
Activated T cells recruit exosomes secreted by dendritic cells ...
Upload
independentCategory
view
2download
0
Expression of Cathepsins B, L, S, and D byGastric Epithelial Cells Implicates Them asAntigen Presenting Cells in Local ImmuneResponses
Carlos A. Barrera, Gang Ye,Rosario Espejo, Shani Gunasena, Ruben Almanza,James F. Leary, Sheila E. Crowe, Peter B. Ernst,and Victor E. Reyes
ABSTRACT: Helicobacter pylori infection is linked tochronic gastritis, peptic ulcer and gastric carcinoma. Dur-ing H. pylori infection, class II MHC expression by thegastric epithelium increases, as does the number of localCD4� T cells, which appear to be important in theassociated pathogenesis. These observations suggestedthat the epithelium might present antigens to T cells.Thus, we sought to determine whether gastric epithelialcells process antigens to establish their function as localantigen presenting cells (APC). We examined a panel ofgastric epithelial cell lines for expression of the antigenprocessing cathepsins B (CB), L (CL), S (CS), and D (CD).The mRNA for these enzymes were detected by RT-PCRand the enzymes in the gastric epithelial cells were iden-tified by various independent methods. We corroborated
the expression of CB and CD on gastric epithelial cellsfrom human biopsy samples. The functions of these pro-teases were confirmed by assessing their ability to digestovalbumin, a conventional dietary antigen, and proteinsfrom H. pylori. In summary, multiple lines of evidencesuggest gastric epithelial cells process antigens for pre-sentation to CD4� T cells. To our knowledge, these arethe first studies to document the antigen processing ca-pacity of human gastric epithelial cells. Human Immu-nology 62, 1081–1091 (2001). © American Society forHistocompatibility and Immunogenetics, 2001. Pub-lished by Elsevier Science Inc.
KEYWORDS: antigen presentation; endosomal pro-teases; Helicobacter pylori; mucosal immunity
ABBREVIATIONSAPC antigen presenting cellsCB cathepsin BCL cathepsin LCS cathepsin SCD cathepsin DEBV Epstein-Barr virus
Ii class II MHC-associated invariant chainECL enhanced chemiluminescenceHLA human leukocyte antigenIFN� interferon gammaMHC major histocompatibility complexOVA ovalbumin
INTRODUCTIONHeliobacter pylori infection is recognized as the majorcause of chronic gastritis and is also associated with most
gastric and duodenal ulcers and gastric carcinomas. H.pylori is the first bacterial agent to be classified as a classI carcinogen [1]. The interactions between H. pylori andthe host that determine disease or protection are not
From the Departments of Pediatrics (C.B., G.Y., R.E., S.G., R.A.,P.B.E., V.E.R.), Internal Medicine (J.F.L., S.E.C.), Pathology (J.F.L.),Microbiology and Immunology (C.B., S.E.C., P.B.E., V.E.R., J.F.L.), andthe Sealy Center for Molecular Sciences (P.B.E.), University of TexasMedical Branch, Galveston, TX, USA.
Address reprint requests to: Dr. Victor E. Reyes, Route 0366, Children’sHospital, 301 University Boulevard, UTMB, Galveston, TX 77555-
0366; Tel: (409) 772-1750/1751; Fax: (409) 772-1761; E-mail:[email protected].
Received December 27, 2000; revised February 21, 2001; accepted May23, 2001.
Human Immunology 62, 1081–1091 (2001)0198-8859/01/$–see front matter© American Society for Histocompatibility and Immunogenetics, 2001
Published by Elsevier Science Inc. S0198-8859(01)00281-6

clear. The human gastric epithelium constitutively ex-presses major histocompatibility complex (MHC) class IImolecules, and this expression increases during infectionwith H. pylori [2–4]. These observations, together with aparallel rise in T lymphocyte numbers within gastricbiopsy specimens [5, 6] from patients with H. pylori-associated diseases, suggested that the gastric epitheliummay actively participate in antigen presentation in theinitiation of local immune responses.
In addition to class II MHC expression, another keycharacteristic of antigen presenting cells (APC) is theirability to proteolytically process endocytosed antigensinto peptide fragments of a size suitable for binding byclass II MHC molecules. Among the proteases impli-cated in the processing of foreign antigens are cathepsinsB, L, S, D, and E [7–11]. The individual niche of thesecathepsins in generating antigenic peptides recognizedby CD4� T cells has been well documented in multipleexperimental systems. The use of selective inhibitors forcysteine and aspartyl proteases independently on APCwas found to prevent formation of ovalbumin (OVA)peptides, which could be recognized by OVA-specificT-cell clones [8]. These data suggested that cysteine andaspartic proteases may be important in generating suit-able peptide epitopes for presentation by class II MHC toactivate specific T cells [12]. Because these proteaseshave an optimal pH in the acidic range, inhibition ofendosomal acidity prevents antigen processing. In addi-tion to their antigen processing functions, cathepsins B,S, and D have been described to play a role in theproteolytic removal of the class II MHC-associated in-variant chain (Ii) that, in turn, permits the binding ofantigenic peptides by class II MHC molecules [11–14].
Because cathepsins B, L, S and D play a prominentrole in the processing functions of APCs, the expressionof these proteases by the human gastric epithelium has tobe established to support the role of the epithelium inregulating local immunity. Although recent studies haveimplicated cathepsin E in presentation of some antigens[15], its overall involvement in APC functions appearslimited and is less well characterized than those of cathe-psins B, L, S, and D. Thus, in the present study weexamined the expression of cathepsins B, L, S, and D andtheir function in human gastric epithelial cells.
MATERIALS AND METHODSCells Lines and Culture ConditionsJesthom, a previously described HLA-DR1 homozygousEBV-transformed B-cell line [13], was used as a proto-typic APC. KATO III, RF1, RF48, and N87 humangastric epithelial cell lines were from ATCC (Rockville,MD, USA). Jesthom, KATO III, and N87 were grown inRPMI 1640 medium supplemented with 10% fetal calf
serum (FCS), glutamine (1 mM), penicillin (1 U/ml), andstreptomycin (100 mg/ml). RF1 and RF48 were grownin Leibovitz’s L-15 (10% FCS).
BacteriaHelicobacter pylori LC-11 (Cag A�) was originally isolatedfrom the antral mucosa of a patient with duodenal ulcersas previously described [16]. The bacteria were grown onblood-agar plates (Becton Dickinson, Mountain View,CA, USA) at 37 °C under microaerophilic conditions for4 days before being metabolically labeled with 3 �Ci of[35S] methionine and cysteine for 4 h in the presence ofRPMI methionine- and cysteine-free medium. The bac-terial pellet was then washed five times with PBS (0.01MNa3 PO4, 0.005 M MgSO4), and then 30 �g/ml ofDNAse was added prior to sonication. The bacteria weresonicated and lysed on ice using four cycles of 50%vibration with intervals of 5 min per set. The bacteriawere then centrifuged at 10,000 rpm for 15 min, and thesupernatant was collected. The extracted proteins werethen measured by optical density (OD) at an absor-bance of 280 nm using a DU-65 spectrophotometer(Beckman).
