
Alpha-Helical Cationic Anticancer Peptides: A Promising Candidate for Novel Anticancer Drugs
Upload
independentCategory
view
0download
0
# 2007 The Authors
Journal compilation# 2007 Blackwell Publishing Ltd
doi: 10.1111/j.1600-0854.2007.00572.xTraffic 2007; 8: 848–866Blackwell Munksgaard
A Comprehensive Model for the Cellular Uptake ofCationic Cell-penetrating Peptides
Falk Duchardt1,†, Mariola Fotin-Mleczek1,†,
Heinz Schwarz2, Rainer Fischer1 and
Roland Brock1,3,*
1Interfaculty Institute for Cell Biology, University ofTubingen, Auf der Morgenstelle 15, 72076 Tubingen,Germany2Max-Planck-Institute for Developmental Biology,Spemannstr. 35, 72076 Tubingen, Germany3Department of Biochemistry, Nijmegen Centre forMolecular Life Sciences, Radboud University NijmegenMedical Centre, PO Box 9101, 6500 HB Nijmegen,The Netherlands*Corresponding author: Roland Brock,[email protected]†These authors contributed equally to this work.
The plasma membrane represents an impermeable
barrier for most macromolecules. Still some proteins and
so-called cell-penetrating peptides enter cells efficiently.
It has been shown that endocytosis contributes to the
import of these molecules. However, conflicting results
have been obtained concerning the nature of the endo-
cytic process. In addition, there have been new findings
for an endocytosis-independent cellular entry. In this
study, we provide evidence that the Antennapedia-
homeodomain-derived antennapedia (Antp) peptide,
nona-arginine and the HIV-1 Tat-protein-derived Tat pep-
tide simultaneously use three endocytic pathways: mac-
ropinocytosis, clathrin-mediated endocytosis and
caveolae/lipid-raft-mediated endocytosis. Antennapedia
differs from Tat and R9 by the extent by which the
different import mechanisms contribute to uptake. More-
over, at higher concentrations, uptake occurs by a mech-
anism that originates from spatially restricted sites of the
plasma membrane and leads to a rapid cytoplasmic
distribution of the peptides. Endocytic vesicles could
not be detected, suggesting an endocytosis-independent
mode of uptake. Heparinase treatment of cells negatively
affects this import, as does the protein kinase C inhibitor
rottlerin, expression of dominant-negative dynamin and
chlorpromazine. Thismechanism of uptakewas observed
for a panel of different cell lines. For Antp, significantly
higher peptide concentrations and inhibition of endocy-
tosis were required to induce its uptake. The relevance of
these findings for import of biologically active cargos is
shown.
Key words: antennapedia homeodomain, cell-penetrating
peptide, endocytosis, oligo-arginine, TAT
Received 1 February 2006, revised and accepted for publi-
cation 26 March 2007, uncorrected manuscript published
online 29 March 2007, published online 10 May 2007
For most polar molecules, the plasma membrane repre-
sents an impermeable barrier. It is therefore highly remark-
able that some proteins and peptides possess the ability to
cross this border and reach the cytoplasm. Among these
are transcription factors belonging to the homeodomain
family, such as theDrosophila melanogaster Antennapedia
homeodomain protein (1), the HIV-1 Tat protein (2) and
fibroblast growth factors 1 and 2 (3). Peptides that possess
this ability were identified as the protein transduction
domains (PTDs) of aforementioned proteins such as the
Antennapedia-homeodomain-derived antennapedia (Antp)
peptide (4) and the HIV-1 Tat-derived Tat peptide, denoted
here as Tat (5). Alternatively, peptides, among them oligo-
arginine peptides (6), were designed de novo based on
structure activity relationships of PTDs. The attractivity of
these peptides, generally defined as cell-penetrating pepti-
des (CPPs), in biomedical research is a consequence of their
ability to mediate the import of membrane-impermeable,
biologically active molecules such as small interfering
RNA, DNA, peptides or entire proteins into the cells ex
vivo and in whole organisms [for reviews, see Dietz and
Bahr (7) and Snyder and Dowdy (8)].
Initially, the import of these cell-penetrating molecules,
proteins and peptides alike, was considered to occur by
direct permeation of the plasma membrane (9). This model
was based on evidence obtained from cell biological as
well as biochemical and biophysical experiments. More
recently, it was shown that endocytosis plays a major role
in the import of the basic and amphiphilic Antp peptide, the
highly basic and arginine-rich R9 and Tat peptides and in
the import of the HIV Tat protein itself (10–12). No specific
receptor has been implicated in the uptake of these
molecules. Instead, in some cases, the initial association
with the plasma membrane was attributed to multivalent
interactions with cell surface heparan sulfate proteo-
glycans (13,14).
To this point, however, conflicting results were obtained
for the involvement of specific endocytic pathways. Data
were presented that supported a role of macropinocytosis
(15), clathrin-mediated endocytosis (CME) (11) and caveo-
lae/lipid-raft-mediated endocytosis (16,17).
Contrary to this uptake through endocytosis, for a fluores-
cein-labeled Tat peptide, Ziegler et al. observed a rapid
cellular import into fibroblasts that was heparan sulfate
dependent (18). Similarly, Tunnemann et al. reported that
the Tat peptide conjugated to a small peptide cargo enters
cells by a rapid and endocytosis-independent process, with
resemblance to the one described by Ziegler et al., while
848 www.traffic.dk

larger conjugates enter cells by endocytosis (19). They
concluded that endocytosis is restricted to high molecular
weight complexes. However, this conclusion is in conflict
with a number of reports observing a vesicular mode of
uptake for the Tat peptide as well (11,20–22).
As an explanation for these apparent discrepancies, in this
article, we provide evidence that the peptides use the
three endocytic pathways simultaneously. Antennapedia
differs from R9 and Tat by the extent that individual
processes contribute to import. Moreover, for R9 and Tat
above a concentration threshold in the lower micromolar
range, both peptides are internalized predominantly
through a process that leads to a rapid distribution of
peptides into the cytoplasm and nucleus. Our observations
suggest that this mechanism is endocytosis independent.
Blockage of macropinocytosis and lipid-raft-dependent
endocytosis lowers this threshold. This rapid cytoplasmic
entry originates from spatially confined zones of the
plasma membrane. For Antp, this uptake was observed
only for cells treated with the inhibitor of lipid-raft-depen-
dent endocytosis methyl-b-cyclodextrin (MbCD) at signifi-
cantly higher peptide concentrations. The analysis of the
mechanistic basis showed that this uptake is heparan sul-
fate dependent and sensitive to the protein kinase C (PKC)
inhibitor rottlerin, the expression of dominant-negative
(DN) dynamin and chlorpromazine (CPZ). The relevance
of these results for the application of CPPs as tools in cell
biological research is shown using a Smac-derived peptide
(23) that potentiates the Fas-dependent induction of
apoptosis.
Results
The intracellular distribution of R9 and Tat is
concentration dependent
For the three cationic CPPs Antp, R9 and Tat (Table 1),
there is a consensus that endocytosis at least strongly
contributes to cellular uptake (24). However, conflicting
evidence has been presented concerning the role of the
individual endocytic pathways. Moreover, some research-
ers observed a rapid entry of peptides into the cytosol for
which endocytosis did not seem to be involved. We
reasoned that the apparent discrepancies were a conse-
quence of the different experimental conditions with
respect to cell lines, incubation times and peptide concen-
trations. For this reason, we performed experiments in the
presence of different peptide concentrations. Experiments
investigating the uptake mechanism of CPP have mostly
used these peptides at lower micromolar concentrations,
while the applications of CPP–peptide conjugates for
interfering with molecular interactions inside the cell have
frequently used these molecules in the mean to upper
micromolar range (25–28). The selected concentration
range for our experiments therefore covered peptide
concentrations used in both types of experiments. As
a cellular model system, we selected HeLa cells because
this cell line has been used in a significant number of
previous reports (11,13,17,20,21,29). All three peptides
were synthesized as fluorescein-labeled analogues. The
cellular uptake was quantified in living cells by flow
cytometry. Trypsinization of cells, required to detach the
cells from the tissue culture plate prior to flow cytometry,
removed peptides merely adsorbed to the outer plasma
membrane (11). In addition, the intracellular peptide distri-
bution was investigated by live cell confocal laser scanning
microscopy.
For Antp, the cellular fluorescence was proportional to the
concentration of peptide in the medium (Figures 1A, 2A).
With respect to the intracellular fluorescence, the cell
population was fully homogeneous for each peptide con-
centration. Neither saturation nor changes in the intracel-
lular distribution of peptide was observed. Antennapedia
was localized predominantly within vesicular structures.
Fluorescence could also be detected in the cytoplasm, and
this fluorescence also increased with peptide concentra-
tion. In contrast, the cellular uptake of R9 and Tat did not
increase linearly with peptide concentration (Figures 1B,C,
2A). Both flow cytometry and fluorescence microscopy
clearly showed that at concentrations higher than 10 mM,
cytoplasmic delivery of peptides was strongly enhanced.
Simultaneously, the cell population became very hetero-
geneous with respect to peptide uptake. Some cells were
fluorescent to the point that they were outside the
detection range. Other cells showed a moderate uptake,
and finally, some cells exhibited only a weak intracellular
fluorescence. This heterogeneity in the level of the cell
population was reflected by a heterogeneity in the distri-
bution of fluorescence on the subcellular level. With in-
creasing peptide concentration, an increasing number of
cells exhibited a clear enrichment of peptide in the nucleus
and cytoplasm, with little vesicular fluorescence, resem-
bling the distribution reported by Tunnemann et al. (19).
To exclude that the highly efficient peptide import was
a result of damage of the plasma membrane, we tested
the membrane integrity using a trypan blue exclusion test
(data not shown) and incubation with peptide in the
presence of propidium iodide (PI) (Figure 1D). Cells
Table 1: Primary structures of the peptides used in this studya
Entry Peptide Sequence
1 Antp Fluo-RQIKIWFQNRRMKWKK-CONH2
2 R9 Fluo-RRRRRRRRR-CONH2
3 Tat Fluo-YGRKKRRQRRR-CONH2
4 Smac–Antp AVPIAQK-RQIKIWFQNRRMKWKK-
eK(Fluo)-CONH2
5 Smac–R9 AVPIAQK-RRRRRRRRR-eK(Fluo)-CONH2
aAll peptides were synthesized as C-terminal peptide amides
(-CONH2). Fluo represents 5(6)-carboxyfluorescein.
Traffic 2007; 8: 848–866 849
Uptake of Cationic Cell-Penetrating Peptides
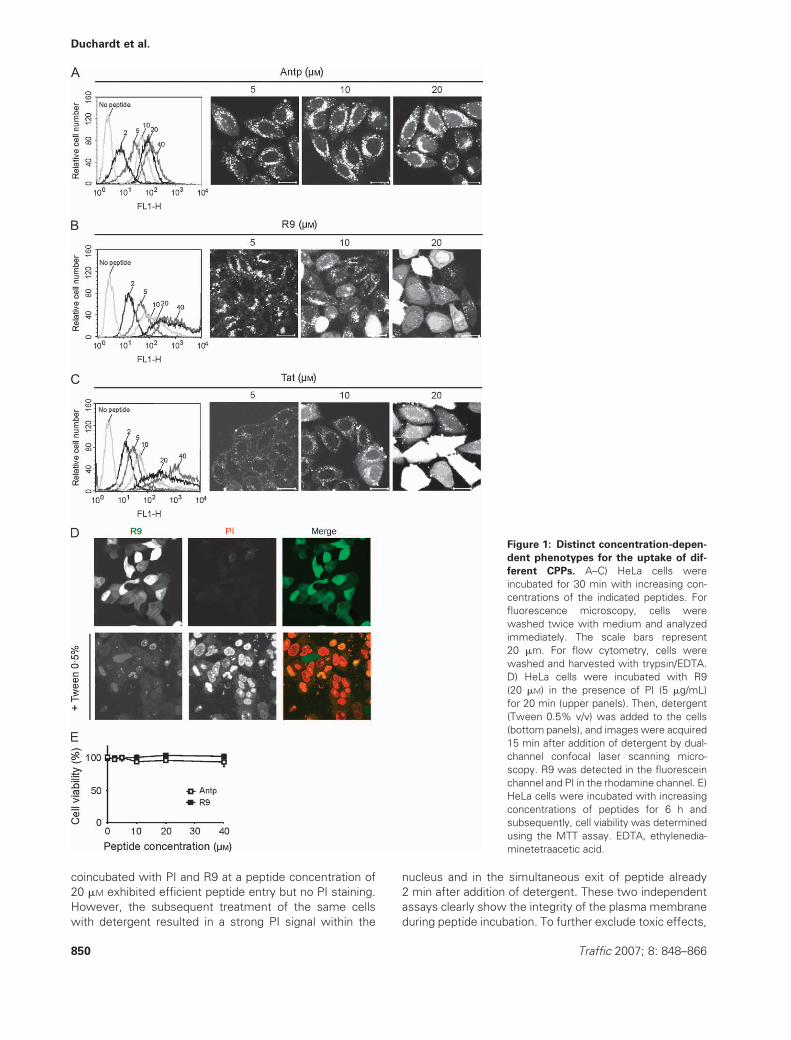
coincubated with PI and R9 at a peptide concentration of
20 mM exhibited efficient peptide entry but no PI staining.
However, the subsequent treatment of the same cells
with detergent resulted in a strong PI signal within the
nucleus and in the simultaneous exit of peptide already
2 min after addition of detergent. These two independent
assays clearly show the integrity of the plasma membrane
during peptide incubation. To further exclude toxic effects,
Figure 1: Distinct concentration-depen-
dent phenotypes for the uptake of dif-
ferent CPPs. A–C) HeLa cells were
incubated for 30 min with increasing con-
centrations of the indicated peptides. For
fluorescence microscopy, cells were
washed twice with medium and analyzed
immediately. The scale bars represent
20 mm. For flow cytometry, cells were
washed and harvested with trypsin/EDTA.
D) HeLa cells were incubated with R9
(20 mM) in the presence of PI (5 mg/mL)
for 20 min (upper panels). Then, detergent
(Tween 0.5% v/v) was added to the cells
(bottom panels), and images were acquired
15 min after addition of detergent by dual-
channel confocal laser scanning micro-
scopy. R9 was detected in the fluorescein
channel and PI in the rhodamine channel. E)
HeLa cells were incubated with increasing
concentrations of peptides for 6 h and
subsequently, cell viability was determined
using the MTT assay. EDTA, ethylenedia-
minetetraacetic acid.
850 Traffic 2007; 8: 848–866
Duchardt et al.

