
1-s2 0-S0733521013000921-main
9
Electrophoretic analysis of malting degradability of major sorghum reserve proteins Gaone Mokhawa a , Charity K. Kerapeletswe-Kruger a , Lewis I. Ezeogu a, b, * a National Food Technology Research Centre, 1840 Mpuutsane Industrial Area, P/Bag 008, Kanye, Botswana b Dept. of Microbiology, University of Nigeria, Nsukka, Enugu State, Nigeria article info Article history: Received 15 January 2013 Received in revised form 24 April 2013 Accepted 6 May 2013 Keywords: Sorghum malting Proteolysis Kafirin Free a-amino nitrogen abstract Proteolysis is vital to the generation of amino acids and short peptides during malting. The qualitative and quantitative effects of malting on proteolytic digestion were investigated for 11 Botswana sorghum cultivars. Protein hydrolysis was influenced by sorghum grain cultivar. All protein fractions were degraded, although the extents of their digestion appeared cultivar-dependent. The most significant changes in total and free kafirins occurred among the HMW aggregates while the 45 kDa dimer appeared the most recalcitrant. Free kafirin monomers were digested in variety-defined manner; a-kafirin faded the most (ca. 82%) in Phofu, but appeared least digested (10%) in Lekgeberwa; b-kafirin faded the most in Phofu and Segaolane (90e94%) but only 46% in Lekgeberwa. Overall, a-kafirin appeared the least digested of the free monomers. The proportion of free kafirin accounted for by the 45 kDa and mono- meric kafirins increased in all malts (except Lars Vyt and Mafia), probably due to protein depolymer- isation. Grain variety significantly influenced (p < 0.001) malt FAN levels. Mean FAN values were highest in Segaolane (225 mg/100 g) followed by Sefofu, Town and BSH-1 , but lowest in Mafia (96.9 mg/100 g). Results indicate wide variability in the proteolytic malting digestibility of the eleven sorghum cultivars. Ó 2013 Elsevier Ltd. All rights reserved. 1. Introduction The hydrolysis of proteins constitutes one of the most important endosperm mobilization processes occurring during seed germi- nation. It provides the growing seedling with raw materials (amino acids and short peptides) for enzyme and seedling structure synthesis and facilitates the enzymatic hydrolysis of starch (Palmer, 1989). In barley, a carboxypeptidase, b-glucan solubilase, is responsible for the initial phases of cell wall hydrolysis and b-D- glucan solubilisation (through hydrolysing the bonds between b- glucan in the grain’s cell wall and its protein matrix) (Palmer, 1989). The most critical function of germination-related endosperm pro- teolysis, vis-á-vis the industrial and nutritional uses of cereal malts, appears to be the provision of free a-amino nitrogen (FAN), a mixture of amino acids, ammonium ions and short peptides. In brewing, FAN are vitally important, both as major sources of assimilable nitrogen for rapid yeast growth and ethanol production and for being centrally involved in the complex biochemical pro- cesses that culminate in beer flavour development (Lekkas et al., 2007). In human nutrition, germination-related proteolysis en- hances protein and starch availabilities while the soluble peptides and amino acids liberated during malting serve as ready sources of assimilable nitrogen. Sorghum (Sorghum bicolor L.) is ranked fifth among the world’s major cereal crops. Across Africa, malted sorghum has grown increasingly in industrial use, as a replacement for malt barley in lager beer brewing (Dufour et al., 1992) and as a source for readily assimilable nutrients in high nutrient density food formulations (Correia et al., 2008). Sorghum endosperm proteins are heterogeneous. Prolamins constitute 77e82% of the total proteins (Hamaker et al., 1995), while albumins and globulins are minor components. Sorghum prolamins (kafirins) are mixtures of monomeric and aggregated (oligomeric and polymeric) proteins. Monomeric kafirins include a- (23 kDa and 25 kDa), b- (20 kDa) and g- (28 kDa) kafirins (Shull et al., 1991; Mazhar et al., 1993). a-Kafirins comprise 60e70% of total sorghum protein (and ca. 80% of total kafirins), followed by the g- (ca. 15% of total kafirins), and b-monomers (ca. 5% of total kafirins) (Hamaker et al., 1995; de Mesa-Stonestreet et al., 2010). Abbreviations: CBB-R250, Coomassie brilliant blue R250 dye; FAN, Free a-amino nitrogen; HMW, high molecular weight; 2-ME, 2-mercaptoethanol; M r , Molecular mass; SDS, sodium dodecyl sulphate; SDS-PAGE, Sodium dodecyl sulphate poly- acrylamide gel electrophoresis. * Corresponding author. National Food Technology Research Centre, 1840 Mpuutsane Industrial Area, P/Bag 008, Kanye, Botswana. Tel.: þ267 5445516; fax: þ267 5440713. E-mail addresses: [email protected], [email protected] (L.I. Ezeogu). Contents lists available at SciVerse ScienceDirect Journal of Cereal Science journal homepage: www.elsevier.com/locate/jcs 0733-5210/$ e see front matter Ó 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.jcs.2013.05.008 Journal of Cereal Science xxx (2013) 1e9 Please cite this article in press as: Mokhawa, G., et al., Electrophoretic analysis of malting degradability of major sorghum reserve proteins, Journal of Cereal Science (2013), http://dx.doi.org/10.1016/j.jcs.2013.05.008
Transcript of 1-s2 0-S0733521013000921-main

at SciVerse ScienceDirect
Journal of Cereal Science xxx (2013) 1e9
Contents lists available
Journal of Cereal Science
journal homepage: www.elsevier .com/locate/ jcs
Electrophoretic analysis of malting degradability of major sorghumreserve proteins
Gaone Mokhawa a, Charity K. Kerapeletswe-Kruger a, Lewis I. Ezeogu a,b,*
aNational Food Technology Research Centre, 1840 Mpuutsane Industrial Area, P/Bag 008, Kanye, BotswanabDept. of Microbiology, University of Nigeria, Nsukka, Enugu State, Nigeria
a r t i c l e i n f o
Article history:Received 15 January 2013Received in revised form24 April 2013Accepted 6 May 2013
Keywords:Sorghum maltingProteolysisKafirinFree a-amino nitrogen
Abbreviations: CBB-R250, Coomassie brilliant bluenitrogen; HMW, high molecular weight; 2-ME, 2-memass; SDS, sodium dodecyl sulphate; SDS-PAGE, Sodacrylamide gel electrophoresis.* Corresponding author. National Food Technol
Mpuutsane Industrial Area, P/Bag 008, Kanye, Botfax: þ267 5440713.