AntibodiesAntihuman CB (rabbit IgG) was from Cortex Biochem(San Leandro, CA, USA). Antihuman CD (goat IgG) wasa gift from Dr. Annette Hille-Rehfeld (Georg-August-Universitat, Gottingen, Germany). Other antibodieswere rabbit antihuman CD (Calbiochem, La Jolla, CA);anticathepsin L (rabbit serum; Athens Research andTechnology, Inc, Athens, GA, USA); Ber-EP4 mouse(IgG) anti-epithelial antigen (DAKO Corporation,Glostrup, Denmark); and FITC-conjugated goat antirab-bit-IgG and R-Phycoerythrin-conjugated goat anti-mouse antibodies (Caltag Laboratories, San Francisco,CA, USA). Antihuman Ii (CD74) monoclonal antibodyVicY-1 was a gift from Dr. Walter Knapp (Institute ofImmunology, University of Vienna, Vienna, Austria).The hybridoma secreting antihuman class II MHCmonoclonal antibody 9.3F10 was obtained from ATCCand used in developing antibody-containing ascites fluid.Antihuman Ii monoclonal antibody BU45 was obtainedfrom The Binding Site (Birmingham, United Kingdom).
Isolation of Epithelial Cells From Gastric BiopsiesGastric antrum biopsy tissues were obtained from con-senting donors undergoing gastro-esophageal-duodenos-copy. The biopsies were obtained from areas that weregrossly inflamed, as well as regions that appeared unin-volved. The tissue was acquired following informed con-sent from patients and volunteers, and with prior ap-proval of the institutional IRB. The biopsy tissue wasplaced in Ca2�- and Mg2�-free HBSS supplemented
1082 C. A. Barrera et al.

with 5% fetal bovine serum (FBS), and transported im-mediately to the laboratory. The tissue was then placedin HBSS containing 0.1-mM EDTA and 0.1-mM dithio-threitol (DTT) and incubated at 37 °C for 15 min withvigorous shaking. The supernatant was decanted andreplaced with fresh HBSS-EDTA-DTT solution and in-cubated another 15 min. Then, the biopsy sample wasplaced in 1�(2.4 U/ml) dispase solution (BoehringerMannheim, Mannheim, Germany) and incubated at 37°C for 15 min with vigorous shaking. The supernatantwas removed and the cells were pelleted by centrifuga-tion at 200�g for 5 min. Fresh dispase solution wasadded to the biopsy tissue and the above procedure wasrepeated. The cells isolated were identified as mostly(� 90%) epithelial cells, following May-Grunwald-Gi-emsa staining (Sigma, St. Louis, MO, USA) and flowcytometry with immunofluorescence staining with Ber-EP4 mouse antihuman epithelial antigen (see below).The freshly isolated cells were used immediately forintra-cytoplasmic staining as described below.
Reverse Transcriptase–Polymerase Chain ReactionTotal RNA was extracted from gastric epithelial cellsand B cells by using TRIzol (Life Technologies Inc.,Grand Island, NY, USA) according to the manufacturer’sinstructions. The RNA preparation was reverse-tran-scribed with 0.1 �M of oligo primers, 1 mM of eachdNTP and 50 U MuLV reverse transcriptase (Perkin-Elmer, Branchburg, NJ, USA) and then incubated at 42°C for 15 min, 92 °C for 5 min, and 5 °C for 5 min. Theresulting cDNA was then amplified by polymerase chainreaction (PCR) in a reaction mixture consisting of 1-mMMgCl2, 50-mM KCl, 0.2 mM of each dNTP, 0.2 �M ofeach primer, and 2.5-U amplitaq DNA polymerase (Per-kin Elmer). All reactions were overlaid with 50-�l min-eral oil to prevent evaporation. PCR was carried out in aprogrammed thermal cycler (Perkin Elmer Cetus, Em-eryville, CA, USA) as follows: CS and CB at 40 cycles of1 min at 92 °C, 1 min at 60 °C, and 1 min at 72 °C; andfor CD and CL at 35 cycles of 1 min at 92 °C, 1 min at43 °C, and 1 min at 72 °C. The primers were designedusing a Gene-runner software program and synthesizedlocally (Dr. T. Wood, UTMB Sealy Center for MolecularScience, Galveston, TX, USA) using an automated DNAsynthesizer (Model 394, Applied Biosystems Inc., FosterCity, CA, USA). The CB primers were 5�-TCGGATGAGCTGGTCAACTATG-3� (sense) and 5�-TCCAAGCTTCAGCAGGATAG-3� (antisense); CL 5�-GACAGGGACTGGAAGAGAG-3� (sense) and 5�-GTTTCCCTTCCCTGTATTC-3� (antisense); CS 5�-GGGTACCTCATGTGACAAG-3� (sense) and 5�-TCACTTCTTCACTGGTCATG-3� (antisense); and CD, 5�-ACTGTATTTCCATGTGAGC-3� (sense) and 5�-CAGAGAGTC(AGCT)ACCTCCCAG-3� (antisense) primers.
The expected products are 400 bp (CB) and 390 bp(CL, CS, and CD). Amplified products were analyzedby 1.5% agarose gel electrophoresis and ultravioletFoto/Prep I (Fotodyne Inc, New Berlin, WI, USA) forvisualization and photography. As an internal control,the RNA preparations were processed by reverse tran-scriptase–polymerase chain reaction (RT-PCR) to am-plify the housekeeping gene of �-actin. The �-actinprimers were 5�-ATCTGGCACCACACCTTCTACAATGAGCTGCG-3� (sense) and 5�-CGTCATACTCCTGCTTGCTGATCCACATCTGC-3� (antisense) synthe-sized locally at UTMB and the expected product is 800bp.
Flow Cytometry StudiesTo detect the intracellular compartments containingcathepsins B, D, and L on gastric biopsies and cell lines,1�106 gastric epithelial cells were treated with monen-sin (10 �M) and incubated at 37 °C, 5% CO2 for 3 h.The cells were then fixed with 2% of paraformaldehydein PBS pH 7.4 for 10 min, followed by permeabilizationwith 0.1% of saponin in PBS, pH 7.4, supplementedwith 1% FBS, 1% BSA and the protease inhibitorsleupeptin (10 mM) and pepstatin-A (5�10�8 M) for 1 h.The cells were incubated on ice for 45 min with Ber-EP4mouse antihuman epithelial antigen and rabbit anti-human CB, rabbit anti-human CL or anti-human CDantibodies, respectively. After washing three times withPBS, the cells were incubated on ice for 45 min withFITC-conjugated goat antirabbit IgG and/or goat anti-mouse IgG-R phycoerythrin-conjugated antibodies.Then, the cells were washed and fixed with 2% parafor-maldehyde in PBS. The cells were then analyzed in aFACScan cytometer (Becton Dickinson, Mountain View,CA, USA).
