
A distinctive repertoire of cathepsins is expressed by juvenile invasive Fasciola hepatica
15
Research paper A distinctive repertoire of cathepsins is expressed by juvenile invasive Fasciola hepatica Martı ´n Cancela a,b , Daniel Acosta c , Gabriel Rinaldi a , Edileusa Silva b , Rosario Dura ´n d , Leda Roche a , Arnaldo Zaha b , Carlos Carmona c , Jose F. Tort a, * a Departamento de Gene ´tica, Facultad de Medicina, Universidad de la Repu ´ blica, UDELAR, Gral. Flores 2125, CP 11800, Montevideo, Uruguay b Laboratorio de Ce ´stodos, Centro de Biotecnologı ´a, Universidad Federal de Rio Grande do Sul, UFRGS, Porto Alegre, Brazil c Unidad de Biologı ´a Parasitaria, Instituto de Higiene, Facultad de Ciencias, Universidad de la Repu ´ blica, UDELAR, Av. Alfredo Navarro 3051, CP 11600, Montevideo, Uruguay d Laboratorio de Bioquı ´mica Analı ´tica, Instituto de Investigaciones Biolo ´ gicas Clemente Estable, Av. Italia Montevideo, Uruguay Received 14 January 2008; accepted 28 April 2008 Available online 5 June 2008 Abstract Secreted cysteine proteases are relevant actors in parasite biology, taking part in critical host colonization roles such as traversing tissue barriers, immune evasion and nutrient digestion. In the trematode Fasciola hepatica, the initial step to successful infection of the mammalian host is the excystment of metacercariae and the invasion through the intestinal wall by the newly excysted juveniles (NEJ). While the cathepsin L-like cysteine proteinases secreted by the adult fluke have been extensively characterized, the cataloguing and description of the cathepsins B and L reported in the invasive stages is only sketchy. To identify the cathepsins expressedduring excystment and early invasion we constructed cDNA libraries encoding NEJ cathepsins B and L. We found two cathepsin L-like cysteine proteinases (CL3, CL4) and three cathepsins B (CB1, CB2, CB3) which are predominantly expressed in NEJ. Phylogenetic analysis showed that NEJ-expressed cathepsins L constitute a well-defined clade separate from the adult enzymes. Excystment induction resulted in a significant increment in activity towards cathepsin-specific fluoro- genic substrates in metacercariae homogenates, consistent with the detection of precursor and mature forms of cathepsins B and L before and after induction. In NEJ culture supernatants, protein and relative activity profiles show subtle changes during the first 48 h, with prevalence of cathepsin L-like activity, although cathepsins CB3 and CL3 were detected by mass spectrometry. Noticeably, the hydrolysis of a substrate with proline in the P2 position was predominant, a property only shared with adult CL2 and vertebrate cathepsin K among the C1A subfamily of cysteine proteases. Collectively these mRNA, protein and enzymatic data demonstrate the existence of a NEJ-specific repertoire of cathepsins expressed early in invasion, distinct to those used by other trematodes, potentially relevant for specific vaccine and chemotherapy design. The diversity of proteases employed by trematodes in the invasion process is discussed. Ó 2008 Elsevier Masson SAS. All rights reserved. Keywords: Cathepsins; Trematode; Invasion; Fasciola hepatica; Cercaria 1. Introduction In order to complete their life cycle, parasitic organisms have to gain access to their definitive location within their hosts, while avoiding the defense mechanisms imposed upon them. At the same time they are committed to obtain the necessary resources from their hosts. Diverse molecules and biochemical pathways have evolved to perform some of these processes, with proteolytic enzymes being major components of the parasitic way of life. Accumulated evidence from Abbreviations: NEJ, newly excysted juveniles; E/S, excreted and/or se- creted products; SDSePAGE, sodium dodecyl sulfateepolyacrylamide gel electrophoresis; AMC, 7-amino-4-methylcoumarin; ORF, open reading frame; RT, reverse transcription; BSA, bovine serum albumin; MSE, metacercariae somatic extract; HEPES, 4-(2-hydroxyethyl)-1-piperazine ethanesulfonic acid; DTT, 1,4-dithiothreitol; MALDI-TOF, matrix-assisted laser desorption ionization time of flight. * Corresponding author. Tel.: þ598 2 924 9562; fax: þ598 2 924 9563. E-mail address: [email protected] (J.F. Tort). 0300-9084/$ - see front matter Ó 2008 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.biochi.2008.04.020 Available online at www.sciencedirect.com Biochimie 90 (2008) 1461e1475 www.elsevier.com/locate/biochi
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of A distinctive repertoire of cathepsins is expressed by juvenile invasive Fasciola hepatica

Available online at www.sciencedirect.com
Biochimie 90 (2008) 1461e1475www.elsevier.com/locate/biochi
Research paper
A distinctive repertoire of cathepsins is expressed by juvenileinvasive Fasciola hepatica
Martın Cancela a,b, Daniel Acosta c, Gabriel Rinaldi a, Edileusa Silva b, Rosario Duran d,Leda Roche a, Arnaldo Zaha b, Carlos Carmona c, Jose F. Tort a,*
a Departamento de Genetica, Facultad de Medicina, Universidad de la Republica, UDELAR, Gral. Flores 2125, CP 11800, Montevideo, Uruguayb Laboratorio de Cestodos, Centro de Biotecnologıa, Universidad Federal de Rio Grande do Sul, UFRGS, Porto Alegre, Brazil
c Unidad de Biologıa Parasitaria, Instituto de Higiene, Facultad de Ciencias, Universidad de la Republica, UDELAR, Av. Alfredo Navarro 3051,
CP 11600, Montevideo, Uruguayd Laboratorio de Bioquımica Analıtica, Instituto de Investigaciones Biologicas Clemente Estable, Av. Italia Montevideo, Uruguay
Received 14 January 2008; accepted 28 April 2008
Available online 5 June 2008
Abstract
Secreted cysteine proteases are relevant actors in parasite biology, taking part in critical host colonization roles such as traversing tissuebarriers, immune evasion and nutrient digestion. In the trematode Fasciola hepatica, the initial step to successful infection of the mammalianhost is the excystment of metacercariae and the invasion through the intestinal wall by the newly excysted juveniles (NEJ). While the cathepsinL-like cysteine proteinases secreted by the adult fluke have been extensively characterized, the cataloguing and description of the cathepsins Band L reported in the invasive stages is only sketchy. To identify the cathepsins expressed during excystment and early invasion we constructedcDNA libraries encoding NEJ cathepsins B and L. We found two cathepsin L-like cysteine proteinases (CL3, CL4) and three cathepsins B (CB1,CB2, CB3) which are predominantly expressed in NEJ. Phylogenetic analysis showed that NEJ-expressed cathepsins L constitute a well-definedclade separate from the adult enzymes. Excystment induction resulted in a significant increment in activity towards cathepsin-specific fluoro-genic substrates in metacercariae homogenates, consistent with the detection of precursor and mature forms of cathepsins B and L beforeand after induction. In NEJ culture supernatants, protein and relative activity profiles show subtle changes during the first 48 h, with prevalenceof cathepsin L-like activity, although cathepsins CB3 and CL3 were detected by mass spectrometry. Noticeably, the hydrolysis of a substrate withproline in the P2 position was predominant, a property only shared with adult CL2 and vertebrate cathepsin K among the C1A subfamily ofcysteine proteases. Collectively these mRNA, protein and enzymatic data demonstrate the existence of a NEJ-specific repertoire of cathepsinsexpressed early in invasion, distinct to those used by other trematodes, potentially relevant for specific vaccine and chemotherapy design. Thediversity of proteases employed by trematodes in the invasion process is discussed.� 2008 Elsevier Masson SAS. All rights reserved.
Keywords: Cathepsins; Trematode; Invasion; Fasciola hepatica; Cercaria
Abbreviations: NEJ, newly excysted juveniles; E/S, excreted and/or se-
creted products; SDSePAGE, sodium dodecyl sulfateepolyacrylamide gel
electrophoresis; AMC, 7-amino-4-methylcoumarin; ORF, open reading frame;
RT, reverse transcription; BSA, bovine serum albumin; MSE, metacercariae
somatic extract; HEPES, 4-(2-hydroxyethyl)-1-piperazine ethanesulfonic
acid; DTT, 1,4-dithiothreitol; MALDI-TOF, matrix-assisted laser desorption
ionization time of flight.
* Corresponding author. Tel.: þ598 2 924 9562; fax: þ598 2 924 9563.
E-mail address: [email protected] (J.F. Tort).
0300-9084/$ - see front matter � 2008 Elsevier Masson SAS. All rights reserved.
doi:10.1016/j.biochi.2008.04.020
1. Introduction
In order to complete their life cycle, parasitic organismshave to gain access to their definitive location within theirhosts, while avoiding the defense mechanisms imposed uponthem. At the same time they are committed to obtain thenecessary resources from their hosts. Diverse molecules andbiochemical pathways have evolved to perform some of theseprocesses, with proteolytic enzymes being major componentsof the parasitic way of life. Accumulated evidence from

1462 M. Cancela et al. / Biochimie 90 (2008) 1461e1475
parasitic protozoa and helminths show that a correlation existsbetween the expanding role of proteases in parasite life cycleand an increase in the number of protease genes, leading to thewidespread presence of multigenic families [1e4].
While several proteases are present in different tissues andcellular compartments, it is reasonable to assume that those re-leased by the parasite are the most relevant in the interplay withhost targets. In multicellular parasites such as platyhelminthsand nematodes, the gut is a major source of secreted proteases.As in other invertebrates, the principal digestive proteases arepapain family cysteine proteases (Merops Clan C1A [5]) andaspartic proteinases, in contrast to the conserved pancreasderived serine proteases found in intestinal digestion in verte-brates [6]. These parasitic proteases, which are essential for nu-trition and protein catabolism, are also critical players infundamental processes for parasite infection such as hosttissue invasion, migration and evasion of host immune mecha-nisms [1,7e9]. A complex repertoire of proteases seems to beused by diverse trematodes in the invasion process. Whileacetabular gland secreted cercarial elastase (a serine protease)is fundamental in skin penetration in Schistosoma mansoni, theenzyme is absent in S. japonicum, where the crucial role in theinitial invasion is performed by gut-secreted cathepsin B-likecysteine proteases [10,11]. In the trematodes of the genus Para-gonimus a subset of cysteine proteases was proposed to haverelevant functions in tissue invasion/migration in the definitivehost [12e15], and in our model organism Fasciola hepatica,gut-secreted cysteine proteases have long been proposed asprincipal players in host duodenal invasion [16,17].