E-mail addresses: [email protected], lewis@nafte
0733-5210/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.jcs.2013.05.008
Please cite this article in press as: MokhawJournal of Cereal Science (2013), http://dx.d
a b s t r a c t
Proteolysis is vital to the generation of amino acids and short peptides during malting. The qualitativeand quantitative effects of malting on proteolytic digestion were investigated for 11 Botswana sorghumcultivars. Protein hydrolysis was influenced by sorghum grain cultivar. All protein fractions weredegraded, although the extents of their digestion appeared cultivar-dependent. The most significantchanges in total and free kafirins occurred among the HMWaggregates while the 45 kDa dimer appearedthe most recalcitrant. Free kafirin monomers were digested in variety-defined manner; a-kafirin fadedthe most (ca. 82%) in Phofu, but appeared least digested (�10%) in Lekgeberwa; b-kafirin faded the mostin Phofu and Segaolane (90e94%) but only 46% in Lekgeberwa. Overall, a-kafirin appeared the leastdigested of the free monomers. The proportion of free kafirin accounted for by the 45 kDa and mono-meric kafirins increased in all malts (except Lars Vyt and Mafia), probably due to protein depolymer-isation. Grain variety significantly influenced (p < 0.001) malt FAN levels. Mean FAN values were highestin Segaolane (225 mg/100 g) followed by Sefofu, Town and BSH-1, but lowest in Mafia (96.9 mg/100 g).Results indicate wide variability in the proteolytic malting digestibility of the eleven sorghum cultivars.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
The hydrolysis of proteins constitutes one of the most importantendosperm mobilization processes occurring during seed germi-nation. It provides the growing seedling with raw materials (aminoacids and short peptides) for enzyme and seedling structuresynthesis and facilitates the enzymatic hydrolysis of starch(Palmer, 1989). In barley, a carboxypeptidase, b-glucan solubilase, isresponsible for the initial phases of cell wall hydrolysis and b-D-glucan solubilisation (through hydrolysing the bonds between b-glucan in the grain’s cell wall and its protein matrix) (Palmer, 1989).The most critical function of germination-related endosperm pro-teolysis, vis-á-vis the industrial and nutritional uses of cereal malts,appears to be the provision of free a-amino nitrogen (FAN), amixture of amino acids, ammonium ions and short peptides.
R250 dye; FAN, Free a-aminorcaptoethanol; Mr, Molecularium dodecyl sulphate poly-
ogy Research Centre, 1840swana. Tel.: þ267 5445516;
c.org (L.I. Ezeogu).
All rights reserved.
a, G., et al., Electrophoretic aoi.org/10.1016/j.jcs.2013.05.00
In brewing, FAN are vitally important, both as major sources ofassimilable nitrogen for rapid yeast growth and ethanol productionand for being centrally involved in the complex biochemical pro-cesses that culminate in beer flavour development (Lekkas et al.,2007). In human nutrition, germination-related proteolysis en-hances protein and starch availabilities while the soluble peptidesand amino acids liberated during malting serve as ready sources ofassimilable nitrogen.
Sorghum (Sorghum bicolor L.) is ranked fifth among the world’smajor cereal crops. Across Africa, malted sorghum has grownincreasingly in industrial use, as a replacement for malt barley inlager beer brewing (Dufour et al., 1992) and as a source for readilyassimilable nutrients in high nutrient density food formulations(Correia et al., 2008).
Sorghum endosperm proteins are heterogeneous. Prolaminsconstitute 77e82% of the total proteins (Hamaker et al., 1995),while albumins and globulins are minor components. Sorghumprolamins (kafirins) are mixtures of monomeric and aggregated(oligomeric and polymeric) proteins. Monomeric kafirins include a-(23 kDa and 25 kDa), b- (20 kDa) and g- (28 kDa) kafirins (Shullet al., 1991; Mazhar et al., 1993). a-Kafirins comprise 60e70% oftotal sorghum protein (and ca. 80% of total kafirins), followed by theg- (ca. 15% of total kafirins), and b-monomers (ca. 5% of totalkafirins) (Hamaker et al., 1995; de Mesa-Stonestreet et al., 2010).
nalysis of malting degradability of major sorghum reserve proteins,8

G. Mokhawa et al. / Journal of Cereal Science xxx (2013) 1e92
The in vitro enzyme-digestibility of kafirins has been studied. a-Kafirin is the most digestible, while g-kafirin is comparativelydigestion-resistant (Oria et al., 1995). Digestibility varies alsoamong the various oligomeric and polymeric kafirins (Nunes et al.,2005). Between-cultivar digestibility differences have been relatedto variations in the proportional and physical distribution of kafirinmonomers in both the endosperm and protein granules (Oria et al.,1995).
The physiology of endosperm reserve protein mobilization hasbeen studied extensively for barley, and results show that this canhave strongly defining influences on the type and quality of bothmalt and beer (Briggs et al., 2004). On the other hand, only verylittle is known of the relationship between malting and the contentand distribution of sorghum’s polypeptides and proteins. As forbarley, a comparative study of the effects of malting on sorghum’sreserve proteins could contribute to the eventual characterizationof sorghum cultivars as malting grains or otherwise. Employingsodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE), Correia et al. (2008) and Taylor (1983), investigatedgermination-induced changes occurring in the reserve proteins ofAustralian and South African sorghum varieties. Using similarmethods, we investigated the protein profiles of eleven commercialBotswana cultivars and how germination-induced changes in thecontents and distributions of their key components could influencetheir maltability.
2. Experimental
2.1. Plant materials
Grains from eight indigenous Botswana sorghum cultivars(Phofu, Segaolane, Lekgeberwa, Town, Sefofu, Kanye Standard, Mar-upantshi and Mafia) and three hybrids (Lars Vyt, BSH-1 and SNK)were obtained from the Department of Agricultural Research,Gaborone, and used for this study. Cultivars BSH-1, Lars Vyt, Lek-geberwa, Phofu, Sefofu were corneous, while the remaining wereintermediate (Kebakile et al., 2007). All the grains had germinativeenergies of �93% while none was water-sensitive (Mokhawa et al.,2009). The grains were all tannin-free.
