
ظ 66 ዜुጆ - 中国农业科学
289
ISSN 0578-1752 CODEN CKNYAR ዐ ࡔ౫ᄽბ SCIENTIA AGRICULTURA SINICA ڼ59 ਝ ڼ28 Wpm/59 Op/28 ዐࡔ౫ᄽბᇾ ዐ ࡔ౫ბ ዷӸ ڼෙ া ࡔ ঃ ఁ ঃ ظ 66 ዜጆ Special Issue for 55th Anniversary 2015.9.2, 2:49 PM
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of ظ 66 ዜुጆ - 中国农业科学

Michael T. Clegg
M. Alley, USA David A. Andow, USA R. Appels, Australia Jill Shore Auburn, USA Bing Yang, USA Bas Bouman, IRRI
Bouzayen M, France John Bower, South Africa Kenneth G. Cassman, USA Chen Xian-ming, USA Chen Z X, USA Michael T. Clegg, USA
Matthew J. W. Cock, Switzerland Nick Costa, Australia Thomas Crenshaw, USA Zhanao Deng, USA C. Robert Dove, USA Lester E. Ehler, USA
Laurie Burkitt
Scientia Agricultura Sinica 1960 (Semimonthly, Started in 1960)
2015 9 1 48 17 Vol.48 No.17 September 1, 2015 Superintended by Ministry of Agriculture, P. R. China Sponsored by Chinese Academy of Agricultural Sciences
Chinese Association of Agricultural Science Societies Published by Editorial Department of Scientia Agricultura Sinica
12 Address No.12 South Street, Zhongguancun, Beijing 100081 Postcode 100081
010-82109808 82106281 Telephone 010-82109808 82106281 010-82106247 Fax 010-82106247 http://www.ChinaAgriSci.com Website http://www.ChinaAgriSci.com
[email protected] E-mail [email protected] Issued by Beijing Newspapers and Periodicals Distribution OfficeSubscription Domestic Post Office Issued Abroad by International Book Trading Corporation,
P.O.Box 399, Beijing, P.R. China 399 Abroad Issued No.BM43
49.50 / 1188.00 / Price US$ 49.50 / Issue US$ 1188.00 / year
ISSN 0578-1752 2-138
CN 11-1328/S BM43 0178
ISSN 0578-1752CODEN CKNYAR
SCIENTIA AGRICULTURA SINICA
Special Issue for 55th Anniversary
中国农业科学2015 2015.9.2, 2:49 PM17

写 在 前 面
创刊于 1960 年的《中国农业科学》,走过了 55 年的风雨历程。55 年来,在各级主管领导、
主管单位的关怀下,在广大读者、作者、审者的热心支持下,在几代编辑们的辛勤努力下,已
由一株名不见经传的幼苗,成长为业内瞩目的有用之材。多年来,我刊秉承传播农业科研成果,
加强国内外农业科学交流,促进我国农业科学研究的宗旨,用科学、求实精神,吸引了我国一
大批高端农业科学家发表其研究成果,其学术影响力已然多年居于国内同类期刊第一,在全国
近 2000 种科技期刊中综合排名第六位。
有鉴于我刊的成长,农业部、国家新闻出版广电总局、国家自然科学基金委员会、中国科
协、中国期刊协会、中国科学技术信息研究所、清华电子期刊(光盘版)杂志社、武汉大学中
国科学评价研究中心、中国农学会等部门,或给予我刊至高的荣誉,或给予宝贵的资金支持。
在此,谨表示我们最衷心的感谢!
办好一份学术期刊,拥有充足的优秀稿件是第一位的。在此,我们要真诚感谢一大批新老
农业科学家对我刊的厚爱!是他们,不仅在百刊林立、多家竞争的态势中选择我刊,而且为我
刊带来了莫大的学术影响力和荣誉!我们极其珍视这种信任与挚爱,竭力维护这种宝贵的情感,
并将以百倍的努力、细心和热情,为这些资深科学家和一批又一批的青年学者提供周到的服务。
作为一份学术刊物,离不开同行评议环节。多年来我国海内外农业科学家为维护我刊学术
形象、净化学术风气、筛选优质稿件、提供宝贵建议等方面付出了辛勤劳动,使我刊成为业内
瞩目、同行托举、综合评分居于最前列或前列位次的期刊。没有同行评议专家,就没有我刊连
续多年多项的学术荣誉和较高的学术影响力。未来我刊要走向世界,深度参与国际农业科学学
术交流,更要依靠一批又一批不断成长的海内外农业专家。这些专家,不仅要做好自身的科学
研究,还要参与国内外同行评议,不仅需要发挥创造力,而且还要有默默无闻的奉献精神和心
力、汗水的付出。在此,我们特向为我刊学术质量把关做出贡献的所有专家们致以崇高的敬意
和深切的感谢!
作为编辑出版的核心、枢纽——编委会和编辑部,几十年来,各位编委和编辑部成员尽职
尽责,尽心尽力,为我刊的发展,发挥了集体智慧和个人创造力,淌下了辛勤的汗水,换回了
学界的信任和上级的褒奖。在未来的发展中,各位编委和编辑部成员,仍要恪尽职守,再接再
厉,贯彻科学发展观,做好整刊和栏目策划,团结好农业科学共同体,抓好优质稿件的策划和
落实,高效做好同行评议,以严谨、严肃、严格的态度,把好编辑出版的每一环节,要为我国
农业科技创新做好服务与引导,为我院、我国农业科学研究“顶天立地”,跻身国际先进行列
做好扎实的信息传播服务。
值此创刊 55 周年之际,我刊编辑出版一期专刊,大略介绍国内外相关学科发展成果与趋
势,展示我国农业科学研究成就之一斑,以飨对我刊寄予热心关注和期待的读者。 编 者 2015 年 8 月 20 日

- I -
中 国 农 业 科 学 ZHONGGUO NONGYE KEXUE 2015年第 48卷第 17期
目 次
作物遗传育种·种质资源·分子遗传学
3303 中国作物种业科学技术发展的评述 盖钧镒 刘 康 赵晋铭
3316 作物基因组学与作物科学革命 贾继增 高丽锋 赵光耀 周文斌
张卫健
3333 基于基因组学的作物种质资源研究:现状与展望 黎 裕 李英慧 杨庆文 张锦鹏
张金梅 邱丽娟 王天宇
3354 作物科学中的环境型鉴定(Envirotyping)及其应用 徐云碧
3372 中国转基因棉花研发应用二十年 郭三堆 王 远 孙国清 金石桥
周 焘 孟志刚 张 锐
3388 黄淮麦区小麦品种的高产潜力与实现途径 茹振钢 冯素伟 李 淦
3394 中麦 175 高产高效广适特性解析与育种方法思考 何中虎 陈新民 王德森 张 艳
肖永贵 李法计 张 勇 李思敏
夏先春 张运宏 庄巧生
耕作栽培·生理生化·农业信息技术
3404 中国水稻高产栽培技术创新与实践 朱德峰 张玉屏 陈惠哲 向 镜
张义凯
3415 数字植物研究进展:植物形态结构三维数字化 赵春江 陆声链 郭新宇 杜建军
温维亮 苗 腾
3429 大数据推动农业现代化应用研究 许世卫 王东杰 李哲敏
植物保护
3439 小麦条锈菌致病性及其变异研究进展 康振生 王晓杰 赵 杰 汤春蕾
黄丽丽
3454 中国农业昆虫基因组学研究概况与展望 张传溪
3463 中国设施蔬菜害虫天敌昆虫应用研究进展 张 帆 李 姝 肖 达 赵 静
王 然 郭晓军 王 甦
土壤肥料·节水灌溉·农业生态环境
3477 植物营养与肥料研究的回顾与展望 白由路
3493 蓝水和绿水视角下划定“中国农业用水红线”探索 李保国 黄 峰

- II -
中 国 农 业 科 学 2015年第48卷第17期
园艺·贮藏·保鲜·加工
3504 基于文献计量的园艺学基础研究发展状况分析 刘 彬 邓秀新
3515 设施园艺发展概况、存在问题与产业发展建议 蒋卫杰 邓 杰 余宏军
3524 主要果树果实品质遗传改良与提升实践 陈学森 郭文武 徐 娟 丛佩华
王力荣 刘崇怀 李秀根 吴树敬
姚玉新 陈晓流
3541 中国粮食加工科技与产业的发展现状与趋势 姚惠源
畜牧·兽医
3547 全球口蹄疫防控技术及病原特性研究概观 刘在新
3565 《中国农业科学》创刊以来被引次数居前 100 的论文

- III -
SCIENTIA AGRICULTURA SINICA 2015 48 (17)
CONTENTS
CROP GENETICS & BREEDING·GERMPLASM RESOURCES·MOLECULAR GENETICS 3303 A Review on Advances in Science and Technology in Chinese Seed
Industry GAI Jun-yi, LIU Kang, ZHAO Jin-ming
3316 Crop Genomics and Crop Science Revolutions JIA Ji-zeng, GAO Li-feng, ZHAO Guang-yao,
ZHOU Wen-bin, ZHANG Wei-jian 3333 Genomics-Based Crop Germplasm Research: Advances and Perspectives LI Yu, LI Ying-hui, YANG Qing-wen, ZHANG
Jin-peng, ZHANG Jin-mei, QIU Li-juan, WANG Tian-yu
3354 Envirotyping and Its Applications in Crop Science XU Yunbi 3372 Twenty Years of Research and Application of Transgenic Cotton in
China GUO San-dui, WANG Yuan, SUN Guo-qing, JIN Shi-qiao, ZHOU Tao, MENG Zhi-gang, ZHANG Rui
3388 High-Yield Potential and Effective Ways of Wheat in Yellow & Huai
River Valley Facultative Winter Wheat Region RU Zhen-gang, FENG Su-wei, LI Gan
3394 Characterization of Wheat Cultivar Zhongmai 175 with High Yielding
Potential, High Water and Fertilizer Use Efficiency, and Broad Adaptability
HE Zhong-hu, CHEN Xin-min, WANG De-sen, ZHANG Yan, XIAO Yong-gui, LI Fa-ji, ZHANG Yong, LI Si-min, XIA Xian-chun, ZHANG Yun-hong, ZHUANG Qiao-sheng
TILLAGE & CULTIVATION·PHYSIOLOGY & BIOCHEMISTRY·AGRICULTURE INFORMATION TECHNOLOGY
3404 Innovation and Practice of High-Yield Rice Cultivation Technology in China
ZHU De-feng, ZHANG Yu-ping, CHEN Hui-zhe, XIANG Jing, ZHANG Yi-kai
3415 Advances in Research of Digital Plant: 3D Digitization of Plant
Morphological Structure ZHAO Chun-jiang, LU Sheng-lian, GUO Xin-yu, DU Jian-jun, WEN Wei-liang, MIAO Teng
3429 Application Research on Big Data Promote Agricultural Modernization XU Shi-wei, WANG Dong-jie, LI Zhe-min
PLANT PROTECTION 3439 Advances in Research of Pathogenicity and Virulence Variation of the
Wheat Stripe Rust Fungus Puccinia striiformis f. sp. tritici KANG Zhen-sheng, WANG Xiao-jie, ZHAO Jie, TANG Chun-lei, HUANG Li-li
3454 Current Research Status and Prospects of Genomes of Insects
Important to Agriculture in China ZHANG Chuan-xi
3463 Progress in Pest Management by Natural Enemies in Greenhouse
Vegetables in China ZHANG Fan, LI Shu, XIAO Da, ZHAO Jing, WANG Ran, GUO Xiao-jun, WANG Su

- IV -
SCIENTIA AGRICULTURA SINICA 2015 48(17)
SOIL & FERTILIZER·WATER-SAVING IRRIGATION·AGROECOLOGY & ENVIRONMENT3477 Review on Research in Plant Nutrition and Fertilizers BAI You-lu 3493 Defining the Baselines for China Agricultural Water Use in Green and
Blue Water Approach LI Bao-guo, HUANG Feng
HORTICULTURE·STORAGE·FRESH-KEEPING·PROCESSING 3504 Basic Research Development Status of Horticulture Based on
Bibliometric Analysis LIU Bin, DENG Xiu-xin
3515 Development Situation, Problems and Suggestions on Industrial
Development of Protected Horticulture JIANG Wei-jie, DENG Jie, YU Hong-jun
3524 Genetic Improvement and Promotion of Fruit Quality of Main Fruit
Trees CHEN Xue-sen, GUO Wen-wu, XU Juan, CONG Pei-hua, WANG Li-rong, LIU Chong-huai, LI Xiu-gen, WU Shu-jing, YAO Yu-xin, CHEN Xiao-liu
3541 The Present Development Status and Tendency of Grain Processing
Technology and Industry in China YAO Hui-yuan
ANIMAL SCIENCE·VETERINARY SCIENCE
3547 Progress and Prospect of the Technologies to Control Foot-and-Mouth Disease and Its Pathogen Characteristics Worldwide
LIU Zai-xin
3565 Top100 Highly-Cited Papers Since the Start of Scientia Agricultura Sinica

中国农业科学 2015,48(17):3303-3315 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.001
收稿日期:2015-04-20;接受日期:2015-06-15 基金项目:国家自然科学基金专项基金软课题研究项目(L1422038)、中国工程院中国工程科技中长期发展战略研究项目(2014-zcq-11)、教育部
“111”项目(B08025)、教育部创新团队项目(PCSRT13073)、江苏省优势学科建设工程专项、江苏省 JCIC-MCP 项目 联系方式:盖钧镒,E-mail:[email protected]
中国作物种业科学技术发展的评述
盖钧镒1,刘 康
2,赵晋铭
1
(1 南京农业大学大豆研究所/国家大豆改良中心/农业部大豆生物学与遗传育种重点实验室(综合)/作物遗传与种质创新国家重点实验室,
南京 210095;2 南京农业大学农学院种业科学系/江苏省现代作物生产协同创新中心,南京 210095)
摘要:《国务院关于加快推进现代农作物种业发展的意见》提出构建以产业为主导、企业为主体、“育繁推一
体化”的现代农作物种业体系,全面提升中国农作物种业发展水平。国内外种业的发展推动了种业科学的形成和
发展。种业科学是围绕“育繁推一体化”种业产业发展而形成的科学技术学科类群,作物遗传育种是其中的一部
分,它和种子生产的理论与技术、种子示范和营销的理论与技术构成了种业科学技术的主体,在相应的遗传、生
理、信息技术、政策法规等学科知识的配合下成为相对集中的学科体系。中国的种业科学技术体系正在形成与完
善之中。文章在回顾作物育种科学技术进展包括传统的作物育种科学技术和现时分子生物育种研究热点的基础上,
归纳出现时重要的 8 个育种理论和技术问题,继而回顾了作物种子生产和示范推广科学技术的进展,并在此基础
上探讨了中国种业科学技术发展的策略和建议。提出要围绕种业发展的需求,建成相对完整的种业科学技术学科
体系;要顶层设计,建设成企业种业科技和公益性种业科技两支相互补充、相互配合的种业科学技术研发力量;
要优先研究和解决种业发展中的重大科学技术问题,其中包括规模化育种技术,资源富集、遗传解析与创新,常
规育种的分子辅助技术,转基因育种与安全技术,品种区域适应性试验制度与品种审定制度的完善,配齐种业基
础性公益性研究并确立种子生产标准化体系,加强作物杂优化研发使杂种化成为中国未来种业的特色等。
关键词:作物种业;育繁推一体化;规模化商业育种;种子生产标准化;分子辅助育种技术
A Review on Advances in Science and Technology in
Chinese Seed Industry GAI Jun-yi1, LIU Kang2, ZHAO Jin-ming1
(1Soybean Research Institute, Nanjing Agricultural University/National Center for Soybean Improvement, Ministry of Agriculture/
Key Laboratory of Biology and Genetic Improvement of Soybean, Ministry of Agriculture/National Key Laboratory for Crop
Genetics and Germplasm Enhancement, Nanjing 210095; 2Department of Seed Industry Sciences, College of Agriculture,
Nanjing Agricultural University/Jiangsu Collaborative Innovation Center for Modern Crop Production, Nanjing 210095)
Abstract: In the suggestions on speeding up the Development of the Modern Crop Seed Industry, the State Council called for the establishment of a modern crop seed industry with an integrated cultivar development, seed production and seed extension system for the enhancement of Chinese seed industry. The growth of international and domestic seed industry has promoted the formation and development of the seed industry science and technology. The seed industry science and technology is a group of sciences and technologies related to the integrated cultivar development, seed production and seed extension system as well as their neighboring sciences and technologies, such as genetics, plant physiology, information techniques, seed laws and rules, etc. The seed industry science and technology in China is on the way at its beginning and development. The present paper summarized a group of key theoretical and technological issues in plant breeding anxiously to be explored for current seed industry based on a review on the
jia
文本框
返回目录

3304 中 国 农 业 科 学 48 卷
advances in conventional plant breeding and on the hot topics in molecular biological breeding as well as in seed production and extension sciences and technologies. Accordingly, strategies and suggestions were presented for the development of seed science and technology in China. They are the establishment of the whole system of disciplinary sciences and technologies related to seed industry, establishment of the two groups of scientists and technologists, one from seed company side, another from public side, with them co-operated each other, and the seven research priorities to solve the urgent problems in Chinese seed industry, including technology system for large scale breeding programs, genetic dissection and enhancement of germplasm collection, molecular assistance of conventional breeding procedures, enhanced transgenic breeding and safety technology, improvement of regional testing system and cultivar certification, establishment of a standardization system of seed production, hybridization of the seed industry in China.
Key words: crop seed industry; integrating cultivar development, seed increase and seed extension; standardization of seed production; molecular-assisted breeding technology
1 国内外作物种业的发展推动了种
业科学的发展
2050 年全球人口将超过 90 亿,作物生产必须在
2050 年前实现翻番才能满足全球人口的需求。要实现
这个目标,必须以 2.4%的年增长率递增,而目前平均
增长率仅 1.3%[1]。耕地资源的开发和生产水平的提高
是解决问题的 2 个关键途径。耕地资源有限,生产水
平则是开放的。品种改良和栽培水平两者间,品种改
良是主动性因素,栽培水平的实现以品种特性和水平
的上限为依据。因而,全世界都十分重视品种改良和
种业发展,从而驱动种业科学技术的发展。 20 世纪 70 年代,玉米小斑病感染 T 型不育系导
致美国玉米毁灭性损失,促使育种学家对于作物品种
遗传脆弱性的深刻反思,由此推动了对作物种质资源
的研究以及作物种业的发展[2]。国际种业从 20 世纪 70年代起就有了新起步,作物种业从公益性部门扩展、
转移到民营企业。迄今美国的种业 90%以上属于民营
企业。种业作为一个产业,是支撑农作物生产的 基
本产业。以种业为背景的科学技术归之为种业科学技
术。种业公司相继从高等学校吸收育种毕业生,聘请
在任或退休教授,建立从育种到种子生产和种子精选、
加工、检验、销售的技术体系。一批包括 Monsanto、Pioneer、Syngenta、Bayer 等在内的全世界跨国种业公
司成长起来[3]。公司竞相发展种业科技并与公益性研
究机构的科技研究相互呼应、补充或结合,推动了现
代种业科学技术体系的建立。种业的竞争深入到品种、
种子质量、销售推广等各个方面,因而推动了育、繁、
推各环节科学技术,尤其是现代分子生物科技在种业
中应用的快速发展。 中国 21 世纪前后才重视种子产业的发展。这以往
种子是公益性事业。品种选育和种子的繁殖、推广都
是政府的行政事业,种子部门的工作重点是组织新品
种选育、试验、示范以及种子的生产和推广。研究部
门主要负责新品种培育,各级原(良)种场负责种子
繁殖和提纯复壮,农民和合作组织对推广品种进行自
繁、自选、自留、自用,辅之以调剂。当时种业的科
学依托是作物育种学和种子学,因为种子未成为产业,
种业科学停留在一个单调的育种理论和技术的体系。
随着种业的发展,种业科学的内涵不断扩展,涉及到
品种选育、品种潜力挖掘、种子生产、种子经营、技
术推广和法律法规等各个方面。 中国是作物生产大国和用种大国。农作物种业已
明确为国家战略性、基础性核心产业,是促进农业长
期稳定发展、保障国家粮食安全和农产品有效供给的
根本。国家十分重视农作物种业的发展,国务院连续
3 年发布了 3 个国家发展农作物种业的文件,即:《国
务院关于加快推进现代农作物种业发展的意见(国发
〔2011〕8 号)》、《全国现代农作物种业发展规划
(2012—2020 年)(国办发〔2012〕59 号)》和《深
化种业体制改革提高创新能力的意见(国办发〔2013〕109 号)》。文件明确国家发展种业的指导思想是“以
科学发展观为指导,推进体制改革和机制创新,完善
法律法规,整合农作物种业资源,加大政策扶持,增
加农作物种业投入,强化市场监管,快速提升中国农
作物种业科技创新能力、企业竞争能力、供种保障能
力和市场监管能力,构建以产业为主导、企业为主体、
基地为依托、产学研相结合、‘育繁推一体化’的现
代农作物种业体系,全面提升中国农作物种业发展水
平”[4]。提出中国发展农作物种业的目标为“到 2020年,形成科研分工合理、产学研相结合、资源集中、
运行高效的育种新机制,培育一批具有重大应用前景
和自主知识产权的突破性优良品种,建设一批标准化、

17 期 盖钧镒等:中国作物种业科学技术发展的评述 3305
规模化、集约化、机械化的优势种子生产基地,打造
一批育种能力强、生产加工技术先进、市场营销网络
健全、技术服务到位的‘育繁推一体化’现代农作物
种业集团,健全职责明确、手段先进、监管有力的种
子管理体系,显著提高优良品种自主研发能力和覆盖
率,确保粮食等主要农产品有效供给”[5]。而且提出
要“加快种子生产基地建设”,“加强种子市场监管”。
“建立种子市场秩序行业评价机制,督促企业建立种
子可追溯信息系统,完善全程可追溯管理……规范种
子营销网络”。“国家级和省部级科研院所和高等院
校要重点开展种质资源搜集、保护、鉴定、育种材料
的改良和创制,重点开展育种理论方法和技术、分子
生物技术、品种检测技术、种子生产加工和检验技术
等基础性、前沿性和应用技术性研究以及常规作物育
种和无性繁殖材料选育等公益性研究。推进实施转基
因生物新品种培育重大专项。完善公共研究成果共享
机制,为种子企业提供科技支撑”[6]。 根据国家对于农作物种业发展的指导方针和种业
科学技术内涵的发展,中国有待在以往作物遗传育种
学科的基础上建立起种业科学技术的完整体系。下文
在评述国内外作物育种、种子生产和种子推广科学技
术进展的基础上,着重就中国种业科学技术的发展提
出一些讨论意见。
2 作物育种科学技术的进展
2.1 传统的作物育种科学技术
植物育种的历史可以追溯到农作物物种的驯化,
作物驯化和选择育种的历史超过了 10 000 年[7]。人类
从众多的个体中,把可能由于突变或者重组而表现非
凡的个体识别并选留下来,然后有意识或者无意识地
繁殖他们 喜爱的类型。 后成为农家地方品种。但
驯化之后,作物的改良非常缓慢,其原因可能是因为
不知道如何创造人类需要的变异。17 世纪和 18 世纪
处于启蒙运动的欧洲,人口增长和城市化刺激提高作
物产量的渴望,人们开始有意识地选择有用植物,但
是植物改良超前于遗传学发展,此时兴起的植物学,
授粉、杂交以及杂种优势现象的阐述对系统育种作出
了贡献。但是直到达尔文和孟德尔学说的问世,才引
发了过去 100 多年作物遗传育种的大发展。 20 世纪之前的育种,基本上是对表现型进行直接
选择,因此,常常又被称为“经验育种”。早期的选
择育种主要是对自然变异进行选择,在 20 世纪上半叶
及以前一直发挥着主导作用,是历史 悠久的育种方
法。在中国,先是从水稻南特品种中选出了南特 16号等品种,进而又选出了矮脚南特,从矮脚南特中又
选出了近 20 个品种[8];从引进的小麦品种阿夫中选出
了扬麦 1 号、白泉 673 等十几个品种[9];从引进的陆
地棉品种岱字 15 中选出了近 40 多个品种[10]。 19 世纪末到 20 世纪初,英国一些种子公司就从
全球收集植物,进行复合杂交,并从中选择“突破类
型”,孟德尔时代之前的这种杂交育种,已经取得很
大成功,甚至这些成功本身阻碍了孟德尔规律的采用。
随着孟德尔遗传规律的广泛传播,育种家可以根据孟
德尔规律有目的地选配不同亲本进行杂交、自交、回
交,再通过后代选择,获得符合人类需求的重组类型
或超亲分离类型。在中国,通过杂交育种途径育成的
品种占所有育成品种的比例从 20 世纪 50 年代的
14.8%、60—70 年代 35.5%和 80 年代 79%,上升到目
前约 100%。 回交育种则是被发明用于对特定的单个目标性状
进行遗传转移的技术。中国用鸡脚棉与推广品种德字
棉 531 杂交,经多次回交后,于 1944 年育成抗卷叶螟
的鸡脚德字棉。美国用抗根腐病的大豆材料与高产品
种回交后,得到抗病品种哈罗索 63 等,在严重发病条
件下,产量为原品种的 3 倍。加拿大利用回交法,得
到了低芥酸和低硫代葡萄糖苷油菜品种托尔。玉米抗
大斑病 Ht 和抗小斑病 rhm 的转育,也应用了回交育
种方法。另外,采取修饰回交技术有利于打破性状的
不良连锁,并实现对多个性状的同步改良[11]。 玉米杂种优势现象的发现和双杂交种的商业化应
用,是 20 世纪初育种工作的突破性成就。直到 20 世
纪 60 年代,得益于培育优良自交系的努力,玉米单交
种逐步取代双交种,产量得到进一步提高的同时,还
节约了大量的用于杂交的时间和工本。1933 年首次发
现的玉米细胞质雄性不育系,则为玉米杂种优势的利
用开辟了新的思路和途径。玉米杂种优势的商业化利
用,推动了其他作物杂种优势利用的研究,先后在高
粱、水稻、油菜、棉花等大田作物上成功应用并大面
积推广。其中,中国科学家在杂交水稻研究和应用方
面作出了突出贡献[12]。 高选择压力、轮回使用优质受体种质以及采取回
交等减少遗传重组的育种策略使品种得到改良的同时
也降低了品种遗传基础的多样性。例如,美国大多数
硬红冬小麦品种主要来源于 2 个东欧品系;栽培水稻
的基因库遗传变异较野生近缘种狭窄;即使在具有高
度多样性的玉米中,也有平均大约 30%位点的多样性

3306 中 国 农 业 科 学 48 卷
比玉米野生近缘种低。此外,往往存在现有种质资源
缺乏目标性状的基因,或者同一作物品种之间存在不
可弥补的缺点。远缘杂交可以在一定范围内有效打破
物种之间的界限,促进不同物种基因的渐渗和交流,
引入异种甚至异属物种的有利基因或等位变异,培育
作物新品种。通过远缘杂交创造的包含不同物种染色
体组的作物新类型,异附加系、置换系、易位系等各
种异染色体系,单倍体或者细胞质雄性不育系等各种
新种质,大大丰富了作物遗传改良的基因资源。小麦
中,有近 30 种不同病害抗性基因从野生近缘种渐渗到
栽培种中,例如利用黑麦 1R 染色体短臂与小麦染色
体易位系培育的品种占17个国家505个面包小麦品种
的 45%。从二倍体栽培一粒小麦渗入到小麦中的 Pin的功能等位基因使小麦表现为软质籽粒。大麦的改良
也得益于野生大麦和中东地方品种大麦的基因。还有
从球茎大麦转移到大麦的白粉病抗性基因和大麦云纹
病抗性基因。中国科学院李振声等将普通小麦与长穗
偃麦草杂交,育成了多种小偃麦杂种新类型(异源八
倍体、异附加系、异代换系和易位系)和小偃 4 号、
小偃 5 号、小偃 6 号等一系列优良小麦新品种,其中
以小偃 6 号表现 为突出,1985 年推广面积超过 6.7×105 hm2,开创了小麦远缘杂交育种在生产上大面积
推广的先例[13]。中国农业科学院鲍文奎等人工合成了
八倍体小黑麦。 诱变育种通过人工采用物理、化学或者太空等因
素的诱发,不仅可以创造全新的变异,还能促进远缘
杂交过程中染色体的变异。另外通过轮回选择等群体
遗传改良手段,可以提高群体优良基因和基因型频率,
增加优良基因的重组,使有利等位基因聚集到同一个
体之中,从而创造新的种质资源,或者改良外来种质
的适应性等。 传统育种取得了巨大的成功,作物产量的提高主
要归功于特异种质发掘、创新和有效利用。据统计,大
多数国家谷物和豆类作物的年遗传增益达到了 10—50 kg·hm-2。利用矮秆基因(来自低脚乌尖水稻的 Sd1,来
自 Norin 10 小麦的 Rht1 和 Rht2)培育的高产、耐肥、
抗倒伏水稻和小麦品种以及可以抵御高密度种植的杂
交玉米新品种在发展中国家的推广使用,短短几年时
间里实现了粮食产量的大幅度增产,被誉为“绿色革
命”,使许多国家基本摆脱了饥饿[14]。近年来,控制
分蘖、分枝以及种子数量的单基因性状也得以应用。 传统育种对单个基因的选择和应用非常有效,但
是对于遗传率低、基因型-环境互作显著的复杂性状,
例如产量和品质性状、生物胁迫和非生物胁迫抗耐性
等的改良进展有限。而这些性状恰恰是提高作物产量
潜力和产量稳定性的关键性状。进一步增产有赖于将
传统育种与远缘杂交、基因组学、转基因技术相结合,
提高作物产量潜力和生物与非生物抗耐性,适应各种
农业生态位。 2.2 现时分子生物育种的研究热点
虽然在过去的一个世纪里传统育种使用丰富的种
质资源取得了巨大的成功,但是遗传增益日渐萎缩是
当前育种面临的一大挑战。栽培品种、地方品种、野
生近缘种等种质资源中还有没有可资利用的优良基因
或者等位基因?通过外源基因渗入和优良等位基因的
累积还能不能有效提高育种的遗传增益?如果有,如
何才能有效鉴定和利用这些基因资源?这些问题的答
案直接关系作物育种的目标和策略的制定。过去 30年里,人们在分子标记技术、DNA 测序技术、数据计
算技术、生物技术、遥感技术、信息技术等领域取得
了突破性进展。全球转基因作物(GM 作物)种植面
积已经超过了耕地面积的 10%。分子标记辅助选择
(marker assisted selection,MAS)为许多育种程序增
添了潜力。随着新一代测序技术的发展和广泛应用,
已经有超过 50 种作物的参考基因组公布。这些基因组
信息正在改变种质资源的研究以及遗传多样性的应
用,反过来,将改变传统育种策略。基因组途径,例
如基因组重测序、等位基因发掘以及基因组选择将整
合传统技术用于基因型鉴定、种质资源筛选以及品种
选育。 2.2.1 分子标记辅助育种 20 世纪 80 年代兴起的
DNA 标记技术,经历了标记开发、遗传图谱、功能和
比较基因组连锁分析及基因组测序等不同发展时期。
应用分子标记可以对种质资源、杂交亲本和后代个体
进行大规模的基因型鉴定,可以构建遗传图谱并与表
型数据建立联系。基因组学的革命,可以大大提高育
种效率和规模。分子标记可以用来(1)区分品种,构
建杂种优势群体;(2)鉴定种质资源冗余度、未充分
发掘的等位基因以及现有种质资源的遗传构成;(3)监测种质资源储存、繁育、驯化以及育种过程中的遗
传飘变;(4)筛选新基因或超等位基因种质资源;(5)构建作物核心种质库或代表性子集等。
虽然所有的分子标记都可以用于标记辅助选择
(MAS),但是 SSR、SNP 因其简易、量大、共显性、
自动化等优点而被广泛应用。大规模二代测序技术
(next generation sequencing,NGS)使测序成本降低

17 期 盖钧镒等:中国作物种业科学技术发展的评述 3307
了 1 000 倍以上,可以应用于作物遗传分析和改良。
基因组测序大大提高了人们对于植物基因组组成、表
达、进化和功能的研究能力,提高了全基因组范围多
态性发现的效率[15]。 探索作物种质资源基因组多样化的驱动力及其多
样性的遗传基础是作物遗传育种的一个挑战性命题。
密集的分子标记遗传图谱和基因组序列可以更精细地
评价作物基因库的多样性。越来越多的证据指出,一
些作物,例如玉米的遗传多样性的很大一部分来自于
整个表达基因的存在 /缺失变异(presence/absence variations,PAV)和数目变异(copy number variant,CNV)。当 PAV 为主要遗传多样性时,参考基因组
序列不能完整描述某个物种的遗传组分。由此提出了
“全基因组”概念。全基因组由包含所有种质共同的
基因组特征的核心基因组,以及只有部分或没有共有
序列的非必需基因组两部分组成。因此,要全面描述
一个物种的基因组组成,需要测定不同种质系的基因
组序列。揭示非必需基因组的成分和功能可能是了解
表型多样性发生机制的关键步骤。显然,这种功能基
因含量的变异预示自交系功能基因的丢失可以在杂交
种中得到互补,从而导致杂种优势效应[16]。分子证据
表明几个世纪的选择育种过程中,作物丢失了大量有
用或者无用的等位基因。要充分利用种质资源中的有
利等位基因资源,需要提高对关键性状分子基础的认
识,拓宽种质资源库的表型和基因型,提高外源染色
体片段的渗入效率和比例。 分子标记的应用可以提高种质资源的鉴定与创新
和育种的效率。MAS 广泛应用于渗入性状和简单遗传
基础的性状选择,尤其是抗病性选择。通过回交和分
子标记辅助选择,将野生种有益等位基因渗入到优良
栽培种时,连锁累赘可能降低了 终效果。对于复杂
的农艺性状,由于存在受环境影响表达的 QTL 之间的
互作,使问题复杂化。这些缺点可以通过大规模开发
基因组资源,从生理水平解析 QTL×环境、QTL×QTL互作得到部分解决。 2.2.2 QTL 与基因组选择 作物育种的主攻性状几
乎都是复杂的数量性状。 新的基因组学,特别是
NGS 技术的发展使人们可用更低的成本获取更多的
植物种质资源遗传组成信息。表型鉴定技术的显著进
步使人们能够以前所未有的准确度、速度和低成本测
定控制条件下或田间多个个体农艺相关表型,其结果
是 QTL 的数目急剧增加而单个 QTL 的贡献却很小。
因而有人认为目前“QTL 在纸上而不是在品种里”[17]。
通过 QTL 元分析(Meta 分析),可以将来自不同群
体、不同环境同一性状的 QTL 以及采用连锁分析或者
全基因组关联分析(genome-wide association study,GWAS)得到的 QTL 进行合成,从而更好地估计 QTL效应。另外,元分析还试图分析 QTL 的上位性、多效
性以及与环境的互作效应。但由于个体数量性状表现
型可能是非线性效应,正确预测等位基因以及 QTL 单
倍型新的重组可能有难度,需要进一步探索[18]。 对于器官、组织、生理、代谢等不同水平的性状
和不同发育时期的动态性状,如叶面积指数、植物生
物量、代谢物含量等,通过分析所谓隐藏的遗传变异
及其 QTL,可以鉴定复杂性状组分的 QTL[19-20]。与不
同组学研究相结合,获得表达 QTL(eQTL)、蛋白
质 QTL(pQTL)以及代谢 QTL(mQTL),可以用
来发现性状形成的分子机制[21]。所有这些都可能为育
种提供新的机会。虽然育种资源中含有少量强效应
QTL,但是一些主效 QTL 已经被定位、确证、克隆甚
至在分子标记辅助育种(marker assisted breeding,MAB)中应用。育种框架中的 QTL 信息可用于回交
育种和设计育种,或者单倍型育种。这些途径谋求通
过标记辅助选择已知的关键位点上的有益等位基因培
养新品种。QTL 一旦被克隆,就可以用来培育转基因
作物。 QTL 效应一般呈 L 形分布,即大效应 QTL 数目
逐渐减少,而小效应 QTL 数目逐渐增多。目前,大多
数研究的检测能力不能鉴定大量数目的小效应 QTL,也就是说大量的小效应 QTL 被埋没在“冰山”之下。
大效应 QTL 容易在育种选择过程中丢失,而小效应
QTL 在育种选择过程中,不太容易被淘汰。因此,除
了从种质资源中“重新获取”那些消失的大效应 QTL用于 MAS,还要考虑如何有效捕获和利用大量的小效
应 QTL。 近提出的基因组选择(genome selection,GS)是 MAS 的延伸。这种方法不考虑单个 QTL,而
是通过对分布于整个基因组的大量数目分子标记基因
型估算个体的育种值,实际上,其模型针对的是无法
检测的小效应 QTL。该法根据多标记基因型计算个体
育种值(genomic estimated breeding value,GEBV)作
为选择值。和 MAS 一样,GS 包括 2 个步骤:练习期
和育种期。在练习期对亚群体进行表现型和基因型的
关联分析,在育种期鉴定育种群体的基因型,按由基
因型估算的 GEBV 进行选择。GS 可望能够解决 MAS不能揭示的微效基因选择问题。但是也有观点认为,
GS 可能并不适用于植物复杂性状的改良[22]。GS 是一

3308 中 国 农 业 科 学 48 卷
个植物育种潜在有用的方法,但是 GS 不是表型选择
的完美替代。它应该与传统育种整合,使传统育种更
加快速高效。 基因组学数据还为设计和培育携带预先设定的性
状的新品种(分子设计育种)提供了可能性。全球基
因组学正集中努力揭示农艺性状的遗传基础,分析相
应位点等位基因变异, 终使育种者能够先模拟设计
理想的类型,再在植物中构建新的基因型。虽然这个
途径目前还未收到育种家的普遍重视,但已有一些报
道。例如具有抗多种病虫害、高营养效率和抗旱的“绿
色超级水稻”新品种的培育中,第一个阶段已经部分
取得成功,已经培育和评估出携带单个基因并对关键
性状的表达有主效作用的优良家系。第二阶段,累积
所有控制目标性状的主效基因,培育具有多重抗性和
适应性的性状,此阶段正在进行。如果全面贯穿,GS不仅将有助于设计具有少数目标性状,还将可能设计
具有基因组所有优良目标性状位点的新植物,完全实
现分子设计育种的概念。 2.2.3 等位基因挖掘:通过测序探究植物多样性 采用 DNA 技术可以在一个代表性的种质库中大规模解
析与特定等位变异关联的基因或候选位点变异。第一
个挖掘等位基因的技术是 Eco-TILLING,通过核酸酶
剪切杂合双链 DNA 分子,测定每个基因型与参考基
因型之间的 SNP。随着成本的不断降低,应用 NGS技术可以进行复等位基因检测,等位基因挖掘正成为
一种探索植物多样性的新常规化方法。而且,NGS 为
分析单倍型结构(包括频率、类型等)、拓展关联作
图铺平了道路。早期的等位基因挖掘研究完全建立在
搜索基因编码区的突变,因此,与蛋白质的结构和功
能的变化有关。目前的重点已经延伸到对基因表达水
平有重要效应的非编码序列,例如内含子、UTR(untranslated region)和启动子。
在种质选择中,对特异基因和明显关联等位基因
的挖掘正发挥越来越重要的作用。种质资源重点鉴定
策略(focused identification of germplasm strategy,FIGS)是结合环境和植物特征来鉴定对某一特定环境
具有潜在意义的种质资源的方案。应用 FIGS,例如,
从一个拥有 16 089 个种质系的种质资源中选出 1 320个面包小麦地方品种,进行等位基因挖掘,搜索 Pm3位点白粉病抗性的变异。分析该位点带有核酸变异种
质系的表型,发现了 7 个新的抗性等位基因。对几百
个大麦基因型进行等位基因挖掘,发现 Mla 位点上有
白粉病抗性新位点、Rdg2a 叶条纹病抗性以及
VRN-H1/H4 春化位点有新等位基因。在水稻中,等位
基因挖掘能够鉴定 Wx 位点编码颗粒结合淀粉合酶等
等。等位基因挖掘主要集中于单个或者几个特别位点,
新的 NGS 可以同时定向测定几百个位点,整个外显
子组(exome),或者整个基因组。将这些技术运用
到大的种质资源库可以在一个平行测定中发现所选择
的若干个种质系的大多数位点的等位基因多样性。例
如对 500 个中国水稻地方品种进行重测序大规模评价
基因组遗传变异,鉴定了 360 万个 SNPs,用于描绘籼
稻和粳稻地方品种的连锁不平衡结构(LD),构建高
密度的单倍型图谱。通过对 14 个农艺性状的全基因组
进行关联分析,表明这个途径是进行遗传作图、复杂
性状遗传分析的重要替代方案。大豆中,综合 GWAS对 31 个栽培种和野生种进行重测序,检测到高水平
LD 以及大约 200 000 个 SNPs,可以用来定位复杂性
状的 QTL[23]。 2.2.4 转基因育种 植物转基因完全打破了物种界
限,为育种者提供了传统方法无法接触的新基因。来
自不同物种的基因可以插入并在农作物背景中表达,
而且没有其他附加效应。通过选择不同的启动子,可
以改变基因时空表达模式和强弱。转基因可以解决许
多在主要作物中缺乏而且难以引进的基因。目前已经
在抗病、虫、草等性状上获得了成功。除此之外,作
物的营养、品质、磷吸收利用效率、固氮效率、对土
壤盐分和铝毒耐性、光合速率、糖淀粉生产、雄性不
育和无融合生殖固定自交作物的杂种优势等性状都被
列为转基因(GM)的目标性状。目前,全世界的实
验室里已经获得了大量的转基因作物,但实际应用的
很少。除了政治和环境方面的考虑之外,单个或者少
数几个基因对于复杂性状表现的影响非常有限,尤其
是释放到大田经过“稀释”之后。植物转基因还容易
产生基因组位置效应和基因沉默现象。然而,科学家
相信,未来 GM 将是育种的重要途径。2012 年全球
GM 作物面积达到 1.7×108 hm2,占总耕地面积的
11%,52%的 GM 作物种植在发展中国家,而且发展
中国家 GM 作物面积的增长率(11%)高于发达国家
(3%)。92%以上的商业化 GM 作物集中在美国、巴
西、阿根廷、加拿大、印度和中国。98%的 GM 作物
是大豆(47%)、玉米(32%)、棉花(14%)和油菜
(5%)。主要性状为抗除草剂(主要为抗草甘膦和草
铵膦)和抗虫[23]。 2.2.5 超 GM 植物——新育种技术 随着分子生物学
和基因组学技术的发展,现在可以利用重组 DNA 技

17 期 盖钧镒等:中国作物种业科学技术发展的评述 3309
术在作物品种早期选择阶段引入新的性状,而 后获
得的植物基因组不含任何外来 DNA 系列。这样的技
术总称为新育种技术。新育种技术促进了 GM 事件的
发生,但是它们的终产物没有外源基因,获得的品种
类似于采用传统点突变技术。考虑到早期阶段 GM 的
介入,在法律上将通过这些技术获得的作物归类为生
物技术来源作物(例如欧盟指引 2001/18/EC)。 目前, 重要的新育种技术是促进座位特异性点
突变技术。由于基因序列的微小变化对于植物基因型
可以产生显著的影响,这种技术试图对基因组特定序
列进行修饰,用来创造新的等位基因。寡核苷酸定点
突变(oligonucleotide-directed mutagenesis,ODM)、
转录激活样效应因子核酸酶(transcription activator- like effector nuclease,TALEN)技术、锌指核酸酶
(zinc-finger nuclease,ZFN)、成簇规律间隔短回文
重复(clustered regulatory interspaced short palindromic repeat,CRISPR)技术可以特异性修饰某个靶向序列,
与传统的点突变相似,但是效率更高,速度更快,没
有多余的随机突变。这些技术对育种的意义和影响还
有待观察,但其有创新性的前景。例如,利用 ODM方法获得了玉米、小麦、水稻、烟草的新除草剂抗性
以及油菜和香蕉抗生物胁迫性;利用 CRISPR 技术可
以提高小麦对白粉病的抗性[24]。 2.3 现时中国需要优先解决的作物育种理论和技术
问题
当前,中国种子企业的育种力量正在形成或发展
壮大,作物种业的育种队伍主力仍然在公益性研究单
位,这两支力量主要从事传统育种工作,新品种几乎
完全出自这两支队伍。公益性研究单位还有一支相当
强的力量也在从事与育种有关的研究,但受 SCI 论文
分数的驱使,虽然研究的命题和育种有关,实际偏向
于分子生物学。这两部分工作正巧分别与以上所述传
统作物育种科学技术和现时分子生物育种研究热点相
对应。目前,中国作物育种这两方面的研究在很大程
度上还处于分离状态,缺少交叉和融合。因而现时分
子生物育种研究热点的工作与实际种业的育种工作间
有待建立起相应的桥梁或通道,在育种实践中检验、
优选、修正和延伸。这中间的路程需要组织科技力量去
打通,当然有些途径可能行得通,有些途径可能行不通。
从目前中国种业现状出发,步入规模化商业育种,需优
先解决的育种理论和技术问题有以下 8 个方面。 2.3.1 种业的目标性状 要研究纳入育种目标性状
的界定,通用型品种与专用型品种性状的配置和生态
适应性的优化。育种目标性状是以生产和利用为基准
的,与生育过程有关,以耐旱性为例,是各种耐旱机
制的综合表现,个别组成型性状不一定能代表真实的
育种要求,作为育种目标性状的要求不一定与个别生
理研究的指标一致[25]。因而必须界定育种目标性状的
要求,确定标准鉴定方法,在此基础上优化与生态区
和产品利用方向一致的综合目标要求。 2.3.2 性状的精准鉴定 育种的关键在于精准表型
鉴定和基因型鉴定。很难设想粗糙的试验评价会给出
精准的结论。要建立大规模育种材料田间产量的精准
鉴定技术(包括试验设计技术)、田间农艺性状的规
模化精准鉴定技术及品质、抗生物逆境、耐非生物逆
境、育性等的精准鉴定技术。这些是规模化育种重要
而必须解决的简单命题,商业化作物育种必然是规模
化育种,优良品种的界定、标记辅助基因型选择是手
段, 终还要落实在表型的真实评价,因而性状的规
模化精准评价是关键。 2.3.3 种业基因的源泉 各种作物国际种质资源的
收集,种质资源群体各类育种性状(产量、品质、抗
生物逆境、耐非生物逆境、育性等)的全基因组遗传
解析,每份资源的 QTL/基因及其等位变异的全基因组
构成分析。探索建立有效的遗传解析方法,用之于各类
作物,并能由资源群体推及各育种计划育种群体每份材
料的遗传构成。在此基础上实现育种的遗传设计,包括
杂交组合 优设计和后裔的 优选择。从而进一步推进
资源的研究与开发利用这项育种中 根本的工作。 2.3.4 性状基因体系的生态区特征 各种作物各类
性状的全基因组单一性状基因网络特征和综合性状基
因网络特征的解析及其在不同生态区的共性与特异性
解析,即性状的基因组学特征和生态特征的解析。这
是实现上述资源研究目标的基础和前提。 2.3.5 育种现代化技术 规模化育种的实施要求有
从田间到实验室的系列机器、仪器和设备、设施,这
是种业发展必须配套的条件建设。以高效的性状检测
为例,现代的高光谱遥感技术可以利用飞行器在短时
间大范围监测作物生长,高通量、快速反射数据与现
代信息技术结合,在规模化育种中具有极大的潜力[26]。
中国作物育种长期停留在手工劳动状态,没有形成相
配合的机械、仪器、设备、设施的研发体系,进口的
机器等不一定适合中国的试验环境。必须推动这方面
的研究和开发。 2.3.6 育种信息技术 要研究建立育种的信息化技
术体系,乃至育繁推一体化的信息技术体系。商业化

3310 中 国 农 业 科 学 48 卷
的规模育种要在季节容许范围内收集、记录、分析大
量数据,不仅育种试验本身,还有大量试验点的数据
收集、记录、分析与决策。因而高效的性状鉴定与数
据采集技术,数据的传输、整理与分析技术,或简言
之,育种信息技术将是未来规模化育种的重要技术。
2.3.7 标记辅助技术 分子标记辅助育种单性状和
综合性状技术方案的制定。包括高通量标记技术和高
通量优化决策技术。标记辅助育种研究采取“简单性
状的先用,复杂性状的试用”的原则,由简到繁、由
浅到深。已有大量积累的作物性状 QTL/基因定位结
果,需要归纳、整合,梳理出可用的技术方案,在实
际育种中试用、改进。同时要结合育种需求推动标记
辅助育种实用技术的研究和应用。 2.3.8 转基因技术体系 转基因技术在作物育种中
具有明显的突破性潜力,可以突破本作物自然界没有
的有利变异。转基因育种是常规育种的重要补充,是
在老品种的遗传背景下导入全新的基因,使老品种产
生突破性的改变。转基因是创新种质的手段,可以通
过转基因技术创造大量新种质、新材料。当务之急,
一是发掘出这类真有突破意义的基因,创造出大批育
种可用的材料,使转基因育种进入一个实质性的制高
点,现在是可用的基因太少了。二是必须严格保证其
安全性。伦理上的转基因障碍只有从社会的意念上去
解决,科学可以解决的是如何保证严格的安全性,例
如,如果转入产品的基因 后全部删除,检测不到,
也就谈不上后患了。这才能免除冗余的无休止的争议。
更重要的是,要严格实施转基因的法律法规,切实防
止转基因材料非法泄漏。
3 作物种子生产和示范推广科学技
术的进展
中国种业的历史较短,有关作物种子生产和示范
推广科学技术的研究还在推动、起步阶段。规模化种
业的产品是优良品种的种子,种子作为商品必须进行
标准化生产,每一批种子应该都可以溯源,可以追查
责任。所以种子生产的关键是建立标准化种子生产
规范,以及研究实现或超越种子生产标准的先进技
术[27]。商业化种业的产品销售自然有竞争,销售的竞
争实质上是种业示范推广科学技术的竞争。 3.1 种子生产的标准化[28]
种子生产标准化涉及以下各环节: (1)品种的标准化 需要对所生产种子的品种身
份进行验证,生产种子所用的原始种子应该是育种家
提供的育种家种子,以保证生产出符合该品种标准的
种子。 (2)种子生产程序和各个环节的标准化 种子生
产程序必须标准化,国际一般都采用育种家种子、基
础种子、注册种子和检定种子四级生产程序,国内河
南省提出育种家种子、原原种、原种和检定种四级生
产程序。种子生产田间为防止串粉必须设有隔离标准。
各级种子生产过程中的操作步骤,包括收获、脱粒、
烘干、清选、包装、标签、贮藏、运输等环节都必须
设有标准操作程序。 (3)种子质量的标准化 不同程序阶段所生产的
种子质量必须设有相应的标准,标准的内容因作物而
异,一般至少有净种子、杂质、其他作物种子量、其
他类型种子、杂草种子量、有毒(有害)杂草种子量、
发芽率、水分含量等。 (4)种子质量检验技术的标准化 为保证种子质
量标准的实现,种子质量检验技术也须建立标准方法。 中国种业规范还在形成之中,目前河南省提出了一
个省标准。全国的种子生产标准,国务院文件已提出需
要研究,但还未见眉目,这应是种业发展的当务之急。 国际发达国家均订有相应的种子生产标准,可以
借鉴。但各国体制、环境不同,一个国家的种子生产
标准应该建立在本国研究的基础上。以往中国对此缺
少系统研究,现在应该加快推动中国各类作物种子生
产标准的研究和制定。 3.2 种子生产的附加技术
国际种业发展中,对种子质量的保证投入了大量
研究。高档种子论粒销售的基础是种子生活力、发芽
率的有效保证。这和种子收获、加工、储藏的技术密
切相关。 国际种业发展中结合种子出售还附加了有益于作
物生长的农业技术,其中,首先是种子的处理和包衣
技术,这些附加的苗期病虫害防治技术、微量肥料的
添加技术、幼苗生长的壮苗技术增添了用户对种业公
司产品的好感和信心[27]。 目前,中国的种业公司在种子生产技术上还没有
努力下功夫,今后的商业竞争机制可能会推动种子生
产新技术的研发。相应地,种业的发展还需要以种业
机械的配套发展为支撑。 3.3 作物种业示范推广的科学技术
种业企业种子产品的销售竞争 终是品种水平和
示范推广科学技术水平上的竞争。

17 期 盖钧镒等:中国作物种业科学技术发展的评述 3311
3.3.1 明确销售品种的最佳适应区域 种业企业
育成或销售的品种,企业本身应十分熟悉其适应的
地区范围,使企业的种子应用在 佳、 适应的地
区,因而有 好的效果和 佳的信誉。所以种业企
业应建立本企业的相当于区域试验的多点试验系
统,明确所销售品种种子的适销范围和该品种种子
的生产规模。 3.3.2 提供销售品种的最佳栽培技术 种业企业应
该有自己的栽培技术专家,品种育成前后对有苗头的
品种研究制定适合该品种特点的栽培技术。销售品种
种子的同时要向用户提供该品种应用的技术指导。品
种种子和其栽培技术同时销售使用户得到的高产水平
将为企业品种的水平增添信誉。 3.3.3 建立品种示范系统 用户选用品种种子,常需
有直观的判断。种子企业必须有示范试验供用户去观
察、遴选。因而企业要在销售地区建立相对固定的示
范点系统,让用户随时去评比,做出自己的判断。 3.3.4 种子售后服务 种子售后的技术咨询服务和
多媒体技术,是巩固企业信誉度的有效手段。 中国的种业刚从公益性事业转成商业化体制,还
没有进入常规秩序。一些不法企业套牌侵权、假冒伪
劣种子混淆市场。国家已明确要采取措施,建立种子
市场秩序行业评价机制,督促企业建立种子可追溯信
息系统,完善全程可追溯管理,从而规范种子营销网
络,保证企业公平竞争,保障用户权益。
4 中国种业科学技术发展展望
4.1 围绕种业发展的需求,建成相对完整的种业科学
技术学科体系
支撑种业战略性产业的科学技术应是一群而不是
单一学科。因为种业的产品是品种和它的种子,作物
育种学和种子学是其核心内涵,但不是种业科学的全
部;遗传学和生理学又是核心内涵的基础,但不是育
种学和种子学的全部;以此类推,分子遗传学目前是
遗传学 为活跃的一部分,但不是遗传学的全部,更
不是种业科学的全部。局部不能代表全部,因而种业
科学要发掘它本身的全面内涵。中国商业化种业的实
践还在起步阶段,种业科学的全面内涵还在形成之中。
图 1 中的下游部分大致代表了种业科学技术的主体部
分,中游和上游部分是以育种为主的构思,围绕种子
生产、加工、检验和推广营销的相关学科内容以及有
关种业的国内和国际法规等都还未包含在内。当然,
这只是种业科学上下游领域初步构架,还有待进一步
完善和深化。
种质
创新Germ
plasmenhanc
ement
种业科学
与技术
Breeding methods and technology
新品种试验New cultivar testing system
种质创新
Germplasm enhancement
种业科学与技术Science and technology in seed industry
种业产业Seed industry
中游
Mid
stre
am
上游
Ups
tream
下游
Dow
nstre
am新品种选育New cultivar development
种子生产与加工储藏Seed production and processing/storage
种子检验与检疫Seed inspection and quarantine
种业推广与营销Extension and marketing of seed industry
种业市场管理Management of seed industry market
种业实验机械与设备 Experimental machinery and equipment in seed industry
种业政策与法规 Policy and regulation of the seed industry
生物学与生物技术Biology and biotechnology
种质资源
Germplasm resources
育种技术方法Breeding methods and technology
转基因技术Transgenic technology
基因QTL定位/标记辅助选择
QTL mapping/Marker-assisted selection
基因、基因组学与-omicsGene, genomics and -omics
图 1 种业科学上下游领域的一种构想
Fig. 1 A conception of the relationship between the seed industry discipline and its upstream and downstream disciplines

3312 中 国 农 业 科 学 48 卷
种业科学技术体系的建立要紧密联系种业生产实
际,在种业发展中逐步积累、完善和发展种业科学体
系。在此过程中,应该从国家的高度进行引导。围绕
建立育繁推一体化的种业体系中心目标,着眼于育繁
推各个环节,开展种业科学技术的设计和研发。管中
窥豹,只见一斑是不能形成一体化的种业科学技术体
系。 4.2 优化顶层设计,建设好企业种业科技和公益性种
业科技两支相互补充、相互配合的种业科学技术
研发力量 中国种业商业化起步晚,种业科技力量正在形成,
要建成育繁推一体化的种业科技体系,高水平专业技
术队伍建设是关键。目前中国作物育种的基本队伍仍
然是公益性占主导地位,在材料和技术上已有长期积
累。要进一步落实好“鼓励‘育繁推一体化’种子企
业整合现有育种力量和资源,充分利用公益性研究成
果,按照市场化、产业化育种模式开展品种研发,逐
步建立以企业为主体的商业化育种新机制”、“引导
和积极推进科研院所和高等院校逐步退出商业化育
种”等文件精神,要通过妥善的顶层设计,将两方面
的种业科技力量协同起来,形成合力。
顶层设计的一方面是种业企业要建立起强有力
的研发队伍。这要在政策上推动企业建立有竞争力
的育繁推一体化技术体系,要推动企业建立以领先
技术参与种业竞争的基本理念。然后要推动培养种
业的科技人才,以充实到种业企业中去。目前种业
人才相当缺少。育繁推一体化的技术链主体是要能
育种、能生产良种、能做田间示范的专家,品种的
优劣只有在田间试验中才能检验,种业的专家,首
先应该是田间试验的专家。但是目前高等学校培养
的作物遗传育种专业学生,因为受发表高分数论文
的驱使,基本以实验室工作和技能训练为主,缺乏
田间育种等实际工作能力的训练。而且目前作物遗
传育种专业的课程体系并不强调种子的生产与推
广,缺少育繁推各环节及其一体化相关技术和能力
的训练,所以国家应该推动农业高校适应种业发展
的需要,建立种业科学技术新专业,尽快培养出在
种业科技方面有开拓性的人才。根据中国目前的课
程设置状况,要专门委托有关高校,建立种业的新
课程并建立相应的实验实习基地和研发平台。图 2为一个作物种业科学专业本科生或专业硕士生必须
具备的科学知识设计。
作物育种学(总论与各论)Crop breeding (General and crop species)
种子生产与加工 (标准化)
Seed production and processing (Standardization)
种业推广与营销Seed industry extension and marketing种子检验与检疫Seed inspection and quarantine
仓储害虫与种子储藏Storage pests and seed storage
种业设备、设施与机械Equipment, facilities and machinery in seed industry
种业科学信息化技术Information technology in seed science
育繁推一体化课程The course system Integrating cultivar development, seed
production and seed extension
现代技术课程The courses of
modern techniques 种业社会管理课程The courses of social management in seed
industry
专业基础课程
Professional basic courses
遗传学——普通遗传学 General geneticsGenetics 细胞遗传学 Cytogenetics
数量遗传学 Quantitative genetics分子遗传学 Molecular genetics
种业市场管理Manegement of seed industry market
种业政策与法规Policy and regulation in seed industry
种质资源学 Germplasm resources试验统计学 Experimental statistics 基因组学 Genomics转基因育种技术 Transgenic breeding technology
种子学与种子生理 Seed science and seed physiology作物栽培学 Crop cultivation sciences作物病虫害防治 Crop disease and pest management
图 2 种业专业必须具备的科学知识
Fig. 2 The basic academic knowledge/course structure for the seed industry discipline
顶层设计的另一方面是要建立起推动公益性种业
科技专家将知识和研究成果(包括育种材料和育种技
术)在种业企业的育繁推一体化进程中及培育优良品
种中发挥作用。这在全世界普遍承认知识产权的今天,
关键是建立起合理的知识产权确认和转移的政策体
系。这需要系统地研究和认证。偏颇的政策起不了实
质性的作用。 顶层设计的再一方面是国家在种业科技研发上的
高瞻远瞩,围绕种业育繁推各环节、各方面全程设置
相互衔接、相互推动的导向性研究计划。当前种业发

17 期 盖钧镒等:中国作物种业科学技术发展的评述 3313
展方兴未艾的时候,国家要投入力量来研究和制定各
类各种作物的种子质量标准,通过标准来规范后续种
业的健康发展。在推动种业科技研发和规范种业标准
的前提下,进一步建设好企业和公益性两支有分工、
有合作的种业科技队伍。 4.3 发展中国种业科学技术的策略和建议
前文已列述一些需优先解决的作物育种理论和技
术问题,此处再就其实现的必要性和策略提出建议。 4.3.1 规模化育种技术 中国种业企业首要是建立
起育繁推一体化的研发体系,真正依靠科技立业。收
购上家品种、销售种子算不上科技型种子企业。这类
企业并无竞争潜力,不是国家要支持的种业企业。已
建有育繁推研发体系的种业,面临的就是快速发展规
模化育种技术的命题。 先锋公司每年测试上百万个杂交组合,只有十万
分之一被选中商业化。如此大规模的育种体系如果没
有标准化的操作技术和育种程序作为支撑,是不可能
领先世界的。因此,规模化、程序化和标准化是现代
商业育种的重要标志。 规模化育种技术的内涵涉及田间试验规模化、试
验操作机械化、鉴定技术现代化、数据采集信息化、
试验布局区域化、良种决选全局化、育繁推一体化等。 4.3.2 资源富集、遗传解析与创新 种业的基础是育
种材料,育种材料衍生自种质资源,种业企业必须掌
握的财富是种质资源和优异素材。种质资源是国家的
财富,国家种质资源库应该是种业原材料的第一源泉,
公益性育种应该是种业原材料的第二源泉。 国家要鼓励、支持种业企业收集全世界的有用资
源。关键是严格检验和检疫手续。目前海关的管理措
施滞后于国家种业发展的需求。
产量遗传体系(
Genetic system conferring yield (
株型(光截取)
Heads or branches
Dry matter accumulation
Genetic system conferring quality traits
选育 优基因型:优良品种(优异基因的集合)
Breeding for optimized genotype: super cultivar ( A collective o
耐非生物逆境(耐荫性)
Adaptability to local environments
Genetic system supporting yield realization
产量遗传体系(QTL)Genetic system conferring yield (QTLs)
株型(光截取)Plant type (light interception)光合效率 Photosynthesis efficiency
生物量 Biomass收获指数 Harvest index
株数 No. plants穗数/分枝数 Heads or branches籽粒数 No. seeds籽粒重 Seed weight
资源高效利用 Efficient utilization of resources杂种优势 Heterosis干物质积累 Dry matter accumulation生长模型 Growth modelling
质量性状遗传体系Genetic system conferring quality traits
育性和其他Fertility and others
选育 优基因型:优良品种(优异基因的集合)Breeding for optimized genotype: Super cultivar ( A collective of elite genes)
抗生物逆境Resistance to biotic stresses
耐非生物逆境(耐荫性)
Tolerance to abiotic stresses (to shadding)
农作效率Farming efficiency
环境适应性Adaptability to local environments
熟期类型Maturity types
支撑遗传体系Genetic system supporting yield realization
图 3 作物育种性状的归类
Fig. 3 Grouping the breeding target traits in crops
种质资源库中的有用基因有待深入发掘。国家要
组织力量对资源库的种质按育种性状进行遗传解析,
明确各性状群体的遗传组成和个体的遗传组成,即
QTL/基因及其等位变异的组成。图 3 大致归纳了各类
作物的性状类别,对于种质群体的遗传解析要落实到
每个性状[29]。穷尽资源群体遗传解析的数据库就可借
以推测到种业企业育种材料的遗传结构,为种业企业
的设计育种奠定基础。现有的分子技术已经为种质资
源的遗传解析提供了有效手段,关键在于国家对相关
科研工作的组织和安排。 作为种业原材料第二源泉的公益性育种,应该明
确以创造突破性材料和创新育种技术为主要任务,使
他们的工作成就惠及全国的种业,并以此为荣。国家
和社会要建立起知识产权权益和荣誉的分配机制。 种业企业的育种特色就体现在本企业特有的创新
种质和材料上,这是种业原材料的第三源泉,也应该

3314 中 国 农 业 科 学 48 卷
是 活跃的创新源泉。未来中国的种业企业间的竞争
若能达到这个境界,中国的种业发展水平也就达到国
际化了。 4.3.3 推动常规育种分子辅助技术的应用 新品种
来自老品种,基本上继承了老品种的遗传体系,但在
继承中有创新,通过重组、诱变、渗入等途径吸收了
新基因。育种的大量工作是要鉴别出哪些新基因纳入
了正在创造的品种,基因看不见,因而发明了许多办
法可以从表型判断新基因的存在。发展了 30 多年的分
子标记技术,从可见的分子标记去判别正在创造的品
种中新基因的存在,称为标记辅助育种,包括标记辅
助优化组合和标记辅助后裔选择。迄今为止,报道了
数以万计的 QTL 和基因定位、标记辅助育种的文章,
也已有数以万计的硕士博士得到学位,但是在种业企
业和公益性的实际育种中鲜见标记辅助育种的成效。
究其原因,关键在于通向实际育种的 后一公里没有
走好。要为育种或性状的改良提出标记辅助的实际方
案,而且有实践检验结果的证实。因而,国家和社会
要推动研究标记辅助育种的实际应用技术和方案。传
统育种是基本的,要赋予传统育种以推陈出新的技术。 4.3.4 品种区域适应性试验制度与品种审定制度的
完善 国务院文件指出:“积极推进构建一批种子产
业技术创新战略联盟,为有实力的“育繁推一体化”
种子企业建立品种审定绿色通道”。品种的审定取决
于区域试验的结果。目前中国现行的区域试验和审
定制度存在一些矛盾:一个种业企业选育的新品种
在本企业要经过若干年的产量试验,交出参加区域
试验又要经过若干年的区域试验,国家区域试验的
点数和环境数有限,虽然经过多年试验,但经历的
考核并不充分。而国际上跨国公司在全球近百个国
家建立育种研发中心或试验站,如孟山都全球育种
试验站有 180 多个,先锋公司有 126 个,并以这些
研发中心为依托,广泛布局试验点,开展大规模育
种材料筛选和品种适应性试验。每个杂交组合在商
业化推广前至少经过 150 多个试验点和 200 多个不
同地区农民的种植测试评价,确保适合当地种植。
参考国际公司的经验,国务院提出的为“有实力的
‘育繁推一体化’种子企业建立品种审定绿色通道”
的意见是有可能实现的。条件是种业企业要承担品
种的全部责任;要建立起如跨国种业公司那样的内
部试验体系;再加上要推动保险业为种业企业建立
种子保险业务的体系,平衡企业的风险,保障用户
的权益。
4.3.5 配齐种业基础性公益性研究,确立种子生产
标准化体系 种业基础性公益性研究除上述种质创新
和育种技术创新等以外,还必须在育种过程、种子生
产和种子推广的设备、设施方面配以基础性、公益性
的科学技术研发,从而提高育繁推一体化的水平。当
前首先要在确立种子生产标准化体系的基础上,研发
种子生产各环节的设备和设施,以全面提高种子的质
量。进一步还必须着手多功能的种子处理和包衣技术,
并将现代栽培技术融于种业之中。 4.3.6 加强作物杂优化研发,使杂种化成为中国未
来种业的特色 比起家系品种,杂种品种可以同时利
用杂合位点的显性效应和纯合位点的加性效应,理论
上有独特的优势。20 世纪初,杂交育种及杂种优势利
用成功应用于农作物育种,利用杂种优势有效地提高
了作物的产量、抗病、抗逆和适应性水平。杂种品种
还能更有效地保护品种权益。中国的水稻、玉米、油
菜和主要瓜菜作物品种已经实现了杂种化,还有一些
作物,如小米、棉花、小麦、大豆等已经研发了雄性
不育、三系(或二系)配套的技术体系,正在向杂种
应用发展,建议中国加强作物全面杂优化的研发,使
杂种化种业成为中国种业的特色。 References
[1] Tester M, Langridge P. Breeding technologies to increase crop
production in a changing world. Science, 2010, 327(5967): 818-822.
[2] Kingsbury N. Hybrid: The History and Science of Plant Breeding.
Chicago: University of Chicago Press, 2009.
[3] Howard P. Visualizing consolidation in the global seed industry:
1996-2008. Sustainability, 2009, 1(4): 1266-1287.
[4] 国务院. 国务院关于加快推进现代农作物种业发展的意见. 中华
人民共和国国务院公报, 2011, 12: 6-9.
The State Council. The State Council on Accelerating the
Development of Modern Crop Seed Industry. State Council Bulletin of
the People's Republic of China, 2011, 12: 6-9. (in Chinese)
[5] 国务院办公厅. 全国现代农作物种业发展规划(2012-2020 年).
(国办发〔2012〕59 号).
The State Council Office. The developmental programs of modern
crop seed industry in China (2012-2021). No. (2012) 59. (in Chinese)
[6] 国务院办公厅. 深化种业体制改革提高创新能力的意见.(国办发
〔2013〕109 号).
The State Council Office. Guidelines on structure reform of crop seed
industry for the enhancement of technical innovation. No. (2013)109.
(in Chinese)

17 期 盖钧镒等:中国作物种业科学技术发展的评述 3315
[7] Palmgren M G, Edenbrandt A K, Vedel S E, Andersen M M, Landes X,
Østerberg J T, Falhof J, Olsen L I, Christensen S B, Sandøe P,
Gamborg C, Kappel K, Thorsen B J, Pagh P. Are we ready for
back-to-nature crop breeding? Trends in Plant Science, 2015. 20(3):
155-164.
[8] 申宗坦, 吕子同, 李壬生, 选育早熟矮稈水稻类型中一些性状的遗
传分析. 作物学报, 1965, 4(4): 391-402.
Shen Z T, Lü Z T, Li R S. Shorter breeding early maturing upland rice
in the genetic analysis of some. Acta Agronomica Sinica, 1965, 4(4):
391-402. (in Chinese)
[9] 司清林, 刘新伦, 刘智奎, 王长有, 吉万全. 阿夫及其衍生小麦品
种(系)的 SSR 分析. 作物学报, 2009, 35(4): 615-619.
Si Q L, Liu X L, Liu Z K, Wang C Y, Ji W Q. SSR analysis of Funo
wheat and its derivatives. Acta Agronomica Sinica, 2009, 35(4):
615-619. (in Chinese)
[10] 杨绪明. 岱字 15 号棉种的历史地位. 作物品种资源, 1985, 2: 4.
Yang X M. The historical status of cotton seed of Daizi 15. China
Seed Industry, 1985, 2: 4. (in Chinese)
[11] 王顺华, 潘家驹, 闵留芳, 宋银富. 修饰回交法培育棉花品种的初
步研究. 南京农业大学学报, 1985(3): 1-9.
Wang S H, Pan J J, Min L F, Song Y F. Modified backcross method
preliminary study on cotton varieties. Journal of Nanjing Agricultural
University, 1985(3): 1-9. (in Chinese)
[12] 袁隆平. 杂交水稻培育的实践和理论. 中国农业科学, 1977(1):
27-31.
Yuan L P. Practice and theory of hybrid rice cultivation. Scientia
Agricultura Sinica, 1977(1): 27-31. (in Chinese)
[13] 张爱民, 童依平, 王道文. 小麦遗传育种学家李振声. 遗传, 2008,
30(10): 1239-1240.
Zhang A M, Tong Y P, Wang D W. Wheat genetics and breeding
scientists Li Zhensheng. Hereditas, 2008, 30(10): 1239-1240. (in Chinese)
[14] Swaminathan M S. An evergreen revolution. Crop Science, 2006.
46(5): 2293-2303.
[15] Varshney R K, Nayak S N, May G D, Jackson S A. Next-generation
sequencing technologies and their implications for crop genetics and
breeding. Trends in Biotechnology, 2009, 27(9): 522-530.
[16] Schnable P S, Springer N M. Progress toward understanding heterosis
in crop plants. Annual Review of Plant Biology, 2013, 64: 71-88.
[17] Bernardo R. Molecular markers and selection for complex traits in
plants: Learning from the last 20 years. Crop Science, 2008, 48(5):
1649-1664.
[18] Salvi S, Tuberosa R. The crop QTLome comes of age. Current
Opinion in Biotechnology, 2015, 32: 179-185.
[19] Ghanem M E, Marrou H, Sinclair T R. Physiological phenotyping of
plants for crop improvement. Trends in Plant Science, 2015, 20(3):
139-144.
[20] Andrade F H, Sala R G, Pontaroli A C, León A J. Integration of
biotechnology, plant breeding and crop physiology. Dealing with
complex interactions from a physiological perspective//Crop
Physiology: Applications for Genetic Improvement and Agronomy.
New York: Elsevier Science, 2009: 267-276.
[21] Langridge P, Fleury D. Making the most of ‘omics’ for crop breeding.
Trends in Biotechnology, 2011, 29(1): 33-40.
[22] Jonas E, de Koning D J. Does genomic selection have a future in plant
breeding? Trends in Biotechnology, 2013, 31(9): 497-504.
[23] Borrelli G M, Orrù L, De Vita P, Barabaschi D, Mastrangelo A M,
Cattivelli L. Integrated views in plant breeding: From the perspective
of biotechnology//Sadras O V, Calderini F D. Crop Physiology:
Applications for Genetic Improvement and Agronomy (2nd Edition).
New York: Elsevier Science, 2015: 467-486.
[24] Wang Y P, Cheng X, Shan Q W, Zhang Y, Liu J X, Gao C X, Qiu J L.
Simultaneous editing of three homoeoalleles in hexaploid bread wheat
confers heritable resistance to powdery mildew. Nature Biotechnology,
2014, 32(9): 947-951.
[25] Langridge P, Reynolds M P. Genomic tools to assist breeding for
drought tolerance. Current Opinion in Biotechnology, 2015, 32: 130-135.
[26] Araus J L, Cairns J E. Field high-throughput phenotyping: The new
crop breeding frontier. Trends in Plant Science, 2014, 19(1): 52-61.
[27] Finch H J S, Samuel A M, Lane G P F. 12-Plant breeding and seed
production//Finch H J S, Samuel A M, Lane G P F. Lockhart &
Wiseman’s Crop Husbandry Including Grassland (Ninth Edition).
Woodhead Press, 2014: 263-283.
[28] Khanal N P, Maharjan K L. Institutionalization of community seed
production//Khanal P N, Maharjan L K. Community Seed Production
Sustainability in Rice-Wheat Farming. Springer, 2015: 163-172.
[29] Ghanem M E, Marrou H, Sinclair T R. Physiological phenotyping of
plants for crop improvement. Trends in Plant Science, 2015, 20:
139-144.
(责任编辑 李莉)

中国农业科学 2015,48(17):3316-3332 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.002
收稿日期:2015-03-06;接受日期:2015-04-29 基金项目:国家自然科学基金(31171548)、农业部“948”重点项目(2011-G9) 联系方式:贾继增,Tel:010-82105831;E-mail:[email protected]
作物基因组学与作物科学革命
贾继增,高丽锋,赵光耀,周文斌,张卫健
(中国农业科学院作物科学研究所,北京 100081)
摘要:作物科学的发展对于世界粮食的增产起着重要的作用。本文回顾了第一次绿色革命的历史。通过比较
中国与国际上矮化育种时间、矮源的来源以及同期主要绿色革命国家产量的提高幅度,证明中国应该是第一次绿
色革命的策源地与发起国之一。包括“中国绿色革命”、杂交水稻等成果在内的中国作物科学研究成果对中国及世
界作物生产作出了突出的贡献。当前中国的作物生产面临着严峻的挑战。第一次绿色革命之后的数十年间,中国
小麦等主要粮食作物遗传改良年增长仅为 0.7%—0.9%,远低于 1.7%的需求;肥水利用效率仅为发达国家的 1/3;
机械化程度虽有较大的提高,但与发达国家相比仍有较大差距;农产品品质不能满足人们的需求,食品安全存在
较大问题。鉴于上述情况,目前迫切需要开展一场以“更高产、更高效、更优质、更环保”为主要目标的新的绿
色革命。尽管传统的作物科学对作物生产作出了巨大的贡献,但事实证明,仅靠传统的作物科学难以完成新的绿
色革命重任。作物基因组学是当前生物科学领域发展最为迅速的一门新兴学科。当前大多数作物基因组测序已经
完成,标志着作物科学已经进入基因组时代,为基于基因组学的作物科学研究奠定了基础。基因组学的发展正在
促进作物科学在以下 4 个方面取得重要进展:(1)促进作物种质资源核心种质构建、重要农艺性状基因的克隆与
种质资源的开发与利用,推动种质资源变异组学科的形成;(2)促进作物育种理论与育种方法的重大突破,推动
育种基因组学的形成与发展;(3)推动环境条件与栽培措施影响基因表达调控的影响机制研究,一批受环境因素
调控的基因将被发现,促进栽培学研究向栽培基因组学的方向发展;(4)迅速提高特色作物的研究水平,缩小作
物间研究差距。当前是作物科学发展史上前所未有的高速发展时期,种质资源变异组学、育种基因组学与栽培基
因组学的发展将引起作物科学的革命,并将由此引发新的绿色革命。文中分析了中国当前在研究队伍的组织、研
究题目与研究材料的选择与研究成果的转化等方面存在的问题,并提出了解决方法及发展方向。
关键词:作物科学;基因组学;作物种质资源;作物育种;作物栽培
Crop Genomics and Crop Science Revolutions
JIA Ji-zeng, GAO Li-feng, ZHAO Guang-yao, ZHOU Wen-bin, ZHANG Wei-jian
(Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081)
Abstract: Development in the field of crop sciences is playing an important role in increasing yield throughout the world. The
history of the first Green Revolution was overviewed in this paper. According to the initiation time, gene donors and increased yield caused by dwarfing breeding over the same period, China should be one of the origins and initiators of the first Green Revolution. The achievements in crop scientific researches including Chinese Green Revolution and hybrid rice have made great contributions to crop production all over the world. However, China is facing serious challenges in crop production. Over the decades after the first Green Revolution, the increasing percentage of production per year attributed to genetic breeding for major crops accounts for only 0.7%-0.9%, far from the required 1.7% in China. The efficiency of fertilization and watering is only one third of that in developed countries. Although agricultural mechanization improved a lot in China, big gaps still existed in comparison with developed countries. The quality of agricultural products can’t meet requirements, and food safety is still a big problem. In view of the above, a new Green
jia
文本框
返回目录

17 期 贾继增等:作物基因组学与作物科学革命 3317
Revolution, aimed at higher yielding, higher efficiency, better quality, and friendly environment, is imperative. Although traditional crop sciences have made great contributions to crop yield, they are not competent enough to complete the new Green Revolution. Crop genomics is booming as a new subject in the biological sciences. The finished genome sequencing for many crops has advanced crop sciences to genomic era, which highlights the genomics-based crop research. Genomics has promoted five distinct improvements in crop sciences: (1) Great progresses in germplasm-based researches have been achieved, such as the construction of core collection of crop germplasms, cloning of agricultural important genes, development and application of germplasm resources, and introduction of new subject of germplasm variome. (2) Breakthroughs in theory and methology for crop breeding resulted in a new subject of breeding genomics. (3) The effect of environment and cultivation management on gene expression and regulation will be clarified. A lot of genes regulated by environmental factors will be discovered. These will direct cultivation research to cultivation genomics. (4) The breeding level for minor crops will be improved rapidly. Therefore, the research gaps between major and minor crops are narrowing. Nowadays, crop sciences are developing at an unprecedented high speed. The development of germplasm variome, breeding genomics and culture genomics will lead to revolutions in crop sciences and hence result in a new green revolution. Some problems in scientific research of crop sciences in China were listed, including organizations, topics, materials used, and achievements transformation. How to deal with them and which are the directions are also discussed in this review.
Key words: crop science; genomics; crop germplasm; crop breeding; crop cultivation
1 中国作物科学的成就与面临的主要
问题
在过去的 60 多年间,中国主要农作物生产在保障
中国的粮食与食品安全、促进国民经济发展与提供就
业岗位等方面取得了举世瞩目的巨大成就。中国以占
世界 9%的土地、6%的水资源养活了占世界 22%的人
口。近 50 年来,中国的粮食总产和单产均提高了 5倍多(FAO,2014)[1],其中,作物科学的发展对于
粮食增产起了关键的作用。迄今,尚未被人们认识的
“中国绿色革命”就是一个典型的事例。全世界都认
为 20 世纪 60—70 年代举世闻名的小麦与水稻“绿色
革命”的发源地是国际玉米小麦改良中心(CIMMYT)与国际水稻研究所(IRRI),绿色革命的代表国是印
度与巴基斯坦。而实际上中国的小麦与水稻的绿色革
命与上述组织(国家)是平行进行的。就矮化育种使
用的矮源而言,水稻矮化育种使用的矮源是来自中
国的地方品种低脚乌尖与自然突变体矮脚南特,其
中,低脚乌尖后来还成了国际水稻所矮化育种的主
要矮源;中国小麦矮化育种使用的矮源与 CIMMYT不同[2];就矮化育种时间而言,中国水稻矮化育种
始于 20 世纪五六十年代,还略早于国际水稻所;就
增产效果而言,中国粮食产量在 1960—1970 年 10 年
间年均增长 6.8%,等于印度(2.3%)和巴基斯坦(4.4%)
2 个国家之总和,是同一时期世界上增长速率最高的
国家(图 1-a,FAO 数据[1])。“中国绿色革命”对
于世界粮食增产所作的贡献远高于世界上任何其他国
家。因此,如果说 CIMMYT 与 IRRI 是绿色革命的发
源地,印度、巴基斯坦、菲律宾等国家是绿色革命的
代表国,中国则更应该是世界绿色革命的起源地与代
表国。中国还是世界上最早培育与推广杂交水稻的国
家。该项成果的推广与利用对于绿色革命之后中国粮
食产量的大幅度提高产生了重要的作用。长期以来,
中国是一个以农业为基础的国家,农业为今天国民经
济的高速发展作出了巨大的贡献。虽然目前农业在国
民经济中的地位有所下降,但仍然起着举足轻重的作
用。中国的种植业是劳动密集型行业,接纳了最多的
从业者,在以往的任何时期都提供了最多的就业岗位,
保障了中国社会的稳定。 当前,中国的国民经济发展在经历了 20 余年的高
速发展之后,进入了一个稳定持续发展时期。新形势
下国民经济的发展对于农作物生产提出了新的要求。 1.1 中国粮、棉、油的单位面积产量迫切需要大幅度
提高
由于中国是一个人口大国,粮食的需求量巨大。
绿色革命、杂种优势利用等成果的推广与利用曾大幅
度提高了中国及世界粮食作物的产量。但此后,中国
的产量进入一个缓慢增长时期,由原来的年增长近 7%降低到 1%左右(图 1-b),降低了 6—7 倍之多(FAO数据)。特别需要指出的是,即使这 1%左右的增长,
也是建立在大量使用水肥资源的基础上,而依靠品种
改良取得的进展则更少。例如,黄淮麦区是中国的小
麦主产区,也是育种水平最高的地区。对该地区近 20多年来育成品种的产量水平进行了统计,结果发现品
种改良的遗传进度仅为每年 0.7%(图 1-c)。由于粮
食生产的增速显著放缓,中国近年来进口粮食的速度
则成倍增长(图 1-d),2013 年谷物进口量高达 1 500
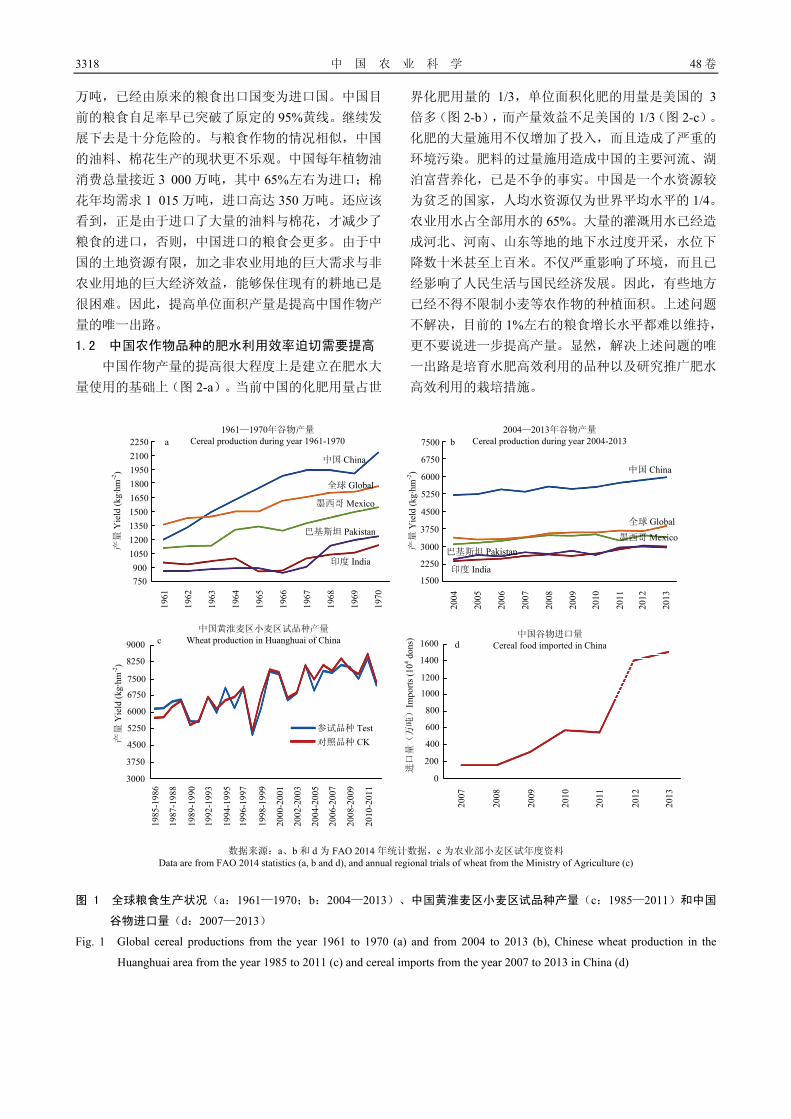
3318 中 国 农 业 科 学 48 卷
万吨,已经由原来的粮食出口国变为进口国。中国目
前的粮食自足率早已突破了原定的 95%黄线。继续发
展下去是十分危险的。与粮食作物的情况相似,中国
的油料、棉花生产的现状更不乐观。中国每年植物油
消费总量接近 3 000 万吨,其中 65%左右为进口;棉
花年均需求 1 015 万吨,进口高达 350 万吨。还应该
看到,正是由于进口了大量的油料与棉花,才减少了
粮食的进口,否则,中国进口的粮食会更多。由于中
国的土地资源有限,加之非农业用地的巨大需求与非
农业用地的巨大经济效益,能够保住现有的耕地已是
很困难。因此,提高单位面积产量是提高中国作物产
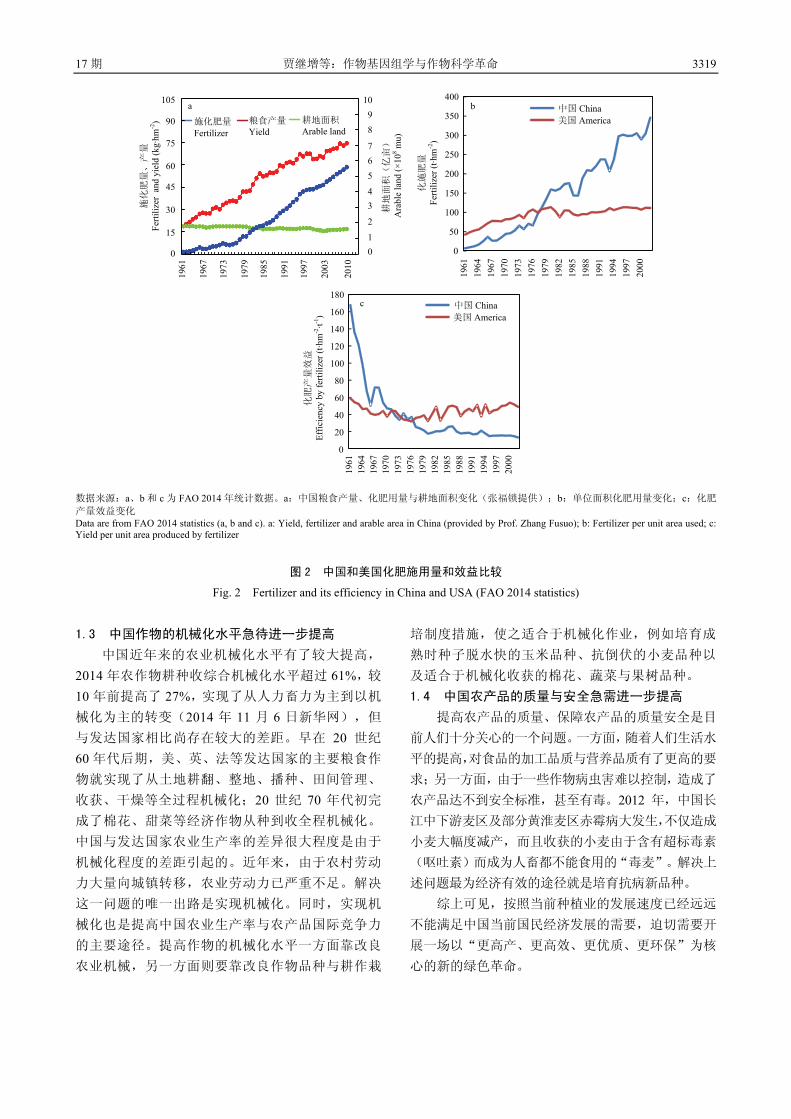
量的唯一出路。 1.2 中国农作物品种的肥水利用效率迫切需要提高
中国作物产量的提高很大程度上是建立在肥水大
量使用的基础上(图 2-a)。当前中国的化肥用量占世
界化肥用量的 1/3,单位面积化肥的用量是美国的 3倍多(图 2-b),而产量效益不足美国的 1/3(图 2-c)。
化肥的大量施用不仅增加了投入,而且造成了严重的
环境污染。肥料的过量施用造成中国的主要河流、湖
泊富营养化,已是不争的事实。中国是一个水资源较
为贫乏的国家,人均水资源仅为世界平均水平的 1/4。农业用水占全部用水的 65%。大量的灌溉用水已经造
成河北、河南、山东等地的地下水过度开采,水位下
降数十米甚至上百米。不仅严重影响了环境,而且已
经影响了人民生活与国民经济发展。因此,有些地方
已经不得不限制小麦等农作物的种植面积。上述问题
不解决,目前的 1%左右的粮食增长水平都难以维持,
更不要说进一步提高产量。显然,解决上述问题的唯
一出路是培育水肥高效利用的品种以及研究推广肥水
高效利用的栽培措施。
900105012001350150016501800195021002250
750
产量
Yie
ld (k
g·hm
-2)
1962
1961
1963
1964
1965
1966
1967
1968
1969
1970
2005
2004
2006
2007
2008
2009
2010
2012
2013
2011
中国 China
全球 Global
墨西哥 Mexico
巴基斯坦 Pakistan
印度 India巴基斯坦 Pakistan
印度 India
墨西哥 Mexico
全球 Global
中国 China
1500
2250
3000
3750
4500
5250
6000
6750
7500
8250
9000
3000
3750
4500
5250
6000
6750
7500
产量
Yie
ld (k
g·hm
-2) 1400
1600
0
200
400
600
800
1000
1200
2007
2008
2009
2010
2012
2013
2011
1987
-198
8
1985
-198
6
1989
-199
0
1992
-199
3
1994
-199
5
1996
-199
7
1998
-199
9
2000
-200
1
2002
-200
3
2004
-200
5
2006
-200
7
2008
-200
9
2010
-201
1
1961—1970年谷物产量Cereal production during year 1961-1970
2004—2013年谷物产量Cereal production during year 2004-2013
中国黄淮麦区小麦区试品种产量Wheat production in Huanghuai of China
中国谷物进口量Cereal food imported in China
参试品种 Test对照品种 CK
a b
c d
产量
Yie
ld (k
g·hm
-2)
进口量(万吨)
Impo
rts (1
04do
ns)
数据来源:a、b 和 d 为 FAO 2014 年统计数据,c 为农业部小麦区试年度资料 Data are from FAO 2014 statistics (a, b and d), and annual regional trials of wheat from the Ministry of Agriculture (c)
图 1 全球粮食生产状况(a:1961—1970;b:2004—2013)、中国黄淮麦区小麦区试品种产量(c:1985—2011)和中国
谷物进口量(d:2007—2013)
Fig. 1 Global cereal productions from the year 1961 to 1970 (a) and from 2004 to 2013 (b), Chinese wheat production in the
Huanghuai area from the year 1985 to 2011 (c) and cereal imports from the year 2007 to 2013 in China (d)
17 期 贾继增等:作物基因组学与作物科学革命 3319
1964
0
50
100
150
200
250
300
350
400
1961
1964
1967
1970
1973
1976
1979
1982
1985
1988
1991
1994
1997
2000
化施
肥量
Ferti
lizer
(t·h
m-2
)
中国 China美国 America
b
0
20
40
60
80
100
120
140
160
180
1961
1967
1970
1973
1976
1979
1982
1985
1988
1991
1994
1997
2000
化肥产
量效
益Ef
ficie
ncy
by fe
rtiliz
er (t
·hm
-2·t-1
)中国 China美国 America
c
施化肥量Fertilizer
粮食产量Yield
耕地面积Arable land
施化肥
量、产
量Fe
rtiliz
er a
nd y
ield
(kg·
hm-2
)
耕地
面积(
亿亩)
Ara
ble
land
(×10
8m
u)
a105
90
75
60
45
30
15
0
765
43
2
10
1098
1961
1967
1973
1979
1985
1991
1997
2003
2010
数据来源:a、b 和 c 为 FAO 2014 年统计数据。a:中国粮食产量、化肥用量与耕地面积变化(张福锁提供);b:单位面积化肥用量变化;c:化肥
产量效益变化 Data are from FAO 2014 statistics (a, b and c). a: Yield, fertilizer and arable area in China (provided by Prof. Zhang Fusuo); b: Fertilizer per unit area used; c: Yield per unit area produced by fertilizer
图 2 中国和美国化肥施用量和效益比较
Fig. 2 Fertilizer and its efficiency in China and USA (FAO 2014 statistics)
1.3 中国作物的机械化水平急待进一步提高
中国近年来的农业机械化水平有了较大提高,
2014 年农作物耕种收综合机械化水平超过 61%,较
10 年前提高了 27%,实现了从人力畜力为主到以机
械化为主的转变(2014 年 11 月 6 日新华网),但
与发达国家相比尚存在较大的差距。早在 20 世纪
60 年代后期,美、英、法等发达国家的主要粮食作
物就实现了从土地耕翻、整地、播种、田间管理、
收获、干燥等全过程机械化;20 世纪 70 年代初完
成了棉花、甜菜等经济作物从种到收全程机械化。
中国与发达国家农业生产率的差异很大程度是由于
机械化程度的差距引起的。近年来,由于农村劳动
力大量向城镇转移,农业劳动力已严重不足。解决
这一问题的唯一出路是实现机械化。同时,实现机
械化也是提高中国农业生产率与农产品国际竞争力
的主要途径。提高作物的机械化水平一方面靠改良
农业机械,另一方面则要靠改良作物品种与耕作栽
培制度措施,使之适合于机械化作业,例如培育成
熟时种子脱水快的玉米品种、抗倒伏的小麦品种以
及适合于机械化收获的棉花、蔬菜与果树品种。 1.4 中国农产品的质量与安全急需进一步提高
提高农产品的质量、保障农产品的质量安全是目
前人们十分关心的一个问题。一方面,随着人们生活水
平的提高,对食品的加工品质与营养品质有了更高的要
求;另一方面,由于一些作物病虫害难以控制,造成了
农产品达不到安全标准,甚至有毒。2012 年,中国长
江中下游麦区及部分黄淮麦区赤霉病大发生,不仅造成
小麦大幅度减产,而且收获的小麦由于含有超标毒素
(呕吐素)而成为人畜都不能食用的“毒麦”。解决上
述问题最为经济有效的途径就是培育抗病新品种。 综上可见,按照当前种植业的发展速度已经远远
不能满足中国当前国民经济发展的需要,迫切需要开
展一场以“更高产、更高效、更优质、更环保”为核
心的新的绿色革命。

3320 中 国 农 业 科 学 48 卷
2 大多数作物基因组测序的完成为基于
基因组学的作物科学研究奠定基础
以基因组学为代表的各类组学是当今发展最为迅
速的一门科学。一门新兴学科的兴起必将促进与之有
关的应用学科的飞跃。例如,孟德尔-摩尔根遗传理论
促进建立了作物育种学;分子生物学的发展促进了包
括分子标记辅助选择和转基因育种在内的分子育种的
发展。基因组学将全面促进作物种质资源、遗传育种
与作物栽培等学科取得突破性进展,从而促成新的绿
色革命。 基因组学研究是近年来世界上发展最为迅速的一
个研究领域。随着基因组测序技术的迅速发展,越来
越多的主要农作物基因组测序取得了突出进展。继水
稻基因组测序完成之后,目前世界各国已完成或接近
完成 64 种作物的基因组测序,其中包括主要粮食作物
(小麦、玉米、高粱、谷子等)、经济作物(棉花、
大豆等)、园艺作物(主要蔬菜、果树等)等(表 1)。
中国在该研究领域中作出了突出的贡献。在已完成或
即将完成的 64 种作物中,有 25 种是由中国独立完成
或参与完成。基因组测序与分析发现:(1)不同作物
虽然基因组大小差异巨大,但其二倍体基因组中的数
量相似,均为 3 万—4 万个,基因组大小的差异主要
是由于重复序列的比例不同所致;(2)物种的特性与
相关基因的数量密切相关。例如高粱的抗旱性与其携
带较多的抗旱基因有关[3],小麦的抗寒性与其携带较
多的抗寒基因有关[4];(3)同一物种不同材料的基因
组大小差异可高达 20%左右[5],而人与黑猩猩的差异
仅为 1%[6]。因此,一个作物物种需要多个参照基因组,
特别是杂合基因组。 农作物基因组测序的完成标志着作物科学已经进
入基因组学时代。基因组测序不仅第一次在全基因组
水平揭示了该物种的组成,而且为之后的种质资源变
异组学研究、育种基因组与栽培基因组奠定了基础。
3 基因组时代作物科学的重要进展
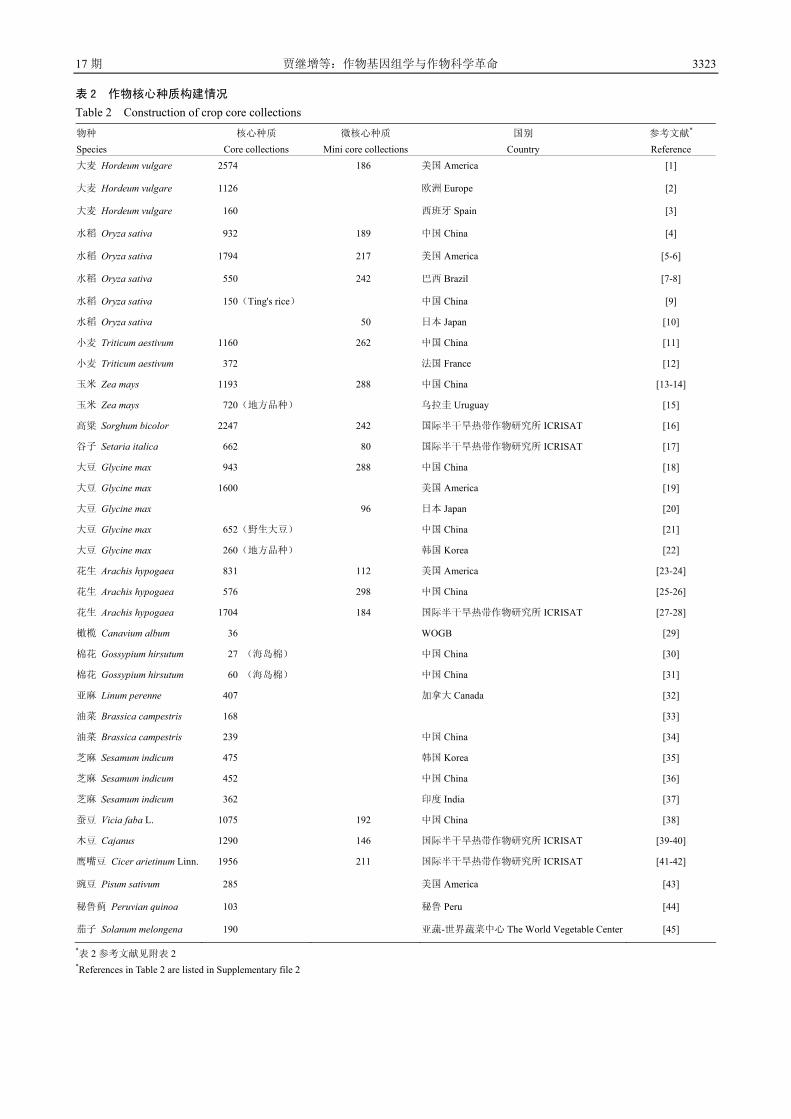
3.1 基因组学对作物种质资源研究的促进——建立
种质资源变异组学
育种亲本是作物育种的根本。第一次绿色革命的
突破即源于小麦、水稻等矮源的发现与利用;中国杂
交稻的突破则是由于“野败”不育系资源的发现与利
用。鉴于种质资源的重要性,世界各国及国际组织都
十分重视种质资源的收集、保存、研究与利用。目前,
世界上保存的各类作物种质资源已达 740 万份,其中
中国保存的种质资源有 44.5 万份,居世界第二[7]。然
而由于研究水平的局限性,这些资源中的重要基因基
本没有得到开发,致使育种进展缓慢。基因组学的发
展为种质资源研究与开发利用提供了全新的理论、技
术与方法,全面促进了种质资源研究的发展。2003 年,
中国农业科学院在北京组织召开了第一届基于基因组
学的种质资源国际学术讨论会(GPGR),目前已召
开了 3 届,标志着国际上对这一新的研究领域的认可。
在基因组学的促进下,各主要农作物都先后建立了核
心种质(表 2)。核心种质的建立解决了种质资源数
量巨大、难以深入进行研究与利用的难题,并为之后
的种质资源重测序、重要新基因的发掘以及资源创新
及开发利用奠定了基础。 变异组学(variome)一词首先出现在人类研究中[8]。
将其引用到作物种质资源研究中,泛指在全基因组水
平研究种质资源变异的种类、分布与功能。变异组学
是连接基因组学与育种学的桥梁。变异组学研究的
最重要进展就是相继建立了玉米[9-11]、水稻[12-14]、
高粱[15]、谷子[16]、大豆[17]、黄瓜[18]、西红柿[19]等作物
的变异组图谱(即单倍型图谱,hapmap)。变异组图
谱的建立为在全基因组水平进行作物的遗传多样性、
起源演化、基因发掘以及作物育种学奠定了基础,推
动作物种质研究进入一个新的发展时期。遗传多样性
是变异组研究的重要内容。各类作物种质资源的多样
性研究发现:(1)多样性在物种间的分布极不平衡,
异花授粉作物的多样性远高于自花授粉作物;(2)多
样性在物种内的分布极不平衡,野生种的多样性远高
于栽培种;(3)多样性在基因组内的分布极不平衡,
染色体两端的多样性远大于着丝点附近的多样性。起
源演化与分类是变异组研究的另一个重要内容,也是
此前研究中存在争议较多的一个研究领域。变异组学
首次在全基因组水平对资源进行了起源演化与分类研
究,为作物的起源演化及分类提供了可靠的分子证据,
取得了前所未有的巨大进展。例如,大豆变异组研究
将野生大豆驯化的时间提前了 2 500 余年[5];通过研究
水稻基因组变异,Huang 等[12]对水稻的起源提出了新
的见解。随着研究的深入,可以预计,许多历史上争
论了上百年的学术问题将得以解决。特别值得注意的
是,变异组学揭示了作物驯化与品种改良中的基因组
变异,将为今后有计划地进行品种改良与品种设计奠
定基础。

17 期 贾继增等:作物基因组学与作物科学革命 3321
表 1 已完成或部分完成基因组测序的作物
Table 1 Genome sequencing progress of crops
物种 Species
分类 Taxon
基因组大小 Genome size (bp)
发表时间 Publish date
完成国别/组织 Country/Org
参考文献*
Reference水稻 Oryza sativa. ssp. indica 禾本科、稻属 Gramineae, Oryza 466M 2002.04 中国 China [1]
水稻 Oryza sativa. ssp. japonica 禾本科、稻属 Gramineae, Oryza 466M 2002.04 美国 America [2]
野生稻 Oryza nivara 禾本科、稻属 Gramineae, Oryza 2014.08 中国 China [3]
短花野生稻 Oryza brachyantha 禾本科、稻属 Gramineae, Oryza 342—362M 2013.03 中国 China [4]
非洲野生稻 Oryza glaberrima 禾本科、稻属 Gramineae, Oryza 316M 2014.08 美国 America [5]
普通小麦 Triticum aestivum 禾本科、小麦属 Gramineae, Triticum 17G 2012.01 英国 England [6]
普通小麦 Triticum aestivum 禾本科、小麦属 Gramineae, Triticum 17G 2014.07 德国 Germany [7]
普通小麦 Triticum aestivum 禾本科、小麦属 Gramineae, Triticum 17G 2015.02 美国 America [8]
乌拉尔图小麦 Triticum urartu 禾本科、小麦属 Gramineae, Triticum 4.94G 2013.03 中国 China [9]
粗山羊草 Aegilops tauschii 禾本科、粗山羊草属 Gramineae, Aegilops 4.36G 2013.03 中国 China [10]
玉米 Zea mays ssp. mays 禾本科、玉蜀黍属 Gramineae, Zea 2300M 2009.11 美国 America [11]
高粱 Sorghum bicolor 禾本科、高粱属 Gramineae, Sorghum 730M 2009.01 美国 America [12]
谷子 Setaria italica 禾本科、狗尾草属 Gramineae, Setaria 510M 2012.05 美国 America [13]
谷子 Setaria italica 禾本科、狗尾草属 Gramineae, Setaria 490M 2012.05 中国 China [14]
二穗短柄草 Brachypodium distachyon
禾本科、短柄草属 Gramineae, Brachypodium 260M 2010.02 美国 America [15]
大麦 Hordeum vulgare 禾本科、大麦属 Gramineae, Hordeum 5.1G 2012.01 德国 Germany [16]
青稞 Hordeum vulgare 禾本科、大麦属 Gramineae, Hordeum 5.1G 2015.01 中国 China [17]
雷蒙德氏棉 Gossypium raimondii 锦葵科、棉属 Malvaceae, Gossypium 775.2M 2012.08 中国 China [18]
雷蒙德氏棉 Gossypium raimondii 锦葵科、棉属 Malvaceae, Gossypium 761.4M 2012.12 美国 America [19]
亚洲棉 Gossypium arboreum 锦葵科、棉属 Malvaceae, Gossypium 1694M 2014.05 中国 China [20]
大豆 Glycine max 豆科、大豆属 Leguminosae, Glycine 1100M 2010.01 美国 America [21]
野生大豆 Glycine soja 豆科、大豆属 Leguminosae, Glycine 868M 2014.07 中国 China [22]
野生大豆 Glycine soja 豆科、大豆属 Leguminosae, Glycine
889.33— 1118.34M
2014.09 中国 China [23]
野生大豆 Glycine soja 豆科、大豆属 Leguminosae, Glycine 915.4M 2010.12 韩国 Korea [24]
菜豆 Phaseolus vulgaris L. 豆科、菜豆属 Leguminosae, Phaseolus 587M 2014.06 美国 America [25]
木豆 Cajanus cajan 豆科、木豆属 Leguminosae, Cajanus
833M 2011.11 国际半干旱热带作物
研究所 ICRISAT [26]
绿豆 Vigna radiata 豆科、豇豆属 Leguminosae, Vigna 543M 2014.01 韩国 Korea [27]
鹰嘴豆 Cicer arietinum 豆科、鹰嘴豆属 Leguminosae, Cicer
738M 2013.01 国际半干旱热带作物
研究所 ICRISAT [28]
油菜 Brassica napus 十字花科、芸薹属 Brassicaceae, Brassica 630M 2014.08 法国 France [29]
蓖麻 Ricinus communis 大戟科、蓖麻属 Euphorbiaceae, Ricinus 350M 2010.08 美国 America [30]
芝麻 Sesamum indicum 胡麻科、胡麻属 Pedaliaceae, Sesamum 274M 2014.02 中国 China [31]
白菜 Brassica rapa 十字花科、芸薹属 Brassicaceae, Brassica 485M 2011.08 中国 China [32]

3322 中 国 农 业 科 学 48 卷
续表 1 Continued table 1
物种 Species
分类 Taxon
基因组大小 Genome size (bp)
发表时间 Publish date
完成国别/组织 Country/Org
参考文献*
Reference甘蓝 Brassica oleracea 十字花科、芸薹属 Brassicaceae, Brassica 630M 2014.05 中国 China [33]
萝卜 Raphanus sativus L 十字花科、萝卜属 Brassicaceae, Raphanus 402M 2014.05 日本 Japan [34]
黄瓜 Cucumis sativus 葫芦科、黄瓜属 Cucurbitaceae, Cucumis 350M 2009.11 中国 China [35]
马铃薯 Solanum tuberosum 茄科、茄属 Solanaceae, Solanum 844M 2011.07 中国 China [36]
番茄 Solanum lycopersicum 茄科、茄属 Solanaceae, Solanum 900M 2012.05 日本 Japan [37]
野生番茄 Solanum pennellii 茄科、茄属 Solanaceae, Solanum 942M 2014.07 德国 Germany [38]
甜菜 Beta vulgaris ssp. vulgaris 藜科、甜菜属 Chenopodiaceae, Beta vulgaris 566.6M 2013.12 西班牙 Spain [39]
辣椒 Capsicum annuum 茄科、辣椒属 Solanaceae, Capsicum 3.48G 2014.01 韩国 Korea [40]
辣椒 Capsicum annuum 茄科、辣椒属 Solanaceae, Capsicum 3.48G 2014.03 中国 China [41]
西瓜 Citrullus lanatus 葫芦科、西瓜属 Cucurbitaceae, Citrullus 425M 2012.11 中国 China [42]
茄子 Solanum melongena 茄科、茄属 Solanaceae, Solanum 1093M 2014.09 日本 Japan [43]
苹果 Malus × domestica 蔷薇科、苹果属 Rosaceae, Malus 742M 2010.09 意大利 Italy [44]
梨 Pyrus bretschneideri 蔷薇科、梨属 Rosaceae, Pyrus 527M 2012.11 中国 China [45]
葡萄 Vitis vinifera 葡萄科、葡萄属 Vitaceae, Vitis 490M 2007.09 法国 France [46]
番木瓜 Carica papaya 番木瓜科、番木瓜属 Caricaceae, Carica 370M 2008.04 美国 America [47]
甜橙 Citrus sinensis 芸香科、柑橘属 Rutaceae, Citrus 367M 2012.11 中国 China [48]
猕猴桃 Actinidia chinensis 猕猴桃科、猕猴桃属 Actinidiaceae, Actinidia 616.1M 2013.01 中国 China [49]
森林树莓 Fragaria vesca 蔷薇科、草莓属 Rosaceae, Fragaria 240M 2010.12 美国 America [50]
八倍体草莓 Fragaria. xananassa 蔷薇科、草莓属 Rosaceae, Fragaria 698M 2013.12 日本 Japan [51]
甜瓜 Cucumis melo 葫芦科、甜瓜属 Cucurbitaceae, Cucumis 450M 2012.07 西班牙 Spain [52]
枣树 Ziziphus jujuba Mill 鼠李科、枣属 Rhamnaceae, Ziziphus 437.65M 2014.10 中国 China [53]
枣椰树 Phoenix dactylifera 棕榈科、刺葵属 Arecaceae, Palmae 685M 2011.05 卡塔尔 Qatar [54]
枣椰树 Phoenix dactylifera 棕榈科、刺葵属 Arecaceae, Palmae 671M 2013.08 沙特阿拉伯 Saudi Arabia
[55]
桃树 Prunus persica 蔷薇科、梨属 Rosaceae, Pyrus 265M 2013.05 意大利 Italy [56]
香蕉 Musa acuminata 芭蕉科、芭蕉属 Musaceae, Musa 523M 2012.07 法国 France [57]
油棕榈 Elaeis guineensis 棕榈科、油棕榈属 Arecaceae, Elaeis 1.8G 2013.07 马来西亚 Malaysia [58]
莲藕 Nelumbo nucifera 睡莲科、莲属 Nymphaeaceae, Nelumbo 879M 2013.08 中国 China [59]
毛竹 Phyllostachys heterocycla 禾本科、刚竹属 Gramineae, Phyllostachys 2.075G 2013.02 中国 China [60]
中果咖啡 Coffea canephora 茜草科、咖啡属 Rubiaceae, Coffea 710M 2014.09 法国 France [61]
啤酒花 Humulus lupulus 大麻科、葎草属 Cannabaceae, Humulus 2.57G 2014.11 日本 Japan [62]
桑树 Morus notabilis 桑科、桑属 Moraceae, Morus 357M 2013.09 中国 China [63]
中国莲 Nelumbo nucifera Gaertn 睡莲科、莲属 Nymphaeaceae, Nelumbo 929M 2013.05 中国 China [64]
橡胶树 Hevea brasiliensis 大戟科、橡胶树属 Euphorbiaceae, Hevea 2.15G 2013.02 马来西亚 Malaysia [65]
可可树 Theobroma cacao 梧桐科、可可属 Sterculiaceae, Theobroma 430M 2010.12 法国 France [66]
*表 1 参考文献见附表 1 *References in table 1 are listed in Supplementary file 1
17 期 贾继增等:作物基因组学与作物科学革命 3323
表 2 作物核心种质构建情况
Table 2 Construction of crop core collections
物种 Species
核心种质 Core collections
微核心种质 Mini core collections
国别 Country
参考文献* Reference
大麦 Hordeum vulgare 2574 186 美国 America [1]
大麦 Hordeum vulgare 1126 欧洲 Europe [2]
大麦 Hordeum vulgare 160 西班牙 Spain [3]
水稻 Oryza sativa 932 189 中国 China [4]
水稻 Oryza sativa 1794 217 美国 America [5-6]
水稻 Oryza sativa 550 242 巴西 Brazil [7-8]
水稻 Oryza sativa 150(Ting's rice) 中国 China [9]
水稻 Oryza sativa 50 日本 Japan [10]
小麦 Triticum aestivum 1160 262 中国 China [11]
小麦 Triticum aestivum 372 法国 France [12]
玉米 Zea mays 1193 288 中国 China [13-14]
玉米 Zea mays 720(地方品种) 乌拉圭 Uruguay [15]
高粱 Sorghum bicolor 2247 242 国际半干旱热带作物研究所 ICRISAT [16]
谷子 Setaria italica 662 80 国际半干旱热带作物研究所 ICRISAT [17]
大豆 Glycine max 943 288 中国 China [18]
大豆 Glycine max 1600 美国 America [19]
大豆 Glycine max 96 日本 Japan [20]
大豆 Glycine max 652(野生大豆) 中国 China [21]
大豆 Glycine max 260(地方品种) 韩国 Korea [22]
花生 Arachis hypogaea 831 112 美国 America [23-24]
花生 Arachis hypogaea 576 298 中国 China [25-26]
花生 Arachis hypogaea 1704 184 国际半干旱热带作物研究所 ICRISAT [27-28]
橄榄 Canavium album 36 WOGB [29]
棉花 Gossypium hirsutum 27 (海岛棉) 中国 China [30]
棉花 Gossypium hirsutum 60 (海岛棉) 中国 China [31]
亚麻 Linum perenne 407 加拿大 Canada [32]
油菜 Brassica campestris 168 [33]
油菜 Brassica campestris 239 中国 China [34]
芝麻 Sesamum indicum 475 韩国 Korea [35]
芝麻 Sesamum indicum 452 中国 China [36]
芝麻 Sesamum indicum 362 印度 India [37]
蚕豆 Vicia faba L. 1075 192 中国 China [38]
木豆 Cajanus 1290 146 国际半干旱热带作物研究所 ICRISAT [39-40]
鹰嘴豆 Cicer arietinum Linn. 1956 211 国际半干旱热带作物研究所 ICRISAT [41-42]
豌豆 Pisum sativum 285 美国 America [43]
秘鲁蓟 Peruvian quinoa 103 秘鲁 Peru [44]
茄子 Solanum melongena 190 亚蔬-世界蔬菜中心 The World Vegetable Center [45]
*表 2 参考文献见附表 2 *References in Table 2 are listed in Supplementary file 2

3324 中 国 农 业 科 学 48 卷
变异组学研究推动了作物种质资源的开发与利
用。研究发现,驯化与品种改良不仅对目标基因进行
了正向选择,同时由于瓶颈效应也造成多样性的严重
丢失,其中包括大量优良基因的丢失,因此,在野生
种质资源中存在着大量的、尚未利用的优良变异。重
新找回这些千百年前被丢失了的优良基因,是现代作
物育种取得突破的重要途径[20]。变异组学正在促进该
研究领域的进展。四川农业科学院与四川农业大学利
用人工合成小麦为亲本率先培育大面积推广的小麦品
种[21],在区试中较对照增产 10%以上,这在多年来的
小麦区试中是极为罕见的。英国、澳大利亚等国有望
培育出比目前推广品种增产 30%以上的“超级”小麦。
水稻[22]、小麦[23]、西红柿[24]等作物的研究已经证明,
通过培育栽培种特别是主栽品种背景下的野生种导入
系,是寻找并利用这些丢失优良基因的有效措施。 在作物基因组测序的促进下,建立起了基于连锁
分析、关联分析、比较基因组学、基因表达等一系列
的克隆基因的新方法,极大地促进了作物重要农艺性
状新基因发掘与功能研究。一批水稻、玉米、小麦等
重要粮食作物的产量性状相关基因(穗数、穗粒数、
千粒重等)、发育相关基因(抽穗期、光周期、春化
等)、抗逆相关基因(抗旱、抗寒、抗盐)、抗病相
关基因已经被克隆。作为禾谷类作物模式材料,水稻
基因组研究一直处于领先地位,克隆了一大批与产量
及产量要素基因[25],在此基础上,笔者收集了近年来
通过图位克隆获得的基因(表 3),如调控穗数的基
因 MOC1、D3、D10、D14、D27、D53、OsSPL14 和
TAD1,调控籽粒发育的基因 LAX1、SPA、Gn1a、OsCKX2、SP1 和 DEP1 等,调控籽粒大小的基因 GS3、GS5、GW2、GW5、qSW5 和 GIF1 等,调控抽穗及开
花的基因 DTH2、DTH3、DTH7、DTH8、Ehd2、Ehd3、Ehd4、Ghd7、Ghd8、Hd1、Hd3a、Hd6、Hd16 和 Hd17等,调控株型、叶型和根型的基因 EP2、IPA1、DWT1、PROG1、Dwarf1、ds-1、OsEBS、NAL1 和 DRO1 等,
调控抗病抗逆的基因 Xa23、Xa26、Pib、Pi37、SKC1、LTG1、COLD1、qUVR-10 和 Sub1A 等。玉米是继水
稻之后完成全基因组测序测定的第二个主要粮食作
物,但由于基因组复杂,基因组组装不太好,因而图
位克隆进展较慢。2011 年以来,已经陆续有 7 个调控
玉米产量、形态建成与抗病的基因通过图位克隆的方
法获得(表 3)。小麦的基因组更大,因而图位克隆
的进展也较慢。已经克隆的基因包括驯化基因 Q,抗
病基因 Sr33、Sr35、Lr1、Lr10、Lr21、Lr34、Yr36、
Pm3b 和 Pm6,春化基因 Vrn1、Vrn2 和 Vrn3。小麦全
基因组测序近年来有较大的进展,将对小麦基因克隆
与功能解析产生重要的促进作用。除主要粮食作物外,
园艺基因组学家克隆了园艺和油料作物特殊的基因,
如瓜类性别决定基因(CmACS-7)、苦味基因(Bi 和Bt)[18, 26]、马铃薯块茎形成相关基因(StCDF1)。由
于中国的结构基因组研究基础较好,自然变异与人工
突变资源丰富,无论是新基因发掘的数量与质量都走
在世界的前列,许多重要的基因都是首先由中国的科
学家克隆的(尤其是水稻)。揭示重要农艺性状基因
的功能与作用机理、发掘其优异等位基因将为分子育
种及其他相关研究奠定基础。 目前变异组研究主要集中在基因组学,尚有一些
作物没有开展,或研究不够深入。今后,变异组将会
向表型变异组、转录变异组、代谢变异组与蛋白变异
组的方向发展。不同层次变异组的结合对作物科学所
产生的巨大作用将很快显现。 3.2 基因组学对育种学的促进——建立育种基因组
学
作物常规育种为作物改良作出了巨大的贡献。迄
今为止,生产上的品种基本上都是由常规育种培育
的。但是,作物育种的基本理论还是百余年前的孟德
尔-摩尔根遗传学理论,上百年来没有大的突破。作
物育种在某种程度上还是基于经验的一门艺术。育种
的成功往往取决于经验与一定程度的机遇,“效率低、
周期长、预见性差”是作物育种长期存在的“老大难”
问题。全世界近 30 余年的实践已经证明,仅靠常规
育种难以取得大的突破。基因组学将在育种材料、理
论与方法上取得重大进展,从而促进作物育种学这门
古老科学取得重大突破,实现由经验到理论的质的飞
跃,建立起基于基因组学理论与技术的育种。在基因
组学的促进下,有关作物育种的理论取得了如下重要
进展:(1)建立了不同作物的重组率图谱(图 3),
明确了不同作物的重组规律及影响重组率的主要因
素,为育种后代的群体设计奠定了理论基础;(2)建
立了主要作物的单倍型图谱(hapmap),发现单倍型、
重组率与多样性三者存在较密切的关系;(3)在全基
因组水平明确了作物驯化与品种改良的主要基因(位
点)以及主栽品种与骨干亲本的基因组组成与遗传多
样性,为未来的基因组设计育种奠定基础;(4)发现
了非孟德尔遗传、表观遗传学及其机理,明确了米丘
林遗传学的基因组学基础,从而解决或部分解决了持
续上百年的孟德尔与米丘林遗传学派的争论。

17 期 贾继增等:作物基因组学与作物科学革命 3325
表 3 图位克隆法获得的作物重要性状基因
Table 3 Genes conferring agronomic important traits cloned by map-based cloning method in crops
基因 Genes 物种 Species 性状 Traits 文献* References
Dwarf 3 水稻 Oryza sativa 株高、分蘖 Plant height, tiller [1]
Dwarf 10 水稻 Oryza sativa 枝梗 Branch [2]
Dwarf 14 水稻 Oryza sativa 枝梗 Branch [3]
Dwarf 27 水稻 Oryza sativa 枝梗 Branch [4]
Dwarf 53 水稻 Oryza sativa 枝梗 Branch [5-6]
Htd1 水稻 Oryza sativa 株高、分蘖 Plant height, tiller [7]
LAX 水稻 Oryza sativa 枝梗 Branch [8]
MOC1 水稻 Oryza sativa 分蘖 Tiller [9]
SPA 水稻 Oryza sativa 枝梗 Branch [8]
TAD1 水稻 Oryza sativa 分蘖 Tiller [10]
OsSPL14 水稻 Oryza sativa 枝梗、产量 Branch, yield [11-12]
DTH2 水稻 Oryza sativa 开花期 Flowering stage [13]
DTH3 水稻 Oryza sativa 开花期 Flowering stage [14]
DTH7 水稻 Oryza sativa 开花期 Flowering stage [15]
DTH8 水稻 Oryza sativa 开花期、株高、产量 Flowering stage, plant height, yield [16]
Ehd2 水稻 Oryza sativa 开花期 Flowering stage [17]
Ehd3 水稻 Oryza sativa 开花期 Flowering stage [18]
Ehd4 水稻 Oryza sativa 开花期 Flowering stage [19]
Ghd7 水稻 Oryza sativa 开花期、株高、产量 Flowering stage, plant height, yield [20]
Ghd8 水稻 Oryza sativa 开花期、株高、产量 Flowering stage, plant height, yield [21]
Hd1 水稻 Oryza sativa 开花期 Flowering stage [22]
Hd3a 水稻 Oryza sativa 开花期 Flowering stage [23]
Hd6 水稻 Oryza sativa 开花期 Flowering stage [24]
Hd16 水稻 Oryza sativa 开花期 Flowering stage [25]
Hd17 水稻 Oryza sativa 开花期 Flowering stage [26]
qSH1 水稻 Oryza sativa 落粒性 Seed shattering [27]
Sdr4 水稻 Oryza sativa 种子休眠 Seed dormancy [28]
sh4 水稻 Oryza sativa 落粒性 Seed shattering [29]
Waxy 水稻 Oryza sativa 籽粒糯性 Grains of waxy [30]
BADH2 水稻 Oryza sativa 籽粒香味 Grain aroma [31]
OsCKX2/Gn1a 水稻 Oryza sativa 产量 Yield [32]
DEP1 水稻 Oryza sativa 产量 Yield [33]
GE 水稻 Oryza sativa 种子发育 Seed development [34]
LOG 水稻 Oryza sativa 茎分生组织活性 Stem meristem activity [35]
GIF1 水稻 Oryza sativa 籽粒灌浆 Grain filling [36]
GS3 水稻 Oryza sativa 籽粒大小 Seed size [37]
GS5 水稻 Oryza sativa 籽粒大小 Seed size [38]