the viability of cells was also determined after incubation
with peptides for 6 h. Using the 3-(4,5-dimethylthiazol-2-yl)-
2,5-diphenyltetrazolium bromide (MTT) test, no toxicity
was observed for peptide concentrations up to 40 mM
(Figure 1E).
R9 and Tat differ from Antp in the concentration
dependence of the uptake mechanism
The surprising observation that for R9 and Tat, different
peptide concentrations led to clearly distinct phenotypes
with respect to intracellular peptide distribution motivated
us to ask whether different import mechanisms may be
responsible for this effect. To test this hypothesis, we
investigated the involvement of the three major endocytic
pathways in peptide uptake in dependence on peptide
concentration. Pharmacological inhibitors were used to
interfere with individual endocytic pathways (30–32).
Chlorpromazine was used for the inhibition of clathrin-
mediated internalization, 5-(N-ethyl-N-isopropyl)amiloride
(EIPA) for inhibition of macropinocytosis and MbCD for
disruption of import through caveolae/lipid rafts. The
specificity of the inhibitors is an issue of debate, also
within the field of CPP uptake (33). For fluorescently
labeled tracer molecules [transferrin, dextran and cholera
toxin beta subunit (CTB)], each of the inhibitors showed
the expected effect (Figure S1). However, for none of the
tracer molecules, import could be fully blocked. All three
inhibitors alone or in combination, applied at concentra-
tions used in this study, had no toxic effect (Figure S2). The
effects of all three inhibitors on the delivery of Antp, R9
and Tat within a concentration range of 2–40 mM were
tested by flow cytometry (Figure 2). The concentration-
dependent peptide import measured in cells not treated
with inhibitor was used as a 100% reference for deter-
mining the reduction in uptake caused by the different
inhibitors (Figure 2A). For Antp, CPZ and EIPA reduced the
cellular fluorescence by about 20% for the entire concen-
tration range. Preincubation of cells with MbCD had no
detectable effect. In contrast, for R9 and Tat, the effect
exerted by each inhibitor was strongly dependent on
peptide concentration. At peptide concentrations <5 mM,
CPZ was without effect. However, with increasing peptide
concentration, the inhibitory activity of CPZ increased,
amounting to about 80% at a peptide concentration of
40 mM. At low peptide concentrations, both MbCD and
EIPA inhibited the import of R9 and Tat only slightly if at
all. Interestingly, at higher peptide concentrations, the
same compounds did not reduce but dramatically
enhanced the uptake of these CPPs. We observed a four-
to sevenfold increase of cellular fluorescence compared
with the fluorescence in the control, not treated with
inhibitors. At a concentration of 40 mM, the fluorescence
was outside the detection range. These results again
confirm a difference between the cellular import of Antp
and one of the other two peptides. Moreover, the
dependence of the inhibitor effects on peptide concen-
tration strongly suggests that the contribution of the
individual uptake mechanisms may in fact depend on
peptide concentration.
To test this hypothesis in more detail, the cellular peptide
distribution was analyzed at low (2 mM) and high (20 mM)
peptide concentrations. For Antp, all three inhibitors led
to distinct phenotypes with respect to cellular peptide
Figure 2: Dependence of the
effect of inhibitors of endocytosis
on peptide concentration. A) Con-
centration dependence of the cellu-
lar uptake of Antp, R9 and Tat. B–D)
Cellular peptide uptake in the pres-
ence of 10 mg/mL CPZ (B), 5 mM
MbCD (C) and 50 mM EIPA (D).
HeLa cells were incubated for 30
min at 378C with the indicated
inhibitor of endocytosis or remained
untreated (A). Then, the cells were
incubated with increasing concen-
trations of CPPs for further 30 min
in the presence or absence of inhibi-
tors, washed, harvested with tryp-
sin/EDTA and analyzed by flow
cytometry. The effect of the inhibi-
tors on cellular fluorescence is
shown relative to the fluorescence
measured in the inhibitor-free
controls incubated with the same
concentration of peptide. EDTA,
ethylenediaminetetraacetic acid.
Traffic 2007; 8: 848–866 851
Uptake of Cationic Cell-Penetrating Peptides
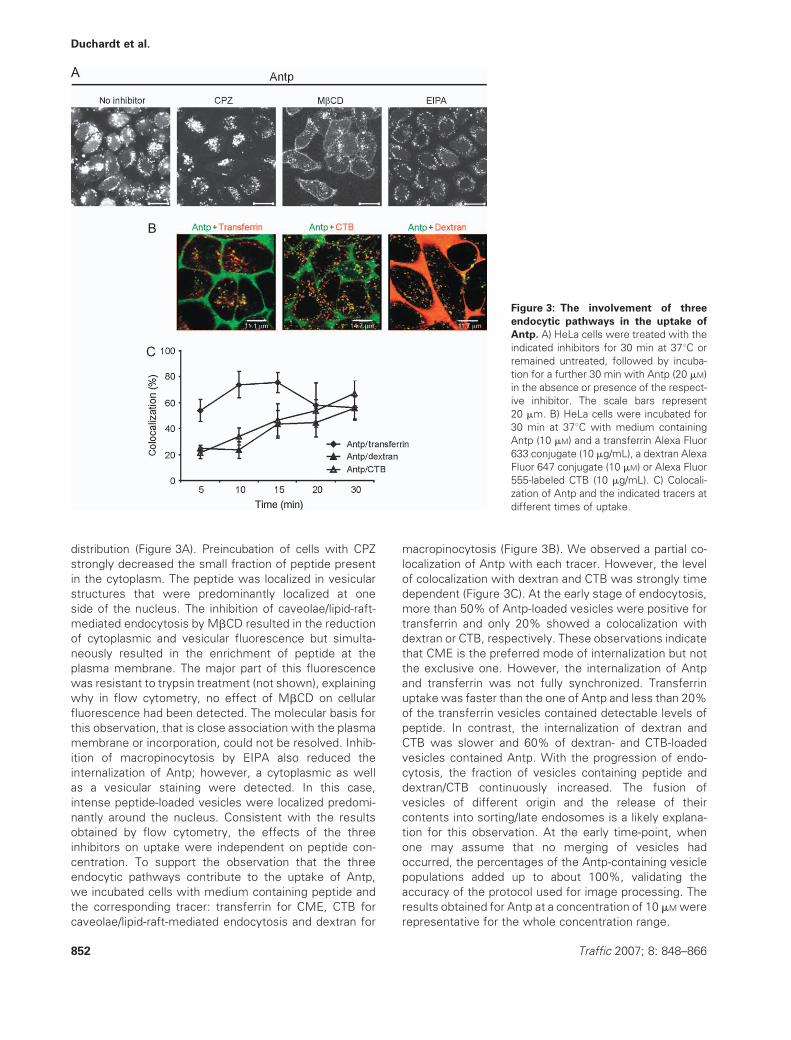
distribution (Figure 3A). Preincubation of cells with CPZ
strongly decreased the small fraction of peptide present
in the cytoplasm. The peptide was localized in vesicular
structures that were predominantly localized at one
side of the nucleus. The inhibition of caveolae/lipid-raft-
mediated endocytosis by MbCD resulted in the reduction
of cytoplasmic and vesicular fluorescence but simulta-
neously resulted in the enrichment of peptide at the
plasma membrane. The major part of this fluorescence
was resistant to trypsin treatment (not shown), explaining
why in flow cytometry, no effect of MbCD on cellular
fluorescence had been detected. The molecular basis for
this observation, that is close association with the plasma
membrane or incorporation, could not be resolved. Inhib-
ition of macropinocytosis by EIPA also reduced the
internalization of Antp; however, a cytoplasmic as well
as a vesicular staining were detected. In this case,
intense peptide-loaded vesicles were localized predomi-
nantly around the nucleus. Consistent with the results
obtained by flow cytometry, the effects of the three
inhibitors on uptake were independent on peptide con-
centration. To support the observation that the three
endocytic pathways contribute to the uptake of Antp,
we incubated cells with medium containing peptide and
the corresponding tracer: transferrin for CME, CTB for
caveolae/lipid-raft-mediated endocytosis and dextran for
macropinocytosis (Figure 3B). We observed a partial co-
localization of Antp with each tracer. However, the level
of colocalization with dextran and CTB was strongly time
dependent (Figure 3C). At the early stage of endocytosis,
more than 50% of Antp-loaded vesicles were positive for
transferrin and only 20% showed a colocalization with
dextran or CTB, respectively. These observations indicate
that CME is the preferred mode of internalization but not
the exclusive one. However, the internalization of Antp
and transferrin was not fully synchronized. Transferrin
uptake was faster than the one of Antp and less than 20%
of the transferrin vesicles contained detectable levels of
peptide. In contrast, the internalization of dextran and
CTB was slower and 60% of dextran- and CTB-loaded
vesicles contained Antp. With the progression of endo-
cytosis, the fraction of vesicles containing peptide and
dextran/CTB continuously increased. The fusion of
vesicles of different origin and the release of their
contents into sorting/late endosomes is a likely explana-
tion for this observation. At the early time-point, when
one may assume that no merging of vesicles had
occurred, the percentages of the Antp-containing vesicle
populations added up to about 100%, validating the
accuracy of the protocol used for image processing. The
results obtained for Antp at a concentration of 10 mMwere
representative for the whole concentration range.
Figure 3: The involvement of three
endocytic pathways in the uptake of
Antp. A) HeLa cells were treated with the
indicated inhibitors for 30 min at 378C or
remained untreated, followed by incuba-
tion for a further 30 min with Antp (20 mM)
in the absence or presence of the respect-
ive inhibitor. The scale bars represent
20 mm. B) HeLa cells were incubated for
30 min at 378C with medium containing
Antp (10 mM) and a transferrin Alexa Fluor
633 conjugate (10 mg/mL), a dextran Alexa
Fluor 647 conjugate (10 mM) or Alexa Fluor
555-labeled CTB (10 mg/mL). C) Colocali-
zation of Antp and the indicated tracers at
different times of uptake.
852 Traffic 2007; 8: 848–866
Duchardt et al.

