
Possibility of using purified pyocins for typing Pseudomonas aeruginosa: Purification of pyocins and...
14
(~) ELSEVIER Paris 1986 Ann. lnst. Pasteur/Microbiol. 1986, 137 A, 253-266 POSSIBILITY OF USING PURIFIED PYOCINS FOR TYPING PSEUDOMONAS AERUGINOSA : PURIFICATION OF PYOCINS AND SENSITIVITY OF P. AERUGINOSA IN DIFFERENT TESTS by D. Jurado Chacon (1), A. Chueca Sancho (2), J.F. Guillen Solvas (1), B. Garcia-Villanova Ruiz (3), and R. Galvez Vargas (3) Q) Medicina Preventiva, Escuela de Enfermeria, Universidad de Granada, (2) Estacidn Experimental Zaidin (CSIC), Granada, and (3) Cdtedra de Medicina Preventiva, Facultad de Medicina, Universidad de Granada (Espaha). SUMMARY Two types of pyocins were simultaneously found in Pseudomonas aerugi- nosa strains HCP2. According to their structures, they belonged to the types classified as R and F, respectively, and were named HCP2-R and HCP2-F. The sensitivity of 87 strains of P. aeruginosa of clinical origin to pyocins of strain HCP2 was compared in different types of tests. Results indicated that the use of the purified pyocins for the typing of P. aeruginosa provides data which are easier to control and interpret. KEY-WORDS: Pseudomonas aeruginosa, Bacteriocin, Pyocin, Pyocin typing; Purification. INTRODUCTION Pyocin, a bacteriocin produced by Pseudomonas aeruginosa, was first reported by Jacob [i5]. Pyocins have been classified into three groups : types R, F and S. R-type and F-type pyocins are very large particles having a struc- ture similar to bacteriophage tails T and ),, respectively [13, 19, 21]. A third group of pyocins, named S-type, is distinguished from the above two types by its sensitivity to proteases and a higher diffusion rate on agar plates [14]. The epidemiological importance of P. aeruginosa in nosocomial infections has encouraged the development of various typing techniques which allow us to observe the evolution of the infections associated with this microorga- Manuscrit re~u le 11 septembre 1985, accept6 le 27 mars 1986.
Transcript of Possibility of using purified pyocins for typing Pseudomonas aeruginosa: Purification of pyocins and...

(~) ELSEVIER Paris 1986
Ann. lnst. Pasteur/Microbiol. 1986, 137 A, 253-266
POSSIBILITY OF USING PURIFIED PYOCINS FOR TYPING
PSEUDOMONAS AERUGINOSA : PURIFICATION OF PYOCINS
AND SENSITIVITY OF P. AERUGINOSA
IN DIFFERENT TESTS
by D. Jurado Chacon (1), A. Chueca Sancho (2), J.F. Guillen Solvas (1), B. Garcia-Villanova Ruiz (3), and R. Galvez Vargas (3)
Q) Medicina Preventiva, Escuela de Enfermeria, Universidad de Granada, (2) Estacidn Experimental Zaidin (CSIC), Granada, and
(3) Cdtedra de Medicina Preventiva, Facultad de Medicina, Universidad de Granada (Espaha).
SUMMARY
Two types of pyocins were simultaneously found in Pseudomonas aerugi- nosa strains HCP2. According to their structures, they belonged to the types classified as R and F, respectively, and were named HCP2-R and HCP2-F. The sensitivity of 87 strains of P. aeruginosa of clinical origin to pyocins of strain HCP2 was compared in different types of tests. Results indicated that the use of the purified pyocins for the typing of P. aeruginosa provides data which are easier to control and interpret.
KEY-WORDS: Pseudomonas aeruginosa, Bacteriocin, Pyocin, Pyocin typing; Purification.
INTRODUCTION
Pyocin, a bacteriocin produced by Pseudomonas aeruginosa, was first reported by Jacob [i5]. Pyocins have been classified into three groups : types R, F and S. R-type and F-type pyocins are very large particles having a struc- ture similar to bacteriophage tails T and ),, respectively [13, 19, 21]. A third group of pyocins, named S-type, is distinguished from the above two types by its sensitivity to proteases and a higher diffusion rate on agar plates [14].
The epidemiological importance of P. aeruginosa in nosocomial infections has encouraged the development of various typing techniques which allow us to observe the evolution of the infections associated with this microorga-
Manuscrit re~u le 11 septembre 1985, accept6 le 27 mars 1986.