3326 中 国 农 业 科 学 48 卷
续表 3 Continued table 3
基因 Genes 物种 Species 性状 Traits 文献* References
GW2 水稻 Oryza sativa 粒宽 Grain width [39]
GW5 水稻 Oryza sativa 粒宽、粒重 Grain width, grain weight [40]
GW8 水稻 Oryza sativa 籽粒大小和形状 Seed size and shape [41]
qSW5 水稻 Oryza sativa 籽粒大小 Seed size [42]
Sp1 水稻 Oryza sativa 穗发育 Spike and floret development [43]
EP2 水稻 Oryza sativa 穗直立性 Panicle erect [44]
Dwarf 1 水稻 Oryza sativa 株高 Plant height [45]
sd-1 水稻 Oryza sativa 株高 Plant height [46]
OsEBS 水稻 Oryza sativa 株高、叶片大小、小穗数 Plant height, leaf size, number of spikelets [47]
IPA1 水稻 Oryza sativa 株型 Plant architecture [11]
DWT1 水稻 Oryza sativa 主茎分蘖发育一致性 Main stem tiller development consistency [48]
PROG1 水稻 Oryza sativa 株型 Plant architecture [49-50]
NAL1 水稻 Oryza sativa 叶型、产量 Leaf shape, yield [51-52]
DRO1 水稻 Oryza sativa 根系结构 Root structure [53]
Phr1 水稻 Oryza sativa 种子保存褪色性 Grain discoloration [54-55]
Rc 水稻 Oryza sativa 籽粒颜色 Grain color [56-57]
Xa23 水稻 Oryza sativa 抗病 Disease resistance [58]
Xa26 水稻 Oryza sativa 抗病 Disease resistance [59]
Pi37 水稻 Oryza sativa 抗病 Disease resistance [60]
Pib 水稻 Oryza sativa 抗病 Disease resistance [61]
SKC1 水稻 Oryza sativa 抗逆 Stress tolerance [62]
LTG1 水稻 Oryza sativa 抗逆 Stress tolerance [63]
qUVR-10 水稻 Oryza sativa 抗逆 Stress tolerance [64]
Sub1A 水稻 Oryza sativa 抗逆 Stress tolerance [65]
COLD1 水稻 Oryza sativa 抗逆 Stress tolerance [66]
OsAM1 水稻 Oryza sativa 细胞减数分裂 Meiosis [67]
Sr33 小麦 Triticum aestivum 抗病 Disease resistance [68]
Sr35 小麦 Triticum aestivum 抗病 Disease resistance [69]
Lr21 小麦 Triticum aestivum 抗病 Disease resistance [70]
Lr10 小麦 Triticum aestivum 抗病 Disease resistance [71]
Lr1 小麦 Triticum aestivum 抗病 Disease resistance [72]
Lr34 小麦 Triticum aestivum 抗病 Disease resistance [73]
Yr36 小麦 Triticum aestivum 抗病 Disease resistance [74]
Pm3b 小麦 Triticum aestivum 抗病 Disease resistance [75]
Pm6 小麦 Triticum aestivum 抗病 Disease resistance [76]
Rht1 小麦 Triticum aestivum 株高 Plant height [77]
Vrn1 小麦 Triticum aestivum 春化 Vernalization [78]
Vrn2 小麦 Triticum aestivum 春化 Vernalization [79]
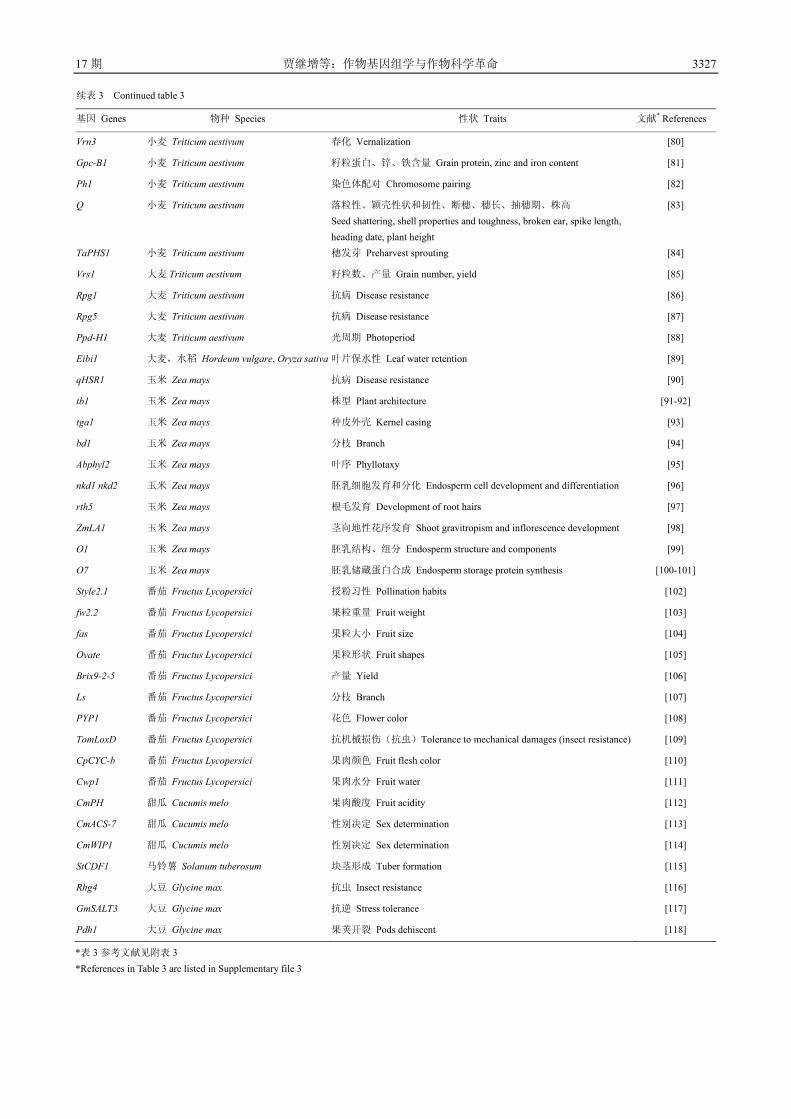
17 期 贾继增等:作物基因组学与作物科学革命 3327
续表 3 Continued table 3
基因 Genes 物种 Species 性状 Traits 文献* References
Vrn3 小麦 Triticum aestivum 春化 Vernalization [80]
Gpc-B1 小麦 Triticum aestivum 籽粒蛋白、锌、铁含量 Grain protein, zinc and iron content [81]
Ph1 小麦 Triticum aestivum 染色体配对 Chromosome pairing [82]
Q 小麦 Triticum aestivum 落粒性、颖壳性状和韧性、断穗、穗长、抽穗期、株高 Seed shattering, shell properties and toughness, broken ear, spike length, heading date, plant height
[83]
TaPHS1 小麦 Triticum aestivum 穗发芽 Preharvest sprouting [84]
Vrs1 大麦 Triticum aestivum 籽粒数、产量 Grain number, yield [85]
Rpg1 大麦 Triticum aestivum 抗病 Disease resistance [86]
Rpg5 大麦 Triticum aestivum 抗病 Disease resistance [87]
Ppd-H1 大麦 Triticum aestivum 光周期 Photoperiod [88]
Eibi1 大麦,水稻 Hordeum vulgare, Oryza sativa 叶片保水性 Leaf water retention [89]
qHSR1 玉米 Zea mays 抗病 Disease resistance [90]
tb1 玉米 Zea mays 株型 Plant architecture [91-92]
tga1 玉米 Zea mays 种皮外壳 Kernel casing [93]
bd1 玉米 Zea mays 分枝 Branch [94]
Abphyl2 玉米 Zea mays 叶序 Phyllotaxy [95]
nkd1 nkd2 玉米 Zea mays 胚乳细胞发育和分化 Endosperm cell development and differentiation [96]
rth5 玉米 Zea mays 根毛发育 Development of root hairs [97]
ZmLA1 玉米 Zea mays 茎向地性花序发育 Shoot gravitropism and inflorescence development [98]
O1 玉米 Zea mays 胚乳结构、组分 Endosperm structure and components [99]
O7 玉米 Zea mays 胚乳储藏蛋白合成 Endosperm storage protein synthesis [100-101]
Style2.1 番茄 Fructus Lycopersici 授粉习性 Pollination habits [102]
fw2.2 番茄 Fructus Lycopersici 果粒重量 Fruit weight [103]
fas 番茄 Fructus Lycopersici 果粒大小 Fruit size [104]
Ovate 番茄 Fructus Lycopersici 果粒形状 Fruit shapes [105]
Brix9-2-5 番茄 Fructus Lycopersici 产量 Yield [106]
Ls 番茄 Fructus Lycopersici 分枝 Branch [107]
PYP1 番茄 Fructus Lycopersici 花色 Flower color [108]
TomLoxD 番茄 Fructus Lycopersici 抗机械损伤(抗虫)Tolerance to mechanical damages (insect resistance) [109]
CpCYC-b 番茄 Fructus Lycopersici 果肉颜色 Fruit flesh color [110]
Cwp1 番茄 Fructus Lycopersici 果肉水分 Fruit water [111]
CmPH 甜瓜 Cucumis melo 果肉酸度 Fruit acidity [112]
CmACS-7 甜瓜 Cucumis melo 性别决定 Sex determination [113]
CmWIP1 甜瓜 Cucumis melo 性别决定 Sex determination [114]
StCDF1 马铃薯 Solanum tuberosum 块茎形成 Tuber formation [115]
Rhg4 大豆 Glycine max 抗虫 Insect resistance [116]
GmSALT3 大豆 Glycine max 抗逆 Stress tolerance [117]
Pdh1 大豆 Glycine max 果荚开裂 Pods dehiscent [118]
*表 3 参考文献见附表 3 *References in Table 3 are listed in Supplementary file 3

3328 中 国 农 业 科 学 48 卷
0
50
100
150
200
0
50
100
150
200
0
50
100
150
200
0
50
100
150
200
0
50
100
150
200
0
50
100
150
200
0 50 100 150 200 250 0 50 100 150 200 250 0 50 100 150 200 250
0 50 100 150 200 250 0 50 100 150 200 250 0 50 100 150 200 250
物理距离 Physical distances
遗传
距离
Gen
etic
dis
tanc
es (c
M)
物理距离 Physical distances 物理距离 Physical distances
遗传
距离
Gen
etic
dis
tanc
es (c
M)
遗传
距离
Gen
etic
dis
tanc
es (c
M)
遗传
距离
Gen
etic
dis
tanc
es (c
M)
遗传
距离
Gen
etic
dis
tanc
es (c
M)
遗传
距离
Gen
etic
dis
tanc
es (c
M)
物理距离 Physical distances 物理距离 Physical distances 物理距离 Physical distances
1D 2D 3D
4D 5D 6D
0
50
100
150
200
0 50 100 150 200 250
遗传距离
Gen
etic
dis
tanc
es (c
M)
物理距离 Physical distances
7D
图 3 粗山羊草基因组重组率图谱
Fig. 3 Recombination map of Aegilops tauschii
作物基因组学正在促进作物育种方法取得重要进
展。长期以来,作物育种都是依据表型进行选择,由
于表型易受环境条件的影响,因此不仅影响选择的效
率,而且影响加代效率。基因组研究发现许多目标性
状基因的功能标记或与目标性状基因紧密连锁的分子
标记,利用这些标记可以不受环境条件影响,对目标
性状进行直接选择,这就是标记辅助选择。近年来,
在重测序的基础上,各种作物都开发了高通量的 SNP芯片。SNP 是继第一代 RFLP 分子标记与第二代 SSR标记之后的第三代分子标记,具有数量多、效率高的
特点。SNP 芯片既可用于新基因发掘、多样性检测、
单倍型图谱绘制,也可直接用于基因组选择育种。目
前,水稻、玉米等作物已经研制出了基因组育种 SNP芯片[27-29]。中国已经研制出了有 63 万个位点的小麦
SNP 芯片 Wheat660,小麦育种芯片正在研制之中。随
着越来越多的育种目标基因的发现与育种规律的揭
示,未来的基因组育种将对作物改良产生巨大的甚至
是革命性的推动作用。 3.3 基因组学正在推动作物栽培学取得重大突破—
建立栽培基因组学
作物产量、品质与水肥利用效率均是其基因表达
的结果。作物育种是通过对基因的选择与重组实现目

17 期 贾继增等:作物基因组学与作物科学革命 3329
标性状基因的最佳表达,而作物栽培是通过对环境的
调控,来实现作物重要农艺性状基因的最佳表达。二
者异曲同工,且作用可相互弥补。作物栽培学家在过
去近百年的实践中积累了丰富的经验,建立了比较系
统全面的研究体系和技术模式,对作物生产作出了重
大贡献。但长期以来,作物栽培研究主要侧重在表型
水平,有关栽培措施对基因表达的影响基本不了解,
基础理论研究严重不足,已经不能满足现代作物生产
的发展需求。栽培基因组学是通过研究环境条件对重
要农艺性状基因表达的影响规律,提出合理的栽培措
施和实时的生长调控技术体系,实现目标基因的最佳
表达。近年来,由于基因组学的飞速发展,在与作物
栽培有关的诸方面取得了重要进展,发现了大量的与
环境因素有关的 QTL,克隆了一批重要的环境相关的
基因,如盐胁迫[30-31]、干旱胁迫[32-34]、温度胁迫[35]以
及营养胁迫如氮胁迫[36-37]、磷胁迫[38-39]、钾胁迫[40]等。
实际上,大部分基因的表达都是受光、温、肥、水、
热、气等环境条件影响。人们在进行多环境 QTL 分
析时,常常发现在不同条件下出现的 QTL,而这些
QTL 很可能就是环境条件影响基因表达的结果。这
些基因的克隆将帮助栽培工作者根据基因表达情况
来制定最佳栽培措施。反过来,在田间环境下从转
录组、蛋白组、代谢组等组学层面上研究不同农艺
措施的调控效果,可以更为全面、系统、真实地阐
明基因与重要农艺性状的关系,为现代作物分子生
物学研究提供新的研究方向。总之,栽培生理学通
过与基因组学的融合,必将推进栽培基因组学达到全
新的水平。 3.4 基因组学促进“小”作物的“飞跃”
据统计,目前人类种植的各类作物共计 1 500 余
种,其中,中国栽培的有 600 余种。就数量而言,水
稻、小麦、玉米、大豆、棉花等种植面积很大的“大”
作物仅占极少数,而谷子、高粱、食用豆及各类蔬菜
等种植面积较小或很小的“小”作物则占绝大多数。
这些小作物之所以能够经历数千年的自然选择与人工
选择而不被淘汰,说明在这些小作物中还蕴藏着丰富
的而且为大作物没有的重要基因,例如,谷子的抗旱
性与营养品质突出,其中很可能存在与抗旱及品质相
关的基因;木薯、马铃薯与甘薯等薯类作物高光效、
水分利用高效与 N、P 利用高效,其中应该有与上述
性状相关的基因。因此,这些小作物也是“特色”作
物。然而由于研究力量、投入、历史等原因,小宗作
物的研究远远落后于五大作物,制约了这些作物的发
展。基因组学大大促进了这些作物研究水平。由于许
多小宗作物的基因组很小,方便进行基因组测序与重
测序,目前谷子、黄瓜等小宗作物已完成了基因组测
序与变异组分析,跨越了大作物花费了近百年走过的
表型遗传、细胞遗传、数量遗传历程,直接跨入基因
组学时代,赶上了一些大作物的研究水平。这些作物
的基因组与变异组研究不仅促进其自身的发展,而且
通过比较基因组学与比较变异组学,对于大作物的发
展也将产生重要的促进作用。这些作物携带的重要功
能基因及其分子机制,对于作物科学的研究也将具有
普遍的指导意义。
4 目前存在的问题、解决途径与展望
在看到基因组学取得重大研究进展的同时,也应
看到当前基因组学研究存在的重要问题。基因组学研
究已进行了 20 余年,发现了上千个农艺性状的 QTL,克隆了数以百计的农艺性状基因,发表了许多高水平
的论文。然而这些结果却很少在育种上发挥作用。迄
今为止,生产上的大面积主栽品种仍然是常规育种培
育的。因此,很多人怀疑基因组学是否真的具有应用
价值。造成上述现象的原因主要有以下几方面:(1)基因组学研究队伍与育种学研究队伍没有很好地结
合,基因组学的研究成果没有有效地转化为育种资源
或育种技术;(2)基因组学研究材料与育种学研究
材料没有很好地结合,基因组学研究材料难以被育种
家所接受;(3)所发现的 QTL 或克隆的基因大多是
在育种上已经被利用,缺乏新的育种价值;(4)许
多所谓的“高产基因”实际上是产量构成因素相关基
因,并非真正的产量基因。而且这些基因不是在群体
条件下,而是在个体条件下获得的,产量构成因素间
的负相关没有暴露出来。一旦到了真正的生产条件
下,由于相互间的显著负相关而失去应用价值;(5)研究分散、重复,无序竞争,缺乏顶层设计;(6)从理论到应用需要一定的时间。可喜的是已有越来越
多的基因组工作者认识到上述问题,越来越多的常规
育种家认识到基因组育种是未来育种的发展方向。基
因组学家、育种家、资源学家与栽培学家的结合,必
将促进作物科学发展的突破与飞跃,促进作物科学进
入一个新的历史发展时期。中国应该在这个新的发展
时期对作物科学作出中国科学家应有的贡献。 在过去的 10 年作物科学在基础研究上取得了重
要的进展。今后应紧密围绕解决当前中国主要农作物
生产存在的“产量增长慢、资源消费高、生产效率低、

3330 中 国 农 业 科 学 48 卷
质量不稳定”等突出问题,综合利用基因组学、代谢
组学和表型组学与基因工程等领域的国际前沿科学技
术,以实现“更高产、更高效、更优质、更环保”为
核心的新的绿色革命为目标,重点开展主要农作物种
质资源、遗传育种与作物栽培等方面的基础研究,促
进中国作物科学取得重大突破,从而进入一个新的发
展时期,促进新的绿色革命率先在中国实现。
致谢:李家洋、朱立煌、杨建昌、陈晓亚、邓秀新、王
汉中、李付广等参与了文章的讨论并提出宝贵意见与建议,
在此表示感谢。
References
[1] http://www.fao.org/home/en/.
[2] Zhang C, Gao L, Sun J, Jia J, Ren Z. Haplotype variation of
Green Revolution gene Rht-D1 during wheat domestication and
improvement. Journal of Integrative Plant Biology, 2014, 56(8):
774-780.
[3] Paterson A H, Bowers J E, Bruggmann R, Dubchak I, Grimwood J,
Gundlach H, Haberer G, Hellsten U, Mitros T, Poliakov A, Schmutz J,
Spannagl M, Tang H, Wang X, Wicker T, Bharti A K, Chapman J,
Feltus F A, Gowik U, Grigoriev I V, Lyons E, Maher C A, Martis M,
Narechania A, Otillar R P, Penning B W, Salamov A A, Wang Y,
Zhang L, Carpita N C, Freeling M, Gingle A R, Hash C T, Keller B,
Klein P, Kresovich S, McCann M C, Ming R, Peterson D G, Mehboob
ur R, Ware D, Westhoff P, Mayer K F, Messing J, Rokhsar D S. The
Sorghum bicolor genome and the diversification of grasses. Nature,
2009, 457(7229): 551-556.
[4] Jia J, Zhao S, Kong X, Li Y, Zhao G, He W, Appels R, Pfeifer M, Tao
Y, Zhang X, Jing R, Zhang C, Ma Y, Gao L, Gao C, Spannagl M,
Mayer K F, Li D, Pan S, Zheng F, Hu Q, Xia X, Li J, Liang Q, Chen J,
Wicker T, Gou C, Kuang H, He G, Luo Y, Keller B, Xia Q, Lu P, Wang
J, Zou H, Zhang R, Xu J, Gao J, Middleton C, Quan Z, Liu G, Wang J,
Yang H, Liu X, He Z, Mao L, Wang J. Aegilops tauschii draft genome
sequence reveals a gene repertoire for wheat adaptation. Nature, 2013,
496(7443): 91-95.
[5] Li Y H, Zhou G, Ma J, Jiang W, Jin L G, Zhang Z, Guo Y, Zhang J, Sui
Y, Zheng L, Zhang S S, Zuo Q, Shi X H, Li Y F, Zhang W K, Hu Y,
Kong G, Hong H L, Tan B, Song J, Liu Z X, Wang Y, Ruan H, Yeung
C K, Liu J, Wang H, Zhang L J, Guan R X, Wang K J, Li W B, Chen S
Y, Chang R Z, Jiang Z, Jackson S A, Li R, Qiu L J. De novo assembly
of soybean wild relatives for pan-genome analysis of diversity and
agronomic traits. Nature Biotechnology, 2014, 32(10): 1045-1052.
[6] Mikkelsen T S, Hiller L W, Eichler E, Zody M C, Jaffe D B, Yang S P,
Enard W, Hellmann I, Lindblad-toh K, Altheide T K, Archidiacono N,
Bork P, Butler J, Chang J I, Cheng Z, Chinwalla A T, de Jong P,
Delehaunty K D, Fronick C, et al. Initial sequence of the chimpanzee
genome and comparison with the human genome. Nature, 2005,
437(7055): 69-87.
[7] 王述民, 张宗文. 世界粮食和农业植物遗传资源保护与利用现状.
植物遗传资源学报, 2011, 12(3): 325-338.
Wang S M, Zhang Z W. The state of the world’s plant genetic
resources for food and agriculture. Journal of Plant Genetic Resources,
2011, 12(3): 325-338. (in Chinese)
[8] Ring H, Kwok P, Cotton R. Human Variome Project: An international
collaboration to catalogue human genetic variation. Pharmacogenomics,
2006, 7(7): 969-972.
[9] Gore M A, Chia J M, Elshire R J, Sun Q, Ersoz E S, Hurwitz B L,
Peiffer J A, McMullen M D, Grills G S, Ross-Ibarra J, Ware D H,
Buckler E S. A first-generation haplotype map of maize. Science, 2009,
326(5956): 1115-1117.
[10] Chia J, Song C, Bradbury P, Costich D, de Leon N, Doebley J, Elshire
R, Gaut B, Geller L, Glaubitz J, Gore M, Guill K, Holland J, Hufford
M, Lai J, Li M, Liu X, Lu Y, McCombie R, Nelson R, Poland J,
Prasanna B, Pyhajarvi T, Rong T, Sekhon R, Sun Q, Tenaillon M, Tian
F, Wang J, Xu X, Zhang Z, Kaeppler S, Ross-Ibarra J, McMullen M,
Buckler E, Zhang G, Xu Y, Ware D. Maize HapMap2 identifies extant
variation from a genome in flux. Nature Genetics, 2012, 44(7):
803-807.
[11] Hufford M, Xu X, van Heerwaarden J, Pyhajarvi T, Chia J, Cartwright
R, Elshire R, Glaubitz J, Guill K, Kaeppler S, Lai J, Morrell P,
Shannon L, Song C, Springer N, Swanson-Wagner R, Tiffin P, Wang
J, Zhang G, Doebley J, McMullen M, Ware D, Buckler E, Yang S,
Ross-Ibarra J. Comparative population genomics of maize
domestication and improvement. Nature Genetics, 2012, 44(7):
808-811.
[12] Huang X, Kurata N, Wei X, Wang Z, Wang A, Zhao Q, Zhao Y, Liu K,
Lu H, Li W, Guo Y, Lu Y, Zhou C, Fan D, Weng Q, Zhu C, Huang T,
Zhang L, Wang Y, Feng L, Furuumi H, Kubo T, Miyabayashi T, Yuan
X, Xu Q, Dong G, Zhan Q, Li C, Fujiyama A, Toyoda A, Lu T, Feng Q,
Qian Q, Li J, Han B. A map of rice genome variation reveals the
origin of cultivated rice. Nature, 2012, 490(7421): 497-501.
[13] Huang X, Zhao Y, Wei X, Li C, Wang A, Zhao Q, Li W, Guo Y, Deng
L, Zhu C, Fan D, Lu Y, Weng Q, Liu K, Zhou T, Jing Y, Si L, Dong G,
Huang T, Lu T, Feng Q, Qian Q, Li J, Han B. Genome-wide
association study of flowering time and grain yield traits in a

17 期 贾继增等:作物基因组学与作物科学革命 3331
worldwide collection of rice germplasm. Nature Genetics, 2012, 44(1):
32-39.
[14] Xu X, Liu X, Ge S, Jensen J, Hu F, Li X, Dong Y, Gutenkunst R, Fang
L, Huang L, Li J, He W, Zhang G, Zheng X, Zhang F, Li Y, Yu C,
Kristiansen K, Zhang X, Wang J, Wright M, McCouch S, Nielsen R,
Wang J, Wang W. Resequencing 50 accessions of cultivated and wild
rice yields markers for identifying agronomically important genes.
Nature Biotechnology, 2012, 30(1): 105-111.
[15] Morris G P, Ramu P, Deshpande S P, Hash C T, Shah T, Upadhyaya H
D, Riera-Lizarazu O, Brown P J, Acharya C B, Mitchell S E,
Harriman J, Glaubitz J C, Buckler E S, Kresovich S. Population
genomic and genome-wide association studies of agroclimatic traits in
sorghum. Proceedings of the National Academy of Sciences of the
United States of America, 2013, 110(2): 453-458.
[16] Jia G, Huang X, Zhi H, Zhao Y, Zhao Q, Li W, Chai Y, Yang L, Liu K,
Lu H, Zhu C, Lu Y, Zhou C, Fan D, Weng Q, Guo Y, Huang T, Zhang
L, Lu T, Feng Q, Hao H, Liu H, Lu P, Zhang N, Li Y, Guo E, Wang S,
Wang S, Liu J, Zhang W, Chen G, Zhang B, Li W, Wang Y, Li H, Zhao
B, Li J, Diao X, Han B. A haplotype map of genomic variations and
genome-wide association studies of agronomic traits in foxtail millet
(Setaria italica). Nature Genetics, 2013, 45(8): 957-961.
[17] Lam H, Xu X, Liu X, Chen W, Yang G, Wong F, Li M, He W, Qin N,
Wang B, Li J, Jian M, Wang J, Shao G, Wang J, Sun S, Zhang G.
Resequencing of 31 wild and cultivated soybean genomes identifies
patterns of genetic diversity and selection. Nature Genetics, 2010,
42(12): 1053-1059.
[18] Qi J, Liu X, Shen D, Miao H, Xie B, Li X, Zeng P, Wang S, Shang Y,
Gu X, Du Y, Li Y, Lin T, Yuan J, Yang X, Chen J, Chen H, Xiong X,
Huang K, Fei Z, Mao L, Tian L, Stadler T, Renner S S, Kamoun S,
Lucas W J, Zhang Z, Huang S. A genomic variation map provides
insights into the genetic basis of cucumber domestication and
diversity. Nature Genetics, 2013, 45(12): 1510-1515.
[19] Lin T, Zhu G, Zhang J, Xu X, Yu Q, Zheng Z, Zhang Z, Lun Y, Li S,
Wang X, Huang Z, Li J, Zhang C, Wang T, Zhang Y, Wang A, Zhang Y,
Lin K, Li C, Xiong G, Xue Y, Mazzucato A, Causse M, Fei Z,
Giovannoni J J, Chetelat R T, Zamir D, Stadler T, Li J, Ye Z, Du Y,
Huang S. Genomic analyses provide insights into the history of
tomato breeding. Nature Genetics, 2014, 46(11): 1220-1226.
[20] Palmgren M, Edenbrandt A, Vedel S, Andersen M, Landes X,
Østerberg J, Falhof J, Olsen L, Christensen S, Sandøe P, Gamborg C,
Kappel K, Thorsen B, Pagh P. Are we ready for back-to-nature crop
breeding? Trends in Plant Science, 2015, 20(3): 155-164.
[21] 李俊, 魏会廷, 胡晓蓉, 李朝苏, 汤永禄, 刘登才, 杨武云. 川麦
42 中源于人工合成小麦的一个高产位点鉴定. 作物学报, 2011,
37(2): 255-262.
Li J, Wei H T, Hu X R, Li C S, Tang Y L, Liu D C, Yang W Y.
Identification of a high-yield introgression locus from synthetic
hexaploid wheat in Chuanmai 42. Acta Agronomica Sinica, 2011,
37(2): 255-262. (in Chinese)
[22] Yoon D B, Kang K H, Kim H J, Ju H G, Kwon S J, Suh J P, Jeong O
Y, Ahn S N. Mapping quantitative trait loci for yield components
and morphological traits in an advanced backcross population
between Oryza grandiglumis and the O. sativa japonica cultivar
Hwaseongbyeo. Theoretical and Applied Genetics, 2006, 112(6):
1052-1062.
[23] Liu S, Zhou R, Dong Y, Li P, Jia J. Development, utilization of
introgression lines using a synthetic wheat as donor. Theoretical and
Applied Genetics, 2006, 112(7): 1360-1373.
[24] Eshed Y, Zamir D. An introgression line population of Lycopersicon
pennellii in the cultivated tomato enables the identification and fine
mapping of yield-associated QTL. Genetics, 1995, 141(3): 1147-1162.
[25] Xing Y, Zhang Q. Genetic and molecular bases of rice yield. Annual
Review of Plant Biology, 2010, 61(1): 421-442.
[26] Shang Y, Ma Y, Zhou Y, Zhang H, Duan L, Chen H, Zeng J, Zhou Q,
Wang S, Gu W, Liu M, Ren J, Gu X, Zhang S, Wang Y, Yasukawa K,
Bouwmeester H J, Qi X, Zhang Z, Lucas W J, Huang S. Biosynthesis,
regulation, and domestication of bitterness in cucumber. Science, 2014,
346(6213): 1084-1088.
[27] Chen H, Xie W, He H, Yu H, Chen W, Li J, Yu R, Yao Y, Zhang W, He
Y, Tang X, Zhou F, Deng X W, Zhang Q. A high-density SNP
genotyping aArray for rice biology and molecular breeding.
Molecular Plant, 2014, 7(3): 541-553.
[28] Unterseer S, Bauer E, Haberer G, Seidel M, Knaak C, Ouzunova M,
Meitinger T, Strom T M, Fries R, Pausch H, Bertani C, Davassi A,
Mayer K F, Schon C C. A powerful tool for genome analysis in maize:
Development and evaluation of the high density 600K SNP
genotyping array. BMC Genomics, 2014, 15: 823.
[29] Yu H, Xie W, Li J, Zhou F, Zhang Q. A whole-genome SNP array
(RICE6K) for genomic breeding in rice. Plant Biotechnology Journal,
2014, 12(1): 28-37.
[30] Kumar K, Kumar M, Kim S R, Ryu H, Cho Y G. Insights into
genomics of salt stress response in rice. Rice, 2013, 6(1): 1-15.
[31] Kawaura K, Mochida K, Yamazaki Y, Ogihara Y. Transcriptome
analysis of salinity stress responses in common wheat using a 22K
oligo-DNA microarray. Functional & Integrative Genomics, 2006,
6(2): 132-142.

3332 中 国 农 业 科 学 48 卷
[32] Tuberosa R, Salvi S. Genomics-based approaches to improve drought
tolerance of crops. Trends in Plant Science, 2006, 11(8): 405-412.
[33] Wang D, Pan Y, Zhao X, Zhu L, Fu B, Li Z. Genome-wide
temporal-spatial gene expression profiling of drought responsiveness
in rice. BMC Genomics, 2011, 12(1): 149.
[34] Degenkolbe T, Do P T, Kopka J, Zuther E, Hincha D K, Köhl K I.
Identification of drought tolerance Mmarkers in a diverse population
of rice cultivars by expression and metabolite profiling. PLoS ONE,
2013, 8(5): e63637.
[35] Qin D, Wu H, Peng H, Yao Y, Ni Z, Li Z, Zhou C, Sun Q. Heat
stress-responsive transcriptome analysis in heat susceptible and
tolerant wheat (Triticum aestivum L.) by using wheat genome array.
BMC Genomics, 2008, 9(1): 432.
[36] Lian X, Wang S, Zhang J, Feng Q, Zhang L, Fan D, Li X, Yuan D,
Han B, Zhang Q. Expression Profiles of 10,422 Genes at early stage
of low nitrogen stress in rice assayed using a cDNA microarray. Plant
Molecular Biology, 2006, 60(5): 617-631.
[37] Gelli M, Duo Y, Konda A, Zhang C, Holding D, Dweikat I.
Identification of differentially expressed genes between sorghum
genotypes with contrasting nitrogen stress tolerance by genome-wide
transcriptional profiling. BMC Genomics, 2014, 15(1): 179.
[38] Li L, Liu C, Lian X. Gene expression profiles in rice roots under low
phosphorus stress. Plant Molecular Biology, 2010, 72(4/5): 423-432.
[39] Zheng L, Huang F, Narsai R, Wu J, Giraud E, He F, Cheng L, Wang F,
Wu P, Whelan J, Shou H. Physiological and transcriptome analysis of
iron and phosphorus interaction in rice seedlings. Plant Physiology,
2009, 151(1): 262-274.
[40] Ma T L, Wu W H, Wang Y. Transcriptome analysis of rice root
responses to potassium deficiency. BMC Plant Biology, 2012, 12(1):
161.
(责任编辑 李莉)

17 期 贾继增等:作物基因组学与作物科学革命
附表 1 [1] Yu J, Hu S, Wang J, Wong G K, Li S, Liu B, Deng Y, Dai L, Zhou Y,
Zhang X, Cao M, Liu J, Sun J, Tang J, Chen Y, Huang X, Lin W, Ye C,
Tong W, Cong L, Geng J, Han Y, Li L, Li W, Hu G, Huang X, Li W, Li
J, Liu Z, Li L, Liu J, Qi Q, Liu J, Li L, Li T, Wang X, Lu H, Wu T,
Zhu M, Ni P, Han H, Dong W, Ren X, Feng X, Cui P, Li X, Wang H,
Xu X, Zhai W, Xu Z, Zhang J, He S, Zhang J, Xu J, Zhang K, Zheng
X, Dong J, Zeng W, Tao L, Ye J, Tan J, Ren X, Chen X, He J, Liu D,
Tian W, Tian C, Xia H, Bao Q, Li G, Gao H, Cao T, Wang J, Zhao W,
Li P, Chen W, Wang X, Zhang Y, Hu J, Wang J, Liu S, Yang J, Zhang
G, Xiong Y, Li Z, Mao L, Zhou C, Zhu Z, Chen R, Hao B, Zheng W,
Chen S, Guo W, Li G, Liu S, Tao M, Wang J, Zhu L, Yuan L, Yang H.
A draft sequence of the rice genome (Oryza sativa L. ssp. indica).
Science, 2002, 296(5565): 79-92.
[2] Goff S A, Ricke D, Lan T H, Presting G, Wang R, Dunn M,
Glazebrook J, Sessions A, Oeller P, Varma H, Hadley D, Hutchison D,
Martin C, Katagiri F, Lange B M, Moughamer T, Xia Y, Budworth P,
Zhong J, Miguel T, Paszkowski U, Zhang S, Colbert M, Sun W L,
Chen L, Cooper B, Park S, Wood T C, Mao L, Quail P, Wing R, Dean
R, Yu Y, Zharkikh A, Shen R, Sahasrabudhe S, Thomas A, Cannings R,
Gutin A, Pruss D, Reid J, Tavtigian S, Mitchell J, Eldredge G, Scholl T,
Miller R M, Bhatnagar S, Adey N, Rubano T, Tusneem N, Robinson R,
Feldhaus J, Macalma T, Oliphant A, Briggs S. A draft sequence of the
rice genome (Oryza sativa L. ssp. japonica). Science, 2002, 296(5565):
92-100.
[3] Zhang Q J, Zhu T, Xia E H, Shi C, Liu Y L, Zhang Y, Liu Y, Jiang W
K, Zhao Y J, Mao S Y, Zhang L P, Huang H, Jiao J Y, Xu P Z, Yao Q Y,
Zeng F C, Yang L L, Gao J, Tao D Y, Wang Y J, Bennetzen J L, Gao L
Z. Rapid diversification of five Oryza AA genomes associated with
rice adaptation. Proceedings of the National Academy of Sciences of
the United States of America, 2014, 111(46): E4954- E4962.
[4] Chen J, Huang Q, Gao D, Wang J, Lang Y, Liu T, Li B, Bai Z, Luis
Goicoechea J, Liang C, Chen C, Zhang W, Sun S, Liao Y, Zhang X,
Yang L, Song C, Wang M, Shi J, Liu G, Liu J, Zhou H, Zhou W, Yu Q,
An N, Chen Y, Cai Q, Wang B, Liu B, Min J, Huang Y, Wu H, Li Z,
Zhang Y, Yin Y, Song W, Jiang J, Jackson S A, Wing R A, Wang J,
Chen M. Whole-genome sequencing of Oryza brachyantha reveals
mechanisms underlying Oryza genome evolution. Nature
Communications, 2013, 4: 1595.
[5] Wang M, Yu Y, Haberer G, Marri P R, Fan C, Goicoechea J L, Zuccolo
A, Song X, Kudrna D, Ammiraju J S, Cossu R M, Maldonado C, Chen
J, Lee S, Sisneros N, de Baynast K, Golser W, Wissotski M, Kim W,
Sanchez P, Ndjiondjop M N, Sanni K, Long M, Carney J, Panaud O,
Wicker T, Machado C A, Chen M, Mayer K F, Rounsley S, Wing R A.
The genome sequence of African rice (Oryza glaberrima) and
evidence for independent domestication. Nature Genetics, 2014, 46(9):
982-8.
[6] Brenchley R, Spannagl M, Pfeifer M, Barker G L, D'Amore R, Allen
A M, McKenzie N, Kramer M, Kerhornou A, Bolser D, Kay S, Waite
D, Trick M, Bancroft I, Gu Y, Huo N, Luo M C, Sehgal S, Gill B,
Kianian S, Anderson O, Kersey P, Dvorak J, McCombie W R, Hall A,
Mayer K F, Edwards K J, Bevan M W, Hall N. Analysis of the bread
wheat genome using whole-genome shotgun sequencing. Nature, 2012,
491(7426): 705-710.
[7] The_International_Wheat_Genome_Sequencing_Consortium. A
chromosome-based draft sequence of the hexaploid bread wheat
(Triticum aestivum) genome. Science, 2014, 345(6194): 1251788.
[8] Chapman J A, Mascher M, Buluc A N, Barry K, Georganas E, Session
A, Strnadova V, Jenkins J, Sehgal S, Oliker L, Schmutz J, Yelick K A,
Scholz U, Waugh R, Poland J A, Muehlbauer G J, Stein N, Rokhsar D
S. A whole-genome shotgun approach for assembling and anchoring
the hexaploid bread wheat genome. Genome Biology, 2015, 16(1): 26.
[9] Ling H Q, Zhao S, Liu D, Wang J, Sun H, Zhang C, Fan H, Li D,
Dong L, Tao Y, Gao C, Wu H, Li Y, Cui Y, Guo X, Zheng S, Wang B,
Yu K, Liang Q, Yang W, Lou X, Chen J, Feng M, Jian J, Zhang X,
Luo G, Jiang Y, Liu J, Wang Z, Sha Y, Zhang B, Wu H, Tang D, Shen
Q, Xue P, Zou S, Wang X, Liu X, Wang F, Yang Y, An X, Dong Z,
Zhang K, Zhang X, Luo M C, Dvorak J, Tong Y, Wang J, Yang H, Li
Z, Wang D, Zhang A, Wang J. Draft genome of the wheat A-genome
progenitor Triticum urartu. Nature, 2013, 496(7443): 87-90.
[10] Jia J, Zhao S, Kong X, Li Y, Zhao G, He W, Appels R, Pfeifer M, Tao
Y, Zhang X, Jing R, Zhang C, Ma Y, Gao L, Gao C, Spannagl M,
Mayer K F, Li D, Pan S, Zheng F, Hu Q, Xia X, Li J, Liang Q, Chen J,
Wicker T, Gou C, Kuang H, He G, Luo Y, Keller B, Xia Q, Lu P, Wang
J, Zou H, Zhang R, Xu J, Gao J, Middleton C, Quan Z, Liu G, Wang J,
Yang H, Liu X, He Z, Mao L, Wang J. Aegilops tauschii draft genome
sequence reveals a gene repertoire for wheat adaptation. Nature, 2013,
496(7443): 91-95.
[11] Schnable P S, Ware D, Fulton R S, Stein J C, Wei F, Pasternak S,
Liang C, Zhang J, Fulton L, Graves T A, Minx P, Reily A D, Courtney
L, Kruchowski S S, Tomlinson C, Strong C, Delehaunty K, Fronick C,
Courtney B, Rock S M, Belter E, Du F, Kim K, Abbott R M, Cotton
M, Levy A, Marchetto P, Ochoa K, Jackson S M, Gillam B, Chen W,
Yan L, Higginbotham J, Cardenas M, Waligorski J, Applebaum E,
Phelps L, Falcone J, Kanchi K, Thane T, Scimone A, Thane N, Henke
J, Wang T, Ruppert J, Shah N, Rotter K, Hodges J, Ingenthron E,

中 国 农 业 科 学 48 卷
Cordes M, Kohlberg S, Sgro J, Delgado B, Mead K, Chinwalla A,
Leonard S, Crouse K, Collura K, Kudrna D, Currie J, He R, Angelova
A, Rajasekar S, Mueller T, Lomeli R, Scara G, Ko A, Delaney K,
Wissotski M, Lopez G, Campos D, Braidotti M, Ashley E, Golser W,
Kim H, Lee S, Lin J, Dujmic Z, Kim W, Talag J, Zuccolo A, Fan C,
Sebastian A, Kramer M, Spiegel L, Nascimento L, Zutavern T, Miller
B, Ambroise C, Muller S, Spooner W, Narechania A, Ren L, Wei S,
Kumari S, Faga B, Levy M J, McMahan L, Van Buren P, Vaughn M W,
Ying K, Yeh C T, Emrich S J, Jia Y, Kalyanaraman A, Hsia A P,
Barbazuk W B, Baucom R S, Brutnell T P, Carpita N C, Chaparro C,
Chia J M, Deragon J M, Estill J C, Fu Y, Jeddeloh J A, Han Y, Lee H,
Li P, Lisch D R, Liu S, Liu Z, Nagel D H, McCann M C, SanMiguel P,
Myers A M, Nettleton D, Nguyen J, Penning B W, Ponnala L,
Schneider K L, Schwartz D C, Sharma A, Soderlund C, Springer N M,
Sun Q, Wang H, Waterman M, Westerman R, Wolfgruber T K, Yang L,
Yu Y, Zhang L, Zhou S, Zhu Q, Bennetzen J L, Dawe R K, Jiang J,
Jiang N, Presting G G, Wessler S R, Aluru S, Martienssen R A, Clifton
S W, McCombie W R, Wing R A, Wilson R K. The B73 maize
genome: complexity, diversity, and dynamics. Science, 2009,
326(5956): 1112-1115.
[12] Paterson A H, Bowers J E, Bruggmann R, Dubchak I, Grimwood J,
Gundlach H, Haberer G, Hellsten U, Mitros T, Poliakov A, Schmutz J,
Spannagl M, Tang H, Wang X, Wicker T, Bharti A K, Chapman J,
Feltus F A, Gowik U, Grigoriev I V, Lyons E, Maher C A, Martis M,
Narechania A, Otillar R P, Penning B W, Salamov A A, Wang Y,
Zhang L, Carpita N C, Freeling M, Gingle A R, Hash C T, Keller B,
Klein P, Kresovich S, McCann M C, Ming R, Peterson D G, Mehboob
ur R, Ware D, Westhoff P, Mayer K F, Messing J, Rokhsar D S. The
Sorghum bicolor genome and the diversification of grasses. Nature,
2009, 457(7229): 551-556.
[13] Bennetzen J L, Schmutz J, Wang H, Percifield R, Hawkins J, Pontaroli
A C, Estep M, Feng L, Vaughn J N, Grimwood J, Jenkins J, Barry K,
Lindquist E, Hellsten U, Deshpande S, Wang X, Wu X, Mitros T,
Triplett J, Yang X, Ye C Y, Mauro-Herrera M, Wang L, Li P, Sharma
M, Sharma R, Ronald P C, Panaud O, Kellogg E A, Brutnell T P,
Doust A N, Tuskan G A, Rokhsar D, Devos K M. Reference genome
sequence of the model plant Setaria. Nature Biotechnology, 2012,
30(6): 555-561.
[14] Zhang G, Liu X, Quan Z, Cheng S, Xu X, Pan S, Xie M, Zeng P, Yue
Z, Wang W, Tao Y, Bian C, Han C, Xia Q, Peng X, Cao R, Yang X,
Zhan D, Hu J, Zhang Y, Li H, Li H, Li N, Wang J, Wang C, Wang R,
Guo T, Cai Y, Liu C, Xiang H, Shi Q, Huang P, Chen Q, Li Y, Wang J,
Zhao Z, Wang J. Genome sequence of foxtail millet (Setaria italica)
provides insights into grass evolution and biofuel potential. Nature
Biotechnology, 2012, 30(6): 549-554.
[15] The International Brachypodium Initiative. Genome sequencing and
analysis of the model grass Brachypodium distachyon. Nature, 2010,
463(7282): 763-768.
[16] Mayer K F, Waugh R, Brown J W, Schulman A, Langridge P, Platzer
M, Fincher G B, Muehlbauer G J, Sato K, Close T J, Wise R P, Stein
N. A physical, genetic and functional sequence assembly of the barley
genome. Nature, 2012, 491(7426): 711-716.
[17] Zeng X, Long H, Wang Z, Zhao S, Tang Y, Huang Z, Wang Y, Xu Q,
Mao L, Deng G, Yao X, Li X, Bai L, Yuan H, Pan Z, Liu R, Chen X,
WangMu Q, Chen M, Yu L, Liang J, DunZhu D, Zheng Y, Yu S,
LuoBu Z, Guang X, Li J, Deng C, Hu W, Chen C, TaBa X, Gao L, Lv
X, Abu Y B, Fang X, Nevo E, Yu M, Wang J, Tashi N. The draft
genome of Tibetan hulless barley reveals adaptive patterns to the high
stressful Tibetan Plateau. Proceedings of the National Academy of
Sciences of the United States of America, 2015.
[18] Wang K, Wang Z, Li F, Ye W, Wang J, Song G, Yue Z, Cong L, Shang
H, Zhu S, Zou C, Li Q, Yuan Y, Lu C, Wei H, Gou C, Zheng Z, Yin Y,
Zhang X, Liu K, Wang B, Song C, Shi N, Kohel R J, Percy R G, Yu J
Z, Zhu Y X, Wang J, Yu S. The draft genome of a diploid cotton
Gossypium raimondii. Nature Genetics, 2012, 44(10): 1098-1103.
[19] Paterson A H, Wendel J F, Gundlach H, Guo H, Jenkins J, Jin D,
Llewellyn D, Showmaker K C, Shu S, Udall J, Yoo M J, Byers R,
Chen W, Doron-Faigenboim A, Duke M V, Gong L, Grimwood J,
Grover C, Grupp K, Hu G, Lee T H, Li J, Lin L, Liu T, Marler B S,
Page J T, Roberts A W, Romanel E, Sanders W S, Szadkowski E, Tan
X, Tang H, Xu C, Wang J, Wang Z, Zhang D, Zhang L, Ashrafi H,
Bedon F, Bowers J E, Brubaker C L, Chee P W, Das S, Gingle A R,
Haigler C H, Harker D, Hoffmann L V, Hovav R, Jones D C, Lemke C,
Mansoor S, ur Rahman M, Rainville L N, Rambani A, Reddy U K,
Rong J K, Saranga Y, Scheffler B E, Scheffler J A, Stelly D M, Triplett
B A, Van Deynze A, Vaslin M F, Waghmare V N, Walford S A, Wright
R J, Zaki E A, Zhang T, Dennis E S, Mayer K F, Peterson D G,
Rokhsar D S, Wang X, Schmutz J. Repeated polyploidization of
Gossypium genomes and the evolution of spinnable cotton fibres.
Nature, 2012, 492(7429): 423-427.
[20] Li F, Fan G, Wang K, Sun F, Yuan Y, Song G, Li Q, Ma Z, Lu C, Zou
C, Chen W, Liang X, Shang H, Liu W, Shi C, Xiao G, Gou C, Ye W,
Xu X, Zhang X, Wei H, Li Z, Zhang G, Wang J, Liu K, Kohel R J,
Percy R G, Yu J Z, Zhu Y X, Wang J, Yu S. Genome sequence of the
cultivated cotton Gossypium arboreum. Nature Genetics, 2014, 46(6):
567-572.

17 期 贾继增等:作物基因组学与作物科学革命
[21] Schmutz J, Cannon S B, Schlueter J, Ma J, Mitros T, Nelson W, Hyten
D L, Song Q, Thelen J J, Cheng J, Xu D, Hellsten U, May G D, Yu Y,
Sakurai T, Umezawa T, Bhattacharyya M K, Sandhu D, Valliyodan B,
Lindquist E, Peto M, Grant D, Shu S, Goodstein D, Barry K,
Futrell-Griggs M, Abernathy B, Du J, Tian Z, Zhu L, Gill N, Joshi T,
Libault M, Sethuraman A, Zhang X C, Shinozaki K, Nguyen H T,
Wing R A, Cregan P, Specht J, Grimwood J, Rokhsar D, Stacey G,
Shoemaker R C, Jackson S A. Genome sequence of the
palaeopolyploid soybean. Nature, 2010, 463(7278): 178-183.
[22] Qi X, Li M W, Xie M, Liu X, Ni M, Shao G, Song C, Kay-Yuen Yim
A, Tao Y, Wong F L, Isobe S, Wong C F, Wong K S, Xu C, Li C,
Wang Y, Guan R, Sun F, Fan G, Xiao Z, Zhou F, Phang T H, Liu X,
Tong S W, Chan T F, Yiu S M, Tabata S, Wang J, Xu X, Lam H M.
Identification of a novel salt tolerance gene in wild soybean by
whole-genome sequencing. Nature Communications, 2014, 5: 4340.
[23] Li Y H, Zhou G, Ma J, Jiang W, Jin L G, Zhang Z, Guo Y, Zhang J, Sui
Y, Zheng L, Zhang S S, Zuo Q, Shi X H, Li Y F, Zhang W K, Hu Y,
Kong G, Hong H L, Tan B, Song J, Liu Z X, Wang Y, Ruan H, Yeung
C K, Liu J, Wang H, Zhang L J, Guan R X, Wang K J, Li W B, Chen S
Y, Chang R Z, Jiang Z, Jackson S A, Li R, Qiu L J. De novo assembly
of soybean wild relatives for pan-genome analysis of diversity and
agronomic traits. Nature Biotechnology, 2014, 32(10): 1045-1052.
[24] Kim M Y, Lee S, Van K, Kim T H, Jeong S C, Choi I Y, Kim D S, Lee
Y S, Park D, Ma J, Kim W Y, Kim B C, Park S, Lee K A, Kim D H,
Kim K H, Shin J H, Jang Y E, Kim K D, Liu W X, Chaisan T, Kang Y
J, Lee Y H, Kim K H, Moon J K, Schmutz J, Jackson S A, Bhak J, Lee
S H. Whole-genome sequencing and intensive analysis of the
undomesticated soybean (Glycine soja Sieb. and Zucc.) genome.
Proceedings of the National Academy of Sciences of the United States
of America, 2010, 107(51): 22032-22037.
[25] Schmutz J, McClean P E, Mamidi S, Wu G A, Cannon S B,
Grimwood J, Jenkins J, Shu S, Song Q, Chavarro C, Torres-Torres M,
Geffroy V, Moghaddam S M, Gao D, Abernathy B, Barry K, Blair M,
Brick M A, Chovatia M, Gepts P, Goodstein D M, Gonzales M,
Hellsten U, Hyten D L, Jia G, Kelly J D, Kudrna D, Lee R, Richard M
M, Miklas P N, Osorno J M, Rodrigues J, Thareau V, Urrea C A,
Wang M, Yu Y, Zhang M, Wing R A, Cregan P B, Rokhsar D S,
Jackson S A. A reference genome for common bean and genome-wide
analysis of dual domestications. Nature Genetics, 2014, 46(7):
707-713.
[26] Varshney R K, Chen W, Li Y, Bharti A K, Saxena R K, Schlueter J A,
Donoghue M T, Azam S, Fan G, Whaley A M, Farmer A D, Sheridan J,
Iwata A, Tuteja R, Penmetsa R V, Wu W, Upadhyaya H D, Yang S P,
Shah T, Saxena K B, Michael T, McCombie W R, Yang B, Zhang G,
Yang H, Wang J, Spillane C, Cook D R, May G D, Xu X, Jackson S A.
Draft genome sequence of pigeonpea (Cajanus cajan), an orphan
legume crop of resource-poor farmers. Nature Biotechnology, 2012,
30(1): 83-89.
[27] Kang Y J, Kim S K, Kim M Y, Lestari P, Kim K H, Ha B K, Jun T H,
Hwang W J, Lee T, Lee J, Shim S, Yoon M Y, Jang Y E, Han K S,
Taeprayoon P, Yoon N, Somta P, Tanya P, Kim K S, Gwag J G, Moon J
K, Lee Y H, Park B S, Bombarely A, Doyle J J, Jackson S A,
Schafleitner R, Srinives P, Varshney R K, Lee S H. Genome sequence
of mungbean and insights into evolution within Vigna species. Nature
Communications, 2014, 5: 5443.
[28] Varshney R K, Song C, Saxena R K, Azam S, Yu S, Sharpe A G,
Cannon S, Baek J, Rosen B D, Tar'an B, Millan T, Zhang X, Ramsay
L D, Iwata A, Wang Y, Nelson W, Farmer A D, Gaur P M, Soderlund
C, Penmetsa R V, Xu C, Bharti A K, He W, Winter P, Zhao S, Hane J
K, Carrasquilla-Garcia N, Condie J A, Upadhyaya H D, Luo M C,
Thudi M, Gowda C L, Singh N P, Lichtenzveig J, Gali K K, Rubio J,
Nadarajan N, Dolezel J, Bansal K C, Xu X, Edwards D, Zhang G,
Kahl G, Gil J, Singh K B, Datta S K, Jackson S A, Wang J, Cook D R.
Draft genome sequence of chickpea (Cicer arietinum) provides a
resource for trait improvement. Nature Biotechnology, 2013, 31(3):
240-246.
[29] Chalhoub B, Denoeud F, Liu S, Parkin I A, Tang H, Wang X, Chiquet
J, Belcram H, Tong C, Samans B, Correa M, Da Silva C, Just J,
Falentin C, Koh C S, Le Clainche I, Bernard M, Bento P, Noel B,
Labadie K, Alberti A, Charles M, Arnaud D, Guo H, Daviaud C,
Alamery S, Jabbari K, Zhao M, Edger P P, Chelaifa H, Tack D,
Lassalle G, Mestiri I, Schnel N, Le Paslier M C, Fan G, Renault V,
Bayer P E, Golicz A A, Manoli S, Lee T H, Thi V H, Chalabi S, Hu Q,
Fan C, Tollenaere R, Lu Y, Battail C, Shen J, Sidebottom C H, Wang
X, Canaguier A, Chauveau A, Berard A, Deniot G, Guan M, Liu Z,
Sun F, Lim Y P, Lyons E, Town C D, Bancroft I, Wang X, Meng J, Ma
J, Pires J C, King G J, Brunel D, Delourme R, Renard M, Aury J M,
Adams K L, Batley J, Snowdon R J, Tost J, Edwards D, Zhou Y, Hua
W, Sharpe A G, Paterson A H, Guan C, Wincker P. Plant genetics.
Early allopolyploid evolution in the post-Neolithic Brassica napus
oilseed genome. Science, 2014, 345(6199): 950-953.
[30] Chan A P, Crabtree J, Zhao Q, Lorenzi H, Orvis J, Puiu D,
Melake-Berhan A, Jones K M, Redman J, Chen G, Cahoon E B, Gedil
M, Stanke M, Haas B J, Wortman J R, Fraser-Liggett C M, Ravel J,
Rabinowicz P D. Draft genome sequence of the oilseed species
Ricinus communis. Nature Biotechnology, 2010, 28(9): 951-956.

中 国 农 业 科 学 48 卷
[31] Wang L, Yu S, Tong C, Zhao Y, Liu Y, Song C, Zhang Y, Zhang X,
Wang Y, Hua W, Li D, Li D, Li F, Yu J, Xu C, Han X, Huang S, Tai S,
Wang J, Xu X, Li Y, Liu S, Varshney R K, Wang J, Zhang X. Genome
sequencing of the high oil crop sesame provides insight into oil
biosynthesis. Genome Biology, 2014, 15(2): R39.
[32] Wang X, Wang H, Wang J, Sun R, Wu J, Liu S, Bai Y, Mun J H,
Bancroft I, Cheng F, Huang S, Li X, Hua W, Wang J, Wang X,
Freeling M, Pires J C, Paterson A H, Chalhoub B, Wang B, Hayward
A, Sharpe A G, Park B S, Weisshaar B, Liu B, Li B, Liu B, Tong C,
Song C, Duran C, Peng C, Geng C, Koh C, Lin C, Edwards D, Mu D,
Shen D, Soumpourou E, Li F, Fraser F, Conant G, Lassalle G, King G J,
Bonnema G, Tang H, Wang H, Belcram H, Zhou H, Hirakawa H, Abe
H, Guo H, Wang H, Jin H, Parkin I A, Batley J, Kim J S, Just J, Li J,
Xu J, Deng J, Kim J A, Li J, Yu J, Meng J, Wang J, Min J, Poulain J,
Wang J, Hatakeyama K, Wu K, Wang L, Fang L, Trick M, Links M G,
Zhao M, Jin M, Ramchiary N, Drou N, Berkman P J, Cai Q, Huang Q,
Li R, Tabata S, Cheng S, Zhang S, Zhang S, Huang S, Sato S, Sun S,
Kwon S J, Choi S R, Lee T H, Fan W, Zhao X, Tan X, Xu X, Wang Y,
Qiu Y, Yin Y, Li Y, Du Y, Liao Y, Lim Y, Narusaka Y, Wang Y, Wang Z,
Li Z, Wang Z, Xiong Z, Zhang Z, Brassica rapa Genome Sequencing
Project C. The genome of the mesopolyploid crop species Brassica
rapa. Nature Genetics, 2011, 43(10): 1035-1039.
[33] Liu S, Liu Y, Yang X, Tong C, Edwards D, Parkin I A, Zhao M, Ma J,
Yu J, Huang S, Wang X, Wang J, Lu K, Fang Z, Bancroft I, Yang T J,
Hu Q, Wang X, Yue Z, Li H, Yang L, Wu J, Zhou Q, Wang W, King G
J, Pires J C, Lu C, Wu Z, Sampath P, Wang Z, Guo H, Pan S, Yang L,
Min J, Zhang D, Jin D, Li W, Belcram H, Tu J, Guan M, Qi C, Du D,
Li J, Jiang L, Batley J, Sharpe A G, Park B S, Ruperao P, Cheng F,
Waminal N E, Huang Y, Dong C, Wang L, Li J, Hu Z, Zhuang M,
Huang Y, Huang J, Shi J, Mei D, Liu J, Lee T H, Wang J, Jin H, Li Z,
Li X, Zhang J, Xiao L, Zhou Y, Liu Z, Liu X, Qin R, Tang X, Liu W,
Wang Y, Zhang Y, Lee J, Kim H H, Denoeud F, Xu X, Liang X, Hua
W, Wang X, Wang J, Chalhoub B, Paterson A H. The Brassica
oleracea genome reveals the asymmetrical evolution of polyploid
genomes. Nature Communications, 2014, 5: 3930.
[34] Kitashiba H, Li F, Hirakawa H, Kawanabe T, Zou Z, Hasegawa Y,
Tonosaki K, Shirasawa S, Fukushima A, Yokoi S, Takahata Y,
Kakizaki T, Ishida M, Okamoto S, Sakamoto K, Shirasawa K, Tabata
S, Nishio T. Draft sequences of the radish (Raphanus sativus L.)
genome. DNA Research, 2014, 21(5): 481-490.
[35] Huang S, Li R, Zhang Z, Li L, Gu X, Fan W, Lucas W J, Wang X, Xie
B, Ni P, Ren Y, Zhu H, Li J, Lin K, Jin W, Fei Z, Li G, Staub J, Kilian
A, van der Vossen E A, Wu Y, Guo J, He J, Jia Z, Ren Y, Tian G, Lu Y,
Ruan J, Qian W, Wang M, Huang Q, Li B, Xuan Z, Cao J, Asan, Wu Z,
Zhang J, Cai Q, Bai Y, Zhao B, Han Y, Li Y, Li X, Wang S, Shi Q, Liu
S, Cho W K, Kim J Y, Xu Y, Heller-Uszynska K, Miao H, Cheng Z,
Zhang S, Wu J, Yang Y, Kang H, Li M, Liang H, Ren X, Shi Z, Wen
M, Jian M, Yang H, Zhang G, Yang Z, Chen R, Liu S, Li J, Ma L, Liu
H, Zhou Y, Zhao J, Fang X, Li G, Fang L, Li Y, Liu D, Zheng H,
Zhang Y, Qin N, Li Z, Yang G, Yang S, Bolund L, Kristiansen K,
Zheng H, Li S, Zhang X, Yang H, Wang J, Sun R, Zhang B, Jiang S,
Wang J, Du Y, Li S. The genome of the cucumber, Cucumis sativus L..
Nature Genetics, 2009, 41(12): 1275-1281.
[36] Xu X, Pan S, Cheng S, Zhang B, Mu D, Ni P, Zhang G, Yang S, Li R,
Wang J, Orjeda G, Guzman F, Torres M, Lozano R, Ponce O, Martinez
D, De la Cruz G, Chakrabarti S K, Patil V U, Skryabin K G,
Kuznetsov B B, Ravin N V, Kolganova T V, Beletsky A V, Mardanov
A V, Di Genova A, Bolser D M, Martin D M, Li G, Yang Y, Kuang H,
Hu Q, Xiong X, Bishop G J, Sagredo B, Mejia N, Zagorski W,
Gromadka R, Gawor J, Szczesny P, Huang S, Zhang Z, Liang C, He J,
Li Y, He Y, Xu J, Zhang Y, Xie B, Du Y, Qu D, Bonierbale M,
Ghislain M, Herrera Mdel R, Giuliano G, Pietrella M, Perrotta G,
Facella P, O'Brien K, Feingold S E, Barreiro L E, Massa G A,
Diambra L, Whitty B R, Vaillancourt B, Lin H, Massa A N, Geoffroy
M, Lundback S, DellaPenna D, Buell C R, Sharma S K, Marshall D F,
Waugh R, Bryan G J, Destefanis M, Nagy I, Milbourne D, Thomson S
J, Fiers M, Jacobs J M, Nielsen K L, Sonderkaer M, Iovene M, Torres
G A, Jiang J, Veilleux R E, Bachem C W, de Boer J, Borm T,
Kloosterman B, van Eck H, Datema E, Hekkert B, Goverse A, van
Ham R C, Visser R G. Genome sequence and analysis of the tuber
crop potato. Nature, 2011, 475(7355): 189-195.
[37] The Tomato Genome Consortium. The tomato genome sequence
provides insights into fleshy fruit evolution. Nature, 2012, 485(7400):
635-641.
[38] Bolger A, Scossa F, Bolger M E, Lanz C, Maumus F, Tohge T,
Quesneville H, Alseekh S, Sorensen I, Lichtenstein G, Fich E A, Conte
M, Keller H, Schneeberger K, Schwacke R, Ofner I, Vrebalov J, Xu Y,
Osorio S, Aflitos S A, Schijlen E, Jimenez-Gomez J M, Ryngajllo M,
Kimura S, Kumar R, Koenig D, Headland L R, Maloof J N, Sinha N,
van Ham R C, Lankhorst R K, Mao L, Vogel A, Arsova B, Panstruga
R, Fei Z, Rose J K, Zamir D, Carrari F, Giovannoni J J, Weigel D,
Usadel B, Fernie A R. The genome of the stress-tolerant wild tomato
species Solanum pennellii. Nature Genetics, 2014, 46(9): 1034-1038.
[39] Dohm J C, Minoche A E, Holtgrawe D, Capella-Gutierrez S,
Zakrzewski F, Tafer H, Rupp O, Sorensen T R, Stracke R, Reinhardt R,
Goesmann A, Kraft T, Schulz B, Stadler P F, Schmidt T, Gabaldon T,

17 期 贾继增等:作物基因组学与作物科学革命
Lehrach H, Weisshaar B, Himmelbauer H. The genome of the recently
domesticated crop plant sugar beet (Beta vulgaris). Nature, 2014,
505(7484): 546-549.
[40] Kim S, Park M, Yeom S I, Kim Y M, Lee J M, Lee H A, Seo E, Choi J,
Cheong K, Kim K T, Jung K, Lee G W, Oh S K, Bae C, Kim S B, Lee
H Y, Kim S Y, Kim M S, Kang B C, Jo Y D, Yang H B, Jeong H J,
Kang W H, Kwon J K, Shin C, Lim J Y, Park J H, Huh J H, Kim J S,
Kim B D, Cohen O, Paran I, Suh M C, Lee S B, Kim Y K, Shin Y,
Noh S J, Park J, Seo Y S, Kwon S Y, Kim H A, Park J M, Kim H J,
Choi S B, Bosland P W, Reeves G, Jo S H, Lee B W, Cho H T, Choi H
S, Lee M S, Yu Y, Do Choi Y, Park B S, van Deynze A, Ashrafi H, Hill
T, Kim W T, Pai H S, Ahn H K, Yeam I, Giovannoni J J, Rose J K,
Sorensen I, Lee S J, Kim R W, Choi I Y, Choi B S, Lim J S, Lee Y H,
Choi D. Genome sequence of the hot pepper provides insights into the
evolution of pungency in Capsicum species. Nature Genetics, 2014,
46(3): 270-278.
[41] Qin C, Yu C, Shen Y, Fang X, Chen L, Min J, Cheng J, Zhao S, Xu M,
Luo Y, Yang Y, Wu Z, Mao L, Wu H, Ling-Hu C, Zhou H, Lin H,
Gonzalez-Morales S, Trejo-Saavedra D L, Tian H, Tang X, Zhao M,
Huang Z, Zhou A, Yao X, Cui J, Li W, Chen Z, Feng Y, Niu Y, Bi S,
Yang X, Li W, Cai H, Luo X, Montes-Hernandez S, Leyva-Gonzalez
M A, Xiong Z, He X, Bai L, Tan S, Tang X, Liu D, Liu J, Zhang S,
Chen M, Zhang L, Zhang L, Zhang Y, Liao W, Zhang Y, Wang M, Lv
X, Wen B, Liu H, Luan H, Zhang Y, Yang S, Wang X, Xu J, Li X, Li S,
Wang J, Palloix A, Bosland P W, Li Y, Krogh A, Rivera-Bustamante R
F, Herrera-Estrella L, Yin Y, Yu J, Hu K, Zhang Z. Whole-genome
sequencing of cultivated and wild peppers provides insights into
Capsicum domestication and specialization. Proceedings of the
National Academy of Sciences of the United States of America, 2014,
111(14): 5135-5140.
[42] Guo S, Zhang J, Sun H, Salse J, Lucas W J, Zhang H, Zheng Y, Mao L,
Ren Y, Wang Z, Min J, Guo X, Murat F, Ham B K, Zhang Z, Gao S,
Huang M, Xu Y, Zhong S, Bombarely A, Mueller L A, Zhao H, He H,
Zhang Y, Zhang Z, Huang S, Tan T, Pang E, Lin K, Hu Q, Kuang H,
Ni P, Wang B, Liu J, Kou Q, Hou W, Zou X, Jiang J, Gong G, Klee K,
Schoof H, Huang Y, Hu X, Dong S, Liang D, Wang J, Wu K, Xia Y,
Zhao X, Zheng Z, Xing M, Liang X, Huang B, Lv T, Wang J, Yin Y,
Yi H, Li R, Wu M, Levi A, Zhang X, Giovannoni J J, Wang J, Li Y,
Fei Z, Xu Y. The draft genome of watermelon (Citrullus lanatus) and
resequencing of 20 diverse accessions. Nature Genetics, 2013, 45(1):
51-58.
[43] Hirakawa H, Shirasawa K, Miyatake K, Nunome T, Negoro S,
Ohyama A, Yamaguchi H, Sato S, Isobe S, Tabata S, Fukuoka H. Draft
genome sequence of eggplant (Solanum melongena L.): The
representative Solanum species indigenous to the old world. DNA
Research, 2014, 21(6): 649-660.
[44] Velasco R, Zharkikh A, Affourtit J, Dhingra A, Cestaro A,
Kalyanaraman A, Fontana P, Bhatnagar S K, Troggio M, Pruss D,
Salvi S, Pindo M, Baldi P, Castelletti S, Cavaiuolo M, Coppola G,
Costa F, Cova V, Dal Ri A, Goremykin V, Komjanc M, Longhi S,
Magnago P, Malacarne G, Malnoy M, Micheletti D, Moretto M,
Perazzolli M, Si-Ammour A, Vezzulli S, Zini E, Eldredge G,
Fitzgerald L M, Gutin N, Lanchbury J, Macalma T, Mitchell J T, Reid
J, Wardell B, Kodira C, Chen Z, Desany B, Niazi F, Palmer M,
Koepke T, Jiwan D, Schaeffer S, Krishnan V, Wu C, Chu V T, King S
T, Vick J, Tao Q, Mraz A, Stormo A, Stormo K, Bogden R, Ederle D,
Stella A, Vecchietti A, Kater M M, Masiero S, Lasserre P, Lespinasse
Y, Allan A C, Bus V, Chagne D, Crowhurst R N, Gleave A P, Lavezzo
E, Fawcett J A, Proost S, Rouze P, Sterck L, Toppo S, Lazzari B,
Hellens R P, Durel C E, Gutin A, Bumgarner R E, Gardiner S E,
Skolnick M, Egholm M, Van de Peer Y, Salamini F, Viola R. The
genome of the domesticated apple (Malus× domestica Borkh.).
Nature Genetics, 2010, 42(10): 833-839.
[45] Wu J, Wang Z, Shi Z, Zhang S, Ming R, Zhu S, Khan M A, Tao S,
Korban S S, Wang H, Chen N J, Nishio T, Xu X, Cong L, Qi K,
Huang X, Wang Y, Zhao X, Wu J, Deng C, Gou C, Zhou W, Yin H,
Qin G, Sha Y, Tao Y, Chen H, Yang Y, Song Y, Zhan D, Wang J, Li L,
Dai M, Gu C, Wang Y, Shi D, Wang X, Zhang H, Zeng L, Zheng D,
Wang C, Chen M, Wang G, Xie L, Sovero V, Sha S, Huang W, Zhang
S, Zhang M, Sun J, Xu L, Li Y, Liu X, Li Q, Shen J, Wang J, Paull R
E, Bennetzen J L, Wang J, Zhang S. The genome of the pear (Pyrus
bretschneideri Rehd.). Genome Research, 2013, 23(2): 396-408.
[46] Jaillon O, Aury J M, Noel B, Policriti A, Clepet C, Casagrande A,
Choisne N, Aubourg S, Vitulo N, Jubin C, Vezzi A, Legeai F,
Hugueney P, Dasilva C, Horner D, Mica E, Jublot D, Poulain J,
Bruyere C, Billault A, Segurens B, Gouyvenoux M, Ugarte E,
Cattonaro F, Anthouard V, Vico V, Del Fabbro C, Alaux M, Di
Gaspero G, Dumas V, Felice N, Paillard S, Juman I, Moroldo M,
Scalabrin S, Canaguier A, Le Clainche I, Malacrida G, Durand E,
Pesole G, Laucou V, Chatelet P, Merdinoglu D, Delledonne M,
Pezzotti M, Lecharny A, Scarpelli C, Artiguenave F, Pe M E, Valle G,
Morgante M, Caboche M, Adam-Blondon A F, Weissenbach J, Quetier
F, Wincker P. The grapevine genome sequence suggests ancestral
hexaploidization in major angiosperm phyla. Nature, 2007, 449(7161):
463-467.
[47] Ming R, Hou S, Feng Y, Yu Q, Dionne-Laporte A, Saw J H, Senin P,

中 国 农 业 科 学 48 卷
Wang W, Ly B V, Lewis K L, Salzberg S L, Feng L, Jones M R,
Skelton R L, Murray J E, Chen C, Qian W, Shen J, Du P, Eustice M,
Tong E, Tang H, Lyons E, Paull R E, Michael T P, Wall K, Rice D W,
Albert H, Wang M L, Zhu Y J, Schatz M, Nagarajan N, Acob R A,
Guan P, Blas A, Wai C M, Ackerman C M, Ren Y, Liu C, Wang J,
Wang J, Na J K, Shakirov E V, Haas B, Thimmapuram J, Nelson D,
Wang X, Bowers J E, Gschwend A R, Delcher A L, Singh R, Suzuki J
Y, Tripathi S, Neupane K, Wei H, Irikura B, Paidi M, Jiang N, Zhang
W, Presting G, Windsor A, Navajas-Perez R, Torres M J, Feltus F A,
Porter B, Li Y, Burroughs A M, Luo M C, Liu L, Christopher D A,
Mount S M, Moore P H, Sugimura T, Jiang J, Schuler M A, Friedman
V, Mitchell-Olds T, Shippen D E, dePamphilis C W, Palmer J D,
Freeling M, Paterson A H, Gonsalves D, Wang L, Alam M. The draft
genome of the transgenic tropical fruit tree papaya (Carica papaya
Linnaeus). Nature, 2008, 452(7190): 991-996.
[48] Xu Q, Chen L L, Ruan X, Chen D, Zhu A, Chen C, Bertrand D, Jiao
W B, Hao B H, Lyon M P, Chen J, Gao S, Xing F, Lan H, Chang J W,
Ge X, Lei Y, Hu Q, Miao Y, Wang L, Xiao S, Biswas M K, Zeng W,
Guo F, Cao H, Yang X, Xu X W, Cheng Y J, Xu J, Liu J H, Luo O J,
Tang Z, Guo W W, Kuang H, Zhang H Y, Roose M L, Nagarajan N,
Deng X X, Ruan Y. The draft genome of sweet orange (Citrus
sinensis). Nature Genetics, 2013, 45(1): 59-66.
[49] Huang S, Ding J, Deng D, Tang W, Sun H, Liu D, Zhang L, Niu X,
Zhang X, Meng M, Yu J, Liu J, Han Y, Shi W, Zhang D, Cao S, Wei Z,
Cui Y, Xia Y, Zeng H, Bao K, Lin L, Min Y, Zhang H, Miao M, Tang
X, Zhu Y, Sui Y, Li G, Sun H, Yue J, Sun J, Liu F, Zhou L, Lei L,
Zheng X, Liu M, Huang L, Song J, Xu C, Li J, Ye K, Zhong S, Lu B R,
He G, Xiao F, Wang H L, Zheng H, Fei Z, Liu Y. Draft genome of the
kiwifruit Actinidia chinensis. Nature Communications, 2013, 4: 2640.
[50] Shulaev V, Sargent D J, Crowhurst R N, Mockler T C, Folkerts O,
Delcher A L, Jaiswal P, Mockaitis K, Liston A, Mane S P, Burns P,
Davis T M, Slovin J P, Bassil N, Hellens R P, Evans C, Harkins T,
Kodira C, Desany B, Crasta O R, Jensen R V, Allan A C, Michael T P,
Setubal J C, Celton J M, Rees D J, Williams K P, Holt S H, Ruiz Rojas
J J, Chatterjee M, Liu B, Silva H, Meisel L, Adato A, Filichkin S A,
Troggio M, Viola R, Ashman T L, Wang H, Dharmawardhana P, Elser
J, Raja R, Priest H D, Bryant D W, Jr., Fox S E, Givan S A, Wilhelm L
J, Naithani S, Christoffels A, Salama D Y, Carter J, Lopez Girona E,
Zdepski A, Wang W, Kerstetter R A, Schwab W, Korban S S, Davik J,
Monfort A, Denoyes-Rothan B, Arus P, Mittler R, Flinn B, Aharoni A,
Bennetzen J L, Salzberg S L, Dickerman A W, Velasco R, Borodovsky
M, Veilleux R E, Folta K M. The genome of woodland strawberry
(Fragaria vesca). Nature Genetics, 2011, 43(2): 109-116.
[51] Hirakawa H, Shirasawa K, Kosugi S, Tashiro K, Nakayama S,
Yamada M, Kohara M, Watanabe A, Kishida Y, Fujishiro T, Tsuruoka
H, Minami C, Sasamoto S, Kato M, Nanri K, Komaki A, Yanagi T,
Guoxin Q, Maeda F, Ishikawa M, Kuhara S, Sato S, Tabata S, Isobe S
N. Dissection of the octoploid strawberry genome by deep sequencing
of the genomes of Fragaria species. DNA Research, 2014, 21(2):
169-181.
[52] Garcia-Mas J, Benjak A, Sanseverino W, Bourgeois M, Mir G,
Gonzalez V M, Henaff E, Camara F, Cozzuto L, Lowy E, Alioto T,
Capella-Gutierrez S, Blanca J, Canizares J, Ziarsolo P, Gonzalez-Ibeas
D, Rodriguez-Moreno L, Droege M, Du L, Alvarez-Tejado M,
Lorente-Galdos B, Mele M, Yang L, Weng Y, Navarro A,
Marques-Bonet T, Aranda M A, Nuez F, Pico B, Gabaldon T, Roma G,
Guigo R, Casacuberta J M, Arus P, Puigdomenech P. The genome of
melon (Cucumis melo L.). Proceedings of the National Academy of
Sciences of the United States of America, 2012, 109(29): 11872-7.
[53] Liu M J, Zhao J, Cai Q L, Liu G C, Wang J R, Zhao Z H, Liu P, Dai L,
Yan G, Wang W J, Li X S, Chen Y, Sun Y D, Liu Z G, Lin M-J, Xiao J,
Chen Y Y, Li X F, Wu B, Ma Y, Jian J B, Yang W, Yuan Z, Sun X C,
Wei Y L, Yu L L, Zhang C, Liao S G, He R J, Guang X M, Wang Z,
Zhang Y Y, Luo L H. The complex jujube genome provides insights
into fruit tree biology. Nature Communications, 2014.
[54] Al-Dous E K, George B, Al-Mahmoud M E, Al-Jaber M Y, Wang H,
Salameh Y M, Al-Azwani E K, Chaluvadi S, Pontaroli A C, DeBarry J,
Arondel V, Ohlrogge J, Saie I J, Suliman-Elmeer K M, Bennetzen J L,
Kruegger R R, Malek J A. De novo genome sequencing and
comparative genomics of date palm (Phoenix dactylifera). Nature
Biotechnology, 2011, 29(6): 521-527.
[55] Al-Mssallem I S, Hu S, Zhang X, Lin Q, Liu W, Tan J, Yu X, Liu J,
Pan L, Zhang T, Yin Y, Xin C, Wu H, Zhang G, Ba Abdullah M M,
Huang D, Fang Y, Alnakhli Y O, Jia S, Yin A, Alhuzimi E M, Alsaihati
B A, Al-Owayyed S A, Zhao D, Zhang S, Al-Otaibi N A, Sun G,
Majrashi M A, Li F, Tala, Wang J, Yun Q, Alnassar N A, Wang L,
Yang M, Al-Jelaify R F, Liu K, Gao S, Chen K, Alkhaldi S R, Liu G,
Zhang M, Guo H, Yu J. Genome sequence of the date palm Phoenix
dactylifera L. Nature Communications, 2013, 4: 2274.
[56] Verde I, Abbott A G, Scalabrin S, Jung S, Shu S, Marroni F,
Zhebentyayeva T, Dettori M T, Grimwood J, Cattonaro F, Zuccolo A,
Rossini L, Jenkins J, Vendramin E, Meisel L A, Decroocq V, Sosinski
B, Prochnik S, Mitros T, Policriti A, Cipriani G, Dondini L, Ficklin S,
Goodstein D M, Xuan P, Del Fabbro C, Aramini V, Copetti D,
Gonzalez S, Horner D S, Falchi R, Lucas S, Mica E, Maldonado J,
Lazzari B, Bielenberg D, Pirona R, Miculan M, Barakat A, Testolin R,

17 期 贾继增等:作物基因组学与作物科学革命
Stella A, Tartarini S, Tonutti P, Arus P, Orellana A, Wells C, Main D,
Vizzotto G, Silva H, Salamini F, Schmutz J, Morgante M, Rokhsar D S.
The high-quality draft genome of peach (Prunus persica) identifies
unique patterns of genetic diversity, domestication and genome
evolution. Nature Genetics, 2013, 45(5): 487-494.
[57] D'Hont A, Denoeud F, Aury J M, Baurens F C, Carreel F, Garsmeur O,
Noel B, Bocs S, Droc G, Rouard M, Da Silva C, Jabbari K, Cardi C,
Poulain J, Souquet M, Labadie K, Jourda C, Lengelle J, Rodier-Goud
M, Alberti A, Bernard M, Correa M, Ayyampalayam S, McKain M R,
Leebens-Mack J, Burgess D, Freeling M, Mbeguie A M D, Chabannes
M, Wicker T, Panaud O, Barbosa J, Hribova E, Heslop-Harrison P,
Habas R, Rivallan R, Francois P, Poiron C, Kilian A, Burthia D, Jenny
C, Bakry F, Brown S, Guignon V, Kema G, Dita M, Waalwijk C,
Joseph S, Dievart A, Jaillon O, Leclercq J, Argout X, Lyons E,
Almeida A, Jeridi M, Dolezel J, Roux N, Risterucci A M,
Weissenbach J, Ruiz M, Glaszmann J C, Quetier F, Yahiaoui N,
Wincker P. The banana (Musa acuminata) genome and the evolution
of monocotyledonous plants. Nature, 2012, 488(7410): 213-217.
[58] Singh R, Ong-Abdullah M, Low E T, Manaf M A, Rosli R, Nookiah R,
Ooi L C, Ooi S E, Chan K L, Halim M A, Azizi N, Nagappan J,
Bacher B, Lakey N, Smith S W, He D, Hogan M, Budiman M A, Lee
E K, DeSalle R, Kudrna D, Goicoechea J L, Wing R A, Wilson R K,
Fulton R S, Ordway J M, Martienssen R A, Sambanthamurthi R. Oil
palm genome sequence reveals divergence of interfertile species in
Old and New worlds. Nature, 2013, 500(7462): 335-339.
[59] Wang Y, Fan G, Liu Y, Sun F, Shi C, Liu X, Peng J, Chen W, Huang X,
Cheng S, Liu Y, Liang X, Zhu H, Bian C, Zhong L, Lv T, Dong H, Liu
W, Zhong X, Chen J, Quan Z, Wang Z, Tan B, Lin C, Mu F, Xu X,
Ding Y, Guo A Y, Wang J, Ke W. The sacred lotus genome provides
insights into the evolution of flowering plants. The Plant Journal,
2013, 76(4): 557-567.
[60] Peng Z, Lu Y, Li L, Zhao Q, Feng Q, Gao Z, Lu H, Hu T, Yao N, Liu
K, Li Y, Fan D, Guo Y, Li W, Lu Y, Weng Q, Zhou C, Zhang L, Huang
T, Zhao Y, Zhu C, Liu X, Yang X, Wang T, Miao K, Zhuang C, Cao X,
Tang W, Liu G, Liu Y, Chen J, Liu Z, Yuan L, Liu Z, Huang X, Lu T,
Fei B, Ning Z, Han B, Jiang Z. The draft genome of the fast-growing
non-timber forest species moso bamboo (Phyllostachys heterocycla).
Nature Genetics, 2013, 45(4): 456-461, 461e1-461e2.
[61] Denoeud F, Carretero-Paulet L, Dereeper A, Droc G, Guyot R,
Pietrella M, Zheng C, Alberti A, Anthony F, Aprea G, Aury J M, Bento
P, Bernard M, Bocs S, Campa C, Cenci A, Combes M C, Crouzillat D,
Da Silva C, Daddiego L, De Bellis F, Dussert S, Garsmeur O,
Gayraud T, Guignon V, Jahn K, Jamilloux V, Joet T, Labadie K, Lan T,
Leclercq J, Lepelley M, Leroy T, Li L T, Librado P, Lopez L, Munoz
A, Noel B, Pallavicini A, Perrotta G, Poncet V, Pot D, Priyono,
Rigoreau M, Rouard M, Rozas J, Tranchant-Dubreuil C, VanBuren R,
Zhang Q, Andrade A C, Argout X, Bertrand B, de Kochko A, Graziosi
G, Henry R J, Jayarama, Ming R, Nagai C, Rounsley S, Sankoff D,
Giuliano G, Albert V A, Wincker P, Lashermes P. The coffee genome
provides insight into the convergent evolution of caffeine biosynthesis.
Science, 2014, 345(6201): 1181-1184.
[62] Natsume S, Takagi H, Shiraishi A, Murata J, Toyonaga H, Patzak J,
Takagi M, Yaegashi H, Uemura A, Mitsuoka C, Yoshida K, Krofta K,
Satake H, Terauchi R, Ono E. The draft genome of hop (Humulus
lupulus), an essence for brewing. Plant & Cell Physiology, 2014.
[63] He N, Zhang C, Qi X, Zhao S, Tao Y, Yang G, Lee T H, Wang X, Cai
Q, Li D, Lu M, Liao S, Luo G, He R, Tan X, Xu Y, Li T, Zhao A, Jia L,
Fu Q, Zeng Q, Gao C, Ma B, Liang J, Wang X, Shang J, Song P, Wu
H, Fan L, Wang Q, Shuai Q, Zhu J, Wei C, Zhu-Salzman K, Jin D,
Wang J, Liu T, Yu M, Tang C, Wang Z, Dai F, Chen J, Liu Y, Zhao S,
Lin T, Zhang S, Wang J, Wang J, Yang H, Yang G, Wang J, Paterson A
H, Xia Q, Ji D, Xiang Z. Draft genome sequence of the mulberry tree
Morus notabilis. Nature Communications, 2013, 4: 2445.
[64] Ming R, VanBuren R, Liu Y, Yang M, Han Y, Li L T, Zhang Q, Kim M
J, Schatz M C, Campbell M, Li J, Bowers J E, Tang H, Lyons E,
Ferguson A A, Narzisi G, Nelson D R, Blaby-Haas C E, Gschwend A
R, Jiao Y, Der J P, Zeng F, Han J, Min X J, Hudson K A, Singh R,
Grennan A K, Karpowicz S J, Watling J R, Ito K, Robinson S A,
Hudson M E, Yu Q, Mockler T C, Carroll A, Zheng Y, Sunkar R, Jia R,
Chen N, Arro J, Wai C M, Wafula E, Spence A, Han Y, Xu L, Zhang J,
Peery R, Haus M J, Xiong W, Walsh J A, Wu J, Wang M L, Zhu Y J,
Paull R E, Britt A B, Du C, Downie S R, Schuler M A, Michael T P,
Long S P, Ort D R, Schopf J W, Gang D R, Jiang N, Yandell M,
dePamphilis C W, Merchant S S, Paterson A H, Buchanan B B, Li S,
Shen-Miller J. Genome of the long-living sacred lotus (Nelumbo
nucifera Gaertn.). Genome Biology, 2013, 14(5): R41.
[65] Rahman A Y, Usharraj A O, Misra B B, Thottathil G P, Jayasekaran K,
Feng Y, Hou S, Ong S Y, Ng F L, Lee L S, Tan H S, Sakaff M K, Teh
B S, Khoo B F, Badai S S, Aziz N A, Yuryev A, Knudsen B,
Dionne-Laporte A, McHunu N P, Yu Q, Langston B J, Freitas T A,
Young A G, Chen R, Wang L, Najimudin N, Saito J A, Alam M. Draft
genome sequence of the rubber tree Hevea brasiliensis. BMC
Genomics, 2013, 14: 75.
[66] Argout X, Salse J, Aury J M, Guiltinan M J, Droc G, Gouzy J, Allegre
M, Chaparro C, Legavre T, Maximova S N, Abrouk M, Murat F, Fouet
O, Poulain J, Ruiz M, Roguet Y, Rodier-Goud M, Barbosa-Neto J F,

中 国 农 业 科 学 48 卷
Sabot F, Kudrna D, Ammiraju J S, Schuster S C, Carlson J E, Sallet E,
Schiex T, Dievart A, Kramer M, Gelley L, Shi Z, Berard A, Viot C,
Boccara M, Risterucci A M, Guignon V, Sabau X, Axtell M J, Ma Z,
Zhang Y, Brown S, Bourge M, Golser W, Song X, Clement D,
Rivallan R, Tahi M, Akaza J M, Pitollat B, Gramacho K, D'Hont A,
Brunel D, Infante D, Kebe I, Costet P, Wing R, McCombie W R,
Guiderdoni E, Quetier F, Panaud O, Wincker P, Bocs S, Lanaud C.
The genome of Theobroma cacao. Nature Genetics, 2011, 43(2):
101-108.
附表 2: [1] Munoz-Amatriain M, Cuesta-Marcos A, Endelman J B, Comadran J,
Bonman J M, Bockelman H E, Chao S, Russell J, Waugh R, Hayes P
M, Muehlbauer G J. The USDA barley core collection: Genetic
diversity, population structure, and potential for genome-wide
association studies. PLoS One, 2014, 9(4): e94688.
[2] Li Y H, Zhou G, Ma J, Jiang W, Jin L G, Zhang Z, Guo Y, Zhang J, Sui
Y, Zheng L, Zhang S S, Zuo Q, Shi X H, Li Y F, Zhang W K, Hu Y,
Kong G, Hong H L, Tan B, Song J, Liu Z X, Wang Y, Ruan H, Yeung
C K, Liu J, Wang H, Zhang L J, Guan R X, Wang K J, Li W B, Chen S
Y, Chang R Z, Jiang Z, Jackson S A, Li R, Qiu L J. De novo assembly
of soybean wild relatives for pan-genome analysis of diversity and
agronomic traits. Nature Biotechnology, 2014, 32(10): 1045-1052.
[3] Igartua E, Gracia M P, Lasa J M, Medina B, Molina-Cano J L,
Montoya J L, Romagosa I. The Spanish barley core collection.
Genetic Resources and Crop Evolution, 1998, 45(5): 475-481.
[4] Zhang H, Zhang D, Wang M, Sun J, Qi Y, Li J, Wei X, Han L, Qiu Z,
Tang S, Li Z. A core collection and mini core collection of Oryza
sativa L. in China. Theoretical and Applied Genetics, 2011, 122(1):
49-61.
[5] Yan W, Rutger J N, Bryant R J, Bockelman H E, Fjellstrom R G, Chen
M-H, Tai T H, McClung A M. Development and evaluation of a core
subset of the USDA rice germplasm collection. Crop Science, 2007,
47(2): 869-876.
[6] Agrama H A, Yan W, Lee F, Fjellstrom R, Chen M-H, Jia M, McClung
A. Genetic assessment of a mini-core subset developed from the
USDA rice genebank. Crop Science, 2009, 49(4): 1336-1346.
[7] Abadie T, Cordeiro C M T, Fonseca J R, Alves R D B D N, Burle M L,
Brondani C, Rangel P H N, Castro E d M d, Silva H T D, Freire M S,
Zimmermann F J P, Magalhães J R. Constructing a rice core collection
for Brazil. Pesquisa Agropecuária Brasileira, 2005, 40: 129-136.
[8] de Oliveira Borba T C, Brondani R P, Rangel P H, Brondani C.
Microsatellite marker-mediated analysis of the EMBRAPA rice core
collection genetic diversity. Genetica, 2009, 137(3): 293-304.
[9] Li X L, Lu Y G, Li J Q, Xu H M, Shahid M Q. Strategies on sample
size determination and qualitative and quantitative traits integration to
construct core collection of rice (Oryza sativa). Rice Science, 2011,
18(1): 46-55.
[10] Ebana K, Kojima Y, Fukuoka S, Nagamine T, Kawase M.
Development of mini core collection of Japanese rice landrace.
Breeding Science, 2008, 58(3): 281-291.
[11] Hao C, Dong Y, Wang L, You G, Zhang H, Ge H, Jia J, Zhang X.
Genetic diversity and construction of core collection in Chinese wheat
genetic resources. Chinese Science Bulletin, 2008, 53(10): 1518-1526.
[12] Balfourier F, Roussel V, Strelchenko P, Exbrayat-Vinson F, Sourdille P,
Boutet G, Koenig J, Ravel C, Mitrofanova O, Beckert M, Charmet G.
A worldwide bread wheat core collection arrayed in a 384-well plate.
Theoretical and Applied Genetics, 2007, 114(7): 1265-1275.
[13] Li Y, Shi Y, Cao Y, Wang T. Establishment of a core collection for
maize germplasm preserved in Chinese National Genebank using
geographic distribution and characterization data. Genetic Resources
and Crop Evolution, 2005, 51(8): 845-852.
[14] Yu Y, Wang E, Shi Y, Song Y, Wang T, Li Y. Genetic diversity and
structure of the core collection for maize inbred lines in China.
Maydica, 2007, 52: 181-194.
[15] Malosetti M, Abadie T. Sampling strategy to develop a core collection
of Uruguayan maize landraces based on morphological traits. Genetic
Resources and Crop Evolution, 2001, 48(4): 381-390.
[16] Upadhyaya H D, Pundir R P S, Dwivedi S L, Gowda C L L, Reddy V
G, Singh S. Developing a mini core collection of sorghum for
diversified utilization of germplasm. Crop Science, 2009, 49(5):
1769-1780.
[17] Upadhyaya H D, Sarma N D R K, Ravishankar C R, Albrecht T,
Narasimhudu Y, Singh S K, Varshney S K, Reddy V G, Singh S,
Dwivedi S L, Wanyera N, Oduori C O A, Mgonja M A, Kisandu D B,
Parzies H K, Gowda C L L. Developing a mini-core collection in
finger millet using multilocation data. Crop Science, 2010, 50(5):
1924-1931.
[18] 邱丽娟, 李英慧, 关荣霞, 刘章雄, 王丽侠, 常汝镇. 大豆核心种
质和微核心种质的构建、验证与研究进展. 作物学报, 2009, 35(4):
571-579.
Qiu L J, Li Y H, Guan R X, Liu Z X, Wang L X, Chang R Z.
Establishment, representative testing and research progress of soybean
core collection and mini core collection. Acta Agronomica Sinica,
2009, 35(4): 571-579. (in Chinese)
[19] Oliveira M F, Nelson R L, Geraldi I O, Cruz C D, de Toledo J F F.

17 期 贾继增等:作物基因组学与作物科学革命
Establishing a soybean germplasm core collection. Field Crops
Research, 2010, 119(2/3): 277-289.
[20] Kaga A, Shimizu T, Watanabe S, Tsubokura Y, Katayose Y, Harada K,
Vaughan D A, Tomooka N. Evaluation of soybean germplasm
conserved in NIAS genebank and development of mini core
collections. Breeding Science, 2012, 61(5): 566-592.
[21] 赵丽梅, 董英山, 刘宝, 郝水, 王克晶, 李向华. 中国一年生野生
大豆(Glycine soja)核心资源构建. 科学通报, 2005, 50(10): 992-999.
Zhao L M, Dong Y S, Liu B, Hao S, Wang K J, Li X H.
Establishment of a core collection for the Chinese annual wild
soybean (Glycine Soja). Chinese Science Bulletin, 2005, 50(10):
992-999. (in Chinese)
[22] Cho G T, Yoon M S, Lee J, Baek H J, Kang J H, Kim T S, Paek N C.
Development of a core set of Korean soybean landraces [Glycine max
(L.) Merr.]. Journal Of Crop Science and Biotechnology, 2008, 11(3):
157-162.
[23] Holbrook C C, Anderson W F, Pittman R N. Selection of a core
collection from the U.S. germplasm collection of peanut. Crop
Science, 1993, 33(4): 859-861.
[24] Holbrook C C, Dong W. Development and evaluation of a mini core
collection for the U.S. peanut germplasm collection. Crop Science,
2005, 45(4): 1540-1544.
[25] 姜慧芳, 任小平, 黄家权, 廖伯寿, 雷永. 中国花生小核心种质的
建立及高油酸基因源的发掘. 中国油料作物学报, 2008, 30(3):
294-299.
Jiang H F, Ren X P, Huang J Q, Liao B S, Lei Y. Establishment of
peanut mini core collection in China and exploration of new resource
with high oleat. Chinese Journal of Oil Crop Sciences, 2008, 30(3):
294-299. (in Chinese)
[26] 姜慧芳 , 任小平 , 廖伯寿 , 黄家权 , 雷永 , 陈本银 , Guo B Z,
Holbrook C C, Upadhyaya H D. 中国花生核心种质的建立及与
ICRISAT 花生微核心种质的比较. 作物学报, 2008, 34(1): 25-30.
Jiang H F, Ren X P, Liao B S, Huang J Q, Lei Y, Chen B Y, Guo B Z,
Holbrook C C, Upadhyaya H D. Peanut core collection established in
China and compared with ICRISAT mini core collection. Acta
Agronomica Sinica, 2008, 34(1): 25-30. (in Chinese)
[27] Upadhyaya H, Ortiz R, Bramel P, Singh S. Development of a
groundnut core collection using taxonomical, geographical and
morphological descriptors. Genetic Resources and Crop Evolution,
2003, 50(2): 139-148.
[28] Upadhyaya H D, Bramel P J, Ortiz R, Singh S. Developing a mini
core of peanut for utilization of genetic resources. Crop Science, 2002,
42(6): 2150-2156.
[29] Belaj A, Dominguez-García M, Atienza S, Martín Urdíroz N, De la
Rosa R, Satovic Z, Martín A, Kilian A, Trujillo I, Valpuesta V, Del Río
C. Developing a core collection of olive (Olea europaea L.) based on
molecular markers (DArTs, SSRs, SNPs) and agronomic traits. Tree
Genetics & Genomes, 2012, 8(2): 365-378.
[30] Mei Y, Zhou J, Xu H, Zhu S. Development of sea island cotton
(Gossypium barbadense L.)Core collection using genotypic values.
Australian Journal of Crop Science 2012, 6(4): 673-680.
[31] Xu H, Mei Y, Hu J, Zhu J, Gong P. Sampling a core collection of
island cotton (Gossypium barbadense L.) based on the genotypic
values of fiber traits. Genetic Resources and Crop Evolution, 2006,
53(3): 515-521.
[32] Diederichsen A, Kusters P, Kessler D, Bainas Z, Gugel R. Assembling
a core collection from the flax world collection maintained by Plant
Gene Resources of Canada. Genetic Resources and Crop Evolution,
2013, 60(4): 1479-1485.
[33] Pino Del Carpio D, Basnet R K, De Vos R C, Maliepaard C, Visser R,
Bonnema G. The patterns of population differentiation in a Brassica
rapa core collection. Theoretical and Applied Genetics, 2011, 122(6):
1105-1118.
[34] Zhao J, Artemyeva A, Del Carpio D P, Basnet R K, Zhang N, Gao J,
Li F, Bucher J, Wang X, Visser R G, Bonnema G. Design of a Brassica
rapa core collection for association mapping studies. Genome, 2010,
53(11): 884-898.
[35] Kang C W, Kim S Y, Lee S W, Mathur P N, Hodgkin T, Zhou M D,
Lee J R. Selection of a core collection of korean sesame germplasm
by a stepwise clustering method. Breeding Science, 2006, 56(1):
85-91.
[36] Zhang X R, Zhao Y Z, Cheng Y, Feng X Y, Guo Q Y, Zhou M D,
Hodgkin T. Establishment of sesame germplasm core collection in
China. Genetic Resources and Crop Evolution, 2000, 47(3): 273-279.
[37] Bisht I S, Mahajan R K, Loknathan T R, Agrawal R C. Diversity in
Indian sesame collection and stratification of germplasm accessions in
different diversity groups. Genetic Resources and Crop Evolution,
1998, 45(4): 325-335.
[38] 姜俊烨, 杨涛, 王芳, 方俐, 仲伟文, 关建平, 宗绪晓. 国内外蚕豆
核心种质SSR遗传多样性对比及微核心种质构建. 作物学报, 2014,
40(7): 1311-1319.
Jiang J Y, Yang T, Wang F, Fang L, Zhong W W, Guan J P, Zong X X.
Genetic diversity analysis of germplasm resources and construction of
mini-core collections for Vicia faba L. at home and abroad. Acta
Agronomica Sinica, 2014, 40(7): 1311-1319. (in Chinese)
[39] Upadhyaya H D, Reddy K N, Gowda C L L, Singh S. Phenotypic

中 国 农 业 科 学 48 卷
diversity in the pigeonpea (Cajanus cajan) core collection. Genetic
Resources and Crop Evolution, 2007, 54(6): 1167-1184.
[40] Upadhyaya H D, Reddy L J, Gowda C L L, Reddy K N, Singh S.
Development of a mini core subset for enhanced and diversified
utilization of pigeonpea germplasm resources. Crop Science, 2006,
46(5): 2127-2132.
[41] Upadhyaya H D, Bramel P J, Singh S. Development of a chickpea
core subset using geographic distribution and quantitative traits. Crop
Science, 2001, 41(1): 206-210.
[42] Upadhyaya H D, Ortiz R. A mini core subset for capturing diversity
and promoting utilization of chickpea genetic resources in crop
improvement. Theoretical and Applied Genetics, 2001, 102(8):
1292-1298.
[43] Kwon S-J, Brown A, Hu J, McGee R, Watt C, Kisha T,
Timmerman-Vaughan G, Grusak M, McPhee K, Coyne C. Genetic
diversity, population structure and genome-wide marker-trait
association analysis emphasizing seed nutrients of the USDA pea
(Pisum sativum L.) core collection. Genes & Genomics, 2012, 34(3):
305-320.
[44] Ortiz R, Ruiz-Tapia E N, Mujica-Sanchez A. Sampling strategy for a
core collection of Peruvian quinoa germplasm. Theoretical and
Applied Genetics, 1998, 96(3/4): 475-483.
[45] Rao E S, Kadirvel P, Symonds R C, Geethanjali S, Ebert A W. Using
SSR markers to map genetic diversity and population structure of
Solanum pimpinellifolium for development of a core collection. Plant
Genetic Resources, 2012, 10(1): 38-48.
附表 3: [1] Ishikawa S, Maekawa M, Arite T, Onishi K, Takamure I, Kyozuka J.
Suppression of tiller bud activity in tillering dwarf mutants of rice.
Plant & Cell Physiology, 2005, 46(1): 79-86.
[2] Arite T, Iwata H, Ohshima K, Maekawa M, Nakajima M, Kojima M,
Sakakibara H, Kyozuka J. DWARF10, an RMS1/MAX4/DAD1
ortholog, controls lateral bud outgrowth in rice. The Plant Journal,
2007, 51(6): 1019-1029.
[3] Nakamura H, Xue Y L, Miyakawa T, Hou F, Qin H M, Fukui K, Shi X,
Ito E, Ito S, Park S H, Miyauchi Y, Asano A, Totsuka N, Ueda T,
Tanokura M, Asami T. Molecular mechanism of strigolactone
perception by DWARF14. Nature Communications, 2013, 4: 2613.
[4] Lin H, Wang R, Qian Q, Yan M, Meng X, Fu Z, Yan C, Jiang B, Su Z,
Li J, Wang Y. DWARF27, an iron-containing protein required for the
biosynthesis of strigolactones, regulates rice tiller bud outgrowth. The
Plant Cell, 2009, 21(5): 1512-1525.
[5] Jiang L, Liu X, Xiong G, Liu H, Chen F, Wang L, Meng X, Liu G, Yu
H, Yuan Y, Yi W, Zhao L, Ma H, He Y, Wu Z, Melcher K, Qian Q, Xu
H E, Wang Y, Li J. DWARF 53 acts as a repressor of strigolactone
signalling in rice. Nature, 2013, 504(7480): 401- 405.
[6] Zhou F, Lin Q, Zhu L, Ren Y, Zhou K, Shabek N, Wu F, Mao H, Dong
W, Gan L, Ma W, Gao H, Chen J, Yang C, Wang D, Tan J, Zhang X,
Guo X, Wang J, Jiang L, Liu X, Chen W, Chu J, Yan C, Ueno K, Ito S,
Asami T, Cheng Z, Wang J, Lei C, Zhai H, Wu C, Wang H, Zheng N,
Wan J. D14-SCFD3-dependent degradation of D53 regulates
strigolactone signalling. Nature, 2013, 504(7480): 406-410.
[7] Zou J, Zhang S, Zhang W, Li G, Chen Z, Zhai W, Zhao X, Pan X, Xie
Q, Zhu L. The rice HIGH-TILLERING DWARF1 encoding an ortholog
of Arabidopsis MAX3 is required for negative regulation of the
outgrowth of axillary buds. The Plant Journal, 2006, 48(5): 687-698.
[8] Komatsu K, Maekawa M, Ujiie S, Satake Y, Furutani I, Okamoto H,
Shimamoto K, Kyozuka J. LAX and SPA: major regulators of shoot
branching in rice. Proceedings of the National Academy of Sciences of
the United States of America, 2003, 100(20): 11765-11770.
[9] Li X, Qian Q, Fu Z, Wang Y, Xiong G, Zeng D, Wang X, Liu X, Teng
S, Hiroshi F, Yuan M, Luo D, Han B, Li J. Control of tillering in rice.
Nature, 2003, 422(6932): 618-621.
[10] Xu C, Wang Y, Yu Y, Duan J, Liao Z, Xiong G, Meng X, Liu G, Qian
Q, Li J. Degradation of MONOCULM 1 by APC/C(TAD1) regulates
rice tillering. Nature Communications, 2012, 3: 750.
[11] Jiao Y, Wang Y, Xue D, Wang J, Yan M, Liu G, Dong G, Zeng D, Lu Z,
Zhu X, Qian Q, Li J. Regulation of OsSPL14 by OsmiR156 defines
ideal plant architecture in rice. Nature Genetics, 2010, 42(6): 541-544.
[12] Miura K, Ikeda M, Matsubara A, Song X J, Ito M, Asano K, Matsuoka
M, Kitano H, Ashikari M. OsSPL14 promotes panicle branching and
higher grain productivity in rice. Nature Genetics, 2010, 42(6):
545-549.
[13] Wu W, Zheng X M, Lu G, Zhong Z, Gao H, Chen L, Wu C, Wang H J,
Wang Q, Zhou K, Wang J L, Wu F, Zhang X, Guo X, Cheng Z, Lei C,
Lin Q, Jiang L, Wang H, Ge S, Wan J. Association of functional
nucleotide polymorphisms at DTH2 with the northward expansion of
rice cultivation in Asia. Proceedings of the National Academy of
Sciences of the United States of America, 2013, 110(8): 2775-2780.
[14] Bian X F, Liu X, Zhao Z G, Jiang L, Gao H, Zhang Y H, Zheng M,
Chen L M, Liu S J, Zhai H Q, Wan J M. Heading date gene, dth3
controlled late flowering in O. Glaberrima Steud. by down-regulating
Ehd1. Plant Cell Reports, 2011, 30(12): 2243-2254.
[15] Gao H, Jin M, Zheng X M, Chen J, Yuan D, Xin Y, Wang M, Huang D,
Zhang Z, Zhou K, Sheng P, Ma J, Ma W, Deng H, Jiang L, Liu S,

17 期 贾继增等:作物基因组学与作物科学革命
Wang H, Wu C, Yuan L, Wan J. Days to heading 7, a major
quantitative locus determining photoperiod sensitivity and regional
adaptation in rice. Proceedings of the National Academy of Sciences of
the United States of America, 2014, 111(46): 16337-16342.
[16] Wei X, Xu J, Guo H, Jiang L, Chen S, Yu C, Zhou Z, Hu P, Zhai H,
Wan J. DTH8 suppresses flowering in rice, influencing plant height
and yield potential simultaneously. Plant Physiology, 2010, 153(4):
1747-1758.
[17] Matsubara K, Yamanouchi U, Wang Z X, Minobe Y, Izawa T, Yano M.
Ehd2, a rice ortholog of the maize INDETERMINATE1 gene,
promotes flowering by up-regulating Ehd1. Plant Physiology, 2008,
148(3): 1425-1435.
[18] Matsubara K, Yamanouchi U, Nonoue Y, Sugimoto K, Wang Z X,
Minobe Y, Yano M. Ehd3, encoding a plant homeodomain
finger-containing protein, is a critical promoter of rice flowering. The
Plant Journal, 2011, 66(4): 603-612.
[19] Gao H, Zheng X M, Fei G, Chen J, Jin M, Ren Y, Wu W, Zhou K,
Sheng P, Zhou F, Jiang L, Wang J, Zhang X, Guo X, Wang J L, Cheng
Z, Wu C, Wang H, Wan J M. Ehd4 encodes a novel and Oryza-
genus-specific regulator of photoperiodic flowering in rice. PLoS
Genetics, 2013, 9(2): e1003281.
[20] Xue W, Xing Y, Weng X, Zhao Y, Tang W, Wang L, Zhou H, Yu S, Xu
C, Li X, Zhang Q. Natural variation in Ghd7 is an important regulator
of heading date and yield potential in rice. Nature Genetics, 2008,
40(6): 761-767.
[21] Yan W H, Wang P, Chen H X, Zhou H J, Li Q P, Wang C R, Ding Z H,
Zhang Y S, Yu S B, Xing Y Z, Zhang Q F. A major QTL, Ghd8, plays
pleiotropic roles in regulating grain productivity, plant height, and
heading date in rice. Molecular Plant, 2011, 4(2): 319-330.
[22] Takahashi Y, Shimamoto K. Heading date 1 (Hd1), an ortholog of
Arabidopsis CONSTANS, is a possible target of human selection
during domestication to diversify flowering times of cultivated rice.
Genes & Genetic Systems, 2011, 86(3): 175-182.
[23] Kojima S, Takahashi Y, Kobayashi Y, Monna L, Sasaki T, Araki T,
Yano M. Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes
transition to flowering downstream of Hd1 under short-day conditions.
Plant & Cell Physiology, 2002, 43(10): 1096-1105.
[24] Takahashi Y, Shomura A, Sasaki T, Yano M. Hd6, a rice quantitative
trait locus involved in photoperiod sensitivity, encodes the alpha
subunit of protein kinase CK2. Proceedings of the National Academy
of Sciences of the United States of America, 2001, 98(14): 7922-7927.
[25] Hori K, Ogiso-Tanaka E, Matsubara K, Yamanouchi U, Ebana K,
Yano M. Hd16, a gene for casein kinase I, is involved in the control of
rice flowering time by modulating the day-length response. The Plant
Journal, 2013, 76(1): 36-46.
[26] Matsubara K, Ogiso-Tanaka E, Hori K, Ebana K, Ando T, Yano M.
Natural variation in Hd17, a homolog of Arabidopsis ELF3 that is
involved in rice photoperiodic flowering. Plant & Cell Physiology,
2012, 53(4): 709-716.
[27] Konishi S, Izawa T, Lin S Y, Ebana K, Fukuta Y, Sasaki T, Yano M.
An SNP caused loss of seed shattering during rice domestication.
Science, 2006, 312(5778): 1392-1396.
[28] Sugimoto K, Takeuchi Y, Ebana K, Miyao A, Hirochika H, Hara N,
Ishiyama K, Kobayashi M, Ban Y, Hattori T, Yano M. Molecular
cloning of Sdr4, a regulator involved in seed dormancy and
domestication of rice. Proceedings of the National Academy of
Sciences of the United States of America, 2010, 107(13): 5792-5797.
[29] Li C, Zhou A, Sang T. Rice domestication by reducing shattering.
Science, 2006, 311(5769): 1936-9.
[30] Wang Z Y, Zheng F Q, Shen G Z, Gao J P, Snustad D P, Li M G,
Zhang J L, Hong M M. The amylose content in rice endosperm is
related to the post-transcriptional regulation of the waxy gene. The
Plant Journal, 1995, 7(4): 613-622.
[31] Bradbury L M, Fitzgerald T L, Henry R J, Jin Q, Waters D L. The
gene for fragrance in rice. Plant Biotechnology Journal, 2005, 3(3):
363-370.
[32] Ashikari M, Sakakibara H, Lin S, Yamamoto T, Takashi T, Nishimura
A, Angeles E R, Qian Q, Kitano H, Matsuoka M. Cytokinin oxidase
regulates rice grain production. Science, 2005, 309(5735): 741-745.
[33] Huang X, Qian Q, Liu Z, Sun H, He S, Luo D, Xia G, Chu C, Li J, Fu
X. Natural variation at the DEP1 locus enhances grain yield in rice.
Nature Genetics, 2009, 41(4): 494-497.
[34] Yang W, Gao M, Yin X, Liu J, Xu Y, Zeng L, Li Q, Zhang S, Wang J,
Zhang X, He Z. Control of rice embryo development, shoot apical
meristem maintenance, and grain yield by a novel cytochrome p450.
Molecular Plant, 2013, 6(6): 1945-1960.
[35] Kurakawa T, Ueda N, Maekawa M, Kobayashi K, Kojima M, Nagato
Y, Sakakibara H, Kyozuka J. Direct control of shoot meristem activity
by a cytokinin-activating enzyme. Nature, 2007, 445(7128): 652-655.
[36] Wang E, Wang J, Zhu X, Hao W, Wang L, Li Q, Zhang L, He W, Lu B,
Lin H, Ma H, Zhang G, He Z. Control of rice grain-filling and yield by
a gene with a potential signature of domestication. Nature Genetics,
2008, 40(11): 1370-1374.
[37] Mao H, Sun S, Yao J, Wang C, Yu S, Xu C, Li X, Zhang Q. Linking
differential domain functions of the GS3 protein to natural variation of
grain size in rice. Proceedings of the National Academy of Sciences of

中 国 农 业 科 学 48 卷
the United States of America, 2010, 107(45): 19579-19584.
[38] Li Y, Fan C, Xing Y, Jiang Y, Luo L, Sun L, Shao D, Xu C, Li X, Xiao
J, He Y, Zhang Q. Natural variation in GS5 plays an important role in
regulating grain size and yield in rice. Nature Genetics, 2011, 43(12):
1266-1269.
[39] Song X J, Huang W, Shi M, Zhu M Z, Lin H X. A QTL for rice grain
width and weight encodes a previously unknown RING-type E3
ubiquitin ligase. Nature Genetics, 2007, 39(5): 623-630.
[40] Weng J, Gu S, Wan X, Gao H, Guo T, Su N, Lei C, Zhang X, Cheng Z,
Guo X, Wang J, Jiang L, Zhai H, Wan J. Isolation and initial
characterization of GW5, a major QTL associated with rice grain
width and weight. Cell Research, 2008, 18(12): 1199-1209.
[41] Wang S, Wu K, Yuan Q, Liu X, Liu Z, Lin X, Zeng R, Zhu H, Dong G,
Qian Q, Zhang G, Fu X. Control of grain size, shape and quality by
OsSPL16 in rice. Nature Genetics, 2012, 44(8): 950-954.
[42] Shomura A, Izawa T, Ebana K, Ebitani T, Kanegae H, Konishi S, Yano
M. Deletion in a gene associated with grain size increased yields
during rice domestication. Nature Genetics, 2008, 40(8): 1023-1028.
[43] Li S, Qian Q, Fu Z, Zeng D, Meng X, Kyozuka J, Maekawa M, Zhu X,
Zhang J, Li J, Wang Y. Short panicle1 encodes a putative PTR family
transporter and determines rice panicle size. The Plant Journal, 2009,
58(4): 592-605.
[44] Zhu K, Tang D, Yan C, Chi Z, Yu H, Chen J, Liang J, Gu M, Cheng Z.
Erect panicle2 encodes a novel protein that regulates panicle erectness
in indica rice. Genetics, 2010, 184(2): 343-350.
[45] Ashikari M, Wu J, Yano M, Sasaki T, Yoshimura A. Rice gibberellin-
insensitive dwarf mutant gene Dwarf 1 encodes the alpha-subunit of
GTP-binding protein. Proceedings of the National Academy of
Sciences of the United States of America, 1999, 96(18): 10284-10289.
[46] Spielmeyer W, Ellis M H, Chandler P M. Semidwarf (sd-1), "green
revolution" rice, contains a defective gibberellin 20-oxidase gene.
Proceedings of the National Academy of Sciences of the United States
of America, 2002, 99(13): 9043-9048.
[47] Dong X, Wang X, Zhang L, Yang Z, Xin X, Wu S, Sun C, Liu J, Yang
J, Luo X. Identification and characterization of OsEBS, a gene
involved in enhanced plant biomass and spikelet number in rice. Plant
Biotechnology Journal, 2013, 11(9): 1044-1057.
[48] Wang W, Li G, Zhao J, Chu H, Lin W, Zhang D, Wang Z, Liang W.
Dwarf Tiller1, a Wuschel-related homeobox transcription factor, is
required for tiller growth in rice. PLoS Genetics, 2014, 10(3):
e1004154.
[49] Jin J, Huang W, Gao J P, Yang J, Shi M, Zhu M Z, Luo D, Lin H X.
Genetic control of rice plant architecture under domestication. Nature
Genetics, 2008, 40(11): 1365-1369.
[50] Tan L, Li X, Liu F, Sun X, Li C, Zhu Z, Fu Y, Cai H, Wang X, Xie D,
Sun C. Control of a key transition from prostrate to erect growth in
rice domestication. Nature Genetics, 2008, 40(11): 1360-1364.
[51] Qi J, Qian Q, Bu Q, Li S, Chen Q, Sun J, Liang W, Zhou Y, Chu C, Li
X, Ren F, Palme K, Zhao B, Chen J, Chen M, Li C. Mutation of the
rice Narrow leaf1 gene, which encodes a novel protein, affects vein
patterning and polar auxin transport. Plant Physiology, 2008, 147(4):
1947-1959.
[52] Fujita D, Trijatmiko K R, Tagle A G, Sapasap M V, Koide Y, Sasaki K,
Tsakirpaloglou N, Gannaban R B, Nishimura T, Yanagihara S, Fukuta
Y, Koshiba T, Slamet-Loedin I H, Ishimaru T, Kobayashi N. NAL1
allele from a rice landrace greatly increases yield in modern indica
cultivars. Proceedings of the National Academy of Sciences of the
United States of America, 2013, 110(51): 20431-20436.
[53] Uga Y, Sugimoto K, Ogawa S, Rane J, Ishitani M, Hara N, Kitomi Y,
Inukai Y, Ono K, Kanno N, Inoue H, Takehisa H, Motoyama R,
Nagamura Y, Wu J, Matsumoto T, Takai T, Okuno K, Yano M. Control
of root system architecture by DEEPER ROOTING 1 increases rice
yield under drought conditions. Nature Genetics, 2013, 45(9):
1097-1102.
[54] Gross B L, Skare K J, Olsen K M. Novel Phr1 mutations and the
evolution of phenol reaction variation in US weedy rice (Oryza sativa).
The New Phytologist, 2009, 184(4): 842-850.
[55] Yu Y, Tang T, Qian Q, Wang Y, Yan M, Zeng D, Han B, Wu C I, Shi S,
Li J. Independent losses of function in a polyphenol oxidase in rice:
differentiation in grain discoloration between subspecies and the role
of positive selection under domestication. The Plant Cell, 2008,
20(11): 2946-2959.
[56] Sweeney M T, Thomson M J, Pfeil B E, McCouch S. Caught
red-handed: Rc encodes a basic helix-loop-helix protein conditioning
red pericarp in rice. The Plant Cell, 2006, 18(2): 283-294.
[57] Sweeney M T, Thomson M J, Cho Y G, Park Y J, Williamson S H,
Bustamante C D, McCouch S R. Global dissemination of a single
mutation conferring white pericarp in rice. PLoS Genetics, 2007, 3(8):
e133.
[58] Wang C, Zhang X, Fan Y, Gao Y, Zhu Q, Zheng C, Qin T, Li Y, Che J,
Zhang M, Yang B, Liu Y, Zhao K. XA23 is an executorr protein and
confers broad-spectrum disease resistance in rice. Molecular Plant,
2015, 8(2): 290-302.
[59] Sun X, Cao Y, Yang Z, Xu C, Li X, Wang S, Zhang Q. Xa26, a gene
conferring resistance to Xanthomonas oryzae pv. oryzae in rice,
encodes an LRR receptor kinase-like protein. The Plant Journal, 2004,

17 期 贾继增等:作物基因组学与作物科学革命
37(4): 517-527.
[60] Lin F, Chen S, Que Z, Wang L, Liu X, Pan Q. The blast resistance
gene Pi37 encodes a nucleotide binding site leucine-rich repeat
protein and is a member of a resistance gene cluster on rice
chromosome 1. Genetics, 2007, 177(3): 1871-1880.
[61] Wang Z X, Yano M, Yamanouchi U, Iwamoto M, Monna L, Hayasaka
H, Katayose Y, Sasaki T. The Pib gene for rice blast resistance belongs
to the nucleotide binding and leucine-rich repeat class of plant disease
resistance genes. The Plant Journal, 1999, 19(1): 55-64.
[62] Ren Z H, Gao J P, Li L G, Cai X L, Huang W, Chao D Y, Zhu M Z,
Wang Z Y, Luan S, Lin H X. A rice quantitative trait locus for salt
tolerance encodes a sodium transporter. Nature Genetics, 2005, 37(10):
1141-1146.
[63] Lu G, Wu F Q, Wu W, Wang H J, Zheng X M, Zhang Y, Chen X, Zhou
K, Jin M, Cheng Z, Li X, Jiang L, Wang H, Wan J. Rice LTG1 is
involved in adaptive growth and fitness under low ambient
temperature. The Plant Journal, 2014, 78(3): 468-480.
[64] Ueda T, Sato T, Hidema J, Hirouchi T, Yamamoto K, Kumagai T, Yano
M. qUVR-10, a major quantitative trait locus for ultraviolet-B
resistance in rice, encodes cyclobutane pyrimidine dimer photolyase.
Genetics, 2005, 171(4): 1941-1950.
[65] Xu K, Xu X, Fukao T, Canlas P, Maghirang-Rodriguez R, Heuer S,
Ismail A M, Bailey-Serres J, Ronald P C, Mackill D J. Sub1A is an
ethylene-response-factor-like gene that confers submergence tolerance
to rice. Nature, 2006, 442(7103): 705-708.
[66] Ma Y D X, Xu Y, Luo W, Zheng X, Zeng D, Pan Y, Lin X, Liu H,
Zhang D, Xiao J, Guo X, Xu S, Niu Y, Jin J, Zhang H, Xu X, Li L,
Wang W, Qian Q, Ge S, Chong K. COLD1 confers chilling tolerance
in rice. Cell, 2015, 160: 13.
[67] Che L, Tang D, Wang K, Wang M, Zhu K, Yu H, Gu M, Cheng Z.
OsAM1 is required for leptotene-zygotene transition in rice. Cell
Research, 2011, 21(4): 654-665.
[68] Periyannan S, Moore J, Ayliffe M, Bansal U, Wang X, Huang L, Deal
K, Luo M, Kong X, Bariana H, Mago R, McIntosh R, Dodds P,
Dvorak J, Lagudah E. The gene Sr33, an ortholog of barley Mla genes,
encodes resistance to wheat stem rust race Ug99. Science, 2013,
341(6147): 786-788.
[69] Saintenac C, Zhang W, Salcedo A, Rouse M N, Trick H N, Akhunov E,
Dubcovsky J. Identification of wheat gene Sr35 that confers resistance
to Ug99 stem rust race group. Science, 2013, 341(6147): 783-786.
[70] Huang L, Brooks S A, Li W, Fellers J P, Trick H N, Gill B S.
Map-based cloning of leaf rust resistance gene Lr21 from the large
and polyploid genome of bread wheat. Genetics, 2003, 164(2):
655-664.
[71] Feuillet C, Travella S, Stein N, Albar L, Nublat A, Keller B.
Map-based isolation of the leaf rust disease resistance gene Lr10 from
the hexaploid wheat (Triticum aestivum L.) genome. Proceedings of
the National Academy of Sciences of the United States of America,
2003, 100(25): 15253-15258.
[72] Cloutier S, McCallum B D, Loutre C, Banks T W, Wicker T, Feuillet
C, Keller B, Jordan M C. Leaf rust resistance gene Lr1, isolated from
bread wheat (Triticum aestivum L.) is a member of the large psr567
gene family. Plant Molecular Biology, 2007, 65(1/2): 93-106.
[73] Krattinger S G, Lagudah E S, Spielmeyer W, Singh R P, Huerta-
Espino J, McFadden H, Bossolini E, Selter L L, Keller B. A putative
ABC transporter confers durable resistance to multiple fungal
pathogens in wheat. Science, 2009, 323(5919): 1360-1363.
[74] Fu D, Uauy C, Distelfeld A, Blechl A, Epstein L, Chen X, Sela H,
Fahima T, Dubcovsky J. A kinase-START gene confers temperature-
dependent resistance to wheat stripe rust. Science, 2009, 323(5919):
1357-1360.
[75] Yahiaoui N, Srichumpa P, Dudler R, Keller B. Genome analysis at
different ploidy levels allows cloning of the powdery mildew
resistance gene Pm3b from hexaploid wheat. The Plant Journal, 2004,
37(4): 528-538.
[76] Qin B, Cao A, Wang H, Chen T, You F M, Liu Y, Ji J, Liu D, Chen P,
Wang X E. Collinearity-based marker mining for the fine mapping of
Pm6, a powdery mildew resistance gene in wheat. Theoretical and
Applied Genetics, 2011, 123(2): 207-218.
[77] Peng J, Richards D E, Hartley N M, Murphy G P, Devos K M,
Flintham J E, Beales J, Fish L J, Worland A J, Pelica F, Sudhakar D,
Christou P, Snape J W, Gale M D, Harberd N P. 'Green revolution'
genes encode mutant gibberellin response modulators. Nature, 1999,
400(6741): 256-261.
[78] Yan L, Loukoianov A, Tranquilli G, Helguera M, Fahima T,
Dubcovsky J. Positional cloning of the wheat vernalization gene
VRN1. Proceedings of the National Academy of Sciences of the United
States of America, 2003, 100(10): 6263-6268.
[79] Yan L, Loukoianov A, Blechl A, Tranquilli G, Ramakrishna W,
SanMiguel P, Bennetzen J L, Echenique V, Dubcovsky J. The wheat
VRN2 gene is a flowering repressor down-regulated by vernalization.
Science, 2004, 303(5664): 1640-1644.
[80] Yan L, Fu D, Li C, Blechl A, Tranquilli G, Bonafede M, Sanchez A,
Valarik M, Yasuda S, Dubcovsky J. The wheat and barley
vernalization gene VRN3 is an orthologue of FT. Proceedings of the
National Academy of Sciences of the United States of America, 2006,

中 国 农 业 科 学 48 卷
103(51): 19581-19586.
[81] Uauy C, Distelfeld A, Fahima T, Blechl A, Dubcovsky J. A NAC Gene
regulating senescence improves grain protein, zinc, and iron content in
wheat. Science, 2006, 314(5803): 1298-1301.
[82] Griffiths S, Sharp R, Foote T N, Bertin I, Wanous M, Reader S, Colas
I, Moore G. Molecular characterization of Ph1 as a major chromosome
pairing locus in polyploid wheat. Nature, 2006, 439(7077): 749-752.
[83] Simons K J, Fellers J P, Trick H N, Zhang Z, Tai Y S, Gill B S, Faris J
D. Molecular characterization of the major wheat domestication gene
Q. Genetics, 2006, 172(1): 547-555.
[84] Liu S, Sehgal S K, Li J, Lin M, Trick H N, Yu J, Gill B S, Bai G.
Cloning and characterization of a critical regulator for preharvest
sprouting in wheat. Genetics, 2013, 195(1): 263-273.
[85] Komatsuda T, Pourkheirandish M, He C, Azhaguvel P, Kanamori H,
Perovic D, Stein N, Graner A, Wicker T, Tagiri A, Lundqvist U,
Fujimura T, Matsuoka M, Matsumoto T, Yano M. Six-rowed barley
originated from a mutation in a homeodomain-leucine zipper I-class
homeobox gene. Proceedings of the National Academy of Sciences of
the United States of America, 2007, 104(4): 1424-1429.
[86] Brueggeman R, Rostoks N, Kudrna D, Kilian A, Han F, Chen J, Druka
A, Steffenson B, Kleinhofs A. The barley stem rust-resistance gene
Rpg1 is a novel disease-resistance gene with homology to receptor
kinases. Proceedings of the National Academy of Sciences of the
United States of America, 2002, 99(14): 9328-9333.
[87] Brueggeman R, Druka A, Nirmala J, Cavileer T, Drader T, Rostoks N,
Mirlohi A, Bennypaul H, Gill U, Kudrna D, Whitelaw C, Kilian A,
Han F, Sun Y, Gill K, Steffenson B, Kleinhofs A. The stem rust
resistance gene Rpg5 encodes a protein with nucleotide-binding-site,
leucine-rich, and protein kinase domains. Proceedings of the National
Academy of Sciences of the United States of America, 2008, 105(39):
14970-14975.
[88] Turner A, Beales J, Faure S, Dunford R P, Laurie D A. The
pseudo-response regulator Ppd-H1 provides adaptation to photoperiod
in barley. Science, 2005, 310(5750): 1031-1034.
[89] Chen G, Komatsuda T, Ma J F, Nawrath C, Pourkheirandish M, Tagiri
A, Hu Y G, Sameri M, Li X, Zhao X, Liu Y, Li C, Ma X, Wang A,
Nair S, Wang N, Miyao A, Sakuma S, Yamaji N, Zheng X, Nevo E.
An ATP-binding cassette subfamily G full transporter is essential for
the retention of leaf water in both wild barley and rice. Proceedings of
the National Academy of Sciences of the United States of America,
2011, 108(30): 12354-12359.
[90] Zuo W, Chao Q, Zhang N, Ye J, Tan G, Li B, Xing Y, Zhang B, Liu H,
Fengler K A, Zhao J, Zhao X, Chen Y, Lai J, Yan J, Xu M. A maize
wall-associated kinase confers quantitative resistance to head smut.
Nature Genetics, 2015, 47(2): 151-157.
[91] Doebley J, Stec A, Hubbard L. The evolution of apical dominance in
maize. Nature, 1997, 386(6624): 485-488.
[92] Wang R L, Stec A, Hey J, Lukens L, Doebley J. The limits of selection
during maize domestication. Nature, 1999, 398(6724): 236-239.
[93] Wang H, Nussbaum-Wagler T, Li B, Zhao Q, Vigouroux Y, Faller M,
Bomblies K, Lukens L, Doebley J F. The origin of the naked grains of
maize. Nature, 2005, 436(7051): 714-719.
[94] Chuck G, Muszynski M, Kellogg E, Hake S, Schmidt R J. The control
of spikelet meristem identity by the branched silkless1 gene in maize.
Science, 2002, 298(5596): 1238-1241.
[95] Yang F, Bui H T, Pautler M, Llaca V, Johnston R, Lee B H, Kolbe A,
Sakai H, Jackson D. A Maize Glutaredoxin Gene, Abphyl2, Regulates
Shoot Meristem Size and Phyllotaxy. The Plant Cell, 2015.
[96] Yi G, Neelakandan A K, Gontarek B C, Vollbrecht E, Becraft P W. The
naked endosperm Genes Encode Duplicate INDETERMINATE
Domain Transcription Factors Required for Maize Endosperm Cell
Patterning and Differentiation. Plant Physiology, 2015, 167(2):
443-456.
[97] Nestler J, Liu S, Wen T J, Paschold A, Marcon C, Tang H M, Li D, Li
L, Meeley R B, Sakai H, Bruce W, Schnable P S, Hochholdinger F.
Roothairless5, which functions in maize (Zea mays L.) root hair
initiation and elongation encodes a monocot-specific NADPH oxidase.
The Plant Journal, 2014, 79(5): 729-740.
[98] Dong Z, Jiang C, Chen X, Zhang T, Ding L, Song W, Luo H, Lai J,
Chen H, Liu R, Zhang X, Jin W. Maize LAZY1 mediates shoot
gravitropism and inflorescence development through regulating auxin
transport, auxin signaling, and light response. Plant Physiology, 2013,
163(3): 1306-1322.
[99] Wang G, Wang F, Wang G, Wang F, Zhang X, Zhong M, Zhang J, Lin
D, Tang Y, Xu Z, Song R. Opaque1 encodes a myosin XI motor
protein that is required for endoplasmic reticulum motility and protein
body formation in maize endosperm. The Plant Cell, 2012, 24(8):
3447-3462.
[100] Miclaus M, Wu Y, Xu J H, Dooner H K, Messing J. The maize
high-lysine mutant opaque7 is defective in an acyl-CoA
synthetase-like protein. Genetics, 2011, 189(4): 1271-1280.
[101] Wang G, Sun X, Wang G, Wang F, Gao Q, Sun X, Tang Y, Chang C,
Lai J, Zhu L, Xu Z, Song R. Opaque7 encodes an acyl-activating
enzyme-like protein that affects storage protein synthesis in maize
endosperm. Genetics, 2011, 189(4): 1281-1295.
[102] Chen K Y, Cong B, Wing R, Vrebalov J, Tanksley S D. Changes in

17 期 贾继增等:作物基因组学与作物科学革命
regulation of a transcription factor lead to autogamy in cultivated
tomatoes. Science, 2007, 318(5850): 643-645.
[103] Frary A, Nesbitt T C, Grandillo S, Knaap E, Cong B, Liu J, Meller
J, Elber R, Alpert K B, Tanksley S D. fw2.2: a quantitative trait locus
key to the evolution of tomato fruit size. Science, 2000, 289(5476):
85-88.
[104] Cong B, Barrero L S, Tanksley S D. Regulatory change in
YABBY-like transcription factor led to evolution of extreme fruit size
during tomato domestication. Nature Genetics, 2008, 40(6): 800-804.
[105] Liu J, Van Eck J, Cong B, Tanksley S D. A new class of regulatory
genes underlying the cause of pear-shaped tomato fruit. Proceedings
of the National Academy of Sciences of the United States of America,
2002, 99(20): 13302-13306.
[106] Fridman E, Carrari F, Liu Y S, Fernie A R, Zamir D. Zooming in on a
quantitative trait for tomato yield using interspecific introgressions.
Science, 2004, 305(5691): 1786-1789.
[107] Schumacher K, Schmitt T, Rossberg M, Schmitz G, Theres K. The
Lateral suppressor (Ls) gene of tomato encodes a new member of the
VHIID protein family. Proceedings of the National Academy of
Sciences of the United States of America, 1999, 96(1): 290-295.
[108] Ariizumi T, Kishimoto S, Kakami R, Maoka T, Hirakawa H, Suzuki Y,
Ozeki Y, Shirasawa K, Bernillon S, Okabe Y, Moing A, Asamizu E,
Rothan C, Ohmiya A, Ezura H. Identification of the carotenoid
modifying gene PALE YELLOW PETAL 1 as an essential factor in
xanthophyll esterification and yellow flower pigmentation in tomato
(Solanum lycopersicum). The Plant Journal, 2014, 79(3): 453-465.
[109] Yan L, Zhai Q, Wei J, Li S, Wang B, Huang T, Du M, Sun J, Kang L,
Li C B, Li C. Role of tomato lipoxygenase D in wound-induced
jasmonate biosynthesis and plant immunity to insect herbivores. PLoS
Genetics, 2013, 9(12): e1003964.
[110] Blas A L, Ming R, Liu Z, Veatch O J, Paull R E, Moore P H, Yu Q.
Cloning of the papaya chromoplast-specific lycopene beta-cyclase,
CpCYC-b, controlling fruit flesh color reveals conserved microsynteny
and a recombination hot spot. Plant Physiology, 2010, 152(4): 2013-2022.
[111] Hovav R, Chehanovsky N, Moy M, Jetter R, Schaffer A A. The
identification of a gene (Cwp1), silenced during Solanum evolution,
which causes cuticle microfissuring and dehydration when expressed
in tomato fruit. The Plant Journal, 2007, 52(4): 627-639.
[112] Cohen S, Itkin M, Yeselson Y, Tzuri G, Portnoy V, Harel-Baja R, Lev
S, Sa'ar U, Davidovitz-Rikanati R, Baranes N, Bar E, Wolf D,
Petreikov M, Shen S, Ben-Dor S, Rogachev I, Aharoni A, Ast T,
Schuldiner M, Belausov E, Eshed R, Ophir R, Sherman A, Frei B,
Neuhaus H E, Xu Y, Fei Z, Giovannoni J, Lewinsohn E, Tadmor Y,
Paris H S, Katzir N, Burger Y, Schaffer A A. The PH gene determines
fruit acidity and contributes to the evolution of sweet melons. Nature
Communications, 2014, 5: 4026.
[113] Boualem A, Fergany M, Fernandez R, Troadec C, Martin A, Morin H,
Sari M A, Collin F, Flowers J M, Pitrat M, Purugganan M D,
Dogimont C, Bendahmane A. A conserved mutation in an ethylene
biosynthesis enzyme leads to andromonoecy in melons. Science, 2008,
321(5890): 836-838.
[114] Martin A, Troadec C, Boualem A, Rajab M, Fernandez R, Morin H,
Pitrat M, Dogimont C, Bendahmane A. A transposon-induced epigenetic
change leads to sex determination in melon. Nature, 2009, 461(7267):
1135-1138.
[115] Kloosterman B, Abelenda J A, Gomez Mdel M, Oortwijn M, de Boer
J M, Kowitwanich K, Horvath B M, van Eck H J, Smaczniak C, Prat S,
Visser R G, Bachem C W. Naturally occurring allele diversity allows
potato cultivation in northern latitudes. Nature, 2013, 495(7440):
246-250.
[116] Liu S, Kandoth P K, Warren S D, Yeckel G, Heinz R, Alden J, Yang C,
Jamai A, El-Mellouki T, Juvale P S, Hill J, Baum T J, Cianzio S,
Whitham S A, Korkin D, Mitchum M G, Meksem K. A soybean cyst
nematode resistance gene points to a new mechanism of plant
resistance to pathogens. Nature, 2012, 492(7428): 256-260.
[117] Guan R, Qu Y, Guo Y, Yu L, Liu Y, Jiang J, Chen J, Ren Y, Liu G, Tian
L, Jin L, Liu Z, Hong H, Chang R, Gilliham M, Qiu L. Salinity
tolerance in soybean is modulated by natural variation in GmSALT3.
The Plant Journal, 2014, 80(6): 937-950.
[118] Funatsuki H, Suzuki M, Hirose A, Inaba H, Yamada T, Hajika M,
Komatsu K, Katayama T, Sayama T, Ishimoto M, Fujino K. Molecular
basis of a shattering resistance boosting global dissemination of
soybean. Proceedings of the National Academy of Sciences of the
United States of America, 2014, 111(50): 17797-17802.

