
Gene profiling in white blood cells predicts infliximab responsiveness in rheumatoid arthritis
Upload
mayoclinicCategory
view
2download
0
of February 9, 2016.This information is current as
Implications for Rheumatoid ArthritisArthritis in HLA-DQ8-Transgenic Mice:Susceptibility/Protection to Collagen-Induced CD4 and CD8 T Cells in
Chella S. DavidMarshall Behrens, Marie Griffiths, Harvinder Luthra and Veena Taneja, Neelam Taneja, Tawatchai Paisansinsup,
http://www.jimmunol.org/content/168/11/5867doi: 10.4049/jimmunol.168.11.5867
2002; 168:5867-5875; ;J Immunol
Referenceshttp://www.jimmunol.org/content/168/11/5867.full#ref-list-1
, 24 of which you can access for free at: cites 51 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2002 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on February 9, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on February 9, 2016
http://ww
w.jim
munol.org/
Dow
nloaded from

CD4 and CD8 T Cells in Susceptibility/Protection toCollagen-Induced Arthritis in HLA-DQ8-Transgenic Mice:Implications for Rheumatoid Arthritis 1
Veena Taneja,2* Neelam Taneja,‡ Tawatchai Paisansinsup,† Marshall Behrens,*Marie Griffiths, § Harvinder Luthra, † and Chella S. David*
To investigate the role of CD4 and CD8 T cells in arthritis, we generated transgenic mice deficient in CD4 and CD8 moleculesexpressing RA-susceptible geneHLA-DQ8. DQ8�CD4�/� mice were resistant to developing collagen-induced arthritis (CIA).However, DQ8�CD8�/� mice developed CIA with increased incidence and more severity than DQ8 mice. Both DQ8�CD8�/� andDQ8 mice produced rheumatoid factor. In addition, DQ8�CD8�/� mice produced antinuclear Abs. The B cell compartment andexpression of DQ8 were normal in all the strains, although frequency of cells expressing DQ8 was less in CD4�/� mice. Anincreased frequency of CD3� double-negative (DN) T cells was found in DQ8�CD8�/� compared with DQ8�CD4�/� and DQ8 mice.These CD3� DN T cells produced high amounts of IL-10 in CD8-deficient mice. Analysis of cell division using a cell cycle trackingdye showed a higher rate of division of CD3� and CD3� DN T cells in DQ8�CD8�/� mice compared with DQ8�CD4�/� and DQ8mice. Decreased apoptosis was seen in CIA-susceptible DQ8 and CD8-deficient mice, indicating a defect in activation-induced celldeath. These observations suggest that CD4 cells are necessary for initiation of CIA in DQ8 mice. We hypothesize that CD8� Tcells are not capable of initiating CIA in DQ8-transgenic mice but may have a regulatory/protective effect. The Journal ofImmunology, 2002, 168: 5867–5875.
R heumatoid arthritis (RA)3 is an autoimmune disordercharacterized by synovial inflammation and erosion ofbone and cartilage leading to destruction of joints. Genes
in the MHC have been shown to contribute to the RA predisposi-tion (1–2). While linkage studies have reported HLA-DRB1*0401association with susceptibility to RA (3–7), there are reports on therole of DR4 haplotype-associated DQB1 alleles, DQB1*0301 and*0302, in RA predisposition (8–10). Published data are consistentwith an involvement of T cells, especially CD4� T cells, in patho-genesis of RA. These include 1) the infiltration of inflamed syno-vium with predominantly T cells, 2) improvement in joint diseasemanifestations of arthritis following treatment with depleting CD4mAb, and 3) multiple oligoclonally expanded CD4 T cells withinthe rheumatoid joint (11–15). Recent studies suggest that RA pa-tients carry expanded CD4 clonotypes, which are not restricted tothe joint but are also found in periphery, thus indicating a systemicnature of the disease (16, 17). The observation that CD8 clono-types are detected in synovial fluids while CD4� clonal expan-sions have been found predominantly in periphery has led to the
notion that CD8 T cells may play a significant role in RA patho-genesis (17, 18). T cells lacking both CD4 and CD8 surface mark-ers have also been isolated from synovial fluid of RA patients (19).Oligoclonal expansion of T cells in RA joints is thought to bedriven in part by type II collagen (CII), a putative RA-associatedautoantigen, and T cells reactive to CII may be mediators of RApathogenesis (20–22).
Collagen-induced arthritis (CIA) is an experimental model ofautoimmune inflammatory polyarthritis sharing clinical and patho-logical features with RA (23). Susceptibility to CIA is associatedwith MHC class II polymorphism (H2Aq and H2Ar), suggesting animportant role of MHC-restricted T cell in development of disease(24). CII-specific CD4� T cells have been reported to be funda-mental in initiation and perpetuation of the disease (25, 26) and areessential for transferring arthritis into SCID mice (27). Furtherevidence is provided by the observations that treatment of CIAmice with either anti-TCR Ab (28) or anti-CD4 Ab (29) abrogatesdisease. In contrast, CII-reactive CD4� T cells have been reportedin some circumstances to protect against CIA (30). DBA/1�mCD4-deficient mice in a model of CIA were susceptible to CIA, sug-gesting that, in the absence of CD4� T cells, CD8 and double-negative (DN) T cells could play a role in initiation of disease (31).While CD8� T cells have been shown to be crucial in the onset ofseveral experimental autoimmune disease models (32–34), thecontribution of CD8� T cells in CIA has yet to be defined.
In this study, we investigated the role of CD4� and CD8�
T cells in initiation of CIA in transgenic mice expressing aRA-associated HLA class II molecule, DQ8 (DQA1*0301,DQB1*0302). Previous studies from our laboratory (35) haveshown that A�o�DQ8 mice elicit a vigorous CD4-mediated, DQ8-restricted cellular response following immunization using CIIwhich progresses to a severe form of CIA. To investigate whetherCD8� T cells or any other cell can initiate CIA in A�o�DQ8 micein the absence of CD4 expression, we generated A�o�DQ8 mice
*Department of Immunology and †Division of Rheumatology, Mayo Clinic, Roch-ester, MN 55905; ‡Department of Radiation Oncology, University of Chicago, Chi-cago, IL 60637; and §Research Services, Veterans Administration Medical Center andDepartment of Medicine, University of Utah, Salt Lake City, UT 84148
Received for publication January 22, 2002. Accepted for publication March 25, 2002.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This study was supported by National Institutes of Health Grants AR 30752 and AI14764. V.T. is the recipient of an Arthritis Foundation’s Investigator Award.2 Address correspondence and reprint requests to Dr. Veena Taneja, Department ofImmunology, Mayo Clinic, 200 First Street Southwest, Rochester, MN 55905. E-mailaddress: [email protected] Abbreviations used in this paper: RA, rheumatoid arthritis; CIA, collagen-inducedarthritis; CII, type II collagen; MsII, mouse CII; RF, rheumatoid factor; ANA, anti-nuclear Ab; AICD, activation-induced cell death; DN, double negative; LNC, lymphnode cell.