Metabolic Radiolabeling and LysisKato III cells were incubated in methionine- and cys-teine-free RPMI-1640 medium for 1 h and then meta-bolically radiolabeled with [35S] methionine and cysteine(0.5 mCi/108 cells). Cells were then washed and lysed inice-cold 10-mM Tris-HCl buffer, pH 8.1, containingprotease inhibitors (2-mM phenyl L-methyl sulfonyl flu-oride [PMSF] and 1.0-mM N-ethyl maleimide [NEM])for 30 min in ice. Nuclei were removed by centrifugationat 500�g for 10 min. This process was repeated anadditional time to ensure that all cells were lysed. Mi-crosomal membranes were obtained from the superna-tants by ultra-centrifugation at 100,000�g for 45 min.The membrane pellet was solubilized in the lysis buffercontaining 0.5% Triton X-100, followed by a secondcentrifugation at 100,000�g for 45 min to remove theunsolubilized material.
1083Antigen Processing Proteases in the Gastric Epithelium

Immunoprecipitation and ElectrophoresesSolubilized membrane samples were precleared with nor-mal rabbit serum and formalinized Staphylococcus aureusCowen strain A (Chemicon, El Segundo, CA, USA).Immunoadsorbants (Iad) were prepared by incubating100 �l of protein A-sepharose (Sigma Chemical Co.)with approximately 1 �g of either normal rabbit serum,rabbit antihuman CB or rabbit anti-human CD. After16-h incubation at 4 °C with solubilized membraneproteins, the Iad were washed five times with Iad buffer(50-mM Tris-HCl, pH 7.4, 150-mM NaCl, 5-mMEDTA, 0.5% [V/V] Triton X-100, and 0.02% [W/V]NaN3) and eluted with sodium duodecyl sulfate (SDS)sample buffer (62.5-mM Tris-HCl, pH 6.8, 10% [W/V]glycerol, 5% 2-mercaptoethanol, and 2.3% SDS). Thesamples were analyzed on 12.5% acrylamide gels byone-dimensional SDS-polyacrylamide gel electrophoresis(SDS-PAGE) as previously reported [13].
Affinity Selection of Cathepsin D by Pepstatin-AAgaroseGastric epithelial cells (1�107) were metabolically la-beled with [35S] methionine and cysteine; their proteinswere obtained by lysis and solubilization with 1% TritonX-100 (W/V) in PBS. After incubation for 30 min at 4°C the unsolubilized material was removed by centrifu-gation at 50000 rpm for 15 min. The supernatants weremixed 1/10 with 1-M sodium acetate buffer pH 4.0.These supernatants were incubated with pepstatin-Aagarose beads (Pierce, Rockford, IL, USA) overnight at 4°C. The beads were washed five times with 0.1-M so-dium acetate, 1-M NaCl pH 4.5; then twice with 0.1-MTris, 1-M NaCl pH 8.6; and an additional five timeswith 0.1-M sodium acetate, 1-M NaCl pH 4.5, used forbinding. Bound proteins were eluted with 0.1-M sodiumacetate, 1-M NaCl pH 8.5, and collected in fractions.The recovered fractions were separated by SDS-PAGE gelelectrophoresis.
Western Blot Identification and Confirmationof CB, CL, and Affinity-Purified CDAfter affinity selection of CD by pepstatin-A agarose, theeluted proteins and commercial, pure human CD (SigmaChemical Co.) were separated by SDS-PAGE gel electro-phoresis. In the case of cathepsin B and L, gastric epi-thelial cells were lysed with 10-mM Tris, 1% TritonX-100, pH 8.4 buffer. Then, proteins, commercial purehuman CB (Sigma Chemical Co.), and pure human ca-thepsin L (Sigma Chemical Co.) were examined on SDS-PAGE gel electrophoresis. In all cases, for western blot-ting purposes proteins were separated using precasted4%–20% acrylamide mini gels (Bio-Rad Laboratories,Hercules, CA, USA), and then transferred onto a nitro-cellulose membranes (Schleicher & Schuell, Inc). Non-
specific binding sites were blocked by incubating themembranes at 4 °C overnight in 0.2% Tween-20 in PBS(T-PBS) containing 10% casein. After washing in T-PBS, the membrane was incubated at room temperaturefor 4 h with rabbit anti-CD (1:30 dil), or rabbit antihu-man CB (1:300 dil), or rabbit antihuman CL (1:30)according to the manufacturer’s instructions. After sev-eral washes with T-PBS, the membranes were incubatedfor an additional 1 h at room temperature with horse-radish peroxidase-conjugated goat antirabbit antibody(1:8000). After extensive washing, the proteins to whichthe antibodies bound were detected with enhancedchemiluminescence (ECL; Amersham, Inc., Piscataway,NJ, USA) and autoradiographed.
Metabolic Pulse-Chase LabelingKato III cells (6.2�106) in the log phase of growth werewashed and incubated in methionine-and serum-freeRPMI 1640 medium for 1 h prior to labeling. Then highconcentrations of [35S] methionine (1.5 mCi/6.2�107
cells; ICN Pharmaceuticals, Irvine, CA, USA) wereadded for 15 min. Subsequently, cells were washed anddivided in five equal parts and chased with cold RPMIsupplemented with 10% FBS in the presence of leupep-tin (100 �g/ml) for 0, 1, 2, 3, and 5 h. At the end of eachtime point, microsomal membrane proteins were pre-pared from [35S] methionine and cysteine labeled cells bysolubilization with 0.01 M Tris-HCl buffer, pH 7.0,containing 0.5% Triton X-100 and finally immunopre-cipitated.
Protease InhibitorsThe cysteine proteases (cathepsin B, L, and S) inhibitorsleupeptin and E-64 ([N-L-3-trans- carboxyoxine-2-car-bonyl]-L-leucine-3-methylbutylamide), and the asparticprotease (cathepsin D) inhibitor pepstatin-A were pur-chased from Sigma Chemical Company.
Protein Digestion by CB, CL, CS, and CD[14C]-labeled ovalbumin (Sigma Chemical Co.) was usedas an antigen. Immediately before digestion ovalbuminwas reduced with 2� SDS sample buffer for 10 min atroom temperature and boiled for 3 min. Then, the pro-tein was dialyzed using a Centricon 10 filter unit(Pierce). The cells were lysed with 20-nM Hepes and 1%(W/V) Triton X-100, pH 6.8 buffer. The proteases in thecytosolic contents were then activated with 1-mM DTT,1.33-mM EDTA, and 2.7-mM cysteine in PBS pH 6.0,for 5 min at 40 °C, and next incubated together with theprotein antigen for 0.5, 1, 3, and 5 h in the presence orabsence of the cysteine protease inhibitor E-64, and theaspartic protease inhibitor pepstatin-A.