2.2. Sorghum malting
Malting was as per Mokhawa et al. (2009). Samples (200 g) weresteeped in distilled water (400 mL) at 30 �C for 51 h using thefollowing steep schedule: 5 cycles of 6 h wet and 3 h dry, followedby a final immersion of 6 h. For each steep cycle, fresh distilledwater was used. Before and after steeping, grains were sterilized byimmersion (20min) in NaOCl solution [1% (w/v) available chlorine],then rinsed with sterile distilled water. Germinationwas conductedin the dark, at 30 �C, in an atmosphere of near water-saturation.During germination, grains were, every 12 h, turned and sprayedwith distilled water (5 mL). Germinated grains were dried (50 �C,24 h) in a forced draught oven. Shoots and rootlets were removedmanually. Malt kernels were milled to pass through a 0.25 mmsieve in awater-cooled coffee grinder (Janke and Kunkel, IKA-Werk,Germany).
2.3. Protein extraction and sodium dodecyl sulphateepolyacrylamide gel electrophoresis (SDS-PAGE)
2.3.1. Protein extractionTotal protein of sorghum grains and malt were extracted by a
modification of the Bean et al. (2000) method. Samples (200 mg)were extracted at 60 �C with 1 mL of 12.5 mM sodium borate (pH10.0 containing 2% (w/v) SDS and 2% 2-ME). Samples were
Please cite this article in press as: Mokhawa, G., et al., Electrophoretic aJournal of Cereal Science (2013), http://dx.doi.org/10.1016/j.jcs.2013.05.0
extracted for 1 h and vortexed every 10 min during extraction.Extracted samples were centrifuged (12,000 � g, 10 min, at 4 �C).Supernatants from two extractions were pulled 1:1 to produce thetotal protein fraction. Albumins and globulins were extracted fromsamples (200 mg) using 1 mL of 50 mM Tris-HCL at pH 7.8 con-taining 0.1 M KCl and 5 mM EDTA as has been previously described(Bean et al., 2000) twice for 5 min with continual vortexing. Pelletsobtained after centrifugation (12,000 � g, 10 min at 4 �C) werewashed using distilled water (1 mL) for 5 min and centrifuged.These were used for the free and total kafirin extraction. Totalkafirins were extracted using 1 mL of 70% (v/v) isopropanol con-taining 0.5% (w/v) sodium acetate and 2% (v/v) 2-ME. Extractionwas for 1 h at 60 �C in a water bath. During extraction, tubes weremixed vigorously every 10 min. Supernatants obtained from twoextractions, and centrifugation (13,200 � g, 10 min, 4 �C) werepooled 1:1 to give the total kafirin fraction. Extraction of the freekafirin fraction proceeded in a similar manner as for total kafirin,except that 2-ME was omitted in the extractant. Extracts (200 mL)were freeze-dried and stored at �70 �C until used.
2.3.2. SDS-PAGE analysisThe discontinuous TriseHCl/glycine buffer system of Laemmli
(1970) was employed, with a 3.9e20% (w/v) polyacrylamidegradient. Electrophoresis was conducted in a Protean II xi verticalcell equipped with a Powerpac 1000 (Bio-Rad Laboratories, Her-cules, CA, USA) used at a constant current of 50mA and 150 V. Driedresidues were each dissolved in 200 mL of electrophoretic samplebuffer [0.3 M TriseHCl, pH 6.8, containing 4% (w/v) SDS and 20%glycerol]. Before loading, samples were heated in a water bathat boiling temperature (z96 �C) for 30 min. Sample loading waswith 20 mL. Gels were run under non-reducing conditions. Gelswere stained with CBB-R250 (0.1%) in a solution of water:-methanol:glacial acetic acid (5:5:2, vol/vol/vol). De-staining waswith a solution containing 30 and 10% (v/v) methanol and aceticacid, respectively, in distilled water. Gels were scanned and thedigital pictures subjected to densitometric analysis using theGeneTools software version 3.07.03 from Syngene (a division ofSynoptics, Cambridge, England).
2.3.3. Free-amino nitrogen (FAN)The method of Taylor and Boyd (1986) was employed. Samples
(1.0 g) were each extracted with 40 ml of 5% (w/v) TCA at 30 �C for1 h. After centrifugation (10 min at 10,000 � g, 4 �C), the clearsupernatant (1 ml) was diluted appropriately in distilled water andused for FAN determination by the EBC ninhydrin method(European Brewing Convention, 1998). FAN levels were expressedin mg/100 g of sorghum grain or malt.
2.3.4. Statistical analysisFAN data were evaluated by analysis of variance (ANOVA) based
on a 0.001 level of significance. Differences between means weredetermined using the least significant difference (LSD) test.
3. Results and discussion
3.1. Total protein fraction of sorghum grain and malt
Results are presented in Fig. 1(A) for the PAGE analysis of thetotal protein fractions of the eleven cultivars. Several polypeptidesofMr 10e25 kDawere separated from both the raw and germinatedgrains; however these differed in their individual abundance, asassessed by their respective absorbances (Fig. 1(B)). The total pro-tein profiles, after malting, varied markedly between all elevencultivars. Compared to the respective raw grains, marked reductionin the intensity, and sometimes total disappearance, of several
nalysis of malting degradability of major sorghum reserve proteins,08

25015010075
50
37
25201510
250150
10075
50
37
25201510
25015010075
50
37
25201510
25015010075
50
37
25201510
(Raw) (Malt)
1 2 3 4 5 6 7 8 9 10 11 12 13 1 2 3 4 5 6 7 8 9 10 11 12 13