Fasciola hepatica and Fasciola gigantica are the causativeagents of fasciolosis, a chronic liver infection of cattle andsheep affecting an estimated 700 million animals worldwide.Furthermore, it has been determined that at least one fourthof the world livestock population grazes at areas where F.hepatica (temperate), F. gigantica (tropical) or both speciesare present. The economic losses caused by the infection areconservatively estimated at US$3.2 billion annually, witha more pronounced impact to rural agricultural communitiesin developing countries [18]. In recent years the incidence offasciolosis in humans has increased significantly, leading toits recognition as an emerging zoonotic disease by the WorldHealth Organization [19]. The infective metacercariae areingested, excysted within the intestine, the newly excysted ju-veniles (NEJ) burrow through the duodenal wall, and migrateeroding the liver parenchyma while maturing, and finallyestablish in the biliary ducts to develop into fertile adults.Studies performed in the 1960s pointed out a relevant rolefor proteolytic enzymes in these processes [16], and morethan two decades later the major proteins in excretionesecretion products (E/S) of adult and immature stages ofF. hepatica were biochemically identified as cysteine protein-ases [17].
Two cathepsin L-like proteinases, cathepsin L1 and L2, arepredominant in adult F. hepatica gut contents and excretion/secretion products [20e23]. Several in vitro assays demon-strated that these enzymes are capable of degrading extracellu-lar matrix and basement membrane components suggesting
a role in invasion [24]. The reported degradation of fibrinogencan contribute to their role in the feeding process [25]. Theirpossible involvement in immune evasion is supported byexperiments demonstrating cleavage of immunoglobulins andthe impairment of antibody-mediated eosinophil attachmentto newly excysted juveniles [26,27]. Moreover, cysteineproteinases are believed to immunomodulate T-cell responsesduring F. hepatica infection by suppressing interferon-gproduction [28] and degrading surface CD4 molecules in Tcells [29]. When used as targets for immunoprophylaxis,significant levels of protection, reduction of viability andfertility both in F. hepatica and the tropical related speciesF. gigantica have been obtained [30e34].
Numerous sequences corresponding to adult F. hepatica ca-thepsins have been reported, and although few sequencesmight correspond to polymorphisms or geographical variationsof the same gene, it is clear that different cathepsins genes ex-ists in liver fluke, constituting a multigenic family [1,35,36].This view is supported by kinetic analysis revealing subtle dif-ferences in substrate specificity between selected cathepsins L[20,37,38]. The diverse repertoire of cathepsins has been pro-posed to be related to specific expression of different genes atvarious stages of the complex life cycle, or also related tofunctional specialization of the isoforms [8,9]. Recently thecathepsins L secreted by the adult stage have been identifiedby proteomic approaches [39].
While the biochemical properties of the main adult cathep-sins have been extensively studied, proteases secreted by theinvasive juvenile stages have been scarcely analyzed. This isdue to limited access to this developmental stage (NEJ) andthe difficulty to obtain sufficient material for mRNA analysisand E/S protein studies. As the initial stages of invasion arethe main objectives of the protective immune responses, andthe fluke pathology is mainly due to liver erosion during juve-nile fluke migration, the NEJ cathepsins would be the most rel-evant targets for vaccine and chemotherapy design. Cathepsinsobtained from NEJ and immature intraperitoneal flukes haveshown to be different from the adult enzymes [26,40,41].Whereas cathepsins L are predominant in adult stages, earlierreports indicate that cathepsin B activity is present in juvenilestages or immature flukes [42,43]. In a recent study in F. gi-gantica, three cathepsin B genes were identified, two ofthem being expressed exclusively in metacercariae and juve-nile stages [44]. However, cathepsin L-like proteinases havealso been detected in somatic extracts of juveniles from F. he-patica [40]. In addition, two genes were reported encodinga stage-specific cathepsin L expressed in NEJ, which corre-sponds to an immunodominant antigen detected by serafrom infected animals that has been termed CL3 [45,46].
The elucidation of the excreted/secreted degradome by theearly invading stage might contribute to discriminate thoseproteinases involved in pivotal functions at the parasiteehostinterface. In this study, we report the cloning of the cDNAscorresponding to cathepsins L and B expressed during theearly invasive stage. At the same time the cysteine proteasessecreted by this stage were identified by a combination of im-munological, enzymatic and proteomic approaches. Finally,

1463M. Cancela et al. / Biochimie 90 (2008) 1461e1475
we correlated the secreted forms with particular sequences,univocally identifying the isoforms secreted by juvenile para-sites. We found that three cathepsin B genes are expressedshortly after excystment, while the cathepsins L secreted byjuveniles are encoded by early divergent stage-specific genesand have different enzymatic properties than the predominantcounterparts in the adult flukes.
2. Materials and methods
2.1. Parasites
Mature F. hepatica worms were extracted from the bileducts of bovine livers obtained at local abattoirs. Liver flukeswere transported to the laboratory in bile at 37 �C, washed for1 h at 37 �C in 10 mM phosphate-buffered saline (PBS) pH 7.3before in vitro culture. Metacercariae were obtained in our lab-oratory from experimentally infected Lymnaea viatrix snailsand maintained on 0.4% carboxymethyl cellulose until use.Excystment of metacercariae was performed as describedpreviously [26] with minor modifications. Briefly, metacercar-iae were placed in a 100-mm filter and incubated 5 min with1% sodium hypochlorite to remove the outer cyst wall. Thenthey were washed exhaustively with PBS at 39 �C andactivated in a medium prepared by mixing equal volumes ofsolution A (0.4% sodium taurocholate, 120 mM NaHCO3,140 mM NaCl pH 8.0 and solution B (50 mM HCl and33 mM L-cysteine). The excystment process was monitoredunder the microscope. After 90e180 min of incubation at39 �C, NEJ were collected in a 20 mm-filter, washed withRPMI-1640 medium and used for RNA extraction or in vitroculture as described below.
2.2. Metacercariae somatic extract (MSE)
To analyze changes in cysteine proteinases during metacer-cariae excystment, we prepared somatic extract of metacercar-iae before and after 1-h incubation in induction medium.Briefly, 500 metacercariae were collected in a 1.5 ml tube,washed with 500 ml of cold PBS and sonicated in ice bathfor 5 min with 60-s bursts at 20% power followed by 30 spauses, using an Ultrasonic Homogenizer 4710 (Cole-PalmerInstrument, IL). The homogenate was centrifuged at10,000 � g for 20 min at 4 �C. The obtained supernatant(MSE) was stored at �20 �C until used.
2.3. Excretionesecretion (E/S) products
Adult fluke E/S products were obtained as previously de-scribed [17]. Briefly, adult flukes were cultured in RPMI-1640, pH 7.3 supplemented with 30 mM HEPES, 2% glucose,25 mg/l gentamycin, (1 adult worm per ml medium) for 6 h at37 �C and the supernatant centrifuged at 20,000 � g for 1 h at4 �C. E/S products were filtered through 0.22-mm and concen-trated by ultrafiltration using a 10-kDa cut-off membrane(Amicon, Millipore Co., MA). NEJ E/S products were ob-tained as previously described [26]. NEJ were cultured in
RPMI-1640 pH 7.3 containing 2 mM L-glutamine, 25 mg/lgentamycin, 2 g/l NaHCO3 and 2 mg/ml amphotericin B (1NEJ per ml medium) at 37 �C in an atmosphere containing5% CO2. Medium was replaced at 2, 4, 6, 8, 24, and 48 h dur-ing in vitro culture. The culture supernatant was centrifugedand filtered as described above. The E/S products were con-centrated in 10-kDa cut-off membrane Amicon Ultra-4 tubes(Millipore, MA).
2.4. RNA extraction and cDNA synthesis
Total RNA from NEJ was extracted using a micro-midiRNA extraction kit (Invitrogen). Total RNA from adult flukeswas prepared by using TRIzol reagent (Life Technologies), ac-cording to the manufacturer’s protocol. First strand synthesiswas performed with the reverse transcriptase Superscript II(Invitrogen) with oligo dT primer using 200 ng of total RNAas template.
2.5. Construction and sequencing of NEJ cathepsins Band L cDNAs mini-libraries
PCR amplifications were performed using 50 ng of NEJcDNA as template and degenerate primers, corresponding tohighly conserved regions of cathepsins B and L sequencesfrom F. hepatica and F. gigantica. Cathepsin B amplificationwas done using a mixture of two overlapping forward primers(proCBF5 50-GARCCNTTYTCNGAYGARCT-30 and proCBF6 50-GAYGARCTNATYCAYTCYAT-30) and a reverseprimer (CBR 50-CCARTANTTNACNCCRTTYT-30). Forcathepsin L amplification the primers used were proCL(50-TGGCAYSARTGGAAGCGAAT-30) and reverse NEJR(50-GAAGTCAGRTTGRGCATCCAC-30). Reaction condi-tions were the same for both amplifications (200 nM eachdNTPs, 2.5 mM MgSO4, 0.05 U/ml HiFi Taq Platinum DNApolymerase (Invitrogen), and primers at a final concentrationof 300 pM each one. Reactions were incubated in a MJResearch PTC-255 thermocycler using five cycles (94 �C,1 min; 45 �C, 45 s and 72 �C, 1 min), five cycles(94 �C, 1 min; 48 �C, 45 s and 72 �C, 1 min) and 20 cycles(94 �C 1 min; 50 �C, 45 s and 72 �C, 1 min). The resultingPCR fragments of the expected sizes (830 bp for cathepsinB amplification and 678 bp for cathepsin L reaction) weregel-purified using the GFX kit (Amersham Pharmacia Biotech,UK) and ligated to TOPO TA cloning vectors (Invitrogen, CA,USA). Cathepsin B products were cloned in pCR2.1 generat-ing the cathepsin B cDNA library, and the cathepsin L librarywas generated in pCR4.0 vector likewise. Recombinant cloneswere identified by colony PCR using universal primers M13forward and reverse to evaluate the quality of the library andinsert size. Selected clones were grown in 2� LB and stockedin 96-well plates after glycerol addition to a final concentra-tion of 15%. Forty-eight clones of each library were selectedat random for plasmid DNA purification and sequencing ofboth strands with vector primers using the Dyenamic ETDye Terminator cycle sequencing kit for MegaBace DNA

1464 M. Cancela et al. / Biochimie 90 (2008) 1461e1475
Analysis Systems (Amersham Pharmacia Biotech, UK) ac-cording to the manufacturer’s instructions.
2.6. Stage-specific expression
Specific PCR primers were designed based on the se-quences obtained (Table 1) and used for PCR amplificationon NEJ and adult stage cDNA. Fifty nanograms of NEJ oradult cDNA were amplified with the corresponding primerpair during 30 cycles (1 min at 94 �C, 45 s at annealingtemperature, 1 min at 72 �C) using Taq DNA polymerase(Biotechnology Center, UFRGS, Brazil). Different annealingtemperatures and MgCl2 concentrations were previously opti-mized for each set of primers using the cloned sequences astemplates (see Table 1). PCR products were analyzed by1.5% agarose gel electrophoresis.
