
Alpha-Helical Cationic Anticancer Peptides: A Promising Candidate for Novel Anticancer Drugs
9
1875-5607/15 $58.00+.00 © 2015 Bentham Science Publishers Send Orders for Reprints to [email protected] Mini-Reviews in Medicinal Chemistry, 2015, 15, 73-81 73 Alpha-Helical Cationic Anticancer Peptides: A Promising Candidate for Novel Anticancer Drugs Yibing Huang 1,2,3 , Qi Feng 1,3 , Qiuyan Yan 1,3 , Xueyu Hao 1,3 and Yuxin Chen 1,2,3* 1 Key Laboratory for Molecular Enzymology and Engineering of the Ministry of Education, Jilin University, Changchun, China; 2 National Engineering Laboratory for AIDS Vaccine, Jilin University, Changchun, China; 3 College of Life Sciences, Jilin University, Changchun, China Abstract: Cancer has become a serious concern in public health. Harmful side effects and multidrug resis- tance of traditional chemotherapy have prompted urgent needs for novel anticancer drugs or therapeutic ap- proaches. Anticancer peptides (ACPs) have become promising molecules for novel anticancer agents because of their unique mechanism and several extraordinary properties. Most α-helical ACPs target the cell mem- brane, and interactions between ACPs and cell membrane components are believed to be the key factor in the selective killing of cancer cells. In this review, we focus on the exploitation of the structure and function of α-helical ACPs, includ- ing the distinction between cancer and normal cells, the proposed anticancer mechanisms, and the influence of physico- chemical parameters of α-helical ACPs on the biological activities and selectivity against cancer cells. In addition, the de- sign and modification methods to optimize the cell selectivity of α-helical ACPs are considered. Furthermore, the suitabil- ity of ACPs as cancer therapeutics is discussed. Keywords: Alpha-helical anticancer peptide, cell selectivity, electrostatic interaction, action mechanism, physicochemical parameter. 1. INTRODUCTION Cancer has become the most malignant disease, threaten- ing human health and life, particularly in the developing countries [1, 2]. According to the World Cancer Report 2014 published by the World Health Organization (WHO), the worldwide prevalence of cancer had risen to approximately 14 million new cases per year and 8.2 million cancer deaths annually in 2012. This figure is expected to rise to 22 million new cases annually within the next 2 decades and cancer deaths are predicted to rise to an estimated 13 million per year. At present, among surgery, chemotherapy or radiother- apy, chemotherapy is still the main strategy to treat most cancer cases. The anticancer agents are mostly based on alkylating agents and antimetabolites [3] which exhibit activ- ity against cancer cells by inhibiting DNA synthesis, DNA replication, or microtubule polymerization [4]; however, both these forms of therapy are not successful in more than 50% of cases because of low therapeutic indices and broad spectrum side effects [5-7]. In addition, the development of multidrug resistance of cancer cells is another major limita- tion [8]. Thus, the development of a novel class of anticancer agents or therapy is urgently required. Cationic antimicrobial peptides (host defense peptides, AMPs) are widely distributed in almost all species including mammals, plants, birds, amphibians, fishes, insects, and microbes, and several reviews have highlighted the structure, *Address correspondence to this author at the Key Laboratory for Molecular Enzymology and Engineering of the Ministry of Education, College of Life Sciences, Jilin University, 2699 Qianjin Street, Changchun 130012, China; Tel: +86-431-85155220; Fax: +86-431-85155200; E-mail: [email protected] activity, and the mechanism of action of AMPs [9-16]. AMPs are highly effective and play a key role in innate im- mune systems against pathogens including bacteria, viruses, parasites, fungi, and even cancer cells [17-22]. Till date, more than 2400 AMPs have been found and 166 peptides have exhibited anticancer activities [named as anticancer peptides (ACPs)], refer to the website: http://aps.unmc.edu/ AP/main.php. Compared with traditional antibiotics and chemotherapy, AMPs display several extraordinary proper- ties such as broad spectrum activity and rapid action and also make it difficult for cancer cells to develop resistance against them. Hence, AMPs have become promising molecules for novel antibiotics and anticancer agents [23-25]. Despite the fact that ACPs are from different sources and have low homologies, they share some common characteris- tics such as a positive charge, an amphipathic conformation, the ability to fold into a secondary structure (α-helical or β - sheet) in a hydrophobic environment, and exhibition of broad spectrum and rapid anticancer activity [26-28]. In this re- view, we present an overview of the α-helical ACPs, includ- ing the distinction between cancer and normal cells, the proposed anticancer mechanism of α -helical ACPs, and the role of physicochemical parameters of α-helical ACPs on the biological activity and selectivity. In addition, we also dis- cuss the design and the modification method to optimize the cell selectivity of α-helical ACPs. 2. MECHANISM OF ACTION The α -helical cationic ACPs are a major class of ACPs that generally exhibit the following properties: (1) they are rich in basic amino acids, such as lysine and arginine, result- ing in a net positive charge of peptides from +2 to +9 in neutral pH; (2) they are usually composed of 5-40 amino Yuxin Chen
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Alpha-Helical Cationic Anticancer Peptides: A Promising Candidate for Novel Anticancer Drugs

1875-5607/15 $58.00+.00 © 2015 Bentham Science Publishers
Send Orders for Reprints to [email protected]
Mini-Reviews in Medicinal Chemistry, 2015, 15, 73-81 73
Alpha-Helical Cationic Anticancer Peptides: A Promising Candidate for Novel Anticancer Drugs Yibing Huang1,2,3, Qi Feng1,3, Qiuyan Yan1,3, Xueyu Hao1,3 and Yuxin Chen1,2,3*
1Key Laboratory for Molecular Enzymology and Engineering of the Ministry of Education, Jilin University, Changchun, China; 2National Engineering Laboratory for AIDS Vaccine, Jilin University, Changchun, China; 3College of Life Sciences, Jilin University, Changchun, China
Abstract: Cancer has become a serious concern in public health. Harmful side effects and multidrug resis-tance of traditional chemotherapy have prompted urgent needs for novel anticancer drugs or therapeutic ap-proaches. Anticancer peptides (ACPs) have become promising molecules for novel anticancer agents because of their unique mechanism and several extraordinary properties. Most α-helical ACPs target the cell mem-brane, and interactions between ACPs and cell membrane components are believed to be the key factor in the selective killing of cancer cells. In this review, we focus on the exploitation of the structure and function of α-helical ACPs, includ-ing the distinction between cancer and normal cells, the proposed anticancer mechanisms, and the influence of physico-chemical parameters of α-helical ACPs on the biological activities and selectivity against cancer cells. In addition, the de-sign and modification methods to optimize the cell selectivity of α-helical ACPs are considered. Furthermore, the suitabil-ity of ACPs as cancer therapeutics is discussed.
Keywords: Alpha-helical anticancer peptide, cell selectivity, electrostatic interaction, action mechanism, physicochemical parameter.
1. INTRODUCTION
Cancer has become the most malignant disease, threaten-ing human health and life, particularly in the developing countries [1, 2]. According to the World Cancer Report 2014 published by the World Health Organization (WHO), the worldwide prevalence of cancer had risen to approximately 14 million new cases per year and 8.2 million cancer deaths annually in 2012. This figure is expected to rise to 22 million new cases annually within the next 2 decades and cancer deaths are predicted to rise to an estimated 13 million per year. At present, among surgery, chemotherapy or radiother-apy, chemotherapy is still the main strategy to treat most cancer cases. The anticancer agents are mostly based on alkylating agents and antimetabolites [3] which exhibit activ-ity against cancer cells by inhibiting DNA synthesis, DNA replication, or microtubule polymerization [4]; however, both these forms of therapy are not successful in more than 50% of cases because of low therapeutic indices and broad spectrum side effects [5-7]. In addition, the development of multidrug resistance of cancer cells is another major limita-tion [8]. Thus, the development of a novel class of anticancer agents or therapy is urgently required. Cationic antimicrobial peptides (host defense peptides, AMPs) are widely distributed in almost all species including mammals, plants, birds, amphibians, fishes, insects, and microbes, and several reviews have highlighted the structure, *Address correspondence to this author at the Key Laboratory for Molecular Enzymology and Engineering of the Ministry of Education, College of Life Sciences, Jilin University, 2699 Qianjin Street, Changchun 130012, China; Tel: +86-431-85155220; Fax: +86-431-85155200; E-mail: [email protected]
activity, and the mechanism of action of AMPs [9-16]. AMPs are highly effective and play a key role in innate im-mune systems against pathogens including bacteria, viruses, parasites, fungi, and even cancer cells [17-22]. Till date, more than 2400 AMPs have been found and 166 peptides have exhibited anticancer activities [named as anticancer peptides (ACPs)], refer to the website: http://aps.unmc.edu/ AP/main.php. Compared with traditional antibiotics and chemotherapy, AMPs display several extraordinary proper-ties such as broad spectrum activity and rapid action and also make it difficult for cancer cells to develop resistance against them. Hence, AMPs have become promising molecules for novel antibiotics and anticancer agents [23-25]. Despite the fact that ACPs are from different sources and have low homologies, they share some common characteris-tics such as a positive charge, an amphipathic conformation, the ability to fold into a secondary structure (α-helical or β -sheet) in a hydrophobic environment, and exhibition of broad spectrum and rapid anticancer activity [26-28]. In this re-view, we present an overview of the α-helical ACPs, includ-ing the distinction between cancer and normal cells, the proposed anticancer mechanism of α -helical ACPs, and the role of physicochemical parameters of α-helical ACPs on the biological activity and selectivity. In addition, we also dis-cuss the design and the modification method to optimize the cell selectivity of α-helical ACPs.
