
A novel Bim-BH3-derived Bcl-XL inhibitor: Biochemical characterization, in vitro, in vivo and...
14
[Cell Cycle 7:20, 3211-3224; 15 October 2008]; ©2008 Landes Bioscience BH3-only members of the Bcl-2 family exert a fundamental role in apoptosis induction. This work focuses on the development of a novel peptidic molecule based on the BH3 domain of Bim. The antiapoptotic molecule Bcl-X L , involved in cancer development/ progression and tumour resistance to cytotoxic drugs, is a target for Bim. According to a rational study of the structural interactions between wt Bim-BH3 and Bcl-X L , we replaced specific residues of Bim-BH3 with natural and non-natural aminoacids and added an internalizing sequence, thus increasing dramatically the inhibi- tory activity of our modified Bim-BH3 peptide, called 072RB. Confocal microscopy and flow cytometry demonstrated cellular uptake and internalization of 072RB, followed by co-localization with mitochondria. Multiparameter flow cytometry demonstrated that the 072RB dose-dependent growth inhibition of leukaemia cell lines was due to apoptotic cell death. No effect was observed when cells were treated with the internalizing vector alone or a mutated control peptide (single aminoacid substitution L94A). Ex-vivo derived leukemic cells from acute myeloid leukaemia (AML) patients underwent cell death when cultured in vitro in the presence of 072RB. Conversely, no significant cytotoxic effect was observed when 072RB was administered to cultures of periph- eral blood mononuclear cells, either resting or PHA-stimulated, and bone marrow cells of normal donors. Xenografts of human AML cells in NOD/SCID mice displayed a significant delay of leukemic cell growth upon treatment with 072RB administered intravenously (15 mg/Kg three times, 48 hours after tumour cell injection). Altogether, these observations support the therapeutic potentials of this novel BH3 mimetic. Introduction Impaired apoptosis is a crucial step in tumorigenesis and also a relevant impediment to a successful cytotoxic therapy. 1,2 Apoptosis can be induced by cell surface death receptors engagement, that stimulate the extrinsic pathway, or by intracellular death signals that perturb mitochondria, triggering the intrinsic pathway of programmed cell death. The intrinsic pathway is tightly regulated by the interplay among proteins of the B cell lymphoma-2 (Bcl-2) family, which comprises anti-apoptotic members (e.g., Bcl-2, Bcl-X L , Bcl-w, Mcl-1), multi-domain pro-apoptotic members (e.g., Bak and Bax) and a large group of single-domain pro-apoptotic members, the BH3-only proteins (e.g., Bim, Bid, Bad, Bmf, Noxa, Puma) 3 . Proteins of the first two classes share at least three of the four conserved structural motifs known as Bcl-2 homology domains (BH1–BH4), while the BH3-only proteins contain only the single BH3 motif (9–16 amino acids). 3 Generally pro-death proteins bind to multi-domain proteins by inserting their BH3 domain, an amphipathic α-helix, into a hydrophobic cleft made up by the folding of BH1, BH2 and BH3 domains. 3 The cell’s fate is governed by a complex and finely tuned network of protein-protein interac- tions that can be either apoptosis-promoting or -blocking. When the *Correspondence to: Silvia Bruno; Department of Experimental Medicine; Human Anatomy Section; University of Genoa; Via De Toni, 14; Genoa 16132 Italy; Phone: +390103537641; Email: [email protected]/ Silvio Parodi; Department of Oncology, Biology and Genetics; University of Genoa; Via Pastore, 3; Genoa 16132 Italy; Phone: 39+3295481011; Fax: 39+010-353-38272; Email: silvio. [email protected] Submitted: 08/06/08; Accepted: 08/20/08 Previously published online as a Cell Cycle E-publication: http://www.landesbioscience.com/journals/cc/article/6830 Report A novel Bim-BH3-derived Bcl-X L inhibitor Biochemical characterization, in vitro, in vivo and ex vivo anti-leukemic activity Raffaella Ponassi, 1,2,† Barbara Biasotti, 1,2,† Valeria Tomati, 1,2 Silvia Bruno, 3, * Alessandro Poggi, 4 Davide Malacarne, 2 Guido Cimoli, 5 Annalisa Salis, 6 Sarah Pozzi, 7 Maurizio Miglino, 8 Gianluca Damonte, 6 Pietro Cozzini, 9 Francesca Spyraki, 9 Barbara Campanini, 9 Luca Bagnasco, 1,2 Nicoletta Castagnino, 1,2 Lorenzo Tortolina, 1,2 Anna Mumot, 1,2 Francesco Frassoni, 7 Antonio Daga, 10 Michele Cilli, 11 Federica Piccardi, 11 Ilaria Monfardini, 12 Miriam Perugini, 12 Gabriele Zoppoli, 12 Cristina D’Arrigo, 13 Raffaele Pesenti 14 and Silvio Parodi 1,2,‡ 1 Department of Oncology, Biology and Genetics; 3 Department of Experimental Medicine; Section of Human Anatomy; 8 Hematology Division, and 12 Department of Internal Medicine; University of Genoa; Genoa Italy; 2 Laboratory of Experimental Oncology; 4 Laboratory of Immunology; 10 Laboratory of Genes Transfer; and 11 Animal Facility; National Cancer Institute (IST); Genoa, Italy; 5 Tecan Italia S.r.l.; Cernusco Sul Naviglio (MI), Italy; 6 Department of Experimental Medicine; Biochemistry section c/o Center of Excellence for Biomedical Research; Genoa Italy; 7 Department of Hemato-Oncology; S. Martino Hospital; Genoa Italy; 9 Biochemistry and Molecular Biology; University of Parma; Parma Italy; 13 Institute for Macromolecules Studies; CNR Genoa Area; Genoa Italy; 14 Department of Applied Mathematics; Ca’ Foscari University; Venice Italy † These authors contributed equally to this work. ‡ Scientific coordinator of the team work Abbreviations: AML, acute myeloid leukaemia; ALL, acute lymphoblastic leukaemia; NBM, normal bone marrow; PBL, peripheral blood lymphocytes; CD, circular dichroism; FA, fluorescence anisotropy; FBS, fetal bovine serum Key words: Bim-BH3, apoptosis, multi-parameter flow cytometry, leukaemia, targeted therapy www.landesbioscience.com Cell Cycle 3211
Transcript of A novel Bim-BH3-derived Bcl-XL inhibitor: Biochemical characterization, in vitro, in vivo and...

[Cell Cycle 7:20, 3211-3224; 15 October 2008]; ©2008 Landes Bioscience
BH3-only members of the Bcl-2 family exert a fundamental role in apoptosis induction. This work focuses on the development of a novel peptidic molecule based on the BH3 domain of Bim. The antiapoptotic molecule Bcl-XL, involved in cancer development/progression and tumour resistance to cytotoxic drugs, is a target for Bim. According to a rational study of the structural interactions between wt Bim-BH3 and Bcl-XL, we replaced specific residues of Bim-BH3 with natural and non-natural aminoacids and added an internalizing sequence, thus increasing dramatically the inhibi-tory activity of our modified Bim-BH3 peptide, called 072RB. Confocal microscopy and flow cytometry demonstrated cellular uptake and internalization of 072RB, followed by co-localization with mitochondria. Multiparameter flow cytometry demonstrated that the 072RB dose-dependent growth inhibition of leukaemia cell lines was due to apoptotic cell death. No effect was observed when cells were treated with the internalizing vector alone or a mutated control peptide (single aminoacid substitution L94A). Ex-vivo derived leukemic cells from acute myeloid leukaemia (AML) patients underwent cell death when cultured in vitro in the presence of 072RB. Conversely, no significant cytotoxic effect
was observed when 072RB was administered to cultures of periph-eral blood mononuclear cells, either resting or PHA-stimulated, and bone marrow cells of normal donors. Xenografts of human AML cells in NOD/SCID mice displayed a significant delay of leukemic cell growth upon treatment with 072RB administered intravenously (15 mg/Kg three times, 48 hours after tumour cell injection). Altogether, these observations support the therapeutic potentials of this novel BH3 mimetic.
Introduction
Impaired apoptosis is a crucial step in tumorigenesis and also a relevant impediment to a successful cytotoxic therapy.1,2 Apoptosis can be induced by cell surface death receptors engagement, that stimulate the extrinsic pathway, or by intracellular death signals that perturb mitochondria, triggering the intrinsic pathway of programmed cell death. The intrinsic pathway is tightly regulated by the interplay among proteins of the B cell lymphoma-2 (Bcl-2) family, which comprises anti-apoptotic members (e.g., Bcl-2, Bcl-XL, Bcl-w, Mcl-1), multi-domain pro-apoptotic members (e.g., Bak and Bax) and a large group of single-domain pro-apoptotic members, the BH3-only proteins (e.g., Bim, Bid, Bad, Bmf, Noxa, Puma)3. Proteins of the first two classes share at least three of the four conserved structural motifs known as Bcl-2 homology domains (BH1–BH4), while the BH3-only proteins contain only the single BH3 motif (9–16 amino acids).3 Generally pro-death proteins bind to multi-domain proteins by inserting their BH3 domain, an amphipathic α-helix, into a hydrophobic cleft made up by the folding of BH1, BH2 and BH3 domains.3 The cell’s fate is governed by a complex and finely tuned network of protein-protein interac-tions that can be either apoptosis-promoting or -blocking. When the
*Correspondence to: Silvia Bruno; Department of Experimental Medicine; Human Anatomy Section; University of Genoa; Via De Toni, 14; Genoa 16132 Italy; Phone: +390103537641; Email: [email protected]/ Silvio Parodi; Department of Oncology, Biology and Genetics; University of Genoa; Via Pastore, 3; Genoa 16132 Italy; Phone: 39+3295481011; Fax: 39+010-353-38272; Email: [email protected]
Submitted: 08/06/08; Accepted: 08/20/08
Previously published online as a Cell Cycle E-publication: http://www.landesbioscience.com/journals/cc/article/6830
Report
A novel Bim-BH3-derived Bcl-XL inhibitorBiochemical characterization, in vitro, in vivo and ex vivo anti-leukemic activity
Raffaella Ponassi,1,2,† Barbara Biasotti,1,2,† Valeria Tomati,1,2 Silvia Bruno,3,* Alessandro Poggi,4 Davide Malacarne,2 Guido Cimoli,5 Annalisa Salis,6 Sarah Pozzi,7 Maurizio Miglino,8 Gianluca Damonte,6 Pietro Cozzini,9 Francesca Spyraki,9 Barbara Campanini,9 Luca Bagnasco,1,2 Nicoletta Castagnino,1,2 Lorenzo Tortolina,1,2 Anna Mumot,1,2 Francesco Frassoni,7 Antonio Daga,10 Michele Cilli,11 Federica Piccardi,11 Ilaria Monfardini,12 Miriam Perugini,12 Gabriele Zoppoli,12 Cristina D’Arrigo,13 Raffaele Pesenti14 and Silvio Parodi1,2,‡
1Department of Oncology, Biology and Genetics; 3Department of Experimental Medicine; Section of Human Anatomy; 8Hematology Division, and 12Department of Internal Medicine; University of Genoa; Genoa Italy; 2Laboratory of Experimental Oncology; 4Laboratory of Immunology; 10Laboratory of Genes Transfer; and 11Animal Facility; National Cancer Institute (IST); Genoa, Italy; 5Tecan Italia S.r.l.; Cernusco Sul Naviglio (MI), Italy; 6Department of Experimental Medicine; Biochemistry section c/o Center of Excellence for Biomedical Research; Genoa Italy; 7Department of Hemato-Oncology; S. Martino Hospital; Genoa Italy; 9Biochemistry and Molecular Biology; University of Parma; Parma Italy; 13Institute for Macromolecules Studies; CNR Genoa Area; Genoa Italy; 14Department of Applied Mathematics; Ca’ Foscari University; Venice Italy
†These authors contributed equally to this work.
