators; F i f t y
encounters (lip contact of predator with the prey at
right angles) were observed between predators and prey
in petri—dish containing agar. For each encounter
fresh individual of predator and prey were used
(irrespective of whether the predator failed or
succeeded). A prey nematode was placed in front of an
active predator with the help of picking needle
without touching (disturbing) the predator in any
manner. Only such observations were recorded where
predators behaved as normally as could be ensured. To
reduce the effects of satiation and prey habituation
A—h day starved adult predators were tested. The
strike rate <SR) of A. americanus and D. silvicolus

341
and prey resistance <PR} and prey 5U5ce?pt i bi 1 i ty <PS>
of prey neroatodes were determined by using the models
proposed by Bilqrami ^ Jsircijpuri <1989c>. fcncounters
resulting in attacks (EA> , attacks resulting in prey
wounding <AI>J}, feeding after wounding <FW) and feeding
time (FT) were also determined.
RESULTS
Prey catchi nq and feedi nq mechani smss Prey catching
and feeding mechani SBIS of All odor yl ai mus smer i canus
snd Pi SCOIai mus silvicolus were comprised of five
recognisable phases viz., encounters with prey, attack
response {probing), attack, extra—corporeal digestion
and ingestion/ feeding. These have been used to
generate quantitative results. Predators and prey
moved randomly in agar and made contact with each
other 4 encounter with prey) . Predators attacked when
a lip contact <at right angle) was established with
prey. Predators did not show preference for any
particular region of prey. When contact had been
established, a predator probed the prey by moving its
lips sideways along the body of the prey (searching
movements. Attack response) . A. Bmeri canus probed
slightly for a longer duration (3—5 m) than D.
silvicQJus (2-3 ro). During probing a few oesophageal

pulsations were observed in predatore>. I-f the contour
and te>:ture o-P prey cuti cl e suited the predator the
prey was attacked, otherwise predators probed sD/ne
other region- After selecting s suitable spot, the
predator attacked with odontostyle and punctured the
prey cuticle (attack >. A. ameri canus required 8-11
odontostyle thrusts compared with D. si 1vi col us which
required 9-14 thrusts to puncture the prey cuticle.
The predators penetrated odontostyle deep into the
body of prey and disorganised body organs in order to
render it immobile. The oral disc of D. si Ivi col us
also helped in holding prey by suction. During
ingestion oesophageal secretions were rel eased in the
body of prey through the odontostyle (ev.tra—corporeal
di qest i on). Prey showed considerable shrinkage when
their body contents were sucked by the predators.
Ingestion was supported by oesophageal pulsations
which occured at • regular intervals <18-23
pulsations/min in A. aroeri canus and 15—20
puisations/min in D. silvicolus) (inqestion/feedinq).
The ingestion was intermittent 43-4 feeding bouts at a
time). During feeding predators, often left prey for
a while, moved around and reverted back to feed again.
Predatare punctured prey at more than one place- Only
the cuticle of prey was left unconsumed when feeding
was completed.

Q '( '13
Some times prey escaped from the suction "grip"
D-f predator. If sn injured prey escaped it became
susceptible -for future attacks and attracted 3—4
predators which then aggregated around the prey Bnd
-fed together. Females of A. amer i canus and D.
silvicolus probed <by rubbing their lips) around
their own eggs but left without initiating feeding.
In contrast, predators coming in contact with the eggs
of Rhabdi ti s sp. attacked, punctured and fed upon
them.
Attracti on towards prey; A. atner i canus and D.
silvicolus did not show attraction towards live prey
<p < 0-05)- Both predators were attracted towards
excised prey and attraction increased significantly
from 2 to 8 h ip < O.D5).
Effect of type pf prey <Fig. Table - I>: M.
i ncoqni ta, H. mothi , A. tri ti ci and T. semi penetrans
were killed most by A. ameri canus and D. si 1vi col us.
Mycopbagous nematodes and epidermal feeders (ecto-
parasitic) were killed in moderate? numbers. Cortical
feeders (ecto-parasitic) were predated upon the least
{p < D,D5>. No specimen of Hopl ol ai mus indie us..
He) icoty]enchus i ndicus and Hemicycl iophora dhi render i
was either killed or injured by the predators.

? / t'i O '1 'T
When compared, the mean killing o-f individuals
belonging to di-fferent prey trophic categories <PTC>
endo—parasitic nematodes were killed most by A.
ameri canus and D» si 1vi col us and cortical feeders the
least <p < 0.05) <Fig. 1>,
Prey sel ect i on i n mi >; ture of prey; Second stage
juveniles of H. i ncpqni t a ., H. mothi and A. tr i t i ci
were roost preferred in all combinations (p < 0 . 0 5 ) .
When tested together they were killed equally and
frequently. X. basi r i and Lonqi dorus sp. were
preferred the least. No individual of Hoplolai mus
i ndi cus, Heli coty1enchus i ndi cus and H. dhi rendri was
killed in any combinations.
Effect of prey densi ty <Fig. 2 , A & B)s There was a
positive correlation between the number of prey killed
by A. ameri canus <Fig. 2 , A) and D. si 1vi col us (Fig.
2, B> and prey density <p < 0 . 0 5 ) . Maximum predation
Dccured in a population of 200 prey and minimum with
25 individuals of prey <p < 0 . 0 5 ) .
Effect of starvati on of predators (Fig. 2 , C & D ) :
Prey r.on —avai 1 - ability did not affect predation by A.
ameri canus (Fig. 2, C) and D. silvicolus (Fig. 2 , D ) .
The number of prey killed by fresh and ten day starved
predators did not differ (p > 0 . 0 5 ) .

" /' ^
E-f -f e c t p< t e m p e r a t u r e ("t^ig. 2 , E h F> i T h e r e wa
s i gni-f i c a n t i n c r e a s e i n p r e d a t i o n by A_.
< F i g . 2 , E> and D- s i 1 v i c o l u s < F i g . 2 , F>
amer j c snu!
as
temperature increased to 2' Both species of P»"ey
were killed maximally at 25 and 3D »C <P < 0 , 0 5 ) .
Predation decreased at temperatures lower than 25 and
higher than 30 "C <P < 0.05)-
Ef-Pect of agar con cent rat i ons (Fig. 3, A & B>j There
was a negative correlation between the riumber of prey
killed by A. ameri canus (Fig, 3, A) and D. si 1vi col us
(Fig. 3, B> snd increasing agar concentrations <p <
D.D5>. Haximum predation occured at i and 2X agar (p
< 0 . 0 5 ) , Predation declined at concentrations higher
than these, D. si 1vi col us did not kill any prey at 5
or h'A and A. amer i canus failed to kill any prey at h7.
water agar.
Effect of age of predators (Fig- 3, C 8* D> Ail
except first stage juveniles feed on nematodes.
Maximum number of prey wers killed by adult and
second stage juveniles the least (p < 0 . 0 5 ) .
Predation was more on H- i ncoqni ta than H. oryz ae,
Feedi nq pattern (Fig. 3, E it F ) ; A. amer i canus and D.
si Ivjcol us did not show any consistent predatory
behaviour over a period of ten days (p > 0 . 0 5 ) .

4 6
Resi st ance and suscept i bi 1 i t y o-f prey to predat i on and
strike rate o-f the predators (Table ~ II ?< I l l ) : A
5igr«iT3cant CDrreiation was observed between
encounters resulting in attacks and attacks resulting
in wounding < r = 4- 0. 70 - 1 .00; p < 0-05) for all
PTCs. Similar correlations. were obtained between
attacks resulting in wounding and -feeding after
wounding ir = + 0.86 - 1.00; p < O-05) except
endoparasites for A. ameri c anus \r = + D-2S; P > 0.05)
and cortical feeders for D. silvicolus ir = -t- 0-34; p
> 0.05) where the relationships were insignificant.
Mi qratory Juveni1es (Sedentary endpparasi tes> <Table —
II, A) s A. amer i canus and D. si 1vi col us acheived
maximum success on T. semi penetrans (BR = 9 2 % ) . A.
ameri canus wounded H. mothi and A. tr i t i ci maximally
(AW = 93X> while D. si 1vi col us wounded M. i ncoqni ta
and T. semi penetrans (AW = 90 ic 9 1 X ) . Maximum feeding
occurred on M, i ncoqni t a by D. si 1 vi col us (FW = 90>C>
and A_^ tri ti ci by D ^ si 1 vi colus (FW = 8 8 % ) . Both
predators took may. i mum time to consume A. t r i t i c i and
minimum to M. mot hi . All prey species were highly
susceptible to predation by A. ameri canus and D.
silvicolus (PR = 7-13"/-).
Epi dermal feeders <Ecto-parasi tic) (Table-II B>:
Hop 1 o 1 a 1 mos i ndi rtis, Hel i cntiyl enchus i n d i c u s and H .

»5 4 /
dh ' r Bn<j er i p v ^ i b i t e d 1D0% r e s i s t a n c e a g a i n s t
p r e d a t i o n , A . e m e r i c an»g i a n d D . s i l v i c o l u s a t t a c k e d
prey ^ 3f< = 7 2 - S G A > a n d f e d m o s t u p o n B a s i r i a 5 p , (FW =
7 6 « 67/L> . B o t h p r e d a t o r s t o o k sjtsximum t i )7»e ( F T = ABm
e n d 4 6 J I ( > t o c o n s u r o e H . o r y z a e a n d m i n i / n u m (FT = 3 5 n»
a n d 3 9 m) t o - f i n i s h B a s i r i B s p .
C o r t i c a l - f e e d e r s ( E c t o - p a r a s i t e s ) ( T a b l e - I I , C ) :
C o r t i c a l f e e d i n g p l a n t n e m a t o d e s s h o w e d h i g h d e g r e e
o f r e s i s t a n c e (PR = 5i—h77.} w i t h l o w s t r i k e r a t e <SR =
i>0-70X> a n d w o u n d i n g iFU = 3 3 — * 9 X ) , B o t h p r e d a t o r s
-Fed m o r e on T r i c h o d o r u s s p . <FW = 4 6 Se 4 7 % ) . A ,
a m e r i c a n u s t o o k roaxiroura t i m e ( F T = 62ffi> t o f i n i s h X .
b a s i r i a n d D . s i l v i c Q l u s <FT = hSm} t o c o n s u m e
L o n q i d o r u s s p .
M y c o p h a q o u s ( T a b l e - I I , D) A. avenae and
Aphelenchoi des sp- were more vulnerable to predation
by A. Bmeri canus and D. si 1vi col us (PS = 75 - 83%) .
Former fed more on these prey (FW = 78—79X) than the
later (FW = 70-74%). Predators took wore time to
consume A. avenae (FT = 38m and Aim) than
Aphel enchoi des sp (FT = 34 m and 37 /n> .
Numeri cal analysi s of prey trophi c cateqori es (PTC)
(Table-Ill)s
Stri ke rate of the predators : A. atner i canus and D.
silvicolus were fnnst successful predators on migratory

3 4 8
juveniles c-f sedentory endopar Hsi t e s , Comparatively
1 e&E. enconnt&rs resulted in attacks when cortical and
epjderfnel -feedf^rs were subjected to predation fay A.
BPiFr i canus end D. silvicolHS.
A11 a c li 5 resulting i n woundi nq? Attacks with migratory
iuvenilee re&ulted in may.imuiTi wounding with low degree
of variation. Fewer encounters on cortical and
epidermal {-se^derm resulted in attacks.
Prey re-si -st ance? Maximum resistance against predation
was observed in epidermal -feeders. The migratory
juveniles o-f endoparasi tes were most susceptible
trophic group. Mycophagous nematodes also showed
comparatively high degree o-f susceptibility.
Feedi nQ after woundi nq; A. ameri canus and D.
si J vico? us. -fed maximum on migratory juveniles with a
low degree of variation- Feeding after wounding on
epidermal feeders was least with a high degree of
\/&r i ati on.
Feedi nq ti me i Both the predators took a minimum of
24—27m ho consume an individual of migratory juveniles
whereas maximum of 58-60 m to finish a cortical
feeder.

3 4 9
DISCUSSION
A. amer i c anus and D, EsilvicolL's ars very similar
in their predatory behf^viour and r equi rement 5 - The
predator—prey relationships measured -for one species
were very similar to those made for other predator.
Predatian depended on physical contacts with the
prey- Excised or isTJured prey attracted predators
(Veates, 1969? Bilgrami, Ahmad ?' Jairajpuri, 1985;
Sha-fqat, Bilgra<>*i ?< Jair^jpuri j 19S7) arid a sifniiar
phenomenon was observed during present investigations-
The pre and post—feeding attraction and aggregation of
A- americanus and D- s i1v i c o1u s is similar to that
described by Bilgrsmi Sf Jairajpuri (193Sb) in H.
fort i denf? and M- lonqicaudatus- In the presence of
eggs of free living nematodes the affinity of A-
ameri canus and D. silvicolus for their own eggs
suggests a recognition of their own chemical
attractants ^Esser, 1963j Bilgrami, Ahmad ?<
Jairajpuri, 1985; Shafqat, Bilgrami & Jairajpuri,
1987; Khan, Bilgrami & Jairajpuri, 1991 > .
The prey catching and feeding mechanisms of A.
ameri canus and D- silvicolus conforms to the five
phases as described by Bilgrami ?< Jairajpuri {1989a).
Aquat i des thornei required 8-10 thrusts (Bilgrami,
Ahmad ?< Jairajpuri , 1985) and Dor y 1 a i mus st annal i s 6—8

350
<Shafqat, Bilgrami ?<• Jai rajpuri , 19S7> to puncture the
cuticle of prey- D. siIvicolus required more stylet
thrusts (9—14} than A. affler i canus <^S—2 1 > and only
probed -for short durations. Aphelenchid predators
paralyst? their ptrey by injecting saliva H-inford ?<
D3iviera, 1937j Hechler, 1 9 6 3 ) , Dip3ogasterids hold
their f>rey with the oesophageal suction and buccal
armature^ Oilgraroi t< Jairajpuri, 19S9a>, nygolairo
predatar^ paralyse prey by disorganising internal
body organs <Silgra«»i, Ah<nad ?f Jairajpuri, 3 9 8 5 ) .
Both sf>ecies of predators paralysed prey sifniiar to
nygolaiw and dorylaijB predators (Shafqat, Bilgraroi ^
Jairajpuri, i987| Khan, Bilgra»i Sr Jairajpuri, 1991.
The movement of oesophagc»al secretions through the
}uman of odontostyie suggests the occurrence of extra
corporeal digestion in A. c>mer i canus and D. si 1 vi col us
<Grootaert, Jaques & EJmal 2 , 1977; Bilgrami &
Jairajpuri, 1989a; Khan, Bi 1 gr ami 8t Jairajpuri, 1 9 9 1 ) .
The rate of predation by both the predators
increased when the number of prey increased as is the
case with D^ nlenteron colobocercus (Yeates, 1969) ,
Pr i onchu 1 ijs punctatus (Nelmes, 1974) , A. t hronei
(Bilgrami, Ahmad Sr Jairajpuri, ?985>, Hon one hoi dus
1 onqicaudat»<s anti M. fort i dens <BJlnrami 8c Jairajpuri,

19S9a) BTid Apof eel ai mel 1 us nival s (rChsn, Bilgra/ni S«
Jairajpuri, 1 991 > . Fo!^ Honon<-hii%. aquftt i Ois i t
remairfed unaltered <Si Igrami , Ahmad ?< Jai ra jpuri ,
i 934 > - Sha+qat, Silgrasji ?f Jairajpuri <1987) obtained
an optifnuTfj ratio between the predato?- and prey number
•for ms>i i mum predation. During the pres^ent observations
the increase in predation may be attributed to
improved pr&dstor- prey encounters, Wallace (1963)
found lower CS-ID" C> and higher temperatures <40" or
more) unfavourable for nematodes, Bilgranii, Abroad ?r
Jairajpuri5 <1933) found temperatures influencing
activity and predation by H, aquat i c u s . The decrease
in predation by the two species of predators at
temperatures lower than 25° and higher than 30''C may
be attributed to their activity which might be
inhibited by the unfavourable temperatures. Agar
concentrations also govern activity (Wallace, 1969;
Azmi ?f Jairajpuri, 1979; Bilgraroi, Ahmad & Jairajpuri,
19S3) and predation by nematodes <Bilgrami, Ahmad ?f
Jairajpuri, I933j Shafqat, Bilgrami ?c Jairajpuri,
1987; Bilgrami &• Jairajpuri, 19S9b>,. During the
present studies the negative correlation between
predation and agar concentrations suggest that the
activity of predators and prey was affected by higher
concentrations of agar above which predation declined-

1 r- ",
More predation by adults than juveniles re-fiects their
predatory potentials <Bilgran»i Sr Jairajpuri
Bilgrami & Jairajpuri; 1 9 8 9 a ) .
I9i
A. ameri canus and D. s i1v i c o1u s preferred
endoparasitic nematodes most when tested singly or in
combinations- No predation on Hslicotylenchus
indie u s , {-«QP I O *, ai fflus i n d i c u s and H, dhi rendr i could be
attributed to -fHictors such as the activity o-f prey,
type o-f prey, contour and texture o+ cuticle, cuticle
thickness Bnd body annuiations etc, <Bilgrami Sf
Jairajpuri, 1939c>,
Various -factors provide resistance to p'^ey
nematodes against predation <Esser, 1963; Small Sf
Grootaert, 1983j Jairajpuri ?< Bilgrami, 1 9 9 0 ) .
Wounding determines predation and degree of prey
susceptibility- The wound may result in the loss of
hydrostatic pressure o-f the body, af-fect locomotion,
make prey individuals more vulnerable to predation and
allow secondary infections <Bilgrami, 1 9 9 3 ) , Esser
<19S7) also suggested loss of resistance in a wounded
or weakened prey. During the present observations a
thick body cuticle (e.g. Hopl ol ai mi.is i ndi cus) and body
annul at i on-=. < N. dhirendri > might have provided 1 00/i
resistance against A. amer i canus a n ci D. silvicolus a s

• r- - 0 ;•» J
wes observed for other pre-detcrs (teser, 1963; Small 3<
Srootaert, 19S3; Bilgrami ?c Jairajpuri, 2939c>. Esser
(1963) BuggeB-fced that r-esistance to predetion in
H P I i cot yl gnchus^ i ndi ci'S vjas chemical which was later
supported by Biigraoti ^ Jairajpuri {1 9SSa 1 9S9c > ^
Bilgrami {1993>. All odoryIai mos amer i c anus and
Di scolai mus s i1v i c o1u s exhibited similar behaviour
towards Hel i cot yi enchus i ndicus. The high degree o-f
susceptibility o-f endo—par asi tes may be attributed to
their relatively so-ft and sroooth cuticle which is
easily pierced (Bilgrajni, Ahmad « Jairajpuri, 19S3;
Jairajpuri Sf Bilgrami, 1 9 9 0 ) , their activity
<Bilgrami, Ahmad Sc Jairajpuri, 19S3) and lack o-f their
anti predation characteristics such as thick body
cuticle annuiations, cuticular -folds, gelatinous
matrix, rapid escape response e t c . , <Bilgrami ?<
Jairajpuri, 1989c)-
The characterization of prey trophic categories
(PTCs) has revealed that migratory juveniles of
sedentory endoparasites were most vulnerable tc
predation as they appeared to lack antipredation
characteristics (Esser, 1963; Groctaert, Jaques &
Small, 19775 Small ?< Grootaert, 1933; Bilgrami &
Jairajpuri, 1989c). The ectoparasitic nematodes

v O 'i
<CDrtical> poBsess physical characteristics which
resisted predation better than other PTCs- The reason
why cortical feed&r-B were more susceptible to
predation could be their inactivity, large size and
inability to take evasive actions when attacked
tBilgrami, i993>,
Acknowledgemeotsi
The first author is grate-ful to the Council of
Scientific ?f Industrial Research, Jvlew Delhi the second
to the Department of Science and Technology, New Delhi
for the financial assistance.

n r- r
REFERENCES
AHMAD, I. 8< JAIRAJPURI, H. S. (1980). Se>i attraction
arid copulation in Chi 1 opl ecus s y Timet ri cus
(Wematoda : Acrobelinae>, NematoloQica^ 26 s
139-148-
AZMI, M. I- ^ JAIRAJPURI, M. S. (1979)- Effect of
Tiechanical stifflulation and crowding on
My 1 onchul t<5 den t atug . Nematoloqi a fltediterranga,
7 I 203-207.
BILSRAHI , A. L- ^1990). Predaceo»»5 doryl aimE: In
Nematode Bio—control: Aspects and Prospects (eds
Jairajpuri et - al . ) CBS publishers ?f Distributors
Pvt, Ltd. New Delhi pp. 143-115. BILGRAMI, A. L.
(1992), Resistance and susceptibility o-f prey
nematodes to predation and strike rate o-f the
predators Mononchus aq»»at i cus , Doryl ai mus
si aqnali s and Aqnat i des thor nei . Fundament al and
Appl i ed Newatoloqy 15 : 265-270.
BILGRAMI, A. L. n 9 9 3 ) . Analysis of relationships
helw<->en predation by Apor ctr'l aimel lus nivalis
and different P^^ey trophic catecjories-
Nt <»a t n^ oqi c PI ,

7 ,; i^ • U
BILSRAMI, A. L. ^ JAIRAJPURI, M. S, <1985>.
Cannibalism in Hononchus aquati cus^ T n d i an
Journal of. Nematoloqy, 14 <19S4) : 282-283-
BILGRAMI, A. L. ^ JAIRAJPLiRI, M- S-<198Sa>. Attraction
o-f Hononchoi des i onqi caudatus and H. fort i dens
ihi&matoda: Di pi ogast er i da) towards prey and
•fecto'^s influencing att-action. Revue de
NemetoloQie, 11:195-202,
BILGRA.MI, A- L. «f JAIRAJPURI, M. S. <198ab>.
Aggregation o* iJononchoi des 1 onqi caudatus and M,
fort i den*:^ (Newatoda: Di pi oga'5ter i da > at feeding
Bites. Nf>mat ol oqi ca . 34 s 119 — 121.
BILSRAMI, A, L. §c JAIRAJPURI , M, S. (1989a>- Predatory
abilities of Hononchoides 1onqi caudatus and H.
forti dens (Nematoda : Dipiogasterida> and
factors influencing predation. Ne<natoI oqi ca ,
35 s 475-48e.
BILGRAMI, A. L. «. JAIRAJPURI, M. S. <19a9b>. Effects
of concentration and thickness of agar on the
predatory behaviour of Mononchoi des
lonqi cauf<atus and M. for t i dens <NefBatoda s
Di pi ogaster i da > . lofH ar> J n'»r u a I of Nematol oqy ,
If^ i 223-227.

337
BILGRAMI, A, L. & JAIRAJPURI, Yi. S. <19S9c}.
Resistance o-f prey to predatian and strike rate
of the predators, Mor> on c h o i d e s 1 on q i c & u d a t u s end
H. -fort i dens < Meroat ode : Di pi ogasr t er i da ) . Revue
de Nematoloqie, 12 : 45-49.
B3LGRAHI, A, L., AHHAD, I- §. JAIRAJpyRI, M. S. <1983>.
Sofne -factors in-Pluencing p^redation by Hononchus
aqM;=it i cu^ ^ Revue de Me^^at o? o<ii e e> ; 325—326,
BILGRAHI, A, L., AHMAD, I. & JAIRAJPURI, M. S. (1984).
Observations on the predatcjry behaviour o-f
Mononchus aquat i c»s. Nematoloqi a Medi terranea,
12 J41-45.
BILGRAMI, A. L., AHMAD, I. ^ JAIRAJPURI, M. 3. <19a5).
Predatory behaviour of Aquat ides thornei
<WygolaiminaI Nematoda). Nematoloqi ca (19S4>, 30
: 457-462-
ESSER, R- P. <1963). Nematode interactions in plates
o-f non-sterile water sqar. Proceedi nq of Soi 1
Crop Science Soci ety F1 or i d -gi , 23 : 121-128.
ESSER, R. P. n 9 8 7 > . Biological control of nematodes
by nematodes I- Dorylaims (Nematoda :
Dnry1 aimi da> . Nematolngy Cirrul ar^ 4 ! pp 4.

3^.8
GRGOTAERT, P., JAQUES, A. ^ SMALL, H. W. {1977>. Prey
selection in But Ieri us 5p, (Rhabditidae :
Di pi ogaster i dae) . Hed , Fair . Landbouww.
Ri i »;sun J. v- Sent . , A'. > X.
HECHLER, H. C, n 9 6 3 > - Description, developmental
biology and -feeding habits o-f Sei nur a
ten»teaudata <de Yian) , J. B. Boodey, i960
<We«»atoda: Aphel enchi da) , a nematode predator,
Pr-Qceedi n<3s o-f He I fflintholoqical Soci et y
Washinotor. , 30 r 182-195-
JAIRAJPURI, M. S. Sc BILGRAMI, A. L. <1990>. Predatory
nematodes. In Nematode Bio—control : Aspects
and Prospects (Eds Jairajpuri et al.> CBS
Publishers Sf Distributers P'vt. Ltd.,New Delhi pp
95-125.
KHAW, 7., BILGRAHI, A. L. ?c JAIRAJPURI, H. S. (1992>.
Bowe observations on the pr^dation ability o-f
Apporcelaimellus nivalis (Altber, 1952> Heyns,
1966 (Nematoda s Dorylaimi da) . Nematoloqi ca,
37 : 333-3-42,
LINFORD, M. B. & OLIVIERA, J. H. (1937). The feeding
o-f hollow spear nematodes on other nematodes.
5ci ence, 85 : 295-297.

359
MELMfcS, A . J . < i 9 7 4 > . E v a i u a t i o n o+ t h e f e e d i n g
b e h a v i o u r o-f Pr -< onch<<l u s r't-'r.c t f i t I ' s ' ' C o b b ) ^ A
n e m a t o d e pf e d a i o r , J o u r o w l o f A n i m a l E c o l o q y .,
4 -7" _ Ertr-:^ i!r » ir-
BHAFGAT, S,, BILGRAMI, A. L. ^ JAIRAJPURI, M, S.
<19S7>, Evaluation o-f the predstion ability of
Dorylai mus staqnal i s Dujerding, 1845 {Kematoda i
Dor yl ai iisi da > . R e v; e de Memet ol oqi e „ 3D : 455—
SHALL, R. W. S* G R O G T A E H T J P. U 9 S 3 > . Observations on
the predation abilities o-f =o<ne soil dwell irfg
predatory newatodes,. Nematol oqica ., 29: 119 —
WALLACE, H. R. ^9^4>3>. The biol oqv of_ pi ant garasi tic
nematodes. Edward Arnold. London, 280 pp.
WALLACE, H. R- (1969). Wave -formation of infective
larvae o-f the plant parasitic nematodes,
Mel oi donvne i ncoqi nt a . Mei?^atoloqica, 15 s 65—75.
WYSS, U. ?f GROOTAERT, P. <1977>- Feeding mechanisms
o-f Lahr onema sp. Hed. Fac, Landbouww.
Ri jl^Huni V. Bent. . 42 j 1521-1527.
YEATFS, G. W. n 9 6 9 ) . Predahioo by Hononchoides
poloh '\ 'KU- ^ (Memattula i Hi pi ogast er i da > in
laboratory culture. Mematolnqi ca. 15 : 1-9.

3 U Q
T a b l e - I
Rate o-f p r e d a t i o n by A l l o d o r y \ ^ ^ <!» < 5 a??;e'''i cf^ouB a n d ,01 scoJ. a i mus s i 3 Vt colt??:- u s i n g o n e s p e c i e s of pi^ey
Pre-y nematodes a n d Bc<{_>rcp No. o-f P'^ey = 2 5
E n d o - p a r a s i t i c neBjatodes 1 , Mgj o i docjyng j nf~r>rini ^:B
<From t o m a t o )
Ha- o-f p r e y f r i l l e d by p - r e d a t o r s
A, liosaifibr^enei B D. s i 1 v i c o l u s
15 ± 1.00 14 ± D.G9 <13-15)
Hete*="ode't'a sno+hi
4 .
(FroTO m o t h a g r a s s )
Ancfuj n a t r j t i c i (FrofB w h e a t g a l l )
Tvl e n c h u 1 u s semi p i p n e t r a n s < F r OR} L e ^ o n >
E c t o - p a r a s i t i c J Epi derma J - f e e d e r s
1 , Hel j c o t y j . e n c h u s i n d i c t j s (Frora r o s e bed>
15 ± 2 , 0 7 <13-1S>
l i ± 2 - 0 7
a3-is>
i t j j : 1 - 5 3 a 4 - 1 8 >
0
14 + .QB <13-16>
15 ± 1 . 1 4 U 4 - } 7 >
15 ± 1 -58 <13-17)
2 . H o p ? . o l a i m u s i n d i c u s ^Froin p a l m
3 - TyA e n c h o r h y n c h u s m e s b h o o d i CFrc?'^ p a d d y )
4 , Hi r s c h t s a n n i e M ^ o r y ? a e (From p a d d y )
5 , Pa s J r i a s p -(Fro<B p a d d y )
ife. Hei7»icycl i o p H p r a d b i r e n d e r j (From r o s e b e d )
0
10 ± 1 -14 { 8 - 1 1 )
11 ± 1 . 5 S <9-13 )
13 + 1-5S { 1 1 - 1 5 )
O
8 ± 1 . 5 8 { 7 - 1 0 )
9 j 1 . 1 4 ( 8 - 1 1 )
li ± 1 . 5 8 ( 9 - 1 3 )
O
E c t i i - p a r a s i t i c t C o r t i c a l - feederB
1 . X iph i neg>a b a s i r j h ± 1.00 (From <w.<l b e r r y ) (5 -7>
2. i u n q i do r t t g F p , ^ i 1 -09 (Frow mi f lbe ry ) ( 4 - 7 )
3 . T r i c b o d o r u s '^p. 8 ± 0 - 6 4 (From r o s e b e d ) ( 7 - 9 )
4 ± 0 . 8 4 ( 3 - 5 )
4 j : 0 . 8 4 ( 3 - 5 )
7 + 1.30 (5 -8 )
Mycopbagous
1 . A<>bt- 1 e n c b n s ^ve»»r<e (From prTjd'Jy)
An)<e) pncb-i i flr-s
12 + 1 . 1 4 ( 1 1 - 1 4 )
t 1 + 1 . 3 0
12 ± 1 . 1 4 ( 1 0 - 1 3 )
9 4- 0 .84
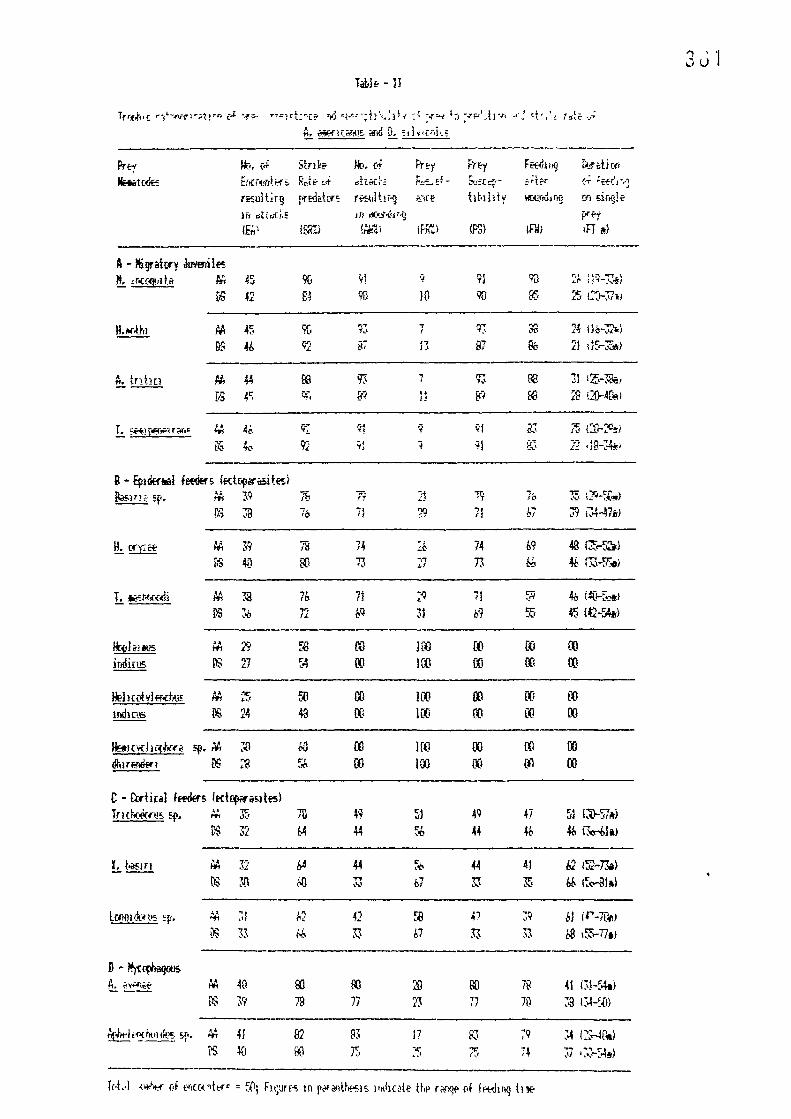
Uh - 33
3u
Tfjoh'C '•"( ' ' •-Wfi ' i t ! ' ' ' ' C^
Prw lfe»t!tC<ie5
A - ^fiyatory iuvea lw
H. ifiCH^/lts fiA
IS
• • f r - " • • = ' ! '
We.. C.+
EficrwiltfE.
(En'
45 42
A, t * f !
Stnlr i i 'd f L't
{"TedtftcfE
iSSX)
% 84
)k: rf
« t iH^ ;
f ^ i )
V] 9fl
3v'€.-k!
fre^ ^^^ii-
i sce
iFS:)
V
J0
'-rf'-'Arf'
hi-/ 'auil^'-
t ibi l i ly
(PS)
?} ?Ci
-•J = 1 ' . \ /
FrtdUig
r f i f
W-iffsuiD^
IFH)
"il)
85
• i t f >.-
i>j-5tic<<
ci ^eet'jVj
!FT»)
2 ;.V5-irvs) 25 ("(-yi ;
H.W4T(1
A. i;nbt3
T. ;r*!i'eri^7''y''=
AA DS
AA i'S
AA S
45 U
44 4' .
4c J
vf. " ^
£8 or.
5-1
92
93 S'
93 ff?
';i 5
T f
33
1
53
9 1
'o 87
93 &v
v3
?}
O P
%
m S8
d." 83
24 i3i-32»)
23 !35-3^>
33 iS -Ss ,
S '.2D-4i:iRi
25 SJ-I-SS;
72 .38-34*'
B - fyjdefMl feeders {«tcfa-a6itetJ B»K?e if', m 3v 7b
DS 38 76
ifrijics
He3Ki?tv3ff«cfa;
23
90 73 A7 39 i34-47ir)
;^ B
« iS
^ DS
AA
es
39 40
7€ :*
?9 27
24
/•a SO
7i
58 54
53 43
74 73
71 y^
CO 00
CD
oc
2i - . 1 i f
1-7 33
m 100
m 100
74 73
73 69
[0 00
00 00
i9 hi.
5? 55
QO
m
OTi CO
48 (35-5^3
46 n3-^«)
46 i4Ct-5(jft)
45 (42-54*)
00 00
00 00
sp. AA
DS
30
28
Trick«:ni5. sp. AA 35
r§ 32
i. basin
La){}i{k<t>5 5j).
A. dV*ficr
A{:.heii«cMiles s?
DS
AA
PS
32
30
3!
33
40
39
43
40
5
70
t4
i4
80
78
82
80
00
00
49
44
44 T7
42
33
80
77
83
7!.
100
53
67
5B
i7
20
23
37
00
00
49
44
44
33
4"
33
77
9Z
7,
00
on
47
4i
43
35
39
33
79
70
79
74
00
53 (3-57*)
Ms (3<ri3a)
A2 iS-7:*> 4i t5<-S3»3
h\ |4"-7C«) ^ i^77»)
41 (33-54*3 75 (34-!tt
34 (S^fti) 37 CC-^s)
Toti'l <i* t< of E-riCHiter" = 50; FUjUre* m pi*f3!)t3i«is iwhcate Uip rftm><» of f«rd!!^ tiie

3 J2
Ui6 - m
fref \n^hii
CatfljCfiK
iuvavile
SitK)
Ccrb£2l
feeders
Ifirto-
parsits)
Epideraal
fEtdETS
<«:tD-
pifaEJtK)
AA
m
fifi
B5
^
IS
AA
DS
E!
K + Si
45+1.-0
45+2.0
(4-
3>2,0
m
32+2-0
iS
33t«.0
(185
32+7.0
S )
4!+5.0
CI
40+1,0
(3
Si S i
K + a i
^ 2 . 0
!2i
95+2.0
i4)
fe+4.G
(6)
&4t3.8
tSJ
i^+}2,0
UB)
^+{3.0
121)
82+1.0
SG+5.0
(2>
MJm
fi + g)
92+1.6
U")
89+2.0
\2)
45+3,G
(8)
37+&.B
HI)
37+4}J
U09)
34+37.8
1109)
82+i.O
n')
78+5.0
«)
miti
K + gi
8+1,0
Il4
11+2,0
!15)
S5+3,0
(71
&3+6.Q
UO)
i3t4L0
i£<)
^-+39,0
(59)
19+2,0
ni)
24+1.0
ii)
PSfi)
fi + 3
92+1.0
(1)
89+2.0
i2'}
45+4,0
(8)
37+6,0
«7)
37+41,0
fill)
34*37,0
«09)
82+2.0
(3)
7i+2.0
{3)
F» \l)
H + S>
87+3.0
(3)
8i+2,0
!2)
42+4.0
OO)
35+7,0
os;
34+S.O
Oil)
31+35.0
ill2)
79+1,0
72+3.0
<4)
FT is)
fi + SJ
27+3.1
<?2V
24+3,2
i}3)
m^A m)
4C+1D.9
«8}
43+7.0
116)
43+3,8
19)
38+4,9
a'3)
38+0,7
C)
Figtyps in jyara-fthefiH iwjjcate a<effk2»t af variaticfi, hll iiqura, are mi/ai to xtele fiinbers.
H+^ = fieafi StiTid*^ Swiatiw; FA s EfftcwdffB rK«Uin§ in e t t s t b | 3? = Strike rate of pfslatw^
fiH = Attacks r«aitnt§ KI »tc«»d!rig; PR = Prey rwiUaice} PS = ft-ey suscsftihility} ffi = Fedifig after
«:«idin95 FT = fe«di^§ t3*e.

^\ '^ ^ oo3
LEGENDS
Fig. 1 Mean killing of P»"ey belonging to di-f-Terent prey trophic categories- A—Endo-parasites; B-HycophagouB5 C-Cortical -feedersj D-Epidermal feeders.
Fig- 2 A. Effect of number of prey on the rate of predaiion by All odorv3 ai mus amer i c anus.
B- Effect of number of prey on the rate of predat ion by Pi scola imus si 1vi col us.
C- Effect of starvation on the rstte of predation by A, amBri c anus.
D- Effect of starvation an the rate of predation by D. si 1vi col us.
E. Effect of temperature on the rate of p*redation by A. amer i canus .
F. Effect of temperature on the rate of predation by D. silvicolus.
Fig- 3 A. Effect of agar concentrations on the rate of predation by All odor ylai mus ameri canus.
B. Effect of agar concentrations on the rate of predation by Pi scolai mus si 1vi col us.
C- Rate of predation by different stages of A. ameri canus.
P. Rate of predation by different stages of P. silvicolus.
E. Predatory pattern of A, ameri canus over a period of ten days-
F. Predatory pattern of P. si 1vi col us over a period of ten days.