2. MECHANISM OF ACTION
The α -helical cationic ACPs are a major class of ACPs that generally exhibit the following properties: (1) they are rich in basic amino acids, such as lysine and arginine, result-ing in a net positive charge of peptides from +2 to +9 in neutral pH; (2) they are usually composed of 5-40 amino
Yuxin Chen

74 Mini-Reviews in Medicinal Chemistry, 2015, Vol. 15, No. 1 Huang et al.
acid residues and the percentage of hydrophobic residues is more than 30%; (3) they usually possess an amphipathic conformation with both polar and nonpolar face in hydro-phobic environments [29]. Although the exact mechanism of anticancer action of α-helical ACPs is not clear, it is believed that the interactions between α-helical ACPs and the mem-brane of cancer cells is the key step leading to cell lysis and cell death [17, 26]. In addition, 2 general effects of α-helical ACPs against cancer cells have been suggested such as the disruption of the cytoplasmic membrane via micellization or pore formation and induction of apoptosis [27].
2.1. Distinction in Cell Membranes
According to Hanahan and Weinberg, there are 6 princi-ple alterations in cell physiology that result in transformation of normal cells to cancer cells; these alterations are as fol-lows: (a) self-sufficiency in growth signals; (b) insensitivity to growth-inhibitory (antigrowth) signals; (c) evasion of programmed cell death (apoptosis); (d) limitless replicative potential; (e) unstained angiogenesis; and (f) tissue invasion and metastasis [30]. Compared with normal mammalian cells, the membrane components of cancer cells have several different properties that are as follows: (a) high-content of negatively charged components exposed on the outer membrane leaflet of cancer cells such as phosphatidylserine (3–9% of total phospholip-ids of membranes) [31-35], glycoproteins [36, 37], glycolip-ids [38], and proteoglycans [39]. This major difference could be used to improve the specificity of α-helical ACPs by electrostatic interactions; (b) increased membrane fluidity of cancer cells because of the difference in the cholesterol to phospholipid ratio which may enhance the lytic activity of ACPs by facilitating membrane destabilization [40-42]. However, it is not clear whether this alteration is uniform in all carcinomas; (c) cancer cells have a greater cell surface area because of microvilli which increases the opportunity of ACPs to bind to cancer cells [27, 43]. Therefore, electrostatic interactions between positively charged ACPs and negatively charged cell membrane components are believed to be the major reason in the selective killing of cancer cells by α-helical ACPs. Furthermore, the differences in membrane composition between cancer and normal cells provide targets for designing and developing novel ACP therapeutics with high-specificity.
2.2. Necrosis
As previously mentioned, the cancer cell membrane and bacterial membrane share several common features such as a negative charge and high-transmembrane potentials. The different properties of the normal cell membrane and cancer cell membrane provide the molecular basis for the high-specificity of α-helical ACPs against cancer cells. Most α-helical ACPs first target the highly negatively charged can-cer cell membrane by electrostatic interactions, then destabi-lize and disrupt the cell membrane by hydrophobic interac-tions leading to cancer cell necrosis, and exhibit a mecha-nism of action similar to α-helical AMPs such as the carpet-like mechanism as well as the barrel-stave model and toroi-dal pore model [22, 44-46]. Depending on the different lipid compositions of the prokaryotic cell membranes, AMPs
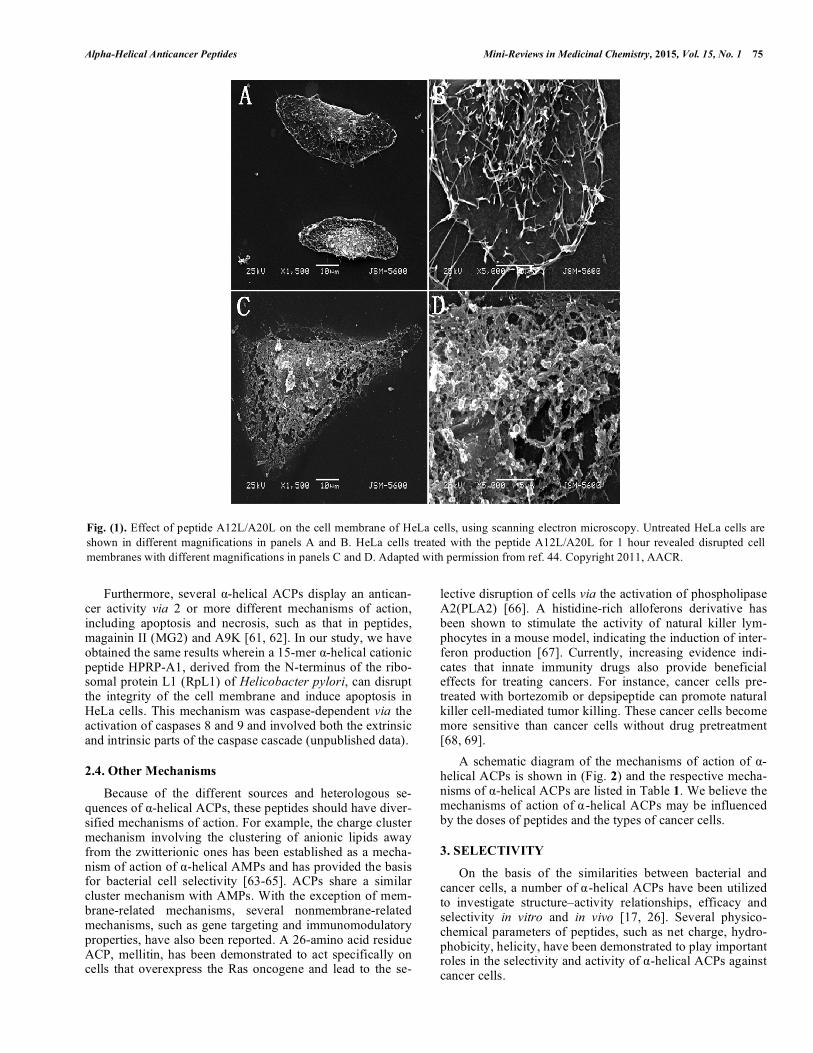
exhibit different mechanisms against the bacterial membrane [47, 48]. In addition, AMPs also exhibit selectivity against bacterial cells; this can be attributed to the presence of abun-dant acidic phospholipids and the absence of cholesterol in bacterial membranes because cholesterol increases the rigid-ity of lipid bilayers and suppresses the membrane disruption of AMPs, particularly for the simple, homogeneous lipid bilayer systems (nonraft domain) [49, 50]. However, cancer cells and normal cells are eukaryotic cells, containing choles-terol in their membrane. According to the aforementioned mechanisms of AMPs, there is a high-content of acidic phosphatidylserine and other negatively charged components in cancer cells which may attract cationic ACPs by electro-static interactions or form heterogeneous lipid bilayers ex-hibiting phase separation (or “lipid rafts”). This results in the susceptibility of the membrane for easy disruption by ACPs and the exhibition of high-selectivity of ACPs against cancer cells. However, this hypothesis needs to be studied further. In addition, ACPs may exhibit strong activity against multidrug resistant cancer cells because of the unique target-ing of the cell membrane by physical interactions without receptors. In the previous study, we found that a 26-residue α-helical ACP A12L/A20L exhibited broad spectrum antican-cer activity and a high-specificity against various cancer cells by a necrotic mechanism of action [44]. The scanning electron microscopy images of the HeLa cells treated with A12L/A20L are shown in (Fig. 1). It could be clearly seen that the peptide caused rapid cell membrane disruption with significant cavity formation and loss of microvilli and mem-brane integrity [51]. Similar results were obtained from other α-helical ACPs, such as polybia-MPI, isolated from the venom of the social wasp Polybia paulista [52], and tem-porin-1CEa, a 17-residue ACP isolated from the skin secre-tions of the Chinese brown frog (Rana chensinensis) [53].