254 D. JURADO CHACON AND COLL.
nism. These techniques are: serological typing, bacteriophage typing, pyocin typing, pyocin sensitivity typing, biotyping and antibiogram. Most authors recommend the use of O-serological typing as the primary epidemiological marker. If a greater differentiation is necessary, this technique should be complemented with another method, such as pyocin typing [1, 6, 24].
Pyocin typing depends on the product ion of pyocin by the strain under investigation. The method of Gillies and Govan is used world-wide [8]. Pyo- cin typing, hovewer, presents certain obstacles not only in terms of its stan- dardization, but also in terms of the technique itself, due to the limited possibilities of guaranteeing the correct typing of all isolated P. aeruginosa strains. The disadvantages associated with this technique, as well as new methods, have been investigated in other studies [2, 4, 6, 23]. Typing by means of sensitivity patterns to a standard set of pyocin lysates involves certain pro- blems as well. First of all, upon inducing the product ion of the pyocins, bac- teriophages are also simultaneously induced [11, 25]. In addition, one must take into account the fact that one strain may produce more than one type of pyocin, and therefore a superposition of reactions may occur [14, 21, 24]. Lastly, the presence of residual growth in the inhibited zones makes it very difficult to offer an objective interpretation of results [7, 16, 23]. Other authors [3, 23] have proposed the use of purified pyocins in pyocin sensitivity typing, whereby the results obtained would be much clearer and easier to read objec- tively. Thus, the disadvantages of this technique could be avoided.
In this work, we describe the purification of two pyocins, HCP2-R and HCP2-F, and a comparative study of the sensitivity of 87 strains of P. aeru- g inosa of clinical origin toward pyocins of the strain P. aeruginosa HCP2 in different types of tests. We first studied the sensitivity of these strains toward the pyocins produced by the strain HCP2 when cultivated on a solid medium according to the pyocin typing technique of Gillies and Govan [8]. Secondly, the sensitivity to pyocins was studied with the following pyocin preparations : a) mitomycin-C-induced lysate obtained from the producer strain HCP2; b) dialysed precipitate of (NH4)2SO 4 of the lysate; c) HCP2-R and HCP2-F pyocins in a purified state; d) << final fraction >> of the purification process. Because the supernatant which constitutes the <~ final fraction >> of the purifi- cation process showed activity toward a group of indicator strains (data not shown), we decided to include it in this study.
MATERIALS AND METHODS
Bacterial strains.
Producer strain. - - P. aeruginosa strain HCP2 from our collection was used for the preparation of pyocins HCP2-R and HCP2-F.
Indicator strains. - - We used P. aeruginosa GG1-8 and GGA-E of Gillies and Govan [8, 9] and Kageyama's strains P. aeruginosa PML1550, PML1516, PML15163, PML15163-b, PML15163-d, PML15163-e and PML15163-f [19].
Examined strains. -- A total of 87 strains of P. aeruginosa, isolated in the Labo- ratory of Microbiology from pathological products of patients in the Hospital Cli-

P Y O C I N T Y P I N G O F P. A E R U G I N O S A 255
nico S. Cecilio, Granada (Spain) were used. Each of the strains was from a different patient, with the exception of a certain number of strains belonging to one patient who presented a different pyocin type.
Media.
The G medium [18] was used as the culture medium for pyocin preparation. The indicator and the examined strains were cultivated in triptic soy agar (Difco).
Production and purification of pyocins.
Both pyocins were purified simultaneously from one litre of the mitomycin-C- induced lysate of P. aeruginosa strain HCP2. The purification of pyocin HCP2-R reported earlier [17] was slightly modified as follows. To one litre of lysate, 500 g of (NH4)2SO 4 were added and it was then left overnight at 4~ The precipitate was collected by centrifugation at 4~ and dissolved in 66 ml of 10 mM sodium phos- phate buffer (pH 6.8) 10 mM Na 3 EDTA. The insoluble material was centrifuged off and the sample was dialysed against 10 litres of 8 mM sodium phosphate 0.5 mM EDTA buffer (pH 6.8) for 10 h, then centrifuged for 1 h at 52,000 g at 4~ The precipitate thus obtained was dissolved in 8 ml of 10 mM sodium phosphate buffer (pH 6.8) and chromatographed on diethylaminoethyl-cellulose (DEAE) column (2.5 • 6 cm), washed beforehand with the same buffered saline and eluted with 400 ml of a sodium chloride gradient (0 to 0.5 M in 10-mM sodium phosphate buffer). Active fractions of DEAE-cellulose chromatography were not concentrated by vacuum fil- tration as described [17], since we observed a loss of activity. The unconcentrated sample was applied to a <~Bio-Gel>> A-15m column (1.6• 55 cm), which had been washed beforehand with l0 mM sodium phosphate buffer (pH 6.8) containing 0.2 M NaC1 and eluted with the same buffer.
