
A Comparative Study of HLA Binding Affinity and Ligand Diversity: Implications for Generating...
7
A Comparative Study of HLA Binding Affinity and Ligand Diversity: Implications for Generating Immunodominant CD8 T Cell Responses 1 Xiangyu Rao,* Ana Isabel C. A. Fontaine Costa, † Debbie van Baarle, † and Can Kes ¸mir 2 * ‡ Conventional CD8 T cell responses against intracellular infectious agents are initiated upon recognition of pathogen-derived peptides presented at the cell surface of infected cells in the context of MHC class I molecules. Among the major MHC class I loci, HLA-B is the swiftest evolving and the most polymorphic locus. Additionally, responses restricted by HLA-B molecules tend to be dominant, and most associations with susceptibility or protection against infectious diseases have been assigned to HLA-B alleles. To assess whether the differences in responses mediated via two major HLA class I loci, HLA-B and HLA-A, may already begin at the Ag presentation level, we have analyzed the diversity and binding affinity of their peptide repertoire by making use of curated pathogen-derived epitope data retrieved from the Immune Epitope Database and Analysis Resource, as well as in silico predicted epitopes. In contrast to our expectations, HLA-B alleles were found to have a less diverse peptide repertoire, which points toward a more restricted binding motif, and the respective average peptide binding affinity was shown to be lower than that of HLA-A-restricted epitopes. This unexpected observation gives rise to new hypotheses concerning the mechanisms underlying immunodominance of CD8 T cell responses. The Journal of Immunology, 2009, 182: 1526 –1532. M HC class I molecules play a crucial role in initiating potentially protective immune responses by presenting intracellular pathogen-derived peptides to CD8 T cells and thus targeting infected cells for elimination. The enor- mous polymorphism of MHC class I genes (1) is most likely pathogen-driven (2– 6). Still, this selection pressure does not seem to be acting homogeneously in all major class I loci. Among the homologous major HLA class I loci, HLA-B seems to possess some distinctive characteristics. HLA-B is the most diverse class I locus and the most polymorphic gene of the human genome (7). Indeed, it seems to be the most rapidly evolving locus and it has an exceptional rate of recombination (8), suggesting that it is under the strongest selective pressure. The latter has also been high- lighted by Prugnolle and colleagues, who have shown that the positive correlation between variation at the classical HLA class I loci and pathogen richness is strongest for HLA-B (5). Further- more, HLA-B-restricted pathogen-derived epitopes seem to be more frequently targeted by CD8 T cells than those presented on HLA-A. This has been shown for viral pathogens such as HIV, EBV, influenza, and CMV (9 –12), as well as for an intracellular bacterial pathogen, Mycobacterium tuberculosis (13). Addition- ally, HLA-B-dictated T cell responses seem to be of higher mag- nitude (10, 13, 14) and have been described to influence infectious disease course and outcome. In HIV infection, a specific group of B alleles (e.g., B*3502, B*3503, B*3504, B*5301, B*5802) has been associated with fast progression toward AIDS (9, 15, 16), while others, like B27, B57, and B*5801, are commonly referred to as “protective”, as they are associated with a longer AIDS-free survival period or other favorable features related to a better prog- nosis, such as a lower viral load (17, 18). Kiepiela and colleagues have also demonstrated a broader dominance of B alleles in influ- encing HIV disease outcome due to their impact on viral setpoint and CD4 T cell counts (9). Also, in hepatitis C virus infection, HLA-B27 seems to be the allele most significantly associated with spontaneous viral clearance (19). Furthermore, the association of B53 with protection from severe malaria has most likely contrib- uted to its high frequencies in West Africa (20). Although the associations of HLA-B molecules with disease outcome suggest a fundamental role of CD8 T cells in responding against intracel- lular pathogens upon recognition of peptide-HLA-B complexes in infected cells, they may also be a consequence of engagement of HLA-B by NK cell receptors and subsequent impact on NK cell function (17, 21). The three features of HLA-B molecules mentioned above— high polymorphism, trend to induce immunodominant CD8 T cell responses, and association with disease susceptibility or pro- tection—are most likely related: HLA-B alleles may be major de- terminants, “for good or for bad”, of the effectiveness of immune responses against pathogens because the responses they elicit are dominant. This immunodominance may, among others, stem from differences in presentation of antigenic peptides via HLA-B or from a more amplified qualitative and/or quantitatively response from specific CD8 T cells to HLA-B-restricted epitopes (22, 23). Intrinsic differences in the amount/diversity and binding affinity (and thus stability at the cell surface) of the peptides they present might provide the grounds for their superiority. These issues on peptide presentation have been addressed in this study. We envi- sioned that HLA-B alleles would have a broader peptide repertoire *Department of Theoretical Biology/Bioinformatics, Utrecht University, Utrecht, The Netherlands; † Department of Immunology, Wilhelmina Children’s Hospital, Univer- sity Medical Center Utrecht, Utrecht, The Netherlands; and ‡ Academic Biomedical Centre, Utrecht University, Utrecht, The Netherlands Received for publication September 24, 2008. Accepted for publication November 16, 2008. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 This work was supported by a HIPO (High Potential) grant from Universiteit Utrecht. 2 Address correspondence and reprint requests to Dr. Can Kesmir, Department of Theoretical Biology/Bioinformatics, Padualaan 8, 3584 CH Utrecht, The Netherlands. E-mail address: [email protected] Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00 The Journal of Immunology www.jimmunol.org
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of A Comparative Study of HLA Binding Affinity and Ligand Diversity: Implications for Generating...

A Comparative Study of HLA Binding Affinity and LigandDiversity: Implications for Generating Immunodominant CD8�
T Cell Responses1
Xiangyu Rao,* Ana Isabel C. A. Fontaine Costa,† Debbie van Baarle,† and Can Kesmir2*‡
Conventional CD8� T cell responses against intracellular infectious agents are initiated upon recognition of pathogen-derivedpeptides presented at the cell surface of infected cells in the context of MHC class I molecules. Among the major MHC class I loci,HLA-B is the swiftest evolving and the most polymorphic locus. Additionally, responses restricted by HLA-B molecules tend tobe dominant, and most associations with susceptibility or protection against infectious diseases have been assigned to HLA-Balleles. To assess whether the differences in responses mediated via two major HLA class I loci, HLA-B and HLA-A, may alreadybegin at the Ag presentation level, we have analyzed the diversity and binding affinity of their peptide repertoire by making useof curated pathogen-derived epitope data retrieved from the Immune Epitope Database and Analysis Resource, as well as in silicopredicted epitopes. In contrast to our expectations, HLA-B alleles were found to have a less diverse peptide repertoire, whichpoints toward a more restricted binding motif, and the respective average peptide binding affinity was shown to be lower than thatof HLA-A-restricted epitopes. This unexpected observation gives rise to new hypotheses concerning the mechanisms underlyingimmunodominance of CD8� T cell responses. The Journal of Immunology, 2009, 182: 1526–1532.