2.3. Apoptosis
The apoptosis of cancer cells via mitochondrial mem-brane disruption by α-helical ACPs induction is another possible mechanism of action [54]. If internalized inside eukaryotic cells, α-helical ACPs can induce the permeation and swelling of mitochondria, resulting in the release of cytochrome c (Cyt c), which leads to apoptosis of cancer cells. The release of Cyt c from damaged mitochondria in-duces Apaf-1 oligomerization, caspase 9 activation and the subsequent conversion of pro-caspase 3 to caspase 3, which is responsible for many of the hallmarks of apoptotic symp-toms [55, 56]. A peptide obtained from Meretrix, a 15-mer, induces apoptosis and increases reactive oxygen species in the K562 cell cycle, destroying the electric potential on the surface of the membrane, breaks down microcracking [57]. Pardaxin exhibits the caspase-3/7-dependent and disrupts the mitochondrial membrane, causing the release of Cyt c and also inducing apoptosis in the HT-1080 cell line [58]. In the mitochondrial pathway, the induction of apoptosis in cancer cells can also be associated with the death receptor pathway [59]. Moreover, the synthetic tachyplesin conjugated to the integrin homing domain RGD has been demonstrated to induce cancer cell apoptosis by the 2 aforementioned path-ways [60].
Alpha-Helical Anticancer Peptides Mini-Reviews in Medicinal Chemistry, 2015, Vol. 15, No. 1 75
Furthermore, several α-helical ACPs display an antican-cer activity via 2 or more different mechanisms of action, including apoptosis and necrosis, such as that in peptides, magainin II (MG2) and A9K [61, 62]. In our study, we have obtained the same results wherein a 15-mer α-helical cationic peptide HPRP-A1, derived from the N-terminus of the ribo-somal protein L1 (RpL1) of Helicobacter pylori, can disrupt the integrity of the cell membrane and induce apoptosis in HeLa cells. This mechanism was caspase-dependent via the activation of caspases 8 and 9 and involved both the extrinsic and intrinsic parts of the caspase cascade (unpublished data).
2.4. Other Mechanisms
Because of the different sources and heterologous se-quences of α-helical ACPs, these peptides should have diver-sified mechanisms of action. For example, the charge cluster mechanism involving the clustering of anionic lipids away from the zwitterionic ones has been established as a mecha-nism of action of α-helical AMPs and has provided the basis for bacterial cell selectivity [63-65]. ACPs share a similar cluster mechanism with AMPs. With the exception of mem-brane-related mechanisms, several nonmembrane-related mechanisms, such as gene targeting and immunomodulatory properties, have also been reported. A 26-amino acid residue ACP, mellitin, has been demonstrated to act specifically on cells that overexpress the Ras oncogene and lead to the se-
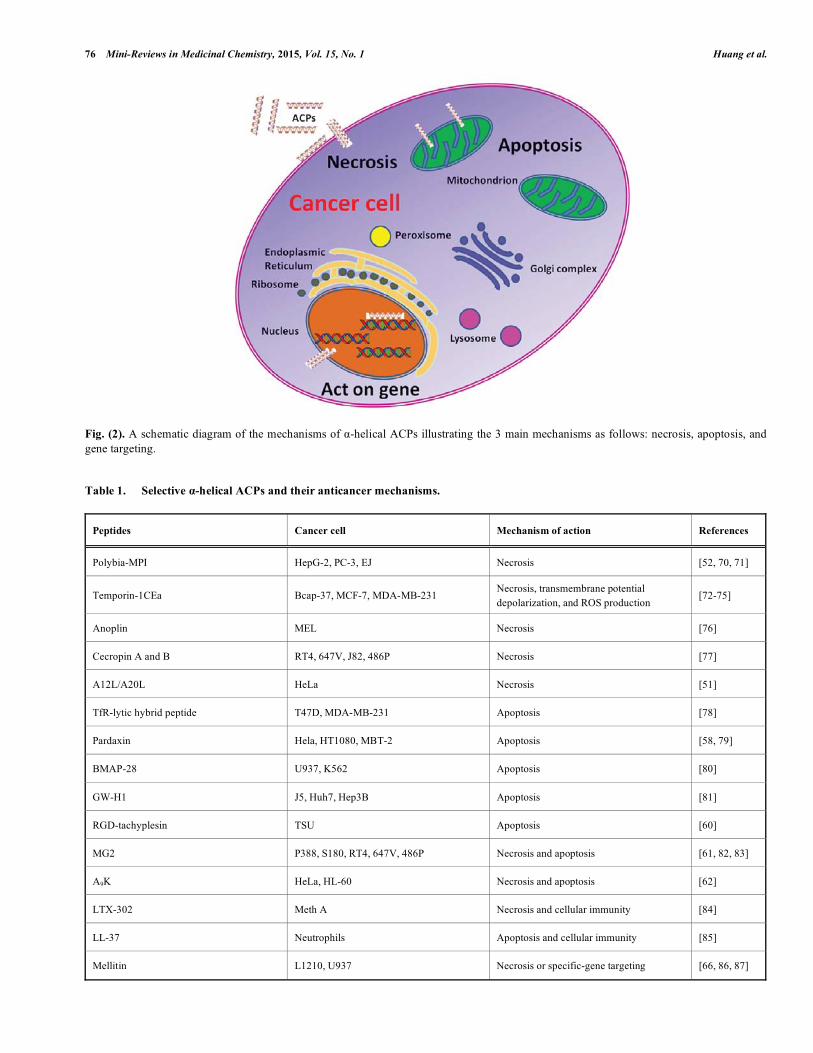
lective disruption of cells via the activation of phospholipase A2(PLA2) [66]. A histidine-rich alloferons derivative has been shown to stimulate the activity of natural killer lym-phocytes in a mouse model, indicating the induction of inter-feron production [67]. Currently, increasing evidence indi-cates that innate immunity drugs also provide beneficial effects for treating cancers. For instance, cancer cells pre-treated with bortezomib or depsipeptide can promote natural killer cell-mediated tumor killing. These cancer cells become more sensitive than cancer cells without drug pretreatment [68, 69]. A schematic diagram of the mechanisms of action of α-helical ACPs is shown in (Fig. 2) and the respective mecha-nisms of α-helical ACPs are listed in Table 1. We believe the mechanisms of action of α-helical ACPs may be influenced by the doses of peptides and the types of cancer cells.
3. SELECTIVITY
On the basis of the similarities between bacterial and cancer cells, a number of α-helical ACPs have been utilized to investigate structure–activity relationships, efficacy and selectivity in vitro and in vivo [17, 26]. Several physico-chemical parameters of peptides, such as net charge, hydro-phobicity, helicity, have been demonstrated to play important roles in the selectivity and activity of α-helical ACPs against cancer cells.
Fig. (1). Effect of peptide A12L/A20L on the cell membrane of HeLa cells, using scanning electron microscopy. Untreated HeLa cells are shown in different magnifications in panels A and B. HeLa cells treated with the peptide A12L/A20L for 1 hour revealed disrupted cell membranes with different magnifications in panels C and D. Adapted with permission from ref. 44. Copyright 2011, AACR.
76 Mini-Reviews in Medicinal Chemistry, 2015, Vol. 15, No. 1 Huang et al.
Fig. (2). A schematic diagram of the mechanisms of α-helical ACPs illustrating the 3 main mechanisms as follows: necrosis, apoptosis, and gene targeting.
Table 1. Selective α-helical ACPs and their anticancer mechanisms.