2.7. Bioinformatics analysis
Sequences obtained from each library were processed usingthe Phred/Phrap/Consed package [47e49]. The resultingclusters were compared to cysteine proteinases sequencesavailable in public databases using TBLASTX [50]. Represen-tative complete sequences of each cluster were deposited inGenBank with the accession numbers DQ534443, DQ534444, DQ534445, DQ534446 and DQ534447. All Fasciolacathepsin sequences deposited in GenBank were retrievedand aligned along with those obtained in the present studyusing the ClustalX package [51]. Alignments were manuallycorrected and used to generate a bootstrapped Neighbor Join-ing tree using ClustalX. Human cathepsins L and B wereincluded as outgroups. Putative glycosylation sites weresearched using the NetNGlyc 1.0 Server (http://www.cbs.dtu.dk/services/NetNGlyc/).
2.8. Gelatin substrate and SDSePAGE
Non-denaturing gelatin substrateePAGE was performed asdescribed [52]. Samples were mixed with an equal volume ofloading buffer (0.5 M TriseHCl pH 6.8, 20% glycerol and0.01% bromophenol blue) and separated by PAGE with 1%gelatin at 4 �C. After electrophoresis, the gel was incubatedfor 18 h at 37 �C in activity buffer consisting in 50 mMsodium phosphate buffer pH 7.0, 1 mM EDTA and 1 mMDTT, in the presence or absence of 5 mM of E64. The gelswere stained with Coomassie Brilliant Blue R250
Table 1
Specific primers for cathepsins B and L amplification
Primer Forward Reverse
FhCB1 50-GGCAGTGGCTGGTGTCGG-30 50-GTGATCTGGATCCAG
FhCB2 50-CACGGCGGCAGCCAGTG-30 50-TTCGAGAGTCACCA
FhCB3 50-GGCTGTCAGCTCTGCTTCG-30 50-ATCTCTTGGACACGG
FhCL1 50-CGTGCCGTACCCGACAAA-30 50-GATTCGGTTTCCAAT
FhCL2 50-CTTGCCGTACCCGAGAGC30 50-TTGGCATATGCCAAC
FhCL3 50-TTTCATATGAAGCCGAAGGC-30 50-GGCTACTCCAAGCTC
FhCL4 50-CCGTATGAGGCAAACGATCG-30 50-GCAACTCCCAACCG
(CBB-R250). Denaturing SDSePAGE was performed as de-scribed and proteins stained using either CBB-R250 [53] orsilver nitrate [54]. The molecular weight markers wereP7702S (New England Biolabs) and RPN 755 (BioRad). Pro-tein concentration was determined by the Bicinchonic acidprotein assay kit (Pierce, IL, USA) in microtiter plates usingbovine serum albumin (BSA) as protein standard. Densitomet-ric analysis of SDSePAGE was performed using Quantity One1D-Analysis Software (BioRad, USA).
2.9. Western blots
For Western blot analysis, proteins were electrotransferredonto a 0.22-mm nitrocellulose membrane (BioRad) for90 min at 1.2 mA/cm2 [55] and the membrane was blockedwith 1% bovine serum albumin (BSA) in PBS-0.1% Tween20, overnight at 4 �C. The strips were probed either with1:7000 sheep anti-procathepsin L1 serum (anti-FhCL1) [56]or with 1:1000 polyclonal sheep serum raised against cathep-sin B from 5-week-old worms (anti-FhCB) [43] for 1 h atroom temperature. Bound antibodies were detected using ananti-sheep IgG horseradish peroxidase conjugate (Sigma)diluted 1:3000 in BSA 1%. The blots were developed by usingthe 3,3-diaminobenzidine (DAB) system (Sigma).
2.10. Proteolytic activity assays
Hydrolysis of substrates Z-Phe-Arg-AMC, Z-Arg-Arg-AMC and t-Gly-Pro-Arg-AMC was monitored in 96-wellplates in a BMG FluoStar Galaxy Spectrophotometer (BMGLabtechnologies, Toronto). Reactions were performed in a vol-ume of 200 ml and final substrate concentration of 50 mM.Samples (20 ng protein) were pre-incubated in 50 mM sodiumphosphate buffer (pH ranging from 5 to 8), 1 mM DTT, 1 mMEDTA, for 10 min at room temperature, before the addition ofthe substrate. The release of AMC was continually monitoredat 355 nm (excitation) and 460 nm (emission) for 40 min at37 �C. For inhibition assays, 5 mM E-64 (a general clan CAcysteine protease inhibitor) was added to the pre-incubationreaction. Activity was calculated from the slope of the fluores-cence vs. time curve and the AMC standard curve.
2.11. Mass spectrometry analysis
Proteins of interest were obtained from an unidimensionalSDSePAGE as previously described [57,58]. After trypsin
Size (bp) Annealing temp. (�C) [MgCl2] (mM)
GATGACG-30 253 66 2
ACGTGATC-30 250 72 1.5
AGGC-30 276 66 2
CCAA-30 262 66 2
CGC-30 327 68 1.5
TTTCC-30 344 62 2
GCTATC-30 340 72 2.4

1465M. Cancela et al. / Biochimie 90 (2008) 1461e1475
digestion, peptides were analyzed by matrix-assisted laser de-sorption ionization time of flight (MALDI-TOF) in a Voyager-DE-PRO mass spectrometer (Perspective Biosystems,Framingham, MA). Masses of each peptide were comparedto theoretical masses of protein sequences available from theNational Center for Biotechnology Information and those ob-tained in this work by in silico translation of cDNA sequences(CL3, CL4, CB1, CB2 and CB3). Theoretical digestion ofprotein sequences with trypsin was performed by using theProtein Prospector 3.4.1 MS-Digest program (http://www.prospector.ucsf.edu).
2.12. Detection of N-glycans in NEJ cysteine proteases
NEJ E/S proteins were heated for 3 min at 95 �C in 0.1%SDS and 1% b-mercaptoethanol and then incubated over-night at 37 �C in 50 mM sodium phosphate buffer pH 7.3,25 mM EDTA, 1% Triton X-100 and 2.5 U N-glycosidase Ffrom Flavobacterium maningosepticum (Roche, Mannheim,Germany). Fetuin (Sigma, USA) was used as positive controlusing the same reaction conditions. Treated and untreatedsamples were separated by 14% SDSePAGE, silver stainedor transferred to nitrocellulose membrane for Western blotanalysis.
3. Results
3.1. NEJs express three different cathepsin B mRNAs
We constructed plasmid minilibraries containing mem-bers of the cathepsin multigenic families by cloning theproducts of PCR amplification with degenerate primersspanning conserved regions. In the case of cathepsin B, forexample, the primers designed were expected to amplifyall the cathepsin B genes of Fasciola species found in thepublic databases. Amplification of NEJ cDNA with cathepsinB primers yielded 830 bp PCR products that were subse-quently cloned to construct the cathepsin B minilibrary,then 48 clones were selected at random for sequencing.The resulting sequences were compared, assembled andgrouped in three different NEJ cathepsin B genes, CB1,CB2 and CB3, comprising open reading frames (ORFs) of834 bp encoding 278 amino acid residues of the proteinase(Fig. 1). Identities between amino acid sequences of theclones ranged from 62% to 66%. Alignment of cathepsinB predicted amino acid sequences showed homology withpreviously described Fasciola cathepsins B, including thehighly conserved active site residues (Glu23, Gly-/Ser27,Cys29, Hys199), and the 23 residue occluding loop. Sequenceanalysis revealed potential N-glycosylation sites at N57 ofCB1, N200 and N211 of CB2, and N200 of CB3. No predictedO-glycosylation sites were found. The three genes are puta-tive orthologues of the three F. gigantica cathepsin B genesreported in databases [44], since they exhibit very high se-quence identity, and CB2 exhibits 99% amino acid identityto the sequence of 5-week immature F. hepatica cathepsinB originally reported by Wilson et al. (1998) as CB1 [43].
The nomenclature used here is consistent with the reportedF. gigantica genes.
3.2. Expression of two NEJ cathepsins L mRNAsdistinct from adult CLs
Using the degenerate primer minilibrary approach de-scribed above, NEJ cathepsins L clones were obtained froma 670 bp amplification product, then 48 clones were selectedat random, sequenced and assembled into groups. We identi-fied two different cDNAs encoding cathepsins L, namedCL3 and CL4, with 678 bp ORFs coding 226 amino acid res-idues of the enzyme (Fig. 2). Amino acid sequence identity be-tween the two sequences was 69% and exhibited the typicalcathepsin L characteristics, including the conserved catalyticactive site residues (Glu19, Gly23, Cys25). CL3 exhibits 96%identity with the NEJ CL3 described by Harmsen et al. [45],which has a putative orthologue in F. gigantica, CATL1G[59]. CL4 had not been found previously in F. hepatica, butit exhibits 90% identity to F. gigantica CATL1H [59], whichis probably its orthologue. Contrasting with the adult ex-pressed CL1 and CL2, CL3 and CL4 display potential N-gly-cosylation sites at N128 in CL3 and N129 in CL4. No predictedO-glycosylation sites were found.
3.3. Phylogenetic analysis of NEJ cysteine proteases
By alignment (Figs. 1 and 2) and construction of phyloge-netic trees (Fig. 3) we compared the sequences obtained withother Fasciola cathepsin sequences available in the publicdatabases. Phylogenetic analysis showed that cathepsins Lfrom both F. hepatica and F. gigantica form four differentclades named CL1, CL2, CL5, and NEJ CLs. The CLs ex-pressed in NEJ (CL3 and CL4) form a distinct group withrespect to CLs expressed in adult stage of F. hepatica(CL1, CL2 and CL5). The four clades include isoformsfrom F. hepatica and F. gigantica, so the group of genespreferentially expressed in NEJ may have diverged fromthe group expressed in adults before the gigantica/hepaticaspeciation.
Two of the three cathepsin B genes detected, CB2 and CB3,grouped with F. gigantica cathepsins B previously identifiedfrom juvenile flukes (CB2 grouped with F. gigantica CB2and CB3 grouped with F. gigantica CB1 and CB3 accordingto the nomenclature by Meemon et al. [44], indicating thatalso in F. hepatica several cathepsin B isoforms are expressedin NEJ as was described in F. gigantica. CB1 exhibited 97%identity with a F. hepatica NEJ protein predicted froma cDNA sequence entered as AJ488928 (Fig 3).