M HC class I molecules play a crucial role in initiatingpotentially protective immune responses by presentingintracellular pathogen-derived peptides to CD8� T
cells and thus targeting infected cells for elimination. The enor-mous polymorphism of MHC class I genes (1) is most likelypathogen-driven (2–6). Still, this selection pressure does not seemto be acting homogeneously in all major class I loci. Among thehomologous major HLA class I loci, HLA-B seems to possesssome distinctive characteristics. HLA-B is the most diverse class Ilocus and the most polymorphic gene of the human genome (7).Indeed, it seems to be the most rapidly evolving locus and it has anexceptional rate of recombination (8), suggesting that it is underthe strongest selective pressure. The latter has also been high-lighted by Prugnolle and colleagues, who have shown that thepositive correlation between variation at the classical HLA class Iloci and pathogen richness is strongest for HLA-B (5). Further-more, HLA-B-restricted pathogen-derived epitopes seem to bemore frequently targeted by CD8� T cells than those presented onHLA-A. This has been shown for viral pathogens such as HIV,EBV, influenza, and CMV (9–12), as well as for an intracellularbacterial pathogen, Mycobacterium tuberculosis (13). Addition-ally, HLA-B-dictated T cell responses seem to be of higher mag-
nitude (10, 13, 14) and have been described to influence infectiousdisease course and outcome. In HIV infection, a specific group ofB alleles (e.g., B*3502, B*3503, B*3504, B*5301, B*5802) hasbeen associated with fast progression toward AIDS (9, 15, 16),while others, like B27, B57, and B*5801, are commonly referredto as “protective”, as they are associated with a longer AIDS-freesurvival period or other favorable features related to a better prog-nosis, such as a lower viral load (17, 18). Kiepiela and colleagueshave also demonstrated a broader dominance of B alleles in influ-encing HIV disease outcome due to their impact on viral setpointand CD4� T cell counts (9). Also, in hepatitis C virus infection,HLA-B27 seems to be the allele most significantly associated withspontaneous viral clearance (19). Furthermore, the association ofB53 with protection from severe malaria has most likely contrib-uted to its high frequencies in West Africa (20). Although theassociations of HLA-B molecules with disease outcome suggest afundamental role of CD8� T cells in responding against intracel-lular pathogens upon recognition of peptide-HLA-B complexes ininfected cells, they may also be a consequence of engagement ofHLA-B by NK cell receptors and subsequent impact on NK cellfunction (17, 21).
The three features of HLA-B molecules mentioned above—high polymorphism, trend to induce immunodominant CD8� Tcell responses, and association with disease susceptibility or pro-tection—are most likely related: HLA-B alleles may be major de-terminants, “for good or for bad”, of the effectiveness of immuneresponses against pathogens because the responses they elicit aredominant. This immunodominance may, among others, stem fromdifferences in presentation of antigenic peptides via HLA-B orfrom a more amplified qualitative and/or quantitatively responsefrom specific CD8� T cells to HLA-B-restricted epitopes (22, 23).Intrinsic differences in the amount/diversity and binding affinity(and thus stability at the cell surface) of the peptides they presentmight provide the grounds for their superiority. These issues onpeptide presentation have been addressed in this study. We envi-sioned that HLA-B alleles would have a broader peptide repertoire
*Department of Theoretical Biology/Bioinformatics, Utrecht University, Utrecht, TheNetherlands; †Department of Immunology, Wilhelmina Children’s Hospital, Univer-sity Medical Center Utrecht, Utrecht, The Netherlands; and ‡Academic BiomedicalCentre, Utrecht University, Utrecht, The Netherlands
Received for publication September 24, 2008. Accepted for publication November 16,2008.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by a HIPO (High Potential) grant from UniversiteitUtrecht.2 Address correspondence and reprint requests to Dr. Can Kesmir, Department ofTheoretical Biology/Bioinformatics, Padualaan 8, 3584 CH Utrecht, The Netherlands.E-mail address: [email protected]
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org

than that of HLA-A alleles, inducing a more diverse CTL re-sponse, and that the epitopes would be of higher affinity, ensuringa longer stimulation of T cell responses and thus further account-ing for the immunodominance observed. Surprisingly, our resultsshow that, in contrast with our initial expectations, HLA-B allelesmay actually have a more restricted binding motif, they maypresent fewer peptides than do their HLA-A homologs, and theirbinding affinity tends to be lower. How these particular HLA-Bfeatures may lead to immunodominance is thus far unclear, yetthey open up new hypotheses for the effects of Ag presentation onimmunodominance.
Materials and MethodsEpitope data
All epitopes previously shown to bind to HLA molecules were downloadedas an XML file from the Immune Epitope Database and Analysis Resource(IEDB3; www.immuneepitope.org/;downloads were made in May 2007).As the chance of two different proteins sharing a peptide larger than a7-mer seems to be maximally 2–3% (24), all epitopes with fewer thanseven amino acids similarity were kept as unique epitopes for each HLAallele. In total, 11,878 unique epitopes for 19 HLA-A alleles and 2353unique epitopes for 16 HLA-B alleles having a length varying from 6 to 16amino acids were obtained from IEDB as our experimental binding data.
The affinity of most (�98%) HLA-A and B epitopes in IEDB wasmeasured by two experimental methods. The first method (competitionassays where purified MHC and radioactive labeling are used) covered94% of HLA-A epitopes and 78% of HLA-B epitopes, while the secondmethod (association studies where purified MHC and fluorescence la-beling are used) covered 5% HLA-A epitopes and 21% HLA-Bepitopes. Eighty-five epitopes were measured by both methods. Usingthis overlap data, we found a significant correlation of measured affin-ities by both methods ( p � 0.0001).
Genomic data
Sequences of the human proteome as a resource of self-peptides, 13 com-mon bacteria, and 17 common virus proteomes (supplemental Table S1)4
as resources of pathogenic (non-self) peptides were downloaded from theEuropean Bioinformatics Institute web site (www.ebi.ac.uk/; downloadswere made in Oct 2006). Peptide fragments of 9 or 10 amino acids longwere generated from each protein in all proteomes using all positions aspossible first positions. All redundant 9-mers or 10-mers that are presentmore than one time in a proteome or contain any ambiguous amino acidssuch as “B, Z, X” were deleted from the dataset. In the end, a total of10,941,519 unique human 9-mers, 11,647,396 unique bacterial 9-mers, and57,433 unique viral 9-mers were retrieved from these proteomes.
Two types of sequence modifications in the original proteomes wereapplied in this study: 1) protein shuffling, that is, shuffling residues withina protein by keeping the frequency of amino acids and the length of theprotein intact; and 2) generating artificial proteomes, that is, artificial pro-teomes were generated by using a uniform amino acid distribution (i.e.,frequency of each amino acid is 0.05) without changing the size of originalproteome. All of these sequence modifications were performed by Perlprogramming.