1084 C. A. Barrera et al.

RESULTSHuman Gastric Epithelial Cell Lines ExpressmRNA of Cathepsins B, L, S, and DRT-PCR reactions were performed to initially establishthe expression of mRNA of CB, CL, CS, and CD. Theseendosomal proteases are present in KATO III and N87gastric epithelial cells. Jesthom B cells were used asinternal controls (Figure 1).
To validate the previous findings, flow cytometryanalysis was performed to detect the intracellular expres-sion of cysteine proteases CB, CL, and aspartic protease
CD in a panel of gastric epithelial cell lines and com-pared with the staining of a B-cell line, Jesthom as aprototypic APC (Figure 2). Although the level of expres-sion was variable, as illustrated by relative fluorescenceintensity, this study confirmed that the expression ofboth proteases was not restricted to one cell line. Inaddition to KATO III and N87, RF-1 and RF-48 gastricepithelial cell lines were examined and confirmed theexpression of these enzymes (data not shown).
To establish the specificity of the antiserum used inboth immunofluorescence and flow cytometry studies,we immunoprecipitated or selectively isolated by affin-ity, the respective proteins from gastric epithelial cells,which were metabolically labeled with [35S] methionine.Anti-cathepsin B antibodies immunoprecipitated pro-teins of 31, 27, and 25 kD (Figure 3). The expression of31kD and 25kD bands is consistent with the molecularweights of the proenzyme and mature forms of the en-zyme respectively previously reported [17]. These bandswere confirmed as isoforms of CB by Western blot usingthe rabbit antiserum (Figure 3). The anti-human CD
FIGURE 1 Cathepsins B, L, S, and D mRNA detected ingastric epithelial cells by RT-PCR analysis. RNA extractedfrom KATO III and N87 gastric epithelial cells were reverse-transcribed, amplified, and separated by electrophoresis. Spe-cific products were detected in these cells by RT-PCR usingthe CB, CS, CL, and CD primers described in the materials andmethods section. As internal control, B (Jesthom) cells exhib-ited mRNA for those cathepsins. The amplification of house-keeping gene �-actin was included as a control for the proce-dure.
FIGURE 2 Detection of Cathepsin B, L, and D by flowcytometry in gastric epithelial cell lines. KATO III and N87gastric epithelial cells and Jesthom B cells were treated withmonensin 3 h prior to fixation and permeabilization with 0.1%saponin and 3% BSA in PBS. The permeabilized cells werethen incubated with rabbit antihuman cathepsin B, cathepsinL, or cathepsin D antibodies, followed by FITC-conjugatedgoat antirabbit secondary antibody. X axis relative fluores-cence intensity; Y axis number of cells.
1085Antigen Processing Proteases in the Gastric Epithelium
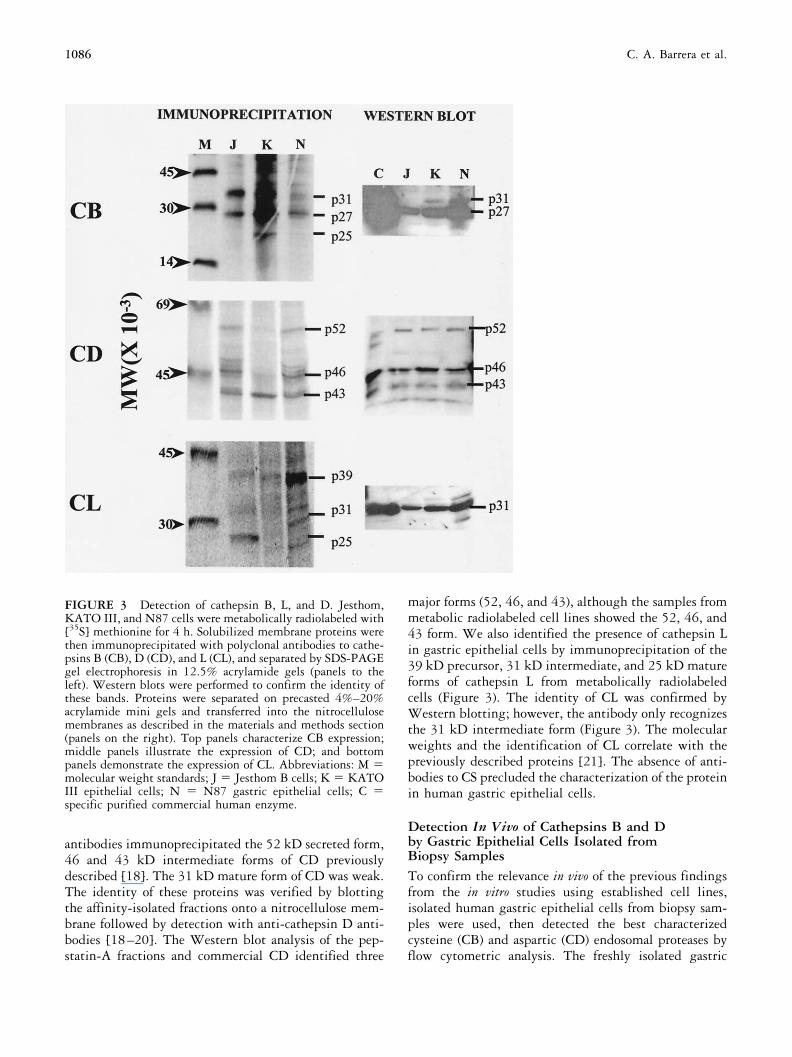
antibodies immunoprecipitated the 52 kD secreted form,46 and 43 kD intermediate forms of CD previouslydescribed [18]. The 31 kD mature form of CD was weak.The identity of these proteins was verified by blottingthe affinity-isolated fractions onto a nitrocellulose mem-brane followed by detection with anti-cathepsin D anti-bodies [18–20]. The Western blot analysis of the pep-statin-A fractions and commercial CD identified three
major forms (52, 46, and 43), although the samples frommetabolic radiolabeled cell lines showed the 52, 46, and43 form. We also identified the presence of cathepsin Lin gastric epithelial cells by immunoprecipitation of the39 kD precursor, 31 kD intermediate, and 25 kD matureforms of cathepsin L from metabolically radiolabeledcells (Figure 3). The identity of CL was confirmed byWestern blotting; however, the antibody only recognizesthe 31 kD intermediate form (Figure 3). The molecularweights and the identification of CL correlate with thepreviously described proteins [21]. The absence of anti-bodies to CS precluded the characterization of the proteinin human gastric epithelial cells.
