
A chicken leptin-specific radioimmunoassay
11
A chicken leptin-specific radioimmunoassay S. Dridi a , J. Williams a , V. Bruggeman b , M. Onagbesan b , N. Raver c , E. Decuypere b , J. Djiane d , A. Gertler c , M. Taouis a, * a Endocrinologie Mole ´culaire et Cellulaire du Me ´tabolisme, SRA, INRA Nouzilly 37380, France b Department of Animal Sciences, Catholic University, Leuven, Belgium c Institute of Biochemistry, Food Science and Nutrition, Faculty of Agriculture, The Hebrew University, Rehovot 76100, Israel d INRA, Domaine de Vilvert, 78352 Jouy-en-Josas, France Received 21 October 1999; accepted 12 January 2000 Abstract Recombinant chicken leptin was used to produce an antiserum in order to develop a specific and sensitive radioimmunoassay (RIA) for chicken leptin in plasma and serum. We have used either murine or chicken leptin as tracer and competition curves were performed using recombinant chicken leptin. Variations in leptin plasma levels in different chicken strains and various nutritional states were correlated with the physiological status. Leptin plasma concentrations were regulated by the nutritional state with higher levels in the fed state as compared to the fasted state (3.36 6 0.13 versus 2.78 6 0.11 ng/ml) and being dependent upon the age. Higher leptin levels were found in 22 week-old as compared to 15 week-old layer chickens (2.709 6 0.172 versus 1.478 6 0.102 ng/ml). We have also shown that the multispecies leptin RIA kit (LINCO Inc.) underestimated leptinemia compared to the chicken leptin- specific RIA reported here. In conclusion the RIA developed in the present study is specific to the chicken and thus may be considered as powerful tool for investigating the physiological significance of leptin in chickens. © 2000 Elsevier Science Inc. All rights reserved. Keywords: Leptin; Chicken; Adiposity * Corresponding author. Tel.: 133-2-47427929; fax: 133-2-47427778. E-mail address: [email protected] (M. Taouis). Domestic Animal Endocrinology 18 (2000) 325–335 0739-7240/00/$ – see front matter © 2000 Elsevier Science Inc. All rights reserved. PII: S0739-7240(00)00053-9
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of A chicken leptin-specific radioimmunoassay

A chicken leptin-specific radioimmunoassay
S. Dridia, J. Williamsa, V. Bruggemanb, M. Onagbesanb, N. Raverc,E. Decuypereb, J. Djianed, A. Gertlerc, M. Taouisa,*
aEndocrinologie Mole´culaire et Cellulaire du Me´tabolisme, SRA, INRA Nouzilly 37380, FrancebDepartment of Animal Sciences, Catholic University, Leuven, Belgium
cInstitute of Biochemistry, Food Science and Nutrition, Faculty of Agriculture, The Hebrew University,Rehovot 76100, Israel
dINRA, Domaine de Vilvert, 78352 Jouy-en-Josas, France
Received 21 October 1999; accepted 12 January 2000
Abstract
Recombinant chicken leptin was used to produce an antiserum in order to develop a specificand sensitive radioimmunoassay (RIA) for chicken leptin in plasma and serum. We have usedeither murine or chicken leptin as tracer and competition curves were performed usingrecombinant chicken leptin. Variations in leptin plasma levels in different chicken strains andvarious nutritional states were correlated with the physiological status. Leptin plasmaconcentrations were regulated by the nutritional state with higher levels in the fed state ascompared to the fasted state (3.366 0.13 versus 2.786 0.11 ng/ml) and being dependent uponthe age. Higher leptin levels were found in 22 week-old as compared to 15 week-old layerchickens (2.7096 0.172 versus 1.4786 0.102 ng/ml). We have also shown that the multispeciesleptin RIA kit (LINCO Inc.) underestimated leptinemia compared to the chicken leptin- specificRIA reported here. In conclusion the RIA developed in the present study is specific to the chickenand thus may be considered as powerful tool for investigating the physiological significance ofleptin in chickens. © 2000 Elsevier Science Inc. All rights reserved.