The Journal of Immunology
Copyright © 2002 by The American Association of Immunologists 0022-1767/02/$02.00
by guest on February 9, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

deficient in mCD4 (DQ8�CD4�/�). To delineate the role of CD8 Tcells we also generated A�o�DQ8 mice deficient in mCD8 mole-cules (DQ8�CD8�/�). While DQ8�mCD4�/� mice were com-pletely resistant to CIA, DQ8�CD8�/� mice developed a severeform of CIA with earlier onset than DQ8 mice, suggesting thatCD8� T cells may have a protective or regulatory role in arthri-togenic processes.
Materials and MethodsMice
A�o�DQ8�CD4�/� and A�o�DQ8�CD8�/� mice were generated by matingA�o�DQ8 mice in B6/129 background with mCD4-deficient and mCD8-deficient mice (generously provided by Dr. T. Mak, University of Toronto,Toronto, Ontario, Canada), also in B6/129 background, respectively. TheF1 mice were intercrossed two to four generations to generateA�o�DQ8�CD4�/� and A�o�DQ8�CD8�/� mice. Thus, all the mice used inthis study had a similar B6/129 background. For all the groups, parentalmice and negative littermates were included as controls. All mice weretyped for phenotypic expression of DQ8, CD4, and CD8 molecules.
Mice of both sexes (8–12 wk of age) used in this study were bred andmaintained in the pathogen-free Immunogenetics Mouse Colony at theMayo Clinic (Rochester, MN). All mice used in this study lack endogenousclass II molecules (A�o).
Flow cytometry
Expression of HLA-DQ, H2A, CD3, CD4, CD8, pan-B, and TCR V� chainmolecules on PBLs was analyzed by flow cytometry using FACS IV (BDBiosciences, Mountain View, CA) as described earlier (36). Abs used forstaining are IVD12 (anti-DQB1), HB163 (anti-Ab), GK1.5 (anti-CD4),53.6.8 (anti-CD8), MR9-4 (anti-V�5.1), MR9-8 (anti-V�5.1.2), 44-22-1(anti-V�6), F23.1 (anti-V�8.1.2.3), KJ-16 (anti-V�8.1.2), F23.2 (anti-V�8.2), and KT11 (anti-V�11).
Induction and evaluation of CIA
Pure native chick CII was obtained by multiple step purification describedpreviously (37). To induce CIA, 8- to 12-wk-old transgenic mice and neg-ative littermates were immunized with 100 �g of CII emulsified 1:1 withCFA H37Ra (100 �l; Difco, Detroit, MI) intradermally at the base of thetail. Animals received a booster of 100 �g of CII emulsified in IFA (100�l) 28 days later. Mice were monitored for the onset and progression ofCIA from 3 to 12 wk postimmunization. The arthritic severity of mice wasevaluated as described previously with a grading system for each paw from0 to 3 (24). The mean arthritic score was determined using arthriticanimals only.
Histopathology
Mice were sacrificed after 10–12 wk of immunization and kidneys wereextracted and snap-frozen in OCT tissue-embedding medium (Sakura Fi-netek, Torrance, CA). Cryostat sections were prepared, fixed, and stainedwith H&E. Paws from these mice were also fixed and sections were stainedwith H&E.
Anti-collagen Abs
Mice were bled on day 35 postimmunization, and the level of anti-mouseCII and anti-chick CII Abs in sera were determined using a standardELISA technique as described previously (38). Briefly, microtiter plateswere coated overnight with chick or mouse CII (6 �g/well in KPO4, pH7.6) at 4°C, washed, and blocked with 1% BSA in PBS/0.05% Tween 20.Sera were added in 4-fold dilutions (1/100–1/65,000) and incubated over-night at 4°C. The plates were washed and peroxidase-conjugated goat anti-mouse IgG (Organon Teknika, West Chester, PA) was added for anotherovernight incubation at 4°C. After washing, O-phenylenediamine wasadded and the colorimetric change was measured at 410 nm. All assayswere performed in duplicate and were quantified against a standard curveobtained with a known positive sera, arbitrarily determined to equal 100 Abunits per milliliter.
Rheumatoid factor
Rheumatoid factor (RF) was measured by ELISA as previously described(39). Briefly, ELISA plates were coated with rabbit IgG overnight at 4°C.After washing, sera (1/40 dilution) were added and incubated for 45 min atroom temperature and washed five times with PBS containing 0.05%Tween 20. Subsequently, wells were incubated with HRP-conjugated rab-
bit anti-mouse IgG (Fc specific; Pierce, Rockford, IL) or rabbit anti-mouseIgM (�-chain specific; Pierce) for 1 h and washed. 3,3�,5,5�-Tetramethyl-benzidine substrate (Sigma-Aldrich, St. Louis, MO) was added, and theabsorbance spectrum was determined with automated spectrophotometer(Bio-Rad, Hercules, CA). Sera from MRL/lpr (39) and B6 mice were usedas positive and negative controls, respectively. Sera from the following groupsof mice were also tested concurrently: DQ8 naive mice, littermates negativefor transgene, B10.Q mice immunized with CII, and DQ8�CD8�/� miceimmunized with only CFA.
Antinuclear Abs
Hep-2 cell line slides (Bio-Rad) were used as substrates for antinuclear Ab(ANA) detection. Sera diluted (1/100) in PBS containing 1% BSA wereadded, incubated for 1 h at room temperature, and washed three times inPBS for 15 min. Bound Abs were detected with FITC-coupled goat anti-mouse IgG (Accurate Chemical & Scientific, Westbury, NY). Stainedslides were washed three times in PBS for 15 min and examined by flu-orescence microscopy. Autoantibodies to ssDNA, dsDNA, and histonewere determined by ELISA as previously described (40).
In vitro T cell proliferation
Mice were immunized with 200 �g of CII emulsified 1:1 in CFA (Difco)intradermally at the base of the tail and one hind footpad. Ten days postim-munization, draining popliteal, caudal, and lumbar lymph nodes were re-moved and prepared for in vitro culture. A total of 1 � 106 lymph nodecells (LNCs) were challenged by adding 100 �l of medium (negative con-trol), Con A (20 �g/ml, positive control), and native collagen (50 �g/ml).For inhibition experiments, culture supernatant containing mAb (25 �g/mlAb) GK1.5 (anti-CD4), IVD12 (anti-HLA-DQ), or Lyt2 (anti-CD8) wasadded to the cells challenged in vitro with CII at 50 �g/ml. The cells wereincubated for 48 h at 37°C. During the last 18 h the cells were pulsed with[3H]thymidine and the tritium incorporation was determined by liquid scin-tillation counting. Results are calculated as �cpm (mean cpm of triplicatecultures containing Ag � mean cpm of medium).
Measurement of cytokines
Capture ELISA was done for measuring cytokines IFN-� and IL-4 (from Gen-zyme, Cambridge, MA), and TNF-�, IL-10, and IL-6 (from BD PharMingen,San Diego, CA) in culture supernatants using commercial kits.
Cell division and cytokine production byCD3�CD4�CD8� cells
Mice were sacrificed on days 7, 14, and 21 after immunization with CII.For every experiment, pooled LNCs from two to three mice were collectedand stained with conjugated Abs CD3-FITC, CD4-PE, CD8-PE, andNK1.1-PE mAbs (BD PharMingen) according to the manufacturer’s in-structions and then sorted by FACS IV (BD Biosciences) for FITC�PE�
cells. CD3� DN cells were collected from both naive and immunized miceand cultured for further analysis. For cell division, an equal number ofCD3� DN cells were stained with cell division tracking dye, CFSE (41),and cultured in vitro for 72–96 h with or without CII. Culture supernatantswere collected and assessed for cytokines.