Detection In Vivo of Cathepsins B and Dby Gastric Epithelial Cells Isolated fromBiopsy SamplesTo confirm the relevance in vivo of the previous findingsfrom the in vitro studies using established cell lines,isolated human gastric epithelial cells from biopsy sam-ples were used, then detected the best characterizedcysteine (CB) and aspartic (CD) endosomal proteases byflow cytometric analysis. The freshly isolated gastric
FIGURE 3 Detection of cathepsin B, L, and D. Jesthom,KATO III, and N87 cells were metabolically radiolabeled with[35S] methionine for 4 h. Solubilized membrane proteins werethen immunoprecipitated with polyclonal antibodies to cathe-psins B (CB), D (CD), and L (CL), and separated by SDS-PAGEgel electrophoresis in 12.5% acrylamide gels (panels to theleft). Western blots were performed to confirm the identity ofthese bands. Proteins were separated on precasted 4%–20%acrylamide mini gels and transferred into the nitrocellulosemembranes as described in the materials and methods section(panels on the right). Top panels characterize CB expression;middle panels illustrate the expression of CD; and bottompanels demonstrate the expression of CL. Abbreviations: M molecular weight standards; J Jesthom B cells; K KATOIII epithelial cells; N N87 gastric epithelial cells; C specific purified commercial human enzyme.
1086 C. A. Barrera et al.
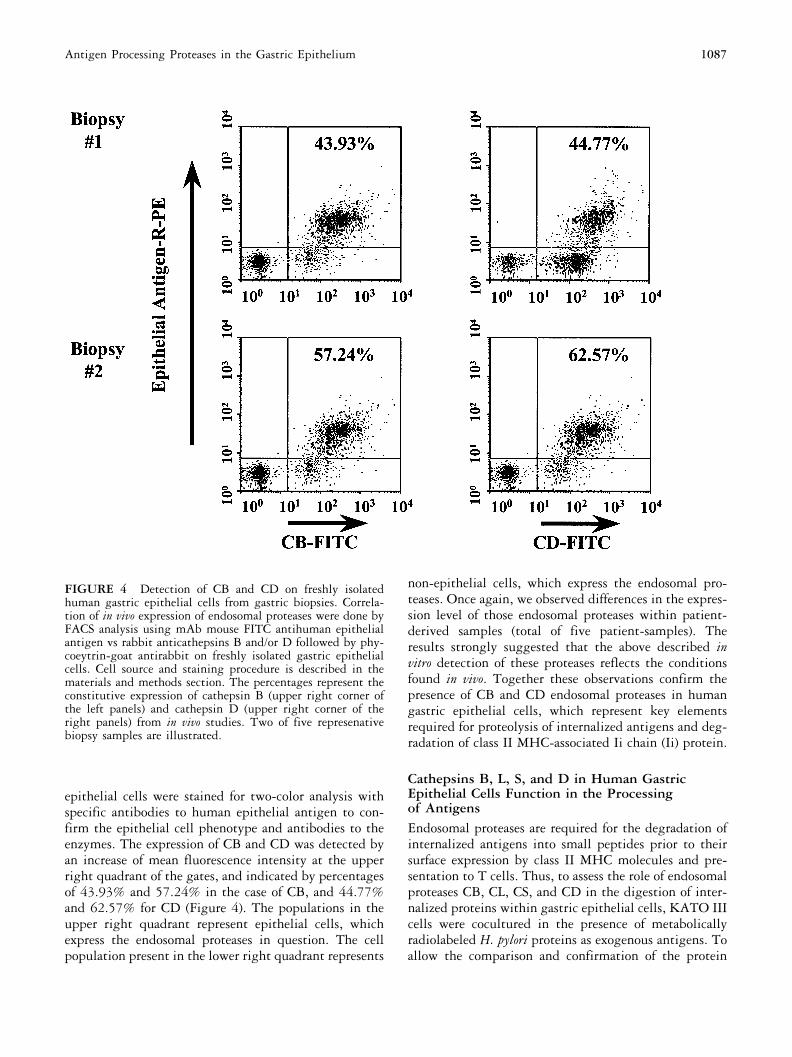
epithelial cells were stained for two-color analysis withspecific antibodies to human epithelial antigen to con-firm the epithelial cell phenotype and antibodies to theenzymes. The expression of CB and CD was detected byan increase of mean fluorescence intensity at the upperright quadrant of the gates, and indicated by percentagesof 43.93% and 57.24% in the case of CB, and 44.77%and 62.57% for CD (Figure 4). The populations in theupper right quadrant represent epithelial cells, whichexpress the endosomal proteases in question. The cellpopulation present in the lower right quadrant represents
non-epithelial cells, which express the endosomal pro-teases. Once again, we observed differences in the expres-sion level of those endosomal proteases within patient-derived samples (total of five patient-samples). Theresults strongly suggested that the above described invitro detection of these proteases reflects the conditionsfound in vivo. Together these observations confirm thepresence of CB and CD endosomal proteases in humangastric epithelial cells, which represent key elementsrequired for proteolysis of internalized antigens and deg-radation of class II MHC-associated Ii chain (Ii) protein.
Cathepsins B, L, S, and D in Human GastricEpithelial Cells Function in the Processingof AntigensEndosomal proteases are required for the degradation ofinternalized antigens into small peptides prior to theirsurface expression by class II MHC molecules and pre-sentation to T cells. Thus, to assess the role of endosomalproteases CB, CL, CS, and CD in the digestion of inter-nalized proteins within gastric epithelial cells, KATO IIIcells were cocultured in the presence of metabolicallyradiolabeled H. pylori proteins as exogenous antigens. Toallow the comparison and confirmation of the protein
FIGURE 4 Detection of CB and CD on freshly isolatedhuman gastric epithelial cells from gastric biopsies. Correla-tion of in vivo expression of endosomal proteases were done byFACS analysis using mAb mouse FITC antihuman epithelialantigen vs rabbit anticathepsins B and/or D followed by phy-coeytrin-goat antirabbit on freshly isolated gastric epithelialcells. Cell source and staining procedure is described in thematerials and methods section. The percentages represent theconstitutive expression of cathepsin B (upper right corner ofthe left panels) and cathepsin D (upper right corner of theright panels) from in vivo studies. Two of five represenativebiopsy samples are illustrated.