HLA peptide binding predictions
There are at present several HLA peptide binding predictors, such as thepolynomial method (25), artificial neural networks (26), the BIMAS algo-rithm (27), a classification and regression tree (28), an additive method(29), and the stabilized matrix method (SMM) (30). The performance ofthese algorithms to identify new epitopes has recently been benchmarkedon experimental data (31). In general, artificial neural networks and SMMmethods were found to be superior to the other ones. Moreover, the resultsof these large-scale benchmark studies show that the accuracy of HLApeptide binding prediction algorithms has increased over time to such anextent that the correlation between predicted and measured binding affinityis as good as the correlation between measurements from different labo-ratories (31). Another demonstration of how powerful these predictions areis a study by Schellens et al. (7), who identified 18 new CTL epitopes outof a set of 22 predicted CTL epitopes using artificial neural networks-based
HLA peptide binding predictors. This suggests that the specificity of thepredictors can be as high as 80%.
SMM was chosen as the main prediction method in this project becauseit has the most broad HLA coverage at the time this project was initiated.For each HLA molecule a specific SMM matrix was generated using theexperimental binding data. The details of this algorithm are given else-where (30). We downloaded SMM prediction software and training data-sets from IEDB (mhcbindingpredictions.immuneepitope.org/) and gener-ated scoring matrices ourselves. For 9-mer prediction, 19 HLA-A and 15HLA-B SMM matrices were generated, while for 10-mer prediction 14HLA-A and 5 HLA-B matrices were generated because of training datalimitation.
As suggested by Ruppert et al. (32), an IC50 threshold of 500 nM wasused to distinguish binding peptides from nonbinding peptides.
Thus, the binding fraction of a particular HLA allele was calculated as:binding fraction � (no. of n-mers/no. of all n-mers) (n � 9 or 10).
Shannon entropy calculation and binding motif visualization
The Shannon entropy (33) was calculated to measure the degree of vari-ability in HLA binding motifs. This entropy measure is defined as:
E�i� � � �L � 1
20
qi log2 qi,
where E(i) is the Shannon entropy at position i, and qi is the probability thata particular amino acid occurs at position i in the alignment. The maximumvalue of the Shannon entropy is obtained when all amino acids occur withsame frequency (�20(1/20)log2(1/20) � 4.3) and the minimum Shannonentropy is 0 if a position is fully conserved. The binding motifs of HLAmolecules were visualized by sequence logo technique (34) in this study(see caption of supplemental Fig. S5 for an explanation).
ResultsHLA-A alleles present a larger set of epitopes than do HLA-Balleles
To explore the differences in the epitope repertoire of HLA mol-ecules, publicly available experimental epitope data were ana-lyzed. At present, IEDB is the most complete database for exper-imentally verified epitopes. All epitopes that can bind to HLA-A orHLA-B alleles were retrieved from IEDB (downloads were madein October 2006). The number of known epitopes per allele showsa very skewed distribution: for the common alleles, thousands ofepitopes are identified, while for the rare alleles as few as only oneepitope is found. In IEDB the average number of unique epitopesfor each HLA-A allele is 216 (median, 15; range, 1–2341) and theaverage number of unique epitopes for each HLA-B allele is 37(median, 8; range, 1–399). Thus, the diversity of HLA-A bindingepitopes seems to be larger than that of HLA-B. After excludingalleles having fewer than 20 epitopes, we calculated the epitopelength distribution for 19 HLA-A and 16 HLA-B alleles (Fig. 1).The epitope length distributions of HLA-A and HLA-B alleleswere very similar and showed a marked preference to bindepitopes of length 9 (9-mers) or length 10 (10-mers) (Fig. 1). Forthe rest of the paper this data set is used and referred to as “ex-perimental binding data”.
The experimental data might have some biases, for example, dueto dominance of certain HLA molecules like HLA-A*02 (www.allelefrequencies.net). Alternatively, one can screen proteomes forepitopes restricted by every HLA molecule for which predictionmethods are available to determine epitope repertoires. We basedour analysis on SMM predictions because at the time we initializedour study the SMM method had the largest allele coverage. Usingpublicly available software and data, we generated 19 HLA-A and15 HLA-B SMM scoring matrices and predicted the binding af-finity of all possible 9-mers from the human, bacterial, and viralproteomes (as explained in Materials and Methods). SMM matri-ces assign to each 9-mer HLA pair an IC50 value that indicates thebinding affinity. The lower IC50 value implies the higher binding
3 Abbreviations used in this paper: IEDB, Immune Epitope Database and AnalysisResource; SMM, stabilized matrix method.4 The online version of this article contains supplemental material.
1527The Journal of Immunology
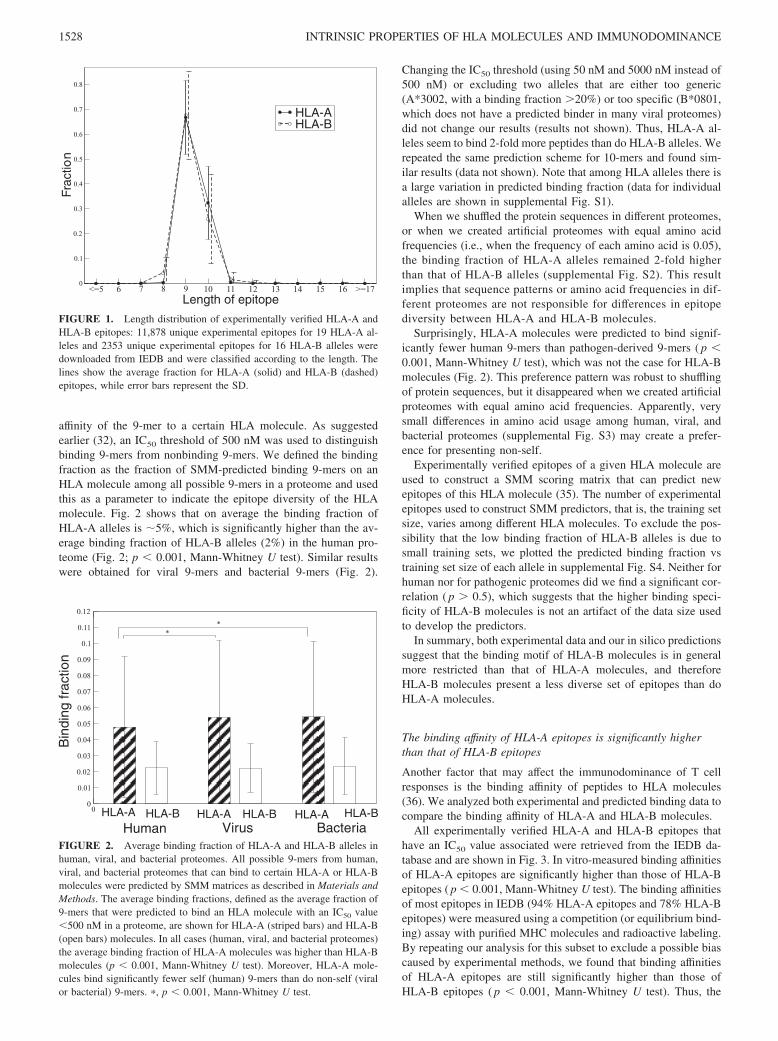
affinity of the 9-mer to a certain HLA molecule. As suggestedearlier (32), an IC50 threshold of 500 nM was used to distinguishbinding 9-mers from nonbinding 9-mers. We defined the bindingfraction as the fraction of SMM-predicted binding 9-mers on anHLA molecule among all possible 9-mers in a proteome and usedthis as a parameter to indicate the epitope diversity of the HLAmolecule. Fig. 2 shows that on average the binding fraction ofHLA-A alleles is �5%, which is significantly higher than the av-erage binding fraction of HLA-B alleles (2%) in the human pro-teome (Fig. 2; p � 0.001, Mann-Whitney U test). Similar resultswere obtained for viral 9-mers and bacterial 9-mers (Fig. 2).