Peptides Cancer cell Mechanism of action References
Polybia-MPI HepG-2, PC-3, EJ Necrosis [52, 70, 71]
Temporin-1CEa Bcap-37, MCF-7, MDA-MB-231 Necrosis, transmembrane potential depolarization, and ROS production
[72-75]
Anoplin MEL Necrosis [76]
Cecropin A and B RT4, 647V, J82, 486P Necrosis [77]
A12L/A20L HeLa Necrosis [51]
TfR-lytic hybrid peptide T47D, MDA-MB-231 Apoptosis [78]
Pardaxin Hela, HT1080, MBT-2 Apoptosis [58, 79]
BMAP-28 U937, K562 Apoptosis [80]
GW-H1 J5, Huh7, Hep3B Apoptosis [81]
RGD-tachyplesin TSU Apoptosis [60]
MG2 P388, S180, RT4, 647V, 486P Necrosis and apoptosis [61, 82, 83]
A9K HeLa, HL-60 Necrosis and apoptosis [62]
LTX-302 Meth A Necrosis and cellular immunity [84]
LL-37 Neutrophils Apoptosis and cellular immunity [85]
Mellitin L1210, U937 Necrosis or specific-gene targeting [66, 86, 87]

Alpha-Helical Anticancer Peptides Mini-Reviews in Medicinal Chemistry, 2015, Vol. 15, No. 1 77
3.1. Net Charge
Most α-helical ACPs contain high-levels of basic amino acids, such as Lys and Arg, resulting in a high-net positive charge of peptides ranging from +2 to +9 [17, 88]. Based on the mechanism of action of α-helical ACPs, electrostatic interactions between cationic α-helical ACPs and negatively charged cell membrane components are believed to be the major force in driving the selective killing of cancer cells. Thus, the net charge of ACPs could represent a key role for anticancer activity and specificity of α-helical ACPs. In our previous study, the effects of net charge on anti-cancer activity and specificity of α-helical ACPs were tested using a 26-residue α-helical ACP, A12L/A20L [89]. The net charge of the peptide was also systematically altered by replacing serine residues with lysine residues to increase the net charge or by substituting lysine residues with serine residues to decrease the net charge on the polar face of the helix. The results indicated that net charge has 2 effects on the anticancer activity of the peptide: at a relatively lower level of net charge, an increase in the net charge of the pep-tide caused an improvement in anticancer activity until an optimal net charge was reached. In contrast, the anticancer activity of the peptide was weakened dramatically with fur-ther increase in the net charge beyond the optimum point which may be because of the strong electrostatic repulsion between lysine residues, resulting in a change in the structure and stability of peptides. However, the hemolytic activity of peptides remained at the same level, indicating that the net charge has no significant effect on normal cells. Lysine (K) and arginine (R) are 2 major positively charged hydrophilic amino acids with a side chain amine group and a guanidinium group, respectively. The pKa val-ues are 10.5 for lysine and 12–13.7 for arginine [90, 91]. Compared with lysine, arginine has a stronger ability to engage in electrostatic interactions and hydrogen bonding. Although the basic side chains of lysine and arginine have a high-affinity for the anionic membrane, the guanidinium group of arginine exhibits a higher affinity for the zwitteri-onic membrane leading to the toxicity against normal cells [92, 93]. In addition, the side chain of lysine is significantly more hydrophobic than that of arginine. The long nonpolar region of the side chain of lysine extends or snorkels into the hydrophobic core of the target membrane and increases the toxicity of α-helical ACPs against cancer cells [94, 95]. Thus, lysine is a more preferred basic amino acid for α-helical ACPs, and this finding is consistent with the previous results of the statistical study [88].
3.2. Hydrophobicity
For some α-helical ACPs, the cell membrane is the sole target of ACPs. According to the structure of the cell mem-brane, excluding the polar head of phospholipids, the nonpo-lar tail of phospholipids form a hydrophobic environment; thus, the hydrophobic interactions between α-helical ACPs and the hydrophobic core of the cell membrane should play the key role in the biological activity. It is generally known that amino acids with different side chains show different intrinsic hydrophobicity and helical propensity; thus, the amino acid substitutions in peptides not
only change the hydrophobicity of a peptide, but also change the helicity and self-association ability [51, 96-98]. In the previous study, a 26-residue α-helical peptide V13K (an amphipathic α-helical AMP [97, 99] was used to study the effects of peptide hydrophobicity on the mechanism of ac-tion of cationic ACPs by amino acid substitutions [51]. The results indicate that hydrophobicity plays an important role in the anticancer activity of α-helical ACPs, and the V13K analogs killed cancer cells via a fast, necrotic mechanism causing cell membrane lysis [51], as described in the “mem-brane discrimination” mechanism [99]. Increasing peptide hydrophobicity on the nonpolar face would enhance the helicity and self-association ability of peptides. Peptides with higher hydrophobicity may be able to penetrate deeper into the hydrophobic core of the cell membrane causing stronger activity for the formation of pores or channels in the cancer cell membrane, and higher hydrophobicity would always be accompanied with greater anticancer and hemolytic activity [51]. Besides substitutions with natural amino acids, substitu-tions with unnatural amino acids were also used to design and modify α-helical ACPs to increase anticancer activity and specificity [100, 101]. Modifications of C-terminal ami-dation and N-terminal acylation were also used to increase the activity, stability, and specificity of α-helical ACPs [102-104]. Furthermore, truncations of several amino acid resi-dues from the N-terminus or C-terminus of peptides were also useful in obtaining novel ACPs [105-108]. Thus, de novo design approaches, such as amino acid substitution and terminus modification, confirm their value in creating novel ACPs with high-specificity against cancer cells. Meanwhile, peptide length and hydrophobicity need to be carefully modi-fied because these parameters play crucial roles in anticancer activity and specificity [109].
3.3. Helicity
Besides hydrophobicity, the helicity of ACPs has also been demonstrated to play a crucial role in the biological activities of α-helical ACPs. Shai et al. designed and investi-gated D-amino acids which were to be incorporated into short model peptides (diastereomers), such as D-K4R2L9 and K6L9, containing varying ratios of D- and L-forms of leucine, lysine, and arginine. The results showed that the diastereo-mers did not retain the α-helical structure but reduced the toxicity against normal mammalian cells such as human erythrocytes. However, the diastereomers maintained both strong antimicrobial and anticancer activities and selectivity against bacterial and cancer cell membranes because of elec-trostatic interactions. In addition, the diastereomers also exhibited high-stability regarding enzymatic degradation by serum components [44, 110-113]. D-amino acids can be used to systematically modulate peptide helicity by replacing the original L-amino acids on either the nonpolar face or the polar face of the helix because of the helix-destabilizing properties without altering the intrinsic amino acid hydrophobicity of the peptide [96, 99, 114]. In the previous study, introduction of D-amino acids to replace the original L-amino acids on the nonpolar face or the polar face of a 26-residue α-helical ACP improved the therapeutic indices of peptide analogs against HeLa cells by

78 Mini-Reviews in Medicinal Chemistry, 2015, Vol. 15, No. 1 Huang et al.
9-fold and 22-fold, respectively [98]. The improvement in the therapeutic index could be attributed to the low-hemolytic activity and high-anticancer activity of peptide derivatives; however, when the number of D-amino acid substitutions on both the polar face and nonpolar face of peptides is more than 3, IC50 values of most peptide analogs increased dramatically (i.e., low-anticancer activity), result-ing in the decrease of specificity. The results indicated that peptide helicity correlated with both peptide hydrophobicity and the number of D-amino acid substitutions, and the helic-ity is important for the biological activity and specificity of α-helical ACPs. Enantiomeric α-helical ACPs also have been designed and investigated. The results showed that the L- and D-enantiomeric peptides exhibited similar biological activities, indicating that the mechanism of action of α-helical ACPs does not include receptors on the cell membrane. However, the D-enantiomers exhibited better potential for application because of its resistance to proteolysis [115, 116]; thus, de-signing and synthesizing D-enantiomers of α-helical ACPs would be a practical method for the systemic application of ACP drugs in clinical practices.
4. CONCLUSIONS AND OUTLOOKS
ACPs display unique mechanisms of action and several extraordinary properties such as broad spectrum activity and rapid action and also makes it difficult for the cancer cells to develop resistance against them. Therefore, these peptides have become promising molecules for novel anticancer agents, particularly for cancer cells with multidrug resis-tance. The cell membrane is a major target of most α-helical ACPs, and the inherent distinctions in lipid compositions between cancer cells and normal cells provide an opportunity to design or modify ACPs to improve the specificity against cancer cells. Electrostatic interactions between positively charged α-helical ACPs and negatively charged cell mem-brane components are believed to be the major force in kill-ing cancer cells. Different ACPs may use different mecha-nisms to kill cancer cells, which is also influenced by the dose of ACPs and the type of cancer cells. An increasing number of technologies have been used to study the mecha-nism of action of ACPs such as atomic force microscope [117], nuclear magnetic resonance spectroscopy [118], and attenuated total reflectance-fourier transform infrared spec-troscopy [119]. The development of peptide-based drugs has several obstacles to overcome such as the systemic toxicity, poor pharmacokinetics, and high-costs. Several methods have been attempted to solve these problems, for example, ligand modification by target molecules [120], RGD linkage [121, 122], or antibodies [123] to improve the specificity against cancer cells. Peptide self-assembly not only improves the stability of the peptide, but also affects the target delivery and release [124-128]. In addition, the design of ultra-short ACPs was a useful method to reduce the cost of industrial production [129]. Furthermore, the combination therapy of ACPs and traditional chemotherapy drugs may be an effi-cient way to treat cancer cells by reducing the toxicity and increasing the efficiency of drugs [130].