AICD
To study the sensitivity of the proliferating cells to CII-induced activation-induced cell death (AICD), CD3� cells were stained with annexin V con-jugated with FITC after in vitro stimulation with CII and analyzedby FACS.
Statistical analysis
Difference in incidence of arthritis between groups was analyzed using the�2 test with Yates’ correction. Ab levels and means scores for arthritic micewere compared using Student’s t test. Differences in T cell types (CD3�
DN, NK1.1) between groups was analyzed using the �2 test.
ResultsCD4-deficient mice are resistant to CIA
Transgenic HLA-DQ8 and DQ8�CD4�/� mice were immunizedwith CII and followed for onset and progression of arthritis for 12wk. The DQ8 mCD4�/� mice did not show any evidence of ar-thritis either phenotypically or histologically, although 70% ofDQ8 mice exhibited severe arthritis. There was no evidenceof disease or infiltration of mononuclear cells in the synovia of
5868 CD8 T CELLS AS IMMUNOREGULATORY CELLS IN CIA IN HLA-DQ8 MICE
by guest on February 9, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

DQ8�CD4�/� mice (Fig. 1). In addition, while all the immunizedDQ8 mice produced anti-CII Abs, none of the DQ8�CD4�/� miceproduced anti-CII Abs, although the B cell numbers in CD4-defi-cient and DQ8 mice are comparable (50 and 56%, respectively;mean values from 10 mice in each strain).
Increased susceptibility to the development of CIA andautoantibodies in CD8-deficient DQ8-transgenic mice
Both DQ8 and DQ8�CD8�/� mice were immunized with CII andfollowed for development of arthritis. Both strains developed se-vere arthritis, although a higher incidence (70 vs 95%, respec-tively; p � 0.05) and an earlier onset (mean � SD onset: 41 � 12vs 35 � 10 days, respectively) of arthritis was detected in the lattergroup (Fig. 2A). A more severe disease was observed in CD8-deficient mice compared with DQ8 mice ( p � 0.05) (Fig. 2B). Todetermine the Ab responses to CII, sera were collected and Abs toCII and mouse CII (MsII) were assayed by ELISA. All mice pro-duced anti-CII and anti-MsII Abs, although the levels were sig-nificantly higher in the DQ8�CD8�/� mice ( p � 0.0008) (Fig. 2C).Mice immunized with adjuvant alone produced no detectable anti-collagen Abs (data not shown).
Transgenic CIA-susceptible mice produce RF
To further determine the clinical relevance of this animal modeland the fact that these mice carry human gene, sera from thesemice were tested for the presence of RF. The result showed thatboth DQ8- and DQ8�CD8�/�-immunized mice produced IgM andIgG RF. The incidence and level of IgM-RF were not significantlydifferent between DQ8�CD8�/� (74%, 25 of 34) and DQ8 (67%,10 of 15) mice. However, more DQ8�CD8�/� mice were positivefor IgG-RF (28 of 34) than DQ8 (6 of 15, p � 0.004) mice. More-over, the levels of IgG-RF were significantly higher inDQ8�CD8�/� compared with DQ8 mice ( p � 0.005) (Fig. 3). Incontrast, none of the naive DQ8, DQ8�CD4�/�, transgene negativelittermates, A�o, and DQ8�CD8�/�-transgenic mice immunizedwith adjuvant alone produced a detectable level of RF. CIA-sus-ceptible B10.Q mice also failed to develop RF following immu-nization with CII (data not shown).
ANAs are present in CD8-deficient mice
Sera obtained at day 35 postimmunization with CII or adjuvantwere used to study ANAs. All DQ8�CD8�/� sera (35 of 35) testedpositive for ANAs with homogenous pattern while none of theDQ8 and DQ8�CD4�/� sera elicited ANA staining (Fig. 4). Inaddition, while DQ8 and DQ8�CD4�/� mice did not produce de-tectable levels of ssDNA, dsDNA, or anti-histone Abs, 66% ofDQ8�CD8�/� mice produced anti-histone Abs and 43% ofDQ8�CD8�/� mice produced anti-dsDNA Abs (Fig. 4). However,
none of the mice positive for ANAs developed proteinuria andglomerulonephritis (data not shown).
CD4 T cells are required for T cell response to CII
LNCs were harvested from mice primed with CII and cultured invitro in the presence or absence of collagen. DQ8 andDQ8�CD8�/� mice gave a robust response to immunizing collagen(Fig. 5A). Inhibition assays using specific Abs demonstrated thatthe response to CII in these mice was mediated by CD4� T cellsand restricted by DQ8 molecules (Fig. 5B). DQ8�CD4�/� micemounted minimal response to CII in vitro, thus indicating that CD4T cells are required to initiate response against CII.
FIGURE 1. Histopathology of hind paws from DQ8�CD4�/� andDQ8�CD8�/� mice. A, H&E staining of a representative hind paw fromDQ8�CD8�/� mice showing complete destruction of the synovial lining(arrow) and infiltration of cells in the synovium. B, No abnormality orinfiltration of cells was seen in hind paws from DQ8�CD4�/� mice.
FIGURE 2. A, Incidence and onset of arthritis after immunization withCII showed an earlier onset of arthritis with increased incidence inDQ8�CD8�/� mice compared with DQ8 mice (p � 0.05), whileDQ8�CD4�/� mice did not develop disease. B, Severity of arthritis wasgraded as described in Materials and Methods. Arthritic score for eachmouse was calculated as the sum of arthritic severity of all four paws.Mean arthritic severity was calculated at the termination of the experimentat 12 wk postimmunization using arthritic mice only. DQ8�CD8�/� micedeveloped much more severe disease than DQ8 mice. C, Abs to self (MsII)and priming CII in all three DQ8-transgenic strains. Sera at day 35 fromprimed mice were used to study anti-CII Abs. Bars depict mean � SD(units per milliliter) of anti-collagen Abs from arthritic mice only.
5869The Journal of Immunology
by guest on February 9, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

Analysis of T cell types
To determine whether there was any alteration in T cell popula-tions in our CD4 and CD8 knockout mice, we analyzed the numberof different CD3� T cell populations. Total numbers of CD3� cellswere decreased in DQ8�CD4�/� and CD8�/� mice by �15 and5%, respectively, when compared with DQ8 mice (Fig. 6). Of theCD3� cells, �80% were positive for CD4 in DQ8�CD8�/� miceand 90% were positive for CD8 in DQ8�CD4�/� mice, 20 and 10%being CD3� DN, respectively. Expression of DQ8 was sim-
ilar in all the transgenics as observed by FACS analysis, althoughthe number of spleen cells positive for DQ molecule was lower inCD4-deficient mice than in the other two strains (Fig. 7). Thenumber of cells positive for CD5 and CD40 were lower inDQ8�CD4�/� mice compared with DQ8 and DQ8�CD8�/�
mice, although differences reached statistical significance onlyfor CD5 (DQ8 vs DQ8�CD4�/�, p � 0.01; DQ8�CD4�/� vsDQ8�CD8�/�, p � 0.05). Interestingly, activation marker CD69was seen with similar frequency in primed mice of all threestrains.