1087Antigen Processing Proteases in the Gastric Epithelium
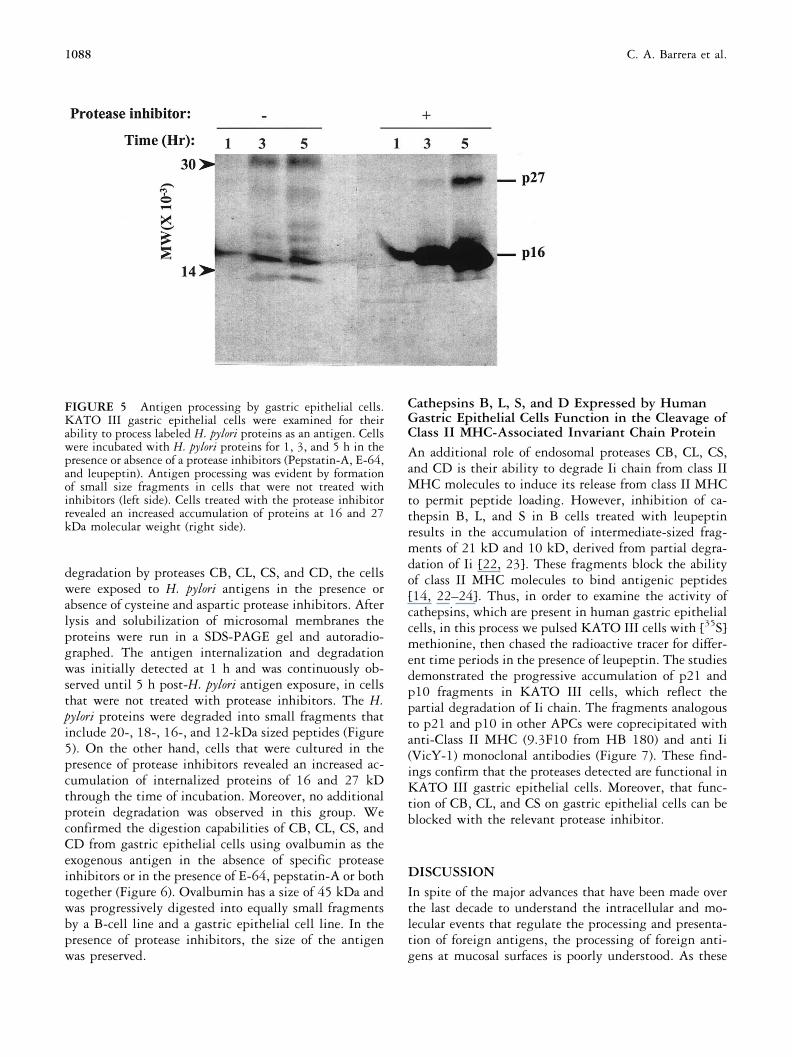
degradation by proteases CB, CL, CS, and CD, the cellswere exposed to H. pylori antigens in the presence orabsence of cysteine and aspartic protease inhibitors. Afterlysis and solubilization of microsomal membranes theproteins were run in a SDS-PAGE gel and autoradio-graphed. The antigen internalization and degradationwas initially detected at 1 h and was continuously ob-served until 5 h post-H. pylori antigen exposure, in cellsthat were not treated with protease inhibitors. The H.pylori proteins were degraded into small fragments thatinclude 20-, 18-, 16-, and 12-kDa sized peptides (Figure5). On the other hand, cells that were cultured in thepresence of protease inhibitors revealed an increased ac-cumulation of internalized proteins of 16 and 27 kDthrough the time of incubation. Moreover, no additionalprotein degradation was observed in this group. Weconfirmed the digestion capabilities of CB, CL, CS, andCD from gastric epithelial cells using ovalbumin as theexogenous antigen in the absence of specific proteaseinhibitors or in the presence of E-64, pepstatin-A or bothtogether (Figure 6). Ovalbumin has a size of 45 kDa andwas progressively digested into equally small fragmentsby a B-cell line and a gastric epithelial cell line. In thepresence of protease inhibitors, the size of the antigenwas preserved.
Cathepsins B, L, S, and D Expressed by HumanGastric Epithelial Cells Function in the Cleavage ofClass II MHC-Associated Invariant Chain ProteinAn additional role of endosomal proteases CB, CL, CS,and CD is their ability to degrade Ii chain from class IIMHC molecules to induce its release from class II MHCto permit peptide loading. However, inhibition of ca-thepsin B, L, and S in B cells treated with leupeptinresults in the accumulation of intermediate-sized frag-ments of 21 kD and 10 kD, derived from partial degra-dation of Ii [22, 23]. These fragments block the abilityof class II MHC molecules to bind antigenic peptides[14, 22–24]. Thus, in order to examine the activity ofcathepsins, which are present in human gastric epithelialcells, in this process we pulsed KATO III cells with [35S]methionine, then chased the radioactive tracer for differ-ent time periods in the presence of leupeptin. The studiesdemonstrated the progressive accumulation of p21 andp10 fragments in KATO III cells, which reflect thepartial degradation of Ii chain. The fragments analogousto p21 and p10 in other APCs were coprecipitated withanti-Class II MHC (9.3F10 from HB 180) and anti Ii(VicY-1) monoclonal antibodies (Figure 7). These find-ings confirm that the proteases detected are functional inKATO III gastric epithelial cells. Moreover, that func-tion of CB, CL, and CS on gastric epithelial cells can beblocked with the relevant protease inhibitor.
DISCUSSIONIn spite of the major advances that have been made overthe last decade to understand the intracellular and mo-lecular events that regulate the processing and presenta-tion of foreign antigens, the processing of foreign anti-gens at mucosal surfaces is poorly understood. As these
FIGURE 5 Antigen processing by gastric epithelial cells.KATO III gastric epithelial cells were examined for theirability to process labeled H. pylori proteins as an antigen. Cellswere incubated with H. pylori proteins for 1, 3, and 5 h in thepresence or absence of a protease inhibitors (Pepstatin-A, E-64,and leupeptin). Antigen processing was evident by formationof small size fragments in cells that were not treated withinhibitors (left side). Cells treated with the protease inhibitorrevealed an increased accumulation of proteins at 16 and 27kDa molecular weight (right side).
1088 C. A. Barrera et al.

are properties ascribed to class II MHC-expressing cells,the role of class II MHC-expressing gastric epithelialcells had to be considered in the handling of H. pyloriantigens. Bearing in mind that the epithelium is themost abundant class II MHC-expressing cell phenotypelocally, it is important to examine their antigen process-ing ability.