Changing the IC50 threshold (using 50 nM and 5000 nM instead of500 nM) or excluding two alleles that are either too generic(A*3002, with a binding fraction �20%) or too specific (B*0801,which does not have a predicted binder in many viral proteomes)did not change our results (results not shown). Thus, HLA-A al-leles seem to bind 2-fold more peptides than do HLA-B alleles. Werepeated the same prediction scheme for 10-mers and found sim-ilar results (data not shown). Note that among HLA alleles there isa large variation in predicted binding fraction (data for individualalleles are shown in supplemental Fig. S1).
When we shuffled the protein sequences in different proteomes,or when we created artificial proteomes with equal amino acidfrequencies (i.e., when the frequency of each amino acid is 0.05),the binding fraction of HLA-A alleles remained 2-fold higherthan that of HLA-B alleles (supplemental Fig. S2). This resultimplies that sequence patterns or amino acid frequencies in dif-ferent proteomes are not responsible for differences in epitopediversity between HLA-A and HLA-B molecules.
Surprisingly, HLA-A molecules were predicted to bind signif-icantly fewer human 9-mers than pathogen-derived 9-mers ( p �0.001, Mann-Whitney U test), which was not the case for HLA-Bmolecules (Fig. 2). This preference pattern was robust to shufflingof protein sequences, but it disappeared when we created artificialproteomes with equal amino acid frequencies. Apparently, verysmall differences in amino acid usage among human, viral, andbacterial proteomes (supplemental Fig. S3) may create a prefer-ence for presenting non-self.
Experimentally verified epitopes of a given HLA molecule areused to construct a SMM scoring matrix that can predict newepitopes of this HLA molecule (35). The number of experimentalepitopes used to construct SMM predictors, that is, the training setsize, varies among different HLA molecules. To exclude the pos-sibility that the low binding fraction of HLA-B alleles is due tosmall training sets, we plotted the predicted binding fraction vstraining set size of each allele in supplemental Fig. S4. Neither forhuman nor for pathogenic proteomes did we find a significant cor-relation ( p � 0.5), which suggests that the higher binding speci-ficity of HLA-B molecules is not an artifact of the data size usedto develop the predictors.
In summary, both experimental data and our in silico predictionssuggest that the binding motif of HLA-B molecules is in generalmore restricted than that of HLA-A molecules, and thereforeHLA-B molecules present a less diverse set of epitopes than doHLA-A molecules.
The binding affinity of HLA-A epitopes is significantly higherthan that of HLA-B epitopes
Another factor that may affect the immunodominance of T cellresponses is the binding affinity of peptides to HLA molecules(36). We analyzed both experimental and predicted binding data tocompare the binding affinity of HLA-A and HLA-B molecules.
All experimentally verified HLA-A and HLA-B epitopes thathave an IC50 value associated were retrieved from the IEDB da-tabase and are shown in Fig. 3. In vitro-measured binding affinitiesof HLA-A epitopes are significantly higher than those of HLA-Bepitopes ( p � 0.001, Mann-Whitney U test). The binding affinitiesof most epitopes in IEDB (94% HLA-A epitopes and 78% HLA-Bepitopes) were measured using a competition (or equilibrium bind-ing) assay with purified MHC molecules and radioactive labeling.By repeating our analysis for this subset to exclude a possible biascaused by experimental methods, we found that binding affinitiesof HLA-A epitopes are still significantly higher than those ofHLA-B epitopes ( p � 0.001, Mann-Whitney U test). Thus, the
<=5 6 7 8 9 10 11 12 13 14 15 16 >=170
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Frac
tion
HLA-AHLA-B
FIGURE 1. Length distribution of experimentally verified HLA-A andHLA-B epitopes: 11,878 unique experimental epitopes for 19 HLA-A al-leles and 2353 unique experimental epitopes for 16 HLA-B alleles weredownloaded from IEDB and were classified according to the length. Thelines show the average fraction for HLA-A (solid) and HLA-B (dashed)epitopes, while error bars represent the SD.
FIGURE 2. Average binding fraction of HLA-A and HLA-B alleles inhuman, viral, and bacterial proteomes. All possible 9-mers from human,viral, and bacterial proteomes that can bind to certain HLA-A or HLA-Bmolecules were predicted by SMM matrices as described in Materials andMethods. The average binding fractions, defined as the average fraction of9-mers that were predicted to bind an HLA molecule with an IC50 value�500 nM in a proteome, are shown for HLA-A (striped bars) and HLA-B(open bars) molecules. In all cases (human, viral, and bacterial proteomes)the average binding fraction of HLA-A molecules was higher than HLA-Bmolecules (p � 0.001, Mann-Whitney U test). Moreover, HLA-A mole-cules bind significantly fewer self (human) 9-mers than do non-self (viralor bacterial) 9-mers. �, p � 0.001, Mann-Whitney U test.
1528 INTRINSIC PROPERTIES OF HLA MOLECULES AND IMMUNODOMINANCE

differences we found between HLA-A and HLA-B epitope affin-ities are not due to the use of different experimental assays.
We then analyzed the in silico-predicted binding affinities fromthe human, viral, and bacterial proteomes. To exclude a potentialbias introduced by the 2-fold higher amount of predicted HLA-Aepitopes as compared with those restricted by HLA-B alleles, weselected the top five predicted epitopes, with the highest predictedbinding affinity, for each HLA molecule from each proteome. Theaverage binding affinity of predicted HLA-A epitopes is signifi-cantly higher ( p � 0.001, Mann-Whitney U test) than that of pre-dicted HLA-B epitopes (Fig. 3). The results were confirmed byrepeating the same analysis for the top 10 epitopes of each HLAmolecule (results not shown).
Although the predicted binding affinities show the same ten-dency as the experimental data (Fig. 3), one can argue that thepredictions are not true validations of the experimental data, be-cause they are not independent, that is, the prediction methodswere trained on the available experimental data. To separate theexperimental data as much as possible from the predictions, weused the following strategy: each time we chose three HLA-A andthree HLA-B alleles randomly from all of the alleles where pre-diction methods were available. Then, we compared the predictedbinding affinities for the randomly chosen three HLA-A andHLA-B alleles, while excluding the six alleles used in the predic-tion analysis from the experimental binding data and comparedHLA-A and HLA-B binding affinities in the rest. In this way, thealleles (and thus the epitope data) tested using the experimentaldata and the predictions are nonoverlapping. We repeated this pro-cess 100 times, and in 92 cases we found that both the predictedand experimentally verified binding affinities of HLA-B alleleswere significantly lower than those for HLA-A alleles. One cantherefore conclude that the significant difference in binding affin-ities of HLA-A and HLA-B alleles found in the experimental datais further supported with our predictions.