For the uptake of R9 and Tat at low peptide concentrations,
CPZ had no effect (Figure 4). In contrast, at high peptide
concentrations, the same inhibitor completely abolished
the cytoplasmic and nuclear localization of the peptides.
Under the assumption that CPZ is a specific inhibitor of
clathrin-dependent endocytosis, this observation provided
strong evidence that at high concentrations, this endocytic
pathway is a key internalization mechanism used by these
peptides, which leads to their cytoplasmic and nuclear
enrichment. The inhibition of transferrin uptake confirmed
that inhibition of CME is at least one activity exerted by
CPZ on HeLa cells. However, CPZ originally was identified
as an antipsychotic drug (34), acting as an antagonist of
dopamine D2 receptors (35). Moreover, CPZ directly
interacts with calmodulin, thereby also interfering with
a number of Ca2þ-dependent signaling pathways (36).
Given the fact that our further experiments (see below)
supported a nonendocytic uptake mechanism, in the
following we refer to this uptake as CPZ sensitive rather
than taking these results as direct evidence for clathrin-
dependent endocytosis.
At low concentrations, the CPZ-sensitive pathway did not
seem to be involved in the uptake of R9 and Tat. MbCD
and EIPA exerted a slight inhibitory effect, leading to
phenotypes similar to the ones observed for Antp. How-
ever, at high peptide concentrations, the same inhibitors
strongly promoted the cytoplasmic and nuclear distribution
of peptides (Figure 4). This result was surprising but
completely consistent with the data obtained by flow
cytometry (Figure 2C,D). Evidently, at high peptide con-
centrations, the inhibition of endocytosis exerted by these
inhibitors directed the peptides toward a more efficient
compensatory mechanism.
Having observed that for R9 and Tat, inhibitors of endocy-
tosis lower the threshold for the rapid entry of peptide into
the cytoplasm, we decided to investigate the uptake of
Figure 4: Dependence of the effects of in-
hibitors of endocytosis on the concentra-
tion of R9 and Tat. HeLa cells were treated
with the indicated inhibitors for 30 min at 378Cor remained untreated. Then, the cells were
incubated for a further 30 min with either 2 or
20 mM of R9 A) or Tat peptide B) in the
presence or absence of the respective inhibi-
tor. The scale bars represent 20 mm. C)
MbCD-induced cytoplasmic uptake of Antp in
HeLa cells. HeLa cells were treated with
MbCD for 30 min at 378C or remained
untreated. Then, the cells were incubated for
a further 30 min with 100 mM of Antp in the
presence or absence of MbCD. The scale bars
represent 20 mm.
Traffic 2007; 8: 848–866 853
Uptake of Cationic Cell-Penetrating Peptides

Antp at even higher concentrations. Antennapedia was
applied to HeLa cells at a concentration of 100 mM. In the
absence of inhibitors, in spite of a further increase of
cytoplasmic fluorescence, vesicular staining was still
prominent (Figure 4C). In contrast, in the presence of
MbCD, some cells showed a homogeneous cytoplasmic
and nuclear fluorescence. However, after about 20-min
incubation, morphological changes of the cells were indica-
tive of cell damage. In contrast to R9 and Tat, EIPA was
without effect. Nevertheless, these data indicate that the
differences between Antp and R9 and Tat may be rather
quantitative than qualitative.
To provide direct evidence that application of EIPA and
MbCD promoted the uptake of R9 and Tat through the
CPZ-sensitive rapid uptake mechanism, HeLa cells were
incubated either with MbCD or with EIPA alone or in
combination with CPZ (Figure 5A). R9 was applied at
a concentration of 10 mM. In control cells, at this concen-
tration, the peptide was predominantly localized within
vesicles. Consistent with our hypothesis, at this peptide
concentration, CPZ had no effect on peptide uptake. As
before, preincubation of cells with MbCD or EIPA led to
a pronounced localization of peptides in the cytoplasm
and nucleus. However, this phenotype was completely
Figure 5: MbCD- and EIPA-mediated induction of a rapid cytoplasmic internalization mechanism. A) HeLa cells were treated with
the indicated inhibitors for 30 min at 378C and then incubated with 5 mM of R9 in the presence of the respective inhibitors for a further
30 min. The scale bars represent 20 mm. B) HeLa cells were treated with the indicated inhibitors as described in (A), followed by incubation
with increasing concentrations of R9 or Tat and then harvested with trypsin/EDTA, and the intracellular fluorescence was quantified with
flow cytometry. The error bars represent the mean deviation of three independent experiments. EDTA, ethylenediaminetetraacetic acid.
854 Traffic 2007; 8: 848–866
Duchardt et al.

reversed when cells were treated with either MbCD or
EIPA in combination with CPZ. The primary inhibitory
effects of both MbCD (membrane enrichment) and EIPA
(vesicles distributed around the nucleus) were detected
again. The MbCD-induced promotion of peptide import
and the reversal of this effect by CPZ were confirmed by
flow cytometry (Figure 5B). For R9 and Tat, applied at
concentrations higher than 5 mM, the inhibition of caveolae/
lipid-raft-mediated endocytosis by MbCD enhanced pep-
tide uptake by up to 800 (R9) and 400% (Tat) of controls
not treated with inhibitor. This increase was almost
completely inhibited when cells were treated with a com-
bination of MbCD and CPZ.
The uptake of R9 at high peptide concentrations
originates from spatially restricted membrane regions
Having shown the remarkable efficiency of the CPZ-
sensitive uptake mechanism used by R9 and Tat at high
peptide concentrations, we aimed at obtaining information
about the time–course of this uptake mechanism. Cells
were incubated with R9 (20 mM), and the internalization of
peptide was imaged by time-lapse confocal fluorescence
microscopy, with the peptide still in the medium (Figure
6A). Image acquisition was started about 60 seconds
after addition of peptide. Frames were recorded every
30 seconds.
In some cells, areas of higher fluorescence intensity were
observed that seemed to emerge from spatially restricted
membrane regions. During the next 10 min, further highly
fluorescent zones (one or two per cell) developed that
were the origin of a rapid spreading of fluorescence
throughout the cytoplasm and nucleus. We chose to call
these highly efficient internalization platforms nucleation
zones (NZs). Almost immediately after formation of an NZ,
the peptide was enriched in the nucleus. In agreement
with our previous observation, the cell population became
heterogeneous with regards to the intracellular fluores-
cence. The efficient peptide entry strictly depended on the
formation of the NZs. Cells lacking these structures did not
Figure 6: The rapid uptake of R9 originates from spatially confined internalization platforms. A) Medium containing R9 at
a concentration of 20 mM was added to the cells and uptake of fluorescence followed by time-lapse confocal microscopy at room
temperature with frames recorded every 30 seconds. Four frames recorded at 1, 5, 10 and 15min are shown. B) Time-lapse images for the
fluorescence intensity graph (C) marked with a star. The scale bar corresponds to 20 mm. C) Increase of fluorescence intensities over time
determined for the time series shown in (B) from circular ROI placed into individual cells. To render the comparison of uptake kinetics for
different days insensitive to differences in laser power and fluorescence detection efficiency, rather than in absolute values, the values are
expressed in fold increase in comparison with the intracellular signal after addition of peptide. For the graph marked with a star, the points
corresponding to the individual frames are represented by white diamonds. NZs are marked with arrows. D) NZ-mediated uptake
accompanied by bleb formation. The individual time-points are given in the images. The scale bar corresponds to 10 mm.
Traffic 2007; 8: 848–866 855
Uptake of Cationic Cell-Penetrating Peptides