Keywords:Leptin; Chicken; Adiposity
* Corresponding author. Tel.:133-2-47427929; fax:133-2-47427778.E-mail address:[email protected] (M. Taouis).
Domestic Animal Endocrinology 18 (2000) 325–335
0739-7240/00/$ – see front matter © 2000 Elsevier Science Inc. All rights reserved.PII: S0739-7240(00)00053-9

1. Introduction
In mammals, leptin has been described as a satiety hormone produced almost exclusivelyby adipose tissue [1,2]. Recently, various studies have described the presence of leptinmRNA in the human placenta [3] and stomach [4]. Leptin is secreted into the bloodstreamas a 16 kDa protein and acts on hypothalamic leptin receptors which are most likely to beone of the key signal transducers controlling appetite. The effect of leptin on reducing foodintake has been confirmed by the administration of recombinant leptin to rodents and humans[5–11]. In these species, leptinemia is highly correlated to the fat pad size or to adiposity[12–14]. Furthermore, using leptinemia as criteria for selection, two strains of mice wereselected which showed a significant change in the level of adipose tissue deposition [15,16].Therefore, a radioimmunoassay (RIA) for leptin may be an important tool for the indirectestimation of adiposity and for the genetic selection of livestock. During the past few yearsseveral commercial kits for human, mouse or multi-species RIA for leptin (from Linco Inc.)have been described and can be used in farm animals such as sheep, cows and pigs. However,an RIA specific for chicken leptin has not been reported. Recently, chicken leptin has beencloned by our group [17] and confirmed by others [18]. Chicken leptin shows a highsimilarity to mammalian leptin but has an extra cysteine residue [17,18] that may affect itsconformational structure. We have shown that leptin is expressed not only in adipose tissuebut also in liver, indicating that hepatic expression of leptin in chicken may be associatedwith the important role of the liver in lipogenesis in this species. Furthermore, insulin,corticosterone and oestrogen [18] regulate chicken leptin expression in the liver. We haverecently prepared recombinant chicken leptin and its C4S analog and have demonstrated itsnegative effect on food intake in two chicken lines [19,20], indicating that the effect of leptinin reducing food intake is similar to that described in mammals. Therefore, the estimation ofleptin plasma levels is an important tool for understanding the physiological role of thishormone in the chicken. Here, we report the first homologous RIA for chicken leptin. Wehave evaluated this assay for accuracy, precision, sensitivity and linearity by comparisonwith the multispecies leptin RIA KIT (LINCO inc.) and also by using radio-labeled murineleptin as tracer.
2. Materials and methods
2.1. Production of recombinant chicken leptin
Recombinant chicken leptin was prepared as previously described [20]. Briefly, chickenleptin cDNA produced by PCR was subcloned into pMON3401 expression vector andexpressed in MON105 cells. Four hours after nalidixic acid induction, E. Coli MON105 cellsexpressed leptin (; 16 kDa) as a major cell protein. The recombinant chicken leptin waspurified using anion-exchange chromatography and its purity and monomer content wasverified by SDS-PAGE and gel-filtration chromatography on a Superdex column.
326 S. Dridi et al. / Domestic Animal Endocrinology 18 (2000) 325–335

2.2. Immunization and preparation of antiserum
One mg recombinant chicken leptin was dissolved in PBS with Freund’s completeadjuvant (Sigma, France) (1:1). Four rabbits were injected intradermally at several locationswith this mixture (100mL). Three weeks later the injection was repeated in the sameconditions except that Freund’s complete adjuvant was replaced by Freund’s incompleteadjuvant. The same treatment was continued at one and two weeks later. Serum was tested10 days following the last injection.