中国农业科学 2015,48(17):3333-3353 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.003
收稿日期:2014-12-16;接受日期:2015-02-09 基金项目:中国农业科学院科技创新工程 联系方式:黎裕,E-mail:[email protected]
基于基因组学的作物种质资源研究:现状与展望
黎 裕,李英慧,杨庆文,张锦鹏,张金梅,邱丽娟,王天宇
(中国农业科学院作物科学研究所,北京 100081)
摘要:作物种质资源工作涉及面很广,包括种质资源的收集、编目、保护、繁种更新、分发利用与信息系统
建立等基础性工作,作物起源、驯化与传播、种质分类、民族植物学与传统知识研究等基础研究,遗传多样性评
价、重要性状表型鉴定、种质资源基因型鉴定、基因发掘和种质创新等应用基础研究。经过近百年的努力,种质
资源的基础性工作已卓有成效,作物种质资源保护与共享利用体系基本建立。但由于主要基于形态学的传统研究
思路和方法存在很多先天不足,种质资源基础研究和应用基础研究一直举步维艰,效率低下。随着分子标记技术
和第二代测序技术的快速发展,基因组学理论和方法不断深入到种质资源研究的多个层面,使种质资源保护和创
新利用发生了研究思路和方法学上的变革。基因组学研究成果为种质资源的有效收集和保护提供了理论指导,也
为阐明作物起源和演化、全面评估种质资源结构多样性提供了核心理论和技术,同时大幅度提高了基因发掘和种
质创新效率。特别是全基因组测序、重测序和简化基因组测序技术不断成熟,使在全基因组水平上比较不同种质
资源基因组变异成为可能;在此基础上,可阐明农作物起源以及驯化、改良和传播对种质资源形成的影响,明确
现有种质资源和野外种质资源群体结构和遗传多样性,提出种质资源异地保存和原生境保护的最佳策略;结合表
型鉴定数据,利用连锁分析和关联分析等基因组学方法,可高效发掘种质资源中蕴含的新基因和有利等位基因,
提出其利用途径和具体方案,并在种质创新过程中充分利用基因组学研究成果提高创新效率。文章评述了基因组
学在种质资源研究中的应用现状,特别是在种质资源基因型鉴定、异地保存和原生境保护、作物起源与进化研究、
结构多样性分析、新基因发掘和种质创新等方面的应用情况及其发展趋势。提出了今后的发展方向和工作重点,
强调把基因组学理论和方法与作物种质资源研究紧密结合,为种质资源的有效保护和高效利用提供强有力的理论、
技术、材料与信息支撑。
关键词:种质资源;基因组学;评述
Genomics-Based Crop Germplasm Research:
Advances and Perspectives LI Yu, LI Ying-hui, YANG Qing-wen, ZHANG Jin-peng, ZHANG Jin-mei, QIU Li-juan, WANG Tian-yu
(Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081)
Abstract: The scale of activities related to crop germplasm is massive, including basic work (germplasm collecting,
documentation, multiplication, conservation, regeneration, distribution and information system establishment, etc.), basic research (studies on crop origin, domestication and dispersal, germplasm classification, ethnobotany, indigenous knowledge, etc.), and applied basic research (genetic diversity assessment, precise phenotypic evaluation of important traits, germplasm genotyping, gene discovery and allele mining, germplasm enhancement etc.). With the efforts in the last century, the basic work on crop germplasm has been very fruitful and the system of crop germplasm conservation and utilization has been established in the world. Because the traditional ideas and methodologies based on morphology had their innate disadvantages, however, the basic research and the applied basic research had developed along at an agonizingly slow pace before genomics appeared. With rapid development of molecular
jia
文本框
返回目录

3334 中 国 农 业 科 学 48 卷
marker technology and next-generation sequencing technology, theories and techniques of genomics have extended to multi-faceted germplasm research, resulting in revolutionary changes of conception and methodology in germplasm conservation, in-depth research and utilization. The achievements made in genomics provide not only theoretical guidance to collecting and conservation of crop germplasm, but also core theories and techniques to clarify crop origin and evolution. Meanwhile, genomics has the power to promote the efficiency of gene discovery and germplasm enhancement. Especially, the rapid development of whole genome sequencing, genome re-sequencing and simplified genome sequencing enable comparisons of genomic variation in different germplasms at the whole-genomic level. Further, crop origin can be illustrated and effects of domestication, improvement and dispersal on germplasm can be clarified. Population structure and genetic diversity of crop germplasm preserved in genebanks and existing in the wild can be evaluated in depth, generating huge amounts of information useful in designing optimum strategies of ex situ preservation and in situ conservation of crop genetic resources. Through integrating phenotypic data, new genes and their favorable alleles can be mined by using various approaches such as linkage mapping and association analysis. In addition, germplasm enhancement can also be benefited from the advances in genomics through the use of alleles discovered and other molecular marker-based techniques such as marker-assisted selection and genomic selection. This paper reviews the advances of genomics-based crop germplasm research, especially the applications of genomics in germplasm genotyping, ex situ and in situ conservation, crop origin and evolution research, structural diversity assessment, gene discovery and allele mining, and germplasm enhancement etc. Finally, future development directions and priorities in the field are proposed, which emphasize the close combination of genomics and crop germplasm research to provide a strong support to effective conservation and efficient utilization of crop germplasm in terms of theories, techniques, materials and information.
Key words: germplasm; genomics; review
0 引言
种质资源是作物遗传改良和相关基础研究的物质
基础。拥有作物种质资源的数量和质量,以及种质资
源研究和创新的深度和广度,直接影响到种质资源利
用效率和现代种业的可持续发展。因此,种质资源保
护和利用已成为世界各国农业科技创新驱动战略的重
要组成部分。
“基因组学”在 1986 年首次提出后发展十分迅
猛,基因组学理论和方法广泛应用于其他学科和不同
行业,催生了生物学科大数据时代,促进了生物技术
产业的蓬勃发展。与其他学科一样,基因组学的发展
对作物种质资源研究思路、技术路线、研究方法等产
生了革命性的影响,种质资源研究进入一个新的历史
发展阶段。特别是分子标记和测序技术的广泛应用使
种质资源的全基因组水平的基因型鉴定成为可能,种
质资源的结构多样性和功能多样性研究愈加深入,对
阐释作物起源、进化和传播、有效保护种质资源、发
掘新基因和高效种质创新将起到重要的推动作用。
1 种质资源的基因型鉴定
对种质资源的认识分 2 个层次,一是某种作物的
所有种质资源;另一是特定种质资源。针对所有种质
资源,需要全面了解这些种质资源的地理分布、群体
结构及其相互关系,也就是结构多样性,还需要了解
同一个基因在不同种质资源中的不同形式(即等位基
因)及其遗传效应,也就是功能多样性。而针对特定
种质资源,需要从 5 个方面了解:(1)这份资源是什
么;(2)这份资源的特性是什么;(3)控制这些特
性的基因或等位基因是什么;(4)这份资源有什么利
用价值;(5)通过什么途径可高效利用这份资源。 种质资源研究涉及到多门学科,特别是近年来生
物组学对其产生了深远影响,其中,基因组学带来的
颠覆性技术之一是基因型鉴定(又称基因分型)技术。
这些技术不仅可用于作物种质资源保护等基础性工
作,还广泛应用于遗传多样性分析、新基因发掘和种
质创新等多个方面。 基因型鉴定的必要手段是利用分子标记。最先开
发的分子标记是限制性片段长度多态性(restriction fragment length polymorphism,RFLP),之后不断出
现新的标记类型及其衍生形式,其中,最主要的包括
随机扩增多态性 DNA(random amplified polymorphic DNA,RAPD)、扩增片段长度多态性(amplified restriction fragment polymorphism,AFLP)、简单序列
重复(simple sequence repeat,SSR)和单核苷酸多态
性(single nucleotide polymorphism,SNP)等。目前,
随着高通量检测和测序技术的发展,除 SSR 和 SNP标记在广泛使用外,其他类型的标记已用得越来越少。
在大多数情况下,SNP 标记与 SSR 标记的效用相差无
几[1]。如 Hamblin 等[2]用 89 个 SSR 标记和 847 个 SNP

17 期 黎裕等:基于基因组学的作物种质资源研究:现状与展望 3335
标记对 259 份玉米自交系进行基因型鉴定,发现 2 种
标记类型的遗传多样性研究结果近似。Van Inghelandt等[3]进一步用 359 个 SSR 和 8 244 个 SNP 对 1 537 份
玉米自交系进行了鉴定,发现在群体结构和遗传多样
性分析时,2 种标记均可正确反映真实结果,但 SNP标记数量需要比 SSR 标记多 7—10 倍。不过,由于
SNP 标记具有覆盖全基因组、高通量、位点特异、共
显性遗传、误检率低、开发和检测成本急剧降低等优
点,将成为未来基因型鉴定的主要标记类型。 SNP 芯片技术可广泛用于种质资源的全基因组水
平基因型鉴定。目前广泛使用的基于芯片技术的 2 个
基 因 型 鉴 定 平 台 是 Affymetrix 的 GeneChipTM microarray 和 Illumina 的 BeadArrayTM technology。近
年来,科学界与企业合作开发了一批针对不同作物的
芯片。例如,在水稻上,Chen 等[4]从 801 份水稻品种
的重测序数据超过1 000万SNP位点中,筛选出51 478个均匀分布的标记(68%位于基因内),开发出了
RiceSNP50 芯片。在小麦上,全球多家研究单位合作,
开发了含 90 000 个基因相关 SNP 的芯片,其中 46 977个 SNP 已定位在染色体上[5]。Ganal 等[6]从 80 万个玉
米 SNP 中,筛选出 49 585 个 SNP 构建芯片,并用该
芯片对玉米的经典群体 IBM(B73×Mo17)和 LHRF(F2×F252)进行了图谱构建,分别定位了 20 913 个
和 14 524 个标记。Unterseer 等[7]开发了一个含 616 201个 SNP 和小的插入缺失变异(Indel)芯片,适合欧洲
和美国温带玉米材料的关联分析。目前,中国也开发
了一个适合同时研究温带和热带玉米的 55K玉米芯片
(徐云碧,私人通讯)。 在过去的几年中,基于第二代测序的全基因组水
平基因型鉴定技术(如全基因组测序、重测序、简化
基因组测序、RNA 测序等)开始出现。全基因组测序
策略适合基因组小的物种(如拟南芥),重测序则对
那些基因组相对较小的物种(如水稻、高粱、谷子等)
是个较好的策略。例如,Lam 等[8]对 17 份野生大豆
和 14 份栽培大豆进行了重测序,检测到 205 614 个
SNP,发现野生大豆中等位基因多样性较高,大豆
基因组中连锁不平衡(linkage disequilibrium,LD)
强。Qi 等[9]对 115 份黄瓜种质资源进行了重测序,
检测到 3 305 010 个 SNP、336 081 个短于 5 bp 的插
入缺失变异(Indel)、594 个有无变异(presence/absence variations,PAVs)。近年来,针对基因组较大的物种,
科学家们开发了一系列低成本、高通量的基于简化基
因组测序(即通过只对非重复或低重复基因组区域进
行测序来降低测序基因组复杂程度)的基因型鉴定方
法,如 RRLs[10]、CRoPS[11]、GBS[12-14]、SBG[15]、RAD[16]、
RESCAN[17]等。其中,GBS(genotyping by sequencing)应用较多,其优势是通过降低基因组复杂程度,变得
简便、可靠、实用,并且目前的使用成本已降到完全
可以接受的程度,但其缺点是需要有参考的基因组序
列[12, 14],缺失数据较多也对进行生物信息学分析造成
较大挑战[18],特别是针对基因组复杂的物种,建库和
测序策略以及生物信息学分析方法可能需要针对性制
定或开发。Glaubitz 等[19]最近开发了基于 GBS 的生物
信息学分析平台 TASSEL-GBS,目前,该平台已用于
多个物种的 GBS 数据分析,如在玉米中对 45 000 个
样品进行了 SNP 鉴定。但也有报道没有参考基因组序
列而直接利用 GBS 进行基因型鉴定,从而用于遗传图
谱的构建和 QTL 的分析[20-21]。 此外,外显子测序[22]、甲基化 DNA 测序[23]、转
录组测序(RNA-seq)[24-25]和序列捕获技术[26]也可用
来鉴定基因型。基于RNA-seq的基因型鉴定成本低廉,
更有可能检测到功能 SNP,但对其进行生物信息学处
理较为困难,需要解决表达丰度的巨大差异性问题和
可变剪切问题,特别是在没有参考基因组的物种中进
行基因型鉴定时,需慎重使用该方法[24]。序列捕获技
术主要针对目标区段进行基因型鉴定,利用
SureSelect、Nimblegen 和 Raindance 等方法,在测序
前对目标区段进行选择或富集,但这种方法更适合有
参考基因组序列的物种[26]。 除全基因组水平的基因型鉴定外,还有一种针对
目标基因进行的基因型鉴定,可以用来鉴定特定的等
位基因或单倍型 haplotype。例如,Pham 等[27]针对控
制大豆亚麻油酸含量的脂肪酸去饱和酶基因 FAD,鉴
定出 FAD3A 的 6.4 kb 缺失以及 FAD3B 和 FAD3C 的
SNP,前者用普通电泳方法即可分辨杂合子,后者用
TaqMan 方法即可做高通量标记选择。De La Fuente 等[28]
对 400 份玉米种质资源的脂氧化酶基因 ZmLOX4 和
ZmLOX5 进行了基因型鉴定,发现遗传多样性和 LD程度很低,并且有 1 个自交系的 ZmLOX5 没有功能,
1 个自交系缺失该基因,5 个自交系该基因有重复。 目前,中国的作物种质资源基因型鉴定大多数还
使用 SSR 标记,基于芯片和测序技术的基因型鉴定
(SNP 标记)仅在玉米、水稻、小麦、大豆、棉花等
作物中刚开始应用。因此,进一步提高种质资源基因型
鉴定的通量和效率,并对中国库存种质资源进行全面鉴
定评价,将是今后种质资源研究的突破口和重点任务。

3336 中 国 农 业 科 学 48 卷
2 种质资源异地保存
据 FAO[29]估计,全球收集保存的 740 万份种质资
源中仅 100 万—200 万份是特异的,其他种质都存在
不同程度的重复。传统的重复判定方法是通过形态学
性状和地理来源来判断,这种方法显然存在缺陷,因
为可利用的形态学标记相对较少。更有效的方法是通
过基因型鉴定来推断种质重复与否,但要注意的是不
同分子标记的分辨率会影响到“重复种质”的判定。
利用高分辨率的分子标记类型或测序技术,可以提高
遗传材料内及材料间遗传相似度和杂合度的鉴别能
力,有利于库存种质资源管理,如国际水稻研究所
(International Rice Research Institute,IRRI)分别利
用 384 个 SNP 位点和 50 对 SSR 分子标记对 2 800 份
水稻种质进行分析,发现 SNP 比 SSR 具有更高的分
辨率,可将之前认定的“重复种质”进一步加以区分,
即认定为非重复种质[30]。 种子保存寿命是种质库管理评价的重要指标。种
子保存寿命研究的经典方法是对通过发芽试验对种子
活力进行监测,并用分子标记等技术监测其遗传完整
性,确定繁殖更新临界值。研究表明,不同作物的种
子寿命是受遗传控制的。近年来,在拟南芥、水稻、
大麦以及油菜等作物中已鉴定出与种子寿命相关的数
量性状位点(quantitative trait loci,QTL)[31-33]。水稻
种子保存寿命 QTL 被定位在 7 条染色体上[31];大麦染
色体 2H、5H 和 7H 携带有种子老化基因 [33];小麦种
子保存寿命受生长和保存环境以及处理过程中种子含
水量的影响,与种子寿命相关的 QTL 定位在染色体
2AS 上[34]。如果克隆了那些在低温条件下(如长期库
-18℃下)种子保存寿命相关的 QTL,并对不同种质资
源中的等位基因进行系统分析,将有助于了解不同材
料种子保存寿命的自然遗传变异,通过检测等位基因
或利用功能标记就可预测繁殖更新时间。 对于无性繁殖作物,对茎尖等分生组织进行超低
温保存(-196℃到-130℃)是重要的长期保存方式。
分子标记或其他基因组学技术可应用于超低温冻存
后遗传稳定性研究[35-36]。研究结果表明,超低温保存
后一般可保持遗传稳定性,但也可能影响到 DNA 甲
基化[37-38]。超低温保存过程中涉及的低温处理和脱水
环节对植物组织超低温保存成活有显著影响,研究超
低温保存过程中基因组和转录组对冷冻和脱水胁迫的
响应,有助于制定最佳保存策略。目前主要以拟南芥
茎尖为材料进行了研究,如 Volk[39]利用 cDNA 芯片技
术,研究了超低温保护剂处理、液氮保存以及恢复培
养条件下的基因表达模式。在作物上的类似研究应该
更具实践意义。 中国种质安全保存相关研究取得了一定进展,如
利用 AFLP 标记分析了繁殖群体量及隔离对蚕豆种质
遗传完整性的影响[40]、大豆中黄 18 种质在种子老化
处理后的遗传完整性[41],利用 SSR 等分子标记研究了
马铃薯种质超低温保存后的遗传稳定性[42],利用转录
组学方法开展不同苗龄的拟南芥茎尖材料超低温保存
过程差异基因表达研究[43]。但总体来看,在种质资源
保存方面基于基因组学水平的研究规模和深度均较欠
缺,与种子保存寿命相关、种子老化相关的相关基因
研究更少。
3 种质资源原生境保护
原生境保护已成为作物野生近缘植物保护的主要
方式,广泛应用于濒危野生近缘植物的遗传多样性保
护。原生境保护的理论基础是通过对地球上物种形成
机制的了解以及调查人类活动对物种、遗传变异和生
态系统的影响,建立可操作的方法来维持野生植物的
遗传多样性,保护和恢复生物群落及其生态功能。但
是,传统原生境保护中的很多科学问题很难用经典遗传
学技术加以解决,特别是群体结构、居群间的基因流和
进化动态及居群间的亲缘关系等受到遗传和环境因素
的双重影响而难以被精确估算[44]。 应用分子标记及基因组数据估计野外居群的群体
结构、确定居群间的基因流和亲缘关系等,可较为准
确地估计生物多样性,结合野外调查统计数据和生态
学信息,即可确定优先保护区域和优先保护居群。例如,
杨庆文等[45]通过对东乡普通野生稻(Oryza rufipogon Griff.)原位保存群体遗传分化进行研究,发现该地的
2 个居群应属于一个大群体,因为人为干扰导致生境
片断化,从而形成了 2 个独立的居群,并据此提出了
将 2 个居群同时保护但侧重于庵家山居群的保护策
略,为该地建立原生境保护点提供了科学依据。王家
祥等[46]利用 SSR 分子标记对中国 15 个普通野生稻原
生境保护居群的 427 份材料和在中国分布区内按照纬
度划分后随机挑选的15个未保护居群的357份普通野
生稻材料进行了遗传多样性分析,发现中国普通野生
稻原生境保护居群具有较高的遗传多样性,已经建立
的 15 个原生境保护点内的普通野生稻基本上能够代
表中国普通野生稻的遗传多样性状况,但广东和广西
保护居群的遗传多样性水平低于未保护居群,说明该

17 期 黎裕等:基于基因组学的作物种质资源研究:现状与展望 3337
地区原生境保护点数量较少,遗传代表性不够,应该
扩大保护范围,增加保护点数量。 新一代测序技术的发展,使得某些基因的序列能
够在更多的样品上进行比对,获取基因序列多样性的
大量信息,从而阐明基于基因序列的谱系关系及其近
缘种间关系,有助于确定物种优先保护顺序。例如,
Qiao 等 [47]对 98 份栽培稻和普通野生稻糯性基因
(waxy)的序列变异进行了研究,从检测到的栽培稻
和野生稻 SNP 插入或缺失的核苷酸序列得出结论,栽
培稻中的糯性基因来源于华南地区的普通野生稻。
Wei 等[48-49]对叶绿体中 atpA 序列、rps16 内含子序列、
trnP—rp133 间隔区、trnG—trnfM 序列、trnG—trnfM间隔区序列,线粒体的 cox3、cox1、orf224 和基因间
序列 ssv-39/178、rps2—trnfM 以及核基因组的 ITS、Ehd1、Hd1 进行测序,利用生物信息学方法进行比对
分析,发现所有与中国栽培稻亲缘关系较近的普通野
生稻均来源于华南地区,而且栽培稻的 2 个亚种籼稻
和粳稻在进化过程中分别由该地区偏籼型的普通野生
稻和偏粳型的普通野生稻进化而来,因此,推断华南
地区是亚洲栽培稻的起源中心,并由此制定了以华南
地区为主的野生稻原生境保护策略。此外,Henry[50]
在 2014 年也明确提出,对野生近缘种材料进行测序和
深入分析,可在全基因组水平上对作物野生近缘植物
的遗传多样性及其变化进行系统调查研究,将有助于
种质资源的原生境保护。 目前,基因组学在作物种质资源的原生境保护中
的应用案例还较少。这里主要存在 2 个问题,一是基
因组学研究取样数量往往有限,并且没有以居群为单
位,不能完全反映现有野生群体的状态;二是对野生
居群的研究仅局限于与栽培种的比较,对其本身多样
性及其形成机制和环境适应机制的研究较少。因此,
在未来的保护基因组学研究中,需首先研究保护基因
组学所需的居群取样原则,在此基础上通过对不同生
境样本的基因组测序获得基因组的平均多样性变化,
有效监测等位基因频率的变化,评估遗传漂变、选择
和基因流对原生境保护群体的影响,阐明群体适应环
境变化的遗传因素,从而提高对野生群体进行原生境
管理的能力。
4 作物起源、驯化与传播
驯化(domestication)是把野生植物变成栽培作
物的过程,经过驯化,栽培作物丧失了野生植物的不
良特性,具备了种子易萌发、直立生长、籽粒或果实
大、不落粒或不炸荚等丰产和适合管理的性状,使其
在人工种植条件下单位面积产量显著提高。以大豆为
例,栽培大豆由其祖先种一年生野生大豆(Glycine soja Sieb.& Zucc)进化而来,驯化前的野生大豆表现为小
粒、植株缠绕生长,驯化后产生植株明显变小、营养
生长降低、植株缠绕性减弱、但仍然易于倒伏的原始
地方品种(Glycine max L. Merr.)。之后,现代大豆
育种家对地方品种进行了定向改良(improvement),
培育出植株直立生长、大粒、高产的优良品种。但与
对多湿、盐碱、低温等不良环境具有较强的适应能力
的作物祖先种相比,人工集约化种植的栽培作物的适
应性往往大为降低,不过也存在一些适应性更广的例
子,如热带起源的栽培玉米现在广泛种植于南北纬 50度之间的辽阔区域。
栽培作物与其祖先野生种在这些表型和适应性上
的差异是由遗传控制的。目前,已成功定位并克隆到
驯化相关性状(又称为“驯化综合特征(domestication syndrome)”性状)的 QTL 和基因,如控制玉米分蘖
的顶端优势基因 Teosinte branched1[51-52]、水稻单分
蘖基因 MOC1[53]、落粒基因 sh4[54]和 qSH1[55]、抗倒
伏基因 PROG1[56]和籽粒大小基因 qSW5[57]、黄瓜苦
味基因[9]等。驯化过程涉及到的复杂表型往往是多基
因作用的结果,基于表现型先验鉴定的分析方法只是
对极少数基因/位点进行分析,不能检测到那些还未测
定表现型的位点;另一方面,由于每个基因/位点受选
择历史、遗传结构等不尽相同,单一基因/位点不能反
映全基因组水平的驯化过程,无法满足全面解析驯化
过程中人工选择导致的全基因组范围内遗传变异的发
生规律及相互作用机制的需要。 随着高通量测序技术和生物信息学的快速发展,
很多作物的参考基因组不断被公布,基因组学数据迅
速积累。特别是全基因组重测序、RNA-Seq、miRNA测序、GBS 等分析技术的出现,产生了大量可利用的
基因组信息,促进了基于全基因组序列的作物比较基
因组学和进化基因组学等学科的发展,为全面理解和
诠释作物驯化历史及全基因组的遗传变异特征奠定了
基础。 4.1 作物驯化历史与地理起源
作物被驯化的次数和地理起源是影响作物遗传结
构和遗传多样性水平及驯化相关性状形成的根本性因
子之一。但由于多方面的原因,迄今为止,大多数作
物驯化次数和起源地等驯化历史问题还没有得到解
决。近年来,基于重测序分析的比较基因组学研究促

3338 中 国 农 业 科 学 48 卷
进了在全基因组范围内了解作物的驯化历史。例如,
对 84 份桃种质的重测序分析,明确了从西藏光核桃、
山桃、甘肃桃到普通桃的分子进化路线[58]。对 44 份
高粱种质的重测序分析发现高粱的驯化历史复杂,至
少发生了 2 次独立的驯化事件,最近一次驯化产生了
guinea-margaritiferum 类群[59]。通过对 60 份普通菜豆
野生材料和 100 份地方品种进行重测序,证实了普通
菜豆在中美洲和安第斯地区是独立起源的,并且发现
这两个基因库是在人类到达前就已分化开来[60]。 亚洲稻有单一起源(粳稻和籼稻皆从野生稻 O.
rufipogon 驯化而来)和多起源(粳稻和籼稻是从亚洲
不同地方驯化而来)两种假说,一直是科学争论的焦
点问题之一。Huang 等[61]通过对 446 份普通野生稻、
1 063 份粳稻和籼稻品种进行重测序,构建水稻基因组
变异图谱,鉴定出驯化过程中发生了 55 个选择清除事
件(selection sweep,即由于作物驯化过程中选择压力
强,累积的遗传重组事件少,导致受选择基因与其邻
近区域等位基因间产生牵连效应,形成多样性大幅降
低的人工选择区段),认为粳稻最先从普通野生稻一
个特定群体中驯化出来,地点发生在珠江流域中游地
区,籼稻是粳稻与野生稻杂交而来,然后传入东南亚
和南亚。而对非洲稻而言,通过对 20 份非洲栽培水稻
(O. glaberrima)及 94 份野生近缘种 O. barthii 的重
测序分析,在分子水平上确认非洲栽培水稻独立起源
于尼日尔河流域[62]。 4.2 作物驯化相关的遗传多样性变化
人类活动在作物驯化过程中扮演着重要角色,其
有目的的选择使作物每繁殖一代都只有个别或少部分
野生植株通过繁殖保留下来,最终形成遗传瓶颈效应。
近年来利用重测序分析方法对水稻、玉米、大豆、黄
瓜等栽培作物及其野生近缘种进行群体基因组学分
析发现,栽培作物在驯化过程中遗传多样性大量丢
失、连锁不平衡水平增高[63-65],人工选择对栽培作物
的种质资源构成产生了重要影响。各种作物驯化过程
中遗传瓶颈效应强度差异明显(表 1),其中,西瓜
瓶颈效应最强,丢失的多样性最多,玉米丢失的多样
性最少,仅为西瓜的 20.8%。对黄瓜、西瓜和西红柿等
果蔬和水稻、玉米、大豆、高粱等谷类作物的比较分析,
发现黄瓜等果实类作物在驯化过程中的遗传多样性下
降程度比后者更大[9]。对水稻 2 个亚种粳稻和籼稻的进
一步分析发现,驯化后粳稻只保留了 O. rufipogon 51%的遗传多样性,而籼稻保留了 O. nivara 91%的遗传多
样性,说明驯化过程中粳稻受人工选择强度更为强烈,
导致粳稻的有效群体规模大幅缩小[63]。
表 1 8 种作物驯化过程中遗传瓶颈效应评估
Table 1 Estimation of domestication bottlenecks in eight crop species
作物 Crop
野生近缘种 Wild relative
栽培种 Cultivated crop
驯化过程中丢失的多样性 Percent of diversity lost during
domestication (%)
文献 Reference
水稻 Rice Oryza rufipogon O. sativa 29.9 [63]
玉米 Maize Zea mays ssp. parviglumis Z. mays ssp. mays 16.9 [64]
大豆 Soybean Glycine soja G. max 33.3 [65]
高粱 Sorghum Sorghum bicolor ssp. verticilliflorum, S. propinquum S. bicolor subsp. bicolor 35.9 [59]
西红柿 Tomato Solanum pimpinellifolium S. lycopersicum 61.9 [66]
桃 Peach Prumus davidiana, P. ferganensis, P. kansuensis, P. mira P. persica - [58]
黄瓜 Cucumber Cucumis sativus var. hardwickii C. sativus L. 48.9 [9]
西瓜 Watermelon Citrullus lanatus ssp. mucosospermus C. lanatus subsp. vulgaris 81.6 [67]
驯化过程中作物在丢失了大量遗传多样性的同
时,因为突变和基因渐渗等原因又有可能产生了新的
遗传多样性。例如,利用全基因组重测序分析,在 25份野生大豆和 30 份栽培大豆中共检测到 5 102 244 个
SNP 位点,其中有 10.4%(529 724 个)为栽培大豆所
特有[65]。同样,驯化后的栽培玉米也获得了新的遗传
多样性[64]。
4.3 人工选择区段的鉴定与驯化相关性状的关系
在经历驯化后,作物的遗传多样性明显降低,是
人类有目的地选择期望性状的结果。在控制驯化性状
的基因组区段,只有极个别等位变异被保留下来,其
他等位变异则逐步被淘汰,使控制这些性状的基因在
驯化过程中逐渐被固定下来[68]。 利用群体基因组学理论和方法对栽培作物及其近

17 期 黎裕等:基于基因组学的作物种质资源研究:现状与展望 3339
缘野生种的基因组序列进行对比分析,可以鉴定出符
合分化系数(Fst)高、多样性差异显著和不符合中性
检验等特点的基因组区段及其包含的重要功能基因,
这些在群体间存在巨大频率变化的区段和基因可能是
驯化过程中人工选择的靶点。利用这种分析方法,在
高粱上鉴定出 725 个人工选择候选基因[59]、大豆 928个[65]、黄瓜 2 054 个[9]、辣椒 511 个[69]、普通菜豆 2 524个[60]、粳稻 1 322 个、籼稻 1 265 个,其中 73 个基因
在粳稻和籼稻上同时被检测到[63]。进一步交叉对比分
析发现,部分人工选择区段位于控制驯化相关性状的
QTL 区间,预示着可能与作物的农艺性状相关。例如,
果实或籽粒重量是重要的驯化综合特征性状之一,驯
化后,现代栽培作物的果实或籽粒重量大大增加。从
野生醋栗番茄到樱桃番茄是个驯化过程,检测到 186个选择清除事件,其中有 5 个 QTL 与之重叠;从樱桃
番茄到大果栽培番茄是个改良过程,发生了 133 个选
择清除事件,有 13 个基因与之重叠,其中包括以前认
为是驯化基因的 fw2.2[66,70]。对黄瓜种质资源重测序分
析发现控制果实苦味的关键基因位于第 5 染色体上一
个物理长度为 442 kb 的基因组区段,包含 67 个注释
基因,该区段也是发生选择清除事件的区段[9]。 以上主要是对重测序数据发掘的微小遗传变异
(SNP 或小于 5 bp 的 Indel)进行分析的结果,在大
片段 Indel、拷贝数变异(copy number variation,CNV)、
有无变异(PAV)等结构变异检测上则由于具有偏向
性和不完整性,无法准确发现那些在人工选择下丢失
和新产生的基因。然而,研究表明,这些结构变异与
作物重要农艺性状密切相关,越来越受关注[65, 71]。
随着高通量测序技术进步和成本降低,为了完整地呈
现基因序列及其变异特征的全貌并全面发掘驯化性
状相关基因,Li 等[72]系统筛选出 7 份国内外代表性
野生大豆,并分别进行从头测序、组装和分析,建立
了可代表物种整体特点的野生大豆泛基因组(Pan- genome)。与栽培大豆参考基因组的比对,鉴定到
328 个野生大豆特有基因、16 个栽培大豆特有基因、
1 978 个拷贝数发生变化基因等结构变异,为研究作
物遗传多样性及驯化历程提供了新的启示,奠定了解
析重要驯化性状建成、发掘优异基因和相关标记的基
础。 4.4 作物的传播与地方品种的形成
作物从一个或多个地点驯化而成后,随着人类活
动特别是人口迁移而传播到世界不同地区。历史上最
突出的作物传播事件是哥伦布发现美洲后,美洲作物
(如玉米、马铃薯、食用豆等)向欧洲、亚洲和非洲
的迅速传播。在传播过程中,由于环境条件发生了改
变,自然选择和人工选择使作物基因组经历了第二次
较大的变化,由此产生了大量适应当地生态环境和人
类生活习惯的地方品种。 利用基因组学理论和方法,可以追踪作物传播轨
迹。最经典的例子是 Mir 等[73]对玉米传播的研究,法国
农业科学院与墨西哥、乌拉圭、泰国等合作,利用 SSR标记,对 799 份来自全球的玉米地方品种(共 11 985 个
植株)进行了基因型鉴定,发现地方品种之间的遗传
关系与地理来源有对应关系,基本澄清了亚非玉米的
来源。有意思的是,研究发现中国南方和黄淮海的地
方品种可能来自从葡萄牙和西班牙传入东南亚后再传
入的墨西哥地方品种,而中国东北的地方品种可能来
自过去 100 多年里从美国传入的地方品种。 需要指出的是,目的性更强的现代作物育种历史
并不长,但所形成的现代品种(或品系)携带的目标
性状更符合人类的需求。从基因组水平来说,强烈的
人工选择使遗传多样性从地方品种到现代品种进一步
降低,基因组中出现了除驯化相关区域之外的新选择
区域。应用基因组学方法,全面分析野生近缘种、地
方品种、现代品种这些种质资源,可更清楚地了解不
同阶段产生的种质资源的遗传变异变化和人工选择区
段及其与表型性状的关系。例如,Wang 等[74]利用 60 K油菜芯片对 472 份来自全球的油菜种质进行了分析,
发现不同来源的种质资源遗传多样性存在差异,中国
和欧洲油菜从 20世纪 50年代到 80年代的遗传多样性
呈增加态势,然后基本处于相似水平。在玉米上,现
代育种对玉米基因组带来了剧烈变化,人工选择影响
到上千的靶点(包括基因和非基因区域),导致核苷
酸多样性降低和稀有等位基因频率提高[75]。在番茄上
的类似研究也取得了突破性进展[66]。
5 作物种质资源的结构多样性分析
全面了解种质资源的遗传结构是其有效保护和高
效利用的前提。遗传多样性(genetic diversity)、群
体结构(population structure)和连锁不平衡分析是作
物种质资源结构多样性分析的 3 个重要方面,彼此之
间既有区别,也有关联。其中,群体结构是结构多样
性分析的核心,也是发掘新基因的重要手段之一关联
分析的重要基础,本文将着重讨论。 一般来说,SSR 等常规标记即可用于群体结构分
析。例如,Dang 等[76]利用 262 个 SSR 标记对 419 份

3340 中 国 农 业 科 学 48 卷
中国水稻材料和 121 份越南材料进行了群体结构分
析,发现该群体可分为 7 个亚群,连锁不平衡水平在
10—80 cM。由于类似论文发表众多,并且涉及很多
物种,本文将不再赘述。近年来,直接利用序列信息
或高密度标记进行群体结构分析越来越普遍,主要方
法包括全基因组测序(如在番茄[77]上)、重测序(在
大豆[8]、黄瓜[9]、高粱[59]、西瓜[67]、芝麻[78]、桃[79]、
鹰嘴豆[80]、大麦[81]、玉米[75,82]上)、RNA 测序[25]及
SNP 芯片等,不仅可检测 Indel、CNV、PAV 等变异,
而且能检测到大量 SNP、SSR 等变异,其中 SNP 变异
广泛用于群体结构分析上。 5.1 群体结构的统计学分析方法
通过群体结构分析可以了解种质资源中的亲缘关
系、特定材料中某位点的来源等多种信息。分析种质资
源遗传结构最常用的方法是层级聚类分析(hierarchical clustering analysis),以前直接用基因型数据估算遗传
差异就可聚类分析,最近的研究发现先进行主成分分
析(principal components analysis,PCA)后再做聚类
分析可以提高功效[83]。主成分分析的优势是直观,缺
点是难以获得数量化指标来判断群体结构数量及其材
料间的相互关系。因此,Pritchard 等[84]提出基于贝叶
斯模型的方法后迅速得到应用,使 STRUCTURE 成为
目前用得最多的群体结构分析软件。此后,为了适应
SNP 大数据的快速计算,还开发出一些如基于最大似
然法等的算法和软件(如 FRAPPE 和 ADMIXTURE);2014 年,Raj 等[85]又开发出 fastSTRUCTURE,不仅功
能强大,而且运算快速,值得推广。 5.2 基于 SNP 数据的种质资源群体结构分析
SNP 芯片数据和基于测序的 SNP 数据均可用于
遗传多样性、群体结构和 LD 分析。但要注意的是,
当利用 SNP 芯片数据分析时,在较少种质材料基础上
开发的 SNP 应用于更广泛的种质资源时,可能出现估
计偏差。例如,Frascaroli 等[86]对 731 个 AFLP、186个 SSR、41 434 个 MaizeSNP50 芯片 SNP 以及该芯片
中 30 068 个 SNP(Panzea 在 14 个玉米和 16 个大刍草
材料基础上开发)和 11 366 个 SNP(Syngenta 专为北
美马齿种质开发)的基因型鉴定数据分别进行分析,
发现尽管分群的总趋势相似,不影响群内种质的亲缘
关系估计,但比较群间种质的关系时出现差异,认为
相对来说 Panzea 的 SNP 标记较好。因此,利用遗传
多样性最高的一套材料来开发的 SNP 芯片应用价值
更高。 近年来,SNP 用于群体结构分析的报道越来越多,
使人们对种质资源认识的深度达到前所未有的程度。
例如,在大麦上,Munoz-Amatriain 等[87]利用含 7 842个 SNP 的大麦 SNP iSelect 平台,对从来自全球 100多个国家的 33 176 份大麦种质资源中挑选出来的
2 417 份大麦核心种质进行了基因型鉴定,STRUCTURE和主成分分析结果表明,这些材料可分为 5 个与地理
来源和穗行数对应的亚群。Comadran 等[88]对 190 份西
北欧洲和北美大麦种质进行了 4 596 个 SNP 标记鉴
定。Hamblin 等[89]利用 1 536 个 SNP 标记对 1 816 份
美国育种项目的大麦种质进行了基因型鉴定,发现存
在 7 个亚群,群体结构与穗类型、生长习性、育种项
目有关。 在小麦上,Cavanagh 等[90]开发了一个含 9 000 个
基因相关的 SNP 芯片,对 2 994 份小麦地方品种和育
成品种进行了基因型鉴定,发现在现代育种中广泛利
用的等位变异可追溯到祖先亲本,通过分析群体间的
遗传分化和单倍型共享程度,可鉴定出遗传改良中受
到选择的性状(主要是开花期和物候期)的等位变异。
此外,Wurschum 等[91]利用 9 K SNP 芯片对 172 份欧
洲冬小麦种质资源进行了基因型鉴定,发现利用 SNP和SSR获得的群体结构和遗传相似性结果存在一定差
异,LD 衰减距离在 5—10 cM,因而认为 9 000 个 SNP不能完全解析其群体结构。
在玉米上,Yan 等[92]利用含 1 536 个 SNP(来自
582 个位点)的玉米芯片,对 632 份来自热带、亚热
带和温带的玉米自交系进行了基因型鉴定,发现存在
亚群结构,LD 衰减距离在染色体间存在差异(1—10 kb),并且随稀有等位基因频率(MAF)增加和样品
减少而变大;温带玉米的 LD 衰减距离比热带和亚热
带玉米高大得多,因为后者多样性高且携带有更多的
稀有等位基因。Lu 等[93]利用 1 943 个 SNP 标记对 287份热带玉米和 160 份温带玉米自交系进行了基因型鉴
定,发现内含子 SNP 变异比外显子变异高,单倍型多
态性信息含量比单 SNP 多样性信息含量高,遗传多样
性水平和亚群结构与种质来源和驯化后选择存在密切
关系;热带玉米比温带玉米遗传多样性高,但没检测
到明显的亚群结构,其 LD 衰减距离(5—10 kb)比
温带玉米小(10—100 kb)。Wu 等[94]利用含 56 110个 SNP的玉米芯片对 367份中国重要自交系进行了基
因型鉴定,发现这些材料可分为国外种质和国内种质
两组,在此基础上可进一步分为对应杂种优势群的 5个亚组,即瑞德、兰卡斯特、P 群、唐四平头和温热
I 群,并且各亚组中的遗传多样性存在较大差异。有

17 期 黎裕等:基于基因组学的作物种质资源研究:现状与展望 3341
意思的是,各个亚组均存在数目不同的保守基因组区
段,平均 LD 距离为 391 kb,且不同染色体区域有差
异。利用该芯片还对其他玉米种质资源如密里苏达大
学培育的玉米自交系进行了基因型鉴定,获得一些重
要种质的遗传信息[95]。一个规模较大的玉米种质资源
结构多样性分析工作是,Romay 等[96]利用 GBS 技术
对美国国家种质库中的 2 815 份玉米自交系进行了基
因型鉴定,获得 681 257 个 SNP 标记,其中半数以上
属稀有等位基因;结果表明,尽管绝大多数稀有等位
基因已被公益性温带育种项目利用,但商业用种质中
并不多。另一方面,这些自交系呈现明显的群体结构,
且 3 个亚群间的遗传分化程度为中等水平,LD 衰减
很快,但其程度与种质来源和基因组区域有关。 此外,在番茄上,Sim 等[97]利用含 7 720 个 SNP
的芯片,对 426 份番茄种质进行了基因型鉴定,发现
其中 410 份自交系可分为 7 个亚群,其中加工番茄亚
群和鲜食番茄亚群可进一步细分,通过分析稀有等位
基因频率,可鉴定出区分不同亚群的基因组区段,LD衰减与染色体和亚群密切相关,结果表明,栽培番茄
种质资源的不同遗传结构是野生近缘种基因渐渗和根
据市场目标的育种选择带来的。Wang 等[98]利用 GBS技术,对 242 份高粱微核心种质进行了基因型鉴定,
最后选出 13 390 个 SNP 进行遗传结构分析,结果发
现这些种质存在与地理来源和类群划分相关的群体结
构,LD 衰减距离为 10—30 kb。 5.3 骨干亲本及其衍生品种的遗传结构与传递
骨干亲本及其衍生品种(或品系)是种质资源的
重要组成部分,对其进行基因组水平的研究具有重要意
义。如在玉米上,Wu 等[94]发现从骨干亲本黄早四到其
衍生系中的基因组区段传递有一定规律,这些材料中存
在 15 个保守区段,它们可能是黄早四成为骨干亲本的
重要原因。肖永贵等[99]利用 90 K SNP 芯片分析了小麦
骨干亲本京 411 及其 14 个衍生品种(系),发现京 411衍生群体平均遗传相似性为 57.9%,该亲本与其衍生一
代和衍生二代相同的等位变异频率分别为 63.9%和
67.9%,显著高于理论值;在 A、B 和 D 基因组间的相
同等位变异频率分别为 62.2%、61.3%和 74.3%。
6 基于种质资源的新基因发掘
6.1 连锁分析
利用 2 个材料作为亲本组配人工作图群体来进行
新基因发掘,在近 20 年来已成为一种广泛采用的方
法,但由于这种连锁分析方法难以体现种质资源的规
模效应,本文将不作为重点进行评述。这里要提到的
是,在连锁分析中构建高密度的 SNP 图谱可提高 QTL位置的定位精度、提高 QTL 效应值估计精度和缩小
QTL 的置信区间,有利于 QTL 精细定位、图位克隆
和候选基因挖掘,特别是由于近年来二代测序技术成
本大幅度降低,构建高密度分子图谱将成为今后重要
的发展方向。一种策略是先对双亲进行重测序,在高
质量 SNP 中筛选部分均匀分布的标记构建 SNP 芯片,
再用芯片对群体进行基因型鉴定[100]。另一种策略是对
所有作图群体个体或家系直接进行测序,再构建 SNP图谱,如 Huang 等[101]利用低倍测序基因型鉴定技术,
对 150 个水稻重组近交系群体进行基因型鉴定,发现
数据采集比常规方法快 20 倍,重组断点确定精度高
35 倍,主效 QTL 可定位到 100 kb 的区间。Xie 等[102]
提出了一种用低倍数测序(0.05×)建立高密度 SNP图谱的方法,并在 238 个重组近交系群体中得到验
证。Spindel 等[18]利用 384 重 GBS 技术,对包括 176个水稻重组近交系的作图群体进行基因型鉴定,定位
了 30 984 个 SNP 标记,通过对叶宽和耐铝性进行 QTL定位,检测到用 1 464 个 SNP 进行作图所检测不到的
位点。Liu 等[103]利用 GBS 技术对大麦重组近交系群体
进行基因型鉴定、遗传作图和株高的 QTL 分析,精细
定位到 3 个置信区间很小的重要 QTL,对比物理图谱,
找到了相关候选基因。 6.2 关联分析
关联分析是利用历史积累的自然变异材料(即种
质资源)来阐明基因型与表型的相互关系,因此,成
为研究种质资源自然变异、发掘有利等位基因最佳的
策略之一。关联分析有 3 种形式,即全基因组水平关
联研究(genome-wide association study,GWAS)、区
域水平关联分析(local or regional association analysis)和候选基因关联分析(candidate gene-based association analysis)。 6.2.1 全基因组关联分析的统计学方法 GWAS 最
简单的统计学模型是 SNP 基因型与表型相关性的单
位点测验,但由于存在群体结构等问题,这种测验结
果往往假阳性率很高。因此,针对 GWAS,目前发展
了多种统计学分析方法。例如,Yu 等[104]发展了考虑
群体结构的混合线性模型;Zhang 等[105]提出了提高运
算能力的新算法,并整合目前关联分析应用最多的软
件 TASSEL;Korte 等[106]又针对多个相关性状发展出
参数化的多性状混合模型(multi-trait mixed model,MTMM),Segura 等[107]发展出多位点混合模型来优

3342 中 国 农 业 科 学 48 卷
化存在群体结构情况下的关联分析;Wang 等[108]则提
出 FaST-LMM 算法,并整合进 GAPIT 软件包。Yang等[109]利用 513 个玉米自交系和 50 万左右的 SNP 标记
以及 17 个表型性状,发展了结合 IBD 预测和 KNN 算
法的大量缺失标记估计方法,认为全基因组关联分析
时使用 Anderson-Darling(A-D)测验对于那些非正态
分布的复杂表型性状有较好效果。另外,更新一代的
统计软件 FARM-CPU 正在开发中(张志武,私人通
讯)。 6.2.2 全基因组关联分析的典型例证 从近年来发
表的关联分析论文来看,其中相当一部分是利用为数
几百的 SSR 或其他类型标记,取得了一些研究进展,
但由于标记密度太少,其结果需要进一步验证。为了
获得更可靠的结果,特别是需要找到目标性状的候选
基因,则有必要开发更多的 SNP 标记才能满足全基因
组关联分析的要求。一般来说,对于异花授粉作物或
常异花授粉作物需要十万数量级的标记,对于自花授
粉作物也需要上万数量级的标记,这取决于所用材料
的 LD 程度大小。因此,这部分限制了关联分析的应
用范围,能应用的作物需要有较多 SNP 数量的芯片,
或者已有全基因组测序草图然后进行重测序。总之,
目前利用关联分析方法较成功的作物还不太多,需要
进一步拓展和加强。 在水稻上,Chen 等[110]对 529 份水稻种质进行了
重测序,获得 640 万左右的 SNP,对 5 叶期叶片的 840种代谢物进行了分析,通过全基因组关联分析确定了
控制代谢物水平的 36 个候选基因。Huang 等[111]对 517份籼稻地方品种进行重测序,鉴定出 360 万个 SNP,对 14 个农艺性状进行全基因组关联分析所检测到的
位点可解释 36%的表型变异。Huang 等[112]进一步把材
料扩大到包括籼稻和粳稻的 950 份种质资源,利用重
测序方法进行基因型鉴定,针对开花期和籽粒相关性
状进行全基因组关联分析,结果表明,研究样本越多,
检测到的位点越多,并且 GWAS 与功能基因组注释结
合起来可快速找到控制目标性状的候选基因。 在小麦上,肖永贵等[99]对小麦骨干亲本京 411 及
其 14 个衍生品种(系)进行了根系性状的精准鉴定,
结合 90 K SNP 芯片数据,发现京 411 衍生品种的主根
长和根干物质重量等性状的改良较为显著;在此基础
上,共发掘出 35 个根系性状位点,来自京 411 的 26个位点对根部性状起正向效应,重要衍生品种中麦
175 和 CA0958 携带正向效应位点最多,占正向效应
位点总数的 73.1%。Zegeye 等[113]利用小麦 9 K SNP
iSelect 芯片对 181 份人工合成小麦进行了基因型鉴
定,对条锈病抗性的 GWAS 结果检测到 9 个基因组区
段的 27 个 SNP 与幼苗期抗性有关,18 个基因组区段
的 38 个 SNP 与成株期抗性有关。Liu 等[114]利用小麦
芯片对 205 份种质进行了基因型鉴定,并鉴定了土传小
麦花叶病毒(soil borne wheat mosaic virus,SBWMV)
的抗性,检测到 5D 染色体上 2 个基因的 6 个 SNP 与
抗病性有关,其中有 2 个 SNP 转成 KASP(kompetitive allele specific PCR)标记后很容易用于标记选择。
在玉米上,Romay 等[96]利用 2 815 份自交系进行
关联分析的结果表明,由于 LD 衰减快,针对复杂性
状的连锁分析需要更多的 SNP 标记。Bouchet 等[115]
利用 50 K SNP 芯片对 375 份欧美热带和温带种质进
行了基因型鉴定,针对开花期进行了关联分析,检测
到 18 个基因组区段,其中有 1 个显著关联的 SNP 与
开花期基因 ZCN8 距离 5 kb,该区段多样性较低,分
化和 LD 程度较高。选择测验表明,还存在着一些其
他与开花期有关的位点,但关联分析没有检测到,因
而建议增加 SNP 数量、改进试验设计和统计学方法。
Riedelsheimer等[116]利用含56 110个SNP的芯片对289份玉米自交系进行了基因型鉴定和叶片 118 种代谢物
表型鉴定,检测到 15 个 SNP-代谢物关联关系,对其
中一些代谢物的遗传控制进行了深入探讨。Strigens等[117]利用该芯片对 375 份玉米自交系进行了基因型
鉴定和耐冷性评价,全基因组关联分析结果检测到 19个显著相关信号。
在谷子上,Jia 等[118]对 916 份谷子种质资源进行
了重测序,获得 258 万 SNP,这些材料可明显分为与
开花期相关的两组;对其在 5 种环境下鉴定了 47 个农
艺性状,GWAS 结果检测到相关的 512 个位点。在番
茄上,Sauvage 等[119]利用 5 995 个 SNP 标记对 163 份
番茄种质资源进行了基因型鉴定,针对果实代谢物进
行了精准的表型评价,GWAS 结果检测到与 19 个性
状关联的 44 个位点。在大麦上,Cockram 等[120]利用
1 536 个 SNP 对 500 个大麦品种进行了基因型鉴定,
针对 15 个形态学性状进行了全基因组关联分析,发现
LD 水平很高,检测到一些已知基因,并把花青素基
因定位到一个含 3 个基因在内的 140 kb 区间内。Wang等[121]利用含 1 536个 SNP的芯片对 615份大麦种质资
源进行了基因型鉴定,并对 32 个形态学性状和 10 个
农艺性状进行了关联分析,发现存在明显的与季节生
长习性和穗行数密切相关的群体结构。Zhou 等[122]为
了发掘美国大麦种质资源对非洲秆锈小种 TTKSK 的

17 期 黎裕等:基于基因组学的作物种质资源研究:现状与展望 3343
抗病基因,幼苗期鉴定了 960 份材料,成株期鉴定了
768 份材料,用 3 072 个 SNP 进行基因型鉴定,分别
在 7H 和 5H 染色体上定位到抗病 QTL。 6.3 基于候选基因的关联分析与有利等位基因发掘
康奈尔大学 Buckler 研究组是最早把关联分析从
人类遗传学引入植物遗传学研究领域的团队,2001 年
针对玉米 Dwarf8 进行候选基因关联分析,发现其与
开花期有关[123],要指出的是由于统计学方法的不断发
展,后来的研究表明这可能是由于群体结构造成的假
阳性结果[124]。之后,候选基因关联分析成为多个作物
验证基因功能和挖掘有利等位基因的常用方法。例如,
Wilson 等[125]利用含 102 份玉米自交系的关联分析群
体,应用基因测序的方法对 6 个淀粉合成相关基因
ae1、bt2、sh1、sh2、su1 和 wx1 进行了基因型鉴定,
发现 bt2、sh1 和 sh2 与籽粒组分性状显著相关,ae1和 sh2 与淀粉糊化特性相关,ae1 和 sh1 与直链淀粉含
量相关。Setter 等[126]筛选出 540 个与 ABA 和碳水化
合物代谢有关的候选基因,利用在此基础上构建的
SNP 芯片,对 350 份热带和亚热带玉米自交系中进行
了基因型鉴定,结合不同水分条件下的性状表现,检
测到其中一些基因的等位变异影响到干旱条件下穗部
组织的 ABA 和碳水化合物代谢。 为了挖掘大豆早熟种质的有利等位基因,Tardivel
等[127]针对成熟期基因 GmPhyA3,利用 GBS 方法鉴定
了 53 份大豆近等基因系和 396 份种质资源,发现围绕
该位点存在 6 种不同的单倍型,加拿大东部大豆种质
中最普遍存在的是功能等位基因 E3Ha 和功能缺失等
位基因 e3-tr。Rodriguez 等[128]针对 4 个控制番茄果实
形状的基因 SUN、OVATE、FAS 和 LC,分析了在 368份种质资源中的等位基因分布,发现这 4 个基因的等
位基因分布与形状分类显著相关,认为选择在亚群分
化中起重要作用。Bhullar 等[129-130]从 1 320 份小麦种
质中,分离到白粉病抗性 Pm3 的 7 个等位基因后,又
从来自20个国家的733份小麦种质中分离抗白粉病基
因 Pm3 的等位基因,发现了 2 个新的功能等位基因,
分别来自中国和尼泊尔。 6.4 连锁分析与关联分析相结合是基于种质资源发
掘新基因的重要策略
由于连锁分析和关联分析各有其优势和缺点,通
过关联分析与连锁分析紧密结合,相互验证彼此结果,
可快速地找到控制目标性状的候选基因[131]。Yu 等[132]
和 McMullen 等[133]提出结合关联分析和连锁分析最好
的策略之一是构建巢式关联分析群体( nested
association mapping,NAM),即用一个共同亲本与有
广泛遗传多样性的其他一系列亲本杂交创建重组近交
系群体。在玉米上,康奈尔大学用 B73 分别与 25 个
自交系杂交,构建了一个含约 5 000 个 RIL 的巢式关
联分析群体,捕获了 136 000 个重组事件。该群体后
来用于多个性状的遗传解析,如叶部结构[134]、小斑病
抗性[135]。Cook 等[136]利用玉米 NAM 群体开展了针对
籽粒组分的连锁分析和关联分析,连锁分析表明籽粒
组分性状由 21—26 个 QTL 控制,GWAS 检测到很多
性状-位点关联关系,同时利用由 282 份自交系组成的
关联分析群体进行的 GWAS 和候选基因关联分析的
结果也证实了这些结果。中国也用黄早四与 11 个自交
系组配创建了一个解析中国玉米骨干亲本形成遗传基
础的 NAM 群体,目前正在数据分析中。 Lu 等[137]利用 3 个 RIL 群体和 305 份关联分析群
体开展玉米抗旱性的 QTL 定位研究,发现由于群体大
小、等位基因多样性和等位基因频率增加而使作图效
率大为提高,所有数据进行联合分析可以检测到连锁
分析和关联分析单独检测时检测不到的 QTL,利用单
倍型数据比单一的 SNP 数据更好。Krill 等[138]利用 282份玉米自交系组成的关联分析群体和 3 个 F2 作图群
体,对耐铝性遗传基础进行了剖析,发现在用于关联
分析的 21个候选基因中,6个与耐铝性有显著相关性,
但只有 4 个被连锁分析所证实。 Li 等[139]利用 RNA 测序技术对 368 份玉米自交系
进行基因型鉴定,获得 103 万 SNP,针对籽粒油分含
量和脂肪酸组分进行全基因组关联分析,检测到 74个显著相关的位点,利用表达 QTL 作图、连锁分析、
共表达分析进行验证,发现 26 个与油分含量相关的位
点可解释 83%的表型变异。Wen 等[140]利用 368 份关
联分析群体和 2 个重组近交系群体(B73/By804 和
Zong3/Yu87-1),关联分析群体用含 56 110 个 SNP的芯片和 RNA 测序(授粉后 15 d 籽粒)获得的 106万 SNP 进行基因型鉴定(含 28 769 个基因的表达数
据),RIL 群体用含 1 536 个 SNP 的芯片进行基因型
鉴定,同时用 LC-ESI-MS/MS 系统分析了 3 种环境下
成熟籽粒的 983 种代谢物特征水平,GWAS 结果检测
到 1 459 个显著的性状-位点关系。 发掘黄瓜苦味基因也是一个综合运用连锁分析和
关联分析的典型例证。Bi 控制全株苦味,该基因与另
一个显性基因 Bt 共同控制果实苦味,Qi 等[9]利用群体
基因组学方法发现在驯化过程中对 Bt 的人工选择使
栽培黄瓜苦味消失。Shang 等[141]应用关联分析方法检

3344 中 国 农 业 科 学 48 卷
测了 Bi 在 115 份黄瓜种质中的遗传变异,发现在含
Bi 的基因组区段存在一个效应非常显著的 SNP,使编
码葫芦素生物合成第一种酶的基因产物第 393 位氨基
酸从半胱氨酸变成酪氨酸,从而产生无苦味表型。Bt定位在第 5 染色体上 442 kb 的区间(含 67 个基因)[10],
其中包含苦叶基因 Bl 及其 2 个同源基因,通过表达分
析,预测有一个同源基因更有可能是 Bt 的候选基因。
进一步进行区域水平关联分析,发现该候选基因的调
控区域存在 11 个强烈信号,特别是一个位于起始密码
子 1 601 bp 上游的一个 SNP 与 F2群体中的 Bt 位点共
分离,因此,判断对该候选基因调控区域的选择使栽
培品种中该基因的表达下调,从而使苦味降低。
7 种质创新
由于长期的驯化和遗传改良,当今的优良作物品
种常遇到遗传基础变窄的瓶颈,迫切需要从育成品种
外部导入新基因或引入新的等位变异。由于野生近缘
种和地方品种的遗传多样性远远高于现代品种,因此,
针对地方品种和野生近缘植物的种质创新研究已成为
热点领域[142]。随着基因组学的发展,种质创新研究工
作正由过去的以表型选择为主转变为以分子标记选择
和全基因组选择等为主,外源优异基因的鉴定和利用
不断加快。 野生稻中蕴含着不少有可能用于水稻遗传改良
的基因(如产量基因[143])。野生稻中的有利等位变
异被广泛应用,基因组学方法起到了重要的推动作
用。比如,将东乡野生稻染色体导入到籼稻品种桂朝
2 号中,构建了 159 个导入系,覆盖了东乡野生稻约
67.5%的基因组[144],并从这些导入系中鉴定出一个来
自东乡野生稻的抗旱渗入系 IL23[145]。构建了在粳稻
品种特青背景下的 120 个云南元江野生稻导入系[146],
并利用图位克隆技术获得一个来源于野生稻并控制
水稻从匍匐生长向直立生长转化的基因 PROG1[56]。
这些导入系中存在丰富的表型变异,为进一步鉴定和
利用野生稻的优异等位变异打下了很好的基础。 小麦地方品种和野生近缘物种是拓宽小麦遗传多
样性的主要基因源。例如, Hou 等[147]通过对 1 520份来源不同的小麦种质中影响淀粉合成的蔗糖合成酶
基因 TaSu1(定位在 7A/7B/7D 染色体上)和 TaSu2(定
位在 2A/2B/2D 染色体上)的序列比对,发现这两个
基因在现代育种过程中被强烈选择,对千粒重的提高
作出了贡献,但大量的稀有等位变异存在于地方品种
中。簇毛麦 6VS 携带的白粉病水平抗性基因 Pm21 被
成功地导入小麦中,在小麦育种中被有效利用[148]。为
了鉴定簇毛麦 6VS 携带的 Pm21,Cao 等[149]利用大麦
基因芯片筛选并克隆出一个簇毛麦抗白粉病基因的关
键成员丝氨酸和苏氨酸蛋白激酶基因 Stpk-V,该研究
提供了一个反向遗传学研究策略,用来解决大片段外
源染色体片段目标基因的分离和鉴定困难的问题。
Zhang等[150]利用类似抗病R基因序列开发出 2个标记
可以有效地检测和跟踪中间偃麦草携带的抗黄矮病
(BYDV)基因。Sr33 从近缘物种粗山羊草引入小麦
中,被发现可以增强小麦对 Ug99 新强毒性生理小种
的抗性,Periyannan 等[151]采用图位克隆方法将 Sr33克隆出来,发现其编码一个含有 R 基因功能域的抗性
蛋白。Munns 等[152]报道将二倍体野生栽培一粒小麦
(Triticum monococcum)的耐盐碱基因 TmHKT1;5-A导入四倍体硬粒小麦中,可以增强其抗盐碱能力,在
盐碱土地上生长较对照增产 25%。 玉米栽培品种中蕴含丰富的遗传变异,因此,野
生近缘种的利用相对较少,但也取得一些进展。例如,
中国科学院遗传研究所利用远缘杂交方法将大刍草导
入自交系 330,选育而成遗单 6 号单交种,提高了茎
秆强度和抗倒性,兼抗大、小斑病、青枯病,具有保
绿性能好的特点。河南省农业科学院将大刍草基因导
入自交系掖 478 等优良自交系,从中选育出抗逆性强
的郑远 36、郑远 37 等新系。Amusan 等[153]报道从二
倍体多年生大刍草(Zea diploperennis)和玉米的回交
后代中选出抵抗寄生杂草菟丝子(Striga hermonthica)的自交系。Chia 等[82]认为可以利用摩擦禾属的多年
生、抗寒、抗旱外源等位基因改良玉米。 中国是世界上保存野生大豆资源最多的国家,野
生大豆具有高蛋白、多花多荚丰产特性、对病虫害和
非生物逆境的环境适应能力和人类需求的特殊功能性
状。Concibido 等[154]报道引入野生大豆种质 PI 407305的 QTL 到栽培大豆中,2 年多环境试验表明可以将商
业推广的大豆品种产量提高 9%。为了鉴定和利用野
生大豆种质的优异基因资源,利用野生大豆种质
N24852 为供体,以栽培大豆优良品种 NN1138-2 为受
体,构建了染色体片段置换系[155]。
8 展望
随着多个重要农作物和一些模式植物全基因组测
序的完成和高通量重测序技术的普及,为作物种质资
源研究提供了跨越式发展的机遇[156]。如何把基因组学
理论、方法及其成果与作物种质资源研究的各个环节

17 期 黎裕等:基于基因组学的作物种质资源研究:现状与展望 3345
有机结合起来,提高种质资源保护的安全性和利用的
高效性,已成为中国作物种质资源工作者的重点任务。 8.1 中国目前的国家作物种质资源保存体系中长期
保存库和种质圃保存了 45 万份种质资源(约 2 300 个
物种),此外还建立原生境保护点 163 个。因此,需要
充分应用基因组学理论和方法,对中国保存的种质资源
和野生近缘种居群进行系统评价,科学制定中国种质资
源下一步的收集策略与保护重点。同时,探索各种基因
组学技术方法,为种质资源的有效保护提供新手段。 8.2 尽管中国在种质资源表型精准鉴定和全基因组
水平的基因型鉴定方面开展了部分工作,但涉及的作
物种类及其资源数量极为有限,对库存种质资源的遗
传多样性缺乏系统和深入研究,很难为育种家和基础
理论研究者提供针对性的资源。因此,一方面,应针
对未来育种需求的重要性状,开展多年多点的或控制
环境条件下的表型鉴定评价;另一方面,应充分利用
高通量的测序技术和 SNP 芯片技术,在全基因组学水
平对中国的作物种质资源进行系统的基因型鉴定,并
在此基础上,开展遗传多样性和群体结构等分析,全
面了解中国的种质资源自然遗传变异“家底”。 8.3 作物种质资源具有数量多、覆盖面广的特点,目
前,中国种质资源的深度发掘力度还较为薄弱。因此,
需要充分利用关联分析和连锁分析等手段,在强化重
要性状表型精准鉴定的基础上,发掘能满足未来育种
需求的优异种质资源和基因,使库存种质资源能得到
高效利用。如果进一步拓展,可把表型组学、转录组
学、蛋白组学、代谢组学、表观组学等有机结合起来,
用系统生物学的思路和方法,开展种质资源的变异组
学研究,阐明控制重要性状的遗传和分子基础,挖掘
有利等位基因并得到应用。 8.4 中国的作物种质创新目前在很大程度上还停留
在现代品种的进一步改良上。因此,需要充分应用基
因组学理论和方法(包括分子标记、全基因组选择、
基因组编辑等),重点突破野生近缘种和地方品种的
创新利用瓶颈,不断拓展现代作物育种的遗传基础。 8.5 目前中国的作物种质信息系统中主要涵盖的是
护照信息和基本农艺性状信息,育种家和其他用户感
兴趣的信息不多。因此,需要重建数据库系统框架,
把基因组信息(包括基因型数据、基因信息、等位基
因信息、标记信息等)、目标性状表型信息、谱系信
息、生态环境信息进行整合,研发种质资源新型统计
分析方法,使信息系统数据更实用、界面更友好、使
用更简单、服务更便捷。
References
[1] Semagn K, Babu R, Hearne S, Olsen M. Single nucleotide
polymorphism genotyping using Kompetitive Allele Specific PCR
(KASP): Overview of the technology and its application in crop
improvement. Molecular Breeding, 2014, 33: 1-14.
[2] Hamblin M T, Warburton M L, Buckler E S. Empirical comparison of
simple sequence repeats and single nucleotide polymorphisms in
assessment of maize diversity and relatedness. PLoS ONE, 2007, 2:
e1367.
[3] Van Inghelandt D, Melchinger A, Lebreton C, Stich B. Population
structure and genetic diversity in a commercial maize breeding
program assessed with SSR and SNP markers. Theoretical and
Applied Genetics, 2010, 120: 1289-1299.
[4] Chen H, Xie W, He H, Yu H, Chen W, Li J, Yu R, Yao Y, Zhang W,
He Y, Tang X, Zhou F, Deng X W, Zhang Q. A high-density SNP
genotyping array for rice biology and molecular breeding. Molecular
Plant, 2014, 7: 541-553.
[5] Wang S, Wong D, Forrest K, Allen A, Chao S, Huang B E,
Maccaferri M, Salvi S, Milner S G, Cattivelli L, Mastrangelo A M,
Whan A, Stephen S, Barker G, Wieseke R, Plieske J, International
Wheat Genome Sequencing Consortium, Lillemo M, Mather D,
Appels R, Dolferus R, Brown-Guedira G, Korol A, Akhunova A R,
Feuillet C, Salse J, Morgante M, Pozniak C, Luo M C, Dvorak J,
Morell M, Dubcovsky J, Ganal M, Tuberosa R, Lawley C, Mikoulitch
I, Cavanagh C, Edwards K J, Hayden M, Akhunov E. Characterization
of polyploid wheat genomic diversity using a high-density 90000
single nucleotide polymorphism array. Plant Biotechnology Journal,
2014, 12: 787-796.
[6] Ganal M W, Durstewitz G, Polley A, Bérard A, Buckler E S,
Charcosset A, Clarke J D, Graner E M, Hansen M, Joets J, Le Paslier
M C, McMullen M D, Montalent P, Rose M, Schon C C, Sun Q,
Walter H, Martin O C, Falque M. A large maize (Zea mays L.) SNP
genotyping array: Development and germplasm genotyping, and
genetic mapping to compare with the B73 reference genome. PLoS
ONE, 2011, 6: e28334.
[7] Unterseer S, Bauer E, Haberer G, Seidel M, Knaak C, Ouzunova M,
Meitinger T, Strom T M, Fries R, Pausch H, Bertani C, Davassi A,
Mayer K F X, Schon C C. A powerful tool for genome analysis in
maize: development and evaluation of the high density 600 K SNP
genotyping array. BMC Genomics, 2014, 15: 823.
[8] Lam H M, Xu X, Liu X, Chen W B, Yang G H, Wong F L, Li M W,
He W M, Qin N, Wang B, Li J, Jian M, Wang J, Shao G H, Wang J,
Sun S S M, Zhang G Y. Resequencing of 31 wild and cultivated

3346 中 国 农 业 科 学 48 卷
soybean genomes identifies patterns of genetic diversity and selection.
Nature Genetics, 2010, 42: 1053-1059.
[9] Qi J, Liu X, Shen D, Miao H, Xie B, Li X, Zeng P, Wang S, Shang Y,
Gu X, Du Y, Li Y, Lin T, Yuan J, Yang X, Chen J, Chen H, Xiong X,
Huang K, Fei Z, Mao L, Tian L, Städler T, Renner S S, Kamoun S,
Lucas W J, Zhang Z, Huang S. A genomic variation map provides
insights into the genetic basis of cucumber domestication and
diversity. Nature Genetics, 2013, 45: 1510-1518.
[10] Gore M A, Wright M H, Ersoz E S, Bouffard P, Szekeres E S, Jarvie
T P, Hurwitz B L, Narechania A, Harkins T T, Grills G S, Ware D H,
Buckler E S. Large-scale discovery of gene-enriched SNPs. Plant
Genome, 2009, 2: 121-133.
[11] Van Orsouw N J, Hogers R C J, Janssen A, Yalcin F, Snoeijers S,
Verstege E. Complexity reduction of polymorphic sequences (CRoPS):
A novel approach for large-scale polymorphism discovery in complex
genomes. PLoS ONE, 2007, 2: e1172.
[12] Elshire R J, Glaubitz J C, Sun Q, Poland J A, Kawamoto K, Buckler E
S, Mitchell S E. A robust, simple genotyping-by-sequencing (GBS)
approach for high diversity species. PLoS ONE, 2011, 6: e19379.
[13] Poland J A, Rife T W. Genotyping-by-sequencing for plant breeding
and genetics. Plant Genome, 2012, 5: 92-102.
[14] Sonah H, Bastien M, Iquira E, Tardivel A, Légaré G, Boyle B. An
improved genotyping by sequencing (GBS) approach offering
increased versatility and efficiency of SNP discovery and genotyping.
PloS ONE, 2013, 8: e54603.
[15] Truong H T, Ramos A M, Yalcin F, de Ruiter M, van der Poel H J A,
Huvenaars K H J, Hogers R C J, van Enkevort L J G, Janssen A, van
Orsouw N J, van Eijk M J T. Sequence-based genotyping for marker
discovery and co-dominant scoring in germplasm and populations.
PLoS ONE, 2012, 7: e37565.
[16] Miller M R, Dunham J P, Amores A, Cresko W A, Johnson E A.
Rapid and cost-effective polymorphism identification and genotyping
using restriction site associated DNA (RAD) markers. Genome
Research, 2007, 17: 240-248.
[17] Monson-Miller J, Sanchez-Mendez D, Fass J, Henry I, Tai T H,
Comai L. Reference genome-independent assessment of mutation
density using restriction enzyme-phased sequencing. BMC Genomics,
2012, 13: 72.
[18] Spindel J, Wright M, Chen C, Cobb J, Gage J, Harrington S, Lorieux
M, Ahmadi N, McCouch S. Bridging the genotyping gap: Using
genotyping by sequencing (GBS) to add high-density SNP markers
and new value to traditional bi-parental mapping and breeding
populations. Theoretical and Applied Genetics, 2013, 126: 2699-2716.
[19] Glaubitz J C, Casstevens T M, Lu F, Harriman J, Elshire R J, Sun Q,
Buckler E S. TASSEL-GBS: A high capacity genotyping by
sequencing analysis pipeline. PLoS ONE, 2014, 9: e90346.
[20] Poland J A, Brown P J, Sorrells M E, Jannink J L. Development of
high-density genetic maps for barley and wheat using a novel
two-enzyme genotyping-by-sequencing approach. PLoS ONE, 2012, 7:
e32253.
[21] Russell J, Hackett C, Hedley P, Liu H, Milne L, Bayer M, Marshall D,
Jorgensen L, Gordon S, Brennan R. The use of genotyping by
sequencing in blackcurrant (Ribes nigrum): Developing high-
resolution linkage maps in species without reference genome
sequences. Molecular Breeding, 2014, 33: 835-849.
[22] Ng S B, Turner E H, Robertson P D, Flygare S D, Bigham A W, Lee
C, Shaffer T, Wong M, Bhattacharjee A, Eichler E E, Bamshad M,
Nickerson D A, Shendure J. Targeted capture and massively parallel
sequencing of 12 human exomes. Nature, 2009, 461: 272-276.
[23] Brunner A L, Johnson D S, Kim S W, Valouev A, Reddy T E, Neff N
F, Anton E, Medina C, Nguyen L, Chiao E, Oyolu C, Schroth G P,
Absher D M, Baker J C, Myers R M. Distinct DNA methylation
patterns characterize differentiated human embryonic stem cells and
developing human fetal liver. Genome Research, 2009, 19: 1044-1056.
[24] Barbazuk W B, Schnable P S. SNP discovery by transcriptome
pyrosequencing. Methods in Molecular Biology, 2011, 729: 225-246.
[25] Fu J, Cheng Y, Linghu J, Yang X, Kang L, Zhang Z, Zhang J, He C,
Du X, Peng Z, Wang B, Zhai L, Dai C, Xu J, Wang W, Li X, Zheng J,
Chen L, Luo L, Liu J, Qian X, Yan J, Wang J, Wang G. RNA
sequencing reveals the complex regulatory network in the maize
kernel. Nature Communications, 2013, 4: 2832, DOI:10.1038.
[26] Kiialainen A, Karlberg O, Ahlford A, Sigurdsson S, Lindblad-Toh K,
Syvänen A C. Performance of microarray and liquid based capture
methods for target enrichment for massively parallel sequencing and
SNP discovery. PLoS ONE, 2011, 6: e16486.
[27] Pham A T, Bilyeu K, Chen P, Boerma H R, Li Z. Characterization of
the fan1 locus in soybean line A5 and development of molecular
assays for high-throughput genotyping of FAD3 genes. Molecular
Breeding, 2014, 33: 895-907.
[28] De La Fuente G N, Murray S C, Isakeit T, Park Y S, Yan Y,
Warburton M L, Kolomiets M V. Characterization of genetic diversity
and linkage disequilibrium of ZmLOX4 and ZmLOX5 loci in maize.
PLoS ONE, 2013, 8: e53973.
[29] FAO. The Second Report on The State of The World’s Plant Genetic
Resources for Food and Agriculture. 2010. FAO, Rome, Italy.
[30] Thomson M J, Zhao K, Wright M, McNally K L, Rey J, Tung C W,

17 期 黎裕等:基于基因组学的作物种质资源研究:现状与展望 3347
Scheffler A R B, Eizenga g, McClung A, Kim H, Ismail A M, de
Ocampo M, Mojica C, Reveche M Y, Dilla-Ermita C J, Mauleon R,
Leung H, Bustamante C, McCouch S R. High-throughput single
nucleotide polymorphism genotyping for breeding applications in rice
using the BeadXpress platform. Molecular Breeding, 2012, 29:
875-886.
[31] Xue Y, Zhang S Q, Yao Q H, Peng R H, Xiong A S, Li X, Zhu W M,
Zhu Y Y, Zha D S. Identification of quantitative trait loci for seed
storability in rice (Oryza sativa L.). Euphytica, 2008, 164: 739-744.
[32] Nagel M, Rosenhauer M, Willner E, Snowdon R J, Friedt W, Boerner
A. Seed longevity in oilseed rape (Brassica napus L.) - genetic
variation and QTL mapping. Plant Genetic Resources: Characterization
and Utilization, 2011, 9: 260-263.
[33] Nagel M, Vogel H, Landjeva S, Buck-Sorlin G, Lohwasser U, Scholz
U, Boerner A. Seed conservation in ex situ genebanks - genetic studies
on longevity in barley. Euphytica, 2009, 170: 5-14.
[34] Arif M A R, Nagel M, Neumann K, Kobiljski B, Lohwasser U, Borner
A. Genetic studies of seed longevity in hexaploid wheat using
segregation and association mapping approaches. Euphytica, 2012,
186: 1-13
[35] Harding K, Benson E E. The use of microsatellite analysis in Solanum
tuberosum L. in vitro plantlets derived from cryopreserved germplasm.
Cryoletters, 2001, 22: 199-208.
[36] Kaity A, Ashmore S E, Drew R A, Dulloo M E. Assessment of
genetic and epigenetic changes following cryopreservation in papaya.
Plant Cell Reports, 2008, 27: 1529-1539.
[37] Peredo E L, Arroyo-Garcia R, Reed B M, Revilla M A. Genetic and
epigenetic stability of cryopreserved and cold-stored hops (Humulus
lupulus L.). Cryobiology, 2008, 57: 234-241.
[38] Johnston J W, Benson E E, Harding K. Cryopreservation induces
temporal DNA methylation epigenetic changes and differential
transcriptional activity in Ribes germplasm. Plant Physiology and
Biochemistry, 2009, 47: 123-131.
[39] Volk G M. Application of functional genomics and proteomics to
plant cryopreservation. Current Genomics, 2010, 11: 24-29.
[40] 刘敏, 辛霞, 张志娥, 陈晓玲, 张金梅, 卢新雄. 繁殖群体量及隔
离对蚕豆种质遗传完整性的影响. 植物遗传资源学报, 2012, 13:
175-181.
Liu M, Xin X, Zhang Z E, Chen X L, Zhang J M, Lu X X. Effect of
sample size and isolation methods on genetic integrity of faba bean
(Vicia faba L.) germplasm. Journal of Plant Genetic Resources, 2012,
13: 175-181. (in Chinese)
[41] 王栋, 张志娥, 陈晓玲, 辛霞, 辛萍萍, 卢新雄. AFLP 标记分析生
活力影响大豆中黄 18 种质遗传完整性. 作物学报, 2010, 36(4):
555-564.
Wang D, Zhang Z E, Chen X L, Xin X, Xin P P, Lu X X. Analysis of
viability affecting on genetic integrity in soybean germplasm
Zhonghuang 18 by AFLP markers. Acta Agronomica Sinica, 2010,
36(4): 555-564. (in Chinese)
[42] Bai J M, Chen X L, Lu X X, Xin X, Zhang Z E, Liu X C, Sun B S,
Zhang J M, Yin G K, Sui Q J. Effects of different conservation
methods on the genetic stability of potato germplasm. Russian Journal
of Plant Physiology, 2011, 58: 728-736.
[43] Ren L, Zhang D, Jiang X N, Gai Y, Wang W M, Reed B M, Shen X H.
Peroxidation due to cryoprotectant treatment is a vital factor for cell
survival in Arabidopsis cryopreservation. Plant Science, 2013, 212:
37-47.
[44] Frankham R. Conservation genetics. Annual Review of Genetics, 1995,
29: 305-327.
[45] 杨庆文, 余丽琴, 张万霞, 时津霞, 任军方, 苗晗. 东乡普通野生
稻(Oryza rufipogon Griff.)原位保存群体的遗传分化和保护策略研
究. 中国农业科学 2007, 40(6): 1085-1093.
Yang Q W, Yu L Q, Zhang W X, Shi J X, Ren J F, Miao H. Genetic
differentiation and conservation strategy of in situ conserved
populations of Dongxiang wild rice (Oryza rufipogon Griff.). Scientia
Agricultura Sinica, 2007, 40(6): 1085-1093. (in Chinese)
[46] 王家祥, 陈友桃, 黄娟, 乔卫华, 张万霞, 杨庆文. 中国普通野生
稻原生境保护与未保护居群的遗传多样性比较. 作物学报, 2009,
35(8): 1474-1482.
Wang J X, Chen Y T, Huang J, Qiao W H, Zhang W X, Yang Q W.
Comparison of genetic diversity between in-situ conserved and non-
conserved Oryza rufipogon populations in China. Acta Agronomica
Sinica, 2009, 35(8): 1474-1482. (in Chinese)
[47] Qiao W H, Chen Y T, Wang R S, Wei X, Cao L R, Zhang W X, Yang
Q W. Nucleotide diversity in waxy gene and validation of single
nucleotide polymorphism in relation to amylose content in Chinese
microcore rice germplasm. Crop Science, 2012, 52: 1689-1697.
[48] Wei X, Qiao W H, Chen Y T, Wang R S, Cao L R, Zhang W X, Yuan
N N, Li Z C, Zeng H L, Yang Q W. Domestication and geographic
origin of Oryza sativa in China: Insights from multilocus analysis of
nucleotide variation of O. sativa and O. rufipogon. Molecular Ecology,
2012, 21: 5073-5087.
[49] Wei X, Wang R S, Cao L R, Yuan N N, Huang J, Qiao W H, Zhang W
X, Zeng H L, Yang Q W. Origin of Oryza sativa in China inferred by
nucleotide polymorphisms of organelle DNA. PloS ONE, 2012, 7:
e49546.

3348 中 国 农 业 科 学 48 卷
[50] Henry R J. Sequencing of wild crop relatives to support the
conservation and utilization of plant genetic resources. Plant Genetic
Resources: Characterization and Utilization, 2014, 12: S9-S11.
[51] Doebley J, Stec A, Gustus C. teosinte branched1 and the origin of
maize: Evidence for epistasis and the evolution of dominance.
Genetics, 1995, 141: 333-346.
[52] Studer A, Zhao Q, Ross-Ibarra J, Doebley J. Identification of a
functional transposon insertion in the maize domestication gene tb1.
Nature Genetics, 2011, 43: 1160-1163.
[53] Li X, Qian Q, Fu Z, Wang Y, Xiong G, Zeng D, Wang X, Liu X, Teng
S, Hiroshi F. Control of tillering in rice. Nature, 2003, 422: 618-621.
[54] Li C, Zhou A, Sang T. Rice domestication by reducing shattering.
Science, 2006, 311: 1936-1939.
[55] Konishi S, Izawa T, Lin S Y, Ebana K, Fukuta Y, Sasaki T, Yano M.
An SNP caused loss of seed shattering during rice domestication.
Science, 2006, 312: 1392-1396.
[56] Tan L, Li X, Liu F, Sun X, Li C, Zhu Z, Fu Y, Cai H, Wang X, Xie D.
Control of a key transition from prostrate to erect growth in rice
domestication. Nature Genetics, 2008, 40: 1360-1364.
[57] Shomura A, Izawa T, Ebana K, Ebitani T, Kanegae H, Konishi S, Yano
M. Deletion in a gene associated with grain size increased yields
during rice domestication. Nature Genetics, 2008, 40: 1023-1028.
[58] Cao K, Zheng Z, Wang L, Liu X, Zhu G, Fang W, Cheng S, Zeng P,
Chen C, Wang X. Comparative population genomics reveals the
domestication history of the peach, Prunus persica, and human
influences on perennial fruit crops. Genome Biology, 2014, 15: 415.
[59] Mace E S, Tai S, Gilding E K, Li Y, Prentis P J, Bian L, Campbell B C,
Hu W, Innes D J, Han X. Whole-genome sequencing reveals untapped
genetic potential in Africa's indigenous cereal crop sorghum. Nature
Communications, 2013, 4: 2320.
[60] Schmutz J, McClean E, Mamidi S, Wu G A, Cannon S B, Grimwood
J, Jenkins J, Shu S, Song Q, Chavarro C, Torrres-Torres M, Geffroy V,
Moghaddam S M, Gao D, Abernathy B, Barry K, Blair M, Brick M,
Chovatia M, Gepts P, Goodstein D M, Gonzales M, Hellsten U, Hyten
D L, Jia G, Kelly J D, Kudrna D, Lee R, Richard M M S, Miklas P N,
Osorno J M, Rodrigues J, Thareau V, Urrea C A, Wang M, Yu Y,
Zhang M, Wing R A, Cregan P B, Rokhsar D S, Jackson S A. A
reference genome for common bean and genome-wide analysis of
dual domestications. Nature Genetics, 2014, 46: 707-713.
[61] Huang X, Kurata N, Wei X, Wang Z X, Wang A, Zhao Q, Zhao Y, Liu
K, Lu H, Li W. A map of rice genome variation reveals the origin of
cultivated rice. Nature, 2012, 490: 497-501.
[62] Wang M, Yu Y, Haberer G, Marri P R, Fan C, Goicoechea J L, Zuccolo
A, Song X, Kudrna D, Ammiraju J S S. The genome sequence of
African rice (Oryza glaberrima) and evidence for independent
domestication. Nature Genetics, 2014, 46: 982-988.
[63] Xu X, Liu X, Ge S, Jensen J D, Hu F, Li X, Dong Y, Gutenkunst R N,
Fang L, Huang L, Li J, He W, Zhang G, Zheng X, Zhang F, Li Y, Yu C,
Kristiansen K, Zhang X, Wang J, Wright M, McCouch S, Nielsen R,
Wang J, Wang W. Resequencing 50 accessions of cultivated and wild
rice yields markers for identifying agronomically important genes.
Nature Biotechnology, 2012, 30: 105-111.
[64] Hufford M B, Xu X, Van Heerwaarden J, Pyhäjärvi T, Chia J M,
Cartwright R A, Elshir R J, Glaubitz J C, Guill K E, Kaeppler S M,
Lai J, Morrell P L, Shannon L M, Song C, Springer N M,
Swanson-Wagner R A, Tiffin P, Wang J, Zhang G, Doebley J,
McMullen M D, Ware D, Buckler E, Yang S, Ross-Ibarra J.
Comparative population genomics of maize domestication and
improvement. Nature Genetics, 2012, 44: 808-811.
[65] Li Y H, Zhao S C, Ma J X, Li D, Yan L, Li J, Qi X T, Guo X S, Zhang
L, He W M, Chang R Z, Liang Q S, Guo Y, Ye C, Wang X B, Tao Y,
Guan R X, Wang J Y, Liu Y L, Jin L G, Zhang X Q, Liu Z X, Zhang L
J, Chen J, Wang K J, Nielsen R, Li R Q, Chen P Y, Li W B, Reif J C,
Purugganan M, Wang J, Zhang M C, Wang J, Qiu L J. Molecular
footprints of domestication and improvement in soybean revealed by
whole genome re-sequencing. BMC Genomics, 2013, 14: 579.
[66] Lin T, Zhu G, Zhang J, Xu X, Yu Q, Zheng Z, Zhang Z, Lun Y, Li S,
Wang X, Huang Z, Li J, Zhang C, Wang T, Zhang Y, Wang A, Zhang Y,
Lin K, Li C, Xiong G, Xue Y, Mazzucato A, Causse M, Fei Z,
Giovannoni J J, Chetelat R T, Zamir D, Stadler T, Li J, Ye Z, Du Y,
Huang S. Genomic analyses provide insights into the history of
tomato breeding. Nature Genetics, 2014, 46: 1220-1226.
[67] Guo S, Zhang J, Sun H, Salse J, Lucas W J, Zhang H, Zheng Y, Mao L,
Ren Y, Wang Z. The draft genome of watermelon (Citrullus lanatus)
and resequencing of 20 diverse accessions. Nature Genetics, 2013, 45:
51-58.
[68] Tanksley S D, McCouch S R. Seed banks and molecular maps:
Unlocking genetic potential from the wild. Science, 1997, 277:
1063-1066.
[69] Qin C, Yu C, Shen Y, Fang X, Chen L, Min J, Cheng J, Zhao S, Xu M,
Luo Y. Whole-genome sequencing of cultivated and wild peppers
provides insights into Capsicum domestication and specialization.
Proceedings of the National Academy of Sciences of the USA, 2014,
111: 5135-5140.
[70] Frary A, Nesbitt T C, Grandillo S, Knaap E, Cong B, Liu J, Meller J,
Elber R, Alpert K B, Tanksley S D. A quantitative trait locus key to

17 期 黎裕等:基于基因组学的作物种质资源研究:现状与展望 3349
the evolution of tomato fruit size. Science, 2000, 289: 85-88.
[71] Cook D E, Lee T G, Guo X, Melito S, Wang K, Bayless A, Wang J,
Hughes T J, Willis D K, Clemente T. Copy number variation of
multiple genes at Rhg1 mediates nematode resistance in soybean.
Science, 2012, 338: 1206-1209.
[72] Li Y H, Zhou G, Ma J, Jiang W, Jin L G, Zhang Z, Guo Y, Zhang J,
Sui Y, Zheng L, Zhang S S, Zuo Q, Shi XH, Li Y F, Zhang W K, Hu
Y, Kong G, Hong H L, Tan B, Song J, Liu Z X, Wang Y, Ruan H,
Yeung CK, Liu J, Wang H, Zhang L J, Guan R X, Wang K J, Li W B,
Chen S Y, Chang R Z, Jiang Z, Jackson S A, Li R, Qiu L J. De novo
assembly of soybean wild relatives for pan-genome analysis of
diversity and agronomic traits. Nature Biotechnology, 2014: 32:
1045-1052 .
[73] Mir C, Zerjal T, Combes V, Dumas F, Madur D, Bedoya C,
Dreisigacker S, Franco J, Grudloyma P, Hao P X. Out of America:
Tracing the genetic footprints of the global diffusion of maize.
Theoretical and Applied Genetics, 2013, 126: 2671-2682.
[74] Wang N, Li F, Chen B, Xu K, Yan G, Qiao J, Li J, Gao G, Bancroft I,
Meng J, King G J, Wu X. Genome-wide investigation of genetic
changes during modern breeding of Brassica napus. Theoretical and
Applied Genetics, 2014, 127: 1817-1829.
[75] Jiao Y, Zhao H, Ren L, Song W, Zeng B, Guo J, Wang B, Liu Z,
Chen J, Li W, Zhang M, Xie S, Lai J. Genome-wide genetic changes
during modern breeding of maize. Nature Genetics, 2012, 44:
812-815.
[76] Dang X, Thi T G T, Dong G, Wang H, Edzesi W M, Hong D. Genetic
diversity and association mapping of seed vigor in rice (Oryza sativa
L.). Planta, 2014, 239: 1309-1319.
[77] The 100 Tomato Genome Sequencing Consortium, Aflitos S, Schijlen
E, de Jong H, de Ridder D, Smit S, Finkers R, Wang J, Zhang G, Li N,
Mao L, Bakker F, Dirks R, Breit T, Gravendeel B, Huits H, Struss D,
Swanson-Wagner R, van Leeuwen H, van Ham R C H J, Fito L,
Guignier L, Sevilla M, Ellul P, Ganko E, Kapur A, Reclus E, de Geus
B, van de Geest H, te Lintel Hekkert B, van Haarst J, Smits L, Koops
A, Sanchez-Perez G, van Heusden A W, Visser R, Quan Z, Min J,
Liao L, Wang X, Wang G, Yue Z, Yang X, Xu N, Schranz E, Smets E,
Vos R, Rauwerda J, Ursem R, Schuit C, Kerns M, van den Berg J,
Vriezen W, Janssen A, Datema E, Jahrman T, Moquet F, Bonnet J,
Peters S. Exploring genetic variation in the tomato (Solanum section
Lycopersicon) clade by whole-genome sequencing. The Plant Journal,
2014, 80: 136-148.
[78] Wang L, Han X, Zhang Y, Li D, Wei X, Ding X, Zhang X. Deep
resequencing reveals allelic variation in Sesamum indicum. BMC
Plant Biology, 2014, 14: 225.
[79] The International Peach Genome Initiative. The high-quality draft
genome of peach (Prunus persica) identifies unique patterns of
genetic diversity, domestication and genome evolution. Nature
Genetics, 2013, 45: 487-494.
[80] Varshney R K, Song C, Saxena R K, Azam S, Yu S, Sharpe A G,
Cannon S, Baek J, Rosen B D, Tar’an B, Millan T, Zhang X, Ramsay
L, Iwata A, Wang Y, Nelson W, Farmer A D, Gaur P M, Soderlund C,
Penmetsa R V, Xu C, Bharti A K, He W, Winter P, Zhao S, Hane J K,
Carrasquilla-Garcia N, Condie J A, Upadhyaya H D, Luo M C, Thudi
M, Gowda C L L, Singh N P, Lichtenzveig J, Gali K K, Rubio J,
Nadarajan N, Dolezel J, Bansal K C, Xu X, Edwards D, Zhang G,
Kahl G, Gil J, Singh K B, Datta S K, Jackson S A, Wang J, Cook D R.
Draft genome sequence of chickpea (Cicer arietinum) provides a
resource for trait improvement. Nature Biotechnology, 2013, 31:
240-246.
[81] Zeng X, Long H, Wang Z, Zhao S, Tang Y, Huang Z, Wang Y, Xu Q,
Mao L, Deng G, Yao X, Li X, Bai L, Yuan H, Pan Z, Liu R, Chen X,
WangMu Q, Chen M, Yu L, Liang J, DunZhu D, Zheng Y, Yu S,
LuoBu Z, Guang X, Li J, Deng C, Hu W, Chen C, TaBa X, Gao L, Lv
X, Abuf Y B, Fang X, Nevo E, Yu M, Wang J, Tashia N. The draft
genome of Tibetan hulless barley reveals adaptive patterns to the high
stressful Tibetan Plateau. Proceedings of the National Academy of
Sciences of the USA, 2015, 112: 1095-1100.
[82] Chia J M, Song C, Bradbury P J, Costich D, de Leon N, Doebley J,
Elshire R J, Gaut B, Geller L, Glaubitz J C, Gore M, Guill K E,
Holland J, Hufford M B, Lai J, Li M, Liu X, Lu Y, McCombie R,
Nelson R, Poland J, Prasanna B M, Pyhajarvi T, Rong T, Sekhon R S,
Sun Q, Tenaillon M I, Tian F, Wang J, Xu X, Zhang Z, Kaeppler S M,
Ross-Ibarra J, McMullen M D, Buckler E S, Zhang G, Xu Y, Ware D.
Maize HapMap2 identifies extant variation from a genome in flux.
Nature Genetics, 2012, 44: 803-807.
[83] Odong T L, van Heerwaarden J, van Hintum T J L, van Eeuwijk F A,
Jansen J. Improving hierarchical clustering of genotypic data via
principal component analysis. Crop Science, 2013, 53: 1546-1554.
[84] Pritchard J K, Stephens M, Donnelly P. Inference of population
structure using multilocus genotype data. Genetics, 2000, 155:
945-959.
[85] Raj A, Stephens M, Pritchard J K. fastSTRUCTURE: Variational
inference of population structure in large SNP data sets. Genetics,
2014, 197: 573-589.
[86] Frascaroli E, Schrag T A, Melchinger A E. Genetic diversity analysis
of elite European maize (Zea mays L.) inbred lines using AFLP, SSR,

3350 中 国 农 业 科 学 48 卷
and SNP markers reveals ascertainment bias for a subset of SNPs.
Theoretical and Applied Genetics, 2013, 126: 133-141.
[87] Munoz-Amatriain M, Cuesta-Marcos A, Endelman J B, Comadran J,
Bonman J M, Bockelman H E, Chao S, Russell J, Waugh R, Patrick
M H, Muehlbauer G J. The USDA barley core collection: Genetic
diversity, population structure, and potential for genome-wide
association studies. PLoS ONE, 2014, 9(4): e94688.
[88] Comadran J, Ramsay L, MacKenzie K, Hayes P, Close T J,
Muehlbauer G, Stein N, Waugh R. Patterns of polymorphism and
linkage disequilibrium in cultivated barley. Theoretical and Applied
Genetics, 2011, 122: 523-531.
[89] Hamblin M T, Close T J, Bhat P R, Chao S, Kling J G, Abraham K J,
Blake T, Brooks W S, Cooper B, Griff ey C A, Hayes P M, Hole D J,
Horsley R D, Obert D E, Smith K P, Ullrich S E, Muehlbauer G J,
Jannink J L. Population structure and linkage disequilibrium in U.S.
barley germplasm: Implications for association mapping. Crop
Science, 2010, 50: 556-566.
[90] Cavanagh C R, Chao S, Wang S, Huang B E, Stephen S, Kiani S,
Forrest K, Saintenac C, Brown-Guedira G L, Akhunova A, See D, Bai
G, Pumphrey M, Tomar L, Wong D, Kong S, Reynolds M, da Silva M
L, Bockelman H, Talbert L, Anderson J A, Dreisigacker S, Baenziger
S, Carteri A, Korzun V, Morrell P L, Dubcovsky J, Morell M K, Mark
E. Sorrells M E, Matthew J. Hayden M J, Akhunov E. Genome-wide
comparative diversity uncovers multiple targets of selection for
improvement in hexaploid wheat landraces and cultivars. Proceedings
of the National Academy of Sciences of the USA, 2013, 110:
8057-8062.
[91] Wurschum T, Langer, S M, Longin C F H, Korzun V, Akhunov E,
Ebmeyer E, Schachschneider R, Schacht J, Kazman E, Reif J C.
Population structure, genetic diversity and linkage disequilibrium in
elite winter wheat assessed with SNP and SSR markers. Theoretical
and Applied Genetics, 2013, 126: 1477-1486.
[92] Yan J, Shah T, Warburton M L, Buckler E S, McMullen M D, Crouch
J. Genetic characterization and linkage disequilibrium estimation of a
global maize collection using SNP markers. PLoS ONE, 2009, 4:
e8451.
[93] Lu Y, Shah T, Hao Z, Taba S, Zhang S, Gao S, Liu J, Cao M, Wang J,
Prakash A B, Rong T, Xu Y. Comparative SNP and haplotype
analysis reveals a higher genetic diversity and rapider LD decay in
tropical than temperate germplasm in maize. PLoS ONE, 2011, 6:
e24861.
[94] Wu X, Li Y, Shi Y, Song Y, Wang T, Huang Y, Li Y. Fine genetic
characterization of elite maize germplasm using high-throughput SNP
genotyping. Theoretical and Applied Genetics, 2014, 127: 621-631.
[95] Schaefer C M, Bernardo R. Population structure and single nucleotide
polymorphism diversity of historical Minnesota maize inbreds. Crop
Science, 2013, 53: 1529-1536.
[96] Romay M C, Millard M J, Glaubitz J C, Jason A, Peiffer J A, Swarts
K L, Casstevens T M, Elshire R J, Acharya C B, Mitchell S E,
Flint-Garcia S A, McMullen M D, Holland J B, Edward S, Buckler E
S, Gardner C A. Comprehensive genotyping of the USA national
maize inbred seed bank. Genome Biology, 2013, 14: R55.
[97] Sim S C, Van Deynze A, Stoffel K, Douches D S, Zarka D, Ganal M
W, Chetelat R T, Hutton S F, Scott J W, Gardner R G, Panthee D R,
Mutschler M, Myers J R, Francis D M. High-density SNP genotyping
of tomato (Solanum lycopersicum L.) reveals patterns of genetic
variation due to breeding. PLoS ONE, 2012, 7: e45520.
[98] Wang Y H, Upadhyaya H D, Burrell A M, Sahraeian S M E, Klein R
R, Klein P E. Genetic structure and linkage disequilibrium in a diverse,
representative collection of the C4 model plant, Sorghum bicolor. G3:
Genes, Genomes, Genetics, 2013, 3: 783-793.
[99] 肖永贵, 路亚明, 闻伟锷, 陈新民, 夏先春, 王德森, 李思敏, 童依
平, 何中虎. 小麦骨干亲本京 411及衍生品种苗期根部性状的遗传.
中国农业科学, 2014, 47(15): 2916-292.
Xiao Y G, Lu Y M, Wen W E, Chen X M, Xia X C, Wang D S, Li S
M, Tong Y P, He Z H. Genetic contribution of seedling root traits
among elite wheat parent Jing 411 to its derivatives. Scientia
Agricultura Sinica, 2014, 47(15): 2916-2926. (in Chinese)
[100] Chen W, Chen H, Zheng T, Yu R, Terzaghi W B, Li Z, Deng X W,
Xu J, He Hang. Highly efficient genotyping of rice biparental
populations by GoldenGate assays based on parental resequencing.
Theoretical and Applied Genetics, 2014, 127: 297-307.
[101] Huang X, Feng Q, Qian Q, Zhao Q, Wang L, Wang A, Guan J, Fan D,
Weng Q, Huang T, Dong G, Sang T, Han B. Highthroughput
genotyping by whole-genome resequencing. Genome Research, 2009,
19: 1068-1076.
[102] Xie W, Feng Q, Yu H, Huang X, Zhao Q, Xing Y, Yu S, Han B,
Zhang Q. Parent-independent genotyping for constructing an
ultrahigh-density linkage map based on population sequencing.
Proceedings of the National Academy of Sciences of the USA, 2010,
107: 10578-10583.
[103] Liu H, Bayer M, Druka A, Russell J R, Hackett C A, Poland J,
Ramsay L, Hedley P E, Waugh R. An evaluation of genotyping by
sequencing (GBS) to map the Breviaristatum-e (ari-e) locus in
cultivated barley. BMC Genomics, 2014, 15: 104.
[104] Yu J, Pressoir G, Briggs W H, Bi I V, Yamasaki M, Doebley J F,

17 期 黎裕等:基于基因组学的作物种质资源研究:现状与展望 3351
McMullen M D, Gaut B S, Nielsen D M, Holland J B, Kresovich S,
Buckler E S. A unified mixed-model method for association mapping
that accounts for multiple levels of relatedness. Nature Genetics, 2006,
38: 203-208.
[105] Zhang Z, Ersoz E, Lai C Q, Todhunter R J, Tiwari H K, Gore M A,
Bradbury P J, Yu J, Arnett D K, Ordovas J M, Buckler E S. Mixed
linear model approach adapted for genome-wide association studies.
Nature Genetics, 2010, 42: 355-360.
[106] Korte A, Bjarni J Vilhjálmsson B J, Segura V, Platt A, Long Q,
Nordborg M. A mixed-model approach for genome-wide association
studies of correlated traits in structured populations. Nature Genetics,
2012, 44: 1066-1071.
[107] Segura V, Vilhjálmsson B J, Platt A, Korte A, Seren U, Long Q,
Nordborg M. An efficient multi-locus mixed-model approach for
genome-wide association studies in structured populations. Nature
Genetics, 2012, 44: 825-830.
[108] Wang Q, Tian F, Pan Y, Buckler E S, Zhang Z. A SUPER powerful
method for genome wide association study. PLoS ONE, 2014, 9:
e107684.
[109] Yang N, Lu Y, Yang X, Huang J, Zhou Y, Ali F, Wen W, Liu J, Li J,
Yan J B. Genome wide association studies using a new nonparametric
model reveal the genetic architecture of 17 agronomic traits in an
enlarged maize association panel. PLoS Genetics, 2014, 10: e1004573.
[110] Chen W, Gao Y, Xie W, Gong L, Lu K, Wang W, Li Y, Liu X, Zhang
H, Dong H, Zhang W, Zhang L, Yu S, Wang G, Lian X, Luo J.
Genome-wide association analyses provide genetic and biochemical
insights into natural variation in rice metabolism. Nature Genetics,
2014, 46: 714-721.
[111] Huang X, Wei X, Sang T, Zhao Q, Feng Q, Zhao Y, Li C, Zhu C, Lu
T, Zhang Z, Li M, Fan D, Guo Y, Wang A, Wang L, Deng L, Li W,
Lu Y, Weng Q, Liu K, Huang T, Zhou T, Jing Y, Li W, Lin Z,
Buckler E S, Qian Q, Zhang Q F, Li J, Han B. Genome-wide
association studies of 14 agronomic traits in rice landraces. Nature
Genetics, 2010, 42: 961-967.
[112] Huang X, Zhao Y, Wei X, Li C, Wang A, Zhao Q, Li W, Guo Y,
Deng L, Zhu C, Fan D, Lu Y, Weng Q, Liu K, Zhou T, Jing Y, Si L,
Dong G, Huang T, Lu T, Feng Q, Qian Q, Li J, Han B. Genome-wide
association study of flowering time and grain yield traits in a
worldwide collection of rice germplasm. Nature Genetics, 2012, 44:
32-39.
[113] Zegeye H, Rasheed A, Makdis F, Badebo A, Ogbonnaya F C.
Genome-wide association mapping for seedling and adult plant
resistance to stripe rust in synthetic hexaploid wheat. PLoS ONE,
2014, 9: e105593.
[114] Liu S, Yang X, Zhang D, Bai G, Chao S, Bockus W. Genome-wide
association analysis identified SNPs closely linked to a gene resistant
to soil-borne wheat mosaic virus. Theoretical and Applied Genetics,
2014, 127: 1039-1047.
[115] Bouchet S, Servin B, Bertin P, Madur D, Combes V, Dumas F, Brunel
D, Laborde J, Charcosset A, Nicolas S. Adaptation of maize to
temperate climates: Mid-density genome-wide association genetics
and diversity patterns reveal key genomic regions, with a major
contribution of the Vgt2 (ZCN8) locus. PLoS ONE, 2013, 8: e71377.
[116] Riedelsheimer C, Lisec J, Czedik-Eysenberg A, Sulpice R, Flis A,
Grieder C, Altmann T, Stitt M, Willmitzer L, Melchinger A E.
Genome-wide association mapping of leaf metabolic profiles for
dissecting complex traits in maize. Proceedings of the National
Academy of Sciences of the USA, 2012, 109: 8872-8877.
[117] Strigens A, Freitag N M, Gilbert X, Grieder C, Riedelsheimer C,
Schrag T A, Messmer R, Melchinger A E. Association mapping for
chilling tolerance in elite flint and dent maize inbred lines evaluated in
growth chamber and field experiments. Plant Cell and Environment,
2013, 36: 1871-1887.
[118] Jia G, Huang X, Zhi H, Zhao Y, Zhao Q, Li W, Chai Y, Yang L, Liu
K, Lu H, Zhu C, Lu Y, Zhou C, Fan D, Weng Q, Guo Y, Huang T,
Zhang L, Lu T, Feng Q, Hao H, Liu H, Lu P, Zhang N, Li Y, Guo E,
Wang S, Wang S, Liu J, Zhang W, Chen G, Zhang B, Li W, Wang Y,
Li H, Zhao B, Li J, Diao X, Han B. A haplotype map of genomic
variations and genome-wide association studies of agronomic traits in
foxtail millet (Setaria italica). Nature Genetics, 2013, 45: 957-961.
[119] Sauvage C, Segura V, Bauchet G, Stevens R, Do P T, Nikoloski Z,
Fernie A R, Causse M. Genome-wide association in tomato reveals 44
candidate loci for fruit metabolic traits. Plant Physiology, 2014, 165:
1120-1132.
[120]Cockram J, White J, Zuluaga D L, Smith D, Comadran J, Macaulay M,
Luo Z, Kearsey M J, Werner P, Harrap D, Tapsell C, Liu H, Hedley P
E, Stein N, Schulte D, Steuernagel B, Marshall D F, Thomas W T,
Ramsay L, Mackay I, Balding D J, AGOUEB Consortium, Waugh R,
O'Sullivan D M. Genome-wide association mapping to candidate
polymorphism resolution in the unsequenced barley genome.
Proceedings of the National Academy of Sciences of the USA, 2010,
107: 21611-21616.
[121] Wang M, Jiang N, Jia T, Leach L, Cockram J, Waugh R, Ramsay L,
Bill Thomas B, Lu Z. Genome-wide association mapping of
agronomic and morphologic traits in highly structured populations of
barley cultivars. Theoretical and Applied Genetics, 2012, 124:

3352 中 国 农 业 科 学 48 卷
233-246.
[122] Zhou H, Steffenson B J, Muehlbauer G, Wanyera R, Njau P, Ndeda S.
Association mapping of stem rust race TTKSK resistance in US barley
breeding germplasm. Theoretical and Applied Genetics, 2014, 127:
1293-1304.
[123] Thornsberry J M, Goodman M M, Doewbley J, Kresovich S, Nielsen
D, Buckler E S. Dwarf8 polymorphisms associate with variation in
flowering time. Nature Genetics, 2001, 28: 286-289.
[124] Larsson S J, Lipka A E, Buckler E S. Lessons from Dwarf8 on the
strengths and weaknesses of structured association mapping. PLoS
Genetics, 2013, 9: e1003246.
[125] Wilson L M, Whitt S R, Ibanez A M, Rocheford T R, Goodman M M,
Buckler E S. Dissection of maize kernel composition and starch
production by candidate gene association. The Plant Cell, 2004, 16:
2719-2733.
[126] Setter T L, Yan J, Warburton M, Ribaut J M, Xu Y, Sawkins M,
Buckler E S, Zhang Z, Gore M A. Genetic association mapping
identifies single nucleotide polymorphisms in genes that affect
abscisic acid levels in maize floral tissues during drought. Journal of
Experimental Botany, 2011, 62: 701-716.
[127] Tardivel A, Sonah H, Belzile F, O’Donoughue L S. Rapid
identification of alleles at the soybean maturity gene E3 using
genotyping by sequencing and a haplotype-based approach. Plant
Genome, 2014, 7: 1-9.
[128] Rodriguez G R, Munos S, Anderson C, Sim S C, Michel A, Causse M,
Gardener B B M, Francis D, van der Knaap E. Distribution of SUN,
OVATE, LC, and FAS in the tomato germplasm and the relationship to
fruit shape diversity. Plant Physiology, 2011, 156: 275-285.
[129] Bhullar N K, Street K, Mackay M, Yahiaouia N, Keller B. Unlocking
wheat genetic resources for the molecular identification of previously
undescribed functional alleles at the Pm3 resistance locus.
Proceedings of the National Academy of Sciences of the USA, 2009,
106: 9519-9524.
[130] Bhullar N K, Zhang Z, Wicker T, Keller B. Wheat gene bank
accessions as a source of new alleles of the powdery mildew
resistance gene Pm3: A large scale allele mining project. BMC Plant
Biology, 2010, 10: 88.
[131] Myles S, Peiffer J, Brown P J, Ersoz E S, Zhang Z, Costich D E,
Buckler E S. Association mapping: Critical considerations shift from
genotyping to experimental design. The Plant Cell, 2009, 21:
2194-2202.
[132] Yu J, Holland J B, McMullen M D, Buckler E S. Genetic design and
statistical power of nested association mapping in maize. Genetics,
2008, 178: 539-551.
[133] McMullen M D, Kresovich S, Villeda H S, Bradbury P, Li H, Sun Q,
Flint-Garcia S, Thornsberry J, Acharya C, Bottoms C, Brown P,
Browne C, Eller M, Guill K, Harjes C, Kroon D, Lepak N, Mitchell S
E, Peterson B, Pressoir G, Romero S, Rosas M O, Salvo S, Yeates H,
Hanson M, Jones E, Smith S, Glaubitz J, Goodman M, Ware D,
Holland J B, Buckler E S. Genetic properties of the maize nested
association mapping population. Science, 2009, 325: 737-740.
[134] Tian F, Bradbury P J, Brown P J, Hung H, Sun Q, Flint-Garcia S,
Rocheford T R, McMullen M D, Holland J B, Buckler E S.
Genome-wide association study of leaf architecture in the maize
nested association mapping population. Nature Genetics, 2011, 43:
159-162.
[135] Kump K L, Bradbury P J, Wisser R J, Buckler E S, Belcher A R,
Oropeza-Rosas M A, Zwonitzer J C, Kresovich S, McMullen M D,
Ware D, Balint-Kurti P J, Holland J B. Genome-wide association
study of quantitative resistance to southern leaf blight in the maize
nested association mapping population. Nature Genetics, 2011, 43:
163-168.
[136] Cook J P, McMullen M D, Holland J B, Tian F, Bradbury P,
Ross-Ibarra J, Buckler E S, Flint-Garcia S A. Genetic architecture of
maize kernel composition in the nested association mapping and
inbred association panels. Plant Physiology, 2012, 158: 824-834.
[137] Lu Y, Zhang S, Shah T, Xie C, Hao Z, Li X, Farkhari M, Ribaut J M,
Cao M, Rong T, Xu Y. Joint linkage–linkage disequilibrium mapping
is a powerful approach to detecting quantitative trait loci underlying
drought tolerance in maize. Proceedings of the National Academy of
Sciences of the USA, 2010, 107: 19585-19590.
[138] Krill A M, Kirst M, Kochian L V, Buckler E S, Hoekenga O A.
Association and linkage analysis of aluminum tolerance genes in
maize. PLoS ONE, 2010, 5: e9958.
[139]Li H, Peng Z, Yang X, Wang W, Fu J, Wang J, Han Y, Chai Y, Guo T,
Yang N, Liu J, Warburton M L, Cheng Y, Hao X, Zhang P, Zhao J,
Liu Y, Wang G, Li J, Yan J. Genome-wide association study dissects
the genetic architecture of oil biosynthesis in maize kernels. Nature
Genetics, 2013, 45: 43-50.
[140] Wen W, Li D, Li X, Gao Y, Li W, Li H, Liu J, Liu H, Chen W, Luo J,
Yan J. Metabolome-based genome-wide association study of maize
kernel leads to novel biochemical insights. Nature Communications,
2014, 5: 3438.
[141] Shang Y, Ma Y, Zhou Y, Zhang H, Duan L, Chen H, Zeng J, Zhou Q,
Wang S, Gu W, Liu M, Ren J, Gu X, Zhang S, Wang Y, Yasukawa K,
Bouwmeester H J, Qi X, Zhang Z, Lucas W J, Huang S. Biosynthesis,

17 期 黎裕等:基于基因组学的作物种质资源研究:现状与展望 3353
regulation, and domestication of bitterness in cucumber. Science, 2014,
346: 1084-1088.
[142]Able J A, Langridge P, Milligan A S. Capturing diversity in the cereals:
Many options but little promiscuity. Trends in Plant Science, 2007, 12:
71-79.
[143] Xiao J, Grandillo S, Ahn S N, McCouch S R, Tanksley S D, Li J,
Yuan L Y. Genes from wild rice improve yield. Nature, 1996, 384:
223-224.
[144] Tian F, Li D J, Fu Q, Zhu Z F, Fu Y C, Wang X K, Sun C Q.
Construction of introgression lines carrying wild rice (Oryza
rufipogon Griff.) segments in cultivated rice (Oryza sativa L.)
background and characterization of introgressed segments associated
with yield-related traits. Theoretical and Applied Genetics, 2006, 112:
570-580.
[145] Zhang X, Zhou S, Fu Y, Su Z, Wang X, Sun C. Identification of a
drought tolerant introgression line derived from Dongxiang common
wild rice (O. rufipogon Griff.). Plant Molecular Biology, 2006, 62:
247-259.
[146] Tan L, Liu F, Xue W, Wang G, Ye S, Zhu Z, Fu Y, Wang X, Sun C.
Development of Oryza rufipogon and O. sativa introgression lines and
assessment for yield-related quantitative trait loci. Journal of
Integrative Plant Biology, 2007, 49: 871-884.
[147] Hou J, Jiang Q, Hao C, Wang Y, Zhang H, Zhang X. Global selection
on sucrose synthase haplotypes during a century of wheat breeding.
Plant Physiology, 2014, 164: 1918-1929.
[148] Chen P D, Qi L L, Zhou B, Zhang S Z, Liu D J. Development and
molecular cytogenetic analysis of wheat-Haynaldia villosa 6VS/6AL
translocation lines specifying resistance to powdery mildew.
Theoretical and Applied Genetics, 1995, 91: 1125-1128.
[149] Cao A, Xing L, Wang X, Yang X, Wang W, Sun Y, Qian C, Ni J, Chen
Y, Liu D, Wang X, Chen P. Serine/threonine kinase gene Stpk-V, a key
member of powdery mildew resistance gene Pm21, confers powdery
mildew resistance in wheat. Proceedings of the National Academy of
Sciences of the USA, 2011, 108: 7727-7732.
[150] Zhang Z, Xu J, Xu Q, Larkin P, Xin Z. Development of novel PCR
markers linked to the BYDV resistance gene Bdv2 useful in wheat for
marker-assisted selection. Theoretical and Applied Genetics, 2004,
109: 433-439.
[151] Periyannan S, Moore J, Ayliffe M, Bansal U, Wang X, Huang L, Deal
K, Luo M, Kong X, Bariana H, Mago R, McIntosh R, Dodds P,
Dvorak J, Lagudah E. The gene Sr33, an ortholog of barley Mla genes,
encodes resistance to wheat stem rust race Ug99. Science, 2013, 341:
786-788.
[152]Munns R, James R A, Xu B, Athman A, Conn S J, Jordans C, Byrt C S,
Hare R A, Tyerman S D, Tester M, Plett D, Gilliham M. Wheat grain
yield on saline soils is improved by an ancestral Na(+) transporter
gene. Nature Biotechnology, 2012, 30: 360-364.
[153] Amusan I O, Rich P J, Menkir A, Housley T, Ejeta G. Resistance to
Striga hermonthica in a maize inbred line derived from Zea
diploperennis. New Phytologist, 2008, 178: 157-166.
[154] Concibido V C, La Vallee B, McLaird P, Pineda N, Meyer J, Hummel
L, Yang J, Wu K, Delannay X. Introgression of a quantitative trait
locus for yield from Glycine soja into commercial soybean cultivars.
Theoretical and Applied Genetics, 2003, 106: 575-582.
[155] Wang W, He Q, Yang H, Xiang S, Zhao T, Gai J. Development of a
chromosome segment substitution line population with wild soybean
(Glycine soja Sieb. et Zucc.) as donor parent. Euphytica, 2013, 189:
293-307.
[156] Morrell P L, Buckler E S, Ross-Ibarra J. Crop genomics: advances
and applications. Nature Reviews: Genetics, 2012, 13: 85-96.
(责任编辑 李莉)