To obtain the pyocin HCP2-F in a purified state, we started from the superna- tant obtained after an ultracentrifugation at 52,000 g for 1 h as indicated in figure 1. After two more centrifugations, one at 72,000 g at 4~ for 1 h and the second at 150,000 g for 3 h at 4~ the precipitate was applied on a <~ Bio-Gel >> A-15m column (1.6 • 55 cm), the same as for pyocin HCP2-R, except that the column was washed with 10 mM sodium phosphate buffer (pH 6.8) containing 0.14M NaC1, and the sam- ple was eluted with the same buffered saline.
Assay of pyocin activity.
Pyocin activity was estimated by spotting serially diluted samples on a lawn of indicator strain (P. aeruginosa PML1516) on tryptic soy agar [18]. The highest dilu- tion which gave a discernible inhibition of growth was taken as the activity unit.
Assays of sensitivity to pyocins.
The sensitivity of the 87 strains to pyocin preparations was determined in the fol- lowing manner.
a) Effect of the pyocins produced on agar medium : the effect of pyocin produ- ced by the pyocinogenic strain P. aeruginosa HCP2 (when the latter was cultivated on solid medium) upon the 87 examined strains was determined using the pyocin typing technique of Gillies and Govan [8]. Of the 87 strains, a maximum of 8 were streaked on each plate.
b) Activity of mitomycin-C-induced lysate, dialysed precipitate of (NH4)2SO4, purified pyocins and << final fraction >>: using bacterial suspensions of 103 to 104 orga- nism in 1 ml of sterile physiological saline, the 87 strains of P. aeruginosa were spread in layers upon the surface of the plate of triptic soy agar. After the plate had dried,

256 D. J U R A D O C H A C O N A N D C O L L .
drops of each cited fraction were deposited o n th e plate. Once the drops were dry, the plates were incubated overnight at 37~ Afterwards, observations regarding activity and the appearance of resistant colonies in some of the inhibition zones were recorded.
E l e c t r o n m i c r o s c o p y .
Each of the fractions obtained in the various stages of purification were studied under the electron microscope. Regarding the chromatographical processes, the frac- tions constituting the peaks of absorption (at 280 nm) and the activity, as well as the fractions next to these, were observed.
Negatively stained specimens were prepared by depositing a drop of each frac- tion on a carbon-coated grid and adding a drop of 1 % phosphotungstate neutrali- zed with KOH. The excess solution was withdrawn by touching the edge of the grid with filter paper. When dried, the specimens were ready for electron microscopy using a type <~ EM-300 Phillips >>.
RESULTS
P u r i f i c a t i o n o f p y o c i n s .
The pyocins H C P 2 - R and H C P 2 - F were pur i f i ed acco rd ing to the proce- du re descr ibed earl ier (fig. 1). T h e y were i sola ted f r o m a litre o f mi tomyc in -
f
Precipitate (discard)
Supernatant (discard)
I
Supernatant I
Centrifugation at 72,000 g
i w Precipitate Supernatant (discard) I
Centrifugation at 150,000 g
I I I
Supernatant Precipitate (final fraction) I Chromatography on
Bio-Gel A-15m +
HCP2-F pyocin
Mitomycin C-induced lysate I
Centrifugation at 5,000 g I !
Supernatant (NH4) 2SO4
Centrifugation at 11,000 g I !
Precipitate I
Dialysis I
Centrifugation at 52,000 g I
I
Precipitate I
Chromatography on DEAE I
Two peaks with pyocin activity
I Collected fractions
of peak I
I Chromatography on '
Bio-Gel A= 15m $
HCP2-R pyocin
FIG. 1. - - Purification procedure fo r pyocins HCP2-R and HC12-F.

PYOCIN TYPING OF P. A E R U G I N O S A 257
C-induced lysate of the P. aeruginosa HCP2. Both pyocins were observed in the lysate, showing a structure similar to that of types R and F. In addi- tion, entire bacteriophages (morphotype B1) were observed with a sheath whose basal plaque structure suggested that of a sheath o f the bacteriophage B1, empty heads, empty sheaths, polysheaths, etc. (fig. 2).
J
D
I=
Eli:
FIG. 2. i Electron micrographs of a mitomycin-C-induced lysate of strain HCP2. A) pyocin HCP2-R; B) pyocin HCP2-R, bacteriophages morphotype B1, empty heads;
C) polysheath ; D) empty sheath ; E) pyocin HCP2-F (x 168,000).