‡Scientific coordinator of the team work
Abbreviations: AML, acute myeloid leukaemia; ALL, acute lymphoblastic leukaemia; NBM, normal bone marrow; PBL, peripheral blood lymphocytes; CD, circular dichroism; FA, fluorescence anisotropy; FBS, fetal bovine serum
Key words: Bim-BH3, apoptosis, multi-parameter flow cytometry, leukaemia, targeted therapy
www.landesbioscience.com Cell Cycle 3211

3212 Cell Cycle 2008; Vol. 7 Issue 20
Anti-leukemic activity of a new Bcl-XL inhibitor
former prevail, activation of the multi-domain pro-apoptotic execu-tioner proteins Bax and Bak takes place, followed by pore formation on the outer mitochondrial membrane, release of cytochrome c and other apoptogenic factors that interact in the cytosol with Apaf-1 to activate death-promoting caspases.4 However, several aspects of the Bcl-2 regulatory pathways are not fully elucidated. In particular, it is debated whether the BH3-only proteins of the Bcl-2 family exert their pro-apoptotic function by activating pro-apoptotic members or by inhibiting anti-apoptotic members. According to the so-called “indirect activation” model, anti-apoptotic proteins hold in check Bax or Bak by sequestering them in a reversible manner.5,6 For apoptosis to occur, BH3-only pro-apoptotic proteins compete for the hydrophobic cleft on the anti-apoptotic proteins, thus triggering the release of active Bax and Bak.6 In the “two-class” or “direct acti-vation” model, the BH3-only proteins are divided into two distinct subsets (“activators”, e.g., Bim, Puma and Bid , and “sensitizers”, e.g., Bad and Noxa), each acting at a different step in a hierarchy of Bcl-2-family interactions.7-9 Briefly, in response to apoptogenic signals the “activator” BH3-only proteins, sequestered by the anti-apoptotic proteins, are freed from this association upon displacement by “sensitizers ” BH3-only proteins (e.g., Bad and Noxa)8 and allowed to bind to and subsequently activate Bax and Bak. It is of note that Bim, tBid (the activated, truncated form of Bid) and Puma, are promiscuous pro-apoptotic proteins in that they avidly bind to all anti-apoptotic proteins, while the others bind only to subsets (e.g., Bad engages Bcl-2 and Bcl-XL but not Mcl-1, and Noxa preferen-tially binds to Mcl-1 and not to Bcl-XL and Bcl-2).6,8,10
Regardless of the different hierarchies attributed to the specific interactions among the above Bcl-2-family members, and despite the fact that a consensus has not been reached for whether or not BH3-only proteins, and which of them, are also able to directly bind and activate Bax and Bak, the two models agree that BH3-only proteins are critical for cell death initiation and exert a fundamental role for apoptosis induction by their direct binding to some or all anti-apoptotic members. Exploitation of the BH3-only protein features provides clues for the development of therapeutic anti-cancer approaches that specifically target anti-apoptotic proteins overex-pressed in tumors. To this end a number of BH3 mimetic drugs was produced in the last decade. They are either peptidic BH3-domains of BH3-only proteins or non-peptidic synthetic small molecules identified from random screening of combinatorial libraries or libraries of natural products, or designed/improved by rational struc-ture-based techniques. The non-peptidic small molecules have been questioned for their specificity for Bcl-2 pathways in several cases. Some of them, including the gossypol-enantiomer11 or HA14-1,12 do not kill cells like BH3-only proteins, but through other unknown pathways.13 The cycloprodigiosin-derivative small molecule GM15-070 (Obatoclax), in addition to releasing Bak from Mcl-1 and liberating Bim from Bcl-2 and Mcl-1, activates apoptosis also by other Bcl-2-independent pathways.14 In addition, it failed to exhibit efficacy in tumour xeno-transplant models in vivo.15 To date, the most studied and promising synthetic small molecule BH3-mimetic is ABT-737.16 It exhibits apoptotic efficacy against several cell tumor cell lines and ex-vivo tumor cells,16-18 potentiates the activity of some commonly used therapeutic agents19,20 and counteracts human tumor growth in mouse xenograft models.17 However, ABT-737, like its second generation orally bio-available compound ABT-263,21 is
effective in unleashing cell killing only if Mcl-1 activity is not crucial for the target cell,13,17 due to its BAD-like BH3 mimetic structure. Indeed, BH3 mimetics often lack overall single-agent apoptogenic ability, and are mostly effective when utilized in conjunction with commonly used therapeutic compounds. Their single-agent effec-tiveness largely depends from their specificity on one or another Bcl-2-family molecule, on the overall tumour basal pattern of Bcl-2-family molecules expression, and on other more complex features of the individual signalling network of a given tumour. As adjuvant to ABT-737 for killing tumors that overexpress both Bcl-XL and Mcl-1,22 a recently published peptidomimetic drug was proposed.22 It derives from the Bim BH3-domain modified to be highly selec-tive for Mcl-1, and cannot be utilized as single agent either, since it promotes cell death only when, in addition to Mcl-1, also Bcl-XL is neutralized.22 No data are available yet about its in vivo efficacy.
Another promising peptidic BH3 mimetic was recently devel-oped, a “hydrocarbon-stapled” Bid-based BH3 domain, modified to become cell permeable and protease resistant through its increased helicity in solution.23 It showed activity against a panel of tumour cell lines and delayed growth of human leukaemia in a xeno-trans-plant model.23 Rather than inhibition of anti-apoptotic molecules, a direct Bax oligomerization seems to account for its pro-apoptotic activity.24 In addition, very little information is available about its effects on healthy cells.
Altogether, although much progress has been made over the last few years, further investigations are needed to achieve sensitive and specific inhibition of anti-apoptotic components of the Bcl-2 machinery.
We focused our attention on the BH3 only protein Bim, which emerged recently as one of the most critical Bcl-2 family proteins in the hematopoietic system.25 This small protein is involved in the regulation of apoptosis and homeostasis in several cell types, including B and T lymphocytes, and thymocytes.25 Several cellular stresses, like survival factors withdrawal, unbalances in onco-protein expression, endoplasmic reticulum (ER) stresses, induce cell death through Bim activation.25,26 Homozigous deletion of Bim is associated with mantle cell lymphoma27 and survival of auto-reactive T and B cells,28,29 and repression of Bim expression by EBV has a significant role in Burkitt lymphoma.30 Several antitumor chemotherapies were demonstrated to kill neoplastic cells through Bim-activated pathways, including a Raf kinase inhibitor in acute myeloid cells,31 glucocorticoids in leukaemia cell lines and primary leukemia cells,32,33 ABT-737 for chronic lymphocytic leukaemia cells and myeloma cell lines18 and paclitaxel in xenografts of epithelial cancer cells.34 Altogether, Bim can be classified as a powerful tumour suppressor, and its strong pro-apoptotic activity is regulated both at transcriptional and post-translational levels.25,35,36
Bim potently engages all anti-apoptotic members of the Bcl-2 family with high affinity.10 Inhibition of Bcl-XL anti-apoptotic activity is particularly attractive since Bcl-XL protein overexpression is linked to the progress of hematologic and solid malignancies, contributes to clonal expansion, metastatic spreading and tumour resistance to common cytotoxic drugs.37-39
In this work we synthesized a series of Bim-BH3 derived peptides. The biochemical activity of each peptide was evaluated through fluorescence anisotropy assays, leading to selection of the molecule with the highest affinity for Bcl-XL. The selected peptide was further

Anti-leukemic activity of a new Bcl-XL inhibitor
www.landesbioscience.com Cell Cycle 3213
Peptides cellular uptake and intracellular localization. We first verified cellular uptake and internalization of the 072RB peptide by flow cytometry and fluorescence microscopy, respectively (Fig. 1). U937, Jurkat and Namalwa human leukemic cell lines were incu-bated with 072RB, or the internalizing sequence Int alone, all labelled with a fluoresceine molecule at their N-terminus. Displayed in Figure 1A are U937 cells incubated with fluoresceinated peptides (1 or 10 μM) and examined after 60 min or 180 min. Compared to the extra-cellular fluorescence significant 072RB uptake was observed already after 60 min of 10 μM incubation (Mean Fluorescence Intensity (MFI): 350 ± 50 a.u., n = 4). The fluorescence further increased during the subsequent two hours, and reached a plateau at 18–24 h (MFI: 1700 ± 150 a.u., n = 3) (Fig. 1B). The amount of fluoresceinated 072RB taken up by the cells was comparable to that of fluoresceinated Int (Fig. 1A). On the contrary, fluoresceinated Bim-BH3YA2Aib, lacking the internalizing sequence, was taken up by the cells at a much slower rate and inefficiently: after 60 min of 10 μM treatment the mean MFI was approximately 15 a.u., and increased to only 50 a.u. after 180 min (Fig. 1A). When 072RB, Int or Bim-BH3YA2Aib, were administered to Jurkat or Namalwa cells, the uptake of the different peptides by these cells was very similar to that shown by U937 cells (Fig. 1B and not shown).
Confocal microscopy measurements were carried out on the tumour cell lines incubated with fluoresceinated 072RB 10 μM for 3 h to determine whether 072RB preferentially co-localized with mitochondria, since Bcl-XL is known to mainly localize on the mito-chondria outer membrane and, to a lesser extent, on the endoplasmic reticulum. Mitotracker red, a mitochondrion-selective probe, was added during the last 30 min of incubation. As shown in Figure 1C, fluoresceinated 072RB mainly co-localized with Mitotracker red. Interestingly, it did not stain lysosomes (Fig. 1D), as shown by lack of co-localization with cellular organelles labelled with Lysotracker Red DND-99, a fluoresceinated probe specific for lysosomes. Conversely, only a dim fluorescence halo at the plasma membrane and no intra-cellular fluorescence were detected when cells were incubated with fluoresceinated Bim-BH3YA2Aib (not shown), Instead, for cells treated with the internalizing peptides the fluorescence halo around the cellular border was a very minor component with respect to the
modified to improve its stability in serum and increase its ability to penetrate cell membranes. The resulting peptide was named 072RB.
The apoptotic effects of 072RB were investigated on leukaemia cell models, either in vitro cell cultures of leukaemia cell lines or ex-vivo bone marrow- and peripheral blood-derived cells from acute myeloid leukemia (AML) patients. As well, 072RB was assayed on peripheral blood and bone marrow cells from healthy donors. In addition, the in vivo antiproliferative activity of 072RB was studied in a mouse model of human AML xenograft.