Next we tested whether the higher binding affinity can be aresult of lower specificity in HLA-A binding motifs. In Fig. 4 we
plotted the binding fraction vs the binding affinity of the top fivepredicted epitopes for all HLA molecules in our in silico analysis.Interestingly, there is a significant positive correlation ( p � 0.001)between the binding fraction and the average peptide binding af-finity of these alleles, which suggests that a less specific bindingmotif would imply the higher binding affinities.
In summary, HLA-A epitopes have significantly higher bindingaffinities than do HLA-B epitopes, and a significant positive cor-relation was found between the binding fraction and the bindingaffinity of HLA molecules.
HLA-A and HLA-B molecules have different restrictions inanchor residues
Sequence patterns in a cohort of epitopes specific for a certainHLA molecule define its binding motif (37). Some positions of
FIGURE 3. Binding affinities of HLA-A epitopes are significantlyhigher than HLA-B epitopes. Left, Binding affinity (IC50 value) for HLA-Aand HLA-B epitopes measured in vitro (data downloaded from IEDB).Right, Binding affinity of top five predicted epitopes in human, viral, andbacterial proteomes for 19 HLA-A and 15 HLA-B molecules. The graylines show the average. In all cases the binding affinities of HLA-A mol-ecules were significantly higher (p � 0.001, Mann-Whitney U test) thanthose of HLA-B molecules.
0.0001 0.001 0.01 0.1
10
100
1000HLA-AHLA-B
Bin
ding
affi
nity
(IC
50)
FIGURE 4. Correlation between binding fraction and binding affinity. Asignificant correlation (correlation of �0.25; p � 0.001, Mann-Whitney Utest) was found between predicted binding fraction of 19 HLA-A and 15HLA-B molecules and predicted binding affinity of their top five epitopesin bacterial proteomes. Filled circles depict HLA-A epitopes and emptycircles depict HLA-B epitopes. Similar results were found in human andviral proteomes (human correlation of �0.32, p � 0.001; virus correlationof �0.31, p � 0.001).
FIGURE 5. Variability in experimentally verified HLA-A and HLA-Bepitopes. The entropy of HLA-A (filled circles) and HLA-B (open circles)epitopes were calculated based on 7238 unique epitopes (length of 9) of 19HLA-A alleles and 1585 unique epitopes (length of 9) of 16 HLA-B allelesdownloaded from IEDB database. Only in P2 and P9 is the variability ofHLA-A and HLA-B epitopes significantly different (both cases p � 0.001,Mann-Whitney U test). Shannon entropy for P2 of HLA-B epitopes issignificantly lower (p � 0.017, Mann-Whitney U test) than that of the bestconserved position of HLA-A, the P9.
1529The Journal of Immunology

HLA binding motif (e.g., P2, P3, P5, P9), which are very importantfor epitope binding, were defined as anchor positions (38–41).
To explore if the difference in binding specificity of HLA-A andHLA-B molecules is due to their binding motifs, all 9-mer epitopesfrom our experimental binding data were used to measure the de-gree of variability at each position in terms of the Shannon entropy(33), as explained in Materials and Methods. The larger Shannonentropy indicates higher level of variation in a position. Only thesequence conservation in P2 and P9 was significantly differentbetween HLA-A and HLA-B epitopes ( p � 0.001, Mann-WhitneyU test; Fig. 5). Interestingly, P2 is the most conserved position forHLA-B alleles, while for HLA-A alleles P9 has the lowest Shan-non entropy (Fig. 5).
Classifying all HLA molecules by their 9-mer binding motifpatterns, we observed three groups of binding motifs. Group 1includes HLA alleles with binding motifs that have a similarlevel of conservation in P2 and P9, and it contains both HLA-Aand HLA-B molecules. Group 2 contains only HLA-B mole-cules, defined by a markedly conserved P2. Group 3 enclosesonly HLA-A molecules, and these share a conservative P9 intheir binding motifs. Supplemental Fig. S5 gives two examplesfor each of these three groups using sequence logos (see figurecaption for explanation).
Thus, in general both HLA-A and HLA-B molecules are moreconserved in their anchor residues (e.g., P2 and P9), where P2 ismore important for HLA-B alleles and P9 is more important forHLA-A alleles.
DiscussionImmune responses restricted by HLA-B alleles to several patho-gens have been previously shown to be immunodominant (9–11,13, 14). Additionally, particular HLA-B alleles seem to be asso-ciated with either protection or susceptibility to infectious diseases(see reviews in Refs. 17–20). It has been suggested that this asso-ciation of HLA-B molecules with a favorable disease outcomemay derive from the fact that they are ligands for NK cell re-ceptors, and thus may promote either NK cell lysis of infectedcells or inhibit NK cell function and diminish chronic immuneactivation (17, 21). Alternatively, the immunodominance ofHLA-B-restricted CD8� T cell responses might be the under-lying reason why particular HLA-B alleles of a given individualtend to determine the effectiveness of the immune responsesthat he/she mounts.
In this study, we undertook two approaches to evaluate if theimmunodominance of HLA-B-restricted responses relates to theintrinsic Ag presentation features of these alleles in terms of di-versity and binding affinity of the epitopes they present. First, wecompared experimental data on pathogen-derived epitopes avail-able in the IEDB database. Second, and to exclude for potentialbiases in the IEDB epitope set, we performed the same analysis onin silico-predicted peptides derived from proteomes of viruses,bacteria, and human genomes. This analysis did not includeepitopes restricted by HLA-C alleles, given that these have beenhistorically less studied, and thus the data are more scarce and,consequently, no good predictors are currently available.
Surprisingly, we show that HLA-B alleles present a less diverseset of epitopes, �5-fold lower than that of HLA-A alleles (basedon experimental data). This disparity does not seem to be exclu-sively explained by the large amount of peptides described to bepresented by HLA-A2, and is narrower (around 2.5 fold) whenderived from prediction data (Fig. 2). The familiar notion that9-mers are the most preferred MHC class I ligands (42, 43) can beconfirmed in this analysis, and it holds true for both loci (Fig. 1).Curiously, our predictions suggest that HLA-A alleles have a pref-
erence for foreign over self-epitopes, which does not hold true forHLA-B molecules (Fig. 2) and is lost when the subtle differencesamong amino acid frequencies are excluded (see supplementalFigs. S2 and S3).
High HLA peptide binding affinity is not strictly correlated withimmunodominance (44). Moreover, Assarsson et al. showed thatdominant epitopes might have lower affinities than subdominantepitopes (36). In line with these findings, we found that the averagebinding affinity of epitopes restricted by HLA-B alleles is signif-icantly lower than that of HLA-A peptides, in both experimentaland predicted data (Fig. 3). This might reflect a tendency forHLA-B molecules to be less dependent on tapasin-ERp57, a com-plex involved in editing the peptide cargo of MHC class I in favorof complexes with longer cell surface half-lives enclosing epitopesof higher binding affinity (45–48). In fact, thus far only B alleleshave been shown to associate poorly with this chaperone, amongwhich B27 and B*4405 (48–50). HLA dependence on tapasinseems to be dictated by polymorphisms at residues 116 and 114located at the base of the F pocket of the binding cleft, whichaccommodates the peptide C terminus (48), and the complex hasbeen proposed to act by disrupting peptide-MHC class I complexformation until high-affinity peptides are bound (47).