We also investigated the V� usage by T cells selected by thetransgenics. All the strains represented most of the V�, althoughthe DQ8�CD4�/� had a decreased frequency of all the V� thatreflects the lower number of total CD3 T cells (data not shown).
Role of CD3� DN T cells in arthritis
A significant role of CD3� DN T cells has been suggested forinitiation of CIA (29). To determine the role of these cells in ini-tiation/progression of arthritis, we studied CD3� DN cells in allmice included in the study. Cells were sorted for CD3�CD4�
CD8� T cells by FACS. Characterization of CD3� T cells showedan increased number of CD4 and DN cells in DQ8�CD8�/� micecompared with the DQ8 mice. DQ8�CD4�/� mice had more CD8�
T cells and CD3� DN cells than did DQ8 mice. However, CD3�
DN cells in DQ8�CD4�/� mice were significantly lower in numberthan those in DQ8�CD8�/� mice. Similar results were observed fornaive and immunized mice (Table I). Moreover, the single-posi-tive cells decreased in LNCs, while a concomitant increase inCD3� DN T cells up to 14 days postimmunization with CII wasseen. After 21 days of immunization, CD3� DN T cells de-creased in all mice except DQ8�CD8�/�, in which they werepersistently elevated and comprised 35– 45% of CD3� cells.
FIGURE 3. RF was tested in sera collected at day 35 after immunizationfor all transgenic, B10.Q, and A�o mice immunized with CII. Sera fromMRL/lpr and B6 mice were used as positive and negative controls, respec-tively. Naive DQ8 mice, DQ8�CD8�/� mice immunized with only CFA,and transgene-negative littermates were also included as controls. Hor-izontal bars for each strain indicate mean values of IgG-RF (A) andIgM-RF (B).
FIGURE 4. The sera obtained at day 35 after immunization with CIIwere used to study for ANAs. Hep-2 cell line was used as substrate forANA detection (upper panel). Only DQ8�CD8�/� mice showed positiveANA. Representative data from a mouse from each group are shown. In-cidence of specific ANAs against ssDNA, dsDNA, and histone in sera ofDQ8, DQ8�CD4�/�, and DQ8�CD8�/� mice were analyzed by ELISA(lower panel). Individual sera were titered with the end points at which theOD readings were about three times greater than those of mice treated withadjuvant alone (cut-off titer for positive value was 0.129).
FIGURE 5. In vitro response of LNCs to CII from primed mice. A, Alltransgenic mice were immunized with CII/CFA. LNCs were harvested onday 10 and cultured in the presence or absence of CII (50 �g/ml). Theresults shown are representative of five experiments. The data are reportedas �cpm. SDs were �15% in all experiments. The background cpm were2240 � 229, 1659 � 408, and 5193 � 560 in DQ8, DQ8�CD4�/�, andDQ8�CD8�/� mice, respectively. B, Inhibition of response was measuredfrom LNCs cultured as above in the presence of one of the Abs specific forDQ (IVD12), mCD4 (GK1.5), and mCD8 (Lyt2). The results are shown asthe percentage of inhibition of response by each specific Ab.
5870 CD8 T CELLS AS IMMUNOREGULATORY CELLS IN CIA IN HLA-DQ8 MICE
by guest on February 9, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

We further analyzed the number of CD3�NK1.1� cells in thesemice. Interestingly, both DQ8 and DQ8�CD8�/� mice had highernumber of these cells (10%) compared with DQ8�CD4�/� (3%),
although the difference did not reach statistical significance.NK1.1� cells increased in number up to 2 wk after immunizationwith a subsequent decrease.
FIGURE 6. Characterization of CD3� T cells.Splenic cells were purified, labeled with conjugatedAbs specific for CD4, CD8, and CD3, and analyzed byFACS. For DN cells, CD3� cells were gated and dotblots were generated by plotting CD4 and CD8 on thex and y axes, respectively. The circled area showsCD3� DN cells in each strain.
FIGURE 7. Splenic cells were labeled with MHC class II-specific mAbs and detected with a specific FITC-conjugated mAbs. The data shown wereobtained from spleen cells pooled from two mice per strain and represented at least three separate assays. Costimulatory molecules CD40 and CD5 andactivation marker CD69 were analyzed from purified spleen cells after labeling with specific mAbs.
5871The Journal of Immunology
by guest on February 9, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

Susceptible mice have a defect in AICD
To study the kinetics of cell division, splenic CD3� and CD3� DNT (sorted from LNCs) cells were isolated from primed mice at days7, 14, and 21, and challenged in vitro with CII after staining themwith CFSE. Higher numbers of dividing CD3� T cells at day 14were observed in DQ8 and DQ8�CD8�/� mice as compared withDQ8�CD4�/� mice after 48 h in response to CII (Fig. 8A). Asimilar phenomenon was seen in 7- and 21-day primed mice (datanot shown). CD3� DN T cells isolated from mice after 14 days ofimmunization also showed a higher number of cells undergoingmultiple divisions in DQ8�CD8�/� and DQ8 mice (80 and 60%,respectively), whereas only 45% cells underwent division in CD4-deficient mice (Fig. 8B). Characterization of the CD3� DN cellsshowed a high number of cells being positive for B220. InDQ8�CD8�/� mice, the CD3�B220� DN cells proliferated ac-tively after immunization. In contrast, there was no significant in-crease in proliferation of CD3�B220� DN cells after immuniza-tion in the other two strains (Fig. 8C).
