
Correction: Dynorphin Activates Quorum Sensing Quinolone Signaling in Pseudomonas aeruginosa

Molecular Microbiology (2006)
59
(4), 1114–1128 doi:10.1111/j.1365-2958.2005.05008.xFirst published online 12 December 2005
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd
Blackwell Science, LtdOxford, UKMMIMolecular Microbiology0950-382X© 2005 The Authors; Journal compilation © 2005 Blackwell Publishing Ltd
? 2005
59
411141128
Original Article
Quorum sensing-regulated DNA releaseM. Allesen-Holm
et al.
Accepted 15 November, 2005. *For correspondence. [email protected]; Tel. (
+
45) 45 25 27 93; Fax (
+
45) 45 8873 28.
A characterization of DNA release in
Pseudomonas aeruginosa
cultures and biofilms
Marie Allesen-Holm,
1
Kim Bundvig Barken,
1
Liang Yang,
1
Mikkel Klausen,
1
Jeremy S. Webb,
2
Staffan Kjelleberg,
2
Søren Molin,
1
Michael Givskov
1
and Tim Tolker-Nielsen
*
1
1
Centre for Biomedical Microbiology, BioCentrum-DTU, Technical University of Denmark, DK-2800 Lyngby, Denmark.
2
University of New South Wales, Sydney, NSW 2052, Australia.
Summary
Pseudomonas aeruginosa
produces extracellularDNA which functions as a cell-to-cell interconnectingmatrix component in biofilms. Comparison of extra-cellular DNA and chromosomal DNA by the use ofpolymerase chain reaction and Southern analysissuggested that the extracellular DNA is similar towhole-genome DNA. Evidence that the extracellularDNA in
P. aeruginosa
biofilms and cultures is gener-ated via lysis of a subpopulation of the bacteria wasobtained through experiments where extracellular
ββββ
-galactosidase released from
lacZ
-containing
P.aeruginosa
strains was assessed. Experiments withthe wild type and
lasIrhlI
,
pqsA
,
pqsL
and
fliMpilA
mutants indicated that the extracellular DNA is gen-erated via a mechanism which is dependent on acylhomoserine lactone and
Pseudomonas
quinolonesignalling, as well as on flagella and type IV pili.Microscopic investigation of flow chamber-grownwild-type
P. aeruginosa
biofilms stained with differentDNA stains suggested that the extracellular DNA islocated primarily in the stalks of mushroom-shapedmulticellular structures, with a high concentrationespecially in the outer part of the stalks forming aborder between the stalk-forming bacteria and thecap-forming bacteria. Biofilms formed by
lasIrhlI
,
pqsA
and
fliMpilA
mutants contained less extracellu-lar DNA than biofilms formed by the wild type, and themutant biofilms were more susceptible to treatmentwith sodium dodecyl sulphate than the wild-typebiofilm.
Introduction
Bacteria in natural, industrial and clinical settings predom-inantly live in surface-associated communities called bio-films (Costerton
et al
., 1995). Bacteria in biofilms areresistant to biocides, antibiotics and host immuneresponses, and biofilm formation therefore leads to a vari-ety of problems in industry and to chronic or lethal infec-tions in human and animals (Costerton
et al
., 1999).During the last 10 years there has been a rapidly increas-ing recognition of microbial biofilms as a highly significanttopic in microbiology with relevance for many importantareas in modern society. It has become apparent thatdetailed knowledge about the developmental processfrom single bacteria scattered on a surface to the forma-tion of thick structured biofilms is essential in order tocreate strategies to control biofilm development.
The opportunistic pathogen
Pseudomonas aeruginosa
has become a model organism in biofilm research. Ofconsiderable interest is the nature of the components thathold the bacteria together in biofilms, and the cues whichinduce their synthesis. It appears that extracellular DNA,exopolysaccharide and proteinaceous compounds can allfunction as matrix components in
P. aeruginosa
biofilms,and that their relative importance as structural componentmay depend on the environmental conditions, on theage of the biofilm and on the particular
P. aeruginosa
strain forming the biofilm. Evidence for a role of extra-cellular DNA as cell-to-cell interconnecting compoundin
P. aeruginosa
biofilms has been presented bothfor the
P. aeruginosa
PAO1 reference strain and forclinical
P. aeruginosa
isolates (Whitchurch
et al
., 2002;Nemoto
et al
., 2003). DNase treatment dissolved young
P. aeruginosa
PAO1 biofilms whereas established PAO1biofilms were only marginally affected by the DNase treat-ment, suggesting that the cells in young PAO1 biofilms areheld together by extracellular DNA whereas the cells inolder PAO1 biofilms are held together primarily by othercompounds (Whitchurch
et al
., 2002). In accord, Mat-sukawa and Greenberg (2004) recently reported that,although DNA makes up a substantial fraction of thematrix material in mature PAO1 biofilms, exopolysaccha-rides encoded by the
psl
genes appears to be the criticalmatrix component. A role for the
psl
-encoded exopolysac-charide as biofilm matrix component was also reported forthe
P. aeruginosa
ZK2870 strain (Friedman and Kolter,
Quorum sensing-regulated DNA release
1115
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd,
Molecular Microbiology
,
59
, 1114–1128
2004a). In addition, it was reported that the
P. aeruginosa
PA14 strain produces a glucose-containing exopolysac-charide encoded by the
pel
genes as the main structuralcomponent of the biofilm matrix (Friedman and Kolter,2004b). In contrast to the findings that extracellular DNAis not the primary cell-to-cell interconnecting compoundin mature biofilms formed by the
P. aeruginosa
referencestrains, Nemoto
et al
. (2003) found that mature biofilmsformed by four independent clinical
P. aeruginosa
isolatescould be dissolved by DNase treatment, suggesting thatextracellular DNA is the primary cell-to-cell interconnect-ing compound in mature biofilms formed by these
P. aeruginosa
strains. Long before biofilms became acentral research area a Japanese group reported thecharacterization of extracellular ‘slime’ produced by
P. aeruginosa
(Murakawa, 1973a,b). They investigatedthe chemical composition of slimes from 20 clinical
P. aeruginosa
isolates, and found that slimes from 18strains consisted primarily of DNA, while two strains witha mucoid phenotype produced slimes composed primarilyof polyuronic acid. Besides extracellular DNA andexopolysaccharide, proteinaceous polymers may beimportant matrix components. Evidence was recently pro-vided that cell appendages, termed CupA fimbria, are animportant constituent of the extracellular matrix for the
P. aeruginosa
strains PAK, PAO1 and PA14 (Vallet
et al
.,2001; D’Argenio
et al
., 2002; Friedman and Kolter,2004b). In addition, type IV pili have various conditionalroles in
P. aeruginosa
biofilm formation (O’Toole andKolter, 1998; Klausen
et al
., 2003a,b).Quorum sensing, a mechanism by which bacteria can
monitor their cell population density through the extracel-lular accumulation of signalling molecules, has beenshown to play a role in structural development and stabi-lization of
P. aeruginosa
biofilms (Davies
et al
., 1998).
P. aeruginosa
employs the quorum-sensing systems
lasRI
and
rhlRI
to control the expression of a wide range ofvirulence factors (Passador
et al
., 1993; Latifi
et al
., 1995;1996). The
rhlRI
system is to some extent controlled bythe
lasRI
system (Latifi
et al
., 1996). The signal moleculesinvolved are primarily 3-oxo-C12-homoserine lactone(3-oxo-C12-HSL) in case of the
lasRI
system, and prima-rily C4-HSL in case of the
rhlRI
system (Latifi
et al
., 1995;1996). Quorum-sensing systems involving such acylatedhomoserine lactone (AHL) signalling molecules can beinhibited by halogenated furanone compounds isolatedfrom the macroalgae
Delisea pulchra
(Givskov
et al
.,1996; Hentzer
et al
., 2002; 2003). A third signalling sys-tem based on 2-heptyl-3-hydroxy-4-quinolone, designatedthe
Pseudomonas
quinolone signal (PQS), has beenshown to be part of the quorum-sensing regulatory net-work in
P. aeruginosa
(Pesci
et al
., 1999). The productionof PQS depends on
lasR
(Pesci
et al
., 1999), and exoge-nous PQS strongly induces expression of elastase B and
rhlI
in a
lasR
mutant background (McKnight
et al
., 2000).The PQS signalling system is therefore believed to oper-ate between the
las
and the
rhl
systems in the quorum-sensing regulatory network (McKnight
et al
., 2000).In the present article we report a characterization of the
release of extracellular DNA in
P. aeruginosa
cultures andbiofilms. Although the study was initiated in order to gaininsight into the role of extracellular DNA in
P. aeruginosa
biofilm development, we found it worthwhile to beginwith a characterization of DNA release in planktonic
P. aeruginosa
cultures. The experiments with planktoniccultures are followed by studies of DNA release in micro-titre trays where the bacteria are present both in the plank-tonic and in the biofilm mode of growth. Finally, DNArelease and the spatial distribution of extracellular DNAare studied in flow chamber-grown biofilms.
Results
Extracellular DNA in
P. aeruginosa
cultures
The accumulation of extracellular DNA in the medium ingrowing
P. aeruginosa
PAO1 cultures was assessed byspectrophotometric measurements of light absorbance byDNA, and by fluorometric measurements of PicoGreenbound to DNA. Both methods showed that the
P. aeruginosa
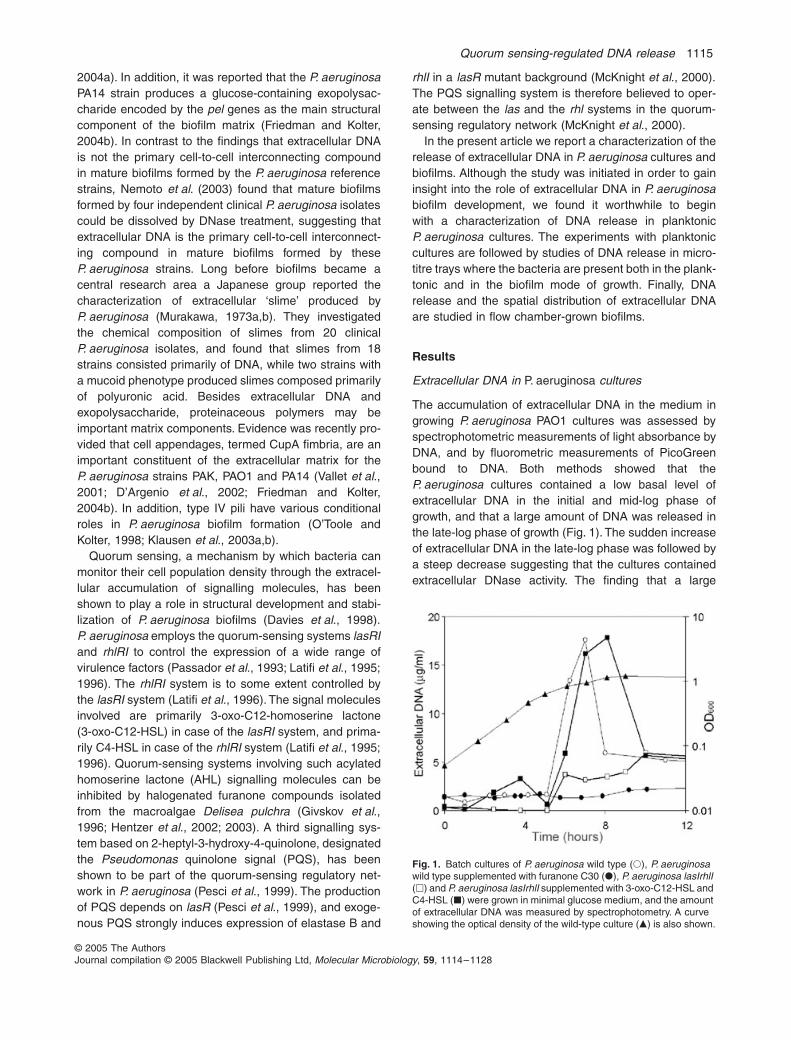
cultures contained a low basal level ofextracellular DNA in the initial and mid-log phase ofgrowth, and that a large amount of DNA was released inthe late-log phase of growth (Fig. 1). The sudden increaseof extracellular DNA in the late-log phase was followed bya steep decrease suggesting that the cultures containedextracellular DNase activity. The finding that a large
Fig. 1.