3.4. Stage-specific expression of the identifiedNEJ cathepsins
Using specific primers designed on the sequences obtained,we tested by RTePCR the presence of each NEJ cathepsin injuvenile and adult cDNA. The three cathepsin B genes andCL3 were specifically detected in juvenile cDNA, and not
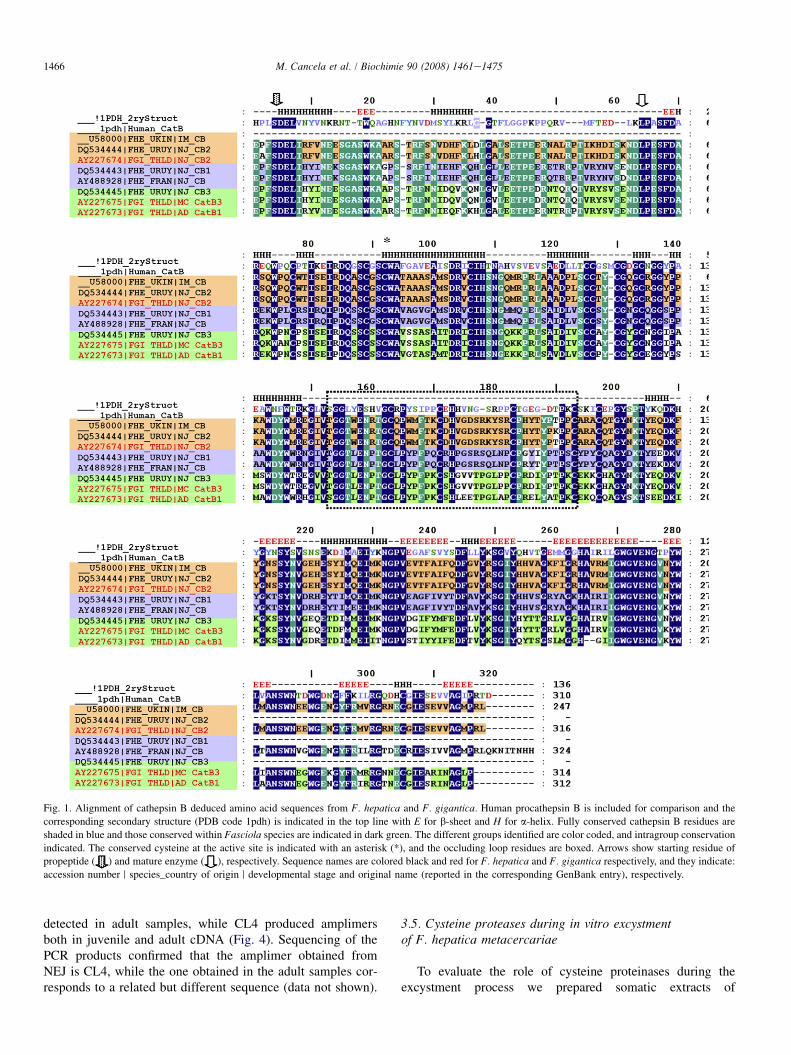
Fig. 1. Alignment of cathepsin B deduced amino acid sequences from F. hepatica and F. gigantica. Human procathepsin B is included for comparison and the
corresponding secondary structure (PDB code 1pdh) is indicated in the top line with E for b-sheet and H for a-helix. Fully conserved cathepsin B residues are
shaded in blue and those conserved within Fasciola species are indicated in dark green. The different groups identified are color coded, and intragroup conservation
indicated. The conserved cysteine at the active site is indicated with an asterisk (*), and the occluding loop residues are boxed. Arrows show starting residue of
propeptide ( ) and mature enzyme ( ), respectively. Sequence names are colored black and red for F. hepatica and F. gigantica respectively, and they indicate:
accession number r species_country of origin r developmental stage and original name (reported in the corresponding GenBank entry), respectively.
1466 M. Cancela et al. / Biochimie 90 (2008) 1461e1475
detected in adult samples, while CL4 produced amplimersboth in juvenile and adult cDNA (Fig. 4). Sequencing of thePCR products confirmed that the amplimer obtained fromNEJ is CL4, while the one obtained in the adult samples cor-responds to a related but different sequence (data not shown).
3.5. Cysteine proteases during in vitro excystmentof F. hepatica metacercariae
To evaluate the role of cysteine proteinases during theexcystment process we prepared somatic extracts of
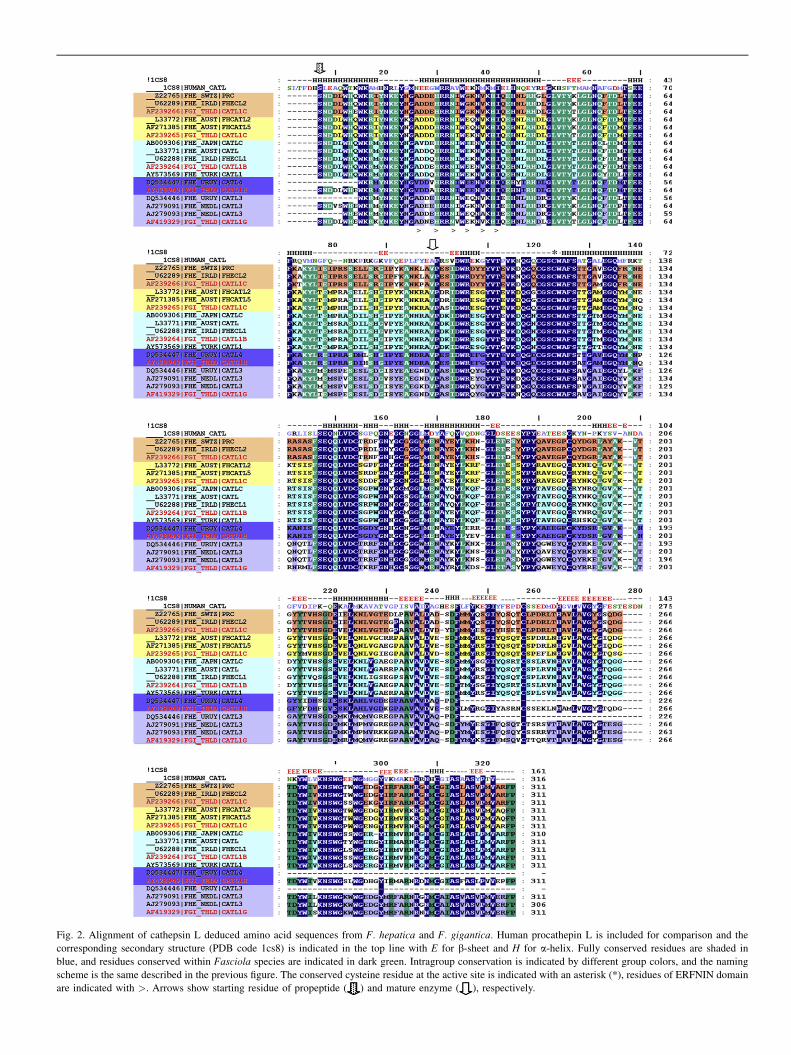
Fig. 2. Alignment of cathepsin L deduced amino acid sequences from F. hepatica and F. gigantica. Human procathepin L is included for comparison and the
corresponding secondary structure (PDB code 1cs8) is indicated in the top line with E for b-sheet and H for a-helix. Fully conserved residues are shaded in
blue, and residues conserved within Fasciola species are indicated in dark green. Intragroup conservation is indicated by different group colors, and the naming
scheme is the same described in the previous figure. The conserved cysteine residue at the active site is indicated with an asterisk (*), residues of ERFNIN domain
are indicated with >. Arrows show starting residue of propeptide ( ) and mature enzyme ( ), respectively.

Fig. 3. Phylogenetic relationship between cysteine proteases of F. hepatica and F. gigantica. Bootstrap neighbor-joining tree of Fasciola cathepsins B and L mem-
bers. The different clades are identified by boxes colored as in the previous figures.
1468 M. Cancela et al. / Biochimie 90 (2008) 1461e1475
metacercariae before and after 1 h of induction. At this timepoint, larvae inside the cyst are highly motile but none ofthem have excysted. Proteolytic cleavage over three substrateswas observed in metacercariae before and after induction(Fig. 5). This activity was inhibited by the specific class inhibitorE-64 and enhanced by DTT. Z-Phe-Arg-AMC was preferen-tially cleaved followed by t-Gly-Pro-Arg-AMC and Z-Arg-Arg-AMC. Interestingly, we found a ten-fold increase incysteine protease activity on the three substrates at 1 h post-induction compared with non-induced metacercariae. In orderto identify the isoforms responsible of this activity we analyzed
the presence of cysteine proteinases in metacercariae somaticextract. By SDSePAGE, we observed a qualitative change inthe protein profile during the induction of metacercariae excyst-ment (Fig. 6A). Western blot analysis revealed that a 45-kDa ca-thepsin L and 35- and 32-kDa cathepsin B proenzymes werepresent in somatic extracts before induction, while processed28 kDa CL and CB mature forms appeared after 1 h of induction(Fig. 6B). By MALDI-TOF analysis we confirmed that thepredominant bands of 41 kDa (band 1)and 35 kDa (band 2)(Fig. 6A) corresponded to the pro-cathepsin CL3 and pro-cathepsin CB3 identified in this work (Fig. 6C).
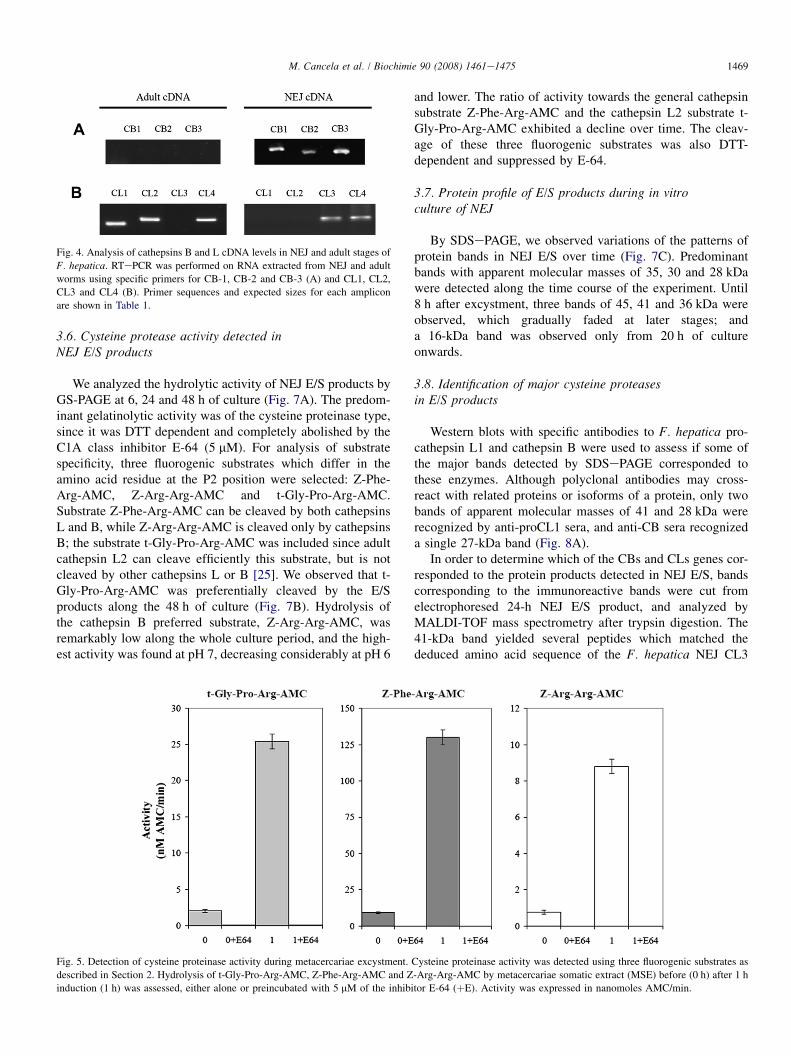
Fig. 4. Analysis of cathepsins B and L cDNA levels in NEJ and adult stages of
F. hepatica. RTePCR was performed on RNA extracted from NEJ and adult
worms using specific primers for CB-1, CB-2 and CB-3 (A) and CL1, CL2,
CL3 and CL4 (B). Primer sequences and expected sizes for each amplicon
are shown in Table 1.