In the present studies, direct evidence that humangastric epithelial cells express the best-characterized pro-cessing enzymes (cathepsins B, L, S, and D) has beenprovided. The presence of these enzymes was recognizedand confirmed by several independent approaches, whichincluded RT-PCR, flow cytometry, immunoprecipita-tion and isolation by affinity chromatography. Polyacryl-amide gel electrophoresis of the immunoprecipitatedsamples or affinity purified samples revealed the presenceof precursor (31 kD), intermediate (27 kD), and mature(25 kD) forms of cathepsin B [17]. We also detected theproform (39 kD) and intermediate form (31 kD) ofcathepsin L previously described [21], as well as thesecreted form (52 kD) and intermediate forms (46 kDand 43 kD) of cathepsin D [18–20]. We also confirmedthe identity of the enzymes by Western blot analysisusing specific antibodies. The relative levels of each formof these enzymes detected might be influenced by themetabolic status of the cells. Although cathepsin S hasbeen documented to play an important role in the anti-gen processing and Ii proteolysis, its expression in gastricepithelial cells was not confirmed, because there is nocommercially available antibody yet. The biologic activ-ity of these proteases from gastric epithelial cells was alsoexamined in the studies presented. Interestingly, cathe-
psins were also found to be involved in both processingof H. pylori and dietary antigens as well as in the degra-dation of the class II MHC-associated invariant chain,which is a required step to allow class II MHC moleculesto bind peptides [11–14, 24].
Although the expression of cathepsins B, L, S, and Dis well characterized in cells other than professional APC,their role in antigen processing by mucosal epithelialcells has been ignored [25]. Such “ectopic” expression ofthese enzymes has been associated with apoptotic eventsin other systems [26] or tumor metastasis [27,28]. Inline with the suggested role of these acidic proteases inapoptosis, in addition to degrading H. pylori antigens,cathepsins B and D expressed by gastric epithelial cellsmay play a role in the tissue damage that occurs duringH. pylori infection, which has been reported to include anincrease in apoptosis while in non-infected tissue apo-ptotic cells were infrequent [29]. In support of thispossibility, cathepsin D mucosal levels were higher in H.pylori-infected patients [30]. Although the source of gas-tric mucosal cathepsin D is unclear, it has been assumedto originate from macrophages, because in immunohis-tochemical studies the mucosal epithelium has shownlittle cathepsin D expression and the strongly stainedcells were macrophages [31–33]. By using freshly iso-lated gastric epithelial cells from human biopsies wedetermined that these cells in fact express these impor-tant cathepsins, once again the in vitro findings correlatedwith the in vivo situation.
Because IFN� is a cytokine known to be producedlocally during infection with H. pylori and because IFN�has been reported to enhance the expression of cathepsinsB, S, and D [26–34], we examined whether the expres-sion of these proteases by the gastric epithelium could beinfluenced by IFN�. However, the effect of IFN� onexpression of cathepsins B, L, and D on gastric epithelialcells was marginal (data not shown). It is possible thatexpression of cathepsins B, L, S, and D by human gastricepithelial cells may be influenced by other inflammatorymediators produced during infection with H. pylori.
Although other endosomal or lysosomal proteases arelikely involved in the processing of foreign antigens,little is known about their identity and their overallcontribution. In addition to cathepsins B, L, S, and D,cathepsin E has also been implicated in antigen process-ing for class II MHC-mediated presentation [9, 10].Interestingly, cathepsin E has been found to be expressedby human gastric epithelial cells and is increased in H.pylori-colonized gastric epithelium [35]. These observa-tions, together with those reported here, strongly sup-port a role for gastric epithelial cells in the digestion ofH. pylori antigens. The resulting peptides could then bepresented by class II MHC molecules expressed by thesecells to the CD4� T cells, which are common in the
FIGURE 6 Protein digestion by CB, CS, CL, and CD fromgastric epithelial cells. The digestion capabilities of CB, CS,CL (cysteine proteases), and CD (aspartyl proteases) from gas-tric epithelial cells were confirmed again, and their identitywas approved by selective inhibition studies. The enzymaticactivity of CB, CL, CS, and CD over ovalbumin was blockedusing specific protease inhibitors (E-64 and pepstatin-A). Thedigestion activity of these endosomal proteases was comparablewith those expressed by Jesthom (B cells) and KATO III(gastric epithelial cells) throughout the indicated time period.
1089Antigen Processing Proteases in the Gastric Epithelium

infected tissue. The role of the gastric epithelium in thepresentation of local antigens is further supported bytheir expression of B7-1 and B7-2 costimulatory mole-cules expressed by APC and required for T-cell activation[36].
ACKNOWLEDGMENTS
This work was supported in part by grants DK 50669 and AI34043 from the National Institutes of Health. Gang Ye wassponsored by the McLaughlin Fellowship foundation. CarlosA. Barrera was sponsored by the Clayton Foundation.
REFERENCES
1. Anonymous. Infection with Helicobacter pylori. IARCMonographs on the Evaluation of Carcinogenic Risks toHumans. 61:177, 1994.
2. Brandtzaeg P, Valnes K, Scott H, Rognum TO, Bjerke K,Baklien K: The human gastrointestinal secretory immunesystem in health and disease. In Polak JM, Bloom SR,Wright NA, Butler AG (eds): Basic Science in Gastroen-terology: Diseases of the Gut. Norwich, CT: Page Bros,1986.
3. Engstrand L, Scheynius A, Pathlson C, Grimelius L,
Schwan A, Gustavsson S: Association of Campylobater pyloriwith induced expression of class II transplantation anti-gens on gastric epithelial cells. Infect Immun 57:827,1989.
4. Valnes K, Huitfeldt HS, Brandtzaeg P: Relation betweenT cell number and epithelial HLA class II expressionquantified by image analysis in normal and inflamedhuman gastric mucosa. Gut 31:647, 1990.
5. D’Elios MM, Manghetti M, De Carli M, Costa F, BaldariCT, Burroni D, Telford JL, Romagnani S, Del Prete G: Thelper 1 effector cells specific for Helicobacter pylori inthe gastric antrum of patients with peptic ulcer disease.J Immunol 158:962, 1997.
6. Ernst PB, Song F, Klimpel GR, Haeberle H, BamfordKB, Crowe SE, Ye G, Reyes VE: Regulation of the mu-cosal immune response. Am J Trop Med Hyg 60:2, 1999.
7. Diment S: Different roles for thiol and aspartyl proteasesin antigen presentation of ovalbumin. J Immunol 145:417, 1990.
8. Mizuochi T, Yee ST, Kasai M, Kakiuchi T, Muno D,Kominami E: Both cathepsin B and cathepsin D arenecessary for processing of ovalbumin as well as for deg-radation of class II MHC invariant chain. Immunol Lett43:189, 1994.
9. Solcia E, Paulli M, Silini E, Fiocca R, Finzi G, Kindl S,Boveri E, Bosi F, Cornaggia M, Capella C: Cathepsin E inantigen-presenting Langerhans and interdigitating retic-ulum cells. Its possible role in antigen processing. EurJ Histochem 37:19, 1993.
10. Bennett K, Levine T, Ellis JS, Peanasky RJ, Samloff IM,Kay J, Chain BM: Antigen processing for presentation byclass II major histocompatibility complex requires cleav-age by cathepsin E. Eur J Immunol 22:1519, 1992.