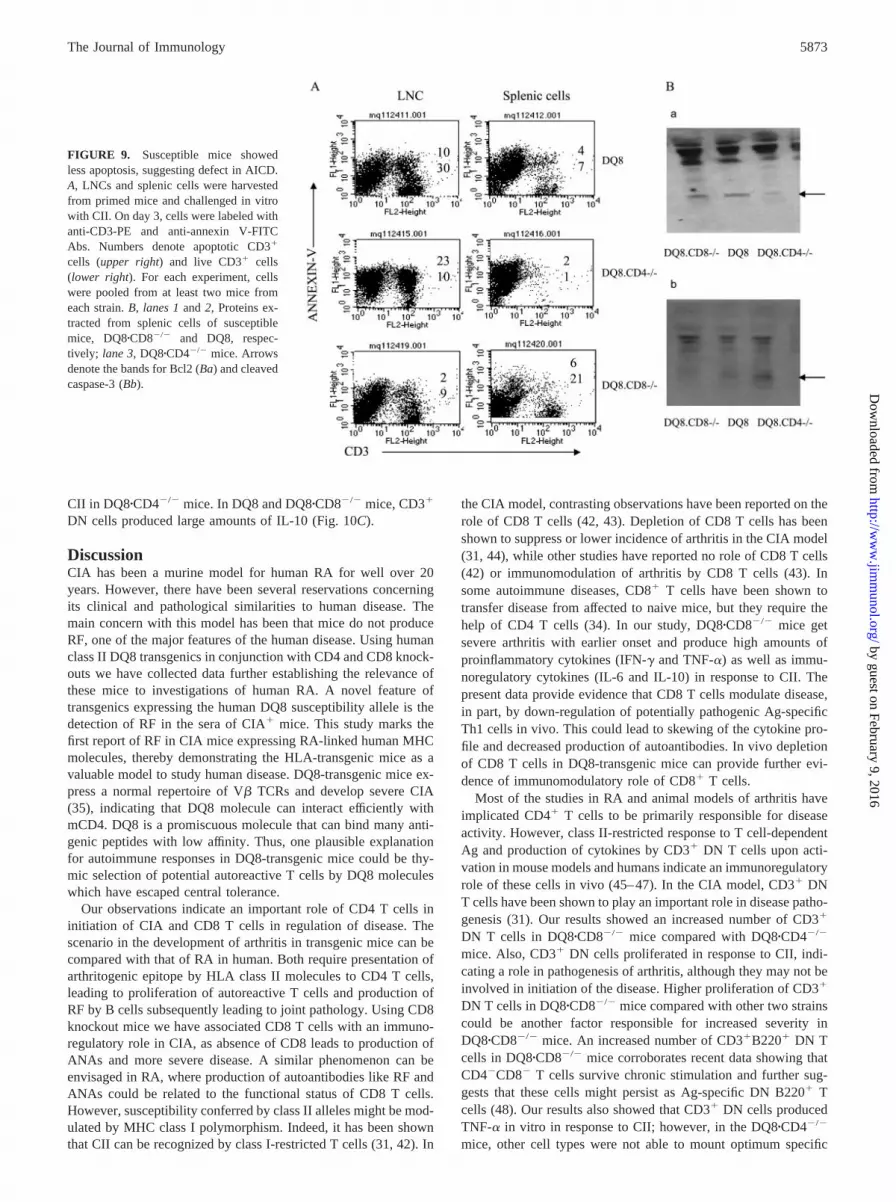
We further investigated whether there is a defect in AICD inmice susceptible to develop arthritis. Splenic cells and LNCs wereisolated from all the primed transgenic mice and cultured in thepresence of CII for 96 h and then stained with annexin V. Highernumbers of apoptotic cells were observed in CIA-resistant micecompared with those of CIA-susceptible mice. Seventy percent ofthe lymph node CD3� T cells were annexin-positive inDQ8�CD4�/� mice, while only 20–25% of CD3� T cells fromDQ8 and DQ8�CD8�/� mice stained positive for annexin. De-creased apoptosis indicates a defect in AICD in DQ8 andDQ8�CD8�/� mice. Similar results were observed with splenicCD3� T cells (Fig. 9A). To confirm the above results, we analyzedthe levels of Bcl2 and caspase-3 in primed mice. Both DQ8 andDQ8�CD8�/� mice had higher Bcl2, with lower caspase-3 levelscompared with DQ8�CD4-deficient mice (Fig. 9B).
CIA-resistant DQ8�CD4�/� mice can produce Th2 cytokines
TNF-� and IFN-� (Th1) cytokines, IL-4 and IL-10 (Th2) cyto-kines, and IL-6 (regulatory) cytokines were studied in all the testand control mice included in this study. High levels of IFN-� andTNF-� were produced by DQ8 and DQ8�CD8�/� mice (Fig. 10A).No detectable levels of IFN-� were observed in DQ8�CD4�/�
mice, although some IL-4 were produced. Both DQ8�CD4�/� andDQ8�CD8�/� mice produced IL-6, although there was a consistenttrend toward increased levels of IL-6 production in DQ8�CD8�/�
compared with DQ8�CD4�/� and DQ8 mice. However, differencesdid not reach statistical significance (Fig. 10B). There was a lateproduction of IL-10 in DQ8�CD4�/� mice after 21 days of immu-nization and 96 h in vitro culture only, while in DQ8 andCD8�DQ8�/� mice IL-10 was detected after 7 days of immuniza-tion. CD3� DN cells did not produce IFN-� in all mouse strains,although a large amount of TNF-� was produced in response to
FIGURE 8. CFSE-labeled LNCs were cultured in vitro with CII. Onday 3, CFSE profiles of activated cells were determined using flow cytom-etry. A, Cells were stained with CD3-FITC and CFSE profiles were gen-erated from CD3� gated cells. DQ8 (solid line) and DQ8�CD8�/� (dashedline) cells proliferated at comparable rates, whereas DQ8�CD4�/� (dottedline) cells proliferated at a slower rate. B, CD3� DN cells were sorted fromLNCs of primed mice. Equal numbers of CFSE-labeled cells were culturedas above and CFSE profiles were determined using flow cytometry. CD3�
DN cells from DQ8�CD8�/� mice underwent more divisions than the othertwo strains. C, Spleen cells were stained for CD3, CD4, CD8, and B220.CD3� DN T cells were gated and analyzed for the presence of B220. Barsdenote the percentage of CD3�B220� DN cells of the total CD3� DN Tcells in each strain.
Table I. Percentage of CD3� DN and NK1.1 cells in DQ8 and DQ8�CD4�/� and DQ8�CD8�/� micea
CD3�CD4�CD8� NK1.1
Naive Day 7 Day 14 Day 21 Day 7 Day 14 Day 21
DQ8 1.7 5.2 4.2 3.6 10.3 9.9 3.0DQ8�CD4�/�b 8.5 18.5 14.5 13.1 3.9 6.0 1.8DQ8�CD8�/�c 11.9 23.9 23.1 36.8 11.3 10.4 5.1
a Equal numbers of cells isolated from LNCs of naive or primed mice were harvested on days 7, 14, and 21 postimmunization with CII and stained with conjugated AbsCD3-FITC, CD4-PE, CD8-PE, and NK1.1-PE. Numbers indicate the percentage of the CD3�CD4�CD8� (FITC�PE�) and NK1.1 (PE�FITC�) cells sorted at indicated times.Differences in NK1.1 did not reach statistical significance.
b DQ8�CD4�/� vs DQ8, p � 0.05 for all comparisons.c For CD3� DN cells, DQ8�CD8�/� vs DQ8, p � 0.005 for all comparisons and for DQ8�CD8�/� vs DQ8�CD4�/�, p � 0.01 for days 14 and 21.
5872 CD8 T CELLS AS IMMUNOREGULATORY CELLS IN CIA IN HLA-DQ8 MICE
by guest on February 9, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
CII in DQ8�CD4�/� mice. In DQ8 and DQ8�CD8�/� mice, CD3�
DN cells produced large amounts of IL-10 (Fig. 10C).
DiscussionCIA has been a murine model for human RA for well over 20years. However, there have been several reservations concerningits clinical and pathological similarities to human disease. Themain concern with this model has been that mice do not produceRF, one of the major features of the human disease. Using humanclass II DQ8 transgenics in conjunction with CD4 and CD8 knock-outs we have collected data further establishing the relevance ofthese mice to investigations of human RA. A novel feature oftransgenics expressing the human DQ8 susceptibility allele is thedetection of RF in the sera of CIA� mice. This study marks thefirst report of RF in CIA mice expressing RA-linked human MHCmolecules, thereby demonstrating the HLA-transgenic mice as avaluable model to study human disease. DQ8-transgenic mice ex-press a normal repertoire of V� TCRs and develop severe CIA(35), indicating that DQ8 molecule can interact efficiently withmCD4. DQ8 is a promiscuous molecule that can bind many anti-genic peptides with low affinity. Thus, one plausible explanationfor autoimmune responses in DQ8-transgenic mice could be thy-mic selection of potential autoreactive T cells by DQ8 moleculeswhich have escaped central tolerance.