Interestingly, binding fraction and binding affinity seem to be pos-itively correlated (Fig. 4). HLA-A alleles seem to have a more per-missive binding motif, where the most conserved residue is theC-terminal (Fig. 5 and supplemental Fig. S5). Consequently, thechance of finding peptides that fit their looser requirements ishigher and will account for a larger pool of possible binders, fromwhich the high-affinity peptides may be selected. Conversely, thebinding motif of HLA-B alleles appears to be more restricted, es-pecially in P2 (Fig. 5 and supplemental Fig. S5). In accordance, theinformation content for P2 of HLA-B epitopes is significantlyhigher ( p � 0.017, Mann-Whitney U test) than that of the bestconserved position of HLA-A, the C terminus. Thus, there is anarrower probability that the small proteome of pathogens willenclose sequences that would perfectly conform to HLA-B pre-requisites. As a result, the peptides loaded in HLA-B complexesmight frequently be suboptimal.
Another factor that determines the available peptides for eachHLA is the specificity of the proteasome. Kesmir et al. have shownthat the immunoproteasome has co-evolved with human MHCmolecules, thus optimizing the process of Ag presentation (51).We, therefore, plan to test next if especially HLA-A alleles haveco-evolved with the proteasome, and thus have “designed” their Fpocket as to favor the C terminus amino acids defined by the cleav-age patterns of the catalytic subunits, while HLA-B alleles, evolv-ing faster, may have a more variable F pocket and not so strictrequirements for the C-terminal amino acids.
In summary, HLA-B alleles seem to present fewer peptides toCD8� T cells, which is consistent with a more restricted bindingmotif, and epitope binding affinity seems to be lower as comparedwith their HLA-A counterparts. Note that experimental data onbinding affinity are generally determined in vitro using syntheticpeptides derived from the protein sequence under study, ratherthan from naturally processed peptides. The in silico analysis,which we herein use to exclude possible sampling biases in theexperimental data, was therefore also performed for all possible9-mers, and it does not take into account the limiting steps of Agprocessing (efficiency of proteasome cleavage, TAP transport, N-terminal trimming). Despite the lack of data on naturally processedpeptides, our results, although unexpected and counterintuitive,call for novel hypotheses to explain why HLA-B alleles are asso-ciated with immunodominance.
1530 INTRINSIC PROPERTIES OF HLA MOLECULES AND IMMUNODOMINANCE

First, if epitopes indeed bind with higher affinity to HLA-Amolecules, these more stable complexes might remain longer at thecell surface. Stimulation of cognate T cells will thus occur longerand may potentially lead to exhaustion of these responses, espe-cially in cases of chronic infection and, thus, Ag persistence. Thiswould be in agreement with the findings of Harari and colleagues,who have shown that HLA-A-restricted epitopes drive high-avid-ity T cell responses by CTLs that have high expression of theexhaustion marker programmed death (PD)-1 (52). As these high-avidity T cells will be preferentially deleted (53–55), the “surviv-ing” HLA-B-restricted responses would become dominant. In con-trast, Almeida and coworkers (56) have found HLA-B T cellresponses to be of higher avidity during HIV infection. Still, thehighest avidity response tested, although showing signs of higherclonal turnover and some features of senescence, was polyfunc-tional and still seemed to be actively involved in controlling HIVinfection.
Second, because the pool of B epitopes is less diverse, the Ag“density” could be higher for B-restricted ligands; that is, HLA-Balleles would present epitopes more abundantly (assuming identi-cal A and B expression at the cell surface). As such, the chancethat a responsive B-restricted CD8� T cell would see its ligandwould then be increased, and clones responding to these Agswould preferentially expand.
Third, one can envision that the difference in immunodominancemay already be defined at the precursor level. In two studies com-paring immunodominant and subdominant responses to lympho-cytic choriomeningitis virus in mice, differences could be ex-plained by a higher precursor frequency of the former (44, 57).Most interestingly, a recent paper by Obar and colleagues hasquantified naive CD8� T cells and showed that their frequenciescan differ broadly, even more than 10-fold, among different spec-ificities, and that this variation can account for a swifter response,as measured by the timing of the peak response, and may play arole in immunodominance (58). Assuming again that A and Balleles will be expressed in the same level in the thymus, the di-versity, abundance, and affinity of the self-epitopes presented viathese molecules might determine the outcome of positive and neg-ative selection, and thus the naive precursor level. However, thenet effect is hard to predict. It seems reasonable to think that thehigher binding affinity of peptides for HLA-A will allow the com-plexes they form to be more stable at the cell surface, hence anincreased chance of promoting survival during the positive selec-tion, but also of being recognized with too high affinity and in-ducing deletion. Additionally, taking into account that HLA-Bmolecules will present a less diverse repertoire of epitopes, butpotentially to a higher cell surface density, it is tempting to spec-ulate that they may select for larger epitope-specific clones.
In this study, we showed that the HLA locus more associatedwith immunodominant T cell responses seems to be, intriguingly,the one exhibiting the least diverse peptide binding repertoire andpresenting peptides with a lower average binding affinity. Al-though one might conclude that features other than epitope diver-sity and binding affinity might determine immunodominance, wehave explored, above, plausible mechanisms on how these traitscan still play a major role. More detailed studies, for example, thedetermination of HLA-A- and HLA-B-restricted T cell precursorfrequencies, may resolve this discrepancy.
AcknowledgmentsWe thank Julie Grenouillet for generating scoring matrices, Rob de Boerfor valuable comments on the manuscript, and Jorg Calis and Boris Schmidfor technical support.
DisclosuresThe authors have no financial conflicts of interest.
References1. Parham, P., and T. Ohta. 1996. Population biology of antigen presentation by
MHC class I molecules. Science 272: 67–74.2. Hughes, A. L., and M. Nei. 1988. Pattern of nucleotide substitution at major
histocompatibility complex class I loci reveals overdominant selection. Nature335: 167–170.
3. Vogel, T. U., D. T. Evans, J. A. Urvater, D. H. O’Connor, A. L. Hughes, andD. I. Watkins. 1999. Major histocompatibility complex class I genes in primates:co-evolution with pathogens. Immunol. Rev. 167: 327–337.
4. Cooke, G. S., and A. V. Hill. 2001. Genetics of susceptibility to human infectiousdisease. Nat. Rev. Genet. 2: 967–977.
5. Prugnolle, F., A. Manica, M. Charpentier, J. F. Guegan, V. Guernier, andF. Balloux. 2005. Pathogen-driven selection and worldwide HLA class I diver-sity. Curr. Biol. 15: 1022–1027.