Phofu Segaolane Lekgeberwa
Town Lars Vyt Sefofu
BSH1 Kanye Standard Marupantsi
Mafia SNK
MW standards MW standards MW standards
A
B
250
150
100 75 50 37 25 20 15 10250
150
100 75 50 37 25 20 15 10
250
150
100 75 50 37 25 20 15 10
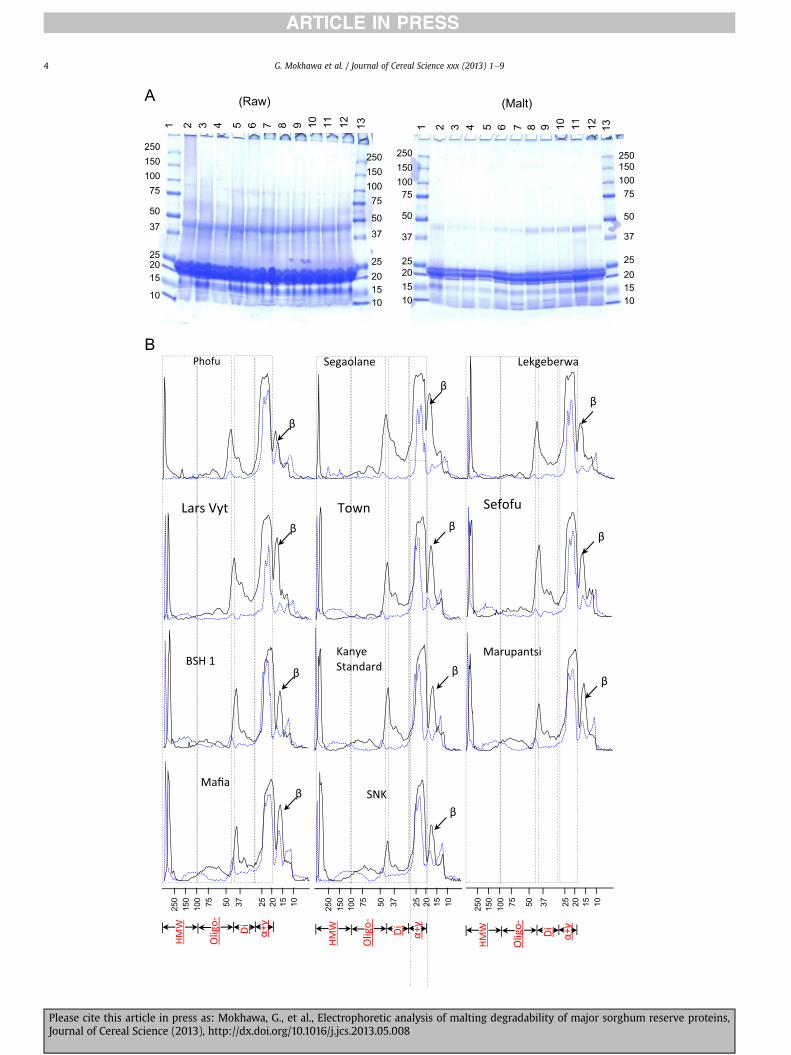
Fig. 1. SDS-PAGE analysis of total proteins from ungerminated grains (Raw) and malts (Malt) of eleven Botswana sorghum cultivars (Fig. 1A). Lanes 1 and 13, protein standards; lane2, Phofu; lane 3, Segaolane; lane 4; Lekgeberwa; lane 5; Lars Vyt; lane 6, Town; lane 7, Sefofu; lane 8, BSH-1; lane 9, Kanye Standard; lane 10, Marupantshi; lane 11, Mafia; lane 12, SNK.Densitometric analysis of the total protein SDS-PAGE gels are presented in Fig. 1B for the ungerminated grains (solid line) and malt (broken blue line). (For interpretation of thereferences to colour in this figure legend, the reader is referred to the web version of this article.)
G. Mokhawa et al. / Journal of Cereal Science xxx (2013) 1e9 3
Please cite this article in press as: Mokhawa, G., et al., Electrophoretic analysis of malting degradability of major sorghum reserve proteins,Journal of Cereal Science (2013), http://dx.doi.org/10.1016/j.jcs.2013.05.008
1 2 3 4 5 6 7 8 9 10 11 12 13
25015010075
5037
252015
10
25015010075
5037
25201510
1 2 3 4 5 6 7 8 9 10 11 12 13
25015010075
50
37
25201510
25015010075
50
37
25201510
(Raw) (Malt)A
B
250
150
100 75 50 37 25 20 15 10250
150
100 75 50 37 25 20 15 10250
150
100 75 50 37 25 20 15 10
G. Mokhawa et al. / Journal of Cereal Science xxx (2013) 1e94
Please cite this article in press as: Mokhawa, G., et al., Electrophoretic analysis of malting degradability of major sorghum reserve proteins,Journal of Cereal Science (2013), http://dx.doi.org/10.1016/j.jcs.2013.05.008

G. Mokhawa et al. / Journal of Cereal Science xxx (2013) 1e9 5
bands, was observed after malting. Most medium-to-high Mrcomponents (Mr z 30e250 kDa) were readily digested duringmalting, sometimes disappearing completely from the total proteinextract. Proteolytic digestion was most pronounced in Phofu (lane2) and BSH 1 (lane 8) (Fig. 1(A)). In both these cultivars, proteins ofMr �30 kDa (Phofu) or Mr �37 kDa (BSH 1) were totally undetect-able in the total protein fractions of the malts, indicating totaldepolymerisation. Medium-to-high Mr proteins were detectable inmalts from the other 9 varieties, although the individual intensitiesof their corresponding bands were considerably diminished, incomparison to the respective raw grains (see Fig. 1(B)). This in-dicates that extensive proteolysis occurred during malting. Theintensities of medium-to-high Mr protein bands changed in acultivar-dependent manner. Mr 70e100 kDa proteins were detect-able only in Mafia malts. In Segaolane, Lekgeberwa, Lars Vyt, Town,Sefofu, Kanye Standard, Marupantsi and SNK, the largest detectableprotein was a polypeptide of Mr z 55 kDa, at absorbances (AU) of2.5, 8.8, 21.3, 5.6, 12.3, 17.3, 26.2 and 19.1, respectively. At 50.9 AU,the intensity of theMr 55 kDa proteinwas highest inMafia, as werethe levels of all the other medium-to-high Mr proteins (especiallythose with Mr z 30e75 kDa) (Fig. 1(B)) when compared to theother cultivars. This suggests that Mafia was probably the leastproteolytic of the eleven sorghum cultivars. Marupantshi, Lars Vyt,and Lekgeberwa displayed relatively high levels of the Mr z 30e55 kDa proteins (Fig. 1(B)), also suggesting low proteolytic poten-tials. Except for Segaolane, all malts exhibited marked increases inthe intensity of the low Mr (�15 kDa) protein bands, compared tothe raw grain.
3.2. Total kafirin fraction of sorghum grain and malt
The total kafirin profiles of sorghum grains and malts are pre-sented in Fig. 2 ((A) and 2 (B)). Polymers, oligomers and monomersof kafirin were detected in all grains and malts. Cultivar-dependentvariations in malt total prolamin profiles were observed. Bandscorresponding to b-, a- and g-kafirins and a kafirin dimer ofMr z 45 kDa, were very intense in the raw grain extracts, butconsiderably fainter in the malts, suggesting extensive digestionduring malting. The largest protein detectable in the total kafirinfraction of the malts consisted of the Mr z 45 kDa kafirin dimer.This contrasted with the raw grains, in which bands of up to Mr100 kDa were clearly visible. The rates at which the intensity of theMr z 45 kDa protein faded appeared also to vary by cultivar. Thisband was faintest in Lars Vyt, Lekgeberwa, Segaolane and Sefofu,where its densitometric peak heights of 5.94, 10.7, 10.5 and 13.1 AUindicated proportional reduction rates of 92.8, 86.8, 87 and 83.7%respectively, in comparison to corresponding raw grain densito-metric peak values of 82.5, 80.6, 80.7 and 80.4 AU. The Mr 45 kDaprotein was also extensively digested in the other varieties,although at much lower and more varied rates (50e79%). However,the 45 kDa kafirin band faded the least inMafia, with a proportionalpeak height reduction of only z32%, after malting. The 45 kDaprotein was probably non-disulphide cross-linked, which isconsistent with the findings of Correia et al. (2008).