258 D. JURADO CHACON AND COLL.
The chromatographic profile on DEAE-52-cellulose for the HCP2-R pyocin is illustrated in figure 3. The activity of the pyocin began to elute at 0.15 M of NaC1. An electron microscopic study showed that both peaks contained a large quantity of the HCP2-R pyocin. In addition, bacteriophages could be seen in the second peak, although the proport ion of these was lesser than
J J
J 10
j
I It \
20 zo 40 5b 8b F r a c t i o n n tuaber
5 I0
10 3 .~
IO I
FIG. 3. - - The elution profile o f the partially purified HCP2-R pyocin on DEAE-52 cellulose. �9 = opt ica l dens i ty at 280 n m ; . . . . act ivi ty o f pyoc in H C P 2 - R . T h e f low rate was 30 m l / h ,
1 f r a c t i o n was 6 ml .
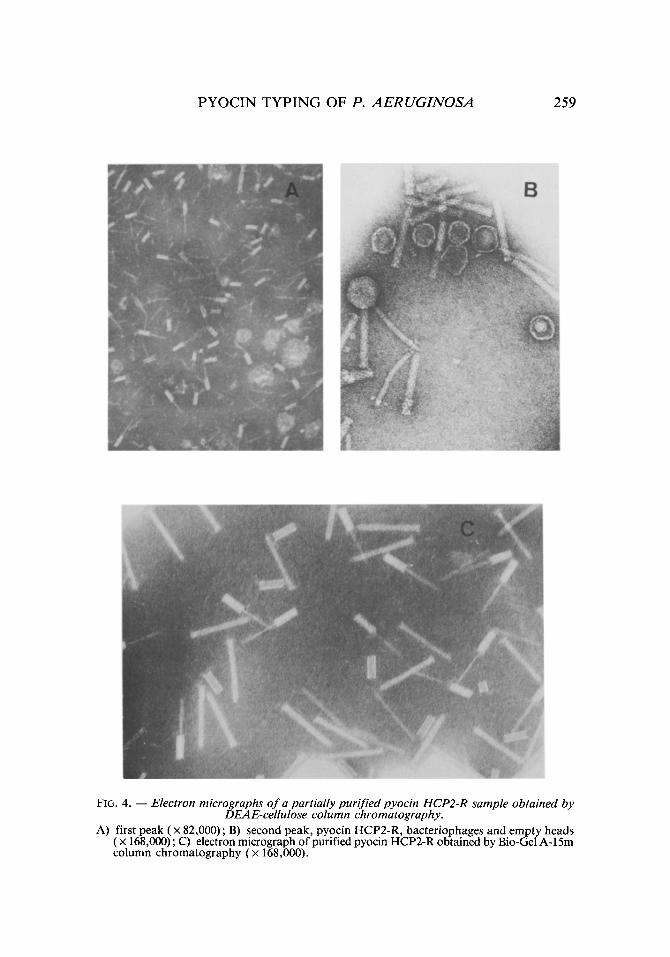
that of fhe pyocins (figs. 4A and 4B). In order to attain a better purification, this peak was discarded in spite of the activity of its fractions. Only the frac- tions of greatest activity forming the first peak were collected. The volume of the resulting sample was applied to a <(Bio-Gel >> A-15m column, eluting the HCP2-R pyocin in the first fractions (figs. 4C and 5a).