中国农业科学 2015,48(17):3354-3371 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.004
收稿日期:2015-05-04;接受日期:2015-06-15 基金项目:国家“973”计划(2014CB138206)、国家“863”计划(2012AA101104)、国家自然科学基金(31271736、31371638)、科技部国际合作
项目(2011DFA31140、2012DFA32290)、科技部科技伙伴计划、比尔盖茨基金会资助 联系方式:徐云碧,E-mail:[email protected]
作物科学中的环境型鉴定(Envirotyping)及其应用
徐云碧
(中国农业科学院作物科学研究所/CIMMYT-中国,北京 100081)
摘要:全球气候变化正在对地球的环境产生日益重要的影响,而作物生产取决于作物基因型和环境之间的相
互作用。利用现代生物学技术可以在分子水平上精细解析作物的基因型及其各个遗传组分对于表现型的贡献。然
而对于作物具有重大影响的环境因子,目前,只能通过作物在不同环境下的表现型来推测其综合作用,或对整个
试验区的个别环境因子进行对比分析,因而无法对各类环境因子进行深入剖析。笔者首次在国际上提出了环境型
鉴定概念,并创造了一个英文新词 etyping 来表示。在本文中,环境型鉴定用 envirotyping 来代替。环境型
(envirotype)用来描述包括所有影响作物不同生长发育阶段的内部和外部环境因子及其各种组合,外部环境因
子主要包括水、肥、气、热、光、土壤、耕作制度和伴生生物等;而环境型鉴定用来表述对所有环境因子的解析
和测定。环境型信息可以通过多种方式采集。作物多年多点区域试验积累了大量相关试验点的环境数据;地理信
息系统(geographic information system,GIS)和土壤信息系统积累了大量气候、天气、土壤的数据;小型气
象站可以监测小范围的天气、降雨、温度、气流等气象因子。众多环境检测仪器的使用,可以大规模采集与植物
冠层、植物周边甚至单个试验小区和单个测试材料有关的土壤、光照、温度、水分、病虫害、伴生生物等外界环
境因子。环境型信息将日益广泛应用于环境及其特征性鉴定、作物基因型-环境型互作、表现型预测、病虫害流行
预测、近等环境型(neariso-envirotype)确定、作物对特定环境的反应研究、农艺组学(agronomic genomics)、
精准高效农业等。展望未来,环境型鉴定需要将研究对象聚焦在单个材料的水平,实现单个材料的相关环境因子
不同阶段的动态鉴定;需要开发和建立与基因型、表现型相结合的综合信息系统以及相应的决策支撑系统;环境
型信息将最终有助于建立基于基因型-表现型-环境型的三维作物生产和研发系统,从而使未来作物育种中的选择
建立在此三维空间概念的基础之上,并推动高产高效作物生产体系的建立。
关键词:作物生产;环境信息;环境型;环境型鉴定;基因型-环境型互作;近等环境型;农艺组学;表现
型预测
Envirotyping and Its Applications in Crop Science
XU Yunbi
(Institute of Crop Science, Chinese Academy of Agricultural Sciences/CIMMYT-China, Beijing 100081)
Abstract: Global climate changes have increasing impacts on worldwide environments. Crop productivity is largely determined by interaction between the genotype a crop has and the environment surrounding the crop plants. With modern biotechnologies, genotypic contribution to a phenotype can be dissected at molecular level into individual genetic components. However, the environmental factors that have significant impacts on crops have not been dissected individually, and thus their contribution to phenotype can be only inferred by their integrative effect under different types of environments, or described for the whole experimental plot by comparing pairwise major environmental factors. The author proposed a concept of environmental assay for the first time by coining a word “etyping”, which represents “envirotyping”, a more suitable word used in this article. The term “envirotype”
jia
文本框
返回目录

17 期 徐云碧:作物科学中的环境型鉴定(Envirotyping)及其应用 3355
is used to describe all internal and external environmental factors and their combinations that affect plants across growth and developmental stages. The external environmental factors include moisture, fertilizers, air, temperature, light, soil properties, cropping system and companion organisms. Envirotyping refers to dissecting and measuring all these environmental factors. Environmental information can be collected through various approaches, including multi-environmental trials with environmental data accumulated related to trial locations; geographic and soil information systems containing environmental data for climate, weather, and soil; and small weather stations that collect factors related to weather, precipitation, temperature and air. Using remote sensing and other instruments, many external environmental factors can be measured for plant canopy, plant surroundings, and even for single plots or individual plants. Environmental information will be increasingly used for environment characterization, genotype-by-environment interaction analysis, phenotype prediction, disease epidemic prediction, near iso-environment construction, understanding of the response of plants to specific environmental factors, agronomic genomics, and precision farming. In the future, envirotyping needs to be improved to zoom into specific plots and individual plants across growth and developmental stages, along with the development of integrative information system and decision support tools to bring genotypic, phenotypic and envirotypic information together. Envirotypic information will finally contribute as a third dimension to the crop research and development system involving genotype-phenotype-envirotype complex. Such efforts will help establish a high-efficient crop breeding and production system based on the concept of the three-dimensional profile.
Key words: crop production; environmental information; envirotype; envirotyping (etyping); genotype-by-environment interaction; near iso-envirotype; agronomic genomics; phenotype prediction
0 引言
在过去 50 年中,世界各国在农作物改良方面作出
了艰辛的努力,显著改进了农作物的产量潜力、品质、
耐逆性,使重要作物的产量在过去 50 年保持了直线增
长。为了养活发展中国家快速增加的人口、应对全球
气候变化及脆弱自然资源所带来的挑战,需要在未来
30—40 年内实现作物产量至少翻番的目标[1]。同时,
在最佳试验条件下可以达到的理论产量与农民田间条
件下所能获得的实际产量之间还存在巨大差异。以水
稻为例,亚洲国家在正常农艺措施管理条件下所获得
的平均产量大约为 5 t·hm-2,还不到育种家在试验站所
获得的产量的 30%[2]。因此,在不断提高作物产量潜
力的同时最大限度地实现其潜力,是满足未来世界作
物生产需求的两项重要技术途径。 作物生产需要理解和操控许多因素,其中包括影
响植物生长、发育的所有环境因素以及这些环境因素
对作物代谢、基因表达等的影响。作物表现型是基因
型和各种环境因子相互作用的结果。表现型
(phenotype)是指一个或一群生物由遗传组成和环境
因子的相互作用所决定的全部可观察特性,包括形态、
生理、生化等性状;利用形态学、生理学、生物化学
等方法对所有这些特性进行区分和测量称为表现型鉴
定(phenotyping)。与表现型的概念相对应,基因型
(genotype)可以定义为一个细胞、生物或个体的全
部遗传构成。基因型的经典概念通常是针对某个特定
性状或基因座位,而其现代概念已扩展到包括某个细
胞、生物或个体的全部 DNA 序列,并以另外某个个
体或参考序列为参照进行比较。基因型鉴定
(genotyping)就是利用生物学方法比较某个体与另一
个体或参考基因组 DNA 的序列以确定该个体遗传组
成(基因型)差异的过程。检测 DNA 差异方法包括
分子杂交法(如 RFLP)、PCR 扩增法(如 SSR)、
芯片分析法(如 SNP)、DNA 测序等。 过去几年,笔者倡导和提出了环境型(envirotype)
和环境型鉴定(envirotyping)的新概念。在第六届国
际作物科学大会上的专题报告中,作者首次正式提出
了环境型鉴定这一新概念,并将其英文名称简化为
“etyping”[3-4]。作者所发展的全基因组策略(whole genome strategies)[4]就是通过整合所有基因型、表现
型和环境型的信息,以实现植物分子标记辅助育种的
高效设计和顺利实施。本文将综合讨论环境型鉴定的
基本概念、环境型信息的分类和采集以及环境型信息
在作物科学研究中的应用。
1 环境型鉴定的基本概念
作物遗传改良的全基因组策略可以定义为在全基
因组水平上对分子植物育种所需的各种有效工具和方
法进行全面集成[4]。全基因组策略的建立涉及到下列
信息的完美整合:所有种质资源(M)的全基因组序
列或高密度分子标记、基因标记和功能等位基因标记
(G)、在多个环境条件下测量的不同目标性状的精
准表现型(P)以及影响基因、基因型和植物表现型
的所有相关环境因素(E)(图 1)。与表现型和基

3356 中 国 农 业 科 学 48 卷
图1 环境型鉴定作为影响整合作物育种的4个关键要素之一
Fig. 1 Envirotyping as one of the four key components for
integrative plant breeding
因型的概念相对应,环境型可以定义为影响某一个
体或一群生物不同生长发育时期的所有环境因子,
而利用各种手段和仪器对所有这些环境因子进行动
态测定和描述的过程则称为环境型鉴定。传统的环
境信息采集是假定同一地点或区域的所有基因型都
处于相同的环境之下且具有相同的环境因子。环境
型与基因型和表现型相对应,环境型鉴定就是要确
定每个基因型所处周边及其内部的所有环境因子,
这些环境因子因每个基因型(品种)所处的环境不
同而异。每个基因型或品种所处的周边环境和内部
环境因子是特定的、互不相同的,从而构成每个基
因型或品种的特定环境型。 环境型鉴定是全基因组分子育种策略的一个重要
组成部分。基于全基因组策略的作物遗传改良程序,
始于自然和人工群体,通过基因型鉴定、表现型鉴定、
环境型鉴定,产生大量的育种相关信息,在一系列决
策支持系统的帮助下,实现对最佳重组体的选择、固
定和测试,最后育成新的作物品种。 作物的表现型是基因型和特定环境因子相互作用
的结果。在经典的数量遗传学研究中,环境因子被作
为影响作物性状遗传、不能被明确定义或分解的整体
环境因素 E 来看待,实际上是作为一个未知的“黑
箱”来处理,因此,无法从本质上了解不同环境因子
对作物生长、发育、遗传、代谢等的影响,更无法通
过设计和调控环境因子,进而通过改变和调控作物来
实现作物的高效生产。 综合理解作物生长发育所处的环境极为重要。首
先,基因表达取决于作物所处的环境条件;其次,QTL定位和基因挖掘取决于精准的表现型鉴定;第三,表
现型鉴定,特别是生物逆境和非生物逆境的评价具有
环境依赖性。因此,精准表现型鉴定必须在精确管控
环境下进行,且与精准的环境型鉴定相结合。同时,
由于全球气候变暖,精确的环境因子测定对于作物生
产的许多环节也变得日益重要。
2 环境型信息的分类
根据对作物生长发育的影响,可以将植物外部的
环境信息区分为气候、天气、土壤和水分、耕作制度
和伴生生物几大类。它们之间存在相互影响和制约。
其中气候和天气又涉及水分、温度、光照、气流等诸
多要素,而土壤本身就是一个影响作物生长发育的复
杂生态系统。极端的环境因子本身成为影响作物生长
发育的非生物逆境胁迫因子。 2.1 植物的内部环境和外部环境
植物的生长发育受到内部和外部环境因子的影
响。内部的环境因子主要是细胞内和细胞间的微环
境,包括水分、矿质营养等影响植物代谢和基因表达
的因子。植物体内微环境的测定需要借助高度精密的
仪器设备,同时需要严格控制植物所处的外部环境条
件。因此,大规模、标准化测定植物体内微环境,目
前还存在许多困难。因此,本文主要讨论针对植物所
处的外部环境。 植物的生长发育主要受外部环境的制约,同时
外部环境对植物体内微环境具有重大影响。外部环
境涉及许多自然和人为的因素,环境型鉴定需要尽
可能地考虑所有环境因素(图 2)。基本因素包括
显著影响植物发育的要素[3],比如土壤、空气、光
照、温度、水分、耕作制度以及伴生生物。根据对
植物发育和生产力的影响,可以进行环境或农田的
分类。例如,土壤类型和降雨量是可以用于分类的
2 个重要指标,据此可以进行大环境的划分。养料
和水分等的田间梯度变化对于非生物胁迫的度量
具有关键作用,它们也会对小区之间的环境变异产
生影响。 2.2 气候因子
气候(climate)是地球上某一地区多年时段大气
的一般状态,表述气候的基本依据是气象要素(温度、
降水、风等)的各种统计量(均值、极值、概率等)。
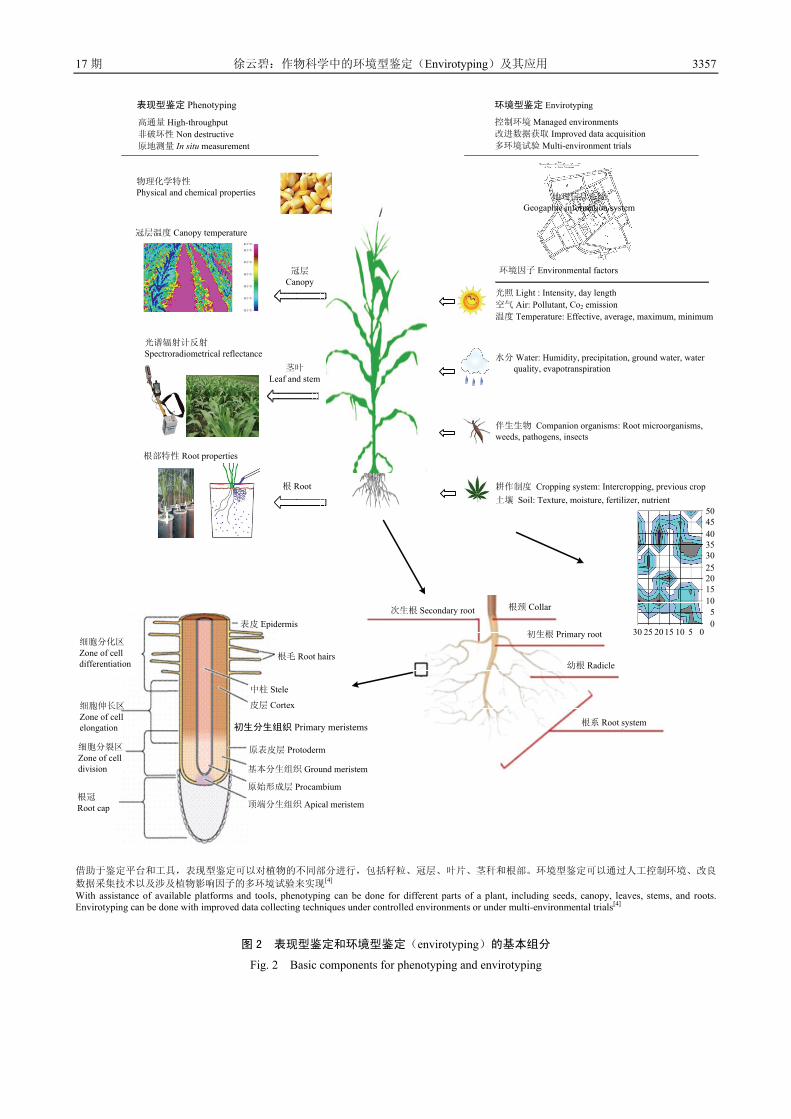
17 期 徐云碧:作物科学中的环境型鉴定(Envirotyping)及其应用 3357
高通量 High-throughput非破坏性 Non destructive 原地测量 In situ measurement
冠层温度 Canopy temperature
环境因子 Environmental factors
茎叶Leaf and stem
根 Root
光谱辐射计反射Spectroradiometrical reflectance
冠层Canopy
根部特性 Root properties
物理化学特性Physical and chemical properties
控制环境 Managed environments改进数据获取 Improved data acquisition多环境试验 Multi-environment trials
地理信息系统Geogaphic information system
光照 Light : Intensity, day length空气 Air: Pollutant, Co2 emission温度 Temperature: Effective, average, maximum, minimum
水分 Water: Humidity, precipitation, ground water, water quality, evapotranspiration
伴生生物 Companion organisms: Root microorganisms, weeds, pathogens, insects
耕作制度 Cropping system: Intercropping, previous crop土壤 Soil: Texture, moisture, fertilizer, nutrient
原表皮层 Protoderm
原始形成层 Procambium
顶端分生组织 Apical meristem
表皮 Epidermis
根毛 Root hairs
中柱 Stele
皮层 Cortex
根冠Root cap
细胞分化区Zone of cell differentiation
细胞伸长区Zone of cell elongation
细胞分裂区Zone of cell division
根颈 Collar
初生根 Primary root
次生根 Secondary root
幼根 Radicle
根系 Root system
50454035302520151050
051015202530
初生分生组织 Primary meristems
基本分生组织 Ground meristem
表现型鉴定 Phenotyping 环境型鉴定 Envirotyping
借助于鉴定平台和工具,表现型鉴定可以对植物的不同部分进行,包括籽粒、冠层、叶片、茎秆和根部。环境型鉴定可以通过人工控制环境、改良
数据采集技术以及涉及植物影响因子的多环境试验来实现[4]
With assistance of available platforms and tools, phenotyping can be done for different parts of a plant, including seeds, canopy, leaves, stems, and roots. Envirotyping can be done with improved data collecting techniques under controlled environments or under multi-environmental trials[4]
图 2 表现型鉴定和环境型鉴定(envirotyping)的基本组分
Fig. 2 Basic components for phenotyping and envirotyping

3358 中 国 农 业 科 学 48 卷
影响气候的主要因素包括纬度位置、海陆位置、地形
因素、洋流因素。地理因素对气候形成的影响归根到
底可以归结为辐射因素。由于太阳辐射在地球表面的
分布差异导致温度大致按纬度分布的特征,同时受
海洋、陆地、山脉、森林等影响产生明显的地域性,
从而形成热带雨林气候、地中海型气候、极地气候、
高原气候等大气候,森林气候、城市气候、山地气
候以及湖泊气候等中气候,贴地气层和小范围特殊
地形下的小气候。影响气候的因子包含气温、雨量、
气压和风。 2.3 天气因子
天气(weather)是在较短时间内特定地区的大气
状况。天气现象是某瞬时内大气中各种气象要素(气
温、气压、湿度、风、云、雾、雨、闪、雪、霜、雷、
雹、霾等)空间分布的综合表现。光照是最重要的天
气要素,光照强度、光照时间和光能利用率直接决定
作物产量的形成。 作为一种无时无处不在起作用的重要生态因子,
气温的主要指标包括月均温、年均温、日温差、月温
差、年温差等。地球表面的温度在空间上随纬度、海
拔高度、生态系统的垂直高度和各种小生境而变化,
在时间上随四季和昼夜变化。 大气层的空气对流,形成风、云、雨、雪、雾等
各种天气现象。同时,大气为作物提供了 CO2、O2、
H2O 及 N 素的来源。大气污染来源于各种废气、粉尘
排放。大气中的有害气体包括 SO2、N2O、NO、NO2、
HF、Cl2、H2S、NH3、甲烷、甲醛、乙烯、丙烯、丁
烯、苯并芘等;而大气中粉尘排放主要是颗粒状污染
物,包括粉尘、烟尘、烟气等,均含有毒重金属元素。
大气中 SO2、NO、NO2、Cl2等溶于水形成含 H2SO4、
HNO3和 HCl 的酸雨。 2.4 土壤和水分
岩石经过物理、化学、生物的侵蚀和风化作用,
以及地貌、气候等诸多因素的长期作用下形成能生长
植物和微生物的疏松表层环境——土壤。土壤环境由
矿物质、动植物残体腐烂分解产生的有机物质以及水
分、空气等固、液、气 3 相组成,由土壤物理性质(土
壤容重、土壤颗粒组成和土壤质地)、土壤化学性质
(酸碱度、有机质、土壤阳离子交换能力、交换性盐
基、交换性氢和铝、碳酸盐或石灰含量、硫酸盐含量、
电导率等)、土壤养分(磷素养分潜力、钾素潜力;
速效磷和速效钾含量;有机质含量、全磷和全钾含量;
土壤微量元素)等决定。
土壤含水量直接影响作物生长、农田小气候以及
土壤的机械性能。土壤水分可以用多种指标来描述,
包括土壤剖面不同深度(例如每 10 cm 间隔)的水分
含量、土壤有效水含量、土壤相位、土壤含水量特征
等。土壤水分受降雨量、灌溉、植被(作物生长发育
状况)和土壤蒸腾作用等的影响。而降雨量涉及最多
雨少雨月及其降水量、年降水量、年际降水变率、蒸
散量、空气湿度等。 2.5 耕作制度
农作物种植制度以及与之相适应的养地制度,称
为耕作制度,影响作物生长发育的环境。种植制度包
括作物组成配置、熟制、作物布局、复种休闲和种植
方式(单作、间作、混作、套作或移栽、轮作和连作)。
养地制度是与种植制度相适应,以提高土壤生产力为
中心的一系列技术措施,包括农田基本建设、土壤培
肥、水肥管理、土壤耕作以及农田保护等。 2.6 伴生生物
作物一生从播种到收获与许多其他生物为伴。有
些伴生生物对作物的生长发育是有益的,而大部分则
与作物存在竞争的关系。有益的伴生生物包括部分根
际微生物和协助作物授粉的虫媒昆虫等。农田杂草与
作物竞争所有与生长发育有关的环境因子。作为许多
微生物的寄主,作物在其一生中可能遭受许多病原细
菌、真菌和病毒等的危害。害虫则直接以作物的组织
和器官作为取食对象,对作物造成最为直接的伤害。
另外,有些伴生生物则是作物生长发育所必需的,例
如帮助作物进行授粉的昆虫。 2.7 生物胁迫和非生物胁迫
外部环境因子的变化,会对作物的生长发育造成
生物胁迫和非生物胁迫。生物因素如真菌、细菌、病
毒、线虫、昆虫等对作物的危害而产生生物胁迫。非
生物胁迫因子包括温度、水分、光周期等关键因素的
极度变化所造成的胁迫。 在有害生物的侵染或不良环境条件的影响下,植
物的正常新陈代谢受到干扰,导致生理机能和组织结
构等发生一系列变化,在外部形态上呈现枯萎、腐烂、
斑点、霉粉、花叶等现象。真菌、细菌、病毒等生物
因素侵入植物体引起寄生性病害;而旱、涝、严寒、
养分失调等非生物因素会影响或损坏生理机能而引起
生理性病害。 非生物因子的变异超出植物正常生长发育所需的
环境因子范围时导致非生物胁迫。冷害引起作物生育
期延迟,或使其生殖器官的生理机能受到损害,最终

17 期 徐云碧:作物科学中的环境型鉴定(Envirotyping)及其应用 3359
导致作物减产。高温会导致植物叶绿素失活、光合作
用受阻、消耗增强、细胞膜半透性丧失、植物器官组
织受损;高温还能使光合同化物输送到穗部和籽粒的
能力下降,酶的活性降低,致使灌浆期缩短而导致产
量下降。重大干旱将导致土壤水分严重不足,植物萎
蔫、叶片干枯,果实脱落,对作物生产和生态环境造
成较严重影响。其他非生物逆境胁迫还包括土壤盐渍
化、涝渍、大风、空气污染、CO2浓度升高等。 同一作物在同一地点不同的发育阶段可能会遭遇
多种逆境胁迫的影响。解决逆境胁迫问题需要深入了
解各种胁迫因子及其对作物生长发育的综合作用和相
互影响。
3 环境型数据的采集
作物育种和生产的许多环节已经产生和积累了大
量的环境型数据。可供利用的环境型数据主要来自多
年多点区域试验、地理信息系统采集的气候和气象数
据、土壤信息系统采集的土壤信息、利用各种土壤水
分测定仪获得的土壤水分数据、利用遥感技术获得的
作物冠层数据等。此外,本文建议将伴生生物有关的
信息也纳入环境型数据之列进行系统采集。 3.1 利用多年多点区域试验采集试验点环境信息
多年多点的多环境试验(multi-environment trial,MET)是环境数据采集的传统方式之一。MET 主要用
于品种区域试验,以确定品种的适应范围和需要采取
的相应农艺措施。在 MET 实施过程中,一般都要针
对 MET 所在的实验站或试验点进行有计划、标准化
的环境数据采集。所采集的环境数据包括主要的天气
数据、土壤和农艺措施等。在发达国家和一些国际跨
国公司,现已累计了长达数十年的 MET 及其相应的
环境数据,成为环境型信息的重要部分。国际农业研
究磋商小组(CGIAR)所属国际研究中心(包括国际
玉米小麦改良中心和国际水稻研究所)和跨国公司组
织的国际育种计划,已开展了多年的多点试验,其代
表性目标环境和多点试验数据正在日益增加。收集、
保存、挖掘这些历史性环境信息是环境型鉴定的一个
重要内容。 3.2 利用地理信息系统采集气候和气象数据
地理信息系统(geographic information system,
GIS)的建立,是为了获取、储存、操作、分析、管
理和展示所有的地理数据。GIS 将制图、统计分析和
数据库技术等融合在一起,可以用于整合、储存、编
辑、分析、分享和展示地理信息。地球空间-时间中的
任何一点及其范围都可以用相关的日期、时间以及经
度-纬度-海拔三维坐标来确定。GIS 可以将图谱信息、
地球观测卫星所获得的实时数据与气候变化模型等不
同来源的数据整合起来,从而有助于了解气候变化对
复杂自然系统的影响。 为了取得气象资料,世界各国建立了各类气象观
测站,如地面站、探空站、测风站、火箭站、辐射站、
农业气象站和自动气象站等。中国已建成类型齐全、
分布广泛的台站网,目前共有地面气象观测站点 2 500个左右,太阳辐射观测站 98 个,高空气象探测站 120个,农业气象试验站 70 个,农业气象基本站 672 个[5]。
国家气象中心每天接收来自国内外主要台站的观测资
料,这些资料日积月累,随时间的推移而成为气候资
料。国家气象中心收集和储存国内一部分台站的观测
记录报表和数字化资料,这些资料或报表成为气候资
料的重要部分。此外,通过其他渠道还可以收集到水
文、地学等气候资料。 3.3 利用土壤信息系统采集土壤信息
中国科学院南京土壤研究所建立了一个较为系统
的中国土壤信息系统( soil information system of China)。中国土壤数据包括数字化的土壤空间数据、
土壤属性数据和土壤参比数据。这些数据包括 3 个来
源。一是来自南京土壤研究所长期以来所积累的各种
土壤数据,包括各种土壤图、土壤属性图和区域土壤
调查报告;二是来自中国第二次土壤普查数据[6];三
是来源于中国土壤信息查询系统和中国土壤参比查询
系统[7]。 数字化土壤空间数据基于原始纸质土壤图件通
过数字化编制而成(图 3)。根据承载土壤信息类
型的不同,这些图件可分为土壤图、土壤基本属性
图(黏粒矿物、侵蚀土壤分布和分区、土壤侵蚀点、
土壤利用现状和分区、成土母质类型、酸碱度和质
地)、土壤养分图(有机质含量、磷素养分潜力、
钾素潜力;速效磷和速效钾含量;有机质含量、全
磷和全钾含量)、土壤微量元素图(有效铜、锌、
锰、硼等的含量)[8]。 WebGIS 建立的中国土壤信息查询系统提供了
全国土壤的类型分布、基本属性、养分元素含量和
微量元素含量在多个尺度下土壤信息的查询。按照
参比出发点的不同,该系统可以分别在全国尺度、
区域尺度(省级)和单个土体尺度下,通过友好的
交互性界面便捷地查询到尽可能精确的土壤分类参
比数据。

3360 中 国 农 业 科 学 48 卷
A B
0 500 1000 2000km
0900 900km
土纲类型 Soil type
铁铝土 Pedalfer淋溶土 Alfisol半淋溶土 Semi-Luvisol钙层土 Pedocal干旱土 Xerosol漠土 Desert soil
初育土 Primitive soil
半水成土 Semi-hydromorphic soil水成土 Hydromorphic soil盐碱土 Saline soil人为土 Anthrosol高山土 Alpine soil
土壤有机碳密度Soil organic carbon density
SOCD (kg·m-2)
<11-55-1010-2020-4040-6060-100>100无数据 No data
图 3 基于中国土壤发生分类的中国 1:100 万数字化土壤图(A)[8]和中国土壤有机碳密度(0—100 cm)分布(B)[9]
Fig. 3 A digital soil map for China soil classification (1:100M) (A) [8] and the distribution of soil organic carbon density in China
(0-100 cm)[9]
3.4 土壤质地和水分测定
土壤表观电导率与黏土含量、土壤水分、离子浓
度等密切相关。Geonics 公司生产的 EM 系列大地电导
率仪是确定地下电导率分布的一种地球物理手段,可
以快速探测不同土壤层的含盐度、水分和磁化率。因
此,可用于测定土壤质地和水分,包括电导率、土壤
强度、盐度、团聚体大小等,由此描绘出有关土壤质
地、结构、水分等的图谱。 主要土壤水分测量技术,包括直接测量土壤的重
量、含水量或容积含水量的称重法和中子仪等标准方
法,测量土壤传导性的时域反射仪( time domain reflectometry,TDR)法,测量土壤基质势的张力计法、
电阻块法、干湿计法等,以及非接触式的间接测定方
法,如远红外遥测法、地面热辐射测量法、声学方法
等。 作为测定土壤水分的主流方法,TDR 是通过测
量土壤表观介电常数来获得土壤容积含水量[10],已
成为研究土壤水分的基本仪器设备和主流方法。土
壤表观介电常数与容积含水量存在非线性关系,而
土壤介电特性与下列因素存在函数关系:电磁频率、
温度和盐度,土壤容积含水量,束缚水与土壤总容
积含水量之比,土壤容重,土壤颗粒形状及其所包
含水的形态。在一定的电磁波频率范围内(50 M—
10 GHz),矿物质、空气和水的介电特性为常数,
因此土体的介电常数主要依赖于土壤容积含水量
(极微弱地依赖于土壤类型、紧实度、束缚水等),
因此,可以建立土壤容积含水量与土壤介电常数的
经验方程。通过测量高频电磁脉冲在土壤中的传播
速度,TDR 可以获得土壤的介电常数,进而计算出
土壤含水量。轻巧、便携式 TDR 可作野外测量,并
与计算机相连,实现对土壤样品快速、连续、准确
地测量,平均分辨率达 0.02—0.005 cm3·cm-3,且含
水量测量范围广(0—100%)。 利用土壤剖面水分探头,可以精确测定土壤剖
面多个深度的土壤水分含量。作为 TDR 类似仪器,
PR2 可以测定不同类型、不同营养成分和酸碱度的
土壤。PR2/6 型仪器可在 6 种不同土壤深度(10、20、30、40、60 和 100 cm)进行水分测定。便携式

17 期 徐云碧:作物科学中的环境型鉴定(Envirotyping)及其应用 3361
Diviner 2000 型探头可在每隔 10 cm 的土壤深度快
速测定水分含量,其手持式数据记录显示元件可以
帮助用户在多达 99 个测试点作出现场管理决策。该
探头有 0.7、1.0 和 1.6 m 3 种不同的长度选择,其
1.6 m 的探头可在几秒钟内完成多达 16 种不同深度
的水分测定。这类仪器已广泛用于监测干旱试验中
水分的动态变化。 3.5 作物冠层特性的测定
利用遥感技术,包括反射的光谱辐射测量、数字
图像、热图象、近红外反射光谱等,可以对冠层温度、
湿度、光照、气流、生物量、覆盖度等作物的冠层特
性进行测定,以描述或推断作物的生长发育状况。许
多冠层特性的测定采用了携带照相设备的空中平台
(unmanned aerial platform),如小型飞机等。为高通
量和精准表现型鉴定发展的自动图像平台和计算机视
觉辅助分析[11],可以用于作物冠层特性的测定。通过
多视角二维图像的堆积构建植物三维图像的计算方
法[12]为作物冠层特性的测定提供了新方法。同时,三
维结构也可以利用激光扫描技术[13-14]和通过各类深度
/时间飞行传感器[15]直接进行测定。 3.6 伴生生物的测定
作物保护研究机构为作物病虫害等的测定发展了
一系列方法,以针对不同的作物,同一作物的不同病
原菌、病毒、害虫、杂草。这些因素的综合测定需要
不同领域的科学家密切合作。作物根际微生物的测定,
需要提取土壤混合样本并对所含各种微生物进行综合
分析和评价。对土壤混合样本进行微生物的混合
DNA/RNA 测序,可以精确了解和测定土壤中微生物
的种类、数量及其相互关系。叶片和冠层混合取样分
析,也可以测定附着在植物表面的各种微生物,包括
病原菌等的种类、数量、分布、发育阶段、共生关系
等。
4 环境型信息的应用
环境型信息在作物科学研究具有广泛的应用。主
要可用于分解 G×E 互作,确定环境特征,评价环境
依赖性试验,控制试验误差,确定作物对特定环境因
子的反应及其相关基因,预测流行性病虫害,在基因
组水平上研究农艺措施对作物生长发育的影响,以及
与基因型信息相结合进行表现型的预测。将环境因子
进行精细的剖分,可以研究作物对不同环境因子的反
应。此外,环境型信息还可用于农艺组学的研究并服
务于精准农业。
4.1 G×E 互作解析与表现型的精准预测
表现型(P)可以分解为基因型效应(G)、环境
型效应(E)、G×E 互作效应以及试验误差: P = G + E + GE + error 其中,G 为基因型信息;E 为环境型信息,包括
影响植物生长、发育和生产力的环境因子; GE 为G×E 互作及其对植物生长、发育和生产力的贡献。
迄今,环境型信息的重要应用之一就是在不解析
环境因子的情况下,将其作为一个整体,以研究基因
型在不同环境下的表现以及 G×E 互作。相关研究大
致可以分为以下几类: (1)利用 QTL 作图研究 G×E 互作。在多个环境
下进行的 QTL 作图为了解各类G×E 互作机制的相对
重要性提供了试验数据。不同环境下 QTL 加性效应存
在 4 种不同类型的作用模式 [16] :拮抗上位性
(antagonistic epistasis;加性效应的符号或方向发生了
改变)、条件中性(加性效应仅在某些特定环境条件
下表现因而具有环境依赖性)、差异敏感性(加性效
应的大小因环境而发生变化)、无 G×E 互作(不同
环境下加性效应保持不变)。最早有关 QTL×E 的评
价之一就是对3个不同地点所检测到的番茄QTL进行
简单的比较[17]。这类研究不涉及有关环境因子的任何
具体信息,而只是假定不同的地点具有不同的环境效
应,把不同环境下检测到不同的 QTL 归结于 QTL×E互作的结果 。
在相对可比的环境下进行的 QTL 作图,是比较
接近真实的 G×E 互作研究。这类 G×E 互作研究大
多与非生物逆境胁迫有关。Des Marais 等[16]从 700 多
个研究报告中,筛选出与非生物逆境胁迫有关、且
QTL 信息比较完整的 37 项研究进行综合分析,发现
大约 60%的 QTL 存在拮抗上位性和环境特异性效应
导致的 G×E 互作。以不同环境下标准化加性效应的
绝对差值为指标所定义的 QTL 效应可朔性,受到这
两类 G×E 互作类型的强烈影响。 (2)利用 eQTL 作图研究 G×E 互作。有两大类
基于基因表达分析的 G×E 互作分析方法。一种是表
达 QTL(eQTL)分析,即某一基因在 2 个不同的基
因型中表现出不同的条件特异性表达模式。另一种是
全基因组-全环境关联分析,即在许多环境下测定一
个物种许多个体所有基因的表达水平。这两类分析均
把生物所处的环境作为一个整体来看待,而没有对构
成复杂环境的个别环境因子进行解析。尽管在一些植
物作图中对群体进行了非生物条件控制和性状表达

3362 中 国 农 业 科 学 48 卷
的定量测定,但真正典型的 eQTL 作图很少。
Hammond 等[18]报道了一个比较正式的 eQTL 作图研
究,其利用 2 种不同磷水平下采集的白菜叶片组织,
鉴定出 3 226个磷反应的转录本和几个富集磷代谢相
关基因的重要热点区域,但不幸的是作者未能进行
G×E 的正式检验。在酵母和果蝇等模式生物中日益
增加的证据表明,G×E 互作或许可以解释大部分的
表现型变异;这种互作是上游调节因子而不是启动子
局部变化的结果;不同的基因表现出不同程度的
G×E 互作,而且启动子结构、表达水平、调节复杂
性等与环境造成的基因差异性调节有关[19]。 (3)通过基因克隆研究 G×E 互作。为了解 G×E
互作的起源、扩散和进化过程,需要鉴定控制影响
G×E 表现型的特定基因和功能等位基因的突变体。
要确定下列因素中什么是控制表现型所需的G×E互
作的常见驱动因子:基因类型(编码环境感应、生物
合成酶、或调节蛋白类型)、基因特性(具有物种内
同源性或复杂顺式调节控制特性)、突变类型(编码
和非编码突变、转座子诱导的突变等)、分子机制(条
件依赖性上位性、基因表达、酶活性)[16]。鉴定 G×E互作的基因还有助于对环境反应相关的平行和趋同
进化进行跨世系和检验的比较分析。主要依赖于光周
期、温度、生理信号等环境因子的驱动,植物已进化
出控制复杂表现型的调控系统[20]。Des Marais 等[16]
表列了一些业已被克隆的 G×E 互作基因,并以开花
期、土壤和水分可用性的基因克隆为例讨论了与
G×E 互作有关的分子生物学,认为导致 G×E 互作
的基因和分子突变是多样性的。在分子水平,G×E互作涉及一些自然突变及其机制,包括受体蛋白的非
同义改变(PhyB 和 Cry2)、翻译抑制子的功能缺失
突变(FRI 和 FLG)、合成酶的剪切变异(P5CS1)、
转录因子的基因重复(Sub1A)。 随着对基因型信息(G)日益深入的了解,可以
精确描述和确定基因、等位基因、单倍型及其对于植
物表现型的综合贡献。而环境型鉴定的结果,可以将
长期以来作为一个整体看待的环境效应(E)进一步
分解为光照、空气、温度、水分、逆境、肥力、湿度
和伴生生物等贡献的各个组分。因此,在精确测定 G和 E 的基础上,结合精准表现型鉴定,可以对 G×E互作进行精细的评价。根据G和E所建立的理论模型,
也可以对 G×E 互作进行预测。另一方面,如果已知
某一植株的大量基因型信息及其所处的各种环境因
子,根据试验或预测的 G×E 互作,可能对该植株的
表现型进行前所未有的精准预测。在一些情况下,根
据少数环境因子的综合分析,就有可能对植物群体的
重要性状进行比较可靠的预测。例如,对非洲大陆长
达 10 年的降雨量和玉米产量的数据分析表明,两者之
间存在高度正相关(CIMMYT,内部交流信息),因
此可以根据降雨量对非洲大陆的玉米产量进行相当准
确的预测。 许多环境型信息本来是可以用来帮助了解植物
对环境因素的响应,但迄今尚未被充分挖掘。通过大
规模环境型鉴定,诸如 GIS 之类的高级信息系统,
改进了数据的采集,已极大地丰富了环境信息的内
涵。可以按照多种方式对环境型鉴定的信息进行挖掘
并用于植物育种,包括:精确测定影响作物特定发育
过程或阶段的环境因素;为特定环境适应性鉴定确定
目标环境;分析单个或多个环境因素对田间试验或试
验区组内表现型变异的贡献;将 G×E 剖分为特定的
环境因素;通过育种改进植物对特定环境、环境因素
及其各种环境综合因子的反应。在引入 G×E 的概念
后,根据多年多点的区域试验数据,可以将育种试验
中的目标环境主要区分为年份间存在可重复的交叉性
G×E 互作、年份间存在不可重复的交叉性 G×E 互
作、以及年份间不存在交叉性互作等 3 种类型。最终,
可以建立包括基因型、表现型和环境型的三维育种图
像及其相应的作物育种计划(图 4),并对基因型进
行设计,以获得对特定环境条件具有最佳适应性的最
优表现型。 野生植物遭遇多种、波动的环境因素的影响,它
们处于复杂的自然环境中,依赖其遗传调控网络发挥
功能并不断进化。全表观基因组的多样性为植物适应
各种环境提供了不需要改变 DNA 序列的快速途径。
未来还需要进行更大规模的全表观遗传组学分析,包
括收集上千份样品。目标是通过分解空间和时间对家
族或自交系的影响,最大限度地减少遗传变异,鉴定
出常见的表观等位基因(epiallele)。 4.2 环境特征分析
环境型鉴定及其所获得的环境信息可以用来揭示
试验环境的一系列重要特征特性。包括: (1)田间特性和试验点内变异的确定:消除新试
验点的潜在变量,了解田间试验点的历史。如前所述,
EM 大地电导仪可以提供关于土壤质地和水分的田间
梯度和均匀性的评价。图像采集技术与无线传感网络、
地理信息系统相结合,可以对土壤空间变异等进行更
为精确的描绘和监测[21]。空中高通量表型鉴定平台与

17 期 徐云碧:作物科学中的环境型鉴定(Envirotyping)及其应用 3363
G
E
P
基因型 Genotype
表现型
Phen
otyp
e
点 Point 线 Line 平面 Plane 空间 Space
单一表型Single-Phenotype
多个表型Multi-Phenotype
基因型辅助下多个表型
Multi-phenotype assisted by genotype基因型辅助下跨环境的多个表型Multi-phenotype across environments assisted by genotype
表现型
Phen
otyp
e
表现型
Phen
otyp
e
图 4 植物育种的三维图像概念[4]
Fig. 4 A three-dimensional profile of plant breeding[4]
归一化植被指数(normalized difference vegetation index,NDVI)相结合,可以快速、非破坏性地测定
作物的生物量,利用单一基因型作物的田间表现(生
物量)可以用于评价田间变异性[22]。 (2)土壤类型和土壤特性的确定:土壤地图集(soil
atlas,图 5-A)可以提供大量有关土壤的信息,包括
土壤板结、土壤流失、有机质伤失、生物多样性下降、
污染、土壤压实、水文地质风险、土壤盐碱化等。 (3)土壤肥力或养分的评价:利用 GIS 进行精准
取样、测试和定位,可以精确评价土壤肥力,包括土
壤的微量和大量养分、pH、盐度和碳含量等。 (4)环境相似性分析:利用最高温度、降雨量、
表土 pH 等指标进行多环境试验的序贯性模式分析
(sequential retrospective pattern analysis)。例如据此将
南部非洲的玉米大环境分成了由 A 到 H 的八大类型[23]
(图 5-B)。 (5)测试地点的选择:综合各类环境信息,可以
精确评价和确定最佳的试验点,包括适合特定环境胁
迫试验的地点,例如 CIMMYT 在非洲的干旱、低氮、
低 pH、螟虫、杂草独角金等的测试点(图 5-C)。这
对于环境依赖性试验极为重要。 4.3 试验误差控制
试验误差主要来源于农田小气候的差异,农事操
作和管理技术的不一致性,土质和肥力的不一致性,
病虫草害、野生动物、鼠雀、大风暴雨冰雹等灾害性
天气的影响,人畜家禽等引起的差异,观察记载的时
间、采用的标准、工具、仪器以及不同的观察者所引
起的差异。 控制试验误差的途径包括:(1)选择土壤质地一
致且肥力均匀的试验地,要求试验田要有土地使用历
史记录,要能代表当地的土质、气候、自然和经济条件,
要有足够的面积,并且合理轮作;(2)选择同质一致
的试验材料,同质试验材料的基因型、个体(种子质量、
幼苗大小等)、生产批次必须一致;(3)对同组试验
管理和操作要实行同一标准;(4)遵守重复、随机和
局部控制的试验设计三原则;(5)采用合理的小区设
计技术,包括优化试验小区的面积和形状、重复次数、
对照设置以及重复和小区排列方式等;(6)设置保护
行并远离村庄、树林、道路,以尽量减少边际效应。 可以利用现代土壤质地检测仪器,进行土壤质地
和肥力的测定,同时根据土壤质地进行田间试验地的
选择、试验误差的局部控制,以最大限度地控制和减
少误差[22,25](图 5-D;根据土壤质地的变化设置 2 个
重复使重复内的误差最小)。这已成为跨国公司和国
际研究机构的普遍做法。 4.4 近等环境型确定
对环境进行精细的划分,将有助于研究作物对不
同环境因子的反应。与育种中的近等基因系(near iso-genic line)概念相类似,作者提出了近等环境(near iso-environment)的概念[26],以描述只存在单个环境
因子差异的 2 种环境:一种环境对植物的影响呈中
性,而另一种对植物具有逆境胁迫作用。一种环境的
条件效应可以通过与另一(中性)环境的比较分析来
获得。由此可以衍生出近等环境型(near iso-envirotype)的概念。因此,表现型的相对性状值可以通过 2 种环
境下的表现型差值来定义。当其中一种环境为中性
时,这个相对值被称为耐逆性,而当 2 种环境均为中
性时则被称为敏感性。

3364 中 国 农 业 科 学 48 卷
0 1600 Km800
干旱 Drought
低氮 Low N
低 pH Low pH
螟虫 Stemborer
独脚金 Striga0.20
0.25
0.45
0.40
0.50
0.35
0.30
A B
CD
A
B
C
D
E
F
G
H
150
100
50
0020406080
A:FAO 2000 非洲土壤分类[24],不同土壤类型由不同颜色表示;B:南部非洲的玉米大环境分类[23];C:CIMMYT 在非洲根据环境信息确定的逆境
测试地点;D:根据土壤质地进行试验误差控制[25] A: FAO soil classes of Africa 2000[24], represented by different colors; B: Mega-environments for maize production in southern African countries [23]; C: Stress-screening sites in Africa determined by CIMMYT based on environment information; D: Control of experimental errors using soil texture information[25]
图 5 土壤特征分析及其应用
Fig. 5 Soil characterization analysis and its applications
耐逆性=中性环境下的表现型
逆境环境下的表现型中性环境下的表现型−×100%
敏感性=型第一中性环境下的表现
型第二中性环境下的表现型第一中性环境下的表现 −×100%
例如,水稻在北京、杭州两地的开花期就是 2种中性环境下的表现型,可以用来测定水稻的光周
期敏感性。研究表明,虽然控制北京和杭州两地开
花期的 QTL 是部分相同的,但由两种开花期所衍生
的光周期敏感性则是由不同的 QTL 控制的[26](图
6)。

17 期 徐云碧:作物科学中的环境型鉴定(Envirotyping)及其应用 3365 D
H系
数目
Nu m
ber o
f DH
l ine
s
DH系
数目
Nu m
ber o
f DH
l ine
s
0
2
4
6
8
10
12
14
16
18
20杭州 Hangzhou
抽穗日数 Days-to-heading (DTH)
65 71 77 83 89 95 101 107 113 119
抽穗日数和光温敏感性的 LOD 值 LODs for DTH and PTS
抽穗日数 DTH 染色体
Chr.
标记区间
Marker interval 北京
Beijing
杭州
Hangzhou
光温敏感性
PTS
1 RG400-RM84 2.41* 1.68 0.87
7 G379A-RM248 1.21 0.56 4.47*
8 RG885-RM44 7.35* 6.56* 1.07
10 C16-RM228 2.67* 3.04* 0.41
12 RG463-RG323 2.30 2.55* 0.82
*LOD 值达到显著水平 Significant LODs
图 6 水稻在北京、杭州两地的开花期及其光周期敏感性的 QTL 分析[26]
Fig. 6 QTL analysis on flowering time and photoperiod sensitivity based on the flowering data collected in Beijing and Hangzhou[26]
4.5 病害流行预测
利用影响病害流行的环境因子及其变化规律,结
合其他相关的各种社会因素等,可以对新的病害进行
流行时间、地点和范围的预测,从而为作物生产提供
预警。在小麦中,综合利用影响小麦 Ug99 可能流向
的相关因子进行了锈病流行的预测[27]。用于预测的各
类因子包括:Ug99 的现状和分布,流行风向,有利于
在小麦生产带生存、产孢和传播的气候因子,小麦产
区的人口,历史上东非锈病小种的迁移模式以及现有
品种的感病性。 4.6 伴生生物的基因改良与农艺组学
作物的伴生生物主要存在于作物的周围,特别是
根际土壤。作物的产量、品质和耐逆性受土壤环境,
特别是根际微生物群落影响极大。因此同步和协调进
行作物及其根际微生物群落的基因改良具有重要的遗
传学、生态学和农学意义。通过根际微生物群落的基
因改良,使其为作物的健康生长创造更良好的生长和
发育条件。这些条件涉及无机和有机的环境条件。无
机条件的改良包括土壤保水保肥性能的提升,土壤无
机营养从无效向有效的转化,土壤有毒物质的毒性降
解等;有机条件的改良包括土壤中有害生物种群的遏
制和有益生物种群的提升。根际微生物群落的基因改
良与目前的作物抗性或耐逆育种具有殊途同归的效
果。因此,伴生生物的基因改良(特别是根际微生物
群落的改良)是一种环境型鉴定指导下的新型生物技
术。用分子生物技术来改造或优化作物生长的环境,
使作物育种从以作物为主体扩展到作物群落系统的整
体基因改良,从而为作物产量、品质和耐逆性的遗传
改良提供新的途径。 近年来,中国农业科学院作物科学研究所的同事

3366 中 国 农 业 科 学 48 卷
们一起讨论有关如何利用基因组学研究各种栽培措施
对作物生长发育的影响。基于这些讨论,最近作者提
出了“农艺组学”(agronomic genomics)的概念。所
谓农艺组学,主要是“利用各种组学技术和方法揭示
环境因子对主要农艺性状相关基因表达的影响,在此
基础上构建调控这些基因表达的高效、节约、环境友
好的农艺措施。”利用 DNA、RNA 和蛋白质等的现
代测序技术,可以通过施肥、灌溉、病虫害防治等农
艺措施所造成的基因表达模式和数量的变化来揭示这
些农艺措施对作物生长发育的影响。可以在生物生长
的复杂波动环境下直接确定基因表达模式的各种驱动
力。目前,农艺措施相关数据分析的主要困难在于同
时影响转录组变化的环境因子具有复杂性、干扰性和
多重性。最终,农艺组学的研究将有助于培育对农艺
措施反应良好的作物新品种。 与作物生长所经历的不断变化的复杂环境相比,
在试验条件下可以更为直接地确定基因表达模式的
驱动力。Nagano 等[28]收集了稻田植株叶片的转录组
数据及其相应的气候资料(包括每一分钟间隔所测定
的风速、气温、降雨、太阳辐射、相对湿度、大气压
力等),同时在气候箱获得了下人工控温、控光条件
下的相应数据。利用统计模型研究影响基因表达的外
源和内源影响因子,结果表明,转录组动力学主要由
内源日常节律、周围温度、植物生长阶段、太阳辐射
所决定。还利用模型预测了温度变化对转录组动力学
的影响。一般来说,这类方法可以用于预测植物为应
对诸如全球变暖所导致的环境变化而可能发生的生
物学改变。 4.7 环境反应相关基因
严格控制的环境条件或人工创造的近等环境,为
深入剖析和研究与环境反应有关的基因提供了条件。
最近,Izawa[29]强调了在田间自然条件下进行植物动
力学的破译和预测的重要性。因此,综合利用环境误
差得到严格控制的环境、人工创造的近等环境、环境
因子得到精确测定和描述的环境,可以将基因型效应
和环境型效应在很大程度上区分开来,从而确定对重
要环境因子作出相关响应的基因。这对于培育和选用
适应特定环境条件的品种具有重要的意义。环境反应
相关基因可以分为两大类,一类是对中性环境产生响
应的基因,如对光周期、正常范围的温度、正常营养
条件的反应等。另一类是对逆境产生响应的基因,如
对各种环境胁迫因子(干旱、淹水、极端温度、营养
极度缺乏等)的反应。
在特定的环境条件下,已实现了对中性环境和
极端逆境产生响应的基因进行克隆和功能分析。首
先,已获得了大量与淹水、干旱、低温、铝毒、盐
胁迫等非生物胁迫耐性有关的基因,其耐性通常与
维持植物自我调节所需基因的特征性调节或功能、
复制和/或新功能化有关[30]。其次,克隆了大量与生
物逆境耐性有关的基因,如抗病虫基因等,已克隆
的水稻抗稻瘟病基因就有 10 余个[31]。第三,已克隆
了大量对常规或中性环境因子如光照、温度等的正
常条件做出相关反应的基因,包括作物的光敏色素
基因、温敏基因等。 4.8 为精准高效农业提供服务
影响作物生长的因素有很多,品种、土壤、气候、
水分、病虫害和杂草等,作物产量是这些因素综合作
用的结果。长期以来,人类收集了海量的农业相关数
据,包括气候、土壤、病虫害爆发、产量和密度之间
的相关性,以及农产品的市场价格等。通过数据整合
和模拟分析,可以为规模化农作物生产提供决策上的
帮助。在美国,农业生产模式正在从机械化向信息化
转变,以精准为特征的农业,正在让作物生产变得更
加容易。 近年来,国际种业巨头纷纷投入巨资,将产品研
发线延伸至大数据支持下的农业决策领域。孟山都近
年来收购了 Precision Planting 和 Climate Corporation两家公司。在这些专业服务公司的指导下,每 4 英亩
设 1 个取样点进行土壤分析测试,以获得各个地块详
细的土壤成分数据,从而确定种植不同作物所需要的
肥料、水分,并对产量进行预测。考虑到种植活动可
能导致的土壤成分的动态变化,每隔 3 年需要重新进
行一次土壤分析。利用 Climate Corporation 的气象数
据软件,可以获得农场范围内的实时天气信息,如温
度、湿度、风力、雨水等,这些信息可以帮助确定每
个地块的播种、收获、耕作时间[32]。 通过历年的气候数据,Climate Corporation 可以提
供更加精确的小范围气象预测。基本模式是将全球种
质资源和海量的产量数据与 Climate Corporation 的数
据库结合,描绘出信息丰富的美国农田地图,并在此
基础上叠加一切可用的气候信息。然后,依靠大数据
分析结果为农民提供农作物保险服务,农民也可以通
过该农田地图查询农场所处的区域,了解应该种植什
么种子、什么条件下长势最好等实用信息。 通过卫星导航系统、自动驾驶系统、计算机设备
以及必要的传感器,可以“理解”大数据分析软件所

17 期 徐云碧:作物科学中的环境型鉴定(Envirotyping)及其应用 3367
给出的信息,实现农业机械的智能化。例如,智能化
的农业机械可根据土地的松软程度,自动调节播种动
作,以便将种子播种于同样的深度。智能化农业机械
也大大提高了作业质量。例如,可以将单粒播种率提
高到 99%。 利用大数据支持下的移动设备,可以掌握实时的
土壤湿度、环境温度和作物状况等信息,从而大幅度
提高农场管理的精确性。Precision Planting 专门制造
的精准农业配套设备,可以固定在大马力拖拉机后部、
同步运行的播种机和其他设备上。利用包括环境数据
在内的海量数据,进行全流程的精准计算。通过内置
的应用软件,可以确定何时下地查看,何时打药、施
肥,以及提供实时的和未来几天的天气数据[32]。大数
据支持下的精准农业可以极大地节约化肥、水、农药
等投入,对各种原料的使用实行非常准确的控制,从
而实现精准高效的规模化经营。
5 展望
环境型鉴定在作物科学中具有广阔的应用前景。
展望未来,精准和高通量的表现型鉴定有赖于与环境
型鉴定相结合。这种结合需要将环境型鉴定聚焦和细
化到单个的试验小区甚至单个植株的水平。处理海量
数据需要建立相应的信息处理和决策支持系统。最终,
高效的作物育种和生产将有可能建立在基因型-表现
型-环境型三维信息支撑的基础之上。 5.1 表现型和环境型的综合鉴定
高通量和精准表现型鉴定是利用分子标记进行
性状的遗传分析、快速经济地实施分子标记辅助选
择的关键。精准表现型鉴定就是要详细地测量植株
性状,对内在基因型的性状表现型进行可靠估计。
然而,作物表现型的准确评价严重落后于对基因型
进行表征的能力。为了发展和利用高通量的表现型
鉴定技术,必须首先认识到表现型组学的重要性[33]。
为了与现行的基因型鉴定系统的能力和成本效率相
匹配,精准表现型鉴定系统必须实现高通量的数据
生成、收集、加工、分析和输出。现代作物科学的
发展趋势就是将高通量的精准表现型鉴定与大规模
的环境型鉴定相结合,进行两大类信息的综合采集、
开发和利用。 首先,目前许多自动化、高通量的表现型鉴定系
统都是建立在可控环境下的。最近开发了可以在重要
环境因素可控的生长箱中进行表现型鉴定的自动化
平台 [34-35],并建立了国际植物表型组学协作网
(international plant phenomics network,IPPN),为
高通量表现型鉴定提供新技术,比如针对植株较小、
寿命较短的模式植物进行跨越作物整个生命周期的
高通量、全自动、非侵入式成像技术,可以用于代
谢组学分析和数量表现型鉴定[36]。在拟南芥的表型
组学研究中,利用 191 个在基因组上相互重叠覆盖
的自交系对 107 个主要数量表现型进行了全基因组
关联分析,涉及病原菌抗性、开花性状以及生活史
习性[37]。要实现将理想表型组学研究与环境型鉴定
结合起来困难重重,这是因为:(1)对于大量基因
型样本的基因组信息,每个基因型都需要在一系列的
环境下进行检测;(2)需要进行跨越生长发育的整
个空间和时间范围、广泛而精细的表现型鉴定;(3)分析成本高。由于成本和表现型鉴定能力的限制,早
期的表型组学研究项目还没有一个能接近全部表型
组学视野的要求[33]。更具有挑战性的是开发适合于
生活周期长、植株高大的植物(比如玉米和高粱)的
高通量表现型鉴定平台,因为要建立适合高大植株长
期生长发育的可控环境比较困难。 与那些精心设计的人工控制环境或生长条件相
比,要模拟植株生长的田间环境是极为困难的。对
于田间精准表现型鉴定,重要的是要在同步进行环
境型鉴定的同时,通过一系列控制环境误差的措施
来降低“信噪比”。这些措施包括:选择在土壤特
性方面空间变异较小的试验小区或试验点;通过良
好的杂草和病虫害防治使不同区域的资源投入均一
化;设置适当的小区保护行;采用适当的试验设计
控制重复内变异;通过数据分析减少或消除空间趋
势;采用新的田间试验技术,比如养料和水分的精
确应用以及利用遥感技术,进行次级性状的检测;
正确选择、校准和使用各类仪器,比如中子探测仪、
射线传感器、光谱辐射计以及叶绿素和光合作用计
量器等。 同步采集的环境型信息与表现型信息,可以互为
控制条件,利用一类信息对另一类信息进行补充和矫
正。首先,利用基因型同质的材料在大规模环境下的
表现型,可以对试验点内和点间环境条件的变异进行
检测,因为显著差异的表现型是显著差异的外界环境
条件及其 G×E 互作的结果。其次,在严格控制环境
误差的试验条件下,基因型异质的材料间所存在的表
现型变异可以归结于遗传因子的结果。第三,如前所
述,在人工创造的近等环境型条件下,表现型变异是
基因型对单个目标环境因子的反应。第四,在表现型

3368 中 国 农 业 科 学 48 卷
鉴定试验中,对各种环境因子进行综合鉴定,可以获
得不同基因型在复杂但已知的环境条件下的综合表
现,由此可以确定基因型和环境型及其相互作用对表
现型的贡献。因此,表现型和环境型的同步鉴定与综
合分析,在未来作物科学试验和研究中具有广阔的应
用前景。 5.2 单个测试小区和单个植株水平的环境型鉴定
目前,对于某些环境因素,已经可以针对试验
的小区、单行、甚至单个植株进行采集。例如,针
对每个试验材料的种植小区进行土壤水分监测,土
壤营养元素和 pH 值的测定等。而对于大部分环境
因子,针对单个植物材料的环境型鉴定,还有待技
术和方法的重要革新和进步。期待未来的新技术和
新方法可以将环境型测定方法从现有的实验站和小
区水平精确、聚焦到单个测试小区甚至单株水平(图
7)。由于技术进步,期待对于每个测试材料,都可
以获得与基因型和表现型信息相对应的环境型信
息。
图 7 未来的环境型鉴定需要聚焦到单个测试小区和单株水平
Fig. 7 Future envirotyping needs to zoom into single plots and individual plants
5.3 环境信息系统与综合信息平台
环境型数据与基因型和表现型数据一起,构成了
未来数据海啸的基础。在理论上,很大部分的基因型
数据,所有的表现型和环境型都随着时间和空间的变
化而发生改变。因此,广义的信息矩阵还应包括第四
维度——时间序列。为满足日益增长的各类环境信息,
为防止生物学家和农学家被数据海啸所吞噬,必须建
立综合的环境信息系统,负责环境信息的采集、整合、
存储、挖掘、分析、利用和管理。同时还要实现环境
信息采集的标准化、环境信息术语的统一化、环境信
息利用的程序化。 综合考虑所有的四维空间(基因型、表现型和
环境型在时间序列下的变化),无疑需要一个综合
的信息平台,将所有的这些信息都整合起来,实现
全部信息的综合管理、挖掘和利用。目前,虽然基
因型和表现型实现了很大程度的整合,但这三类信
息的采集、存储、挖掘、分析、利用和管理等主要
由不同的科学团体来完成,且侧重于各自不同的工
作目标。 环境信息系统和综合信息平台的建立依赖于数据
和信息系统的标准化。信息的标准化已成为信息整合
的巨大障碍[38]。例如,土壤分类是土壤数据集成的基
础,但至今国际上还没有统一的土壤分类。美国农部
的土壤系统分类 ST(soil taxonomy)和 FAO 等国际
组织制定的 WRB(world reference base for soil resources)分类在国际上影响较大,成为主流分类体
系。为方便国际学术交流,涉及有关土壤名称时有必
要使用这两个分类体系。目前中国积累的土壤数据,
绝大多数采用的是“中国土壤发生分类”,如红壤、
水稻土、棕壤、褐土、黄绵土等发生分类的名称。但
这个分类不同于上述 2 个主流分类系统,导致国际交
流的极大困难。因此,有必要建立“中国土壤参比系
统”,即以《中国土种志》和各省土种志的数据资料
为基础,由土壤分类经验丰富的土壤学家对照各个分
类系统的诊断层和诊断特性,检索出每一土种在上述
2 个国际主流分类系统中的归属,从而建立起一个参
比系统[39]。 5.4 决策支持系统
环境型信息与基因型信息、表现型信息的综合
应用涉及到从海量数据挖掘和提取作物生产所需的

17 期 徐云碧:作物科学中的环境型鉴定(Envirotyping)及其应用 3369
有用信息并用于指导生产实践。常规的分析方法和
手段难以满足各种应用的需求。所需要的决策支持
系统包括:整合不同信息的数据库技术和方法;综
合利用基因型、表现型和环境型信息发展和优化作
物生产模型;利用基因型信息和环境型信息进行 G×E(GEI)和表现型的预测;综合利用基因型、表
现型和环境型的信息进行分子设计育种。这些决策
支持工具将促进分子生物学家、农学家、信息学家
之间开展交流和合作,从而做出有效的多学科决策。
最终,农学家和育种家需要有效地将环境型数据的
采集和分析与先进的生物统计分析、GEI 分析、数
量遗传学建模等各种不同的技术辅助措施相结合,
而实现这种结合将需要越来越复杂的辅助软件和工
具[32]。同时,还需要谨慎平衡各种不同元件,以实
现时间、成本以及生产效益之间的最佳结合,其中
包括管理和分析大量的基因型、系谱、表现型以及
环境型数据。 5.5 作物育种的三维概念
标记-性状关联的确定、标记效应的可靠估计、
标记辅助的轮回选择和全基因组选择过程中的表现
型鉴定等需要在特定的环境条件下进行,对标记辅助
选择的准确性和效率产生了重要的影响。如上所述,
精准表现型鉴定在很大程度上取决于环境因素及其
误差能否被有效地管理和控制。这对于需要在控制或
胁迫环境下进行表现型鉴定的性状(比如非生物逆境
耐性)来说尤为重要。另一方面,一直以来 GEI 是
利用特定环境下不同基因型的表现型数据进行度量
的。然而,环境数据本身很少被结合进 GEI 的估计,
也没有用于揭示环境因素对特定表现型的作用。环
境型鉴定概念,可以作为现代植物育种图像的第三
维[4]。环境型鉴定提供了描述一个植株所需要的综合
性环境信息,是对基因型信息和表现型信息的补充。
标记辅助选择导致了植物育种方法的巨大变革,通过
基于表现型的选择转换到基于表现型+基因型的选
择,它代表了植物育种概念从“线”到“面”的变化。
环境型鉴定,与基因型鉴定和表现型鉴定一起,又将
植物育种概念由基因型-表现型确定的“面”转变成
由基因型-表现型-环境型所确定的三维“空间”概念
(图 4)。
6 结论
作物的品种选育和高效生产需要全面了解其基因
型、表现型、环境因子及其相互作用。环境型鉴定,
作为生物第三类重要因子的检测手段,应该用于检测
与植物生长发育有关的所有环境因子。环境型鉴定可
以帮助全面了解环境因子对生物的影响。通过已有的
各种环境信息数据库或利用各种环境信息采集工具可
以进行环境型鉴定。环境信息可以广泛应用于作物科
学研究的许多方面,包括 G×E 互作的分解、环境特
征分析、试验误差控制、流行性病虫害预测、特定环
境反应基因的鉴定等。环境型信息构成了未来植物育
种所需的第三维图像。通过大规模环境型鉴定,结合
已知的基因型信息,未来可以采用生物技术优化作物
生长环境,对特定环境下作物的表现型进行精准的预
测,从而推动作物科学研究的预见性和作物生产的高
效性。 致谢:王天根博士、两位审稿人的建议和评论,美国新
墨西哥州立大学 Paul Bosland 博士关于环境型鉴定
(envirotyping)英文表述的建议,中国农业科学院作物科
学研究所贾继增博士关于利用基因组学研究栽培措施影响
作物生长发育的讨论,在此表示感谢。
References
[1] Tilman D, Cassman K G, Matson P A, Naylor R, Polasky S.
Agricultural sustainability and intensive production practices. Nature,
2002, 418: 671-677.
[2] Chaudhary R C. Strategies for bridging the yield gap in rice: A
regional perspective for Asia. International Rice Commission
Newsletter, 2000, 49: 22-31.
[3] Xu Y. Environmental assaying or e-typing as a key component for
integrated plant breeding platform, Marker-Assisted Selection
Workshop, 6th International Crop Science Congress, August 6-10,
2012, Bento Goncalves, RS, Brazil.
[4] Xu Y, Lu Y, Xie C, Gao S, Wan J, Prasanna B M. Whole-genome
strategies for marker-assisted plant breeding. Molecular Breeding,
2012, 29: 833-854.
[5] http://www.dilikexue.cn/geog/show.aspx?id=2234&cid=35.
[6] 史学正, 于东升, 高鹏, 王洪杰, 孙维侠, 赵永存, 龚子同. 中国土
壤信息系统 (SISChina)及其应用基础研究 . 土壤 , 2007, 39(3):
329-333.
Shi X Z, Yu D S, Gao P, Wang H J, Sun W X, Zhao Y C, Gong Z T.
Soil information system of china (SISChina) and its application. Soils,
2007, 39(3): 329-333. (in Chinese)
[7] http://www.issas.ac.cn/ztwz/200910/t20091015_2551708.html.
[8] Shi X Z, Yu D S, Warner E D, Pan X Z, Petersen G W, Gong Z G,

3370 中 国 农 业 科 学 48 卷
Weindorf D C. Soil Database of 1:1,000,000 digital soil survey and
reference system of the Chinese genetic soil classification system. Soil
Survey Horizons, 2004, 45(4): 129-136.
[9] http://www.issas.ac.cn/ztwz/200910/t20091015_2551708.html.
[10] Smith P, Furse C, Gunther J. Analysis of spread spectrum time
domain reflectometry for wire fault location. IEEE Sensors Journal,
2005, 5: 1469-1478.
[11] Fahlgren N, Gehan M A, Baxter I. Lights, camera, action:
High-throughput plant phenotyping is ready for a close-up. Current
Opinion in Plant Biology, 2015, 24: 93-99.
[12] Pound M P, French A P, Murchie E H, Pridmore T P. Automated
recovery of three-dimensional models of plant shoots from multiple
color images. Plant Physiology, 2014, 166: 1688-1698.
[13] Li Y, Fan X, Mitra N J, Chamovitz D, Cohen-Or D, Chen B.
Analyzing growing plants from 4D point cloud data. ACM
Transactions on Graphics, 2013, 32: 1-10.
[14] Paulus S, Dupuis J, Mahlein A K, Kuhlmann H. Surface feature based
classification of plant organs from 3D laser scanned point clouds for
plant phenotyping. BMC Bioinformatics, 2013, 14: 238.
[15] Chéné Y, Rousseau D, Lucidarme P, Bertheloot J, Caffier V, Morel P,
Belin É, Chapeau-Blondeau F. On the use of depth camera for 3D
phenotyping of entire plants. Computers and Electronics in
Agriculture, 2012, 82: 122-127.
[16] Des Marais D L, Hernandez K M, Juenger T E. Genotype-by-
environment interaction and plasticity: exploring genomic responses
of plants to the abiotic environment. Annual Review of Ecology
Evolution and Systematics, 2013, 44: 5-29.
[17] Paterson A H, Damon S, Hewitt J D, Zamir D, Rabinowitch H D,
Lincoln S E, Lander E S, Tanksley S D. Mendelian factors underlying
quantitative traits in tomato: Comparison across species, generations,
and environments. Genetics, 1991, 127: 181-197.
[18] Hammond J P, Mayes S, Bowen H C, Graham N S, Hayden R M,
Love C G, Spracklen, W P, Wang J, Welham S J, White P J, King G J,
Broadley M R. Regulatory hotspots are associated with plant gene
expression under varying soil phosphorus supply in Brassica rapa.
Plant Physiology, 2011, 156: 1230-1241.
[19] Grishkevich V, Yanai I. The genomic determinants of genotype×
environment interactions in gene expression. Trends in Genetics, 2013,
29: 479-487.
[20] Kim D H, Doyle M R, Sung S, Amasino R M. Vernalization: Winter
and the timing of flowering in plants. Annual Review of Cell and
Development Biology, 2009, 25: 277-299.
[21] Lee W S, Alchanatis V, Yang C, Hirafuji M, Moshou D, Li C. Sensing
technologies for precision specialty crop production. Computers and
Electronics in Agriculture, 2010, 74: 2-33.
[22] Araus J L, Cairns J E. Field high-throughput phenotyping: The new
crop breeding frontier. Trends in Plant Science, 2014, 19: 52-61.
[23] Bänziger M, Setimela P S, Hodson D, Vivek B. Breeding for
improved abiotic stress tolerance in maize adapted to southern Africa.
Agricultural Water Management, 2006, 80: 212-224.
[24] http://eusoils.jrc.ec.europa.eu/.
[25] Prasanna B M, Cairns J, Xu Y. Genomic tools and strategies for
breeding climate resilient cereals// Kole C. Genomics and Breeding
for Climate-Resilient Crops. Berlin Heidelberg: Springer-VerlagPress,
2013: 213-239.
[26] Xu Y. Global view of QTL: Rice as a model//Kang M S. Quantitative
Genetics, Genomics and Plant Breeding. Wallingford, UK: CABI
Publishing, 2002: 109-134.
[27] Singh R P, Hodson D P, Jin Y, Huerta-Espino J, Kinyua M G,
Wanyera R. Current status, likely migration and strategies to mitigate
the threat to wheat production from race Ug99 (TTKS) of stem rust
pathogen. CAB Reviews, 2006, 54: 1-13.
[28] Nagano A J, Sato Y, Mihara M, Antonio B A, Motoyama R, Itoh H,
Nagamura Y, Izawa T. Deciphering and prediction of transcriptome
dynamics under fluctuating field conditions. Cell, 2012, 151:
1358-1369.
[29] Izawa T. Deciphering and prediction of plant dynamics under field
conditions. Current Opinion in Plant Biology, 2015, 24: 87-92.
[30] Mickelbart M V, Hasegawa P M, Bailey-Serres J. Genetic
mechanisms of abiotic stress tolerance that translate to crop yield
stability. Nature Reviews Genetics, 2015, 16: 237-251.
[31] 邱丽娟, 郭勇, 黎裕, 王晓波, 周国安, 刘章雄, 周时荣, 李新海,
马有志, 王建康, 万建民. 中国作物新基因发掘: 现状、挑战与展
望. 作物学报, 2011, 37(1): 1-17.
Qiu L J, Guo Y, Li Y, Wang X B, Zhou G A, Liu Z X, Zhou S R, Li X
H, Ma Y Z, Wang J K, Wan J M. Novel gene discovery of crops in
China: status, challenging, and perspective. Acta Agronomica Sinica,
2011, 37(1): 1-17. (in Chinese)
[32] http://www.agrogene.cn/info-1949.shtml.
[33] Houle D, Govindaraju D R, Omholt S. Phenomics: The next challenge.
Nature Reviews Genetics, 2010, 11: 855-866.
[34] Jansen M, Gilmer F, Biskup B, Nagel K A, Rascher U, Fischbach A,
Briem S, Dreissen G, Tittmann S, Braun S, De Jaeger I, Metzlaff M,
Schurr U, Scharr H, Walter A. Simultaneous phenotyping of leaf
growth and chlorophyll fluorescence via GROWSCREEN FLUORO
allows detection of stress tolerance in Arabidopsis thaliana and other

17 期 徐云碧:作物科学中的环境型鉴定(Envirotyping)及其应用 3371
rosette plants. Functional Plant Biology, 2009, 11: 902-914.
[35] Massonnet C, Vile D, Fabre J, Hannah M A, Caldana C, Lisec J,
Beemster G T S, Meyer R C, Messerli G, Gronlund J T, Perkovic J,
Wigmore E, May S, Bevan M W, Meyer C, Rubio-Díaz S, Weigel D,
Micol J L, Buchanan-Wollaston V, Fiorani F, Walsh S, Rinn B,
Gruissem W, Hilson P, Hennig L, Willmitzer L, Granier C. Probing
the reproducibility of leaf growth and molecular phenotypes: A
comparison of three Arabidopsis accessions cultivated in ten
laboratories. Plant Physiology, 2010, 152: 2142-2157.
[36] Bergelson J, Roux F. Towards identifying genes underlying
ecologically relevant traits in Arabidopsis thaliana. Nature Reviews
Genetics, 2010, 11: 867-879.
[37] Atwell S, Huang Y S, Vilhjálmsson B J, Willems G, Horton M, Li Y,
Meng D, Platt A, Tarone A M, Hu T T, Jiang R, Muliyati N W, Zhang
X, Amer M A, Baxter I, Brachi B, Chory J, Dean C, Debieu M, de
Meaux J, Ecker J R, Faure N, Kniskern J M, Jones J D, Michael T,
Nemri A, Roux F, Salt D E, Tang C, Todesco M, Traw M B, Weigel D,
Marjoram P, Borevitz J O, Bergelson J, Nordborg M. Genome-wide
association study of 107 phenotypes in Arabidopsis thaliana inbred
lines. Nature, 2010, 465: 627-631.
[38] Xu Y. Molecular Plant Breeding, Wallingford. UK: CAB
International. 2010: 734pp.
[39] Shi X Z, Yu D S, Warner E D, Sun W X, Petersen G W, Gong Z T.
Cross-reference system for translating between genetic soil
classification of china and soil taxonomy. Soil Science Society of
America Journal, 2006, 70: 78-83.
(责任编辑 李莉)

中国农业科学 2015,48(17):3372-3387 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.005
收稿日期:2015-01-19;接受日期:2015-06-15 基金项目:国家“863”计划(863-Z17-01-01)、农业部“发展棉花专项资金”(99044)、科技部“转基因植物产业化”专项(J99B-002)、中华农业
科教基金(99-01-04)、国家计委高科技产业化示范工程(计高技-20010482)、转基因生物新品种培育重大专项(ZX08005-004) 联系方式:郭三堆,Tel:010-82106140;E-mail:[email protected]
中国转基因棉花研发应用二十年
郭三堆1,王 远
1,孙国清
1,金石桥
2,周 焘
1,孟志刚
1,张 锐
1
(1 中国农业科学院生物技术研究所/国家农作物基因资源与遗传改良重大科学工程,北京 100081;2 全国农业技术推广服务中心,北京 100125)
摘要:转基因技术是指利用重组 DNA 原理,将优良目的基因整合到靶标生物基因组中,并使靶标生物得以表
达目的性状的技术。这一技术克服了生物有性杂交的限制,使物种间基因交流的范围无限扩大,既可以从原核生
物到真核生物,也可以从单细胞生物到多细胞生物,还可以从低等生物到高等生物,反之亦然。因此,这项技术
自发明以来,即广泛应用于农业、林业、医学等领域,为其研究开辟了一个全新的时代。转基因植物是以农杆菌
等为媒介,将来源于动物、植物或微生物等其他生物甚至人工合成的外源基因转入基因组中,使之稳定遗传并赋
予其靶标性状,如抗病、抗虫、抗逆、高产、优质等的植物。以 1972 年构建第一个重组 DNA 分子为契机,1983
年首次获得转基因烟草为起点,植物转基因技术在近 30 年的时间内发展迅猛,至今已有 200 多种植物已成功获得
转基因株系,40 多种数千例转基因植株进入田间试验。根据国际农业生物技术应用咨询服务中心(ISAAA)统计,
全球转基因植物的种植面积由 1996 年的 260 万 hm2已经迅速增到 2014 年的 1.815 亿 hm
2,累计种植面积大约比中
国国土总面积还多 80%。在全球转基因植物研发和应用迅猛发展的同时,中国也先后批准了 7 种转基因植物的生
产应用,其中,抗虫棉是唯一大规模应用的转基因农作物。从 1994 年中国研制成功国产单价抗虫棉(GK),以及
1995 年美国保铃棉进入中国至今,抗虫棉已经在中国推广应用了近 20 年的时间。文章介绍了这 20 多年来,中国
科学家在抗虫、抗旱耐盐碱、抗除草剂、抗病以及纤维品质改良等性状方面所取得的转基因棉花研究进展;在农
杆菌介导、基因枪轰击、花粉管通道介导、茎尖或芽尖转化、农杆菌液浸染和纳米载体花粉介导等不同转化技术
上所进行的探索;同时,介绍了中国转基因植物的安全性评价状况,并从抗虫棉品种审定、发展趋势和产业化状
况几个方面,介绍了转基因抗虫棉在中国的应用,最后对未来转基因棉花研究方向进行展望。
关键词:抗虫棉;转基因;安全评价;品种;产业化
Twenty Years of Research and Application of
Transgenic Cotton in China GUO San-dui1, WANG Yuan1, SUN Guo-qing1, JIN Shi-qiao2, ZHOU Tao1, MENG Zhi-gang1, ZHANG Rui1
(1Biotechnology Research Institute, Chinese Academy of Agricultural Sciences/National Key Facility for Crop Gene Resources and
Genetic Improvement, Beijing 100081; 2The National Agro-Tech Extension and Service Center, Beijing 100125)
Abstract: Genetic engineering mainly deals with the transfer and expression of functional genes into the targeted genome of an organism to have desired phenotype. This technique has stunned the limitations of sexual hybridization by allowing the transfer of genes among species from prokaryotes to eukaryotes, unicellular to multicellular, lower organisms to higher organisms and Vice versa. This technology has opened the avenues of research, and since its invention scientists are readily using it in the field of agriculture, forestry and medicine. The use of different techniques like agrobacterium mediated transformation has made it possible to transfer different genes to the targeted genomes for deploying the resistance against biotic and abiotic stresses like diseases, insect/pests, drought and salinity as well as for the improvement of yield and quality of plants. Since the birth of first transgenic
jia
文本框
返回目录

17 期 郭三堆等:中国转基因棉花研发应用二十年 3373
tobacco in 1983, more than 200 plant species have been used by the researchers for genetic transformation and some thousands of transgenic plants of 40 species are under field trials. International Service for the Acquisition of Agri-Biotech Applications (ISAAA) has reported the dramatic increase in area under transgenic plant, i.e. 2.6 million hectares in 1996 to 181.5 million hectares in 2014 around the whole globe. The accumulated area of transgenic crops in the world is 80% more than the total land of China. With the global large scale adoption and application of transgenic plants development technology, seven transgenic plants also have been approved by Chinese government for general cultivation. Among these, insect resistant transgenic cotton including series of GK and SGK cotton cultivars developed by the Chinese scientists in 1994 and 1998 respectively, as well as the Boll guard imported from USA in 1995 was the only crop occupying the large cultivated area of China. This paper discussed the course of development of transgenic cotton for resistance against diseases, insects and herbicides as well as for the improvement of fiber yield and quality in China and improved methods of genetic transformation like Agrobacterium mediated, tissue culture, gene gun bombardment, pollen tube pathway, shoot tip, floral-dip and nano-carrier pollen mediated transformations. Finally this paper also discussed the biosafety system regarding the transgenic plants development, Bt cotton seed industry and future prospects of transgenic cotton in China.
Key words: insect resistant cotton; transgene; biosafety evaluation; variety; industrialization
0 引言
从 1994 年中国研制成功单价抗虫棉(GK),以
及 1995 年美国保铃棉进入中国以来,抗虫棉在中国推
广应用了近 20 年的时间。在这期间,虽然中国先后批
准了 7 种转基因植物生产应用,但在中国转基因农作
物产业化应用方面,抗虫棉仍然是一枝独秀[1]。 本文介绍了 20 多年来,中国科学家在抗虫、抗除
草剂、抗旱耐盐碱、抗病、纤维品质改良等转基因棉
花研究方面所取得的进展,以及在农杆菌介导、花粉
管通道、茎尖转化等不同转化技术上所进行的探索。
同时,以国产抗虫棉为例,介绍了中国转基因生物安
全性评价体系,概述了抗虫棉在中国的产业化历程,
并对转基因棉花的研究方向作出进一步展望。
1 不同类型转基因棉花研究进展
利用基因工程技术对棉花进行遗传改良是棉花分
子设计育种的主要途径,有效地弥补了传统育种方法
不能解决的问题,快速地培育出多种新型棉花育种材
料。棉花遗传改良主要涉及抗逆、抗除草剂、纤维品
质和早衰等重要农艺性状改良。其中,抗逆基因工程
培育的抗虫转基因棉花和抗除草剂转基因棉花得到了
广泛的生产应用,并取得了巨大的经济和生态效益。
同时,为解决粮食增产与耕地面积不断缩小的矛盾,
使棉花能够在粮食作物不能种植的干旱地和盐碱地进
行栽培生产,培育具有优质、高产、耐旱耐盐碱、抗
病、抗早衰等重要农艺性状的新型转基因棉花材料成
为当前棉花遗传改良的研究热点。此外,适宜于南方
多雨高湿地区栽培的耐涝转基因棉花等特殊用途的新
材料培育也取得了一定的进展。
1.1 抗虫转基因棉花
中国抗虫基因的研制起始于20世纪90年代初期,
通过遗传转化方法使棉花获得抗虫功能的基因主要包
括三类:来源于苏云金芽孢杆菌的杀虫蛋白基因(Bt)、从植物中分离的昆虫蛋白酶抑制剂基因(PI)和植物
凝集素基因(1ectin)。当前,大规模生产应用的国产
转基因抗虫棉主要为单价抗虫棉和双价抗虫棉,其遗
传转化的抗虫基因为 Bt 杀虫基因 GFM Cry1A 和豇豆
胰蛋白酶抑制剂基因 Cpti。 1992 年,郭三堆等[2]利用分子设计技术人工合成
了具有高杀虫活性的 Bt 杀虫基因 GFM Cry1A 并于
1994 年将该基因导入棉花,在此基础上与育种单位合
作,成功选育出 GK1、GK12、GK19、GKZ1 和晋棉
26 国产单价转基因抗虫棉品种,并大面积推广应用,
使中国成为继美国之后世界上第二个研制成功转基因
抗虫棉的国家。之后,为了提高抗虫棉的杀虫效率、
延缓棉铃虫等害虫对杀虫蛋白产生抗性,郭三堆等[3]
将 2 个不同杀虫机理的抗虫基因 GFM Cry1A 和 Cpti同时导入棉花,创制了双价转基因棉花新种质。通过
与国内多家育种单位合作,成功选育出 sGK321(1998年)、中棉所 41(2001 年)和中棉所 45(2003 年)
等双价转基因抗虫棉新品种。田间抗虫性试验结果表
明,2、3、4 代棉铃虫平均双价抗虫棉百株幼虫数量
分别比常规棉田减少 81.4%、87.1%和 87.0%,分别比
单价抗虫棉田减少 11.1%、33.3%和 57.1%[4]。双价转
基因抗虫棉的研制成功,不仅在抗虫性上比单价抗虫
棉优越,更重要的是丰富了抗虫棉种质资源,加速了
抗虫棉新品种的培育和推广应用[3]。 此外,转 GFM Cry1A 和 Cpti 双价转基因抗虫棉
的大面积推广应用,为高效广谱抗虫棉的研制奠定了

3374 中 国 农 业 科 学 48 卷
理论基础,推动了新抗虫基因的研制和应用,尤其是
在抗棉铃虫和蚜虫等棉花主要害虫的防治方面取得了
突破性的进展。王伟等[5]利用根癌农杆菌介导遗传转
化方法获得含有豌豆外源凝集素基因 P-Lec 和大豆
Kunitz 型胰蛋白酶抑制剂基因 SKTI 的双基因转基因
棉花,对蚜虫表现出高效的抗虫性。刘志等[6]将 Cry1A和雪花莲凝集素基因 GNA 导入棉花,选育获得转
Cry1A+GNA 双基因抗虫棉花纯合株系 TBG,在苗期、
蕾期、开花期和结铃期主茎叶都对棉铃虫幼虫具有很
高杀虫活性。同时,TBG 植株在不同生育时期对棉蚜
群体也表现出明显的抑制效果,其苗期和蕾期抑蚜率
分别达到了 50.31%和 46.61%。吴家和等[7]将入工设计
合成的 Cry1Ac、Bt29K 和慈菇蛋白酶抑制剂基因
API-B 导入棉花,培育出 9 个抗虫性好、农艺性状优
良的转基因棉品系,对棉铃虫抗性均达到 90%以上。
肖松华等[8]将野生荠菜凝集素基因(WSA)导入常规
陆地棉品种,对 9 个高品质转 WSA 棉花株系进行抗蚜
鉴定,结果表明,转基因棉花株系对棉蚜均表现出较
高的抗性。张林水等[9]将 Cry1Ac3 与 Cpti 的融合基因
和 GNA 共转化棉花,获得了同时含有 3 个抗虫基因的
转基因棉花新材料,该材料对棉铃虫具有较高的抗性。
雒瑶瑜等[10]将 Cry1Ac 与 Cry2Ab 导入棉花,获得了双
基因抗虫棉育种材料 2009002。该转基因棉花除了抗
棉铃虫外,对小地老虎、刺吸类害虫、蛀食类害虫和
食叶类害虫有一定的抗虫性。 多基因共转化抗虫棉花的研制为中国后续抗虫基
因的研制和应用研究开拓了新思路。通过构建多基因
植物表达载体共转化不同杀虫机制的抗虫基因,或者
通过基因聚合技术将多个抗虫基因聚合于一个转基因
棉花材料内,培育获得含有多个基因的转基因抗虫棉。
不仅可以使棉花受体获得多种机制的抗虫性,利用基
因互补的方式加强转基因棉花的抗虫能力延缓害虫产
生抗(耐)性,而且可以拓宽棉花抗虫谱,使其获得
能够高效杀死主要害虫的广谱抗虫性[11]。 1.2 抗旱耐盐碱转基因棉花
中国水资源的短缺,土壤盐渍化和频繁的极端天
气严重影响作物的生产,此外,在有限耕地的前提条
件下,粮棉征地的矛盾日益凸显,通过研究抗逆基因
提高棉花的抗逆能力,将有效地提高土地利用率,拓
展可利用土地资源。近年来,中国在抗旱耐盐碱基因
的研究方面取得了一些重要进展。 1.2.1 抗旱转基因棉花 水资源的日益缺乏,全球温
室效应的不断加剧,对棉花生产带来了极大挑战,因
此,抗旱基因的相关研究对于提高棉花产量和加强盐
碱地及干旱地的开发利用具有极其重要的意义[12]。
吕素莲[13]将来自大肠杆菌的编码胆碱脱氢酶(CDH)
基因 betA 导入棉花,发现该基因能够显著提高转基
因棉花的抗旱和耐盐性,通过棉花苗期和蕾期的渗
透(干旱)及盐胁迫试验发现来自于盐芥的 TsVP可以提高转基因棉花的抗渗透(干旱)和盐能力,
该工作一方面为培育抗旱、耐盐棉花新品种创造了
优异材料,为中国棉花生产和大面积的盐碱地开发
利用作出了力所能及的贡献;另一方面为深入了解
棉花抗旱、耐盐的分子机制提供了重要资料,开辟
了新的途径。 曹燕燕等[14]通过对 rolB 的功能研究,发现该基因
不仅使转基因棉的茎粗明显提高,而且还使其生根能
力明显增强,根/冠比显著大于对照,表现为根系更为
强大,抗旱性更强。王娟[15]对转 ZmPIS 棉花植株在苗
期、蕾期和开花期的干旱胁迫研究表明转基因棉花的
一些株系具有较强的渗透调节能力,其抗旱能力显著
提高,转基因棉花植株具有较强的渗透调节能力,此
外,王娟[15]通过杂交,将来自大肠杆菌的编码胆碱脱
氢酶(CDH)基因 betA 和来自于盐芥的 TsVP 聚合在
一起,结果表明,转基因聚合植株与转单基因植株相
比具有较高的耐旱性。吴伟[16]在前人研究的基础上,
对聚合ZmPLC1和betA的转基因聚合棉花进行耐旱性
研究,结果表明,与野生型对照和转单基因对照株系
相比,转基因聚合株系具有更高的干旱耐性,而且相
比于野生型对照株系和转单基因对照株系,转基因聚
合株系具有更高的籽棉和皮棉产量,这表明通过基因
聚合手段提高植物抗逆性是可行的。杨云尧 [17]将
MvP5CS 和 MvNHX1 整合到棉花中,通过研究发现转
基因株系能够提高棉花的抗旱能力。蔡永智[18]在转基
因棉花中对来源于葛兰菜的 CBF1 和来源于大肠杆菌
的 KatG 进行功能研究,发现在干旱胁迫下,转 CBF1和 KatG 棉花在光合作用、叶绿素荧光参数、生理生
化以及农艺性状方面均表现出优良的生长和生理优
势。说明这两个基因能通过提高抗逆能力来提高转基
因棉花的产量。钱进等[19]通过分析转 MvNHX1 的 10个 T5棉花株系在干旱胁迫后的生理指标和农艺性状,
发现经干旱胁迫后转基因植株光合作用更强,根系更
发达,吸收水的能力更强,农艺性状方面,转基因株
系的有效铃数、有效果枝数、单铃重、皮棉、子棉和
衣分等指标明显高于对照,这些株系可以作为后续遗
传育种材料[20]。

17 期 郭三堆等:中国转基因棉花研发应用二十年 3375
1.2.2 耐盐碱转基因棉花 棉花是公认的耐盐、抗旱
作物,克隆优良耐盐碱基因,导入棉花选育耐盐棉花
品种,提高棉花对盐渍环境的适应能力,利用围垦改
造的滩涂地,种植转基因耐盐碱棉花,是减轻土壤盐
渍化危害以及开发利用沿海滩涂等盐渍化土地资源的
重要途径之一,是缓解粮棉争地矛盾的重要策略。张
慧军等 [21]将克隆自山菠菜(Atriplex hortensis)的
AhCMO 导入泗棉 3 号棉花,盐胁迫试验结果表明,
转 AhCMO 的棉花耐盐性显著优于对照组棉株,说明
AhCMO 提高了转基因棉花对盐胁迫的耐受性。大肠杆
菌的 betA 编码的胆碱脱氢酶(choline dehydrogenase,CDH)兼具有胆碱单氧化物酶和甜菜碱醛脱氢酶的功
能。连丽君[22]对转 betA 鲁棉研 19 的耐盐研究表明,
betA 的表达提高了转基因棉花叶片中甘氨酸甜菜碱
(glycinebetaioe,GB)的累积量,转基因棉花的耐盐
能力明显提高。樊文菊[23]对转 ZmPLC1 棉花和聚合
ZmPLC1/betA 棉花的耐盐性进行初步研究,通过水培、
沙培和东营盐碱地种植,发现转 ZmPLC1 棉花的耐盐
性高于野生型对照,转 betA 和 ZmPLC1 的聚合转基因
植株,籽棉产量显著高于转 ZmPLC1 棉花、转 betA 棉
花和野生型对照,以上工作表明,转 ZmPLC1 单个基
因可以在一定程度上提高棉花的耐盐性,且聚合 betA和 ZmPLC1 2 个基因后,棉花的耐盐性又进一步提高。
因此,可以通过基因聚合的方法提高植物的耐盐性,
为培育耐盐棉花新品种提供新的种质资源,也为研究
多个耐盐基因相互作用及其调控机制提供新的试验材
料。陈翠霞等[24]利用花粉管通道技术将耐盐碱的罗布
麻(Apocynum venetum)DNA 导入鲁棉 6 号,从后代
中筛选出棉花耐盐变异体山农 011,对山农 011 及其
杂种后代进行了抗盐遗传和基因效应分析,结果表明,
变异体的耐盐性是由核基因控制的,其遗传基因效应
以加性效应为主,并存在较大的加性互作效应。该耐盐
变异体可作为一良好的抗盐种质材料利用。李雪林[25]
通过过表达对逆境起调控作用的转录因子基因
SNAC1,系统地对转基因棉花植株进行抗逆性分析和
鉴定,验证了 SNAC1 是具有耐旱及耐盐的转录因子基
因。中国农业科学院生物技术研究所郭三堆课题组将
具有自主知识产权的耐盐碱关键基因 GhABF2(专利
号:ZL200910158311.X)导入中国棉花主栽品种苏棉
12 号,创制出耐盐碱转基因棉花新品系 8 个,进入中
间试验阶段。经过连续 3 年分别在山东、新疆两地进
行的耐盐碱试验鉴定,获得 4 个综合农艺性状优良、
耐盐碱性能突出的转基因棉花新品系[26]。在 0.40%—
0.45%的盐碱条件下,受体棉花品种苏棉 12 出苗率只
有 32%,转 GhABF2 棉花新品系出苗率高达 65%,单
株结铃比受体增加 3—4 个, 终两者产量分别比其受
体对照增加 33%以上,具有较大的盐碱地生产应用潜
力,该课题组还通过农杆菌介导棉花下胚轴遗传转化
方法,将溶菌酶 hel(抗病)、CP4-EPSPS(抗除草剂)
和 Na+/H+逆向转运蛋白基因(NhaD k3)同时导入棉
花模式受体 R15 中,再生植物经过连续 3 代自交后,
经抗草甘膦、抗棉花黄萎病及耐盐碱能力筛选,得到
高抗棉花黄萎病、抗除草剂草甘膦及具一定耐盐碱能
力的转基因棉花新品系 T58-22。 1.3 抗除草剂转基因棉花
草害是影响棉花生产的主要因素之一,棉田杂草
种类严重影响棉花的生长。除草剂和抗除草剂棉花的
协同使用是棉田杂草防除的重要对策。目前,抗除草
剂主要有 2 种策略:一是引入降解除草剂的酶或酶系
统,在除草剂发生作用前将其分解。二是修饰除草剂
作用的靶蛋白,使其对除草剂不敏感或促使其过量表
达以使作物吸收除草剂后仍能正常代谢。中国科学家
针对这两种策略均做了大量研究[27]。 1.3.1 抗草甘膦 草甘膦(glyphosate)是世界上使
用 广泛的一种非选择性内吸传导有机膦类除草剂。
其机理是竞争性抑制植株中 5-烯醇丙酮莽草酸-3-磷酸合成酶(EPSPs)的活性,阻断芳香族氨基酸的合
成,造成芳香族氨基酸缺乏,导致植株死亡。国内对
抗草甘膦的基因进行了大量研究,取得了很多可喜的
进展。 雷凯健等[28]2006 年从土壤总 DNA 中克隆到草甘
膦 N-乙酸转移酶基因,并对其表达蛋白的酶学特性进
行了分析,为其在抗草甘膦转基因作物中的应用积累
了理论依据。何鸣等[29]通过对鼠伤寒沙门氏杆菌和大
肠杆菌的 EPSP 合成酶基因多位点突变,克隆了突变
的 aroA,并使之在大肠杆菌中得到表达。赵特[30]从严
重草甘膦污染的土壤中分离出一株高抗草甘膦的假单
胞杆菌菌株,克隆到与 EPSPS 同源的 G6 基因,将其
转化水稻进行功能验证,发现该基因具有一定的抗草
甘膦活性。朱玉[31]从长期污染草甘膦的土壤中分离到
一株草甘膦高耐受性荧光假单胞菌 G2 菌株,从中克
隆到的 epsps 不含国外公司专利保护的保守序列和突
变位点,显示了良好的开发应用潜力。沙纪莹[32]从斯
氏假单胞菌基因组文库中克隆到一个完整的基因,能
在草甘膦浓度为 150 mmol·L-1的 M9 液体培养基中良
好生长,该基因有望成为培育高抗草甘膦转基因作物

3376 中 国 农 业 科 学 48 卷
的基因材料。刘柱[33]从被草甘膦极度污染的土壤中
分离到一株极端抗草甘膦菌 HTG7,该菌株能在草甘
膦浓度为 900 mmol·L-1的 Mops 限制性培养基上生长。
从中克隆到全长 1 350 bp、编码 EPSP 合酶的基因。
该 EPSP 合酶基因与之前报道的 aroA 在核苷酸水平
几乎没有任何同源性,蛋白序列与 22 种已报道的微
生物来源的 EPSP 合酶的同源性也在 46%以下。并
通过错误倾向 PCR 方法,找到了该 EPSP 合酶的关
键活性位点,为培育具有中国自主产权的转基因抗
草甘膦作物提供的基因资源。童旭宏等[34]从陆地棉
中克隆到 EPSPS,对该基因进行了组织特异性表达
分析,为抗草甘膦棉花种质资源的创制提供了理论
基础。程海刚等 [35]从棉田间的杂草青麻中克隆到
epsps,通过转化酵母证明所获得的 epsps 具有生物
学功能。之后,中国农业科学院生物技术研究所郭
三堆课题组将该基因转化棉花,筛选出具有良好草
甘膦抗性的棉花植株。此外郭三堆课题组还和中国
农业科学院生物技术研究所林敏课题组合作,将林
敏课题组从极端抗草甘膦微生物中克隆到的抗草
甘膦新型 EPSP 合酶 GR79 及 N-乙酰转移酶 GAT 进
行了密码子改造和基因结构优化,通过转化烟草和
棉花,初步验证了抗草甘膦基因的抗性,并获得了
对草甘膦有较强抗性的双价抗草甘膦转基因棉花材
料[36-37]。 1.3.2 抗其他除草剂 除草剂 2,4-D 类似植物生长
素,前人从土壤细菌 Alcaligene eutrophus 分离到 tfda,该基因编码的 2,4-D 单氧化酶能将 2,4-D 降解为 2,4-二氧苯酚,其降解产物对植物的毒性比 2,4-D 小 100倍,山西省棉花研究所陈志贤等与澳大利亚 CSIRO 及
中国农业科学院生物技术中心合作,将 tfda 导入晋棉
7 号、冀合 321 等棉花品种,对其后代进行田间抗药
性鉴定表明转基因系对 2,4-D 的耐受性超过了大田使
用浓度[38]。中国农业科学院棉花研究所与中国科学院
上海植物生理研究所合作,将除草剂草丁膦的 bar 导
入棉花主栽品种,取得了阶段性成果[39]。张磊[40]通过
精细定位,从水稻中克隆到 bel,该基因可对苯达松和
磺酰脲类除草剂产生抗性。溴苯腈(bromoxyril)是除
草剂 buctril 的活性成分,其作用机理是抑制光合作用
的电子传递。张金文等[41]2006 年从克雷伯氏臭鼻杆菌
中克隆到编码腈水解酶基因 bxn,通过在大肠杆菌中
表达,证明其可将溴苯腈降解为无毒物质。乙酰乳酸
合成酶(ALS,EC2.2.1.6)在细菌、酵母和植物中催
化支链氨基酸的生物合成,它是磺酰脲类除草剂的作
用靶标。宋贵生等[42]2007 年从水稻中克隆了 als,转
化烟草后发现转基因植株和对照烟草相比其除草剂抗
性高达 50 mg·L-1。卢宗志等[43]2009 年从杂草雨久花中
克隆到 als,研究表明抗药性雨久花对除草剂苄嘧磺的
抗性可能与其第 197 位的突变相关。 1.4 抗病转基因棉花
棉花病害是影响棉花生产的主要因素之一,目前 困扰中国棉花产业的主要病害是黄萎病,棉花黄萎病
是黄萎病菌经土壤传播、侵染到棉花植株 终引发维
管束疾病的一种真菌性病害,具有危害严重、分布范
围大、寄主种类多及存活时间长等特点,可造成棉花
大量减产甚至绝收,被形象地称为棉花的“癌症”。
针对以黄萎病为代表的主要病害,采用传统的防治手
段,如通过传统育种的方法培育抗病品种、农药防治
等方法收效甚微,而且存在培育周期长,严重污染环
境等问题,因此,近年来中国研究人员从生物技术的
角度,对抗棉花黄萎病基因开展了大量的研究,并取
得了重要进展。 高巍[44]以海岛棉品种海 7124 作为研究材料,通
过比较蛋白质组学的方法,一共得到 188 个在海 7124根系中受黄萎病菌侵染后与水处理的平行对照相比表
达量出现差异的蛋白,这些蛋白共同参与了棉花对黄
萎病菌入侵的响应。徐理[45]分别构建了抗黄萎病的海
岛棉海 7124 接种黄萎病菌后的抑制差减杂交 cDNA文库以及通过大规模测序的 RNA-Seq 技术研究棉花
抗黄萎病机制,通过基因表达分析,获得了大批棉花
抗黄萎病相关基因,并对其中的一个 WRKY 类转录
因子进行了较为详细的功能。孟宪鹏[46]克隆得到 2 个
海岛棉 ERF 转录因子基因,将其转化植物后对其抗病
相关功能进行研究,结果表明,在转基因烟草中,海
岛棉 ERF 转录因子可以增强烟草 PR 基因的表达;在
转基因棉花中,可以增强几丁质酶基因、β-1,3-葡聚糖
酶基因和 PR 基因等棉花抗病相关基因的表达。由此
推测,将海岛棉 ERF 基因转入棉花超表达后能够高效
激活下游抗病相关基因的表达。通过对已得到的转基
因棉花部分幼苗进行黄萎病菌侵染后发现:T0 转
EREB1 棉花幼苗的耐病能力高于对照。通过上述研究
可知:海岛棉 ERF 族基因的新成员 EREB1/2 在调控
抗病相关基因表达方面具有重要作用,EREB1/2 在转
基因烟草和棉花中超表达后可有效增强植株体内抗病
相关基因的表达,从而提高了转基因植株的抗病能力。
张文蔚[47]以陆地棉高抗黄萎病新品系中植棉 KV1 为
材料,接种大丽轮枝菌强致病力落叶型菌株 V991 后

17 期 郭三堆等:中国转基因棉花研发应用二十年 3377
0—96 h 棉株幼根总 RNA,构建正、反向抑制差减杂
交文库,筛选到 147 个信号强度差异明显的克隆,建
立了黄萎病菌侵染初期陆地棉的抗病相关基因表达
谱,并证明了 GhUbI1、GhEG 和 GhSCF 在陆地棉抗
黄萎病的过程中起重要作用。赵付安[48]采用 SSH 技术
从转录组水平探讨了陆地棉基因的差异表达,采用
2-DE 技术从蛋白质组水平探讨了栽培棉近缘野生种
瑟伯氏棉的蛋白差异表达,并以此为基础克隆了 2 个
陆地棉结构抗性基因 GhDIR 和 GhSUMO,克隆了瑟
伯氏棉根的GPIP,克隆了瑟伯氏棉根的4个TIR-NBS- LRR 类抗病基因类似物,这七个基因的克隆为进一步
研究其抗病功能奠定了基础。倪萌等[49]采用叶片针刺
接种法从细胞学方面分析转 hpa1xoo 棉花与棉花黄萎
病菌互作产生的微过敏抗病反应,通过细胞显微观察
表明,转 hpa1xoo 棉花 T-34 与非转基因棉花在抗病性
表型方面存在明显差异,转 hpa1xoo 棉花较非转基因
棉花有较强的抗病性。 1.5 纤维品质改良转基因棉花
棉纤维是棉花产量形成的主要部分,其品质决定
经济价值。在生产实践中,高产棉花不优质、优质棉
花不高产是限制棉花种植业发展的主要因素之一。随
着纺织工业的不断发展,对棉花纤维品质不断提出新
的要求,传统的遗传育种技术已经不能解决生产实践
中关于棉花产量和品质之间存在的矛盾。利用基因工
程技术将棉花纤维发育相关基因导入棉花,提高棉花
纤维产量和品质,成为当前棉花增产和品质改良的主
要途径。同时,随着高通量基因克隆和表达分析技术
的不断发展,大量与棉花纤维产量和品质相关基因和
表达调控元件得到了验证,并初步应用于棉花。 据报道,棉花纤维增产基因主要与棉花种子发育
和 IAA 等激素代谢相关。李德谋等[50]将 GhASN-Like导入棉花,提高了转基因植株的单株成铃数和单铃种
子数,增加了籽棉和皮棉的产量。裴炎等[51]利用棉花
纤维细胞特异启动子 FBP7,驱动 IAA 合成基因 iaaM在棉花中表达。使在棉纤维发育起始阶段,胚珠表面
的 IAA 含量迅速增高,增加了能够发育成纤维的细胞
数量。经过连续 4 年测产,转基因棉花纤维产量比对
照提高了 15%以上[52]。 棉花纤维品质与纺织工业的应用密切相关,主要
涉及了棉纤维长度、细度、强度、色泽等指标。目前,
利用基因工程技术将调控棉花纤维发育的相关基因导
入棉花来提高棉花纤维品质,已经取得了初步的进展。
李文彬等[53]利用棉花纤维特异启动子驱动吲哚双加
氧酶(bec)基因在棉花中表达,使转基因棉花纤维获
得了特殊的颜色。左开井等[54]将 GbAnn9 导入棉花,
转基因棉花纤维长度比对照材料增加 2—3.5 mm,纤
维强度增加 0.3—3.3CN/tex。夏桂先等[55]将棉花苯丙
烷类化合物木质素合成基因 GhLIN2 导入棉花,增加
转基因棉花纤维内木质素含量,使棉花纤维长度、细
度和强度均得到显著提高。李晓荣等[56]利用 35S 启动
子驱动棉花尿甘二磷酸葡萄糖焦磷酸化酶基因
GhUGP1 在棉花中表达,获得的转基因棉花材料纤维
长度比对照增加 18.5%,断裂比强度增加 31.85%。 此外,通过对棉花纤维细胞发育机理的简析,分
离获得了多个与棉花纤维品质相关的候选基因。刘进
元等[57]克隆获得棉花纤维可逆糖基化蛋白酶基因,该
基因可以通过控制棉纤维中多糖物质的积累,进一步
提高棉纤维的品质。李学宝等[58]克隆获得棉花纤维特
异表达的水孔蛋白家族基因 GhPIP2;6,其转录产物主
要在伸长期的棉花纤维中高表达,通过在酵母中表达
研究发现可以使细胞长度增加 1.3—1.47 倍。朱玉贤
等[59]克隆获得与棉花纤维伸长密切相关的 β微管蛋白
基因,该基因在棉花纤维伸长期高表达与棉纤维长度
发育密切相关。 为了能够实现棉花纤维发育相关基因的精确定位
表达,在棉纤维特异表达启动子方面的研究也取得了
重要的进展。郭三堆等[60]分离获得了一个在棉花生殖
器官优势表达的启动子 Arf1,该启动子为棉花腺苷酸
核糖基化作用因子基因的启动子。与报告基因 GUS融合后,利用花粉管通道法转化棉花,报告基因在转
基因棉花的生殖器官高表达。张献龙等[61]克隆获得了
棉花纤维特高效表达的类成束蛋白样阿拉伯半乳聚糖
蛋白基因(GhFLA1)启动子,该启动子能够驱动 GUS报告基因在棉花纤维发育 0—20 d 高表达。 1.6 其他性状如早衰、耐涝以及特殊用途等转基因棉
花
目前,生产应用的高产、优质棉花品种均具有早
衰的特征,限制了高产、优质棉花品种的推广应用。
利用基因工程技术延缓棉花的生长发育过程中的早衰
现象,对具有早衰特性的棉花高产品种的培育和推广
应用具有重要意义。李静等[62]克隆获得了异戊烯基转
移酶基因 ipt,该基因编码的蛋白是细胞分裂素生物合
成途径中的关键酶。将该基因导入早衰型陆地棉品种
中棉所 10 号中,通过对转基因棉花进行叶绿素和细胞
分裂素含量的测定及形态观察,发现转基因棉花的早
衰性状得到延迟。

3378 中 国 农 业 科 学 48 卷
在中国南方湿度大或者雨水多的地方,由于棉花
不耐湿和涝的特性,雨季来临时会引起棉花大量落铃
导致减产,限制了棉花种植业的发展。利用基因工程
技术将耐涝基因导入棉花,培育耐涝棉花新材料,不
仅可以保证棉花稳产高产,而且可以通过将其他地区
不耐涝的棉花培育为耐涝材料,为遗传育种提供更加
丰富的资源材料。中国农业科学院生物技术研究所作
物分子育种课题组将来源于透明颤菌血红蛋白基因
(vgb)经过密码子优化设计后导入棉花,培育出高耐
涝的转基因棉花新材料,该材料在地下部分封闭的涝
池内表现良好,比对照材料增产 20%以上[63]。 上述不同类型转基因棉花的获得,极大地丰富了
中国的棉花种质资源,促进了中国棉花的育种研究。
2 棉花的遗传转化方法
遗传转化方法是进行基因功能分析和改良作物性
状的必要手段。主要的植物遗传转化技术可分为两大
类:第一类是直接基因转移技术,包括基因枪法、原
生质体法、脂质体法、花粉管通道法、电激转化法、
PEG 介导转化方法等,以基因枪转化法和花粉管通道
法为代表。第二类是生物介导的转化方法,主要有农
杆菌介导和病毒介导等转化方法,其中农杆菌介导的
转化方法操作简便、成本低、转化率高,广泛应用于
单、双子叶植物的遗传转化。随着科学技术的不断发
展,近年来,逐渐发展了一些新的转化技术,如茎尖
或芽尖转化法,农杆菌菌液浸染法以及纳米载体花粉
介导法等。这些转化方法的兴起及广泛应用对转基因
育种具有重要意义,极大地加快了农作物改良以及新
品种选育的步伐。 2.1 农杆菌介导转化法
农杆菌介导法是 早应用、 实用有效并且具有
很多成功实例的一种植物转基因方法。农杆菌是一类
存在于土壤中的革兰氏阴性细菌。目前,用于植物转
基因介导的农杆菌为根癌农杆菌。根癌农杆菌含有一
种 Ti 质粒,Ti 质粒上有一 DNA 片段,称为转移 DNA。
农杆菌侵染植物时,细菌通过植物伤口进入寄主组织,
但细菌本身不进入寄主植物细胞,只是把 Ti 质粒的
T-DNA 片段导入植物的基因组中,并得以表达,从而
导致冠瘿瘤的发生,并且导入的 T-DNA 在转化植株
后代中通常表现出典型的孟德尔遗传规律。由于 Ti质粒本身能插入大约 50 kb 的 DNA 外源基因,因此,
利用这种转化载体,将 Ti 质粒上致瘤的有害基因切
除,代之以有益的外源基因,使得向植物细胞引入外
源基因已成为植物基因工程研究的一项常规技术。棉
花农杆菌介导转化一般是采用叶盘法。国外 早利用
此法的是 Agracetus 公司,将 NPTII 和 CAT 导入柯字
棉 312 中获得转基因再生棉株。国内 早的是山西省
农业科学院棉花研究所生物技术研究室陈志贤等,将
除草剂基因 Tfda 导入晋棉 7 号获得转基因再生棉株。
目前,中国农业科学院生物研究所、中国科学院遗传
与发育生物学研究所、中国科学院微生物研究所与各
地方农业科学院合作,利用农杆菌介导法将抗棉铃虫
和抗蚜虫的单价、双价及三价等目的基因导入棉花,
获得单价、双价和三价转基因抗虫棉花新品种。 2.2 基因枪轰击转化法
基因枪是 20 世纪 80 年代初由 Sanford 等发明。
该技术原理是:利用基因枪产生的高压动力冲击波将
包裹外源 DNA 的重金属颗粒(如钨粉、金粉等)射
穿植物细胞壁和细胞膜,射入植物细胞,是外源 DNA随机整合到植物细胞染色体中,达到外源 DNA 在受
体植物中正常表达和稳定遗传的目的。经过 10 多年的
改进和提高,基因枪技术已成功应用在烟草、水稻、
小麦、棉花、大豆等许多农作物的品种改良上,并且
该技术被用于瞬时表达研究和培育稳定的转基因植物
等研究领域[64-67]。 2.3 花粉管通道介导转化法
20 世纪 70 年代末期,在 DNA 片段杂交假设理论
和对植物开花受精过程的解剖学及细胞学特征研究的
基础上,中国科学家周光宇等推测外源 DNA 可以通
过花粉管经过的珠心通道进入受精胚囊,转化进入精
卵融合细胞、早期合子及早期胚细胞。随后,建立花
粉管通道技术并通过该技术将外源DNA导入陆地棉,
成功培育出抗枯萎病的新品种。1993 年,中国农业科
学院生物技术研究所与江苏农业科学院经济作物研究
所合作,利用花粉管通道法在国内首次将 GFM Cry1A杀虫蛋白基因导入棉花,培育出转基因抗虫棉花植株。
花粉管通道法具有操作简单、对受体植物以及外源
DNA 无特别要求、无组织培养过程、转化速度较快、
育种周期较短等特点,目前,该方法已成功应用于棉
花、水稻、小麦、大豆等农作物的改良和育种工作[68-71]。 2.4 茎尖或芽尖转化法
传统的农杆菌介导法受体主要是幼胚和胚性愈伤
组织,受体的基因型决定着遗传转化的成功与否;而
愈伤组织形成的再生植株具有无性系变异大、周期长
等不利因素。因此,选择合适的外植体作为农杆菌转
化的受体、建立不受基因型限制、快速而又高效的遗

17 期 郭三堆等:中国转基因棉花研发应用二十年 3379
传转化体系是转基因生物技术研究者的目标。茎尖培
养是植物组织的常规技术,已被广泛应用。目前,幼
胚仍然是转化受体系统中 重要的外植体。而由成熟
胚茎尖分生组织诱导而来的丛生芽与传统的幼胚相
比,具有不受季节限制,诱导率高,再生植株变异少
等优势。通过改善茎尖的遗传转化,利用农杆菌介导
法转化棉花、玉米等农作物的茎尖,缩短转化周期,
从而获得阳性转基因植物的方法称为茎尖/芽尖转化
法。与传统的幼胚及其胚性愈伤组织为受体的遗传转
化体系相比,此种方法具有简单、快速、工作效率高
和试验周期短等优点;与基因枪法相比更加经济适用;
与合子转化相比,取材不受季节限制。但该方法的缺
点是获得的转基因后代嵌合体较多,得到纯合稳定的
转基因品系困难较大[72]。 2.5 农杆菌液浸染法
大多数的转化方法都需要组织培养技术,通过组
织培养技术进行单细胞的选择并再生出整个植株虽然
减少了嵌合体的出现,但是由于后生影响和染色体重
排就会出现体细胞无性系变异。利用农杆菌液浸染植
物组织(多数研究都采用花器官)的方法进行转基因,
其特点是避免了组织培养和再生过程。前人的研究表
明,浸花得到的 T0种子是典型的杂合子,在同一插入
位点只有 2 个等位基因中的一个[73-74]。大量的结果表
明转化发生在花发育的后期[75]。尽管浸花的方法有较
高的转化率,但对于转化的细胞和转化的时间仍然不
太清楚。 2.6 纳米载体花粉介导转化法
随着 20 世纪末纳米科技的兴起,纳米材料以其特
殊的表面效应、小尺寸效应、量子尺寸效应、宏观量
子隧道效应及良好的生物学特性,在生物学领域得到
了成功地应用,尤其为制备高效、靶向的基因载体系
统提供了良好的介质。自从 20 世纪 60 年代末发现二乙
基氨乙基葡聚糖/DNA 复合物能介导基因传递后[76],纳
米材料作为基因载体的研究不断深入。 纳米基因载体在动物和人的细胞转染试验中已经
取得了成功[77-78]。并显示出巨大的应用前景。纳米载
体在植物的遗传转化方面与裸 DNA 分子相比,具有
更高的转导率和基因表达率。日本大阪大学的
Takefumi 等[79]以钙盐粒子微球为基因载体,将质粒
DNA 囊括在微球内部,对 BY-2 烟草细胞原生质体进
行转染,试验结果显示其转染率是裸 DNA 分子转染
效率的 10 倍。美国爱荷华州立大学的 Torney 等[76]利
用蜂窝状介孔二氧化硅纳米粒子为载体,装载基因及
刺激该基因表达的化学物质,使用金纳米粒子覆盖
介空表面,穿过植物细胞壁,同时将两者置入植物
细胞中,并控制在适当的时间和地点释放,成功获
得转基因植株。采用纳米载体基因技术改良花粉管
通道法,有可能大幅度地提高外源基因在花粉管通
道法中的传输、保护外源基因在传输和转导过程中
免受 DNA 酶的降低,提高遗传转化效率,为棉花等
转基因作物新品种培育提供了更为简便、高效的分
子育种新技术。 目前,虽然纳米载体在植物遗传转化上的应用时
间还较短,仍存在许多尚待解决的问题,但与其他植
物基因转化载体相比,纳米载体技术具备生物相容性
好、稳定性高、具有浓缩及保护 DNA 的作用、装载
容量较大、优良的表面性质、无遗传毒性和免疫原性、
制备简单等方面的优势,并且通过不同材料和工艺,
能制备出各种各样具有“特异功能”的载体。这种材
料并不昂贵,能使纳米载体更具市场竞争力,因此,
纳米载体转基因技术必将成为今后植物转基因技术的
有效途径之一。
3 转基因植物安全性评价
一系列具有优良性状(如抗虫、抗除草剂等)的
转基因作物新品种的大面积推广种植在带来直接经济
与生态效益的同时,也为解决未来世界粮食、能源危
机提供了新的途径。但同时,转基因技术也存在一定
的风险。近年来,转基因生物安全问题在全世界范围
内引起了广泛关注。目前对转基因植物的安全性评价
主要集中在 2 个方面,一是环境安全性,另一是食品
安全性。 3.1 转基因植物的环境安全性
转基因植物的环境安全性主要涉及 3 个方面:(1)转基因植物本身可能演变为农田杂草。植物在获得某
种特定基因(如抗病、抗虫、抗除草剂和抗逆等),
可能会增强其生存竞争性,在生长势、越冬性、耐受
性、种子产量等方面强于非转基因植物。若被推广种
植,释放到自然环境中的机会特别大。因其具有野生
植物没有的各种抗性,将会迅速地成为新的优势种群,
进而演变成农田杂草[80]。(2)基因漂流影响其他物
种。在自然条件下,栽培作物和栽培作物之间、栽培
作物与其近缘野生种之间、栽培作物和杂草之间都可
能存在种间基因漂流能力。因此,转基因植物中的
一些抗除草剂、杀虫剂和病毒的抗性基因就有可能
通过花粉杂交等途径向其同种或近缘野生种转移,

3380 中 国 农 业 科 学 48 卷
从而产生出一些可抗除草剂、杀虫剂和病毒的“超
级杂草”[81],造成不可估量的农业损失和生态灾难。
(3)影响生物的多样性。植物在引入抗虫或抗病基因
后,往往具有较强的“选择优势”,大量的转基因植
物进入自然生态系统中,可能会淘汰原来栖息地上的
物种及其他遗传资源,加剧品种的单一化,造成生物
数量剧减,甚至会使原有物种灭绝。如转入 BT 杀虫
基因的抗虫棉,其目标昆虫是棉铃虫和红铃虫等植物
害虫,如大面积和长期种植抗虫棉,昆虫有可能对抗
虫棉产生适应性或抗性,这会影响抗虫棉的应用和 BT农药制剂的防虫效果。因此,在抗虫棉推广时一般要
求种植一定比例的非抗虫棉,以延缓昆虫产生抗性。
(4)可能对非目标生物造成危害,在植物基因工程中
所用的许多基因是与抗虫或抗病有关的,其直接作用
的对象是生物。其插入的抗虫或抗真菌的基因可能对
其他非目标生物起作用,从而杀死了环境中有益的昆
虫和真菌。美国康奈尔大学 Losey 等[82]发现,转基因
Bt 玉米花粉能导致非目标害虫黑麦金斑蝶(Danaus plexippus L.)幼虫死亡。 3.2 转基因植物的食品安全性
转基因食品又称基因修饰食品( genetically modified food,GMF),即用转基因生物制造或产生
的食品。进行转基因食品安全评价时,应从宿主、
载体、插入基因、重组 DNA、基因表达产物及其对
食品营养成分的影响等方面来考虑。评价内容主要
包括 4 个部分,第一部分是基本情况:包括供体与
受体生物的食用安全情况、基因操作、引入或修饰
性状和特性的叙述、实际插入或删除序列的资料、
目的基因与载体构建的图谱及其安全性、载体中插
入区域各片段的资料、转基因方法、插入序列表达
的资料等;第二部分是营养学评价:包括主要营养
成分和抗营养因子的分析;第三部分是毒理学评价:
包括急性毒性试验、亚慢性毒性试验等;第四部分
是过敏性评价,主要依据联合国粮农组织与世界卫
生组织提出的过敏源评价决策树依次评价,禁止转 入已知过敏源。
中国农业科学院生物技术研究所与江苏省农业科
学院经济作物研究所、山西省运城棉花研究所、石家
庄市农业科学院、中国农业科学院棉花研究所等多家
育种单位合作,将构建成功的含有自主知识产权的抗
虫基因植物表达载体采用花粉管通道技术和农杆菌介
导法进行遗传转化,经过后代分子鉴定、室内生测后,
育成了一批转基因抗虫棉品种,为以后国内转基因抗
虫棉品种的培育提供了特异的种质资源,也为国内抗
虫棉的迅猛发展提供了动力。 为了加强农业转基因生物安全管理,保障人类健
康和动植物、微生物安全,保护生态环境,促进农业转
基因生物技术研究,农业部陆续出台了多个条例、办法
和公告,来规范转基因抗虫棉的田间释放和产业化。 自 1996 年开始转基因抗虫棉安全性评价申报以
来,至 2013 年共计申报并通过生产应用的安全性评价
2 016 份,从图 1 中可以看出,随着安全性评价内容的
完善、管理规范化,从 2004 年农业部简化了转基因抗
虫棉安全评价的申报程序以后,获得的安全证书的数
量急剧上升,基本上每年审批的证书数量都在 150 份
以上(图 1)。 拥有中国基因知识产权的转基因抗虫棉安全性评
价共计 1 955 份,其中,转 Cry1Ab/cry1Ac 棉花 1 391
数量
Num
ber
图 1 2002—2013 年转基因抗虫棉获得生产应用的安全证书
Fig. 1 The safety certificate of transgenic insect-resistant cotton with the production in 2002-2013

17 期 郭三堆等:中国转基因棉花研发应用二十年 3381
份,占总安全性评价 2 016 份的 69.00%,转 Cry1Ab/ cry1Ac+CpTI 棉花 553 份,占 27.40%,转 Cry1Ac+API棉花 11 份,占 0.55%;美国孟山都公司申请并通过转
cry1Ac 抗虫棉安全评价 61 份,占 3.05%。 总之,自 1996 年国产抗虫棉进入田间试验,各省
棉花育种相关单位利用抗虫棉种质资源开始大规模以
来,中国抗虫棉的研究及产业化取得了迅速的发展,
国产抗虫棉的种植面积逐渐增加。国产转基因抗虫棉
的应用经历了近 20 年的风风雨雨,为中国棉花产业的
发展和改善生态环境作出了巨大贡献。而中国转基因
抗虫棉安全性评价为转基因抗虫棉的生产应用起到了
保驾护航的作用。 目前,转基因作物的安全性评价技术与政策保障
体系已经建立,今后还应该加大转基因棉花安全生产应
用保驾护航作用的力度,大力发展中国转基因植物分子
育种高新技术,为迅速占领高新技术制高点、提高中国
转基因植物分子育种的国际竞争力再作新贡献。
4 转基因抗虫棉的品种培育及推广
4.1 抗虫棉品种审定情况
中国审定的第一个转基因抗虫棉品种是美国岱字
棉公司的新棉 33B,该品种于 1997 年由河北省审定,
审定编号为冀审棉 97001 号,1997—2006 年美国岱字
棉公司与孟山都公司先后在河北省、河南省、山东省、
安徽省、湖北省和湖南省审定了 7 个转基因抗虫棉品
种,在 2003 年以前无论从品种数量还是种植面积都占
抗虫棉的统治地位。中国从 1998 年开始审定了第一批
转单价抗虫基因国产转基因抗虫棉品种,如:GK12、晋棉 26 和 GKZ1(国抗杂一号),1999 年审定了第
一个国审抗虫棉品种中棉所 38,从此迈开了中国转基
因棉花品种培育和生产推广的步伐[83]。截至到 2013年,中国共通过了 134 个国审转基因抗虫棉品种,黄
河流域审定 96 个,长江流域审定 36 个,西北内陆棉
区审定了 2 个,其中以生产需求量 大的中熟品种为
主,有生育期少于 110 d 的早熟品种 7 个,彩色棉品
种 1 个[84]。目前,中国抗虫棉已经形成了覆盖所有棉
区、不同熟期不同类型搭配合理、常规棉与杂交棉并
重的品种体系。 4.2 抗虫棉品种发展的趋势及存在的问题
4.2.1 杂交品种比例越来越大 1999—2013 年,134个国审品种中有 88 个杂交品种,2006 年以前只有 23个杂交品种,2007—2013 年共审定了 65 个杂交品种。
其中,长江流域 36 个品种中只有 1 个常规品种,杂交
品种有 35 个,从以上的数据可以看出,黄河流域棉区以
常规种和杂交种并重,长江流域棉区以杂交种为主[85]。 4.2.2 品种存在多、乱、杂现象 1996—2012 年,
转基因抗虫棉的品种数量在逐年增加,截至到 2012年,内陆棉区种植面积超过 0.33 万公顷的棉花品种品
系一共有 245 个,其中黄河流域有 166 个,长江流域
品种有 79 个[86],这里面有国审、省审品种,还有些
没有审定的品系。品种的多样性满足了不同中国气候
类型耕作制度的需求,但同时也存在品种同质化品种
多,突破性品种少的问题。因此造成了有些抗虫棉品
种审定后没有推广几年就被淘汰的现象,品种更替过
于频繁,造成人力物力的浪费。 4.2.3 品种的产量性状不断提升,但纤维品质性状
改进不大 随着抗虫棉品种的发展,棉花单产不断提
高,在中国棉花面积没有增加的情况下,产量有了大
幅度提高[87]。但是与纤维品质相关的性状改进不明
显,特别是棉纤维的长度和比强满足不了今后棉花机
采作业和纺织行业技术更新的需求。有待今后抗虫棉
育种单位进行选育方向的调整[88]。 4.3 转基因抗虫棉的产业化
4.3.1 抗虫棉的推广面积 从 1997 年转基因抗虫棉
开始大面积种植,主要是以美国的品种为主,当时占
国内抗虫棉市场份额的 95%以上,国产抗虫棉份额不
到 5%。随着中国自主研发的转基因抗虫棉逐渐成熟,
国产抗虫棉的品种数量和种植面积逐年提高,2001 年
转基因抗虫棉的面积首次超过了非转基因棉面积。之
后国产抗虫棉的数量和推广面积都迅猛增长,2002 年
国产抗虫棉占国内抗虫棉市场份额的 43.3%,2003 年
国产抗虫棉的面积超过了美国抗虫棉面积,占国内抗
虫棉市场份额的 53.9%,到 2007 年,国产抗虫棉面积
已占 96.1%,1998—2013 年国产抗虫棉累计推广面积
超过 3 000 万公顷,打破了美国抗虫棉的垄断,成功
地保护了中国棉花产业及相关产业和民族利益[89]。 4.3.2 抗虫棉产业化对中国棉种行业产生了深远的
影响 抗虫棉在中国市场零售价为 37.2 元/kg,改变了
非抗虫棉种 8.2 元/kg 的低价局面,为提升棉种质量,
改进棉种生产和销售方式提供了可能。由于棉种利润
的增加,催生了一批专业从事棉种或以棉种为主营业
务育繁推一体化的公司。在抗虫棉的品种选育方面也
逐渐由科研单位、大专院校为主向育繁推一体化的种
业公司和民营研究所转变。使得抗虫棉品种的选育更
贴近市场的需求,加快了品种选育和推广的步伐,使
中国棉种行业走上了一条良性循环的道路[90]。

3382 中 国 农 业 科 学 48 卷
随着国产抗虫棉研制技术的成熟,中国抗虫棉优
良的抗虫性和突出的综合性状引起了世界上各产棉国
的关注。中国先后与印度、澳大利亚和巴基斯坦签订
了抗虫棉的合作项目,目前,项目正在进行中。国产
抗虫棉已经走出国门,迈向世界,参与了国际竞争[91]。
5 中国转基因棉花研究方向的展望
如上所述,随着转基因抗虫棉的品种类型日渐丰
富、允许安全应用的区域逐步放开,近年来,转基因
抗虫棉的产业化有了长足的发展,取得了良好的经济
和社会效益。但是,在研究方面,仍然存在一些亟待
解决的问题,也是中国转基因棉花的未来研究方向。 5.1 运用多个抗虫基因提高棉花的综合抗虫能力
抗虫棉从开始应用到现在已经近 20 年的时间。长
期种植转基因单价、双价抗虫棉,虽有效防治了棉铃
虫的发生,但由于农药施用量大大减少,一些次要害
虫,如盲蝽象、蚜虫等的危害日益突出,上升成为棉
花主要害虫;同时,抗虫棉本身也存在生长发育后期
抗虫性下降、可能发生抗性棉铃虫等综合问题,所以
需要加大力度寻找新型有效的抗虫基因资源,研发转
新型双基因、多基因的抗虫棉,以提高转基因抗虫棉
的综合抗虫能力。 5.2 培育自主产权的抗除草剂棉花
随着农村劳动力向城市转移以及农业可持续发展
的需要,全生育期利用除草剂防除杂草成为机械化植
棉的重要需求。针对不同棉区的杂草种群类型和除草
剂使用偏好,尤其针对高效、低毒、低残留的除草剂
类型,积极筛选新的有利用价值的抗除草剂基因,研
发具有中国自主知识产权的抗除草剂棉花。 5.3 培育耐旱耐盐碱转基因棉花
中国人多地少是无法解决的刚性难题,要满足棉
花原料的供应问题,利用中国现有的 2 000 多万公顷
的干旱、盐碱地,将其转变成棉田,扩大植棉面积、增
加原棉总量是解决棉花原料供应问题的重要途径。伴随
着植物耐逆分子通路的解析和对极端生境微生物(如盐
湖微生物等耐逆机理)的研究,耐旱、耐盐碱的基因资
源正在逐步被挖掘和应用于转基因棉花育种。 5.4 培育抗病转基因棉花
枯、黄萎病一直是影响中国棉花生产的主要病害。
随着对枯、黄萎病遗传规律和抗性机理的进一步揭示,
以及抗性相关基因 QTL 定位和基因克隆,为转基因抗
病棉花育种提供了抗性基因资源。同时,由于致病生
理小种的复杂性,还需广泛搜集和挖掘利用野生棉及
棉属近缘物种中高抗枯、黄萎病的基因,并将分子标
记育种、转基因育种与常规育种相结合,利用综合育
种技术聚合多种抗性基因,加快棉花抗病育种的进程,
培育出高抗病的转基因棉花新品种。 5.5 培育优质转基因棉花
受国际市场的影响,棉纺工业正处在一个由低端
产品向中高端产品过渡的转型期。而要提高中国棉纺
企业的国际竞争力,首当其冲的就是要解决优质棉原
料的供应问题。因此,满足纺织企业需求的优质棉生
产必将成为中国棉花生产发展的主要方向。随着棉花
功能基因组学研究的发展,棉纤维的发生、发育机制
正在逐步明晰,一批对棉纤维的品质起决定作用的基
因正在被挖掘和应用于转基因优质棉花的培育。 现代棉花种植方式对棉花品种的要求更加多样,
运用转基因生物育种技术,改善棉花的抗虫、抗除草
剂、耐逆、抗病和优质等性状,并让这些有利于棉花
生产的性状在较短的时间内聚合,才能满足今后棉花
生产的需要。因此,综合利用转基因植物生物育种技
术,培育高产、优质、抗逆的突破性新品种,是中国
转基因棉花育种发展的根本。 总之,中国转基因棉花 20 多年来取得了骄人的成
绩。特别是国产转基因抗虫棉的研发和产业化的成功,
不仅促进了中国棉花及相关产业的发展,打破了美国
的垄断,提升了国际竞争力,它推广应用的速度之快,
影响力之广,所产生的经济社会效益和生态效益之大,
创造了中国近代农业科学发展史上的奇迹。
References
[1] 转基因权威关注: http://www.moa.gov.cn/ztzl/zjyqwgz/spxx/.
GM authority concerned: http://www.moa.gov.cn/ztzl/zjyqwgz/spxx/.
(in Chinese)
[2] 郭三堆, 倪万潮, 徐琼芳. 编码杀虫蛋白融合基因和表达载体及其
应用: 中国, ZL95119563.8.
Guo S D, Ni W C, Xu Q F. Encoding insecticidal protein fusion gene
and expression vectors and its application: China, ZL95119563.8. (in
Chinese)
[3] 郭三堆, 崔洪志, 倪万潮. 两种编码杀虫蛋白质基因和双价融合表
达载体及其应用:中国, ZL98102885.3.
Guo S D, Cui H Z, Ni W C. Two genes encoding insecticidal proteins
and double fusion expression vector and itsapplication: China,
ZL98102885.3. (in Chinese)
[4] 崔金杰, 雒瑶瑜, 王春义, 马艳, 李春花. 转双价基因棉田主要害
虫及其天敌的种群动态. 棉花学报, 2004, 16(2): 94-101.