Results
Data on the experimental strategies used for rationally designing the Bim-BH3 peptides synthesized in the present study are described in the Results section of the Supplemental data. Likewise are reported data on the secondary structure of the different peptides. We also studied their stability at 37°C in complete tissue culture medium and/or 100% FCS, after modifications carried out to improve their stability and resistance to proteases. We studied cell permeability (internalization capability of our molecules). Finally we studied the different potencies of our molecules as inhibitors (Kis) of the refer-ence [(BakF-BH3D84A):(Bcl-XL)] reversible interaction. Briefly, the wild-type human Bim-BH3 domain was first modified by two amino acid substitutions (W89Y and E100A) that increased its affinity for Bcl-XL, followed by substitution of the first and the last Alanine (A91, A103) with α-amino isobutyric acid (Aib) residues, that increased its α-helicity and consequently its affinity for the BH3-binding groove on target protein surface. The resulting peptide, called Bim-BH3YA2Aib, lacked structural features for crossing the cytoplasmic membrane barrier. To allow its cellular internalization, the sequence was modified by adding a mutated version of the third helix of the Antennapedia protein as internalizing tag (Int). We called the resulting peptide 072RB. A negative control peptide, 072RB L94A, was also synthesized. It bears a point mutation that decreased by about 42 times its inhibitory capability of the [(BakF-BH3D84A):(Bcl-XL)] reversible interaction, in the presence of a negligible reduction (≈13%) of the level of α-helicity. The amino acid sequences of the inhibitor peptides and their Ki toward the [(BakF-BH3D84A):(Bcl-XL)] interaction are summarized in Table 1.
Table 1 Synthesized peptides: names, sequences, Kd or Ki values, ± SE
Peptide name Peptide sequence Kd weighted mean Pooled SEBakF-BH3D84A 72 GQVGRQLAIIGDAINR 87 138.0E-09M 15.6E-09 Ki weighted mean Pooled SEBim-BH3 83 DMRPEIWIAQELRRIGDEFNAY 104 334.0E-09M 35.8E-09Bim-BH3W89Y 83 DMRPEIYIAQELRRIGDEFNAY 104 29.2E-09M 5.9E-09Bim-BH3W89Y,A91Aib,E100A,A103Aib 83 DMRPEIYIAibQELRRIGDAFNAibY 104 12.0E-09M 4.2E-09 (“Bim-BH3YA2Aib” in the text)072RB 83 DMRPEIYIAibQELRRIGDAFNAibY 104 -Int 4.3E-09M 1.0E-09072RB L94A 83 DMRPEIYIAibQEARRIGDAFNAibY 104 -Int 180.0E-09M 33.0E-09IntQ50P (“Int” in the text) 58 KKWKMRRNPFWIKIQR 43 - -
Inhibition of the interaction [Bcl-XL]:[BakF-BH3D84A]. Weighted mean ± pooled standard error (SE): fluorescence anisotropy assays. For each individual experimental curve [Anisotropy = f(Bcl-XL concentration)] eight different Bcl-XL concentrations were studied: 1.0E-10M; 2.0E-8M; 6.0E-8M; 1.0E-7M; 2.0E-7M; 3.0E-7M; 5.0E-7M; 1.0E-6M. BakF-BH3D84A was kept constant at 2.0E-8M. The inhibitor molecules were tested at the constant concentration of 2.0E-7M. For each molecule at least three independent curves were obtained. For the same inhibitor molecule, we assumed that the Kis and the SEs obtained from our different curves, all belonged to the same population. According to this assumption a weighted Ki average and a weighted average of the sample variances (pooled variance) was computed (see also in Supplemental Material and Methods: Computation of Kd and Ki values from Fluorescence Anisotropy binding experiments). IntQ50P (“Int” in the text) is an internalization sequence corresponding to a Q50P modified IIIrd α-helix of the protein Antennapedia. The human Bim-BH3 derived peptide amino-acid numeration refers to the murine sequence reported by Liu X et al.56

Anti-leukemic activity of a new Bcl-XL inhibitor
3214 Cell Cycle 2008; Vol. 7 Issue 20
pixel separately for the extracellular milieu, for the cellular cytoplasm and for the mitochondria at 24 h 072RB incubation, and after appro-priate subtraction of background and extracellular fluorescence, led to the result that the relative amount of fluoresceinated 072RB associated to the whole mitochondrial compartment was approximately nine folds higher than the one present in the cytoplasm.
intracellular fluorescence levels (confocal microscopy observations of median cellular sections).
Taken together, the flow cytometric and confocal microscopy data indicate that the internalizing sequence Int is necessary for an efficient internalization and that the peptide 072RB specifically targets mito-chondria. Quantification of the microscopy mean fluorescence intensity/
Figure 1. Cellular internalization of 072RB and co-localization with mitochondria. (A) U937 leukemic cells were incubated at 37°C with 1 or 10 μM of the following fluoresceinated peptides: Bim-BH3YA2Aib (left column), Int (middle column) and 072RB (right column), for 60 min or 180 min as indicated. The green fluorescence intensity was measured in log scale by flow cytometry (arbitrary units, a.u.). Bold black lines: fluorescence intensity of cells incubated with fluoresceinated peptides. Thin black lines: background fluorescence of cells without peptides. (B) Kinetics of 072RB uptake in Jurkat, Namalwa and U937 cell lines. Cells were incubated with fluoresceinated 072RB at a 10 μM concentration for the indicated periods of time and fluorescence analysed by flow cytometry. (C and D) Namalwa cells were stained with fluoresceinated 072RB for 3 hours, incubated with Mitotracker red 580 (C) or Lysotracker red DND-99 (D) and analyzed by confocal microscopy with a 40x oil/1.35 NA objective. The two images show one median z-axis plane.

Anti-leukemic activity of a new Bcl-XL inhibitor
www.landesbioscience.com Cell Cycle 3215
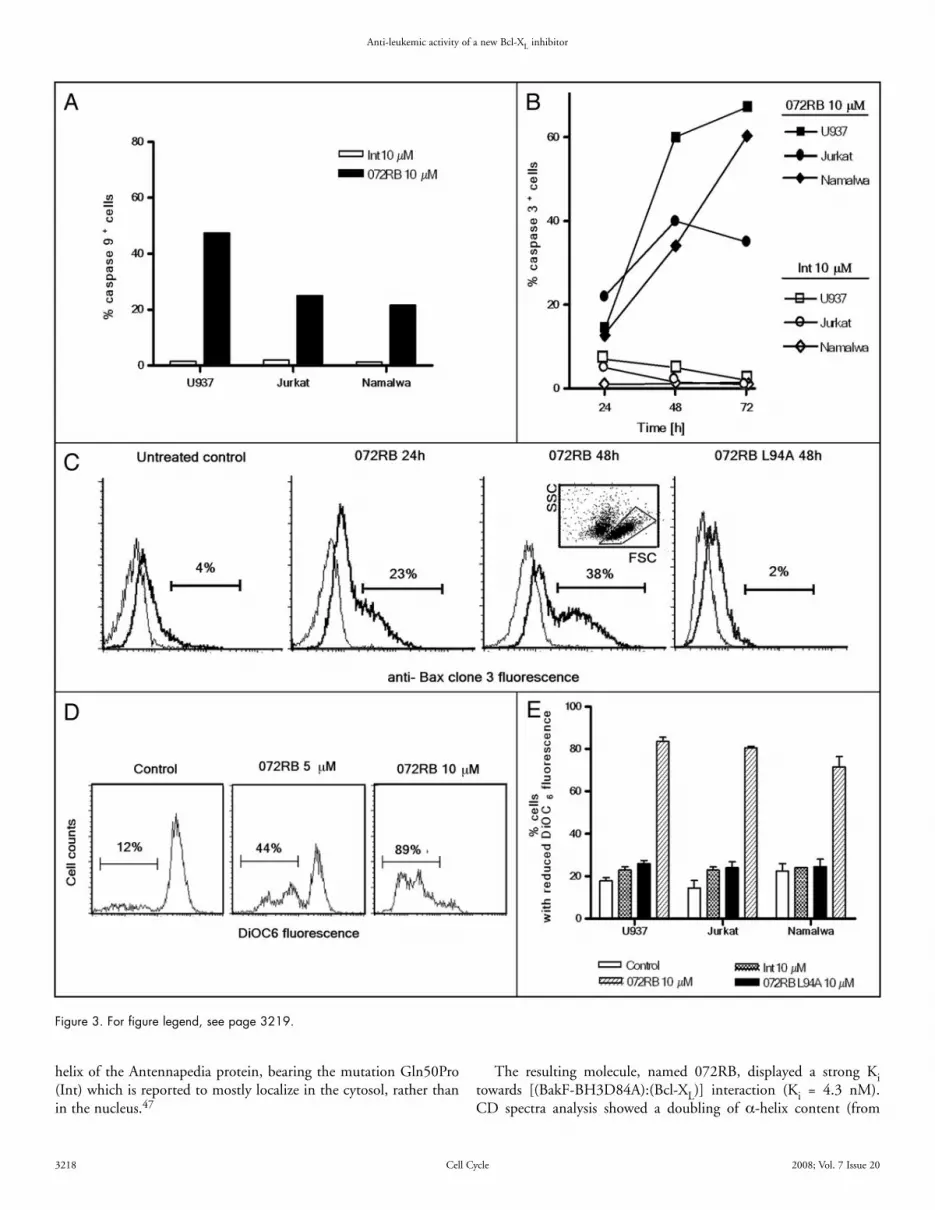
lipophylic cationic probe that accumulates in the mitochondrial matrix of viable cells, because of the relatively high electric potential across the mitochondrial inner membrane. Dissipation of the trans-membrane potential is demonstrated by the drop of cellular DiOC6 fluorescence.40,43 Flow cytometry DiOC6 fluorescence histograms of one representative experiment are shown in Figure 3D. Data on the three leukaemia cell lines are reported in the bar graph of Figure 3E and confirmed trans-membrane potential dissipation after 072RB treatment, but not Int or 072RB L94A.
Ex vivo cellular specimens: 072RB affects leukemic but not normal cell viability. The effects of the peptide on ex vivo cellular specimens from haematological malignancies were investigated. Bone marrow aspirates from seven patients with acute myeloid leukaemia (AML) and two patients with acute lymphoblastic leukaemia (ALL) were plated in tissue culture and treated with 072RB or with the internalizing vector Int. Cellular viability was assessed by the flow cytometric PI exclusion test (Table 2). Except for a mild viability decrease observed in the two M0 samples treated with 10 μM 072RB (relative decrease of 13% and 30%), and an intermediate viability decrease in the two ALL samples (relative decrease of 42% and 46%), all other AML samples displayed significant cytotoxicity (relative decrease of the fraction of viable cells ranging from 50% to 77%). Two normal bone marrow specimens were also treated (Table 2). Notwithstanding results obtained for two samples only, normal bone marrow cells seem much less sensitive to 072RB than leukaemia cells.