Rates of disappearance for a-, b-, and g-kafirins were alsocultivar-dependent. For a- and g-kafirins, the combined rates offading were highest in Lars Vyt (64.8%), Lekgeberwa (58.5%), andSegaolane and Sefofu (55.7 and 53.7%, respectively), based on peakarea (Fig. 2B). b-Kafirin faded most (>77%) in Segaolane,
Fig. 2. SDS-PAGE analysis of total kafirin proteins from ungerminated grains (Raw) and mstandards; lane 2, Phofu; lane 3, Segaolane; lane 4; Lekgeberwa; lane 5; Lars Vyt; lane 6, TowMafia; lane 12, SNK. Densitometric traces of the total kafirin SDS-PAGE gels are presentedinterpretation of the references to colour in this figure legend, the reader is referred to the
Please cite this article in press as: Mokhawa, G., et al., Electrophoretic aJournal of Cereal Science (2013), http://dx.doi.org/10.1016/j.jcs.2013.05.00
Lekgeberwa, and Lars Vyt, but only 21 and 8% in Phofu and Mafia,respectively. For all the remaining cultivars, b-kafirin levels reducedonly 40e58%. Overall, the highest mean reduction in the combinedpeak of all the kafirins occurred in Segaolane, Lekgeberwa and LarsVyt. These varieties may thus have had greater capacity to hydro-lyse kafirin compared to all the other cultivars. Conversely, Mafiagave the lowest reduction in aggregate kafirin content, suggestingthat its kafirins were probably the least degraded during malting.
3.3. Free kafirin of sorghum grain and malt
The free kafirin electrophoretograms displayed bands charac-teristic of high molecular weight (Mr >66 kDa) polymers, a 66 kDatrimer, a 45 kDa dimer and g, a, and b monomers (Fig. 3(A)). Theintensities of all the kafirin bands reduced considerably aftermalting, although in a cultivar-defined manner. Certain kafirinbands also disappeared more readily than the others. With a meanaverage reduction in peak height of only z45%, the 45 kDa olig-omer appeared the most recalcitrant of all the free kafirin bands,while b-, g- and a-kafirins fell in the aggregate by 75, 67 and 57%,respectively. In most of the varieties, except Phofu, the free mono-mer which disappeared the least was a-Kafirin. Malt residual a-kafirin constituted �90% of the level in unmalted Lekgeberwa. Bycontrast, only z18% of the unmalted grain’s free a-kafirin wasdetectable in Phofu malt, while in Lars Vyt, Town and Sefofu, maltfree a-kafirin levels were 75e80% lower than their correspondingraw grains’ contents. Malt residual free a-kafirin content repre-sented 30e39% of the raw grain’s original value in SNK, Mar-upantshi, Kanye Standard, Town and BSH-1, while a-kafirin levelsamounting toz53 and 49% of the original raw grains’ content weredetectable in Segaolane and Mafia malts, respectively. g- and b-kafirins bands also faded the least in Lekgeberwa (43 and 46%respectively), compared to the other varieties. Of all the monomers,the band due to b-Kafirin was the most reduced in Phofu andSegaolane (90e94%), Sefofu and SNK (�70%), Mafia (63%) and Lek-geberwa (46%). Conversely, g-Kafirin faded the most in Lars Vyt,Town, BSH1, Kanye Standard, and Marupantsi. Overall, the kafirinpolymers and the larger sized oligomers faded very readily(Fig. 3(B)), suggesting high susceptibility to malt proteases.
An increase in the levels of monomeric kafirins (especially g anda kafirins), as a proportion of the total amount of detectable freekafirin, was observed in all the malts, except Lars Vyt and Mafia, inwhich these values either fell or showed very minimal change aftermalting (Fig. 3(B)). Levels of the 45 kDa kafirin, as a proportion ofthe total kafirin content, also increased during malting, for all thesorghums, while the proportion of HMW kafirins and the 66 kDaprotein decreased substantially. The HMW kafirin proportion of thefree kafirin fraction was only 11e20.5% in the malts, compared to26.1e36.3% in unmalted sorghum, indicating a decrease of between25 and nearly 70%. The highest reduction in HMW kafirin, as aproportion of total residual free kafirin, was observed in Phofu andSegaolane (z63e70%). Conversely HMW kafirin fell the least inMafia (z25%). Overall, a-kafirin appeared the least digested of allthe kafirin monomers, agreeing with previous reports (Taylor andEvans, 1989; Mazhar and Chandrashekar, 1993; Georget et al.,2012).
b-Kafirin, as a proportion of total free kafirin fell during maltingfor all the sorghum cultivars, except Phofu, Segaolane, and Lekge-berwa. This appears to support the assertion (Oria et al., 1995;
alts (Malt) of eleven Botswana sorghum cultivars (Fig. 2A). Lanes 1 and 13, proteinn; lane 7, Sefofu; lane 8, BSH-1; lane 9, Kanye Standard; lane 10, Marupantshi; lane 11,in Fig. 2B for the ungerminated grains (solid line) and malt (broken blue line). (Forweb version of this article.)
nalysis of malting degradability of major sorghum reserve proteins,8

1 2 3 4 5 6 7 8 9 10 11 12 13
25015010075
5037
252015
10
25015010075
50
37
25201510
1 2 3 4 5 6 7 8 9 10 11 12 13
250
150
10075
503725201510
250
150
10075
503725201510
(Raw) (Malt)
α
αα
γ
γ
β
β
β β
β β
A
B
250
150
100 75 50 37 25 20 15 10250
150
100 75 50 37 25 20 15 10250
150
100 75 50 37 25 20 15 10
Fig. 3. SDS-PAGE analysis of free kafirin from ungerminated grains (Raw) and malts (Malt) of eleven Botswana sorghum cultivars (Fig. 3A). Lanes 1 and 13, protein standards; lane 2,Phofu; lane 3, Segaolane; lane 4; Lekgeberwa; lane 5; Lars Vyt; lane 6, Town; lane 7, Sefofu; lane 8, BSH-1; lane 9, Kanye Standard; lane 10, Marupantshi; lane 11, Mafia; lane 12, SNK.Densitometric traces of the free kafirin SDS-PAGE gels are presented in Fig. 3B for the ungerminated grains (solid line) and malt (broken blue line). (For interpretation of thereferences to colour in this figure legend, the reader is referred to the web version of this article.)