3J4
A.gmen'cQnus
D.silvi'colus
A B c Prey trophic categories
D
-/ - -J

3u5
16-
XI (V
o
u
Z. 8 - .
f
T 1 1 1 \ 1 1 1 50 100 150 200
Number of prey
• • M.incognito
• — • H.oryzae
- 1 — I — I — I — I — 1 — 1 — I — I — I — r 0 2 4 6 8 10
Starvat ion per iod (days)
15n
U-
12
10
50 100 150 200 Number of prey
-I 1 1 — \ 1 1 1 — I 1 1 1 0 2 A 6 8 10
Starvation period (days)
1 6 T
i5l2-
?10-D. o 8-
1 ' 5-
~ T — 10 To" "30"
—1 /.o
16n
U '
12
10'
8H
6
2-\
Temperature C
1 1 1 1 1 s 1 1
0 10 2 0 3 0„ '.0 Temperature C
f/6'

"3 '-• 'A
a?12-
10-
<u 8
2
2
16'
U-•a 01
= 12'
>^10' O)
8 -
E • D
^ 2-
16^
0)
a l 2 H
xi lO E D 2
8-
id A
B M. incogni ta
• _H-oryzae
16
T — I 1 r 2 3 A 5 6
Agar concen l ra i i ons (%)
M.incognita
^ H.oryzae
i 1 i
J2 Feeding rate of adults ^juveniles
I M.incognilo
_H. ory zae
1
T 1 1 1 1 1 \ 1 1 1 2 A 6 6 10
fJumber of days
U
12H
10^
8-
6-
/..
2
16
U
12
10
8
6-
4-
2
1 1 1 1 1 ;
1 2 3 A 5 S Agar concent ra t ions (%)
W 'A
D
^
A JA J3 h Feeding rote of adults S^juveniles
16
1A
12-
10-
I I I I 1 I I I I I
0 2 A 6 8 10 Number cf days

3 J 7
Predatory bcWaviour of a nematode -feeding mite
Tyrophaqus putrescenti ae (Sarcoptiformes s Acridae)
Anwar L. BILGRAMI
Sect! on o-f Nematol oqy , Department of Zool oqy , Al iqarh iJusl i m Uni versi ty Al i qarh-2D2D02, Indi a

' Q
O u O
SUMMARY
Observations on T y r o p h a q u s putrescent!ae revealed that these mites sre predaceous on many species of plant and soil nematodes belonging to -Five prey trophic categories viz., saprophagus, endo-parasitic, cortical -feeders, epidermal feeders and predaceous nematodes- Mites preferred second stage juveniles of endo-parasitic nematodes most but predaceous nematodes resisted predation comparatively better than others-Physical, and behavioural characteristics have been attributed to the cause of resistance. Predation by T. putrescenti ae is density dependent which required essentially a contact between the chelecerae and the body of prey- Chelecerae is the main killing and feeding organs- Legs are used to hold the prey during attack and feeding. An injured prey, if escaped, attracted other mites which aggregated at feeding site. The aggregation of mites at feeding site often resulted in struggle amongst them in order to snatch the pr-ey and feed- Factors like temperature, agar concentrations, agar thicknesses and test arena governed predation by T. putrescent!ae- The mites turned coprophagus (feed on its own excreta) in the absence of prey- No cannibalistic behaviour was observed in T_, putrescent! ae either in the presence or absence of prey nematodes-
Key Words: Tyrophagus putrescent i ae, mites. Predator, Prey, behaviour.
Resume

.-,- g w u
The -feeding by the mites on nematodeB was -first
reported by Linford and Oliviera <193S). Since then,
the work on mites and their possible influence on soil
nematode populations remained very limited. Murphy
and Doncaster <1957> reported injury of Heterodera
cysts by a mite. The most definite association
between mites and nematodes came from the work of
Rodriguez et al. <1962> who cultured Macrocheles
muscaedomest i cae on Rhabdi t i s sp- , and found it to
prefer house fly eggs o\fer nematodes. Its proto— and
deuteo-nyrophs under <&Bn)e conditions, however, prefered
nematodes. Rockett and Woodring <1965) found an
oribited mite Pergalumna sp . feeding on Pelodera
1ambdi ense sn6 Ty1enchorhynchus marti ni in large
numbers. Muraoka and Ishibishi <1976) identified 41
species of mites that fzould feed on Cephal obus sp.
The BBccste adult forms of Hetercdera qlyci nes and
Meloi doqyne i ncoqni ta were not devoured by the mites
but their juveniles were consumed (Muraoka S<
Ishibishi, 1976>. Irobriani and Mankau (1983) observed
voracious feeding by a neostigmatid mite Lasi osei us
s c u1p a t u 5 on Aphelenchus avenae and Cephalobus sp. In
culture, increased population of mites resulted in a
significant decline of Aphel enchus &\/er> ae (Irabriani ?<
Mankau, 1983) and predatory and saprophagus nematodes
<Bilgrami ?< Ta)iBt?eij, 19 9 2 ) . In the present work

370
observationB were made on the predatory behaviour of a
nematode -feeding mite Tyr ophaqus putrescenti ae using
plant and soiJ nematodes as prey,
MATERIALS AND METHODS
Tyrophaqus putrescenti ae was cultured in Petri-
dishes containing 17. water agar using Rhabdi ti s and
Cephal obus spp., as p>rey, Sfnall amount of infant mil};
powder (Lactogen) was spread over the surface of agar
to encourage bacteria to grow- Bacteria served as
food for the prey nematodes.
Observations on predation by T, putrescent i ae
were made in small cavity blocks containing 1'/. water
agar using Hirschmanniella oryz ae and second stage
juveniles of Meloidogyne i ncogni t a as prey separately.
Fifty individuals each of two species of pr'ey were
subjected to predation by five adult mites. Number
of prey killed or injured was counted after 24 h. All
e>!periment5 were made at 28 + 2"C and replicated five
tiroes. The conditions remained same for all the
e>!periment5 unless mentioned otherwise.
Prey c a t c h i n q and feedi nq mechani sms; Prey catching
and feeding mechanisms were studied in culture dishes
and special oljservation chaoibers as designed by

Bilgrami and Tahseen <i992>- A plastic ring 1 cm high
and 2 cm in dia/neter, glued to a coverslip at one end,
was fixed in the middle of the metallic slide- The
chamber, thus formed, was filled with IX water-agar.
The mites Birtd prey nematodes were then inoculated.
The ring was sealed with another coverslip to prevent
air drying and escape of mites. Predsti on was
observed on H- oryz e Af^d M- i ncoqni t a. Observations
were sriEide at more than 100 r, magnifications-
Rate of predati on by T. putrescent i ae on di fferent
spec i es of prey nemat odes: The rate of predation by
T. putrescents ae was observed on prey species
belonging to three different prey trophic categories
t P T O vis. , Saprophagus <Rhabdi t i s, Cephalobus
AcrobeloideS:, Acr obel us < MesorhabditisT Tobr i 1 us spp- ,
and Chi 1oplacus symmetr i cus; plant parsitic (H.
i ncoqni t a, Anqui na tritici , Heterodera mothi; H.
or yr ae Tylenchorhynchus mashhood i Hoplolai mus
i ndi cus , Helicotylenchus indieus, Basi r i a sp- ,
Hemicriconemoi des manga fer ae; Hemicycliophora
dhi render i , Aphelenchoi des sp. ; X i phi nema basi r i ,
Paralonqidorus c i tr i L onq i dortis ., Paratri chodorus
spp., and predaceous nematodes (Mononchus aquaticus.
My] /•» n c li u 1 us den h a <: us t Dor y 1 a i «i)us s t a q it a 1 i s , Aqua tides
t hornei , Mononrhni des 3 onq i r iiacia b us M . f or t i dens and

0 11
Aporcelairoel lus ni vel is) . Predaiior y profi le: Rate of
predation was dst-ermirtedi over a period of ters days-
Each day a-fter observation mites were transferred to
another cavity block containing fresh agar end same
number of prsy nematodes. Ef -feet of prey den si t y i
Effect of prey density on the rate of predation was
observed by placing the mites with 50, IDD, 150, 20D,
250, 300, 350, 400, 450 ar,d 500 individuals of H,
i ncoqni 1:a and H. or ys ae. Ef feet of star vat i on of
mitesi To determine the effect of starvation
(deprivation of food) of T. putrescenti ae 0 (fresh) ,
2, 4, 6, 8, 10, 12 and 14 days starved mites were
released with prey nematodes. Each group of starving
mites was tested separately.
Ef feet of t emperature; Effect of temperature on the
rate of predation was determined by releasing mites
with prey nematodes at temperatures ranging from 5 to
40 "C (with an interval of 5 " C ) .
Ef feet of agar concentrati on; To observe the effect
of agar concentrations on predation, the mites were
placed with prey nematodes in cavity blocks containing
1, 2, 3, 4,5, and &>'/. water agar-
Effect of agar thi ckness: Prey nematodes were
subjected to predation by tlie mites in cavity blocks

•^ rj -N
37
containing 2, 4, 6, 8, ID, 12 and 14 mm thick agar
layers to observe the effect of agar thicknesses on
the rate of predation.
Ef feet of test arf^na; To determine the effect of
surface ares on the rate of predation by mites 4 mm
thick agar blocks of 0,5, 3,0, 1,5, 2.0, 2.5 and 3.0
cm radius were taken in cavity blocks. The area of
each block was calculated by TJ r >; thickness of the
agar -
RESULTS
T. putrescent i ae is predaceous, feed on nematodes
and other micro-organisms in culture dishes. During
routine observations, the culture of many saprophagus
(Rhabdi ti s, Cephalobus, Acrobeloi des Acrobelus,
Hesorhabdi t i s ;, Tobr i 1 us s p p . , Panaqrellus redi vi vus
end C. symmetr i cus and predaceous. nematodes (M.
aquat i cus ., M. dent at us , D. st aqnal i s .. A. thornei ^ M.
1 onqi cuadatus M . fort i dens hi. bast i ani , D.
silvicolt<5 and A. n i vel is) were found heavily
contaminated with T. putrescenti ae, T. putrescenti ae
fed voraciously, restricted their multiplication and
reproduction. As a result of extensive predation,
populatrion of nematodes declined significantly. In
some rases even whole population perished.

'^ .-J ,
J/4
Predation depended upon chance encounters with
prey individualB. T. putrescent!ae could hold its
prey by its pulp (used to identify prey organisms and
other objects) and legs as soon as prey came in
contact. The legs are: used far holding and wounding
prey, Chelecerae is the main killing and feeding
organ, used -for grasping, crushing snd shredding prey
into pieces. The ingestion of prey contents was
intermittent <2-3 feeding bouts at a time; 22-26
feeding bouts/minute) with short periods of resting
activity at regular intervals. Feeding time depended
up«on the type of prey. Mites took longer time to
consume large pf"ey than the small- Physical
characteristics which provide resistance to pf^ey
nematodes against predation governed attaches by the
mites, puncturing of cuticle and time of feeding- No
cannibalistic tendency was observed in T,
putrescenti ae.
An injured prey if escaped, attracted few other
mites. Such kind of attraction led to the aggregation
of mites around the injured prey at feeding site. The
mites fed together on an injured prey. During feeding
struggle to get hold of prey and to feed first on the
prey vtaB observed amongst the mites. Mites pulled the

T 7
prey in opposite directions in order to snatch the
prey. During the process the prey was teared into two
or more pieces one going to each mite- The mites
also consumed its own -feacal matters while moving
rar?domiy, generally in the absence of prey nematodes.
The nymF«hal stages of mites -fed on dead, injured or
inactive prey individuals.
Rate o-f predat i on by T , putrescent i ae on di -f -f erent
speci es of prey nematodes; Migratory juveniles of
sedentary endo—parasites were most susceptible to
predation and preferred most by the mites <p < 0.05)
<Table—1>. Predatory nematodes avoided predation
comparatively well when used as prey against T.
putrescenti ae. They were preferred least by the
mi tes.
SaprophaqL»s nematodes <Table—I, A ) : Maximum predation
occurred on Acrobeloi des sp.^ <MSD 38 +1.2) with a low
degree of variation. Acrobelus sp. was least preferred
(MSD 2 9 + 1 . 3 ) <p < 0.05) .
PI ant p arasi tic nemat odes <Table—I, B ) J The juveniles
o f A • t r i i i c i ., M. incoqni ta< and H. mot hi were hilled
most by the otites. T. mashhnodi , Hopl ol ai mwa i nd i cus,
He] irotylenrhnq i nd i ru'^ , H. manq i ferae and H.

? 7 •
dhi rersdr i resisted predation better than other Bpecies
of plant parasitic nematodes. Predation occurred on
32—54X population o-f these prey- The rest of the
nenjatodes o-f this trophic group we're killed in
moderate numbers <p < 0,05).
PredareouE n ens at odes <Tabie — I, O l Dif-ferent
species of predatory nematodes exhibited resistance
against predation, 24~4B'/. population of p«redaceou5
nematodes was predated upon by the mites- Ha>iimum
being on i±^ aquati cus <MSD 24 + 2-5, CV = 10%) and A.
thornei <MSD 24 + 2-5, CV = 237.) and minimum on D.
staqnal is (HSD 12 + 2-5, CV = 2iy.> <p < 0-05)-
Pr edator V p r o -f i 1 e <Fig- 1 A)j T - putrescent! ae Killed
same number of prey individuals each day over a period
of ten days (p < 0.05)- More individuals of M.
i ncoon i t a were killed than H. oryz ae.
Prey densi ty (Fig. 1 B>. The r ate of killing M-
i ncoqni t a and H. oryz ae b y T. putrescenti ae increased
with the increase in prey density <p < 0-05)- Maximum
predation occurred in a population of 500 P^f^Y
individuals and minimum in 50 <p < 0.05)-
Effert of st arvati on of mi hgs (Fig. 2 A ) : Rate of
predaLiun by T_^ pu Ir e'.c r>" i i o increased with the

'1 7 7
i ncrcBse in the period o-f starvation. Number of prey
killed by 14 €isy starved mites was significantly more
than the fresh (0 day) <p < 0 - 0 5 ) .
Ef feet o-f te«<per at ure <Fig. 2 B) : Temperature also
governed predation by T. putrescenti ae. Rate of
killing H. i ncogni t a and H. oryxae increased
significant!/ opto 25 "C (p < 0.05) but remained
constant thereafter <p > 0 - 0 5 ) .
Effect of atjBr concentr at i ons (Fig. 2 Ot Rate of
predation was directly proportional to agar
concentrations. Maximum killing was recorded Sit
higher concentrations <5-i&/i) (p < 0 . 0 5 ) . Low
concentr at i on^; (3-2%) yielded minimum predation by T.
putrescent i ae.
Effect of a<iar thi r knesses (Fig. 2 D)s Predation
decreased with the increase in agar thickness. Agar
layers more than 6 mm did not favour predation. Mites
killed maximum number of prey in 2-6mm thick agar
layers (p < 0. 0 5 ) .
Effect of test arena (Fig- 3 ) : Surface ar ea affected
predation greatly by T. putrescenti ae. Predator-prey
encounters in small test areas (0.3) cm"^ to 1.3 cm""')
yielded maximum predation whereas in large areas (2-S
Lo 11.3 cm"') predation (.fF>rlJi>ed sharfily (p •. O.D5>-

7 8
DISCUSSION
Dbser vat i ori5 on J, putrescenti ae suggest that
these fijites are predaceous in nature which -feed
voraciously on plant and soil nematodes belonging to
dif-ferent trophic groups. T, putrescent i ae possess
greater prsdai:ory potentials msiinsY because o-f their
abiiit/ to kill variety of nematodes in large numbers
in culture and ex p.sr i mental dishes (Biigrami &
Tahseen, 19*52). RocJ^ett and tvoodring also observed
voracious reeding by an oribited mite Perqalumna sp.
on Pel Oder a 1ambendi ense and Tylenchurhynchus mart i ni .
Probing before attacking prey is a common
phenomenon in predatory nematodes (Bilgrami et al.
1985; Bilgrami ?f Jairajpuri, 199D; Bilgrami, 199Da)
but T. putrescenti ae did not show any such mechanism,
Predation by the mites depended up-on physical contacts
with the prey nematode. Contact o-f prey with the
cheiecerae o-f the mites was necessary to i n i t i at e " and
attacJ;. An injured prey, however, attracted other
mites which were aggregated around an injured prey and
•fed together. This suggest that the mites are able to
percieve prey secretions and respond positively
towards them similar to predatory nematodes which move
in response to prev secretions, aggregate around prey
and feed (Bilgrami, lP90b>.

379
The reason why the second stages o-f hi. i ncogni t a,
H. mothi and A- t r i t i c i killed more than the
parasitic snd predatory nematodes, could be attributed
to phy!sical, chemical and behavioural characteristics
which provides resistance against predation (Bilgrami
?c Jairajpuri , 1989, Bilgrami, 1992). Such anti-
predation characteristics are generally absent in the
juveniles of endo—parasitic nematodes.
Increased predation by Xi. putrescent i ae with
increa^sing number o-f H. oryz ae and N. i ncoqni t a
suggest density dependent predation as is the case
with many predaceous nematodes <BilgraB»i et al . 1934;
19S5; Bilgrami Sc Jairajpuri, 199D>. This increase in
predation may be attributed to improved nhantze: of
contacts . between prey and predators- During present
observations increased pr€?dation by the mites with the
increase in the period of starvation may be due to the
apititic conditions which are governed by the duration
of stajrvation. Differential rate of predation by T.
p u t r e s c en t i a e at different temperatures and agar
concentrations may be attributed to the activity of
mites and prey nematodes (Bilgrami et al. 19S3). More
predation in small test arena suggests that small
surface? area provided mites more nematodes pi^r unit

2 80
area to attac); than the large ones- Thus more
encounters between prey nematodes and mites occurred
which resulted in increased p<redatiDn by the mites.
The rate o-f decline in predation by T. putrescent i ae
in thick agar layers is probably because of the
migration o-f prey nematodes into the agar- The
thickness o^ a^ar provided prey nematodeB more srBa to
avoid predation as the mites could not penetrate agar,
T, putrescenti ae is coprophagus which -feed on its own
feacal matters generally in the absence o - prey
nematodes ^Bilgrami Sc Tabseen, 1992). This kind of
behaviour suggests that mites undergo starvation in
the absence o-f food <prey>- Wo canni bal i si t i c
tendency was observed in mites-
ACKNOWLEDGEMENTS
The author is grateful to the Department o-f
Science and Technology, New Delhi -for providing
•financial assistance-

"ll C 33 1
REFERENCES
BILSRA'II, A. L. <1990a}. Pi plQQa5tf?rid predators; In
Jairajpurij Alam ?f Ahmad (EdB-> Nemat odg h\ o -
control i aspects and prospects CBS publishers
?' Distrihutors Pvt. Ltd New Delhi pp. 135-142.
BILSRAHI, A. L. (199Db). Predaceous doryla ims: In
Jairajpurij A2 am Sc Ahmad (Eds) Nematode Bi o —
control ? aspect s and prospect s. CBS publishers
5. Distributors Pvt. Ltd. New Delhi pp. 143-14S.
BILGRAMI, A. L- <I992>. Resistance and susceptibility
of prey nematodes to predation and strike rate
of the predators Hononchos aquat i cus,
Dor yla i mus staqnal i s and Aquat ides thornei .
Fund. Appl. Nematol.- 15 : 265-270.
BILGRAHI, A. L., AHMAD, I. ?. JAIRAJPURI, M- S. U983>.
Some -factors influencing predation by tJononchus
aquat i cus. Revue Nematol . < 6> : 325— 326.
BILGRAMT, A. L-, AHMAD, I. ?. JAIRAJPURI, M, S. (1934).
Observations on the predatory behaviour of
Mononchus aquati cus. Nemat ol . Medi t. 12 i41 —

382
BILGRAHI, A. L., AHMAD, J. ?, JAIRAJPURI, H, S. <19B5>.
predatory behaviour of Arjuat ides t hornei
(i'iyool ai fiti na ; NefaatDda', Hgma t o I. OQ i C a <19S4>, 30
i 457-462.
BILGRAMI, A. L. ?< TAHSEEN QUDSIA (2 9 9 2 ) . A nematode
-feeding mite, Tyrr^pHarjus putrescent; ae
<BarcDpt i-f orn»i 5 ; A c a r i d a e ) , Fund. App] ,
Nematol,, 15 ; 477-478.
BILSRAMI, A. L. ?< JAIRAJPURI, M. B- ( 1 9 8 9 ) , ReBistarsce
o-f prey to predation and strike rate o-f the
predators Mononchoi des 1onqi caudatus and H.
fort i dens {Ne/natoda ; Di pi ogasteri da ) - Revue
Nematol . .. 12 s 45-49,
BILGRAMI, A. L. & JAIRAJPURI, H. S. (1990). Predatory
abilities o^ Mononchoi des Ionqi caudatus and
H. fort i dens (Nematoda i Dip3ogasterida> and
•factors influencing predation. Ne'nat ol oqi c a ,
35 : 475-488.
IMBRIANI, J. I. ^ MANKAU, R. <1983>. Studies on
Lasi psei ns sculpat u s , a neostigmatid mite
pre'daceoiis on nematod«?B. J. Nematol . , 15 ;
523-528.

•
o 83
LIMFDRD^ H. B, «r OLIVIERA, J, M, (1938). Potential
agent o-f bio —logics*! control o-f plant pe-J^asitic
nefnatodes- Phytopatholociv^ 2Sil4.
HURAOKA, M. ^ ISHJBISHI, N, n 9 7 & > . Nematode feeding
fTiitsB and a their feeding behaviour. App. En t .
Zocl., il : 1-7.
MURPHY, p. W. ?( DONCASTER, C. C. <i957>. A culture
niethod -for soil micro-fauna and its application
to the study of ne<r<atode predation,
Niematoioqi ca , 2 z 202-214.
RDCKETT, C. L. ^ W00DRIN5, J. P. (1966'). Biological
investigation on a new species of ceratozetes
and of Pergalumna. Ac aroloqi a, 8;14 —18.
RODRIGUEZ, T. G. , WADE, C. F. Sf WELLS, C. N. <1962>.
Nematodes as natural food for f^acrocheles
muscaedoroes-t i cae (Acarina J Macrochelidae)
predator of the house fly egg. Ann. Ent. Soc
Affleri ca, 55 : 507-511.

3 3 4
TABLE-I
Rate of predation by Tvropham,<5 putrescent! ae on di-f-ferent species o-F nematodes belonging to
different trophic groups-
Prey specie; H e a n k i l l i n g
P e r c e n t k i 1 1 i n o
CV (•/.}
fits SAPRDPHAGDUS
R h a b d i t i s s p . 3 3 + 3 . ! hh 9 < 3 0 - 3 7 )
C. symmet r i cus 35 + 1. 6; 70 5 { v> - o r /
A c r o b e l o i d e s s p , 3 S + 1 . 2 7«^ 3 { 3 7 - 4 0 }
Acrobelus sp, 2 9 + 1 . 3 58 5 (27-31)
Hesorhabdi tj s sp. 37 + 1.5 74 4 (35-39)
Tobrilus 3D + 2.5 60 8
(28-34)
B: PLANT PARASITIC NEMATODES
Heloi doqyne i nc oqni t a 39 + 4.1 78 11 (34-44)
Anquina triti ci 41 + 2.0 82 5 • (37-44)
Heterodera mothi 40 + 2.9 80 3 (36-44)
Hirschmanniella oryz ae 31 + 3.0 62 10 (28-36)
Tylenchorhynchus mashhnodi 27 + 1.5 54 5 (25-29)
Hop 1 ol a i mus i nd i r ns 21 + 1.1 42 5 (19-22)
Hel icotylenchuB i n iJ i r, u •> 20 + 1-6 40 8 (18-22)
g - s i r i a s p , 3D + 3.5 60 12 (27-36)
HfMit i j i f ' i <" r>f> Hinf) i iit^'>. <lLiLLllLi_f.fJ!-Lilil ^ ^ .t ^ -'^ ^^^ • ^' ' - - . - ( 1 2 - 2 0 )

" p f 0 0 3
Table-I Contd
Prey species Mean k i 1 1 i r; g
18 + 1.2 (17-20)
29 + 2.3 (27-33>
28 + 1.9 (26-31)
20 + A.h (16-28)
22 + 5.0 (14-28)
24 + 6.4 (18-32)
P k ercent i 1 1 i n Q
36
58
59
40
44
48
C'-J (•/.)
7
8
7
23
23
27
Hemicycliophora dhi rendr i
AphelenchoT HF^B sp ,
Xi phi rtgmg bagi r i
Paralonqidorus ci t r i
Lonqi dprus sp.
ParatrichodoruB sp.
Cs PREDACEDUS NEMATODES
Hononchus aquat i C U B 24 + 2.5 48 10 (20-26)
Hylpnchulus dentatus 22 + 3.2 44, 14 (18-26)
Doryla 3 muB staqnal i s 12 + 2.5 24 21 (9-15)
Aquati des thornei 24 + 5.5 48 23 (15-28)
Hononchoi des lonqi c auda t u5_ 21 + 5. 1 42 25 (14-28)
Honpnchoi de'^ -f or t i dens 19 + 2.1 38 11 (17-22)
A p o r c e l a i m e l I I J B n i v a l i s 21 + 2 . 2 4 2 11 ( 1 8 - 2 4 )

386
LEGENDS
Fig. 1. A — Predatory pro^^ile o-f Tyrophaqus pu br escgn t \ ate over c period of ten c5ay& >; = He 1 o 1 dOQ y ne i ncoqn i t a 0 = Hi rschmanni el 1 a o y a a e
B — Ef-fect af prey density on the rate o-f predation by Tyrophaqus putrescenti ae >; = Mel oi dog yrre i ncogn i t a O = Hi rschfr;anni el 1 a or vz ae
Fig. 2. A — Ef-fect of starvstion on the rate o-f predationr by T vrophagits putrescent i ae >: = Hel oi do'^yne incognila 0 = Hirschmanrfiella or yz ae
B - Effect of temperature on the rate of predation by Tyrophtaqus putrescenti ae, '' ~ Mel oi donynB i ncoqni t a D = Hi rschmanni el1 a or yz ae
C — Effect of agar concer? t r at i ons on the rate of predation by Tyrophaqus putrescentiae, X = Meloi doqyne i ncoqni ta D = Hirschf^anniel 1 a or ys ae
D - Effect of agar thicknesses on the rate of predation by Tyrophaqus putrescenti ae. >: = Mel oi doqyne i ncoqna t a O = Hirschmanniell a oryz ae
Fig. 3. — Effect of test arena on the rate of predation by Tyrophaqus putrescenti ae. >: = Mel QJ doqyne ancoqrii ta O = Hi rschmanni el 1 a oryz ae

87
100 A
p E R C E N T
K I L L I N G
80
60
40
20
- « - H.ORYZAE
-+- M, INCOGNITA
1 1 1 — r —I J r-6 6 DAYS
"I—r 8 9 10
P E R C E N T
K
90
80
70
L L
N ^0 G
B
• ^ H. 0RY2AE M. INCOGNITA
6 0 T 1 1 — I 1 — ] — 1 1 1 r — 1 1 1 1 — I 1 1 1 — ! \
50 100 150 200 250 300 360 400 460 500 PREY DENSITY
F\-;N^ \ -

238
;5 •
SOi
30
70
SO-
SO
80
70 p t: H C t M T
K 1 1 L 1 N G
fin
fin
-lU
30
20
-,0-
A
- « - H . ORYZAE -M. INCOGNITA
_ j p^ I ~l ( J I J I I 1
0 2 4 6 8 10
STARVATION (DAYS)
14
-^- a oarzAE H - M, INCOGNITA
"T I ; I 1 I I I I ; I 1 I 1
10 15 20 25 SO 35 " 40
TEMPERy\TURH (°C)
85
p 80 E R C 75 E N T
7 0 -K I L L I N Q 60
65
55 4
60 1
P
R ^° C
, K
n I G
40
"-^ -&-H.. ORYZAH - ^ - M . INCOGNITA
2 3 4 AGAR CONCENTRATiCH {%)
~i 1 f -
6 8 10
AGAR THICKNESS (MM)
1 r 12
V-v.-^

85 1
o n o c o y
p 80 E R
N T
70-K t
I 65-f N G 6 0 - - ^ H. ORYZAE M. INCOGNITA
55 T 1 ( j - — J p
7.8 0.31 1 r
13 2.8 5.0 AREA {CUBIC CM)
T1.3
VKOS^

390 *J w'
EVALUATION OF PREDATION ABILITIES OF MYL0NCHULU5
DENTATUS
By
ANWAR L- BIL5RAHI St RAJEEV KULSHRESHTHA Section of Nematology, Department D-f Zoology,
Aligarh Muslim University, Aligarb-202002
Abstract; Observations on My1onchulus dentatus revealed that it is predaceous in nature. The prey catching and -feeding mechanisms are comparable to other mononcbs. Predation by M_ dent at us depended on chance encounters with the prey. Predators fed by cutting the cuticle and shredding the prey into pieces. There was no set predatory pattern. Females were most active predators than the juveniles. M. dent at us prefered second stage juveniles o ^ Meloi doqyne i ncoqni ta Tylenchulus semi penetrans, Heteroder a mothi and Anqui na tri t i ci most. All •factors vi z . « number o-f prey, temperature, agar concentration, pH o-f the medium, test arena^ starvation of predators and thickness o-f agar in-fiuenced predation by M. dent atus.
Key words: Predation, predator, prey, Mvlonchulus d entatus
The speculation of using mononchs for checking
populations o-f plant parasitic nematodes in soil was
made by Cobb <1917> -followed by other reports on the
use of predatory nematodes in nematode bio-control.
It was only in 1974 when first detailed observation
was made on the predation abilities of a roononch,
Pri onchul us punctatus <Nelme5, 197-4). Cohn 8t
Mordechai <1974> reported density dependent predation
by Mylonchulus si qmaturus upon citrus nematode.

3 31
Tylenchulus semipengtrans. These studies lead to more
detailed obser vat i oris on predation by mononchs.
Investigations were made on the predatory behaviour
<GrDDtaert Zt Maertens, i976j Bilgrami et al . , 1984);
predation abilities (Small & Grootaert, 1933); e-f ect
o-f roechanical stimulation and crowding on predation
iPizmi ?< Jairajpuri, 1979); factors influencing
predation (Nelmes, 1974; Bilgrami et_ al^. , i983j 1984);
strike rate of the predator and resistance and
susceptibility of P^^Y nematodes to predation
<Bilgrami, 1992); life cycle (Maertens, 1976>;
cannibalistic behaviour (Bilgrami Sc Jairajpuri, 1985)
and inter specific interactions (Bilgrami et al.,
1988).
In the present work experiments were made to
evaluate predatory potentials of Hylonchulus dentatus
using plant parasitic nematodes as prey-
MATERIALS AND METHODS
Mylonchulus dentatus was isolated, from the
soil collected, from the agriculture fields near
Jamalpur, Aligarh. The predators were cultured in
Petri-dishes containing 1% water agar (supplemented
with a small quantity of Lactogen roi1k powder) and
Rhabdi t i s sp. as prey. All experiments were performed

392
in small cavity blocks containing IV. water agar at 28
+ 2°C. Twenty -five individuals o-f Meloidoqyne
i ncoqni t a were subjected to predation by -five adult
H. dentatus. The number of prey killed or injured was
counted a-fter 24 h. Each e>:periment was replicated
•five times. Conditions remained same -for all
experiments unless mentioned otherwise.
Prey c at chi ng end -f eedi nq mechani sms; The prey
catching and feeding mechanisms of H. dentatus were
observed by inverting culture dishes containing
predators and prey over the stage of stereoscopic
microscope. Detailed studies on the prey catching and
feeding mechanisms were made at 400 x or higher
magnifications. Observation chambers similar to
Bilgrami et al. (1985) were made. Chambers consisted
of a glass slide with a 2 x 1.5 x 1 cm block of 1%
water agar. Predator and prey nematodes were released
in the agar block. The agar block was then covered
with a cover slip. Nematodes were allowed to
acclimitized for 30 minutes before observations were
started.
Rate of predati on by adult and juveni1es stages; To
find out the rate of predation by female and juvenile
stages of M. dentatus five individuals each of

93
different stage of predator were placed separately
with the juveniles o-f M, i ncpqni t a.
Feedi nq pattern; To determine predatory profile of M^
dentatus over a period of 16 days the predators were
placed with prey individuals. Observations on the
number of prey killed or ingested were made a-Fter 24
h. Each day after observations the predators were
transferred to cavity blocks containing fresh IX water
agar and same number of prey individuals.
Prey preference; To determine prey preference in H.
dentatus second stage juveniles of Heloi doqyne
i ncpqni t a, Tylenchulus semi penetrans, Heterodera mothi
and Anqui na t r i t i ci and adults of Hopl ol ai mus i ndi cus .,
Helicotylenchus i ndi cus, Tylenchorhynchus mashhoodi ,
Hirschmanniella oryz ae Basi ri a sp. , Hemi cri conerooi des
manqi f er ae^ Hewiicycli op h or a dhi render i , Xi phi nema
basi ri , Lonqi dorus sp. , and Paral onqi dorus ci tri were
subjected to predation in separate cavity blocks.
Effect of prey densi ty on the rate of predat i on; To
determine the effect of prey density on the rate of
predation 25, 50, 75, 100, 125, 150, 175 and 200
individuals of N. i ncoqni t a were subjected to
predation by M^ dentatus.

o 9 4
Ef -feet of st ar vat i on of predator on the rate of
predat i oni To observe the effect of starvation of M.
dentat us on the rate of predation the predators were
starved <kept in water without prey) for O (fresh), 2,
4, 65 S, 3D, 12 and 14 days. Each group of starving
predators was tested separately.
Effect of temperature on the rate of predat i on; To
study the effect of temperatures on the rate of
predation the prey individuals were subjected to
predation by H^ dentatus at 5, 10, 15, 20, 25, 30, 35
and 40 "C.
Ef feet of agar concentrat i ons on the rate of
predat i on; To determine the effect of different
concentrations of agar the prey individuals were
subjected to predation by H. dent atus in cavity blocks
containing 1, 2, 3, 4, 5 and 6'i water agar.
Ef feet of agar thi c knesses on the rate of predat i oni
Individuals of M. i ncogni t a were subjected to
predation by M. dentatus at 2, 4, 6, 8 and ID mm thick
agar layers to observe the effect of agar thicknesses
on the rate of predation.
Ef feet of pH on the rate of predat i on: To determine
the effect of pH of the medium on the rate of

'a 0 '
0 b J
predation by H. dent at us the prey irsdividuals were
subjected to predation in cavity blocks containing
water agar with pH 5.0, 5.4, 5.Q, 6.2, i>. h 7-0, 7.4,
7.8, 8.2, 8.6, 9.0-
E-f -feet of test Brena an the rst^ of pr&dat i on: To
determine the e-f-fect o-f sur-face area on the rate of
predation by H. dent at us 4 mm thick agar blocks o-f
0.5, .1.0, 1-5, 2.0, 2.5 and 3.0 cm radius were taken
in cavity blocks. The area of each block was
n calculated by jrr'^ y, thickness of the agar block-
RESULTS
Prey catching and -f eedi nq mechani sms
In the presence o-f prey nematodes M. dent atus
moved in a typical undulatory wave pattern in agar-
Predation depended on chance encounters with the
prey. No chemotactic activity was observed in
predators towards prey. Predators contacted prey
during locomotion, established lip contact and moved
its lip vigorously over the body of prey. To achieve
cuticle perforation predator attacked prey with its
buccal armature after probing as all attacks did not
result in prey wounding. Very often the prey moved
out of the grip of predators. The attacks were

396
supported by the buccal armature and muBculature. The
contracticn and relaxation o-f stomatal muBcles
manipulate the movement of buccal cavity. As a
result o-f contraction o-f labial muscles the lips arB
pulled outwards and backwards to open the mouth and
the vestibule (Coomans Sr Lima, 1965>. The dorsal
tooth and the denticles became fully exposed and ready
to attack and puncture the cuticle of prey» The prey
became immobile as soon as it was punctured. It was
shreded into pieces with the help o-f buccal armaturs
and consumed by the predators. Predator hold pj^&y
by tooth, denticles and lips during feeding. During
feeding hydrostatic pressure of the body of prey
decreased which resulted in a considerable shrinkage
of prey. The food, consisted of body fluids and
internal organs of prey, was ingested by repeated
action of oesophagus and feeding apparatus. The
oesophageal muscle helped the process of opening and
closing of the lumen of oesophagus- With the support
of these structures prey contents were ingested into
the oesophagus for onward movement to intestine. The
predator left the feeding site once the prey is
completely consumed either in search of another prey
or moved randomly in agar.

':; 0 7
Prey selecti on <Fig- 1)
H. dent at us e>:hibited preference when placed with
dif-ferent species of prey nefljatodes- Predators killed
maximum individuals of second stage juveniles o-f endo—
parasitic nematodes in comparison to other species o-f
prey <p < 0 . 0 5 ) . P. c i t r i and H. dhi renderi were
predated least upon by the predators (p < 0 . 0 5 ) .
Dther species of prey were killed in moderate
number s-
Rate o-f Gredati on by adul t snd juveni 1 e stages <Fi g.
2 , A)
All stages of H, de'nt at us were predaceous in
nature. They killed variable number of prey. Female
H. dentatus killed maximum number of prey <p < 0-05)
while the first stage juveniles the least. Feedi nq
pattern (Fig. 2, B)
There was no consistent predatory profile.
Predators killed miximum number of prey after 4th and
16th day and mi ni mum after eight and tenth days of
predation <p < 0 . 0 5 ) . There was no significant
difference in the rate of killing by the predators
between the first and tenth day of predation (p >
0 . 0 5 ) .

3:s
Ef ect of prey densi ty on the rate of predat i on (F i g-
2, C>
Prey density affected predation by M. dentatus.
More prey were killed when prey population was
increased from 25 to 200 individuals (p < 0.D5).
Maximum predation occurred in a population of 200 prey
individuals and isinimuffl in 25 individuals (p < 0.05).
Effect of starvat i on on the rate of predati on (Fig.
2, D)
The prey deprivation influenced the rate of
predation by M. dent atus- The rate of killing prey
increased with the increase in the period of
starvation <p < D.05>-
Ef feet of temperature on the rate of predati on (Fig,
3, A)
The change of temperature affected predation by
the predators. Temperatures between 25 and 30 "C were
optimum where maximum killing was recorded <p < 0-05).
Temperatures lower and higher than thsse declined
predati on -
Ef feet of agar concentrat i on on the rate of predat i on
<Fig. 3, B>
Different concentrations of agar affected rate of
predation by H. dentatus. M. i ncoqn i t a juveniles were

399 v.>
killed more in 1—27, concentration of water agar (p <
0.05>, Agar concentrations higher than these declined
predation Jp < 0.05). <Mo predation was recorded in 6/i
water agar.
E-f-f ect o-f agar thi cknes^s on the rate of predati on
(Fig. 3, C>
Similar to agar concentrations, thickness of agar
layers also in-fiuenced rate of predation by H.
dent at us. H. i ncoqni t a juveniles were killed Hjaximum
at 2-8 mm thick layers of agar <p < D.D5>. Agar
thickness more than these declined rate of predation
by H. dent atus.
Effect of pf-{ of the medi um on the rate of predat i on
(Fig. 3, D)
Rate of predation depended upon the pH of the
medium- Maximum predation occurred at pH 7.4 — 7.8.
More acidic and alkaline pH declined predation
significantly (p < 0.05).
Effect of test arena on the rate of predati on < F i g. 4 >
Test area also governed predation by M. dentatus.
Maximun* predation ocurred in small test areas of 0,31
- 1.3 cubic mm. As the size of test area increased
the rate of predation declined. Minimum predation was
recorded in test area of .11-3 cubic mm (p < O.Q5>)

4 C 0
DISCUSSION
The prey catching and feeding mechanismB are
coBjpareble to other mononchB with -few exceptions.
Predation fay H. dentatus depended upon chance
encDuniers with the prey. The same phenomenon has
si 50 been observed by Nelmes (3974); Cohn & Mordechai
<1974>; Grootaert 8f Haertens <3976>; Bilgrarai et. al . ,
19835 1984). Many mononchs e^ Q_. , Hononchus,
lotonchus, CobbonchuB, Sporonchulus, CIarku^ etc., are
capable of engulfing their prey whole but species of
Hylonchulus were not found to contain any species of
prey intact in their intestine CBilgraflii, et al . ,
1986). H. dent atus e>:hibited preference for the
juveniles of endo—parasitic nematodes- This
preferential behaviour of H. dentatus may be
attributed to lack of anti-predation adaptation. Endo
parasitic nematodes lack characteristics which provide
resisteince against predation (Bilgrarai it Jairajpuri,
1989a; Bilgraroi, 1990a, 1990bj Jairajpuri Sc Bilgrami,
1990). The ectoparasitic nematodes, provided with
physical, chemical and behavioural characteristics,
are able to protect the'mselves from predation by
different predators. Preferential behaviour was also
observed by Sbafqat et_ aj,. <1987}; Bilgrami &

4C1
Jairajpuri (1988); Bilgrarai tt Jairajpuri (19S9a);
Bilgrafiji (1997) . The increased rate of predatiori
by H. dentatus in increasing population of prey
indicates that predation depended on the number of
prey (Bilgrami et al . ., 1985; Yeates, 1969; Bilgrami S<
Jairajpuri, 1989c), Increased predation by W.
dent atus may be attributed to increased chance of
contacts between predmtor and prey. Bilgrami et al.
fi9S4> while working with Hononchus aguati cus did not
observe any effect of prey density on its rate of
predation. The differential rate of predation by M.
dentatus at different teaperatures and agar
concentrations may be the result of inhibited activity
of predators and prey as was observed by Bilgrarai et
al . <1983>; Bilgrami et. al_. <1985); Shafqat et. a^.
(1987); Bilgrami & Jairajpuri <1989b) and Khan et. al .
(1991) with Mononchus aquat i cus, Mononchoi des
1onqi caudataus, M, forti dens, Aquati des thornei ,
DoryIai mus st aqnali s and Aporcelaimellus ni valis.
Less predation in thick agar layers may have resulted
because the predators and prey got more area to move
which allowed fewer contacts to establish between
them.
During present observations adult predators
killed more prey than their younger stages. This

4 C 2
differential rate of predation reflect, their
predatory potentials. Bilgrami et al, (1984) also
r&cor-ded higher rate of predation by adult H.
aquat i cus than juveniles. More predation in small test
arena suggest that small surface area provided
predators more prey per unit area to attack than the
1arge area.
ACKNOWLEDGEMENTS
The authors are grateful to The Department of
Science and Technology, New Delhi for the financial
assi stance.