6. Borghans, J. A., J. B. Beltman, and R. J. De Boer. 2004. MHC polymorphismunder host-pathogen coevolution. Immunogenetics 55: 732–739.
7. Schellens, I. M., C. Kesmir, F. Miedema, D. van Baarle, and J. A. Borghans.2008. An unanticipated lack of consensus cytotoxic T lymphocyte epitopes inHIV-1 databases: the contribution of prediction programs. AIDS 22: 33–37.
8. McAdam, S. N., J. E. Boyson, X. Liu, T. L. Garber, A. L. Hughes, R. E. Bontrop,and D. I. Watkins. 1994. A uniquely high level of recombination at the HLA-Blocus. Proc. Natl. Acad. Sci. USA 91: 5893–5897.
9. Kiepiela, P., A. J. Leslie, I. Honeyborne, D. Ramduth, C. Thobakgale, S. Chetty,P. Rathnavalu, C. Moore, K. J. Pfafferott, L. Hilton, et al. 2004. Dominant in-fluence of HLA-B in mediating the potential co-evolution of HIV and HLA.Nature 432: 769–775.
10. Bihl, F., N. Frahm, G. L. Di, J. Sidney, M. John, K. Yusim, T. Woodberry,K. Sango, H. S. Hewitt, L. Henry, et al. 2006. Impact of HLA-B alleles, epitopebinding affinity, functional avidity, and viral coinfection on the immunodomi-nance of virus-specific CTL responses. J. Immunol. 176: 4094–4101.
11. Boon, A. C., G. De Mutsert, R. A. Fouchier, K. Sintnicolaas, A. D. Osterhaus,and G. F. Rimmelzwaan. 2004. Preferential HLA usage in the influenza virus-specific CTL response. J. Immunol. 172: 4435–4443.
12. Lacey, S. F., M. C. Villacres, R. C. La, Z. Wang, J. Longmate, J. Martinez,J. C. Brewer, S. Mekhoubad, R. Maas, J. M. Leedom, et al. 2003. Relativedominance of HLA-B*07 restricted CD8� T-lymphocyte immune responses tohuman cytomegalovirus pp65 in persons sharing HLA-A*02 and HLA-B*07 al-leles. Hum. Immunol. 64: 440–452.
13. Lewinsohn, D. A., E. Winata, G. M. Swarbrick, K. E. Tanner, M. S. Cook,M. D. Null, M. E. Cansler, A. Sette, J. Sidney, and D. M. Lewinsohn. 2007.Immunodominant tuberculosis CD8 antigens preferentially restricted by HLA-B.PLoS Pathog. 3: 1240–1249.
14. Weichold, F. F., S. Mueller, C. Kortsik, W. E. Hitzler, M. J. Wulf, D. M. Hone,J. C. Sadoff, and M. J. Maeurer. 2007. Impact of MHC class I alleles on the M.tuberculosis antigen-specific CD8� T-cell response in patients with pulmonarytuberculosis. Genes Immun. 8: 334–343.
15. Carrington, M., G. W. Nelson, M. P. Martin, T. Kissner, D. Vlahov, J. J. Goedert,R. Kaslow, S. Buchbinder, K. Hoots, and S. J. O’Brien. 1999. HLA and HIV-1:heterozygote advantage and B*35-Cw*04 disadvantage. Science 283:1748–1752.
16. Gao, X., G. W. Nelson, P. Karacki, M. P. Martin, J. Phair, R. Kaslow,J. J. Goedert, S. Buchbinder, K. Hoots, D. Vlahov, et al. 2001. Effect of a singleamino acid change in MHC class I molecules on the rate of progression to AIDS.N. Engl. J. Med. 344: 1668–1675.
17. Martin, M. P., and M. Carrington. 2005. Immunogenetics of viral infections.Curr. Opin. Immunol. 17: 510–516.
18. Carrington, M., and S. J. O’Brien. 2003. The influence of HLA genotype onAIDS. Annu. Rev. Med. 54: 535–551.
19. Neumann-Haefelin, C., S. McKiernan, S. Ward, S. Viazov, H. C. Spangenberg,T. Killinger, T. F. Baumert, N. Nazarova, I. Sheridan, O. Pybus, et al. 2006.Dominant influence of an HLA-B27 restricted CD8� T cell response in mediatingHCV clearance and evolution. Hepatology 43: 563–572.
20. Hill, A. V., C. E. Allsopp, D. Kwiatkowski, N. M. Anstey, P. Twumasi,P. A. Rowe, S. Bennett, D. Brewster, A. J. McMichael, and B. M. Greenwood.1991. Common West African HLA antigens are associated with protection fromsevere malaria. Nature 352: 595–600.
21. Khakoo, S. I., and M. Carrington. 2006. KIR and disease: a model system orsystem of models? Immunol. Rev. 214: 186–201.
22. Yewdell, J. W., and J. R. Bennink. 1999. Immunodominance in major histocom-patibility complex class I-restricted T lymphocyte responses. Annu. Rev. Immu-nol. 17: 51–88.
23. Yewdell, J. W. 2006. Confronting complexity: real-world immunodominance inantiviral CD8� T cell responses. Immunity 25: 533–543.
24. Burroughs, N. J., R. J. De Boer, and C. Kesmir. 2004. Discriminating self fromnonself with short peptides from large proteomes. Immunogenetics 56: 311–320.
25. Rammensee, H., J. Bachmann, N. P. Emmerich, O. A. Bachor, and S. Stevanovic.1999. SYFPEITHI: database for MHC ligands and peptide motifs. Immunoge-netics 50: 213–219.
26. Gulukota, K., J. Sidney, A. Sette, and C. DeLisi. 1997. Two complementarymethods for predicting peptides binding major histocompatibility complex mol-ecules. J. Mol. Biol. 267: 1258–1267.
1531The Journal of Immunology

27. Parker, K. C., M. A. Bednarek, and J. E. Coligan. 1994. Scheme for rankingpotential HLA-A2 binding peptides based on independent binding of individualpeptide side-chains. J. Immunol. 152: 163–175.
28. Segal, M. R., M. P. Cummings, and A. E. Hubbard. 2001. Relating amino acidsequence to phenotype: analysis of peptide-binding data. Biometrics 57:632–642.
29. Doytchinova, I. A., M. J. Blythe, and D. R. Flower. 2002. Additive method forthe prediction of protein-peptide binding affinity: application to the MHC class Imolecule HLA-A*0201. J. Proteome Res. 1: 263–272.
30. Peters, B., W. Tong, J. Sidney, A. Sette, and Z. Weng. 2003. Examining theindependent binding assumption for binding of peptide epitopes to MHC-I mol-ecules. Bioinformatics 19: 1765–1772.
31. Peters, B., H. H. Bui, S. Frankild, M. Nielson, C. Lundegaard, E. Kostem,D. Basch, K. Lamberth, M. Harndahl, W. Fleri, et al. 2006. A community re-source benchmarking predictions of peptide binding to MHC-I molecules. PLoSComput. Biol. 2: e65.