1469M. Cancela et al. / Biochimie 90 (2008) 1461e1475
3.6. Cysteine protease activity detected inNEJ E/S products
We analyzed the hydrolytic activity of NEJ E/S products byGS-PAGE at 6, 24 and 48 h of culture (Fig. 7A). The predom-inant gelatinolytic activity was of the cysteine proteinase type,since it was DTT dependent and completely abolished by theC1A class inhibitor E-64 (5 mM). For analysis of substratespecificity, three fluorogenic substrates which differ in theamino acid residue at the P2 position were selected: Z-Phe-Arg-AMC, Z-Arg-Arg-AMC and t-Gly-Pro-Arg-AMC.Substrate Z-Phe-Arg-AMC can be cleaved by both cathepsinsL and B, while Z-Arg-Arg-AMC is cleaved only by cathepsinsB; the substrate t-Gly-Pro-Arg-AMC was included since adultcathepsin L2 can cleave efficiently this substrate, but is notcleaved by other cathepsins L or B [25]. We observed that t-Gly-Pro-Arg-AMC was preferentially cleaved by the E/Sproducts along the 48 h of culture (Fig. 7B). Hydrolysis ofthe cathepsin B preferred substrate, Z-Arg-Arg-AMC, wasremarkably low along the whole culture period, and the high-est activity was found at pH 7, decreasing considerably at pH 6
Fig. 5. Detection of cysteine proteinase activity during metacercariae excystment.
described in Section 2. Hydrolysis of t-Gly-Pro-Arg-AMC, Z-Phe-Arg-AMC and Z
induction (1 h) was assessed, either alone or preincubated with 5 mM of the inhibi
and lower. The ratio of activity towards the general cathepsinsubstrate Z-Phe-Arg-AMC and the cathepsin L2 substrate t-Gly-Pro-Arg-AMC exhibited a decline over time. The cleav-age of these three fluorogenic substrates was also DTT-dependent and suppressed by E-64.
3.7. Protein profile of E/S products during in vitroculture of NEJ
By SDSePAGE, we observed variations of the patterns ofprotein bands in NEJ E/S over time (Fig. 7C). Predominantbands with apparent molecular masses of 35, 30 and 28 kDawere detected along the time course of the experiment. Until8 h after excystment, three bands of 45, 41 and 36 kDa wereobserved, which gradually faded at later stages; anda 16-kDa band was observed only from 20 h of cultureonwards.
3.8. Identification of major cysteine proteasesin E/S products
Western blots with specific antibodies to F. hepatica pro-cathepsin L1 and cathepsin B were used to assess if some ofthe major bands detected by SDSePAGE corresponded tothese enzymes. Although polyclonal antibodies may cross-react with related proteins or isoforms of a protein, only twobands of apparent molecular masses of 41 and 28 kDa wererecognized by anti-proCL1 sera, and anti-CB sera recognizeda single 27-kDa band (Fig. 8A).
In order to determine which of the CBs and CLs genes cor-responded to the protein products detected in NEJ E/S, bandscorresponding to the immunoreactive bands were cut fromelectrophoresed 24-h NEJ E/S product, and analyzed byMALDI-TOF mass spectrometry after trypsin digestion. The41-kDa band yielded several peptides which matched thededuced amino acid sequence of the F. hepatica NEJ CL3
Cysteine proteinase activity was detected using three fluorogenic substrates as
-Arg-Arg-AMC by metacercariae somatic extract (MSE) before (0 h) after 1 h
tor E-64 (þE). Activity was expressed in nanomoles AMC/min.

Fig. 6. Identification of cysteine proteases during in vitro excystment of F. hepatica metacercariae. (A) Protein profile during metacercariae activation. Metacer-
cariae somatic extract (MSE) at 0 h (T0) and 1 h (T1) post-induction were analyzed by 12.5% SDSePAGE under reducing conditions. (B) Western blot analysis of
MSE at 0 h and 1 h using anti-procathepsin B and anti-procathepsin L sheep sera. Irrelevant sheep serum was used as negative control. (C) MALDI-TOF analysis
of bands indicated in A by asterisks. Bands were excised, trypsin digested and peptides analyzed by MALFI-TOF. One matching peptide of each band was se-
quenced. MW: molecular weight marker.
1470 M. Cancela et al. / Biochimie 90 (2008) 1461e1475
pro-enzyme [45] (Fig. 8C). Various peptides derived from thein 28-kDa band matched the deduced amino acid sequence ofthe mature CL3 enzyme (Fig. 8C) and other peptides from thesame band matched F. hepatica and F. gigantica CB3 [44]. Westudied the presence of carbohydrate groups in CL3, as a po-tential N-glycosylation site was identified by sequence predic-tions, and the apparent mass determined by SDSePAGEmigration of the proenzyme was 41 kDa compared to the ex-pected 35 kDa from the cDNA sequence. SDSePAGE andWestern blot analyses using anti-CL and anti-CB analysis ofNEJ E/S products showed no difference in migration of the41- and 28-kDa bands before and after deglycosylation treat-ment (data not shown).
4. Discussion
Growth and maturation in helminth parasites are associatedwith substantial changes in the composition of the host-parasite interface resulting from the regulated expression ofsomatic, secretory and/or surface proteins. Fasciola hepaticaundergoes a dramatic transition in the tegument secretorygranules from NEJ-specific T0 granules to T1 and T2 mainlydetected in immature 5-week flukes [60]. Furthermore, rapidswitches in antigenic composition of somatic extracts ofmigrating NEJ have been reported [41], and even whena few potentially important molecules have been identified[40], their relevance in the context of host-parasite interaction
has not been fully addressed yet. The presence of cysteine pro-teases as major components of E/S products in diverse stagesis well documented [40,43,61,62]. While two cathepsins L(CL1 and CL2) are clearly predominant in adult secretions,and a similar pattern of activities has been detected in 3-and 5-week immature flukes [17], a different cathepsin L en-zyme (CL3) and cathepsins B have been detected as majorcomponents of somatic extract of NEJs [40,45]. These prelim-inary data suggest that a distinct repertoire of cathepsins mightbe expressed at diverse stages of the parasite life cycle, switch-ing according to the needs imposed by their interaction withthe host. In order to verify this hypothesis we decided to studythe expression profile of mRNA and secreted cysteine protein-ases using an in vitro excystment model, which mimics thephysico-chemical environment of the stomach and duodenumfound by the parasite at the early stages of invasion.
Analysis of cDNA minilibraries constructed with conserveddegenerate primers for cathepsins B and L allowed us to iden-tify two distinct cathepsins L (CL3 and CL4) and three differ-ent cathepsins B (CB1, CB2 and CB3) expressed in NEJ. Noneof these sequences correspond to proteins previously detectedin the adult stage. While CL3 is very similar to a previouslyreported NEJ enzyme [45], and CB2 corresponds toa 5-week immature F. hepatica previously reported [43], theremaining sequences are novel for F. hepatica and their closerrelatives are the F. gigantica juvenile sequences [44,59],which are their probable orthologues.
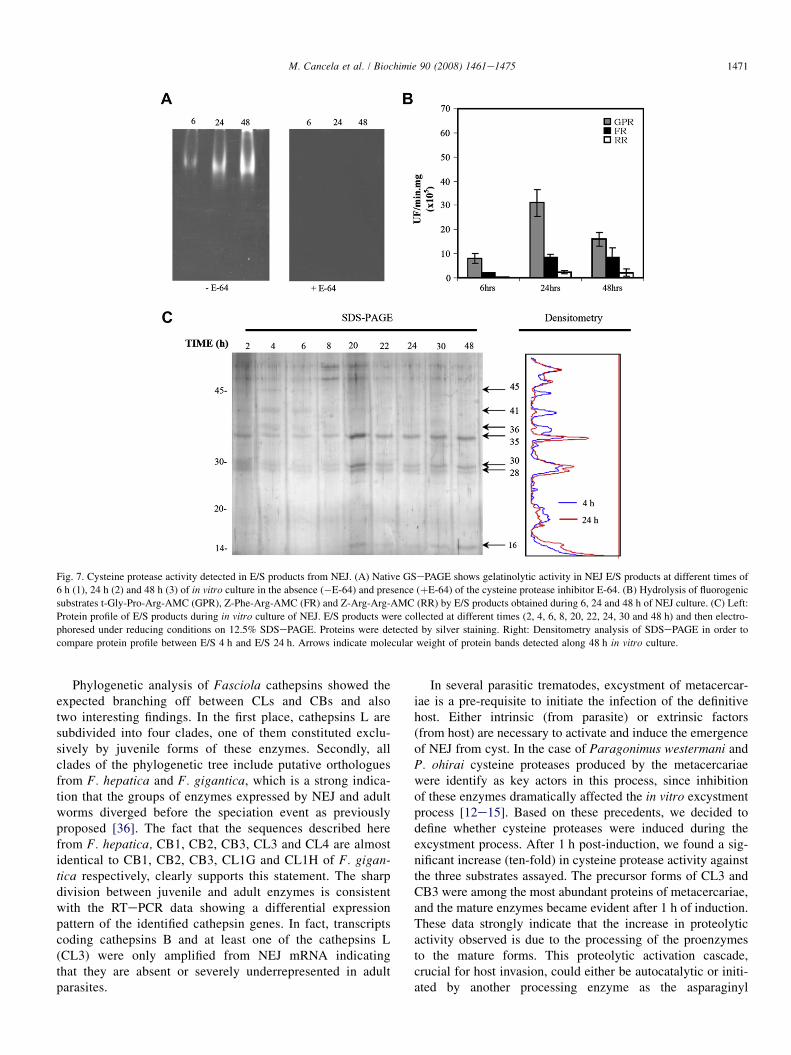
Fig. 7. Cysteine protease activity detected in E/S products from NEJ. (A) Native GSePAGE shows gelatinolytic activity in NEJ E/S products at different times of
6 h (1), 24 h (2) and 48 h (3) of in vitro culture in the absence (�E-64) and presence (þE-64) of the cysteine protease inhibitor E-64. (B) Hydrolysis of fluorogenic
substrates t-Gly-Pro-Arg-AMC (GPR), Z-Phe-Arg-AMC (FR) and Z-Arg-Arg-AMC (RR) by E/S products obtained during 6, 24 and 48 h of NEJ culture. (C) Left:
Protein profile of E/S products during in vitro culture of NEJ. E/S products were collected at different times (2, 4, 6, 8, 20, 22, 24, 30 and 48 h) and then electro-
phoresed under reducing conditions on 12.5% SDSePAGE. Proteins were detected by silver staining. Right: Densitometry analysis of SDSePAGE in order to
compare protein profile between E/S 4 h and E/S 24 h. Arrows indicate molecular weight of protein bands detected along 48 h in vitro culture.