Batch cultures of
P. aeruginosa
wild type (
�
),
P. aeruginosa
wild type supplemented with furanone C30 (
�
),
P. aeruginosa lasIrhlI
(
�
) and
P. aeruginosa lasIrhlI
supplemented with 3-oxo-C12-HSL and C4-HSL (
�
) were grown in minimal glucose medium, and the amount of extracellular DNA was measured by spectrophotometry. A curve showing the optical density of the wild-type culture (
�
) is also shown.
1116
M. Allesen-Holm
et al.
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd,
Molecular Microbiology
,
59
, 1114–1128
amount of extracellular DNA was released specifically inthe late-log phase of growth suggested that quorum sens-ing might have a role in DNA release. Accordingly, mea-surements of the accumulation of extracellular DNA in themedium of growing
P. aeruginosa lasIrhlI
cultures showedthat the quorum-sensing mutant did not release a largeamount of DNA in the late-log phase (Fig. 1). To furthersubstantiate a role of quorum sensing in DNA release,we followed the accumulation of extracellular DNAin a
P. aeruginosa lasIrhlI
culture supplemented with 3-oxo-C12-HSL and C4-HSL signal molecules, and in a
P. aeruginosa
wild-type culture supplemented with thesynthetic compound furanone C-30, which specificallyinhibits quorum sensing (Hentzer
et al
., 2002; 2003). Insupport of the suggestion that generation of extracellularDNA in
P. aeruginosa
late-log phase cultures is a quorumsensing-regulated process, we found that accumulation ofextracellular DNA in the late-log phase was restored in the
P. aeruginosa lasIrhlI
culture supplemented with signalmolecules, and that accumulation of extracellular DNAwas inhibited in the wild-type culture supplemented withfuranone (Fig. 1). Although the signal molecules used forchemical complementation of quorum sensing in the
P. aeruginosa lasIrhlI
culture were present also in the logphase, generation of large amounts of extracellular DNAoccurred only in the late-log phase, suggesting that thequorum sensing-regulated DNA-release process is sub-ject to regulation by other systems than
las
and
rhl, aphenomenon that is also true for other quorum sensing-regulated processes such as rhamnolipid synthesis(Medina et al., 2003). The graphs presented in Fig. 1show the result of a representative experiment; however,for reasons which are addressed below, the amountof extracellular DNA detected in the cultures variedsomewhat between experiments and is probablyunderestimated.
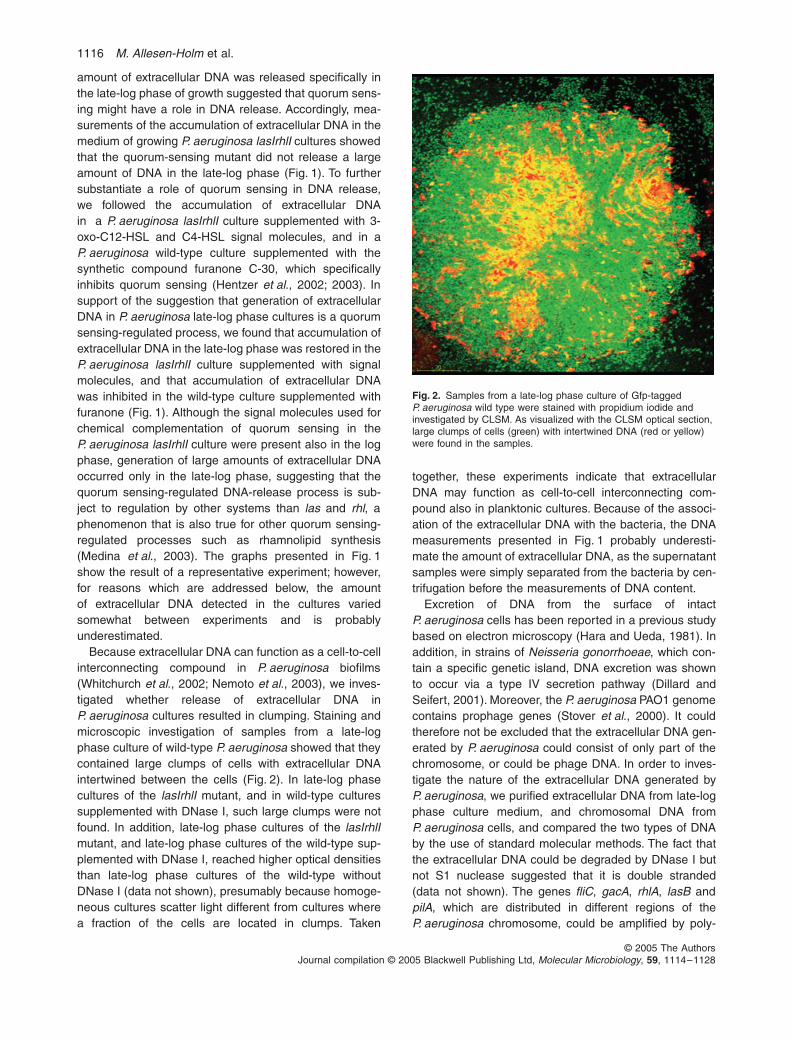
Because extracellular DNA can function as a cell-to-cellinterconnecting compound in P. aeruginosa biofilms(Whitchurch et al., 2002; Nemoto et al., 2003), we inves-tigated whether release of extracellular DNA inP. aeruginosa cultures resulted in clumping. Staining andmicroscopic investigation of samples from a late-logphase culture of wild-type P. aeruginosa showed that theycontained large clumps of cells with extracellular DNAintertwined between the cells (Fig. 2). In late-log phasecultures of the lasIrhlI mutant, and in wild-type culturessupplemented with DNase I, such large clumps were notfound. In addition, late-log phase cultures of the lasIrhlImutant, and late-log phase cultures of the wild-type sup-plemented with DNase I, reached higher optical densitiesthan late-log phase cultures of the wild-type withoutDNase I (data not shown), presumably because homoge-neous cultures scatter light different from cultures wherea fraction of the cells are located in clumps. Taken
together, these experiments indicate that extracellularDNA may function as cell-to-cell interconnecting com-pound also in planktonic cultures. Because of the associ-ation of the extracellular DNA with the bacteria, the DNAmeasurements presented in Fig. 1 probably underesti-mate the amount of extracellular DNA, as the supernatantsamples were simply separated from the bacteria by cen-trifugation before the measurements of DNA content.
Excretion of DNA from the surface of intactP. aeruginosa cells has been reported in a previous studybased on electron microscopy (Hara and Ueda, 1981). Inaddition, in strains of Neisseria gonorrhoeae, which con-tain a specific genetic island, DNA excretion was shownto occur via a type IV secretion pathway (Dillard andSeifert, 2001). Moreover, the P. aeruginosa PAO1 genomecontains prophage genes (Stover et al., 2000). It couldtherefore not be excluded that the extracellular DNA gen-erated by P. aeruginosa could consist of only part of thechromosome, or could be phage DNA. In order to inves-tigate the nature of the extracellular DNA generated byP. aeruginosa, we purified extracellular DNA from late-logphase culture medium, and chromosomal DNA fromP. aeruginosa cells, and compared the two types of DNAby the use of standard molecular methods. The fact thatthe extracellular DNA could be degraded by DNase I butnot S1 nuclease suggested that it is double stranded(data not shown). The genes fliC, gacA, rhlA, lasB andpilA, which are distributed in different regions of theP. aeruginosa chromosome, could be amplified by poly-
Fig. 2. Samples from a late-log phase culture of Gfp-tagged P. aeruginosa wild type were stained with propidium iodide and investigated by CLSM. As visualized with the CLSM optical section, large clumps of cells (green) with intertwined DNA (red or yellow) were found in the samples.