32. Ruppert, J., J. Sidney, E. Celis, R. T. Kubo, H. M. Grey, and A. Sette. 1993.Prominent role of secondary anchor residues in peptide binding to HLA-A2.1molecules. Cell 74: 929–937.
33. Shannon, C. E. 1948. A mathematical theory of communication. Bell SystemTech. J. 27: 379.
34. Schneider, T. D., and R. M. Stephens. 1990. Sequence logos: a new way todisplay consensus sequences. Nucleic Acids Res. 18: 6097–6100.
35. Peters, B., and A. Sette. 2005. Generating quantitative models describing thesequence specificity of biological processes with the stabilized matrix method.BMC Bioinformatics 6: 132.
36. Assarsson, E., J. Sidney, C. Oseroff, V. Pasquetto, H. H. Bui, N. Frahm,C. Brander, B. Peters, H. Grey, and A. Sette. 2007. A quantitative analysis of thevariables affecting the repertoire of T cell specificities recognized after vacciniavirus infection. J. Immunol. 178: 7890–7901.
37. Rammensee, H. G., T. Friede, and S. Stevanoviic. 1995. MHC ligands and pep-tide motifs: first listing. Immunogenetics 41: 178–228.
38. Falk, K., O. Rotzschke, B. Grahovac, D. Schendel, S. Stevanovic, G. Jung, andH. G. Rammensee. 1993. Peptide motifs of HLA-B35 and -B37 molecules. Im-munogenetics 38: 161–162.
39. Takamiya, Y., C. Schonbach, K. Nokihara, M. Yamaguchi, S. Ferrone, K. Kano,K. Egawa, and M. Takiguchi. 1994. HLA-B*3501-peptide interactions: role ofanchor residues of peptides in their binding to HLA-B*3501 molecules. Int. Im-munol. 6: 255–261.
40. DiBrino, M., K. C. Parker, D. H. Margulies, J. Shiloach, R. V. Turner,W. E. Biddison, and J. E. Coligan. 1994. The HLA-B14 peptide binding site canaccommodate peptides with different combinations of anchor residues. J. Biol.Chem. 269: 32426–32434.
41. Lamas, J. R., A. Paradela, F. Roncal, and J. A. Lopez De Castro. 1999. Modu-lation at multiple anchor positions of the peptide specificity of HLA-B27 sub-types differentially associated with ankylosing spondylitis. Arthritis Rheum. 42:1975–1985.
42. Falk, K., O. Rotzschke, S. Stevanovic, G. Jung, and H. G. Rammensee. 1991.Allele-specific motifs revealed by sequencing of self-peptides eluted from MHCmolecules. Nature 351: 290–296.
43. Yewdell, J. W., E. Reits, and J. Neefjes. 2003. Making sense of mass destruction:quantitating MHC class I antigen presentation. Nat. Rev. Immunol. 3: 952–961.
44. Kotturi, M. F., I. Scott, T. Wolfe, B. Peters, J. Sidney, H. Cheroutre,M. G. von Herrath, M. J. Buchmeier, H. Grey, and A. Sette. 2008. Naive pre-cursor frequencies and MHC binding rather than the degree of epitope diversityshape CD8� T cell immunodominance. J. Immunol. 181: 2124–2133.
45. Howarth, M., A. Williams, A. B. Tolstrup, and T. Elliott. 2004. Tapasin enhancesMHC class I peptide presentation according to peptide half-life. Proc. Natl. Acad.Sci. USA 101: 11737–11742.
46. Kienast, A., M. Preuss, M. Winkler, and T. P. Dick. 2007. Redox regulation ofpeptide receptivity of major histocompatibility complex class I molecules byERp57 and tapasin. Nat. Immunol. 8: 864–872.
47. Wearsch, P. A., and P. Cresswell. 2007. Selective loading of high-affinity pep-tides onto major histocompatibility complex class I molecules by the tapasin-ERp57 heterodimer. Nat. Immunol. 8: 873–881.
48. Purcell, A. W., and T. Elliott. 2008. Molecular machinations of the MHC-I pep-tide loading complex. Curr. Opin. Immunol. 20: 75–81.
49. Turnquist, H. R., H. J. Thomas, K. R. Prilliman, C. T. Lutz, W. H. Hildebrand,and J. C. Solheim. 2000. HLA-B polymorphism affects interactions with multipleendoplasmic reticulum proteins. Eur. J. Immunol. 30: 3021–3028.
50. Groothuis, T. A., A. C. Griekspoor, J. J. Neijssen, C. A. Herberts, andJ. J. Neefjes. 2005. MHC class I alleles and their exploration of the antigen-processing machinery. Immunol. Rev. 207: 60–76.
51. Kesmir, C., V. van Noort, R. J. De Boer, and P. Hogeweg. 2003. Bioinformaticanalysis of functional differences between the immunoproteasome and the con-stitutive proteasome. Immunogenetics 55: 437–449.
52. Harari, A., C. Cellerai, F. B. Enders, J. Kostler, L. Codarri, G. Tapia, O. Boyman,E. Castro, S. Gaudieri, I. James, et al. 2007. Skewed association of polyfunctionalantigen-specific CD8 T cell populations with HLA-B genotype. Proc. Natl. Acad.Sci. USA 104: 16233–16238.
53. Van, P. L., and A. K. Abbas. 1998. Homeostasis and self-tolerance in the immunesystem: turning lymphocytes off. Science 280: 243–248.
54. Molldrem, J. J., P. P. Lee, S. Kant, E. Wieder, W. Jiang, S. Lu, C. Wang, andM. M. Davis. 2003. Chronic myelogenous leukemia shapes host immunity byselective deletion of high-avidity leukemia-specific T cells. J. Clin. Invest. 111:639–647.
55. Lichterfeld, M., X. G. Yu, S. K. Mui, K. L. Williams, A. Trocha,M. A. Brockman, R. L. Allgaier, M. T. Waring, T. Koibuchi, M. N. Johnston,et al. 2007. Selective depletion of high-avidity human immunodeficiency virustype 1 (HIV-1)-specific CD8� T cells after early HIV-1 infection. J. Virol. 81:4199–4214.
56. Almeida, J. R., D. A. Price, L. Papagno, Z. A. Arkoub, D. Sauce, E. Bornstein,T. E. Asher, A. Samri, A. Schnuriger, I. Theodorou, et al. 2007. Superior controlof HIV-1 replication by CD8� T cells is reflected by their avidity, polyfunction-ality, and clonal turnover. J. Exp. Med. 204: 2473–2485.
57. De Boer, R. J., D. Homann, and A. S. Perelson. 2003. Different dynamics ofCD4� and CD8� T cell responses during and after acute lymphocytic chorio-meningitis virus infection. J. Immunol. 171: 3928–3935.
58. Obar, J. J., K. M. Khanna, and L. Lefrancois. 2008. Endogenous naive CD8� Tcell precursor frequency regulates primary and memory responses to infection.Immunity 28: 859–869.
1532 INTRINSIC PROPERTIES OF HLA MOLECULES AND IMMUNODOMINANCE




