To further study the sensitivity of normal cells to 072RB, periph-eral blood lymphocytes (PBL) isolated from four different healthy donors were subjected to a four days treatment with 10 μM 072RB. Treatment was performed as usual with two pulses (at t = 0 and 48 h). Lymphocytes were either un-stimulated or activated by PHA (added 48 h before the first 072RB pulse), in order to study the effects of the peptide both on resting and proliferating lymphocytes, respectively. Data on lymphocyte viability, reported in Figure 4A and B for each of four healthy donors, indicate that 072RB exerted no cytotoxic activity both on resting and proliferating lymphocytes derived from Donors 3 and 4 and decreased the fraction of viable cells from Donors 1 and 2 by approximately 20%. Altogether, it seems that the effect of the peptide on normal lymphocytes is minor, especially if compared to the 10 μM effects observed on leukaemia cell lines (Fig. 2) and ex-vivo AML cells (Table 2). Effects on cell cycling of PHA activated lymphocytes were also analyzed by DNA flow cytometry. Data reported in Figure 4C show a slight decrease of the percentage of cells in S, G2 and M phases of the cell cycle in two (Donors 1 and 2) out of four donors, thus suggesting that a possible mild cytostatic effect could be exerted by 072RB on proliferating peripheral blood lymphocytes. Notwithstanding the limited sample size, it is of note that sensitivity to 072RB both in terms of cell viability and prolifera-tion potential was observed in the same two donors.
In vivo anti-leukemic effects of 072RB. Preliminary toxicity experiments on Balb-C four weeks old female mice were carried out to find a convenient peptide dosage and treatment schedule (data not reported). A biologic activity of 072RB toward mouse Bcl-XL has to be experimentally verified, however mouse Bcl-XL protein shares 98% sequence identity with human Bcl-XL, suggesting that 072RB could in principle induce toxicity through apoptotic events even in normal murine tissues. Mice received 072RB at 0, 3 or 5 μmol/Kg (i.e., 0, 15
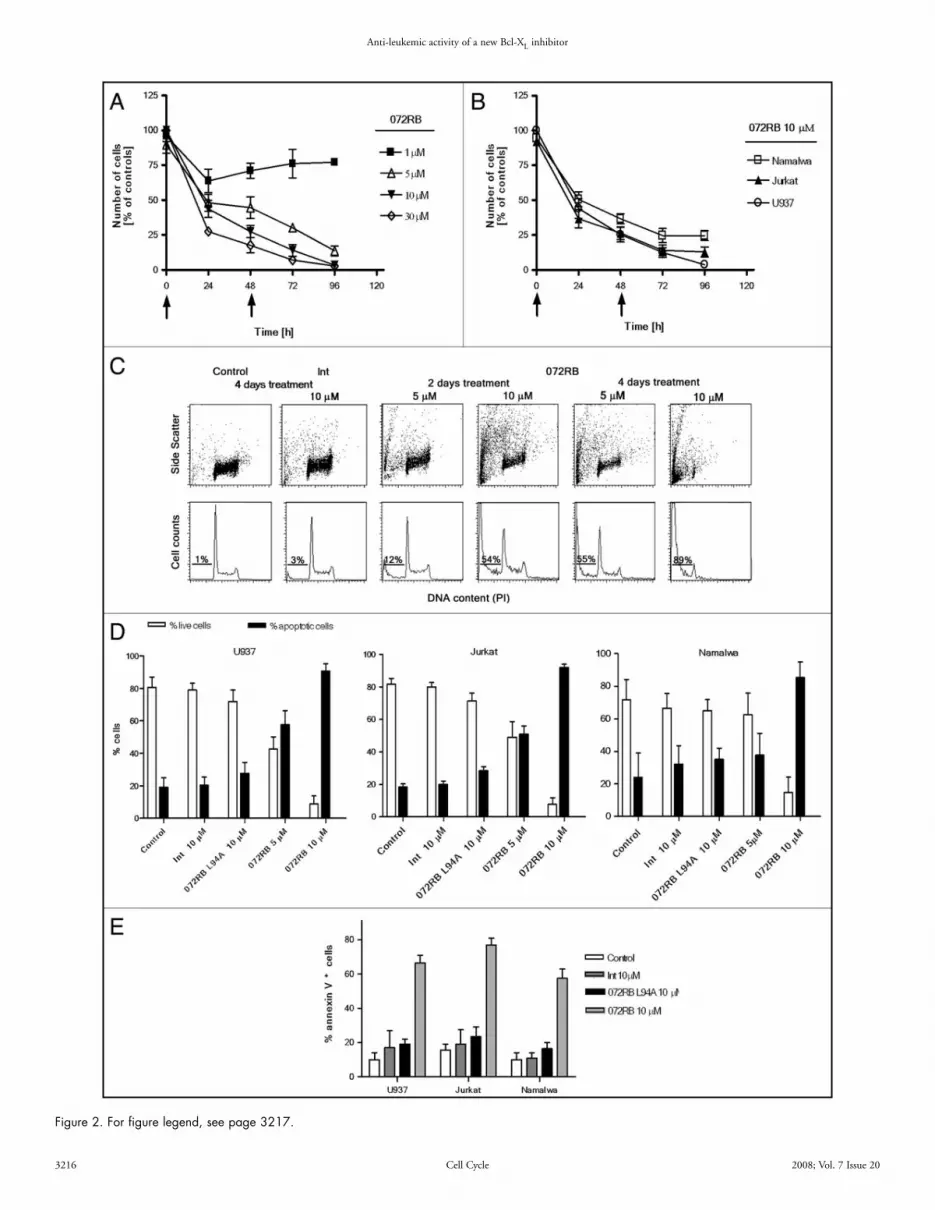
072RB affects leukemic cells growth in vitro, by inducing apop-tosis. To verify whether 072RB affects cell growth of leukemic cell lines and to select the optimum administration protocol, U937 cells were treated with different 072RB concentrations, from 1 μM up to 30 μM, and counted every 24 h. Due to its half-life of 36 h in tissue culture medium containing 10% FBS at 37°C, in all experiments 072RB was re-added to the cell cultures every 48 h. Counts of treated cell cultures, relative to untreated controls, indicate a dose-dependent inhibition of the cell growth (Fig. 2A). Likewise, a progressive decrease of cell counts was observed for U937, Jurkat and Namalwa cell lines exposed to a 10 μM concentration of 072RB (Fig. 2B).
DNA flow cytometry assays were performed to evaluate whether cell growth inhibition was due to cell cycle arrest, to cell death, or both processes.40 As shown in Figure 2C, flow cytometric measure-ments of DNA content and side light scatter confirmed that the main cause of cell growth inhibition was cell death rather than cell cycle perturbation, with an increasing trend dependent on time of treatment and peptide concentration (Fig. 2C). Data collected on the three tumour cell lines treated for four days with 072RB, both at 5 or 10 μM, and the two control peptides Int or 072RB L94A, at a 10 μM concentration, are summarized in Figure 2D. Annexin V measurements confirmed apoptotic cell death40 (Fig. 2E) triggered by 072RB, while the internalizing sequence Int and the mutant peptide 072RB L94A were fully inactive.
Flow cytometric assessment of caspases 9 and 3 activation was achieved by incubating U937, Jurkat and Namalwa cell lines with the cell-permeable fluorochrome-labeled inhibitors of caspases (FLICA) FITC-LEDH-FMK and FITC-DEVD-FMK, that are specific for active centers of caspase 9 and 3, respectively.41 Though we are aware that FLICA binding to apoptotic cells may involve other mechanisms than simply their specific attachment to active caspases,42 neverthe-less we may derive from our observations that caspase 9 was activated in a significant fraction of tumour cells after 48 h 072RB incubation (Fig. 3A) and caspase 3, a downstream substrate of caspase 9, was progressively activated at increasing incubation times with 072RB (Fig. 3B). Finally, apoptosis was confirmed also by alterations of nuclear morphology, such as chromatin condensation and formation of apoptotic bodies which were evident after 36 h and 48 h of incu-bation with 072RB (not shown).
072RB-induced apoptosis is associated to Bax activation and mitochondrial permeability transition (MPT). We then asked whether apoptosis induced by 072RB involves the intrinsic mito-chondrial pathway that culminates with the conformational change and activation of Bax. To this end we performed intracellular staining of leukemic cell lines, treated either with 072RB or its mutant 072RB L94A (both at 10 μM concentrations), with an anti-Bax antibody specific for its active conformational state. Flow cytometric analysis of anti-Bax fluorescence, as reported in Figure 3C, demon-strated a significant increase of active Bax-positive U937 cells upon 072RB treatment, but not with the mutated peptide. Similar results were achieved for Jurkat cells (not shown). It has to be mentioned that the analysis was confined to the cells that displayed still low forward- and low side-light scatter features (inset of Fig. 3C), while cells in late apoptosis were excluded.
We then studied whether dissipation of the inner mitochondrial trans-membrane potential was associated to the 072RB induced apoptosis. To this end we measured the fluorescence of DiOC6, a
Anti-leukemic activity of a new Bcl-XL inhibitor
3216 Cell Cycle 2008; Vol. 7 Issue 20
Figure 2. For figure legend, see page 3217.

Anti-leukemic activity of a new Bcl-XL inhibitor
www.landesbioscience.com Cell Cycle 3217
proteins, are considered promising tailored and target-specific anticancer strategies. As well, tumors that display downregulated BH3-only protein expression, due to factors such as inactivation of the p53 pathway that transcriptionally and post-transcriptionally regulates several BH3-only proteins, BH3-only gene ablation or epigenetic silencing, would strongly benefit from the use of BH3 mimetics.
Here, we report a rationally designed Bim-BH3 derived pepti-domimetic molecule modified to improve stability and resistance to proteases, cell permeability and affinity for Bcl-XL. The apoptogenic features of the molecule are presented.
Starting from a short motif (aa 83–104) surrounding the wild-type human Bim-BH3 domain, several peptidomimetic molecules were first synthesized. Structural investigations were performed to increase the affinity of our lead molecules toward the target, leading to different and subsequent amino acid substitu-tions. Potential Bcl-XL inhibitors were then screened through a biochemical assay based on fluorescence anisotropy and the peptide Bim-BH3YA2Aib resulted the most promising molecule (Ki of the [(BakF-BH3D84A):(Bcl-XL)] interaction = 1.2 E-08M). As suggested by literature data44,45 and FA assays, Bim-BH3YA2Aib was designed by rationally introducing the following substitutions: Trp89Tyr, Glu100Ala, Ala91Aib and Ala103Aib. Molecular modelling of the [Bcl-XL:BimBH3YA2Aib] complex, complemented with peptides CD spectra de-convolution results, explained that the potently increased inhibitory activity, in terms of Ki, of Bim-BH3YA2Aib toward [(BakF-BH3D84A):(Bcl-XL)] interaction (28 times stronger than Bim-BH3wt) was due to the protein-peptide complex stabilizing effect of two new H-bonds (generated from the Trp89Tyr substitu-tion) and its enhanced α-helicity (due to Aib residues). Indeed, the α-helix structure is required for complex formation.44 In 2007, Hinds and co-workers46 carefully investigated Bim secondary and tertiary structures concluding that Bim is mostly unstructured in the absence of binding partners.47 In a binary hetero-dimer [(Bim):(pro-survival-protein)], most residues remain disordered, while only the BH3 motif becomes structured as a short α-helix, resembling a “bead on a string”. Importantly, introduction of non-natural Aib residues led to a longer half-life of Bim-BH3YA2Aib with respect to Bim-BH3W89Y in complete cellular medium.