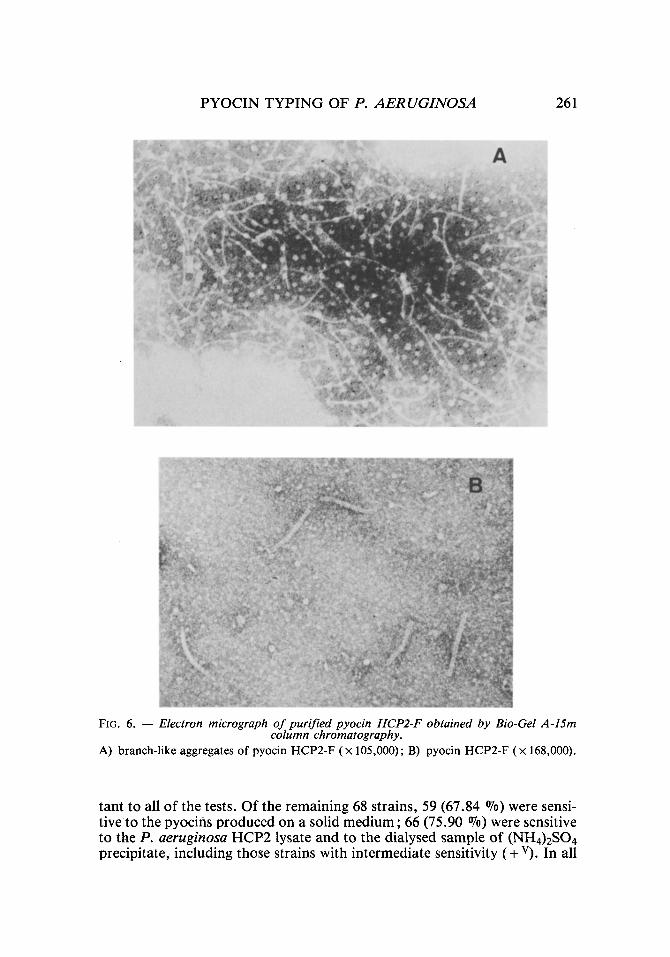
For the purification of the HCP2-F pyocin, centrifugation at 72,000 g was necessary to eliminate the remains of the HCP2-R pyocin and the large quantity of empty sheaths that had appeared in the precipitate. The chromatographic profile on << Bio-Gel >> A-15m column for the HCP2-F pyocin is illustrated in figure 5b. Two peaks of absorption at 280 nm resulted, eluting the HCP2-F pyocin in the first fractions (figs. 5b and 6).
Assays of activity to pyocins.
The sensitivity or resistance of examined strains to pyocins produced by the P. aeruginosa strain HCP2 in the different tests is expressed in
P Y O C I N T Y P I N G O F P. A E R U G I N O S A 259
FIG. 4. - - Electron micrographs of a partially purified pyocin HCP2-R sample obtained by DEAE-cellulose column chromatography.
A) first peak ( • 82,000) ; B) second peak, pyocin HCP2-R, bacteriophages and empty heads ( x 168,000); C) electron micrograph of purified pyocin HCP2-R obtained by Bio-Gel A-15m column chromatography (x 168,000).

260 D . J U R A D O C H A C O N A N D C O L L .
, ~ 0 , 2 '
o
g
} I A
I I
I ' I
10 20 30 Fraction g~
b
i I q t I
/ - I I I I
xo Z~ 30"
~10 5
"4 a
10 3 .~
l o I
FIG. 5. - - Gel filtration o f pyocin HCP2-R (a) and pyocin HCP2-F (b) on Bio-Gel A-15m. �9 = optical density at 280 n m ; . . . . ac t iv i ty o f p y o c i n s ; t h e f low rate was 36 m l / h , 1 f rac-
t i o n was 6 ml .
table I. The sign + v represents inhibitions in which 50-100 resistant colo- nies appeared and which therefore could lead to false interpretations. In tables I and II, one can observe that only 19 of the strains (21.84 ~ proved resis-
TABLE I . - - D i s t r i b u t i o n o f t h e e x a m i n e d s t r a i n s a c c o r d i n g t o t h e s e n s i t i v i t y o r r e s i s t a n c e t o p y o c i n s p r o d u c e d b y t h e P. aeruginosa s t r a i n H C P 2
in t h e d i f f e r e n t t e s t s .
A g a r Mitomycin-C- D i a l y s e d precipi- P y o c i n P y o c i n Final Strains m e d i u m i n d u c e d lysate tate o f (NH4)2SO 4 H C P 2 - R H C P 2 - F fract ion
+ + + + + + 3 7 + + + + + + v 2
+ + + + + - I 1 + v + + + + + 3 + v + + + + - 1 + v + + + + v _ 1 + v + v + v + v _ _ 1 + v + v + v - + v + v l + v + v + v - - - 1 + v - - - + + v 1
- + + - - + 1 - + v + v - - + v 1 _ + v + v - - - 6
. . . . + - 1
. . . . . . 19
S u m 8 7
PYOCIN TYPING OF P. AERUGINOSA 261
FIG. 6. - - Electron micrograph of purified pyocin HCP2-F obtained by Bio-Gel A-15m column chromatography.