Four antisera (L7104, L7105, L7107, L7108) were tested for binding to iodinated chickenleptin. Chicken leptin was radiolabeled as described below. Antisera at various concentrationswere added in a total volume of 100mL to iodinated chicken leptin (about 20000 cpm in 100mL)and 100mL of incubation buffer (0.05 M PBS, 0.1% BSA, 0.1 M NaN3, pH 7.4), this mixturewas incubated for 24 h at 4°C. Following the addition of goat anti-rabbit immunoglobulinantibodies in the presence of 4% Polyethylenglycol (PEG 6000) and 0.1% Tween 20 andcentrifugation (3000 g, 30 min at 4°C), the precipitated immune-complexes were counted.
2.3. Iodination of chicken leptin
Fivemg of chicken leptin was radio-iodinated (IMS 50, Pharmacia-Amersham, France) bythe chloramine T method [21]. Briefly, leptin (5mg/10ml) was diluted with 50ml of 0.5 Msodium phosphate buffer pH 7.4 in a 1.5 mL polypropylene conical tube. 300mCi of 125I(IMS30; Amersham Pharmacia BioTech, France) and 10mL of Chloramine T (3 mg/mL)were introduced and the reaction was allowed to proceed for 20 s then stopped by theaddition of 750ml of Sodium Metabisulfite (2.4 mg/mL) followed by 200ml of SodiumIodide (2 mg/mL). The contents of the tube were transferred to a Sephadex G25 PD-10column (Amersham Pharmacia BioTech, France) previously equilibrated with 25 mL ofincubation buffer (phosphate buffer 0.05 M, pH 7.6 containing 0.1% BSA, 0.15 M NaCl,0.1% Triton X-100 and 0.1% sodium azide). This buffer was used to elute approximately 500mL fractions into collection tubes containing 20mL of 25 mG/mL BSA. Forty fractions werenormally collected and 10mL samples counted on a gamma counter (RIAstar, PackardInstrument Co., Rungis, France) allowing the peak of label and the peak of free iodine to bedefined. The fractions comprising the peak of labeled leptin were checked for bindingcapacity with the antibody and for satisfactory non-specific binding (overnight incubation at4°C followed by separation of bound and free fractions) before use in the RIA. The specificactivity of 125I- chicken leptin was 52.7 mCi/mg and the yield of iodination was 77%.
2.4. Radioimmunoassay (RIA)
The RIA incubation mixture consisted of 100mL of increasing concentrations of recombinantchicken leptin diluted in incubation buffer (phosphate buffer 0.05 M, pH 7.6 containing 0.1%BSA, 0.15M NaCl, 0.1% Triton X-100 and 0.1% sodium azide), 200mL of a final 1: 5000dilution of anti-chicken leptin antiserum (IGg rabbit anti leptin) and 200mL of the incubationbuffer. The mixture was incubated for 24 h at 4°C, then 100mL of 125I-chicken leptin (;20000cpm) was added and the mixture was further incubated for 24 h at 4°C.
327S. Dridi et al. / Domestic Animal Endocrinology 18 (2000) 325–335

Following the addition of PEG (12.5%) and a goat anti-rabbit immunoglobulin antibodyand centrifugation (3750 g, 15 min, 4°C), the supernatants were discarded by aspiration andthe precipitates were counted. The same procedure was applied for the multispecies leptinRIA kit (LINCO) following supplier recommendations.
2.5. Specificity of chicken leptin radioimmunoassay
To demonstrate the specificity of the chicken leptin RIA, the radio-immunoassay wasperformed as described above except that125I chicken leptin binding was displaced byincreasing concentrations of chicken leptin, human IGF-1, human GH or porcine insulin (1pgto 250 ng/ml).
2.6. Administration of recombinant chicken leptin
In order to demonstrate that the developed RIA detects changes in circulating leptin, fiveweek-old ISA brown cockerels (about 600 g) received single intraperitoneal (i.p.) injectionof 1 mg leptin/kg (n 5 4). The control group (n 5 4) received a placebo. Plasma wascollected before and two hours after leptin or placebo treatment.
2.7. Effect of nutritional state and age on plasma leptin level
17 month-old male chickens (Ge´line de Touraine strain) (about 3.8 kg) were housed inindividual cages in standard conditions (temperature, humidity and light) with free access towater and balanced diet. Chickens were divided in two groups (n 5 4), the first was fastedfor 16 h and the second was fed ad libitum. On the day of experiment (16 H later), animalswere sacrificed and plasmas were collected.