Quorum sensing-regulated DNA release 1117
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd, Molecular Microbiology, 59, 1114–1128
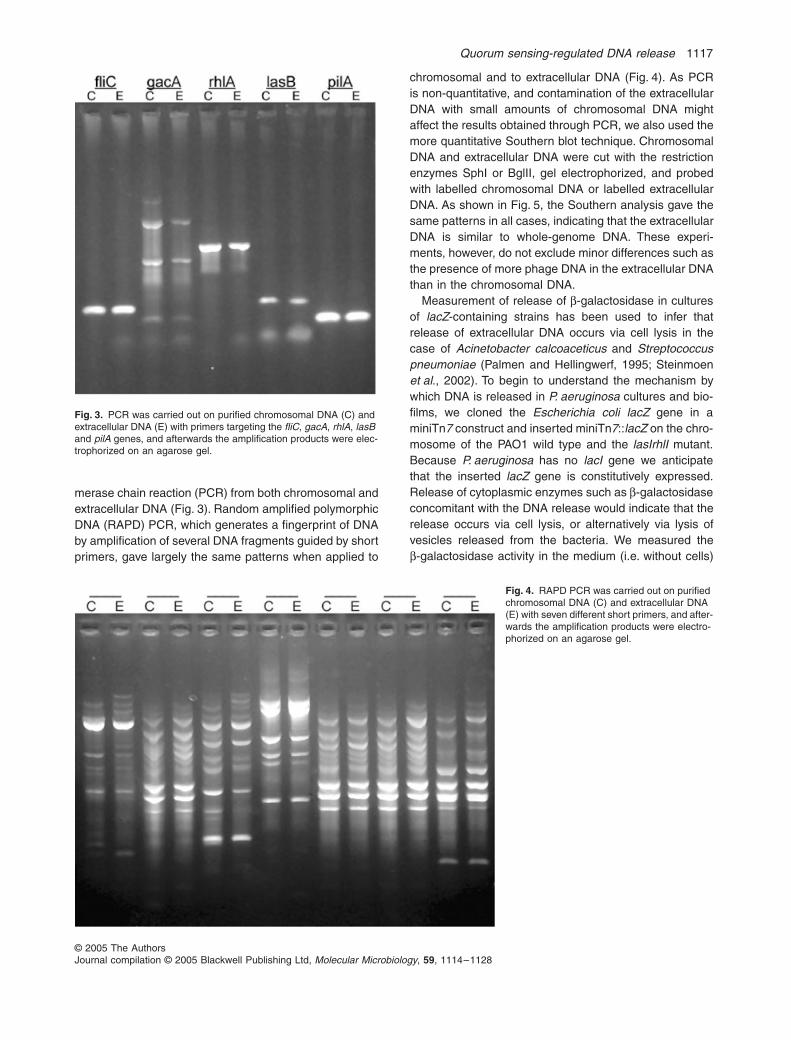
merase chain reaction (PCR) from both chromosomal andextracellular DNA (Fig. 3). Random amplified polymorphicDNA (RAPD) PCR, which generates a fingerprint of DNAby amplification of several DNA fragments guided by shortprimers, gave largely the same patterns when applied to
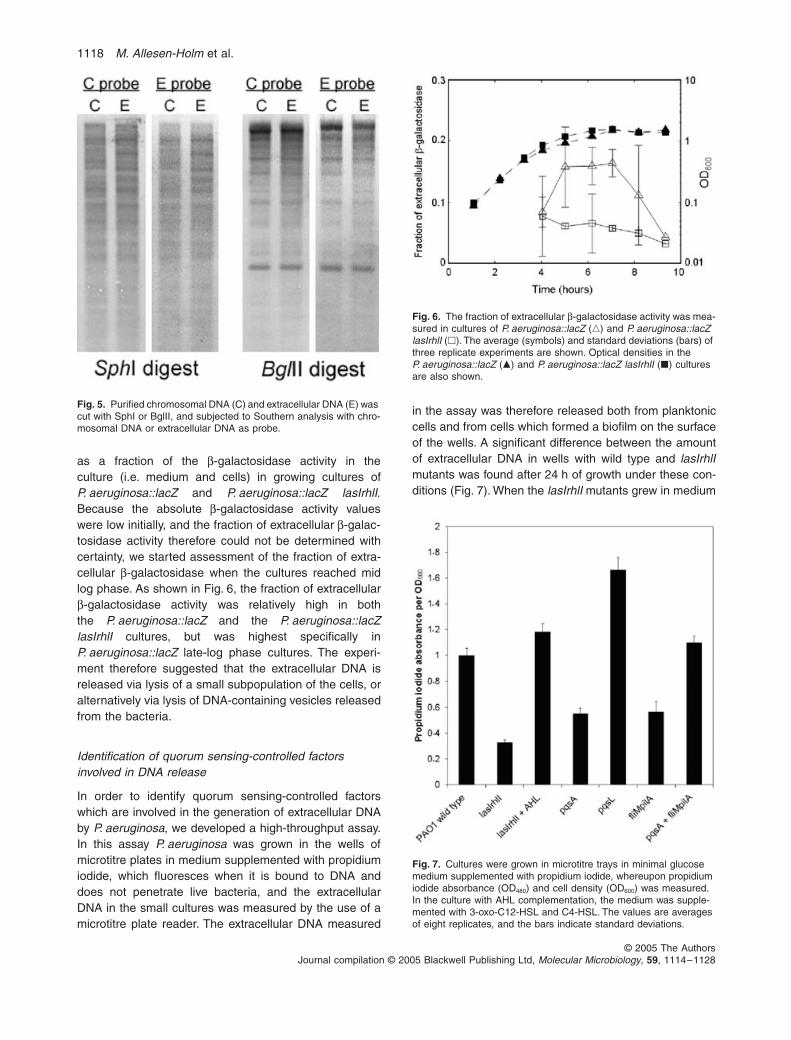
chromosomal and to extracellular DNA (Fig. 4). As PCRis non-quantitative, and contamination of the extracellularDNA with small amounts of chromosomal DNA mightaffect the results obtained through PCR, we also used themore quantitative Southern blot technique. ChromosomalDNA and extracellular DNA were cut with the restrictionenzymes SphI or BglII, gel electrophorized, and probedwith labelled chromosomal DNA or labelled extracellularDNA. As shown in Fig. 5, the Southern analysis gave thesame patterns in all cases, indicating that the extracellularDNA is similar to whole-genome DNA. These experi-ments, however, do not exclude minor differences such asthe presence of more phage DNA in the extracellular DNAthan in the chromosomal DNA.
Measurement of release of β-galactosidase in culturesof lacZ-containing strains has been used to infer thatrelease of extracellular DNA occurs via cell lysis in thecase of Acinetobacter calcoaceticus and Streptococcuspneumoniae (Palmen and Hellingwerf, 1995; Steinmoenet al., 2002). To begin to understand the mechanism bywhich DNA is released in P. aeruginosa cultures and bio-films, we cloned the Escherichia coli lacZ gene in aminiTn7 construct and inserted miniTn7::lacZ on the chro-mosome of the PAO1 wild type and the lasIrhlI mutant.Because P. aeruginosa has no lacI gene we anticipatethat the inserted lacZ gene is constitutively expressed.Release of cytoplasmic enzymes such as β-galactosidaseconcomitant with the DNA release would indicate that therelease occurs via cell lysis, or alternatively via lysis ofvesicles released from the bacteria. We measured theβ-galactosidase activity in the medium (i.e. without cells)
Fig. 3. PCR was carried out on purified chromosomal DNA (C) and extracellular DNA (E) with primers targeting the fliC, gacA, rhlA, lasB and pilA genes, and afterwards the amplification products were elec-trophorized on an agarose gel.
Fig. 4. RAPD PCR was carried out on purified chromosomal DNA (C) and extracellular DNA (E) with seven different short primers, and after-wards the amplification products were electro-phorized on an agarose gel.
1118 M. Allesen-Holm et al.
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd, Molecular Microbiology, 59, 1114–1128
as a fraction of the β-galactosidase activity in theculture (i.e. medium and cells) in growing cultures ofP. aeruginosa::lacZ and P. aeruginosa::lacZ lasIrhlI.Because the absolute β-galactosidase activity valueswere low initially, and the fraction of extracellular β-galac-tosidase activity therefore could not be determined withcertainty, we started assessment of the fraction of extra-cellular β-galactosidase when the cultures reached midlog phase. As shown in Fig. 6, the fraction of extracellularβ-galactosidase activity was relatively high in boththe P. aeruginosa::lacZ and the P. aeruginosa::lacZlasIrhlI cultures, but was highest specifically inP. aeruginosa::lacZ late-log phase cultures. The experi-ment therefore suggested that the extracellular DNA isreleased via lysis of a small subpopulation of the cells, oralternatively via lysis of DNA-containing vesicles releasedfrom the bacteria.
Identification of quorum sensing-controlled factors involved in DNA release
In order to identify quorum sensing-controlled factorswhich are involved in the generation of extracellular DNAby P. aeruginosa, we developed a high-throughput assay.In this assay P. aeruginosa was grown in the wells ofmicrotitre plates in medium supplemented with propidiumiodide, which fluoresces when it is bound to DNA anddoes not penetrate live bacteria, and the extracellularDNA in the small cultures was measured by the use of amicrotitre plate reader. The extracellular DNA measured
in the assay was therefore released both from planktoniccells and from cells which formed a biofilm on the surfaceof the wells. A significant difference between the amountof extracellular DNA in wells with wild type and lasIrhlImutants was found after 24 h of growth under these con-ditions (Fig. 7). When the lasIrhlI mutants grew in medium
Fig. 5. Purified chromosomal DNA (C) and extracellular DNA (E) was cut with SphI or BglII, and subjected to Southern analysis with chro-mosomal DNA or extracellular DNA as probe.
Fig. 6. The fraction of extracellular β-galactosidase activity was mea-sured in cultures of P. aeruginosa::lacZ (�) and P. aeruginosa::lacZ lasIrhlI (�). The average (symbols) and standard deviations (bars) of three replicate experiments are shown. Optical densities in the P. aeruginosa::lacZ (�) and P. aeruginosa::lacZ lasIrhlI (�) cultures are also shown.
Fig. 7. Cultures were grown in microtitre trays in minimal glucose medium supplemented with propidium iodide, whereupon propidium iodide absorbance (OD480) and cell density (OD600) was measured. In the culture with AHL complementation, the medium was supple-mented with 3-oxo-C12-HSL and C4-HSL. The values are averages of eight replicates, and the bars indicate standard deviations.

Quorum sensing-regulated DNA release 1119
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd, Molecular Microbiology, 59, 1114–1128
supplemented with 3-oxo-C12-HSL and C4-HSL, extracel-lular DNA was released to the same level as in wells withwild-type bacteria (Fig. 7). Using this high-throughputassay to screen mutants with defects in quorum sensing-regulated factors we found that a pqsA mutant generatedlow amounts of extracellular DNA (Fig. 7). The pqsAmutant is deficient in the production of PQS, and a pqsLmutant, which overproduces PQS (D’Argenio et al., 2002),was found to release large amounts of extracellular DNA(Fig. 7). PQS has previously been shown to be involvedin lysis of P. aeruginosa cells (D’Argenio et al., 2002), andit was proposed that this might occur via induction of aprophage. Induction of prophage in P. aeruginosa has pre-viously been shown to be quorum sensing-regulated(Hentzer et al., 2004). Because prophage-mediated celllysis in old (9 days or more) P. aeruginosa biofilms hasbeen shown to be dependant on flagella and type IV pili(Webb et al., 2003) we tested a mutant, fliMpilA, devoidof these cell appendages, and found that it released lowamounts of extracellular DNA (Fig. 7). This suggested thatDNA release might occur via induction of a prophage ina few cells followed by flagella/pili-dependent phage prop-agation and lysis of a subpopulation of the cells in theculture. In accordance with the suggestion that the pqsAmutant is deficient in generation of extracellular DNAbecause it lacks PQS-mediated prophage induction, andthat the fliMpilA mutant is deficient in generation of extra-cellular DNA because it lacks flagella/pili-dependentphage propagation, we found that the pqsA and fliMpilAmutants complemented each other in a 1:1 mixture andreleased large amounts of extracellular DNA (Fig. 7). Inorder to obtain more direct evidence for a role of theprophage in DNA release, we have attempted to knockout the chromosomal prophage by the use of allelic dis-placement. So far, however, we have not succeeded inthis, presumably because the prophage also exists in areplicative (plasmid) form.