A n ••) ^ w o
REFERENCES
AZMI, K- I. Z: JAIRAaPLIRI, M. S. <1979). Effect of
mechanical B-timulation and crowding on
Mylonchulus dentatus. Nematol. Medi t. 7 : 203-
207.
BILGRAHI, A. L. (1998a>. Piploqasterid predators; In
nematode bio-controlzAspects and Prospects (eds
Jairajpuri et-al.) CBS publishers It
Distributors, New Delhi pp. 143-148.
BILGRAMI, A. L- <199ab>. Predaceous dorvlaims; In
Nematode Bio-controls Aspects and Prospects ieds
Jairajpuri et. al . > CSS publishers ?f
Distributors Pvt. Ltd. New Delhi pp. 143-148.
BILGRAMI, A. L- <1992>. Resistance and susceptibility
o-f prey nematodes to predation and strike
rate o-f the predators Mononchus aquat i cus,
Dorylai reus staqnali 5 and Aquati des thornei.
Fundamental and Appli ed Newatoloqy 15 : 265—
270.
BILGRAMI, A. L. & JAIRAJPURI, M. S. <19S5>.
Cannibalism in Mononchus aquat i cus. Indi an J.
Nematol. 14 (1984) : 202-203,

40 4
PILGRAMI, A. L. ?< JAIRAJPURI, M. B. (1988).
Attraction o-f Mononchoi des lonqicaudatuB and W.
•fort i dens (Nematada? Di pi ogast er i da ) towards
prey and factors i nil uen{zi ng attraction. Revue
Nematol . lis 195-202.
BILGRA^'.I, A. L. ?. JAIRAJPURI, M. S. (1989a>.
Resistance o-f prey to predation and strike rate
of the predators, Hononchoi des 1onqi caudatus and
M. fort i dens < nerjatoda j Di pi ogasr ter i da > . Revue
Nematol. 12 s 45-49.
BILGRAMI, A. L. ?. JAIRAJPURI, M. S. <1989b). Effects
of concentrations and thicJtness of agar on the
prede-tory behaviour of Wononchoi des
1onqicaudatus and M.fort i dens (Nematoda
JDipiogasterida> and on their attraction
towards prey. Indi an J.Nematol <19B8).IB;222-
227.
BILGRAMI, A. L. f< JAIRAJPURI, M. S. (1989c)-
Predatory abilities of Hononchoi des fort i dens
and H.Ionqi caudatus <NematodasDipiogasterida)
and factors influencing predati on.Nematoloqlea
35 J 475-488.
BILGRAMI, A. L., AHMAD, I. ?< JAIRAJPURI, M- S.
(1983). Some factors influencing predation by

4::5
MononchuB aquati cus. Revue Nematol. 6 i
325- Z'ZL.
BILBRAMI, A. L. , AHMAD, I. «t JAIRAJPURI, M. S.
<1984>. Observations on the predatory
behaviour o-f Mononchus aquat i cus. Nematol .
Hedit. 12 s41-45.
BILGRAMI, A. L-, AHHAD, I. h JAIRAJPURI, M. S.
(1985). Predatory behaviour o-f Aquat i des
thornei <^4ygDl ai mi na : Nematode). Nematol oqi ca
(1984) 30 : 457-462.
BILGRAMI, A. L., AHMAD, I. ?< JAIRAJPURI, M. S.
<193A). A study D-f the intestinal contents of
some mononchs. Revue Nematol. 9 : 191-
194.
BILGRAMI, A. L., AHMAD, I. & JAIRAJPURI, M. S-
(1989). Interaction between mononchs in non-
sterile agar plates. Indi an J. Nematol . 1"?
(19B7) : 339-340.
COBB, N. A. <1917>. The mononchs: a genus of free
living predatory nematodes. Soi1 Sci. 3 : 431-
486.

4r o
COHN, E- ?< MORDECHAI, H. <1974>. Experiments in
suppressing citrus nematode populations by use
of merigold and a predaceous nematode.
Nematol. Hedit. 2 : 43-53.
CODMANS, A. ?< LIMA, B. <i965>. Description oi
Anatonchus ami ci ae n- sp. (Nematoda:
Mcnonchidae) with observations on its juvenile
stages and anatomy. Nematoloqi ca 11 : 413 —
431,
ESSER, R. P. <1963>. Nematode interactions in plates
of non-sterile wiater agar. Proc. Soi 1 Crop
Sci. Soc. Fla. 23 : 121-128.
GROOTAERT, P. S< MAERTENS, D. <1976>. Cultivation and
li-fe cycle of Mononchus aquat i cus.
Nematoloqica 22 j 173-181,
JAIRAJPURI, M. S. ?< BILGRAMI, A. L- <1990}.
Predatory nematodes. In Nematode Bio-control :
Aspects and Prospects (Eds Jairajpuri et al.)
CBS Publishers & Distributers Pvt. Ltd.,New
Delhi pp 95-125.
KHAN, Z., BILGRAMI, A. L. ^ JAIRAJPURI, M. S. <1992).
Some observations on the predation ability of
Apporcelaimellus ni vali s (Altber, 1952) Heyns,

4 C 7
1966 <Nematoda : Dory1aimi d a ) . Nematoloqi ca^
37 : 333-342-
HAERTEiMB, D. <1976>. Observations on the life cycle
of Pri onchulus punctatus (Cobb, 1917) Nematoda
and Bome culture conditions. Bi ol. Jaarb. 43 !
197-21S.
NELMES, A. J. <1974>. Evaluation of the feeding
behaviour of Pri onchulus punctatus (Cobb) , A
nematode predator. J., Ani m Ecol . 43 : 553—565.
SHAFQAT, S., BILGRAMI, A. L. & JAIRAJPURI, r1. S.
(1987). Evaluation of the predation ability of
Dorylai mus st aqnali s Dujarding, 1845
(Nematoda j Dory3aimi da>. Revue Nematol. 10 :
455-461.
SMALL, R. W. S< 6R00TAERT, P. <1983>. Observations on
the predation abilities of some soil dwelling
predatory nematodes. Nematoloqi ca 29: 119-125.
VEATES, G. W. <1969>. Predation by Hononchoides
potohi kus (Nematoda : Dipiogasterida) in
laboratory culture. Nematoloqi ca. 15 : 1-9.

4 C 8
LEGENDS
F a g - 1 , P r e y p r e - f e r e n c e by M-_ d e n t a t u s
A H, i ncoqni ta; B - T . semi penetrans; H. mothi j D - A. tritici ; E - HoplD1ai mus i ndi cus; F - HelicotYlenchus indieus; G - T. mashhoodi j H -H. ory:: ae; I - B a s i r i a s p, ; J — H. manqi -ferae; K - H- dhi render i ; L ~ X. basi ri; M -Lonqi dorus sp. ; N - P. c i tri ,
Fig, 2 A - Rate o-f predation by different stages o-f H. dentatus = Female; = Fourth stage
B
C
D
juveniles; J ^ = Third stage juveniles; Jrj = Second stage" juveniles; Jj = First stage juveni1es.
— Predatory pattern o-f M. dent at us
— Ef-fect o-f prey density on the rate o-f predation by H. dentatus
— E-f-fect o-f starvation o-f predators on the rat
o-f predation
Fig. 3. A -Ef-fect o-f temperature on the rate o-f predation by M. dent atus.
B - Ef-fect o-f agar concentrations on the rate o-f predation by M. dent atus.
C - E-f-fect o-f agar thicknesses on the rate o-f predation by M_^ dentatus.
D - Effect of pH of the medium on the rate of predation by M_^ dentatus.
Fig. 4. Effect of Test arena on the rate of predation by M. dentatus

409
T iX) ^ 0 0 -J
peil!>l Aejd jO'ON
o
LL
LU
Q
o
m
<
Q. if)
CD
K' 6"
U^

4 1 0
2k
x> 18j a*
a.
12
9 -J o
16(JI
80
o 40 <
o
'3
/ •
A 2A
22.
2(lJ
18
J, J' J- J 1 )f
C 24
12
/
25 50 75 100125150175200 Prey density
/ \ / e
•
\ / "
2 4 5 1 — i — 1 — i — I 8 10 12 14 16 Days
• • D
"1 I I I \ 1 1 1
0 2 4 6 8 10 12 K Starvati'on(days)
r

^^] 1
0;
2L
laJ
ii
5. o
24
laJ
11
0
a
1 2 3 A 5 6 Agar concentratfons(%)
D
T 1 — 1 — 1 — I 1 [
2 4 6 8 10 12 1A 16 Agar thicknesses (mm)
1 r-i I 1 1 r 50 5,8 6.6 U
pH
I I I I 8.2 9.0
^

X)
2A"
laJ
4 1 2
12 CD i _
- 6 •
031 13 28 5.0 7.8 113
Test arena (mm3)
^'^3- "-1

4 1 3
OBSERVATIDfaS ON THE PREDATION ABILITIES OE NEOACTINOLAIHUS AGILIS <DORYLAIMIDAi ACTINOLAIMOIDEA)
Z. KHAW, ANWAR L. BILGRAHI and M. SHAMIM JAIRAJPURI
Institute o-f Agriculture, Sha-fi House, Qi 1 a Road, Aligarh Muslim University, Aligarh-202D02,
Abstract: Observations were made on the prey catching and feeding mechanisms, rate o-f predation on different species of plant parasitic nematodes and factors influencing paredation Neoact i nolai mus aqilis. Predation occurred mostly upon contacts with athe prey individuals- N. aqi1i s paralysed its prey by holding the prey with everted onchia and lip region, puncturing cuticle with the odontostyie and disorganizing the internal organs of prey. The attacks were indiscriminate ocuring all over the body. Oesophagus pulsated at regular interval and secreted oesophageal secsretions for extra corporeal digestion. At the end of feeding the whole of the prey contents were ingested and only the cuticle left unconsmed by the predators. The reate of predation remained same over a period of eight days. N. aqilis preferred the second stage juveniles of M. i ncoqni ta , A. tri ti ci and T. smi penetrans. P. ci tri < R. robustus and Helicotylenchus i n d i c u s were killed the least. N. a q i1J s failed to either kill or injure any individual of Hop1olai mus i ndi cus or H. manqi ferae. Predation increased with the increase in prey number but decreased with the increase in concentration of agar. Maximum predation occured at 25—30 "C. Temperatures lower or higher than these declined predation significantly.
Key words: Neoact inolaimus aqilis, predation, behaviour, predators, prey.
Several studies have been made on predation
abilities of mononch, dorylaim and dipiogasterid
predators. Yeates (1969) made a detailed study on

4 1 4
predation by a dipiogasterid nematode, Di pienteron
potohi kus. Nelmes (1974) and Cohn ^ Mordechai <1974>
used mononcbs, Prjonchulus punct atus and HylonchulUP
sigmeturus -for their studies on predation. The prey
catching and feeding mechanisms o-f Labr onema
vulvapapillatum^ Hononchus aquati cs Aquati des
thornei , Hononchoi des lonqi caudatus and M- fort i dens„
Dory1ai mus stapnali s and Aporcelai mel1 us ni vali s were
studied in detail by Wyss & Grootaer (1977)| Bilgrami
et al. <19S4>5 <1985>5 Bilgrami ^ Jairajpuri <1990};
5ha-fqat e ^ ai_. (1987); Khan et al . <i991>. Bilgrami et
al. (1983) and Bilgrami and Jairajpuri <1989a)
observed the e-ffects o-f different factors on the
predation by nematodes- The resistance and
susceptibility of prey nematodes to predation and
strike rate of the predators were measured by Bilgrami
?r Jairajpuri (19a9b>; Bilgrami <1992) while
relationships between predation and different prey
trophic groups were determined by Bilgrami {1993>-
S D far, such studies are lacking on actinolaim
predators and no informations available on their prey
catching and feeding mechanisms, predation abilities
and factors influencing predation. It was therefore,
the present studies were made on an actinolaim

4 1 5
predator, Neoact inolaimus aqilis to evaluate its
predatory poterrtials using plant parasitic nematodes
as prey.
MATERIALS AND METHODS
Neoacti nolai mus aqilis, isolated from the soil
around roots of mango tree, were cultured in petri—
dishes containing 1% water agar using Rhabdi t i s sp- as
prey. The culture dishes were supplemented with a
little amount (10 - 20 mg> of infant milk powder
(Lactogen) in order to encourage bacteria to grow-
Bacteria served as food for the prey nematodes.
In all the experiments 25 individuals of prey
nematodes were subjected to predation by five adult
individuals of N. aqilis. The observations on the
number of prey individuals killed or injured were made
after 24 h. All experiments were made in small cavity
blocks containing IX water agar at 28 + 2"C. The
experiments were replicated five times. During all
experiments except rate of feeding on different pf^Y
species, predation by N. aqilis was tested on the
juveniles of Tylenchulus semi penetrans and adults of
Tylenchorhynchus mashhoodi which were collected from
the soil and roots of citrus and paddy respectively.

A] 6
Prey cat chi nq and -f eedi nq mechani Bms; Prey catching
and -feeding mecbaniB/nB o-f N. aqi lis was studied by
inverting the culture dishes over the stage o-f
stereoscopic microscope at 45 - 60 x magni-f i cat i ons.
To study the -finer details o-f these Biechanisms
observati on chambers similar to Bi Igrami et al_, /I 995)
were made. Observation chambers were consisted o-f a
glass slide with 2 y, 1.5 x 0.5 cm block of 1% water
agar. The predators and prey were le-ft in the block
which was then gently covered with a cover—slip o-f
equal length- The sides o-f the slide was sealed with
tissue or -filter paper. Observations on the prey
catching and -feeding mechanisms of N. aqi lis were made
U51 ng JO individuals of Rhabdi ti s sp. , and ID
individuals of predators. Observations were started
after about half an hour after when the predators and
prey were released in the chamber. Observations were
made at high maginifications ranging from 100 to 400
Rate of feedi nq on di fferent speci es of prey? To
determine the rate of feeding of N. aqilis, five
individuals of predators were released in cavity
blocks containing agar and 25 individuals of prey
nematodes. Rate of predation by N. aqilis was

^\1
determined on plant parasitic nematodes v i z . , second
stage juveniles D = Hel oi doqyne i ncoqni \.a ^ Tyl enchul us
semi penetrans» Anqui r\Si tritici , and the adult
individuals o-f Tyl f?nchQrhvnchu5 washhoodi ,
Hi rschmenni el 1 a oryz ae » Aphelenchoi des sp. , Xi phi nema
ameri canum. Paralonpi dorus ci tri , Paratri chodorus sp . ,
Rotylenchus robustus, Hopl ol ai mus i ndi cus,
Hemi cr i conemoi des gtangi -ferae and Hel i cotyl enchus
i ndi cus- Each species of p>^ey was tested separately.
The prey nenjatodes were isolated -fresh either -fronj the
soil or root samples -for each experiments.
Ef-f ect o-f prey densi ty; To determine the effect o-f
prey density on the rate of predation by N. aqilis
predators were released in cavity blocks containing
25 , 5 0 , 7 5 , 100, 125, 150 175 or 200 individuals of T,
semi penetrans or T. mashhoodi.
Effect of agar concentrat i ons; To find out the effect
of agar concentrations on the rate of predation by N.
aqi lis the predators were r€?l eased with prey nematodes
in cavity blocks containing 0.5, 1, 1-5, 2 , 2.5, 3,
3-5, 4, 4.5, 5, 5.5 or hf. water agar.
Ef feet of temperature; To study the effect of
temperature on the rate of predation by N. aqilis the

418
predators were ie-ft in cavity blocks containing
specimens D-f prey- Predation was tested at different
temperaturs viz-, 5, 10, 15, 20, 25, 30, 35 and 40 C".
Predatory pattern; The predatory profile of the
predators was studied over a period of eight days. To
study predatory pattern the prey nematodes were
subjected to predation by f4. a n i 1 i s in cavity blockr
containing agar. Observations on the number of prey
killed or injured were made after 24 h. After
observations the same predators were transferred to
another cavity block containing equal number of fresh
specimens of the same species- Similarly each day
after observations the predators were changed to fresh
cavity blocks containing fresh prey.
RESULTS
Predators made contact with the prey nematodes
while moving randomly in the* agar. Upon contacting a
prey nematode of a suitable size with the lip region,
the odontostyle was immediately protruded and
penetrated the cuticle at the first thrust. Soon
after thrusting and penetration the prey became
immobile and paralysed. The prey 'at this stage
appeared to have been gripped by the everted onchia

4 1 9
Bnd the lip regiori. Predators did not show any
preference -for body to initiate an attack. The
attacks were made all over the body o-f prey depending
upon the are& first contacted- It was therefore the
initial penetration occurred wherever the prey
happened to be first contacted. The odontostyle was
thrusted about half of its lenght into the body of
prey and simultaneously the oesophageal bulb started
oesophageal pulsations-
The oesophageal bulb pulsataed at regular
intervals of 5-10 contractions over a period of few
seconds. During pumping, granules could be seen
streaming into the odontostyle, down the oesophageal
lumen and finally into the intestine- Secretions from
the oesophageal glands were observed passing into the
prey for the purpose of extra corporeal digestion.
After feeding when the whole of the prey contents were
consumed the odontostyle was withdrawn and predator
left the feeding site. However, if the contents
remain in the body of prey the predators some time
withdrew its odontostyle and penetrated a new spot in
order to consume whole of the prey. Feeding generally
depended upon the size of prey- The body cuticle of
adult Rhabdi t i s sp. was left unconsumed when the
feeding was over but predators shreded the body of

420
small prey individuals (juveniles) into two or three
pieces before -feeding.
Rate o-f -f eedi nq on di f-f erent t^peci es of prey (Fig. i)j
The rate of predation by N. aqilis varied an different
species of plant parasitic nematodes. The second
stages of H. i ncocni ta» A. tri ti ci and T.
setni penetrans were killed in may, i miim nu/nbers (p <
0 . 0 5 ) . Predators killed individuals ofP, ci tr i R,
robustus and Meli cotylenchus i n d i c u s the least (p <
0 . 0 5 ) . The adult individuals of T. mashhoodi, H.
oryg ae, Anhelenchoi des sp- , Paratri chodorus sp. and
X. simf^ri canum, were killed in nioderate numbers by the
predators. N. aqi lis failed to either kill or injure
any individual of HopIolai mus i n d i c u s and H.
B»anqi ferae.
Ef feet of prey densi ty (Fig- 2 A) : The number of
prey individuals affected rate of predation by N.
aqilis. Predation increased with the increase in the
number of prey (p < 0.05) upto a population of 200
individuals. There was a significant difference in
the rate of predation with 25 or 200 prey individuals
<p < 0 . 0 5 ) .
Ef feet of agar concentrat i ons (Fig. 2 B) : Various
cnncenfrat 1ons of aqsr governed predation by N.
aq i } i s. Lower concentrations (1-1.57.) were found most

421
suitable, Ma>! i mum predation took place at these agar
concentrations (p < 0.05). ConcentratiDOB higher than
these declined predation (p > 0,05).
E-f-f ect o'f temperature <Fig, 2 C>, The changes in
temperature regiwe a-ffected rate of predation by N.
aqilis. Maximum predation occurred at temperatures
ranging -from 25 to 30 "C. Temperatures lower or
higher than these declined predation significantly <p
< 0.05).
Predatory pattern (Fig- 2 D>: N. aqilis exhibited
identical predatory pattern over a period o-f eight
days. The rate of predation remai ned ^ame each day.
There was no dif-ference in the rate o-f predation by
the predators after one or eight days (p > 0,05)-
DISCUSSION
It is evident from the present observations that
N. aqi1i s is predaceous in nature- Predation by N-
aqilis occured as the physical contacts between prey
and predators were made. The prey catching and
feeding mechanisms of N. aqilis resembles mostly to
dorylairo predators (Wyss J* Grootaert, 1977; Bilgrami
et SLL- 1, 19fl5; Shafqat et. al_. , 1987; Khan et_ aJL- ,
1991). The actinolaim predators may only differ with
dorylaim predators in respect to the time and number
of odontostyle thrusts and the duration of feeding-

4 2 2
Aphelenchid predators paralyse their prey by
injecting oesophageal secretions I'Lin-ford ?f Oiiviera,
2937; Hechler, 1 9 6 3 ) . Dipiogasterid predators held
their prey with their buccal =''{nettur e as vjell as the
oesophageal suction <Silgrafni S< Jairajpuri, 1 9 9 D ) .
The nygolainj and dorylaiim predators disorganise the
body organs o-f the prey with the help o-f odontostyle
in order to get hold o-F them (Bilgrami et al . , 1 9 8 5 ) .
hi. a q i ? i s paralysed its prey by holding it vyiith
onchi a and lip region,, puncturing cuticle with
odontostyle and disorganising the internal body organs
D-f the prey- Similar to di pi ogast er i d and dorylaim
predators N. agilis also injected oesophageal
secretions into the body of prey, probably for the
purpose of extra-corporeai digestion.
Yeates (1969) j Nel mes {1974) ; Bilgrami et_ aJL-
(1985); Bilgrami Sr Jairajpuri <1990> and Khan e ^ al .
(1991) observed an increase in the rate of predation
by Di pienteron colobocercus, Pr i onchulus punct at us <
Aquat i des t hornei , Mononchoides 1onqi caudatus;
M o n on c h o i d e q fori i dens and Apor eel ai <nel 1 u s nivalis
with the increase in prey number. Present
observations with N. aqi !{ i s also indicate similar
results and conform well with the above observations-
Thir- relationship between the predation and prey

4 2 3
density appeared to be due to the if^proved chance o^
contacts between predators and prey as was suggested
by BHqrami it Jairajpuri (1990). Activity of
predators and prey govern predation. The rate o-f
pr&dBtion increased with the increase and declined
with the decrease in activity o-f predators and prey
<Siigraffli et al . 1 9 3 3 ) , The low rate o-f predation by
!4. a q i 1 i s at high concentrations and at low and high
temperatures may be attributed to the inhibited
activity o-F predators and prey as high concentrations
o-f agar and temperatures sre known to inhibit the
acti-vity o-f nematodes (Bilgrami et aj_, , 1 9 8 3 ) , Due to
this inhibited activity the predators and prey could
have made only fewer contacts among themselves which
resulted in less predation.
The rate o-f predation varied on di-f-ferent
species o-f plant parasitic nematodes as N. a q i 1 i s
showed pre-ference -for dif-ferent prey-. The high rate
of killing of the second stage juveniles of M.
i ncogni t a, A. tr i t i c i and T. semi penetr ans may be
attributed to the lack of anti-predation
charcteristics in these prey individuals (Bilgrami &
Jairajpuri, ]989b>. Mo predation on Hoplolai mus
i nri j cus and H. mangi ffrae could have resulted due to

4 2 4
i f- «=i ^ h i 1 i h y t o a v o i r ? pr- <=>t;? r^i i «*n h ipc i «-« "^f? o f i i: s t h i c «f
h o d y t" M ^ i c 3 6? f*nri Ytrxriy r=* ri n *«1 i=t r j Cj n ^ ( P h y B i c a 3
r- fri c; j c;}- ^,-,,- t^ . p i 1 fj t'" '="** •» ^ ,7 ^ i € a J p !.< r i J 1 9 S 9 b > . V a i i o u B
i-»i-hf3t^ wo i ' ^ j ^ i f «s !>^vf> ;:»lt,», nnV r *-> r: o ••" < J t? rl ^ n y p « r e d a i : i D n o n
^4ort « o l .M i «>>>*=; -i t tH « <-> i^ f Hf'4»< J!-riHf" i- f^ i f=> * . , 1 9 7 7 ; S i l c r a ^ n i
v a ^ ( ? ^ j n . . r i - 1 9 9 r . ; R ; 1 cjr i=.(ni J 1 9 9 2 , B i i g r a m j . 1 9 9 3 ) .
• n i f i t i l ^ r l v . K•=><i«"«•»^'^i'l*=»nr•5^^^c;. i r n i i c u * ? - Eihows- c h e m i c a l « /
«'"6*s^'s<-, Rttc*^ iF'?-5;«?Fj 1 9 A 3 ; B i l t j r s m i ? J a i r a j p u r i ,
1 9 p 9 h * . D«ir i n f j p<-*='^*=>ni: o h s e r v B t i o r j - ^ l o w r a t e o-f
prf=>fl,=»h ^ o n h y M - e n t « < - o n t-i •->» i c o h y 1 «?r» c o < < f i n <j a c J,' B may
a i ? ^ o b«=» a iJ^ ih«^^«?^-5 fco t o . > ; i r o r o n a t t f a c t i v e c h e m i c a l
B e c r - e h i o o B o-f p r e y ss- eiuQoe«?.t ecS b y E s s e r ( 1 9 6 3 5
G r o o V f ^ e r - t eh ^ 1 . { 1 9 7 7 ) j S i l y r a m i ^ J a i r a j p u r i < 1 9 8 8 5
1 9 8 9 b J .
ACKNOWl FDGEMENTS
Tl«e -f i r "^t a u t h o r i "^ g r a t e - f n l t o C o u n c i l o f
F5< i e i « J i f i i « - ar»H T»Hl«««^if i a l R e - ^ ^ e a r r h , N e w D e l h i etttd t h e
" ^ e r o o d t f» P/ep.^i' h mef»} o f R r . i e o r e a n d T e c l m o l o g y , New
D e l h i f«">r t i t e f i i » a r » < - i a l a ' ^ 's i s I: a o r e .

4 2 5
REFERENCES
BILGRAMI, A. L. <1992>. Resistance and susceptifai 1 ity
of prey nematodes to predation and strike rate
of the predators Hononchus aquati cus<
Dorylai mus st aqnali s and Aquat i des thornei,
Fund. App, Newatg?. 15i 265-270.
BILGRAMI, A. L. n993>. Relationship between
predation fay Apor eel ai ?pel 1 us nivalis and
di-f-ferent P^^Y trophic categories,
Nematoloqi ca 39 : 356-365.
BILGRAMI, A. L- ?. JAIRAJPURI, M. S. U9e9a>. Ef + ects
o-f concentrations and thickness o-f agar on the
predatory behaviour o-f Hononchoi des
lonqicaudatus and M. -fort i dens ^Nematoda
:Dipiogasterida> and on their attraction
towards prey. Indi an J. Nematol . <1988) 1_S :
222-227.
BILGRAMI, A. L. ?f JAIRAJPURI, M- S. a9S9b).
Resistance of prey to predation and strike rate
of the predators, Mononchoj des lonqicaudatus and
M. fort i dens (Neroatoda:Dipiogasrterida) . Revue
Neroatol. 12; 45-49.
BILGRAMI,, A. L- ?* JAIRAJPURI, M. S- <1990). Predation
abilities of Hnnonrhoi des }nnqi caudatus and M,

4 ^ 0
•fort i dens iNematoda; Di pi ogast er i da > and
-factors influencing predation. Nemat ol oqi xiSi
35; 475-4S8.
BILGRAMI, A- L., AHMAD, I. & JAIRAJPURI, M. S- (1983).
Some ^ ei^\i!3r<E> i n-f 1 tjenci ng predation by Hononchus
a Qu at i c»^ . Revue Nematol , h_ : 325—326.
BILGRAMI, A- L., AHHAD, I. %< JAIRAJPURI, M. S. <i9S4).
ObBervations on the predatory behaviour of
Hononchus aquat i cus. Nematol . hedi t. 12; 41 —
45.
BILGRAMI, A. L., AHMAD, I. & JAIRAJPURI, H. S. <19S5>.
Predatory he\\&vi our a-f Aquat i des thornei
<Wygolaimina; Neroatoda). Nematoloqi c a (1984)
30; 457-462-
COHN, E. Sc MORDECHAI, M. (1974). Experiments in
suppressing citrus nematode populations by use
of merjgold and a predaceous nematode.
Nematol . Medi te. 2; 43-53.
ESSER, R. P . ( 1 9 6 3 ) . Nematode interactions in plates
of non—sterile water—agar. Proc. Soi1 Crop
5ci. Fla. 23 s 121-138.
GROOTAERT, P., JAQUES, A. ^ SMALL, R. W- (1977). Prey
selection in But 1er i us sp. (Rhabditida :

^27
Di pi Dgasteri dae> . 'J^'SK^'SA , Facu3 t . Landbouww
Ri jttsuni v, Ggnt 42 s 1559,
HECHLER, H. C. n 9 6 3 ) - Description, developmental
biology and -feeding habits of Sei nur a
tenui 'cSkUdB^^st (de Man), J. B. Goadey •, 1960
(Neroatoda i Aphel enchi da > a nenjetode predator-
PrPC. helfl>inthol. Soc. yash. 30 : 102-195.
KHAN, Z. , STLGRAMI, A. L, *r JAIRAJPURI, M- S. <I99i>.
Some studies on the predation abilities of
Aporcelaimellus ni veli s (Nematoda :
Dory1 aimi da> . Nematolopi cs 37 5 333-342.
LINFORD, M. B. ^ Oliviera, J. «. (1937). The deeding
o-f hollow spear nematodes on other nematodes.
Science 85 s 295-297-
NELMES, A. J. <1974>. Evaluation of the -feeding
behaviour of Prionchulus punct atus <Cobb> , A
nematode predator. J. Ani m. Ecol . 43 : 553—565.
SHAFQAT, S., BILGRAMI, A. L. & JAIRAJPURI, M. S-
(1987). Evaluation of the predation ability of
Dor y 1 ai m<,<s st aqnal i s Dujarding, 1845
*'Nematoda s Dor y 1 ai mi da > , Revue Nematol . 1 0 :
455-461 -

4 2 8
WYSS, U- 8c GROOTAERT, P. <1977>- Feeding mechanism of
Labronema sp. Heded, Fac. Landbouwv* Ri jkuniv.
Gent 42 j 1521-1527.
YEATES, G. W- U969>. Predation by Hononchoides
potohi ku^ <Nematoda : Dipiogasteridae) in
laboratory culture, Nematologi ca 15 : 1-9.

429
LEGENDS
Fig. 1. Rate o^ predation by Neoac t i rfol ai mus aqi lis on different species o-f plant parasitic nemat odes
A = JAel oi doqyne i ncoqni t a B = Tvlenchulus semi penetrans C = AnQui na tr i t i c i D = A p h e l e n c h o i d e s s p . E = T y l e n c h o r h y n c h u s m a s h h o o d i F = Hi r s c i ' m i a n n i e l 1 a o r y ? a e G = Paratri chodorus sp, H = Xi phi nema amer- i canum I = P6fr &1 onqi dor »s c i t r i J = R o t y T e n c h t t s r o b u s t t t s K = H e l i c o t y l e n c h u s i n d i C U E L = H o p \ o i a i m u s i n d i c »_ s M = Hemi cr i conewoi des manmiferae
Fig. 2. Factors influencing predation by Neoactinolaimus aqilis
A = Effect of number of prey on the rste of predat i on
B = Effect of aqar concentrations on the rate of predation
C = Effect of temperatures on the rate of predat i on
D = Predatory pattern over a period of ei ght days

4 30
20-
18-
16-
5 U QJ
H2H
^10-
° fl-d ^ ^ 6-
i-
2-LL
n
n
A B C D E F 6 H Type of Pf'ey
I J K L M
./' /^. r J-

4 3
^5i
X3 (U
.^35-1
>^
O
6 ^15H
22i
18'
<v
6-
o 6
2-
- /^
rans e—o T.semipenet
X—^ T.mashhoodi
50 ' 100 ' 150 ' 200 No. of prey
1 I r
10 2 0 30 AO Temperature(°c)
20-
15-
12
8-
A-
T 1 1 1 1 1 1 1 1 I
1 2 3 A 5 Agar concentrat ion (%)
22-1
18-
U
10-
6-
2-
> . ^ -K
n 1 1 1 ! 1 1 1 1
1 3 5 7 9 \^o. of days
f ^v

4 3 2
FEEDING AND ATTRACTION BEHAVIOUR OF TYLENCH0RHYNCHLI5 HA5HH00DI
By
Anwar L- Bilgrami
Section D ^ NematolDgy, Def«artment of Zoology, Aiigarh H U B ! i m University, A2igarb-202002, India
ABSTRACT
Observations on the -feeding and attraction behaviour of Tylenchorhynchus mashhoodi were made using cabbage, carrot, radish and onion roots as host-Feeding of T. fflBshhood i may be divided into five more or less distinct phases viz., widespread e>: p i or at i on , local e>i pi or at i on , stylet thrusting, salivation and feeding. T. mashhoodi attracted positively towards different plant roots but its responses varied with the type of the host. Ma:-!imum attraction occurred towards cabbage seedlings when tested alone or in combination with other hosts. Lower concentrations of cabbage root homogenate attracted lesser number of T. mashhoodi but the degree of attraction increased with the increase in homogenate concentration. hayi i mum attraction was recorded towards SO percent homogenate concentration.
KEY WORDSi Tyl enchorhynchus mashhood i ., plant parasitic nematode, behaviour, attraction, roots.
Several observations were made on the attraction
and feeding behaviour of plan parasitic nematodes
<kbera ?•: Zu kerman, 1963; Armi ?< Jairajpuri, 1977;
Prot, 1980; Wyss, 1981; Prot ?< Van Gundy, 1981a,
1981b; Prot & korprobst, 1 9 S 3 ) . Krusberg (1959)
studied feeding behaviour of Tylenchorhynchus
r a 1 y t tj n i . Wyss <1971> antJ <i973) described different

4 3 3
phases in iibe -feeding o-f Tri ch odor us- si yni 1 i s and
Tyl enchorh vnchus d u b i us respectively. Doncsster S?
Beyroour 0 9 7 3 ) characterized the probable succession
D-f stimuli and the responses leading a nematode to
feed on its host. In He>j at yl us viviparous passive
ingestion was reported by Doncaster ?c Seymour <1974>.
Barbatola S? Bridge <1980) observed -feeding behaviour
o-f H?. >- schmanni el 1 a or yz ae H. spinicaudata and H,
1 mrviBnvr i adults and juveniles on rice. The factors
influencing attraction of Hirschmanniella oryz ae
towards cabbage seedlings was studied by Biigrami et
Sil . <i935a>^ its responses towards different host
roots by Biigrami et al. il9S5b) and attraction and
feeding behaviour by Biigrami et al. <19S5c>-
Bilgrami et al. (1986) studied the effects of
different plants singly and in combinations on the
attraction of adult Xi ph i nema basi r i.
In the present worJt observations were made on the
attraction and feeding behaviour of Tylenchorhynchus
mashhoodi.
MATERIALS AND METHODS
Soil samples containing Tylenchorhynchus
mashhoodi were collected from around the roots of

4 54
cabbage. The nematodeB were extracted by sieving and
decantation and Baermann's funnel methods- The
nematodes prior to ex p«er i ment 5 v;ere surface sterilized
with O.DIX mercuric chloride. The cabbage roots grown
in 0-5/i water sqsir were surface sterilized in 0.1%
mercuric chloride solution. The agar was prepared by
autoclaving at 15 lb pressure for 30 minutes.
Experiments were carried out at 25 + 2''C. For all
ex-periments fresh nematodes and roots were used-
Feedi nq mechani sm; To observe the feeding mechanism
of T. mashhoodi observation chambers similar to those
used by Wyss & Grootaert <I977) and Bilgrami et al-
(1985c> were made- The observation chambers were
consisted of glass slide with a 0.5 cm thick layer of
0-5% water agar. The agar was cut into a block of 2y.
1.5 cm when cooled- A 3 cm long excised root of a 4-6
day old cabbage seedlings was placed in the middle of
agar- Twenty five adult T. mashhoodi were placed
along the edges of the agar. A cover slip was then
gentely placed over the agar and the sides were sealed
with tissue paper to prevent drying-
Attracti on of T. roashhood i towards single host root;
Petri dishes 5.5 cm in diameter were marked at bottom
into three zones by drawing two concentric circles of

0.5 and 2.5 cm diameter. The later was termed as
refererfce cir zle. One cm excised root -frora A—6 day
old t, -&iJlings i-;ft=i pleci=<^ in the inner circle of Petri —
dishes containing 0.5 cm thick layer of 0.5X water
sqsr. After 12 h five specimens of adult T. mashhoodi
were placed at different points on the reference
circle. The distribution of nematodes was recorded
after 4 h and was converted into scores and log scores
<Ah<Bad %c Jairajpuri, 1980; Bilgrami et al_. , 19S5a>.
E>;F«er i ment 5 were rep] icated twenty times. Attraction
of T. mashhoodi was tested towards 4—6 day old
cabbage, carrot, raddish and onion seedlings.
Attraction of T. mashhoodi towards different
concent ratalons of cabbage root homooenates; To
observe the attraction of T. mashhoodi towards
different concentrations of cabbage root homogenates
Petri-dish experiments as described above were
performed. All conditions remained same except that
0.5 ml root homogenate was placed in a cavity made in
the inner circle. Twenty g of 4-6 day old cabbage
seedlings were surface sterilized and homogenized in
20 ml sterile distilled v;ater in a warring blender to
obtain stoclc <100X> root homogenate solution. This
was then filtered to remove the solid materials. From

4 3 6
the stoclj solution 2D, 40, 60 and SO percent dilutions
were made- Attroctiofi of T. mashhoodi was tested
separately towards each concentration. Observations
on the distribution of nematodes were made after A h.
Art ract i on of T. mashhoodi towards combinati ons of
di fferent host root s; To observe the attraction of T,
mashhoodi towards different conibi nat i ons of host roots
7 cm diameter Petri-dishes were marked at the bottom
into 7 zones of 1 cm width each. The zones were
numbered serially as 1, 2^ 3, 4, 5, 6 and 7. An
e>:cised root of one host was placed in zone 1 and that
of the other in zone 7 and left for 12 h for
incubation. Fifty adult nematodes were introduced in
zone 4. The distribution of nematodes was recorded
after 4 h. The following combinations of host roots
were usedi cabbage and carrot; cabbage and radish;
cabbagt? and onion; carrot and radish; carrot and onion
and radish and onion.
RESULTS
Nejmatodes exhibited random movement upon
introduction into the agar within a period of 4—6 h.
Many were found in the vicinity of the roots which
started feeding thereafter. The following feeding
phases were observed in T . n> .-> s h h o o d i .