A) branch-like aggregates of pyocin HCP2-F (x 105,000); B) pyocin HCP2-F (x 168,000).
tant to all of the tests. Of the remaining 68 strains, 59 (67.84 ~ were sensi- tive to the pyocins produced on a solid medium; 66 (75.90 070) were sensitive to theP . aeruginosa HCP2 lysate and to the dialysed sample of (NH4)2SO 4 precipitate, including those strains with intermediate sensitivity (+ v). In all
262 D. J U R A D O C H A C O N AND COLL.
cases, the sensitivity toward both samples was identical. Of these strains, 55 (64.37 %) were sensitive to the HCP2-R pyocin and 58 (66.70 %) to the HCP2-F pyocin. Only one strain showed intermediate sensitivity to the HCP2-R pyocin, but no sensitivity to HCP2-F. However, three strains which were sensitive to HCP2-F pyocin showed no sensitivity to HCP2-R. In addi- tion, 8 strains which were sensitive to the lysate and the dialysed precipitate of (NH4)2SO 4 were resistant to both pyocins produced by strain HCP2 on a solid medium and to the purified pyocins. On the other hand, a high pro- port ion of the strains (52.86 07o) were sensitive to the ~< final fraction >~ of the purification process as well.
As can be seen, the greatest number of strains possessing intermediate sen- sitivity corresponded to those tested with the dialysed precipitate and lysate (11.50 07o) and to the cases when a solid medium was used, as in the first of the assays (10.34 07o). Nevertheless, using the purified HCP2-R and HCP2-F pyocins, the number of strains which showed intermediate sensitivity was only 1.15 070 and 2.30 ~ respectively (table II).
TABLE II. - - Number and frequency of the strains according to sensitivity to pyocins in the different tests.
Test Sensitive strains Intermediate sensitivity Total sensitivity
(+) (+v) Nb % Nb % Nb %
Agar medium 50 57.50 9 10.34 59 67.84 Mitomycin-C-in- 56 64.40 10 11.50 66 75.90 duced lysate Dialysed preci- 56 64.40 10 11.50 66 75.90 pitate of (NH4)2SO 4 Pyocin HCP2-R 55 63.22 1 1.15 56 64.37 Pyocin HCP2-F 56 64.40 2 2.30 58 66.70 Final fraction 41 47.12 5 5.74 46 52.86
Finally, the sensitivity of these 87 strains toward the extracts and purified pyocins cannot be attributed to a lysogenic phenomenon in these strains, as was demonstrated by habitual lysogenic tests.
DISCUSSION
P. aeruginosa strain HCP2 was found to be the producer of two pyocins, HCP2-R and HCP2-F, whose structures correspond to types R and F, res- pectively. They can be separated by means of differential ultracentrifugation at 52,000 g 1 h, 72,000 g 1 h and 150,000 g 3 h. The second centrifugation
P Y O C I N T Y P I N G OF P. A E R U G I N O S A 263
was necessary in order to separate the two pyocins. As the precipitate o f this second centrifugation contained pyocin H C P 2 - R as well as empty sheaths which seemed to lack activity [10], it was discarded. In addition, strain H C P 2 was the producer of a bacter iophage of morpho type B, which was separated from the pyocins by chromatography on DEAE. Our results therefore coincide with those of Ikeda et al. [12], who, in parallel, found that the P. aeruginosa strain PML15 produced pyocin R1 and a phage at the same time. They also separated them using the same method that we used.
The action spectrum of purified H C P 2 - R and H C P 2 - F pyocins on various known indicator strains was different f rom that o f other R- and F-type pyocins [14, 19, 22]. However , since H C P 2 - R and H C P 2 - F possess a very similar action spectrum (table III), the use of both pyocins at the same time in the typing of P. aeruginosa is not recommended.
TABLE III. - - Ac t ion spectra of pyoc ins H C P 2 - R and H C P 2 - F .
Indicator strain Pyocin
GG1 2 3 4 5 6 7 8 A B C D E
PML 1550 1516 15163 15163-b 15163-d 15163-e 15163-f
HCP2-R HCP2-F + + + +
+ +
+
+ + + + +
+ + + + + +
+ +
+ = inhibition; - = no inhibition.
A high propor t ion of the 87 examined strains was sensitive to the pyocins produced by strain H C P 2 in each of the tests carried out. However, the number of strains of intermediate sensitivity ( + v) varies greatly f rom one test to the

264 D. JURADO CHACON AND COLL.
other. In fact, the number of strains of intermediate sensitivity obtained in the first 2 assays (10.34 070 and 11.50 070, respectively) was superior to the results of the assay with the purified pyocins HCP2-R and HCP2-F (1.15 ~ and 2.30 070, respectively). These results could respond to the consi- derations of authors such as Merrikin et al. or Farkas-Himsley et al. [3, 23] regarding the practicality of using purified pyocins for the typing of P. aeru- ginosa.