Our observations indicate an important role of CD4 T cells ininitiation of CIA and CD8 T cells in regulation of disease. Thescenario in the development of arthritis in transgenic mice can becompared with that of RA in human. Both require presentation ofarthritogenic epitope by HLA class II molecules to CD4 T cells,leading to proliferation of autoreactive T cells and production ofRF by B cells subsequently leading to joint pathology. Using CD8knockout mice we have associated CD8 T cells with an immuno-regulatory role in CIA, as absence of CD8 leads to production ofANAs and more severe disease. A similar phenomenon can beenvisaged in RA, where production of autoantibodies like RF andANAs could be related to the functional status of CD8 T cells.However, susceptibility conferred by class II alleles might be mod-ulated by MHC class I polymorphism. Indeed, it has been shownthat CII can be recognized by class I-restricted T cells (31, 42). In
the CIA model, contrasting observations have been reported on therole of CD8 T cells (42, 43). Depletion of CD8 T cells has beenshown to suppress or lower incidence of arthritis in the CIA model(31, 44), while other studies have reported no role of CD8 T cells(42) or immunomodulation of arthritis by CD8 T cells (43). Insome autoimmune diseases, CD8� T cells have been shown totransfer disease from affected to naive mice, but they require thehelp of CD4 T cells (34). In our study, DQ8�CD8�/� mice getsevere arthritis with earlier onset and produce high amounts ofproinflammatory cytokines (IFN-� and TNF-�) as well as immu-noregulatory cytokines (IL-6 and IL-10) in response to CII. Thepresent data provide evidence that CD8 T cells modulate disease,in part, by down-regulation of potentially pathogenic Ag-specificTh1 cells in vivo. This could lead to skewing of the cytokine pro-file and decreased production of autoantibodies. In vivo depletionof CD8 T cells in DQ8-transgenic mice can provide further evi-dence of immunomodulatory role of CD8� T cells.
Most of the studies in RA and animal models of arthritis haveimplicated CD4� T cells to be primarily responsible for diseaseactivity. However, class II-restricted response to T cell-dependentAg and production of cytokines by CD3� DN T cells upon acti-vation in mouse models and humans indicate an immunoregulatoryrole of these cells in vivo (45–47). In the CIA model, CD3� DNT cells have been shown to play an important role in disease patho-genesis (31). Our results showed an increased number of CD3�
DN T cells in DQ8�CD8�/� mice compared with DQ8�CD4�/�
mice. Also, CD3� DN cells proliferated in response to CII, indi-cating a role in pathogenesis of arthritis, although they may not beinvolved in initiation of the disease. Higher proliferation of CD3�
DN T cells in DQ8�CD8�/� mice compared with other two strainscould be another factor responsible for increased severity inDQ8�CD8�/� mice. An increased number of CD3�B220� DN Tcells in DQ8�CD8�/� mice corroborates recent data showing thatCD4�CD8� T cells survive chronic stimulation and further sug-gests that these cells might persist as Ag-specific DN B220� Tcells (48). Our results also showed that CD3� DN cells producedTNF-� in vitro in response to CII; however, in the DQ8�CD4�/�
mice, other cell types were not able to mount optimum specific
FIGURE 9. Susceptible mice showedless apoptosis, suggesting defect in AICD.A, LNCs and splenic cells were harvestedfrom primed mice and challenged in vitrowith CII. On day 3, cells were labeled withanti-CD3-PE and anti-annexin V-FITCAbs. Numbers denote apoptotic CD3�
cells (upper right) and live CD3� cells(lower right). For each experiment, cellswere pooled from at least two mice fromeach strain. B, lanes 1 and 2, Proteins ex-tracted from splenic cells of susceptiblemice, DQ8�CD8�/� and DQ8, respec-tively; lane 3, DQ8�CD4�/� mice. Arrowsdenote the bands for Bcl2 (Ba) and cleavedcaspase-3 (Bb).
5873The Journal of Immunology
by guest on February 9, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

response and produce the right cytokine milieu needed to initiatedevelopment of arthritis.
Our data are consistent with a significant role for CD4 T cells ininitiation of CIA pathogenesis (25–27). We observed an importantrole of CD4� T cells in initiation of arthritis in DQ8-transgenicmice because CD8� T cells as well as CD3� DN cells were notable to initiate CIA in DQ8�CD4�/� mice. In addition,DQ8�CD4�/� mice do not present CII as efficiently as the othertwo strains. However, our results are in contrast with a study inDBA/1 mice suggesting no role of CD4� T cells (31). Unlike CIAin most of the susceptible strains of mice, DBA/1 mice can spon-taneously develop arthritis that phenotypically resembles autoim-mune arthritis but is T cell independent (49) and is therefore notCD4 dependent and MHC class II restricted. Thus, CIA in DBA/1mice might involve both classical and nonclassical pathways fordisease development. DQ8�CD4�/� mice do not produce Abs toCII and IFN-� in vitro, indicating that CIA in transgenic mice ismuch more complex and requires CII-specific CD4 T cell interac-tion with B cells for disease initiation. Low T cell proliferationwith no IFN-� production and lack of autoantibodies are respon-sible for resistance to developing arthritis in DQ8�CD4�/� mice.Even though DQ8�CD4�/� mice did produce IL-6, there was no Igproduction by B lymphocytes, indicating either that there was adefect in B cell function in these mice or that IL-6 did not play a
direct role in initiation/pathogenesis but might be involved in pro-gression of disease. Ohshima et al. (50) demonstrated that IL-6knockout mice develop a more modest form of CIA, which sup-ports the notion that IL-6 may not be involved directly in diseasepathogenesis.
Oligoclonal CD4� T cells from arthritic patients have beenshown to have defects in apoptotic pathways (51). CD3� T cellsfrom CIA-resistant DQ8�CD4�/� mice showed enhanced apopto-sis compared with susceptible DQ8 and DQ8�CD8�/� mice. De-creased apoptosis in susceptible mice was confirmed by poorcleavage of caspase-3, a hallmark of apoptosis. Increased expres-sion of antiapoptotic protein, Bcl2, in CIA-susceptible mice cor-relates with decreased apoptosis in RA patients. This indicates apossible defect in AICD in mice developing arthritis. Activation ofCD3� T cells with subsequent resistance to apoptosis may con-tribute to CIA pathology.
Our data bring forth the following points: 1) transgenic miceexpressing the RA-associated HLA-DQ8 gene develop CIA me-diated by CD4 T cells and also produce RF similar to RA, 2)CD3� DN T cells can undergo chronic stimulation and may persistas Ag-specific T cells, 3) expansion following exposure to Ag aswell as a defect in AICD might lead to pathology seen in RA andCIA, and 4) CD8 T cells play a regulatory role, as their absenceleads to production of higher levels of autoantibodies.
AcknowledgmentsWe thank Dr. Chris Krco for critical comments on the manuscript, JulieHanson and Tad Trejo for maintaining transgenic mice, and Michele Smartfor screening the transgenic mice.
References1. Goldstein, R., and F. C. Arnett. 1987. The genetics of rheumatic disease in man.