exhibit any cytoplasmic or nuclear fluorescence. At micro-
scope settings at which usually endocytic uptake was
followed, for many cells, fluorescence went into satura-
tion. Therefore, further time-lapse recordings were per-
formed in which saturation was carefully avoided
(Figure 6B; Video S1). The uptake kinetics was analyzed
by determining the fluorescence over time for circular
regions of interest (ROI). For nearly all cells showing the
rapid import, the intracellular fluorescence reached a pla-
teau (Figure 6C). However, individual cells strongly varied
in the level of fluorescence at which this plateau was
reached. In addition, there was no correlation between the
uptake kinetics and the increase in fluorescence. In one
case, a strong entry of peptide occurred within 4 min, in
another case, it was within 10 min. A similar variation in
time was observed for cells with a much weaker uptake.
Very remarkably in some cases, import was not continu-
ous. Instead, the increase in cellular fluorescence occurred
in sequential steps, each having similar kinetics and height.
Such a discontinuous stepwise import would be highly
unusual for endocytosis.
A correlation of this increase in cellular fluorescence with
the image sequences showed that for at least some cells,
each step was accompanied with the formation of an NZ
and the dissipation of fluorescence from this NZ into the
cytoplasm (Figure 6B,C). If no NZ was visible during
uptake, this was probably a result of the confocal nature
of the image acquisition that restricted fluorescence
detection to one section through the cell.
In addition, for some cells, after local enrichment of
fluorescence, a bleb-like membrane protrusion was
formed that culminated in the pinching-off of a fluorescent
vesicular structure. An example for a large bleb is shown in
Figure 6D. Concomitant with bleb formation, the rapid
uptake of fluorescence and distribution of fluorescence
into the cytoplasm and nucleus occurred. In some cases,
a fluorescent vesicle was suddenly visible next to an NZ in
only one image frame. It is therefore very likely that in
many of the cases in which no bleb was detected, this was
because of the timing of the image acquisition and/or the
confocality of the detection volume.
The compensatory uptake of R9 in the presence of MbCD
exhibited the same characteristics (not shown). To exclude
(i) that the formation of NZ and rapid cellular uptake were
because of the presence of fluorophore and (ii) that the
cytoplasmic fluorescence was a result of a concentration-
dependent release of proteolytic breakdown products into
the cytoplasm, unlabeled R9 (15 mM) was mixed with
fluorescein-labeled R9 (5 mM) peptide. The formation of
NZs, the import kinetics and the cellular distribution fully
corresponded to the one observed for the fluorescein-
labeled analogue alone (not shown). In addition, the same
formation of NZ, uptake kinetics and distribution were
observed for a tetramethylrhodamine-labeled Tat peptide,
further excluding a role of the fluorophore in our observa-
tions. For MbCD-treated HeLa cells, on incubation with
50 mM Antp, NZ were also observed.
In the NZ, peptides are transiently confined
Fluorescence microscopy had failed to resolve distinct
vesicles at the NZs. As an alternative means to obtain
information about a possible vesicular confinement of the
peptide within the NZ, we compared the mobility of the
peptide within and outside the NZs using fluorescence loss
in photobleaching [FLIP (37)]. The peptide-associated
fluorescein fluorescence was bleached in ROI either inside
or outside the NZ (Figure 7). Bleaching inside the NZ
resulted in the complete destruction of fluorescence not
only in the bleached region but also in the whole cell. In
contrast, bleaching outside the NZ depleted all fluorescein
fluorescence, except the NZ-associated signal, providing
strong evidence for a confinement or association of
peptide within these structures. These FLIP experiments
had been conducted with peptide present in the medium.
To decide whether the NZ-associated fluorescence after
bleaching of fluorescence outside the NZ represented
peptide that was permanently associatedwith a subcellular
structure or only transiently confined peptide, cells were
washed prior to FLIP inside the nucleus. In this case, FLIP
resulted in the depletion of all fluorescence (not shown).
This result confirms that fluorescence inside the NZs
represents a highly dynamic trafficking state of the pep-
tides. This observation also explains why in previous
experiments, in which cells were routinely washed prior
to microscopy, it had been impossible to detect the NZ.
The formation of NZs is CPZ sensitive and dynamin
and heparan sulfate dependent
Having shown that cytoplasmic and nuclear peptide deliv-
ery of R9 and Tat could be inhibited by CPZ, we asked
whether this inhibitor directly affected the development of
NZ. Pretreatment of cells with CPZ abolished the develop-
ment of NZ and the efficient internalization of peptide
(Figure 8A). The rapid import kinetics of the NZ-mediated
Figure 7: In the NZs, peptides are transiently confined. FLIP
inside A) and outside B) an NZ. Fluorescein-labeled peptides were
bleachedwithin the indicated areas (ROIs) for 60 seconds with the
488-nm line of an argon ion laser.
856 Traffic 2007; 8: 848–866
Duchardt et al.

uptake and the failure to detect vesicles questioned
whether the interference of CPZ with this import pathway
was in fact a result of the inhibition of clathrin-dependent
endocytosis. To address this point by an independent
experimental approach, HeLa cells were used in which
endocytosis by clathrin-coated pits and caveolae/lipid rafts
could be disrupted through the inducible expression of
a DN mutant dynamin-2. Dynamin is a guanosine triphos-
phatase, which is critically involved in the scission of
vesicles in these endocytic pathways (38,39). As ex-
pected, the expression of DN dynamin inhibited the
internalization of transferrin, a classical marker for CME
(Figure 8B, lower panels), leading to the enrichment of
receptor-bound transferrin at the plasma membrane.
Figure 8: Uptake through NZ is sen-
sitive to CPZ and is dynamin and
heparan sulfate dependent. A) Inhibi-
tion of NZ formation by CPZ. Cells were
incubated with the compound 30 min
before addition of 20 mM peptide. B)
HeLa dynK44A cells were incubated for
24 h without or with tetracycline (1 mg/
mL). Then, the cells were incubated
with 20 mM R9 or with a transferrin
Alexa Fluor 633 conjugate for 30 min
at 378C. White arrows indicate NZs. C)
Cells were treated for 30 min with
either ammonium chloride (50 mM) or
chloroquine (100 mM) or remained
untreated. The images show the uptake
of R9 after 30 min. Scale bars represent
20 mm. D) HeLa cells were treated for
6 h with heparinases I, II and III (10, 5,
and 2 U/mL, respectively) or remained
untreated. Then, the cells were washed
and incubated with peptide-containing
medium. Images were taken 20 min
after addition of peptide. Removal of
HSPG from the cell surface was con-
firmed by immunofluorescent staining
of treated and untreated cells. Scale
bars correspond to 20 mm.
Traffic 2007; 8: 848–866 857
Uptake of Cationic Cell-Penetrating Peptides

In addition, the induction of DN dynamin abolished the
development of NZs and consequently the delivery of
peptide into the cytoplasm (Figure 8B, upper panels).
For the full-length Tat protein and CPPs, it had been shown
that acidification of the endosomal content is required for
the release of CPPs into the cytoplasm (12,20,21). We
therefore probed for a role of acidification in the NZ-
mediated rapid peptide import into the cytoplasm. Contra-
dictory results were obtained for the two well-established
inhibitors of endosomal acidification, ammonium chloride
and chloroquine (Figure 8C). Treatment of cells with
ammonium chloride completely abolished the develop-
ment of NZs. In contrast, a clear vesicular staining could
be detected. In contrast, chloroquine did not interfere with
the rapid cellular uptake through NZ.
Moreover, we probed for an involvement of heparan sulfate
in peptide uptake. For the uptake of the Tat protein, heparan
sulfate proteoglycans were shown to be involved in uptake
(40–42). In addition, the interaction of R9 with these
molecules was directly shown by biochemical experiments
(43). Heparan sulfate was selectively removed from the cell
surface by treatment of cells with heparinases. Interest-
ingly, different effects on uptake were observed at high and
low concentrations of R9 (Figure 8D). At a low peptide
concentration, heparinase treatment was without effect.
Neither the number of peptide-loaded vesicles nor their
cellular localization was altered. In contrast, at high peptide
concentration, the number of cells showing the develop-
ment of NZs as well as the rapid delivery of R9 into the
cytoplasm and nucleus was reduced. Therefore, the contri-
bution of heparan sulfate to peptide uptake is restricted to
the highly efficient uptake through NZs.
By electron microscopy, NZs are indistinguishable
from the rest of the cell
Light microscopy had failed to provide evidence for an
involvement of vesicular structures in the NZ-mediated
cellular uptake of nona-arginine and Tat. However, the
abolition of NZ formation in cells expressing DN dynamin
and the inhibitory effect of CPZ suggested a clathrin-
dependent endocytic process. To address the presence
of densely packed small vesicles that could not be resolved
by fluorescence light microscopy, a fine structural analysis
of NZ was performed by electron microscopy. Growth of
cells on coverslips structured with co-ordinates enabled
a matching of fluorescence and electron microscopy
images (Figure 9). Paraformaldehyde fixation required for
sample preparation for electron microscopy maintained
the distribution of fluorescence typical for NZ. By electron
microscopy, the NZs were indistinguishable from the
remainder of the cell. Individual clathrin-coated vesicles
could be identified within the cells (not shown). However,
there was no indication of an enrichment of clathrin-
containing structures either inside the cytoplasm or at
the plasma membrane.
The internalization of R9 at high peptide
concentrations is sensitive to the
PKC inhibitor rottlerin
Electron microscopy strongly indicated that uptake
through NZ occurs by an endocytosis-independent mech-
anism. In this case, the activity of CPZ should be because
of interference with processes other than assembly of the
AP-2 coat on nascent clathrin-coated vesicles (44). As CPZ
was shown to interfere with calcium-dependent signaling,
we therefore tested a panel of inhibitors reported to also
interfere with calcium-dependent and calcium-independent
signaling processes. Ro-318220, which inhibits the calcium-
dependent PKCa (45), was without effect on the inter-
nalization of 20 mM R9 through NZ. The PI-3 kinase inhibitor
wortmannin, which had been shown to interfere with
several steps in endosomal trafficking including clathrin-
dependent processes (46,47), was also without effect. In
contrast, preincubation of cells with rottlerin, a compound
that had been reported to be a preferential inhibitor of
PKCd (48), albeit with specificity that was questioned later
(45), abolished the development of the NZ (Figure 10A). R9
uptake at low peptide concentrations remained unaffected
(Figure 10B). Rottlerin had no effect on the endocytosis of
transferrin, providing further evidence that the activity of
CPZ on NZ-dependent import was not a result of interfer-
ence with clathrin-dependent endocytosis. In cells not
treated with rottlerin, the formation of NZ also had no
effect on the endocytosis of transferrin, showing that both
clathrin-dependent processes were spatially and function-
ally distinct (Figure 10A, top panels). Even though these
further inhibitor-based analyses do not provide a conclusive
picture on the molecular mechanism involved in the NZ-
mediated uptake, the inhibition by both CPZ and rottlerin
constitutes a distinct pharmacological profile of this pro-
cess, which may guide the further elucidation of this
uptake mechanism.
The formation of NZs is a characteristic of different
cell types
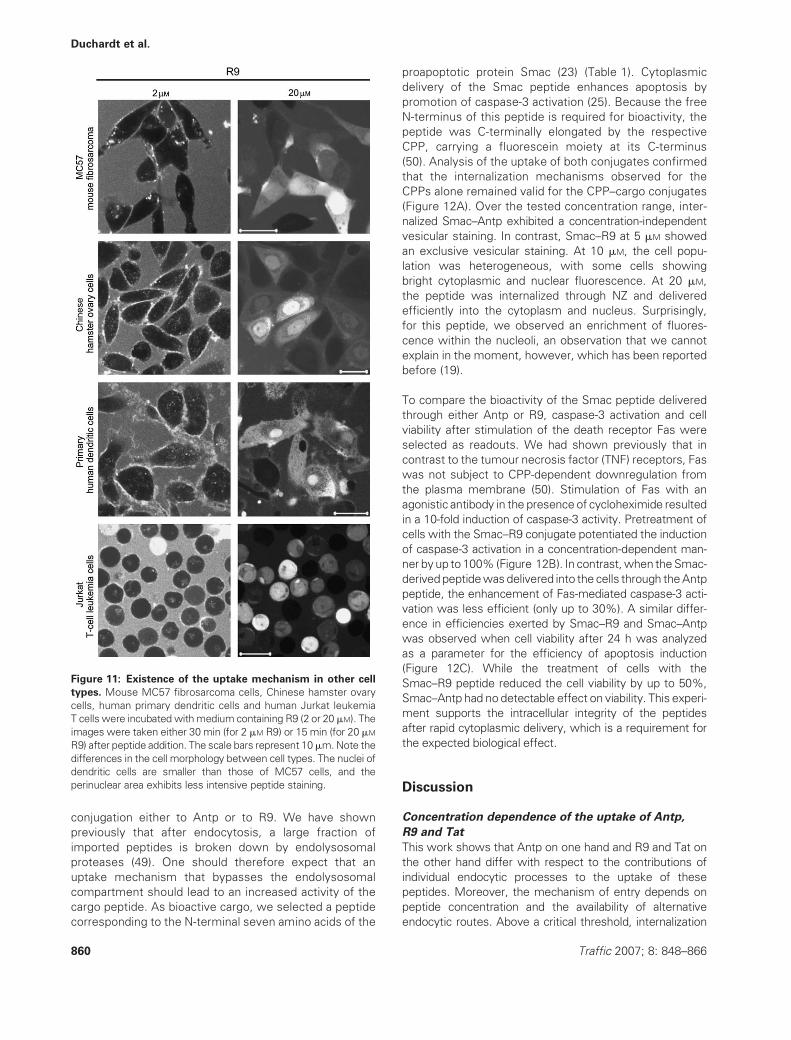
Having shown distinct characteristics of the uptake of Tat
and R9 through NZ, we next wanted to learn whether this
uptake mechanism is a particular characteristic of HeLa
cells. For this reason, we selected a panel of cells derived
from different species, different tissues and with different
growth characteristics: MC57, mouse fibrosarcoma cells;
Chinese hamster ovary cells; primary human dendritic cells
and human Jurkat T-cell leukemia cells. In all cell lines, we
observed a concentration-dependent formation of NZs
(Figure 11). In addition, in all cases, the cell population
was heterogeneous with respect to the formation of NZ.
These results show that the formation of NZ is not only
a mechanism present in cancer cells and furthermore that
cell attachment is not a prerequisite for this uptake
mechanism. For Jurkat cells, we tested whether incuba-
tion with MbCD could also induce cytoplasmic delivery of
Antp. In correspondence to the observation with HeLa
cells, Jurkat cells showed a homogeneous cytoplasmic
fluorescence, albeit for a higher fraction of cells.
858 Traffic 2007; 8: 848–866
Duchardt et al.