Extracellular DNA in P. aeruginosa biofilms
We have previously shown that P. aeruginosa PAO1 bio-films grown in flow chambers on glucose minimal mediumdevelop mushroom-shaped multicellular structures via asequential process which involves a stalk-forming sessilesubpopulation and a cap-forming migrating subpopulation(Klausen et al., 2003a), and were interested in mappingthe location of the extracellular DNA in relation to thesesubpopulations. The extracellular DNA in P. aeruginosaPAO1 biofilms was recently visualized by staining withthe specific fluorescent double-stranded DNA stainPicoGreen (Matsukawa and Greenberg, 2004). BecausePicoGreen penetrates and stains live bacteria it is notsuitable for detailed mapping of the location of extracellu-lar DNA in biofilms. In order to map the extracellular DNA
in the heterogeneous P. aeruginosa biofilm, we grew Gfp-tagged PAO1 biofilm in flow chambers irrigated with glu-cose minimal medium, stained the biofilms with DNAstains that do not penetrate the membrane of live bacteriaand performed microscopic investigations by the use ofconfocal laser scanning microscope (CLSM). Figure 8A–I shows CLSM images acquired in 4-day-old biofilmsstained with propidium iodide (Figure 8A–C), ethidiumbromide (Figure 8D–F) and DDAO [7-hydroxy-9H-(1,3-dichloro-9,9-dimethylacridin-2-one)] (Figure 8G–I). As faras we know, DDAO has not previously been used as aDNA stain, but we tested this compound due to its excel-lent fluorescent properties and its structural resemblancewith the DNA stain acridine orange. The top-down views(Fig. 8A, D and G) show extracellular DNA as a red grid-like structure on the substratum, and mushroom capswhich appear green because they are devoid of visibleextracellular DNA. The horizontal section shown in Fig. 8Bis located just above the substratum so that it visualizesbacteria in the stalk portion of the mushroom-shapedstructure, and it is evident that the propidium iodide-stained extracellular DNA is located in the outer part ofthe stalk. The horizontal section shown in Fig. 8E islocated further away from the substratum and visualizesa thin layer of cap bacteria in the periphery surroundingthe stalk bacteria which have the majority of the ethidiumbromide-stained extracellular DNA located in the outerpart of the stalk forming a border between the stalk sub-population and the cap subpopulation. The horizontal sec-tion shown in Fig. 8H is located even further away fromthe substratum and visualizes more of the cap bacteria inthe periphery surrounding the stalk bacteria with DDAO-stained extracellular DNA of which the majority is locatedin the outer parts of the stalk. The horizontal sections inFig. 8C, F and I visualize only the extracellular DNA, andprovide further evidence that the extracellular DNA islocated in the stalk portion of the mushroom-shapedstructures with an increased concentration in the outerparts of the stalks.
The use of propidium iodide and ethidium bromide tovisualize extracellular DNA in biofilms required that thesensitivity of the confocal laser scanning microscope wasincreased to a very high level, and images of good qualitywere difficult to obtain. DDAO, on the other hand, waseasier to detect, and we therefore used this stain forfurther studies of the spatial distribution of extracellularDNA in our P. aeruginosa biofilms. Figure 8J–L showsmicrocolonies in a 2-day-old biofilm upon which caps havenot yet formed. The microcolonies contain extracellularDNA, and a large amount of extracellular DNA is locatedin their outer part as well as upon them and between themforming a grid-like structure on the substratum. Figure 8Mand N shows mushroom-shaped structures in a 6-day-oldbiofilm. From the top-down view (Fig. 8M) it is evident that
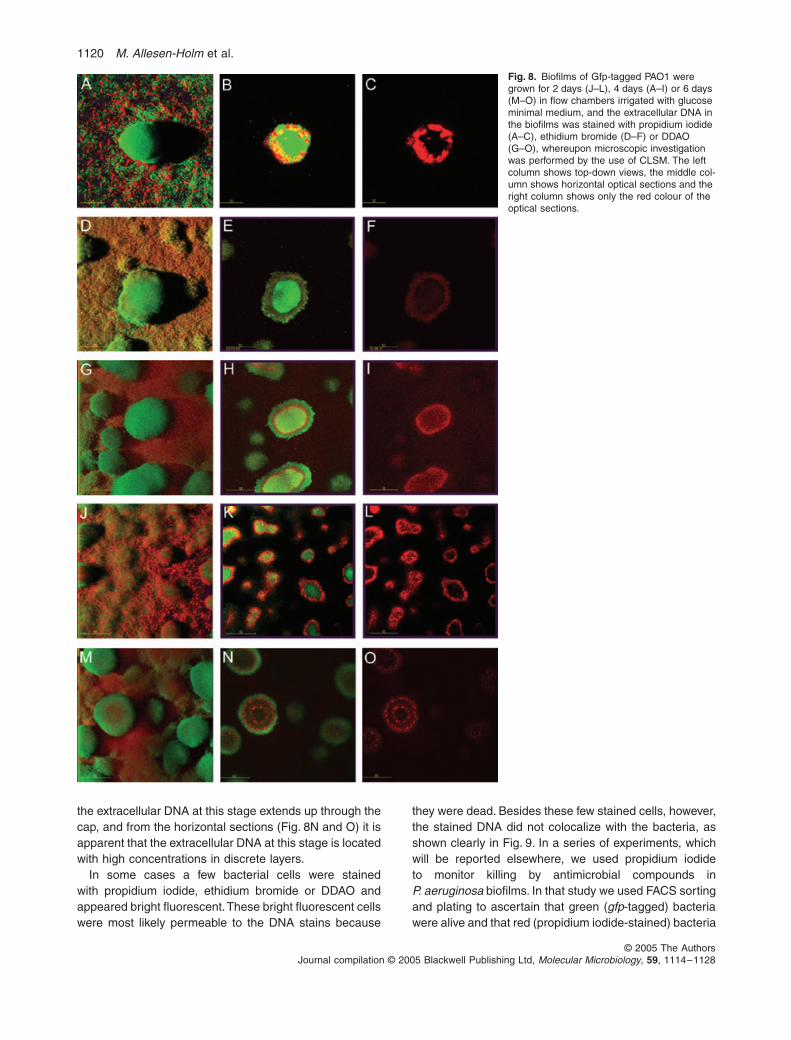
1120 M. Allesen-Holm et al.
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd, Molecular Microbiology, 59, 1114–1128
the extracellular DNA at this stage extends up through thecap, and from the horizontal sections (Fig. 8N and O) it isapparent that the extracellular DNA at this stage is locatedwith high concentrations in discrete layers.
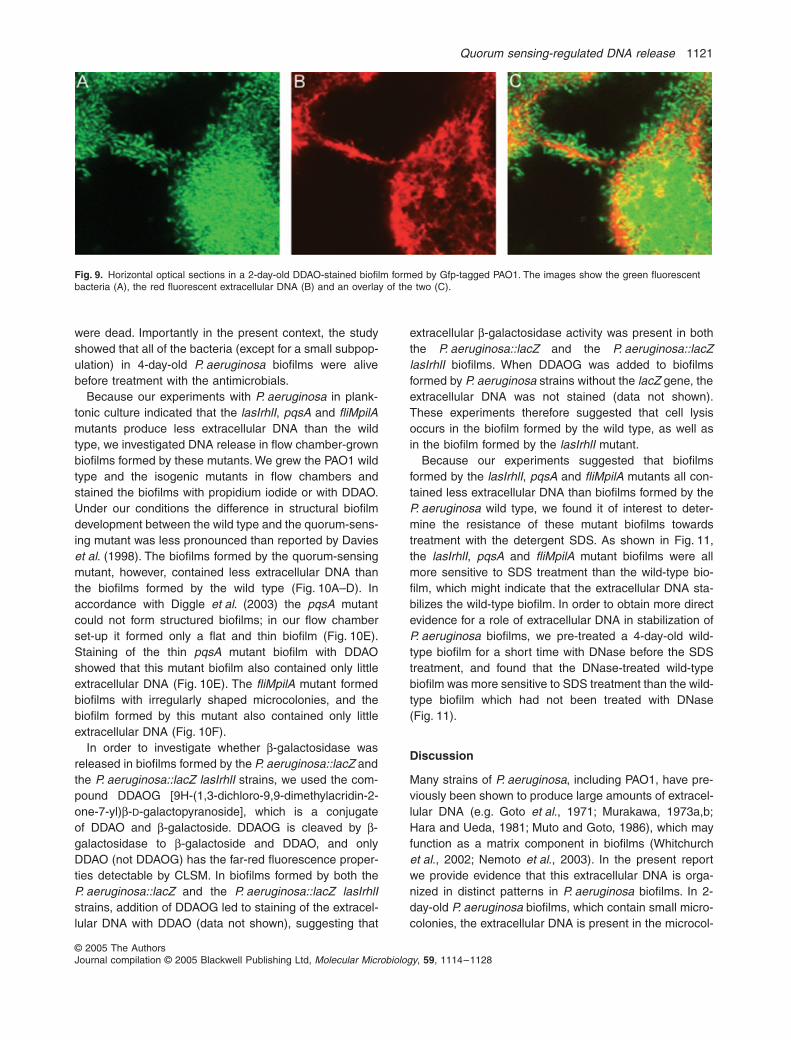
In some cases a few bacterial cells were stainedwith propidium iodide, ethidium bromide or DDAO andappeared bright fluorescent. These bright fluorescent cellswere most likely permeable to the DNA stains because
they were dead. Besides these few stained cells, however,the stained DNA did not colocalize with the bacteria, asshown clearly in Fig. 9. In a series of experiments, whichwill be reported elsewhere, we used propidium iodideto monitor killing by antimicrobial compounds inP. aeruginosa biofilms. In that study we used FACS sortingand plating to ascertain that green (gfp-tagged) bacteriawere alive and that red (propidium iodide-stained) bacteria
Fig. 8. Biofilms of Gfp-tagged PAO1 were grown for 2 days (J–L), 4 days (A–I) or 6 days (M–O) in flow chambers irrigated with glucose minimal medium, and the extracellular DNA in the biofilms was stained with propidium iodide (A–C), ethidium bromide (D–F) or DDAO (G–O), whereupon microscopic investigation was performed by the use of CLSM. The left column shows top-down views, the middle col-umn shows horizontal optical sections and the right column shows only the red colour of the optical sections.
Quorum sensing-regulated DNA release 1121
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd, Molecular Microbiology, 59, 1114–1128
were dead. Importantly in the present context, the studyshowed that all of the bacteria (except for a small subpop-ulation) in 4-day-old P. aeruginosa biofilms were alivebefore treatment with the antimicrobials.