Rheum. Dis. Clin. North Am. 13:587.2. Nepom, G. T. 1989. Determinants of genetic susceptibility in HLA-associated
autoimmune disease. Clin. Immunol. Immunopathol. 53:S53.3. Stastny, P. 1978. Association of the B-cell alloantigen DRw4 with rheumatoid
arthritis. N. Engl. J. Med. 298:869.4. Gregersen, P. K., J. Silver, and R. J. Winchester. 1987. The shared epitope hy-
pothesis: an approach to understanding the molecular genetics of susceptibility torheumatoid arthritis. Arthritis Rheum. 30:1205.
5. Wordsworth, B. P., J. S. Lanchbury, L. I. Sakkas, K. I. Welsh, G. S. Panayi, andJ. I. Bell. 1989. HLA-DR4 subtype frequencies in rheumatoid arthritis indicatethat DRB1 is the major susceptibility locus within the HLA class II region. Proc.Natl. Acad Sci. USA 86:10049.
6. Winchester, R., E. Dawyer, and S. Rose. 1992. The genetic basis of rheumatoidarthritis: the shared epitope hypothesis. Rheum. Dis. Clin. North Am. 18:761.
7. Taneja, V., M. J. Giphart, W. Verduijn, A. Naipal, A. N. Malaviya, andN. K. Mehra. 1996. Polymorphism of HLA-DRB, -DQA1, and -DQB1 in rheu-matoid arthritis in Asian Indians: association with DRB1*0405 and DRB1*1001.Hum. Immunol. 46:35.
8. Singal, D. P., M. D’ Souza, B. Reid, W. G. Benson, Y. B. Kassam, andJ. D. Adachi. 1987. HLA-DQ �-chain polymorphism in HLA-DR4 haplotypesassociated with rheumatoid arthritis. Lancet 2:1118.
9. Lanchbury, J. S., L. I. Sakkas, S. G. Marsh, J. G. Bodmer, K. I. Welsh, andG. S. Panayi. 1989. HLA-DQ�3.1 allele is a determinant of susceptibility toDR4-associated rheumatoid arthritis. Hum. Immunol. 26:59.
10. Taneja, V., N. K. Mehra, A. N. Chadreshekaran, R. K. Ahuja, Y. N. Singh, andA. N. Malaviya. 1992. HLA-DR4-DQw8 but not DR4-DQw7 haplotypes occur inIndian patients with rheumatoid arthritis. Rheumatol. Int. 11:251.
11. Van Boxel, J. A., and S. A. Paget. 1975. Predominantly T-cell infiltrate in rheu-matoid synovial membranes. N. Engl. J. Med. 293:517.
12. Herzog, C., C. Walker, W. Muller, P. Rieber, C. Reiter, G. Riethmuller,P. Wassmer, H. Stockinger, O. Madic, and W. J. Pichler. 1989. Anti-CD4 anti-body treatment of patients with rheumatoid arthritis. I. Effect on clinical courseand circulating T cells. J. Autoimmun. 2:627.
13. Walker, C., C. Herzog, P. Rieber, G. Riethmuller, W. Muller, and W. J. Pichler.1989. Anti-CD4 antibody treatment of patients with rheumatoid arthritis. II. Ef-fect of in vivo treatment on in vitro proliferative response of CD4 cells.J. Autoimmun. 2:643.
14. Strober, S., and J. Holoshitz. 1990. Mechanisms of immune injury in rheumatoidarthritis: evidence for the involvement of T cells and heat-shock proteins. Immu-nol. Rev. 118:233.
15. Khazaei, H. A., C. Lunardi, and A. K. So. 1995. CD4 T cells in the rheumatoidjoint are oligoclonally activated and change during the course of disease. Ann.Rheum. Dis. 54:314.
FIGURE 10. Cytokines produced by CD3� cells isolated from primedmice and cultured in vitro in the presence or absence of CII. A, LNCs fromprimed mice were cultured in vitro as described in Fig. 5. For each exper-iment, cells were pooled from at least two mice per strain. The data arerepresentative of at least three separate assays. Supernatants were collectedafter 48 h and cytokines were measured by ELISA for Th1, IFN-�, andTNF-� (A), Th2/regulatory, IL-4, IL-10, and IL-6 (B). Very low amountsof cytokines, except for IL-6, were detected in DQ8�CD4�/� mice. C,CD3� DN cells were sorted and cultured in vitro as described in Materialsand Methods. IFN-�, TNF-�, and IL-10 cytokines produced in vitro weremeasured in all the strains. None of the strains produced IFN-� (data notshown), whereas TNF-� was produced by all strains. Only DQ8 andDQ8�CD8�/� mice produced IL-10.
5874 CD8 T CELLS AS IMMUNOREGULATORY CELLS IN CIA IN HLA-DQ8 MICE
by guest on February 9, 2016http://w
ww
.jimm
unol.org/D
ownloaded from

16. Goronzy, J. J., P. Bartz-Bazzanella, W. Hu, M. C. Jendro, D. R. Walser-Kuntz,and C. M. Weyand. 1994. Dominant clonotypes in the repertoire of peripheralCD4� T cells in rheumatoid arthritis. J. Clin. Invest. 94:2068.
17. Gonzalez-Quintial, R., R. Baccala, R. M. Pope, and A. N. Theofilopoulos. 1996.Identification of clonally expanded T cells in rheumatoid arthritis using a se-quence enrichment nuclease assay. J. Clin. Invest. 97:1335.
18. Fitzgerald, J. E., N. S. Ricalton, A. C. Meyer, S. G. West, H. Kaplan, C. Behrendt,and B. L. Kotzin. 1995. Analysis of clonal CD8� T cell expansions in normalindividuals and patients with rheumatoid arthritis. J. Immunol. 154:3538.
19. Holoshitz, J., F. Koning, J. E. Coligan, J. De Bruyn, and S. Strober. 1989. Iso-lation of CD4�CD8� mycobacteria-reactive T lymphocyte clones from rheuma-toid arthritis synovial fluid. Nature 339:226.
20. Londei, M., C. M. Savill, A. Verhoef, F. Brennan, Z. A. Leech, V. Duance,R. N. Maini, and M. Feldmann. 1989. Persistence of collagen type II-specificT-cell clones in the synovial membrane of a patient with rheumatoid arthritis.Proc. Natl. Acad. Sci. USA 86:636.
21. Sekine, T., T. Kato, K. Masuko-Hongo, H. Nakamura, S. Yoshino, K. Nishioka,and K. Yamamoto. 1999. Type II collagen is a target antigen of clonally ex-panded T cells in the synovium of patients with rheumatoid arthritis. Ann. Rheum.Dis. 58:446.
22. He, X., A. Kang, and J. M. Stuart. 2000. Accumulation of T cells reactive to typeII collagen in synovial fluid of patients with rheumatoid arthritis. J. Rheumatol.27:589.
23. Trentham, D. E., A. S. Townes, and A. H. Kang. 1977. Autoimmunity to type IIcollagen: an experimental model of arthritis. J. Exp. Med. 146:857.
24. Wooley, P. H., H. S. Luthra, J. M. Stuart, and C. S. David. 1981. Type II col-lagen-induced arthritis in mice. I. Major histocompatibility complex (I region)linkage and Ab correlates. J. Exp. Med. 154:688.