Relevance of the import mechanism for the cellular
activity of cargo peptides
Antennapedia, R9 and Tat have been used as vectors for
the cellular delivery of bioactive cargos. Having shown
that the cellular uptake and cytoplasmic delivery of Antp
are different from the one of the other two peptides, we
finally investigated the relevance of these import charac-
teristics for the bioactivity of a peptide delivered through
Figure 9: Electron microscopy of NZs. A and B) NZs in two different cells within one field of view. HeLa cells were grown on surface-
structured coverslips that enabled a matching of cells in fluorescence and electron microscopy images. Confocal laser scanning
microscopy was performed on paraformaldehyde-fixed cells. For each magnification, a pair of fluorescence and electron microscopy
images is shown. The fluorescence images in the center and bottom panels weremagnified beyond the resolution limit and are intended to
show the correspondence of the electron micrographs with NZ based on larger morphological features, such as the dark spots (left). Scale
bars are 30 mm (top), 6 mm (center) and 3 mm (bottom).
Figure 10: Sensitivity of R9 up-
take at high concentrations to
rottlerin. Cells were either treated
with rottlerin (20 mM) or remained
untreated and then medium contain-
ing either A) 20 mM R9 and transferrin
Alexa Fluor 633 conjugate (10mg/mL)
or B) 5 mM R9 was added to the cells.
Scale bars represent 20 mm. Note
that instrument settings for fluo-
rescein detection at 5 and 20 mM
were different because of the 10-
fold difference in intracellular signal
intensity.
Traffic 2007; 8: 848–866 859
Uptake of Cationic Cell-Penetrating Peptides
conjugation either to Antp or to R9. We have shown
previously that after endocytosis, a large fraction of
imported peptides is broken down by endolysosomal
proteases (49). One should therefore expect that an
uptake mechanism that bypasses the endolysosomal
compartment should lead to an increased activity of the
cargo peptide. As bioactive cargo, we selected a peptide
corresponding to the N-terminal seven amino acids of the
proapoptotic protein Smac (23) (Table 1). Cytoplasmic
delivery of the Smac peptide enhances apoptosis by
promotion of caspase-3 activation (25). Because the free
N-terminus of this peptide is required for bioactivity, the
peptide was C-terminally elongated by the respective
CPP, carrying a fluorescein moiety at its C-terminus
(50). Analysis of the uptake of both conjugates confirmed
that the internalization mechanisms observed for the
CPPs alone remained valid for the CPP–cargo conjugates
(Figure 12A). Over the tested concentration range, inter-
nalized Smac–Antp exhibited a concentration-independent
vesicular staining. In contrast, Smac–R9 at 5 mM showed
an exclusive vesicular staining. At 10 mM, the cell popu-
lation was heterogeneous, with some cells showing
bright cytoplasmic and nuclear fluorescence. At 20 mM,
the peptide was internalized through NZ and delivered
efficiently into the cytoplasm and nucleus. Surprisingly,
for this peptide, we observed an enrichment of fluores-
cence within the nucleoli, an observation that we cannot
explain in the moment, however, which has been reported
before (19).
To compare the bioactivity of the Smac peptide delivered
through either Antp or R9, caspase-3 activation and cell
viability after stimulation of the death receptor Fas were
selected as readouts. We had shown previously that in
contrast to the tumour necrosis factor (TNF) receptors, Fas
was not subject to CPP-dependent downregulation from
the plasma membrane (50). Stimulation of Fas with an
agonistic antibody in the presence of cycloheximide resulted
in a 10-fold induction of caspase-3 activity. Pretreatment of
cells with the Smac–R9 conjugate potentiated the induction
of caspase-3 activation in a concentration-dependent man-
ner byup to 100% (Figure 12B). In contrast,when theSmac-
derived peptidewasdelivered into the cells through theAntp
peptide, the enhancement of Fas-mediated caspase-3 acti-
vation was less efficient (only up to 30%). A similar differ-
ence in efficiencies exerted by Smac–R9 and Smac–Antp
was observed when cell viability after 24 h was analyzed
as a parameter for the efficiency of apoptosis induction
(Figure 12C). While the treatment of cells with the
Smac–R9 peptide reduced the cell viability by up to 50%,
Smac–Antp had no detectable effect on viability. This experi-
ment supports the intracellular integrity of the peptides
after rapid cytoplasmic delivery, which is a requirement for
the expected biological effect.
Discussion
Concentration dependence of the uptake of Antp,
R9 and Tat
This work shows that Antp on one hand and R9 and Tat on
the other hand differ with respect to the contributions of
individual endocytic processes to the uptake of these
peptides. Moreover, the mechanism of entry depends on
peptide concentration and the availability of alternative
endocytic routes. Above a critical threshold, internalization
Figure 11: Existence of the uptake mechanism in other cell
types. Mouse MC57 fibrosarcoma cells, Chinese hamster ovary
cells, human primary dendritic cells and human Jurkat leukemia
T cells were incubated withmedium containing R9 (2 or 20 mM). The
images were taken either 30 min (for 2 mM R9) or 15 min (for 20 mM
R9) after peptide addition. The scale bars represent 10mm.Note the
differences in the cell morphology between cell types. The nuclei of
dendritic cells are smaller than those of MC57 cells, and the
perinuclear area exhibits less intensive peptide staining.
860 Traffic 2007; 8: 848–866
Duchardt et al.

occurs through a highly efficient nonendocytic pathway.
This pathway originates from spatially confined areas of the
plasma membrane and leads to a rapid release of peptides
into the cytoplasm. The integrity of the plasmamembrane is
fully maintained. Antennapedia differs from the arginine-rich
CPPs nona-arginine and Tat in that much higher concen-
trations and at least partial blockade of endocytic uptake are
required for the induction of this pathway.
In the concentration range up to 40 mM, for the Antp
peptide, there is strong evidence that at least three
endocytic pathways, that is macropinocytosis, CME and
caveolae/lipid-raft-mediated endocytosis contribute to
uptake. Neither of these three pathways dominates.
Pharmacological interference with any one pathway yields
a distinct phenotype with respect to the subcellular
distribution of fluorescence and is without effect on the
other two pathways. The simultaneous inhibition of two
endocytic pathways leads to the addition of the respective
inhibitory effects.
For R9 and Tat at concentrations below 10 mM, entry of
these peptides is sensitive to MbCD and EIPA, strongly
indicative of entry through caveolae/lipid-raft-mediated
endocytosis and macropinocytosis. Remarkably, there
was no evidence for an involvement of CME at this
concentration. At a concentration of 5 mM, inhibition of
caveolae/lipid-raft-mediated endocytosis or macropinocy-
tosis induces the rapid NZ-dependent uptake mechanism.
The induction of this mechanism at a concentration
of peptide at which, in the absence of inhibitors, up-
take occurs through endocytosis indicates that a critical
concentration of peptide associated with the plasma
membrane is required. Because of the redirection of pepti-
des toward this pathway, the effect of EIPA and MbCD is
somewhat of a paradox: instead of inhibiting the uptake of
Figure 12: Relevance of the uptake mechanism for the bioactivity of a cargo peptide. A) HeLa cells were incubated with either
Smac–Antp or Smac–R9 at the indicated concentrations for 30 min at 378C. The scale bars represent 20 mm. B and C) After incubation of
cells with peptides for 30 min, the cells were washed and apoptosis was induced by stimulation with an agonistic Fas-specific antibody
(100 ng/mL) and CHX (2 mg/mL). B) For measurements of caspase-3 activity, cells were harvested 4 h after the induction of apoptosis. The
bars represent the results of three independent experiments. C) Determination of cell viability was performed using crystal violet staining
24 h after stimulation of Fas.
Traffic 2007; 8: 848–866 861
Uptake of Cationic Cell-Penetrating Peptides