4 S 7
Hi de spread e>; pi or at i on ; As soon as the nematode made
contact with the root wide spread exploration was
iiiitiotetJ. Nematudeby uioved along the root, made
contact and probed roots with the stylet. The
locomotion o-f nematode stop'ped end nematode made
contacts with the root both at right angle and with
the sides of the head. A right angle contact between
the lips and the root resulted in quick stylet
thrusting. Nenjatodes moved away to locate a suitable
and favourable feeding site if this was not found
suitable. Wide spread exploration which led to contact
with the roots terminated with the selection of
favourable feeding site-
Locel e>! pi or at i on r Local exploration began as soon as
nematodes located a favourable feeding site. Stylet
thrusting started when the lips were at right angle
to the surface of the cell. When the surface of the
roots was rigid the stylet thrusts often resulted in
the lips to lift away from the root surface and the
nematodes started wide espread exploration again.
S t y1e t penetrati on ? Nematodes remained motionless
during penetration of stylet into the cell. At the
onset the stylet thrusting was slow but gradually
increased thereafter. During this period of rapid

4:8
stylet thrusting about hal-f of the stylet was
penetrated. The '-ell wanll v#a5 then punctured and
fi.ule of the stylet was penetrated deep into the eel 1-
Feedi nq; Feeding included ingestion of the cell
contents. Ingestion was intermittent i.e,, ingestion
of ceil contents at short intervals. During feeding
oesDfihageal pulsations started which occurred
irregularly in the begining but becarae regular later.
During this phase oesop'hageal secretions are injected
into the cell for the e>:tra corporeal digestion.
After feeding some nematodes reverted to wide spread
e>;ploratiDn and fed again on the roots while others
moved away.
Retr act i on of s t v1e t and departure f rom feedi nq site;
The oesophageal pulsations became irregular and
ultimately ceased when the feeding was over- Stylet
was retracted and lips were detached from the cell
surface. While moving away nematodes made few stylet
thrusts in the e>:ploratory fashion. After witbdrawl
of stylet the nematodes moved back and forth few times
and left the feeding site.
Attract? of> nf T . mashhonii J t o w a r f I B si nnl g host root
(Fig- 1 A> : T, mashhond i ahlracted poBJtively towards
the roots of all hhf=' plants vir., cabbage, CBrrai^

4 39
radish and onion. The degree of attraction vari ed,
May, i mum attraction occurred towards the roots of
cabbage <p < 0.05>. The roots of onion elicited least
response of T. washhoodi <p < D.D5). Carrot and
radish seedlings attracted T. fflashhoodi moderately-
A11 r a r t i o n of T- mashhoodi towards different
concentr at i ons_ of c a b b a c; e root homogenates (Fig. 1,
3>; Different concentrations of cabbage root
homogenates affected the degree of attraction.
Attraction increased with the increase in the
homogenate concentration. Majiimuro attraction occurred
towards SO percent <p < 0.05) concentration, T.
mashhoodi responded least towards 20 percent
homogenate concentration <p < 0-05>.
At tr sct i on of T. mashhoodi towards combi nat i on of
di f fer en t host root s <Fig. 2>: T. mashhoodi responded
roaximura towards cabbage seedlings in combinations with
onion (Fig. 2, C> and radish (Fig. 2, D) but the
degree of attraction remained at par with carrot (Fig.
2, B>. The seedlings of carrot attracted more T.
mashhoodi when tested in combination with radish (Fig.
2, E> or onion (Fig. 2, F>. Similarly, radish roots
in combination with onion were more attractive to T.
mashhooJ ? (Fig. 2, G>-

4 4 0
DISCUSSION
Host o-F t;<e ^hctses. described in T. mashhoodi ha-^e
also been reported in other plant parasitic nematodes
i,e., Tri chodorus si mi 1i s <WyB5, 1 9 7 1 ) ;
Tylenchorhynchus dubi us ^Wyss, 1 9 7 3 ) ; Nacobbus
batati formes and Heteroder a cruc i f er ae (Done aster ?<
Seymour, 1973) and Hi rschmanni el 1 a o r y z a e (Bilgrarai
et a j . ^ 1985 c ) . The feeding process of Telotylenchus
1 oof 1 as described by Tikyani « k'hera <1969> can be
considered as sn exception, because i t starts feeding
on roots imniediately after puncturing an epidermal
cell- Feeding behaviour of T. mashhoodi also differ
from that of Hex aty1 us vi vi parous in which passive
ingestion of the cell contents was reported by
Doncaster $< Seymour (1974),
Most tylencbids while locating the feeding site
do not show preference for any definite region along
the grov^ing root. However,, T. mashhoodi aggregated
around the root tip and the zone of elongation similar
t o Tyl ertchor hynchus max i mw^* and to T , 1 amel 1 i ferous
^Bridge St Hague, 1 9 7 4 ) ; T. cl ay ton i (Sutherland Sc
Adams, 19^4) and Hirschmanniel1 a o r y ? a e <Bilgrami et
aj_- 5 1 9 S 5 c ) . The exploratory behaviour of T.
mashhoodi comprised of searching movements of the

441
head, lip rubbing and stylet thrusting were more or
less similar to T. d u b i»s (Wyes, 1973) and N.
bat at i -f or^rtes and H. cruc i -P&r £& (Done ester & SeymDLT ,
1973)-
Present observationB on T. mashhoodi showed that
though these nematodes were not host speci-fic but they
showed some degree o-f pre-ferrence towards different
hosts. The occurrence o-f T, mashhoodi in lower ranks
in the presensce of onion and radish seedlings and in
higher ranks in cabbage and carrot seedlings leads to
indicate some degree of host preferrence behaviour,
Khera ?« Zuckerman (19^3) while studying the feeding of
plant parasitic nematodes found that all nematode
species did not respond equally to different hosts-
TV^ mashhoodi also showed similar behaviour with
cabbage end carrot seedlings attracting ma>;ifflum number
of T- mashhoodi and onion the least. Khera &
Zuckerman <19i.3> had also found cabbage to be a good
or an average host and radish as an average or non
host for most species of plant parasitic nematodes
that they had studied. The differences in the
formation of optimum attraction gradient by different
host roots could be the reason for variability in the
degree of attraction. It may be possible that cabbage
and carrot produced stronger stimuli (attractants^

4 42
while onion the weaker stimuli thereby eliciting
different reBponses-
Pbser vat 3 Djf;- on the attraction o-f T, mashboodi
towards various concentrations of cabbage root
homogerfate suggest that neipatodes responded more
quickly and fiiDre vigorously as the intensity o-f
stirauli increased as was also suggested by Doncaster
?< Seymour <i973). Tlve other factors which could be
attributied to the differences in the attraction
behaviour towards different ho/nogenate concentrations
are thf» minimum stimulus thrashold of the nematodes
and the forjrtation of optimum attraction gradient- It
seems possible that lower homogenate concentrations
took more time to develope an optimum attraction
gradient than the higher concentrations and this may
have resulted in the former attracting fewer nematodes
than the later.

4 43
REFERENCES
Abarad, I, ?< M. S. Jairajpuri (198D>. Sex attraction
and copulation in Chi 1oplacus symetr i cus
(Nematoda ; Acr obel i nae) . Nematol oqi c B ., 26 :
139-148.
A2fa-.j M„ I. Sf M- S, Jairajpuri <1977a>. Attraction o^
pjl ant parasitic neroatodes to host roots-
Nt^natol oqica^ 23 : 119-121,
Barbatola, J. O. and J. Bridge <I9S0>, Feeding
b€?haviour and hi st of«ethol ogy of Hirschmanniel 1 a
or yz ae., H, i ismamur i and H. spi ni caudata on
rice, J. Nematol., 12 s 48-53.
Bilgrami, A. L., I. Ahmad & M. S. Jairajpuri (19S5a>-
Factors influencing attraction of adult
Hirschwanniel1 a ory^ ae towards cabbage
seedlings. Revue de Nematol . , S. s 67-75.
Bilgrami, A. L., I. Ahmad & M. S. Jairajpuri <1985b).
Responses of adi?lt Mi rschmanni el 1 a or ys ae
towards different plant roots. Revue Nematol., 8
Bilgrami, A. L., I. Ahmad ?< M. S. Jairajpuri <1995c>-
Attraction and feeding behaviour of adult
Hi r sch'nanni el 1 a or v7 ae . 1 nd i an J . Nematol . , 1 5 :
209-21A.

4 4 4
Bilgrami, A. L., M. B. Jairajpuri & I. Ahmad <1986>.
Effect of different plants, singly and in
combinationB on the attraction of sdult
Xi phi nema basi ri. Indi an J, Nematol , , 16 : 60-
Bridge, J. and W, G- M. Hague (1974). The feeding
behaviour of Tyl enchorhynchus and j^erl i ni us
species and their effect on growth of
prennial ryegrass, Nematoloqi ca^ 20 i 119-130-
Doncaster, C.C- Sc H.K. Seyraour <1973). Exploration and
selection of penetration site by tylenchids,
bLegatol oqi ca , 1 9 ; 137-145.
Doncaster, C. C. and M. K. Seymour (1974). Passive
ingestion in a plant nematode Hex aty1 us
Vj vj parous (Neoty1enchidae : Tylenchida).
Nejgatol oqi ca , 20 s 297-307-
Khera, S- and B. M. iKikerman (1963). l_n_ vi tro studies
of host parasite relationships of some plant
parasitic nematodes. Nematoloqica, 20 : 277-
282.
Krusberg, L- R. (1959). Investigations on t!ie life
cycle, r e?pr oduc t i on , feeding habits and host
range of T y 1 e n r h o r }i y n r h u '5 cl a y t o n i Stein er.
Nemj=>t «T1 Oft < c r> , 4 : 1H7-197,

Prot, J- C. <1980>. Migration of plant parasitic
nematodes towerds plant roots. Revue Nen«aJ;_al_. ,
3 : 305-318-
Prot, J. C. & J- M. Kornprobst (1983).'' Effects of
Azadiracta i ndi c a , Hannoa u n d u1 a t a and Hannoa
klai neana seed extracts on the ability of
Mg> 1 o i d o<3 V n e j a v a n i c a juveniles to penetrate
tomato roots- Revue N e m a t o l , 5 6 ^ : 330-332.
Prot, J- C. ^ B. D. '-Jan Gundy <1932a>- Effect of soil
texture and the clay coinponents on migration of
"eloi donyne i nco^ni t a second stage juveniles.
J. Nematol . . 13 s 213-217..
Prot, J. C. Sf S. D. Van Gundy <19Slb>. Influence of
photoperiod and temperature on migration of
Nel oi donvne j avBni r.a , J . Nematol . , . 1_3 : 2 1 7 -
220.
Sutherland, J. R. and R. E- Adams <1964>. Host range
of Tylenchorhynchus claytoni on some •forest
nursery seedlings. Phytopatholoqy < 54 749.
Tikyani, M. 0. and S, Khera (1969). Iji vi tro feeding
i f Tel ofyl en chits 1 no^ it on great millet (Sorghum
V M I nar e) roots. hJeistat nl oqi c.a ^ 1 5 i 291-293.

*i -'f b
WyB5, U. (1971). Der mechanisms der nahrungsaufnahrae
bei Tr i chodorus si mi 15. Nematologi ca, 17 : SOS-
SIS.
W y s s , U- (1973). Feeding o-f Tyl enchor hyrtchus d u fa 2 u £ .
NeFtatol oqi ca , 19 ; 125-136.
Wyss, U. <1981), Ecto-parasitic root neraatodes :
feeding behaviour and plant cell reBponses. In i
Zuherflian, B. H. and Rohde, H. A. (Eds) Plant
Parasi tic Nematodes Vol. Ill 5 Wew Vorli, Acade/nic
P r e s s , 325-351.
Wyss, y. and P. Grootaert *1977>. Feeding mechanism
of Labronema vulvapapillatum. Hed. Fac.
Landbouww. RiJJiuniv. Gent. 42 : 1521— 1527.

AA7
LEGENDS
Fig. 1. A - Attraction of Tylenchorhynchus mashhoodi towards single host root.
B — AttractioPf of Tyl pnchorhynchus mashhoodi towards different corscent r at i D O B of cabbage root ho/nDgenates.
Fig, 2 Attraction of Tylenchorhynchus mashhoodi towards combinations of different fiost mats.

4 48
(D
2.60!
2.20 O
o
a> -
1.00
A
ca cr rd Root
on CO
s
2.63
a> 2.20 o (J if)
cn o
1.80
§ 1.40 CD
1.00
20
&
AO 60 80 100
Root homogenafe concentrations (%)
/^,o^ i

20
12_
4
H if 9
1 2 3 4 5 6 7
20
12 J
4
ca ct
1 2 3 4 5 6 7
CD
•^ 20"
£ 12.
C
^ 4 o
o
ca D rd
20
12.
4
1 2 3 ^ 5 6 7
ct
on
1 2 3 4 5 6 7 Sectors
20
12
ca
on
1 2 3 4 5 5 7
20
12
4_
rd
=•
Ct
1 2 3 A 5 6 7
20
12_
4
G rd
on
1 2 3 4 5 6 7 Sectors

A •" '1 s ~j U
FACTORS INFLUENCING ATTRACTION OF TVLENCHORHYNCHUS HASHHOODI TOWARDS CA&SAGE SEEDLINGS
By
Anwar L- BILGRAMI
Section o-f Wematology, Department of Zoology, Aiigarh Muslim University, Aligarh-202002, India
SUMMARY
Attraction of Tylenchorhynchus mashhoodi was in-fluenced by all factors except the coloured lights and darkness. Maximum attraction was recorded towards 4—A day old cabbage roots with -fed nematodes as well as 2, 4 and b day un-fed nematodes- Responses of T. mashhood i declined as the age o-f roots and period of food deprivation of nematodes increased. Live roo^s, attracted nematodes but roots killed by heating lost attractiveness- The nematodes showed preference to the region of root tip and aggregated most around it-Maximum number of nematodes were attracted when roots were incubated for 10-12 h in 3—4 mm thick agar layer of 0-55i concentration at 20-25 "C- Attraction decreased with increase in the distance of inoculation of nematodes from the roots.
Key words: Tylenchorhynchus mashhoodi ^ attraction, cabbage, roots, behaviour.
There is ample evidence to suggest that plant
parasitic nematodes Ar& attracted towards roots (
Lownsberg & Viglierchio, 1961; Azroi %c Jairajpuri,
1977a; Bilgrami e ^ al_- » 1985a; i9S5b; 1985c). Few
studies were made on the effects of various biotic and
abiotic factors on the attraction nematodes towards
roots- Weiser <1955) and Lavallee ?< Rohde <1962)
observed attractiveness of roots proportional to its
growth. Linford (1939); Bird (1962) and Bilgrami e^

si . <19S5a) reported I O B B in a tt r at i veness o-f roots i^
killed by heating. Lowensberg 9< Viglierchio (1961)
suggested that roots become unattractive when their
growth is ceased, Moussa e^ al. <1972> and Bilgrami
et B1 . <19S5a) did not observe any signi-ficant
difference in the attraction when coloured lights were
used. Davies & Fisher <1976) found temperature,
initial inoculum, density, duration of exposure and
distance of nematode inoculation from the roots to be
the limiting factors in the invasion of barley
seedlings by Heterodera avenae juveniles, Prot and
Vangundy (i991a; 19Slb) concluded that soil texture,
clay component, photoperiod and temperature influenced
migration of Meloi dogyne i ncoqni ta and M. hapla
towards roots- Bilgrami ej^ al . <1985a) observed that
factors viz., age of roots, food deprivation of
nematodes, incubation period of roots and distance of
inoculation of nematodes from the roots governed
attraction of Hirschmanniella oryzae towards cabbage
seedlings. The attraction of H. oryz ae and Xi phi nema
basi r i depended upon the type of host (Bilgrami et
al., 1985b| 1985c>.
In the present work effects of various factors
were observed on the attraction of Tylenchorhynchus
mashhoodi towards cabbage seedlings (Brassi ca oler acea
L. vsr capi tat a) in agar plates-

An
MATERIALS AMD METHODS
The soil samples containing Tylenchorhynchus
mashhoodi , collected -from cabbage -fields near village
Jamalpur, Aligarh, were processed by decantation and
sieving and modi-fied Baermann's funnel techniques.
All experiments were carried out in 5.5 cm
dianieter Petr i —di shes, divided into inner, middle and
outer zones by drawing two concentric circles of 0.5
and 2,5 cm in diameter at the bottom- The excised
root tip (0-1 cm) o-f 4-6 day old cabbage seedling was
placed in the inner circle of the Petri—dish
containing 0-5% water agar and intiuhated for 12 h. •
Five adult T. mashhoodi were released at the periphery
of the middle circle- The distribution of nematodes
was recored a-fter 4 h in each zone and converted into
scores and log scores (Ahmad %€ Jairajpuri, 1980}
Bilgrami ?< Jairajpuri, 19S8>. All experiments were
carried out at 25 ± 2"^ and replicated 30 times.
Petri-dishes without roots served as control.
Conditions remained same for all experiments unless
mentioned otherwise. In the text MLS stands for Mean
Log Score.
Effect of age of roots on the attracti on? Six age
groups of cabbage roots viz., 2, 4, 6, B, 10 and 12

/ r o 't ::• J
days old were selected to determine their effects on
the attraction of T. mashhoodi-
Effect of age of roots on the attracti on of starvi nq
nematodest The attraction of 2, 4, 6, 8, 10 and 12
day starved (kept in water without roots, the water
was changed every day to restrict growth of any micro—
organism) T. mashhoodi was tested towards 2, 4, 6, S,
ID and 12 day old cabbage seedlings. Each group of
starving nematodes was tested separately towards each
age group of cabbage seedlings.
Effect of 1i ve and dead roots on the attracti on; The
influence of live and dead cabbage roots on the
attraction of T. mashhoodi was observed by incubating
them in the inner circle of Petri-dishe perior to the
introduction of nematodes. Cabbage roots were killed
by boiling in water for 5 min-
Effget of length of i ncubati on peri oda of root» on the
attracti on: The roots were incubated for 2, 4, 6, 8,
10 and 12 h separately prior to nematode inoculation
to observe the influence of different periods of
incubation of roots on the cittraction of nematodes.
Ef f ect of r eqi ons of roots on the at'cracti on; Two
sets of experiments were performed to observe the

4S4
effects of different regions of root on the attraction
of T. mashhoodi, In the first experiment attraction
was tested towards intact non—e>!ci5ed roots- Water
BQar (0-5%> poured on a glass slide was allowed to
cool. The root was placed in the agar parallel to the
length of the slide and divided into three 1 cm
regions by marking lines at the bottom of the slide
(Fig, 1>. The shoot remained outside the agar. After
incubating roots for 12 h, 25 adult T. mashhoodi were
introduced at the edges of the slide opposite to the
middle region of the root. Distribution of nematodes
was observed after 4 b-
In the second experiment the Petri—dish technique
was used- A 3 cm long excised root was cut into three
equal parts viz., root tip <D-1 c m ) ; zone of
elongation (1-2 cm) and zone of maturation (2—3 c m ) .
Ef feet of aqar thi cknesses on the attracti on; T.
mashhoodi was placed with the roots in Petri—dishes
containing 1, 2, 3, 4, 5 and 6 mm thick agar layers to
determine the effect of different agar thicknesses on
the attraction.
Effect of aqar concentrati ons on the attracti on To
observe the influence of different concentrations of
water agar, thf? roots were incubated in separate

45^
Petri-dishes containing 0.,5, 1, 2, 3, 4, 5, and 6/t
water agar prior to nematode incoculation.
E-f-f ect of 3 i qht 5 and darkness on the attract! on;
Roots were incubated in Petri-dishes under dif-ferent
light conditions to determine the e-f + ect of light and
darkness on the attraction of T. meshhoodi. The
nematodes were then introduced and the dishes were
replaced under similar lights. Light -from 60 watt
lamp kept at a distance o-f 65 cm from the Petri—dish
was considered as normal light. Coloured ef-fects were
obtained by covering the lamp with red, yellow, green
and blue cellophane papers.
E-f-feet o-P temperature on thg attracti on; To find out
the effect of temperature on the attraction of T.
mashhoodi the roots were incubated in agar at 5, 10,
15, 20, 25, 30, 35 and 40 "C. The nematodes were then
introduced and Petri-dishes kept again at the same
temperature at which they were stored earlier.
E-f feet of diet ance of root s on the attracti on t The
effect of distance on the attraction of T. mashhoodi
was observed in 7 cm diameter Petri-dishes. The
Petri-dishes were marked at bottom into seven equal
zones similar to Bilgrami et al. (19a5a>. A 1 cm long

4:^0
part of root -from cabbage was placed in zone 1 and
incubated -for 12 h. A-fter incubation 25 T, mashhoodi
were introduced in zone 2, Similarly the nematodes
were introduced in rone 3, 4, 5, 6 and 7 of separate
Petri-disbeB and their distribution was recorded after
4 h.
RESULTS
The attraction of T. mashhoodi was influenced by
all factors except coloured lights and darkness-
Effect of age of roots on the attracti on (Fig. 2>.
Age of seedlings affected attraction- Maxiroum
attraction was recorded towards 4 and h day old
cabbage seedlings. Attraction declined towards
higher age groups of roots 0—12 days). Minimum
attraction was recorded with 12 day old roots <p <
0.05).
Effect of age of roots on the attracti on of st arvi nq
nematodes (Fig. 3) The 4 — 6 day old seedlings
attracted starved nematodes most (p < D.05> (Fig. 3,
A ) . The B - 10 and 12 day old seedlings attracted
less T. mashhoodi (p < 0.05). Minimum attraction of 2
day starved nematodes was r€?corded towards 12 day old
cabbage seedlings (p < 0.05). The 8 day starved

^rj/
nematodes were attracted most towards 2 and 4 day old
cabbage Beedlings (Fig, 3 D>. T. mashhoodi < deprived
of -food -for 10 days, showeo positive response towards
2, 4, 6, 9 snd 10 day old cabbage seedlings <Fig. 3,
E>. f4ematode5 did not respcjnd to 12 day old roots <p
> 0,D5>, T w e l v e d a y s s^tarved nematodes s h o w e d ma^iimum
attraction towards 2 day old roots (Fig. 3, F>.
Attraction declined towards roots of higher age. The
responses of 3 0-12 day starved T, mashhoodi remained
at par with the control (P > 0.05) when tested towards
10 and 12 day old cabbage s€»edlings-
E-f f ec t o-f 1 i ve and dead roots (Fig. 4, A>J Live roots
attracted T. reashhoodi (p<' D.05) while dead roots
•failed to attract then) (p > 0 . 0 5 ) .
Effect of 1enqth of i ncubat i on peri od of root 5 on the
attraction (Fig. 4, D>: Two and four hours
incubation did not attract nematodes (p > 0 . 0 5 ) . The
attraction became positive only after A h (p < 0 . 0 5 ) .
It increased further significantly upto 12 h
incubation (p < 0 . 0 5 ) .
Ef fect of reqi ons of root on the attracti on (Fig. 4, B
?t C>s When excised cabbage roots were used in P e t r i -
dish evperi/nent i?}a>rimiim attraction (P < 0.05) was

4 5 8
observed towards 0-1 cm regions of cabbage roots <Fig.
4, B>. T, raashhoodi responded least towards 1-2 cm (p
< 0-05). SI ass slide experiment also showed maximum
attraction towards 0-1 cm. Nematodes failed to
responsde towards 2-3 cm region of roots (Fig- 4, C ) .
Effect of agar thicknesses on the attracti on (Fig. 5,
B) I i^siy.imi.tm attraction was recorded in 3 and 4 mm
thick agar layers (p < 0.D5), Agar thicknesses less
than and more than these declined attraction <p < 05).
Cabbage roots elicited least response of nematodes in
6 mm thick agar layer <p < 0.05).
Ef feet of agar concentrati on on the attracti on (Fig.
5, C ) . Attraction was greatly influenced by all
concentrations of water agar, except 0.5 and IX
Attraction declined in 2 and 3X (p < 0.05) and there
was no attraction in 4, 5 or is'/, agar dishes (p >
0.05).
Ef feet of temperature on the attract i on (Fig- 5, A ) :
Temperatures less than 20 "C and more than 30 "C
declined attraction markedly- Ma>! i muro attraction was
recorded at temperatures 20-30 "C. No attraction uas
recorded al 5-10 °C (p < 0.C5)- Cabbage roots
elicited least response of nefljatodes at 40 "C.

4 59
E-f f ect of I i qhts and darkng?SB on the attraction (Fig.
5, D ) : Different lights and darkness had no effect on
the attraction behaviour of T. mashhoodi.
Effect of di stanace of roots on the attract!on (Fig.
6 ) ; As the distance of the inoculation of neroatodes
from the roots decreased the number of nematodes
reaching rone 1 increased. Maximum numbers were
recorded in sone 1 when the nematodes were introduced
in zones 2 an6 3 i.e., 1 cm and 2 cm from roots- T.
mashhoodi also showed positive response when placed 3
cm and 4 cm away from the roots i-e., zone 4 and 5 but
not when inoculated in zones 6 and 7 i.e., 5 csnd 6 cm
from the roots-
OISCUSSION
Viglierchio (1961) observed attraction
independent of the age of roots. Weiser <1955> and
Lavallee t< Rohde (1962) reported attractiveness of
roots proportional to their growth. The present
observations are contrary to both- Observations on T.
mashhoodi indicated that as the cabbage roots grew
older, their attractiveness decreased and failed to
elicite response from fed nematodes, suggesting that
they were no longer attractive. This differential

4cja
behaviour could be due to di-f-ferent hosts and nematode
species tested and also as all nematodes are not
equally sensitive to the same stimulus or all plants
do not produce similar attractants (Prot, 1980). The
inhibition of growth o-f roots in Petri—dish containing
only water agar may have also in-fleunced attraction of
T. mashhoodi . As the period o-f food deprivation of
nematodes increased their response towards increasing
age groups of roots decreased. This suggest that
besides the unattractiveness of roots some other
factor<s> within the nematode was (were) also
governing their response <Bilgrami et al_- 1985a>.
Doncaster ?< Seymour <1973} concluded that in starved
nematodes minimum response thrashold decreased and
unfed nematodes could perceive weeker stimuli. These
observations show that in T. mashhoodi, perhaps
deprivation of food caused sn increase in minimum
response thrashold as 12 day unfed nematodes failed to
respond to some of those age groups of roots which
attracted lesser unfed nematodes. Perhaps a loss in
the degree of attraction with an increase in the
period of food deprivation have resulted from loss of
energy, as starving nematodes utilize their reserve
food material (Cooper ?< Van Gundy, 1970; Riversat,
1981a; 1981b>. Hence, both factors, age of roots and

4 J
period of -food deprivation appear to in-fluence
attraction o-f T _ roashhoodi towards cabbage seedlings
independently.
Houssa et. aJL- (1972) did not find any difference
in attraction of juveniles of Globodera rostochi ensi s
under different coloured lights and darkness. The
attraction of T. mashhoodi also remained unaffected
under different coloured lights and darkness.
Morf^over roots killed by heating lost their
attractiveness <Linford, 1939? Bird, 1962; Bilgrami et
al . 1985a> which confirms present results on the
responses of adul t T. mashhoodi towards 1i ve and dead
cabbage roots.
Pitcher St McNamara <1970); Schiltz 8c Cohn < 1975:) 5
Stirling (1976) and Bilgrami et. al . U9S5a) reported
root tips as roost attractive. During present
observations T. mashhood i also exhibited maximum
response towards root tips <both excised and intact).
The excised roots may introduce additional unknown
substances into the medium due to cell breakdown but
these substances did not seem to have any effect on
the attraction behaviour of T. mashhoodi.
Bilgrami et al. (1985a) suggested a minimum
period necfe' isary for the development of perceptible

4 •; '5
attraction gradient by the roots. During present
observations with T. mashhoodi the influence of
different incubation f>eriods of roots on the
attraction suggest the similar phenomenon.
The effect of agar concentrations and te/nperatures
on the at tract ion «Bay be related to the activity of
nematodes. Wallace (1969}, Azmi « Jairajpuri (1977b);
Bilgrami e ^ aj_. (19335 1995a> suggested that activity
of nematodes declined in increasing agar
concentrations. Perhaps this reduced activity
inhibited attraction of T, mashhoodi in more than 2%
water-agar. Wallace (1963) showed that nematodes
became inactive at temperatures 5—10 and 3D-4D"C.
Prot Sc Van Gundy, i981b> reported inhibited activity
of Heloi doqyne i ncoqni ta and M. hapla at low
temperatures. Biigrami et al_. (19S5a) did not find
any response of Hi rschmanni el 1 a oryz ae at temperatures
5 — 10 °C- No attraction of T. mashhoodi at low
temperature and differential attraction behaviour at
others may be attributed to its activity. Besides,
the diffusion of root attractants in the agar have
also been influenced by different temperatures and
agar thicknesses resulting in different responses of
nematodes.

A 63
Globodera rostochi ensi s juveniles migrated upto
a distance of 45 cm in &Di1 <RDhde, 1962); Anqui na
t r i t i c i juveniles travelled 30 cm upwards to reach
their host (Leukel, 1962); t^el oi doqyne javani ca showed
50 cm horizontal and vertical migration in 9 days
towards a susceptible tomato variety (Prot, 1977) and
M, incognita migrated upto 20 cm in soil to reach
tomato roots (Prot Sf Vangundy, 19gia). However, H.
orv2 ae showed a comparatively weak response as it
could reach its host onl / -from a distance of A cm
(Bilgrami et aJ,, 1985a) which is similar to
Di t vl enrhi'B d^ psaci <Bla!<e, 1962), During present
observations T, mashhoodi responded towards the
cabbage roots similar to M. oryz ae and D. di psaci.

4 o 4
REFERENCES
AHnAD, I. ^ JAIRAJPURI, M- S- <198D)- Sex attraction
and copulation in Chi 1oplacus symetri C U B
(Nematoda : Acr ob€fl i nae) . Neroatol oqi ca 2^,
139-148.
AZNI, M. I. S< JAIRAJPURI, ». S. (1977a). Attraction
of plant parasitic nematodes to host roots,
^4ema t o 1 oc; i ca 23 ^ 119 — 121.
A Z M I , M. I. Sc JAIRAJPURI, M. S. (1977b>. Studies on
nematode behaviour III: Locomotion in juveniles
of Hop I ol ai mus i n d i c u s snii Hel i cotyl enchus
i ndi cus in dif-ferent concentrations o-f agar.
Indi an Jounal o-f Mematol oqy 5., 24D—242.
BILGRAMI, A. L, <1983). Attraction of
Hirschraanniel 1 a oryzae towards cabbage
seedlings. Pr oceedi nqs o-f Thi rd Nematol oqy
Symposi urn., Sol an , Indi a (Abstr) , 15.
BILGRAMI, A. L., AHMAD, I. 8f JAIRAJPURI, M. S.
(1985a). Factors influencing attraction of adult
Hirschmanniella oryz ae towards cabbage
seedlings. Revue de Neroatol oqi e 8 , 67-75-
BILGRAMI, A. L., AHMAD, I- & JAIRAJPURI, M, B.
(1985b>. Responses of adult Hirschmanniel1 a

4 u 5
or vz ae towards di-f-ferent plant roots. Revue de
Ngmatoloqi e ^, 265-272.
BILGRAMI, A. L,, AHr^sAD, I. ?< JAIRAJPURI, M- S.
<1985c>. Attraction end feeding behaviour o-f
adult Hi rschmanni el 1 P O r y Z a e , I n d i an Journal o-f
Nem-itol oov 15, 209-216-
BILGRAMI, A. L., AHMAD, I. ^ JAIRAJPURI, M. S.
<19aij>. Response o-F Hirschmanniel 1 a oryx ae
juveniles towards cabbage seedlings.
Helmintholoqi a 23., 119-122-
BILGRAHJ, A. L., JAIRAJPURI, M. B. Sf AHMAD, I.
n 9 B 6 > - Ef-fect o-f di-f-ferent plants, singly and
in co/nbi nat i ons on the attraction o-f adult
Xi phi nema basi ri . Indi an Journal o-f Nematol oqy
\h, 60-64.
BILGRAMI, A. L. 8r JAIRAJPURI, M. S. <19e8>.
Attraction o-f Mononchoi des 1 onqi caudatus and
M . fort i dens (Neroatoda j Di pi ogaster i da)
towards prey and factors influencing attraction.
Revue de NematoJ oqi e Li_» 195-202.
BIRD, A. F. a 9 6 2 > , Drientotion of the larvae of
Mel ai dot; yne javani ca relative to roots.
Nomatol on' ca 7 , 275-287.

466
BLAKE, C. D. <1962>. Some observations on the
orientation of Pi tvlenchus di psaci and
invasion of oat seedlings- Nematoloqi ca 7, ,
177-192-
COOPER, A- F. ?< VAN GUNDY (1970). MetaboliBm of
glycogen and nffutral lipids by Aphel enchus
s-yj^nae and Caenor ha bdi t i s sp- , in aerobic,
ffli cr-o —aerobi c and anaerobic environmentB-
Journal o-P Nematol oqy 2_, 305—315.
DAVIES, K. A. & FISHER, J. «. (1976). Factors
in-fluencing the number o-f larvae o-f Heterodera
avenae invading barley seedlings i n vi tro.
Nematol oqi ca 22., 153-162.
DONCASTER, C. C- ?( SEYMOUR, M. K. <1973>.
Exploration end selction o-f penetration site by
tylenchids. Nematoloqi ca 19, 137-145.
LAVALLEE, W. H. ?c ROHDE, R. A. <1962).
Attractiveness of plant roots to Pratylenchus
penetrans (Cobb). Nematol oqica 8., 252-260-
LEUKEL, R, W. <1962>- Nematode disease o-f wheat and
rye. Farmer 's B»>1 1 . . 1607, 1-16.
LINFORD, M. B. (1939). Attractiveness o+ roots and
excised shoot tissue to certain nematodes.

4 u 7
Proceedi ngs o-f Helminthol oqi cal Soci ety
Washi nqton 6 , IJ-IS.
LDWWSBERY, B. F. «c VIGLIERCHIO, D. R. (19ol>-
Importance of response of Meloi doqyne hapla to
an agent from germinating tomato seeds.
PhvtopatholoQv 51. 219-221.
LUC, H. , LESPINAT, P. ?< SOUCHAUD, B. (1969).
Marquage direct de Hemi cycl i op h or a paradoy.a par
le phospbore radioacti-f. Utilisation pour
1'etude des deplacements des nematodes
phytoperesites dans le sol. NematolOQCS 15, 35—
43.
MOUSSA, F. F., De BRISSE, A. T. & GILLARD, A. 0 972).
The effect of light on the attractiveness of
tomato seedlings to larvae of the golden cyst
nematode, Globodera rostochi ensi s. Mededli nqen
Paculteit Landbouwweten5chappen Ri jksuni ver-
siteit Gent 37., 315-321-
PITCHER, R. S. & McNAMARA, D. C. 0 9 7 0 ) . The effect
of nutrition and season of year on the
reproduction of Tri chodorus viruliferus.
Nematoloqica 1 A, 99-106.
PROT, J. C. (1977). Amplitude et cinetique des
migrations du nematode Melni doqyne javani ca sous

4-.Q
1 ' in-fluence dun plant de tomato- Cah. ORSTOIJ,
ser. Biol 1 I , 157-166-
PRDT, J. C- <19S0>. Migration o-f plant para:=itic
nematodes towards plant roots- Revue de
Nematoloqi e 3_, 305-318.
PRDT, J- C. ?. KORNPROBST, J, M. (1983). E-ffects of
A~ sd j r act i r,d i c a , Har«rfOa »ndu 1 at a and Hannoa
I; 1 a i neao'^ seed extracts on the ability o-f
Heloi dogyne javani ca juveniles to penetrate
tomato roots- Revue de Mematol oqi e 6., 330-332-
PRDT, J. C. ^ NETSCHER, C. <]1978>. Improved
detection o* -flow population densities of
Mel oi doqyni?- Nematol oqi ca 24 , 129 — 132-
PROT, J. C- & VAN GUNDY, S- D- (19aia>. E^^^^ect of
soil texture and the clay components on
migration o-f Mel oi doqyr>e i ncoqni t a second stage
juveniles. Journal o-f Nematol oqy 13, 213-217,
PROT, J. C. Sc VAN GUNDY, S. D. <19aib>- Influence o-f
photoperiod and temperature on migration o-f
Meloi doqyne javani ca. Journal of Nematoloqy
13, 217-220.
REVERSAT, G. (1981a>. Effects of ageing and
starvation on respiration and food conents in

4o9
adult Hi rschmannielI a spa nicaudata. Rgvue de
Nematol oqie 4.* 125-130-
REVER3AT, G. <1981b). Consuroption o-f food re&erves
by starved second stage juveniles of Meloidoqyne
javani cs under conditions inducing osmobiosis-
NemRtolpqica 27, 207-214.
ROHDE, H. U 9 6 2 ) . Untersuchungen uber das
Wanderver'nogen von Larven des Kart o-f f 1 neinat oden
<G1obodera rostochi ensi 5> in Model 1 Versucben
mi t ver schi edenen Bodenerten,. Nematol ogi ca 7 ,
74-82.
SCHILT, H. G. ?« COHN, E. <1975>. Pathogenicity and
population increase of Paratr i chodorus mi nor as
influenced by some environmental factors.
Neroatol oqi ca 2J_? 71-80.
STERLING, G- R. (1976). Paratrichodorus lobatus
asDciated with citrus, peach and apricot trees
in South Australia. Nematol oqi ca 22., 138-144.
VIGLIERCHID, D. R. <19A1>. Attraction of parasitic
nematodes by plant root emanations.
Phytopathology 51, 13A-142.
WALLACE, H. R. <1963>. The biology of plant
parasitic nematodes. Edward Arnold, London 280
pp.

470
WALLACE, H. R. <1969>. Wave formation of infective
larvae of the plant parasitic nematodes
Hsloi doqyne javani ca. Neroatoloqi ca 15, 65-75-
WEISER, W. (1955). The attractiveness of plants to
larvae of rootknot nematode I- The effect of
tomato seedlings and excised roots on
Hel oj. doQvae hapl a Chit wood, Proceedi no
Helmintho1oqi c a 1 Soc i etv Washi nqton 22 < 106-
1 12.
WYSS, U. <1981>. Ecto-parasitic root nematodes i
feeding bc^haviour and plant cell responsBB- In :
Zukerman, B. M. and Rohde, R- A. <Ed5> PI ant
Parasi tic Nematodes Vol. Ill, New York, Academic
Press, 325-351.

47 1 LEGENDS ^ ' '
Fig- 1 E>! per i mental design for testing the attraction of Tylenchorhynchus isashhoodi towards di-f-ferent regions of cabbage roots-0-1 cm = root tip; 1-2 ciB^zone of elongation; 2—3 cm = zone of maturation.
Fig, 2 Effect of different age groups of cabbage root on the attraction of Tylenchorhynchus meshhoodi.
Fig. 3 Effect of different age qroup^ of cabbage root on the attraction of unfed Tylenchorhynchus mashhoodi. A = 2 day unfed adult; B = 4 day unfed adult; C = 6 day unfed adult; D = 8 day unfed adult; E = 10 day unfed adult; F = 12 day unfed adult;
Fig. 4.
A. Effect of live and dead cabbage roots on the attraction of Tylenchorhynchus mashhoodi
B. Attraction of Tylenchorhynchus washhoodi towards different regions of excised cabbage roots (Petri-dish experiments); 0-1 cm = root tip; 1—2 cm = zone of elongation; 2-3 cm — Hone of maturation.
C. Attraction of Tylenchorhynchus mashhoodi towards regions of intact cabbage roots (Glass slide experiment); 0-1 cat »= root tip; 1—2 cm — zone of elongation; 2-3 cm = zone of maturati on.
D. Effect of length of incubation periods of cabbage roots on the attraction of Tylenchorhynchus mashhoodi.
Fig. 5.
A. Effect of temperatures on the attraction of Tylenchorhynchus mashhoodi towards cabbage roots.
B. Effect of agar thicknesses on the attraction of Tylenchorhynchus mashhoodi towards cabbage roots.
C. Effect of agar concentrations on the attraction of Tylenchorhynchus mashhoodi towards cabbage roots.
D- Effect of light snd darfiness on the attraction O'*" Tyl enchorhynchus mashhoodi towards cabbage roots. B = Blue; R = Red; D =• Dark; G = Green; Y = Yellowj N •= Normal; C => Control
Fig. h> Effect of distance* of cabbage roots on the a t t r a c t i cj 1) of Tyl enchor hyrtchus /n a s h h o o d i . Arrows indicate point of inoculation of nematodeB,

47 2
Cover-lip. c
Glass slide
Agar
0-1 1-2 2-3 cm cm cm
Host root
Point of inoculation Of nematodes
f/& '^

473
m o u (/)
C7)
3 c a 2
2.5C
2.2Q
isa
160
130,
inn
X——'\ X—X With roots •—©Without roots
9
X
® Q
8 10 12 Age of roots (days)
/O -Z-

o O !
2.00! en o
"1
CU
120 -« o-1 r
• o
B
© • — «
1 r 2 A 6 8 10 12 i A 6 8 10 12
/ 7 '1
A 6 8 10 12
Age of roots (days)
D
e
\ U 6 8 10 12
x—x With roots p
©-^Without roots
X.
- 9
X- X -o
—^ 1 1 , , 1
2 A 6 8 10 12
Age of roots (days)

475
2-60
2.20 o I/)
c 2uo
l.OOi
A Live roots
^ W i t h o u t roots
Dead roots
Roots
2.60
2.2QJ
1.80
1.00
B
z ^
^ w thout root 0-1 cm l - 2 c m 2-3cnn
Regions of root
12
o 9_ o o if)
^ 6
c a CD 3
:^
cTT iWithout
Regions of root
roots 0-lcm260 l-2cnn 2-3cnn
ZAO
1.80
l.AO
1.00
X—xWith roots n .—aWithoutroot
V — - x -• —— • — •
1 r—\ i r 2 A 6 8 10
Incubation period of roots(h)
12
/Y&-^f
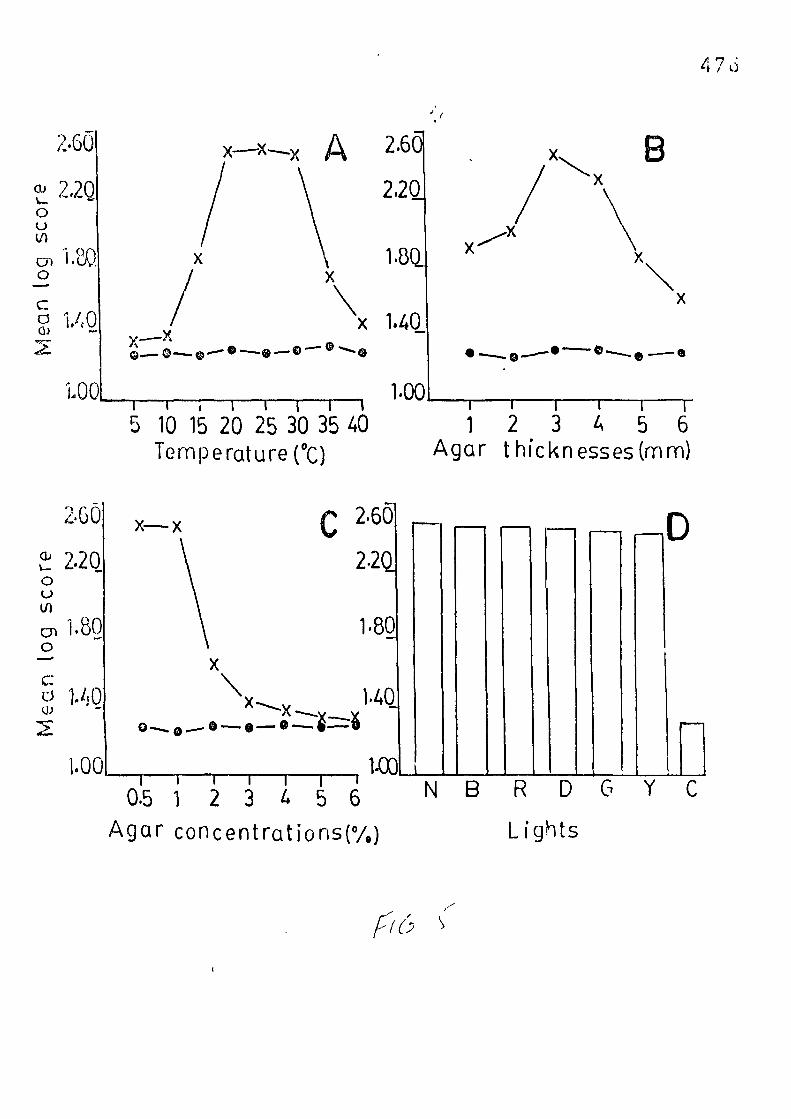
4 7 o
2.60|
2.2Q v_ O O I/)
o
1 '' -a
1.8
LOO
Q © i
2.20
1.801
1.00 1 1 ; [ 1 1 1 1
5 10 15 20 25 30 35 AO Temperature ( t )
x
B X
X.
.©- .©. • o
i I I 1 I r
1 2 3 A 5 6 A g a r t h i c k n e s s e s (mm)
ZGO
2 2.20 o u
O 1.80
1.00
X — X Q 2.60
2.2Q
1.80
A -~— y •
"-1 i 1 1 1 1 1
0.5 1 2 3 A 5 6
LAO
m
Agar concentrations^/,)
1
D
N B R D G Y C
L i g h t s
f^f6 <

18
U iz cu c
q 6
I
^
J I
1 2 3 A 5 6 7
18
12
i 1 2 3 4 5 6 7
477
m 18 a; o 12 "a
c k. o d z:
± J — I
1 2 3 4 5 6 7
18
1il
D
•
1 1
1 2 3 A 5 5 7
12 "O
o a <~ c c
o
E
t 1 •
2 3 4 5 6 7 Sectors
18
12
F
1 2 3 A 5 6 7 Sectors
n6-

4 7 8
ATTRACTION AND FDDO PRFFFRFNCE BFHAVTDDR OF PREDATORY NFMATODFS^ Al } OnORVt ATMUR AMFR TCA.-JliS AND DI5CQLAIHU5
ST? VTCOt US <WFMATODA:DRRVLAIMIDA)
By
Z. KHAN, ANWAR L. BILGRAMI ?. M, SHAHIM JAIRAJPURI
In'3tii-Mte o-f Agriculture, BbaiFi House, Oi 1 a Road, Aligarh M U B 3 i m University, AliQarh-202002, India
SUMMARY
Observations were made on the attraction and food pre-ference behaviour o-f two species o-f dorylairo predators vi?- , Al I odoryl ai wn^s amer i canus and Pi scnl a a <nus silvicnllus, A. amer i canus and D. si Ivico^ «-<s reached prey either by chance or wil^^ul movement (in response to prey secretions). The attraction o-f predators depended upon the type and physical state (live or e>!ci5ed> of pf^Y Bn<i the volume, chemical netiiure and composition o-f prey contents as well as the perceptible attraction gradient o-f the stimuli and the attraction threshold level o-f the predators. Attraction increased with the increase in the period o-f observation and varied with dif-ferent species o-f plant parasitic nematodes. F>;cised individuals attracted mnre^ predators than the live or intact, Hir'=;chmanniel 1 a or y ? ae and Xi ph i nema hasi r i elicited maximum while Mel icohylenchus i nd i cus the least response o-f the two species o-f predators. Other apecies o-f prey were moderately attractive, A. ameT i r ain^\'=, and D. si 1 vi col ».«s also responded positively to secretions of s««ch species of prey (Hopl ol ai mus i f> d i c < t s Hel i cnl y 1 enrV>us i ndi cus and Memi cycl i ophora dh i render i > which t^r^ resistant to predation. Predators failed to injure or kill any individual of these species hut fed when excised, A. amer i c anus and D. silvjcolus prefered the second stage juveniles of Mel oi donate j ncnnn i t a , Anqui nB t. r i t i c i and Net er oder a mot hi most in all combinations in which they were tested. Food preference is attributed to the physical , chemical and behavioural characteristics of prey that provide resistance to predation and the ability of tl»e predators to capture and kill the prey.
Key words; Al 1 odgry 1 ai m<is amer i canus. Pi scol ai mus sjlvirolus, prefJator, prey, predation, attraction, food f>ref erence.