Broiler breeder hens were grown in standard conditions with free access to water andbalanced diet. Breeder hens were divided into three groups that were sacrificed at differentages 15 (n 5 8), 18 (n 5 20) and 22 (age of the first egg,n 5 18) weeks old and plasma werethen collected.
2.8. Statistical analysis
Results are expressed as means6 SEM and statistical comparisons were performed usingStudent’st test. Linear regression analysis was performed with the method of best squares.
For RIA curve analysis Graph pad Software (Version 2, 94–95) was used.
3. Results
3.1. Testing antiserums directed toward chicken leptin
Following the immunization of four rabbits, antiserums were tested as described inmaterials and methods. We have studied the capacity of binding to125I chicken leptin of four
328 S. Dridi et al. / Domestic Animal Endocrinology 18 (2000) 325–335

antiserums: L7104, L7105, L7107 and L7108. Fig. 1 shows a dilution-dependent immuno-precipitation of125I chicken leptin for all antisera tested. At dilution of 1/5000, L7104,L7105, L7107 and L7108 precipitated 54.9%, 42.7%, 47.7% and 32.1% of total125Ichickenleptin added, respectively. This result indicates that the antiserum L7104 is the more
Fig. 2.Comparison of typical standard curves of chicken leptin radioimmunoassay (RIA). The RIA mixture, consistingof increasing concentrations of cold recombinant chicken leptin, antiserum directed against chicken leptin andincubation buffer, was pre-incubated for 24 h at 4°C prior to the addition of labeled chicken (●) or murine (l) leptinand the incubation was performed for an additional 24 h. The Multispecies RIA kit from LINCO Inc. (Œ) was usedas suggested by manufacturer. Each point of the curve represents the mean of triplicate measurements.
Fig. 1. Antiserum from four rabbits were tested for the immunoprecipitation of125I chicken leptin. Increasingdilutions (1/5000 to 1/400000) of L7104, L7105, L7107 and L7108 antiserums were added to125Ichicken leptin(20000 cpm), and following 24 h incubation at 4°C, the immune complexes were precipitated and radioactivitywas counted and reported as B/TA (specific binding/total activity added).
329S. Dridi et al. / Domestic Animal Endocrinology 18 (2000) 325–335

efficient, therefore this antiserum was chosen to develop the chicken leptin radioimmuno-assay.
3.2. Comparison of standard curves
We have determined the standard competition curves for chicken leptin radioimmunoas-say using iodinated recombinant chicken or murine leptin (as tracer), specific anti-chickenleptin antibodies and increasing concentrations of cold recombinant chicken leptin, asdescribed in Materials and Methods. We have compared the chicken specific RIA curve withthe multispecies leptin RIA kit (Linco Inc.) (Fig. 2). The IC50values (defined as theconcentration of unlabeled leptin capable of inhibiting 50% of specific binding to the leptinantibodies) varied between 3 to 8, 10 to 15 and 10 to 20 ng/ml for multispecies LINCO kit,murine and chicken125I leptin, respectively. The specific binding in the three assays variedfrom 30 to 55%.
3.3. Specificity of chicken leptin RIA
To show the specificity of chicken leptin RIA, increasing concentrations of humanGH, IGF-1 and insulin were used to displace125I chicken leptin binding to chickenleptin antibodies, and as positive control recombinant chicken leptin was used. Fig. 3
Fig. 3. Determination of the specificity of chicken leptin radioimmunoassay (RIA). The RIA mixtures, consistingof increasing concentrations of cold recombinant chicken leptin, human IGF-1, human GH or porcine insulin, andantiserum directed against chicken leptin and incubation buffer, were pre-incubated for 24 h at 4°C prior to theaddition of labeled chicken leptin. The incubation was performed for an additional 24 h. Each point of the curverepresents the mean of triplicate measurements.