17 期 郭三堆等:中国转基因棉花研发应用二十年 3383
Cui J J, Luo J Y, Wang C Y, Ma Y, Li C H. Population dynamics of
main pests and enemies in the transgenic Cry1Ac+ CpTI cotton field.
Acta Gossypii Sinica, 2004, 16(2): 94-101. (in Chinese)
[5] 王伟, 朱祯, 高越峰, 石春林, 陈宛新, 郭仲琛, 李向辉. 双价抗虫
基因陆地棉转化植株的获得. 植物学报, 1999, 41(4): 384-388.
Wang W, Zhu Z, Gao Y F, Shi C L, Chen W X, Guo Z C, Li X H.
Obtaining a transgenic upland cotton harboring two insecticidal genes.
Acta Botanica Sinica, 1999, 41(4): 384-388. (in Chinese)
[6] 刘志, 郭旺珍, 朱协飞, 朱祯, 张天真. 转 Bt+GNA 双价基因抗虫
棉花的抗虫性表现特征. 高技术通讯, 2003(1): 37-41.
Liu Z, Guo W Z, Zhu X F, Zhu Z, Zhang T Z. Go Bt+GNA of bivalent
insect-resistant gene zoophobous cotton features. High Technology
Letters, 2003(1): 37-41. (in Chinese)
[7] 吴家和, 田颖川, 罗晓丽, 郭洪年, 石跃进, 陈晓英, 贾燕涛, 肖娟
丽, 张献龙. 转两类抗虫基因棉花优良纯合品系的选育. 中国农业
科学, 2003, 36(6): 651-656.
Wu J H, Tian Y C, Luo X L, Guo H N, Shi Y J, Chen X Y, Jia Y T,
Xiao J L, Zhang X L. Selection of homozygous cotton lines
transformed with two types of insect-resistant genes. Scientia
Agricultura Sinica, 2003, 36(6): 651-656. (in Chinese)
[8] 肖松华, 吴巧娟, 刘剑光, 柏立新, 束春娥, 肖留斌. 转野生荠菜
凝集素基因棉花对棉蚜的抗性鉴定. 江西农业学报, 2006, 18(5):
46-50.
Xiao S H, Wu Q J, Liu J G, Bai L X, Shu C E, Xiao L B. Identification
on resistance of trans- wild shepherd’s purse agglutinin gene cotton
(Gossypium hirsutum) to Aphis (Aphis gossypii). Acta Agriculturae
Jiangxi, 2006, 18(5): 46-50. (in Chinese)
[9] 张林水, 朱祯, 吴霞, 姜艳丽, 徐鸿林, 上官小霞, 李波, 李燕娥.
转三价抗虫基因彩色棉的获得 . 西北植物学报 , 2005, 25(6):
1126-1131.
Zhang L S, Zhu Z, Wu X, Jiang Y L, Xu H L, Shangguan X X, Li B,
Li Y E. Acquisition of transgenic colour cotton with a trivalent
insect-resistant gene. Acta Botanica Boreali-Occidentalia Sinica, 2005,
25(6): 1126-1131. (in Chinese)
[10] 雒瑶瑜, 崔金杰, 王春义, 辛惠江. 转 Cry1Ac+Cry2ab 基因棉花对
小地老虎生长发育的影响. 棉花学报, 2014, 26(5): 179-183.
Luo J Y, Cui J J, Wang C Y, Xin H J. Effects of transgenic Cry1Ac
plus Cry2Ab cotton on blank cutworm (Agrotis ypsilon Rottemberg)
growth and development. Cotton Science, 2014, 26(5): 179-183. (in
Chinese)
[11] McGaughey W H, Whalon M E. Managing insect resistance to
Bacillus thuringiensis toxins. Science, 1992, 258: 1451-1455.
[12] 刘超, 朱彦卓, 代文俊, 任燕萍. 转基因棉花抗逆性研究进展. 湖
南农业科学, 2014(15): 32-35.
Liu C, Zhu Y Z, Dai W J, Ren Y P. Advances in stress resistance
research on transgenic cotton. Hunan Agricultural Sciences, 2014(15):
32-35. (in Chinese)
[13] 吕素莲. 转 betA和TsVP基因提高棉花耐盐、抗旱性的研究[D]. 济
南: 山东大学, 2007.
Lü S L. Go beta and TsVP gene cotton resistant to salt and drought
resistance research [D]. Jinan: Shandong University, 2007. (in Chinese)
[14] 曹燕燕, 廖平安, 葛昌斌, 沈向磊, 杨业华. 转 rolB 基因棉花抗旱
性研究. 湖南农业科学, 2012(15): 1-2, 5.
Cao Y Y, Liao P A, Ge C B, Shen X L, Yang Y H. Drought resistance
of rolB transgenic cotton. Hunan Agricultural Sciences, 2012(15): 1-2,
5. (in Chinese)
[15] 王娟. 转 ZmPIS 基因及聚合 betA/TsVP 基因提高棉花耐旱性的研
究[D]. 济南: 山东大学, 2010.
Wang J. Go ZmPIS betA/TsVP gene and polymerization to improve
drought tolerance of cotton research [D]. Jinan: Shandong University,
2010. (in Chinese)
[16] 吴伟. 转 ZmPLCI-betA 基因棉花的耐旱性研究[D]. 济南: 山东大
学, 2012.
Wu W.Study on drought tolerance of transgenic cotton with ZmPLCI-
betA gene[D]. Jinan: Shandong University, 2012. (in Chinese)
[17] 杨云尧. 转 MVP5CS 和 MvNHX1 基因提髙棉花抗旱性的研究[D].
乌鲁木齐: 新疆农业大学, 2012.
Yang Y Y. Go MVP5CS and MvNHX1 genes of higher drought
resistance of cotton research [D]. Urumqi: Xinjiang Agricultural
University, 2012. (in Chinese)
[18] 蔡永智. 转 CBF1 和 KatG 基因棉花抗旱性分析[D]. 石河子: 石河
子大学, 2013.
Cai Y Z. CBF1 KatG gene and analysis of drought resistance of cotton
[D]. Shihezi: Shihezi University, 2013. (in Chinese)
[19] 钱进, 杨云尧, 任燕萍, 朱超, 陈全家, 张桦. 转 MvNHX1 基因棉
花抗旱性研究和株系变异分析. 分子植物育种, 2014, 12(1): 80-86.
Qian J, Yang Y Y, Ren Y P, Zhu C, Chen Q J, Zhang H. The study on
drought resistance and variation analysis in transgenic MvNHX1
cotton. Molecular Plant Breeding, 2014, 12(1): 80-86. (in Chinese)
[20] Gu L, Zhang H, Guo H, Cheng H. Cloning of Ammopiptanthus
mongolicus C-repeat-binding factor gene and its cold-induced
tolerance in transgenic tobacco. Brazilian Archives of Biology and
Technology, 2013, 56(6): 895-903.
[21] 张慧军, 董合忠, 石跃进, 陈受宜, 朱永红. 山菠菜胆碱单加氧酶
基因对棉花的遗传转化和耐盐性表达. 作物学报, 2007, 33(7):
1073-1078.

3384 中 国 农 业 科 学 48 卷
Zhang H J, Dong H Z, Shi Y J, Chen S Y, Zhu Y. Hong transformation
of cotton (Gossypium hirsutum L.) with AhCMO gene and the
expression of salinity tolerance. Acta Agronomica Sinica, 2007, 33(7):
1073-1078. (in Chinese)
[22] 连丽君. betA 基因的异源表达提高了棉花耐盐和耐低温能力[D].
济南: 山东大学, 2008.
Lian L J. Heterologous expression of beta gene improves salt
tolerance in cotton and low temperature-resistant ability [D]. Jinan:
Shandong University, 2008. (in Chinese)
[23] 樊文菊. 转 ZmPLC1 基因及聚合 ZmPLC1/betA 基因棉花耐盐性的
研究[D]. 济南: 山东大学, 2013.
Fan W J. ZmPLC1 gene and the ZmPLC1/beta study on salt-tolerance
of transgenic cotton [D]. Jinan: Shandong University, 2013. (in
Chinese)
[24] 陈翠霞, 于元杰, 刘风珍, 沈法富, 王洪刚. 棉花耐盐变异体的遗
传分析. 西北植物学报, 2000, 20(2): 234-237.
Chen C X, Yu Y J, Liu F Z, Shen F F, Wang H G. Genetic analysis of
salt-tolerance variant in cotton. Acta Botanica Boreali-Occidentalia
Sinica, 2000, 20(2): 234-237. (in Chinese)
[25] 李雪林. SNAC1 基因在棉花上的应用及盐胁迫下棉花基因组 DNA
甲基化分析[D]. 武汉: 华中农业大学, 2009.
Li X L. Application of SNAC1 in cotton and cotton under salt stress
genome-wide DNA methylation analysis [D]. Wuhan: Huazhong
Agricultural University, 2009. (in Chinese)
[26] 张锐, 郭三堆, 梁成真. 三个棉花 ABF/AREB/ABI5/DPBF 类转录
因子及其编码基因与应用: 中国, ZL200910158311.X.
Zhang R, Guo S D, Liang C Z. Coding genes of three cotton
transcription factors ABF/AREB/ABI5/DPBF and its applications:
China, ZL200910158311.X. (in Chinese)
[27] 王宗文 . 国内外抗除草剂棉花研究应用现状 . 山东农业科学 ,
2011(1): 81-85.
Wang Z W. Research and application status of herbicide- resistant
cotton at home and abroad. Shandong Agricultural Sciences, 2011(1):
81-85. (in Chinese)
[28] 雷凯健, 卢维维, 洪玉枝, 刘子铎. 土壤总 DNA 中草甘膦 N-乙酸
转移酶基因的克隆及酶学特性分析. 农业生物技术学报, 2007,
15(1): 153-156.
Lei K J, Lu W W, Hong Y Z, Liu Z D. Cloning of the glyphosate
N-acetyltransferase gene from soil total DNA and its characterization
of enzyme activity. Journal of Agricultural Biotechnology, 2007,
15(1): 153-156. (in Chinese)
[29] 何鸣, 曾海燕, 徐培林, 叶长明. aroA 基因的克隆和优化. 中山大
学学报: 自然科学版, 2002, 41(2): 76-79.
He M, Zeng H Y, Xu P L, Ye C M. Cloning and mutagenesis of the
aroA gene. Acta Scientiarum Naturalium Universitatis Sunyatseni,
2002, 41(2): 76-79. (in Chinese)
[30] 赵特. 一种抗草甘麟基因的发现和抗草甘麟转基因水稻的培育[D].
杭州: 浙江大学, 2008.
Zhao T. An resistant glyphosate gene discovery and the cultivation of
transgenic rice [D]. Hangzhou: Zhejiang University, 2008. (in Chinese)
[31] 朱玉. 极端污染环境下草甘膦耐受菌株荧光假单胞菌G2的鉴定及
其 EPSP 合成酶墓因的克隆[D]. 北京: 中国农业科学院, 2002.
Zhu Y. Under extreme environmental pollution by glyphosate-tolerant
strains offluorescent Pseudomonas G2, identification and cloning of
EPSP synthase [D]. Beijing: Chinese Academy of Agricultural
Sciences, 2002. (in Chinese)
[32] 沙纪莹. 斯氏假单胞菌 A1501 EPSP 合酶基因的克隆及表达[D].
成都: 四川师范大学, 2008.
Sha J Y. Pseudomonas Stradivari A1501 EPSP synthase gene cloning
and expression [D]. Chengdu: Sichuan Normal University, 2008. (in
Chinese)
[33] 刘柱. 可变盐单胞菌中草甘膦抗性 EPSP 合酶新基因克隆、大肠杆
菌表达及其抗性机制的研究[D]. 成都: 四川大学, 2004.
Liu Z. Cloning a novel 5-enolpyruvylshikimate-3-phosphate synthase
gene conferring increased glyphosate tolerance from Halomonas
variabilis and its expression in Escherichia coli and its glyphosate-
tolerant mechanism [D]. Chengdu: Sichuan University, 2004. (in
Chinese)
[34] 童旭宏, 吴玉香, 祝水金. 陆地棉 epsps 基因的克隆及其组织特异
性表达分析. 棉花学报, 2009, 4: 259-264.
Tong X H, Wu Y X, Zhu S J. Cloning of 5-enolpyruvylshikimate
3-phosphate synthase genes EPSPS from the upland cotton and its
expression in different cotton tissues. Cotton Science, 2009, 4:
259-264. (in Chinese)
[35] 程海刚, 张锐, 罗淑萍, 代培红, 郭三堆. 青麻 ePsPs 基因在酵母
中表达. 新疆农业大学学报, 2007, 30(4): 52-56.
Cheng H G, Zhang R, Luo S P, Dai P H, Guo S D. Expression of
ePsPs gene from Abutilon theophrasti medic in the pichia pastoris.
Journal of Xinjiang Agricultural University, 2007, 30(4): 52-56. (in
Chinese)
[36] 郭三堆, 孙豹, 张锐, 孟志刚, 孙国清, 林敏, 陆伟. 一种含有草甘
膦抗性基因的表达载体及其应用: 中国, 201410204703.6.
Guo S D, Sun B, Zhang R, Meng Z G, Sun G Q, Lin M, Lu W. A
glyphosate resistance gene expression vector and its application:
China, 201410204703.6. (in Chinese)
[37] Guo S D, Sun B, Zhang R, Meng Z G, Sun G Q, Lin M, Lu W. A

17 期 郭三堆等:中国转基因棉花研发应用二十年 3385
glyphosate resistance gene expression vector and its application:
China, PCT/CN2014000620.
[38] 陈志贤, Danny J Liewellyn, 范云六, 李淑君, 郭三堆, 焦改丽, 赵
俊侠. 利用农杆菌介导法转移 tfdA 基因获得可遗传的抗 2, 4 -D 棉
株. 中国农业科学, 1994, 27(2): 31-37.
Chen Z X, Liewllyn D J, Fan Y L, Li S J, Guo S D, Jiao G L, Zhao J
X. 2,4-D resistant transgenic cotton plants produces by Agrobacterium-
mediated gene transfer. Scientia Agricutura Sinica, 1994, 27(2): 31-37.
(in Chinese)
[39] 刘方, 王坤波, 宋国立. 中国棉花转基因研究与应用. 棉花学报,
2002, 14(4): 249-253.
Liu F, Wang K B, Song G L. Application and advances in study on the
improvement of cotton transgene in China. Acta Gossypii Sinica, 2002,
14(4): 249-253. (in Chinese)
[40] 张磊. 水稻苯达松抗性基因Bel的精细定位与克隆[D]. 武汉: 华中
农业大学, 2006.
Zhang L. Rice bentazone fine mapping and cloning of resistance gene
Bel [D]. Wuhan: Huazhong Agricultural University, 2006. (in Chinese)
[41] 张金文, 熊春蓉, 李静雯, 王旺田, 孟亚雄, 陈正华. 臭鼻杆菌腈
水解酶基因(bxn)的克隆及其在原核生物中的表达 . 草业学报 ,
2006, 15(6): 87-92.
Zhang J W, Xiong C R, Li J W, Wang W T, Meng Y X, Chen Z H.
Cloning of bromoxyni-l specific nitrilase gene (bxn) from Klebsiella
ozaenae and expression in prokaryotic cells. Acta Prataculturae
Sinica, 2006, 15(6): 87-92. (in Chinese)
[42] 宋贵生, 冯德江, 魏晓丽, 唐家斌, 朱祯. 水稻乙酰乳酸合成酶基
因的克隆和功能分析. 中国农业科技导报, 2007, 9(3): 66-72.
Song G S, Feng D J, Wei X L, Tang J B, Zhu Z. Isolation and
functional analysis of rice acetolactate-synthase (ALS). Journal of
Agricultural Science and Technology, 2007, 9(3): 66-72. (in Chinese)
[43] 卢宗志, 张朝贤, 傅俊范, 李茂海, 李贵军. 抗苄嘧磺隆雨久花
ALs 基因突变研究. 中国农业科学, 2009, 42(10): 3516-3521.
Lu Z Z, Zhang C X, Fu J F, Li M H, Li G J. Molecular basis of
resistance to bensulfuron-methyl in Monochoria korsakowii. Scientia
Agricultura Sinica, 2009, 42(10): 3516-3521. (in Chinese)
[44] 高巍. 棉花响应黄萎病菌分子机制的蛋白质组学研究及 HDTF1 基
因的功能鉴定[D]. 武汉: 华中农业大学, 2014.
Gao W. Molecular mechanisms of response of cotton verticillium wilt
research onproteomics and functional identification of HDTF1 genes
[D]. Wuhan: Huazhong Agricultural University, 2014. (in Chinese)
[45] 徐理. 棉花与黄萎病菌的分子互作机制研究及 GbWRKY1 基因的
功能鉴定[D]. 武汉: 华中农业大学, 2011.
Xu L. Study on molecular interaction mechanism of verticillium
dahliae of cotton and the functional identification of GbWRKY1
genes [D]. Wuhan: Huazhong Agricultural University, 2011. (in
Chinese)
[46] 孟宪鹏. 海岛棉抗病相关 ERF 转录因子的克隆与鉴定[D]. 安阳:
中国农业科学院, 2009.
Meng X P. Cloning and identification of relevant ERF transcription
factors of Gossypium barbadense [D]. Anyang: Chinese Academy of
Agricultural Sciences, 2009. (in Chinese)
[47] 张文蔚. 陆地棉抗黄萎病相关基因筛选及功能验证[D]. 北京: 中
国农业科学院, 2013.
Zhang W W. Screening and functional verification of genes related to
resistance to Verticillium wilt in Gossypium hirsutum [D]. Beijing:
Chinese Academy of Agricultural Sciences, 2013. (in Chinese)
[48] 赵付安. 棉花黄萎病菌诱导陆地棉和野生棉差异表达分析及抗病
相关基因克隆[D]. 郑州: 河南大学, 2012.
Zhao F A. Differential expression analysis and cloning of disease
resistance-related gene among Wild cotton induced by verticillium
dahlia in cotton [D]. Zhengzhou: Henan University, 2012. (in Chinese)
[49] 倪萌, 缪卫国, 刘海洋, 邵家丽, 努尔孜亚, 乔子辰, 尚衍强, 王金
生. 棉花黄萎病菌诱导转 hpa1xoo 基因棉花产生微过敏防御反应.
新疆农业科学, 2009, 46(1): 46-49.
Ni M, Miao W G, Liu H Y, Shao J L, Nuerziya, Qiao Z C, Shang Y Q,
Wang J S. Micro- HR elicited by Verticillium dahliae kleb. in
transgenic hpa1xoo cotton. Xinjiang Agricultural Sciences, 2009,
46(1): 46-49. (in Chinese)
[50] 李德谋, 裴炎, 罗小英, 侯磊, 李先碧, 肖月华, 罗明, 白文钦, 张
觅, 李忠旺, 梁俊俊. GhASN-Like 基因、表达载体及其在提高棉花
产量中的应用: 中国, 201110104141.4.
Li D M, Pei Y, Luo X Y, Hou L, Li X B, Xiao Y H, Luo M, Bai W Q,
Zhang M, Li Z W, Liang J J. GhASN-Like gene, expression vectors
and its application on cotton yield: China, 201110104141.4. (in Chinese)
[51] 裴炎, 侯磊, 李德谋, 宋水清, 李先碧, 罗明, 肖月华, 郑雪莲, 曾
其伟, 张觅, 邱坤, 罗凤涛. 表达生长素合成相关基因的植物表达
载体及其在棉花纤维性状改良的应用: 中国, CN200810142518.3.
Pei Y, Hou L, Li D M, Song S Q, Li X B, Luo M, Xiao Y H, Zheng X
L, Zeng Q W, Zhang M, Qiu K, Luo F T. Auxin biosynthesis related
genes expression in plant expression vector and itsapplication in
cotton fiber properties improvement: China, CN200810142518.3. (in
Chinese)
[52] Zhang M, Zheng X L, Song S Q, Zeng Q W, Hou L, Li D M, Zhao J,
Wei Y, Li X B, Luo M, Xiao Y H, Luo X Y, Zhang J F, Xiang C B, Pei
Y. Spatiotemporal manipulation of auxin biosynthesis in cotton ovule
epidermal cells enhances fiber yield and quality. Nature Biotechnology,

3386 中 国 农 业 科 学 48 卷
2011, 29(5): 453-459.
[53] 李文彬, 宋晶, 孙勇如, 张利明, 王义琴, 牛恒尧. bec 基因棉花特
异表达载体: 中国, 02103787.6.
Li W B, Song J, Sun Y R, Zhang L M, Wang Y Q, Niu H Y. BEC
cotton-specific expression vector: China, 02103787.6. (in Chinese)
[54] 左开井, 王劲, 陈继军. 提高棉花纤维品质的 GbAnn9 基因及其应
用: 中国, ZL201110253458.4.
Zuo K J, Wang J, Chen J J. GbAnn9 and application of cotton fiber
quality: China, ZL201110253458.4. (in Chinese)
[55] 夏桂先, 韩立波, 焦改丽, 李元宝, 王海云. GhLIN2 蛋白及其基因
在改良棉花纤维品质中的应用: 中国, 201210505575.X.
Xia G X, Han L B, Jiao G L, Li Y B, Wang H Y. GhLIN2 protein and
its gene in the modified application of cotton fiber quality: China,
201210505575.X. (in Chinese)
[56] 李晓荣, 秦咏梅, 李晓东, 范玲, 李波, 胡文冉, 柳皋隽, 杨洋.
GhUGP1 基因在改良棉花纤维品质中的应用方法 : 中国 ,
201310613879.2.
Li X R, Qin Y M, Li X D, Fan L, Li B, Hu W R, Liu G J, Yang Y.
Application of GhUGP1 in improving cotton fibre quality: China,
201310613879.2. (in Chinese)
[57] 刘进元, 赵广荣, 徐振彪. 棉花纤维可逆糖基化蛋白酶基因及其编
码的蛋白质: 中国, CN01142164.9.
Liu J Y, Zhao G R, Xu Z B. Cotton fiber reversible effect of protease
gene and its encoded protein: China, CN01142164.9. (in Chinese)
[58] 李学宝, 李登弟, 王秀兰, 阮想梅, 吴雅洁. 棉花纤维特异表达的
水孔蛋白家族基因 GhPIP2;6: 中国, ZL 200910273363.1.
Li X B, Li D D, Wang X L, Ruan X M, Wu Y J. Specific expression of
Aquaporin family of cotton fiber gene GhPIP2;6: China, ZL
200910273363.1. (in Chinese)
[59] 朱玉贤, 姬生健, 卢迎春, 冯建勋. 棉花纤维的类 β 微管蛋白及其
编码基因与应用: 中国, CN02128986.7.
Zhu Y X, Ji S J, Lu Y C, Feng J X. Coding gene and its application of
β-tubulin protein in cotton fiber: China, CN02128986.7. (in Chinese)
[60] 郭三堆, 任茂智, 张锐. 棉花 Nodulin-like 基因及其启动子: 中国,
CN 2003101137982.
Guo S D, Ren M Z, Zhang R. Nodulin-like gene and its promoter in
cotton: China, CN 2003101137982. (in Chinese)
[61] 张献龙, 胡海燕, 刘迪秋, 涂礼利. 一种棉花纤维伸长期表达的启
动子及制备方法和应用: 中国, CN201210280595.1.
Zhang X L, Hu H Y, Liu D Q, Tu L L. A cotton fibers elongation
expression of promoter and its preparation methodand application:
China, CN201210280595.1. (in Chinese)
[62] 李静, 沈法富, 于海东, 韩秀兰. 转基因抗早衰棉的获得. 西北植
物学报, 2004, 24(8): 1419-1423.
Li J, Shen F F, Yu D H, Han X L. The obtaining of transgenic cottons
resistant to premature senescence. Acta Botanica Boreali-occidentalia
Sinica, 2004, 24(8): 1419-1423. (in Chinese)
[63] 傅荣昭, 孙勇如, 贾士荣. 植物遗传转化手册. 北京: 科学技术出
版社, 1995: 9-11.
Fu R Z, Sun Y R, Jia S R. Plant Genetic Transformation. Beijing:
Science and Technology Press, 1995: 9-11. (in Chinese)
[64] Vasil V, Castillo A M, Fromm M E, Vasil I K. Herbicide resistant
fertile transgenic wheat plants obtained by microprojectile bombardment of
regenerable embryogenic callus. Nature Biotechnology, 1992, 10: 667-674.
[65] Ming X T, Yuan H Y, Wang L J, Chen Z L. Agrobacterium-mediated
transformation of rice with help of bombardment. Acta Botanica
Sinica, 2001, 43(1): 72-73.
[66] Kleint T M, Wlofe E D, Wu R, SANFORD J C. High-velocity
microprojectiles for delievering nucleic acid into living cells. Nature,
1987, 327: 70-73.
[67] Philippe V, Horth Y, Pascal F. Enhancement of production and
regeneration of embryo genictypeII callus in ZeamPaysl by AgNO3.
Plant Cell Tissue and Organ Culture, 1989, 18: 143-151.
[68] 刘娜. vgb 基因在抗虫棉中的功能研究[D]. 乌鲁木齐: 新疆农业大
学, 2007.
Liu N. vgb study of gene function in insect-resistant cotton[D].
Urumqi: Xinjiang Agricultural University, 2007. (in Chinese)
[69] 牟红梅, 刘树俊, 周文娟, 文玉香, 张文俊, 魏荣瑄. 慈菇蛋白酶
抑制剂通过花粉管途径对小麦的导入及转基因植株分析. 遗传学
报, 1999, 26(6): 634-642.
Mu H M, Liu S J, Zhou W J, Wen Y X, Zhang W J, Wei R X.
transformation of wheat with insectide gene arrowhead proteinase
inhibitor by pollen tube pathway and analysis of transgenic plants.
Acta Genetica Sinica, 1999, 26(6): 634-642. (in Chinese)
[70] 曾君祉, 王东江, 吴有强, 张健, 周文娟, 朱小平, 徐乃正. 用花粉
管途径获得小麦转基因植株. 中国科学, 1993, 23(3): 256-262.
Zeng J Z, Wang D J, Wu Y Q, Zhang J, Zhou W J, Zhu X P, Xu N Z.
Obtained by pollen tube pathway transgenic wheat plants. Science in
China: Ser.B, 1993, 23(3): 256-262. (in Chinese)
[71] Langridge P, Brettschneider R, Lazzeri P, Lörz H. Transformation of
cereals via Agrobacterium and the pollen pathway: A critical
assessment. The Plant Journal, 1992, 2(4): 631-638.
[72] 李君, 李岩, 刘德虎. 植物遗传转化的替代方法及研究进展. 生物
技术通报, 2011(7): 31-36.
Li J, Li Y, Liu D H. Alternative methods of plant genetic
transformation and recent advances. Biotechnology Bulletin, 2011(7):

17 期 郭三堆等:中国转基因棉花研发应用二十年 3387
31-36. (in Chinese)
[73] Bechtold N, Ellis J, Pelletier G. In planta Agrobacterium mediated
gene transfer by infiltration of adult Arabidopsis thaliana plants.
Comptes Rendus de l'Académie des Sciences Pairs Life Sciences, 1993,
316: 1194-1199.
[74] Feldmann K. T-DNA insertion mutagenesis in Arabidopsis:
Mutational spectrum. The Plant Journal, 1991, 1: 71-82.
[75] Azpiroz-Leehan R, Feldmann K A. T-DNA insertion mutagenesis in
Arabidopsis: Going back and forth. Trends in Genetics, 1997, 13(4):
152-156.
[76] Torney F, Trewyn B G, Lin V S Y, Wang K. Mesoporous silica
nanoparticles deliver DNA and chemicals into plants. Nature
Nanotechnology, 2007, 2: 295-300.
[77] Yi B, Liu F Y, Zhang H, Liu H, Peng Y M, Liu Y H. Gene therapy of
mice peritoneal fibrosis by nano-carrier pamam mediated pCTGF-
shRNA. Cell Biology International, 2008, 32(3): S7.
[78] Chemin I, Moradpour D, Wieland S, Offensperger W B, Walter E,
Behr J P, Blum H E. Liver-directed gene transfer: A linear
polyethylenimine denvative mediates highly efficient DNA delivery to
primary hepatocytes in vitro and in vivo. Journal of Viral Hepatitis,
1998, 56: 369.
[79] Takefumi S, Eiji N, Tomohiko I, Mizukami A, Takakura Y, Kajiyama
S, Fukusaki E, Harashima S, Kobayashi A, Fukui K. A novel gene
delivery system in plants with calcium alginate micro-beads. Journal
of Bioscience and Bioengineering, 2002, 94(1): 87-91.
[80] 王国英. 转基因植物的安全性评价. 农业生物技术学报, 2001,
9(3): 205-207.
Wang G Y. The safety evaluation of teansgenic plants. Journal of
Agricultural Biotechnology, 2001, 9(3): 205-207. (in Chinese)
[81] 吴志平, 徐步进. 转基因植物释放后在环境中成为杂草的风险性.
生物工程进展, 1999, 19(1): 9-13.
Wu Z P, Xu B J. The risk of transgenic plants changing into wild
weeds after environmental release. Progress in Biotechnology, 1999,
19(1): 9-13. (in Chinese)
[82] Losey J E, Rayor L S, Carter M. Transgenic pollen harms rmnarch
larvae. Nature, 1999(399): 214-419.
[83] 国家农作物品种审定委员会办公室. 中国转抗虫基因棉花品种
(1997~2007). 北京: 中国农业出版社, 2008.
Office of the national crop varieties Committee. Transgenic Cotton
Varieties in China (1997-2007). Beijing: China Agriculture Press,
2008. (in Chinese)
[84] 中华人民共和国农业部公告: 国家审定棉花品种 1997 年-2013 年.
Announcement of the Ministry of agriculture of the people's Republic
of China: National approved cotton varieties from 1997 to 2003. (in
Chinese)
[85] 汪若海, 李秀兰, 杨付新, 付小琼. 从国家棉花品种区试看杂交棉
//中国棉花学会2005年年会暨青年棉花学术研讨会论文汇编. 2005:
117.
Wang R H, Li X L, Yang F X, Fu X Q. The development of hybrid
cotton from the perspective of the national trials for cotton varieties.
//Proceedings of Chinese Cotton Society 2005 Annual Conference &
Youth Symposium. 2005: 117. (in Chinese)
[86] 全国农业技术推广服务中心. 全国农作物主要品种推广情况统计
1996 年-2012 年.
The National Agro-Tech Extension and Service Center. Extension
statistics of main varieties of crops in China from 1996 to 2012. (in
Chinese)
[87] 金石桥, 刘逢举, 刘媛, 杨付新. 黄河流域常规棉品种近几年育种
趋势分析. 中国棉花, 2014, 41(1): 8-11.
Jin S Q, Liu F J, Liu Y, Yang F X. Discussion on the conventional
cotton breeding direction in the yellow river region in recent years.
China Cotton, 2014, 41(1): 8-11. (in Chinese)
[88] 付小琼, 杨付新, 王秀玲. 黄河流域春棉区试纤维品质近 16 年的
变化趋势//中国棉花学会年年会论文汇编. 2008: 99-102.
Fu X Q, Yang F X, Wang X L. Yellow spring trend of cotton fiber
quality test nearly 16 years// China Cotton Association Annual
Proceedings. 2008: 99-102. (in Chinese)
[89] 黄季焜, 米建伟, 林海, 王子军, 陈瑞剑, 胡瑞法, Rozeelle Scott,
Pray Carl. 中国 10 年抗虫棉大田生产:Bt 抗虫棉技术采用的直接效
益和间接外部效应评估. 中国科学: 生命科学, 2010, 40(3): 260-272.
Huang J K, Mi J W, Lin H, Wang Z J, Chen R J, Hu R F, Scott R, Carl
P. A decade of Bt cotton in farmer fields in China: Assessing the direct
effects and indirect externalities of Bt cotton adoption in China.
Scientia Sinica Vitae, 2010, 40(3): 260-272. (in Chinese)
[90] 苏岳静, 胡瑞发, 黄季焜, 范存会. 农民抗虫棉技术选择行为及其
影响因素分析. 棉花学报, 2004, 16(5): 259-264.
Su Y J, Hu R F, Huang J K, Fan C H. The determinants of farmer Bt
cotton technology adoption behavior in China. Acta Gossypii Sinica,
2004, 16(5): 259-264. (in Chinese)
[91] 张锐, 王远, 孟志刚 孙国清, 郭三堆. 国产转基因抗虫棉研究回
顾与展望. 中国农业科技导报, 2007, 9(4): 32-42.
Zhang R, Wang Y, Meng Z G, Sun G Q, Guo S D. Retrospect and
prospect of research on Chinese transgenic insecticidal cotton. Journal
of Agricultural Science and Technology, 2007, 9(4): 32-42. (in Chinese)
(责任编辑 李莉)

中国农业科学 2015,48(17):3388-3393 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.006
收稿日期:2015-01-20;接受日期:2015-04-14 基金项目:河南省重大科技专项(121100111400)、河南省小麦产业技术体系岗位科学家项目 联系方式:茹振钢,E-mail:[email protected]
黄淮麦区小麦品种的高产潜力与实现途径
茹振钢,冯素伟,李 淦
(河南科技学院小麦研究中心/现代生物育种河南省协同创新中心,河南新乡 453003)
摘要:黄淮麦区是中国冬小麦的主产区和高产区,对中国小麦生产以及国家粮食安全起着重要作用。针对中
国人多、地少的基本国情,以及耕地资源非农化、耕地利用非粮化的发展现状,指出未来提高小麦总产的根本出
路在于提高单产。要充分挖掘小麦高产潜力,培育高产品种是进一步提高单产的重要途径。文章根据黄淮麦区的
生产条件及生态特点,分析不同时期高产品种产量结构的发展变化趋势,指出在大田条件下实现小麦高产潜力,
千粒重与穗粒数并重是小麦新品种的发展方向。并从机械化生产对品种的要求出发,探讨了黄淮麦区小麦高产品
种的高产空间与创育思路,提出进一步挖掘黄淮麦区小麦品种高产潜力的有效途径:(1)小麦高产潜力的实现,
应重新认识和定位穗光合在产量形成中的作用,要充分挖掘和利用穗器官的光合优势,培育穗叶高光效品种。小
麦穗器官除具有空间优势外,其光合特点类似于 C4途径或介于 C3—C4中间型,籽粒呼吸释放的 CO2能被穗光合再次
固定。鉴于穗光合对籽粒产量形成的较大贡献,应强化绿穗灌浆特性,发挥穗器官的光合优势,提升穗粒重。(2)
提高单产水平,必须注重群体生物产量的提高。在保持现有收获指数基本不变的情况下,提高茎秆强度,实现植
株高大化、密植化,能有效改善群体穗叶空间结构。通过调节生长发育节律,培育小叶、壮秆、大穗型新品种,
实现小麦高生物产量。高生物产量品种还应拉开穗层,使穗层由一层增至三层,能有效提高单位面积容穗数。(3)
小麦杂种优势利用已日趋成熟,有效利用杂种优势是今后挖掘高产潜力的重要途径。利用杂种优势挖掘高产潜力
需同时兼顾品质性状的优化,充分考虑多个品质性状间的协调稳定。可通过多穗大穗实现高产,通过提高强势小
花结实比例稳定品质,从而实现高产与优质并重发展。挖掘小麦产量潜力是一个长期的动态过程。文章旨在通过
协调各种产量形成影响因素,最大限度地挖掘黄淮麦区小麦的高产潜力,为中国黄淮麦区的小麦高产育种实践提
供思路和方法借鉴。
关键词:小麦;黄淮麦区;高产潜力;高生物产量;高光效
High-Yield Potential and Effective Ways of Wheat in Yellow & Huai
River Valley Facultative Winter Wheat Region RU Zhen-gang, FENG Su-wei, LI Gan
(Center of Wheat Research of Henan Institute of Science and Technology/Henan Province Collaborative Innovation Center of
Modern Biological Breeding, Xinxiang 453003, Henan)
Abstract: Yellow & Huai River Valley Facultative Wheat Region is a major area of wheat production in China. In the present
production condition, improvement of the total production of wheat depends on the increase of yield per unit area. Therefore, the cultivation of high yield varieties is very important for wheat production. In this study, according to the production conditions and ecological characteristics of Yellow & Huai River Valley Facultative Winter Wheat Region, the yield potential under field conditions were analyzed, and pointed out that the two traits, i.e. 1000-grain weight and spike number, will make a greater contribution to wheat production. Furthermore, according to fitting in with the needs of mechanized production in this region, breeding strategies and yield potential of high yield varieties were discussed and pointed out three effective ways to achieve high-yield potential of wheat in future:
jia
文本框
返回目录

17 期 茹振钢等:黄淮麦区小麦品种的高产潜力与实现途径 3389
(1) Making effective utilization of photosynthetic efficiency of spike and cultivating high efficiency varieties. Besides the space superiority, the photosynthetic pathway of spike is C4 or C3 to C4, which can fix CO2 from seed respiration again. So the photosynthetic efficiency of spike plays important roles in grain filing of wheat. Enhancing the photosynthetic efficiency of spike to breed high efficiency varieties is one of the important ways to achieve high-yield potential of wheat. (2) Improving yields of wheat must focus on increasing population biomass. Maintaining the existing harvest index unchanged, improving the spatial structure of populations and breeding new varieties with lobules, strong stalk and large spike can achieve high biomass and improve yields. In addition, high biomass varieties should present different spike layers, which can effectively increase the spike number per unit area. (3) Heterosis is another effective way to realize yield-potential of wheat. However, in the process of utilization of heterosis, both of yield and quality should be considered simultaneously, especially the coordination of different quality traits. Increasing the spike number and improving the proportion of superior florets can realize the coordination of high yield and superior quality. Achieving the yield-potential of wheat is a long-term process and this paper aims to provide ideas and methods for achieving the yield-potential and realizing maximum output in Yellow & Huai River Valley Facultative Wheat Region.
Key words: wheat; Yellow & Huai River Valley Facultative Wheat Region; high-yield potential; high biological yield; high photosynthetic efficiency
中国是世界小麦生产大国,也是小麦消费大国。
国家食物与营养咨询委员会在 2003 年提出了中国食
物发展的阶段性目标,预计到 2020 年人均粮食占有量
应该在 437 kg 左右,到 2030 年要提高到 472 kg 左右
才能满足人们生活对粮食的需求水平[1]。依此推算,
到 2030 年全国粮食消费需求要达到 7.5 亿吨,是现在
粮食生产量的 1.5 倍,提高粮食生产能力刻不容缓。
现阶段,在耕地资源非农化、耕地利用非粮化、人口
持续增加、粮用耕地资源十分有限的情况下,必须通
过提高单产水平才能实现粮食供需平衡。因此,充分
挖掘小麦高产性状的遗传潜力,培育超高产小麦新品
种尤为重要。而要大幅度提高小麦产量,就要打破高
产育种的形态和生理瓶颈,这是中国小麦育种必须着
力解决的重大问题[2]。黄淮麦区是中国最大的小麦适
宜生产区域,常年小麦种植面积约 1 530 万 hm2,通
过遗传改良,充分挖掘该区小麦品种的高产潜力,对
实现全国小麦产量持续稳定增长,保障国家口粮绝对
安全具有重大意义。
1 黄淮麦区小麦品种高产潜力分析
1.1 黄淮麦区的生态特点
黄淮麦区地处温带季风气候区,主体位于北纬
33°—37.5°,光照充足,四季分明,平原辽阔,灌溉
便利,因而非常适宜小麦生长。该区小麦全生育期
225—250 d,光照时数 1 200—2 000 h,太阳辐射总量
52—66 KCal·cm-2,积温 2 000—2 200℃,降水量
300—200 mL,水分亏缺 100—300 mL。该区是中国冬
小麦的主产区和高产区,小麦种植面积约占全国麦播
面积的 55%,总产量约占全国小麦总产的 60%,在中
国小麦生产中具有十分重要的地位。近年来,在全球
气候变化的大背景下,冻害、旱害、倒伏等自然灾害
以及条锈病、白粉病、纹枯病等频繁发生,危害渐趋
加重,给小麦生产带来了严重威胁。 1.2 黄淮麦区小麦的高产潜力
小麦的产量潜力是指某一发展时期所能达到的最
高产量,是各性状相互协调的结果。通过总结分析黄
淮麦区各时期小麦品种的产量水平可知(表 1),从 20世纪 50 年代小麦品种产量水平 3 000 kg·hm-2上升到 20世纪 80 年代的 5 500 kg·hm-2,主要是通过降低株高增
强了群体抗倒性,进而提高了群体密度和经济系数。前
30 年是成穗数和穗粒数并重的多穗多花多实育种阶
段。以碧蚂 1 号、阿勃与百农 3217、济南 2 号为例,
株高由 113 cm 降至 88 cm,平均降低 32%;而成穗数
由 462 万穗/hm2上升至 631 万穗/hm2,增加了 36.7%;
穗粒数由 19.6 粒/穗上升到 28.6 粒/穗,增加了 45.9%。 20 世纪 80 年代之后的 30 年,小麦品种的成穗数
相对稳定,维持在 620 万穗—630 万穗/hm2 高水平状
态,而品种产量水平则由 5 500 kg·hm-2 提高到 8 000 kg·hm-2,主要是通过提高千粒重和穗粒数来实现的。
以矮抗 58、济麦 22、郑麦 9023、烟农 19 为例,千粒
重由 34.5 g 上升至 42 g,增幅为 21.7%;穗粒数由 28.6粒增至 34 粒,增幅为 19.8%。这 30 年是千粒重与穗
粒数并重的多穗多花大粒育种阶段。 参照世界小麦单产水平与产量结构动态变化的发
展趋势(图 1),目前,在黄淮麦区的土壤、气候
条件下,安全产量首先是要由 8 000 kg·hm-2 提升至
10 500 kg·hm-2。在今后相当长的一段时间,继续提高
小麦新品种的产量潜力,千粒重与穗粒数仍需并重发
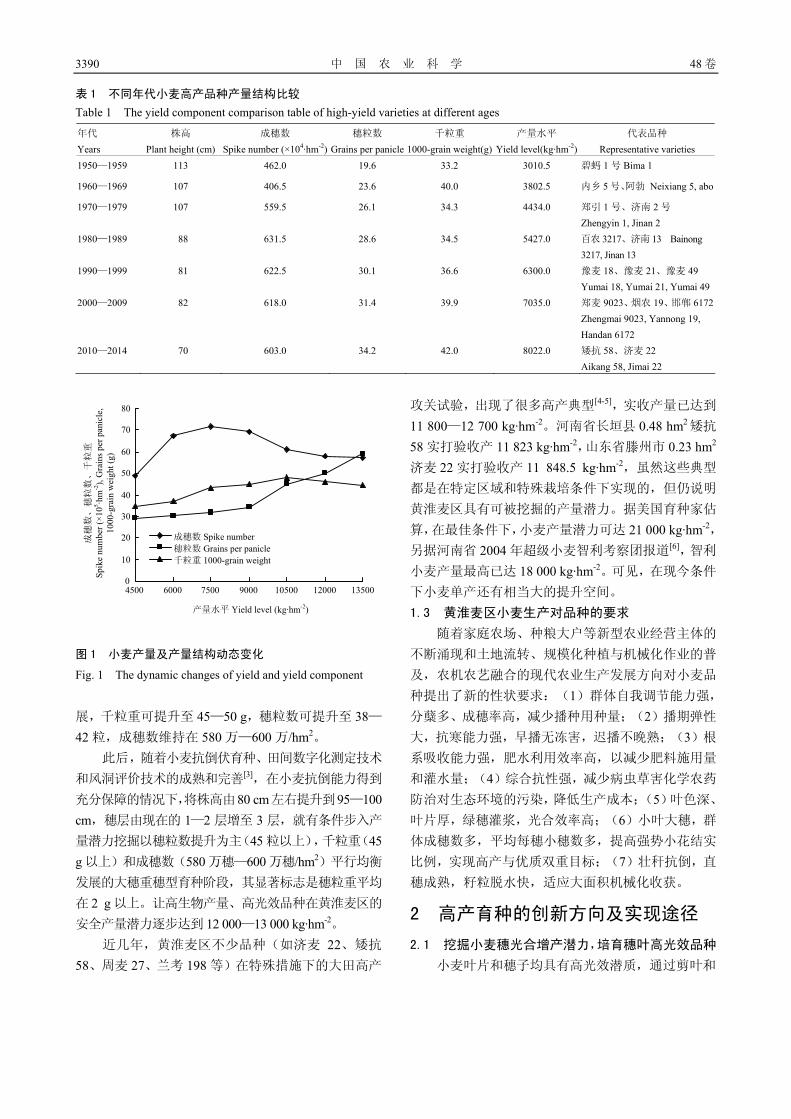
3390 中 国 农 业 科 学 48 卷
表 1 不同年代小麦高产品种产量结构比较
Table 1 The yield component comparison table of high-yield varieties at different ages
年代 Years
株高 Plant height (cm)
成穗数 Spike number (×104·hm-2)
穗粒数 Grains per panicle
千粒重 1000-grain weight(g)
产量水平 Yield level(kg·hm-2)
代表品种 Representative varieties
1950—1959 113 462.0 19.6 33.2 3010.5 碧蚂 1 号 Bima 1
1960—1969 107 406.5 23.6 40.0 3802.5 内乡 5 号、阿勃 Neixiang 5, abo
1970—1979 107 559.5 26.1 34.3 4434.0 郑引 1 号、济南 2 号 Zhengyin 1, Jinan 2
1980—1989 88 631.5 28.6 34.5 5427.0 百农 3217、济南 13 Bainong 3217, Jinan 13
1990—1999 81 622.5 30.1 36.6 6300.0 豫麦 18、豫麦 21、豫麦 49 Yumai 18, Yumai 21, Yumai 49
2000—2009 82 618.0 31.4 39.9 7035.0 郑麦 9023、烟农 19、邯郸 6172Zhengmai 9023, Yannong 19, Handan 6172
2010—2014 70 603.0 34.2 42.0 8022.0 矮抗 58、济麦 22 Aikang 58, Jimai 22
0
10
20
30
40
50
60
70
80
4500 6000 7500 9000 10500 12000 13500
产量水平 Yield level (kg·hm-2)
成穗数 Spike number穗粒数 Grains per panicle千粒重 1000-grain weight
成穗数、穗粒数、千粒重
Spik
e nu
mbe
r (×1
05 ·hm
-2),
Gra
ins p
er p
anic
le,
1000
-gra
in w
eigh
t (g)
图 1 小麦产量及产量结构动态变化
Fig. 1 The dynamic changes of yield and yield component
展,千粒重可提升至 45—50 g,穗粒数可提升至 38—42 粒,成穗数维持在 580 万—600 万/hm2。
此后,随着小麦抗倒伏育种、田间数字化测定技术
和风洞评价技术的成熟和完善[3],在小麦抗倒能力得到
充分保障的情况下,将株高由80 cm左右提升到95—100 cm,穗层由现在的 1—2 层增至 3 层,就有条件步入产
量潜力挖掘以穗粒数提升为主(45 粒以上),千粒重(45 g 以上)和成穗数(580 万穗—600 万穗/hm2)平行均衡
发展的大穗重穗型育种阶段,其显著标志是穗粒重平均
在 2 g 以上。让高生物产量、高光效品种在黄淮麦区的
安全产量潜力逐步达到 12 000—13 000 kg·hm-2。 近几年,黄淮麦区不少品种(如济麦 22、矮抗
58、周麦 27、兰考 198 等)在特殊措施下的大田高产
攻关试验,出现了很多高产典型[4-5],实收产量已达到
11 800—12 700 kg·hm-2。河南省长垣县 0.48 hm2矮抗
58 实打验收产 11 823 kg·hm-2,山东省滕州市 0.23 hm2
济麦 22 实打验收产 11 848.5 kg·hm-2,虽然这些典型
都是在特定区域和特殊栽培条件下实现的,但仍说明
黄淮麦区具有可被挖掘的产量潜力。据美国育种家估
算,在最佳条件下,小麦产量潜力可达 21 000 kg·hm-2,
另据河南省 2004 年超级小麦智利考察团报道[6],智利
小麦产量最高已达 18 000 kg·hm-2。可见,在现今条件
下小麦单产还有相当大的提升空间。 1.3 黄淮麦区小麦生产对品种的要求
随着家庭农场、种粮大户等新型农业经营主体的
不断涌现和土地流转、规模化种植与机械化作业的普
及,农机农艺融合的现代农业生产发展方向对小麦品
种提出了新的性状要求:(1)群体自我调节能力强,
分蘖多、成穗率高,减少播种用种量;(2)播期弹性
大,抗寒能力强,早播无冻害,迟播不晚熟;(3)根
系吸收能力强,肥水利用效率高,以减少肥料施用量
和灌水量;(4)综合抗性强,减少病虫草害化学农药
防治对生态环境的污染,降低生产成本;(5)叶色深、
叶片厚,绿穗灌浆,光合效率高;(6)小叶大穗,群
体成穗数多,平均每穗小穗数多,提高强势小花结实
比例,实现高产与优质双重目标;(7)壮秆抗倒,直
穗成熟,籽粒脱水快,适应大面积机械化收获。
2 高产育种的创新方向及实现途径
2.1 挖掘小麦穗光合增产潜力,培育穗叶高光效品种
小麦叶片和穗子均具有高光效潜质,通过剪叶和

17 期 茹振钢等:黄淮麦区小麦品种的高产潜力与实现途径 3391
套袋试验即可证实[7]。过去研究一直强调叶器官的光
合作用在小麦籽粒产量形成中的作用,而对穗器官光合
在籽粒产量形成中的贡献认识不足。随着研究的逐步深
入,越来越多的证据表明,麦类作物的非叶器官——穗
器官的光合代谢对籽粒的产量形成起着重要作用[8-9],叶
片的光合作用主要是对植株营养器官建成提供光合产
物,只有部分光合产物运往发育着的籽粒[10]。 一些研究认为[11],相对于叶片来讲,在小麦籽粒
产量形成期,穗器官不但具有空间位置优势,极有利
于截获光能和 CO2,而且还具有明显的生理优势。越
来越多的证据表明,穗光合代谢类似于 C4途径或介于
C3—C4中间型,籽粒呼吸释放的大量 CO2能被穗光合
再次固定[12-13]。但麦类作物穗光合产物对籽粒灌浆贡
献率的高低依赖于品种类型和环境条件[14-15],小旗叶
有芒品种其穗光合的贡献较大[16]。Araus 等[17]对旗叶
和穗子采取同样遮光处理,以证明籽粒中的光合产物
大部分来自于穗器官而不是来自于旗叶,说明穗子对
籽粒的贡献率要远大于旗叶。特别是受水分胁迫时,
其光合优势更易发挥[18-19]。据此认为,在当前小麦高
产和超高产育种的探索中,应该充分挖掘和利用穗器
官的光合优势,从而提高小麦品种产量潜力。也有人
将穗光合性能作为麦类作物形态和生理育种的重要指
标[20]。近年来,通过对大量品种的研究,发现穗光合
对粒重的贡献约为 1.5 倍的旗叶,穗是产量形成的重
要光合器官[3]。 实际上,通过对源于不同纬度地区大量的小麦品
种资源经多年研究,发现来自于高纬度地区的品种普
遍表现叶色深,耐弱光性好;来自于低纬度地区的品
种普遍表现叶片厚,耐强光特性好。将小麦叶片的耐
弱光特性和耐强光特性有机结合,将叶光合与穗光合
协调配置,育成了叶色深、叶片厚、小叶形、绿穗灌
浆的高光效小麦品种百农 419,穗光合较普通小麦提
高近 40%,叶光合提高 10%以上,大幅度提高了产量
潜力。 2.2 挖掘生物产量优势,开展高生物产量育种
为了提高小麦产量,人们首先进行了增强抗倒性
的矮化育种。而小麦矮秆品种的增产作用主要是通过
降低植株高度、增加群体密度,从而提高了成穗数和
收获指数来实现的[21]。但是,如今的矮秆品种也有明
显的不足,主要表现在生物产量较低,矮秆品种再次
大幅度提高产量还是很难,加之收获指数已接近上限
(45%—50%),增产余地十分有限。 在保持现有收获指数基本不变的情况下,通过提
高茎秆强度,实现植株高大化、密植化,改善植株空
间结构,从而实现小麦品种生物产量的大幅度提高,
将是提高小麦产量潜力的重要途径。目前,不少育种
单位已开始将株高目标由矮秆变为中高秆,强化穗层
与叶层相分离的小麦株型结构的调整和创新。实践得
出,株高每提升 10 cm,小麦产量就会相应增加 600—750 kg·hm-2。
提高小麦生物产量,主要可以通过以下途径: (1)调节生长发育节律 小麦生长发育进程与产
量有着密切的关系。小麦出苗-拔节期是决定成穗数的
关键时期;拔节-抽穗期是决定穗粒数的关键时期;抽
穗-成熟期是决定粒重的关键时期。因此,根据黄淮麦
区小麦越冬前后的气候特点,在小麦生长的前期要打
好根蘖基础,尽量多的积存生物量;中期要稳健生长,
确保多成穗成大穗;后期要灌浆充分,实现籽粒饱满。
为了防止小麦越冬前无效生长,一定要集聚各类抗冻、
抗寒基因来为大生长量提供安全防护,实现前期生长
量大(干物质量 25%以上)、中期生长快(干物质量
35%以上)、后期籽粒灌浆充分(干物质积累 40%左
右)的高生物产量小麦的遗传调整。 (2)培育小叶、壮秆、大穗型品种 小叶、厚叶、
直叶和壮秆有利于小麦群体内部的光透射与光吸收;
穗层与叶层分离,有利于发挥穗子和叶片的光合潜势,
提高群体光合生产能力,促使光合产物向籽粒及时运
转。地上部植株机能的有效发挥,要靠根系作保障。
一个对土壤 pH 调节能力强、水平根系与垂直根系均
发达的地下根群,是小麦品种地上部正常生长发育和
实现高产的基础。培育高生物产量小麦品种,要地上、
地下协同选择。 (3)注重厚穗层与绿穗灌浆特性的强化配置 提
高小麦产量,不仅要提高茎秆强度、增加植株高度,
还要拉开穗层,增厚穗层。对于高光效绿穗灌浆的品
种培育,高大植株群体第一层穗(顶层穗)占 35%左
右,第二层穗(中间穗)占 45%左右,第三层穗(底
层穗)占 20%左右,有利于降低穗层拥挤度。这样主
茎及大小分蘖抽穗时间拉开,群体穗层拉开,穗层增
厚,不仅有利于提高成穗数,还能减轻穗层郁蔽,减
缓群体养分需求高峰,促使穗光合充分发挥,叶光合
环境趋于改善,整体提高小麦产量。 2.3 有效利用杂种优势,优选杂交小麦制种新途径
与其他作物一样,小麦也存在明显的杂种优势,
特别是生长优势。杂交小麦的突出表现是植株高大,
穗大粒多,千粒重高,明显的劣势是成穗数偏少。将

3392 中 国 农 业 科 学 48 卷
杂交小麦按优势类群进行组配就能很好地克服上述不
足。如利用黄淮类群的矮秆、多穗、强根系亲本,与
西南类群的大穗大粒亲本及南美洲智利的壮秆特大穗
亲本相组配,就能配制出穗多秆壮、穗大粒多的强优
势杂交小麦。经过世界近百年、中国约 50 年的不懈探
讨,目前,杂交小麦研究在中国已日趋成熟,例如河
南省的 BNS 型、云南省和四川省的 C49S 型、新疆的
AL 型等各种类型的杂种优势利用途径,都分别取得
了一些制种产量高、增产幅度大的重要成果,已成为
黄淮麦区今后小麦实现高产的创新途径[22]。
3 黄淮麦区对高产、优质、高效小麦
品种的现实需求
在提高小麦产量潜力、选育高产品种的同时,必
须兼顾优质水平的不断提升。根据多年调查,生产急
需的品种已不再是产量特别高或品质特别优的低效益
品种,而是既高产又优质的、抗倒抗病能力强、管理
简单、适于大规模机械化收获的好品种。高产优质是
农业生产的现实需要。小麦育种家在考虑优质目标时
要特别注意加强品质性状之间的协调稳定性,选择如
蛋白质含量、湿面筋含量、吸水率和面团稳定时间的
协调稳定性。现今通过高分子谷蛋白亚基与低分子谷
蛋白亚基优化组配、强势小花结实性选择和降低黑麦
碱含量等方法,结合氮高效、磷高效的辅助选择,育
成了一批好品种,如郑麦 7698、周麦 26、济麦 23 等,
基本实现了高产、优质、低成本的品种创新目标。
4 结束语
小麦产量潜力的挖掘,要受多方因素的综合作用,
是一个长期的动态过程,不会一蹴而就。不同时期、
不同生态区域可能有不同的主导影响因素,诸如气候
资源、土壤肥力、栽培管理与机械化作业水平等外部
因素和麦类群体株型特点、器官排布、生理生化机能、
网络代谢完善度等内部遗传潜质,再如育种理论与技
术水平、市场需求与比较效益等都需要科学分析、综
合运筹。从科学技术方面,一定要做到穷尽小麦产量
潜力,得出一个极大值;从人类社会的持续发展方面,
永远都不希望资源枯竭,要留下后人对小麦产量潜力
的需求空间。
References
[1] 张晶. 2035 年中国人粮关系情景分析. 中国农业资源与区划, 2011,
32(3): 26-31.
Zhang J. Scenario analysis of relationship between population and
grain in China in 2035. Chinese Journal of Agricultural Resources
and Regional Planning, 2011, 32(3): 26-31. (in Chinese)
[2] 李振声. 我国小麦育种的回顾与展望. 中国农业科技导报, 2010,
12(2): 1-4.
Li Z S. Retrospect and prospect of wheat breeding in China. Journal
of Agricultural Science and Technology, 2010, 12(2): 1-4. (in Chinese)
[3] Niu L Y, Feng S W, Ru Z G, Li G, Zhang Z P, Wang Z W. Rapid
determination of single-stalk and population lodging resistance
strengths and an assessment of the stem lodging wind speeds for
winter wheat. Field Crops Research, 2012, 139: 1-8.
[4] http://newpaper.dahe.cn/hnrb/html/2012-09/11/content_780327.htm?
div =-1.
[5] http://www.saas.ac.cn/news/shownews.asp?id=6605.
[6] 郭天财, 沈天民, 王西成, 茹振钢. 智利的小麦科研与生产. 麦类
作物学报, 2005, 25(2): 132-134.
Guo T C, Shen T M, Wang X C, Ru Z G. Research and production of
wheat in Chile. Journal of Triticeae Crops, 2005, 25(2): 132-134. (in
Chinese)
[7] 刘万代, 尹钧, 朱高纪. 剪叶对不同穗型小麦品种干物质积累及籽
粒产量的影响. 中国农业科学, 2007, 40(7): 1353-1360.
Liu W D, Yin J, Zhu G J. Effects of leaf removal on dry matter
accumulation and grain yield in different spike-type wheat varieties.
Scientia Agricultura Sinica, 2007, 40(7): 1353-1360. (in Chinese)
[8] 冯素伟, 董娜, 姜小苓, 茹振钢, 李笑慧, 程振普. 小麦不同光合
器官对千粒重贡献的研究. 河南科技学院学报: 自然科学版, 2011,
39(4): 1-4.
Feng S W, Dong N, Jiang X L, Ru Z G, Li X H, Cheng Z P. Study on
the contribution of different photosynthesis-organs of wheat to
1000-grain weight. Journal of Henan Institute of Science and
Technology, 2011, 39(4): 1-4. (in Chinese)
[9] 鲁清林, 柴守玺, 张礼军, 周刚. 冬小麦叶片和非叶器官对粒重的
贡献. 草业科学, 2013, 22(5): 165-174.
Lu Q L, Chai S X, Zhang L J, Zhou G. Contribution of winter wheat
leaf and non-leaf organs to grain weight. Acta Prataculturae Sinica,
2013, 22(5): 165-174. (in Chinese)
[10] Gebbing T, Schnyder H. 13C Labeling kinetics of sucrose in glumes
indicates significant refixation of respiration CO2 in the wheat ear.
Australian Journal of Plant Physiology, 2001, 28: 1047-1053.
[11] Morgan J M. Osmotic adjustment in the spikelets and leaves of wheat.
Journal of Experimental Botany, 1980, 31: 655-665.
[12] Ziegler-Jons A. Gas exchange of ears of cereals in response to carbon

17 期 茹振钢等:黄淮麦区小麦品种的高产潜力与实现途径 3393
dioxide and light: II. Occurrence of a C3~C4 intermediate type of
photosynthesis. Planta, 1989, 178: 164-175.
[13] Johnson R, Moss D N. Effect of water stress on 14CO2 fixation and
translocation in wheat during grain filling. Crop Science, 1976, 16:
697-701.
[14] 魏爱丽, 王志敏. 小麦不同光合器官对穗粒重的作用及基因型差
异研究. 麦类作物学报, 2001, 21(2): 57-61.
Wei A L, Wang Z M. A study on the contribution of different organs to
grain weight in different genotype wheat. Journal of Triticeae Crops,
2001, 21(2): 57-61. (in Chinese)
[15] 张英华, 杨佑明, 曹莲, 郝杨凡, 黄菁, 李金鹏, 姚得秀, 王志敏.
灌浆期高温对小麦旗叶与非叶器官光合和抗氧化酶活性的影响.
作物学报, 2015, 41(1): 136-144.
Zhang Y H, Yang Y M, Cao L, Hao Y F, Huang J, Li J P, Yao D X,
Wang Z M. Effect of high temperature on photosynthetic capability
and antioxidant enzyme activity of flag leaf and non-leaf organs in
wheat. Acta Agronomic Sinica, 2015, 41(1): 136-144. (in Chinese)
[16] Blum A. Photosynthesis and transpiration in leaves and ears of wheat
and barley varieties. Journal of Experimental Botany, 1985, 36(3):
432-440.
[17] Araus J L, Brown H R, Febrero A, Bort J, Serret M D. Ear
photosynthesis, carbon isotope discrimination and the contribution of
respiratory CO2 to different in grain mass in durum wheat. Plant Cell
and Environment, 1993, 1(6): 383-392.
[18] 张永平, 王志敏, 黄琴, 谢岷. 不同水分供给对小麦叶与非叶器官
叶绿体结构和功能的影响. 作物学报, 2008, 34(7): 1213-1219.
Zhang Y P, Wang Z M, Huang Q, Xie M. Changes of chloroplast ultra
microstructure and function of different green organs in wheat under
limited irrigation. Acta Agronomic Sinica, 2008, 34(7): 1213-1219. (in
Chinese)
[19] Martinez D E, Luquez V M, Bartoli C G. Persistence of photo-
synthetic components and photochemical efficiency in ears of water-
stressed wheat (Triticum aestivum). Physiology Plant, 2003, 119:
519-525.
[20] 王志敏, 张英华, 张永平, 吴永成. 麦类作物穗器官的光合性能研
究进展. 麦类作物学报, 2004, 24(4): 136-139.
Wang Z M, Zhang Y H, Zhang Y P, Wu Y C. Review on
photosynthetic performance of ear organs in Triticeae crops. Journal
of Triticeae Crops, 2004, 24(4): 136-139. (in Chinese)
[21] 冯素伟, 胡铁柱, 姜小苓, 张自阳, 李小军, 茹振钢. 小麦高产品
种的主要性状比较与相关性分析. 河南科技学院学报, 2012, 40(6):
1-6.
Feng S W, Hu T Z, Jiang X L, Zhang Z Y, Li X J, Ru Z G. Comparison
of main characters and correlation analysis between different
high-yield wheat varieties. Journal of Henan Institute of Science and
Technology, 2012, 40(6): 1-6. (in Chinese)
[22] 李罗江, 茹振刚, 高庆荣, 姜辉, 郭凤芝, 吴世文, 孙哲. BNS 小麦的
雄性不育性及其温光特性. 中国农业科学, 2009, 42(9): 3019-3027.
Li L J, Ru Z G, Gao Q R, Jiang H, Guo F Z, Wu S W, Sun Z. Male
sterility and thermo-photosensitivity characteristics of BNS in wheat.
Scientia Agricultura Sinica, 2009, 42(9): 3019-3027. (in Chinese)
(责任编辑 李莉)

中国农业科学 2015,48(17):3394-3403 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.007
收稿日期:2015-02-12;接受日期:2015-04-14 基金项目:中国农业科学院作物科学研究所中央级公益性科研院所基本科研业务费专项资金、中国农业科学院科技创新工程资助 联系方式:何中虎,E-mail:[email protected]
中麦 175 高产高效广适特性解析与育种方法思考
何中虎1,2,陈新民
1,王德森
1,张 艳
1,肖永贵
1,李法计
1,
张 勇1,李思敏
1,夏先春
1,张运宏
1,庄巧生
1
(1 中国农业科学院作物科学研究所,北京 100081;2CIMMYT 中国办事处,北京 100081)
摘要:解析中麦 175 的高产潜力、水肥高效特性、优良品质及广泛适应性机理将为培育突破性新品种提供理
论指导和实用信息。(1)中麦 175 穗数容易达到 750 个/m2,收获指数高达 0.49,穗数多和收获指数高为实现高产
潜力奠定基础;矮秆及茎秆弹性好与株型紧凑、叶片小且直立为穗多、不倒伏提供保障。(2)千粒重与株高对水分
反应敏感性低,灌浆中后期叶片衰老速度慢,灌浆速度快,这是其水分利用效率高和在旱肥地表现突出的主要原
因;中麦 175 产量的水分敏感指数为 0.86,而京冬 8 号为 1.13。(3)氮肥吸收和利用效率皆高,穗粒数和千粒重
对肥料敏感性低,为肥料高效利用奠定基础;在 6种不同施氮水平下,中麦 175 的产量均高于京冬 17,氮肥施用
量对穗粒数和千粒重影响小。良好的根系为肥水高效利用提供保障。(4)籽粒软质,面筋强度中等偏弱、延展性好,
面粉颜色亮白,为面条和馒头兼用型品质奠定基础;锌高效吸收运转使籽粒锌含量高,具备较高营养价值。(5)
含有光周期不敏感基因 Ppd-D1b、肥水高效、抗寒、耐高温和抗病性等为中麦 175 的广适性和跨区域种植提供保
障。本研究为高产高效广适性新品种培育提供了重要经验和理论支撑。
关键词:小麦品种;中麦 175;高产潜力;水肥高效;广泛适应性
Characterization of Wheat Cultivar Zhongmai 175 with High
Yielding Potential, High Water and Fertilizer Use Efficiency, and Broad Adaptability
HE Zhong-hu1,2, CHEN Xin-min1, WANG De-sen1, ZHANG Yan1, XIAO Yong-gui1, LI Fa-ji1,ZHANG Yong1, LI Si-min1, XIA Xian-chun1, ZHANG Yun-hong1, ZHUANG Qiao-sheng1
(1Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081; 2CIMMYT China Office, Beijing 100081)
Abstract: Documentation of leading cultivars will provide crucially important information for cultivar development. The
objective of this study is to characterize high yield potential, water and fertilizer use efficiency, excellent quality and broad adaptation in wheat cultivar Zhongmai 175. High yield potential was largely due to the increased spike number and high harvest index, while short (around 80 cm) and erect plant type with small leaves contributed to its outstanding lodging resistance. Spike number could easily reach 750/m2, 20%-25% increase in comparison with check cultivar Jingdong 8, while harvest index was 0.49. Insensitivity of plant height and thousand kernel weight to water stress, due to slow leaf senescence and fast grain filling rate, made it high water use efficiency and better performance under rainfed condition. Water sensitive index for yield in Zhongmai 175 was 0.86, while that of Jingdong 8 was 1.13. Nutrient intake and use efficiency contributed to insensitivity of kernel number per spike and thousand kernel weight, and thus better performance under different fertilizer applications. Zhongmai 175 outyielded Jingdong 17 at six different nitrogen levels. It was characterized with soft kernel, weak dough strength and excellent extensibility, and bright flour color, thus conferred excellent dual qualities for Chinese noodles and steamed bread. High Zn content was due to its efficiency in Zn intake and transportation, thus excellent nutritional quality. Presence of Ppd-D1b, water and fertilizer use efficiency, tolerance to cold in winter
jia
文本框
返回目录

17 期 何中虎等:中麦 175 高产高效广适特性解析与育种方法思考 3395
and high temperature during grain filling stage, and resistance to yellow rust, provided basis for its broad adaptation in three wheat ecological zones. This study will provide very important information and experience for developing new cultivars with high yield potential and broad adaptability.
Key words: wheat cultivar; Zhongmai 175; high yield potential; water and fertilizer use efficiency; broad adaptation
0 引言
北部冬麦区曾是我国小麦育种与研究的中心,中
国农业科学院与中国农业大学等单位不仅在育种方法
研究方面走在国内前列,培育的新品种也曾在生产中
发挥过重大作用。但自 1990 年以来,本麦区的育种工
作遇到前所未有的挑战,主要表现在 4 个方面。一是
北部冬麦区东片包括北京、天津、冀东与冀中北和晋
中地区(基本为水浇地)受产业结构调整及减少农业
用水等政策的影响,小麦面积由过去的 200 多万 hm2
下降到目前的不足 100 万 hm2,其中北京市的面积由
过去的 18.7 万 hm2降到目前的 3.3 万 hm2,同时节水、
节肥、易管理已成为生产的普遍要求,育种力量明显
下降。北京市农林科学院于 1990 年育成的京冬 8 号曾
为生产发展作出了重要贡献,但其产量偏低、秆高易
倒伏、品质较差、感白粉病等,难以满足市场需求。
1995—2005 年虽然各单位审定了不少品种,但大面积
推广的很少。总体来说,小麦育种没有实质性突破,
与主产区的差距有所拉大。二是由于气候变化的影响
日益显著,冷冬年份出现频率减少,山东和石家庄等
地育成的新品种矮秆高产特性突出,颇受农民青睐,
尽管存在冬季冻害等风险,仍然形成了南部品种北移
的明显趋势,如济麦 20 和济麦 22 在天津大面积推广。
三是以旱地为主的北部冬麦区西片及黄淮旱肥地育种
力量有所下降,造成现有品种抗病性差,植株偏高,
虽然在旱灾年份表现较好,但丰水年份倒伏十分严重,
往往是丰产不丰收,这些地区迫切需要秆子偏矮、抗
旱、抗寒和灌浆期抗高温的高产广适性品种。虽然历
史上本区东片育成的品种也曾在西片大面积推广,如
丰抗 8 号和中优 9507 先后成为陇东地区的主栽品种,
但总体来说,东片品种的抗条锈病能力偏差,水分利
用效率偏低,不易在西片推广。四是 20 世纪 90 年代
初,针对气候变化的可能影响,农业部曾启动了冬麦
北移和西延计划,但受气候和品种的双重影响,冬麦
替代春麦的设想在当时未能实现[1];但近 10 多年来的
气候变暖进一步加速,春麦改冬麦技术在不少地区已
大面积推广,生产上迫切需要抗寒、高产、矮秆抗倒、
抗病、广适性新品种。
针对这一情况,我们将育种工作的重点逐步转向
主产区如河北中南部和河南省,同时不放弃甚至加强
北部冬麦区的育种工作,期望育成的新品种能在北部
冬麦区东片大面积推广的同时,还能在西片和新疆及
黄淮旱肥地扩大阵地,为小麦生产作出应有贡献。为
此,育成并推广了以中麦 175 为代表的高产高效优质
广适性新品种,基本扭转了中国农业科学院作物科学
研究所从 20 世纪 80 年代中期以来小麦育种下滑的局
面。通过株型等的改良,2001 年育成的中麦 175 实现
了高产潜力、水肥高效、抗病抗逆、优良面条品质、
广适性与早熟性的良好结合,分别通过北部冬麦区水
地(国审麦 2008016)、黄淮旱肥地(国审麦 2011018)、北京市(京审麦 2007001)、山西省(晋审麦 2007007)、河北省(冀审麦 2009017)、青海省(青审麦 2011001)和甘肃省(甘审麦 2012010)的品种审定,且已成为
上述地区的主栽品种及北部冬麦区、北京市、山西省
与河北省冀中北地区区试的对照品种。本文采用育种、
生理、栽培、谷物化学与分子标记相结合的方法,较
全面地解析了中麦 175 的优异特性,目的是为培育突
破性新品种提供理论支撑。
1 产量潜力高
中麦 175 的高产潜力主要表现在两个方面。一是
在各级区域试验中均比对照增产,平均增产 4.8%,增
产点率为 67%—100%,在多数试验中名列前三位,在
所有生产试验中均名列第一,平均较对照增产 8.9%(表
1)。株型紧凑和小区面积小是区试增幅小于生产试验
增幅的重要原因。二是大面积生产示范表现更突出,河
北、天津和北京等地将中麦 175 作为高产创建的主要品
种,大面积容易达到 8 250 kg·hm-2,部分测产和实收结
果列于表 2。如河北省保定市徐水农场二分场示范 6.7 hm2,2005—2009 年连续 4 年度超过 8 250 kg·hm-2,这
在北部冬麦区是很不容易的,或者说以前的品种不具备
这一产量潜力。2007—2008 年度北京市房山区琉璃河
镇实收产量高达 9 169.5 kg·hm-2,创本麦区高产记录。
另外在青海省小面积达到 10 665 kg·hm-2,在石家庄全
生育期仅浇 1 水的条件下产量达 10 609 kg·hm-2,在新
疆阿克苏产量高达 10 440 kg·hm-2。
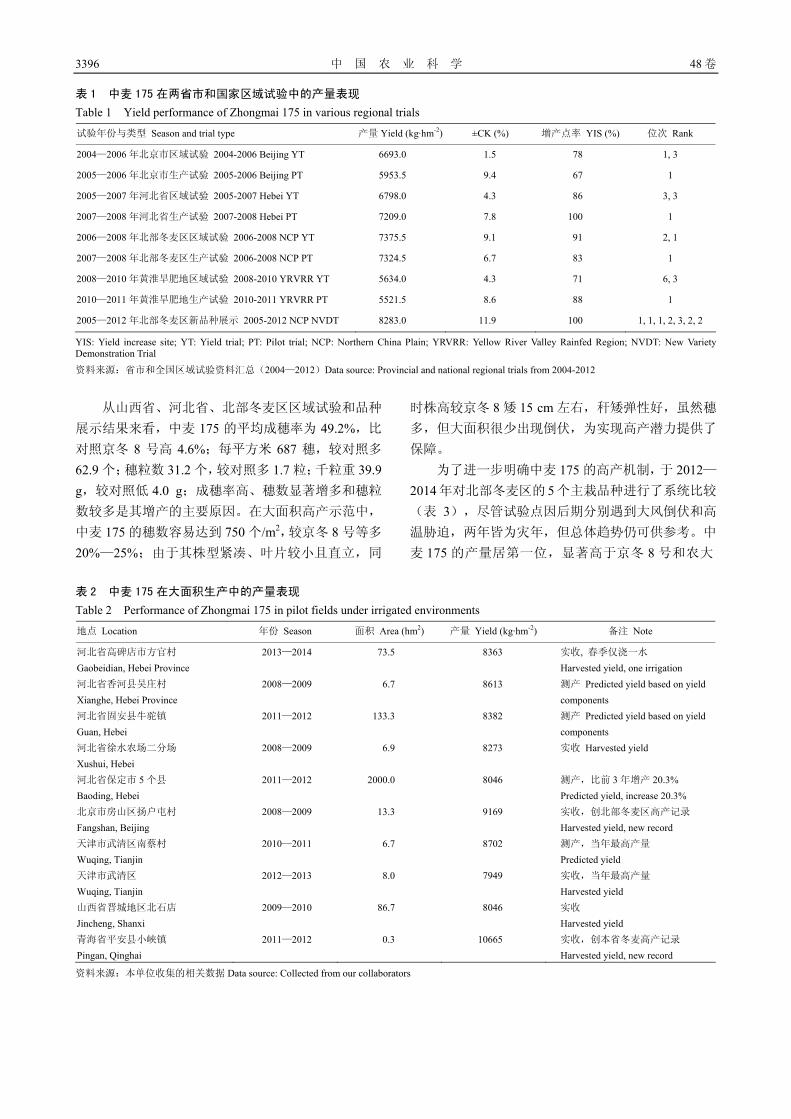
3396 中 国 农 业 科 学 48 卷
表 1 中麦 175 在两省市和国家区域试验中的产量表现
Table 1 Yield performance of Zhongmai 175 in various regional trials
试验年份与类型 Season and trial type 产量 Yield (kg·hm-2) ±CK (%) 增产点率 YIS (%) 位次 Rank
2004—2006 年北京市区域试验 2004-2006 Beijing YT 6693.0 1.5 78 1, 3
2005—2006 年北京市生产试验 2005-2006 Beijing PT 5953.5 9.4 67 1
2005—2007 年河北省区域试验 2005-2007 Hebei YT 6798.0 4.3 86 3, 3
2007—2008 年河北省生产试验 2007-2008 Hebei PT 7209.0 7.8 100 1
2006—2008 年北部冬麦区区域试验 2006-2008 NCP YT 7375.5 9.1 91 2, 1
2007—2008 年北部冬麦区生产试验 2006-2008 NCP PT 7324.5 6.7 83 1
2008—2010 年黄淮旱肥地区域试验 2008-2010 YRVRR YT 5634.0 4.3 71 6, 3
2010—2011 年黄淮旱肥地生产试验 2010-2011 YRVRR PT 5521.5 8.6 88 1
2005—2012 年北部冬麦区新品种展示 2005-2012 NCP NVDT 8283.0 11.9 100 1, 1, 1, 2, 3, 2, 2
YIS: Yield increase site; YT: Yield trial; PT: Pilot trial; NCP: Northern China Plain; YRVRR: Yellow River Valley Rainfed Region; NVDT: New Variety Demonstration Trial
资料来源:省市和全国区域试验资料汇总(2004—2012)Data source: Provincial and national regional trials from 2004-2012
从山西省、河北省、北部冬麦区区域试验和品种
展示结果来看,中麦 175 的平均成穗率为 49.2%,比
对照京冬 8 号高 4.6%;每平方米 687 穗,较对照多
62.9 个;穗粒数 31.2 个,较对照多 1.7 粒;千粒重 39.9 g,较对照低 4.0 g;成穗率高、穗数显著增多和穗粒
数较多是其增产的主要原因。在大面积高产示范中,
中麦 175 的穗数容易达到 750 个/m2,较京冬 8 号等多
20%—25%;由于其株型紧凑、叶片较小且直立,同
时株高较京冬 8 矮 15 cm 左右,秆矮弹性好,虽然穗
多,但大面积很少出现倒伏,为实现高产潜力提供了
保障。 为了进一步明确中麦 175 的高产机制,于 2012—
2014年对北部冬麦区的 5个主栽品种进行了系统比较
(表 3),尽管试验点因后期分别遇到大风倒伏和高
温胁迫,两年皆为灾年,但总体趋势仍可供参考。中
麦 175 的产量居第一位,显著高于京冬 8 号和农大
表 2 中麦 175 在大面积生产中的产量表现
Table 2 Performance of Zhongmai 175 in pilot fields under irrigated environments
地点 Location 年份 Season 面积 Area (hm2) 产量 Yield (kg·hm-2) 备注 Note
河北省高碑店市方官村 Gaobeidian, Hebei Province
2013—2014 73.5 8363 实收, 春季仅浇一水 Harvested yield, one irrigation
河北省香河县吴庄村 Xianghe, Hebei Province
2008—2009 6.7 8613 测产 Predicted yield based on yield components
河北省固安县牛驼镇 Guan, Hebei
2011—2012 133.3 8382 测产 Predicted yield based on yield components
河北省徐水农场二分场 Xushui, Hebei
2008—2009 6.9 8273 实收 Harvested yield
河北省保定市 5 个县 Baoding, Hebei
2011—2012 2000.0 8046 测产,比前 3 年增产 20.3% Predicted yield, increase 20.3%
北京市房山区扬户屯村 Fangshan, Beijing
2008—2009 13.3 9169 实收,创北部冬麦区高产记录 Harvested yield, new record
天津市武清区南蔡村 Wuqing, Tianjin
2010—2011 6.7 8702 测产,当年 高产量 Predicted yield
天津市武清区 Wuqing, Tianjin
2012—2013 8.0 7949 实收,当年 高产量 Harvested yield
山西省晋城地区北石店 Jincheng, Shanxi
2009—2010 86.7 8046 实收 Harvested yield
青海省平安县小峡镇 Pingan, Qinghai
2011—2012 0.3 10665 实收,创本省冬麦高产记录 Harvested yield, new record
资料来源:本单位收集的相关数据 Data source: Collected from our collaborators

17 期 何中虎等:中麦 175 高产高效广适特性解析与育种方法思考 3397
表 3 北部冬麦区 5个主栽品种产量相关性状比较
Table 3 Trait comparison of five major cultivars in North Winter Wheat Region
品种 Cultivar
产量 Yield (kg·hm-2)
穗数
Spikes/m2
穗粒数 Grains/spike
千粒重 TKW (g)
株高 PH (cm)
生物量 Biomass (kg·hm-2)
收获指数 HI
中麦 175 Zhongmai 175 4913a 627a 29.3a 36.4b 77.7b 11040a 42.1a
轮选 987 Lunxuan 987 4759ab 623a 29.1a 36.6b 75.3bcd 11072a 38.3bc
京冬 17 Jingdong 17 4569ab 532b 30.1a 36.2b 76.8bc 9951a 39.8abc
京冬 8 号 Jingdong 8 4480b 534b 27.3a 42.2a 89.2a 10557a 37.4c
农大 211 Nongda 211 4445b 597ab 28.0a 36.6b 74.7cd 11209a 38.2bc
TKW:千粒重;PH:株高;HI:收获指数。下同。同一列不同字母表示差异达 5%显著水平 TKW: Thousand kernel weight; PH: Plant height; HI: Harvest index. The same as below. Different letters in the same column indicate significant difference at 5% probability level
211,也高于公认的高产品种轮选 987;中麦 175 和轮
选 987 穗数显著多于京冬 8 号和京冬 17;中麦 175 的
收获指数为 42.1%,显著高于除轮选 987 外的其他 3个品种。中麦 175 的株高与现有品种接近,但比对照
京冬 8 号降低约 10—15 cm,秆子弹性好,抗倒伏能
力显著提高。进一步分析表明,中麦 175 成熟期干物
质分配到籽粒中的比例高达 39.9%,显著高于其他品
种,如轮选 987 和农大 211 仅为 34.7%和 33.1%。由
于中麦 175 的灌浆期较其他品种仅长 1—2 d,灌浆速
度快是收获指数高的主要原因。在充分灌溉的条件下,
由于中麦 175 株型紧凑、叶片较小且直立,在抽穗前
和灌浆中期的植被指数、抽穗前的叶面积指数显著低
于京冬 8 号,但灌浆中期的叶绿素含量显著高于京冬
8 号,而灌浆中期的叶片衰老速度则显著低于京冬 8号,这可能是中麦 175 收获指数和产量皆高的生理基
础[2]。分子检测表明,中麦 175 不含矮秆基因 Rht1、Rht2 及 Rht8,根据其株高判断,应含有其他矮秆基因,
这有待将来验证;另外它含有 3 个可以增加粒重的等
位基因 TaCwi-A1a、TaSAP1-A1-2606C 和 TaGS-D1a(国家小麦改良中心资料)。因此,株高显著降低、
株型直立、穗数多和收获指数高是其高产的主要原因。 综上所述,株型紧凑、叶片较小且直立,成穗率
高、穗数较一般品种多 10%—25%,株高 80 cm 左右,
抗倒伏能力强,后期叶片衰老速度慢,灌浆速度快,
收获指数高,这是中麦 175 高产的主要原因。
2 水分利用效率高,为水旱兼用型品
种
中麦175的水分利用效率高主要表现在3个方面。
一是在黄淮旱肥地区试中增产显著(表 1),分别通
过北部冬麦区水地和黄淮旱肥地两次国家审定,在建
立国家品种审定制度的 30 多年中,它是唯一一个同时
通过水地和旱地国家审定的品种。二是在石家庄仅浇
1 水的条件下,产量高达 10 609.5 kg·hm-2。石家庄市
农业科学院于 2012—2013 和 2013—2014 年在赵县高
肥力条件下,对近 10 年国家审定的 100 个冬性及半冬
性品种进行了 0 水(全生育期不浇水)、1 水(拔节
期)和 2 水(拔节、抽穗期)试验(3 次重复,小区
面积 6 m2),在全生育期仅浇 1 水的条件下,中麦 175较对照石 4185 增幅高达 9.7%,居 100 个参试品种首
位,石麦 19、西农 558 和石麦 22 的增幅也在 7%以上,
分居 2—4 位。在全生育期浇 2 水的条件下,中麦 175增幅为 5.3%,居 100 个参试品种的第 12 位(郭进考,
2014,个人交流)。这说明中麦 175 不仅产量潜力达
到了黄淮麦区的高产水平,而且实现了高产潜力与节
水性能的良好结合,其产量水平和节水能力皆居参试
品种前列。三是在甘肃和陕西等地的旱地大面积示范
中突破 8 000 kg·hm-2,2014 年还创造了陕西省(9 079.5 kg·hm-2)和甘肃省(9 262.5 kg·hm-2)旱地高产记录(表
4)。甘肃省泾川县大面积示范表明,中麦 175 在大旱
之年和丰水年份皆表现突出,灾年减产少,丰年抗倒
伏、增产幅度大,是解决地膜小麦倒伏的理想品种,非
常适合陇东地区种植[3]。当地老百姓说,种植中麦 175,灾年有粮吃,丰年创高产。在一般干旱的 2010—2011年,中麦 175 与当地主栽品种西峰 27 产量接近,增产
1.4%。在丰水年份如 2011—2012 年,在地膜覆土栽
培条件下,比当地种植多年的长 6359 增产 17.2%;在
露地栽培条件下,比当地主栽品种西峰27增产29.3%,
增产的主要原因是株高较低,抗倒性显著优于当地抗
旱品种。在严重干旱的 2012—2013 年(60 年不遇),
露地条件下的产量与对照品种西峰 27 接近,减产
1.7%;但在地膜覆土栽培条件下,比对照长 6359 增
产 16.1%。据计算,2012—2013 年中麦 175 在地膜栽

3398 中 国 农 业 科 学 48 卷
培条件下的水分利用效率为 1.1 kg·mm-1,而当地主
栽品种泾川 1 号在相同条件下为 0.92 kg·mm-1,西峰
27 在露地栽培下仅为 0.54 kg·mm-1;2013—2014 年
中麦 175 高水分利用效率达 1.50 kg·mm-1,对照品
种仅为 0.85 kg·mm-1。这充分说明中麦 175 为水分高
效及水旱兼用型品种。 为了进一步明确中麦 175 的水分高效利用机理,
在不浇水(W0)、仅浇拔节水(W1)和同时浇拔节
水和开花水(W2)3 种条件下,比较了中麦 175 与北
部冬麦区代表性国审品种京冬 17 的产量等性状(表
5)。在 3 种处理中,中麦 175 的产量和水分利用率皆
高于京冬 17(图 1 及表 5);在不浇水和仅浇拔节水
时,中麦 175 的千粒重和收获指数差异不显著,穗粒
数在 3 种条件下差异不显著,而京冬 17 的所有性状
表 4 中麦 175 在旱肥地的高产表现
Table 4 Performance of Zhongmai 175 in pilot fields under rainfed environments
地点 Location 年份 Season 面积 Area (hm2) 产量 Yield (kg·hm-2) 备注 Note
陕西省永寿县永寿村 Yongshou, Shaanxi
2011—2012 3.0 8745.0 实收,创陕西省旱地高产记录 Harvested yield, new record
陕西省省永寿县 Yongshou, Shaanxi
2013—2014 0.2 9079.5 实收,创陕西省旱地高产纪录 Harvested yield, new record
陕西麟游县九成宫镇 Linyou, Shaanxi
2013—2014 6.7 8053.5 实收,比对照晋麦 47 增 32.7% Harvested yield, increase 32.7%
甘肃省泾川县 Jingchuan, Gansu
2012—2013 6.7 6090.0 实收,比对照泾麦 1 号增 20.8% Harvested yield, increase 20.8%
甘肃省泾川县太平乡 Jingchuan, Gansu
2013—2014 200 7951.5 测产,比对照晋麦 79 增 42.9% Predicted yield, increase 42.9%
甘肃崇信县黄寨乡 Chongxin, Gansu
2013—2014 0.2 9262.5 实收,比对照晋麦 79 增产 51.4%,省旱地高产纪录
Harvested yield, increase 51.4%, new record
资料来源:本单位收集的相关数据 Data source: Collected from our collaborators
表 5 不同灌溉量对中麦 175 和京冬 17 产量相关性状的影响
Table 5 Effect of different irrigations on yield and related traits of Zhongmai 175 and Jingdong 17
灌溉量 Irrigation
穗数 Spikes/m2
穗粒数 Grains/spike
千粒重 TKW (g)
收获指数 HI
生物量 Biomass (kg·hm-2)
产量 Yield (kg·hm-2)
水分利用效率 WUE (kg·mm-1)
0, W0 520b/488b 30.2a/32.4b 45.7b/48.4b 42.9b/39.5c 15827c/14949c 7510c/6620c 21.9b/18.9b
1, W1 637a/619a 30.4a/33.2ab 45.2b/47.7c 44.5b/43.4b 17942b/17352b 8830b/8160b 23.8a/21.3a
2,W2 653a/635a 30.5a/33.9a 46.9a/49.7a 49.4a/45.1a 19078a/18341a 9350a/9050a 22.4b/22.2a
“/”前为中麦 175,后为京冬 17。WUE:水分利用效率。同一列不同字母表示差异达 5%显著水平 Zhongmai 175 is before “/” and Jingdong 17 is below. WUE: Water use efficiency. Different letters in the same column indicate significant difference at 5% probability level
产量
Yie
ld (k
g·hm
-2)
图 1 不同灌溉条件下中麦 175 和京冬 17 产量比较
Fig. 1 Yield comparison between Zhongmai 175 and Jingdong
17 at different irrigations
差异都达显著水平,说明中麦 175 对水分反应相对不
敏感,这与郭进考的结果一致(郭进考,2014,个人
交流)。大面积示范表明,中麦 175 为水分高效型品
种,如河北省高碑店市在 2013—2014 年度种植 73.5 hm2,春季浇 1 水和春季浇 2 水的产量(8 363 kg·hm-2
和 8 520 kg·hm-2)相差无几,但前者的水分利用效率
为 1.21 kg·mm-1,而后者仅为 1.05 kg·mm-1。 在充分灌溉和仅浇冬水条件下的系统比较表明,
中麦 175 产量的水分敏感指数为 0.86,而京冬 8 号为
1.13,主要原因是千粒重与株高对水分反应敏感性低,
虽然穗数对水分的敏感性较高,但其数值仍高于京冬
8 号,这是其在有限灌溉条件下高产的主要原因[2]。中
麦 175 携带较多的京 411 优异根系遗传区段,主根长

17 期 何中虎等:中麦 175 高产高效广适特性解析与育种方法思考 3399
和根干物质重量等的改良较为显著,3DL 和 5BL 携带
控制根长的主效位点[4]。优良的生理和根系特性为水
分高效利用奠定了基础。 综上所述,中麦 175 穗数多,株高、千粒重、穗
粒数和收获指数对水分胁迫敏感性低,灌浆中后期叶
绿素含量高,叶片衰老速度慢,这是其水分高效、在
节水条件下表现突出的原因,中麦 175 为水旱(肥地)
兼用型高产新品种。
3 肥料利用效率高
据中国科学院遗传发育研究所对北部和黄淮麦区
64 份主要品种氮和磷利用效率的研究,中麦 175 在不
施氮、不施磷及氮和磷皆不施的 3 种处理中,产量分
别居参试品种的第 1、第 9 和第 1 位,说明中麦 175为肥料高效型品种(童依平,2012,个人交流)。
为了进一步确认这一特性并明确其机理,设置 6种氮肥处理,对中麦 175 与京冬 17 进行系统比较。在
不同施氮水平下,中麦175的产量均高于京冬17(图2)。从表 6 可以看出,氮肥施用量对中麦 175 的穗数影响
大,但穗粒数和千粒重在不同处理间差异不显著,而氮
肥施用量对京冬 17 的产量三要素均有显著影响。在不
同施氮水平下,中麦 175 氮肥利用率即吸收效率都高于
京冬 17;在 60 kg 和 120 kg 2 个低氮条件下,中麦 175氮肥利用效率和农学效率显著高于京冬 17,其中前者
氮肥利用效率分别为 25.2 和 18.3 kg·kg-1,后者分别为
6.6 和 7.3 kg·kg-1。另有研究表明,京 411 及其 14 个衍
生品种在正常施肥和常年不施肥条件下进行比较,中麦
175 的产量皆为 高,主要原因是穗数多,产量三因素
对肥料的敏感性相对较低[5]。上述结果皆说明中麦 175的确是一个肥料高效型品种,其氮肥吸收和利用效率都
较高。研究还表明,中麦 175 在苗期具有较强的耐低磷
能力,6A、3B 和 4BS 染色体对苗期氮和磷利用效率均
有影响,1B和2AL染色体位点表现较强的耐低磷能力,
4B 染色体 BS00022177_51 附近的标记密集区与氮利用
表 6 不同氮肥水平对中麦 175 和京冬 17 产量及氮肥利用效率相关性状的影响
Table 6 Effect of different N fertilizer levels on yield and nitrogen use efficiency related traits of Zhongmai 175 and Jingdong 17
施氮肥量 N rate (kg·hm-2)
穗数 Spikes/m2
穗粒数 KPS
千粒重 TKW (g)
产量 Yield (kg·hm-2)
氮肥利用率 NRE (%)
氮肥利用效率 NUtE (kg·kg-1)
氮肥农学效率 NAE (kg·kg-1)
0 634c/606b 30.5a/32.4c 47.3a/49.9ab 8970c/8410d 0/0 0/0 0/0
60 692a/613ab 30.8a/33.9ab 47.2a/50.3ab 9380b/8510cd 0.27a/0.24a 25.2a/6.6c 6.9a/1.6b
120 703a/609ab 31.5a/33.2abc 47.4a/49.7b 9410b/8570cd 0.20bc/0.18b 18.3b/7.3c 3.7b/1.3bc
120+60 653bc/635a 30.5a/33.9ab 46.9a/49.7b 9350b/9050b 0.21b/0.18b 9.9c/19.3b 2.1bc/3.5a
120+120 704a/624ab 31.5a/34.4a 47.1a/51.0a 9720a/9390a 0.18cd/0.17b 17.5/b23.4a 3.1bc/4.1a
120+180 678ab/599b 30.5a/32.9bc 47.3a/50.2ab 9330b/8650c 0.16d/0.15b 7.4c/5.2c 1.2c/0.8c
同一品种的不同处理间的不同字母表示在 0.05 水平差异显著。KPS:穗粒数;NRE:氮肥利用率;NUtE:氮肥利用效率;NAE:氮肥农学效率 Values followed by different letters are significantly different at the 0.05 probability level. KPS: Kernels per spike; NRE: Apparent recovery efficiency of applied N; NUtE: N utilization efficiency; NAE: N agronomy efficiency
产量
Yie
ld (k
g·hm
-2)
图 2 中麦 175 和京冬 17 在不同施肥条件下的产量
Fig. 2 Grain yield of Zhongmai 175 and Jingdong 17 in
different N treatments
效率、耐低氮能力和磷利用效率等有关[6]。 综上所述,中麦 175 氮肥吸收即利用率高,在低
氮条件下的氮肥利用效率和农学效率都高;在不同施
肥水平下的产量都高于对照京冬 17,说明它实现了高
产潜力与肥料高效利用率的良好结合,是肥料高效型
高产品种。
4 加工品质和营养品质优良
4.1 面条和馒头兼用型品质
14 份大田样品测试表明,中麦 175 籽粒蛋白质含
量(11.6%,14%湿基)中等,籽粒硬度较低(21.1SKCS单位),出粉率较高(平均 73.7%),吸水率、形成
时间、稳定时间、延展性、 大抗延阻力分别为 52.7%、

3400 中 国 农 业 科 学 48 卷
1.7 min、2.6 min、183.4 mm 和 77.0 BU,属于软质中
弱筋、延展性好的类型[7]。分子标记检测表明,中麦
175 不含 1BL·1RS 易位系,高分子量麦谷蛋白亚基分
别为 null、7+9 和 2+12,低分子量麦谷蛋白亚基分别
为 Glu-A3c 和 Glu-B3h,这是面筋强度弱、延展性好
的主要原因。面粉 L*值较高(92.26)、a*值(-1.27)和 b*值(7.13)较低,面片 L*值较高(82.74)、a*值(0.30)和 b*值(21.23)较低,说明面粉和面片白
度均较高。中麦 175 含有黄色素含量较高和较低基因
Psy-A1a 和 Psy-B1b,表现良好的面粉和面片色泽。 中麦 175 馒头品质优异,特点是体积大、形状好、
质地优良,14 份样品平均总分为 85.3 分,而对照品种
京 411 馒头总分为 75.0 分。面条品质优良,特点是口
感、颜色和黏弹性均好,平均总分 70.9 分,高于对照
样品商业雪花粉(70.0 分)[7]。从表 7 可以看出,在
同一环境下,中麦 175 的馒头和面条品质均显著优于
表 7 中麦 175、京冬 8号和轮选 987 的面条和馒头加工品质比较
Table 7 Comparison of noodle and steamed bread qualities of Zhongmai175, Jingdong 8, and Lunxuan 987
类型 Food type
特性 Character
中麦 175 Zhongmai 175
京冬 8 号 Jingdong 8
轮选 987 Lunxuan 987
雪花粉 Xuehua
色泽(15) Color 9.8 7.5 9.0 10.5
表面状况(10) Appearance 7.5 7.0 6.0 7.0
软硬度(20) Firmness 13.0 13.0 14.0 14.0
粘弹性(30) Viscoelasticity 21.0 16.5 18.0 21.0
光滑性(15) Smoothness 11.3 9.0 8.3 10.5
食味(10) Flavor 8.0 6.5 8.5 7.0
面条 Noodle
总分(100) Total score 70.6 59.5 63.8 70.0
比容(20) Specific volume 19.0 19.0 16.0 15.0
外形(10) Shape 9.0 7.0 8.0 8.0
表面光滑(10) Smoothness 8.0 9.0 8.0 8.0
表面色泽(10) Skin color 10.0 8.0 4.0 10.0
压缩张弛性(35) Stress relaxation 31.0 33.0 33.0 35.0
结构(15) Structure 13.5 10.5 10.5 12.0
馒头 Steamed bread
总分(100) Total score 90.5 86.5 79.5 88.0
北部冬麦区的代表性品种京冬 8 号和轮选 987。
中麦 175 的糯蛋白基因皆为野生型,即 Wx-A1a、Wx-B1a 和 Wx-D1a,但其大淀粉粒重量占 61.5%,小
淀粉粒重量占 38.5%,适宜的淀粉粒度分布可能是其
馒头和面条品质优良的重要原因。用中麦 175 进行的
大小淀粉粒重组试验表明,小淀粉粒含量为 30%—
35%的面粉制作的馒头内部结构 好、总评分 高,
小淀粉粒含量为 30%—40%的面粉制作的面条软硬度
适宜[8]。中麦 175 的抗性淀粉(63.0%)显著高于北
京 0045(57.3%)[9]和其他品种,说明它具有一定的
保健功能。 4.2 锌含量高,营养健康价值高
缺铁性贫血在很大程度上是由于缺锌引起的,锌 缺乏是终极意义上的“隐性饥饿”,培育和推广富锌
作物品种具有重要意义[10]。将 Zn 含量较高的 24 个品
种在 7 个地点种植两年,河农 326(58.2±7.3)、冀麦
26(51.9±4.1)、农大 3197(47.8±3.5)、京冬 8 号
(46.1±2.5)和中麦 175(45.8±2.7)的锌含量居前 5位[11]。除中麦 175 外,其他 4 个品种目前的种植面积
皆很小。 在不施锌肥的条件下,中麦 175 和良星 99 开花期
的根长和根表面积及籽粒锌收获指数差异不显著,但
在施锌肥的条件下,中麦 175 开花期的根长和根表面
积及籽粒锌收获指数均显著高于良星 99,说明土壤施
锌肥可显著提高锌向其籽粒转移,而良星 99 则变化不
明显,因而中麦 175 籽粒锌浓度(51.7 mg·kg-1)高于
良星 99(46.8 mg·kg-1)(邹春琴,个人交流)。这表
明中麦 175 对土壤中锌的吸收转运效率较高。 总之,中麦 175 的特点是软质,面筋强度中等偏
弱、延展性好,大小淀粉粒分布比例适宜,面粉颜色
白,为面条和馒头兼用型优质品种,对土壤中锌的吸
收转运效率高,营养健康价值高。

17 期 何中虎等:中麦 175 高产高效广适特性解析与育种方法思考 3401
5 适应性广泛
中麦 175 的审定和推广区域包括北京、天津、河
北、山西、山东、河南西部、陕西、甘肃及青海,跨
9 个省(市)4 个麦区,已成为北部冬麦区水地及甘肃
和青海春麦改冬麦地区的第一大品种,为黄淮旱肥地
及陇东旱地的主栽品种,累计推广面积近 120 万 hm2,
其中,2014 年秋播约 33 万 hm2,这足以说明它具有
非常广泛的适应性。分子检测表明,中麦 175 含有效
应较大的光周期不敏感基因 Ppd-D1b,这可能是其广
适性的分子基础。 中麦 175 为强冬性品种,春化基因组成为 vrn-A1、
vrn-B1、vrn-D1 和 vrn-B3,抗寒性居目前本麦区推广
品种之首。2009—2010 年北部冬麦区遇到了罕见的低
温危害,大幅度减产,但中麦 175 仍获好收成,天津
武清区 6.7 hm2示范方实收产量高达 7 890 kg·hm-2,为
天津市当年 高记录。随着全球气候变暖,在春麦区
改种冬麦不仅能大幅度提高产量,而且熟期显著提前,
更重要的是能避开春小麦用水高峰,便于统筹安排作
物生产。春麦改种冬麦的基本要求是品种越冬性过
关[12]。由于抗寒性和产量等表现突出,中麦 175 已成
为青海省黄河谷地和湟水河流域的川水地区及甘肃张
掖、武威等地的第一大冬麦品种。 中麦 175 含 Eps,早熟性突出,比黄淮旱肥地品
种早熟 3—5 d;籽粒偏小,灌浆速度特别快,能避开
后期高温胁迫。采用花后 12 d 覆盖塑料薄膜进行高温
胁迫处理,在高温处理与非处理间白天(6:00—18:00)温差为 2.2℃(中午极端温差 高可达 10℃以上,一
般可达 5℃以上)的情况下,中麦 175 灌浆速度受高
温影响很小(图 3),其热敏感指数显著低于石优 17和衡观 35,与耐高温品种石麦 15 和京冬 8 号接近。
条锈病、叶锈病和白粉病轻,同时株高较当地品种矮
20—30 cm 且在不同水分条件下变幅小,茎秆强度好,
在灾年和丰水年皆表现突出,丰水年抗倒伏、创高产,
这是它在黄淮旱肥地及陇东大面积推广的重要原因。
还表现很强的耐晚播能力,2013—2014 年度河南洛宁
县播期较正常年份推迟 1 个月,株高仍达 75 cm,较
其他品种增产 10%以上。 中麦 175 苗期对当前流行小种条中 32、条中 29
表现高抗,成株期对水 4、条中 29、条中 32 表现免疫
至中抗,对水 7、HY8、条中 33 表现中抗。对条中 29的抗性由一对位于 2AS 染色体的显性基因控制,目前
正在寻找与其紧密连锁的 SNP 标记。
千粒重
TK
W (g
)
图 3 中麦 175 在田间正常温度和高温胁迫下灌浆速率的
Logistic 拟合曲线
Fig. 3 Logistic curve of grain filling rate of Zhongmai 175
under normal and heat stress conditions
6 育种经验分析与未来设想
6.1 育种经验分析
自 20 世纪 80 年代末育成京 411 和京冬 8 号以后,
北部冬麦区的育种工作没有太大进展,主要表现在新
品种植株偏高易倒伏,产量没有明显突破,同时对肥
水要求过高。本区灌浆期光温充足,有利于增加粒重;
但穗分化时间短,提高穗粒数主要依靠减少不孕小穗
数和提高结实性来实现;同时对抗寒性和后期抗高温
的要求很高,白粉病和条锈病则是主要病害[13-14]。据
此,确定的具体策略是,以中高水肥地区为主,通过
降低株高和株型改良来显著提高抗倒性;在保持较多
穗数的基础上,通过增加粒数或粒重来提高产量潜力;
加强育成品系的节水节肥性能筛选,保证新品种既能
大面积创高产,又能够适应目前的粗放管理。要实现
这一目标,需抓住 2 个关键环节,一是组合配置,二
是后代选择与鉴定。 京 411 的抗寒性居本区推广品种的前列,产量潜
力也高于京冬 8 号,穗数多(600—680 穗/m2),株
高 90—95 cm,株型较紧凑,穗层整齐,适应性广[14]。
虽然其面筋强度较弱,但手工制作的馒头和面条品质
优良。缺点是植株偏高,在高产地块有倒伏现象,后
期易早衰,造成粒重不稳,同时感白粉病和条锈病,
机器收获时籽粒破碎率较高。自 20 世纪 80 年代后期
的 10 多年,京 411 一直是高产骨干亲本[13]。针对京
411 的不足,选用了中国农业大学培育的新品系
BPM27(组合为 20102//津 441/Fr81-4)为母本,其突
出特点是秆硬、矮秆抗倒伏(约 70 cm)、株型紧凑、

3402 中 国 农 业 科 学 48 卷
穗较大码密、早熟、兼抗白粉和条锈病,于 1995 年配
置单交组合,双亲的优点多且互补性很好。采用系谱
法经6年选择育成,于2000—2001年度参加产量鉴定,
编号为 CA0175 即中麦 175,它聚合了双亲的主要优
点,基本实现了预期目标[14]。意外收获是水肥利用效
率和适应性远远超过了双亲,原因还有待进一步研究。
当然,中麦 175 也继承了京 411 的一些缺点,如机器
收获时籽粒破碎率较高和白粒易穗发芽。 在早期世代,注重株型和穗育性的选择,即要求
株型紧凑、叶片小且直立,以便容纳更多穗数;穗部
性状突出码多、结实性和多粒的选择,为高产稳产提
供保障。重点加强了育成品系的多点鉴定与大面积示
范,不断扩大审定和推广地区。2002—2003 年度的产
量比较试验在 4 个地点进行,中麦 175 全部增产,比
对照京 411 平均增产 9.9%,故将其作为重点品系参加
各级区域试验。由于在北京、山西和北部冬麦区的区
域试验中皆表现突出,均提前一年参加生产试验,在
尚未审定之前,在河北徐水农场进行大面积示范,发
现其产量表现和适应性明显超过同期的其他品种。另
外,在山西晋城的大面积示范中发现其水分利用效率
高,有可能在黄淮旱肥地利用,于是当年秋播试着
参加黄淮旱肥地区域试验并 终通过审定,使推广
区域显著扩大。中麦 175 在青海省的发展则是借鉴
京 411 的推广经验,在甘肃省河西的发展主要得益
于当地春麦改冬麦对品种抗寒性的要求。基于中麦
175 的成功经验,目前的产量比较试验在 6 个地点
进行,覆盖北京市、天津市、河北省和山西省 4 省
区,还在昌平增设 1 个仅浇越冬水的旱地环境,
近又在新疆增加了 2 个试验点,这样就形成了较完
善的品种比较网络,这是近年来育种工作取得较快
进展的重要因素。中麦 175 的育种和推广实践表明,
通过株型改良培育多穗型高产抗倒伏广适性品种完
全可行,培育水旱兼用的半矮秆品种也是可行的,
其关键指标是株型直立、株高适中和穗数多,这与
玉米的直立耐密性杂交种有其相似之处[15]。 6.2 高产广适性品种培育的初步设想
中国小麦生产面临严峻挑战,在气候变化和纹枯
病及赤霉病日益加剧的情况下[16],既要保证较高产量
和可接受的品质,还要降低水肥等投入,以提高产业
竟争力并保护环境,这在缺水十分突出的北部冬麦区
和黄淮北片显得更为重要。因此,除部分地区继续加
强高产更高产甚至超高产品种培育之外,大部分地区
应把培育高产、水肥高效、抗病、抗逆、广适性品种
作为主要任务[17],当然优质面条和馒头及专用面包品
质等还要继续重视。就育种技术而言,尽管小麦测序
已取得较大进展,用于品质和抗病性的分子育种技术
已经成熟并在国内外开始应用[18],全基因组选择在其
他作物中也已取得阶段性进展[19],但考虑到产量、水
肥利用效率及抗热性等的复杂性及表型鉴定的巨大工
作量,在近期内发掘出效应较大、育种家可用的基因
特异性标记的难度相当大,因此,高产高效广适性品
种的培育仍需主要依靠常规育种技术。如前所述,常
规育种成功的关键是组合配置和后代的准确选择与鉴
定,考虑到实际育种中水肥管理的调节难度不小,鉴
于国内外的广适性品种包括适于旱地的大面积品种多
是从水地品种中筛选出来的现实,加强亲本和育成品
系品种的系统鉴定至关重要。初步设想是,首先根据
大面积生产表现和系统可靠的鉴定,明确高产、水肥
高效、抗热及抗病等亲本,在此基础上合理配置组合。
需要说明的是,石家庄市农业科学院对主要品种节水
性能的系统鉴定值得学习和参考,产量、品质和抗病
性等都需做类似的工作。除适当扩大分离群体的规模
外,在早期分离世代如 F2—F4仍可沿用过去的成熟做
法。其次,要尽量扩大产量鉴定和品种比较试验的规
模和点次,可在分离世代的后期如 F5—F6尤其是产量
鉴定及品种比较阶段加强多点和多环境鉴定,至少一
个乃至几个点可为 1—2 水的有限灌溉和中低肥/不施
肥条件,目的是发现既高产又高效广适的“超级品
种”。这是 CIMMYT 及国际小麦育种的成功经验,
国内应借鉴学习。第三,在高世代材料鉴定中,应及
时大胆采用一些可用的分子标记,以便为表型鉴定补
充有益信息。经过长期不懈努力,全国定能培育出一
批高产高效广适性新品种,为小麦产业发展作出新的
贡献。
References
[1] 万富世. 冬小麦品种越冬性改良理论与实践. 北京: 中国农业科技
出版社, 2006.
Wan F S. Theory and Practice of Cold Tolerance Improvement in
Winter Wheat. Beijing: China Agri-technology Press, 2006. (in Chinese)
[2] 李兴茂, 倪胜利. 不同水分条件下广适性小麦品种中麦 175的农艺
和生理特性解析. 中国农业科学, 2015, 已接受.
Li X M, Ni S L. Agronomic and physiological characterization of
wide adaptable wheat cultivar Zhongmai 175 under two different
irrigation conditions. Scientia Agricultura Sinica, 2015, in press. (in
Chinese)

17 期 何中虎等:中麦 175 高产高效广适特性解析与育种方法思考 3403
[3] 史聚宝, 李兴茂, 孟治岳. 小麦全膜覆土穴播技术存在的问题与对
策. 作物杂志, 2014, 1: 129-131.
Shi J B, Li X M, Meng Z Y. Constrains and strategy for plastic mulch
technology in wheat. Crops, 2014, 1: 129-131. (in Chinese)
[4] 肖永贵, 路亚明, 闻伟锷, 陈新民, 夏先春, 王德森, 李思敏, 童依
平, 何中虎. 小麦骨干亲本京 411 及衍生品种苗期根部性状的遗传.
中国农业科学, 2014, 47(15): 2916-2926.
Xiao Y G, Lu Y M, Wen W E, Chen X M, Xia X C, Wang D S, Li S M,
Tong Y P, He Z H. Genetic contribution of seedling root traits among
elite wheat parent Jing 411 to its derivatives. Scientia Agricultura
Sinica, 2014, 47(15): 2916-2926. (in Chinese)
[5] 肖永贵, 李思敏, 李法计, 张宏燕, 陈新民, 王德森, 夏先春, 何中
虎. 两种施肥环境下冬小麦京 411 衍生系的产量和生理性状分析.
作物学报, 2015, 41(9): 1333-1342.
Xiao Y G, Li S M, Li F J, Zhang H Y, Chen X M, Wang D S, Xia X C,
He Z H. Genetic analysis of yield and physiological traits among
genotypes derived from elite parent Jing 411 under two under
fertilizer environments. Acta Agronomica Sinica, 2015, 41(9):
1333-1342. (in Chinese)
[6] 李法计, 肖永贵, 金松灿, 夏先春, 陈新民, 汪洪, 何中虎. 京 411
衍生系苗期氮和磷利用效率相关性状的遗传解析. 麦类作物学报,
2015, 35(6): 737-746.
Li F J, Xiao Y G, Jin S C, Xia X C, Chen X M, Wang H, He Z H,
Genetic analysis of nitrogen and phosphorus use efficiency related
traits at seeding stage of Jing 411 and its derivatives. Journal of
Triticeae Crops, 2015, 35(6): 737-746. (in Chinese)
[7] 张艳, 陈新民, 绍凤成, 王德森, 何中虎. 中麦 175 馒头和面条品
质稳定性分析. 麦类作物学报, 2012, 32(3): 440-447.
Zhang Y, Chen X M, Shao F C, Wang D S, He Z H. Analysis on
quality stability of steamed bread and noodle by wheat cultivar
Zhongmai 175. Journal of Triticeae Crops, 2012, 32(3): 440-447. (in
Chinese)
[8] Guo Q, He Z H, Xia X C, Qu Y Y, Zhang Y. Effects of wheat starch
granule size distribution on qualities of Chinese steamed bread and
raw white noodle. Cereal Chemistry, 2014, 91(6): 623-630.
[9] Yu J L, Wang S J, Wang J R, Li C L, Xin Q W, Huang W, Zhang Y, He
Z H, Wang S. Effect of laboratory milling on properties of starches
isolated from different flour millstreams of hard and soft wheat. Food
Chemistry, 2015, 172: 504-514.
[10] 郝元峰 , 张勇 , 何中虎 . 作物锌生物强化研究进展 . 生命科学 ,
2015, 27(8): 1047-1054.
Hao Y F, Zhang Y, He Z H. Progress in Zinc biofortification of crops.
Life Science, 2015, 27(8): 1047-1054. (in Chinese)
[11] Zhang Y, Song Q C, Yan J, Tang J W, Zhao R R, Zhang Y Q, He Z H,
Zou C Q, Ortiz-Monasterio I. Mineral element concentrations in
grains of Chinese wheat cultivars. Euphytica, 2010, 174: 303-313.
[12] 何中虎, 夏先春, 陈新民, 庄巧生. 中国小麦育种进展与展望. 作
物学报, 2011, 37(2): 202-215.
He Z H, Xia X C, Chen X M, Zhuang Q S. Progress of wheat breeding
in China and the future perspective. Acta Agronomica Sinica, 2011,
37(2): 202-215. (in Chinese)
[13] 周阳, 何中虎, 陈新民, 王德森, 张勇, 张改生. 30 余年来北部冬
麦区小麦品种产量改良遗传进展 . 作物学报 , 2007, 33(9):
1530-1535.
Zhou Y, He Z H, Chen X M, Wang D S, Zhang Y, Zhang G S. Genetic
gain of wheat breeding for yield in northern winter wheat zone over 30
years. Acta Agronomica Sinica, 2007, 33(9): 1530-1535. (in Chinese)
[14] 陈新民, 何中虎, 王德森, 庄巧生, 张运宏, 张艳, 张勇, 夏先春. 利
用京 411 为骨干亲本培育高产小麦新品种. 作物杂志, 2009, 4: 1-5.
Chen X M, He Z H, Wang D S, Zhuang Q S, Zhang Y H, Zhang Y,
Zhang Y, Xia X C. Experience in developing high yielding varieties
from core parent Jing 411. Crops, 2009, 4: 1-5. (in Chinese)
[15] Li Y, Ma X L, Wang T Y, Li Y X, Liu C, Liu Z Z, Sun B C, Shi Y S,
Song Y C, Carlone M, Bubeck D, Bhardwaj H, Whitaker D, Wilson W,
Jones E, Wright K, Sun S K, Niebur W, Smith S. Increasing maize
productivity in China by planting hybrids with germplasm that
responds favorably to higher planting densities. Crop Science, 2011,
51: 2391-2400.
[16] He Z H, Joshi A K, Zhang W J. Climate Vulnerability and Wheat
Production. Climate Vulnerability: Understanding and Addressing
Threats to Essential Resources, Elsevier Inc., Academic Press,
Waltham, USA, 2013: 57-67.
[17] He Z H, Xia X C, Peng S B, Lumpkin T. Meeting demands for
increased cereal production in China. Journal of Cereal Sciences,
2014, 59: 235-244.
[18] Liu Y N, He Z H, Appels R, Xia X C. Functional markers in wheat:
Current status and future prospects. Theoretical and Applied Genetics,
2012, 125: 1-10.
[19] Huang X H, Wei X H, Sang T, Zhao Q, Feng Q, Zhao Y, Li C Y, Zhu
C R, Lu T T, Zhang Z W, Li M, Fan D L, Guo Y L, Wang A H, Wang
L, Deng L W, Li W J, Lu Y Q, Weng Q J, Liu K Y, Huang T, Zhou T
Y , Jing Y F, Li W, Lin Z, Buckler E S, Qian Q, Zhang Q F, Li J Y,
Han B. Genome-wide association studies of 14 agronomic traits in
rice landraces. Nature Genetics, 2010, 42: 961-969.
(责任编辑 李莉)