1471M. Cancela et al. / Biochimie 90 (2008) 1461e1475
Phylogenetic analysis of Fasciola cathepsins showed theexpected branching off between CLs and CBs and alsotwo interesting findings. In the first place, cathepsins L aresubdivided into four clades, one of them constituted exclu-sively by juvenile forms of these enzymes. Secondly, allclades of the phylogenetic tree include putative orthologuesfrom F. hepatica and F. gigantica, which is a strong indica-tion that the groups of enzymes expressed by NEJ and adultworms diverged before the speciation event as previouslyproposed [36]. The fact that the sequences described herefrom F. hepatica, CB1, CB2, CB3, CL3 and CL4 are almostidentical to CB1, CB2, CB3, CL1G and CL1H of F. gigan-tica respectively, clearly supports this statement. The sharpdivision between juvenile and adult enzymes is consistentwith the RTePCR data showing a differential expressionpattern of the identified cathepsin genes. In fact, transcriptscoding cathepsins B and at least one of the cathepsins L(CL3) were only amplified from NEJ mRNA indicatingthat they are absent or severely underrepresented in adultparasites.
In several parasitic trematodes, excystment of metacercar-iae is a pre-requisite to initiate the infection of the definitivehost. Either intrinsic (from parasite) or extrinsic factors(from host) are necessary to activate and induce the emergenceof NEJ from cyst. In the case of Paragonimus westermani andP. ohirai cysteine proteases produced by the metacercariaewere identify as key actors in this process, since inhibitionof these enzymes dramatically affected the in vitro excystmentprocess [12e15]. Based on these precedents, we decided todefine whether cysteine proteases were induced during theexcystment process. After 1 h post-induction, we found a sig-nificant increase (ten-fold) in cysteine protease activity againstthe three substrates assayed. The precursor forms of CL3 andCB3 were among the most abundant proteins of metacercariae,and the mature enzymes became evident after 1 h of induction.These data strongly indicate that the increase in proteolyticactivity observed is due to the processing of the proenzymesto the mature forms. This proteolytic activation cascade,crucial for host invasion, could either be autocatalytic or initi-ated by another processing enzyme as the asparaginyl

Fig. 8. Identification of cysteine proteinases in E/S products of NEJ. (A) Western blot analysis of NEJ E/S products with anti-FhCB and anti-FhCL sera. (B) Protein
profile of NEJ E/S products from 24 h in vitro culture (1) analyzed by 15% SDSePAGE and stained with CBB-R250. (C) MALDI-TOF analysis of the 41-kDa and
28-kDa bands. MW: molecular weight; neg: irrelevant sheep sera.
1472 M. Cancela et al. / Biochimie 90 (2008) 1461e1475
endopeptidase [63]. This enzyme is a predominant protein inNEJ somatic extracts [40] and NEJ E/S (Cancela et al., unpub-lished data). Interestingly, the proCL3 identified in this workhas an asparagine at position P1 of the propeptide cleavagesite. Recombinant expression of a CL3 cDNA in Saccharomy-ces cerevisiae resulted in secretion of the proenzyme form[45], thus suggesting that the enzyme does not auto-processas observed for other recombinant F. hepatica cathepsins[22,23]. Therefore proCL3 maturation could be dependenton an asparaginyl endopeptidase activity probably expressedat the very early stages of invasion.
The NEJ-secreted protein profile shows gradual changesalong the 48 h time course of in vitro culture. ProCL3 andmature CL3 and CB3 are principal components of the 24 hE/S. These enzymes were previously identified by N-terminalprotein sequencing in somatic extracts of NEJ [40,45]. Thepro-cathepsin B sequence described in Tkalcevic el al. [40]could correspond either to CB1 or CB3, but based on ourdata we can now classify it as the orthologue of the F. gigan-tica metacercariae cathepsin B3 [44]. No peptides correspond-ing to adult cathepsins were detected, thus suggesting that theyare either absent or poorly represented in NEJ E/S products. Inaddition, although CL4 was obtained from NEJ RNA, nomatches to CL4 were revealed, so this protein is not presentin the E/S products or its expression and/or stability is low.On the other hand, proteomic analysis of adult E/S have not
found any of the juvenile forms described here [39,64], point-ing to the stage-specific expression of the diverse familymembers.
The changes in the protease activity profile observed duringand after metacercariae excystment are consistent with the ex-pression of diverse enzymes at different times, and confirmthat NEJ E/S cysteine proteases are different of those secretedby the adult fluke. Activity towards the cathepsin B substrateZ-Arg-Arg-AMC is only detected in metacercariae, while t-Gly-Pro-Arg-AMC is the preferred substrate in early NEJ,switching progressively to Z-Phe-Arg-AMC. The predominantactivity during the 48 h time course was the hydrolysis of t-Gly-Pro-Arg-AMC, but the hydrolysis of Z-Phe-Arg-AMCincreased at the 48 h time point. Until now, mammaliancathepsin K and adult liver fluke cathepsin L2 were the onlycysteine proteases reported to be able to cleave substrateswith Pro in P2, an activity related to collagen breakdown formammalian cathepsin K [23,65]. Since we did not find anyof the adult cathepsins by RTePCR nor by mass spectrometry,we suggest that this activity may be due to the CL3. The abil-ity to accept Pro in the P2 position has been thoroughly stud-ied by sequence comparisons and site-directed mutagenesis ofthe amino acid residues that form the S2 active subsite [65].While adult CL1 and most mammalian cathepsins havea Leu residue in position 69 at the entrance of the S2 subsite,FhCL2 and mammalian cathepsin K have a Tyr residue. CL3

1473M. Cancela et al. / Biochimie 90 (2008) 1461e1475
has a Trp residue that could be in part responsible for the dif-ferent substrate specificity. The functional expression of therecombinant NEJ cathepsins will help to elucidate the possiblerole of these residues in substrate specificity.
We propose the hypothesis that cysteine proteasesexpressed during invasion have a similar stage-specific expres-sion pattern in both Fasciola species, in agreement with thefindings in F. gigantica cathepsins B and L [44,59]. Theenzymes expressed in metacercariae could be involved inexcystment, those cathepsins expressed in early NEJ wouldbe involved in invasion, whereas immature and adult variantsmay participate in liver migration and feeding. While the dif-ferent patterns detected in juvenile (this work and [40,43,46]),immature [41,43] and adults [39,64,66] are evident, it remainsto be addressed the precise timing of the changes detected andmore interestingly what signals either endogenous or from thehost trigger these variations, and to compare the in vitro pat-tern with an in vivo model.
Although differences undoubtedly exist between the cyste-ine proteases present in metacercariae, NEJ and adults, therelevance of these enzymes individually and collectively inthe diverse host invasion mechanisms still needs to be ad-dressed. Very recently the role of cathepsins in invasion hasbeen tested using RNA interference [67]. A reduction of gutpassage was detected in NEJ soaked with adult CL1 and a ca-thepsin B that corresponds to the enzyme here referred to asCB2. The fact that CL1 is not produced by NEJ points to a gen-eral unspecific effect affecting the whole family, and confirm-ing that some degree of redundancy or overlappingspecificities exists within liver fluke cathepsins. More detaileddissection of the invasion process is needed to assess the effectof inhibiting or silencing individual cathepsins. These studiescombined with precise transcriptomic and proteomic analysiswould allow demarcating the role of the diverse proteases inthe initial steps of invasion, and offering consequently moreprecise targets for antiparasitic intervention.
Interestingly, the process of invasion in different trematodesseems to involve a diverse set of proteases. The invasionproteases of four species of schistosomes have been compara-tively studied recently [10,11]. While the serine type cercarialelastase produced in the acetabular glands is the principal en-zyme in Schistosoma mansoni and Schistosomatium douthitti,in Schistosoma japonicum this role is performed by cathepsinB [10]. These authors proposed that the cysteine proteinasesare the archetypal proteases that facilitate invasion and tissuepenetration by trematode parasite larvae. The class inhibitorof the subfamily C1A family E-64 has been shown to inhibitmolecular events related to invasion in different experimentalmodels in schistosomes [10,11], Paragonimus [12,14], and F.hepatica. Notably, different subsets of the C1A subfamilyparticipate in the diverse groups of trematode parasites. Whilein schistosomatids the main histolytic enzymes might be ca-thepsins B [10,11,68], in Fasciola both cathepsins B and Lhave been detected, but our data point to a more relevantrole of L type cathepsin in invasion. In Paragonimus[12,13,69,70], Opisthorchis [71] and Clonorchis [72,73] thepredominant cysteine proteases belong to the cruzipain type
(also denominated cathepsins F by similarity to mammalianenzymes), although their role in invasion has only been testedin Paragonimus westermani [12]. It comes into sight that eachparasite group evolved a diverse set of cysteine proteases to in-vade their hosts, and the evolution and expansion of the mul-tigenic families took different routes. Additionally, in somecases similar enzymes are used for cercariae/NEJ invasionand nutrition of adult parasites, but in other cases, typicallyFasciola species, different enzymes evolved for each process.These observations stress the uniqueness of each parasiteehost relationship, emphasizing the need to be cautious atcomparative analysis, even between closely related species.
In conclusion, we have identified cathepsins L and B notpreviously reported in adult flukes, and found that NEJ pro-duce a different set of cysteine proteinases which may assisttissue degradation and immuno-evasion during invasion. Wecorrelated secreted proteins with cDNA sequences and enzy-matic activities for the first time, providing the frameworkfor the dissection of the early stages of the parasite invasionto their mammalian hosts using newly generated functional ge-nomic tools [67,74]. Functional recombinant expression andcharacterization of these enzymes and validation of the datawith in vivo models and gene knock-down strategies wouldbe relevant in order to design rational strategies for diagnosis,prevention and treatment of fasciolosis in the early stages ofinfection.
Acknowledgments
We thank Dr. Carlos Cerve~nansky at the Institut PasteurMontevideo/IIBCE/Facultad de Ciencias for expert technicalassistance on the MALDI-TOF work. This work wassupported by AMSUD-PASTEUR, RTPD-Network (SIDA),CSIC, FCE (Dinacyt, BID). Anti cathepsin B serum waskindly provided by Dr. Terry Spithill. McGill University,Montreal, Canada.