Please cite this article in press as: Mokhawa, G., et al., Electrophoretic analysis of malting degradability of major sorghum reserve proteins,Journal of Cereal Science (2013), http://dx.doi.org/10.1016/j.jcs.2013.05.008

G. Mokhawa et al. / Journal of Cereal Science xxx (2013) 1e9 7
El Nour et al., 1998; Nunes et al., 2005) that far smaller amounts ofb-kafirin (compared to both a- and g-kafirins) are involved in theformation of polymeric kafirin aggregates, while none seems to befound in kafirin oligomers. In contrast, the trend observed in Phofu,Segaolane and Lekgeberwa raises the possibility that these 3 culti-vars probably differed from the other eight both in context of theproportions of a-, b-, and g-kafirins making up their oligomeric andpolymeric kafirin aggregates and in the context of their proteolyticenzyme profiles. The monomer composition of oligomeric andpolymeric kafirins has been well known to vary by grain cultivar(Nunes et al., 2005).
While studying the digestion of maize prolamins, Mitsuhashiand Oaks (1994) had observed that individual zein bands dis-appeared at different rates during germination; g-Zein was first todisappear, followed by b- and k-zeins, while a-zeins persisted longafter the other zeins had been degraded. Garcia-Villalba et al.(2006) showed that barley hordeins disappeared at different ratesduring malting, with C-hordein disappearing significantly faster,while Holopainen et al. (2005), working on 27 barley cultivars,demonstrated the existence of cultivar-related preferences in hor-dein degradation, with D-hordein and B-hordein being the mostdegraded hordeins in Barke and Scarlett, respectively. Preferentialdigestion has similarly been demonstrated for u-gliadin earlyduring wheat germination (Bigiarini et al., 1995). It is apparent inthe present work that the digestion of individual sorghum pro-lamins occurred at different rates during malting, with b- and g-kafirin disappearing significantly more (on the aggregate) than a-kafirin. The slower disappearance of a-kafirin, compared to g- andb-kafirins, during malting has been reported previously by Taylorand Evans (1989), Mazhar and Chandrashekar (1993) and Georgetet al. (2012), who also ascribed the faster disappearance of b-and g-kafirins to their more peripheral physical location insorghum’s protein granule and, therefore, to their greater physicalaccessibility to digesting proteases. We acknowledge that thephysical arrangement of kafirin monomers could have played a rolein regulating the rates at which these disappeared during malting.Experience of protein reserve mobilisation in other cereals (Poulleand Jones, 1988; Mitsuhashi and Oaks, 1994; Mikola et al., 2001)however, suggests the possibility that other additional factors couldhave played equally, if not more, critical roles in the rates at whichindividual monomer bands faded during germination in sorghum.Of these, substrate preference in the activities of key proteasesappears to be quite critical.
Specific proteolytic activities are required for hydrolysis ofspecific prolamin molecules during germination. Major oat pro-teinases appearing at different times during germination hydro-lyse different avenin types (Mikola et al., 2001); a-avenin ishydrolysed by proteases appearing after 1 day of germination,while b2- and b1-avenins are digested by enzymes appearing after5 and 8 days, respectively. In barley, B, C, and D hordeins differ intheir individual susceptibilities to a key protease isolated from thegerminated grain (Poulle and Jones, 1988), while maize cysteineendoproteases appearing early after imbibition hydrolyse g- butnot a-zeins, which are preferably hydrolysed by later-appearingproteinases (Mitsuhashi and Oaks, 1994). Protease preference forspecific kafirin monomers can substantially explain the observeddifferences in the relative rates of digestion/disappearance of a-, b-and g-kafirin monomers. In this sense, across-cultivar differencesin the relative rates of disappearance of individual kafirin mono-mers may reflect cultivar-dependent variations in the compositionof their proteolytic enzyme cocktails. Mokhawa et al. (2009)showed that protease profiles differed significantly among thesorghum grains employed in the current study. Since substratepreference and specific activity vary by protease, it is conceivablethat the extents to which individual kafirin monomers are
Please cite this article in press as: Mokhawa, G., et al., Electrophoretic aJournal of Cereal Science (2013), http://dx.doi.org/10.1016/j.jcs.2013.05.00
digested during malting be related to each monomer’s intrinsicsusceptibility vis-à-vis each grain’s malt proteases and theaggregate of their activities.
Nunes et al. (2005) have shown that the composition of indi-vidual prolamin oligomer and polymer aggregates, in sorghum,differed among the varieties, whereby the 66 kDa prolamin con-tained the g and a monomers in KLW and KAT 369 varieties, butconsisted only of g-kafirin in NK 283 and PAN 8569. The monomercomposition of individual kafirin polymers and oligomers was notstudied in this work for our eleven sorghum cultivars. It is never-theless conceivable that dissimilarities in the composition of thediverse HMW and oligomeric prolamin aggregates of the variouscultivars contributed to observed varietal differences in the rates atwhich the various kafirin bands were digested.
The free kafirin content and profile of malt represents the bal-ance between the amount of kafirin freed through protein depo-lymerisation and that lost through digestion to amino acids, shortpeptides and ammonium. In barley, at least five endoproteases andfive carboxypeptidases, along with four neutral proteases, havebeen associated with seed reserve proteolysis (Mikola, 1983; Jones,2005). At least 15 endoproteinases have been identified in maltedsorghum (Mokhawa et al., 2009), while sorghum malt endo- andexo-protease potentials vary markedly across variety (Ezeogu andOkolo, 1995). Inherent differences in sorghum cultivars’ abilitiesto depolymerise kafirin aggregates to monomers or to hydrolysethe latter to FAN could result in varying rates of disappearance forkafirin monomers. Polymeric and oligomeric aggregates of kafirinare made upmostly of a- and g-kafirin monomers, with a far higheramount of a-monomer being involved (Oria et al., 1995; Nuneset al., 2005; de Mesa-Stonestreet et al., 2010). It is thus highlyprobable that kafirin depolymerisation would release largeramounts of a- and g-kafirin monomers, compared to b-kafirin. Ifthe rate at which the a- and g-kafirin molecules are degraded is notas high as that of their release, these would accumulate. Thisprobably explains why the proportions of a- and g-kafirin rose formost of the sorghum malts.