4 7 9
Sever-aJ studies were made on predation by
new.^todR=i (Melmes, 1974; Cohn ic Mordecbai , 1974;
Bilgrenni et. el_. , 1904; 1 9 Q 5 ) ; resistance and
susceptibility of P»"ey nematodes to predation
(Grootaert et al_. , 1977; Bilgrami it Jairajpuri, 19S9a;
Bilnrami, 1992, 1 9 9 3 ) , gut contents <Bilgrami «f
Jairajpuri, 198o> Bnd factors influencing predation
(Bilgrami et aj_- , 1983; Bilgrami it Jairajpuri, 1989b;
1990; {•:han et. al_. , 1991; k'ul shresbtha e ^ al_. » 1 9 9 3 ) .
Studies arf, bnwever, lacking on the attraction and
food preference bebavi o«ir of predaceous nematodes in
general flrtd dorylaim predators in particular. Esser
(19A3> reported cbemoreception in dorylaim predators
but virtually no work has been done until Bilgrami ?<
Jairajpuri, (1988, 1989c; 1990) made detailed studies
on two species of dipiogasterid predators,
Hononcboi des 3 onqi rattdatus and M. f ort i dens. They
(I.e.) have reported attraction in these predators
towards prey and determined their preference for
prey. Information about the chemoattraction and food
preferences Art^ still larking on dorylaim predators
except those of Bilgrami et^ aj_ <1985); and Shafqat et
al. <1987> who reported the occurrence of attraction
in a nygnlaim and dorylaim predators only in the form
of aggregahion >=*y feeding sites.

4 8 0
In the present work the attraction and -food
preference behaviour o-F Al 1 odoryl ai mus amer i canus and
Di scolai mus si 1vi colos were studied using plant
parasitic nematodes as prey.
MATERIALS AND METHODS
Allodorylai mus aroeri canus and Di scolai mus
si 1vi col us were cultured in Petri-dishes containing
1% water agar using Rhabdi t i s sp as prey. A small
amount of in-fant milk powder <LactQ<3en) was spread
over the sur-face of agar to encourage bacteria to
grow. Bacteria served as the food for prey nematodes.
Attraction of predators towards live and excised preys
The attraction of A. ameri canus and D. si 1vi col us was
tested 5.5 cm diameter Petri-dishes which were marked
into inner, middle and outer zones by drawing two
concentric circles of 0.5 and 2.5cra in diameter at the
bottom <Bilgrami St Jairajpuri, 1988). The petri-
dishes were then filled with 5 mm thick layer of IX
water agar. A plastic straw pipe, 8 mm high and 5 mm
in diameter, sealed at one end with a piece of filter
paper was placed vertically at the centre of the inner
circle. The sealed end remained within the agar.

481
touching the bottom of the petri—dish. The straw pipe
was also -filled with 1/1 water agar. To test
attraction 50 live or excised <cut in two pieces)
individuals o-f Hi rschmanniel 1 a or yz ae or Mel oi doQvne
i ncoqni ta v#ere inoculated into the straw pipe and
incubated for 24 h. A^^ter 24 h -five individuals o-f A.
ameri canus or D. si 1vi col us were released at different
points of the periphery of the middle circle- The
distribution of nematodes was recorded after 6, 12, 13
and 24 h. Scores were then obtained by summing up
the products of the number of worms present in each
zone with their corresponding weighting factors- The
weighting factors were obtained by dividing the srea
of outer zone by that of each of the three zones.
Scores were then converted into log scores. All
experiments were carried out at 28 + 2'C and
replicated 20 times. The attraction of A- ameri canus
and D. si 1vi col us towards live or excised individuals
of H. oryz ae and M. i ncoqni ta was tested separately.
Attraction of predators towards different species of
preys To test the attraction of A. ameri canus and D-
silvicolus towards different types of prey, similar
experiments were carried out in Petri dishes as
described above. All conditions remained same except

4 82
that observations on nematode distribution were made
after 24 h. The attraction of the two species of
predators was tested towards the live or excised
individuals of HopIolai mus i ndi cus Helicotylenchus
i ndi CUB H. oryz ae. H. dhi renderi , X. basi ri and atbe
second stage juveniles of M. i ncogni ta, H. mothi and
A. tri t i ci separately-
Food preference! Species of p.lant parasitic nematodes
(Table—I) were collected fresh from respective hosts
for each experiment- The preference for prey by A.
am&ri canus and D. silvicolus was determined in small
cavity blocks containing 1% water agar at 28 + 2"C. To
determine food preference the plant parasitic
nematodes were subjected to predation by A. ameri canus
and D- silvicolus in all possible paired combinations
(i.e., two species of prey at a time). Each prey
combination thus, contain two species of plant
parasitic nematodes. Twentyfive individuals of each
prey were inoculated in a combination, thus, making
each of 50 individuals. Each combination of prey was
subjected to predation by 5 individuals of A.
amer i canus or D. silvicolus in cavity blocks
containing agar. The number of prey (belonging to
each species) killed or injured by the predators was
recorded after 24 h.

/' 0 1
RESULTS
Attraction of predators towards live and excised prey
(Fig. 1 ) :
H. oryz ae attracted more predators than M.
i nconni ta whether tested intact (live) or excised cut
in two pieces). The excised prey individuals
attracted CTore predators than the live.
There was no attraction of A. ameri canus and D.
silvicolus towards live H. oryz ae and M, i ncoqni ta
after h or 12 h. Attraction became significant after
12 h which increased upto 24 h (p < O.05)(Fig. 1 A,
C) - Similarly, when A. smericanus and D, silvicolus
were tested towards excised prey individuals the
attraction was evident only after 6 h which increased
thereafter (p < 0-05). At h h the degree of attraction
remained same as in control (Fig. 1 B, D ) .
Attraction of predators towards different species of
prey (Fig. 2 ) :
Both the predators showed differential degree of
responses towards different prey. They also exhibited
differential attraction behaviour when tested towards
live (fig- 2 A) and excised (Fig- 2 B> individuals of
prey. Thr? attraction was roore towards excised

4 84
individuals, of prey. H, or yz ae and Xi phi nema basi r i
attracted maximum number of A. ameri canus and D.
silvicolus when tested either intact or excised (p <
0.05). Predators responded least towards
He} i cotyl enchus i ndi cus (p < 0.05). The degree o-f
attraction towards other species o-f plant parasitic
nematodes was njoderate.
Food pre-ference (Table-I2)s Both species o-f
predators viz. , A. ameri canus and D. si 1vi col us showed
identical food preferential behaviour. They have
prefered mostly the second stage juveniles of H.
i ncoqni t a , Anqui na tr i t i ci and hi&t&rodera mothi and
killed them in maximum numbers when placed in one
combination with other species of prey <p < 0.05>.
The adults of H. oryzae and Tylenchorhynchus mashhoodi
were killed in moderate numbers (p < 0,05). A.
amera canus and D. si 1vi col us prefered individuals X.
basi ri T Lonqi dorus sp, and Tri chodorus sp. the least,
killing minimum number of them <P < 0.D5). No
individual of Hoplolai mus i ndi cus, Helicotylenchus
i ndi CUB and Hemi cycli ophora dhi renderi was either
killed or injured by A. ameri canus or D. si 1vi col us in
any combination. Hence, these are not included in
Table II).

4 85
DISCUSSION
Observations on A. ameri canus and D. silvicolus
showed that these species of predators were alike in
their attraction behaviour and food requirements.
Mononchs eare unable to detect their pr-ey unless a
physical contact between the prey and the predator is
estabalished (Nelmes, 1974; Bilgrami et_ a]^. , 1984).
Esser <19^3>; Bilgrami et. al_. i <1985>; Sbafqat et_ al .
(1987) and Khan et. al_. <1991) observed aggregation o-f
dorylaim predators around the previosly injured prey
and attributed this phenomenon to prey attractants.
Bilgrami ?< Jairajpuri <1989c> in their studies on
Hononchoi des Ion qicaudatus and M. fort i dens have
attributed the pre— and post —-f eedi ng aggregation of
predators at feeding sites to the lingering effects of
prey secretions/attractants. Bilgrami & Jairajpuri
<19S8> have also observed positive attraction of these
predators towards the live and excised pr"By and
factors influencing attraction. The present
observations on A. smeri canus sn D. si 1vi col us reveal
that these predators respond positively towards intact
and excised individuals of different species of plant
parasitic nematodes in response to their
secretions/attractants- Both the predators appeared to

4 8u
have reached the prey either by chance or wilful
movement CPoinar & Hansen, 1996), their orientation is
determined by probing movements of the head <Ward,
1973; Seymaour et_ a L- » 1983) and location of the prey
which depended upon the papillae and amphids (McLaren,
1976j Wright, 1983).
The differential response of predators towards
different prey suggests their preferential behaviour-
It could be possible that some nematodes secrete
repellents which are undesirable for predators
(Esser, 1963; Bilgrami & Jairajpuri, 1988) and hence,
affected their chemotactic behaviour. This appears to
be the case with Helicotylenchus i ndi cus which
elicited Jieast response of predators when tested
either intact or excised. A phenomenon which is
supported by the fact that predators did not prefer to
feed upon them and repelled from the site (Bilgrami 8c
Jairajpuri, 1989a; Bilgrami, 1992; 1993). The
attraction of A. ameri canus and D. si 1vi col us appeared
to have been affected by the period of observation and
the physical state of prey (intact or excised).
Attraction towards intact prey became significant only
after 12 h towards intact (live) prey and after 6 h
towards excised individuals. This difference could be

4 8 7
due to attractants of ewcised prey which dispersed
more quickely in a more quantity, -forming stronger
attraction gradient than those o-f the live and intact
i ndi vi dua}s.
It may also be related to predators attraction
threshold levels to perceive chemical stimuli as
predators could have -failed to percieve attractants o-f
weaker stimuli (intact prey) e&rlier than the
attractants o-f stronger stifljuii (excised prey).
Observations on -food pre-ferences of A.
ameri canus and D. si 1vi col us suggest that they have
certain degree o-f preference. The degree o-f
preference may be correlated to various physical,
chemical and behavioural characteristics which provide
resistance against predation and the type, volume and
chemical composition of prey contents (Jairajpuri St
Bilgrami, 1990). The second stage juveniles of H.
i ncoqni ta, A. tri t i ci and H. mothi were prefered most
possibly because of soft bodies and lack of anti-
predation characteristics (Bilgrami, 1993). No
predation on Hoplolai mus i n d i c u s and Hemi cycliophora
dhi renderi sp. may be attributed more to physical
characters like thick body cuticle and body
annulations as also suggested by experiments on the

488
attraction of predators which responded positively
towards these prey and fed consequently upon excised
individuals than the live and intact. A. ameri canus
and D. silvicolus also did not injure or kill any
individual o-f Hel icotylenchus i ndi cus. A phenomenon
which could be attributed to the
repellent/unattractive prey contents (Esser, 1963;
Grootaert et_ §LL- i 1977; Bilgraroi ic Jairajpuri, 19SS>.
The lesser deqree o-f attraction o-f both the predators
towards live and excised Hf?l icotyl enchus i ndi cus and
low predatory pro-file indicate resistance of chemical
nature in Heli cotylenchus i ndi cus <Es5Br, 1963;
Grootaert e?t al . . 1977; Bilgrami tt Jairajpuri, 1989;
Jairajpuri %t Bilgrarai, 1990),
ACKNOWLEDGEMENTS
The first author is grateful to the Council of
Scientific and Industrial Research New Delhi and the
second to the Department of Science and Technology,
New Delhi for the financial assistance.

!-" ^89
REFEREWCeS
BILGRAMI, A- L- <1992). Resistance and susceptibility
o-f prey nematodes to predation and strike rate
D-f the predators Mononchus aquat i c»s,
Dorylai mus staqnali s and Aquat i des thornei.
Fundamental and Appli ed Nematolopy 15, 265-270.
BILGRAHI, A. L. <1993)- Relationship betv^een
predation by Aporcelaimellus ni vali s and
different prey trophic categories. NeTiatoloqica
(in press).
BILGRAMI, A. L. & JAIRAJPURI, M. S, (1988). Attraction
of Mononchoi des lonqicaudatus and M. fort i dens
<Nematoda: Dipiogasterida) towards prey and
factors influencing attraction. Revue de
Nematoloqie 11, 195-202.
BILGRAMI, A- L. & JAIRAJPURI, M. S. <1989a).
Resistance of prey to predation and strike rate of
the predators, Mononchoides lonqicaudatus and M.
fort i dens (Neroatoda;Dipiogasrterida>, Revue de
Neroatoloqie 12, 45-49.
BILGRAMI, A. L. S< JAIRAJPURI, M- S. <19a9b). Effects
of concentrations and thickness of agar on the
predatory behaviour of Mononchoides

4 90
1 onqi caudatuB and M. -fort i dens <Nen)atoda
:Dipiogasterida> and on their attraction towards
prey- Indi an Journal of Nematology (1988) 1_8., 222-
227.
BILBRAHI, A. L.& JAIRAJPURI, M. S.<19e9c). Aggregation
o* t^ononchoi des lonqicaudatus and K. f or t i dens
(NematodaiDipiogasterida> at feeding sites.
Nematolociica <19SS) 34.* 119-121.
BILGRAMI, A. L. ?c JAIRAJPURI, M. 3. (1990). Predation
abilities of Hononchoi des lonqicaudatus and W.
forti dens (Neroatoda; Dipiogasterida> and
factors influencing predation- Nematoloqi c a
35. 475-488.
BILSRAMI, A. L., AHMAD, I- ?c JAIRAJPURI , M. S. <1983>.
Borne factors influencing predation by Mononchus
aquaticus.Revue de Nematol oqi e i , 325- 326.
BILGRAMI, A. L., AHMAD, I. S. JAIRAJPURI, M. S- <1984).
Observations on the predatory behaviour of
Mononc bus aquat i cus- Nematoloqi a Med i terr anea
12., 41-45.
BILGRAMI, A. L., AHMAD, I. ?< JAIRAJPURI, M- S. (1985).
Predatory behaviour of Aquati des thornei
(Nygolaimina: Nematoda). Nematoloqi ca (1984) 30,
457-462.

^3 I
BILGRAMI, A. L-, AHMAD, I. & JAIRAJPURI, M. S. <19S6)-
A study o-f the intestinal contents o-f so/ne
fBononchs. Revue de Nematol oqi e 9 , 191- 194.
COHN, E. & MORDECHAI, H. (1974). E>; peri/nent s in
suppressing citrus nematode populations by use of
merigold and a predaceous nematode.
Neroatol oqi a Medi terranea 2_, 43-53.
E3SER, R. P . ( 1 9 6 3 ) . Nematode interactions in plates
or non-sterile water —agar. Proceedi nqs 3oi1 Crop
Science FIorida 2 3 , 121-138,
GRDOTAERT, P., JAQUES, A. 8c SMALL, R- W- <1977>. Prey
selction in But1eri us sp. <Rhabditida ;
Dipi ogasteridae> . Mededelinqen Facultei t
Landbouwwetenschapen Ri j k suni versiteit Gent 4 2 ,
1559.
JAIRAJPURI, M. S. ^ BILGRAMI, A. L. (19 9 0 ) . Predatory
nematodes. In Nematode Bio—control: Aspects and
Prospects (Eds Jairajpuri et.al.> CBS
Publishers 8f Distributers Pvt- Ltd., New Delhi pp
95-125.
KHAN, 2., BILGRAMI, A. L- S< JAIRAJPURI, M. S. (1991).
Some studies on the predation abilities o-f
Apor C Q I ai me I 1 us rti vel i 5 (Nematoda : Dory 1 ai mi da ) .
Nematoloqi ca 3 7 , 333-342-

4 3 2
KULSHRESTHA, R-, BILGRAMI, A- L. h KHAN, Z. <1993>,
Predation abilities of Hy 1 onchul us mi nor r^nc,
•factors influencing predation. Annal s o-f PI ant
Protect i on 5c i ences <in press).
MACLAREN, D, L. <J976}. Nematode sense organs. In
Dawes, B (Ed) Advance Parasi tolOQV 14, Academic
Press, London ; 195-265-
NELME3, A. J. (1974). Evaluation of the feeding
behaviour of Pri onchulus punct tus (Cobb) , A
nematode predator. Journal o-P Ani mal Ecol oqy 43,
553-565.
POINAR, G- 0. Jr. & HANSEN, E. L. (1986). Association
between nematodes and bacteria. Helmintholoqical
Abstracts Series B 55, 65-31
SHAFQAT, S. , BILGRAMI, A. L. 9< JAIRAJPURI, M. S.
(1987). Evaluation of the predation ability of
Dory1ai mus staqn^li s Dujarding, 1845 (Nematoda :
Dory!aimi da) . Revue de Nematoloqi e 10, 455-461-
WARD, S. (1973). Chemotaxis by athe nematode
Caenorhabdi t i s eleqans J Identification of
attractants and analysis of the response by use of
mutants. Proceedi nqs of Natural Academi of
Science. USA 70. 817-821.
WRIGHT, K. A. (1983). Nematode chemosensi11 a : form
and function. Journal of Nematoloy 15, 151-158.

493
Table-I
List of plant parasitic nematodes used as prey and their hosts
Prey speci es Hosts
Meloi doqyne i ncoqni ta 'rifBiier Oder a mot hi Anqui na tr i t i c i Hel i cot yl ench<<s i ndi cus Hoplolai mus i ndi cus Ty ?. enchor hynchu s mashhoodi Hirschmanni el 1 a or yz ae Hemi cycli ophorg dhi render i X i phi nema basi ri Lonqi dorus sp. Tri chodorus sp.
Tomato Motha grass Wheat gal 1s Rose bed Palm Paddy Paddy Rose bed Hulberry Mulbery Rose bed

4 34 T a b l e - I I
P r e y s e l e c t i o n by A^ f^mer i c anu ' s i=<ric? D-_ s i 1 vi c o l u s i n d i - f - f e r e n t cc«r«bi n e t i o n <Pairf=-d c o m b i n a t i o r i s )
C o m b i n a t i o n s fio, o f p r e y J c i i l e d by p r e d a t o r s o-f p rey n e m a t o d e t i A ^ a m e r i c e n u s D . s i 1 v i c o l u s
P> P o P , P o
^ ' i p"^ H t ??D R a n e e H ± SD R a n g e M ± PD R a n g e M ± SD R a n g e
MI + AT l l i O . ? ? J 0 - 1 2 3 0 ± 1 - 1 9 - 1 2 9 + 1 . 0 8 - 3 0 9 ± 0 . 8 8 - 1 0 MI -i- HM 1 1 + 1 . 1 1 0 - 1 2 1 1 + 1 . 0 1 0 - 1 2 1 0 + 1 . i 9 - 1 2 1 0 + 1 . 1 9 - 1 2 Ml +{- ' '0 1 3 + 2 . 1 1 1 - 1 6 6 + r . O 5 - 7 11 + 1 .1 1 0 - 1 3 5 + 0 . 8 4 - 6 MI + TH 1 3 + 1 . 0 1 2 - 1 4 6 + 0 . S 5 - 7 1 1 + 1 . 0 1 0 - 1 2 5 + 0 . 8 4 - 6 MI + TP M i l . ? 1 2 - 1 6 5 + 0 . 8 4 - 6 1 3 + 1 . 1 1 2 - 1 5 3 + 0 . 8 2 - 4 MI + XB 1 5 + 1 . 0 1 4 - 1 6 3 + 0 . 6 2 - 3 1 4 + 1 , 1 1 3 - 1 5 2 + 1 . 3 0 - 3 MI + LN 1 4 + 1 . 1 1 3 - 1 6 3 + 1 . 0 2 - 4 1 4 + 1 - 1 1 3 - 1 6 2 + 0 . 8 1 - 3 HO + AT 7 + T . 2 5 - S 1 3 + 1 - 3 4 - 6 5 + 0 . 8 - - 6 1 3 + 0 . 8 1 2 - 1 4 HO + HH 6 + 0 . 9 5 - 7 1 3 + 0 . 8 1 2 - 1 4 4 + 1 . 1 3 - 6 1 4 + 0 . 9 1 3 - 1 5 HO + TM 7 + 1 . 3 6 - 9 7 + 0 . 8 6 - 8 7 + 1 . 3 5 - 8 6 + 2 . 2 5 - S HO + TR 9 + i . i 8 - 1 1 5 + 1 . 1 4 - 7 7 + 0 . 9 6 - 8 4 + 0 . 8 3 - 5 HO + XB 1 0 + 1 . 1 8 - 1 1 3 + 0 . 7 2 - 4 8 + 0 . 7 7 - Q 2 + 1 . 1 0 - 3 HO + LN 9 + i . ! 8 - 1 1 3 + 0 . 7 2 - 4 8 + 1 . 1 6 - 9 2 + 0 - 8 1 - 3 XB + AT 3 + 1 - 3 1 - 4 ' 1 3 + 1 . 1 1 2 - 1 5 2 + 1 . 1 0 - 3 1 3 ± l . i 1 1 - 1 4 XB + HM 3 + 0 . 8 2 - 4 1 3 + 0 . 8 1 2 - 1 4 2 + 1 - 1 0 - 3 1 3 + 0 . 8 1 2 - 1 4 XB + TM 4 + 0 . 8 3 - 5 9 + 3 . 0 8 - 1 0 2 + 1 - 1 1 - 4 7 + 0 . 8 6 - 3 XB + TR 4 + 1 . 3 3 - 6 8 + 0 . 8 7 - 9 3 + 1 - 3 1 - 4 6 + 0 . 8 5 - 7 XB + U4 4 + 1 . 3 3 - 5 5 + 0 . 8 4 - 6 3 + 0 - 8 2 - 4 3 + 0 . 8 2 - 4 AT + HM 1 0 + 0 . 8 9 - 1 1 1 1 + 0 . 8 1 0 - 1 2 9 + 1 . 1 8 - 1 1 1 0 + 0 . 8 9 - 1 1 AT + TM 1 2 + 1 . 3 1 0 - 1 3 6 + 0 . 8 5 - 7 1 3 + 0 - 3 1 2 - 1 4 7 + 1 - 1 5 - 8 AT + TR 1 3 + 1 . 3 ii-iA 5 + 0 . 9 4 - 6 1 2 + 0 . 3 1 1 - 1 3 4 + 0 . 8 3 - 5 AT + LH 1 5 + 0 . 8 1 4 - 1 6 3 + 0 . 6 2 - 3 1 4 + 1 . 0 1 3 - 1 5 2 + 0 . 8 1 - 3 TM + HM 6 + 0 . 8 5 - 7 1 3 + 0 . 8 1 2 - 1 4 4 + 0 - 8 3 - 5 1 3 + 1 - 0 1 2 - 1 4 TM + TR 9 + 1 . 1 8 - 1 1 6 + 1 - 0 5 - 7 7 + 1 . 1 5 - 8 4 + 1 . 1 3 - 5 TM + LN 1 0 + 1 - 0 9 - 1 1 2 + 0 . 6 2 - 3 8 + 0 . 8 7 - 9 2 + 0 . 8 1 - 3 HM + TR 1 4 + 0 . 8 1 3 - 1 5 5 + 0 - 8 5 - 7 1 2 + 1 . 6 1 0 - 1 4 4 + 0 - 8 3 - 5 HM + TR 1 5 + 1 . 3 1 3 - 1 6 3 + 1 . 0 2 - 4 1 4 + 1 - 6 1 0 - 1 4 2 + 0 . 8 0 - 2 LN + TR 4 + 0 - 8 3 - 5 7 + 1 . 0 6 - 8 3 + 0 . 8 2 - 4 4 + 1 . 1 3 - 6
C o m b i n a t i o n s w i t h H o p ) o \ a i m u s i n d : t C L ' S < H e l i c c t y l e n c h u s i n d i o j s a n d H e r e i c y c l i o p h o r a d h i r e n d e r i are not i n c l u d e d as n o pr&dation o c c u r r e d o n t h e s e p r e y ,
1 "^ Key; P = F i r s t p r e y ; P^ = S e c o n d p r e y ; MI - M^ i n c o g n i t a ; AT = A. t r i t i c i ; HM = | i ^ mot h i ; HO = Hi. o r y z a e ; TM = J_^ m a s h h o o d i ; TR = T r i c h o d c r u s s p ; XB = X_ b a s i r t ; LN = L o n q i d o r u s s p .

^ 95
LEGENDS
Fig- 1. Attraction o-f predators towards live and
excised prey
A. Attraction of AI lodorylai mus ameri canus towards live and intact prey.
B. Attraction o-f Al 1 odoryl ai mus amer i canus towards excised (cut in two pieces) prey.
C. Attraction of Di scolai mus si 1vi col us towards live and intact prey.
D. Attraction of Si scoi ai reus si 1vi col us towards excised (cut in two pieces) prey.
Fig. 2 Attraction towards different species of prey.
A. Attraction of Allodorylai mus ameri canus and Di scolai mus silvicolus towards live and intact prey- <A—Hoplolai mus i ndi cus; 5—Heli cotylenchus indi cus; C Hirschmanniella orys ae; D-Heloi doqyne i ncoqni t a; E—Heterodera mothi ; F — Anqui na tri ti ci; 8 - Hemi cycli ophora dhi rendrei ; G -Xi phi nema basi r i .
B. Attraction of Allodorylai mus ameri canus and Di scolai mus si 1vicolus towards excised <cut in two pieces) individuals of prey. (A— Hoplolai mus indieus; B - Heli cotylenchus i ndi cus; C — Hirschmanniella oryz ae; P—Meloidogyne i ncoqni ta; E—Heterodera mothi; F — Anqui na tritici; G -Hemi cycliophora dhi renderi ; G —Xi phi nema basi ri.

/i 0
; . / .
1,20 <v o u in
§"1.00-
c 03 CJ z
i./,o
0.80-
0.60
«—° H.oryzae <—X M.incognita
^ —^ Control
12 ^ 18 Tinne (h)
o (J \n en.
1.20-
S1.00
c fXJ QJ
0.80-
0.60 2L 6 12 18
Tinne (h) 2/.
2 o (J 1/1
UOi
1.20
?1.oo^
c ft) <v
0.80-
0.6 OL
C
12 18 Time (h)
UO
a.1.20 i _ o o in
51.00-c a;
—I
24
0.80-
0.50 - T 1 1 1
6 12 18 7L Time(h)
•1^;) •

130-1
o u U1
120- i ° 1.10'
c QJ
2.
I 100-
0.90 I
A
UO 1
1.30-
fe 1.20 o if)
en o
c (D
1.30-
1.00-
0.90
b^
A
i B
1 B
W
i I
I
A.americanus 4 97 D.siLvicolus ^
i I I $
^
I c D E
Type of prey
I
:ld
P 4 i i
I
i I
I
I 1
i c D
Type E
of prey F
I I
1 H
i
I i i I H

4 9 8
Sect i on o-f Nematol oqy , Department of Zool oqy Al i qarh riusl i m Uni versi ty^ Al i o^r h-202002 - Indi a
NUMERICAL ANALYSIS OF THE PREDATORY RELATIONS BETWEEN HE50D0RVLAIMU5 BASTIANI<NEMATODA;DORYLAIMIDA)
AND DIFFERENT PREY TROPHIC CATEGORIES
by
ANWAR L. BILGRAMI
Summary Numerical analysis of relations between the predator Mesodorylai mus basti ani and prey nematodes belonging to -five trophic groups viz., saprophagous, endoparasitic, ectoparasitic—epidermal feeders, ectoparasitic—cortical feeders and predaceous nematodes was made. Predators were most successful on endc-parasitic nematodes- Predaceous nematodes as prey e>!hibited high degree of resistance against predation by M. basti ani. Only IIX attacks resulted in wounding out of 31X encounters that resulted in attacks. Physical characteristics viz., thick body cuticle, body annulations, thick longitudinal cuticular folds are hypothesises as the cause of lOOX resistance of Hoplolai rous indi cus, Hemi cri conemoi des mangi ferae, Hemi cycliophora dhi renderi and Dorylai mus stagnalis. Mononchoides fort i dens, M. 1onqi caudatus and Hylonchulus mi nor also appeared to have resisted predation by their cuticular characteristics and behavioural patterns. Low strike rate of H. bast i ani and less feeding after wounding on Meli cotvlenchus i ndi cus has been attributed to repulsive/unattractive prey secretions (chemical resistance). Feeding time required to consume an individual depended upon the size of the prey- Maximum being on Paralonoi dorus ci tr i and the minimum on Cephalobus sp. The body cuticle, cuticle texture, quality, quantity and chemical composition of prey contents also influenced feeding by predators on different prey.
Studies have been made on predation by different
groups of nematodes (Esser, 1963j 1987j Bilgrami,
1990b> but no apparent effort has been mrndB so far to
identify ways to quantify predation except that of
Bilgrami and Jairajpuri (1989). They proposed

4 9 9
mathematical models to measure and quantify the strike
rate o-f the predators, Wononchoides 1 onqi caudatus and
M, -Fort i dens and the degree o-f resistance and
susceptibility of different prey nematodes. Earlier
observations made by Grootaert et al. (1977) on
But1eri us sp. and later by Small and Grootaert (I933>
on MononchuB aquat i cus ., LsbronemB vul vapapi 1 1 atum and
But1eri us dgqri sse have provided some significant
information in this regard- Still studies are lacking
which analyse predatory relations of nematodes with
different prey trophic categories.
In the present work the numerical analysis of the
relationship between predation by a dorylaimid
predator, Mesodorylai mus bast i ani (Thorne and Swanger,
1936) Andrassy, 1959 and nematode belonging to
different prey trophic categories have been made.
These studies may be helpful in establishing predator-
prey relationships of M. basti ani. The most
effective and suitable combinations of predator and
prey could be the most efficient predator <in terms of
highest strike rate) and most susceptible prey (i.e.,
most vulnerable to predation).
Materials and methods
Hesodoryl ai mus bast i ani was collected from the
Victoria Gate, Aligarh Muslim University, Aligarh and

500
cultured on sparophagous nematodes in petri—dishes
containing 1% water-agar at 28 + 2 °C <Bilgra«)i and
Jairajpuri, 1938). Prey neniatodes belonging to five
trophic categories we»^e selected for analysis of
predation (Table - I ) .
Fifty encounters were observed between each
combination of predator and prey. The encounters
(i.e., lip contact of predators with the prey at right
angles) between predators and prey were observed in
Petri-disbes containing i'/. water agar using a
stereoscopic binocular microscope. For each encounter
a fresh combination of predator and prey individuals
were used (irrespective of the predators failure or
success in attacking and wounding prey). To reduce
the effect of satiation and prey habituation, 4-6 day
starved adult predators were tested. A prey
nematode was placed in front of the head of an active
predator with the help of a fine needle without
touching (disturbing) the predator in any manner.
Only those observations were recorded where the
predators behaved as normally as could be ensured. The
strike rate (SR) of M. basti ani and the degree of
resistance (PR) and susceptibility (PS) of different
prey were determined and analysed by the methods of
Bilgrami and Jairajpuri (1989).

501
EA EA - AW (BRy.) = X 100 ; (PR-/.) = y, lOD ;
E EA
(PS%> = 100 - PR
where, E = Total number of encounters made by the
predators, EA = Total number of encounters resulting
in atteichi AW = Total number of attacks resulting
into wounding.
Feeding after wounding the prey (FW (X) = was
determined by the total number of prey individuals
wounded (AW) by the predators- Prey individuals left
unconsumed (PL'i) by the predators were also
determined. Feeding time ( F T ) , i.e., the time taken by
a single predator to consume an individual prey was
recorded.
The untransforroed results were analysed to obtain
coefficient of correlation ( r ) , standard deviation
( S D ) , standard error (SE) and coefficient of variation
(CV) using following statistical tests.
)!y
'3 2 ' x ^ y^ SE( r) ;
(x - x) SD =
N r N
SE = SD
N CV (X)
SD
M X 100

5C2
Wh«=>re >t flnd y BTP fhf deviations measured -from
their respective mean, 4 i B the total number of
replicates and W the mean o-f the data.
Result5
There was no correlation between encounters
resulting in attacks and a^ttac^^s resulting in wounding
by H. bast i ani on -free living nematodes but -feeding
a-fter wowunding depended upon attacks resulting in
wounding <r = 0.98, SE = + 0.006, p < 0 . 0 5 ) . There
was also no correlation between encounters resulting
in attacks and attacks resulting in wounding on endo—
parasitic nematodes-j cortical feeders and predaceous
nematodes when they were used as prey. Similarly the
relationship between attacks resulting in wounding
snd feeding a-fter wounding was i nsi gni-f i cant when
endopar asj t i r nematodes; cortical -feeders and
predators were used as prey for M. bast i ani . A
significant correlation was observed between
encounters resuslting in attacks and attacks
resulting wounding <r = 0.88; BE = + 0.05j p < 0.05)
and Attacks resulting in wounding and feeding after
wounding (r = 0.90j BE= + D-OAj p< 0.05) when
ejiidermal -feeders were used as prey. Hel oi doqyne
i nrnn,rti t a was the most susceptible prey <PS = 8 7 X ) .
Predators fed maximum <FW = 89%) on it. Aphelenchus
» r H i < i r e« e» ve.J mor e aJ } arks < RR = 84r.) with PR

17%. MaxjLmum individuals o-f A. tr i t i ci were le-ft
unconsufned (PL = 13%> by the predators. The minimum
deeding time (FT) was recorded on H. i ncogni ta (FT =
33 m) and ma>;imum on A. tr i t i c i (FT = 36 m) (Table —
II A) .
H. bast i ani was moBt BUCcesB^^ull on Xi phi neroa
i n s i q n e (SR = 727.). It wounded maximum individuals,
-fed most upon them but left maximum individuals of
Lonqi dorus sp. unconsumed (PL = 32X>. The cortical
•feeders were comparatively less resistant to predation
(PR = 18-19X> except Paratrichodorus sp (PR = 36%) and
Lonqi dorus sp. (PR = 29X). H. bast i ani took 102 m to
consume an individual of Paralonqi dorus ci tr i but
required less time to consume an individual of
Paratrichodorus sp. (FT = 40 m) (Table-II B>.
Hoplolai mus indieus, Hemi cri conemoi des
wanqi ferae , Hemi cycli ophora dhi renderi and
Scutel 1 onema sp,, were totally resistant (PR = 100%).
Interactions with Hirschmanniel1 a orys ae yielded
maximum SR = 72Ti. Most individuals of
Ty1enchorhynchus mashhoodi , H. orvz ae and
Aphelenchoi des were wounded (FW = 78-79%). Though,
75% of attacks on Helicotylenchus i ndi cus resulted in
wounding, predators continued feeding only upon 33% of
individuals. All wounded specimens were left

5 n /!
unconsumed <PL = lOOX). Ma>:imum feeding <FW = 83-
89X> was recorded on H. oryz ag and Aphelenchoides sp.
H. bast i ani required a n>a>iin)um o-f 53 m I D consume H.
pryz ae and a minimum of 35 m to -feed upon Basi r i a sp.
(Table-II C>.
Panaqrellus redi vi vus was attacked most <SR =
70%) by M. basti ani. The attacks on Cephalobus sp,
and Chi 1 ppl acus syrometr i cus yielded ffla>'. imum wounding
as the two nematodes were highly susceptible to
predation (PS = 71%-75X>. Rhabdi ti s sp. was reost
resistant species (PR = 84X>. Predators fed maximum
upon Q_ sywfnetr i cus (FW = 89%) and Acrobel oi des sp. ,
<FW = 95X> and left 10-15% individuals of P.
redi vi vus, Cephalobus sp. Acrobeloi des sp. and C.
symmetri cus unconsumed but none of Rhabdi ti s and
Acrobel oi des sp. Hini/oum FT = 33 m was required to
finish Cephalobus sp., and maximum FT = 57 m for
Acrobeloi des sp- (Table-II D ) -
Wylonchulus mi nor, Dory}ai mus staqnalis,
Mononehoi des fort i dens and M. lonqicaudatus were
totally resistant to predation by M. basti ani (PR =
1007.). The rate of wounding A_^ ni val is, M. bast i ani
and A^ thornei was low (AW = 21-267.) but that of
feeding was maximum (FW = 1007.). M^ basti ani left 25%

505
D-f their own members uncDnsumed but none of others-
It required a n>a>fimum of 70 m to consume A. nivalis
and a minimum of 43 m to finish h, aquat i cus. (Table —
II E) ,
Maximum encounters resulted in attacks with
endoparasites <SR = MSD 83'/. + 2.3: CV = 37.; p < D.05>.
Host encounters failed to p^rovoke predators to attack
other species of predators <prey> (SR = MSD 31% +
10-8? CV = 35y.j p < 0-05) (Table-III>.
The endoparasitic nematodes were most susceptible
as they were wounded in large numbers {AW = MSD
85 + 2.15, CV = 3'i; p < 0.05) - Predaceous group was
most res:i5tant to wounding <AW = MSD 11 + 2.4, CV
21%; p < 0.05) (Table-Ill).
Predaceous nematodes when used as prey formed the
most resistant trophic group (PR = MSD 84 + 15.9; CV
= 19%; p < O.D5>- Endo—parasitic nematodes showed
high degree of susceptibility to predation (PR = MSD
15% + 2 . 1 ; CV = 47%; p < 0.05) (Table-Ill).
Maximum variation in feeding after wounding
occurred epidermal feeders (CV = 99%) and the least
with endo-parasitic nematodes (CV = 2 % ) . Cortical
feeders were fed upon most by M. bast i ani (FW = MSD

c: Ob
eS ' i + 3 . 2 j CV = 4 % ; P < 0 - 0 5 > . The epidermal f e e d e r s
w e r e n o t p r B - F e r e d much b y t h e p r e d a t o r s a s o n l y 4DX +
3 9 . 1 o f w o u n d e d i n d i v i d u a l s w e r e c o n s u m e d ( T a b l e - I I I ) .
M. b a s t i e n i l e f t ina>iimun> c o r t i c a l f e e d e r s
u n c o n s u m e d (PL = MSD 2 6 + 4 - 2 $ CV = 1 6 ) b u t c o n s u m e d
m o r e i n d i v i d u a l s o f p r e d a c e o u s s p e c i e s <PL = MSD 3 +
2 . 8 ; CV =: 99'/. > .
M. b a s t i a n i t o o k R}a>jifrfum t i m e <FT = MSD 7 2 + 2 4 , 2
rain; CV = 3 4 % ; p < 0 . 0 5 > t o c o n s u m e art i n d i v i d u a l o f
c o r t i c a l f e e d e r s - T h e p r e d a t o r s r e q u i r e d a f n i n i m u m
a v e r a g e t i m e <FT = MSD 3 4 + 1 . 5 m i n ; CV = 4%j p <
O.D5> t o c o n s u m e c o m p l e t e l y an e n d o - p a r a s i t i c
n e m a t o d e s < T a b l e — I I I ) .
D i s c u B s i o n
The nematodes &re provided with physical
(Grootaert et. al . , 1977; Small ?c Grootaert , 1983?
Bilgrami, 1990a; 1990b>; chemical (Esser, 19<b3; 1987;
Bilgrami 8t Jairajpuri, 1988) and behavioural
characteristics to avoid predation by nematodes
(Bilgrami et. B1_. , 1984; Bilgrami 8c Jairajpuri, 1989;
1990). Besides, environmental and soil factors i.e
temperature, moisture, type, pH and chemical
composition also govern predation (Wallace, 1969;

sc?
Bilgraroi et. al . ., 1983; Azmi & Jairajpuri, 2979;
Bilgrami & Jairajpuri, 1985; Small, 1979>. Bilgranii ?f
Jairajpuri (1989> proposed models to quanti-fy
predaceous activities of H. fort i dens and M.
1onqicaudatus and considered wounding as an important
factor to determine the degree of predation by
predators (Small ?f Grootaert, 1983). H. bast i ani is
predaceous on nematodes belonging to different trophic
categories. Its relationship with individual pf^Y
species varied. The second stages of endo-parasitic
nematodes remained vulnerable to predation by H.
basti ani as they lack protective covering (Grootaert
et . al , ^ 19771 Jairajpuri ?< Silgrami, 1990; Bilgrami S<
Jairajpuri, 1990).
No predation on Hoplolai mus i ndi cus < Scutel1onema
sp. , H. manqi ferae ., H. dhi rendr i , Acr obel us sp. , M.
mi nor^ H. fort i dens, M. 1onqi caudatus and D. staqnali s
is attributed to physical charcter- istics e.g., thick
cuticle, body annulations and thick cuticular folds
(Esser, 1963; Small & Graotsiert , 1983; Bilqrami &
Jairajpuri, 1986, 1990). The saprophagous (Rhabdi t i s
sp,. , and C. symmetr i cus ) and predatory nematodes, <M.
for fc i dens, H. lonqicaudatus and M. mi nor) avoided
predatior* by behavioural characteristics (Bilgrami
efc. al., 1983; 19a5a; Bilgrami & Jairajpuri, 1989).