To assess Bim-BH3YA2Aib activity on cell systems, the peptide sequence was further modified by adding an internalizing tag at the C-terminus, because Bim-BH3YA2Aib sequence lacks structural features for crossing the cytoplasmic membrane barrier. Since Bcl-XL protein is mostly cytosolic or loosely anchored to the mitochondrial outer membrane,3 the selected internalizing sequence was the third
or 25 mg/kg) twice, at days 0 and 2. By gross inspection, at day seven all mice were found in good general conditions. All animals were euthanized to perform histological examinations of different tissues. No evident tissue toxicity was observed in spleen and liver samples, even in animals where the peptide was administered at the maximum dosage (Poggi, Cilli, Piccardi personal communication).
The pro-apoptotic activity of 072RB was then investigated in NOD/SCID mice transplanted with xenografts of AML (U937luc+)]. On day 0, mice were injected with AML cells, stably expressing a luciferase gene, U937-luc. Leukaemia cells spreading inside the mouse body was monitored using the In Vivo Imaging System, which quantitated total body luminescence after intra-peritoneal injection of D-luciferin. On day 2 after injection, leukemic mice were treated intravenously with 072RB or with the internalizing sequence Int (both administered 3 μmol/kg = 15 and 6.5 mg/kg, respectively) and subsequently for two more times, every 48 h, i.e., at day 4 and day 6. To measure leukemic cells growth, whole body images for each animal have been obtained weekly, over a 28-day time course experi-ment, starting taking pictures at day 14 after tumour cells injection. At day 21 untreated mice, as well as mice treated with Int alone, demonstrated progressive leukemic growth as quantified by increased bioluminescence (Fig. 5). 072RB treatment slowed or suppressed leukemic growth throughout the whole observation period. A non parametric statistical analysis (one tailed Mann-Whitney U test) of bioluminescence values was performed, confirming a significant leukemic growth inhibition in 072RB treated animals (# = 6) (Fig. 6). No difference was assessed between PBS (# = 8) and Int (# = 6) treated animals (p = 0.30 at 21 and p = 0.33 at 28 days acquisitions), while values of p =0.0003 and p = 0.0093 were obtained when comparing PBS and 072RB treated mice at 21 days and 28 days, respectively. Likewise, statistical analysis of Int and 072RB treated mice showed a significant difference both at 21 and 28 days of acquisition (p = 0.013 and p = 0.026, respectively). Considering that a good tumour growth took place in only about 50% of transplanted animals (Fig. 6), we selected the three best growing tumors for each treatment group and report in Figure 5 photon count pictures at 14, 21 and 28 days after leukemic cells injection. It is of note that gross inspection and preliminary histological examination of mice organs like liver, kidneys and lungs did not show any sign of evident tissue suffering.
Discussion
Overexpression of Bcl-2 family anti-apoptotic proteins is a common feature in several tumors and, therefore, drugs mimicking the function of their pro-apoptotic counterparts, the BH3-only
Figure 2. 072RB inhibits leukaemia cell line growth by inducing apoptotic cell death. (A) Cell counts of U937 cell cultures treated for four days every 24 h at a 1 uM 072RB concentration and every 48 h at 5, 10 and 30 μM 072RB concentration. Arrows indicate time treatment for 072RB at concentra-tions ≥5 μM. Cells were counted in a Burker chamber and values are expressed as percentage of the untreated control cell number. Only live, trypan blue excluding cells were counted. Mean values (±SD) from triplicate experiments are reported. (B) U937, Jurkat and Namalwa cells are treated twice, every 48 h, with 10 μM 072RB and followed for four days. Cell counts are normalized to untreated control culture counts. (C) Flow cytometry measurements of DNA content (evaluated by PI fluorescence intensity) and physical laser light scattering (evaluated by side light scatter SSC) for U937 cells after 2 and 4 days treatment with 5 or 10 μM 072RB, or after 4 days with the internalization vector alone Int at 10 μM. Drugs were administered to the cultures every 48 h. Bivariate dot plots of DNA/SSC are reported in the first row and the respective DNA content univariate distributions underneath (projection of the dot plots on the x-axis). The percentage of cells with fractional DNA content (“sub-G1”) is reported, and provides information on the extent of apoptotic cell death. (D) The percentage of live and apoptotic (“sub-G1”) cells in leukaemia cell line cultures (U937, Jurkat and Namalwa) treated twice, at t = 0 and t = 48 h, with 072RB, Int alone or 072RB L94A, at the indicated concentrations, was assessed by flow cytometric DNA content histograms after 4 days (t = 96 h). At least three independent experiments were performed and mean ± SD values are reported. (E) U937 and Jurkat cells were treated once with 072RB, 072RB L94A, or Int (10 μM), and stained after 48 h with annexin V-FITC. The fraction of annexin V positive cells is reported.
Anti-leukemic activity of a new Bcl-XL inhibitor
3218 Cell Cycle 2008; Vol. 7 Issue 20
The resulting molecule, named 072RB, displayed a strong Ki towards [(BakF-BH3D84A):(Bcl-XL)] interaction (Ki = 4.3 nM). CD spectra analysis showed a doubling of α-helix content (from
helix of the Antennapedia protein, bearing the mutation Gln50Pro (Int) which is reported to mostly localize in the cytosol, rather than in the nucleus.47
Figure 3. For figure legend, see page 3219.

Anti-leukemic activity of a new Bcl-XL inhibitor
www.landesbioscience.com Cell Cycle 3219
Since 072RB lacks a mitochondrial localization sequence, we could speculate that it may specifically bind to its target on mitochondrial membrane. Cells seem not to sequester and degrade 072RB into lysosomes, at least within a 24 h observation period, since the peptide did not co-localize with lysosomes.
The apoptogenic features of 072RB were then investigated in in vitro, ex vivo and in vivo cell models. Leukemic cell lines underwent apoptosis in a dosage and time dependent fashion, as assessed by DNA flow cytometry, phosphatidylserine externaliza-tion, activation of caspases 9 and 3. Associated with 072RB-induced apoptosis, we recorded the dissipation of the inner mitochondrial membrane potential. It has to be determined whether it represents an upstream event triggered by our peptidomimetic, as recently described for chronic lymphocytic leukemia cells where the mito-chondrial inner membrane permeabilization induced by ABT-737 resulted in subsequent mitochondrial matrix swelling and rupture of the outer mitochondrial membrane.48 Alternatively, the collapse of mitochondrial trans-membrane potential could be an effect of the permeabilization of the outer mitochondrial membrane, which can be evinced in the 072RB-treated cells by Bax activation and rise of activated caspases 9 and 3. It was indeed demonstrated that caspase activation occurring after rupture of the outer mitochondrial membrane can lead to the cleavage of a subunit of complex I of the electron transport chain, that participates in the potential differ-ence across the inner mitochondrial membrane.49 Nevertheless, it is evident that 072RB leads to mitochondrial membrane permeabiliza-tion (MMP) that is the “point of no return” in the cascade of events leading to apoptosis.4
No cytotoxic activity was detected when cells were exposed to the point mutant 072RBL94A, in any of the experiments. This strengthens the high sequence specificity of our peptidomimetic inhibitor, since in the mutant peptide α-helicity remained substantially invariant and the mutant was even more stable (half-life in FBS at 37°C) than the parent 072RB molecule. This rules out that toxicity of 072RB is due to the relatively high helical content of the peptide47,53 or, else, to an unspecific cytotoxic activity of the antennapedia internal-izing sequence, as previously observed on one antennapedia-linked BAD-derived BH3-like peptide, that was however administered to cells at the high concentration of 50 μM.53
072RB was tested in vivo on a human leukaemia xenograft model. Notwithstanding the limited cohort of engrafted mice, the results suggest that 072RB mono-therapy, administered intravenously (only thrice, every 48 h, starting 48 h after the inoculum) at a dosage of 15 mg/Kg (3 μmol/Kg) inhibited or drastically slowed leukemic cell growth. When tested on a similar in vivo model, i.e., a human xeno-transplant of Raf-transformed myeloid cells, the synthetic small
15% to 30%) in 072RB structure, compared to Bim-BH3YA2Aib. It is likely that these structural and functional improvements are due, to some extent, to the α-helical Int contribution. The internalizing sequence seems not directly to participate in the complex formation, since it is fused to Bim-BH3YA2Aib C-terminus, which apparently lies outside the interaction region (dynamic molecular modelling data). Molecular modelling of the [(Bcl-XL):(Bim-BH3YA2Aib)] complex suggested that Asn102, Aib103 and Tyr104 (the last three aa immediately preceding Int) do not keep contact with the protein, being “too far” from the cleft (Cozzini P, Spiraki F, personal commu-nication).
Although Glu100Ala mutation lies within the Bim-BH3 motif, it did not affect BH3 derived peptide “behaviour”. 072RB inhibi-tion of [(BakF-BH3D84A):(Bcl-XL)] interaction dropped down only when the conserved residue Leu94, critical for the interaction with the hydrophobic cleft, was substituted with an Ala (a shorter aliphatic residue), as suggested from previous observations on the Bak BH3 motif.45 This means that in spite of all substitutions 072RB is still a “BH3-like peptide”.
Cellular uptake of 072RB by tumour cell lines was demonstrated to be fast and efficient. The peptidomimetic localized preferentially in the cytosol and in various organelles, especially mitochondria.
Figure 3. Apoptotic activity of 072RB involves Bax oligomerization, transmembrane potential dissipation, and caspase 9 and 3 activation. (A) Leukemic cell lines were incubated for 48 h with either 072RB (10 μM) or Int (10 μM) and activation of caspase 9 was analyzed by flow cytometry. The plot reports the percentage of cells bound by the fluorescent compound FITC-LEHD-FMK which is specific for active caspase 9. (B) Kinetics of activation of caspase 3 in leukemic cell lines incubated for the indicated periods of time with 10 μM 072RB or Int. The plot reports the percentage of cells bound by the fluorescent compound FITC-DEVD-FMK which is specific for active caspase 3. (C) U937 cells were treated with 072RB or 072RB L94A for the times indicated, processed for intracellular indirect immunofluorescence staining with anti-Bax clone 3 antibody specific for the active form of Bax and analyzed by flow cytometry. Bax histograms are gated on the “low Forward Light Scatter (FSC)/low Side Light Scatter (SSC)” cell population as shown in the inset, in order to exclude late apoptotic cells that might stain unspecifically. Thick lines: frequency histograms for Bax expression. Thin lines: histograms from samples stained with isotypic control antibody. (D) Flow cytometric DiOC6 fluorescence histograms of U937 cells treated with 072RB, 5 or 10 μM, respectively. Decreased fluorescence of the dye indicates dissipation of the mitochondrial transmembrane potential. The percentage of cells undergoing dissipation is reported. (E) Percentage of cells with reduced DiOC6 fluorescence in cultures of U937, Jurkat and Namalwa cell lines treated with 072RB, 072RBL94A or Int for 2 days. Results are expressed as the mean (±SD) from at least two independent experiments.