peptides into the cells, at this concentration, these inhibi-
tors actually promote the uptake. This pathway mediates
a more rapid peptide uptake than the other two pathways,
one indication for a nonendocytic nature of this import
mechanism. When the rapid uptake mechanism is blocked
by CPZ, the phenotypes observed for cells treated with
MbCD or EIPA are recovered.
At higher concentrations, uptake occurs primarily by a
mechanism leading to rapid entry of the peptides into the
cytoplasm and nucleus. The EIPA- and MbCD-sensitive
mechanisms still contribute to peptide uptake as shown
by the fact that the inhibition of one of these pathways
still increases the cellular fluorescence even more. Con-
sidering our previous observations on the CPP-induced
endocytosis of TNF receptors by these three cationic CPPs
(50), one should note that Antp was more effective in
inducing the internalization of the receptors than R9 and
the Tat peptide. At low peptide concentrations, Antp was
in fact the only peptide inducing receptor internalization.
The internalization of EGF receptors suggested that this
internalization occurred through a clathrin-dependent mech-
anism. Consistently, at low concentrations, only entry of
Antp was CPZ sensitive. The internalization induced by Tat
and R9 at higher concentrations suggests that a fraction of
these peptides, in addition to the rapid NZ-mediated
uptake, was also internalized by clathrin-dependent endo-
cytosis, albeit with a much slower kinetics. For Antp, this
rapid cytoplasmic uptake was only observed at sig-
nificantly higher peptide concentrations for cells treated
with MbCD.
Uptake through NZs
The highly efficient uptake of R9 through NZs was heparin
sulfate and dynamin dependent; sensitive to ammonium
chloride, CPZ and rottlerin and insensitive to chloroquine.
The dynamin dependence and sensitivity to CPZ, when
considered in isolation, are strongly indicative of clathrin-
dependent endocytosis. However, the failure to detect
vesicles by electron microscopy; the rapid, stepwise
and spatially confined uptake kinetics and finally the con-
comitant formation of membrane blebs all point toward a
nonendocytic mechanism.
The ability to abolish this uptake by pharmacological
interventions that inhibit molecular processes inside the
cell indicates that association of peptide with the plasma
membrane alone is insufficient to initiate this process.
Instead, accumulation of peptide at the plasma membrane
induces an active process inside the cell. Apparently, this
process requires a certain minimum concentration of
membrane-associated peptide. At medium peptide con-
centrations, peptides are steadily removed from the
plasma membrane by endocytosis. Different import rates
of Antp and the other two peptides by the various
endocytic processes may therefore be one factor contri-
buting to the lower propensity of Antp to enter along this
pathway – a hypothesis that is supported by the ability to
induce this uptake by MbCD in HeLa and Jurkat cells.
Once endocytosis is blocked, peptides accumulate and the
threshold is reached. The initial formation of the NZ, bleb
formation and rapid release into the cytoplasm are tightly
coupled processes. All interventions that blocked this
uptake also blocked the formation of a NZ. It will be
interesting to seewhether an intervention can be identified
that only blocks the second step.
Considering the absence of vesicles in the electron
microscopy images, the blockade of the uptake by expres-
sion of DN dynamin is somewhat surprising. However, the
formation of blebs is a process involving rapid changes of
membrane morphology. Dynamins couple to a large num-
ber of cellular signaling processes (51), and even though it
is not clear why the peptides induce the formation of
outward protrusions, dynamins could well be involved in
such a process.
Interestingly, for any given cell for which uptake occurred
by distinctive steps, these steps frequently were similar
in size. One may hypothesize that the peptides are
enriched on specific domains of the plasma membrane
and that these domains have a certain capacity to bind
peptide that varies from cell to cell. At this point, neither
the nature of these domains nor the details of the uptake
mechanism is clear. At least, our observations favor an
‘accumulate and discharge’ mechanism over a mecha-
nism involving a transient formation of pores or continu-
ous transfer of peptides across the lipid bilayer. The FLIP
experiments also strongly support a transient association
of the peptides with molecules inside and on the surface
of the cell.
Heparan sulfates are candidate molecules for mediating
this association. Cell surface heparan sulfates have been
considered to play a role as multivalent, low-affinity
receptors of cationic CPPs. Remarkably, however, at low
peptide concentrations, removal of cell surface heparan
sulfates was without effect on uptake, at least as detect-
able by fluorescence microscopy. Consistent with our
results, for the Tat peptide at 100 mM, a requirement for
cell surface heparan sulfate for peptide was shown (14),
while at 1 mM, heparan sulfates were not required (52). It is
difficult to understand why these molecules should func-
tionally act as receptors only at high concentrations. One
possible mechanism may be through cross-linking by
polyvalent binding of peptides. Guanidinium groups have
been proposed to engage in bidentate interactions, pos-
sibly with heparan sulfates on the cell surface (53,54). The
higher fraction of arginine side chains in the R9 and Tat
oligopeptides in comparison with Antp may explain the
higher propensity of the former to induce this uptake
mechanism. Moreover, such a mechanism would be
consistent with ammonium chloride acting as an agent
that shields off negative charges on the cell surface.
862 Traffic 2007; 8: 848–866
Duchardt et al.

Reassessment of previous results
To this point, a number of publications have addressed the
mechanisms contributing to the cellular uptake of CPPs,
with sometimes conflicting results. Observations of a rapid
endocytosis-independent mechanism have been the latest
addition in this field (18,19). Our model provides a frame-
work to accommodate these previous observations. First
of all, our data show that the individual CPPs, albeit all
cationic in nature, possess remarkable differences in their
import pathways. Moreover, it is now clear that peptide
concentration is a key experimental condition when ana-
lyzing peptide uptake. Potocky et al. investigated the
cellular delivery of the Tat peptide at a concentration of
7 mM. Three different phenotypes with respect to the
distribution of the peptide in HeLa cells were observed
(21). While some cells exclusively exhibited a vesicular
staining, some cells showed a combination of vesicular
and cytoplasmic fluorescence, and finally in some cells,
fluorescence was only present in the cytoplasm and
nucleus. We show that this heterogeneity in the distribu-
tion of the peptide is likely related to the fact that
a concentration higher than 5 mM had been used. When
cells were incubated with the Tat peptide at concentra-
tions of 1–5 mM, the peptide was located only in vesicles
(20). The partial colocalization of the Tat peptide with
transferrin (11,21) as well as with dextran (20) may now
readily be explained by the finding that the peptides
simultaneously use several endocytic pathways.
To this point, we restricted ourselves to CPPs, two ofwhich
correspond to PTDs of the respective full-length proteins. It
needs to be resolved to which degree the import mecha-
nisms of the PTDs correspond to the ones of the full-length
proteins. Still observations reported for the Antp and Tat
peptide conjugated to protein cargos as well as for the full-
length Tat protein are also fully consistent with our model.
Antennapedia and Tat peptides differed substantially in
their capacity to deliver avidin into HeLa cells (29). For
a Tat-derived peptide (aa 11) and the Tat protein, conjugated
to green fluorescent protein (GFP), evidence was pre-
sented that both conjugates are internalized through the
same lipid-raft-dependent mechanism (17). The concentra-
tion of the peptide–GFP conjugate was less than 1 mM. At
this concentration, lipid-raft-mediated endocytosis as a key
uptake mechanism is fully consistent with our model.
Moreover, for a Tat–Cre conjugate used at a concentration
of less than 1 mM, both lipid-raft-mediated endocytosis and
macropinocytosis were identified as mechanisms contrib-
uting to internalization (15). Finally, both the Tat peptide and
the full-length Tat protein induce the internalization of TNF
receptors from the plasma membrane (50), indicating the
same mechanism of uptake.
The relevance of our findings for the application of CPPs is
supported by the fact that the uptake mechanisms identi-
fied for CPPs alone retain their validity also for cargo
peptides that are introduced into the cells through coupling
to CPPs. Moreover, the demonstration that the uptake
mechanism determines the cargo bioactivity stresses the
importance of understanding in detail the mode of inter-
nalization of CPPs for their application as delivery vectors.
In conclusion, our results provide a comprehensive frame-
work to encompass most previous observation on the
cellular uptake of cationic CPPs. One given CPP may not
only use different endocytic pathways for cellular entry but
may also use nonendocytic entry routes. Considering the
differences between Antp and the other two peptides, the
fraction of arginine side chains may be a relevant structural
characteristic, influencing the contribution of individual
entry pathways to uptake, possibly by induction of cross-
linking of cell surface heparan sulfates. Concerning the
rapid cytoplasmic uptake, a distinctive pharmacological
profile was defined. The data indicate that the uptake is
a specific response of the cell to a local enrichment of
cationic molecules at the plasma membrane. It is not clear
so far whether the CPPs initiate the enrichment of the
cellular molecules to which they associate or whether the
NZ represent preformedmembrane domains. Finally, it will
be interesting to investigate whether a physiological role
for this pathway exists.
Materials and Methods
Cells and reagentsThe human cervical carcinoma cell line HeLa was obtained from the
American Type Culture Collection (Manassas, VA, USA). HeLadynK44A
expressing a DN form of dynamin-2 under the control of a tet on/off
promoter, cultured in medium containing 1 mg/mL tetracycline, was a kind
gift from Bo van Deurs (University of Copenhagen, Copenhagen, Denmark).
The expression of mutant dynamin was induced by tetracycline deprivation
for at least 24 h. Transferrin Alexa Fluor 633 conjugate, dextran Alexa Fluor
647 conjugate and CTB Alexa Fluor 555 conjugate and Zenon mouse IgG1
labeling kit (specific for the Fc part of immunoglobulin G1 antibodies) were
purchased from Mobitech (Gottingen, Germany). Fluorogenic caspase-3
substrate (Ac-DEVD-AMC), wortmannin, Ro-318220, rottlerin and CPZ
were from Calbiochem (Bad Soden, Germany); ammonium chloride, EIPA,
MbCD, MTT and heparinase I, II and III were obtained from Sigma
(Deisenhofen, Germany). The anti-Fas-activating antibody was obtained
from Upstate (Hamburg, Germany). BSA was obtained from SIGMA
(Steinheim, Germany). The anti-heparan sulfate proteoglycan (HSPG)
vesicular stomatitis virus (VSV)-tagged single-chain antibody HS4C3 (55)
and the mouse anti-VSV (clone P5D4) antibody were a kind gift of Toin van
Kuppevelt (Radboud University Nijmegen Medical Centre, Nijmegen, The
Netherlands).
Peptide synthesisAutomated peptide synthesis was performed by solid-phase Fmoc/tBu-
chemistry using an automated peptide synthesizer for multiple peptide
synthesis (RSP5032; Tecan, Hombrechtlikon, Switzerland) in 2-mL syringes
according to a protocol described elsewhere (20). Smac–Antp and Smac–
R9 (Table 1) were synthesized using a Na-carboxyfluorescein-labeled lysyl-
Rink amide resin (56). The purity of all peptides was determined by
analytical high-performance liquid chromatography (HPLC). The identity of
the peptides was confirmed by MALDI-TOF mass spectrometry. Peptides
with a purity of less than 95% were purified by preparative HPLC.
Flow cytometryHeLa cells were incubated with medium containing peptides at the
indicated concentrations for 30 min at 378C. After incubation, the cells
Traffic 2007; 8: 848–866 863
Uptake of Cationic Cell-Penetrating Peptides