Correia et al. (2008) had suggested that the sorghum Mr 66 kDakafirin oligomer might be recalcitrant to proteolytic digestion. Thatwas not the case in the present study. The Mr 66 kDa kafirin bandsfaded quite readily after malting, suggesting otherwise. These dif-ferences between our results and those of Correia et al. (2008)indicate the possibility that the eleven sorghum cultivars used inthe present study were physiologically different from the Austra-lian variety of sorghum used by those authors.
Throughout this study, variations occurred in the rates atwhich individual sorghum cultivars digested their kafirins, sup-porting findings that sorghum cultivars differed markedly intheir individual proteolytic capacities and proteinase profiles. Ina zymographic study of the eleven sorghum cultivars employedin the current investigation, Mafia had been identified as havingboth the fewest proteinase bands and the lowest in-gel activity(Mokhawa et al., 2009). Consistent with that finding, Mafiaappeared the least proteolytic, its proteins fading the least,throughout the current study. Wu and Wall (1980) indicated thatthe total prolamin content of sorghum decreased during germi-nation. The overriding attenuation in the peak heights and in-tensities of individual kafirin bands observed in this work isconsistent with that finding and indicates this to be also true ofthe eleven Botswana sorghum cultivars. In addition, the currentreport shows that decreases in the prolamin levels affected allthe prolamin fractions, including the various high molecularweight kafirins, each of the diverse oligomers, as well as the a-,b- and g-monomers. In the aggregate, the most extensive fadingof all kafirin bands, in any of the eleven sorghum cultivars,during malting occurred in Phofu.
nalysis of malting degradability of major sorghum reserve proteins,8

G. Mokhawa et al. / Journal of Cereal Science xxx (2013) 1e98
3.4. Free amino nitrogen (FAN)
Availability of adequate levels of free amino nitrogen is highlycritical to the sustenance of rapid yeast growth and fermentation,the rapid and effective utilization of sugars and the optimumdevelopment of flavor during beer brewing (Lekkas et al., 2007).The effect of variety on the FAN potentials of the eleven sorghumvarieties was studied (Fig. 4). Evidently, grain variety significantly(p< 0.001) influenced the amount of FAN formed duringmalting inthe eleven Botswana sorghum cultivars. Mean malt FAN contentwas highest with Segaolane (225.6 mg/100 g) followed by Sefofu,Town and BSH1 in that order. On the other hand, FAN developmentwas by far the lowest in malt from Mafia (at 96.9 mg/100 g), rep-resenting between only 41 and 65% of the FAN content of malt fromthe other sorghum cultivars. In previous reports (Ezeogu and Okolo,1995), grain cultivar had been shown to be a significant determi-nant of the patterns of low molecular weight nitrogen compounddevelopment (including FAN) during sorghum malting. This isapparently also true of the eleven Botswana sorghum cultivars. Atbetween>190 and 225 mg/100 g, the levels of FAN observed in thisstudy for some of the sorghumvarieties (viz. Segaolane, Sefofu, Townand BSH 1) were quite high and fall well within ranges reported formalt made from Nigerian (Ezeogu and Okolo, 1995), Ethiopian(Bekele et al., 2012) and other sorghums (Chiba et al., 2012). On theother hand, the significantly (p < 0.001) wide range of differencesin FAN potentials observed in this study for malts from the elevenBotswana sorghum cultivars reinforces the assertion (Dufour et al.,1992) that considerably high malt FAN could be obtained fromsorghum malts if appropriate cultivars were selected. Our resultssuggest that the four cultivars Segaolane, Sefofu, Town and BSH 1could be good candidate varieties for making high FAN sorghummalt.
Malt FAN levels, at any time, depend on several factors, princi-pally, the rates of proteolysis and the rates of consumption, inanabolic processes, of the freed amino acids and small peptides bythe growing seedling. The fact that Mafia produced the lowest FANlevel, of all the cultivars studied, is consistent with observations oflower reserve protein disappearance during electrophoresis and
Fig. 4. Free amino (FAN) content (mg/100 g) of malts from 11 Botswana sorghums.
Please cite this article in press as: Mokhawa, G., et al., Electrophoretic aJournal of Cereal Science (2013), http://dx.doi.org/10.1016/j.jcs.2013.05.0
also reinforces findings by Mokhawa et al. (2009) ascribing a verylow proteolytic potential to this grain.
4. Conclusions
The predominant pattern of seed proteolysis in cereals is basedon an initial limited proteolytic cleavage by specific proteases fol-lowed by a second, more rapid phase of proteolysis catalysed by lessspecific enzymes (Shutov and Vaintraub, 1987). Results indicatethat varietal factors play significant roles in the manner in whichmajor sorghum proteins are digested during malting. With an R2
value of 0.0193, there was no correlation between visual hardnessand extent of reduction in sorghum grain HMW aggregate levels,showing that factors other than grain texture played more impor-tant roles in sorghum grain protein digestion. Cultivar-based dif-ferences in individual kafirin digestion patterns could be related topossible differences in proteolytic enzyme content and activities.Differences in preferential digestion of kafirin monomers amongsorghum cultivars is probably related to the mean substrate spec-ificity of their individual proteolytic enzyme cocktails.
References
Bean, S.R., Lookhart, G.L., Bietz, J.A., 2000. Acetonitrile as a buffer additive for freezone capillary electrophoresis separation and characterization of maize (Zeamays L.) and sorghum (Sorghum bicolor Moench.) storage proteins. Journal ofAgricultural and Food Chemistry 48, 318e327.
Bekele, A., Bultosa, G., Belete, K., 2012. The effect of germination time on maltquality of six sorghum (Sorghum bicolor) varieties grown at Melkassa, Ethiopia.Journal of the Institute of Brewing 118, 76e81.
Bigiarini, L., Pieri, N., Grilli, I., Galleshi, L., Capochi, A., Fontanini, D., 1995. Hydrolysisof gliadin during germination of wheat seeds. Journal of Plant Physiology 147,161e167.
Briggs, D.E., Boulton, C.A., Brookes, P.A., Stevens, R., 2004. Brewing. Science andPractice. CRC Press, New York.
Chiba, Y., Bryce, J.H., Goodfellow, V., MacKinlay, J., Agu, R.C., Brosnan, J.M.,Bringhurst, T.A., Harrison, B., 2012. Effect of germination temperatures onproteolysis of the gluten-free grains sorghum and millet during malting andmashing. Journal of Agricultural and Food Chemistry 60, 3745e3753.