Peptide anticancer therapeutics is a promising field for reducing cancer problems. Although the development of peptide therapeutics has numerous challenges, the marketing approval of 6 peptides in 2012 has immensely encouraged the researchers [131]. The era of peptides as novel anticancer drugs will come soon when more peptide molecules, with an optimal therapeutic index, are used in clinical practices.
CONFLICT OF INTEREST
The authors confirm that there were no conflict of inter-est with regard to this article.
ACKNOWLEDGEMENTS
This work was supported by the National Natural Science Foundation of China (No. 81373445, Y.X.C., No. 21442001, Y.B.H.), the Innovative Team of Peptide Drugs of the Jilin Province (No. 20121807, Y.X.C.), and the Natural Science Foundation of Jilin Province (No. 20140101042JC, Y.B.H.).
REFERENCES [1] Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman,
D., Global cancer statistics. CA Cancer J Clin, 2011, 61(2), 69-90. [2] Beaglehole, R.; Bonita, R.; Magnusson, R., Global cancer prevention:
an important pathway to global health and development. Public Health, 2011, 125(12), 821-831.
[3] Lind, M.J., Principles of cytotoxic chemotherapy. Medicine, 2008, 36(1), 19-23.
[4] Shewach, D.S.; Kuchta, R.D., Introduction to cancer chemotherapeutics. Chem. Rev., 2009, 109(7), 2859-2861.
[5] Andersen, C.; Adamsen, L.; Moeller, T.; Midtgaard, J.; Quist, M.; Tveteraas, A.; Rorth, M., The effect of a multidimensional exercise programme on symptoms and side-effects in cancer patients undergoing chemotherapy--the use of semi-structured diaries. Eur. J. Oncol. Nurs., 2006, 10(4), 247-262.
[6] Breidenbach, M.; Rein, D.T.; Schondorf, T.; Schmidt, T.; Konig, E.; Valter, M.; Kurbacher, C.M., Hematological side-effect profiles of individualized chemotherapy regimen for recurrent ovarian cancer. Anticancer Drugs, 2003, 14(5), 341-346.
[7] Di Franco, R.; Sammarco, E.; Calvanese, M.G.; De Natale, F.; Falivene, S.; Di Lecce, A.; Giugliano, F.M.; Murino, P.; Manzo, R.; Cappabianca, S.; Muto, P.; Ravo, V., Preventing the acute skin side effects in patients treated with radiotherapy for breast cancer: the use of corneometry in order to evaluate the protective effect of moisturizing creams. Radiat. Oncol., 2013, 8, 57.
[8] Gottesman, M.M., Mechanisms of cancer drug resistance. Annu. Rev. Med., 2002, 53, 615-627.
[9] Dennison, S.R.; Wallace, J.; Harris, F.; Phoenix, D.A., Amphiphilic alpha-helical antimicrobial peptides and their structure/function relationships. Protein Peptide Lett., 2005, 12(1), 31-39.
[10] Zasloff, M., Antimicrobial peptides of multicellular organisms. Nature, 2002, 415(6870), 389-395.
[11] Giuliani, A.; Pirri, G.; Nicoletto, S.F., Antimicrobial peptides: an overview of a promising class of therapeutics. Cent. Eur. J. Biol., 2007, 2(1), 1-33.
[12] Toke, O., Antimicrobial peptides: new candidates in the fight against bacterial infections. Biopolymers, 2005, 80(6), 717-735.
[13] Reddy, K.V.R.; Yedery, R.D.; Aranha, C., Antimicrobial peptides: premises and promises. Int. J. Antimicrob. Ag., 2004, 24(6), 536-547.
[14] Ramamoorthy, A., Beyond NMR spectra of antimicrobial peptides: Dynamical images at atomic resolution and functional insights. Solid State Nucl. Mag., 2009, 35(4), 201-207.
[15] Gottler, L.M.; Ramamoorthy, A., Structure, membrane orientation, mechanism, and function of pexiganan - A highly potent antimicrobial peptide designed from magainin. BBA-Biomembranes, 2009, 1788(8), 1680-1686.
[16] Dhople, V.; Krukemeyer, A.; Ramamoorthy, A., The human beta-defensin-3, an antibacterial peptide with multiple biological functions. BBA-Biomembranes, 2006, 1758(9), 1499-1512.

Alpha-Helical Anticancer Peptides Mini-Reviews in Medicinal Chemistry, 2015, Vol. 15, No. 1 79
[17] Hoskin, D.W.; Ramamoorthy, A., Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta., 2008, 1778(2), 357-375.
[18] Marr, A.K.; Gooderham, W.J.; Hancock, R.E.W., Antibacterial peptides for therapeutic use: obstacles and realistic outlook. Curr. Opin .Pharmacol., 2006, 6(5), 468-472.
[19] Boman, H.G., Antibacterial peptides: key components needed in immunity. Cell, 1991, 65(2), 205-207.
[20] Fox, J.L., Antimicrobial peptides stage a comeback. Nat Biotechnol, 2013, 31(5), 379-382.
[21] Pinheiro da Silva, F.; Machado, M.C., Antimicrobial peptides: clinical relevance and therapeutic implications. Peptides, 2012, 36(2), 308-314.
[22] Huang, Y.; Huang, J.; Chen, Y., Alpha-helical cationic antimicrobial peptides: relationships of structure and function. Protein Cell, 2010, 1(2), 143-152.
[23] Schweizer, F., Cationic amphiphilic peptides with cancer-selective toxicity. Eur J Pharmacol, 2009, 625(1-3), 190-194.
[24] Gaspar, D.; Veiga, A.S.; Castanho, M.A., From antimicrobial to anticancer peptides. A review. Front Microbiol, 2013, 4, 294.
[25] Riedl, S.; Zweytick, D.; Lohner, K., Membrane-active host defense peptides--challenges and perspectives for the development of novel anticancer drugs. Chem Phys Lipids, 2011, 164(8), 766-781.
[26] Dennison, S.R.; Whittaker, M.; Harris, F.; Phoenix, D.A., Anticancer alpha-helical peptides and structure/function relationships underpinning their interactions with tumour cell membranes. Curr Protein Pept Sci, 2006, 7(6), 487-499.
[27] Papo, N.; Shai, Y., Host defense peptides as new weapons in cancer treatment. Cell Mol Life Sci, 2005, 62(7-8), 784-790.
[28] Zelezetsky, I.; Tossi, A., Alpha-helical antimicrobial peptides - Using a sequence template to guide structure-activity relationship studies. BBA-Biomembranes, 2006, 1758(9), 1436-1449.
[29] Liberio, M.S.; Joanitti, G.A.; Fontes, W.; Castro, M.S., Anticancer Peptides and Proteins: A Panoramic View. Protein Peptide Lett, 2013, 20(4), 380-391.
[30] Hanahan, D.; Weinberg, R.A., The hallmarks of cancer. Cell, 2000, 100(1), 57-70.
[31] Ran, S.; Downes, A.; Thorpe, P.E., Increased exposure of anionic phospholipids on the surface of tumor blood vessels. Cancer Res, 2002, 62(21), 6132-6140.
[32] Utsugi, T.; Schroit, A.J.; Connor, J.; Bucana, C.D.; Fidler, I.J., Elevated expression of phosphatidylserine in the outer membrane leaflet of human tumor cells and recognition by activated human blood monocytes. Cancer Res, 1991, 51(11), 3062-3066.
[33] Rao, L.V.; Tait, J.F.; Hoang, A.D., Binding of annexin V to a human ovarian carcinoma cell line (OC-2008). Contrasting effects on cell surface factor VIIa/tissue factor activity and prothrombinase activity. Thromb Res, 1992, 67(5), 517-531.
[34] Kirszberg, C.; Lima, L.G.; de Oliveira, A.D.; Pickering, W.; Gray, E.; Barrowcliffe, T.W.; Rumjanek, V.M.; Monteiro, R.Q., Simultaneous tissue factor expression and phosphatidylserine exposure account for the highly procoagulant pattern of melanoma cell lines. Melanoma Research, 2009, 19(5), 301-308.
[35] Schroder-Borm, H.; Bakalova, R.; Andra, J., The NK-lysin derived peptide NK-2 preferentially kills cancer cells with increased surface levels of negatively charged phosphatidylserine. FEBS Letters, 2005, 579(27), 6128-6134.
[36] Yoon, W.H.; Park, H.D.; Lim, K.; Hwang, B.D., Effect of O-glycosylated mucin on invasion and metastasis of HM7 human colon cancer cells. Biochem Biophys Res Commun, 1996, 222(3), 694-699.
[37] Kufe, D.W., Mucins in cancer: function, prognosis and therapy. Nature Reviews Cancer, 2009, 9(12), 874-885.