中国农业科学 2015,48(17):3404-3414 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.008
收稿日期:2015-06-25;接受日期:2015-08-07 基金项目:国家公益性行业(农业)专项(201203029)、国家现代农业产业技术体系建设专项资金(CARS-01-26)、国家自然科学基金(31471416)、
浙江省自然科学基金(Y13C130013) 联系方式:朱德峰,Tel:13388608096;E-mail:[email protected]
中国水稻高产栽培技术创新与实践
朱德峰,张玉屏,陈惠哲,向 镜,张义凯
(中国水稻研究所,杭州 310006)
摘要:水稻是中国主要粮食作物,也是单产最高的粮食作物。文章利用中国农业统计年鉴数据和国内外文献
资料,分析了中国水稻高产栽培技术创新与实践的成功经验、存在问题和稻作技术发展趋势。探讨了中国水稻生
产经历的 4 个阶段的特点,良种良法配套对水稻增产的贡献,不同阶段创新的稻作技术;简述了全球水稻栽培技
术发展的特点;回顾了中国矮秆品种、杂交稻及超级稻品种更替及其配套栽培技术创新;剖析了水稻高产栽培存
在的新问题;对水稻高产栽培技术的未来发展进行了展望。创新与品种生育特性配套的水稻种植制度、生产模式
和环境协调的栽培技术,发挥品种产量潜力和应用,可以实现增产增效,促进水稻产业发展,改善生态环境和提
高资源利用效益。
关键词:水稻;高产;株型;栽培;技术
Innovation and Practice of High-Yield Rice Cultivation
Technology in China ZHU De-feng, ZHANG Yu-ping, CHEN Hui-zhe, XIANG Jing, ZHANG Yi-kai
(China National Rice Research Institute, Hangzhou 310006)
Abstract: Rice is a staple grain crop in China and also the highest yielding one in grain crops. Using Chinese agricultural statistical data and the literature, this paper analyzed the successful experience of innovation and practice, problems and development trend in high-yield cultivation technology in rice, the characteristics of four periods of rice production in China, the contribution of supporting cultivation technology for variety to yield increase, and high-yield cultivation techniques developed in different periods. The characteristics of rice cultivation technology in world are briefly expounded. Supporting cultivation technology for dwarf varieties, hybrid rice, and super rice in the evolution of rice variety and rice industry development was reviewed. The current problems of high yielding cultivation of rice was analyzed. The development of high-yield rice cultivation technology was also discussed. Innovation of supporting cultivation technology based on the growth characteristics of new variety, rice cropping system, rice production models, and ecological environment exploits yield potential of variety and realizes the increase of yield and profit, promotes the development of the rice industry, and improves the environment and raises the efficiency of resource utilization.
Key words: rice; high yield; plant type; cultivation; technology
0 引言
水稻是中国主要粮食作物,2004 年以来的 10 年
中,水稻种植面积和总产量分别占粮食作物面积和总
产的 27.4%和 36.1%。水稻、玉米和小麦是中国的三
大粮食作物,其中水稻是单产 高的作物,统计表明
全国水稻平均单产较玉米和小麦分别提高 39%和
19%。稻米作为中国主要口粮,全国有 60%的人口食
用。全国除西藏和青海的水稻种植面积较小,其他各
省均种植一定面积的水稻,其种植区域广泛、种植制
jia
文本框
返回目录

17 期 朱德峰等:中国水稻高产栽培技术创新与实践 3405
度多种、品种类型多样,对中国粮食安全、生态安全
和稻农增收具有重要作用。中国也是全球主要的水稻
生产国,近年其水稻面积占全球 18.5%,仅次于印度,
水稻总产占全球 27.7%,居全球首位,水稻单产高于
全球平均单产 50%。中国的稻作技术进步和发展对全
球水稻生产发展做出了重要贡献。
中国水稻面积自 1949 年到 2013 年从 2 571×104 hm2提高到 3 031×104 hm2,增长 460×104 hm2,增幅
18%;单产从 1.89 t·hm-2 提高到 6.72 t·hm-2,提高 4.83 t·hm-2,增幅 255%;总产从 4 860×104 t 提高 2 0340×104 t,增长 15 440×104 t,增幅 319%。良种良法配
套及栽培技术创新对水稻单产提高和总产增长作出重
要贡献。近几十年来,中国社会经济发展、农业结构
调整及农村劳动力向其他产业转移,水稻生产面临稻
田面积和双季稻面积下降、水资源短缺、土壤结构变
差及肥力下降、自然灾害频发、生产规模小且分散,
制约了水稻生产高产高效发展。稻作技术需要转型
升级,适应水稻品种更替、生产规模、经营方式和
种植制度变化,适应提高肥料、农药和水资源利用
效率的要求及应对自然灾害的发生。分析中国水稻
高产栽培技术创新与实践的成功经验、存在问题及
发展趋势,探讨水稻高产栽培技术的发展历程,为
创新现代水稻生产经营方式,高产高效和生态安全
的稻作技术提供借鉴。
1 高产栽培技术的贡献
1.1 中国水稻生产发展
根据中国农业统计年鉴数据分析[1],中国水稻生
产经历 4 个阶段(表 1),1949—1961 年为水稻面积、
单产和总产缓慢增长阶段,也是水稻生产波动较大的
时期,这一期间水稻面积、单产和总产年增长分别为
4.73×104 hm2,12.5 kg·hm-2 和 41.7×104 t。1961—1997年为水稻面积、单产和总产快速增长阶段,这一期间
水稻面积、单产和总产年增长分别为 15.25×104 hm2,
118.9 kg·hm-2 和 408.6×104 t。1997—2003 年为水稻面
积、单产和总产下降阶段,这一期间水稻面积、单产
和总产年下降分别为 87.63×104 hm2,53.1 kg·hm-2
和 668.0×104 t。从 2003—2013 年为水稻面积、单产
和总产恢复增长阶段,水稻面积、单产和总产年增长
分别为 38.04×104 hm2,65.7 kg·hm-2 和 429.6×104 t。从 1997—2003 年的 6 年中,水稻面积下降 548.7×104 hm2,其中双季稻面积下降 548.5×104 hm2,单季稻
增长 22.7×104 hm2,种植季节上主要是双季稻面积
下降。进一步分析稻田面积变化和双季稻面积下降
表明,65%水稻面积减少来自稻田面积下降,35%来自双季稻改种单季稻引起双季稻面积下降。从
2004—2013 年,水稻种植面积由 2 837.88×104 hm2
增加到 3 031.17×104 hm2,增长 6.8%;稻谷单产
从 6 310.5 kg·hm-2 增加到 6 717 kg·hm-2,增长 6.4%;
稻谷总产量从17 908.8×104 t增加到20 361.2×104 t,增长 13.7%。从水稻单产提高贡献因素看,一是水稻
品种改良贡献。品种株型、抗病虫性、耐肥抗倒性、
农艺性状及适应性改良,单产潜力提高。二是水稻栽
培技术创新贡献。包括育秧技术、种植方式、群体调
控、肥水管理、病虫草害及灾害防控及机械化装备等
改进。三是水稻生产农资保障共享。化肥、农药、调
节剂、农膜及机械装备等的进步,应用效果和效率提
高。四是农业政策的支撑,大幅提高种粮积极性和生
产效益。近 10 年来,随着种植结构的调整、区域性
灾害的频发、种稻农民的变化,水稻种植面临巨大的
挑战,栽培技术的创新对水稻面积和单产的稳定作用
尤为突出。
表 1 中国不同阶段水稻面积、单产和总产年均增减量
Table 1 Rice planting area, yield and product in different periods in China
阶段 Period 年数 Years 面积 Area(×104 hm2) 单产 Yield(kg·hm-2) 总产 Production(×104 t)
1949-1961 12 4.73 12.5 41.7
1961-1997 36 15.25 118.9 408.6
1997-2003 6 -87.63 -43.1 -668.0
2003-2013 10 38.04 65.7 429.6
1.2 水稻栽培技术的稳定和提高水稻产量的作用
中国水稻单位面积产量从 1949 年的不足 2 t·hm-2
提高到 2013 年 6.72 t·hm-2,单产提高接近 3.5 倍。随
科学技术的进步,良种是增产的内因,水稻产量的提
高需要产量潜力更大的品种,但品种增产潜力的发挥
需要栽培技术的配套才能实现。
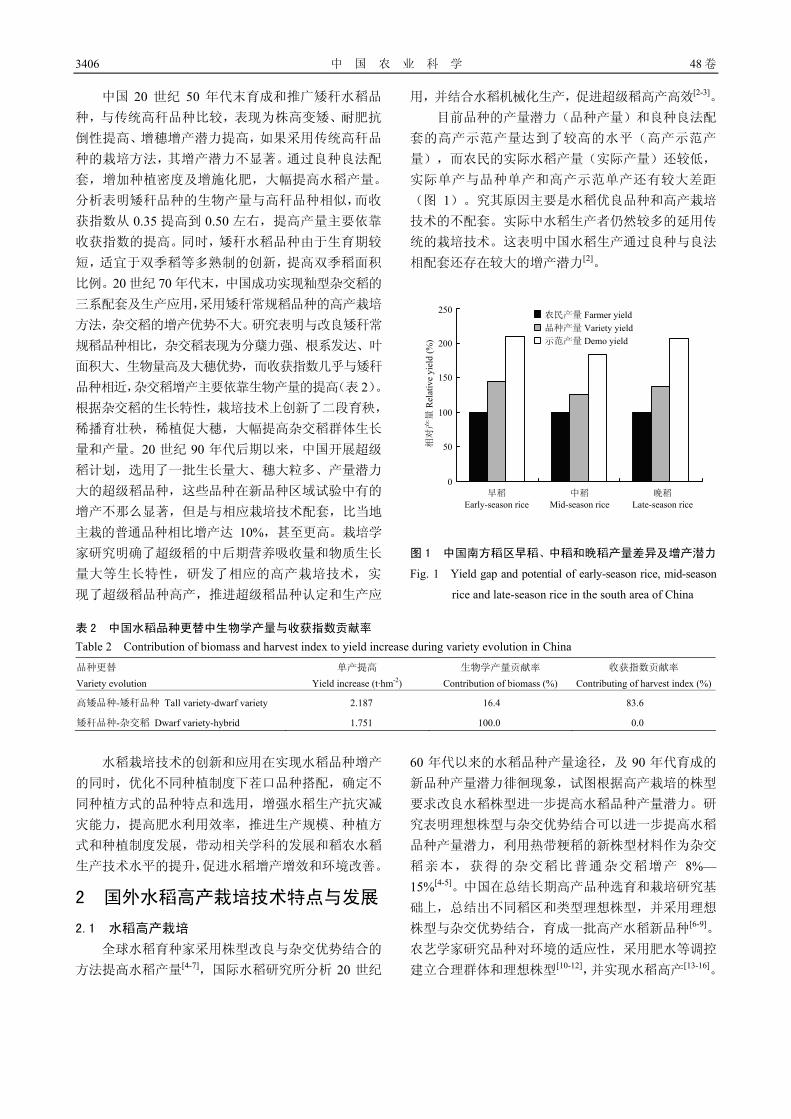
3406 中 国 农 业 科 学 48 卷
中国 20 世纪 50 年代末育成和推广矮秆水稻品
种,与传统高秆品种比较,表现为株高变矮、耐肥抗
倒性提高、增穗增产潜力提高,如果采用传统高秆品
种的栽培方法,其增产潜力不显著。通过良种良法配
套,增加种植密度及增施化肥,大幅提高水稻产量。
分析表明矮秆品种的生物产量与高秆品种相似,而收
获指数从 0.35 提高到 0.50 左右,提高产量主要依靠
收获指数的提高。同时,矮秆水稻品种由于生育期较
短,适宜于双季稻等多熟制的创新,提高双季稻面积
比例。20 世纪 70 年代末,中国成功实现籼型杂交稻的
三系配套及生产应用,采用矮秆常规稻品种的高产栽培
方法,杂交稻的增产优势不大。研究表明与改良矮秆常
规稻品种相比,杂交稻表现为分蘖力强、根系发达、叶
面积大、生物量高及大穗优势,而收获指数几乎与矮秆
品种相近,杂交稻增产主要依靠生物产量的提高(表 2)。根据杂交稻的生长特性,栽培技术上创新了二段育秧,
稀播育壮秧,稀植促大穗,大幅提高杂交稻群体生长
量和产量。20 世纪 90 年代后期以来,中国开展超级
稻计划,选用了一批生长量大、穗大粒多、产量潜力
大的超级稻品种,这些品种在新品种区域试验中有的
增产不那么显著,但是与相应栽培技术配套,比当地
主栽的普通品种相比增产达 10%,甚至更高。栽培学
家研究明确了超级稻的中后期营养吸收量和物质生长
量大等生长特性,研发了相应的高产栽培技术,实
现了超级稻品种高产,推进超级稻品种认定和生产应
用,并结合水稻机械化生产,促进超级稻高产高效[2-3]。 目前品种的产量潜力(品种产量)和良种良法配
套的高产示范产量达到了较高的水平(高产示范产
量),而农民的实际水稻产量(实际产量)还较低,
实际单产与品种单产和高产示范单产还有较大差距
(图 1)。究其原因主要是水稻优良品种和高产栽培
技术的不配套。实际中水稻生产者仍然较多的延用传
统的栽培技术。这表明中国水稻生产通过良种与良法
相配套还存在较大的增产潜力[2]。
0
50
100
150
200
250
早稻Early-season rice
中稻Mid-season rice
晚稻Late-season rice
相对产
量 R
elat
ive
yiel
d (%
)
农民产量 Farmer yield品种产量 Variety yield示范产量 Demo yield
图 1 中国南方稻区早稻、中稻和晚稻产量差异及增产潜力
Fig. 1 Yield gap and potential of early-season rice, mid-season
rice and late-season rice in the south area of China
表 2 中国水稻品种更替中生物学产量与收获指数贡献率
Table 2 Contribution of biomass and harvest index to yield increase during variety evolution in China
品种更替 Variety evolution
单产提高 Yield increase (t·hm-2)
生物学产量贡献率 Contribution of biomass (%)
收获指数贡献率 Contributing of harvest index (%)
高矮品种-矮秆品种 Tall variety-dwarf variety
矮秆品种-杂交稻 Dwarf variety-hybrid
2.187
1.751
16.4
100.0
83.6
0.0
水稻栽培技术的创新和应用在实现水稻品种增产
的同时,优化不同种植制度下茬口品种搭配,确定不
同种植方式的品种特点和选用,增强水稻生产抗灾减
灾能力,提高肥水利用效率,推进生产规模、种植方
式和种植制度发展,带动相关学科的发展和稻农水稻
生产技术水平的提升,促进水稻增产增效和环境改善。
2 国外水稻高产栽培技术特点与发展
2.1 水稻高产栽培
全球水稻育种家采用株型改良与杂交优势结合的
方法提高水稻产量[4-7],国际水稻研究所分析 20 世纪
60 年代以来的水稻品种产量途径,及 90 年代育成的
新品种产量潜力徘徊现象,试图根据高产栽培的株型
要求改良水稻株型进一步提高水稻品种产量潜力。研
究表明理想株型与杂交优势结合可以进一步提高水稻
品种产量潜力,利用热带粳稻的新株型材料作为杂交
稻亲本,获得的杂交稻比普通杂交稻增产 8%— 15%[4-5]。中国在总结长期高产品种选育和栽培研究基
础上,总结出不同稻区和类型理想株型,并采用理想
株型与杂交优势结合,育成一批高产水稻新品种[6-9]。
农艺学家研究品种对环境的适应性,采用肥水等调控
建立合理群体和理想株型[10-12],并实现水稻高产[13-16]。

17 期 朱德峰等:中国水稻高产栽培技术创新与实践 3407
针对非洲及东南亚水稻生产中秧龄过长、密度过
高及淹水灌溉等问题,在马达加斯加首先创新了水稻
强化栽培技术[17]。其高产基本原理通过干湿灌溉,在
节水的同时改善土壤理化特性,根据水稻品种生长特
性,采用增施有机肥料和通气除草等措施,中小苗移
栽和适当降低密度,发挥品种分蘖和大穗优势[18-21]。虽
然该技术在不同地区和技术出现不一致的结果[22-23],但
是,这些高产基本原理和技术与全球的水稻高产栽培
技术一致,在中国水稻高产示范中的应用也取得增产
效果。 2.2 水稻肥水高效利用
施肥是水稻增产的重要措施,增施肥料提高产量
的同时,提高肥料的利用效率引起全球关注。国际水
稻研究所研究建立了水稻实地氮肥管理技术
(SSNM),根据水稻生产氮肥需求量和土壤供氮能
力,并利用 SPAD 等方法建立叶片氮含量诊断指标,
指导水稻氮肥施用量和时期,提高产量和氮肥效率。 水稻是用水量 大的粮食作物,全球水稻生产受
到水资源的制约突出,研究水稻节水栽培技术实现高
产节水是各国研究方向。研发灌溉稻干湿交替(AWD)
水分管理技术,以减少水稻水分用量,提高水分利用
效率,该技术在亚洲主要产稻国广泛应用。在东南亚
及南美等地区旱稻和雨灌稻面积较大,传统的旱稻和
雨灌稻产量低,生产方式比较落后。巴西等国采用保
护性耕作方法,种植旱稻取得很好的增产节水效果,
在不需灌溉稻的灌溉设施和消耗大量灌溉用水的情况
下解决稻米需求[24]。国际水稻研究所针对传统旱稻产
量低,而灌溉稻虽然产量高但用水量大的问题,研发
通气水稻品种,并采用水稻通气栽培,大幅减少水稻
生产的用水量[25-26]。通气水稻具有旱稻的耐旱性及灌
溉稻的产量潜力,与灌溉稻比较可节水 50%。由于
水稻资源的制约,东南亚灌溉稻生产受到限制,稻
农选择通气水稻和通气水稻栽培,提高水资源生产
效率[24-28]。科学家也发现通气水稻栽培管理不当也会
出现连作障碍,导致产量下降。这主要由于土壤大量
和微量元素,如 N、P、K、Fe 和 Zn 缺乏及土壤根结
线虫病等引起,只要管理得当可以消除连作障碍[28-29]。
2.3 水稻抗逆栽培
东南亚水稻生产的主要灾害之一是洪涝灾害[30],
为解决这个难题,科学家在印度耐淹水品种 FR13A 中
发现 SUB1 基因,并将其转到热带主导水稻品种[31]。
这些带有 SUB1 基因的水稻品种能耐两周的淹水,并
正常生长,而淹水敏感品种在淹水一周后生长受抑制
并死亡。在没有淹水情况下,带有 SUB1 基因的水稻
与没有 SUB1 基因的对照品种产量相当,但在淹水条
件下,带有 SUB1 基因的品种产量比没有 SUB1 基因
的品种高出 1 t·hm-2或更高[30, 32]。耐淹水稻及其生产
应用有效解决了易涝地区的水稻生产。在防御水稻洪
涝灾害栽培中,除选择耐淹及高产的水稻品种外,还
提出水稻受淹后采用清水洗苗,补施肥料,湿润灌溉
等栽培技术,加快水稻恢复生长。 欧美及南美主要产稻国的水稻种植方式为直播,
直播稻多年种植后往往会出现红稻,也叫杂草稻,严
重困扰水稻生产。由于红稻和栽培品种具有亲缘关系,
具有水稻属性,普通除草剂对红稻没有灭杀效果。红
稻造成水稻减产,还影响品质,生产上只能采用机械
方法和灭生性除草剂控制红稻。栽培技术上控制杂草
稻的主要方法是在水稻播种前通过耕作和灌溉促使杂
草稻种子萌发出苗,再采用耕作和除草剂灭杀,或采
用移栽方法减少杂草稻发生。美国研究人员发现非转
基因抗咪唑啉酮(imidazoline)等除草剂水稻,利用
该材料选育了抗咪唑啉除草剂水稻品种,这些品种称
为 Clearfield 水稻。种植 Clearfield 水稻的稻田中,当
发现红稻时,可施用咪唑啉酮(imidazoline)等除草
剂将红稻灭杀,而 Clearfield 水稻不受影响[33-34]。这类
除草剂在控制红稻的同时,能有效防除稗草、杂草稻
及其他大多数阔叶杂草与莎草科杂草,从而降低除草
剂用量。该技术的发明有效解决红稻对水稻产量和品
质的影响[33]。 在非洲水稻生产中主要问题是干旱、土壤地力低、
杂草严重,育种和栽培科学家通过亚洲栽培稻与非洲
栽培稻杂交育成非洲新水稻(NERICA),非洲新水
稻保留非洲栽培的抗旱、耐低营养和与较强的杂草
竞争能力等特性,且表现亚洲栽培稻的高产性状,
该类型品种及其栽培技术应用提高了西非和中非的
水稻产量[35-37]。 2.4 水稻生产机械化
社会经济发展及农村劳动力向城镇转移,发达国
家及部分发展中国家推进水稻生产机械化,提高劳动
效率和节约成本。水稻生产的耕作和收获环节,很多
国家基本实现了机械化,但水稻种植环节,不同国家
的机械化水平和方式存在很多差异[38]。欧美、澳大利
亚、日本、韩国等基本实现了从播种到收割各环节的
信息技术和机械化生产相结合的现代水稻生产。目前
欧洲、美国、澳大利亚等国家和地区以机械化直播为
主,其特点是稻田田块面积较大,为防止鸟类等危害

3408 中 国 农 业 科 学 48 卷
种子、控制杂草和防止土壤返盐对出苗影响,采用水
直播方式,播种量高,一般用种量为 150—225 kg·hm-2,
也有旱直播。而日本、韩国及中国台湾水稻种植机械
化以机插秧为主,近年来逐步发展多功能插秧机,
如带平田装置插秧机,深施肥机插一次完成的插秧
机[39]。近年,这些国家和地区与水稻机插配套,发展
水稻机插集中育秧模式和技术,开展机插育供社会化
服务,提高水稻生产的专业化、规模化和机械化水平。
虽然,机插秧已经应用多年,且技术配套完善,但由
于水稻机械育插秧作业环节多、成本相对较高,一直
在研究发展直播稻的可能性。当前,日本和韩国的直
播稻占水稻面积分别在 1.5%和 4%—6%左右,且存在
较大的减产风险。
3 中国水稻高产栽培技术发展回顾
3.1 矮秆品种配套高产栽培
中国水稻高产栽培理论与技术体系,初步形成于
20 世纪 50—60 年代陈永康等针对矮秆晚粳稻老来青
等品种“三黄三黑”高产栽培经验的总结分析[40-41],
所谓水稻“三黄三黑”是指运用肥水管理措施实现水
稻生长过程中叶色三次浓黑、淡黄交替变化,从而构
建合理的高产群体结构。“三黄三黑”高产栽培的核
心是在分蘖期、节间开始形成到幼穗分化期、穗发育
期等三个时期,通过肥水等栽培措施,实现水稻生长
发育的三次黑黄交替变化,分别促进“发棵”、“壮
秆”、“长穗”,从而达到高产。南方的早稻和双季
晚稻以及北方的中早熟品种,因其生育期短,穗分化
和节间形成同时开始,甚至与分蘖末期相重叠,所以
一般只出现“两黑两黄”,即没有圆秆拔节期的一次
黄黑变化。“三黄三黑”高产栽培技术针对当时矮秆
水稻品种生产一般 240—255 万穗/hm2,每穗结实 50—60 粒,穗数和穗粒数均不足,限制高产取得的状况,
提出控制合理苗数,蘖数和穗数的稳定发展,从而培
育高产群体。利用“三黄三黑”高产栽培技术,矮秆
老来青的穗数达到 306—336 万穗/hm2,每穗结实粒数
76—104 粒,百亩示范方高产达 6 840 kg·hm-2,比同
期的全国水稻平均单产增产超过 157.0%,并首次将肥
水技术与水稻生育进程联系起来,对指导水稻大面积
增产发挥了重要作用。 随着中国矮秆品种的选育与推广,相配套的高产
栽培技术以增密、增肥、增穗为主导。为保障粮食安
全,扩大水稻种植面积,中国南方稻区大力发展双季
稻生产,提高稻田单位面积的产量,针对长江中下游
双季稻生长季节紧张等问题,创新了薄膜育秧等技术,
利用薄膜覆盖育秧,增温保湿、防止烂秧、提高成秧
率,实现早稻提早播种,缓解了双季稻生产的季节紧
张和品种搭配问题。同时,采用壮秧密植、增施化肥
等技术,解决了双季稻高产形成过程的穗数及粒数不
足等问题,为促进双季稻在中国南方稻区的推广应用
及高产打下了基础。 3.2 杂交稻品种配套高产栽培
矮秆品种高产主要通过增大群体的办法,来增加
穗数,但群体搞得很大,成穗率不高、结实率较低、
千粒重偏小、病害重、易倒伏,水稻不一定能增产,
却更耗费种子、肥料及农药。蒋彭炎在 20 世纪 70 年
代提出了“稀少平”高产栽培技术,通过稀播、少本
插和平稳促进的肥水管理技术,在一定群体基础上主
攻大穗,避免在一个生育时期猛促猛长,另一个生育
时期又狠控受抑,使群体得到平稳的发展,个体能够
良好的生长生育,实现协调高产[14,42-43]。“稀少平”
高产栽培的核心:(1)大幅度降低秧田播种量,培育
分蘖壮秧;(2)大幅度降低本田用种量,减少抽秧本
数;(3)减少基面肥用量,增加中后期肥料比重;(4)提早搁田,多次轻搁,干干湿湿灌溉,以水调气。随
着中国杂交水稻品种的选育及推广应用,杂交稻需要
通过少本稀植,充分扩大水稻地上部和地下部的生长
与生存空间,达到壮个体、促大穗、得高产的目的,
与“稀少平”栽培不谋而合,该技术研发实现了品种
与技术的配套,对杂交稻品种推广起到了重要作用。
针对双季稻生产晚稻育秧时间长,秧龄过大,不利于
高产,研发创新了二段育秧方法,这是减少专用秧田,
培育矮壮秧的好办法。同时两段育秧秧龄弹性大,有
利于提早播种,早栽夺高产。二段育秧技术在双季晚
稻上应用,对促进双季稻高产栽培起到了重要作用。
中国种植水稻的地域辽阔、生态条件复杂,各地水
稻种植的方法不同,产量表现也不同。邹应斌[44]研究
指出中国东北稻区因低温、四川盆地因高湿和多雾,
水稻栽培上适宜稀植;云贵高原等高海拔稻区由于气
候干燥和光照较强, 水稻栽培上适宜密植;而长江中
下游稻区高温多湿且昼夜温差小, 水稻栽培上适宜合
理密植或适度稀植。
水稻高产栽培强调群体与个体,穗数与粒数、粒
重等因素的协调发展,杂交稻稀植大穗高产,需要适
当控制群体数量、提高群体的质量,因此,需建立水
稻群体质量指标体系,通过高产群体空间结构的指标
合理诊断,优化群体结构。凌启鸿等[50]结合对中国水

17 期 朱德峰等:中国水稻高产栽培技术创新与实践 3409
稻叶龄模式的研究,建立了水稻不同品种类型的生育
进程的叶龄模式,使高产栽培研究由定性向定量并向
模式化、指标化、规范化方向发展。并提出了在适当
压缩群体的前提下,充分发挥水稻个体分蘖能力来确
保群体适宜的穗数,使群体内个体数量和质量达到高
度的协调统一,逐步建立起后期具有高光合生产率和
高物质积累能力的高光效群体,在适宜穗数的基础上
形成大穗,进而提高结实率、粒重和经济系数而获得
高产,并提出了与之相关的高产群体质量指标,如抽
穗期适宜的叶面积指数和与伸长节间数相等的绿色叶
片数,总颖花量,粒叶比,有效叶面积率及高效叶面
积率,抽穗期单茎茎鞘重,抽穗至成熟期颖花根活量、
分藥成穗率等,围绕群体质量的优化,提出了相应的
调控途径与技术,对中国水稻高产栽培起到了很好的
指导作用[45]。
3.3 超级稻品种配套高产栽培
水稻矮秆品种和杂交稻选育及栽培技术配套应用
实现了中国水稻产量的二次飞跃,对水稻生产发展做
出了重要贡献。为进一步提高水稻产量,实现第三次
突破,20 世纪末中国实施了超级稻研究与推广计划,
超级稻品种推广对中国水稻增产做出了重要贡献。研
究超级稻品种特性,发挥其增产潜力,及近年来中国
农村劳动力转移和老龄化,促使水稻种植方式由传统
手插秧向机械化、省工节本种植方式转变,要求水稻
品种与先进的现代种植方式配套。针对稻作技术转型,
种植方式变化,结合中国超级稻不同区域的生态和生
产条件,提出了不同稻区超级稻品种的种植方式适应
性和区域布局。 与普通稻相比,超级稻生长量大、穗大粒多、产
量潜力大。通过不同稻区、季别和类型的超级稻与普
通水稻品种生长特性及产量形成比较研究,表明二者
收获指数差异不大,而总物质生产量差异大,与普通
稻相比,超级稻总生物产量平均提高 10.6%。其中,
拔节前提高 1.3%,拔节-抽穗和抽穗-成熟期分别提高
8.6%和 19.9%。根据超级稻中后期物质生产优势对提
高总生物量有重要贡献(表 3),提出了稳定收获指
数,提高总物质生产量的超级稻高产栽培途径。明确
了超级稻物质生产量高的重要生理基础,其中后期叶
面积指数大,下降缓慢,光合势大。明确超级稻通过
大穗实现高产,提出了在稳定单位面积适量穗数的同
时,通过增加每穗二次枝梗数及着粒数,增加穗粒数
形成大穗是超级稻群体扩库增产的基本途径。根据超
级稻高产形成规律,创立的超级稻“前期早发够穗苗、
中期壮秆扩库容、后期保源促充实”的高产栽培关键
技术,为超级稻区域化高产栽培技术体系建立提供了
支撑。
表 3 超级稻品种与普通水稻品种各生育阶段干物质生产量(kg·hm-2)比较
Table 3 Comparison between super rice and normal rice in dry matter accumulation at different growth stages 品种类型 Variety type
拔节 Elongation
拔节-抽穗期 Elongation-booting
抽穗-成熟期 Elongation-maturity
总生产量 Total biomass
超级稻 Super rice(n=13) 4039.5 7659.0 6517.5 18216.0
普通水稻 Normal rice(n=3) 3987.0 7050.0 5434.5 16471.5
超级稻/普通水稻 Super rice/normal rice 1.01 1.09 1.20 1.11
长期以来,水稻栽培粗放化管理,肥水等盲目投
入,水稻生产污染加重,制约了水稻增产增效与可持
续发展。张洪程等[46]在群体质量栽培等研究基础上,创立了以生育进程、群体动态指标、栽培技术措施“三
定量”和作业次数、调控时期、投入数量“三适宜”
为核心的水稻精确定量栽培技术,创立了水稻高产共
性生育模式与形态生理精确定量指标及其实用诊断方
法,实现了栽培方案优化设计与生产过程实时实地准
确诊断;邹应斌等[16]针对超级稻特点,提出了一种定
目标产量、定群体指标、定技术规程的三定栽培法,
提出了超级稻“因地定产、依产定苗、测苗定氮”栽
培技术,有利于充分发挥超级稻的大穗优势。从而促
进了超级稻栽培技术由定性为主向精确定量的跨越,为统筹实现水稻“高产、优质、高效、生态、安全”
提供了重大技术支撑。 随着社会的发展,技术的更新,水稻机插秧技术
快速发展。针对中国传统的毯状秧苗机插存在的播种
量大机插本数不均匀,漏秧率高;机插对秧苗损伤大,
缓苗期长,返青起发慢等问题,中国水稻研究所朱德
峰等首创了水稻钵形毯状秧苗机插技术,结合机插毯
苗和抛秧钵苗的优势,采用钵形毯状秧盘, 精量定位
播种,培育具有上毯下钵壮苗,及定位定量按钵取秧
机插,有效解决了传统毯状秧苗机插存在的机插定量
定位性差、漏秧率高、伤秧伤根严重、每丛苗数不均

3410 中 国 农 业 科 学 48 卷
匀,及返青慢等问题,实现水稻钵苗机插,大幅提高
水稻机插产量和效益[47-48]。
4 水稻高产栽培存在的问题
4.1 水稻种植机械化水平低,农机与农艺措施不配套
中国社会经济发展、农村劳动力转移和成本提高
及水稻生产规模化的新情况下,迫切需要水稻生产机
械化。近年来,中国水稻机械化发展很快[49],主产区
水稻生产耕作和收获基本实现机械化,但是,存在种
植机械化水平和程度还较低,区域间及季节间差异大,
水稻施肥和植保机械化水平还较低,及耕作和收获的
机械化与种植机械化不配套等问题[50-51]。近年来,在
国家政策的支持下,中国的水稻生产机械化发展迅速,
其中整地和水稻收获环节机械化水平较高,种植环节
机械化水平还很低,2014 年全国水稻机械化种植水平
只达到 38%左右。“十二五”期间,农业部要求水稻
机械化种植水平每年提高 4—5 个百分点[52]。水稻种
植环节机械化主要是机械化移栽和机直播,水稻钵形
毯状机插技术作为水稻机插新技术,其年推广面积超
过 4.5×108 hm2。机直播和机械化摆栽约占水稻种植
面积 2%左右。 目前中国水稻种植逐渐向机械化、规模化和社会
化服务方向发展。水稻种植机械化存在农机与农艺不
配套的问题。机械化种植和耕作技术结合的不够紧密,
水稻田机械平地质量较差,从而造成机插效果不佳。
由于生长季节比较紧张,双季稻机插主要存在早晚稻
品种搭配问题,需选择生育期适宜的品种,减少早稻
苗期倒春寒和晚稻中后期寒露风的危害。单季稻中杂
交稻品种面积大,发展杂交稻机插秧技术必须解决好
目前育秧用种量过多的问题,降低育秧用种量,实现
杂交稻双本精量机插。 4.2 水稻肥水利用效率低
化肥的施用是农业发展史上一次重大变革,它极
大地提高了农作物的产量,缓解了全球的粮食紧张局
面。水稻生产中肥水是高产栽培的主要手段,但是,
过量的肥水不仅会造成减产、减低品质,增加成本,
还污染环境。中国氮肥消费量占世界氮肥总量的 30%,水稻生产所消耗的氮肥占世界水稻氮肥总消耗量的 37%。中国水稻生产氮肥施用量较生产水平相当的产
稻国高 30%—50%,造成其利用率较低[53]。中国水稻
生产氮肥用量大及利用较低,与水稻品种耐肥性较
强、土壤地力、肥料类型、施肥及水分管理等多方
面有关[53-54]。研究表明,在中国稻田碳酸氢铵氮肥的
吸收利用率低于 30%,尿素为 30%—40%[55],中国稻
田氮肥吸收利用率为 30%—35%[56],部分省份如江苏
水稻的氮肥吸收利用率仅 19.9%,远低于全国平均水
平[57]。南方六省农田氮素盈余高达 50%—190%,且有
继续增大的趋向[58]。中国肥料利用效率不高的主要原
因在于长时间以高产为导向,大部分品种高产性能只
有在高肥水条件下才能表现出来,另外肥料类型比较
单一,大部分属于速效肥,肥料损失很大。 中国淡水资源整体上是南多北少,水资源南北分
布差异很大。北方地区土地面积占全国的 63.7%,耕
地面积为全国的 62.2%,而水资源总量却只占全国的
20.11%。近年来,北方水稻面积在不断增加,限制北
方水稻发展的关键问题是水资源短缺,因此如何合理
有效利用淡水资源是北方水稻稳产的关键问题。 4.3 水稻灾害防控能力差
中国水稻种植区域跨度大,种植制度多样,易遭
受多种灾害天气的影响,主要包括高低温、干旱和洪
涝灾害。长江中下游稻区水稻开花结实期间经常遇到
异常高温,易造成早稻高温逼熟,单季稻结实率大幅
度下降。2003 年长江流域的高温灾害导致稻谷损失就
达 5.18×107 t。南方地区水稻遭遇低温灾害主要是早
稻苗期的倒春寒和中晚稻后期的寒露风。2007 年,浙
江、福建、江苏等东南沿海地区中晚稻灌浆结实期间
受台风低温影响,水稻结实率大幅度下降,造成严重
减产。北方稻区低温伤害主要分两种,即抑制性低温
和障碍性低温,如 2009 年,黑龙江水稻移栽后遭受较
长时间的低温导致水稻生育期推迟,而开花结实期又
受低温影响,导致水稻结实率大幅度下降。干旱也是
制约水稻生产的重要因素之一,中国南方尤其是西南
地区干旱发生频率较高,华中和华南地区水稻因干旱
导致的单产损失达 143—250 kg·hm-2。南方稻区雨热
同季,水稻生长季也易遭受洪涝灾害,仅 2010 年全国
水稻因洪涝灾害造成的直接产量损失达 50×108 kg。高低温、干旱和洪涝等自然灾害严重威胁到了中国水
稻生产,但是目前针对水稻灾害的预警措施较少,抗
性品种少,针对性的防御措施也较少[59]。增强水稻对
灾害气候防御能力是进一步提高水稻产量的重要措施
之一。
5 水稻高产栽培技术发展展望
5.1 水稻良种良法配套栽培
中国水稻生产从高秆品种到矮秆品种,从矮秆品
种到杂交稻及近年的超级稻对产量提高和稳定的贡献

17 期 朱德峰等:中国水稻高产栽培技术创新与实践 3411
均得益于良种良法配套。育种技术的进步加快水稻品
种类型和品种的选育及应用[60-61],不同类型品种生育
特性差异较大,且中国稻作技术和生产方式转型,更
加需要良种良法的配套,使品种适用于适宜生态区、
种植制度和种植方式[12,62]。水稻良种良法配套已从传
统的先有品种,后配套栽培技术的方式,转变为育种
与栽培研究互动,按需求目标选育品种。近几十年来,
随着水稻规模化生产和机械化作业的发展,全球气候
变化引起的自然灾害频发,土壤结构和肥力衰退,水
资源短缺,肥药用量大及利用效率不高等新状态,更
加需要加强育种与栽培学科联合来解决问题。水稻栽
培技术研究也正在从传统的高产高效优质为主要目
标,发展到高产高效优质生态安全的综合生产目标。 5.2 水稻生产机械化
水稻机械化重点解决水稻育插秧模式、装备和技
术,双季稻和杂交稻的机插技术,杂交稻制种技术,
机械化施肥施药技术,及主要环节机械化作业的配套。
特别是创新精量播种、取秧和机插,深施肥与机插一
次作业,机插集中育供秧模式和技术,及选育适宜机
插品种,探索适宜地区的机械直播技术,加快推进水
稻规模化生产、机械化作业和社会化服务。 5.3 水稻肥水高效管理
水稻生产降低氮肥用量,提高氮肥效率,实现增
产高效,需要选育氮敏感和高效品种,改善和培育稻
田结构和地力,研发新型肥料和施肥方法,创新通气
耕作、肥水一体化管理和合理密植等栽培技术。中国
的水稻水分管理研究已经取得很好进展,但是,水稻
种植方式、品种特性、作业方式的发展和演变,还有
盐碱地、重金属地区的水稻生产,需要创新配套的新
型水分管理模式和技术,提出定量化、指标化的水分
管理模式。
5.4 水稻灾害防控体系
中国水稻种植区域广阔,全球气候变化及水稻种
植制度的演变,及新类型品种育成和推广,呈现水稻
生产灾害频发重发。中国水稻生产自然灾害主要有高低
温、干旱和洪涝。重点建立水稻高低温、干旱和洪涝灾
害品种耐性鉴定方法、评价标准和灾害损失评估方法,
选育抗灾品种,创新避灾抗灾水稻种植制度,研究水稻
高低温、干旱和洪涝等灾害的预警和抗灾减灾技术,建
立品种、环境和技术结合的灾害防控技术体系。
References
[1] 中国农业年鉴. 北京: 中国农业出版社, 2014.
China Agriculture Yearbook. Beijing: China Agriculture Press, 2014.
(in Chinese)
[2] 朱德峰. 超级稻栽培技术. 北京: 金盾出版社, 2006.
Zhu D F. Cultivation Techniques of Super Rice. Beijing: Jindun
Publishing House, 2006. (in Chinese)
[3] 何强, 邢俊杰, 舒服, 张武汉, 邓华凤. 水稻收获指数研究概况与
展望. 杂交水稻, 2012, 27(6): 3-6.
He Q, Xing J J, Shu F, Zhang W H, Deng H F. Review and prospect
for the research on harvest index in rice. Hybrid Rice, 2012, 27(6): 3-6.
(in Chinese)
[4] Khush G S. Strategies for increasing the yield potential of rice. Studies
in Plant Science, 2000, 7: 207-212.
[5] Peng S, Khush G S, Virk P, Tang Q, Zou Y. Progress in ideotype
breeding to increase rice yield potential. Field Crops Research, 2008,
108(1): 32-38.
[6] 袁隆平. 选育超高产杂交水稻的进一步设想. 杂交水稻, 2012,
27(6): 1-2.
Yuan L P. Conceiving of breeding further super-high-yield hybrid rice.
Hybrid Rice, 2012, 27(6): 1-2. (in Chinese)
[7] 徐正进, 陈温福, 张文忠, 刘丽霞, 周淑清, 张龙步, 杨守仁. 水
稻的产量潜力与株型演变 . 沈阳农业大学学报 , 2000, 31(6):
534-536.
Xu Z J, Chen W F, Zhang W Z, Liu L X, Zhou S Q, Zhang L B, Yang
S R. The yield potentiality of rice and it's development of plant type.
Journal of Shenyang Agricultural University, 2000, 31(6): 534-536.
(in Chinese)
[8] 程式华, 翟虎渠. 水稻亚种间超高产杂交组合若干株型因子的比
较. 作物学报, 2000, 26(6): 713-718.
Cheng S H, Zhai H Q. Comparison of some plant type components in
super high-yielding hybrids of inter-subspecies rice. Acta Agronomica
Sinica, 2000, 26(6): 713-718. (in Chinese)
[9] 周开达, 马玉清, 刘太清, 沈茂松. 杂交水稻亚种间重穗型组合选
育-杂交水稻超高产育种的理论与实践. 四川农业大学学报, 1995,
13(4): 403-407.
Zhou K D, Ma Y Q, Liu T Q, Shen M S. The breeding of subspecific
heavy ear hybrid rice-exploration about super-high yield breeding of
hybrid rice. Journal of Sichuan Agricultural University, 1995, 13(4):
403-407. (in Chinese)
[10] 程式华, 翟虎渠. 杂交水稻超高产育种策略. 农业现代化研究,
2000, 21(3): 147-150, 154.
Cheng S H, Zhai H Q. Breeding strategies for superior high-yielding
in hybrid rice. Research of Agricultural Modernization, 2000, 21(3):
147-150, 154. (in Chinese)

3412 中 国 农 业 科 学 48 卷
[11] 杨惠杰, 杨仁崔, 李义珍, 姜照伟, 郑景生. 水稻超高产品种的产
量潜力及产量构成因素分析. 福建农业学报, 2000, 15(3): 1-8.
Yang H J, Yang R C, Li Y Z, Jiang Z W, Zheng J S. Yield potential
and yield components of super high-yielding rice cultivars. Fujian
Journal of Agricultural Sciences, 2000, 15(3): 1-8. (in Chinese)
[12] 苏柏元, 朱德峰. 超级稻甬优 12 机插单产 1000kg/667m2的产量结
构与配套栽培技术. 中国稻米, 2013, 19(4): 97-100.
Su B Y, Zhu D F. The yield components and cultivation technology of
Yongyou 12 yielding over 1000 kg per 667 m2 through mechanical
transplanting. China Rice, 2013, 19(4): 97-100. (in Chinese)
[13] 颜振德. 杂交水稻高产群体的干物质生产与分配的研究. 作物学
报, 1981, 7(1): 11-18.
Yan Z D. Studies on the production and distribution of dry matter in
high-yielding populations of hybrid rice. Acta Agronomica Sinica,
1981, 7(1): 11-18. (in Chinese)
[14] 蒋彭炎, 冯来定, 姚长溪. 从水稻稀少平栽培法的高产效应看栽培
技术与株型的关系. 中国水稻科学, 1987, 1(2): 111-117.
Jiang P Y, Feng L D, Yao C X. The effect of cultivation techniques on
plant type in view of the high yield result induced by TFS cultivation
method in paddy rice. Chinese Journal of Rice Science, 1987, 1(2):
111-117. (in Chinese)
[15] 凌启鸿, 苏祖芳, 张海泉. 水稻成穗率与群体质量的关系及其影响
因素的研究. 作物学报, 1995, 21(4): 463-469.
Ling Q H, Su Z F, Zhang H Q. Relationship between earbearing tiller
percentage and population quality and its influential factors in rice.
Acta Agronomica Sinica, 1995, 21(4): 463-469. (in Chinese)
[16] 凌启鸿, 张洪程, 蔡建中, 苏祖芳, 凌励. 水稻高产群体质量及其
优化控制探讨. 中国农业科学, 1993, 26(6): 1-11.
Ling Q H, Zhang H C, Cai J Z, Su Z F, Ling L. Investigation on the
population quality of high yield and its optimizing control programme
in rice. Scientia Agricultura Sinica, 1993, 26(6): 1-11. (in Chinese)
[17] Tsujimoto Y, Horie T, Randriamihary H, Shiraiwa T, Homma K. Soil
management: The key factors for higher productivity in the fields
utilizing the system of rice intensification (SRI) in the central highland
of Madagascar. Agricultural Systems, 2009, 100(1-3): 61-71.
[18] 朱德峰, 林贤青, 陶龙兴, 张玉屏, 郑家国, 吕世华, 马均. 水稻强
化栽培体系的形成与发展. 中国稻米, 2003(2): 17-18.
Zhu D F, Lin X Q, Tao L X, Zhang Y P, Zheng J G, Lü S H, Ma J.
The formation and development of system of rice intensification.
China Rice, 2003(2): 17-18. (in Chinese)
[19] 林贤青, 朱德峰, 张玉屏. 水稻强化栽培体系的原理及其应用效
果. 中国稻米, 2003(3): 23-24.
Lin X Q, Zhu D F, Zhang Y P. Principle and Application Effect
of the system of rice intensification. China Rice, 2003(3): 23-24.
(in Chinese)
[20] Sinha S K, Talati J. Productivity impacts of the system of rice
intensification (SRI): A case study in West Bengal, India. Agricultural
Water Management, 2007, 87(1): 55-60.
[21] Stoop W A, Uphoff N, Kassam A. A review of agricultural research
issues raised by the system of rice intensification (SRI) from
Madagascar: Opportunities for improving farming systems for
resource-poor farmers. Agricultural Systems, 2002, 71(3): 249-274.
[22] Dobermann A. A critical assessment of the system of rice
intensification (SRI). Agricultural Systems, 2004, 79(3): 261-281.
[23] Latif M A, Ali M Y, Islam M R, Badshah M A, Hasan M S.
Evaluation of management principles and performance of the system
of rice intensification (SRI) in Bangladesh. Field Crops Research,
2009, 114(2): 255-262.
[24] Pinheiro B d S, Castro E M, Guimarães CM. Sustainability and
profitability of aerobic rice production in Brazil. Field Crops
Research, 2006, 97(1): 34-42.
[25] Bouman B A M, Peng S, Castañeda A R, Visperas R M. Yield and
water use of irrigated tropical aerobic rice systems. Agricultural Water
Management, 2005, 74(2): 87-105.
[26] Kato Y, Okami M, Katsura K. Yield potential and water use efficiency
of aerobic rice (Oryza sativa L.) in Japan. Field Crops Research, 2009,
113(3): 328-334.
[27] Balasubramanian V, Sie M, Hijmans R J, Otsuka K. Increasing rice
production in Sub-Saharan Africa: Challenges and opportunities.
Advances in Agronomy, 2007: 55-133.
[28] Prasad R. Chapter four-aerobic rice systems. Advances in Agronomy,
2011: 207-247.
[29] Kreye C, Bouman B A M, Reversat G, Fernandez L, Vera Cruz C,
Elazegui F, Faronilo J E, Llorca L. Biotic and abiotic causes of yield
failure in tropical aerobic rice. Field Crops Research, 2009, 112(1):
97-106.
[30] Ismail A M, Singh U S, Singh S, Dar M H, Mackill D J. The
contribution of submergence-tolerant (Sub1) rice varieties to food
security in flood-prone rainfed lowland areas in Asia. Field Crops
Research, 2013(152): 83-93.
[31] Mackill D J, Ismail A M, Singh U S, Labios R V, Paris T R. Chapter
six - development and rapid adoption of submergence-tolerant (Sub1)
rice varieties. Advances in Agronomy, 2012: 299-352.
[32] Singh S, Mackill D J, Ismail A M. Responses of SUB1 rice
introgression lines to submergence in the field: Yield and grain quality.
Field Crops Research, 2009,113(1): 12-23.

17 期 朱德峰等:中国水稻高产栽培技术创新与实践 3413
[33] Sudianto E, Beng-Kah S, Ting-Xiang N, Saldain N E, Scott R C,
Burgos N R. Clearfield (R) rice: Its development, success, and key
challenges on a global perspective. Crop Protection, 2013(49): 40-51.
[34] Rosas J E, Bonnecarrère V, Pérez de Vida F. One-step, codominant
detection of imidazolinone resistance mutations in weedy rice (Oryza
sativa L.). Electronic Journal of Biotechnology, 2014, 17(2): 95-101.
[35] Rodenburg J, Saito K, Kakaï R G, TouréA, Mariko M, Kiepe P. Weed
competitiveness of the lowland rice varieties of NERICA in the
southern Guinea Savanna. Field Crops Research, 2009, 114(3):
411-418.
[36] Rodenburg J, Cissoko M, Kayeke J, Dieng I, Khan Z R, Midega C A
O, Onyuka E A, Scholes J D. Do NERICA rice cultivars express
resistance to Striga hermonthica (Del.) Benth. and Striga asiatica (L.)
Kuntze under field conditions. Field Crops Research, 2015(170):
83-94.
[37] Ekeleme F, Kamara A Y, Oikeh S O, Omoigui L O, Amaza P,
Abdoulaye T, Chikoye D. Response of upland rice cultivars to weed
competition in the savannas of West Africa. Crop Protection, 2009,
28(1): 90-96.
[38] 朱德峰, 陈惠哲, 徐一成. 中国水稻种植机械化的发展前景与对策.
北方水稻, 2007(5): 13-18.
Zhu D F, Chen H Z, Xu Y C. Countermeasure and perspective of
mechanization of rice planting in China. North Rice, 2007(5): 13-18.
(in Chinese)
[39] 李耀明, 徐立章, 向忠平, 邓玲黎. 日本水稻种植机械化技术的
新研究进展. 农业工程学报, 2005, 21(11): 190-193.
Li Y M, Xu L Z, Xiang Z P, Deng L L. Research advances of rice
planting mechanization in Japan. Transactions of the Chinese Society
of Agricultural Engineering, 2005, 21(11): 190-193. (in Chinese)
[40] 凌启鸿. 中国特色水稻栽培理论和技术体系的形成与发展. 江苏
农业学报, 2008, 24(2): 101-113.
Ling Q H. Formation and development of theory and technological
system of rice cultivation with chinese characteristics. Jiangsu
Journal of Agricultural Sciences, 2008, 24(2): 101-113. (in Chinese)
[41] 杨立烱, 汤玉庚, 王嘉训, 陈永康. 晚粳稻“三黑三黄”高产栽培经
验的初步分析. 作物学报, 1964, 3(2): 113-132.
Yang L J, Tang Y G, Wang J X, Chen Y K. Analysis on the high
yielding technique of late rice. Acta Agronomica Sinica, 1964, 3(2):
113-132. (in Chinese)
[42] 蒋彭炎, 姚长溪, 任正龙, 冯来定. 论早稻稀少平高产栽培法. 浙
江农业大学学报, 1983, 9(2): 127 -129.
Jiang P Y, Yao C X, Ren Z L, Fen L D. Discussion of the “TFS”
cultivation method for early rice. Journal of Zhejiang Agricultural
University, 1983, 9(2): 127 -129. (in Chinese)
[43] 谢振宇, 沈建凯, 尹明, 贺治洲. 杂交水稻高产栽培研究进展. 热
带农业科学, 2014, 34(7): 39-45.
Xie Z Y, Shen J K, Yin M, He Z Z.Research progress of high yielding
cultivation of hybrid rice. Chinese Journal of Tropical Agriculture,
2014, 34(7): 39-45. (in Chinese)
[44] 邹应斌. 长江流域双季稻栽培技术发展. 中国农业科学, 2011,
44(2): 254-262.
Zou Y B. Development of cultivation technology for double cropping
rice along the Changjiang River Valley. Scientia Agricultura Sinica,
2011, 44(2): 254-262. (in Chinese)
[45] 付景, 杨建昌. 中国水稻栽培理论与技术发展的回顾与展望. 作物
杂志, 2010(5): 1-4.
Fu J, Yang J C. Retrospect and prospect for the development of theory
and technology of rice cultivation in China. Crop, 2010(5): 1-4. (in
Chinese)
[46] 张洪程, 吴桂成, 吴文革, 戴其根, 霍中洋, 许轲, 高辉, 魏海燕,
黄幸福, 龚金龙. 水稻“精苗稳前、控蘖优中、大穗强后”超高产定
量化栽培模式. 中国农业科学, 2010, 43(13): 2645-2660.
Zhang H C, Wu G C, Wu W G, Dai Q G, Huo Z Y, Xu K, Gao H,
Wei H Y, Huang X F, Gong J L. The SOI model of quantitative
cultivation of super-high yielding rice. Scientia Agricultura Sinica,
2010, 43(13): 2645-2660. (in Chinese)
[47] 徐一成, 朱德峰, 赵匀, 陈惠哲. 超级稻精量条播与撒播育秧对秧
苗素质及机插效果的影响. 农业工程学报, 2009, 25(1): 99-103.
Xu Y C, Zhu D F, Zhao Y, Chen H Z. Effects of broadcast sowing and
precision drilling of super rice seed on seedling quality and effectiveness
of mechanized transplanting. Transactions of the Chinese Society of
Agricultural Engineering, 2009, 25(1): 99-103. (in Chinese)
[48] 陈惠哲, 朱德峰, 徐一成. 水稻钵形毯状秧苗机插技术及应用效果.
中国稻米, 2009, 15(3): 5-7.
Chen H Z, Zhu D F, Xu Y C. Mechanized planting technology of rice
bowl-shaped blanket seedling and application effect. China Rice, 2009,
15(3): 5-7. (in Chinese)
[49] 牛盾. 中国农业机械化的新形势和水稻生产机械化问题. 农业工
程学报, 2000, 16(4): 7-10.
Niu D. New situation of agricultural mechanization and the
development of paddy production mechanization in China. Transactions
of the Chinese Society of Agricultural Engineering, 2000, 16(4): 7-10.
(in Chinese)
[50] 朱德峰, 陈惠哲. 水稻机插秧发展与粮食安全. 中国稻米, 2009, 6:
4-7.
Zhu D F, Chen H Z. Rice transplanting development and food security.

3414 中 国 农 业 科 学 48 卷
China Rice, 2009(6): 4-7. (in Chinese)
[51] 朱德峰, 陈惠哲, 徐一成. 中国水稻机械种植的发展前景与对策.
农业技术与装备, 2007, 1(133): 14-15.
Zhu D F, Chen H Z, Xu Y C. Development prospect and
countermeasures of rice machinery plant in China. Agricultural
Technology & Equipment, 2007, 1(133): 14-15. (in Chinese)
[52] 李斯华. 中国水稻生产机械化的发展态势和目标任务. 北方水稻,
2011, 41(2): 1-2.
Li S H. Development trend and target of rice roduction mechanization
in China. North Rice, 2011, 41(2): 1-2. (in Chinese)
[53] 彭少兵, 黄见良, 钟旭华, 杨建昌, 王光火, 邹应斌, 张福锁, 朱庆
森, Buresh C W. 提高中国稻田氮肥利用率的研究策略. 中国农业
科学, 2002, 35(9): 1095-1103.
Peng S B, Huang J L, Zhong X H, Yang J C, Wang G H, Zou Y B,
Zhang F S, Zhu Q S, Buresh C W. Research strategy in improving
fertilizer-nitrogen use efficiency of irrigated rice in China. Scientia
Agricultura Sinica, 2002, 35(9): 1095-1103. (in Chinese)
[54] 李忠芳, 徐明岗, 逄焕成, 张会民, 孙楠, 娄翼来, 李慧. 中国南方
潴育性水稻土产量演变及其肥力驱动因素分析. 土壤学报, 2014,
51(5): 953-962.
Li Z F, Xu M G, Pang H C, Zhang H M, Sun N, Lou Y L, Li H. The
rice yield trends in three hydragric paddy soils in South China and its
fertility factors. Acta Pedologica Sinica, 2014, 51(5): 953-962. (in
Chinese)
[55] 朱兆良. 中国土壤供氮和化肥氮去向研究的进展. 土壤, 1985,
17(1): 2-9
Zhu Z L. Advances in investigations of soil nitrogen supply and fate
fertilizer nitrogen in soils of China. Soils, 1985, 17(1): 2-9. (in Chinese)
[56] 李庆逵. 中国农业持续发展中的肥料问题. 南昌: 江西科学技术出
版社, 1997.
Li Q K. Fertilizer Issues in the Sustainable Development of China
Agriculture. Nanchang: Jiangxi Science and Technology Press, 1997.
(in Chinese)
[57] 李荣刚. 高产农田氮索肥效与调控途径-以江苏太湖地区稻麦两熟
农区为例推及全省[D]. 北京: 中国农业大学, 2000.
Li R G. Efficiency and regulation of fertilizer nitrogen in high-yield
farmland -a case study on rice and wheat double maturing system
agriculture area of Tai Lake for deducing to Jiangsu Province[D].
Beijing: China Agricultural University, 2000. (in Chinese)
[58] 鲁如坤, 时正元, 施建平. 中国南方 6 省农田养分平衡现状评价和
动态变化研究. 中国农业科学, 2000, 33(2): 63-67.
Lu R K, Shi Z Y, Shi J P. Nutrient balance of agroecosystem in six
provinces in Southern China. Scientia Agricultura Sinica, 2000, 33(2):
63-67. (in Chinese)
[59] 程式华. 中国水稻产业发展报告. 北京: 中国农业出版社, 2011.
Cheng S H. The Development Report of Rice Industry in China.
Beijing: China Agriculture Press, 2011. (in Chinese)
[60] 孙宗修, 鄂志国, 王磊, 朱德峰, 张玉屏, 胡国成, 刘文真, 付亚萍.
对中国水稻骨干亲本评定方法的探索. 作物学报, 2014, 40(6):
973-983.
Sun Z X, E Z G, Wang L, Zhu D F, Zhang Y P, Hu G C, Liu W Z, Fu
Y P. Exploring assessment method of Chinese rice backbone parents.
Acta Agronomica Sinica, 2014, 40(6): 973-983. (in Chinese)
[61] 杨仕华, 程本义, 沈伟峰. 中国南方稻区杂交水稻育种进展. 杂交
水稻, 2004, 19(5): 4-8.
Yang S H, Cheng B Y, Shen W F. Progress of hybrid rice breeding in
Southern China. Hybrid Rice, 2004, 19(5): 4-8. (in Chinese)
[62] 陈惠哲, 朱德峰, 杨仕华, 张玉屏, 林贤青. 中国南方稻区水稻产
量差异及增产潜力. 中国稻米, 2004(4): 9-10.
Chen H Z, Zhu D F, Yang S H, Zhang Y P, Lin X Q. Rice yield gap
and potential in southern rice zone in China. China Rice, 2004(4):
9-10. (in Chinese)
(责任编辑 张晶,杨鑫浩)

中国农业科学 2015,48(17):3415-3428 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.009
收稿日期:2015-05-04;接受日期:2015-06-18 基金项目:国家自然科学基金(31171454)、国家“863”计划(2013AA102405)、北京市科技新星计划(XX2013022) 联系方式:赵春江,Tel:010-51503411;E-mail:[email protected]
数字植物研究进展:植物形态结构三维数字化
赵春江,陆声链,郭新宇,杜建军,温维亮,苗 腾
(北京农业信息技术研究中心/国家农业信息化工程技术研究中心/农业部农业信息技术重点开放实验室/数字植物北京市重点实验室,北京 100097)
摘要:数字植物围绕农林植物生命、生产和生态系统的多维信息高效感知和认知的理论、技术和方法,通过
多学科交叉合作,研究农林植物-环境 3D 数字化、高通量信息获取、情景感知、信息融合、结构和功能模拟、数
字化设计和精准管理决策等数字农业的关键性、基础性以及共性理论和技术问题。植物形态结构的三维数字化是
数字植物研究的重要组成部分,近年来很多学者从植物组织、器官、植株和群体等不同尺度,或者从植物根系和
地上部等不同视角,围绕植物形态结构的参数测量、几何结构解析、三维模型构建、结构与功能建模,以及三维
植物模型真实感展现等需要,开展了更深入的研究。在组织尺度方面,随着 MRI、CT、显微成像等技术产品的不
断成熟,使得利用这些先进测量仪器获取和测量植物组织内部结构数据成为可能,并被越来越多的研究者采用,
成为进行植物内部形态结构测量和分析的有效手段。在植物根系的三维数字化方面,由于植物的根普遍生长在土
壤里,观察和测量十分困难。虽然近年来 XCT、MRI 等穿透射线成像技术已越来越多地用于根系的形态结构探测,
但这类技术往往仅能获取范围较小的根系局部数据,且价格昂贵。因此植物根系形态结构的准确、无损(原位)、
快速测量仍然是一个挑战。在群体尺度方面,基于实测数据的三维重建逐渐成为植物群体三维重建的主要途径,
研究者正试图从激光三维扫描仪获取的植物群体三维点云中提取群体的形态参数并实现群体的三维重构。而在三
维植物模型的真实感绘制方面,如何准确地测量各种植物器官的光学特性并建立相应的数学模型是当前的研究重
点,虽然已有不少研究者提出了相应的解决方案,但这些方法在便捷性和普适性方面仍然难以令人满意,有待更
多深入研究。笔者最后结合相关领域的技术进展对数字植物的进一步研究进行了展望。
关键词:数字植物;植物形态结构建模;三维重建;可视化
Advances in Research of Digital Plant: 3D Digitization of Plant
Morphological Structure ZHAO Chun-jiang, LU Sheng-lian, GUO Xin-yu, DU Jian-jun, WEN Wei-liang, MIAO Teng
(Beijing Research Center for Information Technology in Agriculture/National Engineering Research Center for Information
Technology in Agriculture/Key Laboratory for Information Technology in Agriculture, Ministry of Agriculture/Beijing Key
Laboratory of Digital Plant, Beijing 100097)
Abstract: Digital plant focuses on research of the theories, technologies, and methods of efficient perception and cognition to
the multi-dimensional information in plant life, agricultural production, and ecological system. The critical, basic, common theories, and technical problems in digital agriculture, including the 3D digitization of plants and its environment, high-throughput information acquisition, context awareness, information fusion, structural-functional simulation, digital design, and accurate management decision, can be studied in depth through the interdisciplinary cooperation. The study of 3D digitization to plant structure is one of the most important topics in the area of digital plant. Recently many in-depth studies on the 3D digitization of plant structure had been reported by considering the real requirements from parameter measurement of plant structure, analysis of morphological structure, 3D reconstruction, structural-functional modeling and realistic rendering of 3D plant models. These studies
jia
文本框
返回目录

3416 中 国 农 业 科 学 48 卷
were done not only from tissue, organ, plant to colony scale, but also from aboveground to underground. In organ scale, with the advance of MRI, CT, microscopic imaging techniques and the corresponding products, using these advanced instruments to measure internal structure data of plant has become possible and popular in more and more researchers. And this also provides an effective method for measuring and analyzing the microcosmic morphological structure of plant. To the 3D digitization of root system, the observation and measurement of plant root are very difficult for root always grows deep in soil. Although XCT, MRI and X-ray imaging techniques have been more and more widely used in recent years for detecting the morphological structure of root, these techniques can often only be used to measure a small part of a whole root. So the accurate, nondestructive and fast measurement of root structure is still a challenge. At colony scale, real measured data-based 3D reconstruction has become a main method for the three-dimensional reconstruction of plant colony. Currently, some researchers are trying to extract the morphological parameters of plant colony directly from 3D point cloud then realize the 3D reconstruction. While at the research of realistic rendering of 3D plant model, how to accurately measure the optical properties of various plant organs and to establish a corresponding mathematical model is the focus in this area. However, although some solutions had been reported, these proposed methods are still not satisfactory when considering the convenience and general applicability, and more in-depth studies are expected to be done. Lastly, the further study of digital plant was also discussed based on the analysis of the related technical progress.
Key words: digital plant; modeling of plant morphology and structure; 3D reconstruction; visualization
0 引言
“数字植物”是在数字农业研究和应用蓬勃发展背
景下提出的研究专题[1],是指综合运用数字化技术,
通过高通量信息获取及智能处理研究农林植物的生
命、生产和生态系统,实现对复杂系统行为定量化、
可视化高效感知和认知的理论、方法与技术体系,以
及相应的支撑服务平台和应用系统。数字植物研究的
主要目标是对农林植物-环境的形态结构、物理属性、
生理生态过程等进行定量化、可视化表达,通过结构-功能相互作用过程的并行模拟解释农林植物-环境的
关系,具有多维性、可视性、交互性、科学计算等技
术优势,将深刻改变农业科学研究活动和认知植物的
方式,促进农业科学研究和生产方式向数字化、智能
化和可视化方向转变。 由于农林植物-环境系统的复杂性,构建纯粹意义
上的“数字植物”无疑是一项巨大的工程。因此,“数
字植物”在实现上需要经历“数字可视植物”、“数
字物理植物”、“数字生理植物”和“数字智能植物”
四个阶段。通过“数字可视植物”建立植物的形态结
构,然后将植物的物理属性、生理属性和智能性加以
数字化,并与其形态结构相结合,构建外在形态与内
在功能过程都与真实植物相一致的数字植物模型。利
用这样的模型能够在计算机构建的虚拟空间中再现植
物的运动功能、环境响应功能、物质生产和分配功能、
细胞进化现象、群体竞争现象。建立这样的数字植物
是一个艰巨的任务,难以一蹴而就。为此,近几十年
来研究者通常围绕某个主题、某个层次或从某个角度
开展研究。例如在植物生长过程模拟[2-4]、植物器官三
维建模[2-4]、植物形态结构计算机辅助设计[8-10]、植物
功能-结构并行模拟[11-12]、冠层生理生态功能仿真计
算[13-15]、植物生物力学建模[16-17]等方面均开展了广泛
研究并取得了显著的进展。近几年,对于植物生长过
程模拟、植物功能-结构建模、植物形态结构交互设计
等方面的研究进展报道较多。相对而言,在植物组织
层次可视化、根系三维建模、群体三维重建、植物表
观材质建模等方面的研究综述较少。另一个较为明显
的趋势是,随着三维扫描技术及设备的广泛普及,基
于三维扫描数据进行植物形态结构的测量、分析和三
维重建已成为研究热点[18-20]。 数字可视植物是数字植物研究的重要组成部分,
是实现数字植物可计算、可直观展示的载体。关于植
物器官、植株形态结构的建模和生长过程模拟方面的
进展报道并不鲜见,本研究旨在通过对植物组织层次
可视化、根系三维建模、群体三维重建等近年来综述
较少的数字可视植物关键技术进行介绍,并结合相关
领域的 新发展趋势,对数字植物的进一步发展进行
分析并提出展望,以期为不断深化这一跨学科课题的
研究与应用提供有益参考。
1 植物组织层次三维建模和可视化
植物内部具有复杂的组织结构,探测、分析植物
器官的内部构造不仅有助于了解不同物种、不同品种
的形态结构和物理特性差异,而且是探明物质资源在
植物内部的运输利用过程的 直接手段。随着计算机
仿真技术的进步,部分研究者通过计算机仿真方法模

17 期 赵春江等:数字植物研究进展:植物形态结构三维数字化 3417
拟植物组织的生长发育过程[21-22](图 1)。这种方法
能直观展示肉眼难以观察到的植物器官内部的形态结
构变化过程,为传统植物学研究带来了新的技术手段。
但近年来随着各种先进成像设备,如断层扫描成像
(CT)、核磁共振成像(MRI)和光学投影层析技术
(OPT)等的成熟和普及,更多研究者采用这些仪器
设备辅助进行植物组织显微结构研究,成为研究植物
组织结构形态、功能特征的热点。 在植物组织形态研究中,共聚焦显微镜应用 广
并取得了很多重要发现,可对特定组织提供高分辨率
三维成像,成像分辨率已能达到微米层级[23-24] ,但对
样本厚度有一定限制(约 100μm)[25]。在国外,已有
一些基于 CT 和 MRI 等复杂成像设备对植物器官组织
进行功能-结构关系的研究报道。如 Windt 等[26]使用核
磁共振设备研究番茄在不同生长期和生长条件下茎内
木质部和韧皮部中水流量的变化。Horigane 等[27]使用
核磁共振成像检测米粒在浸泡期间的透水性等特征。
Lee 等[28]利用 CT 成像技术可视化显示植物内部结构,
以此研究木质部中水分输运机制及其中栓塞管道的水
填充过程。Brodersen 等[29]利用 CT、光学显微镜、扫
描电镜和染料注射方法研究葡萄藤茎中维管束,表明
植物的某些解剖特征易导致疾病和栓塞。Lee 等[30]利
用 OPT 获取植物器官光学图像,进而进行器官内部形
态结构和基因表达的定量分析和可视化展示,其研究
表明,OPT 适用于苗、叶片、花、根、种子、胚胎、
分裂组织等大多数植物器官。Dercksen 等[31]通过连续
组织切片图像实现了谷粒的三维体素重建(图 2),
这种通过精细网格建立的三维体素模型能进一步与植
物组织的标记数据整合,从而在三维空间中对植物器
官的内部结构进行可视化探测分析。
EmbryonicExtra-embryonic
Upper tierLower tierProtodermInnerHypophysis
Vascular tissueGround tissueQCColumella
InitialDaughter
图 1 拟南芥胚胎发育过程中的形态分化过程模拟[22]
Fig. 1 Geometric division in Arabidopsis embryogenesis[22]
图 2 基于组织切片的谷粒三维体素重建流程[31]
Fig. 2 3D volume reconstruction of grains based on serial sections[31]
上述采用 MRI、CT 成像技术能原位获取组织断
层信息和一些生理参数,但试验条件要求较高,而且
成像分辨率往往难以满足植物维管束精确建模要求;
相比而言,传统组织切片和光学显微成像技术在获取
植物组织显微图像的像素对比度和分辨率、可用样本
尺寸等方面仍具有不可替代的优势[30]。使用石蜡切片
和显微成像相结合获取植物器官系列二维图像数据的
技术方案简单易行、成本低廉,为定量分析整株植物

3418 中 国 农 业 科 学 48 卷
木质部解剖结构特征提供了可靠手段。虽然组织切片
制作的工艺要求较高,过程颇为繁琐,但一些实用实
验技术手段能保证切片制作效率和精度[31]。而光学显
微成像技术具有数据获取便利、无损等优势,也是目
前进行植物内部形态结构测量和分析的有效手段。
2 根系三维建模和可视化
根是植物 重要的器官之一。根系构型不仅直接
影响植物的水分和养分吸收能力,同时通过与土壤环
境相互作用影响植物生产效率,成为影响植物生长的
关键通道,也是认识、分析和评价作物与土壤适应程
度的重要指标[1-2]。此外,由于根生长在土壤里,观察、
测量极不方便,这也为植物根系形态结构研究带来了
极大挑战。因此,植物根系的测量和三维建模研究吸
引了诸多研究者的关注。 2.1 植物根系探测方法
为了解植物根系在土壤中的生长过程、获取根系
形态结构数据,人们开展了很多尝试。其中直接在土
壤中挖掘无疑是 直接的一种手段,例如有研究者用
根钻法直接从土壤中挖掘出根系并清洗或直接在挖掘
过程中对根系的形态结构进行测量[32-33];也有人将植
物种植在根系保护装置中,于根系生长一定时期将装
置与根系作为一个整体取出后再进行人工清洗得到无
土壤的根系[34]。不过,直接挖掘法属于破坏性测量,
根系挖掘后植物即便重新栽植并继续生长,其根系形
态和生长发育都会受到影响,因此更适用于植物某个
特定生长时期根系构型的测量。 针对直接挖掘法的缺点,研究者提出了各种非破
坏性植物根系探测方法,一般这类方法统称为原位测量
法,包括根箱法[35]、微根管法[36]、地面穿透雷达法[37]、
水培法[38]、冷凝胶培养法[39]、穿透射线成像法[40]等。
这些方法各有优缺点,其中微根管法操作简单、成本
低,是目前大田进行根系动态生长监测 常用的方法
之一,其原理是通过在根系生长位置埋插微根管,在
根系生长时期于微根管内放置旋转扫描装置进行根系
扫描并获得根系的图像信息。但这种方法获得的每张
图像中只能得到根系的少量局部信息,如何从多幅图
像中恢复全局信息并重建根系三维形态属技术难点。
雷达法针对近地面粗壮根系,采用地面穿透雷达扫描
根系,其局限性在于仅适用于粗壮根系物如树木等,
对于形态结构较小的作物根系效果不佳。而水培、冷
凝胶培养等特殊培养环境法虽然可使根系生长发育全
过程透明可见,但由于这种方法使植物脱离了其自然
的土壤生长环境,其根系的生长发育可能较自然环境
存在较大差异。X 射线计算机断层扫描(XCT)[41]、
核磁共振(MRI)[42]等穿透射线成像技术是近年来应
用较多也较为成熟的根系探测方法,与其他原位测量
方法相比,该方法更方便、准确、快捷,但目前通过
穿透射线仍仅能获取范围较小土块中的立体几何构型
参数,且其所用仪器较昂贵,难以普及。 在以上介绍的根系探测方法中,大多数方法都获
取了根系部或整体图像,因此,如何从图像中提取根
系的形态结构信息是另一个技术难题。研究者在这方
面开展了广泛研究,并开发了诸多软件工具。早期的
根系图像分析软件仅进行图像中根系统计参数的计
算,如总根长、根面积、根尖数量等,典型软件如 DT-
A
B C
0° 72° 144° 216°
A:植物在圆筒中的凝胶中培养;B:通过放大根部区域获取图像;C:从不同角度获取的裁剪后的根系图像 A: Crop growing in gel in a cylinder; B: Images were acquired by zooming; C: Cropped images from multiple angles
图 3 凝胶培养法获取根系图像过程[39]
Fig. 3 Gel-based growth platform for obtaining images of root system[39]

17 期 赵春江等:数字植物研究进展:植物形态结构三维数字化 3419
SCAN、DigiRootTM[43]。后来,为了能处理背景更复杂
的图像并具有更自动、快速的处理过程,研究者们考
虑在图像分析中加入人类视觉系统方法,即采用半自动
或完全手动的方式进行根系识别,因此,出现了如
EZ-Rhizo[44]、DART[45]等更先进软件。RootReader3D[46]
软件则能够通过序列图像进行根系空间分析和三维重
构,实现了植物苗期根系表型的高通量信息获取。 2.2 根系三维建模与可视化
对植物根系进行三维建模的主要目的是以三维可
视方式了解植物根系的表型特征,或者展示根系形态
结构的生长发育过程。 由于根系生长在土壤中,人们难以直观获得其三
维形态信息,因此,部分研究者通过计算机模拟的方
法建立根系三维结构、或者模拟根系生长发育过程。
张吴平等[47]采用功能-结构植物模型 GreenLab 模拟了
作物的根系拓扑结构,并以三维可视化方式给出根系
的形态结构空间分布。邓旭阳等[48]采用粒子系统方法
对玉米根系进行了三维可视化模拟。Han 等[49]利用经
验数据,采用四象限法(简称 RULD 方法)对大豆根
系及根瘤进行三维建模,并利用信号传递机制模拟大
豆根系生长过程。Casson 等[50]基于细胞分裂与细胞间
信号传递模拟了拟南芥根系的动态生长过程。Leitner等 [51]基于 L-系统,通过定义生长规则建立了植物根
系生长模型。Dupuy 等[52]通过密度分布函数描述根系
分布,并结合与环境之间的反馈,建立了根系生长模
型。Pagès 等[53]提出了 ArchiSimple 模型简化根系空间
分布的模拟算法,从而提高根系建模效率。部分研究
者则采用参数化建模和计算机辅助设计方法进行植物
根系的三维建模和可视化[54-55]。 与计算机模拟法相比,基于测量数据的根系三维
重建是近年来研究更多的方法。例如,基于 CT 图像、
X 光图像的根系三维重建已有大量研究 [56-58]。但由于
成像范围限制,这种基于实测图像的根系三维重建方
法往往仅能针对局部根系进行三维重建,且往往由于
植物生长后期根系内部过于密集,测量图像难以准确
恢复根系的三维形态。三维扫描法[59]是另一种根系三
维重建方法,但由于根系的不可见性,很难对土壤中
的根系进行三维扫描,因此,该方法并不适用于自然
生长植物根系的三维重建。相对地,三维数字化方法
是一种更为直观的三维重建方法。该方法首先通过三
维数字化设备获取根系的空间拓扑结构数据,然后从
这些数据中重建根系的三维形态结构[33]。这种方法虽
然存在破坏性测量和受人为因素干扰等缺点,但实用
a:黏土种植;b:砂壤土种植 a: Grown in clay loam; b: Grown in loamy sand
图 4 采用 X光扫描图像重建的西红柿根系三维模型[58]
Fig. 4 3D models of tomato root reconstructed from X-ray
scanned images[58]
性强、重建成本低、范围大,仍不失为目前进行植物
根系三维重建较为有效的方法之一。 总之,植物根系测量和三维重建是数字植物的重
要内容。虽然国内外在根系探测和三维建模方面已有
较多研究,但这些方法都有一定局限性。造成这一问
题的根本原因在于植物根系深埋于地下的不可见性及
其与生长环境相互作用的复杂性。因此,在植物根系
形态数据获取手段难以取得显著突破的条件下,采用
新的方法解决有限数据条件下根系构型的测量与解析
仍是一个值得深入探索的问题[60]。
3 群体三维重建与定量分析
在农林生产实践中,植物往往群体栽培。因此,
对植物群体分析建模与生产实际更为接近。 早期人们采用三维数字化的方法进行植物群体形
态结构的空间信息采集并实现群体的三维重建[61-62]。
尽管这种方法能够较好地实现植物形态结构的三维重
构,但由于植物群体往往具有成千上万个器官,利用
三维数字化仪进行数据获取仍然是一件极耗时耗力的
工作,特别是结构复杂的群体。 为了克服直接采用三维数字化测量方法进行植物
群体重建的不足,Louarn 等 [63]提出了一种结合植物
冠层形态特征参数模型的群体重建方法。他们首先通
过田间试验获取的数据拟合枝条与叶的空间分布模
型,然后利用随机方法产生枝条,并采用一个类似于
混沌介质(turbid-medium)的包围体描述每根枝条的
大体轮廓,在包围体内叶子的空间分布模型生成叶子,
建立单株三维模型进而生成群体三维模型(图 5),

3420 中 国 农 业 科 学 48 卷
其在图像、形态特征和冠层光截获等方面与真实葡萄
群体相比误差较小,且群体三维重建所需的数据获取 工作量比传统三维数字化方法大大减少。
与此类似,Lu 等[64]也提出了基于统计模型的园
艺植物群体三维重建方法,以减少基于三维数字化
测量数据进行植物群体三维重建的数据采集工作
量。该方法首先通过真实植物栽培试验获取植株和
群体形态特征数据,包括叶长、叶宽、节间长度、
叶方位角、叶倾角等,然后通过数学分析构建群体
尺度形态特征统计模型, 后通过这些统计模型自
动生成群体三维模型(图 6-a)。在此基础上,还可
以三维激光扫描仪获取的叶片精细三维模型为模
板,替代群体植株中的叶片曲面,生成更高精度的
群体三维模型(图 6-b)。Qian 等[65-66]通过进一步
分析植物群体冠层的形态异质性特征,改进了植物
群体冠层的形态特征统计模型,实现了准确性更高
的群体三维重建,并将三维群体模拟方法用于作物
株行距的 优化计算分析,初步证明了基于冠层形
态特征模型的植物群体三维建模方法能为农业管理
措施的决策优化提供辅助决策支持。
图 5 真实葡萄群体和计算机重建的虚拟葡萄群体的对比[63]
Fig. 5 Comparison of real and simulated grapevine canopy[63]
a:不采用叶片模板的结果;b:采用叶片模板的结果 a: Result without using leaf templates; b: Result using leaf templates
图 6 黄瓜群体重建结果[64]
Fig. 6 3D simulated result of soybean root with nodulation[64]
此外,10 多年来基于三维扫描点云的植物三维重
建也得到广泛研究。但早期的研究主要集中在单株树
木,主要原因在于树木形态结构较复杂,冠层枝叶
之间的相互遮挡很容易造成三维点云上枝条信息的
缺失,因此,单株树木的三维重建已是一个技术难
题。直到 近,一些研究者才尝试直接从三维扫描
仪获取的树木群体三维点云中进行多植株的形态信
息提取和场景三维重建。Livny 等[67]利用一种全局优
化方法从激光扫描得到的自然场景三维点云中实现
了植物群体的自动三维重建,该方法无需经过常规
的单树分割策略,因此重建速度快、无需手工交互。
不过,该方法的重建精度尚待提高,更适用于景观

17 期 赵春江等:数字植物研究进展:植物形态结构三维数字化 3421
展示等偏重可视化展示的应用场合。Leeuwen 等[68]
为比较分析不同树木群体冠层的光照特性,在三维
点云中提取树木的枝干骨架,基于一些形态特征参
数,采用 Arbaro 软件生成每棵树的初始三维模型,
根据树木的理论生长空间对树的三维模型进行缩
放,实现了大规模树木群体的三维重建(图 7)。该
方法很好地利用了树木的三维点云信息,同时树木
三维模型缩放策略也能够较好地减少树木之间的相
互穿插现象。由于远程三维激光扫描仪能够快速大
范围获取树木群体的三维形态数据。这种方法近年
来正越来越多地用于林业调查、树木生理生态建模
等领域。自然树林中植物之间的相互遮挡十分普遍,
因此开发更准确的形态特征参数提取方法、更自动
的计算过程仍然是亟待解决的问题。
图 7 两个树木群体的骨架提取和冠层重建结果[68]
Fig. 7 Results of extracted skeleton and reconstructed 3D
canopy of two plots[68]
4 植物表观材质建模
自然界中,植物的多样性和复杂性除了体现在
其形态结构外,另一个重要因素在于其器官材质表
面丰富的颜色和纹理。一方面,不同物种、品种的
植物有不同的颜色和纹理,甚至同一株植物同种器
官的颜色和纹理也会因其生长位置不同而存在较大
差异;另一方面,植物的颜色和纹理随着其年龄的
变化而不断变化。同时,土壤、水分和肥料等环境条
件植物的颜色和纹理有着重要影响。可以说,植物器
官的颜色和纹理是其自身喜怒哀乐的 直接表达通
道。因此,对于植物表观材质的建模是数字植物不可
或缺的研究内容之一。近年来,研究者围绕植物表观
材质建模的研究主要包括静态建模和动态建模。 4.1 静态建模
静态建模是指建立植物器官某个时期的瞬时表观
质感模型,重点关注如何在计算机上逼真地展现植物
器官表面材质的光学特征(如叶片的半透明现象)和
表面细节(如纹理、绒毛等)。 通过算法生成逼真的植物器官彩色图像是虚拟
现实领域的一种流行方法[69]。但更多的研究者则关
注如何在计算机上展现部分植物器官特有的光学特
性,其中,关于植物叶片的次表面散射模拟是 引
入关注的问题。考虑到植物叶片的生理结构特征,
Hanrahan等[70]构建了多层模型并采用Monte-Carlo积分
计算出叶片表面的双向反射分布函数(bidirectional reflectance distribution function,BRDF)以及双向透
射分布函数(bidirectional transmittance distribution function,BTDF)对植物叶片的次表面散射现象进
行模拟。此后,研究者进一步对以 BRDF+BTDF 为
基础的植物叶片光学特性模拟方法进行了改进,并取
得了非常逼真的模拟效果。例如,Wang 等[71]提出了
一种实时叶片渲染算法,利用该方法推导出一个参数
模型计算叶片的 BRDF 和 BTDF,并通过采集真实叶
片的空间 BRDF 和 BTDF 数据,拟合参数模型中的参
数,使其更符合真实叶片表观,同时通过扩展光辐射
传输预计算(precomputed radiance transfer,PRT)算
法以获得全局光照效果,在运行效率上也达到了实时
性,其模拟结果见图 8。但在以上方法中,植物叶片
的 BRDF 和 BTDF 的测量和拟合是一件费时费力的工
作,使得该方法难以广泛普及。
图 8 植物叶片半透明可视化模拟效果[71]
Fig. 8 Real-time rendering of semi-translucent plant leaves[71]

3422 中 国 农 业 科 学 48 卷
4.2 动态建模
植物表观材质动态建模目的主要在于模拟植物器
官老化过程或在某些环境胁迫条件下表面颜色纹理的
变化过程。同样,叶片作为植物 重要的器官之一,
其对植物的形态构造和视觉效果都有十分重要的影
响。因此,大部分关于植物表观变化过程的建模和仿
真研究都是以叶片为对象。 叶片颜色变化主要是由其内部色素含量变化而引
起,很多学者基于这种思想进行叶色老化模拟。
Braitmaier 等[72]通过改变叶片内部叶绿素、胡萝卜素
含量控制 终光谱,从而绘制出四季不同的叶色变化,
但是由于缺少数据,对于各种色素对光谱的影响采用
了简单的指数形式,实际上并不准确。Zhou 等[73]不仅
考虑了色素的影响,还对温度、光照、降水量等环境
因素进行整合,构建了基于生物学驱动的系统,可以
模拟枫叶由绿到红的颜色老化过程。Desbenoit 等[74]
应用马尔可夫链来描述叶片老化过程,其设置一个初
始状态以及一些老化状态,通过环境函数(由温度、
湿度决定)以及时间决定向不同状态的转化概率,
从而得到叶片的老化过程。Wang 等[75]提出了一种
基于表观流形的老化模拟方法,先利用线性光源反
射计获取物体的表观材质数据,对于被采样物体的
要求是样本中需包含老化过程出现的所有表观特
征,得到材质数据之后,构建表观的流形,并通过
人工交互指定老化度及时间轴,这样表观上的每个
点会根据时间以流行为依据进行老化模拟,以此方
法实现了植物叶片、果实老化中表面颜色变化过程
的可视化仿真,效果逼真。迟小羽等[76]从多叶片样
本上获取了表观材质,并利用样条曲线进行老化插
值,可以反演、外推得到不同老化程度下的表观。
Gu 等[77]提出了一种时间-空间变化的双向反射分布
函数,在一段时间内采集物体老化的 BRDF 数据,
并进行参数化控制物体的老化过程。田原等[78]提出
了基于监督 BRDF 描述的植物叶片表观绘制模型,
通过系统分析线性光源变化下拍摄的叶样本图像序
列计算得到定量的反射与透射特性,然后通过对植
物叶片的老化表观做独立成分分析(ICA),分离
出两种独立成分:绿色色素和老化色素的相对量,
并建立了植物叶片表观合成模型,实现了植物叶片
老化的正向和逆向可视化模拟。 以上植物器官表观材质变化模拟研究大都集中在
叶片物理材质属性建模上,却忽略了更为本质的生理
属性,个别研究虽然考虑了生理因子,但多是简单的
定性分析,并且获得的真实感效果较差。同时,由于
目前方法缺少对生理因子的融合,导致其很难与农
业研究或者农业应用进行整合。此外,已有的研究
仅关注正常状态下的植物表观现象,而对于环境胁
迫条件下的表观变化过程却很少关注。为此, 近
陆声链等 [79] 提出了一种基于相对叶绿素含量
(SPAD)的植物叶片颜色变化过程仿真方法,该方
法通过连续对正常生长状态下的植物叶片图像采集
及 SPAD 值测定,建立了叶片颜色分量与 SPAD 值
的数学关系模型, 后结合叶片色素浓度分布图对
叶色空间分布进行仿真,获得了较好的真实感效果。
苗腾等[80]将叶绿素、胡萝卜素、叶片结构参数等生
理因子引入 BRDF 和 BTDF 的计算中,实现了生理
因素驱动植物叶片表观颜色变化仿真。同时,利用
该方法,通过改变色素变化速率纹理的方式,可模
拟植物叶片在环境胁迫条件下的表观变化效果,如
图 9 所示。
a:真实图片;b:模拟效果 a: Real photo; b: Simulated image
图 9 黄瓜叶片锰过剩症状真实图片与模拟效果对比[80]
Fig. 9 Comparison of real image of manganese deficiency
cucumber leaf and simulated image[80]
病害也是造成植物器官表观发生显著变化的主
要原因,同时病害也是农学研究和生产中重点关注
的问题。为实现病害条件下的植物表观变化过程仿
真。苗腾[81]以白粉病为对象,基于病斑运动分布及
运动扩散假设,采用计算机图形学的 Shell 渲染算
法,实现了植物叶片感染白粉病过程的逼真可视化
模拟(图 10)。 总之,在植物表观材质建模和可视化仿真方面,
近年来研究者开展了广泛的研究并取得了长足的进
步。尤其是在静态模型方面,采用 BRDF+BTDF 的植
物表观材质模型已能很好地模拟植物叶的高光、半透

17 期 赵春江等:数字植物研究进展:植物形态结构三维数字化 3423
图 10 模拟白粉病在叶片上的扩展过程[80]
Fig. 10 Simulating the growing process of powdery mildew[80]
明等光学特性。这类材质模型中的参数数值对 后的
模拟效果有重要作用,因此,设定符合植物表观光学
特征的模型参数是模拟的关键。当前研究多基于数据
对参数进行拟合,但对叶片表观数据的采集过程复杂、
对采集环境要求高、工作量大,因此,在实际中的推
广应用会受到限制。而在植物表观动态变化过程模拟
方面,虽然围绕某种特定植物器官表观变化现象提出
了一些较好的模拟方法,但目前的方法还有很大局限
性,很多环境因素,包括温度、水分、肥料、阳光等
都十分重要。只有重复考虑这些环境因素,才能更好
地模拟植物表观在各种环境条件下,包括环境胁迫和
病害下的变化过程和结果。
5 总结和展望
笔者从植物组织层次可视化、根系三维测量、植
物群体三维重建和植物材质建模等方面介绍了数字可
视植物 近几年的新进展。需要指出的是,从虚拟植
物到数字植物,植物建模和仿真的研究已有几十年的
历史, 初也曾得到诸多应用预期[82-83]。但直到今天,
这项研究的现实意义尚未得到很好的体现,其实用价
值远未达到人们预期。当前,植物建模 实际的应用
成果是植物形态结构的三维建模,其已被广泛应用于
影视和动漫游戏制作、产品展示、广告宣传等领域。
但在曾被人给予厚望的作物株型选育、栽培措施优化
等农学应用方面,实际应用很少,大部分还停留在研
究探索阶段。 究其原因,主要还在于已有的关于植物形态结构
发展过程、植物生理生态过程、植物与环境的相互作
用关系等的模型还过粗、过于简化,尚无法准确模拟
植物在不同真实环境条件下的自然发展过程和结果。
另一个客观现实是,当前数字植物研究大部分仅在物
种尺度上进行,且缺乏不同地理环境条件的对比试验
和验证。而实际生产中,往往是以品种为单位。同一
植物物种,不同品种在形态、生理和生产能力等方面
均存在较大差异,同一品种在不同地理环境条件下也
可能表现出显著差异。 因此,以下几方面的研究尤为迫切并预期成为未
来几年的研究重点: 5.1 基于高通量信息获取的植物表型组学研究
在基因和分子尺度上,研究者围绕植物的结构
和功能已经开展了广泛研究。但我们对植物生理过
程及其功能的认识远远不够,更难以满足人类对植
物生命、生产过程进行干预和控制的需要。植物的
形态结构与其内部生理有着密切联系,为此还需要
在器官、植株和群体等尺度上对植物的表型结构信
息进行解析,深入分析和挖掘植物表型、基因及生
理功能的联系。而高光谱相机、X 光、激光扫描等
先进自动化检测技术装备的快速发展进一步促进了
植物表型信息的自动、快速获取和分析的研究与应
用。 当前,植物表型的高通量检测和分析是植物学、
作物遗传育种等领域的研究热点,其目的是通过集
成应用可见光、近红外、荧光、红外等成像技术以
及图像处理、特征提取、三维重建等智能算法,实
现植物表型、生理和物理信息的连续获取和快速解
析。德国 Leibniz 遗传学会与作物科学研究所、法国
农科院、澳大利亚 Adelaide 大学、英国草地研究所、
中国科学院、中国农业科学院、浙江大学等国际著
名研究机构均围绕这一主题开展了广泛研究。目前
也已有一些研究成果进行了商业化,如 Scanalzer HTS 、PlantEye、PlantScan 等。同时,包括 Bayer CropScience、BASF SE、Pioneer Seed Company、Monsanto Company、北大荒种业集团等在内的国内
外大型种业企业也纷纷利用高通量植物表型检测技
术开展辅助育种工作。 可以预期,随着高通量植物表型学研究的深入,
信息提取方法将更自动、快速和高精度,更多的植
物表型和生理参数高通量获取成为可能,这将进一
步促进人们更深入、透彻地认识植物形态结构形成
机制及其与功能的关系,成为创新作物遗传育种、
株型筛选、植物生理学、植物病理学等领域的研究
手段。

3424 中 国 农 业 科 学 48 卷
5.2 根冠一体化建模 由于植物生理过程和形态结构的复杂性,现有数
字植物研究中,往往将植物的地上部和根系作为两个
部分分别考虑,以简化建模难度。然而,根系是植物
与外界环境进行交互的 重要器官之一,脱离地下而
仅考虑地上部或分而治之的建模思想都可能因考虑
不周而导致 终的模型与现实脱节。因此,未来数字
植物研究的重点任务之一,就是进行植物根-冠一体
化建模。特别是面向株型设计、产量预测、生长过程
模拟等应用场合,需要同时考虑植物根系和地上部。
这样建立的模型才能更好地与真实植物接近,得到更
好的模拟结果并能够在现实农业科研和生产中投入
应用。 5.3 大数据驱动的知识发现和建模
随着物联网技术的快速发展,各种传感器广泛
普及于农田、温室、果园等生产基地。随之而来的
是大量、连续的植物生长过程和环境数据。但目前
这些数据尚未得到很好挖掘和利用。例如,目前植
物形态变化过程建模研究很多还在采用手工测量或
三维扫描等需要大量人工参与的操作进行数据采
集。而通过网络摄像头拍摄到的植物生长过程视频
尚没有得到很好利用。若能够从这些连续视频中提
取解析植物生长中各种形态指标,将极大减少传统
方法的工作量,同时还能够得到粒度更细、更连续
的数据。因此,如何从农业物联网的大数据中提取
出植物形态和生理指标,并结合环境监测数据,发
现植物形态和生理、功能间的知识规则,进而构建
用于植物生长、产量模拟的数学模型,将是未来值
得深入研究的课题。 5.4 品种尺度的建模和仿真
当前的数字植物研究几乎都是在物种尺度上进
行。但现实农业生产中,往往是以品种为尺度。相同
物种的植物,不同品种间不但外观形态上相异,生理
属性、物理性能和产量等方面也有较大差异。这表明,
在进行植物模型构建时,需要考虑品种特征。或者说,
只有具有品种识别度的模型,才能更好地满足现实生
产的应用需要。 5.5 功能-结构模型与表观材质模型的结合
已有研究中,对植物功能-结构模型和表观材质模
型往往单独考虑。事实上,植物的功能与其形态结构
和表观纹理都有重要联系。例如,缺水、缺肥、病害
等条件下,植物的形态和表观颜色都会随之变化。或
者说,形态和表观都是植物对环境条件响应的显示通
道。因此,在植物功能-结构并行模拟中,需要考虑植
物表观的同步响应。 总之,数字植物研究是一项具有重要研究意义
和现实价值,同时也是十分复杂的科学任务,涉及
农学、植物学、数学、物理等多个学科,需要多学
科的交叉合作和长期、大量试验积累以及知识发
现,才有可能使其研究成果更好地服务于农业科研
和生产实践。期待更多研究者关注并持续投入数字
植物研究,共同推动这项研究向深层次和实用化发
展。
References
[1] 赵春江, 陆声链, 郭新宇, 肖伯祥, 温维亮. 数字植物及其技术体
系探讨. 中国农业科学, 2010, 43(10): 2023-2030.
Zhao C J, Lu S L, Guo X Y, Xiao B X, Wen W L. Exploration of
digital plant and its technology system. Scientia Agricultura Sinica,
2010, 43(10): 2023-2030. (in Chinese)
[2] Prusinkiewicz P, Lindenmayer A. The Algorithmic Beauty of Plants.
New York: Springer-Verlag, 1990: 228.
[3] Kaandorp J A. Fractal Modeling Growth and Form in Biology. New
York: Springer-Verlag, 1994.
[4] 赵星, de Reffye Philippe, 熊范纶, 胡包钢, 展志岗. 虚拟植物生长
的双尺度自动机模型. 计算机学报, 2001, 24(6): 608-615.
Zhao X, de Reffye Philippe, Xiong F L, Hu B G, Zhan Z G. Dual-scale
automaton model for virtual plant development. Chinese Journal of
Computers, 2001, 24(6): 608-615. (in Chinese)
[5] Prusinkiewicz P, Mündermann L, Karwowski R, Lane B. The use of
positional information in the modeling of plants//In Proceedings of
SIGGRAPH, ACM Press. MiamiBeach, USA, 2001: 289-300.
[6] 郑文刚, 郭新宇, 赵春江, 王纪华. 玉米叶片几何造型研究. 农业
工程学报, 2004, 20(1): 152-154.
Zheng W G, Guo X Y, Zhao C J, Wang J H. Geometry modeling of the
maize leaf canopy. Transactions of the Chinese Society of Agricultural
Engineering, 2004, 20(1): 152-154. (in Chinese)
[7] Hong S M, Simpson B, Baranoski G V G. Interactive venation-based
leaf shape modeling. Computer Animation and Virtual Worlds, 2005,
6(3-4): 415-427.
[8] Deussen O. Digital Design of Nature-Computer Generated Plants and
Organics. New York: Bernd Lintermann Springer-Verlag, 2005.
[9] Wither J, Boudon F, Cani M P, Godin C. Structure from silhouettes: A
new pradigm for fast sketch-based design of trees. Computer Graphics
Forum, 2009, 28(2): 541-550.
[10] Zhu X, Jin X, You L. High-quality tree structures modeling using local

17 期 赵春江等:数字植物研究进展:植物形态结构三维数字化 3425
convolution surface approximation. The Visual Computer, 2015, 31(1):
69-82.
[11] Hu B G, de Reffye P, Zhao X, Yan H P, Kang M Z. GreenLab: A new
methodology towards plant functional-structural model-structural
aspect//2003' International Symposium on Plant Growth, Modeling,
Simulation, Visualization and Their Applications. Beijing, China. 2003:
21-35.
[12] Vos J, Marcelis L F M, Visser J B. Functional-Structural Plant
Modelling in Crop Production. Netherlands: Wageningen University
and Research Center, 2007: 1-12.
[13] Chelle M, Andrieu B. The nested radiosity model for the distribution
of light within plant canopies. Ecological Modeling, 1998, 111: 75-91.
[14] Da Silva D, Boudon F, Godin C, Sinoquet H. Multiscale framework
for modeling and analyzing light interception by trees. Multiscale
Modelling and Simulation, 2008, 7(2): 910-933.
[15] Dragoni D, Laksa A N, Piccioni R M. Transpiration of apple trees in a
humid climate using heat pulse sap flow gauges calibrated with
whole-canopy gas exchange chambers. Agricultural and Forest
Meteorology, 2005, 130: 85-94.
[16] Goriely A, Neukirch S. Mechanics of climbing and attachment in
twining plants. Physical Review Letters, 2006, 97: 184-302.
[17] 梁莉, 郭玉明. 作物茎秆生物力学性质与形态特性相关性研究. 农
业工程学报, 2008, 24(7): 1-6.
Liang L, Guo Y M. Correlation study of biomechanical properties and
morphological characteristics of crop stalks. Transactions of the
Chinese Society of Agricultural Engineering, 2008, 24(7): 1-6. (in
Chinese)
[18] Livny Y, Yan F, Olson M, Chen B, Zhang H, EI-Sana J. Automatic
reconstruction of tree skeletal structures from point clouds. ACM
Transactions on Graphics, 2010, 29(6): 151-158.
[19] Li Y, Fan X, Mitra N J, Chamovitz D, Cohen-Or D, Chen B.
Analyzing growing plants from 4D point cloud data. ACM
Transactions on Graphics, 2013, 32(6): 2504-2507.
[20] Delagrange S, Jauvin C, Rochon P. Pypetree: A tool for reconstructing
tree perennial tissues from point clouds. Sensors, 2014, 14: 4271-4289.
[21] Smith C, Prusinkiewicz P. Simulation modeling of growing tissues. In
4th International Workshop on Functional-Structural Plant Models,
2004: 365-370.
[22] Yoshida S, de Reuille B P, Lane B, Bassel G W, Prusinkiewicz P,
Smith R S, Weijers Dolf. Genetic control of plant development by
overriding a geometric division rule. Developmental Cell, 2014, 29(1):
75-87.
[23] Betz O, Wegst U, Weide D, Heethoff M, Helfen L, Lee W K, Cloetens
P. Imaging applications of synchrotron X-ray phase-contrast
microtomography in biological morphology and biomaterials science.
I. General aspects of the technique and its advantages in the analysis
of millimetre-sized arthropod structure. Journal of Microscopy, 2007,
227(1): 51-71.
[24] Kerwin J, Scott M, Sharpe J, Puelles L, Robson S C, Margaret M,
Ferran J L, Feng G, Baldock R, Strachan T, Davidson D, Lindsay S. 3
dimensional modelling of early human brain development using
optical projection tomography. BMC Neuroscience, 2004, 5(1): 27.
[25] Sands G B, Gerneke D A, Hooks D A, Green C R, Smaill B H,
Legrice I J. Automated imaging of extended tissue volumes using
confocal microscopy. Microscopy Research and Technique, 2005,
67(5): 227-239.
[26] Windt C W, Gerkema E, Van As H. Most water in the tomato truss is
imported through the xylem, not the phloem: A nuclear magnetic
resonance flow imaging study. Plant Physiology, 2009, 151(2):
830-842.
[27] Horigane A K, Takahashi H, Maruyama S, Ohtsubo K. Water
penetration into rice grains during soaking observed by gradient echo
magnetic resonance imaging. Journal of Cereal Science, 2006, 44(3):
307-316.
[28] Lee S, Kim Y. In vivo visualization of the water-refilling process in
xylem vessels using X-ray micro-imaging. Annals of Botany, 2008,
101(4): 595-602.
[29] Brodersen C R, Choat B, Chatelet D S, Shackel K A, Matthews M A,
McElrone A J. Xylem vessel relays contribute to radial connectivity in
grapevine stems (Vitis vinifera and V. arizonica; Vitaceae). American
Journal of Botany, 2013, 100(2): 314-321.
[30] Lee K, Avondo J, Morrison H, Blot L, Stark M, Sharpe J, Bangham A,
Coena E. Visualizing plant development and gene expression in three
dimensions using optical projection tomography. The Plant Cell, 2006,
18: 2145-2156.
[31] Dercksen V J, Brüß C, Stalling D, Gubatz S, Seiffert U, Hege H C.
Towards automatic generation of 3D models of biological objects
based on serial sections. Visualization in Medicine and Life Sciences.
Heidelberg: Springer, 2008: 3-25.
[32] Neumann G, George T S, Plassard C. Strategies and methods for
studying the rhizosphere-the plant science toolbox. Plant Soil, 2009,
321: 431-456.
[33] Wu J, Guo Y. An integrated method for quantifying root architecture
of field-grown maize. Annals of Botany, 2014(114): 841-851.
[34] 管建慧, 郭新宇, 王纪华, 刘克礼, 郭晓东. 玉米不同部位根系生
长发育规律的研究. 玉米科学, 2007, 15(6): 82-85, 88.

3426 中 国 农 业 科 学 48 卷
Guan J H, Guo X Y, Wang J H, Liu K L, Guo X D. Study on growth
and development rule of different parts of maize root. Journal of
Maize Sciences, 2007, 15(6): 82-85, 88. (in Chinese)
[35] Hawkins H, George E. Effect of plant nitrogen status on the
contribution of arbuscular mycorrhizal hyphae to plant nitrogen
uptake. Physiologia Plantarum, 1999, 105(4): 694-700.
[36] 廖荣伟, 刘晶淼, 安顺清, 牛俊丽, 梁宏, 任三学, 乐章燕, 曹玉静,
李文静. 基于微根管技术的玉米根系生长监测. 农业工程学报,
2010, 26(10): 156-161.
Liao R W, Liu J M, An S Q, Niu J L, Liang H, Ren S X, Le Z Y, Cao
Y J, Li W J. Monitor of corn root growth in soil based on
minirhizotron technique. Transactions of the Chinese Society of
Agricultural Engineering, 2010, 26(10): 156-161. (in Chinese)
[37] 崔喜红, 陈晋, 关琳琳. 探地雷达技术在植物根系探测研究中的应
用. 地球科学进展, 2009, 24(6): 606-611.
Cui X H, Chen J, Guan L L. The application of ground penetrating
radar to plant root system detection. Advances in Earth Science, 2009,
24(6): 606-611. (in Chinese)
[38] Fang S, Yan X, Liao H. 3D reconstruction and dynamic modeling of
root architecture in situ and its application to crop. The Plant Journal,
2009, 60: 1096-1108.
[39] Iyer-Pascuzzi A S, Symonova O, Mileyko Y, Hao Y, Belcher H, Harer
J, Weitz J S, Benfey P N. Imaging and analysis platform for automatic
phenotyping and trait ranking of plant root systems. Plant Physiology,
2010, 152: 1148-1157.
[40] Gregory P J, Hutchison D J, Read D B, Jenneson P M, Gilboy W B,
Morton E J. Non-invasive imaging of roots with high resolution X-ray
micro-tomography. Plant and Soil, 2003, 255(1): 351.
[41] Mairhofer S, Zappala S, Tracy S R, Sturrock C, Bennett M, Mooney S
J, Pridmore T. RooTrak: Automated recovery of three-dimensional
plant root architecture in soil from X-ray microcomputed tomography
images using visual tracking. Plant Physiology, 2012, 158(2):
561-569.
[42] Schulz H, Postma J A, van Dusschoten D, Scharr H, Behnke S. Plant
root system analysis from MRI Images, computer vision, imaging and
computer graphics. Theory and Application, 2013: 411-425.
[43] Stefanelli D, Fridman Y, Perry R L. DigiRoot™: New software for
root studies. European Journal of Horticultural Science, 2009, 74(4):
169-174.
[44] Armengaud P, Zambaux K, Hills A, Sulpice R, Pattison R J, Blatt M R,
Amtmann A A. EZ-Rhizo: Integrated software for the fast and
accurate measurement of root system architecture. The Plant Journal,
2009, 57: 945-956.
[45] Le Bot J, Serra V, Fabre J, Draye X, Adamowicz S, Pagès L. DART: A
software to analyse root system architecture and development from
captured images. Plant and Soil, 2010, 326(1-2): 261-273.
[46] Clark R T, MacCurdy R B, Jung J K, Shaff J E, McCouch S R,
Aneshansley D J, Kochian L V. Three-dimensional root phenotyping
with a novel imaging and software platform. Plant Physiology, 2011,
156(2): 455-465.
[47] 张吴平, 郭焱, 李保国. 小麦苗期根系三维生长动态模型的建立与
应用. 中国农业科学, 2006, 39(11): 2261-2269.
Zhang W P, Guo Y, Li B G. Development and application of
three-dimensional growth model of root system in wheat seedling.
Scientia Agricultura Sinica, 2006, 39(11): 2261-2269. (in Chinese)
[48] 邓旭阳, 周淑秋, 郭新宇, 赵春江, 王纪华. 玉米根系几何造型研
究. 工程图学学报, 2004, 25(4): 62-66.
Deng X Y, Zhou S Q, Guo X Y, Zhao C J, Wang J H. Study on the
geometry modeling for corn root system. Journal of Engineering
Graphics, 2004, 25(4): 62-66. (in Chinese)
[49] Han L, Gresshoff P M, Hanan J. A functional-structural modeling
approach to autoregulation of nodulation. Annals of Botany, 2011,
107(5): 855-863.
[50] Casson S A, Lindsey K. Genes and signaling in root development.
New Phytologist, 2003, 158(2): 11-38.
[51] Leitner D, Klepsch S, Bodner G, Schnepf A. A dynamic root system
growth model based on L-systems. Plant and Soil, 2010, 332(1-2):
177-192.
[52] Dupuy L, Gregory P J, Bengough A G. Root growth models: Towards
a new generation of continuous approaches. Journal of Experimental
Botany, 2010, 61(8): 2131-2143.
[53] Pagès L, Moreau D, Sarlikioti V, Boukcim H, Nguyen C. ArchiSimple:
A parsimonious model of the root system architecture, 2012 IEEE 4th
International Symposium on Plant Growth Modeling, Simulation,
Visualization and Applications, 2012: 297-303.
[54] 徐其军, 汤亮, 顾东祥, 姜海燕, 曹卫星, 朱艳. 基于形态参数的
水稻根系三维建模及可视化 . 农业工程学报 , 2010, 26(10):
188-194.
Xu Q J, Tang L, Gu D X, Jiang H Y, Cao W X, Zhu Y. Architectural
parameter-based three dimensional modeling and visulization of rice
roots. Transactions of the Chinese Society of Agricultural Engineering,
2010, 26(10): 188-194. (in Chinese)
[55] 赵春江, 王功明, 郭新宇, 陈立平, 王纪华. 基于交互式骨架模型
的玉米根系三维可视化研究. 农业工程学报, 2007, 23(9): 1-6.
Zhao C J, Wang G M, Guo X Y, Chen L P, Wang J H. 3D visualization
of corn root system based on interactive framework model.