Correia, I., Nunes, A., Barros, A.S., Delgadillo, I., 2008. Protein profile and malt ac-tivity during sorghum germination. Journal of the Science of Food and Agri-culture 88, 2598e2605.
de Mesa-Stonestreet, N.J., Alavi, S., Bean, S.R., 2010. Sorghum proteins: the con-centration, isolation, modification, and food applications of kafirins. Journal ofFood Science 75, R90eR104.
Dufour, J.P., Melotte, L.A., Srebrink, S., 1992. Sorghum malts for the production of alager beer. Journal of the American Society of Brewing Chemists 50, 110e119.
El Nour, I.N.A., Peruffo, A.D.B., Curioni, A., 1998. Characterisation of sorghumkafirins in relation to their cross-linking behaviour. Journal of Cereal Science28, 197e207.
European Brewing Convention, 1998. Free Amino Nitrogen of Malt by Spectro-photometry, fifth ed.. In: Method 4.10. Analytica EBC Fachverlag Hans Carl,Nuremburg.
Ezeogu, L.I., Okolo, B.N., 1995. Effects of air rest periods on malting sorghumresponse to final warm water steep. Journal of the Institute of Brewing 101,39e45.
Garcia-Villalba, R., Cortacero-Ramirez, S., Segura-Carretero, A., Ndez-Gutiearrez, A.F., 2006. Free-Zone capillary electrophoresis analysis of hordeinpatterns at different stages of barley malting. Journal of Agricultural and FoodChemistry 54, 6713e6718.
Georget, D.M.R., Elkhalifa, A.E.O., Belton, S.P., 2012. Structural changes in kafirinextracted from a white type II tannin sorghum during germination. Journal ofCereal Science 55, 106e111.
Hamaker, B.R., Mohamed, A.A., Habben, J.E., Huang, C.P., Larkins, B.A., 1995.Efficient procedure for extracting maize and sorghum kernel proteins revealshigher prolamin content than the conventional method. Cereal Chemistry 72,583e588.
Holopainen, U.R.M., Wilhelmson, A., Salmenkallio-Marttila, M., Peltonen-Sainio, P.,Rajala, A., Reinikainen, P., Kotaviita, E., Simolin, H., Home, S., 2005. Endospermstructure affects the malting quality of barley (Hordeum vulgare L.). Journal ofAgricultural and Food Chemistry 53, 7279e7287.
Jones, B.L., 2005. Endoproteases of barley and malt. Journal of Cereal Science 42,139e156.
Kebakile, M.M., Rooney, L.W., Taylor, J.R.N., 2007. Effects of hand pounding abrasive,decortication-hammer milling, roller milling, and sorghum type on sorghummeal extraction and quality. Cereal Foods World 52, 129e137.
Laemmli, U.K., 1970. Cleavage of structural protein during assembly of the head ofbacteriophage T4. Nature 227, 680e685.
nalysis of malting degradability of major sorghum reserve proteins,08

G. Mokhawa et al. / Journal of Cereal Science xxx (2013) 1e9 9
Lekkas, C., Stewart, G.G., Hill, A.E., Taidi, B., Hodgson, J., 2007. Elucidation of the roleof nitrogenous wort components in yeast fermentation. Journal of the Instituteof Brewing 113, 3e8.
Mazhar, H., Chandrashekar, A., 1993. Differences in kafirin composition duringendosperm development and germination in sorghum cultivars of varyinghardness. Cereal Chemistry 70, 67.
Mazhar, H., Chandrashekar, A., Shetty, H.S., 1993. Isolation and immunochemicalcharacterization of alcohol-extractable proteins (kafirins) of Sorghum bicolor (L.)Moench. Journal of Cereal Science 17, 83e93.
Mikola, L., 1983. Germinating barley grains contain five acid carboxypeptidaseswith complementary substrate specificities. Biochimica et Biophysica Acta747, 241e252.
Mikola, M., Brinck, O., Jones, B.L., 2001. Characterization of endoproteinases thathydrolyze oat avenins. Cereal Chemistry 78, 55e58.
Mitsuhashi, W., Oaks, A., 1994. Development of endoproteolytic activity in maizeendosperm. Plant Physiology 104, 401e407.
Mokhawa, G., Ralefala, M.P., Bulawayo, B.B., Ezeogu, L.I., 2009. Qualitative zymo-graphic studies of major proteinases from malted sorghum (Sorghum bicolor L.)and effects of cultivar. Cereal Chemistry 86, 575e581.
Nunes, A., Correia, I., Barros, A., Delgadillo, I., 2005. Characterization of kafirin andzein oligomers by preparative sodium dodecyl sulfateepolyacrylamide gelelectrophoresis. Journal of Agricultural and Food Chemistry 53, 639e643.
Please cite this article in press as: Mokhawa, G., et al., Electrophoretic aJournal of Cereal Science (2013), http://dx.doi.org/10.1016/j.jcs.2013.05.00
Oria, M.P., Hamaker, B.R., Axtell, J.D., Shull, J.M., 1995. Resistance of sorghum a-, b-,and g-kafirins to pepsin digestion. Journal Agricultural Food Chemistry 1995(43), 2148e2160.
Palmer, G.H., 1989. Cereals in malting and brewing. In: Palmer, G.H. (Ed.), CerealScience and Technology. Aberdeen University Press, Scotland, pp. 61e242.
Poulle, M., Jones, B., 1988. A proteinase from germinating barley. Plant Physiology88, 1454e1460.
Shull, J.M., Watterson, J.J., Kirleis, A.W., 1991. Proposed nomenclature for thealcohol-soluble proteins (kafirins) of Sorghum bicolor (L. Moench) based onmolecular weight, solubility, and structure. Journal of Agricultural and FoodChemistry 39, 83e87.
Shutov, A.D., Vaintraub, I.A., 1987. Degradation of storage proteins in germinatinggrains. Phytochemistry 26, 1557e1566.
Taylor, J.R.N., 1983. Effect of malting on the protein and free amino nitrogen compo-sition of sorghum. Journal of the Science of Food and Agriculture 34, 885e892.
Taylor, J.R.N., Boyd, H.K., 1986. Free a-amino nitrogen production in sorghum beermashing Journal of the Science of Food and Agriculture 37, 1109e1117.
Taylor, J.R.N., Evans, D.J., 1989. Action of sorghum proteinase on the protein bodiesof sorghum starchy endosperm. Journal of Experimental Botany 40, 763e768.
Wu, Y.V., Wall, J.S., 1980. Lysine content of protein increased by germination ofnormal and high-lysine sorghums. Journal of Agricultural and Food Chemistry28, 455e458.
nalysis of malting degradability of major sorghum reserve proteins,8




