330 S. Dridi et al. / Domestic Animal Endocrinology 18 (2000) 325–335

shows that GH, IGF-1 and insulin did not crossreact with chicken leptin RIA whencompared to recombinant chicken leptin which inhibits tracer binding with an EC50 of10 ng/ml.
3.4. Validation of specific chicken leptin radioimmunoassay
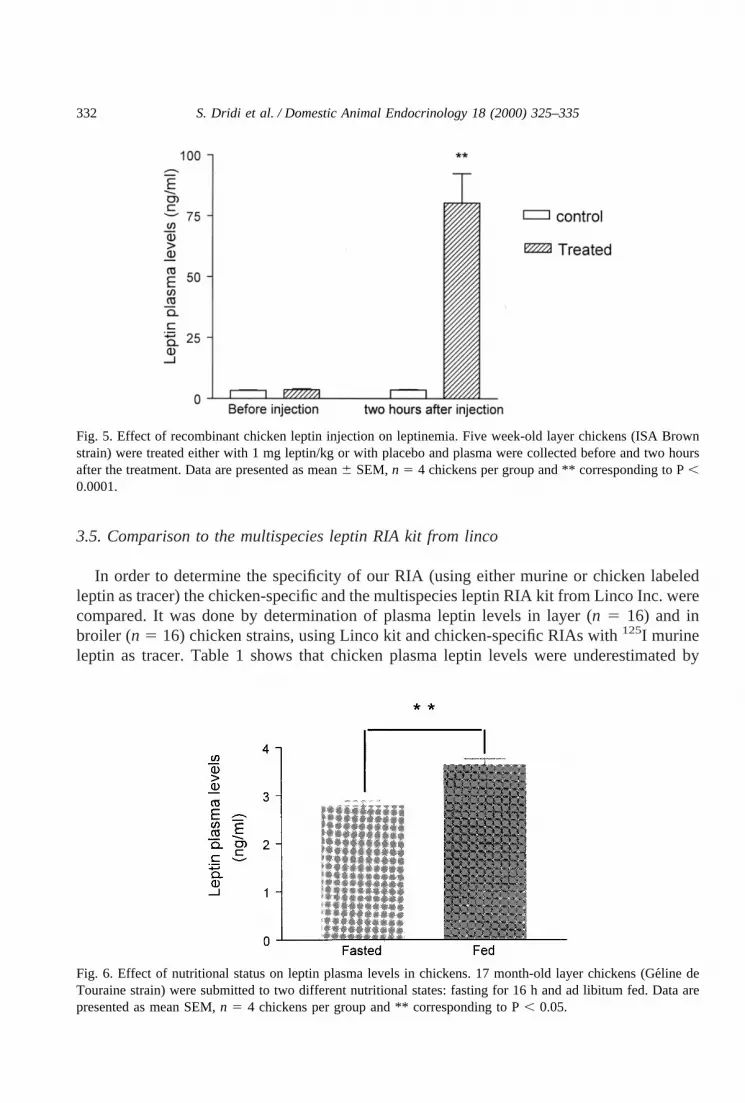
In order to demonstrate leptin recovery and specificity, five week-old chicks (ISA Brownstrain) were treated either with 1 mg leptin/kg or with placebo and plasma collected beforeand two hours after the treatment. Plasma from leptin treated chicken were diluted to 1, 1/2and 1/4, with RIA buffer (Fig. 4). Fig. 5 shows that plasma leptin concentrations were linearand correlated with the dilution factor (d15 83.35, d1/25 36.3 and d1/45 19.89 ng/ml).In addition, using the standard curve obtained with labeled murine leptin, we show asignificant increase, at least 23-fold, in leptin plasma levels in leptin treated chicks comparedto chicks treated with placebo (806 5.34 ng/ml versus 3.436 0.23 ng/ml; mean6 S.E.M,n 5 4) (Fig. 5) this indicates that this assay is able to detect leptin plasma variations. We havealso determined the impact of the nutritional state of chicken on plasma leptin levels of 17month-old layer chickens (Ge´line de Touraine strain). Fig. 6 shows that plasma leptin levelswere significantly (P, 0.05) reduced from 3.366 0.13 ng/ml (measured at the fed state) to2.786 0.11 ng/ml in the fasted state.