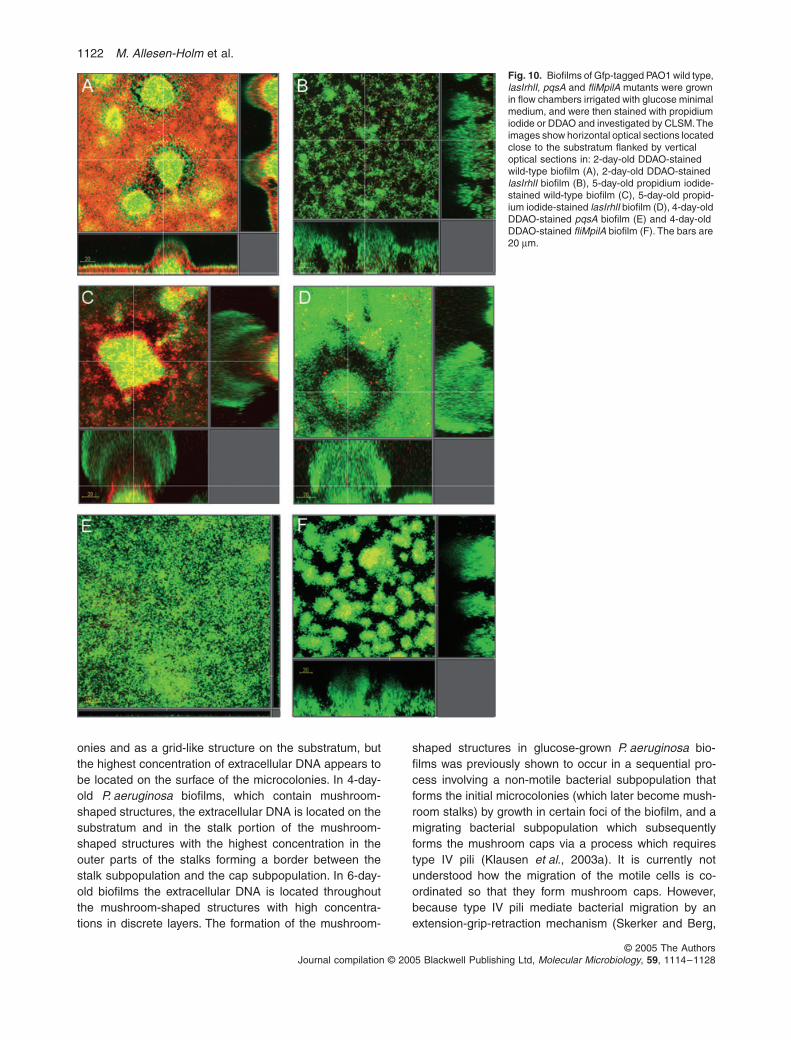
Because our experiments with P. aeruginosa in plank-tonic culture indicated that the lasIrhlI, pqsA and fliMpilAmutants produce less extracellular DNA than the wildtype, we investigated DNA release in flow chamber-grownbiofilms formed by these mutants. We grew the PAO1 wildtype and the isogenic mutants in flow chambers andstained the biofilms with propidium iodide or with DDAO.Under our conditions the difference in structural biofilmdevelopment between the wild type and the quorum-sens-ing mutant was less pronounced than reported by Davieset al. (1998). The biofilms formed by the quorum-sensingmutant, however, contained less extracellular DNA thanthe biofilms formed by the wild type (Fig. 10A–D). Inaccordance with Diggle et al. (2003) the pqsA mutantcould not form structured biofilms; in our flow chamberset-up it formed only a flat and thin biofilm (Fig. 10E).Staining of the thin pqsA mutant biofilm with DDAOshowed that this mutant biofilm also contained only littleextracellular DNA (Fig. 10E). The fliMpilA mutant formedbiofilms with irregularly shaped microcolonies, and thebiofilm formed by this mutant also contained only littleextracellular DNA (Fig. 10F).
In order to investigate whether β-galactosidase wasreleased in biofilms formed by the P. aeruginosa::lacZ andthe P. aeruginosa::lacZ lasIrhlI strains, we used the com-pound DDAOG [9H-(1,3-dichloro-9,9-dimethylacridin-2-one-7-yl)β-D-galactopyranoside], which is a conjugateof DDAO and β-galactoside. DDAOG is cleaved by β-galactosidase to β-galactoside and DDAO, and onlyDDAO (not DDAOG) has the far-red fluorescence proper-ties detectable by CLSM. In biofilms formed by both theP. aeruginosa::lacZ and the P. aeruginosa::lacZ lasIrhlIstrains, addition of DDAOG led to staining of the extracel-lular DNA with DDAO (data not shown), suggesting that
extracellular β-galactosidase activity was present in boththe P. aeruginosa::lacZ and the P. aeruginosa::lacZlasIrhlI biofilms. When DDAOG was added to biofilmsformed by P. aeruginosa strains without the lacZ gene, theextracellular DNA was not stained (data not shown).These experiments therefore suggested that cell lysisoccurs in the biofilm formed by the wild type, as well asin the biofilm formed by the lasIrhlI mutant.
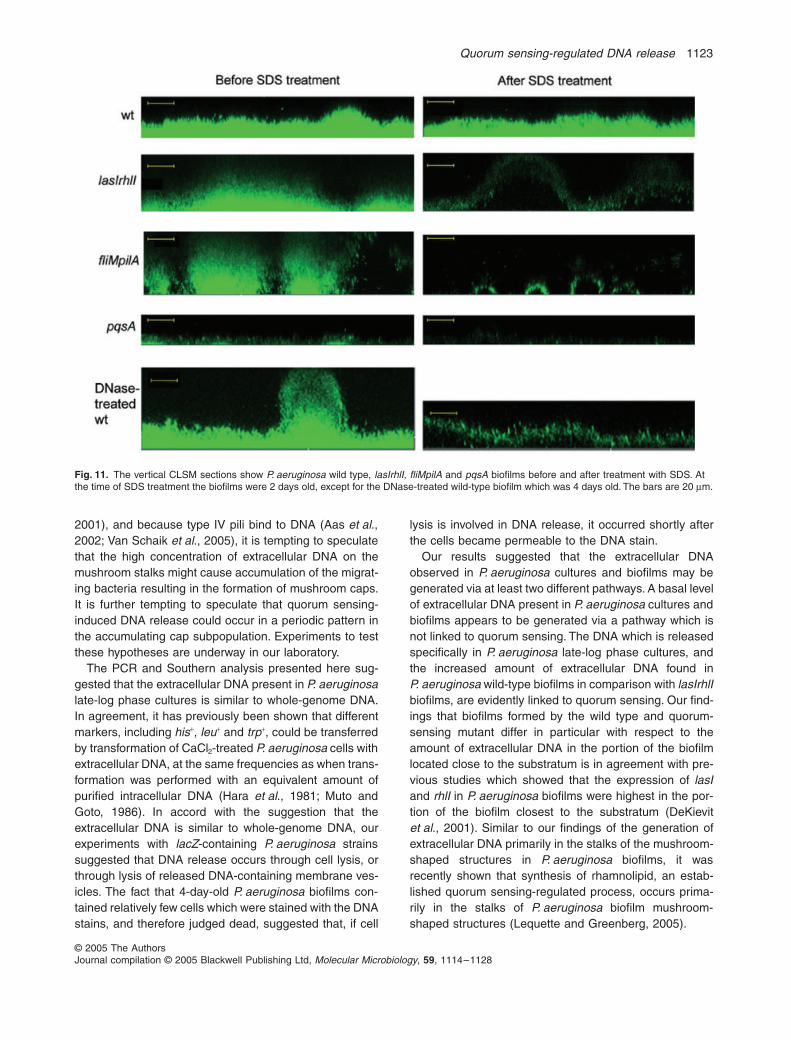
Because our experiments suggested that biofilmsformed by the lasIrhlI, pqsA and fliMpilA mutants all con-tained less extracellular DNA than biofilms formed by theP. aeruginosa wild type, we found it of interest to deter-mine the resistance of these mutant biofilms towardstreatment with the detergent SDS. As shown in Fig. 11,the lasIrhlI, pqsA and fliMpilA mutant biofilms were allmore sensitive to SDS treatment than the wild-type bio-film, which might indicate that the extracellular DNA sta-bilizes the wild-type biofilm. In order to obtain more directevidence for a role of extracellular DNA in stabilization ofP. aeruginosa biofilms, we pre-treated a 4-day-old wild-type biofilm for a short time with DNase before the SDStreatment, and found that the DNase-treated wild-typebiofilm was more sensitive to SDS treatment than the wild-type biofilm which had not been treated with DNase(Fig. 11).
Discussion
Many strains of P. aeruginosa, including PAO1, have pre-viously been shown to produce large amounts of extracel-lular DNA (e.g. Goto et al., 1971; Murakawa, 1973a,b;Hara and Ueda, 1981; Muto and Goto, 1986), which mayfunction as a matrix component in biofilms (Whitchurchet al., 2002; Nemoto et al., 2003). In the present reportwe provide evidence that this extracellular DNA is orga-nized in distinct patterns in P. aeruginosa biofilms. In 2-day-old P. aeruginosa biofilms, which contain small micro-colonies, the extracellular DNA is present in the microcol-
Fig. 9. Horizontal optical sections in a 2-day-old DDAO-stained biofilm formed by Gfp-tagged PAO1. The images show the green fluorescent bacteria (A), the red fluorescent extracellular DNA (B) and an overlay of the two (C).
1122 M. Allesen-Holm et al.
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd, Molecular Microbiology, 59, 1114–1128
onies and as a grid-like structure on the substratum, butthe highest concentration of extracellular DNA appears tobe located on the surface of the microcolonies. In 4-day-old P. aeruginosa biofilms, which contain mushroom-shaped structures, the extracellular DNA is located on thesubstratum and in the stalk portion of the mushroom-shaped structures with the highest concentration in theouter parts of the stalks forming a border between thestalk subpopulation and the cap subpopulation. In 6-day-old biofilms the extracellular DNA is located throughoutthe mushroom-shaped structures with high concentra-tions in discrete layers. The formation of the mushroom-
shaped structures in glucose-grown P. aeruginosa bio-films was previously shown to occur in a sequential pro-cess involving a non-motile bacterial subpopulation thatforms the initial microcolonies (which later become mush-room stalks) by growth in certain foci of the biofilm, and amigrating bacterial subpopulation which subsequentlyforms the mushroom caps via a process which requirestype IV pili (Klausen et al., 2003a). It is currently notunderstood how the migration of the motile cells is co-ordinated so that they form mushroom caps. However,because type IV pili mediate bacterial migration by anextension-grip-retraction mechanism (Skerker and Berg,
Fig. 10. Biofilms of Gfp-tagged PAO1 wild type, lasIrhlI, pqsA and fliMpilA mutants were grown in flow chambers irrigated with glucose minimal medium, and were then stained with propidium iodide or DDAO and investigated by CLSM. The images show horizontal optical sections located close to the substratum flanked by vertical optical sections in: 2-day-old DDAO-stained wild-type biofilm (A), 2-day-old DDAO-stained lasIrhlI biofilm (B), 5-day-old propidium iodide-stained wild-type biofilm (C), 5-day-old propid-ium iodide-stained lasIrhlI biofilm (D), 4-day-old DDAO-stained pqsA biofilm (E) and 4-day-old DDAO-stained fliMpilA biofilm (F). The bars are 20 µm.