25. Mauri, C., C. Q. Chu, D. Woodrow, L. Mori, and M. Londei. 1997. Treatment ofa newly established transgenic model of chronic arthritis with nondepleting anti-CD4 monoclonal antibodies. J. Immunol. 159:5032.
26. Doncarli, A., L. M. Stasiuk, C. Fournier, and O. Abehsira-Amar. 1997. Conver-sion in vivo from an early dominant Th0/Th1 response to a Th2 phenotype duringthe development of collagen-induced arthritis. Eur. J. Immunol. 27:1451.
27. Kadowaki, K. M., H. Matsuno, H. Tsuji, and I. Tunru. 1994. CD4� T cells fromcollagen-induced arthritic mice are essential to transfer arthritis into severe com-bined immunodeficient mice. Clin. Exp. Immunol. 97:212.
28. Yoshino, S., and L. G. Cleland. 1992. Depletion of �/� T cells by a monoclonalAb against the �/� T cell receptor suppresses established adjuvant arthritis, butnot established collagen-induced arthritis in rats. J. Exp. Med. 175:907.
29. Chu, C. Q., and M. Londei. 1996. Induction of Th2 cytokines and control ofcollagen-induced arthritis by nondepleting anti-CD4 antibodies. J. Immunol. 157:2685.
30. Kakimoto, K., M. Katsuki, T. Hirofuji, H. Iwata, and T. Koga. 1988. Isolation ofT cell line capable of protecting mice against collagen-induced arthritis. J. Im-munol. 140:78.
31. Tada, Y., A. Ho, D. K. Koh, and T. W. Mak. 1996. Collagen-induced arthritis inCD4- or CD8-deficient mice. J. Immunol. 156:4520.
32. Wang, B., A. Gonzalez, C. Benoist, and D. Mathis. 1996. The role of CD8� Tcells in the initiation of insulin-dependent diabetes mellitus. Eur. J. Immunol.26:1762.
33. Zhang, G. X., B. G. Xiao, M. Bakhiet, P. van der Meide, H. Wigzell, H. Link, andT. Olsson. 1996. Both CD4� and CD8� T cells are essential to induce experi-mental autoimmune myasthenia gravis. J. Exp. Med. 184:349.
34. Deming, S., J. N. Whitaker, Z. Huang, D. Liu, C. Coleclough, H. Wekerle, andC. S. Raine. 2001. Myelin antigen-specific CD8� T cells are encephalitogenicand produce severe disease in C57BL/6 mice. J. Immunol. 166:7579.
35. Nabozny, G. H., J. S. Baisch, D. Cheng, D. Cosgrove, M. M. Griffiths,H. S. Luthra, and C. S. David. 1996. HLA-DQ8 transgenic mice are highlysusceptible to collagen-induced arthritis: a novel model for human polyarthritis.J. Exp. Med. 183:27.
36. Taneja, V., M. M. Griffiths, H. S. Luthra, and C. S. David. 1998. Modulation ofHLA-DQ-restricted collagen-induced arthritis by HLA-DRB1 polymorphism.Int. Immunol. 10:1449.
37. Griffiths, M. M., E. J. Eichwald, J. H. Martin, C. B. Smith, and C. W. DeWitt.1981. Immunogenetic control of experimental type II collagen induced arthritis.Arthritis Rheum. 24:781.
38. Griffiths, M. M., G. H. Nabozny, J. Hanson, J. S. Harper, S. McCall, K. G. Moder,G. W. Cannon, H. S. Luthra, and C. S. David. 1994. Collagen-induced arthritisand TCRs in SWR and B10.Q mice expressing EaK transgene. J. Immunol. 153:2758.
39. Watson, M. L., J. K. Rao, G. S. Gilkeson, P. Ruiz, E. M. Eicher, D. S. Pisetsky,A. Matsuzawa, J. M. Rochelle, and M. F. Seldin. 1992. Genetic analysis ofMRL-lpr mice: relationship of the Fas apoptosis gene to disease manifestations.J. Exp. Med. 176:1645.
40. Paisansinsup, T., A. N. Vallejo, H. S. Luthra, and C. S. David. 2001. HLA-DRmodulates autoantibody repertoire, but not mortality, in a humanized mousemodel of systemic lupus erythematosus. J. Immunol. 167:4083.
41. Lyons, A. B. 1999. Divided we stand: tracking cell proliferation with carboxy-fluorescein diacetate succinimidyl ester. Immunol. Cell Biol. 77:509.
42. Gao, X.-M., and A. J. McMichael. 1996. Cytotoxic T lymphocytes specific formurine type II collagen do not trigger arthritis in B10 mice. Clin. Exp. Immunol.103:89.
43. Chiocchia, G., M. C. Boissier, B. Manoury, and C. Fournier. 1993. T cell regu-lation of collagen-induced arthritis in mice. II. Immunomodulation of arthritis bycytotoxic T cell hybridomas specific for type II collagen. Eur. J. Immunol. 23:327.
44. Arai, K., S. Yamamura, T. Hanyu, H. E. Takahashi, H. Umezu, H. Watanabe, andT. Abo. 1996. Extrathymic differentiation of resident T cells in the joints of micewith collagen-induced arthritis. J. Immunol. 157:5170.
45. Rahemtullah, A., T. M. Kundig, A. Narendran, M. F. Bachmann, M. Julius,C. J. Paige, P. S. Ohashi, Z. M. Zinkernagel, and T. W. Mak. 1994. Class II majorhistocompatibility complex-restricted T cell function in CD4-deficient mice. Eur.J. Immunol. 24:2213.
46. Zlotnik, A., D. I. Godfrey, M. Fischer, and T. Suda. 1992. Cytokine productionby mature and immature CD4�CD8� T cells: ���TCR�CD4�CD8� T cellsproduce IL-4. J. Immunol. 149:1211.
47. Niehues, T., D. Eichelbauer, and E. M. Schneider. 1999. Functional characteris-tics of human peripheral blood �/� TCR�, CD4� and CD8� double-negative(DN) T cells. Microbiol. Immunol. 43:153.
48. Hamad, A. R., A. Srkrishnan, P. Mirmonsef, P. M. Broeren, C. H. June,D. Pardoll, and J. P. Schneck. 2001. Lack of coreceptor allows survival of chron-ically stimulated double-negative �/� T cells: implications for autoimmunity.J. Exp. Med. 193:1113.
49. Corthay, A., A. S. Hansson, and R. Holmdahl. 2000. T lymphocytes are notrequired for the spontaneous development of entheseal ossification leading tomarginal ankylosis in the DBA/1 mouse. Arthritis Rheum. 43:844.
50. Ohshima, S., Y. Saeki, T. Mima, M. Sasai, K. Nishioka, S. Nomura, M. Kopf,Y. Katada, T. Tanaka, M. Suemura, and T. Kishimoto. 1998. Interleukin 6 playsa key role in the development of antigen-induced arthritis. Proc. Natl. Acad. Sci.USA 95:8222.
51. Vallejo, A. N., M. Schirmer, C. M. Weyand, and J. J. Goronzy. 2000. Clonalityand longevity of CD4�CD28null T cells are associated with defects in apoptoticpathways. J. Immunol. 165:6301.
5875The Journal of Immunology
by guest on February 9, 2016http://w
ww
.jimm
unol.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