Fig. 4. Typical standard curve of chicken leptin RIA (●) and dilution curve of layer chicken plasma (Œ). Fiveweek old layer chickens (ISA brown strain) were treated with 1 mg leptin/kg and plasma was collected two hourslater and leptin levels determined in various plasma dilutions: d1, d1⁄2 and d1⁄4. Each experimental point of thecurve represents the mean6 SEM of triplicate measurements. Values reported on the figure represent the leptinplasma concentration for each dilution.
331S. Dridi et al. / Domestic Animal Endocrinology 18 (2000) 325–335
3.5. Comparison to the multispecies leptin RIA kit from linco
In order to determine the specificity of our RIA (using either murine or chicken labeledleptin as tracer) the chicken-specific and the multispecies leptin RIA kit from Linco Inc. werecompared. It was done by determination of plasma leptin levels in layer (n 5 16) and inbroiler (n 5 16) chicken strains, using Linco kit and chicken-specific RIAs with125I murineleptin as tracer. Table 1 shows that chicken plasma leptin levels were underestimated by
Fig. 5. Effect of recombinant chicken leptin injection on leptinemia. Five week-old layer chickens (ISA Brownstrain) were treated either with 1 mg leptin/kg or with placebo and plasma were collected before and two hoursafter the treatment. Data are presented as mean6 SEM, n 5 4 chickens per group and ** corresponding to P,0.0001.
Fig. 6. Effect of nutritional status on leptin plasma levels in chickens. 17 month-old layer chickens (Ge´line deTouraine strain) were submitted to two different nutritional states: fasting for 16 h and ad libitum fed. Data arepresented as mean SEM,n 5 4 chickens per group and ** corresponding to P, 0.05.
332 S. Dridi et al. / Domestic Animal Endocrinology 18 (2000) 325–335

about 30 to 56% in both strains by using the Linco kit. We have also compared the Linco kitto the chicken-specific assay using125I chicken leptin as tracer by measuring the leptinplasma levels of broiler breeder hens at various ages. Using our assay, we found that leptinplasma levels are 1.4786 0.102 ng/ml (n 5 11), 1.4856 0.057 ng/ml (n 5 40) and 2.70960.172 ng/ml (n 5 38) at 15 and 18 weeks of age and at the age of first egg, respectively(Table 2). Multispecies leptin RIA kit (LINCO), underestimated the leptin levels by 10 to40% (Table 2). However, using both assays leptin plasma levels were significantly higher inthe 22 week-old hens as compared to 15 or 18 week-old hens.
4. Discussion
Recombinant chicken leptin given by IP or IV reduced food intake in various chickenstrains [19,20]. This indicates a similar role of leptin in the chicken to that described inmammals as a regulator of appetite. Therefore, the determination of leptin plasma levels isa useful tool in the understanding of the control of food intake in the chicken. We have
Table 1Leptin plasma levels in layer (59-week-old) and broiler (9-week-old) chickens measured using eithermultispecies RIA kit (Linco Inc) or chicken leptin RIA (with chicken or murine leptin as tracer)a
Multispecies RIA kit(Linco Inc.)
125I chicken or murine leptin
Plasma leptin ng/ml (mean6 SEM)Layer hens 1.536 0.067 (n 5 16) 2.226 0.44 (n5 16)Broiler chicken 1.376 0.10 (n 5 16) 3.156 0.14 (n5 14)
Minimal quantity detectable 1 ng/ml 0.1 ng/mlEC50 3–7.6 ng/ml 10–15 ng/mlLinearity 1–50 ng/ml 0.5–100 ng/ml
(r2 5 0.981; p , 0.0001) (r2 5 0.958; P, 0.0001)CV 3–8% ,10%
a Minimal quantity detectable, EC50, linearity and CV (coefficient of variation) were determined using GraphPad software (Version 2.01).