References
[1] J. Tort, P.J. Brindley, D. Knox, K.H. Wolfe, J.P. Dalton, Proteinases and
associated genes of parasitic helminths, Adv. Parasitol. 43 (1999)
161e266.
[2] Y. Wu, X. Wang, X. Liu, Y. Wang, Data-mining approaches reveal hidden
families of proteases in the genome of malaria parasite, Genome Res. 13
(2003) 601e616.
[3] N.M. El-Sayed, P.J. Myler, G. Blandin, M. Berriman, J. Crabtree,
G. Aggarwal, E. Caler, H. Renauld, E.A. Worthey, C. Hertz-Fowler,
E. Ghedin, C. Peacock, D.C. Bartholomeu, B.J. Haas, A.N. Tran,
J.R. Wortman, U.C. Alsmark, S. Angiuoli, A. Anupama, J. Badger,
F. Bringaud, E. Cadag, J.M. Carlton, G.C. Cerqueira, T. Creasy,
A.L. Delcher, A. Djikeng, T.M. Embley, C. Hauser, A.C. Ivens,
S.K. Kummerfeld, J.B. Pereira-Leal, D. Nilsson, J. Peterson,
S.L. Salzberg, J. Shallom, J.C. Silva, J. Sundaram, S. Westenberger,
O. White, S.E. Melville, J.E. Donelson, B. Andersson, K.D. Stuart,
N. Hall, Comparative genomics of trypanosomatid parasitic protozoa,
Science 309 (2005) 404e409.
[4] M. Klemba, D.E. Goldberg, Biological roles of proteases in parasitic
protozoa, Annu. Rev. Biochem. 71 (2002) 275e305.
[5] N.D. Rawlings, F.R. Morton, A.J. Barrett, MEROPS: the peptidase
database, Nucleic Acids Res. 34 (2006) D270eD272.

1474 M. Cancela et al. / Biochimie 90 (2008) 1461e1475
[6] M. Delcroix, M. Sajid, C.R. Caffrey, K.C. Lim, J. Dvorak, I. Hsieh,
M. Bahgat, C. Dissous, J.H. McKerrow, A multienzyme network func-
tions in intestinal protein digestion by a platyhelminth parasite, J. Biol.
Chem. 281 (2006) 39316e39329.
[7] M. Sajid, J.H. McKerrow, Cysteine proteases of parasitic organisms,
Mol. Biochem. Parasitol. 120 (2002) 1e21.
[8] J. Dalton, C. Caffrey, M. Sajid, C. Stack, S. Donnelly, A. Loukas, T. Don,
J. McKerrow, D.W. Halton, P.J. Brindley, Proteases in trematode biology,
in: A.G. Maule, N.J. Marks (Eds.), Parasitic Flatworms: Molecular Biol-
ogy, Biochemistry, Immunology and Physiology, CAB International,
2006, pp. 348e368.
[9] C.P. Brady, A.J. Dowd, J. Tort, L. Roche, B. Condon, S.M. O’Neill,
P.J. Brindley, J.P. Dalton, The cathepsin L-like proteinases of liver fluke
and blood fluke parasites of the trematode genera Fasciola and Schisto-
soma, Biochem. Soc. Trans. 27 (1999) 740e745.
[10] J. Dvorak, S.T. Mashiyama, S. Braschi, M. Sajid, G.M. Knudsen,
E. Hansell, K.C. Lim, I. Hsieh, M. Bahgat, B. Mackenzie,
K.F. Medzihradszky, P.C. Babbitt, C.R. Caffrey, J.H. McKerrow, Differ-
ential use of protease families for invasion by schistosome cercariae, Bi-
ochimie 90 (2008) 345e358.
[11] M. Kasny, L. Mikes, J.P. Dalton, A.P. Mountford, P. Horak, Comparison
of cysteine peptidase activities in Trichobilharzia regenti and Schisto-
soma mansoni cercariae, Parasitology 134 (2007) 1599e1609.
[12] B.K. Na, S.H. Kim, E.G. Lee, T.S. Kim, Y.A. Bae, I. Kang, J.R. Yu,
W.M. Sohn, S.Y. Cho, Y. Kong, Critical roles for excretory-secretory cys-
teine proteases during tissue invasion of Paragonimus westermani newly
excysted metacercariae, Cell. Microbiol. 8 (2006) 1034e1046.
[13] Y.B. Chung, T.S. Kim, H.J. Yang, Early cysteine protease activity in ex-
cretory bladder triggers metacercaria excystment of Paragonimus wester-
mani, J. Parasitol. 91 (2005) 953e954.
[14] T. Ikeda, Involvement of cysteine proteinases in excystment of Parago-
nimus ohirai metacercariae induced by sodium cholate and A23187, J.
Helminthol. 77 (2003) 21e26.
[15] Y.B. Chung, Y. Kong, I.J. Joo, S.Y. Cho, S.Y. Kang, Excystment of Para-gonimus westermani metacercariae by endogenous cysteine protease,
J. Parasitol. 81 (1995) 137e142.
[16] R.M. Howell, Collagenase activity of immature Fasciola hepatica, Na-
ture 209 (1966) 713e714.
[17] J.P. Dalton, M. Heffernan, Thiol proteases released in vitro by Fasciola
hepatica, Mol. Biochem. Parasitol. 35 (1989) 161e166.
[18] T.W. Spithill, P.M. Smooker, B. Copeman, Fasciola gigantica: epidemi-
ology, control, immunology and molecular biology, in: J.P. Dalton (Ed.),
Fasciolosis, CAB International Publishing, Wallingford, 1999, pp.
465e525.
[19] S. Mas-Coma, M.D. Bargues, M.A. Valero, Fascioliasis and other plant-
borne trematode zoonoses, Int. J. Parasitol. 35 (2005) 1255e1278.
[20] A.J. Dowd, A.M. Smith, S. McGonigle, J.P. Dalton, Purification and
characterisation of a second cathepsin L proteinase secreted by the
parasitic trematode Fasciola hepatica, Eur. J. Biochem. 223 (1994)
91e98.
[21] A.M. Smith, A.J. Dowd, S. McGonigle, P.S. Keegan, G. Brennan,
A. Trudgett, J.P. Dalton, Purification of a cathepsin L-like proteinase
secreted by adult Fasciola hepatica, Mol. Biochem. Parasitol. 62
(1993) 1e8.
[22] L. Roche, A.J. Dowd, J. Tort, S. McGonigle, A. MacSweeney,
G.P. Curley, T. Ryan, J.P. Dalton, Functional expression of Fasciolahepatica cathepsin L1 in Saccharomyces cerevisiae, Eur. J. Biochem.
245 (1997) 373e380.
[23] A.J. Dowd, J. Tort, L. Roche, T. Ryan, J.P. Dalton, Isolation of a cDNA
encoding Fasciola hepatica cathepsin L2 and functional expression in
Saccharomyces cerevisiae, Mol. Biochem. Parasitol. 88 (1997) 163e174.
[24] P. Berasain, F. Goni, S. McGonigle, A. Dowd, J.P. Dalton, B. Frangione,
C. Carmona, Proteinases secreted by Fasciola hepatica degrade extracel-
lular matrix and basement membrane components, J. Parasitol. 83 (1997)
1e5.
[25] A.J. Dowd, S. McGonigle, J.P. Dalton, Fasciola hepatica cathepsin L
proteinase cleaves fibrinogen and produces a novel type of fibrin clot,
Eur. J. Biochem. 232 (1995) 241e246.
[26] C. Carmona, A.J. Dowd, A.M. Smith, J.P. Dalton, Cathepsin L proteinase
secreted by Fasciola hepatica in vitro prevents antibody-mediated eosin-
ophil attachment to newly excysted juveniles, Mol. Biochem. Parasitol.
62 (1993) 9e17.
[27] P. Berasain, C. Carmona, B. Frangione, J.P. Dalton, F. Goni, Fasciolahepatica: parasite-secreted proteinases degrade all human IgG
subclasses: determination of the specific cleavage sites and identification
of the immunoglobulin fragments produced, Exp. Parasitol. 94 (2000)
99e110.
[28] S.M. O’Neill, K.H. Mills, J.P. Dalton, Fasciola hepatica cathepsin L
cysteine proteinase suppresses Bordetella pertussis-specific interferon-
gamma production in vivo, Parasite Immunol. 23 (2001) 541e547.
[29] R.K. Prowse, P. Chaplin, H.C. Robinson, T.W. Spithill, Fasciola hepatica
cathepsin L suppresses sheep lymphocyte proliferation in vitro and
modulates surface CD4 expression on human and ovine T cells, Parasite
Immunol. 24 (2002) 57e66.
[30] G.V. Hillyer, Fasciola antigens as vaccines against fascioliasis and schis-
tosomiasis, J. Helminthol. 79 (2005) 241e247.
[31] L. Piacenza, D. Acosta, I. Basmadjian, J.P. Dalton, C. Carmona, Vaccina-
tion with cathepsin L proteinases and with leucine aminopeptidase
induces high levels of protection against fascioliasis in sheep, Infect.
Immun. 67 (1999) 1954e1961.
[32] J.P. Dalton, P.J. Brindley, D.P. Knox, C.P. Brady, P.J. Hotez, S. Donnelly,
S.M. O’Neill, G. Mulcahy, A. Loukas, Helminth vaccines: from mining
genomic information for vaccine targets to systems used for protein
expression, Int. J. Parasitol. 33 (2003) 621e640.
[33] N.J. Kennedy, T.W. Spithill, J. Tennent, P.R. Wood, D. Piedrafita, DNA
vaccines in sheep: CTLA-4 mediated targeting and CpG motifs enhance
immunogenicity in a DNA prime/protein boost strategy, Vaccine 24
(2006) 970e979.
[34] N. Reszka, J.B. Cornelissen, M.M. Harmsen, K. Bienkowska-Szewczyk,
J. de Bree, W.J. Boersma, F.A. Rijsewijk, Fasciola hepatica procathepsin
L3 protein expressed by a baculovirus recombinant can partly protect rats
against fasciolosis, Vaccine 23 (2005) 2987e2993.
[35] V.T. Heussler, D.A. Dobbelaere, Cloning of a protease gene family of
Fasciola hepatica by the polymerase chain reaction, Mol. Biochem.
Parasitol. 64 (1994) 11e23.
[36] J.A. Irving, T.W. Spithill, R.N. Pike, J.C. Whisstock, P.M. Smooker, The
evolution of enzyme specificity in Fasciola spp, J. Mol. Evol. 57 (2003)
1e15.
[37] P.M. Smooker, J.C. Whisstock, J.A. Irving, S. Siyaguna, T.W. Spithill,
R.N. Pike, A single amino acid substitution affects substrate specificity
in cysteine proteinases from Fasciola hepatica, Protein. Sci. 9 (2000)
2567e2572.