Table 2 Effect of 072RB on haematological malignancies: Percentage of viable cells
Int 072RB Malignancy FAB Control 5 μM 10 μM 5 μM 10 μM classificationAML M0 67 68 62 64 58AML M0–M1 86 - - 78 60AML M2 76 - - 34 -AML M2 48 - - 40 20AML M3 77 74 72 36 18AML M4–M5 50 - - 25 -AML M6 85 - - - 33ALL 57 - - 33 -ALL Pro-B 68 68 57 44 37NBM* 85* - 82* - 79*NBM 61 - 63 - 61
Ex vivo isolated cells were treated with either Int or 072RB at day 0 and 2 (5 and/or 10 μM concentration) and their level of viability measured by Propidium Iodide exclusion tests at day 4. Mean values from dupli-cate experiments are reported. The difference between the two individual values never exceeded 15% of the mean value. *This sample was treated with 15 μM both for Int and 072RB. (FAB, French-American-British classification; AML, acute myeloid leukemia; ALL, acute lymphoid leukemia; NBM, normal bone marrow).

Anti-leukemic activity of a new Bcl-XL inhibitor
3220 Cell Cycle 2008; Vol. 7 Issue 20
and ex vivo bone marrow-derived tumour cells from AML patients. The selectivity for tumour cells has been observed also for other BH3 mimetics, but is still unexplained. It could be envisaged that some cancer cells have a higher degree of “addiction” to a prevalently anti-apoptotic balance than normal cells do.9
Altogether, we may conclude that the peptidic BIM-derived BH3 mimetic molecule 072RB displays promising apoptogenic features and a differential sensitivity in cancer vs. normal cells. Nonetheless, only preliminary data are available as for its mechanisms of action and target specificity. 072RB was selected among different Bim-derived BH3-like peptides based on its maximal inhibitory activity against [(BakF-BH3D84A):(Bcl-XL)] interaction. However, it killed cells that markedly express also Bcl-2 and Mcl-1, in addition to Bcl-XL, according to flow cytometric intracellular immunodetec-tion in the leukaemia cell lines here utilized (Bruno S and Poggi A, personal communication). Bim is a promiscuous BH3-only protein, targeting all multi-domain proteins of the Bcl-2 family, and in silico alignments revealed that all interaction hot spots between 072RB and Bcl-XL are conserved also in Bcl-2 and Mcl-1 sequences (see molecular modelling results in supplementary materials). Therefore, it has to be experimentally determined whether 072RB interacts also with the BH3-binding cleft of Bcl-2 and Mcl-1, and thus kills cells via interaction with all Bcl-2-like anti-apoptotic proteins. Moreover,
molecule ABT-737 required heavier treatment schedules to achieve levels of leukaemia tumour burden suppression comparable to ours.17 ABT-737 had to be given on a continuous daily schedule for the entire observation period, at doses of 20–30 mg/kg. We believe that the good serum stability of 072RB might be, at least in part, responsible for its higher antitumor efficacy. In addition, ABT-737 could only be given by i.p. injection, because of its low aqueous solubility that hampers i.v. delivery. On the contrary, 072RB displays good water solubility because of its amino-acidic sequence. This limitation has been very recently overcome by the second generation ABT compound ABT-263, that is orally bio-available and inhibits human xeno-transplant growth of small-cell lung carcinoma, but not of B-cell lymphoma and multiple myeloma, as a single agent, when given daily at 100 mg/kg.21
No tissue toxicity was observed by histologic examinations of organs like liver, kidneys and lungs from mice treated with 072RB at doses up to 25 mg/kg. Likewise, the mice were found in good general conditions by gross inspection. Thus, it seems that 072RB is much more active on cancer cells than on the normal tissues of treated animals. This is further confirmed by data on human ex-vivo normal hemopoietic cells, either derived from the bone marrow or from the peripheral blood, that were almost unaffected by 072RB at the same treatment schedule shown to be toxic against both leukemic cell lines
Figure 4. 072RB exerts negligible effects on human normal lymphocytes. Quiescent (A), or PHA stimulated peripheral lymphocytes (B and C). After 4 days treatment (twice, at t = 0 and t = 48 h, 072RB 10 μM) the lymphocytes were harvested and processed for flow cytometric DNA content measurements. The percentage of non apoptotic cells (A and B) and the percentage of cells in the [S + G2M] cell cycle phases (C) are reported. Experiments on each donor were carried out in triplicate, and column bars represent the mean values. Error bars are not reported since the difference between maximum and minimum values never exceeded 10% of the mean value.

Anti-leukemic activity of a new Bcl-XL inhibitor
www.landesbioscience.com Cell Cycle 3221
RPMI 1640 medium supplemented with 10% FBS, glutamine and antibiotics.
Human peripheral blood lymphocytes from healthy volunteers were isolated by lymphocyte separation medium (Ficoll) density gradient centrifugation. Mononuclear cells from the interphase were washed twice with PBS and resuspended in RPMI 1640 medium containing 10% FCS, glutamine and antibiotics. Phytohemagglutinin (PHA) was added at 1.5 μg/ml final concentration.
We obtained, after patients informed consent and clinical approval, from bone marrow aspirates that were collected for diag-nostic procedures, bone marrow derived cells, either from patients with Acute Lymphoblastic Leukemia (ALL) and Acute Myeloid Leukemia (AML), or from healthy volunteers. Pathologic samples containing at least 80% blasts among their mononuclear cells were selected. Mononuclear cells were isolated by density centrifugation over Ficoll. Primary cells were cultured as above.
For total body luminescence experiments, U937 cells exhibiting stable luciferase expression (U937-luc) were generated as follows. For the retroviral plasmid production, the firefly luciferase gene
it cannot be excluded that 072RB prompts apoptosis also by directly binding and activating Bax and Bak, according to the “direct acti-vation” model of mitochondrial cell death, and described for a hydrocarbon-stapled BID-derived BH3 domain that bears a stabi-lized α-helix and triggers apoptosis by directly binding and activating Bax.23,24 The relevant degree of α-helicity observed for our modified Bim-like BH3 peptide (even before its interaction with the target) directs future investigations also along this latter hypothesis.
Materials and Methods
Technical details about Molecular Modelling, expression and purification of the recombinant Bcl-XL protein, synthesis and modification of our Bim BH3-like peptides, their analysis by Mass Spectrometry, Circular Dichroism and Fluorescence Anisotropy assays, and the evaluation of their stability in tissue culture media and sera, are all provided in the Supplemental Data section.
Cell lines, clinical samples and xenograft cells. The leukemic myelomonocytic cell line U937, the T cell leukaemia cell line Jurkat, the Burkitt lymphoma cell line Namalwa, were grown in
Figure 5. 072RB activity on a human leukaemia xenograft model. NOD/SCID mice injected with U937luc+ cells, and imaged in vivo at days 14, 21, 28 after cells injection. Controls: PBS treated mice. Int or 072RB: mice treated with internalizing sequence alone or 072RB peptide, respectively (cumulative dosage for three treatments: 9 μmol/Kg). Three animals with maximal tumour growth are shown for each group. The signal intensity is quantified within manual ROIs over the whole mouse, as the flux of all photons/s/cm2/sr. Acquisition time: 300 sec.

Anti-leukemic activity of a new Bcl-XL inhibitor
3222 Cell Cycle 2008; Vol. 7 Issue 20
was obtained as a SalI/BglII fragment from pGL3-control vector (Promega), and cloned into the XhoI/BamHI sites of the retroviral pLXIN bicistronic vector (Clontech), (previously modified inserting the restriction sites NotI, BglII, XhoI, ClaI, SalI, BstxI, BamHI into the multiple cloning site) to obtain pL-Luc-IN. Retroviruses were produced as reported by Finer et al.51 and used to infect U937 cells with 8 μg/ml Polybrene (Sigma). After 48 h, transduced cells were selected with G418 (Calbiochem).
Flow cytometry analysis of peptides uptake. Cell lines were incubated with different concentrations of fluoresceinated peptides (0.1, 1.0, 10 μM) for 30 min, 60 min, 120 min, 180 min, 18 h, 24 h, 48 h, 72 h, in complete tissue culture medium at 37°C, and subsequently analysed on a flow cytometer (FACSort, Becton Dickinson). Data are expressed as Log green fluorescence intensity (arbitrary units, a.u.) frequency histograms.
Confocal microscopy of 072RB intracellular localization. Cell lines were incubated with different concentrations of fluoresceinated peptides for increasing time lengths in tissue culture medium at 37°C. During the last 30 min cells were incubated with either Mitotracker Red 580 (MT, 100 nM) or Lysotracker red DND-99 (LTR, 50 nM) (Molecular Probes, Poortgebouw, Leiden, The Netherlands). A 50 μl cell suspension volume was seeded on a micro-slide, covered with a 5 cm2 cover slip (we formed a sealed micro-chamber 0.1 mm deep). We analysed our samples with an IX81 confocal microscope equipped with a 488 nm argon laser and a He-Neon 543 nm laser (FV500, Olympus BioSystem). Ten to fifteen fields were analysed with 40x oil/1.35 NA or 60x oil/1.40 NA objectives and images acquired using the FlowView 4.2b computer program. We compared relative concentrations of our fluoresceinated-peptides, present in different intracellular compartments, with the concentration present in the extra-cellular compartment (where fluoresceinated-peptide concentration is known). Results were plotted as mean fluorescence intensity (MFI) expressed in arbitrary units (a.u.).
DNA flow cytometry for apoptosis and cell proliferation evaluation. Cells were permeabilized with 0.05% Triton X-100 and incubated with 30 μg/ml PI (Propidium Iodide) (Sigma Chemical Co.) and 0.5 mg/ml RNase for 30 min at room temperature and in the dark. Flow cytometric measurements of excitation laser Forward Light Scatter (FSC) and Side Light Scatter (SSC), and of PI emission fluorescence, were performed on a FACSCalibur (Becton Dickinson) flow cytometer. At least 10,000 events were acquired on a linear scale from each sample. Frequency distributions of DNA content were analysed for the evaluation of proliferation and apoptosis as previ-ously described.52
Annexin V and PI exclusion assays. Apoptosis-associated cellular externalization of phosphatidylserine molecules and plasma membrane damage were assessed by flow cytometric measurements of annexin V and PI uptake, respectively. To this end, cells were stained with annexin V-FITC and PI, according to standard manu-facturer instruction (BioVision, Mountain View, CA). For bone marrow derived specimens, cell death was assessed by PI uptake only, after 5 minutes of 1 μg/ml PI incubation at room temperature. Cells were run on a FACSCalibur (Becton Dickinson) flow cytometer. For each sample 10,000 events were collected on a logaritmic scale, both for annexin V-FITC and PI fluorescence.