Table 2Leptinemia variations upon age in broiler breeder hensa
Leptin plasma levels (ng/ml; mean6 SEM) n
Specific chicken RIA Multispecies RIA kit(Linco Inc.)
Animals: broiler breeder hens of15 week old 1.4786 0.102 1.3456 0.093 818 week old 1.4856 0.057 1.1966 0.050 2022 week old (at the First Egg) 2.7096 0.172** 1.5146 0.108* 18
a Leptin plasma levels were determined in 15, 18, and 22 week-old hens using specific leptin chicken RIA orMultispecies RIA kit (Linco Inc.). Results are presented as mean6 SEM, ** corresponding to P, 0.001 and * toP , 0.01, when leptinemia of 22-week-old hens was compared to that of 15- or 18-week-old hens.
333S. Dridi et al. / Domestic Animal Endocrinology 18 (2000) 325–335

developed a specific chicken leptin RIA based on using either iodinated chicken or murineleptin as tracer. It was shown that the standard curve is similar to that found using the LINCOkit. However, using our assay (with both chicken or murine leptin as tracer), leptin plasmalevels were consistently higher than those obtained with the Linco kit. This may be attributedto the use of specific antibodies directed against chicken leptin and recombinant chickenleptin for the standard curve. Despite its high conservation compared to that of mammals,chicken leptin has an extra cysteine that may result in a specific spatial configurationprobably leading to the creation of an additional antigenic sites that are absent in mammalianleptin. We have validated this assay by measuring leptin plasma levels in various chickenstrains and also in various physiological states. The variations observed are highly correlatedwith the nutritional status. Chicken leptin levels are lower in the fasted state than in the fedstate. Chicken leptin plasma levels increased with age in hens with a maximum level in adulthens at the first egg, which is also correlated with the developed adiposity at this stage. Thisstudy also demonstrates the accuracy of this assay when leptin plasma levels measured inrecombinant leptin treated chickens at various dilutions of the plasma. Moreover, the assaypermits the measurement of leptin plasma levels in ducks, pheasants and turkey (data notshown) indicating that this assay could be used for several if not all avian species.
In conclusion, this is the first study reporting a specific non-mammalian leptin radioim-munoassay, a specific chicken leptin RIA that may further be proven to be an avian RIA. Thisprovides a new tool that may be used as an indicator of chicken physiological state and/orfood intake level or capacity. Whether its physiological role is related to the cross talkbetween brain (appetite-satiety) and body metabolic status (fatness, liver lipogenic capacity)or perhaps related to still other physiological functions as a true endocrine or even paracrinefactor remains totally unknown and has to be further elucidated. This however could hardlybe done without the necessary tools for measuring being developed.
Acknowledgments
We are grateful to A. Bordas for providing the R chicken strain, we also thank M. Derouetfor his technical help and K. Geraert for animal care.
References
[1] ZhangY, Proenca R, Maffei M, Barone M, Leopold L, Freidman JM. positional cloning of the obese geneand its human homologue. Nature 1994;372:425–32.
[2] Tsuruo Y, Sato I, Lida M, Murakami T, Ishimura K, Shima K. Immunohistochemical detection of the obgene product (leptin) in rat white and brown adipocytes. Horm Metab Res 1996;28:753–5.
[3] Senaris R, Garcia-caballero T, Casabiell X, Gallego R, Castro R, Considine RV, Dieguez C, Casanueva FF.Synthesis of leptin in human placenta. Endocrinology 1997;138:4501–4.
[4] Bado A, Levasseur S, Attoub S, Kermorgant S, Laigneau JP, Bortoluzz MN, Moizo L, Lehy T, Guerre-MilloM, Le Marchand-BrustelY, Levin MJM. The stomach is a source of leptin. Nature 1998;394:790–3.
[5] Halaas JL, Gajiwala KS, Maffei M, Cohen SL, Chait BT, Robinowitz D., Lallone RL, Burley SK, FreidmanJM. Weight-reducing effects of the plasma protein encoded by the obese gene. Science 1995;269:543–6.