CP C8
Ec toftar asi t i c nematodes should be more
vulnerable to predation, specially by mononchs
<Bilgrami et . eil . , 1986; Jairajpuri ic Bilgrami, 1990)
but species o-F mononchs had contained intact,
undigested and whole individuals o-f Hopl ol ai mus,
Scutel1onema, Hemi cri conemoi des, Hemi cycli ophora <
Rhabdi ti s and Chi 1oplacus spp., which were resistant
to predation by dorylaimid (Sha-fqat et . al . , 1987;
Khan et. al.< 1991) and dipiogasterid predators
{Bilgrami ^ Jairajpuri, 1988; 2989; 1990). During
present *>tudies also ect opar asi t i c nematodes showed
high degree of resistance. The cortical feeders were
more susceptible than epidermal. This disparity may
be due to their large body size, inactiveness, soft
cuticle and inability to take evasive actions upon
being attacked by the predators,
Dorylaim predators repulsed from feeding and
regurgitaire the contents of Hel i cotyl enchus sp-
(Esser, 31963; Srootaert et. al . - 1977; Small, 1987;
Bilgrami & Jairajpuri, 1989; 1990). M_j_ bast i ani also
showed repulsive behaviour when wounded
Helicotylenchus i ndi cus and initiated feeding. The
duration of feeding depended upon the size of prey,
prey volume and the chemical composition of the prey
contents (Bilgrami et. al., 1985b>,

\j J 3
Acknowledgements! The author is grateful to the
Department of Science and Technology, New Delhi -for
providing -financial assistance.
Literature cited
AZMI M, I. and JAIRAJPURI M . S . , 1979.Effect of
mechanical stimulation and crowding on
Hyl anchul us dent at us. Nematol - medi t . , /[s 2 0 3-
20 7 .
BILGRAHI A. L., 1990a. Pi pioqasterid predators; In
Jairajpuri, Alaro ^ Ahmad (eds> Nematode Bio-
control s Aspect s and Prospects CBS publishers ?<
Distributors, New Delhi pp. 135-142.
BILGRAMI A. L., 1990b. Predaceous dorylaims; In
Jairajapuri, Al am and f^hmad (eds) Nematode Bio-
controls Aspects and Prospects CBS
publishers ?< Distributors Pvt. Ltd. New Delhi
pp. 143-148-
BILGRAMI A.L. and JAIRAJPURI M. S., 1985. The
resiponses of Hononchus aquat i cus to chemicals
and pH. Indian J. Nematol.,14;<1984>, 171-174.
BILBRAMl A. L. and JAIRAJPURI M. S., 1988.
Attraction of Hononchoi des lonqicaudatus and H.

5 1 Q
•f {3rt i dens (Nematode: Di pi ogaster i da) towards
prey and factors influencing attracti on-Revue
Nematol . , 1J_: 195-282.
BILGRAni A. L, and JAIRAJPURI M. S,, 1989.
Resistance o-f prey to predation and strike rate
oi- the predators, Honoochoi des 1 onqi caudatus and
H. fort i dens <NematodasDipiogasterida). Revue
N€?fflatol . .12; 45-49.
BILGRAHl A. L. and JAIRAJPURI M. S., 1990.Predation
by Mononchoi des forti dens and M.1onqi caudatus
<MeroatodaJDipiogasterida). Nematoloqi CB ^ 35;
475-488.
BILGRAHl A. L., AHAMD I. and JAIRAJPURI «. S.,
lS'83.SDffie factors influencing predation by
Hononchus aquat i cus. Revue Nematol . , 6_: 325—326.
BILGRAMI A.L., AHAMD I. and JAIRAJPURI M. S.,
1984,Dbservations on the predatory behaviour of
Hononchus aquat i cus. Nematol . Medi t. , 12; 41—45.
BILGRAMI A. L., AHMAD I. and JAIRAJPURI M. 5.,
1985a. Predatory behaviour of Aquati des thornei
<Nygolaiminai Nematoda). Nematoloqi ca, <1984>
30: 457-4A2.

51 1
9ILGRAMI A. L-, AHMAD I. and JAIRAJPURI M, S.,
19a5b. Factors in-fluence attraction o-f
adult Hirschmanniella oryz ae towards cabbage
B€?edling5- Revue Nematol . ^Qt 67—75.
BILGRAHI A. L., AHMAD I. and JAIRAJPURI M.
S.,19S6- A study of the intestinal contents of
sctme mononchs. Revue Mematol , , 9s 191—194-
ESSER R. P- •% 1963. Nematode interactions in plates
a-f non-sterile water-agar- Proc. Soi 1 Crop Sci .
Soc. Fla. , 23.1 121-138.
ESSER R. P., 19S7. Biological control o-f nematodes
by nematodes I- Dorylaims (Nematoda :
Dor vl aimida) . Nematol oqy Ci rcul ar , 4_: pp 4.
GROOTAERT P., JAOIJES A. and SMALL R. W. , 1977.
Prey selection in But 1eri us sp- <Rhabditida t
Dipiogasteridae>. Weded. F a c u1t. Landb.
Ri jksuni v, Gent, 42: 1559-1563.
JAIRAJPURI M. S. and BILGRAMI A. L., 1990.
Predatory nematodes. In Jairajpuri, Al an: it Ahmad
<ed5) Nematode Bio-control > Aspects and
Prospects CBS Publishers 8< Distributers Pvt.
Ltd., New Delhi pp 95- 125.

5 1 2
KHAN 2., BILGRAMI A. L.and JAIRAJPURI H. S., 1991-
ObsE'rvat i ons on the predation ability o-f
Aporeelaimellos ni vali s (Altherr, 1952)
Heyns, 1966 (Nematoda s Dorylaimi da) .
Nefftatoloqi ca, 37; 333—342.
SHAFPAT S. , PIl.GRAMI A. L. and JAIRAJPIJRI M- S-,
1987,Evaluation of the predation ability of
Dory1ai mus staqnal i s Dujarding, 1S45
(Nematoda s Dorylaimi da>- Revue Neffnatol. , ID;
455-461.
SMALL R. W., 1979. The effect of predatory
nemsiodes on population of plant parasitic
nematodes in pots. NematoloQica 25, 94—103-
BMALL R.W., 1987. A review of the prey nematodes.
PedobioJ oqi a. 30; 179-206.
SMALL R. V'J- and BRDOTAERT 8., 1983. Observations
on the predation abilities of some soil dwelling
predatory nematodes. Nematoloqica^ 29; 109—118.
WALLACE H. R., 1969. Wave formation of infective
larvae of the plant parasitic nematode,
Mel oi donyne javar^ica. Nematol oqica, 15 : 65 — 75.

5 1 3
T a b 3 e - 1 : C I a c s i f i c a t i on o f p i ant p a r a s i t i c nesatDdes used as p r e y and t h e i r h o s t s
prey t rophic rategoriBS (PTO
Prpy species Source of col l ec t j ori
Saprophagus Panaqre l lus redi vi vus^ rteaatodes lh11 op 1 f-cus sygjaetf i c u s ,
Cephfil obiis, Acrobel oi des , Rhabdi 115< Acrobelus spp.
(Linn,1767) Goode/, 1945 Laboratory nhorne ,1925) Thorne, 1937 c u l t u r e
M1 ijf ator y He} oi dogyfie incogni ta juvenileE
f u!Jo-par dEi tes) Aijjiujis tr i lit. i_
(Kofoid and White,1890) Chitwood, 1949 Khan and Hussain (1965) (Steinbuch, 1799) Filipjev, 1936
lo f f ia to
Hot ha qr a'E Wheat qalJ s
Epiderffiel He]icoty)enchus indicus S i d d i q i , 1963 feeders Hopl r>] ai aus indicus Sher, 1963 ( ec to - Tylerichorhynchus: ffiashhoodi Siddiqi and Bas i r , 1959 p a r a s i t e s ) Hirsc hsianni el 1 a oryjae (Breda de Haan,19D2)
Luc and Goodey, 1962 B a s i n a sp, HeBicycliophora dhi render i Hussain and Khan, 1967 Heai cr i confooides ixanqiferap S i d d i q i , 1961 Scutel1onena, Aphelenchoi des spp.
Rose bed Palft Paddy Paddy
Paddy Rosp bsd Flose bed Grass Paddy
Corti cal feeders (ecto-paresi te5>
Xiphinema smeri canum X. 1nsiqne Par atri chodof us sp. Paralonqi doros ci tri
Lonqi dof us sp.
Cobb, 1913 Loos, 1949
(Siddiqi, 1959) Siddiqi, Hooper and Khan,1963
Hedge Mulber / Mulbery Citrus
Mulbery
iV edaceous nt-watodes
Coetzee, 1968 (Cobb, 1913) Andrassy, 1958 Laboratory Dujardin, 3845 culture (Altherr, 1952) Heyns, 1965 (Butschlii,1873) AndraE5y,1959 (Schneider, 1937) Ahftad and Jairajpuri, 1982
(Schuurisans Stekhoven, 1951) Taylor and Hechler , 1966
tiononclioi des 3 onqi caudat us (Khera, 3 965) Andrassy, 1984
tinnonchus aquat i ' u s (^•iflonchu3 us ibi ucr DPI- yl ai I>UB st aqfial i s Aporcel ai ffiel 1 us n i v a l i s y£?5'iS'-1iiil.l_5iLL bast 1 ani Aquat1jes thornei
>k<nonchoi des f or t i dens

5 1 4
TABLE - I h Strike ra te of the predator,KfiodorYlaiBUS bastisrii and, resis tance end. MisceptibiHty of di f ferent prey
Prey species Ho. of
Wicr.uii-
ters
E
A i EHMPARASITIC NENATODEB ^loidc-fiyne incoqnata 50 Heterndera BotKi 50 fianqviina U i t i t i 50
trophic caleqories
Attacks
Encounters resulted into attack
EA (1)
40 42 42
B > ECTWARASITIC) CORTICAL FEEDERS ">r<hir.P%5 f<fiTn?r,>.M " 'P
)iir<hjnp»»a insi^ne 50 Lonqidorus 5p, 5!} Parelwxiidorus c i t r i 50 Psratricho'lorus sp. 50
?? Zh 35 34 33
Strike rate of predators SR a)
80 84 84
i4 72 70 63 bh
Attacks resiOted wounding
A» m
88 84 83
8i 81 71 82 i4
Wounding
Prey resistance
PR (I)
13 14
n
19 19 ?9 )8 Jh
Prey suEce-t i b j -l i t y PS<I>
87 U 83
81 81 71 82 64
Feed
Feeding after Houndi
FH
89 86 86
85 93 88 86 86
ing
(11
ing
Prey left uncon-suted PL(2>
ID 10 13
27
22 32 25 22
Duration of feeding (FT«)
33 34 36
67 61 89 102 ^0
C » ECTOPARASITIC i EPIDERHAL FEEDERS Ty^erichorhYnchtiS «S5hhOC<di
50 64 78 19 81
g^ftqi ferae He'sicYcliophora
Sasir ia sp.
50
50
24
33 30
48
66 60
79 60
100
21 40
79
60
80
89 72
20 41
Hirsch^snnieUft mt^r-r
Hriplolai^i's iridiru«^
Helicotylpnchus ind ic js
ScHteUonesa sp.
He*icricone«tDJdes
5ri 5n 50 50 50
36 ?! 24 31 21
72 56 48 62 42
78 0
75 0 0
22 100 25 100 100
78 0
75 0 0
83 0
33 0 0
20 0
100 0 0
C 7
Fe« sin — —
0
22 45 23 35
D I SAPROPHASOUS NEHATODES
Perts.jrelk'S redivivu=
Cephslfbiis sp.
firrc'bplffifles i p .
Rhsh'-'.itis sp.
Acrobelus rp.
Chiloplacus 5yf**»^tfuu?
E J PREOACEOOS NEHAT[IDES
Hononchus aquaticus
Hvlc'nchitl'js ftinnr
Dwylaifus '^Isanalis Aporcp!a!a»l]us ni'val n Hes<-<dorvlei8ws bs'itij'ifi Aquatide-i tbornei !lctiiO-iiiio'd~5
lor-qiraudaliis
'1orioni:h''!df'5 fDftide?iB
50 50 50 50 50 50
50 50 50 50 50 50 50
50
35 34 ?8 31 30 2^
23 83 11 16 17 23 12
!4
70 68 56 62 60 48
46 16 22 32 34 46 24
28
65 71 61 16 33 75
13 0 0 21 24 26 0
0
34 29 32 84 67 25
87 100 100 81 76 74 100
100
66 71 68 16 33 75
13 0 0 19
24 26 0
0
87 83 95 80 GO 89
67 0 D 100 100 100 0
0
10 15 0 0 13 13
0 0 0 0 25 0 0
0
4} T 7
57 39 51 36
43 — —
70 46 47 —
—
Pred&ii'' fed t'poii only one ir-ilividiisl of preyj E = Tola) nuabers of ericounters »ade between predators and pr(')5 EA= Eiicotjifters rPEultifnj in attacdsj SR = Strike ra le of Predators) AW = Attacks resul t ing in noundiniiiPR = Pri-y reristance) PS = Prey FusceptibiJityi FH = Feeding after Hounding preyi PL » Prey lef t unconsuaedj FT = Feeding t i t e .

5 1 5
TABLE — III: Numerical analysis o£ relationships between Mesodoryl aimus bastiani and di f-ferent prey trophic cataqories
Prey~frQphrc SR AW PR FW PL FT catagories
M + SD K + SD b5 + BD K + BD « + SD M + SD
Baprophagoue Aii;8.1 5^+73.5 45+24.0 8^+5.9 9+6.8 43+9.2 <13) <43) <54) <7> (75) <22>
E n d o p a r a s i - 8 3 + 2 - 3 8 5 + 2 - 5 1 5 + 2 . 1 8 5 + 2 . 1 1 1 + 1 - 7 3 4 + 1 . 5 t i c (3> (3> <47> (2> ( 1 6 > <4>
E c t o p a r a - ^ 8 + 3 , 2 7<t,+7.00 2 4 + 8 . 0 8 8 + 3 . 2 2 6 + 4 . 2 7 2 + 2 4 . 2 B i t i c : c o r t - <5> (11> <33) <4> {16) <34) ical feeders
Ectopara-sitic i Epi- 58+9.8 41+39,4 59+39.7 40+39,10 21+20.7 44+7.5 dermal feeder U 7 ) (96) (69> (99) (99) (17)
Predeceous 31+10.8 11+2-4 84+15.9 46+45.2 3+2.8 52+13.9 (35) (21) <19) <9S> (93) (27)
SR = StriV<e rate of Predators; AW = Attacl;s resulting in wounding; PR = Prey resistance; FW = Feeding after wD%.indijig prey; PL = Prey left unconsumed; FT = Feeding time; M = Mean; BD = Standard Deviation. All figures Brer nearest to whole numbers. Figures in parantheses are the CV values in percent.

51b HELMINTHOLOGIA, 23 179—188,1986
Studies on variability in Xiphinema radicicola Goodey, 1936 and Xiphinema brasilieme Lordelio, 1951
M Farooque RAHMAN*, Aowar I BILGRAMl, M Shamim JAIRAJPURI
Section of hematology. Department of Zoology, Ahgarh Muslim University, Aligarb-202001,
India
Received April 28, 1V84
Summary
The study of variability in Xiphinema radicicola and X brasihense indicate that all characters are either least or moderately variable except for the lengths of posterior gonad and prerectum which are highly variable None of the allometnc characters is highly variable in the adults of the two species The studies on the variability of different characters suggest that c(length of body/tail length) and c'(tail length/anal body diameter) values can be used as distinguishing characters between X radicicola and X brasi/iense
Key words vanabilny, morphometnc, allometnc, Xiphinema radicicola , Xiphinema brasilieme
Introduction
Variations in morphological characters occur in different populations and also within a single population of nematode species In taxonomy the study of variability of morphometnc and allometnc characters is very useful for the differentiation of closely related species The variations may be influenced by host plants (Goodey, 1952, Bird and Mai, 1965, Fisher, 1965), soil temperature (Rohde and Jenkins, 1957, Malek and Jenkins, 1964) and geographical location (Golden and Epps, 1965, Tarjan, 1969) In the following the variability in two very closely related species, Xiphinema radicicola Goodey, 1935 and X brasiliense Lordelio, 1951 was studied and the results were analysed These two species are very widely distributed in the world and in identification it is quite common to confuse one for the other The present study is not only aimed at determining the extent of variability of the two species but also to classify their taxonomic status and remove difficulties in their identification
* Present addiess Department of Zoology-Entomology, College of Agriculture, Assam Agricultural University, Joihat, Assam, India
179

517
Fig. 1. Variation in tail shapes of Xiphinema radicicola: A—C — Adult female tails; D—F — Fourth stage juvenile tails; G—I — Third stage juvenile tails; J—L — Second stage juvenile tails; M—O—First stage juvenile tails
180

D i 8
Fig.2. Variations in tail shapes of Xiphinema brasiliense: A—C—Adult female tails; D—F—Fourth stage juvenile tails; G—I — Third stage juvenile tails; J—L — Second stage juvenile tails; M—O — First stage juvenile tails
181

519
I
IX ^
m m
+1 I . ,, I
00 2S-
. +1 I I +1 s-^ • ^ • -^ ^ vo ro r; rt O " ' 00
, . . . +1 I +1 I o O Q i ^ . ^ r J o . ' *
oo +1 I
•s
^H V© t-^ „ „ ^ . . — ^-N 0 0 ^ - , r M , — , ( N , — , 0 0
+' 0I3 +1 I
« - ^
00 00 , ^ ,—, 00 ^ _: (S a \ -—V r~ rsi O 2 <^ rS "^
„ . , , +1 I +1 I +1 I +1 I +1 I +1 I +1 I +1 I
i o o~ <^ r5 "^ i T * ^ "^ •"- 7 < a <=> 5 r ° ^ +1 ' +1 I +1 I +1 I
•^ O tN CS
• ^^ 00 - ^ ; ' t o t^ •
so " I^ „ T-, , -r, . -ri , +1 I • o SO o o, °£. -^ "
' o i t^ I +1 I +1 . . . +1 I +1
_, 00 —. m
•~- r-j "^ t s ^ so +1 I
T ) - S " - ) ! 2 - > o ^ s o ^
•g
as 00 <N rn fN
f^ - ^ ,—, Q C T S O S ^ N S D O ^ N ' ^ ^ - V - , . — . - - " / ^ T j - c a c s i ^ ^ C T s ( N " o s ^ m i o ^ s O ' ^ - * ' * ° ^ r s l ' ^ > r i +1 I +1 1 +i I +1 1 +1 I +1 I +1 I +1 I +1 I +1 I
' ^ 3 ' ~ ^ ^ > n S s o l C ; , T f C i ^ 3 f N ° « n ! 2 .
i - • O O f O , ^ « ^ , t ^ ^ ^ O s
+1 I +1 I +1 I +1 I 1+ I +1 I +1
r; S - C
u-1 " ro
^ S ^
,—, (N 10 -^ ~ • T-< '^ ai ^ -r^
+1 I +1 I +1 I +1 I +1 I
^ ^ ° d^ s5^ e?i c,° ;:;
^ "
f u
JC a.
u
0 c 0
•0 0 "3 c
. 0 u c
s c 0
•T3 0 C u E 8 _« "E. (U
05
u ^-l 0 0. 0 C 0 s
00 , c 'C cm B
•3 •3 0
1 a 0
0
0 .
-0 a
c _2
•< S
182

6
in r-i r* I +1
o^ ^ o^ • ON ^ — r^
(N CN m o ; -+1 I +1 I +1 I +1 I +1 ^ m _ 00 _ u^ _ bR (-g " ^ " •* T
+1 I
S 3 i J2 „• "
I 4.1 I +1 I _L, , j_, I j_, m _L, 1/^ _ u-i -^
"" S <-, +1 I +11+1'^
1 f*^ ^ ^ r-j ^ ^ ^ ^ • —
r<-i --y
«S " ° = m
^ -^ _f '' ^'
f S • : in <o
,. I +1 . +1 I •n a -o ^.
!> CN sO 1/-1 \D "n vO
O , -H S O tN ;^
- " tn • o -;
+1 1 +1 I +1 I +1 I +1 1 +1 I +1 I +1 I
eg • — • ^ ^
+1 I +1 I +1 I +1 I +1 I +1 I vc (^ 1^ —1 — _ 1*
Zi <^ Z. ^ O T-l '^ ^—'
+ 1 1 + 1 1 + 1 1 + 1 1 . rsl rj- °
+ 1 1 + 1 . : o ?; o "^
+1 . . , + 1 1 ^ <N CO IT)
^ ^ o r ^ ^ v n ^ o ^ o v T ) ^
:-<~H ^ ^ 00
+1 I +1 I +1 I +1 I +1 I +1 I +1 I +1 I
^ 2 -H Ct>o ^ c n Ci , , ; r j , - ^ 2 ,
u E .S '•5 ><
c E U Id
3 00
ij "o Ji
+1 _!. (N CM
^ a e •5 Of 00 C
£ 2 « 5 T3 O
c J> -5 . §
u a;
c <
= S i B S S s >
n I 11
I E
II E J : c *
= >
520
183

521
C +1 I +1 I +1 I +1 I r^ ^^ ^ ' +1 I +1 I +1 I +1 I
OS V-) • VI ,—^
O >0 'O .—. 0^ t^ T— sC " ^
«= 7 c 5) d <N d c;> ri 2 -• +1 . i +1 I - ' " - ' ' -L' I
rts ._ rr
_ " s +' vi +1 ] +1 7 +1 Y +1 in « " ^ "
l / ^ ON , _ ^ 0 0
'— -.O CJS ^ r ^ S ^ 2 ^ so ON - ^
. . . +1 I +1 I +1 I +1 O N i / ^ S , m 2 > 0 < ^ — ' " "^
O 21 <N "
I +i I O ' ' 1 < 2 " !
•* o vo 12, en J2.
!
• NO '. ^~ in ^ -. „ ~ " <> S~ <^ , -- S d ;: ° ^ ' - ' S
; , +' I +1 . +1 s :q , 00
+1 ON Tl- C l o O 2 ~ ^ oo S£, w
I +i u-> QO ' ^ r o
. " 00 j : : ^ ^
- " S "^ S f^ ^ . , . , , + 1 1 + 1
00 5 - :C; = " o m ci
- . rn S I +1 I •• +1 I +1
i s
I fl
2 ^
r j o +1 +1 I +1
i:; '^ t- m ui o m - , ^
2 rsi --i rn ON lo
+1 I +1 I +1 •* J ^ NO 2 00
fN g ON r< in
I +1 I +1 I O) JO T t • *
o o t ~ - T f ^ > ? ) ! 2 , ' * O r ? , 2 ,
} (SI
f-> {:: - r; d r t (Nl .—^ O ^ ^ —I
+1 I +1 I +1 1 +1 I +1 I +1 I +1 I O N ? ^ O N : 2 r - i - m C < N ? o
m
r^ ^ -^r .^1 "* I +1 I
- . _ m ,£; 00
+1 I +1 I +1 I < N 2 £ T > O N s O < N - i - 9 0 u - l ' * '~ •^ 2 00 S
tie c a
o c 0
T3 O
15 c ,2 t j c 3
c o
T3 O
c u E
u
O. «:
0
a o c o 2 0
00 c
on c
•5 '3 0
3 00 CO
x: a o C/3
u
( ^ M
o * d
CS
a
' 1 U4 a>
184

0 0
+1 I +1 I +1 I +1 I +1 I +1 I +1 I +1 I • r ^
+1 I
m 0 0 Ov 1 / ^
, - ; v O 0 0 , ^ O ^ ^ • » ^ - ^ ^ ; - ^ ^ ^ ^ ~ ^ - H ^
O )
vO O 2 '~- 0 0 0 0
, •* .^O -—, r
+1 I +1 I +1 I +1 I +1 I +1 00 O „ VO o 2 < >n 2 f ^ f C- "1
. • r^ ^ ^
^ ~' S ° m I +1 1 +1 1
o 9 o S w "
22 ° 2 " s ^ 5 ° 5 - ^ =; 2 I +1 I +1 I +1 I +1 I +1 I +1 I
' ^ ; r d 2 ' » 2 < N S d w d O*, 1-H 1-H - ^ r j "
. . , + 1 1 + 1 1 + 1 1 o o o p o o f r - ' " ^ " ' ^
+1 ^"S
00 ON ° So
I +1 I +1 I o o ° o o ^ r r - : « ) C ^ > - ; 2 -
I +1 ' ••
s " s •* +1
« •3
2 m >,-a
2 f -3 = OC
•O O u O —
- S "o S 2
"I
11 is
1
c
li II If
C Q V
B 2 u eg
S ^ E >
R 2 2 o ^ ^
185

523 Material and methods
The soil samples containing X. radicicola were collected from around roots of tiamboo from Cinnamara Tea Estate, Jorhat, Assam and those of X. brasi/iense from around roots of MachiY/us sp. from Botanical Gardens, Shillong, Meghalaya. These samples were processed by decantation and sieving and modified Baermann's funnel method. The nematodes were killed in hot X % (double strength) formaldehyde and dehydrated by the slow method. Measurements and observations were made on specimens mounted in anhydrous glycerine.
The comparison of characters of the two species were made after determining their CV values. ("V was obtained by the formula MSD x 100 M; where MSD is the mean standard deviation and M is the mean value of the character. CV (Coefficient of variability) is expressed in terms of percent. The basis of categorization of variability is as follows: least variable CV = 0—5, moderately variable CV - 6—lOand highly variable CV = 11 or more. Forty specimens of each juvenile stage and adults of the two species were examined for this study.
Results
Morphometric and allametric variations in Xiphinema radicicola ind X. brasiliensedifier to varying degrees for almost every character (Tab. I. II).
In adult X, radicicola and X. brasilierise the lengths of odontostyle. odontophore, tail, position of guiding ring, vulva and anal body width are least variable (CV = I—5). The total body length, body width and width of lip region is moderately variable in X. radicicola (CV = 6—7) but least variable (CV = 4—5) in X. hrasiliense The width of vagina {15—18 nm in X, radicicola and 15—22 ^m in X. hrasiliense) and the length of rectum is moderately variable (CV = 6—10) but lengths of posterior gonad and prerectum are highly variable (CV = 13—26)inbothspecies.Noneof the allometric characters is highly variable in the adults of the two species. The values of a, band V are least variable (CV = 5) but cis moderately variable (CV = 7) in X. radicicola. while all these ratios are moderately variable (CV = 6—9) in X. hrasiliense. There is no correlation between the body length and the lengths of odontostyle, odontophore,gonad (136—322 ,um in X. radicico/aand 116—368 iim in X. hrasiliense) and tail in adults and the juvenile stages of both species.
The morphometric and allometric characters also vary differently in different juvenile stages of both the species. The prerectum of all, except third stage juvenile of X. radicicola showed highest variation (CV = 12—14) while those of X. brasi/iense exhibit most variation in prerectum and in the height of the lip.
The shape of tails of the first stage juveniles of X. hrasiliense and X radicicola similar except that it is much longer in the former (Figs 1, 2). However, in the later stages the convexity on the ventral side of tail in X. hrasiliense become fairly apparent.
Discussion
Studies on the variability reveal that the morphometric and allometric characters vary to different degree in the two species of Xiphinema. The present population of X. radicicola has a smaller body, smaller a and V values than the type population (Good e y, 1936) and smaller body and lesser total stylet length than the populations described by McLeod and Khair (1971), Baja j and Ja i r a jpu r i (1979) and Luc (1981). From Luc's population of X. radicicola it also differs in having a lower V value. The X. brasi/tensepopulation is identical to the type population (Lorde l lo , 1951), the Ceylon population described by C o h n a n d S h e r (1972) and Brazil population by Loof and Sharma (1979), but it differs from the
186

5 2 4
populations described from Ivory Coast and Australia (Luc, 1981) mainly in its body length
and value of V The two speties are very much alike except for differences in values c and c', shape of tails
and in the number of caudal panillae (two m X radicico/a and three in X brasihense) X brasi/iense has greater c but smaller c' and tht tail is pegshaped with its ventral surface distinctly convex (the ventral surface is eithei •-traight or concave in X radiacola) This agrees with the observations of C ohn and Sher (1972), Bajaj and Ja i r a jpu r i (1979) and Luc (1981) Herce, c and c values can be used as distinguishing characters between the two species The total stylet length is different in the two species (144—ISZ \im in X radiacola, 185- 197 |Lin in X brasihense) which does not conform with L uc (1981). According to McLeod and Khair (1971), the total stylet 'ength m X radiacola is extremely variable which IS not the case in the present population of X radiacola The V ratio overlaps in the two species which is in agreement with Cohn and Sher (1972) and hence, cannot be used as a distinguishing rhar Kter
jVcknowledgcments
We thank P'of Nawab H Khan Chairman of the department for providing research facilities and to Prof AtherH Siddiqitorhisvaluablesuggestions l-irstandsecondauthorsare thankful to the U G C New Delhi 'or providing tmanuai assistance
References
BAJAJ, H K, JAIRAJPURI, M S (1979) A review of the genus Xiphinema Cobb, 1913 with descriptions of species from India Rec Zool Sur\/ India,! 5 25S—325
BIRD, G W , MAI, W F (196*>) Plant species in relation to morphometnc variations of the New York population of Tnchodorm christiei Nematologica, 11 34
COHN. E SHER, S A (1972) A contribution to the taxonomy of the genus Xiphinema Cobb, 1913 J Nemato/,^ 36—4S
FISHER.J M (196'^, Studies on Paraty/ency7us nanus 1 Effects of variation in environment on several morphometnc rharactcs of adults Sematologica, 11 269 -279
GOLDEN, A M, FPPS. J Vi (196'>) Morphological variations in the soybean-cyst nematode Nematoiogica, 11 38
GOODEY, J B (I95z) The influence of the host on the dimensions of the plant parasitic nematode, Ditylencbus destructor Ann App Biol, 39 468—474
GOODEY, T (1936) A new dorylaimid nematode Xip/ijnemarac/icico/a n sp J Helminth ,\4 69—72 LOOP,? A A,SHARMA, R D (1979) Plant parasitic nematodes from Bahia State, Brazil The
genus Xip/iinemaCobb, 1913 (Dorylaimoidea) Nemato/ogica, 25 HI—127 LORDELLO, L G E (1951) Xip/i/nema brasi/iense, nova especiede nematoide do Brasil,parasitade
Solanum tuberosum L Bragantia, 11 87—90 LUC, M (1981) Observations on some Xiphmema species with the female anterior genital branch
reduced or absent (Nematoda Longidoridae) Revue NematoJ, 4 IS''—167 MALEK, R B , JENKINS, W R (1964) Aspects of host-parasite relationships of nematodes and hairy
vetch N J Agric Expt Sta Bu/;,831 1—31 McLEOD, R W,KHAIR,G T (1971) Xiphinema australiaei\ sp , its host range, observations on X
radicicoJa Goodey, 1936 and X monohysterum Brown, 1968 and a key to monodelphic Xiphmema spp (Netiatoda Longidoridae) Nemato/ogica, 17 58—68
ROHDE, R A, JENKINS, W R (1957) Effect of temperature on the life cycle of stubby root nematode Phytopatclogy, 47 29
TARJAN, A C (1969) Variation within the X/phinema americanum group (Nematoda Longidoridae) Nematoiogica, i 5 241—252
187

c " ^ Di-
Mccjie;(OBaHHe BapHaSejibHocTH Xiphinema radicola Goodey, 1936 a Xiphinema brasiliense Lordello, 1951
BbWOubl
Hi MccjiedOBc.HHH aapMaScibHoc'M Xiphineuid raditoln M X. brasiliense BbrreKaeT, 4ro BCC JHaKH B HH'iKOH Hcpe/iHcM cTeocHH aBJiMion's rcptMKHHbJMH. « MCKJiiOHeHHeM sajjHOH fOHaj]binripepcKTyMa, Koropbie HMeioT 3bicoKyio creneHb BapMaSt.ibHoctH. IlpH nojroBope.ibix 4)opMax o5eHx HeMaxofl HHKaKOH m aji.ioh-eTpuHeamx jwaKoB He riojjaer Bbjciixyw creneHb BapHa6e.ibHocTH. HcMeaoBaHMe pa-jHbix iHaKOB noKaahiBaer, ^ro napaMfxp c (;(.iHHa re.ia a.iHHa XBOcra) H c'(a.iMHa XBOCTa aHawerp rejia Ha ypoBHC ana-ibHoio oTBepciHH) MO>KHO ynoipe&iai'b KHK OTJiHqHTe.ibHbie iHaKH Me>K;iy X. radicola H X. brasiliense
M. (PapyK PaMan AHBap n, EmrpaMH
M. ZliaMHM fliipaHnypH
.Sludiuin der Vaiiubilitat vun Xiphimeiita radicimla Gtiodey, 1936 und Xiphinema brasiliense Lordello, 1951
Zusammenfbssijng
Aus dem Studium der Vanabiiitat von Xiphinema radiacolaund X. brasiliensegehi hervor. dass alle Zeichen in der niedcren und mittleren Stufe veranderlrc*! sind, mit Ausnahme der hinteren (jonade und des Vorrektums, die eine hohe Sture der Variabilitat haben. Bei geschlechtlich reifen Furmen beider Nematoden weist keines der allometrischen Zeichen eine hohe Stufe der Variabilitat auf. Das Studium der Variabilitat verschiedener Zeichen zeigt, dass der Parameter c(K6rperlange/Schwanzlange) und c' (Schwanzlange/Durchmesser des Korpers in der Ebene der Analoffnung) als Unterscheidungsmerkmale zwischen X. radic'co/a ind X. brasi/wnegebraucht werden konnen.
M. Faroque Rahman Anwar L. Bilgrami
JVf. Sbamim Jairajpuri
Studium variability Xiphinema radicicola Goodey, 1936 a Xiphinema brasiliense Lordello, 1951
Siihrn
2a stiidia va-iability Xiphinema radicicola a X. 6rasi7)ensevyplyva, ze vsetky znaky sti v nizkom az strednom stupni premenlive, s vynimkou zadnej gonady a predrekta, ktore majii vysoky stupen variabilnosti. Pri pohlavne zrelych formach oboch nematodov ziadny z alometrickych znakov nejavi vysoky stupefi variabilnosti. Studium variabilnosti roznych znakov ukazuje, ze parameter c (dizka tela/dfzka chvosts.) a c'(dlzka chvosta^priemer tela v urovni analneho otvoru) mozu sa pouzr'vat' ako rozlisovacie znak> medzi X. radicicola a X. brasiliense.
M. Faroque Rahman Anwar L. Bilgrami
M. Shamim Jairajpuri
188

SHORT COMMUNICATIONS 2 3 9
Macrolaimellus longicauda (Rashid et al , 1985) comb. n. syn. Cephalonema longicauda Rashid et al., 1985 nee Cephalonema longicauda Cobb, 1893.
Because of this transfer, we consider Macrolaimellus as belonging to the Cephalobidae.
We thank Prof. Dr. Andrassy and P. A. A. Loof for their Icind information.
REFERENCES
ANDRASSY, I (1966) Erd- und Susswasser-Nematoden aus Ghana Klasse Secernentea (Phasmidia) Annales Umversitahs Scientanum Budapestinensis de Rolando Eotvos Nominatae 8, 5-24
ANDRASSY, I (1984) Klasse Nematoda G Fischer Verlag, Stuttgart, 509 pp COBB, N A (1893) Nematode worms found attackmg sugar cane Agricultural Gazelle of New
South Wales 4, 808-833 RASHID, F , GERAERT, E & SHARMA, R D (1985) Morphology, taxonomy and morphometry
of some Cephalobidae (Nematoda Rhabditida) from Brazil, with descriptions of two new genera and four new species Nemalologica 30 (1984), 251-299
A. L. BiLGRAMi, S. M. ALAM and I. AHMAD') ; Histochemical localization of some enzymes of carbohydrate metabolism in a sod nematode Dorylaimus stagnalis.
Various enzymes of carbohydrate metabolism have been detected biochemically by several workers in free living and plant parasitic nematodes (Read, 1960; Krusberg, 1960; Barrett et al , 1971; Castillo & Krusberg, 1971; Lee & Atkinson, 1976) but few studies have been made on histochemical localization and differential distribution of some of these enzymes in the body organization of these nematodes (Deubert & Zuckerman, 1968; Dietze & Kampfe, 1973). In this study some enzymes of carbohydrate metabolism have been histochemically localized in different parts of the soil nematode Dorylaimus stagnalis Dujardin, 1845.
Adult D. stagnalis freshly isolated from paddy fields were incubated in toto in medium containing an appropriate substrate of a particular enzyme for 20-24 h at 25 + 1°C. The nematodes were then dehydrated in ascending grades of alcohol (3-4 min in each grade) and cleared in xylol (1-2 min). The material was embedded in 56°C wax and 5 fim thick transverse sections of the body were cut by rotary microtome and mounted in DPX. Controls were run in the same incubating medium but without the appropriate substrate. The following enzymes were localized: aldolase, phosphoglucomutase, fructose 1-6 diphosphatase, alcohol dehydrogenase, malate dehydrogenase, succinate dehydrogenase and glucose 6-phosphate dehydrogenase. Succinate dehydrogenase was localized by the technique of Pearse (1968) and the other enzymes by the methods of Shaw & Prasad (1970).
') Department of Zoology, Aligarh Muslim University, Aligarh-202001, India

527 240 SHORT COMMUNICATIONS
• J m <
a bo
3 g
o Q
s
c o
O jj "a
11
0-g.
2-S c o
Q "
S O
5 "J CO 3
a. E
C 3
61
N C
I I I
I I
: I
+ + + +
I I
I I I
lo re
I I I
ID O " U g I - 0
re
re s ^ c ° c bo O- bo ° Ln O
> « > J3 O J3
O
+ +
re
re bo
3^ £ < S c/: O

SHORT COMMUNICATIONS 2 4 1
Table I summarizes the concentration of various enzymes in different parts of D. stagnalis. Malate dehydrogenase, succinate dehydrogenase, glucose 6-phosphate dehydrogenase and fructose 1-6 diphosphatase (Fig. 1, A-E) did not differ much in intensity from each other in different parts of the nematode. These enzymes showed weak to intense activity in the region of the inner cuticle, hypodermis, somatic muscle cells, hypodermal chords, oesophagus, and intestine. No enzymatic activity was detected in the outer cuticle. In the gonads of both sexes malate and succinate dehydrogenases were weakly present but the activity of glucose 6-phosphate dehydrogenase and fructose 1-6 diphosphatase was high. Phosphoglucomutase (Fig. 1, F) was not detected in the outer and inner cuticle but in other regions it was weakly or only moderately present except the intestine where it was maximum. Weak to moderate activity of alcohol dehydrogenase and aldolase was observed in hypodermis, somatic muscle cells and the two lateral chords. The other regions did not show positive reactions for these enzymes. No enzyme activity was observed in the control.
The Embden Meyerhof pathway (glycolysis) has been reported present in different species of free-living and plant-parasitic nematodes (Krusberg, 1960; Barrett et ai, 1971; Lee & Atkinson, 1976). In D. stagnalis it also seems to operate as suggested by the presence of aldolase, phosphoglucomutase, fructose 1-6 diphosphatase and alcohol dehydrogenase. Glycogen is known to be stored in the hypodermis, somatic muscle cells and in the four hypodermal chords (Lee & Atkinson, 1976). It may be because of this that the four enzymes were present in comparatively higher concentrations in the above regions than others. The presence of glucose 6-phosphate dehydrogenase, an enzyme of the pentose phosphate shunt (an alternative route to the tricarboxylic acid cycle), suggests that probably this nematode is also capable of bypassing the main metabolic pathways to obtain its required energy as suggested by Krusberg (1960). Because of the possible occurrence of the pentose phosphate shunt, malate and succinate dehydrogenases might be involved in gluconeogenesis (Krusberg, 1960) or the TCA cycle (Barrett et al, 1971). Malate and succinate dehydrogenase involvement in gluconeogenesis seems likely because the end product of this cycle is glucose which is utilized in the pentose phosphate shunt. However, it is equally possible that both the cycles i.e., pentose phosphate shunt and the TCA cycle occur simultaneously or alternatively according to the energy requirements of D. stagnalis.
Thanks are due to Professor Ather H. Siddiqi for reading the manuscript and for his valuable suggestions.
REFERENCES
BARRETT, J., WARD, C . W . & FAIRBAIRN, D . (1971). Lipid metabolism in the free-living nematodes Panagrellus redivivus and Turbatrix aceti. Comparative Biochemistry & Physiology 38, 279-284.
528

5?.9. 242 SHORT COMMUNICATIONS
E
in N
smc
« ;aK r
Fig. 1. Histochemical localization of some enzymes in different parts ai Dorylaimus stagnahs A: Malate dehydrogenase; B: Malate dehydrogenase; C: Succinate dehydrogenase; D: Glucose 6-phosphate dehydrogenase; E: Fructose 1-6 diphosphatase; F; Phosphoglucomutase. Key: cu = cuticle; hp = hypodermis; smc = somatic muscle cells; Ihc = lateral hypodermal chords; oes = oesophagus; cr = cardia; int = intestine; ov = ovary; od = oviduct; oc = oocyte; ut = uterus;
vg = vagina; vu = vulva.