17 期 赵春江等:数字植物研究进展:植物形态结构三维数字化 3427
Transactions of the Chinese Society of Agricultural Engineering, 2007,
23(9): 1-6. (in Chinese)
[56] 向子云, 罗锡文, 周学成, 严小龙, 罗良平, 赵相胜. 多层螺旋 CT
三维成像技术观测植物根系的实验研究. CT 理论与应用研究,
2006, 15(3): 1-5.
Xiang Z Y, Luo X W, Zhou X C, Yan X L, Luo L P, Zhao X S. CT
theory and applications experimental study on observation of original
shape of crop root system with multi helical CT technology. CT
Theory and Applications, 2006, 15(3): 1-5. (in Chinese)
[57] Flavel R J, Guppy C N, Tighe M, Watt M, McNeill A, Young I M.
Non-destructive quantification of cereal roots in soil using high-
resolution X-ray tomography. Journal of experimental botany, 2012:
1-9.
[58] Mairhofer S, Zappala S, Tracy S, Sturrock C, Bennett M J, Mooney S
J, Pridmore T P. Recovering complete plant root system architectures
from soil via X-ray μ-Computed Tomography. Plant Methods, 2013, 9:
8.
[59] Fang S, Yan X, Liao H. 3D reconstruction and dynamic modeling of
root architecture in situ and its application to crop. The Plant Journal,
2009, 60: 1096-1108.
[60] 温维亮,郭新宇,赵春江,王传宇,肖伯祥. 作物根系构型三维探
测与重建方法研究进展. 中国农业科学, 2015, 48(3): 436-448.
Wen W L, Guo X Y, Zhao C J, Wang C Y, Xiao B X. Crop roots
configuration and visualization: A review. Scientia Agricultura Sinica,
2015, 48(3): 436-448. (in Chinese)
[61] Sinoquet H, Rivet P, Godin C. Assessment of the three-dimensional
architecture of walnut trees using digitising. Silva Fennica, 1997, 31:
265-273.
[62] 郭焱, 李保国. 玉米冠层三维结构研究. 作物学报, 1998, 24(6):
1006-1009.
Guo Y, Li B G. Studies on three dimensional structures of maize
canopy. Acta Agronomica Sinica, 1998, 24(6): 1006-1009. (in
Chinese)
[63] Louarn G, Lecoeur J, Lebon E. A three-dimensional statistical
reconstruction model of grapevine simulating canopy structure
variability within and between cultivar/training system pairs. Annals
of Botany, 2008, 101: 1167-1184.
[64] Lu S L, Guo X Y, Zhao C J, Qian T T, Wen W L, Du J J. Multi-scale
reconstruction of crop canopy//The 4th International Symposium on
Plant Growth Modeling, Simulation, Visualization and Applications.
Shanghai, China, 2012: 262-269.
[65] Qian T T, Lu S L, Zhao C J, Guo X Y, Wen W L. Heterogeneity
analysis of cucumber canopy in a solar greenhouse under natural
sunlight. Journal of Integrative Agriculture, 2014, 13(12): 2645-2655.
[66] Qian T T, Zhao C J, Guo X Y, Lu S L. Introducing an azimuth model
into functional-structural modeling of cucumber canopy. Sensor
Letters, 2014, 12(3-5): 892-896.
[67] Livny Y, Yan F, Olson M, Chen B, Zhang H, El-Sana J. Automatic
reconstruction of tree skeletal structures from point clouds. ACM
Transactions on Graphics, 2010, 29(6): 1-8
[68] Leeuwen M V, Nicholas C C, Thomas H, Michael A W, Glenn J N,
Darius S C. Automated reconstruction of tree and canopy structure for
modeling the internal canopy radiation regime. Remote Sensing of
Environment, 2013, 136: 286-300.
[69] Mochizuki S, Horie D, Cai D. Stealing Autumn Colors//ACM
SIGGRAPH 2005 Posters. Los Angeles, California, USA: ACM Press,
2005.
[70] Hanrahan P, Krueger W. Reflection from layered surfaces due to
subsurface scattering//Proceedings of the 20th annual conference on
Computer graphics and interactive techniques, Anaheim, CA, 1993.
[71] Wang L, Wang W, Dorsey J, Guo B, Shum H Y. Real-time rendering
of plant leaves//SIGGRAPH '06, New York, NY, USA, 2006.
[72] Braitmaier M, Diepstraten J, Ertl T. Real-time rendering of seasonal
influenced trees//Proceedings of Theory and Practice of Computer
Graphics, Los Alamitos: IEEE Computer Society Press, 2004:
152-159.
[73] Zhou N, Dong W M, Mei X. Realistic simulation of seasonal variant
maples//Proceedings of the 2th Plant Growth Modeling and
Applications, Los Alamitos: IEEE Computer Society Press, 2006:
295-301
[74] Desbenoit B, Galin E, Akkouche S, Grosjean J. Modeling autumn
sceneries//Proceedings of Eurographics’06, Vienna, Austria:
Eurographics Association, 2006: 107-110.
[75] Wang J P, Tong X, Lin S, Pan M, Wang C, Bao H, Guo B, Shum H Y.
Appearance manifolds for modeling time-variant appearance of
materials. ACM Transactions on Graphics, 2006, 25(3): 754-761.
[76] 迟小羽, 盛斌, 杨猛, 陈彦云, 吴恩华. 秋季植物叶子表观的模拟.
软件学报, 2009, 20(3): 702-712.
Chi X Y, Sheng B, Yang M, Chen Y Y, Wu E H. Simulation of autumn
leaves. Journal of Software, 2009, 20(3): 702-712. (in Chinese)
[77] Gu J, Tu C, Ramamoorthi R, Belhumeur P, Matusik W, Nayar S K.
Time-varying surface appearance: Acquisition, modeling and
rendering. ACM Transactions on Graphics, 2006, 25(3): 762-771.
[78] 田原. 小麦与玉米叶部表观建模与应用[D]. 合肥: 中国科技大学,
2011.
Tian Y. Appearance modeling of wheat and maize leaves[D]. Hefei:

3428 中 国 农 业 科 学 48 卷
University of Science and Technology of China, 2011. (in Chinese)
[79] 陆声链, 汪丽萍, 何火娇, 郭新宇. 基于相对叶绿素含量的黄瓜叶
色仿真. 农业机械学报, 2014, 45(3): 250-254.
Lu S L, Wang L P, He H J, Guo X Y. Visual simulation of cucumber
leaf color based on the relative content of chlorophyll. Transactions of
the Chinese Society for Agricultural Machinery, 2014, 45(3): 250-254.
(in Chinese)
[80] 赵春江, 苗腾, 郭新宇, 陆声链. 融合生理因子的植物叶片表观建
模. 计算机辅助设计与图形学学报, 2014, 26(4): 597-608.
Zhao C J, Miao T, Guo X Y, Lu S L. Plant appearance modeling of
plant leaf using physiological parameters. Journal of Computer-Aided
Design & Computer Graphics, 2014, 26(4): 597-608. (in Chinese)
[81] 苗腾. 植物叶片表观及其行为特性建模与可视化仿真技术研究[D].
北京: 中国农业大学, 2012.
Miao T. Research on techniques for modeling and visual simulating
characteristic behaviors of plant leaves[D]. Beijing: China Agricultural
University, 2012. (in Chinese)
[82] 郭焱 , 李保国 . 虚拟植物的研究进展 . 科学通报 , 2001, 46(4):
273-280.
Guo Y, Li B G. New advances in virtual plant research. Chinese
Science Bulletin, 2001, 46(4): 273-280. (in Chinese)
[83] 曹卫星, 朱 艳, 田永超, 姚霞, 刘小军. 数字农作技术研究的若干
进展与发展方向. 中国农业科学, 2006, 39(2): 281-288.
Cao W X, Zhu Y, Tian Y C, Yao X, Liu X J. Research progress and
prospect of digital farming techniques. Scientia Agricultura Sinica,
2006, 39(2): 281-288. (in Chinese)
(责任编辑 张晶,杨鑫浩)

中国农业科学 2015,48(17):3429-3438 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.010
收稿日期:2015-03-20;接受日期:2015-07-07 基金项目:农业部农业信息预警专项、中国农业科学院科技创新工程 联系方式:许世卫,Tel:010-82109902;E-mail:[email protected]
大数据推动农业现代化应用研究
许世卫,王东杰,李哲敏
(中国农业科学院农业信息研究所/中国农业科学院智能化农业预警技术与系统重点开放实验室/农业部农业信息服务技术重点实验室,北京 100081)
摘要:中国农业发展面临资源环境约束日益趋紧,农产品国内外市场“双重挤压”,农业稳产、农民增
收困难等严峻挑战,亟需转型升级、优化结构和提质增效。大数据作为一种与材料和能源一样重要的新型战
略资源,在生产要素耦合、农业系统协同等方面发挥着重要作用,正成为驱动农业现代化发展的重要力量。
目前,大数据技术在农业信息获取、分析处理和综合服务等方面取得了多方面的研究进展,并在精准生产决
策、食品安全保障、消费需求挖掘和市场贸易引导等方面开始应用。面对当前及未来一段时期内农业转型升
级的艰巨性和漫长性,应该建立大数据技术创新与组织创新的双重驱动机制。技术上,重点加强数据科学与
农业科学交叉融合,夯实现代农业基准数据基础,快速突破适农智能模型分析处理技术以及推动数据服务持
续创新;组织上,树立数据意识,加强数据立法,完善数据顶层治理结构,尤其要处理好数据安全与数据开
放共享等关系。
关键词:大数据;现代农业;转型升级;应用研究
Application Research on Big Data Promote
Agricultural Modernization XU Shi-wei, WANG Dong-jie, LI Zhe-min
(Agricultural Information Institute of Chinese Academy of Agricultural Sciences/Key Laboratory of Intelligent Agricultural Early
Warning Technology and System of Chinese Academy of Agricultural Sciences/Key Laboratory of Agro-Information Services
Technology, Ministry of Agriculture, Beijing 100081)
Abstract: Chinese agriculture is facing increasing challenges: the constraints of resources and the environment is increasingly tightening, the price difference between domestic and international markets is enlarging, and the growth in agriculture output and farmers' income is becoming hard to achieve. So, Chinese agriculture has a urgent need of transformation and upgrading, optimizing the structure, and improving the quality and efficiency. Big data, as a strategic resource such as material and energy, which plays an important role in the production factors coupling, collaboration, and other aspects of agricultural systems, is becoming an important driving power of agricultural modernization development. At present, big data technology has made progress in agricultural information acquisition, analysis, processing, services, and applied in the precise production decision, food security, mining consumer’s demand, guide trade and market. Facing the difficulties of agriculture transformation and upgrading, both of technology innovation and organization innovation need to be formed. On the one hand, developing a data cross fusion, laying solid foundation of modern agricultural benchmark data, breaking an agricultural intelligent model analysis processing system and development and promoting the continuous innovation in data service; On the other hand, setting up data consciousness, strengthening legislation, and improving the governance structure on top of the data, in particular, to deal with data security and data sharing.
Key words: big data; modern agriculture; transformation and upgrading; application
jia
文本框
返回目录

3430 中 国 农 业 科 学 48 卷
0 引言
在中国经济进入新常态,增速放缓背景下,农业
面临如何继续强化基础地位、促进农民持续增收的重
大课题;在国内生产成本快速攀升,大宗农产品价格
普遍高于国际市场的“双重挤压”下,农业面临如何
创新支持保护政策、提高农业竞争力的重大考验;在
资源短缺,污染加重“双重约束”下,农业面临如何
保障农产品有效供给和质量安全、提升农业可持续发
展能力的重大挑战;在城乡资源要素流动加速、城乡
互动联系增强、城镇化深入发展背景下,农业面临如
何加快新农村建设步伐、实现城乡共同繁荣的重大问
题。新常态、新背景下,这些系统性难题能否解决,
直接关系到农业的转型升级和持续发展。
纵观农业发展历史,农业每一次大的跨越都离不
开基础科学的飞跃,孟德尔、摩尔根的遗传学理论和
李比希的植物矿质营养学说的出现,推动了现代育种
技术和农业化学技术的发展,给世界农业发展带来了
第一次农业技术革命[1]。如今现代信息技术和生物技
术成为了推动农业发展的新型主导力量。信息化和农
业现代化成为“四化同步”战略的重要组成部分。但
是农业现代化是“四化同步”中的短板,需要信息化
这个新型杠杆的强力撬动。据相关研究测算,信息化
投入(信息化发展指数)增长 1.00%,GDP 增长 1.14%,
是资本投入贡献率的 1.60 倍,是劳动投入的 4.50 倍[2]。
从 1936 年图灵机的发明到 1945 年冯·诺依曼机的出
现,再到万维网的发明,人类仅仅用了几十年的时间
就进入了 IT 互联时代。而随着物联网、云计算、移动
互联,“互联网+”等的发展,海量数据爆炸式增长,
人类社会正在快步进入 DT 大数据时代。信息化已经
成为中国经济转型升级的重要力量,以大数据为代表
的信息生产力已经成为支撑国家经济发展的新型动
力。
大数据时代的到来并非偶然发生,而是信息技术
指数级发展,农业数字化不断推进的必然结果,信息
化的基础是数字化,数字化过程中产生了大数据。首
先,以互联网技术、传感感知技术、射频识别技术等
为代表的现代信息技术的快速发展,使得数据的产生
无处不在,无时不有,人类生产数据的能力空前加强;
其次,信息技术的增长超越了线性约束,摩尔定律发
挥了重要作用,18 个月左右计算性能提高一倍、存储
价格下降一半、带宽价格下降一半,使得人类保存数
据的能力显著增强;再次,随着联网用户和设备数量
的急速攀升,联入网络的价值显著增加(梅特卡夫定
律),进一步推动了信息技术的快速成长[3],数据挖
掘、机器学习等智能方法和算法的应用,使得数据的
处理分析显著增强。可以这么说,大数据的应运而生
得益于数据产生、存储和挖掘等一系列技术的升级。
根据国际数据公司(IDC)预测,到 2020 年,全球数
据量将达到 40 ZB。如果把物理客观世界看成一个维
度,那么数据就是物理世界在另一个纬度信息空间上
的映射和痕迹,物理世界的一切就都可以通过大数据
反映出来。海量的数据在运动中形成了数据流,帮助
我们更好的认识物理世界和我们自身。
究竟什么是大数据呢?从学术界和企业界的定义
来看,目前从数据特征出发对大数据进行定义的占据
主流,认为大数据是涵盖规模(volume)、类型
(variety)、价值(value)、速度(velocity)、精度
(veracity)和复杂度(complexity)等基本特征的数
据集及其相关的一系列技术体系[4-5]。从技术发展的角
度来看,大数据是一个动态、复合、发展的概念,是
一种未来发展的技术趋势,是一种科研范式的改变,
是一种与材料和能源一样重要的新型战略资源。
目前,互联网行业是大数据应用的领跑者,农业
正在与大数据加速融合拓展。大数据已经得到政府、
学术界和企业界普遍的重视,2015 年国务院相继发布
《国务院办公厅关于运用大数据加强对市场主体服务
和监管的若干意见》和《国务院关于积极推进“互联
网+”行动的指导意见》。大数据发展的环境得到持续
完善[6]。从技术发展的角度来看,大数据正在从热炒
概念向实际应用转化,其发展前景依然广阔。根据
Gartner 在 2014 年发布的《技术成熟度曲线特别报告》
(Hype Cycle Special Report),“数据科学”成为了
新面孔,大数据相关技术的演进在未来一段时间内仍
将展现出强大的生命力,相关市场的营收也将不断放
大(图 1)[7]。未来,数据资源将成为国家新型战略资
源,数据能力将成为现代农业发展的新型力量和推动
国家进步的新型竞争力。但同时值得注意的是,大数
据在异构性、规模性、时间性、复杂性、隐私性等方
面仍面临巨大挑战[8],真正的科学大数据如农业科学
大数据能在农业科学中发挥作用还有很长的路要走。
1 大数据对现代农业发展的作用
中国农业的根本问题是效率不高、效益不强、效
能不够,原因在于各生产要素缺乏耦合效应,产业链
衔接不紧,农业大系统循环性、协同性不够。这导致

17 期 许世卫等:大数据推动农业现代化应用研究 3431
图 1 2014 技术成熟度曲线[7]
Fig. 1 The hype cycle in 2014[7]
了农业发展较为粗放,而这种粗放是与长期以来农业
基准数据资源薄弱、数据结构不合理、数据细节程度
不够、数据标准化、规范化水平差等原因紧密相连。
随着物联网、云计算、移动互联等技术的突破,更多
的数据得到收集,数据流动性得到了最大程度释放,
数据分析和服务能力得到显著增强,大数据逐渐成为
了农业生产的定位仪、农业市场的导航灯和农业管理
的指挥棒。 1.1 大数据是现代农业生产经营的“定位仪”
中国要强,农业必须强。2014 年,中国粮食生产
实现创纪录的“十一连增”,总产达到 6 071 亿公斤。
但这个“十一连增”代价很大,效率很低。中国农业
从业人员 2.7 亿,但劳动生产率仅为世界的 64%;有
效灌溉面积 0.63 亿公顷,但农田灌溉水有效利用系数
仅为 0.52,远低于发达国家 0.7—0.8 的水平;喷洒农
药 180 万吨,但利用率仅为 35%,比发达国家低 10个百分点;化肥施用折纯量 5 900 万吨,但综合利用
率大概在 30%左右;农机总动力 10.7 亿千瓦,农作物
耕种收综合机械化水平达到 61%,农业科技进步贡献
率 55.6%[9] ,但仍低于大多数发达国家(75%以上)。
显然,这种拼资源、拼消耗的粗放发展方式在资源约
束日益趋紧的情况下难以为继。 今后要做强农业,依靠什么?靠土地?1.2 亿公顷
耕地的红线已经岌岌可危。靠劳动力?十多年来农村
劳动力减少了 8 000 多万人,谁来种地成为新的问题。
靠政策?最低收购价和临时收储政策在国内外市场价
格倒挂的背景下,同样面临新的挑战。靠科技?单项
科技面临边际效益递减的趋势。当前中国农业生产就
像一艘巨型航母,外在装备都已具备,但是缺乏精准
“定位和导航”,无法精准生产。中国已近 30 年未组
织全国性土壤肥力普查和肥料效益研究,对水资源的
调查评估也较为欠缺。想要立足国土资源整体,布局
优势生产区,但“家底”不清,基准数据缺乏。 大数据的兴起,恰恰为改变这种困境找到了出路。
农业大数据可以挖掘农业资源间的发展潜力、搭配关
系和最佳使用途径,精确计算最优化配置模式,帮助
农业实现生产需求变化与资源变化的深度耦合,做到
农业“全要素、全过程、全系统”生产的一体化。依
靠数据驱动,才能使传统农业从主要追求产量和依赖
资源消耗的粗放经营转到数量质量效益并重、注重提
高竞争力、注重农业科技创新、注重可持续的集约发
展上来,才能走产出高效、产品安全、资源节约、环
境友好的现代农业发展道路。 1.2 大数据是现代农业市场消费的“导航灯”
中国要富,农民必须富。2014 年农民增收实现
“十一连快”,全年农民人均纯收入 9 892 元,同比

3432 中 国 农 业 科 学 48 卷
增长 9.2%。但是与城镇居民相比,仍存在很大差距。
从整体情况来看,中国一家一户的传统经营模式仍
占主导地位,大市场小生产背景下,农产品“滞销
卖难”仍然频繁出现,增产不增收,已渐成当前国
内农业生产的头号杀手和影响农民增收的重要障
碍。究其原因是信息滞后,信息利用程度不高,农
业生产难以与市场对接,难以与消费匹配。一方面,
农民缺乏需求信息,市场需要什么不知道,只能凭
经验种植,上年什么赚钱种什么,结果扎堆种植,
生产过剩,造成“滞销卖难”;另一方面,即使有
信息,但是因为信息更新慢、推送不畅、针对性差,
农民面对这些信息往往收不到、看不懂、用不上。
据不完全统计,中国有 3 000 多个涉农网站,国家
发展改革委员会、农业部、商务部等纷纷建立了价
格监测系统,但是实际利用效果仍待提高。 大数据技术的兴起,在实时捕捉消费需求、跟踪
市场变化、个性化推送等方面,为现代农业的发展开
启了“导航灯”。近几年,电子商务、微商营销等大
数据应用从星星之火发展到燎原之势。2014 年阿里平
台上经营农产品的卖家数量达到 76.21 万个,农产品
销售 483.02 亿元,同比增长 69.83%[10]。农产品电子
商务与大数据技术的融合正在成为农民增收的新业
态,促使农业生产从“生产导向”向“消费导向”转
变,帮助农民念好“山海经”、唱好“林草戏”、打
好“果蔬牌”。 1.3 大数据是现代农业管理决策的“指挥棒”
当前,国内外农业环境发生了重大变化,中国与
世界的联系日益一体化,中国已经成为农产品净进口
国,但是中国农业市场信息数据资源研究与建设仍然
滞后,大量“三农”信息的缺失、滞后、封闭严重制
约了全球视角下开展农业管理决策的科学性、系统性、
高效性和精准性。以农业部为例,截至 2014 年 5 月底,
共有12个司局和9家部属事业单位直接开展农业信息
监测统计工作,已经建立了 21 套统计报表制度,针对
75 项监测统计科目,每年开展 300 张报表,5 万个(次)
的指标调查,加上行业管理监测数据、农业地理空间
数据,已经形成了复合型数据获取组织模式。但是随
着社会主义市场经济体制改革的深入,也出现了诸如
消费数据缺失,信息交叉重复、匹配性差,部门内外
信息难以充分共享等问题,导致农产品价格大幅上涨
或质量安全出现问题时,国际环境发生重大变化时,
不能及时发现源头,不能迅速制定应急方案,以致于
延误了解决事件的最佳时机。 大数据时代要求数据做到公开共享,强调数据的
挖掘和利用,数据只有在使用中才能产生巨大的价值。
2012 年美国启动了“大数据研究与发展计划”,2013年启动了“从数据到知识到行动:建立新的伙伴关系”
的大数据计划;日本和英国卡梅伦政府也相继启动了
旨在促进大数据研究和应用的计划。强调数据驱动决
策,用大数据支撑话语权,用大数据支撑决策权,真
正把数据作为管理决策的指挥棒。中国要想在国际市
场中掌握主动权,就必须抓紧建立数据采集、利用、
共享的体系。
2 大数据核心技术在农业领域的研
究进展
从各大 IT 公司的大数据处理流程来看,基本上可
以分成数据获取、数据存储、数据分析处理和数据服
务应用等几大环节。农业作为信息技术的应用部门,
其生产、流通、消费、市场贸易等过程,分别融入在
大数据的流程之中,根据大数据的获取、分析处理和
服务应用等方面开展了大量集成创新,取得了重要研
究进展(图 2)。
图 2 大数据分析流程图
Fig. 2 The flow chart of data analysis

17 期 许世卫等:大数据推动农业现代化应用研究 3433
2.1 大数据获取技术
根据农业大数据来源的领域分类,大致可以分为
农业生产数据、农业资源与环境数据、农业市场数据
和农业管理数据[11]。针对不同领域的农业大数据,大
数据获取技术主要包括感知技术[12](传感器、遥感技
术等)、识别技术(RIFD、光谱扫描、检测技术)、
移动采集技术[13](智能终端、APP)等。第一,感知
技术主要是从不同尺度感知动植物生命与环境信息。
在地域范围,重点考虑对地观测的资源宏观布局,需
要遥感、便携式 GPS 面积测绘仪、农业飞行器等;在
区域范围,重点考虑动植物生长信息的时空变异性,
需要基于 WebGIS 的动植物生长信息的动态检测平台
等;在视域范围,重点考虑动植物生态环境的复杂性,
需要动植物营养、病害及周围环境污染信息的采集测
试传感器;在个域范围,重点考虑动植物信息探测中
环境因素干扰,需要动植物营养病害快速无损测试仪、
活体无损测量仪等。第二,识别技术主要是针对农产
品质量安全开展监测。包括食品安全溯源的 RFID 技
术[14-15],主要保证农产品原料、加工、销售全环节的
追踪可溯。农产品质量安全快速无损检测技术[16],主
要是应用红外光谱、X 射线、计算机视觉等无损检测
技术在农产品品质分析、产地环境监测、农业投入品
评价和商品流通监控等环节应用。第三,智能移动采
集技术主要针对农产品市场、营销、管理信息的采集。
如农信采[17]采集农产品价格信息,农业管理信息系
统[18]的应用等。 传统的大数据获取技术在材料选择、结构设计、
性能指标上相对单一,如种植业中的传感技术只能测
量气温、湿度、CO2 等信息,而随着物联网技术的发
展,其传感器材料已经从液态向半固态、固态方向发
展,结构更加小型化、集成化、模块化、智能化;性
能也向检测量程宽、检测精度高、抗干扰能力强、性
能稳定、寿命长久方向发展[19],目前研发的一些传感
器已经可以用来监测植物中的冠层营养状态、茎流、
虫情等。未来中国的大数据获取技术改进的重点将是
在信息技术与农业的作物机理、动物的行动状态和市
场的实时变化紧密结合,将在提升信息获取的广度、
深度、速度和精度上突破。 2.2 大数据分析处理技术
在大数据环境下,由于数据量的膨胀,数据深度
分析以及数据可视化、实时化需求的增加,其分析处
理方法与传统的小样本统计分析有着本质的不同。大
数据处理更加注重从海量数据中寻找相关关系和进行
预测分析。例如,谷歌做的流行病的预测分析[20],亚
马逊的推荐系统[21],沃尔玛的搭配销售[22],都是采用
相关分析的结果。数据分析技术在经历了商务智能、
统计分析和算法模型之后,目前进入到了大平台处
理的阶段,主要是基于 MapReduce、Hadoop 等分析
平台[23],同时结合 R、SAS 等统计软件,进行并行计
算。近两年来,内存计算逐渐成为高实时性大数据处
理的重要技术手段和发展方向。它是一种在体系结构
层面的解决办法,它可以和不同的计算模式相结合,
从基本的数据查询分析计算到批处理和流式计算,再
到迭代计算和图计算,目前比较典型的有 SAP 的
HANA,微软 Trinity,UC Berkeley AMPLab 的 Spark等。
在农业领域,数据处理正从传统的数据挖掘、机
器学习[24]、统计分析向着动植物数字化模拟与过程建
模分析、智能分析预警模型系统等演进(如图 3)。
在生物学领域,大数据的分析作用已经凸显,基因测
序,数字育种[25]已经采用了大数据算法和模型;作物
模型方面,国际上获得广泛认可的通用作物生长模型
有荷兰 WOSOFT 系列,美国 DSSAT 系列,澳大利亚
APSIM 系列,FAO 的 AQUACROP 等。在植物数字
化模拟方面,国际上已经有了 OpenAlea、GroIMP、VTP 等用于植物建模和分析的开源项目。农产品市场
监测预警模型系统方面,具有代表性的是经合组织和
联合国粮农组织(OECD-FAO)的 AGLINK-COSIMO模型、 FAO 全球粮食和农业信息及预警系统
(GIEWS)、美国农业部(USDA)的国家-商品联系
模型与美国粮食和农业政策研究所(FAPRI)的 FAPRI模型和中国农业科学院农业信息研究所的 CAMES 模
型等。 总体来看,由于农业生产过程发散,生产主体复
杂,需求千变万化,与互联网大数据相比,针对农业
的异质、异构、海量、分布式大数据处理分析技术依
然缺乏,今后农业大数据的分析处理应该将信息分析
处理技术与农业生理机理关键期结合、市场变化过程
紧密结合。 2.3 大数据服务应用技术
目前大数据服务技术已在互联网广告精准投放、
商品消费推荐、用户情感分析、舆情监测等广泛应用。
在农业上,随着农业部“信息进村入户”工程、“物
联网区试工程”、12316 热线、国家农业云服务平台
等的建设和推动,中国的农业信息服务体系逐步得到
完善,“三农”对信息的需求也更加迫切。

3434 中 国 农 业 科 学 48 卷
原始数据:资源环境、生产、流通、消费、管理数据Raw Data: Resources and Environment, Production, Circulation, Consumption, Management
数据导入:提取-转换-加载Data Import: Extract-Transform-Load
批处理Batch
交互分析Transactional analysis
流处理Stream processing
数据存储 (Data Storage): SQL和No SQL
数据挖掘 (Data Mining) (Data Warehouse, BI, OLAP, BPM) 作物生长机理Crop growth mechanism
动物生长轨迹Animal growth
trajectory农产品市场规律
Agricultural products market
rule
动植物数字化模拟、过程建模分析、智能分析预警模型系统Digital simulation of animals and plants, Process modeling analysis, Intelligent analysis of early warning system model
可视化分析与呈现Visual analysis and rendering
内存分析In-memory analytics
图 3 农业大数据分析流程图
Fig. 3 Agricultural big data analysis process
国际上有关农业信息服务技术的研究主要集中在
农业专家决策系统、农村综合服务平台和农业移动服
务信息终端、农业信息资源与增值服务技术以及信息
可视化等方面[26]。国内近些年,先后开展了智能决策
系统、信息推送服务、移动终端等。在大数据时代,
针对农业产前、产中、产后各环节的关联,开发大数
据关联的农业智能决策模型技术;针对大众普遍关注
食品安全的状况,开发大数据透明追溯技术;针对农
民看不懂、用不上等问题,结合移动通信技术、多媒
体技术,开发兼具语音交互、信息呈现、多通道交互
的大数据可视化技术。
3 大数据在农业现代化发展中的应
用成效
大数据的应用,一方面可以全息立体反映客观事
物,洞悉全样本数据特征,促进事物之间的深度耦合,
提升效能;另一方面是通过数据间的关联特征,预测
事物未来发展趋势,增强预见性。目前,从农业生产、
经营、消费、市场、贸易等不同环节来看,大数据在
精准生产决策、食品安全监管、精准消费营销、市场
贸易引导等方面已经有了较为广泛的应用。
3.1 发挥要素耦合效应,提升精准生产决策
大数据的作用不仅仅在于更好发现自身价值,还
在于帮助其他要素更好认识自身,发挥要素间耦合作
用,提升他物价值,促进“价值双增”。国内外在改
变农业粗放生产上,围绕气象预报、水肥管理、作物
育种、病虫害预报、高效养殖等方面已经开展了大量
的应用。美国天气意外保险公司(The Climate Corporation)利用 250 万个采集点获取的天气数据,
结合大量天气模拟、海量植物根部构造和土质分析等
信息对意外天气风险做出综合判断。美国硅谷土壤抽
样分析服务商 Solum 利用软、硬件系统实现精准土壤
抽样分析,给土壤“号脉开方”[27]。在中国农业部印
发《2014 年种植业工作要点》中,要求各地推进配方
肥到田和施肥方式转变,利用“大数据”与相关化肥
生产企业合作推广配方肥。泰国、越南、印度尼西亚
等国基于遥感信息与作物保险的监测计划(Remote sensing-based Information and Insurance for Crops in Emerging economies,RIICE)在水稻上得到广泛应用,
通过采用欧洲航天局卫星实时获取水稻的生长数据,
进行生长跟踪、产量预测[28]。美国农业部研究所开始
在部分农场采用高光谱航空遥感影像和地面观测数据
结合的方式进行面状病虫害监测,利用全球的病虫害

17 期 许世卫等:大数据推动农业现代化应用研究 3435
数据发现害虫的传播规律[29]。国际种业巨头如美国杜
邦先锋、孟山都、圣尼斯公司及瑞士先正达等纷纷采
用现代信息技术开展智能育种,加快“经验育种”向
“精确育种”的转变[30]。在英国,大多数的养牛、养
猪和养鱼场都实现了从饲料配制、分发、饲喂到粪便
清理、圈舍等不同程度智能化、自动化管理。 3.2 跟踪流通全程,保障食品安全质量
受制于传统农产品流通渠道复杂,层级繁多,监
管不透明,公众缺乏知情权和监督权,中国食品安全
事件频发,给消费者造成了重大伤害。大数据技术的
发展使得全面、多维感知农产品流通成为可能。目前,
技术层面上,在产地环境、产品生产、收购、储存、
运输、销售、消费全产业链条上,物联网、RFID 技术
得到广泛应用,一批监测新技术如“食品安全云”和
“食安测”等应用软件陆续开发[31];制度层面上,中
国利用大数据开展食品安全监管的力度不断加强,
2015 年 6 月国务院新出台了《关于运用大数据加强对
市场主体服务和监管的若干意见》,明确提出建立产
品信息溯源制度,对食品、农产品等关系人民群众生
命财产安全的重要产品加强监督管理,利用物联网、
射频识别等信息技术,建立产品质量追溯体系,形成
来源可查、去向可追、责任可究的信息链条,方便监
管部门监管和社会公众查询;商业层面上,阿里巴巴,
京东商场等电商企业利用大数据保障食品溯源。如辽
宁省大洼县盛产稻田米和稻田蟹,2015 年加入阿里农
业满天星计划,开始农产品溯源探索,针对不同类型
农产品的成长特点,通过二维码来承载产品名、产品
特征、产地、种植人、生长周期、生长期施肥量、农
药用量、采摘上市日期等不同的溯源信息。 3.3 挖掘用户需求,促进产销精准匹配
传统的农业发展思维更多关注生产,在乎的是够
不够吃的问题,而在消费结构升级的情况下,应该转
向怎么才能吃得健康,吃得营养。大数据在这方面正
在驱动商业模式产生新的创新。利用大数据分析,结
合预售和直销等模式创新,国内电商企业促进了生产
与消费的衔接和匹配,为农产品营销带来了新的机遇。
截止 2014 年 12 月,全国涌现了 19 个淘宝镇,211 个
淘宝村,以淘宝村为代表的农村电子商务正在深刻改
变中国农村的面貌[32],变革着中国传统农产品营销的
模式。连锁型的社区生鲜超市 M6 于 2005 年前开始了
数据化管理,物品一经收银员扫描,总部的服务器马
上就能知道哪个门店,哪些消费者买了什么。2012 年,
M6 的服务器开始从互联网上采集天气数据,然后,
从中国农历正月初一开始推算,分析不同节气和温度
下,顾客的生鲜购买习惯会发生哪些变化,进而实现
精准订货、存储和精准配货[33]。未来还可以将食品数
据,与人体的健康数据、营养数据连接起来,这样可
以根据人体的健康状况选择适当的食物。 3.4 捕捉市场变化信号,引导市场贸易预期
市场经济中最重要的是信息,利用信息引导市场
和贸易有助于控制国际市场话语权和掌握世界贸易主
导权。以美国为例,其收集信息、利用信息的做法值
得借鉴。19 世纪 60 年代,为了弥补农村市场中出现
的信息不对称,美国农业部在全国雇用了几万名监测
员,形成了一个农情监测网络,每月定期发布各种农
产品的交易情况和价格波动,同时通过免费邮寄、张
贴海报的方式把信息送到各大农场。时至今日,美国
已经形成了一套庞大、规范的农业信息发布体系,其
定期发布的年度《农业中长期展望报告》、月度《世界
农产品供需预测报告》和周度《农作物生长报告》[34],
成为引导全球农产品市场变化的风向标。美国的农民
仅占全国人口的 2%,但 2%的农民不仅养活了 3 亿多
美国人,而且其农产品出口还位居全球第一。目前美国
政府仍在致力于数据的开放和共享,创建了 data.gov网站,链接到 348 个农业数据集,提供政府采集的原
始数据,供私人领域开发者分析数据、提出决策。 与发达国家相比,中国的信息发布和数据利用仍
有很大前进空间。2003 年起,农业部推出《农业部经
济信息发布日历》制度,主要发布生产及市场经济信
息。2014 年中国召开了中国农业展望大会,发布了《中
国农业展望报告(2014—2023)》,结束了中国没有
展望会议的历史,开启了提前发布市场信号、有效引
导市场、主动应对国际变化的新篇章。在中国成为世
界农产品进口大国的背景下,如何有效利用信息,把
握市场和贸易话语权和定价权是必须修炼的功课。
4 主要结论和建议
4.1 主要结论
大数据已经成为一种新兴的战略资源。大数据是
一种以数据驱动农业现代化发展的新兴战略资源,通
过与其他实体要素的耦合,能够进一步提高农业生产
力。随着信息技术进步的加速,未来数据将同物质、
能量一样,成为现代农业转型升级的重要动力。从目
前的发展与应用来看,大数据仍是一个非常年轻而富
有前景的研究领域,在资源效率提升、生产布局优化、
产业安全监管、市场有效引导等方面已经展现了强大

3436 中 国 农 业 科 学 48 卷
的能力。没有信息化就没有农业现代化,所以,要重
视大数据时代的到来,加强理论与方法的深入研究,
发挥大数据的驱动作用,助力信息化成为现代农业发
展的制高点,帮助现代农业实现弯道超车。 与此同时,我们也应该看到数据的爆炸,在创造
重要机遇的同时也带来了巨大的挑战。大数据科学对
数据模型、服务软件和系统能力建设等方面提出了挑
战。开展适农大数据技术研究迫在眉睫。鉴于涉农数
据的大量涌现,中国亟需开展以下技术研究:第一,
多元数据标准融合技术。针对耕地、育种、播种、施
肥、植保、收获、储运、农产品加工、销售、畜牧业
生产等环节,针对数字、文字、视频、音频等不同格
式、不同业务载体的海量数据,建立基准数据工程,
整合成标准统一的数据源;第二,海量数据组织管理
技术。完成海量数据的存储、索引、检索和组织管理,
突破了农业异质数据转换、集成与调度技术,实现海
量数据的快速查询和随时调用;第三,加强适农大数
据分析挖据技术。围绕病虫害综合防治、粮食产量预
测等重点领域,开展并行高效农业数据挖掘算法,建
立智能机理预测分析模型;围绕农产品品种、气象、
环境、生产履历、产量、空间地理、遥感影像等数据
资源建立农业协同推理和智能决策模型;围绕农产品
市场信息开展多品种市场关联预测技术和农产品市场
预警多维模拟技术。 4.2 相关建议
农业从转型升级到迈向现代化,必然是一个漫长
和艰难的过程,要推动其发展必须依靠技术创新和组
织创新的双轮驱动。一方面,以大数据为核心,围绕
数据学科与农业学科体系建设、基础理论研究、基准
数据构建、智能模型研发、系统平台搭建,形成以大
数据为核心的适合现代农业发展的技术系统体系;另
一方面,要加强数据意识,完善数据立法,在保证信
息安全的前提下实现数据开放共享。 4.2.1 加强数据科学与农业科学的融合,完善数据科
学理论方法体系 国内外实践表明,农业信息学科的
新概念、新理论、新方法的创新,是引领农业信息技
术重大变革,促使农业生产发生巨大飞跃的重要引擎。
数据密集型科学成为科研的第四范式,将加速信息技
术科学与现代农业相关学科的融合发展。但数据要形
成一门科学还需要更加注重大数据的基础理论研究、
科学方法创新,更加注重大数据学科体系的建设,目
前国内外一些大学和研究机构已经开展了大数据专
业,并且开始招生。今后应该在大数据生命周期、演
化与传播规律,数据科学与农业相关学科之间的互动
融合机制,以及大数据计算模型、作物模型与模拟、
智能控制理论与技术、农业监测预警技术,大数据可
视化呈现与精准化推送等方面加强研究,形成系统性、
全面性、深入化的理论支撑。 4.2.2 构建农业基准数据,夯实现代农业发展基础支
撑 现代农业基准数据,是指现代农业建设过程中涉
及的生产、经营、管理等各种活动所依赖的标准化、
基础性、系统性数据。当前,中国农业数据尚存在农
业基准数据资源薄弱、数据结构不合理、数据细节程
度不够、数据标准化、规范化水平差等问题,针对这
些问题,建议结合农业部大田长期监测工作,建立现
代农业自然资源基准数据、现代农业生产基准数据、
现代农业市场基准数据、现代农业管理基准数据。以
基准数据建设为契机,制定和完善数据采集、传输、
存储和汇交标准,对数据采集的内容、方式、时间、
地点,数据传输的速率、方式、冗余和编码标准,数
据存储格式、存储方式、存储安全、数据结构汇交方
法以及数据汇交内容、汇交分类、汇交范围等制定标
准和规范,只有这样,才能为现代农业发展决策提供
坚实的基础支撑。
4.2.3 加强智能模型系统研发,推动农业智能转型
数据的处理和分析能力是大数据技术的核心。针对农
业领域数据海量、分散、异构等现象而难以集成,不
能挖掘其巨大潜在价值现状。重点开展农业大数据智
能学习与分析模型系统关键技术研究,利用人工智能、
数据挖掘、机器学习、数学建模、深度学习等技术,
在充分理解农业领域知识的基础上,针对所要解决的
实际问题,建立有效的数学模型对数据进行处理,并
根据处理的结果对模型进行修正,以完成自主学习校
正模型的过程,并利用最终形成的模型对海量数据进
行处理,以进行预测,发现其中蕴含的模式和规律,
辅助农业决策,实现决策的智能化、精确化和科学化。 4.2.4 倡导数据开放,服务引领农业发展 数据的应
用是大数据的最终目的,数据开放是社会管理创新的
一种有效手段和助推器。目前,数据公开、软件开源
与数据共享已经成为了全球发展的重要潮流[35]。数据
的开放共享有助于新常态下中国农业的健康发展。以
农产品为例,中国已经成为世界上最重要的农产品进
口国,但是在很多品种上缺乏定价权和话语权,一个
重要原因就是主要的贸易国对中国主要农产品的产
量、面积,甚至消费、库存等情况甚至比我们还清楚,
他们通过这些基本信息,了解到供求情况,摸清了我

17 期 许世卫等:大数据推动农业现代化应用研究 3437
们的底线,所以在国际贸易中我们总是处于被动地位。
未来农业的发展,信息对市场的服务和引领是一门必
修课。一是要加强数据立法,为农业信息公开提供法
律保障;二是形成数据开发的体制和机制,保证在数
据会商、开放标准、发布规范等方面的切实可行;三
是以召开中国农业展望大会和发布中国农业展望报告
为契机,形成具有中国特色的农产品监测预警和信息
发布制度,最终通过数据的开放,为生产决策、市场
监测、农业管理提供信息支撑,引领现代农业发展。
References
[1] 张润志, 王大生. 农业新技术革命与中国农业发展. 中国科学院院
刊, 1998, 2: 113-117.
Zhang R Z, Wang D S. New technology revolution of agriculture and
China’s agricultural development. Bulletin of Chinese Academy of
Sciences, 1998, 2: 113-117. (in Chinese)
[2] 杨培芳. 信息生产力发展前景及其重大影响. 边缘革命 2.0, 2013.
12
Yang P F. Information productivity and its significant impact on the
development prospects. Edge Revolution 2.0, 2013.12. (in Chinese)
[3] 阿里研究院. 信息经济呈现十大浪潮. http://www.aliresearch.com/
blog/article/detail/id/20046.html
Ali Research. Information economy presents ten wave. http://www.
aliresearch.com/blog/article/detail/id/20046.html (in Chinese)
[4] Manyika J, Chui M, Brown B, Bughin J, Dobbs R, Roxburgh C, Hung
Byers A. Big data: The next frontier for innovation, competition, and
productivity. http://www. mckinsey. com/Insights/MGI/Research/
Technology_and_Innovation/Big_data_The_next_frontier_ for_innovation.
[5] Hey T. The Fourth Paradigm–Data-Intensive Scientific Discovery.
E-Science and Information Management. Berlin: Springer Berlin
Heidelberg, 2012: 1-1.
[6] GigaOM. Putting Big Data to Work: Opportunities for Enterprises.
2011.3 http://en.wikipedia.org/wiki/Big_data
[7] http://www.jifang360.com/news/2014822/n693761045.html
[8] 工信部电信研究院大数据白皮书 (2014 年 ). http://www.cctime.
com/html/2014-5-12/20145121139179652.htm
Big Data White Paper of Telecommunications Research Institute of
MIIT. http://www.cctime.com/html/2014-5-12/20145121139179652.
htm (in Chinese)
[9] 李家洋 . 2014 年我国农业科技进步贡献率达 56%. 新华网 .
http://big5.qstheory.cn/gate/big5/news.xinhuanet.com/legal/2015-01/2
8/c_127428873.htm
Li J Y. China's agricultural science and technology contribution rate of
56% in 2014. Xinhuanet. http://big5.qstheory.cn/gate/big5/ news.
xinhuanet.com/legal/2015-01/28/c_127428873.htm (in Chinese)
[10] 阿里农产品电子商务白皮书(2014). http://www.aliresearch.com/
blog/article/detail/id/20459.html
Ali agricultural e-commerce White Paper (2014). http://www.
aliresearch. com/blog/article/detail/id/20459.html (in Chinese)
[11] 许世卫. 农业大数据与农产品监测预警. 中国农业科技导报, 2014,
16(5): 14-20.
Xu S W. Agricultural big data and monitoring and early warning of
agricultural products. Journal of Agricultural Science and Technology,
2014, 16(5): 14-20. (in Chinese)
[12] 姚元森, 廖桂平, 赵星, 陈艳, 赵丽敏, 雷梦龙. 农作物生长环境
信息感知技术研究进展. 作物研究, 2013, 27(1): 58-63.
Yao Y S, Liao G P, Zhao X, Chen Y, Zhao L M, Lei M L. Reseach
progress of crop growth environment information perception
techniques. Crop Reseach, 2013, 27(1): 58-63. (in Chinese)
[13] 罗锡文, 臧英, 周志艳. 精细农业中农情信息采集技术的研究进
展. 农业工程学报, 2006, 22(1): 167-173.
Luo X W, Zang Y, Zhou Z Y. Reseach progress in farming information
acquisition technique for precision agriculture. Transactions of the
Chinese Society of Agricultural Engineering, 2006, 22(1): 167-173.
(in Chinese)
[14] 杨天和, 褚保金. “从农田到餐桌”食品安全全程控制技术体系研究.
食品科学, 2005, 26(3): 264-268.
Yang T H, Zhu B J. Study on comtrol system of food safety farm to
table. Food Science, 2005, 26(3): 264-268. (in Chinese)
[15] 侯春生, 夏宁. RFID 技术在中国农产品质量安全溯源体系中的应
用研究. 中国农学通报, 2010, 26(3): 296-298.
Hou C S, Xia N. RFID technology in the application of China's
agricultural product quality safety traceability system research.
Chinese Agricultural Science Bulletin, 2010, 26(3): 296-298. (in
Chinese)
[16] 潘立刚, 张缙, 陆安祥, 马智宏, 韩平. 农产品质量无损检测技术
研究进展与应用. 农业工程学报, 2008, 24(S2): 325-330.
Pan L G, Zhang J, Lu A X, Ma Z H, Han P. Review on non-destructive
determination technology for agricultural product quality.
Transactions of the Chinese Society of Agricultural Engineering, 2008,
24(S2): 325-330. (in Chinese)
[17] Xu S W, Kong F T, Li Z M, Li Z Q, Zhang Y E, Lv Y, Tian Z W, Wang
D J, Yu H P. Design and implementation of portable agricultural
product holographic market information collection terminal. Advanced
Materials Research, 2013, 2162(628): 359-366.
[18] 刘忠. 中国农业管理信息系统发展现状、问题、趋势与对策. 农业

3438 中 国 农 业 科 学 48 卷
工程学报, 2005, 21(S1): 201-206.
Liu Z. Status, questions, trend and strategy of management information
system for agriculture in China. Transactions of the Chinese Society of
Agricultural Engineering, 2005, 21(S1): 201-206. (in Chinese)
[19] 中国传感器产业发展白皮书. 2014. http://www.iotcn.org.cn/uploadfile/
2014/1024/20141024034220988.pdf
China's white paper of sensor industry development. http://www.
iotcn.org.cn/uploadfile/2014/1024/20141024034220988.pdf (in Chinese)
[20] Lazer D, Kennedy R, King G, Vespignani A. The parable of Google
Flu: Traps in big data analysis. Science, 2014, 343 (6176): 1203-1205.
[21] Linden G, Smith B, York J. Amazon.com recommendations:
Item-to-item collaborative filtering. Internet Computing, Institute of
Electrical and Electronics Engineers, 2003, 7(1): 76-80.
[22] Waller M A, Fawcett S E. Data science, predictive analytics, and big
data: a revolution that will transform supply chain design and
management. Journal of Business Logistics, 2013, 34(2): 77-84.
[23] Chen H, Chiang R H L, Storey V C. Business Intelligence and
Analytics: From Big Data to Big Impact. MIS quarterly, 2012, 36(4):
1165-1188.
[24] Ma C, Zhang H H, Wang X. Machine learning for Big Data analytics
in plants. Trends in plant science, 2014, 19(12): 798-808.
[25] Marx V. Biology: The big challenges of big data. Nature, 2013,
498(7453): 255-260.
[26] 陈威, 郭书普. 中国农业信息化技术发展现状及存在的问题. 农业
工程学报, 2013, 29(22): 196-205.
Chen W, Guo S P. Current situation and existing problems of
agricultural informatization in China. Transactions of the Chinese
Society of Agricultural Engineering, 2013, 29(22): 196-205. (in Chinese)
[27] 大数据吹响智慧农业的号角. http://www.china-cloud.com/yunzixun/
yunjisuanxinwen/20130807_21011.html
Blowing the wisdom agriculture horns of big data. http://www.china-
cloud.com/yunzixun/yunjisuanxinwen/20130807_21011.html (in Chinese)
[28] Sonka S, IFAMR I. Big Data and the Ag sector: More than lots of
numbers. International Food and Agribusiness Management Review,
2014, 17(1): 1.
[29] Garrett K A. Agricultural impacts: Big data insights into pest spread.
Nature Climate Change, 2013, 3(11): 955-957.
[30] 康乐, 王海洋. 我国生物技术育种现状与发展趋势. 中国农业科技
导报, 2014, 16(1): 16-23.
Kang L, Wang H Y. Status and prospect of agrobiotechnology- based
breeding in China. Journal of Agricultural Science and Technology,
2014, 16(1): 16-23. (in Chinese)
[31] 形成全国首朵“食品安全云”贵州利用大数据保群众舌尖上的安全.
http://gzrb.gog.com.cn/system/2014/12/18/013983006.shtml
As the first cloud of “food safety”, Guizhou use big data to protect the
food safety. http://gzrb.gog.com.cn/system/2014/12/18/013983006.
shtml (in Chinese)
[32] 全国淘宝村增至 211 个 , 阿里推出淘宝村三大扶持措施 .
http://www.aliresearch.com/blog/article/detail/id/20048.html
The taobao village increased to 211 in the country, Ali launched
taobao village three supportive measures. http://www.aliresearch.
com/blog/article/detail/id/20048.html (in Chinese)
[33] 生鲜超市 M6 的大数据实验 . http://tech.ccidnet.com/art/40911/
20130513/4927167_1.html. 2013-05-13
The big data experiment of fresh supermarket M6. http://tech.ccidnet.
com/art/40911/20130513/4927167_1.html. 2013-05-13 (in Chinese)
[34] 许世卫. 美国农产品信息分析预警工作考察报告. 农产品市场周
刊, 2012 (21): 58-60.
Xu S W. US agricultural information analysis and early warning
investigation report. Agricultural Market Weekly, 2012(21): 58-60. (in
Chinese)
[35] 诸云强, 孙九林, 王卷乐, 杨雅萍. 论地球数据科学与共享. 国土
资源信息化, 2015, (1): 3-9.
Zhu Y Q, Sun J L, Wang J L, Yang Y P. Study on Earth data science
and data sharing. Land and Resources Informatization, 2015, (1): 3-9.
(in Chinese)
(责任编辑 张晶,杨鑫浩)

中国农业科学 2015,48(17):3439-3453 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.011
收稿日期:2015-03-16;接受日期:2015-06-08 基金项目:国家“973”计划(2013CB127700)、国家现代农业产业技术体系建设专项资金(CARS-3-1-11)、高等学校学科创新引智计划(B07049) 联系方式:康振生,E-mail:[email protected]
小麦条锈菌致病性及其变异研究进展
康振生,王晓杰,赵 杰,汤春蕾,黄丽丽
(西北农林科技大学植物保护学院/旱区作物逆境生物学国家重点实验室, 陕西杨凌 712100)
摘要:小麦条锈病(wheat stripe rust or yellow rust)是一种气传性的低温真菌病害,在所有小麦产区
均可暴发流行,造成巨大的产量和经济损失,严重威胁着小麦的安全生产。其病原条形柄锈菌小麦专化型(Puccinia
striiformis West. f. sp. tritici Eriks. & Henn.,Pst)为严格专性寄生真菌,致病性变异频繁,常造成小麦
品种抗条锈性“丧失”,引发病害大流行。因而,从 20 世纪 50 年代起,条锈菌致病性变异的研究一直备受关注,
并已开展了大量的研究工作。本文从小麦品种抗病性丧失、条锈菌致病性变异途径、条锈菌群体遗传、条锈菌基
因组和功能基因组以及条锈菌效应蛋白等不同方面概述了近年来取得的重要研究进展,提出了未来条锈病防治上
面临的问题和挑战;以期通过准确预测条锈菌优势小种、合理利用和布局抗病基因、利用新策略创制新型持久广
谱抗病材料,不断优化和完善小麦条锈病综合治理技术体系,实现小麦条锈病可持续控制。
关键词:小麦条锈菌;致病性;变异
Advances in Research of Pathogenicity and Virulence Variation of
the Wheat Stripe Rust Fungus Puccinia striiformis f. sp. tritici KANG Zhen-sheng, WANG Xiao-jie, ZHAO Jie, TANG Chun-lei, HUANG Li-li
(College of Plant Protection, Northwest A&F University/State Key Laboratory of Crop Stress Biology for Arid Areas,
Yangling 712100, Shaanxi)
Abstract: Wheat stripe rust, caused by Puccinia striiformis West. f. sp. tritici Eriks. and Henn. (Pst), is an air-borne cryogenic fungal disease, which causes serious yield and economic losses and severely threatens the security of wheat production worldwide due to its wide transmission and high frequent epidemics. As an obligate parasitic pathogen, the rapid virulence variation of Pst leads to resistance breakdown of the existing resistant wheat cultivars, which then leads to the constant epidemic of the disease. Thus, the study on virulence variation mechanism of Pst has been paid much attention since 1950s. Until now, great efforts have been made and significant progress has been achieved. The progress in research of the disease made in recent years and the relationship between resistance breakdown of wheat cultivar and virulence variation, the variation ways of Pst pathogenicity, genetic structure analyses of Pst population and the molecular mechanism underlying the rapid virulence variation of Pst were summarized. Based on this review, the issues and challenges were put forward to accurately predict main predominant races, rationally use and distribute the resistance genes, and to create the new kind of disease-resistance materials through novel strategies, which will optimize and improve the comprehensive management system for wheat stripe rust, and help to realize the sustainable control of wheat stripe rust.
Key words: Puccinia striiformis f. sp. tritici (Pst); pathogenicity; variation 小麦条锈病(wheat stripe rust or yellow rust)是严
重威胁世界主要产麦国家小麦生产安全的重要真菌病
害[1],在世界各小麦产区均有发生,而在夏季湿润的
温带地区或夜间冷凉的高海拔地区尤为严重。其病原
为条形柄锈菌小麦专化型(Puccinia striiformis West. f. sp. tritici Eriks. & Henn.),具活体专性寄生、喜冷凉
jia
文本框
返回目录

3440 中 国 农 业 科 学 48 卷
的特性,可通过高空气流远距离传播,随降雨或结
露侵染小麦引发病害,通常可造成 0.5%—5%的产
量损失,大发生时可造成 5%—25%的产量损失,甚
至绝收[2-3]。1958—2003 年,美国有 12 个年份小麦条
锈病严重发生,造成高达 25%的产量损失[2]。1999—2009 年期间,在中亚和高加索地区,小麦条锈病 4 次
流行性暴发并造成 20%—40%的小麦产量损失[4-5]。澳
大利亚 1979 年首次发现小麦条锈病,1984—1986 年因
该病害流行性暴发,造成均约 80%的产量损失[6-8]。1980—1981 年,小麦条锈病造成新西兰小麦减产幅度高达
60%[9]。南非自 1996 年小麦条锈病发生以来,该病
不断暴发流行,仅 1998 年就造成自由邦(Free State)东部 4.2×104 hm2 冬小麦受害,导致重大的经济损
失(约 2.25×106 美元)[10]。 而中国是世界上小麦条锈病 大的流行区域。
1949 年以来,小麦条锈病在中国每年均有不同程度
的发生和危害,年均发生面积约 4×106 hm2。历史上
小麦条锈病 4 次(1950、1964、1990 和 2002 年)大
流行均导致 1.0×109 kg 以上的产量损失,损失小麦总
计约 1.2×1010 kg。仅 1950 年,条锈病造成的产量损
失就占全国小麦总产量的 41.4%[11-12]。因而,条锈
病是严重威胁世界小麦粮食安全生产的重大真菌病
害,而且一直是世界上小麦主产国 具威胁性的生
物灾害。
1 小麦品种抗病性丧失与病原菌致
病性变异
利用抗病品种是防治病害 为经济、有效且环
保的措施。然而,由于条锈菌致病性变异频繁,新
致病小种不断出现,常导致小麦抗病品种“丧失”
其抗条锈性,而失去利用价值。一般情况下,小麦
品种在生产上使用 3—5 年便“丧失”其抗锈性[12-13]。
在中国,每次新致病小种的出现与品种抗条锈性“丧
失”均首先发生在甘肃的南部与四川的西北部等地,
这些区域被称为条锈菌“越夏易变区”或条锈菌新
小种产生的策源地[11,14-15]。1950—2010 年,由于小
麦条锈菌致病性变异,先后导致中国小麦品种碧蚂 1号(Yr1)、农大 183 (1955—1956);玉皮、甘肃
96(1960—1962);南大 2419(1962);阿勃(YrAbb1、YrAbb2)、甘麦 8 号(1969);中梁 11、丰产 3 号、
泰山 1 号、农大 139(1975-1977);洛夫林(Yr9)及其衍生品种(1977)、繁 6(Yr3b、Yr4b)系列品
种(1985)、中梁系列品种(2005)、贵农 22(Yr10、YrMor)系列(2010);川麦 42(Yr24)(2010)、
兰天 17(Yr26,Yr26=Yr24)、Moro(Yr10)(2009)等“丧失”抗条锈性[14-19](表 1),造成小麦条锈病
主要流行区 7 次大规模的品种更替。值得注意的是,
2009 年分离到的 V26 新菌系(avrYr10/24/26/ch42),
对贵农 22、川麦 42、Moro(Yr10)、近等基因系 NILS12(Yr10)、NILS16(Yr24)与 NILS17(Yr26)以及
38 个四川小麦新品种均具有致病性,已引起我国相关
部门高度重视。在中亚和高加索地区,2002—2009 年
由于小麦条锈病大流行(小种 6E20、6E148、7E140、7E148、7E150、7E156、15E150 和 39E158),导致
乌孜别克斯坦 24 个主要小麦品种中的 22 个“丧失”
抗病性以及哈萨克斯坦南部的大部分品种高度感病[4]。
在北美,20 世纪 60 年代,条锈病大流行先后导致小
麦抗条锈品种 Suwon 92[20]和小麦品种 Moro(含 Yr10)“丧失”其抗条锈性[21]。2002 年,美国加州大面积种
植的具有小种特异性抗性的小麦品种被条锈菌新小种
PST-78 克服,而造成条锈病大流行和不同程度的小麦
产量损失[2]。因而,从全球范围来看,条锈菌新致病
性小种的出现和发展,均会导致抗病品种抗性的“丧
失”,给小麦安全生产造成巨大威胁。
2 小麦条锈菌致病性变异途径
小麦条锈菌致病性是指条锈菌对具一定抗病基因
品种的专化致病能力。条锈菌致病性变异是导致小麦
品种抗条锈性“丧失”的主要因素。小麦条锈菌致病
性变异的途径主要有突变、异核作用和有性生殖。 2.1 突变
突变一般是指基因的变化,广义的突变还包括染
色体畸变。突变的原因包括细胞分裂时遗传基因的复
制发生错误,或受化学物质、辐射或病毒的影响。 突变是锈菌致病性变异的一种重要途径[22]。早
在 1930 年,Newton 等发现了禾柄锈菌小麦专化型
(P. graminis f. sp. tritici),即小麦秆锈菌突变,引
起病菌致病力和颜色的变化 [23]。随后,1932 年
Gassner 等[24]首先证实了小麦条锈菌通过突变可以
产生新小种,并估算出其突变率约为 1.6×10-5—2.0×10-5。1968 年,Kajiwara 等[25]获得一个致病力减弱
的条锈菌突变菌系,该突变菌系对小麦品种
Chinese166(Yr1)表现无致病性(反应型为 1),
而其原始菌系(42A2)对 Chinese 166 表现高度致病
(反应型为 4)。

17 期 康振生等:小麦条锈菌致病性及其变异研究进展 3441
表 1 中国主栽小麦品种或种质资源抗病性“丧失”与小麦条锈菌新小种出现的关系
Table 1 Relationship between the resistance breakdown of main wheat cultivars or germplasms and new races of the wheat stripe rust pathogen Puccinia striiformis f. sp. tritici in China
年代 Years
抗性“丧失”的主栽小麦品种或种质资源 Main wheat cultivars or germplasms of breakdown resistance
小种 Corresponding races
1950—1960 碧蚂 1 号(Yr1)、西北 54(Yr1)、西北丰收、农大 183 等 Bima 1 (Yr1), Xibei 54 (Yr1), Xibeifengshou, Nongda 183, etc.
条中 1 号 CYR1
1960—1962 玉皮、甘肃 96、Shaannong9 号、西北 134 等 Yupi, Gansu96, Shaannong 9, Xibei 134, etc.
条中 8 号、条中 10 号 CYR8, CYR10
1962—1970 南大 2419 及其衍生系 Mentana and its derivates 条中 13 号、条中 16 号 CYR13, CYR16
1970—1976 北京 8 号(Yr1)、北京 10 号、石家庄 54、阿勃等 Beijing 8 (Yr1), Beijing 10, Shijiazhuang 54, Abbondanza, etc.
条中 17 号、条中 18 号 CYR17, CYR18
1976—1985 丰产 3 号、官村 1 号、泰山 1 号、泰山 4 号、卫东 8 号、农大 139、阿夫(YrA)、白泉 40、偃大 24、尤皮 II 号、天选 15、中梁 5 号、武农 132 等 Fengchan 3, Guancun 1, Taishan 1, Taishan 4, Weidong 8, Nongda 139, Funo (YrA), Baiquan 40, Yanda 24, Jubilejina II, TianXuan 15, Zhongliang 5, Wunong 132, etc.
条中 19 号、条中 23 号、条中
25 号 CYR19, CYR23, CYR25
1985—1995 洛夫林及其衍生品种:洛夫林 10(Yr9)、洛夫林 13(Yr9,+)、丰抗 8 号(Yr9)、鲁麦 1 号(Yr9,+)、
鲁麦 5、7、11 号、鲁麦 14(Yr9,+)、鲁麦 15(Yr+)、鄂恩 1 号(Yr9)、白农 3217(Yr1,2,3,+)、
小偃 6 号、京麦 33 号(Yr1,+)、陕 7859、皖 7107(Yr+)、济麦 26、济麦 30(Yr+)、济麦 36、豫
麦 13、豫麦 18(Yr1,+)、豫麦 21(Yr9,+)、豫麦 25(Yr+)、扬麦 5 号、扬麦 158 等 Lovrin and its derivates: Lovrin 10 (Yr9), Lovrin 13 (Yr9,+), Fengkang 8 (Yr9), Lumai 1 (Yr9,+), Lumai 5, 7, 11, Lumai 14 (Yr9,+), Lumai 15 (Yr+), Een 1 (Yr9), Bainong 3217 (Yr1,2,3,+), Xiaoyan 6, Jinmai 33 (Yr1,+), Shaan 7859, Wan 7107 (Yr+), Jimai 26, Jimai 30 (Yr+), Jimai 36, Yumai 13, Yumai 18 (Yr1,+), Yumai 21(Yr9,+), Yumai 25 (Yr+), Yangmai 5, Yangmai 158, etc.
条中 28 号、条中 29 号、洛夫
林 10 致病类群和洛夫林 13 致
病类群 CYR28, CYR29, Lv 10, and Lv 13 pathotype group
1995—2000 繁 6 及其衍生系:水源 11 系列、绵阳系列(绵阳 11、绵阳 15、绵阳 19、绵阳 20)、川麦系列[川麦
22、川麦 23(Yr9)]等 Fan 6 and its derivates: Suwon 11 lines, Mianyang 11 lines (Mianyang 11, Mianyang 15, Mianyang 19, and Mianyang 20), Chuanmai lines [Chuanmai 22, and Chuanmai 23 (Yr9)], etc.
条中 30 号、条中 31 号、条中
32 号、杂 46 致病类群、水源
11 致病类群 CYR30, CYR31, CYR32, Hy46, and Su11 pathotype group
2000— 贵农 21、贵农 22、兰天 17(Yr26, Yr26=Yr24)、川麦 42(Yr24)、92R137(Yr26)及其衍生系、Moro(Yr10、YrMor)、洮系品种(洮 157、洮 153、洮 718)、清农系品种(清农 3 号、中梁 4 号)、兰
天系品种(兰天 3 号、兰天 4 号、兰天 6 号)、中梁系品种(中梁 16 号、17 号、18 号、20 号、25 号、
29 号)、天选系品种(天选 40 号、41 号、42 号)、绵阳系品种(绵阳 26 号、27 号、28 号、30 号)、
陇鉴系品种(陇鉴 64、陇鉴 196)、冀麦 30 号、北京 837、晋麦 47、鲁麦 22 号、高优 50、武都 12号、15 号、济南 17 号、皖麦 19 号、豫麦 34 号、豫麦 49 号、豫麦 54 号、豫麦 70 号、郑麦 9023、烟
优 361、陇原 061、陇原 931、杂 46、抗引 655 等 Guinong 21, Guinong 22, Lantian 17 (Yr26, Yr26=Yr24), Chuanmai 42 (Yr24), 92R137 (Yr26) and its derivates, Moro (Yr10, YrMor), Tao lines: Tao 157, Tao 153, and Tao718, Qingnong lines: Qingnong 3, Zhongliang 4, Lantian lines: Lantian 3, Lantian 4, and Lantian 6, Zhongliang lines: Zhongliang 16, Zhongliang 17, Zhongliang 18, Zhongliang 20, Zhongliang 25, and Zhongliang 29, Tianxuan lines: Tianxuan 40, Tianxuan 41, and Tianxuan 42, Mianyanglines: Mianyang 26, Mianyang 27, Mianyang 28, and Mianyang 30, Longjian lines: Longjian 64, and Longjian 196, Jimai 30, Beijing 837, Jinmai 47, Lumai 22, Gaoyou 50, Wudu 12, Wudu 15, Ji’nan 17, Wanmai 19, Yumai 34, Yumai 49, Yumai 54, Yumai 70, Zhengmai 9023, Yanyou 361, Longyuan 061, Longyuan 931, Hybrid 46, Kangying 655, etc.
CYR33、中四致病类群、贵农
22 致病类群、Yr10 致病类群
CYR33, Z4,G22, and Yr10 pathotype group
人工射线诱变也可导致小麦条锈菌突变产生新致
病菌系。1968 年,Stubbs[26]通过利用 X 射线照射条锈
菌 3 个致病型的夏孢子,成功地筛选了不同致病力的
突变体,其中一个突变菌系对当时条锈菌普遍表现抗
病的斯卑尔脱小麦(Triticum spelta album,Yr5)表现
致病;1994 年,中国学者商鸿生等[27]利用紫外线处理
小麦条锈菌条中 29 号(CYR29,CYR=Chinese Yellow Rust)单孢菌系,获得了 7 个致病力变异菌系,并指
出突变是中国小麦条锈菌重要的变异途径;2009 年,
王欣丽等[28]利用紫外线对小麦条锈菌小种条中 23 号
(CYR23)进行诱变,获得了致病力明显增强的突变
菌系尤 II-23 和水源 11,其中尤 II-23 经转接 4 代仍保

3442 中 国 农 业 科 学 48 卷
持致病性,而水源 11 菌系经转接两代后回复突变,致
病力与突变前相近。2005 年黄丽丽等[29]对小麦条锈菌
小种 CYR29 的单孢菌系进行紫外线诱变获得了两个
致病力变化的新菌系 Jubi 菌系和 Funo 菌系,前者经
诱变后在小麦品种尤 II 的致病力由突变前的高抗(反
应型为 0)变为高感(反应型为 4);而后者在小麦品
种阿夫的致病力由突变前的高感(反应型为 4)变为
中抗(反应型为 2)。同时,还获得了一个对小麦品
种 Hybrid 46 致病的突变菌系,但经转接两代后发生
回复突变。对突变菌系 DNA 多态性分析发现,突变
菌系与野生型菌系存在明显的多态性差异(分别为
10.58%和 11.57%),表明紫外线诱变可使条锈菌基因
组 DNA 发生较大变化[29]。综上,紫外线诱导小麦条
锈菌突变是不定向的,即可以发生正向的致病力突变,
也可以发生反向的致病力突变,还可发生回复突变。 化学诱变也可导致锈菌致病性发生变异。化学诱
变是指通过化学诱变剂处理生物体来诱发突变。植物
病原菌中常用的化学诱变剂主要有甲基磺酸乙酯
(ethyl methane sulfonate,EMS)、硫酸二乙酯(diethyl sulfate,DES)和亚硝基胍(nitrosoguanidine,NTG)
等。在锈病研究中,利用 EMS 对燕麦秆锈菌(P. graminis f. sp. avenae)[30]和小麦秆锈菌[31]、NTG 对亚
麻锈菌(Melampsora lini)[32]进行了化学诱变其对锈
菌致病力的影响研究。在利用化学诱变剂研究小麦条
锈菌突变方面,姚秋燕等[33]利用 EMS 诱变小麦条锈
菌小种条中 17 号(CYR17)和条中 31 号(CYR31),
获得了 4 个 CYR17 和 1 个 CYR31 的突变菌系,且突
变频率明显不同,介于 2.14×10-6—3.68×10-5。 近些年来,随着分子生物学技术的发展,利用生
物诱变技术(转座子诱变)获得突变体进行致病性变
异研究得到了快速发展。通常用于生物诱变的转座子
主要有插入序列、转座子和 Mu 噬菌体。基因枪法是
一种插入诱导突变的方法,对无法进行原生质体培养
的活体寄生菌有较高的转化效率[34]。锈菌方面在小麦
秆锈菌[35]、小麦叶锈菌(P. triticina)[36]、菜豆锈菌
(Uromyces appendiculatus)[37]都有应用实例。2007年,张如佳等[38]利用基因枪转化技术,以小麦条锈菌
白化菌系为转化受体,以含有潮霉素抗性基因(Hyg)的质粒为插入载体,对小麦条锈菌进行了插入突变研
究,获得了两个稳定的致病力突变菌系 MM-2 和
MM-3,它们对小麦品种南大 2419 的致病力均减弱,
但突变菌系 MM-3 同时出现致病力分化。2013 年,王
阳等[39]通过基因枪转化GUS成功获得了3个小麦条锈
致病性突变体,证实外源 GUS 在突变菌株中可稳定遗
传,外源基因片段整合到条锈菌基因组中引起条锈菌
致病性变异,且外源基因的插入位点与条锈菌的致病
性基因相关。这些尝试为小麦条锈菌致病性变异及其
相关的基础研究奠定了基础。 2.2 异核作用
异核现象(heterokaryosis)是指一个细胞中或单
一菌丝细胞中含有两种或两种以上遗传特性不同的细
胞核的现象[40],条锈菌可以通过两个小种的芽管结合
(fusion of germ tube)或菌丝融合(hyphal fusion)进
行整个核的交换和重新组配。 通过异核作用产生锈菌新致病型 早在小麦秆锈
菌中发现[41]。其后,异核现象在锈菌新小种产生中的
作用受到国内外学者的高度关注。研究表明小麦条锈
菌可通过异核作用进行基因组的重组产生致病力新小
种[42-43]。1969 年,Little 等[43]利用小麦条锈菌小种 2B和 8B 单孢菌系的夏孢子,混合接种两个菌系均可感
染的小麦品种,然后进行单孢分离和筛选,结果产生
了 3 个新小种,其致病性与原始菌系明显不同;1971年,Volin[44]用 BFMO 和 Bw(a)菌系在小麦品种 Lemhi上接种筛选,发现一个对小麦 Red River68 有致病力
的新小种;1976 年,Taylor[45]利用条锈菌白化菌研究
发现体细胞重组(somatic recombination)也可产生新
小种。而值得注意的是,经异核作用产生的小麦条锈
菌新小种,夏孢子芽管具 3 或 4 个细胞核的比例明显
高于原始菌系[42,46]。1976 年,Goddard 用小麦条锈菌
小种 104E137 和 41E136 进行混合接种,从 50 个单孢
子分离物中筛选获得了 3 个有双亲特性的新菌系。并
且由此认为,细胞核的交换与重组是条锈菌致病性变
异的基础[46]。 在中国,1964 年赵来顺等[47]发现了条锈菌夏孢子
芽管融合的现象;康振生等[48-49]利用生物学和细胞学
的方法研究证实中国小麦条锈菌通过异核作用产生新
菌系及异核新菌系的多核现象。从条锈菌不同小种 38个单孢菌系建立的 120 个组合中,筛选出 3 个新菌系,
即阿夫菌系、尤 II 菌系和 R.R68 菌系。通过小麦条锈
菌(条中 28-2)的黄色夏孢子与白化菌系(BW-2)的
白色夏孢子混合接种后,筛选出对小麦南大 2419 致病
的白色新菌系-阿夫菌系,其致病性明显不同于两个亲
本菌系。通过对条中 28-2(CYR28 的单孢菌系)与条
中 20-3(CYR20 的单孢菌系)混合接种获得侵染小麦
品种尤皮 II 号的新菌系-尤 II 菌系,两亲本菌系均不
侵染尤皮 II 号。通过条中 22 号(CYR22)与条中 25

17 期 康振生等:小麦条锈菌致病性及其变异研究进展 3443
号(CYR25)混合接种,筛选出侵染小麦品种 Red River68 的新菌系-R. R68 菌系。同时,发现新菌系存
在多核现象,即新菌系夏孢子芽管多核率高于两亲本
菌系,异核体的多核率随继代繁殖而降低,但有的菌
系的致病性仍保持稳定,并发展成为新致病菌系。 小麦条锈菌多核现象在其整个侵染过程中极为普
遍。1994 年,康振生等研究发现条锈菌夏孢子萌发形
成的芽管、胞间菌丝、吸器母细胞以及吸器中均存在
多核现象,认为小麦条锈菌的细胞核相状况较为复杂,
并且不同于其他锈菌,为研究小麦条锈菌异核作用提
供了线索[50]。他们的研究发现,小麦条锈菌夏孢子通
常为双核体,夏孢子萌发的芽管可为双核、三核或四
核,但仍以双核芽管为主。尽管在胞间菌丝和吸器母
细胞中可观察到典型的双核细胞,但胞间菌丝和吸器
母细胞中的多核现象极为普遍。吸器的细胞核存在双
核、三核、四核、五核和六核,但以四核为主,而夏
孢子堆中不同类型的细胞均为双核。条锈菌胞间菌丝
间的融合十分常见,并可观察到细胞核在菌丝细胞间
的转移。这种异核菌丝体经细胞核的重新组合与双核
化作用,无疑可导致异核新菌系的形成。 条锈菌的多核现象同样也存在于小麦条锈菌的自
然群体中。1994 年,康振生等[48]对田间条锈菌自然群
体中夏孢子的核相研究发现,在甘肃陇南等地,自然
条件下夏孢子不仅存在双核体,还存在三核体
(0.100%)和四核体(0.009%)。这种条锈菌的自然
多核现象,在 2004 年王阳等[51]对甘肃天水不同海拔
地区小麦条锈菌夏孢子多核率的研究中也得到了进一
步证实,但多核率(0.42%—0.55%)明显更高。同时,
研究发现,经继代培养后,有些夏孢子的多核率随之
下降,只是在多核率下降频率上有所差异,有的菌系
多核率下降得快(如菌系 97-33),有的菌系下降得
慢(如 97-44),这一发现与康振生等早期的研究结
论一致。1991 年,马青等[52]研究表明小麦条锈菌小种
内、小种间及条锈菌专化型间(小麦条锈菌白化菌系
与大麦条锈菌)均可发生芽管融合,但不同小种间及
专化型间芽管结合率均较低。 2.3 有性生殖
禾谷类锈菌是具有 5 种不同类型孢子(0 性孢子;
I 锈孢子;II 夏孢子;III 冬孢子;IV 担孢子)的大循
环锈菌(macrocyclic rust),需要在两种不同属的
寄主上才能完成其生活史。早期证实,小麦秆锈菌
与叶锈菌都存在有转主寄主(alternate host),而小
麦条锈菌并未证实存在转主寄主[53-56]。直到 2009 年,
通过人工室内接种才证实灌木小檗(Berberis spp.)是
小麦条锈菌的转主寄主[57]。随后,十大功劳(Mahonia spp.)[58]也被证实是小麦条锈菌的转主寄主。而自然
条件下条锈菌是否存在有性生殖及其在病害流行中的
作用目前已成为全球备受关注的热点。 近,笔者研究室对中国小麦条锈病主要流行区
的小檗资源进行了调查,发现堆花小檗(Berberis aggregata)、短柄小檗(B. brachypoda)、假豪猪刺
(B. soulieana)、陕西小檗(B. shensiana)等 35 种
小檗广泛分布在甘肃、四川、陕西等地,在小麦条锈
菌“越夏易变区”十分常见,并且毗邻小麦田块[58-60]。
目前,通过接种鉴定,证实中国已有 28 种小檗可以作
为小麦条锈菌的转主寄主。根据《中国植物志》
(http://frps.eflora.cn/frps/ Berberidaceae)记载,中国
有丰富的小檗(约 250 种)和十大功劳(约 35 种)资
源。因而,未来或将有更多小檗或十大功种类被鉴定
为小麦条锈菌的转主寄主。 有性繁殖被认为是锈菌致病性变异的重要途径之
一。早期研究证实,小麦秆锈菌与叶锈菌通过有性
生殖可导致其后代群体发生致病性变异产生新的小
种[24,61-63],而有性繁殖在小麦条锈菌致病性变异中的
作用一直无法被证实。直到小麦条锈菌转主寄主小檗
的发现,使得研究条锈菌致病性的遗传学研究成为可
能。通过室内实验,Wang 等[64]揭示了条锈菌小种经
过有性繁殖后发生遗传重组与分离,有性生殖在条锈
菌致病性变异中起重要作用,从遗传学角度初步揭示
了条锈菌毒性基因(virulence)与无毒基因(avirulence)的关系。通过对美国条锈菌 PST-127 的单孢子堆菌系
经有性生殖后的 19 个单夏孢子堆菌系进行了致病性
分析,认为对 Yr1、Yr2 和 Yr9 致病的以及对 Yr5、Yr15、Yr24 和 YrSP 不致病的后代菌系在这些抗病位点是纯
合的,而在其他 14 个 Yr 基因抗病位点发生分离,在
除 Yr17 和 Yr43 的其他单基因位点上 virulence/ avirulence 分离比约为 1﹕1。而笔者研究室通过对小
麦条锈菌单一小种有性生殖后代群体的致病力分析,
获得了与亲本菌系致病力不同的新致病类型 41 个,表
明小麦条锈菌经有性生殖可发生致病性变异并产生新
小种。 早期研究发现小檗在小麦秆锈病的病害流行中起
着重要作用。在欧洲、北美一些国家,小麦秆锈菌在
春季可侵染小檗产生锈孢子,并向邻近小麦提供初始
菌源引起秆锈病的发生[65],而且秆锈菌在小檗上的有
性生殖可导致新小种的产生。那么,小檗在小麦条锈

3444 中 国 农 业 科 学 48 卷
病的病害流行中是否具类似的作用是一个亟待回答的
问题。为此,近年来笔者研究室开展了系统的研究工
作,首次证实小麦条锈菌在自然条件下不仅能侵染小
檗完成有性生殖,而且在小麦条锈病的病害循环中起
重要作用:一是能够提供一定数量的菌源引起小麦条
锈病的发生,二是可导致条锈菌致病性变异而产生新
小种[60]。2013 年,Zhao 等[60]从中国甘肃、陕西、西
藏等地自然条件下受侵染的不同种类小檗上先后分离
到 20 个小麦条锈菌菌系,首次获得了条锈菌自然条件
下进行有性生殖的直接证据。通过在中国鉴别寄主的
致病力鉴定发现,在这 20 个从小檗上分离到的小麦条
锈菌菌系中,既有当前流行的小种,也有新菌系,表
明在自然条件下,小麦条锈菌有性过程与田间条锈菌
群体存在着联系;随后,利用这 20 个菌系建立了单夏
孢子堆菌系群体,经在单基因系(Yr)鉴别寄主进行
致病力鉴定,发现这些单夏孢子堆菌系间存在明显的
致病性多态性,进一步证实条锈菌自然条件下有性生
殖导致其致病性变异。同时,在不同省份的调查中发
现,感病小檗与小麦毗邻生长的状况非常普遍,在有
的年份,不仅小檗感病严重,而且邻近的小麦植株上
也观察到条锈病症状。通过在田间自然条件下用小麦
条锈菌冬孢子侵染小檗,随后从受侵染的小檗上及与
其邻近的小麦上分离获得了多个相同的小麦条锈菌小
种与新小种,从而证实在感病小檗、小麦、存活的小
麦条锈菌冬孢子共同存在的情况下,如果气候条件适
宜,小麦条锈菌可以在转主寄主小檗上顺利完成有性
生殖,释放锈孢子并引起小麦条锈病的发生,同时产
生致病性变异形成新小种。上述这些新的发现逐步揭
开了小麦条锈菌“易变区”的神秘面纱,即转主寄主
小檗的广泛分布与条锈菌有性繁殖的发生无疑是中国
小麦条锈菌“易变区”形成的主要原因。 条锈菌有性生殖的存在使得中国小麦条锈菌自然
群体表现出丰富的遗传多态性[66-68]。然而,美、欧等
国小麦条锈菌自然群体的研究结果则截然不同。2014年,Cheng 等通过对美国太平洋西北部的 26 个田块
(204 个样本)与其他 20 个州(66 个样本)共 270个条锈菌系的群体分析发现,美国条锈菌群体为两个
遗传群组[68]。经基因型比率、多位点连锁不平衡与杂
合性的分析发现,美国的太平洋西北部与其他州的条
锈菌群体为无性繁殖。同时研究证实,在美国太平洋
西北部地区,小檗对小麦条锈病的发生不起作用,对
小麦秆锈病的发生起重要作用[69-70]。对欧洲[69,71-72]和
澳洲[70]自然界条锈菌群体的研究分析也表明,条锈菌
群体均为无性繁殖而来,不存在有性繁殖。 不同专化型的条锈菌有性杂交可否导致小麦条锈
菌致病性变异亦是一值得探索的问题。条形柄锈菌的
寄主十分广泛,存在不同的专化型[3,73]。目前,条形
柄锈菌小麦专化型有性阶段的寄主(即转主寄主)已
经被证实,但其他条锈菌专化型的有性阶段的寄主尚
未被发现。 近,笔者研究室通过人工接种证实,小
檗是包括早熟禾在内的其他几种禾本科草的条锈菌专
化型的共同转主寄主。几种不同的条锈菌专化型均可
成功侵染小檗并产生性子器、锈子器,在小檗上完成
其整个有性生殖过程。据此推测,在自然条件下,不
仅小檗上的锈菌组成非常复杂,而且不同的条锈菌专
化型有可能同时侵染小檗并发生有性杂交,产生新
的条锈菌致病类型,给研究小麦条锈菌致病性变异
成因带来了巨大的挑战。目前,不同条锈菌专化型
间的有性杂交正在研究中,以期揭示不同的条锈菌
专化型的有性杂交在小麦条锈菌致病性变异中所起
的作用。
3 条锈菌群体遗传与毒性变异
3.1 全球小麦条锈菌流行体系
小麦条锈病是一种气传病害,小麦条锈菌的夏孢
子能随气流远距离传播,通过气流远程扩散(long distance dispersal,LDD)迅速在新地区定殖或进行冬
夏栖息地之间的迁移[67]。LDD 使得小麦条锈病的流行
范围包括不同的自然地理区域、不同的国家、甚至不
同的大陆。在一个相当大的地域内,条锈病流行过程
是一个不可分割的整体,称为“大区流行”。对条锈
菌群体遗传结构进行大规模的分析,有助于揭示其传
播迁徙规律、遗传结构多样性及毒性变异方式,明确
现有群体的进化轨迹,进而预测新致病小种的产生与
流行。目前,对全球范围的条锈菌群体遗传结构进行
分析,发现在流行区域内主要存在 6 个遗传群体[74],
包括中亚地中海、中东-东非、欧洲西北部、巴基斯
坦、尼泊尔和中国。中东-东非是产生高温菌系的主要
发源地;欧洲是南美、北美和澳大利亚群体的发源地;
南非条锈菌则主要来源于地中海亚洲群体。通过不同
遗传分子标记对不同地理范围条锈菌群体结构分析
发现,全球范围内条锈菌群体整体呈较低遗传多样
性[72,75-77],但中国和巴基斯坦条锈菌群体却呈显著的
多样性[71,78]。喜马拉雅山区域(主要包括尼泊尔、巴
基斯坦和中国)条锈菌的遗传多样性、重组群体和
有性重组机率明显高于其他地区,推测喜马拉雅山及

17 期 康振生等:小麦条锈菌致病性及其变异研究进展 3445
侵染引起小麦条锈病,在受侵组织(主要
为叶片)产生夏孢子堆与夏孢子(Ⅱ)
春季—锈孢子或越冬后产生的夏孢子随气流传播侵染返青冬小麦
或春播小麦
侵染小麦进行无性繁殖
侵染转主寄主(小檗)进行有性繁殖
秋、冬季—越夏菌源
侵染秋播小麦并越冬
夏孢子随气流传
播进行重复侵染
小麦生长后期,形成冬孢子堆与
冬孢子(Ⅲ)夏季—在冷凉山区晚熟小麦或自生麦苗上或杂草寄主上越夏存活,秋季在杂
草寄主上产生冬孢子堆
冬孢子萌发产生
担孢子(Ⅳ)
担孢子侵染转主寄主小檗产
生性孢子器与性孢子(0)
在转主寄主小檗受侵叶(有时侵染幼茎或
叶柄)背面产生锈孢子器与锈孢子(Ⅰ)
图 1 小麦条锈菌生活史与病害循环
Fig. 1 Life history and disease cycle of P. striiformis f. sp. tritici
附近区域是条锈菌的发源地,是新致病小种产生的起
源[74]。 3.2 中国小麦条锈菌“越夏易变区”
欧洲和澳大利亚的小麦条锈菌群体毒性变异快,
却呈现较低的遗传多样性。与之不同的是,中国条锈
菌群体呈现高度的遗传多样性,这种遗传多样性可能
起源于条锈菌“越夏区”[71]。条锈菌对低温具有较强
的适应能力,却不耐高温,因而,越夏便成为条锈菌
侵染循环中的关键环节。一般来讲,小麦条锈菌只能
在特定的地区越夏和越冬,菌源必须在两者间进行规
律性的转移,方能完成其周年循环。中国小麦条锈
菌主要是以夏孢子世代连续侵染的方式在海拔 1 400 m 以上,特别是 2 000 m 以上的高山山坡和高原
地区的冬、春麦自生麦苗和晚熟春麦上越夏。经过锈
病工作者几十年的努力协作研究,已经揭示了中国小
麦条锈病的流行体系,明确了西北、四川西北、华北、
云贵、新疆等 5 大越夏区[11],特别是陇南、陇东和川
西北为中国小麦条锈菌 重要的“越夏区”,其中尤
以陇南 为关键。该地区不仅向中国东部广大麦区
提供初始菌源,还是中国小麦条锈菌新致病小种的
重要策源地[79]。多年来,新小种的出现和品种抗锈
性的丧失大多首先发生在这里,一直被称为“易变
区”。新致病小种的出现和群体数量的增长,病原
群体结构发生改变,导致一些品种的抗病性丧失,
引发病害的发生和流行。因而,认识“越夏易变区”
小麦条锈菌自然群体毒性结构对小麦抗锈育种、抗
病基因的合理布局及制定有效的小麦条锈病防治策
略是十分必要的。 3.3 “越夏易变区”小麦条锈菌群体遗传
利用传统病原菌毒性鉴定及病害调查方法,根据
病菌的毒性特征推断小种的演化关系并不能全面反映
条锈菌的遗传结构和中国条锈菌大区流行体系。因此,
各种分子遗传标记被开发并应用于条锈菌群体遗传的
研究,大大推动了条锈菌群体遗传学的发展。

3446 中 国 农 业 科 学 48 卷
早在 1998 年,Shan 等[80]以小麦条锈菌基因组特
异重复序列(P. striiformis repeat,PSR)为探针对采
自全国六省的 160 个小麦条锈菌标样进行了 DNA 指
纹分析,表明条锈菌群体存在很高的遗传多样性,不
同地区之间存在多样性,揭示了甘肃天水可能是 重
要的条锈菌越夏区。之后,陈长卿等开发了条锈菌
EST-SSR 标记,结合 TP-M13-SSR 技术建立了适合中
国小麦条锈菌群体遗传研究的微卫星分子标记体系。
利用该微卫星分子标记体系揭示了中国小麦条锈菌
群体具有高度的遗传多样性,种群间和种群内部存
在遗传分化,遗传变异主要存在于群体内部,不同
群体间分化水平有所差异,其中天水种群是中国小
麦条锈菌遗传多样性水平 高的种群。UPGMA 聚
类分析进一步明确了西北-川西北越夏区是中国小
麦条锈菌遗传多样性 为丰富的地区,是病原菌遗
传多样性的中心区域,且西北越夏区和川西北越夏
区存在一定的基因流,为中国小麦条锈菌远距离传
播提供了分子证据[81]。 基于西北和川西北越夏区在中国条锈菌传播与流
行中的重要地位,郑文明等[82]应用 PSR 探针对采自中
国条锈菌越夏关键区甘肃天水的 244 个标样进行指
纹分析,揭示该地区群体内存在丰富的遗传多态性。
进一步采用 SSR 分子标记技术对甘肃和青海 1 447个菌系进行 DNA 指纹分析,表明甘肃省小麦条锈菌
遗传多样性比较丰富,亚群体间遗传多样性有显著
差异。其中,天水种群和陇南种群的遗传多样性
高。Liu 等[83]采用 AFLP 对甘肃和云南两省的条锈
菌群体遗传结构进行比较分析,发现甘肃省条锈菌
群体结构遗传多样性(0.55)明显高于云南省(0.27);Mboup 等[71]对甘肃天水高山区的 412 个菌系进行
AFLP 和 SSR 分子标记,发现该区域条锈菌群体表
型和基因型呈现显著的多态性,分别高达 38%和
89%;Duan 等[69]通过 AFLP 分析发现,甘肃地区一
年中条锈菌样品的遗传多样性甚至是法国 20 年条
锈菌多样性的 7 倍之高。这些结果进一步明确了甘
肃作为中国小麦条锈菌越夏区的重要性。此外,陇
南地区不同生态区域小麦条锈菌呈现不同的群体遗
传多样性与分化程度,高山区与半山区种群多样性
丰富,分化程度也较大,川道区群体遗传多样性
则较低且分化程度较小。在不同海拔区域的条锈菌
群体之间发现基因流的存在,证实小麦条锈菌在高
海拔山区与川地之间进行移动,可就地完成周年循
环[84]。
川西北山区也是中国小麦条锈病重要越夏地区之
一,条锈菌的变异个体或形成的新菌系在这里也容易
保存、繁殖和积累[1]。TP-M13-SSR 分析表明,四川省
小麦条锈菌群体遗传多样性比较丰富,地区之间存在
明显的差异,川西北和四川盆地的种群遗传多样性相
对较高,而四川西南部和四川东南部的种群遗传多样
性较低。基因流和共享基因型从分子水平证实了四川
小麦条锈菌在地区间的传播,且川西北和四川盆地之
间的菌源交流 为广泛[85]。 伴随着高度的遗传多样性,SSR 标记技术检测到
陇南地区小麦条锈菌群体存在体细胞遗传重组现象,
平均频率为 16.6%,表明陇南地区条锈菌体细胞结合
现象十分普遍,推测自然条件下,中国小麦条锈菌可
通过遗传重组导致毒性变异[66]。Mboup 等[71]在甘肃越
夏区发现遗传重组现象,在高度多样性的尼泊尔和
巴基斯坦条锈菌群体中,也检测到丰富的遗传重组
现象[74],推测除却体细胞重组,条锈菌可能通过有性
生殖产生新的致病小种,导致致病性变异。然而,对
有性生殖、无性重组与病菌致病性变异,转主寄主小
檗与致病菌系的演化及其与病害发生、流行之间的关
系等仍有待进一步研究。
4 条锈菌基因组研究及其致病性调控
研究小麦条锈菌的致病性及其变异成为挖掘条锈
病防控新策略的一个重要课题。然而,条锈菌作为一
种专性活体寄生真菌,不能人工培养,缺乏稳定的遗
传转化体系,条锈菌致病机理研究进展一直比较缓慢。
随着近年来生物技术、信息和计算机技术的发展,小
麦条锈菌致病机理的研究得以迅速推进和发展。 4.1 条锈菌基因组和比较基因组
随着第二代测序技术的发展,2009 年 Broad Institute 组织开展了锈菌基因组测序及比较基因组学
研究。2011 年,Cantu 等[86]利用 Illumina 测序技术完
成了美国条锈菌强致病性小种 PST-130 的基因组测
序。基因组大小为 64.8 Mb,编码 22 815 个基因,含
29 178 contigs,覆盖率 88%以上[86]。由于条锈菌具有
杂核现象,2013 年,Zheng 等 [87]利用“Fosmid to Fosmid”策略对条锈菌中国优势小种 CYR32 进行了
基因组测序和组装,该策略相对于传统的测序方法在
分析杂核方面更具有优势。条锈菌 CYR32 基因组大
小 110 Mb,编码 25 288 个蛋白,基因组高度杂合。
同时,为分析条锈菌的进化和致病性变异,他们对来
自不同地区(中国、美国、澳大利亚、匈牙利、巴基

17 期 康振生等:小麦条锈菌致病性及其变异研究进展 3447
斯坦)的 6 个小种进行了重测序,根据 SNP 和 InDel分析发现,不同国家地区的条锈菌小种存在较大的遗
传变异,但这些小种的进化关系与它们的致病性特征
和地理分布并没有必然联系;另外,在基因编码区 SNP约为 35%,且半数为非同义替换,意味着有性生殖在
不同地域小种的起源上具有重要作用[87]。条锈菌转主
寄主小檗在中国西部和中亚广泛分布,因而,这些区
域小麦条锈菌群体间巨大的遗传变异暗示了有性生殖
在条锈菌致病性变异及进化方面的重要作用。 4.2 条锈菌功能基因研究
表达序列标签(expressed sequence tags,ESTs)和转录组测序(transcriptome sequencing)的发展揭开
了锈菌功能基因研究的序幕。2007 年,Ling 等 早开
始了小麦条锈菌功能基因组的研究,构建了条锈菌
PST-78 生理小种的夏孢子 cDNA 文库[88],该文库包含
42 240 个克隆,cDNA 平均长度为 1.9 kb。随后,Zhang等[89-90]分别构建了条锈菌萌发芽管和吸器的 cDNA 文
库,分别获得了 4 798 个和 5 126 条 EST 序列,获得
了一系列病菌功能基因,主要涉及细胞代谢、信号转
导及致病性等相关基因。随着转录组测序技术的诞生,
2013 年,Garnica 等[91]通过 illumina RNAseq 测序技术
完成了条锈菌吸器和夏孢子的转录组测序,明确了病
菌不同发育阶段的基因表达特征,获得了 400 个分泌
蛋白基因。其中 2/3 在吸器中上调表达,主要为真菌
细胞壁修饰蛋白和潜在的致病效应蛋白。并且,夏孢
子及吸器的转录组比较发现,条锈菌夏孢子的主要功
能是储存生长发育所需的能量,而吸器则利用寄主大
量能量进行生物合成和产孢。 对条锈菌不同发育阶段基因转录表达的认识,推动
了条锈菌功能基因的鉴定和功能分析。2009 年,梁晓
飞等 [92]克隆了条锈菌几丁质合成酶基因 PstChsII,其在
条锈菌萌发过程中上调表达,可能参与了条锈菌细胞壁
的合成;2011 年,Guo 等[93]克隆到条锈菌第一个 MAPK家族基因 PsMAPK1,它属于 Fus3/Kss1 家族。该基因在
条锈菌侵染寄主早期诱导表达并在吸器形成阶段达到
高峰,推测在病菌致病过程中具有作用,并借助异源体
系证明了该基因可以影响病菌的生长和致病性,为条锈
菌基因功能的研究打开了一条新的道路。随后,2013年 Guo 等[94]克隆了一个条锈菌夏孢子特异表达基因
PsCon1,该基因在条锈菌休眠的夏孢子中表达 高,随
着夏孢子的萌发、侵染,其表达量持续降低。寄主诱导
的基因沉默(host-induced gene silencing,HIGS)技术
的诞生,实现了通过在病原菌侵染寄主植物过程中抑制
病原菌基因的表达,分析病原菌基因的功能。2011 年,
Yin等[95]建立了利用大麦条纹花叶病毒系统沉默锈菌基
因分析条锈菌基因的功能。随后,2012 年 Zhang 等[96]
利用 HIGS 技术证明了小麦条锈菌钙调磷酸酶基因
PsCNA1 和 PsCNB1 可以调控条锈菌形态分化及吸器形
成,进而参与条锈菌致病过程;2014 年 Cheng 等[97]发
现了锈菌特异的蛋白激酶 PsSRPKL,证明它是锈菌的一
类重要致病因子。因而,随着条锈菌全基因组测序及基
因功能分析技术的突破,加快了条锈菌等专性寄生真菌
功能基因的研究步伐。然而,小麦条锈菌遗传转化体系
仍然没有建立,仍需进一步探索。而条锈菌遗传转化体
系的建立势必会为锈菌基因功能的验证带来革命性的
变化,为揭示条锈菌致病机理插上腾飞的翅膀。 4.3 条锈菌效应蛋白组
效应蛋白(effector)作为一类非常重要的致病因
子,主要通过抑制寄主的防卫反应,引发寄主植物感
病;然而当病原菌效应蛋白被植物抗病蛋白直接或间
接识别时,可以激活植物免疫系统,植物表现为抗病。
因此,效应蛋白是植物病原菌致病的重要武器,也是
植物免疫系统的重要靶标。为此,效应蛋白也成为植
物病理学领域研究的前沿和热点。2011 年,Dong 等[98]
克隆了 3 个条锈菌吸器特异表达的分泌蛋白基因,它
们仅在锈菌中有同源基因,被认为是潜在的效应蛋白。
然而它们是否在条锈菌-小麦互作中起作用并没有研
究报道。Gu 等[99]克隆到条锈菌分泌蛋白基因 Ps87,其与卵菌和其他真菌分泌蛋白具有相似的转运结构域
RXLR,但是并未直接表现对寄主防卫反应的抑制作
用。因此,Ps87 可能是条锈菌一类新的效应蛋白,具有
进入寄主细胞的能力然而尚不能影响寄主的生理反应。 条锈菌基因组信息的释放大大推进了条锈菌分泌
蛋白组的研究。2012 年,Saunders 等[100]建立了从基
因组中综合分析预测锈菌效应蛋白的流程。该流程是
基于目前已知的病原真菌效应蛋白的特征,这些特征
包括(1)包含信号肽;(2)在病菌与寄主互作中诱
导表达;(3)吸器表达蛋白;(4)小分子蛋白并且
富含半胱氨酸;(5)包含已知的效应蛋白结构域或核
定位信号;(6)编码该蛋白的基因具有长的基因间隔
区;(7)包含内部重复序列;(8)不含 PFAM 预测
结构域或预测的结构域与致病性相关 [100]。运用
Markov 分层聚类方法对锈菌蛋白进行分类分级,确定
了 有可能是效应蛋白的 8 个家族作为候选锈菌效应
蛋白,为效应蛋白的研究奠定了基础[100]。2013 年,
Zheng 等在研究中国条锈菌小种 CYR32 时发现,锈菌

3448 中 国 农 业 科 学 48 卷
与玉米黑粉菌(Ustilago maydis)、小麦赤霉菌
(Fusarium graminearum)和双色蜡蘑菌(Laccaria bicolor)相比具有更为庞大的分泌组,包含 2 092 个
分泌蛋白,92%的分泌蛋白在真菌(除去锈菌)中没
有直系同源基因。同时,发现卵菌 RXLR 和白粉菌
Y/F/WxC 转运结构域在条锈菌基因组的分泌蛋白组
和非分泌蛋白中的比例没有明显差异,因而条锈菌
是否利用与卵菌等类似的转运结构域仍需要进一步
证明[87];Cantu 等[101]分析了 4 个条锈菌小种的效应蛋
白,发现在不同致病性小种中具有多态性的分泌蛋白,
并将它们作为后续效应蛋白功能研究的重点。另外,
他们还发现了两个不同的候选基因在一个 contig 上,
它们的 N 端序列具有较高的相似度,推测两个基因可
能来源于基因复制,表明条锈菌致病性因子也有可能
是由于基因复制所致[101]。在效应蛋白功能研究方面。
Yin 等[102-103]建立了利用细菌三型分泌系统将外源蛋
白导入小麦细胞分析效应蛋白功能的技术体系,加快
了条锈菌效应蛋白功能研究的步伐。
5 问题与展望
小麦条锈菌致病性变异,新致病性小种产生,是
引致小麦品种抗锈性“丧失”,从而造成病害大流行
的主要原因。如何降低条锈菌变异速率,延缓新小种
产生,持续有效控制条锈病将是未来锈病研究工作者
亟需解决的重大科学问题。通过几十年的努力攻关,
中国条锈病的综合防治已经取得了重大成就,走在了
国际的前列。然而小麦条锈菌致病性不断发生变异,
条锈病的防治仍旧是一个国际性的难题,条锈病的研
究任重而道远。 目前已证实小麦条锈菌可在其转主寄主植物小檗
上完成有性生殖,初步揭示了自然条件下小檗在条锈
菌致病性变异及病害流行中的作用。然而,自然界中
转主寄主小檗对条锈菌致病性变异、条锈菌群体结构
和病害流行的影响并不清楚。尽管中国条锈菌“越夏
区”和“冬繁区”小麦田块附近分布有大量的小檗,
但自然界中条锈菌致病性变异的方式主要是有性生殖
还是体细胞重组,铲除小檗能否阻止条锈菌致病性的
快速变异还需不断地去探索研究。此外,加强有性生
殖与条锈菌优势小种产生和预测的研究,对完善小麦
条锈菌生理小种的监测和预测预报将具有积极重要的
推动作用。 中国在利用抗病品种防治条锈病方面取得了一定
成效,然而抗病品种的利用还存在诸多问题。小麦生
产品种携带的抗病基因不清楚,许多品种携带同一抗
病基因,导致大规模单一化使用同一抗病基因,无法
真正实现在不同流行区合理布局抗病品种,条锈菌受
到单一抗病基因的高度选择压力,3—5 年便可产生新
致病性小种,而培育一个新的抗病品种一般需要 8—10 年甚至更长时间。因而,抗锈育种速度严重滞后于
病菌小种变异速度的形势依旧严峻。未来,如何在条
锈病不同流行区合理布局抗病基因将是利用抗病品种
防治条锈病的关键。 目前,中国 90%以上的小麦生产品种因条锈菌致
病性变异而“丧失”了其抗性,抗病资源日渐匮乏,
形势严峻,寻找新型抗病资源已迫在眉睫。2014 年,
Wang 等 [104]利用 CRISPR/Cas9 技术编辑小麦感白粉
病基因 TaMLO,创制了具有广谱抗性的小麦材料,为
新型持久广谱抗病材料的创制提供了思路。因而,在
科学研究快速发展的今天,研究者要抓住机遇,加快
对条锈菌致病性及其变异机理的研究,挖掘条锈菌关
键致病基因,为利用新技术新策略修饰和改造病菌或
寄主关键基因创制新型抗病材料奠定基础。 小麦条锈病综合防控是一项复杂的系统工程,面
临众多亟待解决的科学问题,必须依靠全国锈病工作
者长期不懈地协同攻关,结合传统方法与现代技术,
不断优化和完善小麦条锈病综合治理技术体系,才能
实现小麦条锈病的可持续控制。
References
[1] 李振岐, 商鸿生. 小麦锈病及其防治. 上海: 上海科学技术出版杜,
1989.
Li Z Q, Shang H S. Wheat Rust Disease and Its Control. Shanghai:
Shanghai Scientific and Technical Publishers, 1989. (in Chinese)
[2] Chen X M. Epidemiology and control of stripe rust [Puccinia
striiformis f. sp tritici] on wheat. Canadian Journal of Plant
Pathology, 2005, 27(3): 314-337.
[3] Wellings C R. Global status of stripe rust: a review of historical and
current threats. Euphytica, 2011, 179(1): 129-141.
[4] Ziyaev Z, Sharma R, Nazari K, Morgounov A, Amanov A,
Ziyadullaev Z, Khalikulov Z, Alikulov S. Improving wheat stripe rust
resistance in Central Asia and the caucasus. Euphytica, 2011, 179(1):
197-207.
[5] Morgounov A, Yessimbekova M, Rsaliev S, Baboev S, Mumindjanov
H, Djunusova M. High-yielding winter wheat varieties resistant to
yellow and leaf rust in Central Asia//Proceedings of 11th International
Cereal Rusts and Powdery Mildews Conference, Norwich UK, 2004.

17 期 康振生等:小麦条锈菌致病性及其变异研究进展 3449
[6] Wellings C, Mcintosh R. Puccinia striiformis f. sp. tritici in
Australasia: pathogenic changes during the first 10 years. Plant
Pathology, 1990, 39(2): 316-325.
[7] Wellings C, Kandel K. Pathogen dynamics associated with historic
stripe (yellow) rust epidemics in Australia in 2002 and 2003//
Proceedings of 11th International Cereal Rusts and Powdery Mildews
Conference, Norwich UK, 2004.
[8] Murray G, Ellison P, Watson A, Cullis B. The relationship between
wheat yield and stripe rust as affected by length of epidemic and
temperature at the grain development stage of crop growth. Plant
Pathology, 1994, 43(2): 397-405.
[9] Beresford R. Stripe rust (Puccinia striiformis), a new disease of wheat
in New Zealand. Cereal Rusts Bulletin, 1982, 10(2): 35-41.
[10] Pretorius Z. The impact of wheat stripe rust in South Africa//
Proceedings of 11th International Cereal Rusts and Powdery Mildews
Conference, Norwich UK, 2004.
[11] 李振岐, 曾士迈. 中国小麦条锈病. 北京: 中国农业出版社, 2002:
180-190.
Li Z Q, Zeng S M. Stripe Rust in Chinese Wheat. Beijing: China
Agriculture Press, 2002: 180-190. (in Chinese)
[12] 陈万权, 康振生, 马占鸿, 徐世昌, 金社林, 姜玉英. 中国小麦条
锈病综合治理理论与实践 . 中国农业科学 , 2013, 46(20):
4254-4262.
Chen W Q, Kang Z S, Ma Z H, Xu S C, Jin S L, Jiang Y Y. Integrated
management of wheat stripe rust caused by Puccinia striiformis f. sp.
tritici in China. Scientia Agricultura Sinica, 2013, 46(20): 4254-4262.
(in Chinese)
[13] 汪可宁, 洪锡午, 吴立人, 谢水仙, 孟庆玉, 陈善铭. 1951-1983 年
我国小麦品种抗条锈性变异分析. 植物保护学报, 1986, 13(2):
117-124.
Wang K N, Hong X W, Wu L R, Xie S X, Meng Q Y, Chen S M. The
analysis of the resistance of varieties in the wheat stripe rust nurseries
on 1951-1983. Acta Phytophylacica Sinica, 1986, 13(2): 117-124. (in
Chinese)
[14] 邝文静, 张贵胜, 章振羽, 姬红丽, 沈丽, 倪建英, 王宗华, 彭云良.
小麦条锈病菌 avrYr10/24/26/ch42 突变体的毒性研究. 西南农业学
报, 2013, 26(6): 2323-2331.
Kuang W J, Zhang G S, Zhang Z Y, Ji H L, Shen L, Ni J Y, Wang Z
H, Peng Y L. Studies on virulence of avrYr10 /24 /26 /ch42 mutants
of Puccinia striiformis f. sp. tritici. Southwest China Joumal of
Agricultural Sciences, 2013, 26(6): 2323-2331. (in Chinese)
[15] 贾秋珍, 黄瑾, 曹世勤, 张勃, 王晓明, 金社林. 感染我国重要小
麦抗源材料贵农 22 的条锈菌新菌系的发现及致病性初步分析. 甘
肃农业科技, 2012(1): 3-5.
Jia Q Z, Huang J, Cao S Q, Zhang B, Wang X M, Jin S L. Discovery
of new stripe rust strain infected to China’s major wheat resistant
material Guinong 22 and its preliminary pathogenicity analysis.
Gansu Agricultural Science and Technology, 2012(1): 3-5. (in
Chinese)
[16] 胡小平, 王保通, 康振生. 中国小麦条锈菌毒性变异研究进展. 麦
类作物学报, 2014, 34(5): 709-716.
Hu X P, Wang B T, Kang Z S. Research progress on virulence
variation of Puccinia striiformis f. sp. tritici in China. Journal of
Triticeae Crops, 2014, 34(5): 709-716. (in Chinese)
[17] Liu T G, Peng Y L, Chen W, Zhang Z. First detection of virulence in
Puccinia striiformis f. sp. tritici in China to resistance genes Yr24 (=
Yr26) present in wheat cultivar Chuanmai 42. Plant Disease, 2010,
94(9): 1163.
[18] 康振生, 李振岐. 洛夫林 10 常温致病新菌系的发现. 西北农林科
技大学学报: 自然科学版, 1984(3): 106.
Kang Z S, Li Z Q. Discovery of new strains of normal type pathogenic
wheat stripe rust to Lov-rin 10. Journal of Northwest A&F University:
Natural Science Edition, 1984(3): 106. (in Chinese)
[19] 康振生, 李振岐, 商鸿生, 陆和平, 刘学东. 绵阳系统小麦条锈菌
新菌系的发现. 西北农林科技大学学报: 自然科学版, 1994, 22(3):
7-11.
Kang Z S, Li Z Q, Shang H S, Lu H P, Liu X D. Discovery of new
isolates virulent to wheat cultivars of Mianyang system in wheat stripe
rust. Journal of Northwest A&F University: Natural Science Edition,
1994, 22(3): 7-11. (in Chinese)
[20] Purdy L, Allan R. A stripe rust race pathogenic to ‘Suwon 92’ wheat.
Plant Disease Report, 1966, 50: 205-207.
[21] Beaver R, Powelson R. A new race of stripe rust pathogenic on the
wheat variety Moro, CI 13740. Plant Disease, 1969, 53: 91-93.
[22] Bartos P, Fleischmann G, Samborski D, Shipton W. Studies on asexual
variation in the virulence of oat crown rust, Puccinia coronata f. sp.
avenae, and wheat leaf rust, Puccinia recondita. Canadian Journal of
Botany, 1969, 47(9): 1383-1387.
[23] Newton M, Johnson T, Brown A. A preliminary study on the
hybridization of physiologic forms of Puccinia graminis tritici.
Scientific Agriculture, 1930, 10(11): 721-731.
[24] Gassner G, Straib W. Über Mutationen in einer biologischen Rasse
von Puccinia glumarum tritici (Schmidt) Erikss. und Henn. Molecular
and General Genetics, 1933, 63(1): 154-180.
[25] Kajiwara T, Ueda I, Ivata Y. Susceptibility of wheat varieties to the
Japanese isolate of wheat yellow rust (Puccinia striiformis f. sp.

3450 中 国 农 业 科 学 48 卷
tritici). Bulletin of the National Institute of Agricultural Sciences
Series C, 1968, 22: 243-257.
[26] Stubbs R. Artificial mutation in the study of the relationship between
races of yellow rust of wheat//Proceedings of European and
Mediterranean Cereal Rusts Conference, Lisbon, 1968: 60-62.
[27] 商鸿生, 井金学, 李振岐. 紫外线诱导小麦条锈菌毒性突变的研究.
植物病理学报, 1994, 24(4): 347-351.
Shang H S, Jing J X, Li Z Q. Mutations induced by ultraviolent
radiation affecting virulence in Puccinia striiformis. Acta
Phytopathologica Sinica, 1994, 24(4): 347-351. (in Chinese)
[28] 王欣丽, 朱飞, 黄丽丽, 魏国荣, 康振生. 紫外线诱变对小麦条锈
菌致病性突变的影响. 核农学报, 2009, 23(3): 375-379.
Wang X L, Zhu F, Huang L L, Wei G R, Kang Z S. Effects of
ultraviolent radiation on pathogenicity mutation of Puccinia
striiformis f. sp. tritici. Journal of Nuclear Agricultural Sciences,
2009, 23(3): 375-379. (in Chinese)
[29] 黄丽丽, 王欣丽, 康振生, 赵杰. 紫外线诱导小麦条锈菌毒性突变
及突变体的 RAPD 分析. 菌物学报, 2005, 24(3): 400-406.
Huang L L, Wang X L, Kang Z S, Zhao J. Mutation of pathogenicity
induced by ultrviolet radiation in Puccinia striiformis f. sp. tritici and
RAPD analysis of mutants. Mycosystema, 2005, 24(3): 400-406. (in
Chinese)
[30] Teo C, Baker E. Mutagenic effects of ethyl methanesulphonate on the
oat stem rust pathogen (Puccinia graminis f. sp. avenae)//Proceedings
of the Linnean Society of New South Wales, 1975.
[31] Luig N. A high reversion rate for yellow urediospore color in Puccinia
graminis f. sp. tritici. Phytopathology, 1967, 57(10): 1091-1093.
[32] Statler G. Mutations to virulence and avirulence in Melampsora lini.
Phytopathology, 1985, 75(7): 771-773.
[33] 姚秋燕, 王国芬, 徐智斌, 王美南, 王阳, 井金学. EMS 诱导小麦
条锈菌毒性突变的研究. 西北农林科技大学学报: 自然科学版,
2006, 34(6): 120-123.
Yao Q Y, Wang G F, Xu Z B, Wang M N, Wang Y, Jing J X. Virulent
mutant in Puccinia striiformis induced by ethyl methyl sulfomar
(EMS). Journal of Northwest A&F University: Natural Science
Edition, 2006, 34(6): 120-123. (in Chinese)
[34] Mullins E, Kang S. Transformation: a tool for studying fungal
pathogens of plants. Cellular and Molecular Life Sciences, 2001,
58(14): 2043-2052.
[35] Schillberg S, Groß P, Tiburzy R. Isolation and characterization of the
EF-1α gene of the filamentous fungus Puccinia graminis f. sp. tritici.
Current Genetics, 1995, 27(4): 367-372.
[36] Webb C A, Szabo L J, Bakkeren G, Garry C, Staples R C, Eversmeyer
M, Fellers J P. Transient expression and insertional mutagenesis of
Puccinia triticina using biolistics. Functional & Integrative Genomics,
2006, 6(3): 250-260.
[37] Bhairi S, Staples R. Transient expression of the ß-glucuronidase gene
introduced into Uromyces appendiculatus uredospores by particle
bombardment. Phytopathology, 1992, 82(9): 986-989.
[38] 张如佳, 王阳, 王美南, 井金学, 李振岐. 基因枪介导小麦条锈菌
毒性突变的研究. 西北农林科技大学学报: 自然科学版, 2007,
35(5): 87-91.
Zhang R J, Wang Y, Wang M N, Jing J X, Li Z Q. Mutations induced
by biolistics affecting virulence in Puccinia striiformis. Journal of
Northwest A&F University: Natural Science Edition, 2007, 35(5):
87-91. (in Chinese)
[39] 王阳, 马东方, 张亮, 卢丽丽, 王美南, 井金学, 康振生. GUS 基因
插入导致的小麦条锈菌突变体遗传稳定性和毒性变异研究. 植物
病理学报, 2013, 43(1): 42-49.
Wang Y, Ma D F, Zhang L, Lu L L, Wang M N, Jing J X, Kang Z S.
Hereditary stability and pathogenicity of wheat stripe rust mutants
with GUS gene insertion. Acta Phytopathologica Sinica, 2013, 43(1):
42-49. (in Chinese)
[40] Hansen H, Smith R E. The mechanism of variation in imperfect fungi:
Botrytis cinerea. Phytopathology, 1932, 22(12): 953-964.
[41] Nelson R, Christensen W R. Heterokaryosis as a basis for variation in
Puccinia grαminis f. sp. tritici. Phytopathology, 1955, 45: 639-643.
[42] Wright R, Lennard J. Origin of a new race of Puccinia striiformis.
Transactions of the British Mycological Society, 1980, 74(2): 283-287.
[43] Little R, Manners J. Somatic recombination in yellow rust of wheat
(Puccinia striiformis): The production and possible origin of two new
physiologic races. Transactions of the British Mycological Society,
1969, 53(2): 251-258.
[44] Volin R B. Physiological race determination and environmental
factors affecting the development of infection type in stripe rust
(Puccinia striiformis West.)[D]. Bozemanpp: Montana State University,
1971.
[45] Taylor E. The production and behaviour of somatic recombinants in
Puccinia striiformis//Proceedings of European and Mediterranean
Cereal Rusts Conference, 1976.
[46] Goddard M V. The production of a new race, 105E137 of Puccinia
striiformis in glasshouse experiments. Transactions of the British
Mycological Society, 1976, 67(3): 395-398.
[47] 赵来顺, 王焕如. 小麦条锈菌夏孢子芽管结合现象的初步观察. 中
国农业科学, 1964(1): 48-49.
Zhao L S, Wang H R. Preliminary observation of the fussion of

17 期 康振生等:小麦条锈菌致病性及其变异研究进展 3451
uredinospore germ tubes in Puccinia striiformis. Scientia Agricultura
Sinica, 1964(1): 48-49. (in Chinese)
[48] 康振生, 李振岐, 商鸿生. 小麦条锈菌异核新菌系的筛选及核游离
试验研究. 植物病理学报, 1994, 24(2): 101-105.
Kang Z S, Li Z Q, Shang H S. On the screening of new heterocaryons
of wheat stripe rust and its nucleal dissociation. Acta Phytopathologica
Sinica, 1994, 24(2): 101-105. (in Chinese)
[49] 康振生, 李振岐, 商鸿生. 小麦条锈菌异核作用产生的一新菌系.
西北农林科技大学学报: 自然科学版, 1993, 21(1): 97-99.
Kang Z S, Li Z Q, Shang H S. A new isolate produced by the
heterokaryosis of wheat stripe rust. Journal of Northwest A&F
University: Natural Science Edition, 1993, 21(1): 97-99. (in Chinese)
[50] 康振生, 李振岐, 商鸿生. 小麦条锈菌夏孢子阶段核相状况的研究.
植物病理学报, 1994, 24(1): 26-31.
Kang Z S, Li Z Q, Shang H S. Nucleal condition of uredinial stage of
wheat stripe rust. Acta Phytopathologica Sinica, 1994, 24(1): 26-31.
(in Chinese)
[51] 王阳, 康振生, 李振岐. 天水地区高山、半山和川区小麦条锈菌夏
孢子多核率研究. 麦类作物学报, 2004, 24(2): 110-113.
Wang Y, Kang Z S, Li Z Q. Study on heterokaryon distribution of
Puccinia striiformis f. sp. tritici West. in the different mountain areas
of Tianshui. Journal of Triticeae Crops, 2004, 24(2): 110-113. (in
Chinese)
[52] 马青, 李振岐, 商鸿生. 小麦条锈菌夏孢子芽管结合现象的研究.
西北农林科技大学学报: 自然科学版, 1991, 19(增刊): 49-53.
Ma Q, Li Z Q, Shang H S. A study on the fusion of urediospore germ
tubes in Puccinia striiformis f. sp. tritici West. Journal of Northwest
A&F University: Natural Science Edition, 1991, 19(Suppl.): 49-53. (in
Chinese)
[53] Tranzschel W. Promezutocnye chozjaeva rzavciny chlebov i ich der
UdSSR (The alternate hosts of cereal rust fungi and their distribution
in the UdSSR). Bulletin of Plant Protection Series, 1934, 2: 4-10.
[54] Straib W. Untersuchungen über das Vorkommen physiologischer
Rassen des Gelbrostes (Puccinia glumarum) in den Jahren 1935-1936
und über die Agressivitt einiger neur Formen auf Getreide und
Grasern. Arb. Biology Reichsanst. LandForstwirtsch. Berlin-Dahlem,
1937, 22: 91-119.
[55] Mains E B. Studies concerning heteroecious rusts. Mycologia, 1933,
25(5): 407-417.
[56] Hart H, Becker H. Beiträge zur Frage des Zwischenwirtes für
Puccinia glumarum. Zeits. Pflanzenk, 1939, 49: 559-566.
[57] Jin Y, Szabo L J, Carson M. Century-old mystery of Puccinia
striiformis life history solved with the identification of Berberis as an
alternate host. Phytopathology, 2010, 100(5): 432-435.
[58] Wang M, Chen X. First report of Oregon grape (Mahonia aquifolium)
as an alternate host for the wheat stripe rust pathogen (Puccinia
striiformis f. sp. tritici) under artificial inoculation. Plant Disease,
2013, 97(6): 839.
[59] 赵杰, 张宏昌, 姚娟妮, 黄丽丽, 康振生. 中国小麦条锈菌转主寄
主小檗的鉴定. 菌物学报, 2011, 30(6): 895-900.
Zhao J, Zhang H C, Yao J N, Huang L L, Kang Z S. Confirmationof
Berberis spp. as alternate hosts of Puccinia striiformis f. sp. tritici on
wheat in China. Mycosystema, 2011, 30(6): 895-900. (in Chinese)
[60] Zhao J, Wang L, Wang Z Y, Chen X M, Zhang H C, Yao J N, Zhan G
M, Chen W, Huang L L, Kang Z S. Identification of eighteen Berberis
species as alternate hosts of Puccinia striiformis f. sp. tritici and
virulence variation in the pathogen isolates from natural infection of
barberry plants in China. Phytopathology, 2013, 103(9): 927-934.
[61] Stakman E, Levine N, Leach J. New biologic forms of Puccinia
graminis (preliminary paper). Journal of Agricultural Research, 1919,
16: 103-105.
[62] Jin Y. Role of Berberis spp. as alternate hosts in generating new races
of Puccinia graminis and P. striiformis. Euphytica, 2011, 179(1):
105-108.
[63] Craigie J. Discovery of the function of the pycnia of the rust fungi.
Nature, 1927, 120: 765-767.
[64] Wang M, Wan A, Chen X. Genetic characterization of virulence/
avirulence genes of Puccinia striiformis f. sp. tritici. Phytopathology,
2012, 102: S4.
[65] Waterhouse W L. A preliminary account of the origin of two new
Australian physiologic forms of Puccinia graminis tritici. Proceedings
of the Linnean Society of New South Wales, 1929, 54: 96-106.
[66] 陆宁海, 詹刚明, 王建锋, 黄丽丽, 康振生. 我国小麦条锈菌体细
胞遗传重组的分子证据. 植物病理学报, 2009, 39(6): 561-568.
Lu N H, Zhan G M, Wang J F, Huang L L, Kang Z S. Molecular
evidence of somatic genetic recombination of Puccinia striiformis f.
sp. tritici in China. Acta Phytopathologica Sinica, 2009, 39(6):
561-568. (in Chinese)
[67] Brown J K, Hovmøller M S. Aerial dispersal of pathogens on the
global and continental scales and its impact on plant disease. Science,
2002, 297(5581): 537-541.
[68] Cheng P, Chen X. Virulence and molecular analyses support asexual
reproduction of Puccinia striiformis f. sp. tritici in the US Pacific
Northwest. Phytopathology, 2014, 104(11): 1208-1220.
[69] Duan X, Tellier A, Wan A, Leconte M, De Vallavieille-Pope C,
Enjalbert J. Puccinia striiformis f. sp. tritici presents high diversity

3452 中 国 农 业 科 学 48 卷
and recombination in the over-summering zone of Gansu, China.
Mycologia, 2010, 102(1): 44-53.
[70] Steele K A, Humphreys E, Wellings C, Dickinson M. Support for a
stepwise mutation model for pathogen evolution in Australasian
Puccinia striiformis f. sp. tritici by use of molecular markers. Plant
Pathology, 2001, 50(2): 174-180.
[71] Mboup M, Leconte M, Gautier A, Wan A, Chen W, De
Vallavieille-Pope C, Enjalbert J. Evidence of genetic recombination in
wheat yellow rust populations of a Chinese oversummering area.
Fungal Genetics and Biology, 2009, 46(4): 299-307.
[72] Hovmøller M, Justesen A, Brown J. Clonality and long-distance
migration of Puccinia striiformis f. sp. tritici in northwest Europe.
Plant Pathology, 2002, 51(1): 24-32.
[73] Liu M, Hambleton S. Taxonomic study of stripe rust, Puccinia
striiformis sensu lato, based on molecular and morphological evidence.
Fungal Biology, 2010, 114(10): 881-899.
[74] Ali S, Gladieux P, Leconte M, Gautier A, Justesen A F, Hovmoller M
S, Enjalbert J, De Vallavieille-Pope C. Origin, migration routes and
worldwide population genetic structure of the wheat yellow rust
Puccinia striiformis f. sp. tritici. Plos Pathogens, 2014, 10(1):
e1003903.
[75] Enjalbert J, Duan X, Leconte M, Hovmøller M S, De
Vallavieille-Pope C. Genetic evidence of local adaptation of wheat
yellow rust (Puccinia striiformis f. sp. tritici) within France.
Molecular Ecology, 2005, 14(7): 2065-2073.
[76] Wellings C. Puccinia striiformis in Australia: a review of the incursion,
evolution, and adaptation of stripe rust in the period 1979-2006. Crop
and Pasture Science, 2007, 58(6): 567-575.
[77] Chen X, Line R F, Leung H. Relationship between virulence variation
and DNA polymorphism in Puccinia striiformis. Phytopathology,
1993, 83(12): 1489-1497.
[78] Bahri B, Shah S, Hussain S, Leconte M, Enjalbert J, De
Vallavieille-Pope C. Genetic diversity of the wheat yellow rust
population in Pakistan and its relationship with host resistance. Plant
Pathology, 2011, 60(4): 649-660.
[79] Lu N, Wang J, Chen X, Zhan G, Chen C, Huang L, Kang Z. Spatial
genetic diversity and interregional spread of Puccinia striiformis f. sp.
tritici in Northwest China. European Journal of Plant Pathology,
2011, 131(4): 685-693.
[80] Shan W X, Chen S Y, Kang Z S, Wu L R, Li Z Q. Genetic diversity in
Puccinia striiformis Westend. f. sp. tritici revealed by pathogen
genome-specific repetitive sequence. Canadian Journal of Botany,
1998, 76(4): 587-595.
[81] 陈长卿. 中国小麦条锈菌分子群体遗传结构研究[D]. 杨凌: 西北
农林科技大学, 2008.
Chen C Q. Molecular population genetic structure of Puccinia
striiformis f. sp. tritici in China[D]. Yangling: Northwest Agriculture
& Forestry University, 2008. (in Chinese)
[82] 郑文明, 刘峰, 康振生, 陈受宜, 李振岐, 吴立人. 中国小麦条锈
菌主要流行菌系的 AFLP 指纹分析. 自然科学进展, 2000, 10(6):
532-537.
Zheng W M, Liu F, Kang Z S, Chen S Y, Li Z Q, Wu L R. AFLP
analyses of the fingerprinting of the major epidemic strains of
Puccinia striiformis. Progress in Natural Science, 2000, 10(6):
532-537. (in Chinese)
[83] Liu X, Huang C, Sun Z, Liang J, Luo Y, Ma Z. Analysis of population
structure of Puccinia striiformis in Yunnan Province of China by using
AFLP. European Journal of Plant Pathology, 2011, 129(1): 43-55.
[84] 陆宁海, 王建锋, 詹刚明, 黄丽丽, 康振生. 陇南地区不同海拔区
域内小麦条锈菌群体遗传多样性的分析. 菌物学报, 2009, 28(4):
496-503.
Lu N H, Wang J F, Zhan G M, Huang L L, Kang Z S. SSR analysis of
population genetic diversity of Puccinia striifornis f. sp. tritici in
different altitude regions of Longnan, Gansu. Mycosystema, 2009,
28(4): 496-503. (in Chinese)
[85] 王建锋, 陈长卿, 陆宁海, 彭云良, 詹刚明, 黄丽丽, 康振生. 四川
省小麦条锈菌群体遗传多样性的 SSR分析. 菌物学报, 2010, 29(2):
206-213.
Wang J F, Chen C Q, Lu N H, Peng Y L, Zhan G M, Huang L L, Kang
Z S. SSR analysis of population genetic diversity of Puccinia
striiformis f. sp. tritici in Sichuan Province, China. Mycosystema,
2010, 29(2): 206-213. (in Chinese)
[86] Cantu D, Govindarajulu M, Kozik A, Wang M N, Chen X M, Kojima
K K, Jurka J, Michelmore R W, Dubcovsky J. Next generation
sequencing provides rapid access to the genome of Puccinia
striiformis f. sp tritici, the causal agent of wheat stripe rust. Plos One,
2011, 6(8): e24230.
[87] Zheng W M, Huang L L, Huang J Q, Wang X J, Chen X M, Zhao J,
Guo J, Zhuang H, Qiu C Z, Liu J, Liu H Q, Huang X L, Pei G L, Zhan
G M, Tang C L, Cheng Y L, Liu M, Zhang J S, Zhao Z T, Zhang S J,
Han Q M, Han D J, Zhang H C, Zhao J, Gao X N, Wang J F, Ni P X,
Dong W, Yang L F, Yang H M, Xu J R, Zhang G Y, Kang Z S. High
genome heterozygosity and endemic genetic recombination in the
wheat stripe rust fungus. Nature Communications, 2013, 4: Article
number 2673.
[88] Ling P, Wang M N, Chen X M, Campbell K G. Construction and

17 期 康振生等:小麦条锈菌致病性及其变异研究进展 3453
characterization of a full-length cDNA library for the wheat stripe rust
pathogen (Puccinia striiformis f. sp. tritici). BMC Genomics, 2007, 8:
145.
[89] Zhang Y H, Qu Z P, Zheng W M, Liu B, Wang X J, Xue X D, Xu L S,
Huang L L, Han Q M, Zhao J, Kang Z S. Stage-specific gene
expression during urediniospore germination in Puccinia striiformis f.
sp. tritici. BMC Genomics, 2008, 9: 203.
[90] Yin C T, Chen X M, Wang X J, Han Q M, Kang Z S, Hulbert S H.
Generation and analysis of expression sequence tags from haustoria of
the wheat stripe rust fungus Puccinia striiformis f. sp. tritici. BMC
Genomics, 2009, 10: 626.
[91] Garnica D P, Upadhyaya N M, Dodds P N, Rathjen J P. Strategies for
wheat stripe rust pathogenicity identified by transcriptome sequencing.
Plos One, 2013, 8(6): e67150.
[92] 梁晓飞, 刘博, 朱琳, 张晓羽, 王晓杰, 黄丽丽, 康振生. 小麦条锈
菌Ⅱ类几丁质合成酶基因 PstChsⅡ的克隆及表达特征. 微生物学
报, 2009, 49(12): 1621-1627.
Liang X F, Liu B, Zhu L, Zhang X Y, Wang X J, Huang L L, Kang Z
S. Cloning and expression of a class chitin synthase gene PstChs Ⅱ
from the rustⅡ fungus Puccinia striiformis. Acta Microbiologica
Sinica, 2009, 49(12): 1621-1627. (in Chinese)
[93] Guo J, Dai X W, Xu J R, Wang Y L, Bai P F, Liu F R, Duan Y H,
Zhang H, Huang L L, Kang Z S. Molecular characterization of a
Fus3/Kss1 type MAPK from Puccinia striiformis f. sp. tritici,
PsMAPK1. Plos One, 2011, 6(7): e21895.
[94] Guo J, Duan Y H, Zhang J S, Shi X X, Chen Y Y, Zhang H, Huang L
L, Kang Z S. A conidiation-related gene is highly expressed at the
resting urediospore stage in Puccinia striiformis f. sp. tritici. Journal
of Basic Microbiology, 2013, 53(8): 695-702.
[95] Yin C T, Jurgenson J E, Hulbert S H. Development of a host-induced
RNAi system in the wheat stripe rust fungus Puccinia striiformis f. sp.
tritici. Molecular Plant-Microbe Interactions, 2011, 24(5): 554-561.
[96] Zhang H, Guo J, Voegele R T, Zhang J S, Duan Y H, Luo H Y, Kang Z
S. Functional characterization of calcineurin homologs PsCNA1/
PsCNB1 in Puccinia striiformis f. sp. tritici using a host-induced
RNAi system. Plos One, 2012, 7(11): e49262.
[97] Cheng Y L, Wang X J, Yao J N, Voegele R T, Zhang Y R, Wang W M,
Huang L L, Kang Z S. Characterization of protein kinase PsSRPKL, a
novel pathogenicity factor in the wheat stripe rust fungus.
Environmental Microbiology, 2014, doi: 10.1111/1462-2920.12719.
[98] Dong Y L, Yin C T, Hulbert S, Chen X M, Kang Z S. Cloning and
expression analysis of three secreted protein genes from wheat stripe
rust fungus Puccinia striiformis f. sp. tritici. World Journal of
Microbiology & Biotechnology, 2011, 27(5): 1261-1265.
[99] Gu B A, Kale S D, Wang Q H, Wang D H, Pan Q N, Cao H, Meng Y L,
Kang Z S, Tyler B M, Shan W X. Rust secreted protein Ps87 is
conserved in diverse fungal pathogens and contains a RXLR-like
motif sufficient for translocation into plant cells. Plos One, 2011,
6(11): e27217.
[100] Saunders D G, Win J, Cano L M, Szabo L J, Kamoun S, Raffaele S.
Using hierarchical clustering of secreted protein families to classify
and rank candidate effectors of rust fungi. Plos One, 2012, 7(1):
e29847.
[101] Cantu D, Segovia V, Maclean D, Bayles R, Chen X M, Kamoun S,
Dubcovsky J, Saunders D G O, Uauy C. Genome analyses of the
wheat yellow (stripe) rust pathogen Puccinia striiformis f. sp tritici
reveal polymorphic and haustorial expressed secreted proteins as
candidate effectors. BMC Genomics, 2013, 14: 270.
[102] Yin C T, Hulbert S. Prospects for functional analysis of effectors from
cereal rust fungi. Euphytica, 2011, 179(1): 57-67.
[103] Upadhyaya N M, Mago R, Staskawicz B J, Ayliffe M A, Ellis J G,
Dodds P N. A bacterial type III secretion assay for delivery of fungal
effector proteins into wheat. Molecular Plant-Microbe Interactions,
2014, 27(3): 255-264.
[104] Wang Y, Cheng X, Shan Q, Zhang Y, Liu J, Gao C, Qiu J L.
Simultaneous editing of three homoeoalleles in hexaploid bread wheat
confers heritable resistance to powdery mildew. Nature Biotechnology,
2014, 32(9): 947-951.
(责任编辑 岳梅)

中国农业科学 2015,48(17):3454-3462 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.012
收稿日期:2015-01-05;接受日期:2015-04-03 基金项目:国家“973”计划(2010CB126205) 联系方式:张传溪,Tel:0571-88982991;E-mail:[email protected]
中国农业昆虫基因组学研究概况与展望
张传溪
(浙江大学昆虫科学研究所,杭州 310058)
摘要:昆虫种类繁多,变异惊人,是地球上最大的动物类群,其中不少种类是农业上的重要害虫或益虫。自
从 2000 年黑腹果蝇(Drosophila melanogaster)基因组发表以来,当代的昆虫学研究已经越来越多地依赖于从
基因组和转录组所获得的信息。为了更好地理解昆虫生物学特性和增强应对那些严重影响人类健康、粮食供应和
经济安全的各种卫生、农林害虫的能力,更好地利用那些在维持自然和农业生态系统不可或缺的昆虫以及那些为
人类提供丝、蜂蜜和药品的昆虫,需要了解它们的基因组和转录组信息。目前世界上已经有 50 多种昆虫的基因组
被报道,近 10 年来中国科学家也已经完成了家蚕(Bombyx mori)、小菜蛾(Plutella xyllostella)、东亚飞蝗
(Locusta migratoria manilensis)、榕小蜂(Ceratosolen solmsi)和褐飞虱(Nilaparvata lugens)的基因
组测定和分析,还有一批重要农业昆虫的基因组正在分析中,这其中包括烟粉虱(Bemisia tabaci)、斜纹夜蛾
(Spodoptera litura)、二化螟(Chilo suppressalis)、白背飞虱(Sogatella furcifera)、灰飞虱(Laodelphax
striatellu)和若干寄生蜂种类。自从 2010 年中国率先在国际上报道了 2种昆虫(烟粉虱和褐飞虱)的转录组和
表达谱以来,短短几年仅国内就已经有上百个昆虫转录组得到分析。同时,家蚕、东亚飞蝗和褐飞虱的功能基因
组研究也取得了很大的进展, 例如,用 40 个品系重测序揭示了家蚕在驯化过程中相关的基因变化,通过信号通路
分析解析了东亚飞蝗散居型和群居型转换的分子机制,以及发现了 2 个胰岛素受体是控制稻飞虱长、短翅型可塑
性发育的“分子开关”。在基因组和转录组基础上,通过基因功能的研究和挖掘,结合应用新的基因组定点编辑技
术和 RNAi 技术,将使害虫的防治和益虫的利用出现革命性的变化。
关键词:农业昆虫;基因组;功能基因组学;展望
Current Research Status and Prospects of Genomes of
Insects Important to Agriculture in China ZHANG Chuan-xi
(Institute of Insect Science, Zhejiang University, Hangzhou 310058)
Abstract: Insects are the largest class of all living things with overwhelming diversity in the earth. Many of them are of agricultural importance: serious crop pests or beneficial insects. Modern entomological researches are more and more relying on information obtained from different insect genomes and transcriptomes since Drosophila melanogaster genome being sequenced in 2000. To better understand insect biology and transform our ability to manage insects that threaten our health, food supply, and economic security and to use beneficial insects that are essential to the maintenance and productivity of natural and agricultural ecosystems or provide us with silk, honey, medicine and other insect products, we need to know their genomic and transcriptomic information. Up to date, genomes of more than 50 insect species have been sequenced and analyzed around the world, and genomes of five insects of agricultural importance, including the domestic silkworm (Bombyx mori), the diamondback moth (Plutella xylostella), the oriental migratory locust (Locusta migratoria manilensis), the fig wasp (Ceratosolen solmsi) and the brown planthopper (Nilaparvata lugens), have been analyzed during the past decade in China, and genome sequencing for several
jia
文本框
返回目录

17 期 张传溪:中国农业昆虫基因组学研究概况与展望 3455
agricultural insects including the whitefly (Bemisia tabaci), the oriental leafworm moth (Spodoptera litura), the rice stem borer (Chilo suppressalis), the white backed planthopper (Sogatella furcifera), the small brown planthopper (Laodelphax striatellu) and several parasite wasps, are in progress. Transcriptomes of the whitefly and brown planthopper were reported in China in 2010, first two insect transciptomes reported in the world. Hundreds of insect transcriptomes have been reported in China since that year. Many important progresses in the functional genomics of the silkworm, the locust and the brown planthopper have been achieved, including the resequencing of 40 varieties of silkworm genomes which revealed domestication events and genes in silkworm, the uncovering of the precise mechanism of phase changes of the migratory locust, and the finding of that two insulin receptors determine alternative wing morphs in planthoppers. Data mining of insect genomes and transcriptomes, together with newly developed targeted genome editing and RNAi technologies, will lead to a revolutionary change in insect pest control and beneficial insect utilization.
Key words: agricultural insects; genome; functional genomics; prospect 昆虫种类繁多,已知约 180 万种,占动物种类的
3/4 以上,是地球上数量 多的动物群体,其踪迹几乎
遍布世界的每一个角落。除种类繁多外,昆虫生物量
也十分惊人。昆虫与人类生活息息相关,许多种类是
农牧业的害虫和益虫(农林害虫、卫生害虫,生态平
衡因子、传播花粉、工业原料、中药宝库、美味食品、
清洁环境、工艺观赏、科研材料、基因宝库、生物反
应器等)。例如,75%以上的显花植物依赖于昆虫为
其授粉,如果没有昆虫,自然生态系统和农业生态系
统就会崩溃。又如,虽然经过人们的努力防治,还是
有 20%以上的农林牧产品因昆虫危害而损失。昆虫严
重影响人类的另一重要方面是虫传疾病,因此而死亡
的人数众多。长期以来,人们对重要卫生害虫和农业
昆虫的发生规律、防治方法或利用途径进行了多维度
的研究,取得了一系列重要成果。2000 年,果蝇基因
组完成[1],使昆虫学研究进入了一个新时代,为昆虫
科学研究提供了新机遇,对全面了解昆虫的遗传、进
化、系统发育、生长、发育、变态和繁殖等各方面信
息起到了不可替代的作用,并对研究者利用昆虫的遗
传信息在分子水平上改造和应用昆虫提供了 基本的
依据。
1 不同昆虫基因组研究意义
昆虫不同种类生物学特性差异巨大,在基因组组
成上表现巨大多样性。仅依赖模式生物果蝇的基因组
信息,无法解释和深入研究不同类群昆虫生物学特性
的巨大差异,很多情况下也难以仅利用果蝇基因组信
息进行不同昆虫特异、重要生物学特性的分子机理研
究。随着昆虫科学从传统观测分析进入分子机理水平
研究,迫切需要不同类群代表性昆虫的基因组信息。
笔者将以水稻 主要害虫褐飞虱为例说明重要基因组
信息的重要性。首先,褐飞虱不耐低温,不能在我国
绝大部分稻区越冬,却通过迁飞连续频繁暴发危害近
半个世纪。为了实现其源头控制,迫切需要了解每年
的虫源地及其迁飞的具体路径,遗憾的是目前我们还
只能主要靠大气环流推测,缺乏确切的遗传分子标记
检测。而有效的分子标记开发,就需要了解褐飞虱不
同地理种群的基因组差异。另一方面,褐飞虱可以在
不同环境条件下产生长翅型和短翅型,长翅型适合于
长距离迁飞和扩散以逃避不良环境和发现新的适合资
源,而短翅型由于发育历期短产卵量大而适于大量快
速繁殖,在褐飞虱迁入新稻区后可以快速扩大种群,
因此翅二型可变发育是稻飞虱成灾的重要机制。尽管
数十年来很多昆虫研究者进行了大量研究,确定激素
和很多环境因子会影响翅型分化,但对昆虫翅型控制
的分子“开关”这个谜一直未能解开,其信号通路解
析也需要基因组和转录组信息。其次,褐飞虱适应水
稻品种抗性能力很强(所谓“致害性变异”),一个
抗褐飞虱的水稻品种推广后,短则一年,多者三载,
褐飞虱就能克服其抗性。这对抗性育种提出了极大的
挑战。为了为抗性育种提供指导,研究者需要了解褐
飞虱致害性变异的分子机理。褐飞虱致害性变异是种
群遗传结构变异吗?还是表观遗传问题?或是共生微
生物在作祟?这无不要求有其基因组信息。再次,褐
飞虱寄主高度专一性,只取食水稻汁液,但已有研究
表明水稻汁液营养成分组成是十分不平衡的。像水稻
的韧皮部汁液中含有大量的蔗糖,却缺少必需氨基酸
以及只含有少量的氮素化合物,这对于单食性的褐飞
虱来说是远远不够的。那在如此营养不平衡的食物来
源下,共生微生物是如何与其“狼狈为奸”,让褐飞
虱繁殖如此之快以致大暴发?这也需要褐飞虱和共生
菌基因组的代谢通路信息加以解释,并通过发现共生
体系的薄弱环节为通过“抑菌治虫”提供潜在途径。
此外,褐飞虱繁殖能力很强,有些农药能刺激生殖,
对多种农药适应性很快,抗药性很强,还传播多种水
稻病毒病。这些与其频频成灾相关的重要特性的机理