Quorum sensing-regulated DNA release 1123
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd, Molecular Microbiology, 59, 1114–1128
2001), and because type IV pili bind to DNA (Aas et al.,2002; Van Schaik et al., 2005), it is tempting to speculatethat the high concentration of extracellular DNA on themushroom stalks might cause accumulation of the migrat-ing bacteria resulting in the formation of mushroom caps.It is further tempting to speculate that quorum sensing-induced DNA release could occur in a periodic pattern inthe accumulating cap subpopulation. Experiments to testthese hypotheses are underway in our laboratory.
The PCR and Southern analysis presented here sug-gested that the extracellular DNA present in P. aeruginosalate-log phase cultures is similar to whole-genome DNA.In agreement, it has previously been shown that differentmarkers, including his+, leu+ and trp+, could be transferredby transformation of CaCl2-treated P. aeruginosa cells withextracellular DNA, at the same frequencies as when trans-formation was performed with an equivalent amount ofpurified intracellular DNA (Hara et al., 1981; Muto andGoto, 1986). In accord with the suggestion that theextracellular DNA is similar to whole-genome DNA, ourexperiments with lacZ-containing P. aeruginosa strainssuggested that DNA release occurs through cell lysis, orthrough lysis of released DNA-containing membrane ves-icles. The fact that 4-day-old P. aeruginosa biofilms con-tained relatively few cells which were stained with the DNAstains, and therefore judged dead, suggested that, if cell
lysis is involved in DNA release, it occurred shortly afterthe cells became permeable to the DNA stain.
Our results suggested that the extracellular DNAobserved in P. aeruginosa cultures and biofilms may begenerated via at least two different pathways. A basal levelof extracellular DNA present in P. aeruginosa cultures andbiofilms appears to be generated via a pathway which isnot linked to quorum sensing. The DNA which is releasedspecifically in P. aeruginosa late-log phase cultures, andthe increased amount of extracellular DNA found inP. aeruginosa wild-type biofilms in comparison with lasIrhlIbiofilms, are evidently linked to quorum sensing. Our find-ings that biofilms formed by the wild type and quorum-sensing mutant differ in particular with respect to theamount of extracellular DNA in the portion of the biofilmlocated close to the substratum is in agreement with pre-vious studies which showed that the expression of lasIand rhlI in P. aeruginosa biofilms were highest in the por-tion of the biofilm closest to the substratum (DeKievitet al., 2001). Similar to our findings of the generation ofextracellular DNA primarily in the stalks of the mushroom-shaped structures in P. aeruginosa biofilms, it wasrecently shown that synthesis of rhamnolipid, an estab-lished quorum sensing-regulated process, occurs prima-rily in the stalks of P. aeruginosa biofilm mushroom-shaped structures (Lequette and Greenberg, 2005).
Fig. 11. The vertical CLSM sections show P. aeruginosa wild type, lasIrhlI, fliMpilA and pqsA biofilms before and after treatment with SDS. At the time of SDS treatment the biofilms were 2 days old, except for the DNase-treated wild-type biofilm which was 4 days old. The bars are 20 µm.

1124 M. Allesen-Holm et al.
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd, Molecular Microbiology, 59, 1114–1128
Our screen of DNA release from mutants which carrylesions in quorum sensing-regulated genes identifiedpqsA as a gene involved in DNA release. The pqsAmutant is deficient in production of PQS, and in supportof a direct role of PQS in DNA release, a pqsL mutant,which overproduces PQS, showed elevated release ofextracellular DNA. Quinolone compounds have previ-ously been shown to induce prophages in bacteria(Phillips et al., 1987; Froshauer et al., 1996), and recentstudies by Webb et al. (2003) and Hentzer et al. (2004)suggested that quorum sensing-regulated DNA releasemight be linked to phage induction in biofilms causing celllysis. In support of a role of phage lysis in DNA releasewe found that a fliMpilA mutant, which has been shownpreviously not to undergo phage-mediated cell lysis(Webb et al., 2003), was deficient in DNA release. Insupport of the suggestion that PQS-mediated prophageinduction and flagella/pili-dependent phage propagationare involved in DNA release, we found that the pqsA andfliMpilA mutants complemented each other in a 1:1 mix-ture and released the same amount of extracellular DNAas the wild type. We propose that PQS produced by thefliMpilA mutant induced the prophage, and that flagella/pili-dependent phage propagation in the pqsA mutantresulted in lysis of a subpopulation of the cells and DNArelease. However, two very recent findings suggest thatmembrane vesicles might have a role in DNA releaseas an alternative to prophages. P. aeruginosa releasesmembrane vesicles which have bacteriolytic effects andcontain DNA (Kadurugamuwa and Beveridge, 1996;Renelli et al., 2004), and extracellular DNA might bereleased either from vesicles that eventually lyse, orthrough the bacteriolytic activity of the vesicles whichmight lyse a small subpopulation of the P. aeruginosacells. Very recently it was shown that PQS is necessaryfor vesicle formation in P. aeruginosa (Mashburn andWhiteley, 2005), and evidence was presented that typeIV pili and flagella are necessary for quorum sensing inP. aeruginosa (Hassett, 2005). The involvement of PQSand type IV pili and flagella in DNA release thereforeshould not be regarded as strong evidence for theinvolvement of prophage.
Contrary to our suggestion, a DNA release mechanismwhich does not involve lysis was suggested in a studywhich employed electron microscopy to visualize whatwas interpreted as excretion of double-stranded DNA fromthe surface of intact P. aeruginosa cells (Hara and Ueda,1981). Studies of other bacterial species indicate thatalthough they all produce extracellular DNA (for a review,see Lorenz and Wackernagel, 1994), the DNA-releasemechanisms vary between species. While DNA releasecaused by lysis of a fraction of the bacteria was reportedto occur in case of S. pneumoniae (Steinmoen et al.,2002) and A. calcoaceticus (Palmen and Hellingwerf,
1995), DNA release without cell lysis was reported tooccur in case of Bacillus subtilis (Lorenz et al., 1991) andN. gonorrhoeae (Dillard and Seifert, 2001). In strains ofN. gonorrhoeae, which contain a specific genetic island,DNA excretion was shown to occur via a type IV secretionpathway (Dillard and Seifert, 2001).
On top of a basal level of DNA release it appears thatmany bacteria, especially those that are able to developnatural competence, possess a specific DNA-release pro-gramme. For example, a correlation between DNA releaseand competence development has been establishedin S. pneumoniae (Steinmoen et al., 2002), B. subtilis(Lorenz et al. (1991), A. calcoaceticus (Palmen and Hell-ingwerf (1995), N. gonorrhoeae (Dillard and Seifert, 2001)and Pseudomonas stutzeri (Stewart et al., 1983). In allthese cases DNA release and competence developmentoccur in the late-log phase in liquid cultures, and in someof the cases competence development has been shownto be regulated through a quorum-sensing mech-anism (Magnuson et al., 1994; Pestova et al., 1996).P. aeruginosa is not known to be naturally transformable,but as the other members of the P. aeruginosa DNAhomology subgroup (i.e. P. stutzeri, Pseudomonas men-docina, Pseudomonas alcaligenes and Pseudomonaspseudoalcaligenes) were all found to be naturally trans-formable (Carlson et al., 1983), it is possible that someP. aeruginosa strains has this capability. It is possibletherefore that DNA release in some cases both allowsexchange of genetic material to take place and inducesbiofilm structure formation and stabilization. The relativelylong-lasting physical proximity of bacteria in biofilmsenable the constituent cells to establish long-term rela-tionships with each other, and biofilms appear to beoptimal environments for gene transfer to occur via trans-formation. Biofilm-grown Streptococcus mutans cells wereshown to be transformed at rates 10- to 600-fold higherthan planktonic S. mutans cells (Li et al., 2001). The trans-fer of chromosomal genes in P. stutzeri cell mats were1000-fold more efficient when the DNA was released fromdonor cells in comparison with when the same amount ofDNA was provided from a DNA preparation (Stewart et al.,1983). Transfer of a non-conjugative plasmid from Tre-ponema denticola to Streptococcus gordonii growing in amixed species biofilm was demonstrated to occur withhigh efficiency (Wang et al., 2002), and finally Wuertz andco-workers showed that transformation occurs with highefficiency in biofilms of an Acinetobacter sp. (Hendrickxet al., 2003).
Quorum sensing has previously been shown to play arole in structural development of P. aeruginosa biofilms(Davies et al., 1998), but with the exception of rhamno-lipid, which has been shown to play a role in maintainingthe channels between the mushroom-shaped structures(Davey et al., 2003), the actual quorum sensing-controlled

Quorum sensing-regulated DNA release 1125
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd, Molecular Microbiology, 59, 1114–1128
factors that play a role in P. aeruginosa biofilm develop-ment have not been identified. Several recent studieshave demonstrated that biofilms which develop in theabsence of functional quorum-sensing systems (causedby either inhibition or mutation) are vulnerable to SDStreatment, shear forces, kanamycin treatment, tobramycintreatment, H2O2 treatment and polymorphonuclear neutro-phils phagocytosis (Davies et al., 1998; Hassett et al.,1999; Hentzer et al., 2002; 2003; Shih and Huang, 2002;Bjarnsholt et al., 2005; Rasmussen et al., 2005). In thepresent study we found that release of large amounts ofextracellular DNA in P. aeruginosa biofilms is dependenton quorum sensing, and that P. aeruginosa biofilms with-out large amounts of extracellular DNA, formed by thelasIrhlI, fliMpilA and pqsA mutants or obtained throughDNase treatment of wild-type biofilm, are vulnerable totreatment with SDS. Although several hundred genes inP. aeruginosa have been shown to be quorum sensing-regulated (Wagner et al., 2003), and it is possible thatmutations in the quorum-sensing control systems mightaffect many gene activities which have roles in biofilmdevelopment, the present study suggests that one of thequorum sensing-controlled factors that might have a rolein biofilm development by P. aeruginosa is programmedDNA release resulting in extracellular DNA which func-tions as a biofilm matrix component.