\j \j V* SHORT COMMUNICATIONS 2 4 3 O \J
CASTILLO, J M & KRUSBERG, L R (1971) Organic acids of Ditylenchus triformis and Turbatrtx aceti Journal of Nematology 3 , 284-288
DEUBERT, K H & ZucKERMAN, B M (1968) The histochemical demonstration of cytochrome oxidase in fresh frozen sections of nematodes Nemalologica 14, 435-455
DiETZE, H & KAMPFE, L (1973) Histochemical demonstration of various enzymes in normal and nematicidal treated nematodes Wissenschafthche Zeitschnft der Ernst Moritz Arndt-Untversttat Gretfswald, Mathematisch-Naturwissenschafibche Reitre 22, 1 7
KRUSBERG, L R (1960) Hydrolytic and respiratory enzymes of species of Ditylenchus and Pratylenchus Phytopathology 50, 9-22
LEE, D L & ATKINSON, H J (1976) Physiology of Nematodes The MacmiUan Press Ltd London, pp 215
PEARSE, A G E (1968) Histochemistry, theoretical and applied Vol I, I l i rd edition Churchill, London, pp 759
READ, C P (1960) The carbohydrate metabolism in worms In Comparative physiology of carbohydrate metabolism in heterothermic animals (edited by Martm, A W ) University of Washington Press Seattle, 3-34 pp
SHAW, C R & PRASAD, R (1970) Starch gel electophoresis of enzymes - A compilation of recipes Biochemistry and Genetics 4, 297-320
I. J . PARUTHI and D S. BHATTI* ' ) : Comparative morphology and contents of galls formed by Anguina tritici {Nematoda Anguinidae) on various parts of wheat plants
The juveniles of Anguina tritici normally form galls in place of wheat grains (Byars, 1920; Leukel, 1924; Gupta & Swarup, 1968). But galls on other parts like leaves (Byars, 1920) and awns and glumes (Gupta & Swarup, 1968) have also been reported. In addition to these parts we have noted galls on lemma and palea, but there are no details on galls formed on these tissues. This study provides such information.
Green (immature) galls from leaf, glume, palea, awn and floral tissue were collected on March 7, 1980 and ruptured in water to determine their contents. Egg counts were made by a dilution method. Similarly black (mature) galls were collected on May 7, 1980 and their contents noted
Galls on the glumes were found at the base over the median vein; these were more or less conical with a lobed tip (Fig. lA) and were formed at the base of the glume either laterally (on the sides) or centrally One gall was found on the anterior side of the glume.
Galls were normally observed on the apical side of the palea (Fig. IB) but were sometimes found on the central portion; they were normally round to oval in shape.
Galls were observed at the base of leaf blades adjacent to the portion where the leaf sheath was attached. The galls developed on the abaxial side and these were oval shaped and some were lobed. Lobes were on the apical portion of the galls (Fig. 10).
') Department of Nematology, Haryana Agricultural University, Hisar-125 004 (India) *) Director Extension Education, Haryana Agricultural University, Hisar-125 004 (India)

bo 244 SHORT COMMUNICATIONS
Galls on the lemma were observed at the base either laterally or centrally; they were also conical in shape having a pointed tip (Fig. ID).
Galls developed at the base, centre and tip of the awns; they were almost globular in shape with conspicuous lobes. Thickness and length of such awns were significantly reduced (Fig. IE).
The data in Table 1 show that the average number of juveniles found in the green galls collected from non-floral tissues ranged from 3.5-10.3 compared with 59.8 in galls formed in floral tissues. More juveniles in galls formed from floral tissues may indicate that these organs are a preferential site for penetration providing more food and space for development and reproduction of the nematodes. At the time of examination, green galls formed on any other than floral tissues did not contain eggs (except for a very few in galls on the palea) whereas the galls of floral organs, on average had 19500 eggs/gall. Corresponding increases in the juvenile content of black galls over green galls of nonfloral tissues indicates late oviposition in these galls. This delayed oviposi-tion may be due to fewer juveniles penetrating, fewer juveniles undergoing development to the stage of oviposition and reduction in ovulation and period of oviposition.
Fig. 1. Galls on: A-glumes; B-palea; C-leaf blade; D-lemma; E-awns.

•v* w >~
Nematode Pest Identification. (Ea. l.S. Jairajpun, 1939)
SPECIALIZED TECHNIQUES IN NEMATODE TAXONOMY:
NUME.^ICAL TAXONOMY
ANWAR L. BILGRAM! Section of Nematology, Department of Zoology,
Aiigarh iMuslim University, Aligarh - 202 002, India
Ta:.onomy as we all know is the science that concerns v<ith the theory and praaice of anirral classification. Thus, it deals with the description of species and placing thenn into higher taxa on the basis of their degree of similarity or dissinniiarity. Altho^'gn, new techniques are continuously being employed for studvirg living creatures, in discovering new characters, in descriDing new species, and in revising the cassiiication of previously knov n organ,snns, little work has been directed towards the conceptual basis of classification. Indeed, the taxonomic methods of tccay are but little aavanced from these of a hundred, or even t'AO hundred years ago. Biologists have amassed a wealth of material, both of museum specimens and of new taxonomic charaaers. However, todays taxonomy or the theory of ciassii.cation still needs modifications to make it experimental, mathematical, stat^-tical £ind more importaniiy aemonstraole to errors. Tnus, durj- g the last few years there has been an increasing awareness of the proolems m the aims ana practices of taxonomy. There has been an increasing interest in the development of various new techniques. These techniques have proved :o be successful tools for the taxonomists m putting the science of taxonomy on firm footing. Besides, these methccs may also help in filling the lacunae o: the conventional taxonomic procedures whicn depend more on qual.tative estimations rather than quantitative values. Numieri-cai Tax'3nomy is one of these technicues. There has been an increasing interest m developing numerxal methods in taxonomy as an aid to make systematics a Qualitative science.
The ideas on whicn Numerical Taxonomy rests go back to Anderson, a ccntemoorary to L.nnaeus. Tne present mac develcr-ment of th.s mef^oa is cresL-~aDl\ a result o: n e cexeiopme-: o: comouter tec"^n,c:ues. Numerical Taaonomy a ms :c deve :: methods wnicn are obiectne anc '•eceatable. bo-n ,n esaiuat-v--^ 01 taxoncm'C aif.n^t.es anc .n the e-'ection ot :x\a. In addit.v?-', ' 'uTiO'-:'ai mcr.ocs r ax ocon „:: a wic.'' i.eld .n rn. e \^i : "oasu ?-mont 01 e\ou.; onc.r\ rates arj, proN uie a more d t ; . . ! uOpro_^ • to ph\logenet:J oroo.e^^s.

v> C 0
e
1'12 issmaioae fnu lueiuiiiLunun
Definition: Numerical Taxonomy may be defLned as ;he numerical e /aluation of "Lhe affinity or similarity between taxonomic units arid the ordering of these units into taxa on the basis of their aJfinity, It mav Lnclude the drawing of phylogenetic inference from the data by stat^tical and mathematical methods to the extent to which these should prove possible. These methods almost involve the conversion of information about taxonomic entities into numerical quantities.
Numerical Taxonomy involves the measurement of a large number of spec.mi^ns followed by statistical and mathematical enumerations and finally computer analysis. All these processes establish the numerical degree of affinity among unit characters, individuals, specimens, species and higher taxa. The affinity values when achieved can be used to erect hierarchial order of taxonomic categories. So, a specimen may be placed in a certain species or a species in a certain genus when it has a numerical value falling w.thin the range of that particular category or into different species or genus when it has other range of values.
THE AIMS OF NUMERICAL TAXONOMY
Numerical Taxcxiomy would have no claim to the serious attention of biologists unless it could overcome some of the faults found in the conventional taxonomy. Although, we cannot expect scientists ahvavs to ag- ee on the interpretation facts, it is the ain-i of scientific meihodolcgy to reach agreement on the facts themselves througn the repeatability and the objectivity of the observations. Hence, the outstanding aims of Numerical Taxonomy are the repeatability and objectivity. Besides, the goal of Numerical Taxonomy is also to oDtain accurate and identical estimates of the resemblence between two forms of organisms given the same cha-acters b\ different workers. So, by including many characters without previous arbitrary selection or elimination and by providing stanaard metnoos of processing the data and evaluating the results, we reduce sub|e:tive bias and hence, increase ODjectivit\. The obiectivity and repeatability togetner lead to ve -y staole taxonomies which may not be altered by later discove'-ies.
EasicalK, there are eight necessary steps invoi\ed in Numerical Taxonomy i.e.. Choice of specimens; Discovery and measurement of cnaracters; Coding of characters; Calculaticr. of affinities between species; Clustering of specimens into phenons; Extraction of data from taxa; Identification of specimen; and Computer analysis.

5 3 4
iYU;/ie( n-ui iu.<.u<'vjiM/ j l j
CHOICE OF SPECIMENS
The choice of specimens for identification is rather an important aspsa. Utiile selecting specimens for numerical analysis the following points may be taken into consideration:
a) A very large number of specimens should be collected to establish a species; b) A large number of species should be taken into consideration while erecting a higher taxon; c) A preliminary selection of specimens must be made in order to restrict the study to a manageable taxonomic group; d) Selection of specimens should be based on rather few important characters; and e) Tne collection of specimens should be made from large areas, different habitats, geographical areas and ecological locations.
DISCOVERY AND MEASUREMENT OF CHARACTER
The procedure in TcLxonomy, whether orthodox or numerical is based on taxcnomic characters. The term character has been employed in at least two distinct senses by systematists. Its commonest usage is as a distinguishing feature of taxa i.e., a characteristic (or feature) of one kind of organism which distinguishes it from other kind. Thus, a taxonomic character is any attribute of an organism or of a group of organisms by which it differs from an organism belonging to a different taxcnomic category or resembles an organism belonging to the same category (Usinger, 1953). Another frequent meaning of the term character, which has been used by numerical taxonomists, as being the more useful one in their work, is that a character is a property or "feature" which varies from one kind to another (Michener & Sokal, 1957) or anything that can be considered as a variable independent of any other thing considered at the same time (Cain ic Harrison, 1958). The Taxonomic Characters can be grouped mto following types;
aj Morphological characters (externa!, internal, microscopic, cytolo-gical and developmental characters); b) F'hysiological characters; c) Beha.viourai characters; d) Ecological and Distributional characters (habitats, food, hosts, parasites, population dynamics, geographical distribution etc.)
A taxonomic character may be a two state character when a unit character is comprised of two charaaeristics (or states) or a muitistate chiracter when a unit charaaer is comprised of more than two characteristics. The proper selection of characters is clearly a critical point in the application of Numerical Taxonomy, There are, however, certain kinds of characters whose nature clearly disqualifies them from employment in a Numerical

C "I T 0 0 :;>
i-vK/.-jf-uuB rai l laeniificaiion
Taxonomy study. Tnese characters may be referred to as inadmissible characters-
INADMIS5IBLE CHARACTERS
Meaningless characters: It is undesirable to use attributes which are not a reflection of the genotypes of the organisms themselves. For this reason taxonomists do not include the names or num.bers given to species or spec:rr,en, nor do they em.ploy characters whose response to the environment is so variable that it is not possible to decide what is environmentally and what is genetically determined.
Logically correlated characters: There are certain characters which are logically interdependent and hence, they cannot be considered in Numerical Taxonomy. The length and half of the length of an organ can not be employed together or haemoglobin and its colour cannot be used simultaneously for the purpo.-<e of differentiation.
PartisJ logical characters: Situation: may arise- when the dependence of (V.- haracter upon another is not total -but only partial e.g., the circumference and the radius of the body or organs.
Invarljible characters: All those characters which do not vary within the entire sample of organism or taxa.
After the selection each character is measured in terms of numerical values and weightings are given to them depending upon their degree of similarity and occurrence. While giving weign-tagf to different features, each feature should be given equal weightage. Before creating a taxonomic group a decision should be made as how to weigh the features which are to be employed for classification. If somehow.differential weightings are to be given, exact rules for estimating them should be framed.
CODING OF CHARACTERS
In a two st3te character positive characters ma\' be mari^ea as prctjcnt or - c: 1 wni.e negative characters as absent or -or 0. Besides, pro\ision must also be mace tor entries unich cannot 3e scored, either becau- e tne character? are missmg or unscoi',-'G or inapplicable. For e\am?;e the data mas' be unlvnewn. or the specimen is camacec or there mav be .oj^ical bar \\r,;oh prevents scoring oi tho o' trw SU^T, entries ma\' be scorcc as \ C (standmi- f ir "no comparison is to be made with this entPr'").

535
3^5
The multi-state character can each be expressed bv a single numerical value i.e., they can be arranged m order of their magnitude along one directional axis e.g., 1, 2, 3, u, 5 (corresponding to their range of variation).
CALCULATION OF AFFINITY BETWEEN SPECIMENS
The affinity betv,een specimens can be obtained by three different types of Coefficients: a) Coefficient of Association; b) Coefficient of Correlation and c) Coefficient of Distance-
a) Coefficient of Association:
Tl-iese Coefficients are also known as Coefficient of Similarity or Coefficient of Relationohips or Matching Coefficient. Review of the different types of coefficients whicn have been proposed and u;^d can be found in Cole (19 +9; 1957); Dagnelie (1960); Soka! & Sneath (1963).
Tlie basic arrangement of data for computation of Coefficient of Association is the familiar 2 x 2 type table. An example of such an arrangement is given below.
OTU (Taxon) j
OTU (Taxon) k nJK r.,K
nJk njk
nJ nj
nK
nk
T^e above data consist of "n" characters scored for two OTU's (Operational Taxonomic Units) labelled as "3" and "k". They are suDdivided into positive and negative classes for each of the two OTL's. The capital letters mav be used to ircicate positive or 1 Stat? and lower case letters to show negative or O states. The num.oer of cnaracters m'whicn both OTL's are positive in the a:30\e table is :a::e.!ed as ''nJK", the numre'' n wn.c"; both are negative as njk. Tie numcer cl characters :^o^.::\e :c: cne OTL' and negative :cr a^^other are "nJk" and "niK" respectively. Th.e marginal totals :cr pobitive cnaracters "3" anc "K" are "nJ" and "nK" robcective.v. Similarlv "nj" and "nk" are *"e mar-;nal tetai> for negative craract fs " j " and "k". Thus, the "!i.. ~-3er 0: cnaracter-m "matched cells-" or m will be m-nJK - ni s anc :no nu^re-^ of charact?rs \r unmatched cells or u vvi , ;n. - n iK".

f.: •;> 7
KemaiQde Pest Iceniificaz.on
the number of characters or n will equal to
n = m + u = nJK + njk -'r.lk ^ njK = nJ -r nj = nK + nk
Thie 2 x 2 type set up of the data in Numerical Taxonomy is primarily a convenient arrangement and may not be confused with the conventional 2 x 2 tables used for tests of independence in Statistics. Besides the above, there are siill other possi:;ie kinds of Coefficients of Associations which have been used for determining the degree of similarity between two characters or species. These coefficients are named after the author wno proposed the formula to compute the Coefficient of Association. Some of the important and commonly used coefficients are mentioned below.
The Coefficient of Jaccard: The Coefficient of 3accard '^j) omits the consideration of negative characters. It is representetl as follows:
nJK 3 ~ (nJK + u)
The Simple Matching Coefficient; The Simple Matching Coefficient (S ) i: ,sm charactf ed as;-
(S^ ) is equivalent to Jaccard except that this includes negative ch'aracters also. The Simple Matching Coefficient may be represent-
S sm
m (m - u)
The Coefficient of Rogers and Tanimoto: The Coefficient of Rogers ard Tanimoto irciudes cnaracters with mere than two states ana alio ccnsice'-s missirg informations. The Coefficient of Rogers and Tanimoto (Snj) can be rcoresentea as follows:-
^RT m
m - 2u
m tn - u)

Numerical Taxonomy Yl
Tne Coefficient cf Hamann: Tne Coefficient of Hamann (S ^ n
employs the difference between matc'-^ed and unmatched '"a -• 25 3 r r ' - ^ o r t o r t o f 3f;cr,r- " - '^.">. T r i e r^.^^-^n^c^n-*- i—^ ^— i_ . ,
to 1. as a cn t enop of assoc.a:.o-^- Tnis coeffcient ranges fce'weT
c _ m - u H ~ n
The Coefficient of Yule: The Coefficient of Yule (5 ) has bee-described as the simplest possible and most aavantageous methoc to calculate the degree of assoc.ation (Brisbane ic Rovira, 1961). However, its limit is only i .
c _ (nJKnik - niKnJk) Y ~ (nJKnjk -r njKn.lk)
The Pii Coefficient: The Phi Coefficient (SC) is also called focr fold point correlation coefficient. The limits of Phi Coefficient is same as that of Yules.
5- (nJKn)k - njKnJk) " ~ 1/9
(nJnjnKnk) '
b) Coefficient of Correlation:
These coefficients have been employed in Numerical Taxonomy by Micherer and Sokal (1957), Sokal & Michener (1958), Sokai (1958). The coefficient may be computed between taxcn " j " and "k". Thus,
•"ik " 1 = 1
n £ (xii - Xj) {\ik - Xk)
Where;
r — Coeffic ent of Correlation
X = Character state value of charact. 'r V m OTU V-
X = Ciiaracter star? value o: character 'i ' in OTL' ^ • 1 N
5:8

539
313 Nentatode Pest Identification
X] = Mean of all sta^e values for OTU j ,
Xk - Mean of all state values for OTU k,
n r Nurnber of characters sampled.
Since this forn ,ula is bas d on moments around the mean, It takes into account the magnitudes of mismiatch' d cnaracters betvveen taxa with more than two states. It is more s'^penor cceffic.ent than the Coefficient of Association discussed above.
Besides the above, the following Coefficient of Correlation is also beneficial to determine the degree of correlation as this coefficient also possess properties similar to the above.
Thus, < XY
r =
2 2
Where X and Y a/e the deviations measured from their respective means.
c) Coefficient of Distance:
It is also called as Taxonomic Distance. The distance between two OTUs can be obtained by the following formula:
Ai = f.^h--^)'] m
CLUSTERING OF SPECIMENS INTO PHENONS
There are se\eral techniques of clustering specimens into phencns or describing taxonomic structures e.g.. Frequency Distri-but on ol Correlation Coefficient: Differential shading of similarity mainx; Cluster analysis and Factorial analysis.
Frequency distribution of coefficient of correlation: Ilium.'^atin:; ins^nt into pnenetic reiarionsnio among OTUi in a Numerical Taxonomy stuJ\ can be cDtamea b\ in insp<"'ction of frec'..enc\ distriDuti.^n o: the Coefficient Correlation (r). F;Pv:iinf out tne Drinai\, ,oco:var\. tert:j.r\ n w e etc., amon-; tr.o Correlation Coctficicnt.s .v'- t;v» bK'WJ.litv or mulnrrodalitv ol fv.- Corolation Ci.K'fIic;ent n\i\ be helDiul in clustering OTUs in a guen ^:Jd\.

540
humenccl Taxono"'}' .^^
Differential shading of the similarity matrix: This is the rrcs* ODVicus techriGop for recognu.ng an overall glance of the groupings among the OTUs of a similarity cceif.cient matox. Tne'methix! consists of accot "g a s' st^-^ oi g rouc-g the similarity coeffic"ie"i into f.ve to ten ^veniy spacea classes arranged b , orce ' of macni-t jde and reorese^'s eacn oi these classes bv a diftprg-^ de^°'-p° of shading in the sauares of a half m.atrix. G°^erally, t -e hi-^host value is shown carkest ana lowest value ligntest. Tre shading patterns neea to be (cnoseni wiih care so t r a t th°y present an even progression. Thus, a snaded sim.ila'it) coefncient matrix exhiDiting reascraolv good init.al clustering can be made ana by careful rearra-'g'=meni of the sequences oi OTLs the clusters can b? more sna-oiy denned and areas of lignt and aark can be separated (Sokal i Sneath, 1963).
CLUSTER ANALYSIS
Tie cluster analybis means numencal tecnniques for defining grouDS of relatec OTLs based on hign similarity cceffidents. Clustering may be acnieved by the following methods.
Elenfwntary cluster analysis: This is the simplest of the various metnods avaiiaOie for clustering. It mav be obtamea with the help 01 similant% matrix and aooears as the reilection in a mirror set along diagona.lv .However, these techniques o: clustering require, primal ily an arbitrary selection of a level on the sca'e oi similarity coeificient.
Clustering by single linkage: This method oi clustering differs from elementarv cluster analvsis in tnat the a rb i fa r ) select.on of the level ot similar,tv matrix is not required in th.s case. Instead, OTUs v.rxh have higrest possible similaritv coefficients are cluste-'ed f.rst and then it successiveh lowers tne level of admission bv stecs of equal magnituaes. Thus, ii the firs: criterion IS a similarity c:>^!fic e^t ot r-j. '^°, the next ones mav be r = 0.9S: r r 0.97; r . 2.^6 and so on.
' r ' OTUs
O.QQ 1, 2
o.'^s 1 , : . 3 o. -^: 1 . 2 . 3, ^
0..S0

541
520 Nematode Pest Identificz
This indicates that OTUs 1 a' d 2 joir at a similar valL° of r - 0.99, OTU 3 joints them at r - 0.98; OTU 'f at r - 0.97 and OTU 5-7 at r - O.SO.
Clustering by complete linkage; Tnis corresponds in most deta.is to the aoove method except that admission of an OTU into cluster is by a complete linkage.
Centrjii or nodal clustering: Tiis method as proposed by Rogers 6c Tanimoto (1960), is a clustetxrg method but its actual procec-ire IS markedly different from ot^er methods of clustering. In tnis type the matching coefficients are expressed in logrith'mic form. The nost typical OTU heie is called the primary node of the study and around it is formed a cluster of OTUs having high similarity coefficient with it.
FACTORIAL ANALYSIS
Sokal (1958) has been the first to employ FactoriaJ Analysis to indicate taxonomic relationships from similarity matrix. Factorial Analysis, when applied to correlations among taxa, may be interpreted as a statistical method for describing the complex interrelationships among taxa in terms of the smallest number of factors. Factorial Analysis also predicts which OTXJ is incorrectly placea by the weighted g''oup metrcd. This limitation of Factorial Analyijs might be an advantage in that it prevents attempts ot the _irterpretation of differences v-hich are probably not reliable.
REPRESENTATION OF RESULTS OF CLUSTERING
Siniiarity matrices which have been clustered by sirole shading methods are not generally represented in any other form exceoi as the dark triangles. Howe\er, if the clusters are grcured again one or mere secondary jnatrices nnav be shosvn in wn.ch new OTL reprebe^is a cluster oi former OTU. One could conceive of suci shaaed ciagrams as coss-^ectional transects throug". a dend'-0£;ram. Tne most common and convenient representat.on of the results o; nu^-.-ncal bt d es is by denarogra.-^s. Exceot for the'r rectan^ ar nature :he\ a::Dear ve;v much like. Lusto~arv pnvlocenetic tro-^. The absc-ssa ot such a dendogran has no special meaning, v?r\mg onK to seoarate the OTUs. \\ne''eas, the ordinates are in some Similarity coefficient scale, us-^a.u from ;oro to oro ana trequc^riv multiplied b\ 100 or b\ 1-33 m orc"- to avoia decimal places.

5 4 2
Numerical Taxonomy 32i
EXTRACTION Of DATA ON TAXA
The extraction of data' from taxa is yet another important aspect of Numerical Taxonomy. It requires accuracy in measurements and judgements. The informations collected from taxa should be arranged in such a manner that it is easy to understand easy to manage, easy to compare etc. Preparation of keys is perhaps the best way for. grouping informations. Dichotomous keys a^e not always reliable tools for identification of nematode species particularly in genera with a -large number of species showing high intra-specific variability. The dichotomous keys are rapidly outdated and difficult to update. A new key is needed after a number of new species have been added. However, identification of species by means of polytomous keys permits a range of characters to be considered simultaneously. A description of a new species can be simply added to the polytomous type of key. Besides, polytomous keys have been proved a good basis for preparing computer programmes for Numerical Taxonomy.
COMPUTER PROGRAMMING FOR NUMERICAL TAXONOMY
As computer get faster and the so called automatic coding systems become more flexible and powerful, it is likely that programmes for Numerical Taxonomy will no longer be written in Machine' languages but in the automatic coding systems. Ini the Un.ted States of America FORTRAN, developed by IBM and in Europe ALGOL seem to be the most widely used languages for addressing a great variety of computers. Fortran programmes are currently being written for Numerical Taxonomy procedures, but these will have to be recompiled at different computation centres in order to adapt them to the specific installations. The learning of a language such as FORTRAN is Extremely simple and that a little acquaintance A'ith the computational aspects of the work would pay copious dividends.
Programmes that are to be written for Numerical Taxonomy should have detailed write-ups to enable machine operators, unfamiliar with the computations and taxonomists unfamiliar with the computers to do the work with maximum facility. Therelore, each programme write up should -
1) des<:ribe the general idea of what the programme will do;
2) describe the maximum .capacity oi the prcjranim.e-how many OTUs can be processed and how many characters;
3) de:s."ribe in detail the actual algebra of the programme, unless

543
322 Nematode Pest Icentijication
It IS very standard proced-ure in which reference to the publication would be adequate;
^) give detaibd operating instructions of what to do c" given computer to execute this programme;
5) specify the exact format in which the data is to be presented to the computer;
6) specify in detail the format in which the output of the programme will De presented;
7) add time estimates for executing the programme on a given computer;
8) provide some examples of prepared data input in order to amplify point 5;
9) provide some examples of prepared data output in order to amplify point 6.
COMPUTER PROGRAMMES IN NUMERICAL TAXONOMY
Eignt different kinds of computer programmes for Numerical Taxonomy are en\isaged at present:
Grcup-I Control Programmes: These are the master programmes which control the succeeding (i.e., Group II to Group VII) progi'ammes.
Group-ll Language translation Programmes; These programmes take cescriptions of characters in words and be pro-gramred in such a way as to convert these into numerical ccces in some logical and consistent fashion. Sucn programmes eventually may be able to remove sucn a teG...m of coding characters.
Grcup-n Processes ard Convert Character Programmes: 5uch prcgra-.imes preparf^ characters for the calculation of bir..arity coe:licients.
Group-iV Calculate Affinity or Similarity Coefficient Programmes: Th' se .''":'i;iammob ar" required to compute the de ' ee ot .'.:'.'iit\ OT -MtniiaritN coeiricionts between pairs ot Ori 's accorc.p-; to the forruias discus'^'d ear:.er.

5 4 4
Numerical Taxonomy 323
Group-V Cluster Analysis Programmes: Tne cluster analysis programmes take the output of Group-IV programm.es and make clusters from this data. Tne Cluster Analyslj Programmes also convert the clusters into dendrograms, printing these in some suitable forms.
Group-VI Data Extraction Programmes: To extract out informations from old and new literature.
Group VII Inter Study Coordination Programmeii: Such programmes store and sort " previous studies, establish reference taxa and their characters and correlate different studies.
Group-VIII Publication Programmes: These programmes convert the output into logical and pubiishable form such as diagnostic keys with descriptions of characters and organisms.
APPLICATION OF NUMERICAL METHODS
. IN NEMATODE TAXONOMY
In plant parasitic nematodes the importance and analysis of quantitative characters in describing nematode species has been recognized since de Man (18S0) vvhen he introduced a system which designated three principal body ratio's 'a' 'b' and 'c' fcr the nemalode identificatiai. Later, many other.ratios and quantitative characters have been included in the- taxonomy of nematodes (Rehman et al., 19S6). However, these methods fail, at times, to establish the degree of similarity and affinity and to quantify the degree of relationship between individuals, species, genera, populations etc. Thus^ to achieve these aims the use of numerical methods has gained importance during the last 10-15 years and yielded satisfactory and convincing results. B.ra (1967) and Bird ik Mai (1967) obtained satisfactory results from numerical studies of 2- species of Trichodorus. Rey et al. (19SS) used numerical methods to identify spec.?'' of Longidorus. Whereas Rey i \lahajan (19SS) identified species of Tylenchcrtiynchus and Merlinius by numerical methods and computer prcgranmes.
Tne rumerical methods ma\ help :^emato!o;•.^;s in the ident:-ficati.-'n cl ncnafcdes (Lima. I'^iO; -3oa; J. 5rith, 19S3; 19S-; Fortuner, 19S ': Fort'.inor & Won;. l'><-; "lancaaa .V Lrna. 19S^; MobS A- Webiior, l^~\j) an>i t!io ro- , wonre o; sporie-i to or,-" anot'v^r ina\ be quaniified (•r.o.\.^r, ',•";; Sn^ui' , .*c " Oval. 19/ -; R. y, 1- S7; Piiikhain 5c "carson. \')Jh\ \'arioib ao.npiKor .n^nhoas

c: 45
iVK/ziuiOue i'est Identification
have been developed for identifying nennatode species (Rey et al-,19S8; Rey & Mahajan, 1988).
ACKNOWLEDGEMENTS
Tlie author is grateful to Professor M. Shamim Jairajpuri and Dr. M. Hayat for going through this manuscript and also for their suggestions.
REFERENCES
BIRD, G.W. (1967). Numerical Analysis of the genus Trichodorus. (abstr) Phytopathology 51, 804.
BIRD, G.W. &. MAI, W.F. (1968). Morphometric and Allometric variations of Trichodorus christieLi Nematologica (1967), 13, 617-632.
BOAG, B. & SMITH, P. (1983). Computer assisted identification of nematodes. Systematic Parasitology 5, 235-239.
BOAG, B. <5c SMITH, P. (1984). Advances in the use of the computer as an aid to the identification of nematodes. Proceedings of the 1st International Congress of Nematology, Guelph, Canada, 5-10, August, 1984.
BRISBANE, P.G. & ROVIRA, A.D. (1961).A comparision of methods for classifying rhizospere bacteria. J. Gen. MicrobioL 26, 379-392.
CAIN, A.J. &. HARRISON, G.A. (1955). An analysis of taxonomists judgement of affinity. Proceedings of the Zoological Society of London 131, 85-98.
COLE, L.C. (1949). The measurement of interspecific association. Ecology 30, 411-424.
COLE, L.C. (1957). The measurement of partial interspecific associaticn. Ecologir 38, 223-223.
DAG\'ELIE, P. (1960). Contribution a I et..de d:s communcutes vegetales par V anal\se facto--^lle. Bull Serj. Cirte. Phyto-geogr. (B). 5, 7-71, 93-195.

546
Numerical Taxonomy 323
FORTUNER, R. (19S3). Computer assisted semi automata fication of Helicotylenchus species. The programme NE.\...iD. California Plant Past Disease Reporter. 2, 45-48.
FORTUNER, R. &. WONG. Y. (1984). R^viev of the genu. Hsl • cotylenchus Sieiner, 1945 !. A computer programme for 'c>ent:-fication of the species. Revue NematoL 7, 385-392.
COWER, J.C. (1971). • A general coefficient of similarity and some of its properties. Biometrics 27, 857-871.
LIMA, M.B. (1965). A numerical approach to the Xiphinema americaniun complex. 8th International Symposium. Society of European Nemaiologists, Antibes, Frances, September, 1965, 30.
DE MAN, J.G. (1880). Die einheimischen frei in der reinen Erde unci im sussenWasser lehende Nematodes. Tijdschr. ned. dierk. Ver. 5, 1-104.
MICHENER, CD. & SOKAL, R.R. (1957). A quantitative approach to a problem in classification. Evolution, 11, 130-162.
MOSS, W.W. & WEBSTER, W.A. (1970). Phenetics and numerical taxonomy applied to systematic nematology. J. NematoL 2, 16-25.
PINKHAM, C.F.L. & PEARSON, J.L. (1976). Applications of a new coefficient of similarity to population surveys. J. Water Poll Contr. Fe± 48, 717-723.
REHMAN, M.F., BILGRAMI, A.L. & JAIRAJPURI, M.S. (1986). Studies on variability in Xiphinema radicicola Gcodey, 193'. and X. brasiliense Lordello, 1951. Helminthologia 2X 179-lSc.
REY, J.M. (1987). Missing values in Cowers Index and its use in Nematode Identification. Proceedings of the 7th Congress of Mediterranean Phytopathology Union, Granada, Spain, September, 1987, 96-99.
'.I'-', J ^.., A ' T T r •'< ^ r.P.'-Z .. '1 method '"cr icevj.:f'y:nc '::mat:z ycedz-(Lcngidcridae) Rewe Nemidol, 1.. 129-i .^
REY, J.M. i MAH.\JAS, R. (198'^). A compucer method for i'den::-'Vjitj nemciGc: species 11. Genera Tylenchorhynchus c'-.d Mcrlmius (Senuitcda: T\lcncnidc). Revue NematoL 11, 2c~-

5 4 7
11 Z. I I iU. k, L/l_
ROGERS, D.J. & TANIMOTO, T.T. (1960). A computer programme for classifying plants. Science 132, 1115-1118.
SNEATH, P.H.A. & SOKAL, R.R. (1973). Numerical Taxonomy: San Francisco, Freeman &.Co. 573 p.
SOKAL, R.R. (1958). Quantification of systematic relationships and of phylogenetic trends. Proceedings of the Xth International Congress of Entomology 1, 409-415.
SOKAL, R.R. &. MICHESER, CD. (1958). A statistical method for evaluating systematic relationships. University of Kansas Science Bulletin 38, 1409-1433.
SOKAL. R.R. & Sy^EATH, P.H.A. (1963). Principles of Numerical taxonomy. Freeman &. Co. San Francisco &. London, 359 P-
ZANCADA, M.C. & LIMA, M.B. (1985). Numerical Taxonomy of the genera Rotylenchus Filipjev, 1936 and Orientylus Jairaj-puri &. Siddiqi, 1977 (Nematoda : Tylencnida). Nematologica 31, 44-61.