were washed with medium, detached by trypsinization for 5 min, washed,
suspended in PBS and measured immediately by flow cytometry (BD
FACSCalibur System; Becton Dickinson, Heidelberg, Germany). In each
case, the fluorescence of 10 000 vital cells was acquired. Vital cells were
gated based on sideward and forward scatter.
Confocal laser scanning microscopyConfocal laser scanning microscopy was performed on an inverted LSM510
laser scanning microscope (Carl Zeiss, Gottingen, Germany) using a Plan-
Apochromat 63� 1.4 N.A. lens. All measurements of peptide uptake were
performed with living, nonfixed cells grown in eight-well chambered cover
glasses (Nunc, Wiesbaden, Germany). Cells were seeded at a density of
4 � 104/well 1 day before the experiment and cultured in RPMI-1640
supplemented with 10% fetal calf serum. For detection of fluorescein-
labeled peptides, the 488-nm line of an argon ion laser was directed over an
HFT UV/488 beam splitter, and fluorescence was detected with a BP 505–
550 band pass filter. For the simultaneous detection of fluorescein-labeled
peptides and Alexa Fluor 633 or -647 conjugates, the 488-nm line of an
argon ion laser and the light of a 633-nm helium neon laser were directed
over an HFT UV/488/633 beam splitter, and fluorescence was detected
using an NFT 545 beam splitter in combination with a BP 505–530 band
pass filter for fluorescein detection and an LP 650 long pass filter for Alexa
Fluor 633 and Alexa Fluor 647 detection. Life cell microscopy was
performed at room temperature.
Quantitative analysis of colocalizationFirst, color channels containing the signal of fluorescein-labeled Antp and
the respective tracer were extracted from the multichannel confocal
images. After low-pass filtering to reduce image noise, for both channels,
binary masks corresponding to vesicle-associated fluorescence were
generated (MaskAntp and MaskTracer). Threshold levels were selected by
visual inspection. A third mask containing those pixels in which vesicle-
associated fluorescence was present in both channels was generated by
applying an AND-operation to the masks for peptide- and tracer-associated
fluorescence (MaskColoc). Because of the presence of peptide and tracer in
the medium, all three masks also included all pixels outside the cells. For
this reason, individual cells were selected as ROI and the number of objects
(NAntp, NTracer and NColoc) within each cell corresponding to individual
vesicles or small groups of vesicles in all three binary masks was counted
using the object analysis routines within Image Pro Plus 4.5 (Media
Cybernetics Inc., Silver Spring, MD, USA). The fraction of tracer-positive
vesicles colocalizing with Antp was calculated by dividing NColoc by NTracer,
and the fraction of Antp-positive vesicles colocalizing with tracer was
calculated by dividing NColoc by NAntp.
Electron microscopyHeLa cells were grown on CELLocate 5245, with a square size of 55 mm
(Eppendorf, Hamburg, Germany). After addition of medium containing R9 in
a concentration of 20 mM, formation of NZ was followed by confocal laser
scanning microscopy as described previously. Directly after appearance of
NZ, cells were washed with ice-cold PBS and subsequently fixed with 4%
formaldehyde in PBS for 30 min at room temperature. For electron
microscopy, cells were stored for 1 h at 48C, postfixed with 1% osmium
tetroxide in 100 mM phosphate buffer at pH 7.2 for 1 h on ice, washed with
H2O, treated with 1% aqueous uranyl acetate for 1 h at 48C, dehydratedthrough a graded series of ethanol and embedded in Epon. Ultrathin
sections were stained with uranyl acetate and lead citrate and viewed in
a Philips CM10 electron microscope. The imprint of the cellocate coverslip
in the Epon resin was used as an orientation for the matching of
fluorescence and electron microscopy images.
Incubation with inhibitorsCells were treated with the indicated inhibitors for 30 min at 378C. Then,the medium was removed and medium containing peptide as well as the
corresponding inhibitor was added. After 30 min of incubation at 378C,the cells were washed twice with medium and analyzed by fluorescence
microscopy or flow cytometry.
ImmunofluorescenceHeLa cells were seeded at a density of 4 � 104/well in eight-well
chambered covered glasses and incubated with medium containing 10 U/
mL of heparinase I, 5 U/mL heparinase II and 2 U/mL heparinase III for 6 h at
378C. Then, the cells were washed with ice-cold PBS containing 0.1% (w/v)
BSA and incubated with the anti-HSPG antibody HS4C3 on ice. After 1.5 h
of incubation, cells were washed with ice-cold PBS/BSA. To visualize bound
antibodies, the cells were incubated for 1 h with an anti-VSV antibody
(P5D4)/Zenon Alexa Fluor 647 conjugate on ice. The antibody staining was
analyzed by confocal laser scanning microscopy.
Caspase-3 activity assayHeLa cells were incubated with medium containing peptides for 30 min at
378C. After one washing step, cells were treated as indicated with agonistic
Fas antibody and cycloheximide (CHX) for the induction of apoptosis,
followed by incubation for further 3 h. Cells were harvested by scraping,
washed with ice-cold PBS and lysed in lysis buffer [1% Triton, 150 mM
NaCl, pH 7.7, supplemented with protease inhibitor cocktail tablets (Roche
Diagnostics, Mannheim, Germany)] for 30min on ice. The protein content in
lysates was determined using a commercially available Bradford protein
assay kit (Bio Rad Laboratories, Munchen, Germany). Equivalents of 30 mg
protein for each sample were diluted in caspase activity buffer (20 mM
HEPES, 10 mM dithiothreitrol, 10% glycerol, 100 mM NaCl, pH 7.5).
Caspase-3 substrate was added to the samples to a final concentration of
2 mM. The efficiency of the substrate cleavage by active caspase-3 was
analyzed immediately after substrate addition and after 1 h of incubation at
378C using a luminescence spectrometer LS50B (PerkinElmer, Norwalk,
CT, USA).
MTT assayHeLa cells were seeded in 96-well microtiter plates (1.5 � 104/well) and
cultivated over night. The next day, cells were incubated with peptides for
6 h at 378C. Cell viability was measured using the colorimetric MTT dye.
Cells were incubated with MTT at a concentration of 1 mg/mL for 4 h. The
formazan product was solubilized with SDS [10% (w/v) in 10 mM HCl]. Cell
viability was determined by measuring the absorbance of each sample at
570 nm using a microplate reader (Molecular Devices SpectraMax 340;
GMI, Ramsey, Minnesota, USA).
Cytotoxicity assayHeLa cells were treated with the indicated peptides for 30 min at 378C,washed and stimulated with the agonistic Fas antibody (100 ng/mL) and
CHX (2 mg/mL) for the induction of apoptosis. After an additional 24-h
incubation at 378C, cells in both groups were washed with PBS, followed by
crystal violet staining [20% (v/v) methanol, 0.5% (w/v) crystal violet] for
15 min. The wells were washed with H2O and air-dried. The dye was dis-
solved in methanol and the optical density at 550 nm measured with an
enzyme-linked immunosorbent assay plate reader.
Acknowledgments
We thank Viktoria Wolf for the isolation and cultivation of human dendritic
cells, Brigitte Sailer for oriented cutting of ultrathin sections and Anne
Spang and Mark Trautwein for helpful discussions. F. D. is a scholar of the
Graduiertenkolleg 794. R. B. gratefully acknowledges financial support
from the Volkswagen Foundation (‘‘Nachwuchsgruppen an Universitaten’’
I/77 472).
Supplementary Materials
Figure S1: The effect of inhibitors of endocytosis on the internaliza-
tion of tracer molecules. HeLa cells were incubated for 30 min at 378C
with the indicated inhibitors of endocytosis (CPZ 10 mg/mL, MbCD 5 mM
864 Traffic 2007; 8: 848–866
Duchardt et al.