Experimental procedures
Bacterial strains and growth conditions
Pseudomonas aeruginosa PAO1 (Holloway and Morgan,1986) was used as the model organism in this study. Allexperiments involving wild-type PAO1 were performed withthree different sublines, one obtained from the Pseudomo-nas Genetic Stock Center (strain PAO0001), one obtainedfrom the laboratory of Barbara Iglewski and one obtainedfrom the laboratory of John Mattick. The three PAO1 wild-type sublines did not differ with respect to the investigatedphenotypes. The lasIrhlI derivative was constructed by allelicdisplacement in the PAO0001 subline as described (Hentzeret al., 2003). The fliMpilA derivative was constructed in thePAO0001 subline and in the PAO1 subline from John Mat-tick’s laboratory as described (Klausen et al., 2003b), andboth fliMpilA derivatives had the same phenotypes. ThepqsA and pqsL mutants were constructed by transposoninsertion in the PAO1 subline from Barbara Iglewski’s labora-tory as described (D’Argenio et al., 2002). The strains werefluorescently tagged at an intergenic neutral chromosomallocus with gfp in a miniTn7 construct (Klausen et al., 2003b).AB medium (Clark and Maaløe, 1967) was used supple-mented with 30 mM glucose for batch cultures, and with0.3 mM glucose for biofilm cultivation. Biofilms and batchcultures were grown at 30°C. DNase I (Sigma) was used ata concentration of 90 Kunitz units per ml in medium supple-mented with MgCl2 (5 mM). Propidium iodide, ethidium bro-mide and DDAO were used at a concentration of 1 µM to
stain extracellular DNA in biofilms. DDAOG was used at aconcentration of 1 µM to assess β-galactosidase activity inbiofilms. 3-oxo-C12-HSL and C4-HSL were used at a con-centration of 1 µM. Furanone C-30 was used at a concentra-tion of 10 µM.
Construction of P. aeruginosa strains with lacZ inserted on the chromosome
A miniTn7-PUV5–lacZ delivery plasmid was constructed bycloning a 3.3 kb NotI fragment containing a PUV5–lacZcassette into a NotI-digested miniTn7-strepR vector basedon pUC19. The miniTn7-strepR-PUV5–lacZ transposon wasinserted into the chromosome of the PAO1 wild type andlasIrhlI mutant by four parental matings using E. coli HB101with the delivery plasmid, and the helper strains E. coliHB101/RK600 and E. coli HB101/pUX-BF13 as describedpreviously (Klausen et al., 2003b).
Measurements of extracellular DNA in planktonic cultures
Culture samples were centrifuged (3 min, 10.000 r.p.m.) andthe supernatant was transferred to a new eppendorf tube.NaCl was added to the supernatant to a concentration of0.25 M, and the extracellular DNA was precipitated by add-ing 2:1 volume of ethanol. The precipitated extracellularDNA was dissolved in TE buffer, and the DNA concentrationwas determined by spectrophotometry (OD260/OD280), or byusing the PicoGreen® dsDNA Quantitation Kit (MolecularProbes).
Measurements of extracellular DNA in microtitre tray cultures
Overnight cultures grown in AB (0.5% glucose) medium werediluted to OD600 = 0.001 in AB medium supplemented with0.5% glucose and 0.05 mM propidium iodide. The dilutedcultures were transferred to wells of polystyrene microtitreplates (150 µl of cultures per well) and incubated for 24 h at37°C, whereupon propidium iodide absorbance was mea-sured at OD480 and cell density was measured at OD600 bythe use of a Wallac microplate reader. In the experiment withhomoserine lactone complementation, 3-oxo-C12-HSL andC4-HSL were added to the medium to a final concentrationof 10 µM.
Measurements of β-galactosidase activity in planktonic cultures
The fraction of extracellular β-galactosidase in planktoniccultures was assessed by measuring β-galactosidase activityin culture samples and in culture supernatants essentially asdescribed by Steinmoen et al. (2002).
Cultivation of biofilms
Biofilms were grown in flow chambers with individual channeldimensions of 1 × 4 × 40 mm. The flow system was assem-bled and prepared as described previously (Møller et al.,

1126 M. Allesen-Holm et al.
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd, Molecular Microbiology, 59, 1114–1128
1998). The flow chambers were inoculated by injecting 350 µlof overnight culture diluted to an OD600 of 0.001 into eachflow channel with a small syringe. After inoculation flow chan-nels were left without flow for 1 h, after which medium flow(0.2 mm s−1) was started using a Watson Marlow 205S peri-staltic pump.
Microscopy and image acquisition
All microscopic observations and image acquisitions wereperformed with a Zeiss LSM 510 CLSM (Carl Zeiss, Jena,Germany) equipped with detectors and filter sets for monitor-ing of Gfp, propidium iodide, ethidium bromide and DDAOfluorescence. Images were obtained using a 63×/1.4objective or a 40×/1.3 objective. Simulated three-dimensionalimages and sections were generated using the IMARIS soft-ware package (Bitplane AG, Zürich, Switzerland).
SDS treatment of biofilms
In order to assess the sensitivity of wild type and mutantP. aeruginosa biofilms to SDS treatment, 2-day-old biofilmswere irrigated for 2 h with medium containing 0.01% SDS,and CLSM images were acquired before and after the SDStreatment. In order to investigate the effect of pre-treatmentwith DNase I on the sensitivity of the wild-type biofilm toDNase treatment, a 4-day-old wild-type biofilm was treatedwith DNase I (100 µg ml−1) for 45 min before the SDStreatment.
PCR and RAPD PCR
All PCR reactions were performed using Taq DNA Poly-merase (Sigma) and 10× reaction buffer with MgCl2(Sigma) + 0.25 mM each of ATP, CTP, GTP and TTP in thepresence of 5% DMSO (Merck). Primers for amplification offliC, gacA, rhlA, lasB and pilA sequences were used at aconcentration of 5 µM. The PCR reaction was performed ona T3 Thermocycler (Biometra); 45 s at 95°C, 1 min at 52°C,1 min at 72°C and cycled 30 times. The RAPD PCR reactionswere performed with 10-mer primers, the annealing temper-ature was 31°C and the reactions were run for 45 cycles.Sequences of the used primers are available on request.
Southern blot analysis
Southern analysis was performed using standard protocols.Chromosomal and extracellular DNA probes were preparedwith the DIG DNA Labeling Kit (Roche), which inserts digox-igenin-dUTP by random primed DNA labelling using Klenowenzyme. Before use the probe was denatured by boiling for10 min. The probes were mixed with hybridization buffer andallowed to hybridize with the membrane overnight at 55°C.The membrane was then washed with decreasing concen-trations of SSC buffer at 55°C, and the hybridized DIG-labelled probes were bound by anti-DIG alkaline phos-phates. Finally the bands were visualized with NBT/BCIPdissolved in Tris buffer at pH = 9.5 according to the manufac-turer (Roche).
Acknowledgements
We thank Janus Haagensen for help with confocal micros-copy and Anne Nielsen for performing Southern blots. Thiswork was supported by a grant from the Danish TechnicalResearch Council to T.T.N. (26-03-0234).
References
Aas, F.E., Wolfgang, M., Frye, S., Dunham, S., Lovold, C.,and Koomey, M. (2002) Competence for natural transfor-mation in Neisseria gonorrhoeae: components of DNAbinding and uptake linked to type IV pilus expression.Mol Microbiol 46: 749–760.
Bjarnsholt, T., Jensen, P.O., Burmolle, M., Hentzer, M.,Haagensen, J.A.J., Hougen, H.P., et al. (2005) Pseudomo-nas aeruginosa tolerance to tobramycin, hydrogen perox-ide and polymorphonuclear leukocytes is quorum-sensingdependent. Microbiology 151: 373–383.
Carlson, C.A., Pierson, L.S., Rosen, J.J., and Ingraham, J.L.(1983) Pseudomonas stutzeri and related species undergonatural transformation. J Bacteriol 153: 93–99.
Clark, J.D., and Maaløe, O. (1967) DNA replication and thecell cycle in Escherichia coli cells. J Mol Biol 23: 99–112.
Costerton, J.W., Lewandowski, Z., Caldwell, D.E., Korber,D.R., and Lappin-Scott, H.M. (1995) Microbial biofilms.Annu Rev Microbiol 49: 711–745.
Costerton, J.W., Stewart, P.S., and Greenberg, E.P. (1999)Bacterial biofilms: a common cause of persistent infec-tions. Science 284: 1318–1322.
D’Argenio, D.A., Calfee, M.W., Rainey, P.B., and Pesci, E.C.(2002) Autolysis and autoaggregation in Pseudomonasaeruginosa colony morphology mutants. J Bacteriol 184:6481–6489.
Davey, M.E., Caiazza, N.C., and O’Toole, G.A. (2003) Rham-nolipid surfactant production affects biofilm architecture inPseudomonas aeruginosa PAO1. J Bacteriol 185: 1027–1036.
Davies, D.G., Parsek, M.R., Pearson, J.P., Iglewsky, B.H.,Costerton, J.W., and Greenberg, E.P. (1998) The involve-ment of cell-to-cell signals in the development of a bacterialbiofilm. Science 280: 295–298.
DeKievit, T.R., Gillis, R., Marx, S., Brown, C., and Iglewski,B.H. (2001) Quorum-sensing genes in Pseudomonasaeruginosa biofilms: their role and expression patterns.Appl Environ Microbiol 67: 1865–1873.
Diggle, S.P., Winzer, K., Chhabra, S.R., Worrall, K.E.,Camara, M., and Williams, P. (2003) The Pseudomonasaeruginosa quinolone signal molecule overcomes the celldensity-dependency of the quorum sensing hierarchy,regulates rhl-dependent genes at the onset of stationaryphase and can be produced in the absence of LasR. MolMicrobiol 50: 29–43.
Dillard, J.P., and Seifert, H.S. (2001) A variable genetic islandspecific for Neisseria gonorrhoeae is involved in providingDNA for natural transformation and is found more often indisseminated infection isolates. Mol Microbiol 41: 263–277.
Friedman, L., and Kolter, R. (2004a) Two genetic loci producedistinct carbohydrate-rich structural components of thePseudomonas aeruginosa biofilm matrix. J Bacteriol 186:4457–4465.

Quorum sensing-regulated DNA release 1127
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd, Molecular Microbiology, 59, 1114–1128
Friedman, L., and Kolter, R. (2004b) Genes involved in matrixformation in Pseudomonas aeruginosa PA14 biofilms. MolMicrobiol 51: 675–690.
Froshauer, S., Silvia, A.M., Chidambaram, M., Sharma, B.,and Weinstock, G.M. (1996) Sensitization of bacteria todanofloxacin by temperate prophages. Antimicrob AgentsChemother 40: 1561–1563.
Givskov, M., de Nys, R., Manefield, M., Gram, L., Maximilien,R., Eberl, L., et al. (1996) Eukaryotic interference withhomoserine lactone-mediated prokaryotic signalling. JBacteriol 178: 6618–6622.
Goto, S., Enomoto, S., Takahashi, Y., and Motomatsu, R.(1971) Slime production by Pseudomonas aeruginosa I.Conditions for slime production by the cellophane platemethod. Jpn J Microbiol 15: 317–324.
Hara, T., and Ueda, S. (1981) A study on the mechanism ofDNA excretion from P aeruginosa KYU-1 – effect of myto-mycin C on extracellular DNA production. Agric Biol Chem45: 2457–2461.
Hara, T., Aumayr, A., and Ueda, S. (1981) Genetic transfor-mation of Pseudomonas aeruginosa with extracellularDNA. J Gen Appl Microbiol 27: 109–114.
Hassett, D.J. (2005) Tails, hairs, and speech in Pseudomo-nas aeruginosa. Pseudomonas, 10th international con-gress. Marseille, France, August 27–31, 2005. Oralpresentation and Abstract S19.