11. Chapman HA: Endosomal proteolysis and MHC class IIfunction. Cur Opin Immunol 10:93, 1998.
12. Chapman HA, Riese RJ, Shi GP: Emerging roles forcysteine proteases in human biology. Ann Rev Physiol59:63, 1997.
13. Reyes VE, Lu S, Humphreys RE: Cathepsin B cleavage ofIi from class II MHC alpha- and beta-chains. J Immunol146:3877, 1991.
14. Roche PA, Cresswell P: Invariant chain association withHLA-DR molecules inhibits immunogenic peptide bind-ing. Nature 345:615, 1990.
15. Bennett K, Levine T, Ellis JS, Peanasky RJ, Samloff IM,Kay J, Chain BM: Antigen processing for presentation byclass II major histocompatibility complex requires cleav-age by cathepsin E. Eur J Immunol 22:1519, 1992.
16. Dytoc M, Gold B, Louie M, Heusca M, Fedorko L, CroweSE, Lingwood C, Brunton J, Sherman PM: Comparison ofHelicobacter pylori and attaching-effacing Escherichia coliadhesion to eukaryotic cells. Infect Immun 61:448, 1992.
17. Moin K, Day NA, Sameni M, Hasnain S, Hirama T,Sloane BF: Human tumour cathepsin B. Comparison withnormal liver cathepsin. Biochem J 285:427, 1992.
FIGURE 7 Evaluation of biologic activity of cysteine pro-teases on invariant chain digestion. KATO III cells werestimulated with IFN-� 48 h prior to a 30-min pulse with [35S]methionine, and chased in indicated times in the presence ofthe protease inhibitor leupeptin (100 mM). The solubilizedmembrane proteins were then immunoprecipitated withmonoclonal antibodies anti-class II MHC (9.3F10 from HB180) and anti-Ii (VicY-1) to see if 21- and 10-kDa fragmentsresulted from the partial degradation of Ii chain. Those frag-ments are labeled to the right of the gel. The accumulation ofthese fragments confirms the biologic activity of the relevantproteases in gastric epithelial cells.
1090 C. A. Barrera et al.

18. Gieselman V, Pohlmann R, Hasilik A, von Figura K:Biosynthesis and transport of cathepsin D in culturedhuman fibroblasts. J Cell Biol 97:1, 1983.
19. Fujita H, Tanaka Y, Noguchi Y, Kono A, Himeno M,Kato K: Isolation and sequencing of a cDNA clone en-coding rat liver lysosomal cathepsin D and the structure ofthree forms of mature enzymes. Biochem Biophys ResComm 179:190, 1991.
20. Williams KP, Smith JA: Isolation of a membrane-associ-ated cathepsin D-like enzyme from the model antigenpresenting cell, A20, and its ability to generate antigenicfragments from a protein antigen in a cell- free system.Arch Biochem Biophys 305:298, 1993.
21. Bando Y, Kominami E, Katunuma N: Purification andproperties of a new cathepsin from rat liver. J Biochem84:659, 1978.
22. Demotz S, Danieli C, Wallny HJ, Majdic O: Inhibition ofpeptide binding to DR molecules by a leupeptin- inducedinvariant chain fragment. Mol Immunol 31:885, 1994.
23. Villadangos JA, Riese RJ, Peters C, Chapman HA,Ploegh HL: Degradation of mouse invariant chain: rolesof cathepsins S and D and the influence of major histo-compatibility complex polymorphism [published erra-tum appears in J Exp Med 1997;186:5945]. J Exp Med186:549, 1997.
24. Roche PA, Cresswell P: Proteolysis of the class II-associ-ated invariant chain generates a peptide binding site inintracellular HLA-DR molecules. Proc Natl Acad SciUSA 88:3150, 1991.
25. Barrera CA, Almanza RJ, Ogra PL, Reyes VE: The role ofthe invariant chain in mucosal immunity. Int Arch Al-lergy Immunol 117:85, 1998.
26. Deiss LP, Galinka H, Berissi H, Cohen O, Kimchi A:Cathepsin D protease mediates programmed cell deathinduced by interferon-gamma, Fas/APO-1 and TNF-al-pha. EMBO J 15:3861, 1996.
27. Leto G, Tumminello FM, Russo A, Pizzolanti G, Bazan V,
Gebbia N: Cathepsin D activity levels in colorectal cancer:correlation with cathepsin B and L and other biologicaland clinical parameters. Int J Oncol 5:509, 1994.
28. Rozhin J, Sameni M, Ziegler G, Sloane BF: PericellularpH affects distribution and secretion of cathepsin B inmalignant cells. Cancer Res 54:6517, 1994.
29. Moss SF, Calam J, Agarwal B, Wang S, Holt PR: Induc-tion of gastric epithelial apoptosis by Helicobacter pylori.Gut 38:498, 1996.
30. Plebani M, Basso D, Rugge M, Vianello F, Di Mario F:Influence of Helicobacter pylori on tryptase and cathepsin Din peptic ulcer. Dig Dis Sci 40:2473, 1995.
31. Orlowski M, Orlowski R, Chang JC, Wilk E, Lesser M: Asensitive procedure for determination of cathepsin D: ac-tivity in alveolar and peritoneal macrophages. Mol CellBiochem 64:155, 1984.
32. Chambon M, Rebillard X, Rochefort H, Brouillet JP,Baldet P, Guiter J, Maudelonde T: Cathepsin D cytosolicassay and immunohistochemical quantification in prostatetumors. Prostate. 24:320, 1994.
33. Watanabe M, Higashi T, Hashimoto M, Tomoda I, To-minaga S, Hashimoto N, Morimoto S, Yamauchi Y, Na-katsukasa H, Kobayashi M: Elevation of tissue cathepsin Band L activities in gastric cancer. Hepatogastroenterology34:120, 1987.
34. Nadler SG, Rankin BM, Morandavis P, Cleaveland JS,Kiener PA: Effect of interferon-gamma on antigen pro-cessing in human monocytes. Eur J Immunol 24:3124,1994.
35. Fiocca R, Luinetti O, Villani L, Chiaravalli AM, CapellaC, Solcia E: Epithelial cytotoxicity, immune responses,and inflammatory components of Helicobacter pylori gastri-tis. Scand J Gastroenterol 29:11, 1994.
36. Ye G, Barrera C, Fan XJ, Gourley WK, Crowe SE, ErnstPB, Reyes VE: Expression of B7-1 and B7-2 costimulatorymolecules by human gastric epithelial cells — potentialrole in CD4 (�) T cell activation during Helicobacter pyloriinfection. J Clin Invest 99:1628, 1997.
1091Antigen Processing Proteases in the Gastric Epithelium
Copyright © 2022 FDOKUMEN




