[38] Lopez, P.H.H.; Schnaar, R.L., Gangliosides in cell recognition and membrane protein regulation. Curr Opin Struc Biol, 2009, 19(5), 549-557.
[39] Vynios, D.H.; Papageorgakopoulou, N.; Sazakli, H.; Tsiganos, C.P., The interactions of cartilage proteoglycans with collagens are determined by their structures. Biochimie, 2001, 83(9), 899-906.
[40] Sok, M.; Sentjurc, M.; Schara, M., Membrane fluidity characteristics of human lung cancer. Cancer Lett, 1999, 139(2), 215-220.
[41] Nakazawa, I.; Iwaizumi, M., A role of the cancer cell membrane fluidity in the cancer metastases: an ESR study. Tohoku J Exp Med, 1989, 157(3), 193-198.
[42] Schroeder, F.; Gardiner, J.M., Membrane lipids and enzymes of cultured high- and low-metastatic B16 melanoma variants. Cancer Res, 1984, 44(8), 3262-3269.
[43] Chaudhary, J.; Munshi, M., Scanning electron microscopic analysis of breast aspirates. Cytopathology, 1995, 6(3), 162-167.
[44] Shai, Y., Mechanism of the binding, insertion and destabilization of phospholipid bilayer membranes by alpha-helical antimicrobial and cell non-selective membrane-lytic peptides. Biochim Biophys Acta, 1999, 1462(1-2), 55-70.
[45] Oren, Z.; Shai, Y., Mode of action of linear amphipathic alpha-helical antimicrobial peptides. Biopolymers, 1998, 47(6), 451-463.
[46] Nguyen, L.T.; Haney, E.F.; Vogel, H.J., The expanding scope of antimicrobial peptide structures and their modes of action. Trends in Biotechnology, 2011, 29(9), 464-472.
[47] Lee, D.K.; Brender, J.R.; Sciacca, M.F.M.; Krishnamoorthy, J.; Yu, C.S.; Ramamoorthy, A., Lipid Composition-Dependent Membrane Fragmentation and Pore-Forming Mechanisms of Membrane Disruption by Pexiganan (MSI-78). Biochemistry, 2013, 52(19), 3254-3263.
[48] Hallock, K.J.; Lee, D.K.; Ramamoorthy, A., MSI-78, an analogue of the magainin antimicrobial peptides, disrupts lipid bilayer structure via positive curvature strain. Biophysical Journal, 2003, 84(5), 3052-3060.
[49] McHenry, A.J.; Sciacca, M.F.M.; Brender, J.R.; Ramamoorthy, A., Does cholesterol suppress the antimicrobial peptide induced disruption of lipid raft containing membranes? BBA-Biomembranes, 2012, 1818(12), 3019-3024.
[50] Ramamoorthy, A.; Lee, D.K.; Narasimhaswamy, T.; Nanga, R.P.R., Cholesterol reduces pardaxin's dynamics-a barrel-stave mechanism of membrane disruption investigated by solid-state NMR. BBA-Biomembranes, 2010, 1798(2), 223-227.
[51] Huang, Y.B.; Wang, X.F.; Wang, H.Y.; Liu, Y.; Chen, Y., Studies on mechanism of action of anticancer peptides by modulation of hydrophobicity within a defined structural framework. Mol Cancer Ther, 2011, 10(3), 416-426.
[52] Zhang, W.; Li, J.; Liu, L.W.; Wang, K.R.; Song, J.J.; Yan, J.X.; Li, Z.Y.; Zhang, B.Z.; Wang, R., A novel analog of antimicrobial peptide Polybia-MPI, with thioamide bond substitution, exhibits increased therapeutic efficacy against cancer and diminished toxicity in mice. Peptides, 2010, 31(10), 1832-1838.
[53] Wang, C.; Zhou, Y.; Li, S.; Li, H.B.; Tian, L.L.; Wang, H.; Shang, D.J., Anticancer mechanisms of temporin-1CEa, an amphipathic alpha-helical antimicrobial peptide, in Bcap-37 human breast cancer cells. Life Sciences, 2013, 92(20-21), 1004-1014.
[54] Mai, J.C.; Mi, Z.B.; Kim, S.H.; Ng, B.; Robbins, P.D., A proapoptotic peptide for the treatment of solid tumors. Cancer Research, 2001, 61(21), 7709-7712.
[55] Li, H.; Kolluri, S.K.; Gu, J.; Dawson, M.I.; Cao, X.H.; Hobbs, P.D.; Lin, B.Z.; Chen, G.Q.; Lu, L.S.; Lin, F.; Xie, Z.H.; Fontana, J.A.; Reed, J.C.; Zhang, X.K., Cytochrome c release and apoptosis induced by mitochondrial targeting of nuclear orphan receptor TR3. Science, 2000, 289(5482), 1159-1164.
[56] Cory, S.; Adams, J.M., Matters of life and death: programmed cell death at Cold Spring Harbor. Biochim Biophys Acta, 1998, 1377(2), R25-44.
[57] Liu, M.; Zhao, X.Z.; Zhao, J.; Xiao, L.; Liu, H.Z.; Wang, C.C.; Cheng, L.Y.; Wu, N.; Lin, X.K., Induction of Apoptosis, G(0)/G(1) Phase Arrest and Microtubule Disassembly in K562 Leukemia Cells by Mere15, a Novel Polypeptide from Meretrix meretrix Linnaeus. Mar Drugs, 2012, 10(11), 2596-2607.
[58] Huang, T.C.; Lee, J.F.; Chen, J.Y., Pardaxin, an antimicrobial peptide, triggers caspase-dependent and ROS-mediated apoptosis in HT-1080 cells. Mar Drugs, 2011, 9(10), 1995-2009.
[59] Kaufmann, S.H.; Earnshaw, W.C., Induction of apoptosis by cancer chemotherapy. Exp Cell Res, 2000, 256(1), 42-49.
[60] Chen, Y.; Xu, X.; Hong, S.; Chen, J.; Liu, N.; Underhill, C.B.; Creswell, K.; Zhang, L., RGD-Tachyplesin inhibits tumor growth. Cancer Res, 2001, 61(6), 2434-2438.
[61] Liu, S.; Yang, H.; Wan, L.; Cheng, J.Q.; Lu, X.F., Penetratin-Mediated Delivery Enhances the Antitumor Activity of the Cationic Antimicrobial Peptide Magainin II. Cancer Biother Radio, 2013, 28(4), 289-297.
[62] Xu, H.; Chen, C.X.; Hu, J.; Zhou, P.; Zeng, P.; Cao, C.H.; Lu, J.R., Dual modes of antitumor action of an amphiphilic peptide A(9)K. Biomaterials, 2013, 34(11), 2731-2737.

80 Mini-Reviews in Medicinal Chemistry, 2015, Vol. 15, No. 1 Huang et al.
[63] Epand, R.F.; Maloy, L.; Ramamoorthy, A.; Epand, R.M., Amphipathic Helical Cationic Antimicrobial Peptides Promote Rapid Formation of Crystalline States in the Presence of Phosphatidylglycerol: Lipid Clustering in Anionic Membranes. Biophysical Journal, 2010, 98(11), 2564-2573.
[64] Epand, R.F.; Maloy, W.L.; Ramamoorthy, A.; Epand, R.M., Probing the "charge cluster mechanism" in amphipathic helical cationic antimicrobial peptides. Biochemistry, 2010, 49(19), 4076-4084.
[65] Thennarasu, S.; Huang, R.; Lee, D.K.; Yang, P.; Maloy, L.; Chen, Z.; Ramamoorthy, A., Limiting an antimicrobial peptide to the lipid-water interface enhances its bacterial membrane selectivity: a case study of MSI-367. Biochemistry, 2010, 49(50), 10595-10605.
[66] Sharma, S.V., Melittin-induced hyperactivation of phospholipase A2 activity and calcium influx in ras-transformed cells. Oncogene, 1993, 8(4), 939-947.
[67] Chernysh, S.; Kim, S.I.; Bekker, G.; Pleskach, V.A.; Filatova, N.A.; Anikin, V.B.; Platonov, V.G.; Bulet, P., Antiviral and antitumor peptides from insects. Proc Natl Acad Sci USA, 2002, 99(20), 12628-12632.
[68] Lundqvist, A.; Abrams, S.I.; Schrump, D.S.; Alvarez, G.; Suffredini, D.; Berg, M.; Childs, R., Bortezomib and depsipeptide sensitize tumors to tumor necrosis factor-related apoptosis-inducing ligand: A novel method to potentiate natural killer cell tumor cytotoxicity. Cancer Research, 2006, 66(14), 7317-7325.