334 S. Dridi et al. / Domestic Animal Endocrinology 18 (2000) 325–335

[6] Pelleymounter MA, Cullen MJ, Baker MB, Hecht R, Winters D, Boone T, Collins F. Effects of the obesegene product on body weight regulation in ob/ob mice. Science 1995;269:540–3.
[7] Campfield LA, Smith FJ, Guisez Y, Devos R, Burn P. Recombinant mouse ob protein: Evidence forperipheral signal linking adiposity and central neural networks. Science 1995;269:546–9.
[8] Rentsch J, Levens N, Chiesi M. Recombinant ob-gene product reduces food intake in fasted mice. BiochemBiophys Res Commun 1995;214:131–6.
[9] Jacob RJ, Dziura J, Medwick MB, Leone P, Caprio S, During M, Shulman GI, Sherwin RS. The effect ofleptin is enhanced by microinjection into the ventromedial hypothalamus. Diabetes 46:1997:150–2.
[10] Wang Q, Bing C, Al-Barazangi K, Mossakowaska DE, Wang XM, McBay DL, Neville WA, Taddayon M,Pickavance L, Dryden S, Thomas ME, McHale MT, Gloyer IS, Wilson S, Buckingham R, Arch JR,Trayhurn P, Williams G. Interactions between leptin and hypothalamic neuropeptide Y neurons in thecontrol of food intake and energy homeostasis in the rats. Diabetes 46:1997:335–41.
[11] Cusin I, Rohner-Jeanrenaud F, Stricker-Krongrad A, Jeanrenaud B. The weight-reducing effect of anintracerebro-ventricular bolus injection of leptin in genetically obese fa/fa rats. Reduced sensitivity com-pared with lean animals. Diabetes 1996;45:1446–50.
[12] McGregor GP0, Desaga JF, Ehlenz K, Fisher A, Heese F, Hegele A, Lammer C, Peiser C, Lang RE.Radioimmunological measurement of leptin in plasma of obese and diabetic human subjects. Endocrinology1996;137:1501–4.
[13] Takahashi M, Funahashi T, Shimomura I, Miyaoka K, Matsuzawa Y. Plasma leptin levels and body fatdistribution. Horm Metab Res 1996;28:751–2.
[14] Moller N, O’brien P, Nair KS. Disruption of the relationship between fat content and leptin levels with agingin humans. JCEM 1998;38:931–4.
[15] Bunger L, Nicolson M, Hill WG. Leptin levels in lines of mice developed by long-term divergent selectionon fat content. Genet Res 1999:73:37–44.
[16] Bunger L, Hill WG. Effects of leptin administration on long-term selected fat mice. Genet Res 1997;:69:215–25.
[17] Taouis M, chen JW, Daviaud C, Dupont J, Derouet M, Simon J. Cloning the chicken leptin gene. Gene1998;208:239–42.
[18] Ashwell CM, Czerwinski SM, Brocht DM, McMurtry JP. Hormonal regulation of leptin expression inbroiler chicken. American Journal of Physiology 1999;276:R226–32.
[19] Dridi S., Raver N, Derouet M., Picard M., Gertler A, Taouis M. Preparation of C4S analogue of recombinantchicken leptin and comparison of its in vitro and in vivo biological activities with unmodified chicken andovine leptins. Am J Physiol 2000; in press.
[20] Raver N, Taouis M, Dridi S, Derouet M, Simon J, Robinzon B, Djiane J, Gertler A. Large-Scale. Preparationof biologically active recombinant chicken obese protein (leptin). Protein expression and purification1998;14:403–8.
[21] Sugawara A, Morii N, Sakamoto M, Suda M, Shimokura M, Kiso Y, Kihara M, Yamori Y, Nishimura K,Soneda J, Ban T, Nakao K. Alpha-human atrial natriuretic polypeptide is released from the heart andcirculates in the body. Biochem Biophys Res Commun 1984;129:439–46.
335S. Dridi et al. / Domestic Animal Endocrinology 18 (2000) 325–335




