and EIPA 100 mM) or remained untreated (control groups).Then, the
medium was removed and cells were incubated for further 30 min with
a transferrin Alexa Fluor 633 conjugate (10 mg/mL), a dextran Alexa Fluor
647 conjugate (10 mM) or CTB Alexa Fluor 555 (10 mg/mL) in the presence or
absence of the corresponding inhibitor. The scale bars represent 20 mm.
Figure S2: The effect of endocytic inhibitors on cell viability. HeLa cells
were incubated with inhibitors of endocytosis, either alone or in combina-
tion, at the indicated concentrations for 2 h at 378C. Cell viability was
determined using crystal violet staining. The frames indicate the concen-
trations of inhibitors applied in the experiments.
Video S1: Endocytosis of R9 through NZs. Medium containing R9 at
a concentration of 20 mM was added to HeLa cells and uptake of
fluorescence was followed by time-lapse confocal microscopy. The time
series was started 1 min after addition of peptide. The video comprises 50
images, with a time interval of 10 seconds between each image.
Supplemental materials are available as part of the online article at http://
www.blackwell-synergy.com
References
1. Joliot A, Pernelle C, Deagostini-Bazin H, Prochiantz A. Antennapedia
homeobox peptide regulates neural morphogenesis. Proc Natl Acad Sci
USA 1991;88:1864–1868.
2. Frankel AD, Pabo CO. Cellular uptake of the tat protein from human
immunodeficiency virus. Cell 1988;55:1189–1193.
3. Malecki J, Wesche J, Skjerpen CS, Wiedlocha A, Olsnes S. Trans-
location of FGF-1 and FGF-2 across vesicular membranes occurs
during G1-phase by a common mechanism. Mol Biol Cell 2004;15:
801–814.
4. Derossi D, Joliot AH, Chassaing G, Prochiantz A. The third helix of
the Antennapedia homeodomain translocates through biological
membranes. J Biol Chem 1994;269:10444–10450.
5. Vives E, Brodin P, Lebleu B. A truncated HIV-1 Tat protein basic domain
rapidly translocates through the plasma membrane and accumulates in
the cell nucleus. J Biol Chem 1997;272:16010–16017.
6. Mitchell DJ, Kim DT, Steinman L, Fathman CG, Rothbard JB. Poly-
arginine enters cells more efficiently than other polycationic homo-
polymers. J Pept Res 2000;56:318–325.
7. Dietz GPH, Bahr M. Delivery of bioactive molecules into the cell: the
Trojan horse approach. Mol Cell Neurosci 2004;27:85–131.
8. Snyder EL, Dowdy SF. Cell penetrating peptides in drug delivery.
Pharm Res 2004;21:389–393.
9. Prochiantz A. Messenger proteins: homeoproteins, TAT and others.
Curr Opin Cell Biol 2000;12:400–406.
10. Drin G, Cottin S, Blanc E, Rees AR, Temsamani J. Studies on the
internalization mechanism of cationic cell-penetrating peptides. J Biol
Chem 2003;278:31192–31201.
11. Richard JP, Melikov K, Vives E, Ramos C, Verbeure B, Gait MJ,
Chernomordik LV, Lebleu B. Cell-penetrating peptides: a reevaluation
of the mechanism of cellular uptake. J Biol Chem 2003;278:585–590.
12. Vendeville A, Rayne F, Bonhoure A, Bettache N, Montcourrier P,
Beaumelle B. HIV-1 Tat enters T cells using coated pits before trans-
locating from acidified endosomes and eliciting biological responses.
Mol Biol Cell 2004;15:2347–2360.
13. Console S, Marty C, Garcia-Echeverria C, Schwendener R, Ballmer-
Hofer K. Antennapedia and HIV TAT ‘‘protein transduction domains’’
promote endocytosis of high Mr cargo upon binding to cell surface
glycosaminoglycans. J Biol Chem 2003;278:35109–35114.
14. Ziegler A, Seelig J. Interaction of the protein transduction domain of
HIV-1 TAT with heparan sulfate: binding mechanism and thermody-
namic parameters. Biophys J 2004;86:254–263.
15. Wadia JS, Stan RV, Dowdy SF. Transducible TAT-HA fusogenic peptide
enhances escape of TAT-fusion proteins after lipid raft macropino-
cytosis. Nat Med 2004;10:310–315.
16. Ferrari A, Pellegrini V, Arcangeli C, Fittipaldi A, Giacca M, Beltram F.
Caveolae-mediated internalization of extracellular HIV-1 tat fusion
proteins visualized in real time. Mol Ther 2003;8:284–294.
17. Fittipaldi A, Ferrari A, Zoppe M, Arcangeli C, Pellegrini V, Beltram F,
Giacca M. Cell membrane lipid rafts mediate caveolar endocytosis of
HIV-1 tat fusion proteins. J Biol Chem 2003;278:34141–34149.
18. Ziegler A, Nervi P, Durrenberger M, Seelig J. The cationic cell-
penetrating peptide CPP(TAT) derived from the HIV-1 protein TAT is
rapidly transported into living fibroblasts: optical, biophysical, and
metabolic evidence. Biochemistry 2005;44:138–148.
19. Tunnemann G, Martin RM, Haupt S, Patsch C, Edenhofer F, Cardoso
MC. Cargo-dependent mode of uptake and bioavailability of TAT-
containing proteins and peptides in living cells. FASEB J 2006;20:
1775–1784.
20. Fischer R, Kohler K, Fotin-Mleczek M, Brock R. A stepwise dissection
of the intracellular fate of cationic cell-penetrating peptides. J Biol
Chem 2004;279:12625–12635.
21. Potocky TB, Menon AK, Gellman SH. Cytoplasmic and nuclear delivery
of a TAT-derived peptide and a beta-peptide after endocytic uptake into
HeLa cells. J Biol Chem 2003;278:50188–50194.
22. Richard JP, Melikov K, Brooks H, Prevot P, Lebleu B, Chernomordik LV.
Cellular uptake of unconjugated TAT peptide involves clathrin-
dependent endocytosis and heparan sulfate receptors. J Biol Chem
2005;280:15300–15306.
23. Wu G, Chai J, Suber TL, Wu JW, Du C, Wang X, Shi Y. Structural
basis of IAP recognition by Smac/DIABLO. Nature 2000;408:
1008–1012.
24. Fotin-Mleczek M, Fischer R, Brock R. Endocytosis and cationic
cell-penetrating peptides – a merger of concepts and methods. Curr
Pharm Des 2005;11:3613–3628.
25. Fulda S, Wick W, Weller M, Debatin KM. Smac agonists sensitize for
Apo2L/TRAIL- or anticancer drug-induced apoptosis and induce regres-
sion of malignant glioma in vivo. Nat Med 2002;8:808–815.
26. Arnt CR, Chiorean MV, Heldebrant MP, Gores GJ, Kaufmann SH.
Synthetic Smac/DIABOLO peptides enhance the effects of chemo-
therapeutic agents by binding XIAP and cIAP1 in situ. J Biol Chem
2002;277:44236–44234.
27. Torgerson TR, Colosia AD, Donahue JP, Lin Y-Z, Hawiger J. Regula-
tion of NF-kappa B, AP-1, NFAT, and STAT1 nuclear import in T
lymphocytes by noninvasive delivery of peptide carrying the nuclear
localization sequence of NF-kappa B p50. J Immunol 1998;161:
6084–6092.
28. Borsello T, Clarke PGH, Hirt L, Vercelli A, Repici M, Schorderet DF,
Bogousslavsky J, Bonny C. A peptide inhibitor of c-Jun N-terminal
kinase protects against excitotoxicity and cerebral ischemia. Nat Med
2003;9:1180–1186.
29. Saalik P, Elmquist A, Hansen M, Padari K, Saar K, Viht K, Langel U,
PoogaM. Protein cargo delivery properties of cell-penetrating peptides.
A comparative study. Bioconjug Chem 2004;15:1246–1253.
30. Subtil A, Hemar A, Dautry-Varsat A. Rapid endocytosis of interleukin 2
receptors when clathrin-coated pit endocytosis is inhibited. J Cell Sci
1994;107:3461–3468.
31. West MA, Bretscher MS, Watts C. Distinct endocytotic pathways
in epidermal growth factor-stimulated human carcinoma A431 cells.
J Cell Biol 1989;109:2731–2739.
32. Keller P, Simons K. Cholesterol is required for surface transport of
influenza virus hemagglutinin. J Cell Biol 1998;140:1357–1367.
Traffic 2007; 8: 848–866 865
Uptake of Cationic Cell-Penetrating Peptides

33. Fretz M, Jin J, Conibere R, Penning NA, Al Taei S, Storm G, Futaki S,
Takeuchi T, Nakase I, Jones AT. Effects of Naþ/Hþ exchanger
inhibitors on subcellular localisation of endocytic organelles and intra-
cellular dynamics of protein transduction domains HIV-TAT peptide and
octaarginine. J Control Release 2006;116:247–254.
34. Kielholz P, Labhardt F. Treatment of mental disorders with chlorprom-
azine. J Clin Exp Psychopathol 1956;17:38–44.
35. Seeman P. Atypical antipsychotics: mechanism of action. Can J
Psychiatry 2002;47:27–38.
36. Marshak DR, Lukas TJ, Watterson DM. Drug-protein interactions:
binding of chlorpromazine to calmodulin, calmodulin fragments, and
related calcium binding proteins. Biochemistry 1985;24:144–150.
37. Lippincott-Schwartz J, Altan-Bonnet N, Patterson GH. Photobleaching
and photoactivation: following protein dynamics in living cells. Nat Cell
Biol 2003;5(Suppl.):S7–S14.
38. van der Bliek AM, Redelmeier TE, Damke H, Tisdale EJ, Meyerowitz
EM, Schmid SL. Mutations in human dynamin block an intermediate
stage in coated vesicle formation. J Cell Biol 1993;122:553–563.
39. Henley JR, Krueger EW, Oswald BJ, McNiven MA. Dynamin-mediated
internalization of caveolae. J Cell Biol 1998;141:85–99.
40. Argyris EG, Kulkosky J, Meyer ME, Xu Y, Mukhtar M, Pomerantz RJ,
Williams KJ. The perlecan heparan sulfate proteoglycan mediates
cellular uptake of HIV-1 Tat through a pathway responsible for biological
activity. Virology 2004;330:481–486.
41. Tyagi M, Rusnati M, Presta M, Giacca M. Internalization of HIV-1 tat
requires cell surface heparan sulfate proteoglycans. J Biol Chem 2001;
276:3254–3261.
42. Rusnati M, Coltrini D, Oreste P, Zoppetti G, Albini A, Noonan D, d’Adda
di Fagagna F, Giacca M, Presta M. Interaction of HIV-1 Tat protein with
heparin. Role of the backbone structure, sulfation, and size. J Biol
Chem 1997;272:11313–11320.
43. Goncalves E, Kitas E, Seelig J. Binding of oligoarginine to membrane
lipids and heparan sulfate: structural and thermodynamic characteriza-
tion of a cell-penetrating peptide. Biochemistry 2005;44:2692–2702.
44. Wang LH, Rothberg KG, Anderson RG. Mis-assembly of clathrin
lattices on endosomes reveals a regulatory switch for coated pit
formation. J Cell Biol 1993;123:1107–1117.
45. Davies SP, Reddy H, Caivano M, Cohen P. Specificity and mechanism
of action of some commonly used protein kinase inhibitors. Biochem J
2000;351:95–105.
46. van Dam EM, Ten Broeke T, Jansen K, Spijkers P, Stoorvogel W.
Endocytosed transferrin receptors recycle via distinct dynamin and
phosphatidylinositol 3-kinase-dependent pathways. J Biol Chem
2002;277:48876–48883.
47. Hunyady L, Baukal AJ, Gaborik Z, Olivares-Reyes JA, Bor M, Szaszak
M, Lodge R, Catt KJ, Balla T. Differential PI 3-kinase dependence of
early and late phases of recycling of the internalized AT1 angiotensin
receptor. J Cell Biol 2002;157:1211–1222.
48. Gschwendt M, Muller HJ, Kielbassa K, Zang R, Kittstein W, Rincke G,
Marks F. Rottlerin, a novel protein kinase inhibitor. Biochem Biophys
Res Commun 1994;199:93–98.
49. Fischer R, Bachle D, Fotin-Mleczek M, Jung G, Kalbacher H, Brock R.
A targeted protease substrate for a quantitative determination of pro-
teolytic activities in the endolysosomal pathway. Chembiochem 2006;
7:1428–1434.
50. Fotin-Mleczek M, Welte S, Mader O, Duchardt F, Fischer R, Hufnagel
H, Scheurich P, Brock R. Cationic cell-penetrating peptides interfere
with TNF signaling by induction of TNF receptor internalization. J Cell
Sci 2005;118:3339–3351.
51. Praefcke GJ, McMahon HT. The dynamin superfamily: universal
membrane tubulation and fission molecules? Nat Rev Mol Cell Biol
2004;5:133–147.
52. Silhol M, Tyagi M, Giacca M, Lebleu B, Vives E. Different mechanisms
for cellular internalization of the HIV-1 Tat-derived cell penetrating
peptide and recombinant proteins fused to Tat. Eur J Biochem
2002;269:494–501.
53. Fuchs SM, Raines RT. Pathway for polyarginine entry into mammalian
cells. Biochemistry 2004;43:2438–2444.
54. Rothbard JB, Jessop TC, Lewis RS, Murray BA, Wender PA. Role of
membrane potential and hydrogen bonding in the mechanism of
translocation of guanidinium-rich peptides into cells. J Am Chem Soc
2004;126:9506–9507.
55. van Kuppevelt TH, Dennissen MA, van Venrooij WJ, Hoet RM,
Veerkamp JH. Generation and application of type-specific anti-heparan
sulfate antibodies using phage display technology. Further evidence for
heparan sulfate heterogeneity in the kidney. J Biol Chem 1998;273:
12960–12966.
56. Fischer R, Mader O, Jung G, Brock R. Extending the applicability of
carboxyfluorescein in solid-phase synthesis. Bioconjug Chem 2003;14:
653–660.
866 Traffic 2007; 8: 848–866
Duchardt et al.
Copyright © 2022 FDOKUMEN




