The appearance of residual growth in the inhibition zones when the pro- ducer strain was cultivated on a solid medium (according to the pyocin typing technique) could be due to various mechanisms, such as a low pro- duction of pyocins [4], an instability in this production [2] or the secre- tion of certain enzymes produced by the strain itself, which inactivate the pyocins [26].
It is difficult to explain the cause of the activity in the cases of interme- diate sensitivity, as obtained for the lysate and the dialysed precipitate of (NH4)2SO4, considering the fact that 8 of the strains with this sensitivity were resistant to the pyocins produced on a solid medium as well as to those in a purified state. This activity could be due to multiple metabolites present in the ~ final fraction)). In the other 6 strains, the intermediate sensitivity could be explained by the presence of bacteriophages in the above extracts. These phages were later eliminated in the process of purification of the two pyocins as mentioned before.
In summary, we should underline the fact that the typing of P . aerugi- nosa using purified pyocins presents some advantages compared to other typing methods considering the criteria of Ashenov [24] or of Brokopp et al. [1]. For instance, the results of this method can be interpreted with greater cer- tainty. In addition, the activity of the assay and its reproductivity are easier to quantify and therefore, more controllable. This greater degree of control is due to the stability of the pyocins [18, 21]. We have managed to maintain the activity of the purified pyocins HCP2-R and HCP2,F at 4~ for 4 months and the activity of the pyocin HCP2-F by lyophilization of a sample of puri- fied pyocin, to which an equal part of milk was added (data not shown). These results do not coincide with those obtained by Kageyama et aL [18] in which the activity of the pyocin R1 was destroyed by lyophilization, We are cur- rently studying the effectiveness of this method in our laboratory, Finally, typing with purified pyocins could be easily standardized, on the one hand by the use of multi-inoculators and, on the other hand, using simple spectro- photometric assays such as described recently by Kumazaki et al. [20]. We can thus conclude that once the efficiency of a concrete number of pyocins on the typing of P. aeruginosa and their stability in conservation have been determined, these pyocins could be commercialized and their utilization thereby facilitated.
In our opinion, this method, together with that o f serotyping (given the simplicity of the latter), could resolve the majori ty of the problems currently encountered in the typing of P. aeruginosa.

PYOCIN TYPING OF P. A E R U G I N O S A 265
RI~SUMI~
UTILISATION DE PYOCINES PURIFII~ES POUR LE TYPAGE DE PSEUDOMONAS AERUGINOSA : PURIFICATION DES PYOCINES
ET SENSIBILITI~ DE P. AERUGINOSA I~TUDII~E AVEC DIVERS TESTS
La souche Pseudomonas aeruginosa HCP2 produit simultan6ment deux types de pyocines: HCP2-R et HCP2-F. Elles appartiennent par leur struc- ture, l 'une au type R et l 'autre au type F. Nous avons 6tudi6 la sensibilit6 de 87 souches de P. aeruginosa d'origine clinique aux pyocines de la souche HCP2, par le moyen de tests divers. L'emploi des pyocines purifi6es fournit des donn6es mieux interpr6tables et plus facilement contr61ables que celles obtenues avec les cultures de souches pyocinog~nes.
MOTS-CLI~S: Pseudomonas aeruginosa, Bact6riocine, Pyocine, Pyocino- typie; Purification.
ACKNOWLEDGMENTS
This work was supported by an << Investigaciones Sanitarias ~> fellowship (FISSS) to D.J.C.
REFERENCES
[1] BROKOPP, C.D. & FARMER, J.J., III, Typing methods for Pseudomonas aeru- ginosa, in ~(Pseudomonas aeruginosa. Clinical manifestations of infection and current therapy~ (R.G. Dogget) (p. 89). Academic press, New York, London, 1979.
[2] CHADWICK, P., The significance of pattern variations in pyocine typing of Pseu- domonas aeruginosa. Canad. J. Microbiol., 1972, 18, 1153-1158.
[3] FARKAS-HIMSLEY, H. t~r PAGEL, A., Bacteriocin typing by leakage of ultraviolet light-absorbing material. Infect. Immun., 1977, 16, 12-19.
[4] FARMER, J.J. III & HERMAN, L.L.G., Epidemiological fingerprinting of Pseu- domonas aeruginosa by the production of and sensitivity to pyocin and bacteriophage. AppL MicrobioL, 1969, 18, 760-765.
[5] FARMER, J.J. III& HERMAN, L.L.G., Pyocin typing of Pseudomonas aerugi- nosa. J. infect. Dis., 1974, 130 (suppl.), 43-45.