5^8
A scanning electron microscopic study on Dorylaimus stagnalis Dujardin, 1845
Samina SHAFQAT, Irfan AHMAD, Wasim AHMAD and Anwar L. BILGRAMI
Section of Nematology, Department of Zoology, Ahgarh Muslim University, Ahgarh - 202002, India
SLMMARY
Detailed observations have been made on Dorylaimus stagnalis with the scanning electron microscope The cuticle on the lip region is sculptured, lips are amalgamated and provided with the usual sixteen sensiUa The longitudinal ndges (bands) on the cuticle are made up of four to seven discrete cuticular folds The ndges which extend from the neck region to the anus/cloaca may frequently bifurcate or terminate at any point or form loops The entire body including the ridges is finely transversely striated In males, the posterior region has a midventral groove divisible into pre-, post- and supplement regions Four additional rows of papillae are present on each side of the groove
RESLME
Etude de Dorylaimus stagnalis Dujardin, 1845 en microscopie electronique a balayage
Des observations detaillees ont ete faites sur Dorylaimus stagnalis en microscopic electronique a balayage La cuticule de la region labiale est ornementee, les levres sont soudees et pourvues des seize sensilles normales Les cotes (bandes) longitudinales sont constituees par quatre a sept replis cuticulaires peu dcveloppes Les cotes, qui s'ctendent de la region cervicale a celle de I'anus — ou du cloaque —, peuvent frequemment etre bifurquees, se terminer a n'lmporte quel endroit du corps, ou former des boucles Chez le male, la region posteneure presente un siUon medioventral differencie en trois zones celle des supplements et celles situees antcneurement et posteneurement a ceux-ci Quatre rangees supplementaires de papilles sont presentes de chaque cote du siUon
In recent years SEM has provided much detailed informations on the surface structure of different nematode groups (Sher & Bell, 1975; Sher, Bell & Rodriguez, 1977). Studies by Jaira)puri and Southey (1984), Baldwin (1986), Geraert and Raski (1988), and Fortuner and Luc (1987), have highlighted the utility of SEM observations in elucidating concepts in taxonomy and phy-logeny
On dorylaims, except for a few studies on longidonds (Lamberti & Martelh, 1971, Lamberti, 1975, Hogger & Estey, 1976, Eveleigh & Allen, 1982; Swart & Heyns, 1987; Coomans & Carbonell, 1987, Vmciguerra & Coomans, 1988), SEM studies have remained largely neglected. Sauer (1985) and Coomans, Vmciguerra and Loof (1990) provided scanning electron micrographs of several dorylaim genera. Although the dorylaims in general offer relatively few surface structures for observations, we studied Dorylaimus stagnalis Dujardin, 1845 to charactense, in particular, the longitudinal ndges, transverse stnations and the caudal supplements and papillae of the males
Materials and methods
Freshly collected D stagnalis were fixed in 3 % glutaraldehyde solution for 90 min, washed in 0.05 M
sodium phosphate buffer several times then post-fixed in 2 " 0 osmium tctraoxide for 2 h at room temperature and finally washed again in buffer. The specimens were dehydrated in a six grade alcohol series and cntical point dned using carbon dioxide as the transitional fluid. Dried specimens were mounted on stubs using a double sided adhesive tape, coated with 30 nm layer of gold and examined with a Hitachi S 2300 scanning electron microscope at 15 and 25 kV Spicules were extracted and prepared as described by Eisenback (1986).
Results
L I P REGION
Oral aperture circular surrounded by six closely amalgamated, equal sized lips. Antenor sensilla arranged in a 6 -I- 4 + 6 pattern. Cephalic sensilla situated at a level between the inner and outer circlet. Each sensillum is distinctly elevated and provided with a pore. The outer and inner labial sensilla are more prominent than the cephalic ones. Cuticle on the lip region sculptured to from four to six broken concentnc circumoral nngs and a variable number of nngs on each lip surrounding the papillae (Fig. 1 A, C).
Revue Nematol 14 14) 511-515 11991) 511

^ '4 Q ^ '' '•'S. Shafqat, I Ahmad, W Ahmad & A L Bilgrami
Fig. 1. Dorylaimus stagnahs Du)ardin, 1845 — Female — A, C : Anterior ends; B : Protruded odontostyle (dorso-ventral); D : Ridges at midbody; E : Ridge formation in anterior region; F, G : Ridges and transverse stnations; H : Bifurcating ridges; I : Ridges forming loops; J : Vulval region; K : Female postenor region. (Bars = 5 jim in B, C, 25 ,um in K.)
ODONTOSTYLE
The odontostyle is hollow, cylindroid with a smoothly rounded tip (Fig. 1 A, B). The aperture is about 13 \iva. long and continues as a very narrow slit for some distance as the sleeves of the spear apparently fail to meet immediately below the aperture (Fig. 1 B).
AMPHIDS
Amphidial apertures slit-like about 5 |j,m across, situated at the junction of the lip region with the body.
CUTICLE
Cuticle marked with fine transverse striations and longitudinal ridges. Transverse striations present over the entire body. The longitudinal ridges start at the neck region and gradually fade out slightly anterior to the
level of anus/cloaca. Each ridge begins to form by irregular and uneven elevations of the cuticle (Fig. 1 E), gradually these elevations become uniform and grouped into longitudinal bundles forming the longitudinal ridges (Fig. 1 D). At the midbody, each ridge is about 2 |i,m wide and appears like a band consisting of four to seven small and discrete cuticular folds (Fig. 1 F, G). The folds forming the ridges are of variable sizes. Except for some of the outer folds, which may occasionally be relatively straight, each follows a slightly wavy pattern almost parallel to the adjacent one. The folds of the ridges may terminate abruptly or bifurcate (Fig. 1 F, G). A longitudinal ridge may not always traverse the entire length of the body but may terminate at any point. On the other hand, a ridge may also bifurcate (Fig. 1 H). Occasionally ridges bifurcate and then reunite forming loops (Fig. 1 I). Not infrequently, very small aberrant ridges may be formed at any part of the body (Fig. 2 A).
512 Revue Nematol. 14 (4) : 511-515 (1991)

SEM study on Dorylaimus stagnalis
Fig. 2. Dorylaimus stagnalis Dujardin, 1845 — A : Pre-supplement region (showing termination of ridges and one small aberrant ndge); B : Ventromedian supplements and subventral papillae; C . Postenor region showing cloacal opening and papillae; D : Single lateral papilla; E : Posterior region showing post-supplement groove and four rows of papillae; F : Spicule; G, H, I : Ventromedian supplements. tBars = 20 um in A, B, C, E, F.)
In males, the ridges begin to narrow from a region slightly anterior to the beginning of the supplements (Fig. 2 A, B).
Transverse striations are very fine, about 0.5 i m apart at midbody and pass through the ridges (Fig. 1 G). The transverse stnations generally span the distance between the ridges but may, occasionally, terminate in between. As the longitudinal ridges are elevated regions, the transverse striations generally form a shallow concavity between them (Fig. 1 D, E, F). At low magnifications the transversely striated body appears to have alternating light and dark bands (Fig. 1 D). At high magnifications the light bands themselves appear like two light bands sandwiching a dark band... the striae (Fig. 1 F, G). The margins of the interstriae region adjacent to the striae are slightly raised.
VULVA AND ANUS
The vulva is small, circular, generally less than 5 |im in diameter, located between two longitudinal ridges (Fig. 1 J). Some amount of shrinkage was always observed in the vulval region. The anal opening is a crescent-shaped slit measuring 12-13 ^m across (Fig. 1 K).
MALE POSTERIOR REGION
On the posterior region of males are present the ventromedian supplements and an assortment of papillae. The ventral longitudinal ridges terminate well before the beginning of the supplements (Fig. 2 A) and extending from this region to the cloaca is a midventral groove
Revue Nematol 14 14) 511-515 (1991) 513

551 S Shafqat, I Ahmad, W Ahmad & A L Bilgrami
that IS clearly divisible into a pre- supplement, supplement and a post-supplement region. The pre-sup-plement groove (Fig. 2 A) is slightly more than 5 ^m long and trench like The margins are attenuated forming distinct edges which merge with the body anteriorly and posteriorly (Fig. 2 A, G) The ventrome-dian supplements which begin from just beyond the pre-supplement region are situated on a ndge within the supplement groove (Fig. 2 G, H, I). Each supplement is prominent with a centrally located papilla (Fig. 2 G, H). The papilla has a small depression but whether it is a pore or not could not be ascertained Towards the posterior region the supplements are slightly smaller (Fig. 2 B) and the last two or three become progressively degenerate (Fig 2 I) From about the region of the last few supplements, the margins of the groove again form an edge and continue into the post-supplement region (Fig. 2 E, I). Beyond the ventromedian supplements the groove continues postenorly, gradually widens and merges with the body contour just antenor to the cloaca as also do the outer edges of the groove (Fig. 2 C, E).
CLOACAL OPENING
A C-shaped slit with thin antenor lip and a thick cushion-like postenor lip (Fig. 2 C).
ADANAL PAPILLAE
These papillae are paired, extremely small and about 3 |im apart (Fig 2 C).
O T H E R PAPILLAE
In addition to the supplements, the male posterior region bears at least four rows of comparatively smaller papillae on each side. The subventral begin from antenor to the pre-supplement groove and continue up to the cloacal opening, terminating below it in a disorganized manner (Fig. 2 C, E). Of the three other rows on each side, one is lateral, one ventrosublateral and one dorsosublateral. The laterals are a consistant feature found along the entire body length, although widely spaced anterior to the supplements. In the postenor region they are more closely arranged (Fig 2 E). The ventrosublaterals and the dorsosublaterals anse at about the region where the supplements begin and continue postenorly, being more or less regularly arranged, towards the tail tip. The tail terminus presents a disorganized aggregation of papillae as three rows from the two sides of the body converge towards it (Fig. 2 C, E). Each papilla is button shaped, slightly more than 1 |j,m in diameter and with a distinct opening (Fig. 2 D)
Discuss ion
Although few studies are available for comparison, certain features of D. stagnahs appear to be quite in
teresting. What looks like ndges in the light microscope are in effect longitudinal bands made up of discontinuous longitudinal cuticular folds. Because of the nature of the longitudinal bands (ndges) their use as a qualitative character should be judicious taking into account its extreme vanability However, provided information is available, qualitative aspects of these bands may provide a more reliable character. Further studies are needed to reveal the nature of longitudinal ndges (bands) in Dory-laimus species as well as in other dorylaims. The longitudinal ndges of Etamphidelus acucephalus and E fueguensis are smooth and made up of a single cuticular fold, being irregularly, obliquely or transversely stnated in the former species (Coomans & Raski, 1988). The sculptunng of the cuticle on lip region is also found (as seen m SEM photomicrographs) in Eudorylaimus sp., and Aporcelaimellus sp. (Sauer, 1985). The odontostyle though typically dorylaimoid, is different from that of Aporcelaimellus (Lippens et al, 1974) in that the sleeves do not appear to overlap and the aperture continues as a narrow slit for about 3/4 of its length.
The caudal region of males seems to be well adapted for a sensory and copulatory role. The ventromedian supplements are numerous and although situated in a groove, they may be fully exposed once the copulatory muscles contract pulling outwards laterally the margins of the groove. In the same context it may be tempting to suggest that when the edges of the pre- and post-supplement regions are pulled outwards they could act as grasping surfaces, performing a function somewhat analogous to the caudal alae. The numerous papillae (laterals, ventrosublaterals, dorsosublaterals and subven-trals), sometimes partly referred to as body pores, are apparently glandular sensory structures of the somatic sensory system and may play a role dunng reproductive activities.
Ac KNOWLbDGFMENlS
This research work was supported by the grants from the Indian Council of Agricultural Research, New Delhi, and the Departmental Research Support Programme of the University Grants Commission, New Delhi.
REFERENCFS
BALDWIN, J. G. (1986). Testing h>pothesis of phylogeny of Heterodenderae In Lamberti & Taylor, C E (Eds) Cyst Nematodes New York, Plenum Publishing Co 75-100
CooMANS, A & CJIRBONELL, E (1987) The status of the family Thomenematidae Siddiqi, 1969 (Nematode Dory-laimida) Nematologica, 33 375-386
CooMANS, A & RASKI, D J (1988) Three new species of Etamphidelus Andrassy, 1977 (Nemata Alaimidae) in Southern Chile Journal of Nematology, 20 9-22
COOMANS, A, ViNC ICUERRA, M - T & LOOE, P A A (1990) Status of the genera Paractinolaimus Meyl, 1957, Trachy-
514 Revue Nematol 14 (4) 511-515 11991}

SEM study on Dorylaimus stagnalis R r. 9 J -J (»
pleurosum Andrassy, 1959 and Trachactinolaimus Andrassy, 1963 (Nematoda : Actinolaimidae) with description of Tra-chypkurosum venezolanum n. sp. Revue de Nematologie, 13 . 143-154.
EiSbNB'vrK, J. D. (1985). Techniques for prepanng nematodes for scanning electron microscopy. In : Barker, K. R., Carter, C. C. & Sasser, J. N. (Eds). An advanced Treatise on Meloidogyne, Vol. II Raleigh, Dept. PI. Pathol. & USAID : 79-105.
EvbLEiGH, E. S. & Ai I EN, W R. (1982). Description of Lon-gidorus diadecturus n. sp. (Nematoda : Longidondae), a vector of the peach rosette mosaic virus in peach orchards in South Western Ontano, Canada. Canadian Journal of Zoology, 60 : 112-115.
FoRiLNER, R. & Luc, M. (1987). A reappraisal of Tylenchma (Nemata). 5. The family Belonolaimidae Whitehead, 1960. Revue de Nematologie, 10 . 183-202.
GFR,AERI, E & R^SKi, D. J. (1987). Study of some Aglenchus and Coslenchus species. Nematologica, 34 : 6-46.
HoGGER, C. H. & EsiEY, R. H. (1976). Scanning electron microscopy of a plant parasitic nematode : Xtphinema amencanum Phytoprotectwn, 57 : 150-154.
JAIRAJPLRI, M . S. & SoLTHEY, J. F. (1984). Nothocnconema shepherdae n. sp. with observations on extra-cuticular formation. Revue de Nematologie, 7 : 73-79.
LAMBERII, F . (1975). Taxonomy of Longidorus (Micoletzky)
Filipjev and Paralongidorus Siddiqi, Hooper & Khan. In : Lamberti, F., Taylor, C. E. & Seinhorst, J. W. (Eds). Nematodes Vectors of Plant Viruses New York, Plenum Publishing Co. : 71-79.
LAMBERII, F . & MARIELLI, G . P. (1971). Notes on Xiphmema meduerraneum (Nematoda : Longidondae). Nematologica, 17 : 75-81.
Lii'i'ENS, P. L., CooM^NS, A., D F GRISSE, A. T . & LAGASSF, A. (1974). Ultrastructure of the antenor body region in Aporce-laimellus ohtusicaudatus and Aporcelaimus obscurus Nematologica, 20 : 242-256.
SAUER, M . R. (1985). A scanning electron microscope study of plant and soil nematodes. CSIRO, AustraUa, 64 p.
SHER, S. A. & BELI, A. H . (1975). Scanning electron micrographs of the anterior region of some species of Tylenchoi-dea (Tylenchida : Nematoda). Journal of Nematology, 1 : 69-83.
SHER, S. A., BELL, A. H. & RODRIGLEZ, R. (1977). The face view of Tnchodoridae. Journal of Nematology, 9 : 254-257.
SWAR r, A. & HEYNS, J. (1987). Comparative morphology of the head regions of some longidond nematodes from South Afnca using SEM. Phytophylactica, 19 : 99-101.
VlK( IGUERRA, M. T. & CoOMANS, A. (1988). Parastomacho-glossa perplexa (Heyns et Argo, 1969) n. comb, with a definition of the genus. Nematologia Mediterranea, 16 : 205-212.
Accepte pour publication le 28 aout 1990.
Revue Nematol. 14 (4) : 511-515 11991) 515

553 Nematologica 38 (1992) 296-303, E J Brill, Leiden
A SCANNING ELECTRON MICROSCOPE STUDY ON MONONCHOIDES FORTIDENS (SCHUURMANS-STEKHOVEN, 1951)
TAYLOR & HECHLER, 1966
BY
QUDSIA TAHSEEN, IRFAN AHMAD, ANWAR L BILGRAMI and WASIM AHMAD
Section of NematologN, Department of Zoology, Aligarh Muslim University, Aligarh-202002, India
Detailed SEM observations were made on Momnchoidesfortidens (Schuurmans Stekhoven, 1951) Tavlor & Hechler 1966 The cuticle is marked with fine transverse stnations and prominent longitudinal ridges or folds The lip region is continuous and the lips are not prominent Each lip bears a single papilla but in males four additional setose cephalic papillae are present The stomal opening is slit-like, bordered by 13-15 bifid cheilorhabdial filaments Amphids are small and elliptical and situated post-labially The stoma has a prominent dorsal tooth with an opening of the oesophageal gland Other structures in the stoma include a subventrai pyramidal tooth, ventro-lateral toothlets or denticles, a dorsal plate and paired dorso-lateral plates The cheilorhab-dions are apparently not fused but rib-hke, appearing as bifid structures in face view The vulva is circular, situated between two longitudinal ridges The male posterior region has nine pairs of papillae of which three are pre-cloacal and six post-cloacal Phasmids are large pore-like Keywords SEM morphology, stoma, ridges, caudal papilla, Mononchoides fortidens
Scanning electron microscopic studies have provided valuable information on the surface structures of several nematode groups (Sher & Bell, 1975; Sher et ai, 1977). Until now, the relatively few SEM studies made on the diplogasterids include those of Gibhn & Kaya (1984) on Aduncospiculum halicti, Ghitambar & Noffsinger (1989) on Odontopharynx longicaudata, Geraert et al. (1989) on Pris-tionchus Ihentien and Ghitambar (1990) on Monobutlenus macrogubernaculum. This study deals with SEM observations on the diplogasterid predator, Mononchoides
fortidens (Schuurmans Stekhoven, 1951) Taylor & Hechler, 1966. The study includes detailed observations on the cuticular structures, lip region, stoma and the caudal papillae.
MATERIALS AND METHODS
Soil samples contammg M. fortidens collected from Burdwan, West Bengal, India, were processed by sieving and decanting and the modified Baermann funnel technique. To maintain a population of the species, the nematodes were cultured m 1% water agar on a bacterial diet. For SEM, freshly isolated specimens were fixed in 3% glutaraldehyde for 2 h, washed in sodium phosphate buffer several times then post-fixed in 2% osmium tetroxide for 2 h at room temperature and finally washed in buffer. The specimens were then dehydrated in a six-grade acetone series and critical point dried in carbon

554 MONONCHOIDES FORTIDENS 2 9 7
dioxide. Dried specimens were mounted on stubs, coated with 30 nm gold and observed in a Hitachi S-2300 SEM at 15 KV.
RESULTS
Cuticle: The cuticle is marked with fine transverse striations and prominent longitudinal ridges which have the appearance of longitudinal folds (Fig. 1 G). The crest of the ridges are marked by very fme longitudinal striations (Fig. 1 H). The longitudinal ridges, 24-26 in number, arise from the base of the lip region (Fig. 1 E, F) and traverse the entire body length. In some females, however, additional short cuticular ridges, eight in number, terminating about two lip region widths from the anterior end may also be seen (Fig. 1 F). In males the ridges terminate about two anal body widths behind the cloaca and beyond this point the tail continues as a long whip-like structure (Fig. 3 C, E). In females these ridges continue onto the narrow filamentous part of the tail (Fig. 3 G). Longitudinal ridges do not bifurcate or terminate abruptly along most of the body length and therefore their number tends to remain constant. Some ridges, particularly those on the ventral and subventral sides, terminate slightly anterior or posterior to the anus/cloaca (Fig. 3 B, E). The transverse striations are 0.5-0.75 |im apart at midbody and pass over the longitudinal ridges (Fig. 1 G, H). The excretory pore is obliquely located between two ridges (Fig. 3 A). Lip region: The labial region is continuous with the body contour and the lips are discernable only when relaxed (Fig. 1 A). In en face view the labial region shows a deep dorso-ventral cleft which divides the lip region into two lateral sectors (Fig. 1 A). Laterally in the region of amphids there are two shallower crescentic indentations. Lip dimorphism is apparent with the laterals being distinctly grooved and consequently their papillae are placed aborally at the end of the groove, while the non-grooved submedians have their papillae placed orally (Fig. 1 A). Only six labial papillae are visible in females while in males four additional setose cephalic papillae are present at the level of the amphids (Fig. 1 A, C, E). The oral aperture is slit-like when relaxed, bordered by 13-15 processes of cheilorhabdions (rugae as described by Jairajpuri et al. (1973)) arranged in two rows along the dorso-ventral axis. Each cheilorhabdial filament consists of two closely placed finger-like processes, free apically (Fig. 1 A, B). During feeding, as the lips and cheilorhabdions are pulled outwards and posteriorly, the margins of the denticulate stoma and the stomal armature are exposed (Fig. 1 B).
Stoma: The stoma is well armed for a predatory mode of life. A massive tooth is present on the dorsal metarhabdion. The duct of the oesophageal gland opens subapically on to the ventral side of the tooth (Fig 1 B; 2 A, C). Lateral to the dorsal tooth on the right side is a large pyramidal tooth tilted towards the apex of the dorsal tooth (Fig. 1 B, D; 2 A, B, C). Anterior to the dorsal tooth and forming a canopy over it, is a dorsal plate, the free edge of which is denticulate (Fig. 2 A-C). At the lateral margins of this plate, and on the dorso-lateral aspect

l 'S'5 298 TAHSEEN, A H M A D , BILGRAMI & A H M A D
Fig 1 Mononchoidesfortidens A fn/aw view (female) B Open stoma showing dorsal and pvramidal tooth, C En face wtw (male) D Open stoma showing dorsal and pvramidal tooth and triradiate lumen, E interior region male, F Anterior region female, G & H Stnations and ridges (Scale
Bar = 2 (jm in A-D H 5 urn in F G 10 |um in E)

51 — . - >
MONONCHOIDES FORTIDENS 299
Fig. 2. Everted stoma oiM. forhdens showing internal structures. A. End-on view; B. & C:. oblique view. (Scale: Bar = 5 |iml
of the stoma, are two smaller plates (Fig. 2 A, C). Opposite the dorsal tooth are two rows of denticles (toothlets) forming a semi-circle ventro-laterally (Fig. 2 B). Where the double row ends are a few slightly larger teeth somewhat lateral in position (Fig. 2 A). The pro-mesorhabdions are fused and the edge is denticulate (Fig. 2 A, B, C). At the base of the stoma is a triradial opening which leads into the lumen of the oesophagus (Fig. 1 D). Vulva and anus: The vulval opening is circular, located between two longitudinal folds (Fig. 3 D). Because of the location it sometimes appears to be somewhat longitudinally ovoid. The ridges in the region of the vulva project outwards (laterally) and those adjacent to it may be shghtly thickened (Fig. 3 D). The anal opening is transverse and oval in shape. Some of the ventral/subventral longitudinal ridges terminate in this region (Fig. 3 B). Tail: The tail of both sexes is long and filiform and can be distinguished in two parts—an anterior rapidly tapering region which is about 1.5-2.0 anal body widths long, and a posterior whip-like part which is 50% or more of the body length. In males the longitudinal ridges terminate from where the whip-like posterior part commences, while in females the ridges continue posteriorly onto the whip-like part of the tail (Fig. 3 C, E, G) and gradually disappear. The phasmids are large, circular pore-hke structures (Fig. 3 E, F). Male genital papillae: In the posterior region of males nine pairs of genital papillae are present (Fig. 3 F). Three pairs are pre-cloacal and six post-cloacal. The last post-cloacal pair is dorso-lateral (Fig. 3 C) and the others are placed variably from nearly lateral to subventral and those at the end of the tapering part of the tail nearly ventromedian (Fig. 3 E, F). Each pair of papillae occurs at the same level on both the sides of the body except the third, fourth and fifth post-cloacals which are slightly obHquely placed (Fig. 3 E). The fourth pair is

£S7
3 0 0 TAHSEEN, AHMAD, BILGRAMI & AHMAD
interesting in tiiat it is smaller and bifid (Fig. 3 E). Each papilla arises from a distinct socket.
DISCUSSION
The most salient morphological features of M. fortidens are the longitudinal ridges which arise as outfoldings of the cuticle. Through most of the body length the ridges show no variation and hence their number is considered to be a reliable character. The ridges are not marked by prominent longitudinal striations and hence the transverse striations that cut across them do not create a blocked pattern as is visible in Aduncospiculum halicti (Giblin & Kaya, 1984) and Pristionchus Iheritieri (Geraert et ai, 1989). The short cuticular folds restricted to the anterior region, which have also been reported in AI. changi (Goodrich et ai, 1968) apparently do not represent any special feature, as variations of the cuticular patterns in the anterior region are known in other diplogasterids. Ridges similar to those o[ M. fortidens have been seen in A. halicti (Giblin & Kaya, 1984) while in Monobutlerius macrogubernaculum (Chitambar, 1990) the longitudinal and transverse striations are present only in the anterior region. What may be important is that the additional cuticular folds in the anterior region of M. fortidens is an exclusive though not a permanent feature of females only.
There appears to be quite a lot of variability in the number and arrangement of the anterior sensilla in diplogasterids. Six labial papillae occur in A. halicti (Giblin & Kaya, 1984), while in P. Iheritieri, females have two circlets of six labial and four cephalic sensilla (Geraert et ai, 1989). Although the number of head papillae of M. fortidens is similar to that of M. macrogubernaculum where females have six labial papillae and the males four additional cephalic setae (Chitambar, 1990), differences nevertheless still exist. While all papillae are setose in M. macrogubernaculum, only the four cephalic papillae present in males oiM. fortidens are setose. Further, the labial papillae are symmetrically arranged in the former while in the latter, the papillae on the submedian lips are located at the oral end and those on the lateral lips occur aborally at the end of a furrow that bisects these lips.
The opening of the dorsal oesophageal gland on the ventral aspect of the dorsal tooth in M. fortidens confirms the earlier observations of Jairajpuri et ai (1973) on Tridontus longicaudatus (= M. longicaudatus). Calaway & Tarjan (1973) had noted with precision the spatial relationship between the tips of the dorsal and the right subventral (pyramidal) tooth as well as the presence of toothlets (denticles) on the latero-ventral metarhabdions. Although the tips of the two large teeth are close to one another, they point in opposite directions. This arrangement coupled with the posterio-anterior movement of the dorsal tooth could make the non-moveable pyramidal tooth an effective cutting surface. Also, the two undescribed structures viz., the denticulate dorsal plate and the smaller latero-dorsal plates may provide an additional means of gripping/

5:8 MONONCHOIDES FORTIDENS 301
Fig 3 Monomhoides forlidens A Excretor> pore, B \na l opening, C Male posterior region, D Vulva, E Male post-cloacal region (ventral;, F Male posterior region (lateral), G Female poste
rior region (Scale Bar = 5 Jim m A-E, G, 10 |am in F)

I-' r, 9 J -J V
3 0 2 TAHSEEN, AHMAD, BILGRAMI & AHMAD
puncturing the prey cuticle in tlie stoma. Regarding the cheilorhabdionb, we are inclined to concur with the observations of Calaway & Tarjan 1973' that the bifurcated tip filaments may actualh represent this structure
The uncharacteristic nature of the fourth pair of post-cloacal papillae and the slightly ob ique arrangement of the third, fourth and fifth pairs could possibly cause some confusion in ascertaining their exact number in light microscopy.
This research was supported b) a grant from the Departmental Research Support Programme of the UGC and a fellowship of the CSIR to the first author.
RESl ME
/s(«f/f rff Mononc loides fortidens fSihuurmam Stekhoien, 1951) laylor & Hethler, 1966 en muroscopie eleclromque a balaya^e
Des observation' detdillets au MLB out ete fanes Mir MonomhoidesJortiden^ Sthuurmans Stek-ho\en, 1951 Tavlor & Hechler 1966 I a cuticule e t marquee par de fines stries transversales et des tretes ou replis longitudmaux tres en relief La region labiale tsl continue d\et le reste du corps et Its levres ne fort pas saillie Chaquc le\re porte uiie seule papille, mats thez le mak quatrc papilles eephaliqu( s setacets sont presentes en supplement L'ou\eiture buccale en fcntc est bordee par 13-15 filaments bifides appartenant au cheilorhabdion Les amphides petites et elliiHiques sont situces post-labialement La ca\itc buccale compoite une dent doisale importante av ec debouc he de 1 i glande oesophagienne Les autrcs structures dc la ca\ ite buc cale comprennent une dent subventrale pyramidale, de petites dents ou denticules \entro-laterales, une plaque dorsale et une pai e de plaques dorso-lateraies Les t heilorhabdions ne sont appareminent pas fusionnes mats en forme de crete. apparaissant comme une structure bifidc en \ue de face La vuKe, circulaire, est situee entre deux cotes longitudinales La partie posteiicure du male com-portc neuf paires de papilles trois prec loacales, six postcloacales Les phasmides apparaissent comme des ports dssez larges
RLFERENCLS
CALAVVA'I,\\ T & TARJAN A C 1973 A compendium of tlie genus .l/ono/KAuJc/ci Rahm 1928 Diplogasterinae \ ema toda Journal of \ematolo^} 5. 107-116
CHIT^MBAR, J J 1990 Description of a new predaceous nematode, Monobutknus macro^uber naculum n sp Nemata Diplogasttridae Reiue de \ematolo^u 13, 369-373
CHIT wiBAR, J J & NOFFSI\GER, E M 1989 Taxoiiom\ and the postembr\onic stages ol the nematode predator 0(/on(o/i/iannx/on^!(aM(/a(a de Man 1912 Uiplogasteiida Journal uj \ema tology 21 , 189-201
Gi'RAFRT, E , MRACEK, Z & SuDHAi b, \V 1989 The facultative insect parasite Pnstionchui Ihentien s}n P pseudolkeritieri ' \ematoda Diplogastenda \ematologica 35 133-141
GiBi i\, R M & K A \ A , H K 1984 Aduncospuuhim haluti n gen , n sp Diplogastenda Diplogastc roididae , an associate of bees m the genus Halutus Hvmciioptera Halutidae Revue de \ema'otogie 7, 189-197
JAIRAJPURI, M S , BA^RI, Q H & "VKMAD, S 197 5 .\ematodes of high altitudes in India II Studies on the genus Tndonlus Khera, 1965 with notes on the s\non\iTi\ ol the genus S)edella Sur\avvanshi 1971 Diplogasteridae \ematologua 19 69-74
Sent LRMANS Sii-KnovtN, J H 1951 Nematodes saprozoaircs ct librc s du Congo Beige \limoire de rimlttul Royale dei Sitencei \alurellei de Belgique 39, 5-79
SHFR, h \ &. Bn , A H 1975 Scanning electron micrographs ol the antcrioi region of some species of l}ienchoidea Fvlenchida Nematoda Journal if \em(iiolog) 1 69-83

5 GO MONONCHOIDES FORTIDENS 3 0 3
SHLR, S A BELL, A H & RODRIGIE? , R 1977 I he late view oi I ru hodondae Journal oj \eniatolog) 9, 2o4-2')7
IAYIOR, D P & HttHiER, H C 19bb Monotnhuidc Joitidens n comb and the tiansltr of Mononihoides ob^iuridem to the s^tnu^ Diplue^aslei \emaliiliigica 12, -545
\iceptedJoi publuttliun iBJantiarv l'-!92

561
LIST DF PUBLICATIONS
1. BILGRAMI, A. L., AHflAD, I. & JAIRAJPURI, M. B.
(1983). Some factors influencing predation by
Mononchus aquaticus. Revue Nematol., 6: 325-
326.
2. BILGRAMI, A. L., AHMAD, I. ?< JAIRAJPURI,
M.S.<19S4>. Observations on the predatory
bebaviour of Mononchus aquaticus. Nematol.
Medit., 12 s 41-45-
3. BILGRAMI, A. L- 8< JAIRAJPURI, M. S. (1985). The
responses of Mononchus aquaticus to chemicals
and pH. Indian J. Nematol., 14 (1984): 171-
174.
4- BILGRAMI, A. L. %< JAIRAJPURI, M- S. (19B5>.
Cannibalism in Mononchus aquaticus. Indian J.
Nematol. 14 U984):202-2B3.
5. BILGRAMI, A. L., AHMAD, I. ?< JAIRAJPURI, M. S.
(1985)- Factors influencing attraction of
adult Hirscbmanniel 1 a oryzae towards cabbage
seedlings. Revue Nematol., 8:67-75.
6. BILGRAMI, A. L., AHMAD, I. S< JAIRAJPURI, M, S.
(1985). Responses of adult Hirschmanniei1 a
Dryzae towards different plant roots. Revue
Nematol., 8 : 265-272.

5^2
1. BILGRAMI, A. L-, AHMAD, I. S< JAIRAJPURI, H. S.
(1985)- Predatory behaviour of Aquatides
tbornei iNygoiaimina i Kematoda),Nematologica,
<;i984>! 30 : 457-462.
8. BILGRAMI, A. L,, AHHAD, I . ?< JAIRAJPURI, M. S.
<].9S6>. Attraction and feeding behaviour o-f
adult Hirschmanniel1 a oryzae. Indian J.
Nematol., 15 < 1 985> : 21259-21 6.
9. BILGRAMI, A. L., AHMAD, I. & JAIRAJPURI, M. S.
{1936). Ef-fect D-f Boroe chemicals on the
activity o-f Hi rschmanni el 1 a oryzae,
Helminthologia, 23 : 49-54.
10. BILGRAMI, A. L., AHMAD, I. ?< JAIRAJPURI, M. S.
(1996). ResponsBB o-f Hi r schmanni el 1 a oryzae
juveniles towards cabbage seedlings.
Helrointhologia, 23 : 119-122-
11. BILGRAMI, A. L., AHMAD, I. & JAIRAJPURI, M. S.
(1986). A study on the intestinal contents of
some mononchs. Revue Neroatol., 9 s 191—194.
12. BILGRAMI, A. L., JAIRAJPURI, M. S. & AHMAD,I-
(1986). Ef-fect of different plants, singly and
in combinations on the attraction of Xiphinema
basiri. Indian J. nematol-, 16 : 60-64.

533
13. RAHMAN, M. F., BILGRAMI, A. L. Sc JAIRAJPURI, M.
S., (19S6>. Study of variability in Xiphinema
radicicDla Goodey snd X- brasiliense Lordello,
1951- Helminthologia, 23 : 179-1B8.
14. BILGRAMI, A. L., ALAM, S. M. ?< AHMAD, I. (1987).
Histochemical localization of some enzymes of
the carbohydrate metabolism in a soil
nematode Dorylaimus stagnalis. Nematologica
(1986). 32 : 239-243.
15. BHAFDAT, S., BILGRAMI, A. L. ^ JAIRAJPURI, M. B.
(1987). Evaluation of the predatory behaviour
of Dorylairous. s-tagnalis <Ne««atodaj
Dorylaimi da). Revue Nematol. , IB: 455-461.
ih. BILGRAMI, A. L. & JAIRAJPURI, M. B. n 9 8 3 ) .
Attraction of Mononchoides 1ongicaudatus and
M. fortidens (Nematode s Dipiogasterida)
towards prey and factors influencing
attraction. Revue Nematol., 11 J 195-202.
17. BILGRAMI, A. L., AHMAD, I. ?< JAIRAJPURI, M. S.
(1988). Interaction between mononchs in non-
stefrile agar plates. Indian J. Nematol., 17
<19S7). 339-340.

D vj 4
13. AHMAD, W. , KHAN, T. H. & BILGRAMI, A-L. <19QS>.
PI arit parasitic nematodes associated with
paddy crops in Bihar, India. Inter. Nematol-
News Letter, 5 : 4 .
19. PARVEEM, R. , BILGRAMI , A. L. ?< JAIRAJPURI, D. S.
(1989). Observations on the osmoregulation
in an insect nematode Schewenchiel1 a atheri
(Thaiastomatidae j Oxyuroidea). Indian J.
NematDl-, (1988), IS : 218-212.
20- BILGRAMI, A. L. & JAIRAJPURI, «. S. (1989).
Resistance of prey to predation and strike
rate o-f the predators Mononchoides
1 ongi caudatuB and M. -fortidens (Nematodai
Dipiogasterida). Revue Neroatol. , 12 : 45-49.
21. BILGRAMI, A. L. ^ JAIRAJPURI, H. S. (1989).
E-f + ects of concentrations and thickness of
agar on the predatory behaviour of
Morionchoi des 1 ongi caudat us and M- fortidens
(Nematoda:Dipiogasterida) snd on their
attraction towards prey. Indian J. Ne/natol . ,
(1988),18 J 222-227.
22. BILGRAMI, A. L. ?. JAIRAJPURI, M. S. (1989).
Aggregation of Mononchoides 1ongicaudatus and
M. fortidens (Nematoda : Dip1ogasterida) at
feeding sites. Nematologica (1988),34:119-121.

23. BILGF^AMI, A. L., AHMAD, I. ^ JAIRAJPURI, H- S.
(1939), RespDnseB of adult and juvenile
Hirschmanniel 1 a oryzae towards dirferent
sizes D-f the source of attraction. Indian J.
Nematol- 18 (19SS> j 357-360.
24. BILSRAMI, A. L. U989>. Specialised Techniques in
Nematode Taxonomy. In Nematode Pest
Ident i fi cati on, (Ed. Jeirajpuri, M- S. >.,pp
155-168.
25. BILGRAMI, A. L. <1990>. Dipiogasterid predators.
In Nematode Bio—control; Aspects and Prospects
(Eds. Jsirajpuri, Alam Sc Ahmad),CBS Publishers
and Distributors Pvt.,Ltd., New Delhi, pp 135-
142.
26. BILGRAMI, A. L. (1993). Predatory Dorylairos. In
Nematode Bio-Control;Aspects and Prospects
(Ecis. Jairajpuri, Alam §< Ahmad) CBS Publishers
and Distributors Pvt., Ltd., New Delhi pp
143-148.
27. JAIRAJPURI,M. B., BILGRAMI, A. L. (1990).
Predatory nematodes. In Nematode Bi o—control
1_ Aspects and Prospects (Eds. Jairajpuri ,
Alam 8( Ahmad) CBS Publishers and Distributors
Pvt-, Ltd., t4pw Delhi pp 95-125.

b u 0
28. BILGRAMI, A. L. S< JAIRAJPURI, K. B. (1990).
Predatory abilitieE o-f MononchcideB longic-
at'datus end M. •fortidens (Nematcde :
Di pi ogest eri da > and -factors influericing
predation. Nematologica, 35 (1989): 475-488,
29. BILGRAMI, A. L. ?< AMHAD, W. <1990)- Plant
parasitic nematodes associsted with paddy
crops in Uttarpradesh, India. Int. Nematol.
Nf?w5 Letter. 7: pp 25.
30. SHAF9AT, S. ,JAIRAJPURI , M. S, ?< BILGRAMI, A.
L. <}991>. Developmental biology o-f Dorylairous
stagnalis Dujardin 1845 (Neinatoda :
Dorylaimi da) . Revue Nematol., 14 : 61-71.
31. SHAFDAT, S., AHMAD, I., AHMAD, W. & BILGRAMI, A.
L. (1991). A scanning electron microBCop study
on Dorylaimus stagnalis Dujardin, 1845
(Nematoda ; Dor yl ai foi da > . Revue Nematol. 14 :
511-515.
32. AHMAD, W., AHMAD, I., BILGRAMI, A.L. 5. KHAN, Z.
(1991)-Thaiassogenus shamimi n.sp-(Nematoda:
Thai assogener i dae) , a nematode predator -from
India. Fundamental S< Applied Nematology. 15:
127-131 .

33. KHAW, Z-, BILGRAMI, A. L. ?f JAIRAJPURI, M. S.
<1991>. Some studies on the predation
abilities o ^ Aporcel ai mel 1 us. nivalis
(Nematoda: Dorylaimi da>, Nematologica. 37
34. BILGRAHI, A. L. <1992). Resistance end
susceptibility of prey ne/natodes to predation
and stri ?:B rate of the predators, Mononchus
aqusticus, Dorylaimus stagnalis and Aquatides
thornei. Fundamental & Applied Nemaiology. 15
: 265-270.
35. BILGRAMI, A. L. <1992>. Biocontroi agents and
thE»ir role in nematode management, In Recent
Advances in Nematology iEd. B. K. Dwivedi>
Pub. Bioved Research Society, Allahabad,
203-218.
36. TAHSEEH, Q., AHMAD, I., BILGRAMI , A. L. S< AHMAD,
W- <1992>- SEM studies on Mononchoides
•fortidens (Nematoda : Di pi ogast er i da) .
Neroatologica 38 s 296-303.
37. BILGRAMI, A. L. ?c TAHSEEN, Q. <1992>- A nematode
Feeding mite, Tyrophagus putrescentiae
< Sarcopt i-f ormi s : Acaridae). Fundamental ?<
Applied Nematology, 15: 477-478.

5 u 8
3S. SHAFDAT, S., BIL8RAKI, A.L. ^ JAIRAJPURI, H.
5.(1993). Effect of chemicals on the activity and survival of Dorylaifnus stganalis
Dujardin, 1845 and their juvenile stages-
Indian J. Nematol- 22 <1992) : 19-24-
39- BILGRAMI, A. L. <1993>. AnalyseB of
relationships between predation by
Aporcelaimel1UB nivalis and different prey
trophic categories. Nematologica - 39 i 356-
40. BILGRAMI, A. L- <1994>. Predation abilities of
nematodes and protoz oan and thei r role i n
nematode management. In Pesticide and
Environment Vol. II- (Eds- Gaur, H- C. ^
Prasad, D->- (In press).
41. TAHSEEN, OUDSIA & BILGRAMI, A. L. (199 >. Chemo-
oriented sex attraction in a plant parasitic
nematode. Hirschmanniel 1 a oryzae- Indian- J.
Nematol. (in press)-
42. KULSHRESTHA, R., BILGRAMI, A- L. S< KHAN, 2.
(1993). Predation abilities of MylonchuluB
minor and factors influencing predation.
Annals Plant Protection Sciences- (in press) -

5u9
43- KHAN, Z,, BILGRAMI, A. L. S< JAIRAJPURI, H. 5.(199)
A comparative study on the pr sds^i an by
A] 1 Ddoryl ai mu5 americanus n. 5p<- arsd
DiscolaimuB silvicolus (rvematoda: Dor yl ai mi da )
om di-f-ferent species of plant parasitic
nematodes. Fundamental and Applied Nematology.
(in press).
44- BILGRAKI, A. L. <1994>- Predatory behaviour of a
nematode -feeding mite Tyrophagus putrescent! ae
(Sarcopti-formes : Acridae). Fundamental and
Applied Nematology. <in press).
45. BILGRAMI, A. L. & KULSHERESHTHA, R. (199 ) .
Evaluation of predation abilities o-f
Mylonchulus dentatus. Indian J. Nematol.
< Communi cated).
46. KHAH, 7.,BILGRAMI, A. L. 8. JAIRAJPURI, N. S.(1994>
Observations on the predation abilities of
Neoactinolaimus aqilis (Dorylaimida :
Actinolaimoidea). Indian J. Nematol.
47. BILGRAMI, A. L. (1994). Attraction and feeding
b€?haviour of Ty 1 enchorhynchus roashhoodi .
Annals Plant Protection Sciences, (in press).

570
4 8 - B ILGRAMI , A. L- H 9 9 > . F s c t o r s i n f l u e n c i n g
a t t r a c t i o n o-f Hi r schmanni e l 1 a o ryxae to^jqards
cabbagp* SJ^edl i nn=.. Nfjmet o l og j c a . < CcTifrtuni c a t e iJ )
49- KHAt4, Z.jBILGRAMI, A. L. ^ JAIRAJPURI, M. S,<199>.
A t t r a c t i o n &nd food p r e f e r e n c e behaviour i n
p r e d a t o r y nefri.at odes , AI 1 odor y 1 fe i mu s
£<mericBnus end PiBcolai fsuB s i i v i c o i u B
^'Nematoda i Dor y 1 ^ i fni da ) . Nemat ol og i ca
50 . BILGRAKI, A. L. (199 >- Numer ica l a n a l y s i s of t he
r e l a t i o n s h i p between the p r e d a t i o n fay
Mesodoryl ai mus b s s t i a n i (<Nematoda :
Dory l a i mi da) sind d i f f e r e n t prey t r o p h i c
ca tegor ies . Nematol . pvedit . (Cominuni cated > .
Copyright © 2022 FDOKUMEN




