Detection of mitochondrial permeability transition. Induction of mitochondrial permeability transition (MPT) was evaluated by
Figure 6. In vivo imaging data analysis. Plots of bioluminescence assessed at days 14, 21 and 28 for each animal. Bioluminescence values are expressed as: Log[(photons/mouse at 14 or 21 or 28 days acquisition)/(photons medi-an 14 days acquisition)]. (A) Control (PBS) animals; (B) Int treated animals; (C) 072RB treated animals.

Anti-leukemic activity of a new Bcl-XL inhibitor
www.landesbioscience.com Cell Cycle 3223
adopted for obtaining our Kd and Ki estimations. They are described in detail in the SUPPLEMENTAL MATERIAL section.
A Kd value for the reference reversible interaction [(BakF-BH3D84A):(Bcl-XL)] was calculated using a nonlinear regression analysis [NLIN SAS] to fit the Lundblad equation.53 For our Ki estimations we calculated the expected Anisotropy values solving a third degree equation obtained extending the simple binding equa-tion to a competitive binding-inhibition assay.54 To analyze the results of the competitive binding inhibition assay, we performed a mixed parametric/non-parametric bootstrap analysis.55 Comparing experimental data with expected anisotropy, we obtained a Ki value minimizing the Sum of Squared Deviations (SSD) for the bootstrap sample. Point estimates were evaluated weighting each experimental result for its estimated variance. Standard error values reported in Table 1 are computed as squared roots of pooled variance.
Statistical analysis of in vivo results. A non-parametric one-tailed Mann-Whitney U-test (performed using GraphPad Prism version 4.00 for Windows, GraphPad Software, San Diego California USA, www.graphpad.com) was applied to compare in vivo biolumines-cence data, for PBS, Int and 072RB treated animals, at 21 and 28 days after tumour cells inoculum. A probability value of p < 0.05 was considered significant.
Acknowledgements
This work was supported from: MIUR PRIN 2005-prot. 2005061408_005; national project Compagnia di San Paolo 2489 IT/CV, 2006.1997, Genoa Operative Unit (O.U.); Regione Liguria, Cap. 5296 F.S.R. Nr. Prov. 23, Prot. Gen. 616, 2007; Istituto Superiore di Oncologia, O.U.s of an “Istituto Oncologico Veneto” project, and a “Regione Campania project; project Compagnia di San Paolo 4998 IT/CV, 2007.0087, Genoa University.
Note
Supplemental materials can be found at:www.landesbioscience.com/supplement/PonassiCC7-20-Sup.pdf
References 1. Reed JC. Dysregulation of apoptosis in cancer. J Clin Oncol 1999; 17:2941-53. 2. Johnstone RW, Ruefli AA, Lowe SW. Apoptosis: a link between cancer genetics and chemo-
therapy. Cell 2002; 108:153-64. 3. Youle RJ, Strasser A. The BCL-2 protein family: opposing activities that mediate cell death.
Nat Rev Mol Cell Biol 2008; 9:47-59. 4. Green DR, Kroemer G. The pathophysiology of mitochondrial cell death. Science 2004;
305:626-9. 5. Willis SN, Chen L, Dewson G, Wei A, Naik E, Fletcher JI, Adams JM, Huang DC.
Proapoptotic Bak is sequestered by Mcl-1 and Bcl-xL, but not Bcl-2, until displaced by BH3-only proteins. Genes Dev 2005; 19:1294-305.
6. Willis SN, Fletcher JI, Kaufmann T, van Delft MF, Chen L, Czabotar PE, Ierino H, Lee EF, Fairlie WD, Bouillet P, Strasser A, Kluck RM, Adams JM, Huang DC. Apoptosis initi-ated when BH3 ligands engage multiple Bcl-2 homologs, not Bax or Bak. Science 2007; 315:856-9.
7. Kuwana T, Bouchier-Hayes L, Chipuk JE, Bonzon C, Sullivan BA, Green DR, Newmeyer DD. BH3 domains of BH3-only proteins differentially regulate Bax-mediated mitochondrial membrane permeabilization both directly and indirectly. Mol Cell 2005; 17:525-35.
8. Kim H, Rafiuddin-Shah M, Tu HC, Jeffers JR, Zambetti GP, Hsieh JJ, Cheng EH. Hierarchical regulation of mitochondrion-dependent apoptosis by BCL-2 subfamilies. Nat Cell Biol 2006; 8:1348-58.
9. Certo M, Del Gaizo Moore V, Nishino M, Wei G, Korsmeyer S, Armstrong SA, Letai A. Mitochondria primed by death signals determine cellular addiction to antiapoptotic BCL-2 family members. Cancer Cell 2006; 9:351-65.
10. Chen L, Willis SN, Wei A, Smith BJ, Fletcher JI, Hinds MG, Colman PM, Day CL, Adams JM, Huang DC. Differential targeting of prosurvival Bcl-2 proteins by their BH3-only ligands allows complementary apoptotic function. Mol Cell 2005; 17:393-403.
11. Kitada S, Leone M, Sareth S, Zhai D, Reed JC, Pellecchia M. Discovery, characterization, and structure-activity relationships studies of proapoptotic polyphenols targeting B-cell lymphocyte/leukemia-2 proteins. J Med Chem 2003; 46:4259-64.
the decrease in mitochondrial inner membrane potential. Briefly, cells were incubated with 40 nM 3,3'-dihexyloxacarbocyanine iodide (DiOC6) for 15 min at 37°C and analysed by a FACSCalibur (BD Biosciences) equipped with a 488 nm laser suitable for DiOC6 excitation and cut-on optical filters suitable for DiOC6 emission-fluorescence (peak at 552 nm). At least 10,000 events were acquired per sample on a logarithmic scale. Dissipation of the mitochondrial trans-membrane potential causes a shift of DiOC6 fluorescence to lower levels.
Detection of Bax conformational changes. Cells were fixed with 2% paraformaldehyde and permeabilized with 0.1% saponin, incu-bated in the presence of 2 μg/ml of anti-Bax clone 3 monoclonal antibody specific for the active form of Bax (BD Biosciences). After 30 min at room temperature, the cells were washed and incubated with Alexa Fluor 488-conjugated Goat anti-Mouse IgG (Molecular Probes), counterstained with DAPI and either cytospun on cover-splips and mounted with Mowiol (for microscopy imaging on the laser scanning spectral confocal microscope TCS SP2 AOBS from Leica (Heidelberg, Germany)) or directly measured on the FacsCalibur flow cytometer (BD Biosciences), after verifying that no DAPI contribution was collected by the photomultiplier FL-1.
Evaluation of caspases 9 and 3 activation and apoptosis. Caspase 9 and caspase 3 activation was evaluated using the caspase-9 and caspase-3 Detection Kit (Calbiochem, Darmstadt, Germany). Samples were treated as reported in the data sheet and run on a flow cytometer (FACSort, Becton Dickinson). At least 10,000 events were analysed for each sample using the Cell Quest computer program. Data are expressed as Log green fluorescence intensity (a.u.) frequency histograms.
Human leukemic xenografts mice and in vivo bioluminescence imaging. The experiment involving animals were reviewed and approved by Licensing and Ethical Committee of the National Cancer Research Institute, Genoa, Italy and by the Italian Ministry of Health. 30 NOD-SCID female mice (4 weeks old, Charles River Laboratories), housed under specific pathogen-free conditions, were injected intravenously (i.v.) through the tail vein with 5 x 106 U937-luc. Starting on day 2 after cancer cells injection, mice were treated as follows: peptides dissolved in PBS pH 7.3 were adminis-tered i.v. every 48 h for three times. Six mice received the 072RB peptide at the dose of 3 μmol/Kg [15 mg/Kg body weight], six mice the internalizing sequence Int at 3 μmol/Kg [6,8 mg/Kg body weight], and eight mice were PBS treated (controls). At days 14, 21, 28 after injection, mice were imaged for leukemic cells spreading. To this end the mice were anesthetized with 2.5% isofluorane in oxygen, delivered via a nose cone system, and treated with 15 mg/ml of D-luciferin firefly (potassium salt from Xenogen, Almeda, CA USA) in sterile PBS, intraperitoneally injected at a dose of 150 mg/Kg body weight. In vivo imaging was performed with the IVIS 100 animal imaging system (Xenogen, Almeda, CA USA) using the Living Image software version 2.50 (Xenogen), along with IGOR pro (Wawe-Metrics, Seattle, WA USA). Acquisition parameters are: Bin: M (8), FOV25 and time of acquisition 300 seconds. The signal intensity was quantified using the Living Image software package, within manual Regions Of Interest (ROIs) over the whole mouse, and expressed as the flux of photons/s/cm2/sr.
Computation of Kd and Ki values from fluorescence anisotropy binding experiments. We briefly summarize here the procedures we

Anti-leukemic activity of a new Bcl-XL inhibitor
3224 Cell Cycle 2008; Vol. 7 Issue 20
34. Tan TT, Degenhardt K, Nelson DA, Beaudoin B, Nieves-Neira W, Bouillet P, Villunger A, Adams JM, White E. Key roles of BIM-driven apoptosis in epithelial tumors and rational chemotherapy. Cancer Cell 2005; 7:227-38.
35. Adams JM, Cory S. Bcl-2-regulated apoptosis: mechanism and therapeutic potential. Curr Opin Immunol 2007; 19:488-96.
36. Danial NN, Korsmeyer SJ. Cell death: critical control points. Cell 2004; 116:205-19. 37. Minn AJ, Rudin CM, Boise LH, Thompson CB. Expression of bcl-xL can confer a multi-
drug resistance phenotype. Blood 1995; 86:1903-10. 38. Amundson SA, Myers TG, Scudiero D, Kitada S, Reed JC, Fornace AJ Jr. An informatics
approach identifying markers of chemosensitivity in human cancer cell lines. Cancer Res 2000; 60:6101-10.
39. Martin SS, Ridgeway AG, Pinkas J, Lu Y, Reginato MJ, Koh EY, Michelman M, Daley GQ, Brugge JS, Leder P. A cytoskeleton-based functional genetic screen identifies Bcl-xL as an enhancer of metastasis, but not primary tumor growth. Oncogene 2004; 23:4641-5.
40. Darzynkiewicz Z, Huang X, Okafuji M, King MA. Cytometric methods to detect apoptosis. Methods Cell Biol 2004; 75:307-41.
41. Grabarek J, Du L, Johnson GL, Lee BW, Phelps DJ, Darzynkiewicz Z. Sequential activation of caspases and serine proteases (serpases) during apoptosis. Cell Cycle 2002; 1:124-31.
42. Pozarowski P, Huang X, Halicka DH, Lee B, Johnson G, Darzynkiewicz Z. Interactions of fluorochrome-labeled caspase inhibitors with apoptotic cells: a caution in data interpreta-tion. Cytometry A 2003; 55:50-60.
43. Kroemer G, Galluzzi L, Brenner C. Mitochondrial membrane permeabilization in cell death. Physiol Rev 2007; 87:99-163.