[69] Krieg, S.; Ullrich, E., Novel immune modulators used in hematology: impact on NK cells. Front Immunol, 2012, 3, 388.
[70] Wang, K.R.; Yan, J.X.; Zhang, B.Z.; Song, J.J.; Jia, P.F.; Wang, R., Novel mode of action of polybia-MPI, a novel antimicrobial peptide, in multi-drug resistant leukemic cells. Cancer Lett, 2009, 278(1), 65-72.
[71] Wang, K.R.; Zhang, B.Z.; Zhang, W.; Yan, J.X.; Li, J.; Wang, R., Antitumor effects, cell selectivity and structure-activity relationship of a novel antimicrobial peptide polybia-MPI. Peptides, 2008, 29(6), 963-968.
[72] Wang, C.; Zhou, Y.; Li, S.; Li, H.; Tian, L.; Wang, H.; Shang, D., Anticancer mechanisms of temporin-1CEa, an amphipathic alpha-helical antimicrobial peptide, in Bcap-37 human breast cancer cells. Life Sci, 2013, 92(20-21), 1004-1014.
[73] Wang, C.; Li, H.B.; Li, S.; Tian, L.L.; Shang, D.J., Antitumor effects and cell selectivity of temporin-1CEa, an antimicrobial peptide from the skin secretions of the Chinese brown frog (Rana chensinensis). Biochimie, 2012, 94(2), 434-441.
[74] Wang, C.; Tian, L.L.; Li, S.; Li, H.B.; Zhou, Y.; Wang, H.; Yang, Q.Z.; Ma, L.J.; Shang, D.J., Rapid cytotoxicity of antimicrobial peptide tempoprin-1CEa in breast cancer cells through membrane destruction and intracellular calcium mechanism. PLoS One, 2013, 8(4), e60462.
[75] Yang, Q.Z.; Wang, C.; Lang, L.; Zhou, Y.; Wang, H.; Shang, D.J., Design of potent, non-toxic anticancer peptides based on the structure of the antimicrobial peptide, temporin-1CEa. Arch Pharm Res, 2013, 36(11), 1302-1310.
[76] Zhu, L.N.; Fu, C.Y.; Zhang, S.F.; Chen, W.; Jin, Y.T.; Zhao, F.K., Novel cytotoxic exhibition mode of antimicrobial peptide anoplin in MEL cells, the cell line of murine Friend leukemia virus-induced leukemic cells. J Pept Sci, 2013, 19(9), 566-574.
[77] Suttmann, H.; Retz, M.; Paulsen, F.; Harder, J.; Zwergel, U.; Kamradt, J.; Wullich, B.; Unteregger, G.; Stockle, M.; Lehmann, J., Antimicrobial peptides of the Cecropin-family show potent antitumor activity against bladder cancer cells. BMC Urol, 2008, 8, 5.
[78] Kawamoto, M.; Horibe, T.; Kohno, M.; Kawakami, K., A novel transferrin receptor-targeted hybrid peptide disintegrates cancer cell membrane to induce rapid killing of cancer cells. BMC Cancer, 2011, 11, 359.
[79] Hsu, J.C.; Lin, L.C.; Tzen, J.T.C.; Chen, J.Y., Pardaxin-induced apoptosis enhances antitumor activity in HeLa cells. Peptides, 2011, 32(6), 1110-1116.
[80] Risso, A.; Braidot, E.; Sordano, M.C.; Vianello, A.; Macri, F.; Skerlavaj, B.; Zanetti, M.; Gennaro, R.; Bernardi, P., BMAP-28, an antibiotic peptide of innate immunity, induces cell death through opening of the mitochondrial permeability transition pore. Mol Cell Biol, 2002, 22(6), 1926-1935.
[81] Chen, Y.L.S.; Li, J.H.; Yu, C.Y.; Lin, C.J.; Chiu, P.H.; Chen, P.W.; Lin, C.C.; Chen, W.J., Novel cationic antimicrobial peptide GW-
H1 induced caspase-dependent apoptosis of hepatocellular carcinoma cell lines. Peptides, 2012, 36(2), 257-265.
[82] Baker, M.A.; Maloy, W.L.; Zasloff, M.; Jacob, L.S., Anticancer efficacy of Magainin2 and analogue peptides. Cancer Res, 1993, 53(13), 3052-3057.
[83] Lehmann, J.; Retz, M.; Sidhu, S.S.; Suttmann, H.; Sell, M.; Paulsen, F.; Harder, J.; Unteregger, G.; Stockle, M., Antitumor activity of the antimicrobial peptide magainin II against bladder cancer cell lines. Eur Urol, 2006, 50(1), 141-147.
[84] Berge, G.; Eliassen, L.T.; Camilio, K.A.; Bartnes, K.; Sveinbjornsson, B.; Rekdal, O., Therapeutic vaccination against a murine lymphoma by intratumoral injection of a cationic anticancer peptide. Cancer Immunol Immun, 2010, 59(8), 1285-1294.
[85] Nagaoka, I.; Suzuki, K.; Niyonsaba, F.; Tamura, H.; Hirata, M., Modulation of neutrophil apoptosis by antimicrobial peptides. ISRN Microbiol, 2012, 2012, 345791.
[86] Killion, J.J.; Dunn, J.D., Differential cytolysis of murine spleen, bone-marrow and leukemia cells by melittin reveals differences in membrane topography. Biochem Biophys Res Commun, 1986, 139(1), 222-227.
[87] Sui, S.F.; Wu, H.; Guo, Y.; Chen, K.S., Conformational changes of melittin upon insertion into phospholipid monolayer and vesicle. J Biochem, 1994, 116(3), 482-487.
[88] Dennison, S.R.; Whittaker, M.; Harris, F.; Phoenix, D.A., Anticancer alpha-helical peptides and structure/function relationships underpinning their interactions with tumour cell membranes. Curr Protein Pept Sc, 2006, 7(6), 487-499.
[89] Huang, Y.B.; Zhai, N.C.; Gao, G.; Chen, Y.X., Effects of Net Charge on Biological Activities of Alpha-helical Anticancer Peptides. Chem J Chinese U, 2012, 33(6), 1252-1258.
[90] Harms, M.J.; Schlessman, J.L.; Chimenti, M.S.; Sue, G.R.; Damjanovic, A.; Garcia-Moreno, B., A buried lysine that titrates with a normal pK(a): Role of conformational flexibility at the protein-water interface as a determinant of pK(a)values. Protein Science, 2008, 17(5), 833-845.
[91] Li, L.B.; Vorobyov, I.; Allen, T.W., The Different Interactions of Lysine and Arginine Side Chains with Lipid Membranes. J Phys Chem B, 2013, 117(40), 11906-11920.
[92] Harris, F.; Dennison, S.R.; Singh, J.; Phoenix, D.A., On the selectivity and efficacy of defense peptides with respect to cancer cells. Med Res Rev, 2013, 33(1), 190-234.
[93] Fromm, J.R.; Hileman, R.E.; Caldwell, E.E.; Weiler, J.M.; Linhardt, R.J., Differences in the interaction of heparin with arginine and lysine and the importance of these basic amino acids in the binding of heparin to acidic fibroblast growth factor. Arch Biochem Biophys, 1995, 323(2), 279-287.
[94] Johansson, A.C.; Lindahl, E., Position-resolved free energy of solvation for amino acids in lipid membranes from molecular dynamics simulations. Proteins, 2008, 70(4), 1332-1344.
[95] Dennison, S.R.; Dante, S.; Hauss, T.; Brandenburg, K.; Harris, F.; Phoenix, D.A., Investigations into the membrane interactions of m-calpain domain V. Biophys J, 2005, 88(4), 3008-3017.
[96] Chen, Y.; Mant, C.T.; Hodges, R.S., Determination of stereochemistry stability coefficients of amino acid side-chains in an amphipathic alpha-helix. J Pept Res, 2002, 59(1), 18-33.
[97] Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S., Role of peptide hydrophobicity in the mechanism of action of alpha-helical antimicrobial peptides. Antimicrob Agents Chemother, 2007, 51(4), 1398-1406.
[98] Huang, Y.B.; He, L.Y.; Jiang, H.Y.; Chen, Y.X., Role of helicity on the anticancer mechanism of action of cationic-helical peptides. Int J Mol Sci, 2012, 13(6), 6849-6862.
[99] Chen, Y.; Mant, C.T.; Farmer, S.W.; Hancock, R.E.; Vasil, M.L.; Hodges, R.S., Rational design of alpha-helical antimicrobial peptides with enhanced activities and specificity/therapeutic index. J Biol Chem, 2005, 280(13), 12316-12329.