[38] C.M. Stack, C.R. Caffrey, S.M. Donnelly, A. Seshaadri, J. Lowther,
J.F. Tort, P.R. Collins, M.W. Robinson, W. Xu, J.H. McKerrow,
C.S. Craik, S.R. Geiger, R. Marion, L.S. Brinen, J.P. Dalton, Structural
and functional relationships in the virulence-associated cathepsin L
proteases of the parasitic liver fluke, Fasciola hepatica, J. Biol. Chem.
283 (2008) 9896e9908.
[39] M.W. Robinson, J.F. Tort, E. Wong, S.M. Donelly, J. Lowther, W. Xu,
C. Stack, M. Padula, B. Herbert, J.P. Dalton, Proteomic and phylogenetic
analysis of the cathepsin L protease family of helminth pathogen
Fasciola hepatica: expansion of a repertoire of virulence-associated
factors, Mol. Cell Proteomics 7 (2008) 1111e1123.
[40] J. Tkalcevic, K. Ashman, E. Meeusen, Fasciola hepatica: rapid identifi-
cation of newly excysted juvenile proteins, Biochem. Biophys. Res.
Commun. 213 (1995) 169e174.
[41] J. Tkalcevic, M.R. Brandon, E.N. Meeusen, Fasciola hepatica: rapid
switching of stage-specific antigen expression after infection, Parasite
Immunol. 18 (1996) 139e147.
[42] R.H. Law, P.M. Smooker, J.A. Irving, D. Piedrafita, R. Ponting,
N.J. Kennedy, J.C. Whisstock, R.N. Pike, T.W. Spithill, Cloning and
expression of the major secreted cathepsin B-like protein from juvenile
Fasciola hepatica and analysis of immunogenicity following liver fluke
infection, Infect. Immun. 71 (2003) 6921e6932.
[43] L.R. Wilson, R.T. Good, M. Panaccio, G.L. Wijffels, R.M. Sandeman,
T.W. Spithill, Fasciola hepatica: characterization and cloning of the

1475M. Cancela et al. / Biochimie 90 (2008) 1461e1475
major cathepsin B protease secreted by newly excysted juvenile liver
fluke, Exp. Parasitol. 88 (1998) 85e94.
[44] K. Meemon, R. Grams, S. Vichasri-Grams, A. Hofmann, G. Korge,
V. Viyanant, E.S. Upatham, S. Habe, P. Sobhon, Molecular cloning and
analysis of stage and tissue-specific expression of cathepsin B encoding
genes from Fasciola gigantica, Mol. Biochem. Parasitol. 136 (2004) 1e10.
[45] M.M. Harmsen, J.B. Cornelissen, H.E. Buijs, W.J. Boersma,
S.H. Jeurissen, F.J. van Milligen, Identification of a novel Fasciola hepat-ica cathepsin L protease containing protective epitopes within the
propeptide, Int. J. Parasitol. 34 (2004) 675e682.
[46] F.J. van Milligen, J.B. Cornelissen, B.A. Bokhout, Fasciola hepatica: an
antigen fraction derived from newly excysted juveniles, containing an
immunoreactive 32-kDa protein, induces strong protective immunity in
rats, Exp. Parasitol. 94 (2000) 163e171.
[47] B. Ewing, L. Hillier, M.C. Wendl, P. Green, Base-calling of automated
sequencer traces using phred. I. Accuracy assessment, Genome Res. 8
(1998) 175e185.
[48] B. Ewing, P. Green, Base-calling of automated sequencer traces using
phred. II. Error probabilities, Genome Res. 8 (1998) 186e194.
[49] D. Gordon, C. Abajian, P. Green, Consed: a graphical tool for sequence
finishing, Genome Res. 8 (1998) 195e202.
[50] S.F. Altschul, T.L. Madden, A.A. Schaffer, J. Zhang, Z. Zhang,
W. Miller, D.J. Lipman, Gapped BLAST and PSI-BLAST: a new gener-
ation of protein database search programs, Nucleic Acids Res. 25 (1997)
3389e3402.
[51] J.D. Thompson, T.J. Gibson, F. Plewniak, F. Jeanmougin, D.G. Higgins,
The CLUSTAL_X windows interface: flexible strategies for multiple
sequence alignment aided by quality analysis tools, Nucleic Acids Res.
25 (1997) 4876e4882.
[52] C. Carmona, S. McGonigle, A.J. Dowd, A.M. Smith, S. Coughlan,
E. McGowran, J.P. Dalton, A dipeptidylpeptidase secreted by Fasciola
hepatica, Parasitology 109 (Pt 1) (1994) 113e118.
[53] E. Harlow, D. Lane, Antibodies: A Laboratory Manual, Cold Spring
Harbor Laboratory Press, Cold Spring Harbor, 1988.
[54] J. Heukeshoven, R. Dernick, Improved silver staining procedure for fast
staining in PhastSystem Development Unit. I. Staining of sodium
dodecyl sulfate gels, Electrophoresis 9 (1988) 28e32.
[55] J. Kyhse-Andersen, Electroblotting of multiple gels: a simple apparatus
without buffer tank for rapid transfer of proteins from polyacrylamide
to nitrocellulose, J. Biochem. Biophys. Methods 10 (1984) 203e209.
[56] S. Carnevale, M.I. Rodriguez, E.A. Guarnera, C. Carmona, T. Tanos,
S.O. Angel, Immunodiagnosis of fasciolosis using recombinant proca-
thepsin L cystein proteinase, Diagn. Microbiol. Infect Dis. 41 (2001)
43e49.
[57] F. Battistoni, R. Platero, R. Duran, C. Cervenansky, J. Battistoni,
A. Arias, E. Fabiano, Identification of an iron-regulated, hemin-binding
outer membrane protein in Sinorhizobium meliloti, Appl. Environ. Micro-
biol. 68 (2002) 5877e5881.
[58] U. Hellman, Sample preparation from SDS-PAGE and in-gel digestion, in:
P. Jolles, H. Jornvall (Eds.), Proteomics in Functional Genomics. Protein
Structure Analysis, Birkhauser, Basel, Switzerland, 2000, pp. 43e54.
[59] R. Grams, S. Vichasri-Grams, P. Sobhon, E.S. Upatham, V. Viyanant,
Molecular cloning and characterization of cathepsin L encoding genes
from Fasciola gigantica, Parasitol. Int. 50 (2001) 105e114.
[60] C.E. Bennett, L.T. Threadgold, Fasciola hepatica: development of
tegument during migration in mouse, Exp. Parasitol. 38 (1975) 38e55.
[61] J. Creaney, L. Wilson, M. Dosen, R.M. Sandeman, T.W. Spithill,
J.C. Parsons, Fasciola hepatica: irradiation-induced alterations in carbo-
hydrate and cathepsin-B protease expression in newly excysted juvenile
liver fluke, Exp. Parasitol. 83 (1996) 202e215.
[62] J.P. Dalton, S.O. Neill, C. Stack, P. Collins, A. Walshe, M. Sekiya,
S. Doyle, G. Mulcahy, D. Hoyle, E. Khaznadji, N. Moire, G. Brennan,
A. Mousley, N. Kreshchenko, A.G. Maule, S.M. Donnelly, Fasciola
hepatica cathepsin L-like proteases: biology, function, and potential in
the development of first generation liver fluke vaccines, Int. J. Parasitol.
33 (2003) 1173e1181.
[63] S.A. Beckham, R.H. Law, P.M. Smooker, N.S. Quinsey, C.R. Caffrey,
J.H. McKerrow, R.N. Pike, T.W. Spithill, Production and processing of
a recombinant Fasciola hepatica cathepsin B-like enzyme (FhcatB1)
reveals potential processing mechanisms in the parasite, Biol. Chem.
387 (2006) 1053e1061.
[64] R.M. Morphew, H.A. Wright, E.J. Lacourse, D.J. Woods, P.M. Brophy,
Comparative proteomics of excretory-secretory proteins released by the
liver fluke Fasciola hepatica in sheep host bile and during in vitro culture
ex host, Mol. Cell Proteomics 6 (2007) 963e972.
[65] F. Lecaille, S. Chowdhury, E. Purisima, D. Bromme, G. Lalmanach, The
S2 subsites of cathepsins K and L and their contribution to collagen
degradation, Protein Sci. 16 (2007) 662e670.
[66] J.R. Jefferies, A.M. Campbell, A.J. van Rossum, J. Barrett, P.M. Brophy,
Proteomic analysis of Fasciola hepatica excretory-secretory products,
Proteomics 1 (2001) 1128e1132.
[67] L. McGonigle, A. Mousley, N.J. Marks, G.P. Brennan, J.P. Dalton,
T.W. Spithill, T.A. Day, A.G. Maule, The silencing of cysteine proteases
in Fasciola hepatica newly excysted juveniles using RNA interference
reduces gut penetration, Int. J. Parasitol. 38 (2008) 149e155.
[68] J. Dvorak, M. Delcroix, A. Rossi, V. Vopalensky, M. Pospisek,
M. Sedinova, L. Mikes, M. Sajid, A. Sali, J.H. McKerrow, P. Horak,
C.R. Caffrey, Multiple cathepsin B isoforms in schistosomula of Tricho-
bilharzia regenti: identification, characterisation and putative role in
migration and nutrition, Int. J. Parasitol. 35 (2005) 895e910.
[69] Y.B. Chung, Y. Kong, H.J. Yang, S.Y. Kang, S.Y. Cho, Cysteine protease
activities during maturation stages of Paragonimus westermani, J. Para-
sitol. 83 (1997) 902e907.
[70] H. Park, S.I. Kim, K.M. Hong, M.J. Kim, C.H. Shin, J.S. Ryu, D.Y. Min,
J.B. Lee, U.W. Hwang, Characterization and classification of five cyste-
ine proteinases expressed by Paragonimus westermani adult worm, Exp.
Parasitol. 102 (2002) 143e149.
[71] N. Kaewpitoon, T. Laha, S. Kaewkes, P. Yongvanit, P.J. Brindley,
A. Loukas, B. Sripa, Characterization of cysteine proteases from the car-
cinogenic liver fluke, Opisthorchis viverrini, Parasitol. Res. 102 (2008)
757e764.
[72] B.K. Na, J.M. Kang, W.M. Sohn, CsCF-6, a novel cathepsin F-like
cysteine protease for nutrient uptake of Clonorchis sinensis, Int. J. Para-
sitol. 38 (2008) 493e502.
[73] T.H. Kang, D.H. Yun, E.H. Lee, Y.B. Chung, Y.A. Bae, J.Y. Chung,
I. Kang, J. Kim, S.Y. Cho, Y. Kong, A cathepsin F of adult Clonorchis
sinensis and its phylogenetic conservation in trematodes, Parasitology
128 (2004) 195e207.
[74] G. Rinaldi, M.E. Morales, M. Cancela, E. Castillo, P.J. Brindley, J.F.
Tort, Development of functional genomic tools in trematodes: RNA inter-
ference and luciferase reporter gene activity in Fasciola hepatica, PLoS
Neglected Trop. Dis., in press.


