44. Shangary S, Oliver CL, Tillman TS, Cascio M, Johnson DE. Sequence and helicity require-ments for the proapoptotic activity of Bax BH3 peptides. Mol Cancer Ther 2004; 3:1343-54.
45. Sattler M, Liang H, Nettesheim D, Meadows RP, Harlan JE, Eberstadt M, Yoon HS, Shuker SB, Chang BS, Minn AJ, Thompson CB, Fesik SW. Structure of Bcl-xL-Bak peptide complex: recognition between regulators of apoptosis. Science 1997; 275:983-6.
46. Hinds MG, Smits C, Fredericks-Short R, Risk JM, Bailey M, Huang DC, Day CL. Bim, Bad and Bmf: intrinsically unstructured BH3-only proteins that undergo a localized con-formational change upon binding to prosurvival Bcl-2 targets. Cell Death Differ 2007; 14:128-36.
47. Derossi D, Calvet S, Trembleau A, Brunissen A, Chassaing G, Prochiantz A. Cell internal-ization of the third helix of the Antennapedia homeodomain is receptor-independent. J Biol Chem 1996; 271:18188-93.
48. Vogler M, Dinsdale D, Sun XM, Young KW, Butterworth M, Nicotera P, Dyer MJ, Cohen GM. A novel paradigm for rapid ABT-737-induced apoptosis involving outer mitochondrial membrane rupture in primary leukemia and lymphoma cells. Cell Death Differ 2008; 15:820-30.
49. Ricci JE, Munoz-Pinedo C, Fitzgerald P, Bailly-Maitre B, Perkins GA, Yadava N, Scheffler IE, Ellisman MH, Green DR. Disruption of mitochondrial function during apoptosis is medi-ated by caspase cleavage of the p75 subunit of complex I of the electron transport chain. Cell 2004; 117:773-86.
50. Schimmer AD, Hedley DW, Chow S, Pham NA, Chakrabartty A, Bouchard D, Mak TW, Trus MR, Minden MD. The BH3 domain of BAD fused to the Antennapedia peptide induces apoptosis via its alpha helical structure and independent of Bcl-2. Cell Death Differ 2001; 8:725-33.
51. Finer MH, Dull TJ, Qin L, Farson D, Roberts MR. kat: a high-efficiency retroviral trans-duction system for primary human T lymphocytes. Blood 1994; 83:43-50.
52. Bruno S, Del Bino G, Lassota P, Giaretti W, Darzynkiewicz Z. Inhibitors of proteases pre-vent endonucleolysis accompanying apoptotic death of HL-60 leukemic cells and normal thymocytes. Leukemia 1992; 6:1113-20.
53. Lundblad JR, Laurance M, Goodman RH. Fluorescence polarization analysis of protein-DNA and protein-protein interactions. Mol Endocrinol 1996; 10:607-12.
54. Zhang R, Mayhood T, Lipari P, Wang Y, Durkin J, Syto R, Gesell J, McNemar C and Windsor W. Fluorescence polarization assay and inhibitor design for MDM2/p53 interac-tion. Analytical Biochemistry 2004; 331:138-46.
55. Davison AC, Hinkley DV. Bootstrap methods and their application. Cambridge University Press, Cambridge, United Kingdom 1997.
56. Liu X, Dai S, Zhu Y, Marrack P, Kappler JW. The structure of a Bcl-xL/Bim fragment complex: implications for Bim function. Immunity 2003; 19:341-52.
12. Wang JL, Liu D, Zhang ZJ, Shan S, Han X, Srinivasula SM, Croce CM, Alnemri ES, Huang Z. Structure-based discovery of an organic compound that binds Bcl-2 protein and induces apoptosis of tumor cells. Proc Natl Acad Sci USA 2000; 97:7124-9.
13. van Delft MF, Wei AH, Mason KD, Vandenberg CJ, Chen L, Czabotar PE, Willis SN, Scott CL, Day CL, Cory S, Adams JM, Roberts AW, Huang DC. The BH3 mimetic ABT-737 targets selective Bcl-2 proteins and efficiently induces apoptosis via Bak/Bax if Mcl-1 is neutralized. Cancer Cell 2006; 10:389-99.
14. Konopleva M, Watt J, Contractor R, Tsao T, Harris D, Estrov Z, Bornmann W, Kantarjian H, Viallet J, Samudio I, Andreeff M. Mechanisms of antileukemic activity of the novel Bcl-2 homology domain-3 mimetic GX15-070 (obatoclax). Cancer Res 2008; 68:3413-20.
15. Trudel S, Li ZH, Rauw J, Tiedemann RE, Wen XY, Stewart AK. Preclinical studies of the pan-Bcl inhibitor obatoclax (GX015-070) in multiple myeloma. Blood 2007; 109:5430-8.
16. Oltersdorf T, Elmore SW, Shoemaker AR, Armstrong RC, Augeri DJ, Belli BA, Bruncko M, Deckwerth TL, Dinges J, Hajduk PJ, Joseph MK, Kitada S, Korsmeyer SJ, Kunzer AR, Letai A, Li C, Mitten MJ, Nettesheim DG, Ng S, Nimmer PM, O’Connor JM, Oleksijew A, Petros AM, Reed JC, Shen W, Tahir SK, Thompson CB, Tomaselli KJ, Wang B, Wendt MD, Zhang H, Fesik SW, Rosenberg SH. An inhibitor of Bcl-2 family proteins induces regression of solid tumors. Nature 2005; 435:677-81.
17. Konopleva M, Contractor R, Tsao T, Samudio I, Ruvolo PP, Kitada S, Deng X, Zhai D, Shi YX, Sneed T, Verhaegen M, Soengas M, Ruvolo VR, McQueen T, Schober WD, Watt JC, Jiffar T, Ling X, Marini FC, Harris D, Dietrich M, Estrov Z, McCubrey J, May WS, Reed JC, Andreeff M. Mechanisms of apoptosis sensitivity and resistance to the BH3 mimetic ABT-737 in acute myeloid leukemia. Cancer Cell 2006; 10:375-88.
18. Del Gaizo Moore V, Brown JR, Certo M, Love TM, Novina CD, Letai A. Chronic lym-phocytic leukemia requires Bcl-2 to sequester prodeath BIM, explaining sensitivity to Bcl-2 antagonist ABT-737. J Clin Invest 2007; 117:112-21.
19. Trudel S, Stewart AK, Li Z, Shu Y, Liang SB, Trieu Y, Reece D, Paterson J, Wang D, Wen XY. The Bcl-2 family protein inhibitor, ABT-737, has substantial antimyeloma activity and shows synergistic effect with dexamethasone and melphalan. Clin Cancer Res 2007; 13:621-9.
20. Kang MH, Kang YH, Szymanska B, Wilczynska-Kalak U, Sheard MA, Harned TM, Lock RB, Reynolds CP. Activity of vincristine, L-ASP, and dexamethasone against acute lympho-blastic leukemia is enhanced by the BH3-mimetic ABT-737 in vitro and in vivo. Blood 2007; 110:2057-66.
21. Tse C, Shoemaker AR, Adickes J, Anderson MG, Chen J, Jin S, Johnson EF, Marsh KC, Mitten MJ, Nimmer P, Roberts L, Tahir SK, Xiao Y, Yang X, Zhang H, Fesik S, Rosenberg SH, Elmore SW. ABT-263: a potent and orally bioavailable Bcl-2 family inhibitor. Cancer Res 2008; 68:3421-8.
22. Lee EF, Czabotar PE, van Delft MF, Michalak EM, Boyle MJ, Willis SN, Puthalakath H, Bouillet P, Colman PM, Huang DC, Fairlie WD. A novel BH3 ligand that selectively tar-gets Mcl-1 reveals that apoptosis can proceed without Mcl-1 degradation. J Cell Biol 2008; 180:341-55.
23. Walensky LD, Kung AL, Escher I, Malia TJ, Barbuto S, Wright RD, Wagner G, Verdine GL, Korsmeyer SJ. Activation of apoptosis in vivo by a hydrocarbon-stapled BH3 helix. Science 2004; 305:1466-70.
24. Walensky LD, Pitter K, Morash J, Oh KJ, Barbuto S, Fisher J, Smith E, Verdine GL, Korsmeyer SJ. A stapled BID BH3 helix directly binds and activates BAX. Mol Cell 2006; 24:199-210.
25. Labi V, Erlacher M, Kiessling S, Villunger A. BH3-only proteins in cell death initiation, malignant disease and anticancer therapy. Cell Death Differ 2006; 13:1325-38.
26. Puthalakath H, O’Reilly LA, Gunn P, Lee L, Kelly PN, Huntington ND, Hughes PD, Michalak EM, McKimm-Breschkin J, Motoyama N, Gotoh T, Akira S, Bouillet P, Strasser A. ER stress triggers apoptosis by activating BH3-only protein Bim. Cell 2007; 129:1337-49.
27. Tagawa H, Karnan S, Suzuki R, Matsuo K, Zhang X, Ota A, Morishima Y, Nakamura S, Seto M. Genome-wide array-based CGH for mantle cell lymphoma: identification of homozygous deletions of the proapoptotic gene BIM. Oncogene 2005; 24:1348-58.
28. Bouillet P, Purton JF, Godfrey DI, Zhang LC, Coultas L, Puthalakath H, Pellegrini M, Cory S, Adams JM, Strasser A. BH3-only Bcl-2 family member Bim is required for apoptosis of autoreactive thymocytes. Nature 2002; 415:922-6.
29. Enders A, Bouillet P, Puthalakath H, Xu Y, Tarlinton DM, Strasser A. Loss of the pro-apoptotic BH3-only Bcl-2 family member Bim inhibits BCR stimulation-induced apoptosis and deletion of autoreactive B cells. J Exp Med 2003; 198:1119-26.
30. Anderton E, Yee J, Smith P, Crook T, White RE, Allday MJ. Two Epstein-Barr virus (EBV) oncoproteins cooperate to repress expression of the proapoptotic tumour-suppressor Bim: clues to the pathogenesis of Burkitt’s lymphoma. Oncogene 2008; 27:421-33.
31. Zhang W, Konopleva M, Ruvolo VR, McQueen T, Evans RL, Bornmann WG, McCubrey J, Cortes J, Andreeff M. Sorafenib induces apoptosis of AML cells via Bim-mediated activa-tion of the intrinsic apoptotic pathway. Leukemia 2008; 22:808-18.
32. Erlacher M, Michalak EM, Kelly PN, Labi V, Niederegger H, Coultas L, Adams JM, Strasser A, Villunger A. BH3-only proteins Puma and Bim are rate-limiting for gamma-radiation- and glucocorticoid-induced apoptosis of lymphoid cells in vivo. Blood 2005; 106:4131-8.
33. Iglesias-Serret D, de Frias M, Santidrian AF, Coll-Mulet L, Cosialls AM, Barragan M, Domingo A, Gil J, Pons G. Regulation of the proapoptotic BH3-only protein BIM by glucocorticoids, survival signals and proteasome in chronic lymphocytic leukemia cells. Leukemia 2007; 21:281-7.




