[6] FYEE, J.A.M., HARRIS, G. & GOVAN, J.R.W., Revised pyocin typing method for Pseudomonas aeruginosa. J. cfin. MicrobioL, 1984, 20, 47-50.
[7] GARCIA-QuINTANA, H.G. & FLORES, M.E., Inhibition de la croissance de popu- lations bact6riennes par des suspensions obtenues de Pseudomonas aerugi- nosa pyocinog6niques. Canad. J. Microbiol., 1983, 29, 471-475.
GILLIES, R.R. & GOVAN, J.R.W., Typing of Pseudomonas pyocyanea by pyo- cine production. J. Path. Bact., 1966, 91, 339-345.
GOVAN, J.R.W. & GILLIES, R.R., Further studies in the pyocin typing of Pseu- domonas pyocyanea. J. reed. Microbiol., 1969, 2, 17-25.
HIGERD, T., BAECHLER, C.A. & BERK, R.S., In vitro and in vivo characteriza- tion of pyocin. J. Bact., 1967, 93, 1976-1986.
[8]
[9]
[lO]

266 D. J U R A D O C H A C O N A N D COLL.
[11] HOLLOWAY, B.W. & KRISHNAPILLAI, V., Bacteriophages and bacteriocins, /n ~ Genetics and biochemistry of Pseudomonas ~ (P.H. Clarke and M.H. Rich- mond) (p. 99). John Wiley and Sons Ltd, London, 1975.
[12] IKEDA, K., KAGEYAMA, M. & EGAMI, F., Studies of a pyocin. II. - - Mode of production of the pyocin. J. Biochem., 1964, 55, 54-58.
[13] ISHII, S., NISHI, Y. & EGAMI, F., The fine structure of a pyocin. J. tool Biol., 1965, 13, 428-431.
[14] ITO, S., KAGEYAMA, M. & EGAMI, F., Isolation and characterization of pyocins from several strains of Pseudomonas aeruginosa. J. gen. appL Microbiol., 1970, 16, 205-214.
[15] JACOB, F., Biosynth~se induite et mode d'action d'une pyocine, antibiotique de Pseudomonas pyocyanea. Ann. Inst. Pasteur, 1954, 86, 149-160.
[16] JONES, L.F., ZAr, ANYCZ, J.P., THOMAS, E.T. & FARMER, J.J. III, Simplified method for producing pyocin from Pseudomonas aeruginosa: a simplified method. Appl. Microbiol., 1973, 26, 120-121.
[17] JURADO CHACON, D., CHUECA SANCHO, A., MARISCAL LARRUBIA, A., CUETO ESPINAR, A. & GALVEZ VARGAS, R., Purificaci6n de una bacteriocina de estructura contr~ictil obtenida de Pseudomonas aeruginosa. Laboratorio, 1985, 79, 315-324.
[18] KAGEYAMA, M. & EGAMI, F., On the purification and some properties of a pyocin. A bacteriocin produced by Pseudomonas aeruginosa. Life Sci., 1962, 9, 471-476.
[19] KAGEYAMA, M., Bacteriocins and bacteriophages in Pseudomonas aeruginosa, in t~Microbial drug resistances> (S. Mitsuhashi & J. Hashimoto) (p. 291). University Park Press, Baltimore, 1975.
[20] KUMAZAKI, T. & ISHII, S., A simple photometric method for determination of the activity of pyocin R1. J. Biochem., 1982, 91, 817-823.
[21] KURODA, K. & KAGEYAMA, M., Biochemical plJoperties of a new flexuous bacteriocin pyocin F1 produced by Pseudomonas aeruginosa. J. Biochem., 1979, 85, 7-19.
[22] KURODA, K. & KAGEYAMA, M., Comparative study on F-type pyocins of Pseu- domonas aeruginosa. J. Biochem., 1981, 89, 1721-1736.
[23] MERRIKIN, D.J. & TERRY, C.S., Variability of pyocine type and pyocine sensi- tivity in some strains of Pseudomonas aeruginosa. J. appl. Bact., 1972, 35, 667-672.
[24] PITX, T.L., State of the art: typing Pseudomonas aeruginosa. J. Hosp. Infect., 1980, 1, 193-199.
[25] RAMPLING, A., WHITBY, J.L. & WILDY, P., Pyocin-sensitivity testing as a method of typing Pseudomonas aeruginosa: use of ~ phage-free >> prepara- tions of py0cin. J. reed. Microbiol., 1975, 8, 531-541.
[26] WHABA, A.H., The production and inactivation of pyocines. J. Hyg. (Camb.), 1963, 61, 431-441.




