[100] Staykova, S.; Naydenova, E.; Wesselinova, D.; Kalistratova, A.; Vezenkov, L., Synthesis and in vitro study of the anticancer activity of new analogs of octreotide. Prot Pept Lett, 2012, 19(12), 1257-1262.
[101] Khoury, G.A.; Smadbeck, J.; Tamamis, P.; Vandris, A.C.; Kieslich, C.A.; Floudas, C.A., Forcefield_NCAA: Ab Initio Charge Parameters to Aid in the Discovery and Design of Therapeutic Proteins and Peptides with Unnatural Amino Acids and Their Application to Complement Inhibitors of the Compstatin Family. ACS Synth Biol, 2014.

Alpha-Helical Anticancer Peptides Mini-Reviews in Medicinal Chemistry, 2015, Vol. 15, No. 1 81
[102] Copier, J.; Dalgleish, A.G.; Britten, C.M.; Finke, L.H.; Gaudernack, G.; Gnjatic, S.; Kallen, K.; Kiessling, R.; Schuessler-Lenz, M.; Singh, H.; Talmadge, J.; Zwierzina, H.; Hakansson, L., Improving the efficacy of cancer immunotherapy. Eur J Cancer, 2009, 45(8), 1424-1431.
[103] Wang, G.; Li, X.; Wang, Z., APD2: the updated antimicrobial peptide database and its application in peptide design. Nucleic Acids Res, 2009, 37(Database issue), D933-937.
[104] Dennison, S.R.; Harris, F.; Bhatt, T.; Singh, J.; Phoenix, D.A., The effect of C-terminal amidation on the efficacy and selectivity of antimicrobial and anticancer peptides. Mol Cell Biochem, 2009, 332(1-2), 43-50.
[105] Chen, H.C.; Brown, J.H.; Morell, J.L.; Huang, C.M., Synthetic magainin analogues with improved antimicrobial activity. FEBS Lett, 1988, 236(2), 462-466.
[106] Kim, H.S.; Yoon, H.; Minn, I.; Park, C.B.; Lee, W.T.; Zasloff, M.; Kim, S.C., Pepsin-mediated processing of the cytoplasmic histone H2A to strong antimicrobial peptide buforin I. J Immunol, 2000, 165(6), 3268-3274.
[107] Yan, J.X.; Wang, K.R.; Chen, R.; Song, J.J.; Zhang, B.Z.; Dang, W.; Zhang, W.; Wang, R., Membrane active antitumor activity of NK-18, a mammalian NK-lysin-derived cationic antimicrobial peptide. Biochimie, 2012, 94(1), 184-191.
[108] Li, X.; Li, Y.; Han, H.; Miller, D.W.; Wang, G., Solution structures of human LL-37 fragments and NMR-based identification of a minimal membrane-targeting antimicrobial and anticancer region. J Am Chem Soc, 2006, 128(17), 5776-5785.
[109] Zhao, L.J.; Huang, Y.B.; Gao, S.; Cui, Y.; He, D.; Wang, L.; Chen, Y.X., Comparison on effect of hydrophobicity on the antibacterial and antifungal activities of alpha-helical antimicrobial peptides. Sci China Chem, 2013, 56(9), 1307-1314.
[110] Papo, N.; Shahar, M.; Eisenbach, L.; Shai, Y., A novel lytic peptide composed of DL-amino acids selectively kills cancer cells in culture and in mice. J Biol Chem, 2003, 278(23), 21018-21023.
[111] Papo, N.; Shai, Y., New lytic peptides based on the D,L-amphipathic helix motif preferentially kill tumor cells compared to normal cells. Biochemistry, 2003, 42(31), 9346-9354.
[112] Mai, J.C.; Mi, Z.; Kim, S.H.; Ng, B.; Robbins, P.D., A proapoptotic peptide for the treatment of solid tumors. Cancer Res, 2001, 61(21), 7709-7712.
[113] Papo, N.; Oren, Z.; Pag, U.; Sahl, H.G.; Shai, Y., The consequence of sequence alteration of an amphipathic alpha-helical antimicrobial peptide and its diastereomers. J Biol Chem, 2002, 277(37), 33913-33921.
[114] Krause, E.; Beyermann, M.; Dathe, M.; Rothemund, S.; Bienert, M., Location of an amphipathic alpha-helix in peptides using reversed-phase HPLC retention behavior of D-amino acid analogs. Anal Chem, 1995, 67(2), 252-258.
[115] Huang, J.; Hao, D.; Chen, Y.; Xu, Y.; Tan, J.; Huang, Y.; Li, F., Inhibitory effects and mechanisms of physiological conditions on the activity of enantiomeric forms of an alpha-helical antibacterial peptide against bacteria. Peptides, 2011, 32(7), 1488-1495.
[116] Chen, Y.; Vasil, A.I.; Rehaume, L.; Mant, C.T.; Burns, J.L.; Vasil, M.L.; Hancock, R.E.; Hodges, R.S., Comparison of biophysical and biologic properties of alpha-helical enantiomeric antimicrobial peptides. Chem Biol Drug Des, 2006, 67(2), 162-173.
[117] Shan, Y.; Huang, J.; Tan, J.; Gao, G.; Liu, S.; Wang, H.; Chen, Y., The study of single anticancer peptides interacting with HeLa cell membranes by single molecule force spectroscopy. Nanoscale, 2012, 4(4), 1283-1286.
[118] Su, Y.; Waring, A.J.; Ruchala, P.; Hong, M., Membrane-bound dynamic structure of an arginine-rich cell-penetrating peptide, the protein transduction domain of HIV TAT, from solid-state NMR. Biochemistry, 2010, 49(29), 6009-6020.
[119] Arkin, I., Special Issue FTIR in membrane proteins and peptide studies Preface. BBA-Biomembranes, 2013, 1828(10), 2255-2255.
[120] Lu, Y.; Yang, J.; Sega, E., Issues related to targeted delivery of proteins and peptides. AAPS J, 2006, 8(3), E466-478.
[121] Arap, W.; Pasqualini, R.; Ruoslahti, E., Cancer treatment by targeted drug delivery to tumor vasculature in a mouse model. Science, 1998, 279(5349), 377-380.
[122] Millard, M.; Odde, S.; Neamati, N., Integrin targeted therapeutics. Theranostics, 2011, 1, 154-188.
[123] Alley, S.C.; Okeley, N.M.; Senter, P.D., Antibody-drug conjugates: targeted drug delivery for cancer. Curr Opin Chem Biol, 2010, 14(4), 529-537.
[124] Zha, R.H.; Sur, S.; Stupp, S.I., Self-assembly of Cytotoxic Peptide Amphiphiles into Supramolecular Membranes for Cancer Therapy. Adv Healthc Mater, 2013, 2(1), 126-133.
[125] Chang, D.K.; Lin, C.T.; Wu, C.H.; Wu, H.C., A novel peptide enhances therapeutic efficacy of liposomal anti-cancer drugs in mice models of human lung cancer. PLoS One, 2009, 4(1), e4171.
[126] Standley, S.M.; Toft, D.J.; Cheng, H.; Soukasene, S.; Chen, J.; Raja, S.M.; Band, V.; Band, H.; Cryns, V.L.; Stupp, S.I., Induction of cancer cell death by self-assembling nanostructures incorporating a cytotoxic peptide. Cancer Res, 2010, 70(8), 3020-3026.
[127] Sadatmousavi, P.; Soltani, M.; Nazarian, R.; Jafari, M.; Chen, P., Self-Assembling Peptides: Potential Role in Tumor Targeting. Curr Pharm Biotechno, 2011, 12(8), 1089-1100.
[128] Hartgerink, J.D.; Beniash, E.; Stupp, S.I., Self-assembly and mineralization of peptide-amphiphile nanofibers. Science, 2001, 294(5547), 1684-1688.
[129] Yang, C.Y.; Dantzig, A.H.; Pidgeon, C., Intestinal peptide transport systems and oral drug availability. Pharm Res, 1999, 16(9), 1331-1343.
[130] Johnstone, S.A.; Gelmon, K.; Mayer, L.D.; Hancock, R.E.; Bally, M.B., In vitro characterization of the anticancer activity of membrane-active cationic peptides. I. Peptide-mediated cytotoxicity and peptide-enhanced cytotoxic activity of doxorubicin against wild-type and p-glycoprotein over-expressing tumor cell lines. Anticancer Drug Des, 2000, 15(2), 151-160.
[131] Kaspar, A.A.; Reichert, J.M., Future directions for peptide therapeutics development. Drug Discov. Today, 2013, 18(17-18), 807-817.
Received: September 04, 2014 Revised: October 23, 2014 Accepted: October 24, 2014




