Hassett, D.J., Ma, J.-F., Elkins, J.G., McDermoth, T.R.,Ochsner, U.A., West, S.E.H., et al. (1999) Quorum-sensingin Pseudomonas aeruginosa controls expression of cata-lase and superoxide dismutase genes and mediates bio-film susceptibility to hydrogen peroxide. Mol Microbiol 34:1082–1093.
Hendrickx, L., Hausner, M., and Wuertz, S. (2003) Naturalgenetic transformation in monoculture Acinetobacter sp.BD413 biofilms. Appl Environ Microbiol 69: 1721–1727.
Hentzer, M., Riedel, K., Rasmussen, T.B., Heydorn, A.,Andersen, J.B., Parsek, M.R., et al. (2002) Inhibition ofquorum sensing in Pseudomonas aeruginosa biofilm bac-teria by a halogenated furanone compound. Microbiology148: 87–102.
Hentzer, M., Wu, H., Andersen, J.B., Riedel, K., Rasmussen,T.B., Bagge, N., et al. (2003) Attenuation of Pseudomonasaeruginosa virulence by quorum sensing inhibitors. EMBOJ 22: 3803–3815.
Hentzer, M., Eberl, L., and Givskov, M. (2004) Quorum sens-ing in Biofilms: gossip in the slime world? In MicrobialBiofilms. Ghannoum M., and O’Toole, G. (eds). Was-hington, DC: American Society for Microbiology Press,pp. 118–140.
Holloway, B.W., and Morgan, A.F. (1986) Genome organiza-tion in Pseudomonas. Annu Rev Microbiol 40: 79–105.
Kadurugamuwa, J.L., and Beveridge, T.J. (1996) Bacteri-olytic effect of membrane vesicles from Pseudomonasaeruginosa on other bacteria including pathogens: concep-tually new antibiotics. J Bacteriol 178: 2767–2774.
Klausen, M., Aaes-Jorgensen, A., Molin, S., and Tolker-Nielsen, T. (2003a) Involvement of bacterial migration inthe development of complex multicellular structures inPseudomonas aeruginosa biofilms. Mol Microbiol 50: 61–68.
Klausen, M., Heydorn, A., Ragas, P., Lambertsen, L., Aaes-
Jørgensen, A., Molin, S., and Tolker-Nielsen, T. (2003b)Biofilm formation by Pseudomonas aeruginosa wild-type,flagella, and type IV pili mutants. Mol Microbiol 48: 1511–1524.
Latifi, A., Winson, M.K., Foglino, M., Bycroft, B.W., Stewart,G.S., Lazdunski, A., and Williams, P. (1995) Multiplehomologues of LuxR and LuxI control expression of viru-lence determinants and secondary metabolites throughquorum sensing in Pseudomonas aeruginosa PAO1. MolMicrobiol 17: 333–343.
Latifi, A., Foglino, M., Tanaka, K., Williams, P., and Laz-dunski, A. (1996) A hierarchical quorum-sensing cascadein Pseudomonas aeruginosa links the transcriptionalactivators LasR and RhIR (VsmR) to expression of thestationary-phase sigma factor RpoS. Mol Microbiol 21:1137–1146.
Lequette, Y., and Greenberg, E.P. (2005) Timing and local-ization of rhamnolipid synthesis gene expression inPseudomonas aeruginosa biofilms. J Bacteriol 187: 37–44.
Li, Y.-H., Lau, P.C.Y., Lee, J.H., Ellen, R.P., and Cvitkovitch,D.G. (2001) Natural genetic transformation of Streptococ-cus mutans growing in biofilms. J Bacteriol 183: 897–908.
Lorenz, M.G., and Wackernagel, W. (1994) Bacterial genetransfer by natural genetic transformation in the environ-ment. Microbiol Rev 58: 563–602.
Lorenz, M.G., Gerjets, D., and Wackernagel, W. (1991)Release of transforming plasmid and chromosomal DNAfrom two cultured soil bacteria. Arch Microbiol 156: 319–326.
McKnight, S.L., Iglewski, B.H., and Pesci, E.C. (2000) ThePseudomonas quinolone signal regulates rhl quorum sens-ing in Pseudomonas aeruginosa. J Bacteriol 182: 2702–2708.
Magnuson, R., Solomon, J., and Grossman, A.D. (1994) Bio-chemical and genetic characterization of a competencepheromone from B. subtilis. Cell 77: 207–216.
Mashburn, L.M., and Whiteley, M. (2005) Membrane vesiclestraffic signals and facilitate group activities in a prokaryote.Nature 437: 422–425.
Matsukawa, M., and Greenberg, E.P. (2004) Putativeexopolysaccharide synthesis genes influence Pseudomo-nas aeruginosa biofilm development. J Bacteriol 186:4449–4456.
Medina, G., Juarez, K., and Soberon-Chavez, G. (2003) ThePseudomonas aeruginosa rhlAB operon is not expressedduring the logarithmic phase of growth even in the pres-ence of its activator RhlR and the autoinducer N-butyryl-homoserine lactone. J Bacteriol 185: 377–380.
Møller, S., Sternberg, C., Andersen, J.B., Christensen, B.B.,and Molin, S. (1998) In situ gene expression in mixed-culture biofilms: evidence of metabolic interactionsbetween community members. Appl Environ Microbiol 64:721–732.
Murakawa, T. (1973a) Slime production by Pseudomonasaeruginosa. III. Purification of slime and its physicochemi-cal properties. Jpn J Microbiol 17: 273–281.
Murakawa, T. (1973b) Slime production by Pseudomonasaeruginosa. IV. Chemical analysis of two varieties of slimeproduced by Pseudomonas aeruginosa. Jpn J Microbiol17: 513–520.

1128 M. Allesen-Holm et al.
© 2005 The AuthorsJournal compilation © 2005 Blackwell Publishing Ltd, Molecular Microbiology, 59, 1114–1128
Muto, Y., and Goto, S. (1986) Transformation by extracellularDNA produced by Pseudomonas aeruginosa. MicrobiolImmunol 30: 621–628.
Nemoto, K., Hirota, K., Murakami, K., Taniguti, K., Murata,H., Viducic, D., and Miyake, Y. (2003) Effect of Varidase(streptodornase) on biofilm formed by Pseudomonasaeruginosa. Chemotherapy 49: 121–125.
O’Toole, G.A., and Kolter, R. (1998) Flagellar and twitchingmotility are necessary for Pseudomonas aeruginosa bio-film development. Mol Microbiol 30: 295–304.
Palmen, R., and Hellingwerf, K.J. (1995) Acinetobacter cal-coaceticus liberates chromosomal DNA during induction ofcompetence by cell lysis. Curr Microbiol 30: 7–10.
Passador, L., Cook, J.M., Gambello, M.J., Rust, L., andIglewski, B.H. (1993) Expression of Pseudomonas aerug-inosa virulence genes requires cell-to-cell communication.Science 260: 1127–1130.
Pesci, E.C., Milbank, J.B., Pearson, J.P., McKnight, S.,Kende, A.S., Greenberg, E.P., and Iglewski, B.H. (1999)Quinolone signaling in the cell-to-cell communication sys-tem of Pseudomonas aeruginosa. Proc Natl Acad Sci USA96: 11229–11234.
Pestova, E.V., Havarstein, L.S., and Morrison, D.A. (1996)Regulation of competence for genetic transformation inStreptococcus pneumoniae by an auto-induced peptidepheromone and a two-component regulatory system.Mol Microbiol 21: 853–862.
Phillips, I., Culebras, E., Moreno, F., and Baquero, F. (1987)Induction of the SOS response by new 4-quinolones.J Antimicrob Chemother 20: 631–638.
Rasmussen, T.B., Bjarnsholt, T., Skindersoe, M.E.,Hentzer, M., Kristoffersen, P., Köte, M., et al. (2005)Screening for quorum-sensing inhibitors (QSI) by use ofa novel genetic system, the QSI selector. J Bacteriol187: 1799–1814.
Renelli, M., Matias, V., Lo, R.Y., and Beveridge, T.J. (2004)DNA-containing membrane vesicles of Pseudomonasaeruginosa PAO1 and their genetic transformation poten-tial. Microbiology 150: 2161–2169.
Shih, P.-C., and Huang, C.-T. (2002) Effects of quorum-sensing deficiency on Pseudomonas aeruginosa biofilm
formation and antibiotic resistance. J AntimicrobChemother 49: 309–314.
Skerker, J.M., and Berg, H.C. (2001) Direct observation ofextension and retraction of type IV pili. Proc Natl Acad SciUSA 98: 6901–6904.
Steinmoen, H., Knutsen, E., and Håvarstein, L.S. (2002)Induction of natural competence in Streptococcus pneumo-niae triggers lysis and DNA release from a subfraction ofthe cell population. Proc Natl Acad Sci USA 99: 7681–7686.
Stewart, G.J., Carlson, C.A., and Ingraham, J.L. (1983) Evi-dence for an active role of donor cells in natural transfor-mation of Pseudomonas stutzeri. J Bacteriol 156: 30–35.
Stover, C.K., Pham, X.Q., Erwin, A.L., Mizoguchi, S.D., War-rener, P., Hickey, M.J., et al. (2000) Complete genomesequence of Pseudomonas aeruginosa PAO1, an opportu-nistic pathogen. Nature 406: 959–964.
Vallet, I., Olson, J.W., Lory, S., Lazdunski, A., and Filloux, A.(2001) The chaperone/usher pathways of Pseudomonasaeruginosa: identification of fimbrial gene clusters (cup)and their involvement in biofilm formation. Proc Natl AcadSci USA 98: 6911–6916.
Van Schaik, E.J., Giltner, C.L., Audette, G.F., Keizer, D.W.,Bautista, D.L., Slupsky, C.M., et al. (2005) DNA binding:a novel function of Pseudomonas aeruginosa type IV pili.J Bacteriol 187: 1455–1464.
Wagner, V.E., Bushnell, D., Passador, L., Brooks, A.I., andIglewski, B.H. (2003) Microarray analysis of Pseudomonasaeruginosa quorum-sensing regulons: effects of growthphase and environment. J Bacteriol 185: 2080–2095.
Wang, B.Y., Chi, B., and Kuramitsu, H.K., (2002) Geneticexchange between Treponema denticola and Streptococ-cus gordonii in biofilms. Oral Microbiol Immunol 2: 108–112.
Webb, J.S., Thompson, L.S., James, S., Charlton, T., Tolker-Nielsen, T., Koch, B., et al. (2003) Cell death in Pseu-domonas aeruginosa biofilm development. J Bacteriol 185:4585–4592.
Whitchurch, C.B., Tolker-Nielsen, T., Ragas, P.C., and Mat-tick, J.S. (2002) Extracellular DNA is required for bacterialbiofilm formation. Science 295: 1487.
Copyright © 2022 FDOKUMEN




