3456 中 国 农 业 科 学 48 卷
都还不很清楚,其深入研究也迫切需要基因组信息支
撑。褐飞虱在分类上属于不完全变态的半翅目昆虫,
口器为刺吸式,其生活周期短,有系统的 RNAi 现象,
其基因组解析可使之成为半翅目昆虫乃至不完全变态
昆虫的一个研究模式。
2 昆虫基因组学研究概况
昆虫全基因组测序研究,大致可以分为 3 个阶段:
第 1 阶段是开拓阶段,黑腹果蝇[1]的全基因组测序,
是一个里程碑,开创了包括昆虫在内所有动物基因组
研究的新纪元;第 2 阶段是 2002—2009 年零星阶段,
全基因组测序集中在完全变态昆虫的几个主要目,包
括双翅目的冈比亚按蚊(Anopheles gambiae,2002年)[2]和埃及伊蚊(Aedes aegypti,2007 年)[3]以及
12 种果蝇(2007 年)[4]、鳞翅目家蚕(Bombyx mori,2004 年)[5]、膜翅目意大利蜜蜂(Apis mellifera,2006年)[6]和赤拟谷盗(Tribolium castaneum,2008 年)[7],
此阶段测序成本很高,只有少量研究机构的零星昆虫
基因组报道;第 3 阶段是 2010 年后的暴发阶段,随着
二代测序技术发展,成本下降,更多数量和来自更多
不同分类地位的昆虫基因组被不同研究机构解析,至
2014 年底膜翅目昆虫有 3 种丽蝇蛹集金小蜂(Nasonia spp.,2010 年) [8]、佛罗里达弓背蚁(Camponotus floridanus)和印度跳蚁(Harpegnathos saltator,2010年)[9]、顶切叶蚁(Acromyrmex echinatior,2011 年)[10]、
大头切叶蚁(Atta cephalotes,2011 年)[11]、阿根廷蚁
( Linepithema humile, 2011 年) [12] 、红收割蚁
( Pogonomyrmex barbatus , 2011 年) [13] 、火蚁
(Solenopsis invicta,2011 年)[14]等 7 种蚂蚁,叶榕
小蜂( Ceratosolen solmsi , 2013 年) [15] 、隧蜂
(Lasioglossum albipes,2013 年)[16]; 双翅目有致倦
库蚊(Culex quinquefasciatus,2010 年)[17]、达氏按
蚊(Anopheles darlingi,2013 年)[18]、中国按蚊(A. sinensis,2014 年)[19]、南极摇蚊(Belgica antarctica,2014 年)[20]、范氏摇蚊(Polypedilum vanderplanki,2014 年)和云足多集摇蚊(P. nubifer,2014 年)[21]、
采采蝇(Glossina morsitans,2014 年)[22];鳞翅目有
帝王蝶(Danaus plexippus,2011 年)[23]、红带袖蝶
(Heliconius melpomene,2013 年)[24]、小菜蛾(Plutella xylostella,2013 年)[25]。鞘翅目增加了 1 种,即中欧
山松大小蠹(Dendroctonus ponderosae,2013 年)[26];
还有半翅目的豌豆蚜(Acyrthosiphon pisum,2010 年)[27]
和褐飞虱(Nilaparvata lugens,2014 年)[28];直翅
目的东亚飞蝗(Locusta migratoria manilensis,2014年)[29];虱目的人体虱(Pediculus humanus humanus,2011 年) [30];等翅目的湿木白蚁(Zootermopsis nevadensis nuttingi,2014 年) [31]和纳塔尔大白蚁
(Macrotermes natalensis,2014 年)[32];也包括了一
些小目昆虫,如捻翅目 1 种捻翅虫(Mengenilla moldrzyki,2012 年)[33];樇目 1 种竹节虫(Timema cristinae,2014 年)[34]。已经发表的昆虫基因组情况
见表 1。此外,还有多种昆虫基因组测序已经完成,
序列数据已经部分或全部公开,但文章尚未发表,如
吸血蝽(Rhodnius prolixus)、东美洲熊蜂(Bombus impatiens)、欧洲熊蜂(B. terrestris)和烟草天蛾
(Menduca sexta)等数十种昆虫,还有更多的昆虫基
因组在测序组装或进一步完善中。目前,已经报道基
因组分析的昆虫中,一些基因组较小、杂合率低、容
易组装、测序成本低的昆虫,如膜翅目和双翅目昆虫
占了 大的比例,但也包括了许多有重要经济意义的
昆虫,甚至包括基因组很大的昆虫如基因组 6G 的东
亚飞蝗,此外研究者对在进化上有特殊意义的一些小
目昆虫也开始关注。2011 年由美国伊利诺香宾大学昆
虫学家 Gene Robinson 领衔的多位国际著名昆虫学家
联合提出雄心勃勃的 5 000 种昆虫全基因组测序计划
(i5K,http://www.arthropodgenomes.org/wiki/i5K)。
随着测序成本的进一步降低,会有更多的实验室加入
到对不同类群昆虫的基因组分析,而先进的实验室会
更多地关注重要昆虫的功能基因组研究。
3 昆虫的基因组大小和基因数量
昆虫基因组大小差异显著[35],目前已经测定的 50多种昆虫基因组大小约在 89—6 500 Mb,其中基因组
大的东亚飞蝗,达 6 525 Mb,基因组 小的南极摇
蚊只有 89.6 Mb。昆虫基因组大小不同,主要由基因
组重复序列数量、基因间隔区长度和平均内含子大小
决定。比如,同属于半翅目的豌豆蚜和褐飞虱,豌豆
蚜组装的基因组大小只有 464 Mb,而褐飞虱为 1 141 Mb,比较两者的基因组发现,重复序列即称为转座元
件(transposable elements)的可移动 DNA 片段所占比
例是昆虫种类之间基因组大小不同的 主要原因,如
褐飞虱基因组重复序列有 554.4 Mb(占基因组
48.6%),而豌豆蚜只有 143.8 Mb(占基因组 33.3%);
此外,内含子长度和基因间隔区长度也是影响基因组
大小的主要因素,褐飞虱的内含子总长 281.4 Mb,豌
豆蚜只有 114.1 Mb,褐飞虱基因间隔区 319.8 Mb,
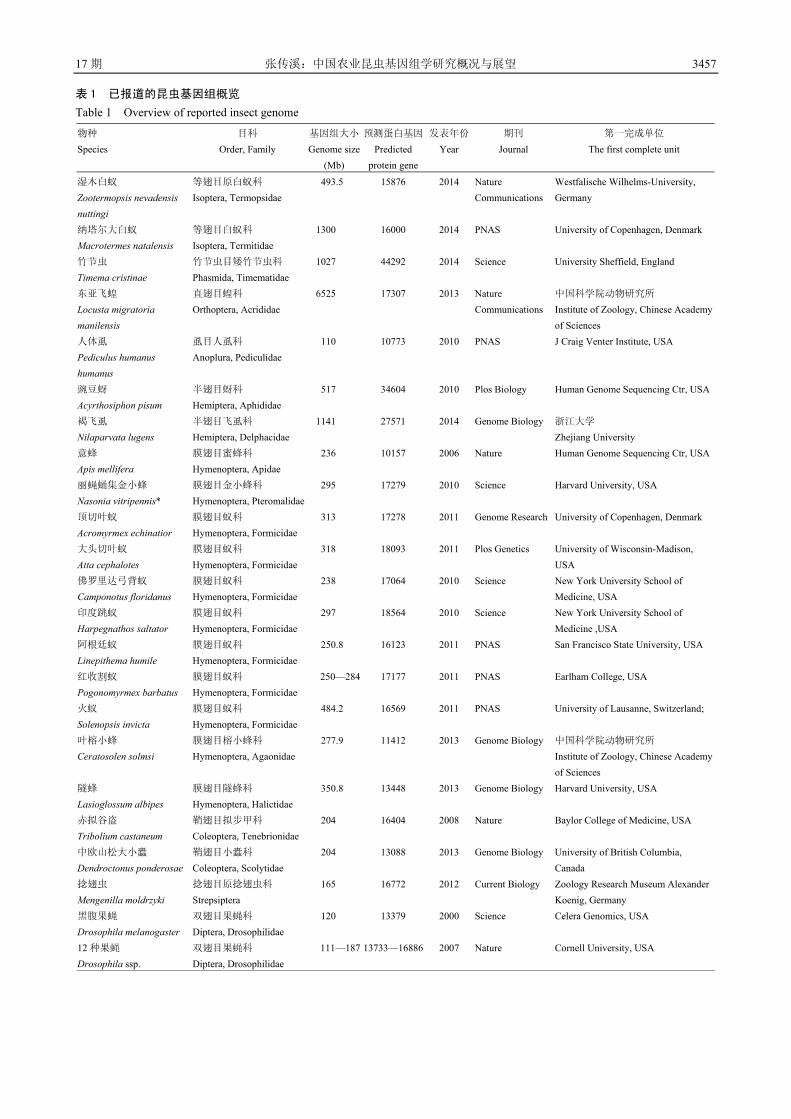
17 期 张传溪:中国农业昆虫基因组学研究概况与展望 3457
表 1 已报道的昆虫基因组概览
Table 1 Overview of reported insect genome
物种 Species
目科 Order, Family
基因组大小
Genome size (Mb)
预测蛋白基因
Predicted protein gene
发表年份
Year 期刊
Journal 第一完成单位
The first complete unit
湿木白蚁 Zootermopsis nevadensis nuttingi
等翅目原白蚁科 Isoptera, Termopsidae
493.5 15876 2014 Nature Communications
Westfalische Wilhelms-University, Germany
纳塔尔大白蚁 Macrotermes natalensis
等翅目白蚁科 Isoptera, Termitidae
1300 16000 2014 PNAS University of Copenhagen, Denmark
竹节虫 Timema cristinae
竹节虫目矮竹节虫科
Phasmida, Timematidae 1027 44292 2014 Science University Sheffield, England
东亚飞蝗 Locusta migratoria manilensis
直翅目蝗科 Orthoptera, Acrididae
6525 17307 2013 Nature Communications
中国科学院动物研究所 Institute of Zoology, Chinese Academy of Sciences
人体虱 Pediculus humanus humanus
虱目人虱科 Anoplura, Pediculidae
110 10773 2010 PNAS J Craig Venter Institute, USA
豌豆蚜 Acyrthosiphon pisum
半翅目蚜科 Hemiptera, Aphididae
517 34604 2010 Plos Biology Human Genome Sequencing Ctr, USA
褐飞虱 Nilaparvata lugens
半翅目飞虱科 Hemiptera, Delphacidae
1141 27571 2014 Genome Biology 浙江大学 Zhejiang University
意蜂 Apis mellifera
膜翅目蜜蜂科 Hymenoptera, Apidae
236 10157 2006 Nature Human Genome Sequencing Ctr, USA
丽蝇蛹集金小蜂 Nasonia vitripennis*
膜翅目金小蜂科
Hymenoptera, Pteromalidae 295 17279 2010 Science Harvard University, USA
顶切叶蚁 Acromyrmex echinatior
膜翅目蚁科 Hymenoptera, Formicidae
313 17278 2011 Genome Research University of Copenhagen, Denmark
大头切叶蚁 Atta cephalotes
膜翅目蚁科 Hymenoptera, Formicidae
318 18093 2011 Plos Genetics University of Wisconsin-Madison, USA
佛罗里达弓背蚁 Camponotus floridanus
膜翅目蚁科 Hymenoptera, Formicidae
238 17064 2010 Science New York University School of Medicine, USA
印度跳蚁 Harpegnathos saltator
膜翅目蚁科 Hymenoptera, Formicidae
297 18564 2010 Science New York University School of Medicine ,USA
阿根廷蚁 Linepithema humile
膜翅目蚁科 Hymenoptera, Formicidae
250.8 16123 2011 PNAS San Francisco State University, USA
红收割蚁 Pogonomyrmex barbatus
膜翅目蚁科 Hymenoptera, Formicidae
250—284 17177 2011 PNAS Earlham College, USA
火蚁 Solenopsis invicta
膜翅目蚁科 Hymenoptera, Formicidae
484.2 16569 2011 PNAS University of Lausanne, Switzerland;
叶榕小蜂 Ceratosolen solmsi
膜翅目榕小蜂科 Hymenoptera, Agaonidae
277.9 11412 2013 Genome Biology 中国科学院动物研究所 Institute of Zoology, Chinese Academy of Sciences
隧蜂 Lasioglossum albipes
膜翅目隧蜂科 Hymenoptera, Halictidae
350.8 13448 2013 Genome Biology Harvard University, USA
赤拟谷盗 Tribolium castaneum
鞘翅目拟步甲科 Coleoptera, Tenebrionidae
204 16404 2008 Nature Baylor College of Medicine, USA
中欧山松大小蠹 Dendroctonus ponderosae
鞘翅目小蠹科 Coleoptera, Scolytidae
204 13088 2013 Genome Biology University of British Columbia, Canada
捻翅虫 Mengenilla moldrzyki
捻翅目原捻翅虫科 Strepsiptera
165 16772 2012 Current Biology Zoology Research Museum Alexander Koenig, Germany
黑腹果蝇 Drosophila melanogaster
双翅目果蝇科 Diptera, Drosophilidae
120 13379 2000 Science Celera Genomics, USA
12 种果蝇 Drosophila ssp.
双翅目果蝇科 Diptera, Drosophilidae
111—187 13733—16886 2007 Nature Cornell University, USA
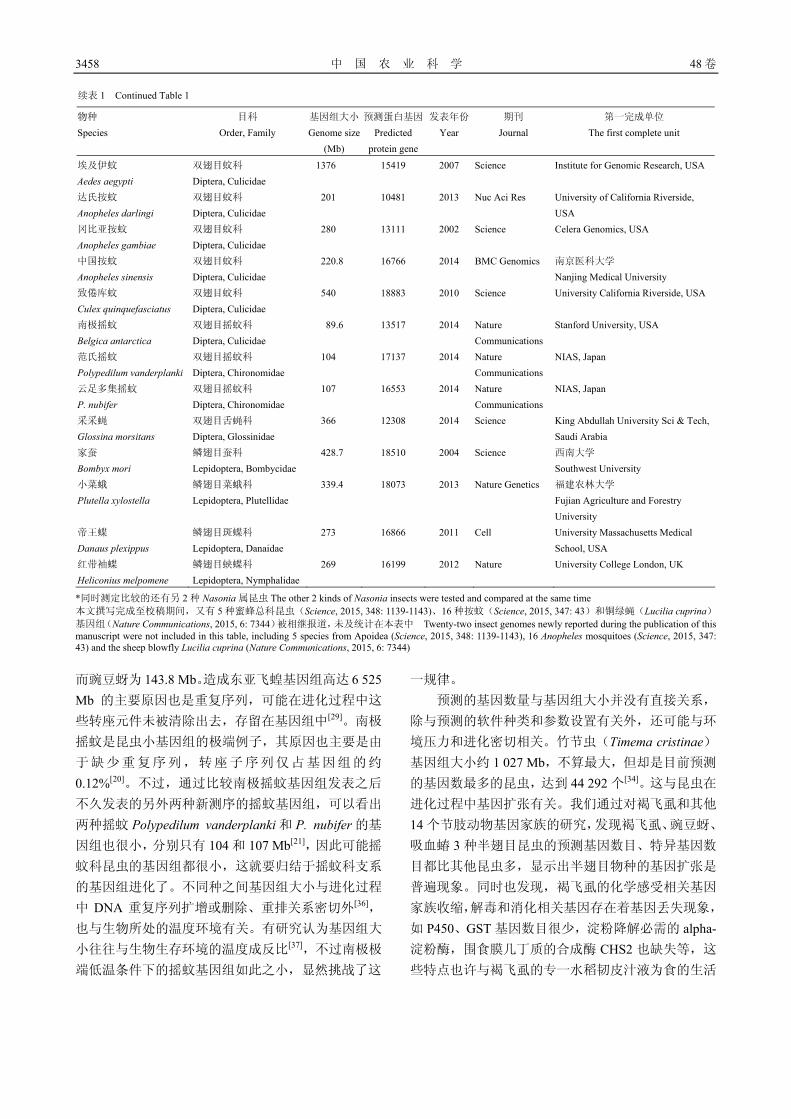
3458 中 国 农 业 科 学 48 卷
续表 1 Continued Table 1
物种 Species
目科 Order, Family
基因组大小
Genome size (Mb)
预测蛋白基因
Predicted protein gene
发表年份
Year 期刊
Journal 第一完成单位
The first complete unit
埃及伊蚊 Aedes aegypti
双翅目蚊科 Diptera, Culicidae
1376 15419 2007 Science Institute for Genomic Research, USA
达氏按蚊 Anopheles darlingi
双翅目蚊科 Diptera, Culicidae
201 10481 2013 Nuc Aci Res University of California Riverside, USA
冈比亚按蚊 Anopheles gambiae
双翅目蚊科 Diptera, Culicidae
280 13111 2002 Science Celera Genomics, USA
中国按蚊 Anopheles sinensis
双翅目蚊科 Diptera, Culicidae
220.8 16766 2014 BMC Genomics 南京医科大学 Nanjing Medical University
致倦库蚊 Culex quinquefasciatus
双翅目蚊科 Diptera, Culicidae
540 18883 2010 Science University California Riverside, USA
南极摇蚊 Belgica antarctica
双翅目摇蚊科 Diptera, Culicidae
89.6 13517 2014 Nature Communications
Stanford University, USA
范氏摇蚊 Polypedilum vanderplanki
双翅目摇蚊科 Diptera, Chironomidae
104 17137 2014 Nature Communications
NIAS, Japan
云足多集摇蚊 P. nubifer
双翅目摇蚊科 Diptera, Chironomidae
107 16553 2014 Nature Communications
NIAS, Japan
采采蝇 Glossina morsitans
双翅目舌蝇科 Diptera, Glossinidae
366 12308 2014 Science King Abdullah University Sci & Tech, Saudi Arabia
家蚕 Bombyx mori
鳞翅目蚕科 Lepidoptera, Bombycidae
428.7 18510 2004 Science 西南大学 Southwest University
小菜蛾 Plutella xylostella
鳞翅目菜蛾科 Lepidoptera, Plutellidae
339.4 18073 2013 Nature Genetics 福建农林大学 Fujian Agriculture and Forestry University
帝王蝶 Danaus plexippus
鳞翅目斑蝶科 Lepidoptera, Danaidae
273 16866 2011 Cell University Massachusetts Medical School, USA
红带袖蝶 Heliconius melpomene
鳞翅目蛱蝶科 Lepidoptera, Nymphalidae
269 16199 2012 Nature University College London, UK
*同时测定比较的还有另 2 种 Nasonia 属昆虫 The other 2 kinds of Nasonia insects were tested and compared at the same time 本文撰写完成至校稿期间,又有 5 种蜜蜂总科昆虫(Science, 2015, 348: 1139-1143)、16 种按蚊(Science, 2015, 347: 43)和铜绿蝇(Lucilia cuprina)基因组(Nature Communications, 2015, 6: 7344)被相继报道,未及统计在本表中 Twenty-two insect genomes newly reported during the publication of this manuscript were not included in this table, including 5 species from Apoidea (Science, 2015, 348: 1139-1143), 16 Anopheles mosquitoes (Science, 2015, 347: 43) and the sheep blowfly Lucilia cuprina (Nature Communications, 2015, 6: 7344)
而豌豆蚜为 143.8 Mb。造成东亚飞蝗基因组高达 6 525 Mb 的主要原因也是重复序列,可能在进化过程中这
些转座元件未被清除出去,存留在基因组中[29]。南极
摇蚊是昆虫小基因组的极端例子,其原因也主要是由
于缺少重复序列,转座子序列仅占基因组的约
0.12%[20]。不过,通过比较南极摇蚊基因组发表之后
不久发表的另外两种新测序的摇蚊基因组,可以看出
两种摇蚊 Polypedilum vanderplanki 和 P. nubifer 的基
因组也很小,分别只有 104 和 107 Mb[21],因此可能摇
蚊科昆虫的基因组都很小,这就要归结于摇蚊科支系
的基因组进化了。不同种之间基因组大小与进化过程
中 DNA 重复序列扩增或删除、重排关系密切外[36],
也与生物所处的温度环境有关。有研究认为基因组大
小往往与生物生存环境的温度成反比[37],不过南极极
端低温条件下的摇蚊基因组如此之小,显然挑战了这
一规律。 预测的基因数量与基因组大小并没有直接关系,
除与预测的软件种类和参数设置有关外,还可能与环
境压力和进化密切相关。竹节虫(Timema cristinae)基因组大小约 1 027 Mb,不算 大,但却是目前预测
的基因数 多的昆虫,达到 44 292 个[34]。这与昆虫在
进化过程中基因扩张有关。我们通过对褐飞虱和其他
14 个节肢动物基因家族的研究,发现褐飞虱、豌豆蚜、
吸血蝽 3 种半翅目昆虫的预测基因数目、特异基因数
目都比其他昆虫多,显示出半翅目物种的基因扩张是
普遍现象。同时也发现,褐飞虱的化学感受相关基因
家族收缩,解毒和消化相关基因存在着基因丢失现象,
如 P450、GST 基因数目很少,淀粉降解必需的 alpha-淀粉酶,围食膜几丁质的合成酶 CHS2 也缺失等,这
些特点也许与褐飞虱的专一水稻韧皮汁液为食的生活

17 期 张传溪:中国农业昆虫基因组学研究概况与展望 3459
习性相关[28]。意大利蜜蜂是目前预测的基因数量 少
的昆虫,只有 10 157 个,其他膜翅目昆虫基因数量也
都不到 19 000 个,膜翅目、鞘翅目、双翅目等目昆虫
基因数量普遍较少,显然与其在进化过程中没有经历
过基因大规模扩张的过程有关。
4 我国农业昆虫基因组研究概况
如前所述,在目前世界上已经报道的 50 余种昆虫
基因组中,我国科学家为主研究的有 5 种,包括家蚕、
小菜蛾、东亚飞蝗、榕小蜂和褐飞虱。西南农业大学
向仲怀院士领导的团队在 2004 年完成的家蚕基因组
是 早的一个[5],其结果为这一重要产丝昆虫遗传改
造利用和鳞翅目昆虫研究方面提供了重要基础。此后
随着 2 代测序技术发展和成本下降,2009 年开始,我
国科学家相继探索利用 2 代测序技术开展重要害虫褐
飞虱、二化螟、小菜蛾等基因组研究,但意料不到的
是,与已测双翅目、膜翅目不同,多数农业重要害虫
杂合度很高,基因组拼接很困难,使得项目进展迟缓。
此后,通过多代连续自交纯化,并采用新的测序和组
装策略,即 Fosmid-to-Fosmid 结合全基因组鸟枪法
(WGS),先后完成了小菜蛾和褐飞虱 2 种复杂基因
组,同时通过更大通量的测序,也完成了目前基因组
大小达 6.5 G 的东亚飞蝗的组装。其中,福建农林
大学尤民生教授领导的团队完成的小菜蛾基因组分
析[25],对于揭示世界性蔬菜害虫小菜蛾与十字花科植
物协同进化及其抗药性的适应进化与治理等均具有重
要的科学价值。中国科学院动物研究所康乐院士团队
完成的东亚飞蝗基因组[29],为蝗虫蝗灾治理和直翅目
昆虫研究提供了重要基因组信息基础。笔者团队完成
的褐飞虱及其 2 种内共生菌代谢互补基因组[28],揭示
了位于宿主脂肪体内的一种共生真菌和一种共生细菌
是如何与宿主褐飞虱“狼狈为奸”,使褐飞虱能仅依
赖水稻汁液就能快速繁殖成灾,研究成果为通过切断
褐飞虱-共生真菌-共生细菌营养途径防治害虫提供了
新视野,也给后续的不同种群间差异、迁飞途径、致
害性变异和抗药性的研究提供了基因组基础。此外,
中国科学院动物研究所黄大卫研究团队完成的榕小蜂
基因组[15],对于榕小蜂-榕树的特殊共进化研究有重要
价值。值得指出的是,上述昆虫基因组测序、组装和
注释均在华大基因公司完成,这段时间华大基因公司
为我国多种生物基因组发展起到了技术支撑作用。目
前,我国多个研究单位正在开展烟粉虱、斜纹夜蛾、
二化螟、白背飞虱、多种寄生蜂等一批重要农业昆虫
的基因组学研究。
5 我国农业昆虫基因组学应用
随着我国几种昆虫基因组的测定进展,这几种昆 虫的研究也同时进入了后基因组时代。家蚕本身是支
撑蚕丝产业的物种基础,也是开发新一代生物反应器
的材料,同时也是鳞翅目昆虫研究的模式种类。自家
蚕基因组框架图完成后,又对家蚕 40 品系基因变异进
行了重测序研究[38],目前在重要经济性状基因定位克
隆、抗病毒病育种、转基因新型有色茧实用蚕品种均
已经取得重要进展。在东亚飞蝗基因组信息基础上,
康乐院士团队在东亚飞蝗功能基因组方面取得了一系
列进展,特别是在飞蝗群居型和散居型两型转变分子
机制方面有重要突破[39-40]。在褐飞虱基因组序列基础
上,我们通过进一步分析和实验,表明褐飞虱存在系
统 RNAi[41],这为利用基于 RNAi 技术防治稻飞虱提
供了基础。对与迁飞和成灾密切相关的稻飞虱翅型相
关基因研究也已取得了很大进展[28,42],并进一步突破
性地发现确证了 2 个序列同源性很高的胰岛素受体是
控制长短翅型可塑性发育的“分子开关”[43]。同时,
在褐飞虱基因组基础上,我们分析了褐飞虱几丁质合
成和降解所需要的关键基因体系,提出了一种昆虫完
成几丁质代谢所需参与关键酶的“1+5+1+3”模式,
即褐飞虱几丁质合成需要 1 个合酶的 2 个可变剪切本
(NlChs1a、NlChs1b),几丁质降解至少需要 5 个
NlCht、1 个 NlHex 和 3 个 NlCDA 的共同参与[44-46],
由于几丁质是昆虫表皮的主要成分,也是重要的新农
药靶标,褐飞虱几丁质合成和降解体系的解析不仅可
提升对昆虫蜕皮变态的认识,对害虫防治也有重要参
考价值。
6 昆虫转录组学与基因组关系
谈到昆虫基因组,不得不说与之密切相关的转录
组分析。自从 2010 年我们在国际上率先开展烟粉虱和
褐飞虱 2 种昆虫的转录组和表达谱分析[47-48]以来,近
几年可以说是极速发展,如用“insect”、“transcriptome”和“China”搜索 ISI 数据库,短短 4 年多时间,我国
已经发表昆虫转录组相关论文 267 篇。特别是第二代
高通量测序技术高通量、快速、低成本的特点,使
RNA-seq 有了超强的覆盖度和灵敏性,还可检出许多
不曾被预测到的由可变剪接或可变 3′多聚腺苷化位点
选择导致的 mRNA isoforms,以及新的 ncRNA 和
antisense RNA。因此,RNA-seq 是基因组研究必不可

3460 中 国 农 业 科 学 48 卷
少的伴侣,转录组是连接基因组遗传信息-蛋白质组-基因生物学功能的必然纽带。同时与昆虫全基因组相
比,转录组测序得到的序列不含有内含子及其他非编
码序列,因此转录组测序有着极高的性价比优势。对
更多目前还没有测定全基因组的昆虫来说,要深入研
究千差万别的昆虫生物学特性的分子机理,先测转录
组更是不二的选择。因此,无论对已测基因组还是没
有测定基因组的昆虫来说,转录组和表达谱都是昆虫
功能基因和功能基因组研究的重要手段,许多昆虫生
物学特性解析都可以从转录组分析和表达谱差异比较
入手。
7 展望
随着测序技术快速发展,测序通量越来越大,读
长越来越长,基因组测序成本急剧下降,“旧时王谢
庭前燕,飞入寻常百姓家”,越来越多的实验室开展
了各种昆虫的基因组学研究。昆虫基因组学之“热”,
从 近几次相关学术活动可窥一斑。在康乐院士组织
下,2013 年 12 月 15 日在中国科学院动物研究所举办
了“首届中国昆虫基因组学及国际 5 000 种昆虫基因
组(i5k)计划研讨会”,有 39 位学者发言,从全基
因组测序、功能基因组学、比较和进化基因组学、生
物信息学分析技术等多个主题,热烈探讨了昆虫基因
组学未来发展趋势,以及如何推进基因组学在害虫防
治、资源昆虫利用、药物靶点开发及进化生物学等方
面的应用问题。此后,中国昆虫学会昆虫基因组学专
业委员会应运而生,并于 2014 年 10 月 22—25 日在河
北保定中国昆虫学会学术年会期间组织了第一次学术
活动。上述两次学术会议吸引了来自我国高校和科研
院所昆虫学各领域的 200 多昆虫研究者参与交流讨
论,人数之多均超过会议组织方预料。这也从一个侧
面说明昆虫基因组学研究,契合了我国昆虫学当前的
发展趋势,昆虫学者都意识到基因组学的重要性。值
得指出的是昆虫基因组学研究目前毕竟还是颇费经费
的研究,宜切忌一哄而上,更不能发篇文章就了事,
不然有“钱多人傻”之嫌。Robinson 等提出的“i5k”计划[49],笔者看来也只是美国人提法大胆而已,要在
其计划的 5 年内全部启动几无可能。我国参与“i5k”计划,也务必从实际需要出发,重点选择那些对我国有
重要经济和生态价值的昆虫种类。更为重要的是,基因
组学序列研究仅仅是万里长征的第一步,要使基因组信
息资源在我国科学研究和国民经济发展中充分发挥应
有的作用,特别需要后续功能基因组研究的跟进。
随着越来越多的昆虫基因组得到解析,昆虫学的
研究发生了前所未有的深刻变化,它不仅为不同昆虫
生物学特性的机理研究提供了序列基础,也使我们能
从更全局的角度研究一种昆虫的生命活动规律,特别
是结合转录组、蛋白组、代谢组等 “组学”,使我们
对昆虫与环境关系的内在机理在整体上有了更好把
握,而不再是“盲人摸象”。生物信息学为基础的基
因功能预测和同源性分析比对、靶标基因的敲除和敲
入、基因 RNAi 技术等功能基因组研究手段,使昆虫
基因组资源利用研究突飞猛进。特别是在基因组和转
录组基础上,通过基因功能的研究和挖掘,应用
CRISPR-Cas9 介导的基因组定点编辑技术和基因
RNAi 技术,将使害虫的防治和益虫的利用出现革命
性的变化。
References
[1] Adams M D, Celniker S E, Holt R A, Evans C A, Gocayne J D,
Amanatides P G, Scherer S E, Li P W, Hoskins R A, Galle R F. The
genome sequence of Drosophila melanogaster. Science, 2000,
287(5461): 2185-2195.
[2] Holt R A, Subramanian G M, Halpern A, Sutton G G, Charlab R,
Nusskern D R, Wincker P, Clark A G, Ribeiro J M C, Wides R. The
genome sequence of the malaria mosquito Anopheles gambiae.
Science, 2002, 298(5591): 129-149.
[3] Nene V, Wortman J R, Lawson D, Haas B, Kodira C, Tu Z, Loftus B,
Xi Z, Megy K, Grabherr M. Genome sequence of Aedes aegypti, a
major arbovirus vector. Science, 2007, 316: 1718-1722.
[4] Drosophila 12 Genomes Consortium. Evolution of genes and genomes
on the Drosophila phylogeny. Nature, 2007, 450: 203-218.
[5] Xia Q, Zhou Z, Lu C, Cheng D, Dai F, Li B, Zhao P, Zha X, Cheng T,
Chai C. A draft sequence for the genome of the domesticated
silkworm (Bombyx mori). Science, 2004, 306(5703): 1937-1940.
[6] The Honeybee Genome Sequencing Consortium. Insights into social
insects from the genome of the honeybee Apis mellifera. Nature, 2006,
443(7114): 931-949.
[7] Richards S, Gibbs R A, Weinstock G M, Brown S J, Denell R,
Beeman R W, Gibbs R, Bucher G, Friedrich M, Grimmelikhuijzen C J.
The genome of the model beetle and pest Tribolium castaneum.
Nature, 2008, 452: 949-955.
[8] The Nasonia Genome Working Group. Functional and evolutionary
insights from the genomes of three parasitoid Nasonia species.
Science, 2010, 327(5963): 343-348.
[9] Bonasio R, Zhang G, Ye C, Mutti N S, Fang X, Qin N, Donahue G,

17 期 张传溪:中国农业昆虫基因组学研究概况与展望 3461
Yang P, Li Q, Li C, Zhang P, Huang Y, Berger S L, Reinberg D, Wang
J, Liebig J. Genomic comparison of the ants Camponotus floridanus
and Harpegnathos saltator. Science, 2010, 329(5995): 1068-1071.
[10] Nygaard S, Zhang G, Schiøtt M, Li C, Wurm Y, Hu H, Zhou J, Ji L,
Qiu F, Rasmussen M, Pan H, Hauser F, Krogh A, Grimmelikhuijzen C
J P, Wang J, Boomsma J J. The genome of the leaf-cutting ant
Acromyrmex echinatior suggests key adaptations to advanced social
life and fungus farming. Genome Research, 2011, 21(8): 1339-1348.
[11] Suen G, Teiling C, Li L, Holt C, Abouheif E, Bornberg-Bauer E,
Bouffard P, Caldera E J, Cash E, Cavanaugh A. The genome sequence
of the leaf-cutter ant Atta cephalotes reveals insights into its obligate
symbiotic lifestyle. PLoS Genetics, 2011, 7(2): e1002007.
[12] Smith C D, Zimin A, Holt C, Abouheif E, Benton R, Cash E, Croset V,
Currie C R, Elhaik E, Elsik C G. Draft genome of the globally
widespread and invasive Argentine ant (Linepithema humile).
Proceedings of the National Academy of Sciences of the United States
of America, 2011, 108(14): 5673-5678.
[13] Smith C R, Smith C D, Robertson H M, Helmkampf M, Zimin A,
Yandell M, Holt C, Hu H, Abouheif E, Benton R. Draft genome of the
red harvester ant Pogonomyrmex barbatus. Proceedings of the
National Academy of Sciences of the United States of America, 2011,
108(14): 5667-5672.
[14] Wurm Y, Wang J, Riba-Grognuz O, Corona M, Nygaard S, Hunt B G,
Ingram K K, Falquet L, Nipitwattanaphon M, Gotzek D. The genome
of the fire ant Solenopsis invicta. Proceedings of the National
Academy of Sciences of the United States of America, 2011, 108(14):
5679-5684.
[15] Xiao J H, Yue Z, Jia L Y, Yang X H, Niu L H, Wang Z, Zhang P, Sun B
F, He S M, Li Z. Obligate mutualism within a host drives the extreme
specialization of a fig wasp genome. Genome Biology, 2013, 14(12):
R141.
[16] Kocher S D, Li C, Yang W, Tan H, Yi S V, Yang X, Hoekstra H E,
Zhang G, Pierce N E, Yu D W. The draft genome of a socially
polymorphic halictid bee, Lasioglossum albipes. Genome Biology,
2013, 14(12): R142.
[17] Arensburger P, Megy K, Waterhouse R M, Abrudan J, Amedeo P,
Antelo B, Bartholomay L, Bidwell S, Caler E, Camara F.
Sequencing of Culex quinquefasciatus establishes a platform for
mosquito comparative genomics. Science, 2010, 330(6000): 86-88.
[18] Marinotti O, Cerqueira G C, de Almeida L G P, Ferro M I T, Loreto E
L D, Zaha A, Teixeira S M R, Wespiser A R, Silva A A E, Schlindwein
A D. The genome of Anopheles darlingi, the main neotropical malaria
vector. Nucleic Acid Research, 2013, 41(15): 7387-7400.
[19] Zhou D, Zhang D, Ding G, Shi L, Hou Q, Ye Y, Xu Y, Zhou H, Xiong
C, Li S. Genome sequence of Anopheles sinensis provides insight into
genetics basis of mosquito competence for malaria parasites. BMC
Genomics, 2014, 15: 42.
[20] Kelley J L, Peyton J T, Fiston-Lavier A S, Teets N M, Yee M C,
Johnston J S, Bustamante C D, Lee R E, Denlinger D L. Compact
genome of the Antarctic midge is likely an adaptation to an extreme
environment. Nature Communications, 2014, 5: Article number 4611.
[21] Gusev O, Suetsugu Y, Cornette R, Kawashima T, Logacheva M D,
Kondrashov A S, Penin A A, Hatanaka R, Kikuta S, Shimura S.
Comparative genome sequencing reveals genomic signature of
extreme desiccation tolerance in the anhydrobiotic midge. Nature
Communications, 2014, 5: Article number 4784.
[22] International Glossina Genome Initiative. Genome sequence of the
tsetse fly (Glossina morsitans): vector of African trypanosomiasis.
Science, 2014, 344(6182): 380-386.
[23] Zhan S, Merlin C, Boore J L, Reppert S M. The Monarch butterfly
genome yields insights into long-distance migration. Cell, 2011,
147(5): 1171-1185.
[24] The Heliconius Genome Consortium. Butterfly genome reveals
promiscuous exchange of mimicry adaptations among species. Nature,
2012, 487: 94-98.
[25] You M, Yue Z, He W, Yang X, Yang G, Xie M, Zhan D, Baxter S W,
Vasseur L, Gurr G M. A heterozygous moth genome provides insights
into herbivory and detoxification. Nature Genetics, 2013, 45(2): 220-225.
[26] Keeling C I, Yuen M M S, Liao N Y, Docking T R, Chan S K, Taylor
G A, Palmquist D L, Jackman S D, Nguyen A, Li M. Draft genome of
the mountain pine beetle, Dendroctonus ponderosae Hopkins, a major
forest pest. Genome Biology, 2013, 14(3): R27.
[27] The International Aphid Genomics Consortium. Genome sequence of
the pea aphid Acyrthosiphon pisum. PLoS Biology, 2010, 8(2):
e1000313.
[28] Xue J, Zhou X , Zhang C X, Yu L L, Fan H W, Wang Z, Xu H J, Xi Y,
Zhu Z R , Zhou W W. Genomes of the rice-pest brown planthopper
and its endosymbionts reveal complex complementary contributions
for host adaptation. Genome Biology, 2014, 15(12): 521.
[29] Wang X, Fang X, Yang P, Jiang X, Jiang F, Zhao D, Li B, Cui F, Wei J,
Ma C. The locust genome provides insight into swarm formation and
long-distance flight. Nature Communications, 2014, 5: 2957.
[30] Kirkness E F, Haas B J, Sun W, Henk R, Braig H R, Perotti M A,
Clark J M, Lee S H, Robertson H M, Kennedy R C, Elhaik E. Genome
sequences of the human body louse and its primary endosymbiont
provide insights into the permanent parasitic lifestyle. Proceedings of

3462 中 国 农 业 科 学 48 卷
the National Academy of Sciences of the United States of America,
2011, 108(15): 6335-6336.
[31] Terrapon N, Li C, Robertson H M, Ji L, Meng X H, Booth W, Chen Z
S, Childers C P, Glastad K M, Gokhale K. Molecular traces of
alternative social organization in a termite genome. Nature
Communications, 2014, 5: 3636.
[32] Poulsen M, Hu H F, Li C, Chen Z S, Xu L H, Otani S, Nygaard S,
Nobre T, Klaubauf S, Schindler P M. Complementary symbiont
contributions to plant decomposition in a fungus-farming termite.
Proceedings of the National Academy of Sciences of the United States
of America, 2014, 111(40): 14500-14505.
[33] Niehuis O, Hartig G, Grath S, Pohl H, Lehmann J, Tafer H, Donath A,
Krauss V, Eisenhardt C, Hertel J, Petersen M, Mayer C, Meusemann
K, Peters R S, Stadler P F, Beutel R G, Bornberg-Bauer E, McKenna
D D, Misof B. Genomic and morphological evidence converge to
resolve the enigma of Strepsiptera. Current Biology, 2012, 22(14):
1309-1313.
[34] Soria-Carrasco V, Gompert Z, Comeault A A, Farkas T E, Parchman T
L, Johnston J S, Buerkle C A, Feder J L, Bast J, Schwander T, Egan S
P, Crespi B J, Nosil P. Stick insect genomes reveal natural selection’s
role in parallel speciation. Science, 2014, 344(6185): 738-742.
[35] 薛建, 程家安, 张传溪. 昆虫基因组及其大小. 昆虫学报, 2009,
52(8): 901-906.
Xue J, Cheng J A, Zhang C X. Insect genomes and their sizes. Acta
Entomologica Sinica, 2009, 52(8): 901-906. (in Chinese)
[36] Biemont C. Genome size evolution: within-species variation in
genome size. Heredity, 2008, 101(4): 297-298.
[37] Hessen D O, Daufresne M, Leinaas H P. Temperature-size relations
from the cellular-genomic perspective. Biological Reviews, 2013,
88(2): 476-489.
[38] Xia Q, Guo Y, Zhang Z, Li D, Xuan Z, Li Z, Dai F, Li Y, Cheng D, Li
R. Complete resequencing of 40 genomes reveals domestication
events and genes in silkworm (Bombyx). Science, 2009, 326(5951):
433-436.
[39] Ma Z Y, Guo W, Guo X J, Wang X H, Kang L. Modulation of
behavioral phase changes of the migratory locust by the catecholamine
metabolic pathway. Proceedings of the National Academy of Sciences of
the United States of America, 2011, 108(10): 3882-3887.
[40] Yang M L, Wei Y Y, Jiang F, Wang Y L, Guo X J, He J, Kang L.
MicroRNA-133 inhibits behavioral aggregation by controlling
dopamine synthesis in locusts. PLoS Genetics, 2014, 10(2): e1004206.
[41] Xu H J, Chen T, Ma X F, Xue J, Pan P L, Zhang X C, Cheng J A ,
Zhang C X. Genome-wide screening for components of small
interfering RNA (siRNA) and micro-RNA (miRNA) pathways in the
brown planthopper, Nilaparvata lugens (Hemiptera: Delphacidae).
Insect Molecular Biology, 2013, 22(6): 635-647.
[42] Xue J, Zhang X Q, Xu H J,Fan H W, Huang H J, Ma X F, Wang C Y,
Chen J G, Cheng J A, Zhang C X. Molecular characterization of the
flightin gene in the wing-dimorphic planthopper, Nilaparvata lugens,
and its evolution in Pancrustacea. Insect Biochemistry and Molecular
Biology, 2013, 43(5): 433-443.
[43] Xu H J, Xue J, Lu B, Zhang X C, Zhuo J C, He S F, Ma X F, Jiang Y
Q, Fan H W, Xu J Y, Ye Y X, Pan P L, Li Q, Bao Y Y, Nijhout H F,
Zhang C X. Two insulin receptors determine alternative wing morphs
in planthoppers. Nature, 2015, 519(7544): 464-467.
[44] Wang Y, Fan H W, Huang H J, Xue J, Wu W J, Bao Y Y, Xu H J, Zhu
Z R, Cheng J A, Zhang C X. Chitin synthase 1 gene and its two
alternative splicing variants from two sap-sucking insects, Nilaparvata
lugens and Laodelphgax striatellus (Hemiptera: Delphacidae). Insect
Biochemistry and Molecular Biology, 2012, 42(9): 637-646.
[45] Xi Y, Pan P L, Ye Y X, Yu B, Zhang C X. Chitin deacetylase family
genes in the brown planthopper, Nilaparvata lugens (Hemiptera:
Delphacidae). Insect Molecular Biology, 2014, 23(6): 695-705.
[46] Xi Y, Pan P L, Ye Y X, Yu B, Xu H J, Zhang C X. Chitinase-like
family genes in the brown planthopper, Nilaparvata lugens. Insect
Molecular Biology, 2015, 24(1): 29-40.
[47] Wang X W, Luan J B, Li J M, Bao Y Y, Zhang C X, Liu S S. De novo
characterization of a whitefly transcriptome and analysis of its gene
expression during development. BMC Genomics, 2010, 11: 400.
[48] Xue J, Bao Y Y, Li B L, Cheng Y B, Peng Z Y, Liu H, Zhu Z R, Lou Y
G, Cheng J A, Zhang C X. Transcriptome analysis of the brown
planthopper Nilaparvata lugens. PLoS ONE, 2010, 5(12): e14233.
[49] Robinson G E, Hackett K J, Purcell-Miramontes M, Brown S J, Evans
J D, Goldsmith M R, Lawson D, Okamuro J, Robertson H, Schneider
D J. Creating a buzz about insect genomes. Science, 2011, 331(6023):
1386.
(责任编辑 岳梅)

中国农业科学 2015,48(17):3463-3476 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.013
收稿日期:2015-03-24;接受日期:2015-05-27 基金项目:国家“973”计划(2013CB127600)、国家桃产业技术体系(nycytx-31-02)、北京市农林科学院科技创新能力建设专项(KJCX20140101)、
北京市科技计划(D141100000914000) 联系方式:张帆,E-mail:[email protected]。通信作者张帆,E-mail:[email protected]。通信作者王甦,E-mail:[email protected]
中国设施蔬菜害虫天敌昆虫应用研究进展
张 帆,李 姝,肖 达,赵 静,王 然,郭晓军,王 甦
(北京市农林科学院植物保护环境保护研究所,北京 100097)
摘要:设施蔬菜是现代农业生产中极为重要的组成部分,而且设施蔬菜的发展不仅使蔬菜生产的品种和产量
得到快速增长,也为有机蔬菜的发展创造了有利条件。但是,设施蔬菜种植条件也给害虫提供了适宜生长、繁殖
和危害的生态环境,严重影响了蔬菜的质量和产量,成为制约设施蔬菜产业进一步发展的关键因素。化学农药的
长期使用带来了一系列的生态环境和食品安全问题,而治理化学农药污染不仅要求逐步减少其使用量,更需要寻
求优化升级与替代传统防治方法的技术。天敌昆虫作为传统的生物防治产品,在控制设施蔬菜虫(螨)害,保证
其产量和品质中起着不可替代的作用。随着人们环境保护意识的加强和绿色农业的发展,以天敌昆虫释放为主的
生物防治技术在害虫综合治理(IPM)中发挥着越来越重要的作用。中国的天敌资源非常丰富,但目前在设施蔬菜
生产中应用的种类相当有限。经过几十年的努力,中国在设施蔬菜害虫生物防治及应用领域开展了大量的研究工
作,在天敌昆虫资源和应用基础、技术研发和配套技术及应用等方面取得了较大进展。本文概述了设施蔬菜害虫
的主要发生种类及其危害特点、主要害虫天敌资源及其生物防治的技术途径等,重点介绍了蚜虫、粉虱、叶螨、
蓟马等几种重大害虫的生物防治实例,并分析了中国天敌昆虫人工繁殖(人工饲料、规模化生产)的研究进展。
此外,还综述了中国温室蔬菜害虫的天敌应用技术研究成果,从天敌昆虫保护利用、释放技术、控害效果评价、
规模化生产等方面对设施蔬菜应用天敌昆虫进行生物防治中存在的问题进行了讨论,并展望了该领域的未来发展
方向。
关键词:设施蔬菜;害虫;天敌昆虫;人工繁殖;生物防治
Progress in Pest Management by Natural Enemies in
Greenhouse Vegetables in China ZHANG Fan, LI Shu, XIAO Da, ZHAO Jing, WANG Ran, GUO Xiao-jun, WANG Su
(Institute of Plant and Environment Protection, Beijing Academy of Agriculture and Forestry Sciences, Beijing 100097)
Abstract: Greenhouse vegetables are very important part of modern agricultural production and the development of vegetables produced in facilities not only makes the rapid growth of variety and production output of vegetables, but also creates favorable conditions for the development of organic vegetables. However, planting conditions of greenhouse vegetables have a serious impact on the quality and yield of vegetables through supplying a suitable environment for pest insect growth and reproduction. It has become the key factor restricting the further development of the vegetable industry. A series of environmental and food safety problems are produced with the long-term use of chemical pesticides. In order to manage the chemical pesticides pollution, not only their application amount should be reduced gradually, but also the technology for optimizing and upgrading the traditional control method should be developed. As traditional biological control products, the natural enemies of insect pests play an irreplaceable role in controlling vegetable insect pests and guaranteeing the yield and quality of vegetables. With the strengthening of people’s consciousness of environment protection and development of green agriculture, the biological control technology plays an important
jia
文本框
返回目录

3464 中 国 农 业 科 学 48 卷
role in the integrated pest management (IPM) which is based on natural enemy insect release. In China, the resources of natural enemy insect are very rich; however, the application of natural enemies in greenhouse vegetable is quite limited. After decades of efforts, lots of researches focus on vegetable pest biological control and application field. A great progress has been made in the resources of natural enemy insects, application basis, technology research development and supporting technology. In this paper, an overview that related to the damage characteristics of vegetable pest insects and biological control of main pest insects using natural enemy technique was summarized. Examples of biological control of aphids, whiteflies, spider mites and thrips were listed. The research progress in insect natural enemies artificial rearing (artificial feed, the scale of production) in China was analyzed. In addition, the results in technology research on greenhouse vegetables control using natural enemies were also studied. The problems existing in biological control including the protection use of natural enemies, release technology, control effect evaluation and scale production were also discussed. And the prospects of the development direction of this field in the future were pointed out.
Key words: greenhouse vegetable; pest; natural enemy; artificial rearing; biological control 设施蔬菜生产是现代农业生产中极为重要的组成
部分,可以高效利用环境因子增加蔬菜生产周期,提
高蔬菜产量和质量[1]。中国是世界上 大的温室蔬菜
种植国家,近年来每年以 10%左右的速度增长[2]。据
新华网报道,2014 年设施蔬菜面积已达到 386 万
hm2[3],蔬菜产值已超过 7 000 亿元,行业从业人员达
到 4 000 万以上,已成为许多区域农业的支柱产业[4-5]。
经过近 30 年的发展,中国设施蔬菜生产在不同地区形
成了各具特色的类型,例如小拱棚、大中棚、日光温
室和连栋温室等[1]。 随着设施蔬菜产业的迅速发展,使蔬菜生产的品
种和产量得到快速增长,但同时也给害虫提供了适宜
的生长、繁殖和危害的生态环境。各类植食性有害生
物的危害不断加剧,造成极大的经济损失[6]。如蚜虫、
蓟马类、粉虱类等小型害虫得到充分有利的发育条件,
有猖獗危害的趋势,发生面积不断扩大(约占播种面
积的 50%—80%),产量损失一般 15%—30%,严重
的减产 50%以上,甚至会造成绝收或使产品失去其商
品价值,严重影响着蔬菜产品的产量和品质。 由于蔬菜生长周期较短,害虫的发生种类多、危
害重,目前尚难于提出既能有效控制其发生又符合食
品安全生产要求的防治措施,化学农药仍占较大比重,
农药不合理的使用造成生态环境污染,影响食物安全。
因此设施农业重要害虫的生态、生物控制措施已成为
首选技术和发展方向。中国生物防治技术研究在 20世纪 80 年代开始列入了国家攻关计划。经过几十年的
努力,中国的生物防治研究把引进外来天敌昆虫防治
外来有害生物、保护和利用本地天敌放在首位,大力
发展规模化天敌生产,取得了较好的结果。设施蔬菜
害虫天敌昆虫研究应用主要涉及天敌资源筛选评价、
重要天敌大量繁殖技术和田间释放及保护利用策略等
方面。
1 设施蔬菜害虫主要种类
中国设施蔬菜重要害虫种类主要包含6目20种昆
虫(表 1)。在相对封闭、环境温暖湿润、作物种植
密度高的设施温室中,发育速率快、世代多、繁殖能
力强的害虫极易定殖与发展,而且比大田作物更易暴
发成灾[7-8]。其中以蚜虫、粉虱、叶螨和蓟马在设施蔬
菜生产中发生 为普遍、危害 重。如桃蚜(Myzus persicae),寄主范围很广,可在 50 多科近 400 种植
物上取食,并可传播 200 多种植物病毒。桃蚜通过直
接刺吸植物,造成植物营养不良,使寄主植物生长缓
慢甚至停滞 [9-10]。粉虱(温室白粉虱 Trialeurodes vaporariorum 和烟粉虱 Bemisia tabaci)的危害不仅降
低了果蔬的产量同时也严重影响果蔬的品质。由于粉
虱抗药性发展迅速,已对常用杀虫剂产生高水平的抗
性和交互抗性。中国设施蔬菜生产中常见叶螨大多是
叶螨属(Tetranychus),主要包括二斑叶螨(Tetranychus urticae)、朱砂叶螨(Tetranychus cinnabarinus)、截
形叶螨(Tetranychus truncatus)和侧多食跗线螨
(Polyphagotarsonemus latus)等,极易暴发成灾,严
重影响蔬菜的品质和质量[11]。蓟马是全球性重要的杂
食性昆虫,黄蓟马(Thrips flavus)和葱蓟马(Thrips tabaci)是危害蔬菜生产的主要蓟马种群。2003 年,
在北京郊区的蔬菜大棚内发现外来入侵种西花蓟马
(Frankliniella occidentalis)[12]。西花蓟马是一种世界
性检疫害虫,具有繁殖速度快和寄主范围广等特点,
危害极其严重,常常造成作物减产甚至绝收,现已成
为危害中国温室蔬菜生产的主要蓟马种类。
此外,许多害虫在温室条件下改变了发生规律,
如小菜蛾(Plutella xylostella)、菜粉蝶(Pieris rapae)等有冬眠习性的害虫,可以一年四季不间断地发育、
繁殖,发生基数大增,危害加重[13]。蚜虫及螨类害虫

17 期 张帆等:中国设施蔬菜害虫天敌昆虫应用研究进展 3465
也改变了其在木本植物上以卵越冬的习性,以孤雌生
殖的方式不断周年存在并持续危害[14]。害虫发生面积
不断扩大(约占播种面积的 50%—80%)、危害频率
增加(一年发生十几代以上)、危害程度加重(总虫
株率 80%,叶片受害率 60%,产量损失一般达 15%—
30%,严重的减产 50%以上,甚至造成绝收或使产品
失去商品价值),严重影响蔬菜的产量和品质[6]。在
温室蔬菜农业生态系统中,由外来入侵害虫和本地害
虫构成的复合群落大大降低了化学杀虫剂的控害效
率。
表 1 中国设施蔬菜害虫主要种类
Table 1 The primary pests of greenhouse vegetables in China
目 Order 害虫 Pest 寄主植物 Host plant
半翅目 Hemiptera 烟粉虱 B. tabaci 茄科、十字花科、葫芦科、豆科等 Solanaceae, Brassicaceae, Cucurbitaceae, Leguminosae, etc.
温室白粉虱 T. vaporariorum 茄科 Solanaceae
桃蚜 M. persicae 十字花科、茄科 Brassicaceae, Solanaceae
菜缢管蚜 Rhopalosiphum pseudobrassicae 甘蓝、花椰菜 Brassica oleracea, Brassica oleracea var. botrytis
甘蓝蚜 Brevicoryne brassicae 大白菜、萝卜等 Brassica rapa pekinensis, Raphanus sativus, etc.
棉蚜 Aphis gossypii 葫芦科、茄科、豆科、菊科及十字花科 Cucurbitaceae, Solanaceae, Leguminosae, Asteraceae, Brassicaceae
瓜蓟马 Thrips flavus 葫芦科、茄科、豆科 Cucurbitaceae, Solanaceae, Leguminosae 缨翅目 Thysanoptera
西花蓟马 F. occidentalis 草莓、茄科等 Fragaria ananassa, Solanaceae, etc.
双翅目 Diptera 美洲斑潜蝇 Liriomyza sativae 茄科、豆科等 Solanaceae, Leguminosae, etc.
鞘翅目 Coleoptera 黄曲条跳甲 Phyllotreta striolata 十字花科(白菜、萝卜、芥菜等) Brassicaceae (Brassica campestris ssp. chinensis, Raphanus sativus, Brassica juncea, etc.)
黄足黄守瓜 Aulacophora indica 葫芦科 Cucurbitaceae
鳞翅目 Lepidoptera 小菜蛾 P. xylostella 十字花科(甘蓝、花椰菜、白菜、萝卜) Brassicaceae (Brassica oleracea, Brassica oleracea var. botrytis, Brassica campestris ssp. chinensis, Raphanus sativus)
菜粉蝶 P. rapae 十字花科(甘蓝、花椰菜和球茎甘蓝等) Brassicaceae (Brassica oleracea, Brassica oleracea var. botrytis, Brassica oleracea var. caulorapa, etc.)
豆野螟 Maruca testulalis 豆科 Leguminosae
小地老虎 Agrotis ypsilon 十字花科、茄科、葫芦科及豆科 Brassicaceae, Solanaceae, Cucurbitaceae, Leguminosae
甘蓝夜蛾 Mamestra (Barathra) brassicae 甘蓝、白菜、萝卜、烟草、菠菜、甜菜及豆科 Brassica oleracea, Brassica campestris ssp. chinensis, Raphanus sativus, Nicotiana tabacum, Spinacia oleracea, Beta vulgaris, Leguminosae
斜纹夜蛾 Prodenia litura 十字花科、茄科 Brassicaceae, Solanaceae
银纹夜蛾 Argyrogramma agnata 十字花科、豆科、茄科 Brassicaceae, Leguminosae, Solanaceae
真螨目 Acariformes 二斑叶螨 Tetranychus urticae 茄科 Solanaceae
侧多食跗线螨 P. latus 茄科、豆科、葫芦科及萝卜 Solanaceae, Leguminosae, Cucurbitaceae, Raphanus sativus
2 天敌昆虫应用历史
中国是 早人工开展生物防治的国家,早在公元前
304 年,就有利用捕食性昆虫——黄猄蚁(Oecophylla smaragdina)来防治柑橘园害虫的记录[15]。自 20 世纪
50 年代起,中国从国外引入赤眼蜂(Trichogrammatid)、澳洲瓢虫(Rodolia cardinalis)、孟氏隐唇瓢虫
(Cryptolaemus montrouzieri)等天敌昆虫,继而开启
了传统生物防治的序幕[13]。1979 年从英国引入丽蚜
小蜂(Encarsia formosa)成功控制了温室白粉虱的
危害,并在北京、天津、辽宁省(市)推广[16]。而
据 Van Lenteren[17]的统计,目前全球天敌应用产
生的效益 80%集中在温室农业系统中,丽蚜小蜂
的收益占 25% ,智利小植绥螨( Phytoseiulus persimils)和黄瓜钝绥螨(Amblyseius cucumeris)的收益占 12%。近年来,中国不少学者研究了在

3466 中 国 农 业 科 学 48 卷
温室条件下释放丽蚜小蜂 [18-19]、东亚小花蝽(Orius sauteri)[20-21]、异色瓢虫(Harmonia axyridis)[22]、
巴氏钝绥螨(Amblyseius barkeri)[23]控制温室害虫
的效果(表 2)。 天敌昆虫作为生物防治的一项重要组成部分在农
业可持续发展和无公害蔬菜生产中具有明显的专业优
势[13]。在应用中主要有经典性、增强性、保护性生物
防治 3 种方式。 2.1 经典型生物防治
从国外或外地引进本地没有的天敌昆虫和有益生
物,可加强对本地害虫的控制。例如从美国和加拿大
分别引进的丽蚜小蜂和食蚜瘿蚊( Aphidoletes abietis),分别在对温室白粉虱及菜蚜(Lipaphis erysimi)的控制上显示出良好的效果[14]。1996 年从英
国引进的胡瓜钝绥螨(Amblyseius cucumeris)可有效
控制蓟马、螨类的危害,2006 年在新疆生产建设兵团
应用面积达到 9 000 hm2[6]。近年来对于引入式天敌的
安全评价成为生物防治的热点,因此提倡外来引入式
天敌与本地天敌混合防治成为趋势。将从美国引进的
海氏桨角蚜小蜂(Eretmocerus hayati)和浅黄恩蚜小
蜂(Encarsia sophia)在田间笼罩中组合释放,比单独
释放对烟粉虱的控制效果好,两者之间的竞争反而增
强了生物防治效果[24]。 2.2 增强型生物防治
人工大量繁殖天敌,接种式释放到田间,促进已
有天敌的控害作用,是目前设施蔬菜生产中天敌应用
主要的生物防治模式。中国成功利用替代猎物粉螨
等饲养捕食螨防治叶螨、蓟马等小型刺吸性害虫[25]。
此外,温室害虫生长周期短、种群结构复杂,单一天
敌释放很难控制在经济阈值以下[26-27],利用多种天敌
复合释放技术已经成为现代生物防治应用体系中的研
究重点。 2.3 保护型生物防治
近年来基于物种多样性和稳定性等生态理论,利
用辅助性助迁功能植被对生境进行调控使天敌能够稳
定增殖起到增效作用,生态调控的综合治理开始受到
较多关注[28-29]。大量实例证明,通过适宜增加植被多
样化、周边生境多样性,或为天敌提供食物、补充寄
主和补充营养;或有助于改善天敌栖息环境、越冬场
所、休息地或产卵场所,为天敌提供逃避农药施用、
耕作干扰等恶劣条件的庇护场所,利于天敌的生存和
繁衍;提高天敌昆虫群落多样性的抗逆性和补偿能力,
增强天敌昆虫对害虫的生态控制能力[30-33]。依据维持
和增强天敌的不同功能可分为蜜源植物、储蓄植物、
栖境植物、诱集植物等。在生产前期增加储蓄植物,
引入替代猎物及植物,为天敌提供食物使其提前在温
室中定殖。在生产后期主栽蔬菜上害虫种群数量较低
时,引入诱集植物或蜜源植物等,诱集天敌,为其提
供补充营养,减少天敌因食物短缺死亡的损失。同时
应用天敌防控时,除应考虑更加合理的益害比,维持
二者的低密度下的平衡外,还需要研发天敌田间保育、
诱集回收再利用等技术模式,探索增加天敌植物支持
系统、调整天敌释放策略等技术措施,降低应用成本,
提高控害效益。 国际上将储蓄植物(banker plant)作为一种天敌
增殖和释放方法在害虫防治中应用已经有近 40 年的
历史,不同时期的系统累积百余种,多种天敌如寄生
蜂、小花蝽、捕食螨等均有应用,防治害虫主要为蚜
虫、粉虱和蓟马等[34],在欧洲和加拿大主要天敌公司
均提供储蓄植物产品[35]。近年来,中国在果园、小麦
等大田作物生态系统中增加功能性植物,建立和保护
生态环境,提高有益昆虫多样性,为害虫生态调控提
供保障。而设施蔬菜生产方面的应用研究相对局限。
“木瓜载体植物系统”防治烟粉虱[36]、“玉米载体植
物系统”防治二斑叶螨[37]、“蓖麻载体植物系统”防
治烟粉虱[38]等储蓄植物系统已取得试验成功。此外,
蜜源植物、栖境植物、诱集植物在设施蔬菜生产中的
应用还在研发中。
3 设施蔬菜害虫的主要天敌昆虫及
其应用
天敌昆虫作为传统的生物防治产品,在控制设施
蔬菜虫(螨)害,保证其产量和品质中起着不可替代
的作用。随着人们环境保护意识的加强和绿色农业的
发展,天敌昆虫在设施蔬菜害虫生物防治中的作用越
来越受到人们的重视。中国的天敌资源非常丰富,但
目前在设施蔬菜生产中应用的种类相当有限。
3.1 主要天敌昆虫资源
表 2 中列出了目前中国设施蔬菜害虫主要天敌资
源种类,蚜虫天敌主要有捕食性瓢虫、小花蝽、寄生
性天敌蚜茧蜂和卵形异绒螨(Allothrombium ovatum)。
粉虱类害虫的天敌主要有捕食性瓢虫、草蛉、小花蝽
和寄生蜂等。蔬菜害虫叶螨的天敌主要以捕食螨为主,
捕食性瓢虫对叶螨也具有一定的控制效果。菜蛾类害
虫的天敌以寄生性天敌为主。

17 期 张帆等:中国设施蔬菜害虫天敌昆虫应用研究进展 3467
表 2 中国设施蔬菜害虫天敌昆虫资源种类
Table 2 The main natural enemies of greenhouse vegetables in China
靶标害虫 Target pest insect 天敌昆虫 Natural enemies 参考文献 Reference
卵形异绒螨 A. ovatum Dong, 2001[39]
多异瓢虫 Adonia variegata 杨芳等,2007[40]
异色瓢虫 H. axyridis 郭丽娜等,2008[41]
六斑月瓢虫 M. sexmaculatus 吴红胜等,2010[42]
龟纹瓢虫 Propylea japonica Zhang 等,2007[43]
大突肩瓢虫 Synonycha grandis 欧善生,2008[44]
南方小花蝽 Orius similis 周兴苗等,2002[45]
烟蚜茧蜂 Aphidius gifuensis 吴兴富, 2007[46]
蚜虫 Aphid
菜少脉蚜茧蜂 Diaeretiella rapae 余明恩等,1993[47]
异色瓢虫 H. axyridis 沈平等,2009[48]
龟纹瓢虫 P. japonica Lin 等,2008[49]
小黑粉虱瓢虫 Delphastus catalinae Lin 等,2008[49];罗宏伟,2005[50]
六斑月瓢虫 M. sexmaculatus 吴红胜等,2010[42]
粉虱小毛瓢虫 Nephaspis oculatus Lin 等,2008[49]
黑襟毛瓢虫 Scymnus hoffmanni Lin 等,2008[49]
刀角瓢虫 Serangium japonicum Lin 等,2008[49]
沙巴拟刀角瓢虫 Serangiella sababensis Lin 等,2008[49]
东亚小花蝽 O. minutus 汤方等,2007[51]
烟盲蝽 Nesidiocoris tenuis 李令蕊等,2008[52]
丽草蛉 Chrysopa formosa 李志刚等,2010[53]
大草蛉 Chrysopa pallens 刘爽等,2011[54]
丽蚜小蜂 E. formosa 张世泽等,2003[55]
浅黄恩蚜小蜂 E. Sophia Xiao 等,2011[36]
粉虱 Whitefly
海氏桨角蚜小蜂 E. hayati Yang 等,2011[56];邱宝利等,2004[57]
真桑钝绥螨 Amblyseius makuwa 蒲天胜等,1991[58]
尼氏钝绥螨 Amblyseius nicholsi 胡展育等,2007[59]
芬兰真绥螨 Euseius finlandicus 关秀敏等,2002[60]
智利小植绥螨 P. persimilis 李丽娟等,2004[61]
异色瓢虫 H. axyridis 郭丽娜等,2008[41]
拟小食螨瓢虫 Stethorus parapauperculus 程立生等,1989[62]
叶螨 Spider mites
食螨瓢虫 Stethorus siphonulus 黄阿兴等,1997[63]
黄瓜钝绥螨 A. cucumeris 孙月华等,2009[64]
东亚小花蝽 O. sauteri 王恩东等,2010[23]
蓟马 Thrips
南方小花蝽 O. similis 周兴苗等,2002[45]
烟盲蝽 Nesidiocoris tenuis 李令蕊等,2008[52] 小菜蛾 Diamond back moth 黄斑粗喙椿象 Eocanthecona furcellata Zhang 等,2001[65]
3.2 应用的主要天敌昆虫
3.2.1 蚜虫天敌 中国设施蔬菜生产中,危害其产量
和品质的蚜虫以桃蚜为主。目前,捕食性瓢虫、食蚜
蝇、蚜茧蜂和食蚜瘿蚊是对设施蔬菜蚜虫防治时释放
的主要天敌昆虫[1]。对设施蔬菜蚜虫控制效果较好的
捕食性瓢虫有异色瓢虫、多异瓢虫、七星瓢虫
(Coccinella septempunctata)和龟纹瓢虫[43,66]。其中,
具有多色型的异色瓢虫在生物防治起着主导作用,它

3468 中 国 农 业 科 学 48 卷
不仅可以取食桃蚜,还可以取食梨二叉蚜(Toxoptera piricola)、桃大尾蚜(Hyaloperus arundinis)和棉蚜
(Aphis gossypii)[67]。利用异色瓢虫可以成功防治温
室黄瓜和草莓上的蚜虫[68-69]。近期研究显示,释放异
色瓢虫对北京温室甜椒和圆茄上的桃蚜均可以达到较
高的防效[22]。食蚜蝇是双翅目中相对较大的类群,对
蚜虫的捕食能力很强,部分种类的食蚜蝇还能捕食粉
虱、飞虱和蚧壳虫等害虫 [70]。例如黑带食蚜蝇
(Episyrphus balteatus)、大灰食蚜蝇(Eupeodes corolla)的幼虫对桃蚜均具有较强的捕食能力[71]。1986年,中国从加拿大引进烟蚜茧蜂后,分别在北京市、
河北省和福建省的温室中释放用来防治蚜虫。上海市
和辽宁省的相关研究人员用桃蚜饲养烟蚜茧蜂,可以
成功防治番茄、黄瓜和辣椒上的桃蚜和棉蚜[46,72-73]。
近期研究表明,烟蚜茧蜂和异色瓢虫混合释放对烟蚜
的防效高于单一释放[74]。将食蚜瘿蚊按益害比 1﹕20的比例释放 10 d 后,蚜虫的种群密度可以减少 70%—
90%[75]。 3.2.2 粉虱天敌 粉虱的天敌种类繁多,包括寄生性
天敌、捕食性天敌和虫生真菌等。目前,在温室中释
放寄生性天敌昆虫可以对粉虱起到很好的控制效果。
目前,对烟粉虱和温室白粉虱有效的寄生性天敌有 27种均属于蚜小蜂科(Aphelinidae),其中有 21 种属于
恩蚜小蜂属(Encarsia),6 种属于桨角蚜小蜂属
(Eretmocerus)[76]。这些寄生性天敌中丽蚜小蜂在防
治温室粉虱中取得了显著的成果。中国于 1978 年从英
国引进丽蚜小蜂,随后中国农业科学院生物防治研究
所和蔬菜花卉研究所研究人员对其生物学特性和应用
方面进行了深入研究并研发了烟草苗繁蜂法[77]。随后
在河北、辽宁、山东、内蒙古等省(市)释放用来防
治温室白粉虱。丽蚜小蜂防治烟粉虱的效果较好,可
在设施蔬菜生产中大面积推广应用[78]。近期研究结
果显示,丽蚜小蜂和东亚小花蝽混合释放对烟粉虱的
防治效果高于单独释放[79]。除丽蚜小蜂以外,双斑
恩蚜小蜂(Encarsia bimaculata)、浅黄恩蚜小蜂和
裸盾恩蚜小蜂(Encarsia aseta)也是防治温室白粉虱
的重要寄生性天敌[76]。防治粉虱的捕食性天敌主要
包括小黑瓢虫(Delphastus cataliane)、刀角瓢虫、
草 蛉 Chrysoperla rufilabris 及 盲 蝽 Macrolophus caliginosus[80]。以捕食性瓢虫为主,例如从国外引进
的刀角瓢虫、沙巴拟刀角瓢虫和小黑粉虱瓢虫对温室
蔬菜上的粉虱均可以起到很好的控制效果[49]。近期研
究结果表明捕食螨也可以用于对烟粉虱的生物防治
中,例如在甜椒温室释放胡瓜钝绥螨对烟粉虱的控制
效果可以达到 94%[81]。 3.2.3 叶螨天敌 中国于 20 世纪 70 年代起开始对
叶螨进行生物防治的研究工作,取得了一定的成效。
叶螨的天敌可以分为捕食性天敌和寄生性天敌两
类。1975 年,从瑞典引入智利小植绥螨,此后通过
利用引入种和本土优势的捕食螨成功地防治了这些
害虫。在这些本土优势种中,长毛钝绥螨(Amblyseius longispinosus ) 和 拟 长 毛 钝 绥 螨 ( Amblyseius pseudolongispinosus)已经广泛应用控制朱砂叶螨。智
利小植绥螨是当前害螨生物防治中 有效的捕食螨,
已成功应用于温室蔬菜、热带水果和观赏园艺植物的
生物防治中[82]。例如,将长毛钝绥螨按益害比 1﹕100的比例释放,3 周后茄子上二斑叶螨的数量显著降低[14]。
拟长毛钝绥螨是叶螨的专性捕食性天敌,可以有效控
制冬瓜上二斑叶螨的数量[83]。捕食性瓢虫也可以用来
控制叶螨,深点食螨瓢虫(Stethorus punctillum)和腹
管食螨瓢虫(Stethorus siphonulus)可以有效地控制温
室中的柑橘全爪螨(Panonychus citri)[63,84]。拟小食
螨瓢虫可以协助智利小植绥螨控制朱砂叶螨[62]。 3.2.4 蓟马天敌 目前,蓟马的捕食性天敌主要包括
食虫蝽、草蛉和捕食螨[85-86]。研究结果显示释放胡瓜钝
绥螨对日光大棚甜椒上西花蓟马的控制效果可达
86.7%[87]。释放巴氏钝绥螨可以控制温室茄子上的西花
蓟马高峰期的数量[23]。将巴氏钝绥螨和剑毛帕厉螨
(Stratiolaelaps scimitus)混合释放对彩椒上蓟马的防效
达到 47.16%[88]。将捕食螨和食虫蝽混合释放后可以提
高对西花蓟马的防控效果[89]。半翅目花蝽科小花蝽属
东亚小花蝽对蓟马也具有很强的控制能力[90]。例如在茄
子生产过程中释放东亚小花蝽,对蓟马的控制效果可以
达到 94.46%[21]。
4 天敌昆虫的人工繁殖
天敌昆虫大量释放的首要前提是在一定的时间内
积累足够数量的个体,因此天敌昆虫的规模化生产一
直以来都是害虫生物防治应用中的关键性环节[91],也
是制约以天敌昆虫应用为基础的生物防治成败的重要
因素[92]。 4.1 人工饲料
目前,绝大多数天敌昆虫的繁殖仍采用自然寄主,
造成其规模化生产受到生产周期长、占用空间大和成
本高等因素的影响。因此,天敌昆虫人工饲料的制作
已成为其大规模扩繁的重要关键技术,且人工饲料对

17 期 张帆等:中国设施蔬菜害虫天敌昆虫应用研究进展 3469
于规模化生产流程是至关重要的。 早期研究表明,花粉等水溶性植物类物质作为人
工饲料在小花蝽的生长过程中起到了显著的作用。此
外,用于大量饲养小花蝽的人工饲料也包括动物的内
脏组织、昆虫的细胞系以及半纯卵黄蛋白。尽管上述
研究较为充分,但是通过评价从而寻找到相对 优的
人工饲料仍是一个未解难题。Tan 等[93]通过试验发现,
通过研制并使用一种微胶囊技术可以解决东亚小花蝽
人工饲料存在的缺点。目前,赤眼蜂[94]、捕食性瓢虫
如七星瓢虫[95]、孟氏隐唇瓢虫[96]、异色瓢虫[97]、龟纹
瓢虫[98]、黑缘红瓢虫(Chilocorus rubidus)[99]等、大
草蛉[100-101]和捕食性蝽类[102]、捕食螨[103]等天敌昆虫的
人工饲料研究已相当成熟,广泛应用于规模化生产。 4.2 规模化生产
中国天敌昆虫规模化扩繁技术研究已取得显著
成就,国内主要天敌昆虫人工饲养开始走向规模化、
商品化。自 20 世纪 80 年代以来,先后获得“赤眼蜂
的应用基础、工厂化中试生产新工艺及大面积防治示
范的建立、改进米蛾饲养技术研究”、“草蛉大量饲
养繁殖技术研究”、“丽蚜小蜂防治温室白粉虱的研
究”等多项科研成果,这为天敌昆虫商品化生产和应
用的新技术和新工艺的开发奠定了良好的基础[92,104]。
到目前为止,已能成功地规模化饲养赤眼蜂、丽蚜小
蜂、七星瓢虫、小花蝽、捕食螨等捕食性或寄生性天
敌昆虫。北京市农林科学院植物保护环境保护研究所
天敌昆虫研究室成功地研制出了利用柞蚕卵、米蛾卵、
人造卵繁殖多种赤眼蜂的技术与工艺流程,研制出了
规模化繁殖天敌昆虫的多套机械化及半机械化生产机
械,并已成功建立生产线。此外,研发的异色瓢虫饲
养技术,可以年产异色瓢虫 200 万头左右(王甦等,
未发表)。 但中国的天敌昆虫产业化还处在起步阶段, 总体
上讲生产规模较小、产品单一,受季节性影响,技术
服务滞后。天敌昆虫的生产大多依附于有关大专院校、
科研单位和技术推广部门。与发达国家相比,中国天
敌昆虫生产及应用的技术手段和天敌种类的多样性方
面差距还较大,无论是天敌的种类,还是天敌的生产
规模,都不能满足农业生产精品化、无公害化趋势的
要求。
5 问题与展望
设施栽培方式为人类带来了更为丰富的蔬菜产
品,同时其特殊的环境也加重了昆虫的发生与危害,
长期的化学防治已引起了害虫的抗药性、农药残留等
问题。毫无疑问,生物防治在温室病虫害的综合治理
中逐渐会上升至重要地位,利用天敌昆虫捕食或寄生
害虫在温室中的应用虽已取得了一定成效,但还有很
多问题亟待解决。 5.1 天敌保护利用
通过建设农业生态景观结构,增加植被多样性,
从而提高农业生态景观自身的病虫害防治能力,是近
年来新兴的生物防治应用辅助策略之一。但针对景观
植被结构及规模管理对天敌昆虫适生性及群落动态影
响的研究还十分有限。随着农业生产向自动化、精细
化和量化调控等方面的飞跃式发展,如何尽量减少植
物保护投入品(包括天敌昆虫),而通过主动调节生
态系统所依托的自然和农业景观来实现以生物防治为
核心的现代绿色循环型植物保护体系,才是我们今后
为之投入与发展的目标。如何设计农业生态群落的结
构,加入功能性植被景观,并且顺应害虫与天敌互作
的基本规律,发挥保护天敌昆虫与抑制单一害虫暴发
成灾的作用,将会是未来保护型生物防治研究中的核
心问题。 欧美国家已经建立以天敌昆虫为核心的生物防治
综合生态功能体系。众多生态因子,如辅助植物多样
性管理、土壤腐殖质的调控、温室及保护地的量化防
治策略等均作为生物防治的增强因素而加以整合。因
此,如何针对中国现有的农业生态系统结构,利用本
地丰富的天敌资源,开发研究出适于本地应用的多层
次复合量化“订制型”生物防治新模式,已经是一个
刻不容缓的问题。 5.2 天敌释放技术
温室害虫生长周期短、种群结构复杂,单一天敌
释放很难控制在经济阈值以下,尽量利用占据不同生
态位的天敌组合,多种天敌复合释放技术已经成为现
代生物防治应用体系中的研究热点。随着提高综合生
态效应与保护农业生态安全逐步成为农业植物保护工
作的基准,人们开始着眼于针对指定的靶标农业生态
系统,而不是单一作物上的单一害虫开展天敌昆虫的
选择与释放工作。通过这种释放方式,不但可以形成
“天敌+害虫”的稳定生态群落,并且可以针对不同
时期的优势害虫,自我调节天敌群落的结构规模,主
动防御害虫,抑制害虫的暴发。此外,这种复合释放
方式,增加了靶标生态系统中的多样性水平,延长了
生态竞争的缓冲条件,也有效地减少了天敌昆虫种群
发展后如非靶标作用等的生态风险。

3470 中 国 农 业 科 学 48 卷
5.3 控害效果评价
控害效果评价是害虫生物防治措施推广应用的重
要依据。然而,目前使用的方法都是建立在一些假设
的或人为设计的条件下进行评价,较少综合考虑取样、
条件控制、模型设计和天敌的行为学等多方面的因素。
通过开展针对天敌昆虫实际防控效果的评价,可以增
强对于现有生物防治应用模式及方法的认知与反馈,
及时对生物防治技术体系自身进行修正和优化,保持
和提升其对害虫的防控能力。 5.4 天敌规模化生产技术
中国天敌昆虫大量繁殖释放方面进展缓慢,与发
达国家相比差距甚大,特别是天敌的规模化繁育及与
其相关的基础研究方面的不足。天敌昆虫人工饲料的
应用也存在一些问题,其中 值得注意的是饲养昆虫
的“活力”问题。例如,连续饲养中有时种群会过早
地衰退,在大量饲养天敌进行田间释放中,其生存能
力和竞争能力不及自然种群等,随着昆虫食性和营养
研究的深入以及饲养技术的发展,人工饲料的研制也
将更加合理,饲养中所产生的过早衰退和“活力”不
强等疑难问题将逐步解决。
天敌昆虫产品的产业化需瞄准当前及未来发展可
持续农业、无公害食品必需的关键技术,并在原有生
产和应用的基础上,发展天敌昆虫扩繁的技术体系。
着眼于优势种高效天敌昆虫的大规模饲养与工厂化生
产的技术体系,对有发展潜力和良好工作基础的天敌
昆虫半人工饲料配方、规模化饲养技术及其与饲养有
关的应用基础工作(如昆虫行为学、营养学与化学生
态学),开展深入研究与技术改进,以适应工厂化生
产的需要,以期快速开发出实用的天敌扩繁技术体系,
形成天敌扩繁的规模化与产业化的模式。 References
[1] Yang N W, Zang L S, Wang S, Guo J Y, Xu H X, Zhang F, Wan F H.
Biological pest management by predators and parasitoids in the
greenhouse vegetables in China. Biological Control, 2014, 68:
92-102.
[2] 喻景权. “十一五” 中国设施蔬菜生产和科技进展及其展望. 中国
蔬菜, 2011(2): 11-23.
Yu J Q. Progress in protected vegetable production and research
during ‘the eleventh five-year plan’ in China. China Vegetables,
2011(2): 11-23. (in Chinese)
[3] http://news.xinhuanet.com/fortune/2014-04/01/c_1110051027.htm.
[4] 张真和, 陈青云, 高丽红, 王娟娟, 沈军, 李中民, 张雪艳. 中国设
施蔬菜产业发展对策研究(上). 蔬菜, 2010(5): 1-3.
Zhang Z H, Chen Q Y, Gao L H, Wang J J, Shen J, Li Z M, Zhang X Y.
Development tendency and countermeasures of vegetable protected
production in China (Ⅰ). Vegetables, 2010(5): 1-3. (in Chinese)
[5] 张真和, 陈青云, 高丽红, 王娟娟, 沈军, 李中民, 张雪艳. 中国设
施蔬菜产业发展对策研究 (下). 蔬菜, 2010(6): 1-3.
Zhang Z H, Chen Q Y, Gao L H, Wang J J, Shen J, Li Z M, Zhang X Y.
Development tendency and countermeasures of vegetable protected
production in China (Ⅱ). Vegetables, 2010(6): 1-3. (in Chinese)
[6] 任顺祥, 陈学新. 生物防治. 北京: 中国农业出版社, 2011.
Ren S X, Chen X C. Biological Control. Beijing: China Agriculture
Press, 2011. (in Chinese)
[7] 吕要斌, 张治军, 吴青君, 杜予州, 张宏瑞, 于毅, 王恩东, 王鸣华,
王满囷, 童晓立, 吕利华, 谭新球, 付卫东. 外来入侵害虫西花蓟
马防控技术研究与示范. 应用昆虫学报, 2011, 48(3): 488-496.
Lü Y B, Zhang Z J, Wu Q J, Du Y Z, Zhang H R, Yu Y, Wang E D,
Wang M H, Wang M K, Tong X L, Lü L H, Tan X Q, Fu W D.
Research progress of the monitoring, forecast and sustainable
management of invasive alien pest Frankliniella occidentalis in China.
Chinese Journal of Applied Entomology, 2011, 48(3): 488-496. (in
Chinese)
[8] 黄保宏, 林桂坤, 王学辉, 张先锋. 防虫网对设施蔬菜害虫控害作
用研究. 植物保护, 2013, 39(6): 164-169.
Huang B H, Lin G K, Wang X H, Zhang X F. Control effects of fly
nets to the pests of vegetables in greenhouses. Plant Protection, 2013,
39(6): 164-169. (in Chinese)
[9] Emden H F, Eastop V F, Hughes R D, Way M J. The ecology of
Myzus persicae. Annual Review of Entomology, 1969, 14(1): 197-270.
[10] 宫亚军, 王泽华, 石宝才, 康总江, 朱亮, 郭晓军, 刘建华, 魏书军.
北京地区不同桃蚜种群的抗药性研究 . 中国农业科学 , 2011,
44(21): 4385-4394.
Gong Y J, Wang Z H, Shi B C, Kang Z J, Zhu L, Guo X J, Liu J H,
Wei S J. Resistance status of Myzus persicae (Sulzer) (Hemiptera:
Aphididae) populations to pesticide in Beijing. Scientia Agricultura
Sinica, 2011, 44(21): 4385-4394. (in Chinese)
[11] 蔡仁莲, 郭建军, 金道超. 蔬菜叶螨发生特点及其生物防治的研究
进展. 贵州农业科学, 2014, 41(1): 81-86.
Cai R L, Guo J J, Jin D C. Review on occurrence characteristics and
biocontrol of vegetable pest mites. Guizhou Agricultural Sciences,
2014, 41(1): 81-86. (in Chinese)
[12] 吴青君, 徐宝云, 张友军, 张治军, 朱国仁. 西花蓟马对不同颜色
的趋性及蓝色粘板的田间效果评价 . 植物保护 , 2007, 33(4):
103-105.

17 期 张帆等:中国设施蔬菜害虫天敌昆虫应用研究进展 3471
Wu Q J, Xu B Y, Zhang Y J, Zhang Z J, Zhu G R. Taxis of western
flower thrips to different colors and field efficacy of the blue sticky
cards. Plant Protection, 2007, 33(4): 103-105. (in Chinese)
[13] 张帆, 张君明, 罗晨, 王甦. 蔬菜害虫的生物防治技术概述. 中国
蔬菜, 2011(1): 23-24.
Zhang F, Zhang J M, Luo C, Wang S. A summary of biological control
of vegetable pest. China Vegetables, 2011(1): 23-24. (in Chinese)
[14] 田毓起. 蔬菜害虫生物防治. 北京: 金盾出版社, 2000.
Tian Y Q. Biological Control of Vegetable Pests. Beijing: Jindun
Press, 2000. (in Chinese)
[15] 包建中 , 古德祥. 中国生物防治. 太原: 山西科学技术出版社 ,
1998.
Bao J Z, Gu D X. Biological Control in China. Taiyuan: Shanxi
Science and Technology Publishing House, 1998. (in Chinese)
[16] 程洪坤, 田毓起, 魏炳传. 丽蚜小蜂商品化生产技术. 生物防治通
报, 1989, 5(4): 178-181.
Cheng H K, Tian Y Q, Wei B C. A technique for commercial-scale
production of Encarsia formosa. Chinese Journal of Biological
Control, 1989, 5(4): 178-181. (in Chinese)
[17] van Lenteren J C. Internet Book of Biological Control. www.IOBC-
Global.org. Wageningen, The Netherlands, 2007.
[18] 张世泽. 丽蚜小蜂 Encarsia formosa 生态学及其对烟粉虱 Bemisia
tabaci 控制效能的研究[D]. 杨凌: 西北农林科技大学, 2003.
Zhang S Z. Ecology of two Encarsia formosa strains and their control
efficacy on tobacco whitefly, Bemisia tabaci[D]. Yangling: Northewest
Sci-Tech University of Agriculture and Forestry, 2003. (in Chinese)
[19] 张君明, 张帆, 王兵. 以释放丽蚜小蜂为主的保护地番茄温室粉虱
的控制技术. 蔬菜, 2010(7): 34-35.
Zhang J M, Zhang F, Wang B. Control effects of releasing density and
time of Encarsia formosa on Trialeurodes vaporariorum. Vegetables,
2010(7): 34-35. (in Chinese)
[20] 蒋月丽, 武予清, 段云, 高新国. 释放东亚小花蝽对大棚辣椒上几
种害虫的防治效果. 中国生物防治学报, 2011, 27(3): 414-417.
Jiang Y L, Wu Y Q, Duan Y, Gao X G. Control efficiencies of
releasing Orius sauteri (Heteroptera: Anthocoridae) on some pests in
greenhouse pepper. Chinese Journal of Biological Control, 2011,
27(3): 414-417. (in Chinese)
[21] 尹健, 高新国, 武予清, 蒋月丽, 刘顺通, 段爱菊, 张自启, 刘长营.
释放东亚小花蝽对茄子上蓟马的控制效果. 中国生物防治学报,
2013, 29(3): 459-462.
Yin J, Gao X G, Wu Y Q, Jiang Y L, Liu S T, Duan A J, Zhang Z Q,
Liu C Y. Thrips control on the greenhouse eggplant by releasing Orius
sauteri (Heteroptera: Anthocoridae). Chinese Journal of Biological
Control, 2013, 29(3): 459-462. (in Chinese)
[22] 李姝, 王甦, 赵静, 杨丽文, 高希武, 张帆. 释放异色瓢虫对北京
温室甜椒和圆茄上桃蚜的控害效果. 植物保护学报, 2014, 41(6):
699-704.
Li S, Wang S, Zhao J, Yang L W, Gao X W, Zhang F. Efficacy of
multicolored Asian lady beetle Harmonia axyridis (Coleoptera:
Coccinellidae) against green peach against green peach aphid Myzus
persicae (Hemiptera: Aphididae) on vegetables under greenhouse
conditions. Acta Phytophylacica Sinica, 2014, 41(6): 699-704. (in
Chinese)
[23] 王恩东, 徐学农, 吴圣勇. 释放巴氏钝绥螨对温室大棚茄子上西花
蓟马及东亚小花蝽数量的影响. 植物保护, 2010, 36(5): 101-104.
Wang E D, Xu X N, Wu S Y. Control effects of Amblyseius barkeri on
Frankliniella occidentalis on the eggplants and their natural enemy
Orius sauteri in the greenhouse. Plant Protection, 2010, 36(5):
101-104. (in Chinese)
[24] 徐海云. 浅黄恩蚜小蜂与海氏桨角蚜小蜂的致死干扰竞争作用[D].
北京: 中国农业科学院, 2011.
Xu H Y. Lethal interference competition between Encarsia and
Eretmocerus hayati (Hymenoptera: Aphelinidae)[D]. Beijing: Chinese
Academy of Agricultural Sciences, 2011. (in Chinese)
[25] 曾凡荣, 陈红印. 天敌昆虫饲养系统工程. 北京: 中国农业科学技
术出版社, 2009.
Zeng F R, Chen H Y. The System Engineering for Rearing Insect
Natural Enemies. Beijing: China Agricultural Science and Technology
Press, 2009. (in Chinese)
[26] Bouyer J, Seck M T, Sall B, Ndiaye E Y, Guerrini L, Vreysen M J.
Stratified entomological sampling in preparation for an area-wide
integrated pest management program: the example of Glossina
palpalis gambiensis (Diptera: Glossinidae) in the Niayes of Senegal.
Journal of Medical Entomology, 2010, 47(4): 543-552.
[27] Wilby A, Thomas M B. Natural enemy diversity and pest control:
patterns of pest emergence with agricultural intensification. Ecology
Letters, 2002, 5(3): 353-360.
[28] Cook S M, Khan Z R, Pickett J A. The use of push-pull strategies in
integrated pest management. Annual Review of Entomology, 2007,
52(1): 375-400.
[29] 陈学新, 刘银泉, 任顺祥, 张帆, 张文庆, 戈峰. 害虫天敌的植物
支持系统. 应用昆虫学报, 2014, 51(1): 1-12.
Chen X X, Liu Y Q, Ren S X, Zhang F, Zhang W Q, Ge F.
Plant-mediated support system for natural enemies of insect pests.
Chinese Journal of Applied Entomology, 2014, 51(1): 1-12. (in
Chinese)

3472 中 国 农 业 科 学 48 卷
[30] Landis D A, Wratten S D, Gurr G M. Habitat management to conserve
natural enemies of arthropod pests in agriculture. Annual Review of
Entomology, 2000, 45: 175-201.
[31] Heimpel G E, Jervis M A. Does floral nectar improve biological
control by parasitoids//Plant-Provided Food for Carnivorous Insects:
a Protective Mutualism and Its Applications. New York: Cambridge
University Press, 2005: 267-304.
[32] 魏永平, 花蕾, 张雅林. 生境调节对苹果园害虫可持续控制作用.
中国农学通报, 2004, 20(1): 204-206.
Wei Y P, Hua L, Zhang Y L. Sustainable control technique of
protecting natural enemies by habitat regulation. Chinese Agricultural
Science Bulletin, 2004, 20(1): 204-206. (in Chinese)
[33] 肖英方, 毛润乾, 万方浩. 害虫生物防治新概念——生物防治植物
及创新研究. 中国生物防治学报, 2013, 29(1): 1-10.
Xiao Y F, Mao R Q, Wan F H. New concept of biological control:
bio-control plants used for management of Arthropod pests. Chinese
Journal of Biological Control, 2013, 29(1): 1-10. (in Chinese)
[34] Frank S D. Biological control of arthropod pests using banker plant
systems: Past progress and future directions. Biological Control, 2010,
52(1): 8-16.
[35] Huang N X, Enkegaard A, Osborne L S, Ramakers P M J, Messelink
G J, Pijnakker J, Murphy G. The banker plant method in biological
control. Critical Reviews in Plant Sciences, 2011, 30(3): 259-278.
[36] Xiao Y F, Chen J J, Cantliffe D, Mckenzie C, Houben K, Osborne L S.
Establishment of papaya banker plant system for parasitoid, Encarsia
sophia (Hymenoptera: Aphilidae) against Bemisia tabaci (Hemiptera:
Aleyrodidae) in greenhouse tomato production. Biological Control,
2011, 58(3): 239-247.
[37] Xiao Y F, Osborne L, Chen J J, Mckenzie C, Houben K, Irizarry F.
Evaluation of corn plant as potential banker plant for supporting
predatory gall midge, Feltiella acarisuga (Diptera: Cecidomyiidae)
against Tetranychus urticae (Acari: Tetranychidae) in greenhouse
vegetable production. Crop Protection, 2011, 30(12): 1635-1642.
[38] Huang H, Zhao H, Zhang Y M, Zhang S Z, Liu T X. Influence of
selected host plants on biology of castor whitefly, Trialeurodes ricini
(Hemiptera: Aleyrodidae). Journal of Asia-Pacific Entomology, 2014,
17(4): 745-751.
[39] Dong Y C. Laboratory experiments on the effect of Allothrombium
ovatum larvae (Acari: Thrombidiidae) on the aphid Rhopalosiphum
padi (Homoptera: Aphididae). Systematic and Applied Acarology,
2001, 6: 61-64.
[40] 杨芳, 王芳, 张蓉, 贺达汉. 小十三星瓢虫对苜蓿斑蚜捕食功能反
应的研究. 草业科学, 2007, 24(10): 80-84.
Yang F, Wang F, Zhang R, He D H. Study on predatory function of
Adonia variegata on Therioaphis trifolii. Pratacultural Science, 2007,
24(10): 80-84. (in Chinese)
[41] 郭丽娜, 王洪平. 异色瓢虫对麦蚜和红蜘蛛的捕食功能反应. 河南
农业科学, 2008(4): 72-74.
Guo L N, Wang H P. The functional response of Harmonia axyridis
(Pallas) to wheat aphids and Tetranychidae. Journal of Henan
Agricultural Sciences, 2008(4): 72-74. (in Chinese)
[42] 吴红胜, 韩诗畴, 张宇宏, 李志刚, 郑基焕. 双带盘瓢虫与六斑月
瓢虫成虫对螺旋粉虱幼虫的捕食作用. 环境昆虫学报, 2010, 32(3):
380-383.
Wu H S, Han S C, Zhang Y H, Li Z G, Zheng J H. Predation of
Lemnia buplagiata and Menochilus sexmaculatus adults to larvae of
Aleurodicus dispersus. Journal of Environmental Entomology, 2010,
32(3): 380-383. (in Chinese)
[43] Zhang S Z, Zhang F, Hua B Z. Suitability of various prey types for the
development of Propylea japonica (Coleoptera: Coccinellidae).
European Journal of Entomology, 2007, 104: 149-152.
[44] 欧善生. 大突肩瓢虫成虫对桃蚜的捕食作用. 安徽农业科学, 2008,
36(3): 974-975.
Ou S S. Predation of Synonycha grandis (Thunberg) adult on peach
aphid. Journal of Anhui Agricultural Sciences, 2008, 36(3): 974-975.
(in Chinese)
[45] 周兴苗, 雷朝亮. 南方小花蝽对不同猎物捕食作用及利用效率. 生
态学报, 2002, 22(12): 2085-2090.
Zhou X M, Lei C L. Utilization efficiency and functional response of
Orius similis (Hemiptera: Anthocoridae) to different preys. Acta
Ecologica Sinica, 2002, 22(12): 2085-2090. (in Chinese)
[46] 吴兴富 . 烟蚜茧蜂繁殖利用概述 . 中国农学通报 , 2007, 23(5):
306-308.
Wu X F. Summarize on the using and reproducing of Aphidius
gifuensis (Aphidiidae). Chinese Agricultural Science Bulletin, 2007,
23(5): 306-308. (in Chinese)
[47] 余明恩, 古德就, 张维球. 温度对菜蚜茧蜂行为功能反应影响的研
究. 华南农业大学学报, 1993, 14(4): 20-25.
Yu E M, Gu D J, Zhang W Q. The effects of temperature on the
foraging behavior of parasitoid Diaeretiella rapae (Hym., Braconidae).
Journal of South China Agricultural University, 1993, 14(4): 20-25.
(in Chinese)
[48] 沈平, 常承秀, 朱惠英, 王新东, 田炜, 张永强, 韩淑琴. 异色瓢虫
成虫对槐豆木虱的捕食作用. 植物保护, 2009, 35(1): 66-69.
Shen P, Chang C X, Zhu H Y, Wang X D, Tian W, Zhang Y Q, Han S
Q. Predation of Cyamophila willeti by the adults of Harmonia axyridis.

17 期 张帆等:中国设施蔬菜害虫天敌昆虫应用研究进展 3473
Plant Protection, 2009, 35(1): 66-69. (in Chinese)
[49] Lin K J, Wu K M, Zhang Y J, Guo Y Y. Naturally occurring
population of Bemisia tabaci, biotype B and associated natural
enemies in agroecosystem in northern China. Biocontrol Science and
Technology, 2008, 18: 169-182.
[50] 罗宏伟. 小黑瓢虫人工繁殖技术的研究[D]. 福州: 福建农林大学,
2005.
Luo H W. Studies on the artificial rearing technology of Delphastus
catalinae (Horn)[D]. Fuzhou: Fujian Agriculture and Forestry
University, 2005. (in Chinese)
[51] 汤方, 李生臣, 刘玉升, 王开运, 赵克思. 微小花蝽对温室白粉虱
的捕食作用. 昆虫知识, 2007, 44(5): 703-706.
Tang F, Li S C, Liu Y S, Wang K Y, Zhao K S. Predation of predatory
bug, Orius minutus on the greenhouse whitefly, Trialeurodes
vaporariorum. Chinese Bulletin of Entomology, 2007, 44(5): 703-706.
(in Chinese)
[52] 李令蕊, 薛仁风, 王晓婧, 曾凡荣. 烟盲蝽对温室白粉虱和小菜蛾
捕食作用研究. 河北农业大学学报, 2008, 31(1): 84-87.
Li L R, Xue R F, Wang X J, Zeng F R. Predation of Nesidiocoris
tenuis to Trialeurodes vaporariorum and Plutella xylostella. Journal
of Agricultural University of Hehei, 2008, 31(1): 84-87. (in Chinese)
[53] 李志刚, 郑基焕, 叶静文, 韩诗畴, 严珍, 陈泽坦. 丽草蛉对螺旋
粉虱的捕食作用. 环境昆虫学报, 2010, 32(4): 516-519.
Li Z G, Zheng J H, Ye J W, Han S C, Yan Z, Chen Z T. Predation of
the green lacewing Chrysopa formosa Brauer on the spiraling whitefly,
Aleurodicus dispersus Russell. Journal of Environmental Entomology,
2010, 32(4): 516-519. (in Chinese)
[54] 刘爽, 王甦, 刘佰明, 周长青, 张帆. 大草蛉幼虫对烟粉虱的捕食
功能反应及捕食行为观察. 中国农业科学, 2011, 44(6): 1136-1145.
Liu S, Wang S, Liu B M, Zhou C Q, Zhang F. The predation function
response and predatory behavior observation of Chrysopa pallens
larva to Bemisia tabaci. Scientia Agricultura Sinica, 2011, 44(6):
1136-1145. (in Chinese)
[55] 张世泽, 万方浩, 张帆, 花保桢. 丽蚜小蜂两个品系对烟粉虱若虫
的寄生适宜性. 中国生物防治, 2003, 19(4): 149-153.
Zhang S Z, Wan F H, Zhang F, Hua B Z. Parasitic suitability of two
strains of Encarsia formosa on Bemisia tabaci. Chinese Journal of
Biological Control, 2003, 19(4): 149-153. (in Chinese)
[56] Yang N W, Wan F H. Host suitability of different instars of Bemisia
tabaci biotype B for the parasitoid Eretmocerus hayati. Biological
Control, 2011, 59(2): 313-317.
[57] 邱宝利, 任顺祥, 林莉, 王兴民. 放蜂密度对桨角蚜小蜂 Eretmocerus
sp.控制烟粉虱效果的影响. 华东昆虫学报, 2004, 13(2): 27-30.
Qiu B L, Ren S X, Lin L, Wang X M. Effects of release density of
Eretmocerus sp. (Hymenoptera: Aphelinidae) on the control effectiveness
of Bemisia tabaci (Homoptera: Aleyrodidae). Entomological Journal
of East China, 2004, 13(2): 27-30. (in Chinese)
[58] 蒲天胜, 曾涛, 韦德卫. 20 种植物花粉对真桑钝绥螨饲养效果的综
合评判. 生物防治通报, 1991, 7(3): 111-114.
Pu T S, Zeng T, Wei D W. An integrated evaluation on 20 plant pollen
as diet for mass rearing Amblyseius makuwa (Acari: Phytoseiidae).
Chinese Journal of Biological Control, 1991, 7(3): 111-114. (in
Chinese)
[59] 胡展育, 郅军锐, 熊继文. 尼氏钝绥螨对二斑叶螨的捕食作用. 安
徽农业科学, 2007, 35(15): 4556-4557.
Hu Z Y, Zhi J R, Xiong J W. Predatory impact of Amblyseius nicholis
to Tetranychus urticae. Journal of Anhui Agricultural Sciences, 2007,
35(15): 4556-4557. (in Chinese)
[60] 关秀敏, 孙续艮. 芬兰真绥螨捕食功能的研究. 山东农业大学学报:
自然科学版, 2002, 33(3): 297-301.
Guan X M, Sun X G. A study of predation of Euseius finlandicus.
Journal of Shandong Agricultural University: Natural Science, 2002,
33(3): 297-301. (in Chinese)
[61] 李丽娟, 鲁新, 刘宏伟, 张国红. 中国智利小植绥螨的繁殖及应用
研究. 吉林农业大学学报, 2004, 26(4): 402-405, 410.
Li L J, Lu X, Liu H W, Zhang G H. Studies on reproduction and
application of Phytoseiulus persimilis in China. Journal of Jilin
Agricultural University, 2004, 26(4): 402-405, 410. (in Chinese)
[62] 程立生, 刘君成, 宋国敏. 拟小食螨瓢虫成虫对朱砂叶螨捕食作用
的研究. 热带作物学报, 1989, 10(2): 99-105.
Cheng L S, Liu J C, Song G M. Studies on predation of Stethorus
parapauperculus to Tetranychus cinnabarinus. Chinese Journal of
Tropical Crops, 1989, 10(2): 99-105. (in Chinese)
[63] 黄阿兴, 蔡衍龙. 腹管食螨瓢虫开发与利用. 华东昆虫学报, 1997,
6(2): 119-110.
Huang A X, Cai Y L. Development and utilization of Stethorus
siphonulus Kapur. Entomological Journal of East China, 1997, 6(2):
119-110. (in Chinese)
[64] 孙月华, 郅军锐, 李景柱, 袁成明. 花粉在胡瓜钝绥螨控制西花蓟
马中的作用. 山地农业生物学报, 2009, 28(3): 260-263.
Sun Y H, Zhi J R, Li J Z, Yuan C M. The role of pollen in the effect of
Amblyseius cucumeris (Acri: Phytoseiidae) on the control of western
flower thrips, Frankliniella occidentalis (Thysanoptera: Thripidae).
Journal of Mountain Agriculture and Biology, 2009, 28(3): 260-263.
(in Chinese)
[65] Zhang J B, Xie F G. The predatory effects of Eocanthecona furcellata

3474 中 国 农 业 科 学 48 卷
(Wolff) (Hemiptera: Pentatomidae) to several preys. Taiwan Insects,
2001, 21: 257-267.
[66] 侯茂林, 万方浩. 七星瓢虫成虫对烟蚜的捕食作用. 昆虫知识,
2004, 41(4): 347-350.
Hou M L, Wan F H. Predation of Myzus persicae by adults of
Coccinella septempunctata. Entomological Knowledge, 2004, 41(4):
347-350. (in Chinese)
[67] 王小艺 , 沈佐锐 . 异色瓢虫的应用研究概况 . 昆虫知识 , 2002,
39(4): 255-261.
Wang X Y, Shen Z R. Progress of applied research on multicolored
Asian ladybird beetle. Entomological Knowledge, 2002, 39(4):
255-261. (in Chinese)
[68] 孙兴全, 陈文龙, 陈志兵, 何继龙, 叶文娟. 异色瓢虫的人工饲料
及防治棚栽草莓蚜虫的初步研究. 上海农学院学报, 1996, 14(2):
133-137.
Sun X Q, Chen W L, Chen Z B, He J L, Ye W J. A preliminary study
on the artificial diet of an aphidophagous coccinellid, Harmonia
axyridis (Pallas) and its use to control strawberry aphids in the plastic
covering. Journal of Shanghai Agricultural College, 1996, 14(2):
133-137. (in Chinese)
[69] 王延鹏, 吕飞, 王振鹏. 异色瓢虫开发利用研究进展. 华东昆虫学
报, 2007, 16(4): 310-314.
Wang Y P, Lü F, Wang Z P. Progress of Harmonia axyridis (Pallas)
utilization. Entomological Journal of East China, 2007, 16(4):
310-314. (in Chinese)
[70] 兰鑫, 罗辑, 成新跃. 东北大豆生态区捕食性食蚜蝇种类及其对大
豆蚜的控害效应分析. 应用昆虫学报, 2011, 48(6): 1625-1630.
Lan X, Luo J, Cheng X Y. Control of soybean aphids by predatory
syrphids in northeast China. Chinese Journal of Applied Entomology,
2011, 48(6): 1625-1630. (in Chinese)
[71] 杨友兰, 王红武, 王全寿, 曹琳, 梁定栋, 张志勇. 山西菜田捕食
性食蚜蝇优势种及其对菜蚜的控制效果. 中国生物防治, 2002,
18(3): 124-127.
Yang Y L, Wang H W, Wang Q S, Cao L, Liang D D, Zhang Z Y.
Control aphids in vegetable fields with syrphid flies. Chinese Journal
of Biological Control, 2002, 18(3): 124-127. (in Chinese)
[72] 忻亦芬, 李学荣, 王洪平, 王桂清, 唐永红. 用萝卜苗作桃蚜植物
寄主繁殖烟蚜茧蜂. 中国生物防治, 2001, 17(2): 49-52.
Xin Y F, Li X R, Wang H P, Wang G Q, Tang Y H. Studies on the use
radish seedling to propagate peach aphid for mass rearing Aphidius
gifuensis. Chinese Journal of Biological Control, 2001, 17(2): 49-52.
(in Chinese)
[73] 邓建华, 吴兴富, 宋春满, 黄江梅, 刘光辉, 杨硕媛. 田间小棚繁
殖烟蚜茧蜂的繁蜂效果研究. 西南农业大学学报: 自然科学版,
2006, 28 (1): 66-69, 73.
Deng J H, Wu X F, Song C M, Huang J M, Liu G H, Yang S Y.
Rearing effect of Aphidius gifuensis with nylon-net covered cages in
tobacco fields. Journal of Southwest Agricultural University: Natural
Science, 2006, 28(1): 66-69, 73. (in Chinese)
[74] 王夸平, 詹莜国, 潘悦, 毛春堂, 高福宏, 孔宁川. 烟蚜茧蜂和异
色瓢虫综合防治烟蚜的效果评价. 湖北农业科学, 2013, 52(7):
1567-1570.
Wang K P, Zhan Y G, Pan Y, Mao C T, Gao F H, Kong N C.
Evaluation on the effects of releasing Aphidius gifuensis and Leis
axyridis to control Myzus persicae. Hubei Agricultural Sciences, 2013,
52(7): 1567-1570. (in Chinese)
[75] 程洪坤, 赵军华, 谢明, 魏淑贤, 宋兴甫, 王锦祯. 利用食蚜瘿蚊
防治保护地蔬菜桃蚜的试验. 生物防治通报, 1992, 8(3): 97-100.
Chen H K, Zhao J H, Xie M, Wei S X, Song X F, Wang J Z. Tests on
the effect of releasing Aphidoletes aphidimyza [Dip: Cecidomyiidae]
to control the aphid, Myzus persicae in greenhouse and plastic tunnels.
Chinese Journal of Biological Control, 1992, 8(3): 97-100. (in
Chinese)
[76] Li S J, Xue X, Ahmed M Z, Ren S X, Du Y Z, Wu J H, Cuthbertson A
G S, Qiu B L. Host plants and natural enemies of Bemisia tabaci
(Hemiptera: Aleyrodidae) in China. Insect Science, 2011, 18(1):
101-120.
[77] 朱国仁, 乔德禄, 徐宝云. 丽蚜小蜂防治白粉虱的应用技术. 中国
农学通报, 1993, 9(3): 52-53.
Zhu G R, Qiao D L, Xu B Y. The applied technology of Encarsia
formosa control Trialeurodes vaporaiorum. Chinese Agricultural
Science Bulletin, 1993, 9(3): 52-53. (in Chinese)
[78] 何笙, 吴晓云, 郑金竹, 韩振芹, 赵晨霞, 王月英, 陈卉, 陈晨, 韩
杰. 丽蚜小蜂防治设施番茄烟粉虱效果研究. 安徽农业科学, 2013,
41(14): 6244-6245, 6248.
He S, Wu X Y, Zheng J Z, Han Z Q, Zhao C X, Wang Y Y, Chen H,
Chen C, Han J. A study of control effect of Bemisia tabaci (Gennadius)
on greenhouse tomatoes using parasitoid Encarsia formosa Gahan.
Journal of Anhui Agricultural Sciences, 2013, 41(14): 6244-6245,
6248. (in Chinese)
[79] 李姝, 劳永兵, 王甦, 郭晓军, 张帆. 东亚小花蝽和丽蚜小蜂对烟
粉虱的协同控制效果研究. 环境昆虫学报, 2014, 36(6): 978-982.
Li S, Lao Y B, Wang S, Guo X J, Zhang F. Control effect of Orius
sauteri collaborated with Encarsia formosa on Bemisia tabaci in the
greenhouse. Journal of Environmental Entomology, 2014, 36(6):
978-982. (in Chinese)

17 期 张帆等:中国设施蔬菜害虫天敌昆虫应用研究进展 3475
[80] 任顺祥, 黄振, 姚松林. 烟粉虱捕食性天敌研究进展. 昆虫天敌,
2004, 26(1): 34-42.
Ren S X, Huang Z, Yao S L. Advances in studies on predators of
Bemisia tabaci gennadius. Natural Enemies of Insects, 2004, 26(1):
34-42. (in Chinese)
[81] 张艳璇, 林坚贞, 张公前, 陈霞, 季洁, 唐清. 胡瓜钝绥螨控制大
棚甜椒烟粉虱的研究. 福建农业学报, 2011, 26(1): 91-97.
Zhang Y X, Lin J Z, Zhang G Q, Chen X, Ji J, Tang Q. Research and
application of Neoseiulus cucumeris (Oudemans) for control of
Bemisia tabaci (Gennadius) on sweet pepper in plastic greenhouse.
Fujian Journal of Agricultural Sciences, 2011, 26(1): 91-97. (in
Chinese)
[82] 董慧芳, 郭玉杰, 牛离平. 应用智利小植绥螨防治温室四种花卉上
二斑叶螨的研究. 生物防治通报, 1986, 2(2): 59-62.
Dong H F, Guo Y J, Niu L P. Biological control of the two spotted
spider mites with Phytoseiulus persimilis on four ornamental plants in
greenhouse. Chinese Journal of Biological Control, 1986, 2(2): 59-62.
(in Chinese)
[83] 候爱平, 张艳璇, 杨孝泉, 陈宇航, 陈浩然. 利用长毛钝绥螨控制
冬瓜上二斑叶螨研究. 昆虫天敌, 1996, 18(1): 29-33.
Hou A P, Zhang Y X, Yang X Q, Chen Y H, Chen H R. Integrated
control of two-spotted spider mite in white ground. Natural Enemies
of Insects, 1996, 18(1): 29-33. (in Chinese)
[84] 姜廷荣, 张立本, 魏新田. 深点食螨瓢虫的生物学及其利用. 昆虫
天敌, 1982, 4(4): 34-36.
Jiang T R, Zhang L B, Wei X T. Biological characteristics and
application of Stethorus punctillum (Coleoptera: Coccinellidae).
Natural Enemies of Insects, 1982, 4(4): 34-36. (in Chinese)
[85] Castane C, Alomar O, Riudavets J. Management of western flower
thrips on cucumber with Dicyphus tamaninii (Heteroptera: Miridae).
Biological Control, 1996, 7(1): 114-120.
[86] Shipp J L, Wang K. Evaluation of Amblyseius cucumeris (Acari:
Phytoseiidae) and Orius insidious (Hemiptera: Anthocoridae) for
control of Frankliniella occidentalis (Thysanoptera: Thripidae) on
greenhouse tomatoes. Biological Control, 2003, 28(3): 271-281.
[87] 张艳璇, 单绪南, 林坚贞, 张公前, 季洁, 陈霞, 唐清. 胡瓜钝绥螨
控制日光大棚甜椒上的西花蓟马的研究与应用. 中国植保导刊,
2010, 30(11): 20-22.
Zhang Y X, Shan X N, Lin J Z, Zhang G Q, Ji J, Chen X, Tang Q. The
studies and applications of Amblyseius cucumeris (Acari:Phytoseiidae)
controlling Frankliniella occidentalis (Thysanoptera: Thripidae).
China Plant Protection, 2010, 30(11): 20-22. (in Chinese)
[88] 谷培云, 马永军, 焦雪霞, 尹哲, 郭书臣, 国洋, 郭明阁, 田学伟.
释放捕食螨对彩椒上蓟马防效的初步评价. 生物技术进展, 2013,
3(1): 54-56.
Gu P Y, Ma Y J, Jiao X X, Yin Z, Guo S C, Guo Y, Guo M G, Tian X
W. Primary evaluation of the control efficacy of releasing predatory
mites to thrips in color sweet pepper. Current Biotechnology, 2013,
3(1): 54-56. (in Chinese)
[89] 徐学农, Borgemeister C, Poehling H M. 联合释放黄瓜钝绥螨或小
花蝽和尖狭下盾螨防治西方花蓟马//迈入二十一世纪的中国生物
防治. 北京: 中国农业科学技术出版社, 2005: 35-40.
Xu X N, Borgemeister C, Poehling H M. Biocontrol of western flower
thrips Frankliniella occidentalis (Pergande) by combined releases of
plant-inhabiting predatory mite, Amblyseius cucumeris Oudemans or
bug, Orius insidious Say with soil-dwelling mite, Hypoaspis aculeifer
Canestrini//Biological Control of China in Twenty-first Century.
Beijing: China Agriculture Science and Technology Press, 2005:
35-40. (in Chinese)
[90] 张安盛, 于毅, 李丽莉, 张思聪. 东亚小花蝽(Orius sauteri)成虫对
入侵害虫西花蓟马(Frankliniella occidentalis)成虫的捕食作用. 生
态学报, 2007, 27(3): 1903-1909.
Zhang A S, Yu Y, Li L L, Zhang S C. Predation of Orius sauteri adult
on adults of western flower thrips (Frankliniella occidentalis), an
invasive insect pest. Acta Ecologica Sinica, 2007, 27(3): 1903-1909.
(in Chinese)
[91] van Lenteren J, Tommasini M. Mass production, storage, shipment
and quality control of natural enemies//Albajes R, Gullino M L, van
Lenteren J C, Elad Y. eds. Mass Production, Storage, Shipment and
Quality Control of Natural Enemies, Integrated Pest and Disease
Management in Greenhouse Crops. Springer, Netherlands, 2002:
276-294.
[92] 万方浩, 叶正楚, 郭建英, 谢明. 中国生物防治研究的进展及展望.
昆虫知识, 2000, 37(2): 65-74.
Wan F H, Ye Z C, Guo J Y, Xie M. The research progress of biological
control and prospect in our country. Entomological Knowledge, 2000,
37(2): 65-74. (in Chinese)
[93] Tan X L, Wang S, Zhang F. Optimization an optimal artificial diet for
the predatory bug Orius sauteri (Hemiptera: Anthocoridae). PLoS
ONE, 2013, 8(4): e61129.
[94] 巫之馨, 钦俊德, 李同修, 张志品, 刘德岷. 以无昆虫物质的人工
饲料培育松毛虫赤眼蜂幼虫. 昆虫学报, 1982, 25(2): 128-134.
Wu Z X, Qin J D, Li T Z, Zhang Z P, Liu D M. Culturing
Truchogramma dendrolimi in vitro with artificial media devoid of
insect materials. Acta Entomologica Sinica, 1982, 25(2): 128-134. (in
Chinese)

3476 中 国 农 业 科 学 48 卷
[95] 孙毅, 万方浩. 七星瓢虫人工饲料的研究现状及发展对策. 中国生
物防治, 1999, 15(4): 169-173.
Sun Y, Wan F H. Mass rearing Coccinella septempunctata L. (Col.:
Coccinellidae) with artificial diets: problems and substitute diets.
Chinese Journal of Biological Control, 1999, 15(4): 169-173. (in
Chinese)
[96] 庞虹, 汤才, 李丽英. 孟氏隐唇瓢虫成虫的人工饲料保种技术. 昆
虫天敌, 1996, 18(2): 64-66.
Pang H, Tang C, Li L Y. The technology for preservation of
Cryptolaemus montrouzieri adult with the artificial diets. Natural
Enemies of Insects, 1996, 18(2): 64-66. (in Chinese)
[97] 杨洪, 熊继文, 张帆. 异色瓢虫人工饲料研究进展. 山地农业生物
学报, 2003, 22(2): 169-172.
Yang H, Xiong J W, Zhang F. Advances of artificial diet for Harminia
axyridis. Journal of Mountain Agriculture and Biology, 2003, 22(2):
169-172. (in Chinese)
[98] 王利娜, 陈红印, 张礼生, 王树英, 杨海霞. 龟纹瓢虫幼虫人工饲
料的研究. 中国生物防治, 2008, 24(4): 306-311.
Wang L N, Chen H Y, Zhang L S, Wang S Y, Yang H X. Improved
formulations of an artificial diet for larvae of Propylea japonica.
Chinese Journal of Biological Control, 2008, 24(4): 306-311. (in
Chinese)
[99] 黄保宏, 邹运鼎, 毕守东, 郭义. 黑缘红瓢虫成虫人工饲料研究.
中国农业大学学报, 2005, 10(3): 4-9.
Huang B H, Zou Y D, Bi S D, Guo Y. Artificial diet for adults of
Chilocorus rubidus. Journal of China Agricultural University, 2005,
10(3): 4-9. (in Chinese)
[100] 林美珍, 陈红印, 王树英, 仝赞华. 大草蛉幼虫人工饲料的研究.
中国生物防治, 2007, 23(4): 316-321.
Lin M Z, Chen H Y, Wang S Y, Tong Z H. Development of artificial
diet for Chrysopa pallens larva. Chinese Journal of Biological Control,
2007, 23(4): 316-321. (in Chinese)
[101] 党国瑞, 张莹, 陈红印, 张礼生, 王孟卿, 刘晨曦. 人工饲料对大
草蛉生长发育和繁殖力的影响 . 中国农业科学 , 2012, 45(23):
4818-4825.
Dang G R, Zhang Y, Chen H Y, Zhang L S, Wang M Q, Liu C X.
Effect of different artificial diets on the development and fecundity of
the green lacewing Chrysopa pallens (Rambur). Scientia Agricultura
Sinica, 2012, 45(23): 4818-4825. (in Chinese)
[102] 刘丰姣, 曾凡荣. 捕食性蝽类昆虫人工饲料研究进展. 中国生物防
治学报, 2013, 29(2): 294-300.
Liu F J, Zeng F R. Advances in artificial diet for rearing of predatory
bugs. Chinese Journal of Biological Control, 2013, 29(2): 294-300.
(in Chinese)
[103] 徐学农, 吕佳乐, 王恩东. 国际捕食螨研发与应用的热点问题及启
示. 中国生物防治学报, 2013, 29(2): 163-174.
Xu X N, Lü J L, Wang E D. Hot spots in international predatory mite
studies and lessons to us. Chinese Journal of Biological Control, 2013,
29(2): 163-174. (in Chinese)
[104] 万方浩, 王韧, 叶正楚. 中国天敌昆虫产品产业化的前景分析. 中
国生物防治, 1999, 15(3): 135-138.
Wan F H, Wang R, Ye Z C. Prospects of commercial products of
insect natural enemies in China. Chinese Journal of Biological
Control, 1999, 15(3): 135-138. (in Chinese)
(责任编辑 岳梅)

中国农业科学 2015,48(17):3477-3492 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.014
收稿日期:2015-06-01;接受日期:2015-06-29 基金项目:国家科技支撑计划(2015BAB23B02) 联系方式:白由路,Tel:010-82108673;E-mail:[email protected]
植物营养与肥料研究的回顾与展望
白由路
(中国农业科学院农业资源与农业区划研究所,北京 100081)
摘要:论文回顾了 1840 年以来植物营养与肥料科学的重大成就和有影响的事件,并针对目前有歧义的问题
进行了详细论述。在植物营养生理与生物学方面对植物营养必需元素、营养元素的吸收与转运、营养元素的转移
与再利用、菌根在植物营养中的作用、植物营养与植物抗病性,化学诊断中的植物有效养分提取、养分测定和目
前进行的光谱诊断等进行了回顾。论述了施肥模型、精准施肥技术和灌溉施肥技术的发展。分析了施肥模型中养
分分级模型、肥料效应函数模型、养分平衡模型和 DRIS 方法。综述了施肥对温室气体排放和环境的影响等。介绍
了目前主流的肥料品种和国际上的主要肥料资源。论文还对未来植物营养与肥料科学发展进行了展望,指出提高
肥料利用率是解决肥料问题的关键;解决蔬菜施肥过量需要理论与技术的支撑;养分高效基因的开发与利用需要
在正常养分条件下进行;施肥新技术是提高肥效的重要措施;同时还需要立足当前可用肥料资源发展肥料产业。
植物营养与肥料学科的发展与人们所面临的人口、资源和环境息息相关,肥料的施用既是保证食物安全的需要,
同时也给环境造成了巨大的压力。使用最少肥料,获得最多的食物是今后植物营养与肥料学科努力的方向。
关键词:植物营养;肥料;植物营养生理;营养诊断;施肥技术
Review on Research in Plant Nutrition and Fertilizers
BAI You-lu
(Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences, Beijing 100081)
Abstract: This article reviewed the major achievements and influential events in plant nutrition and fertilizer since 1840. In plant nutrition physiology and biology fields, the essential plant nutrition elements, the process of nutrient absorption and translocation, the nutrient transfer and re-utilization, the role of mycorrhizal in plant nutrition, and plant nutrition and disease resistance were reviewed. In nutrition diagnosis field, available nutrient elements extraction and determination in chemical diagnosis, and spectrum diagnosis were reviewed. In the fertilization technology field, the fertilization models including nutrient classification model, fertilizer effecting function model, nutrients balance model and DRIS method were reviewed. The article also described the development of precision fertilization and irrigation technology. The pollutions caused by fertilization, including greenhouse gas emissions, groundwater pollution and surface water pollution were summarized. This article also introduced current main fertilizer products and fertilizer resources worldwide. In this article, it was pointed out that first, the key to solve fertilizer related problems is to improve fertilizer use efficiency. Secondly, to solve the problem of excessive fertilization on vegetables, new and improved theory and technology on vegetable fertilization are needed. Thirdly, the selection of nutrient efficiency gene should have to be taken place at common nutrient conditions in field. Fourthly, new fertilization technology is an important step to improve the fertilizer efficiency. Lastly, fertilizer industry development needs to be based on currently available fertilizer resources. The development of plant nutrition and fertilizer science is closely related to the population, resources and environmental problems that the people are facing. Fertilizer secures food security, but at the same time causes tremendous pressure to the environment. “Get more food with scientific use of fertilizer” is the direction of plant nutrition and fertilizer research in the future.
Key words: plant nutrition; fertilizer; plant nutrition physiology; nutrition diagnosis; fertilization technology
jia
文本框
返回目录

3478 中 国 农 业 科 学 48 卷
0 引言
肥料是粮食的粮食,支撑着人类食物的生产和供
给。虽然肥料的应用可追溯到几千年前[1],但是现代
施肥理论体系的建立与化学肥料的应用还是 19 世纪
40 年代以后的事情。1840 年德国科学家李比希(Justus Von Liebig)提出的“矿物质营养学说”、“ 小养
分律”、“归还学说”等[2]为现代施肥技术与化学肥
料的发展奠定了理论基础。一百多年来,经过数代人
的努力,人们在植物营养生理、营养诊断、施肥技术
与肥料创制等方面取得了重大进展,为保障人类的粮
食供应做出了卓越的贡献。目前,随着全球人口的增
长,对粮食的需求越来越大,农业开发强度越来越高,
施肥与环境的问题日益突出。因此,保证粮食充分供
应、满足植物营养需求、保护生态环境、节约肥料资
源成为了世界植物营养与肥料研究的主题。笔者结合
国内外的研究文献,对植物营养生理、植物营养诊断、
施肥技术、施肥与环境及肥料产品等方面里程碑事件
及其容易产生歧义的问题进行了回顾,同时展望了植
物营养与肥料科学未来发展方向,旨在为中国植物营
养与肥料研究提供参考。
1 植物营养生理与生物学
1.1 植物营养必需元素
1840 年之后,Leibig 提出“植物矿物质营养学说”
被人们广泛接受。然而,植物生长发育过程中究竟需
要什么元素成了当时研究的热点。1930 年之前,人们
认识到的植物营养必需元素仅有 10 种,即氮(N)、
磷(P)、钾(K)、钙(Ca)、镁(Mg)、硫(S)、
碳(C)、氢(H)、氧(O)和铁(Fe)[3]。到 1938年,人们认识到的植物营养必须元素增加到了 14 种,
除上述 10 种外,又增加了硼(B)、铜(Cu)、锰(Mn)和锌(Zn)[4]。1939 年,美国加里福尼亚大学农学院
的 Arnon 和 Stout 提出了植物必需元素的 3 个原则,
即(1)缺乏该元素,植物不能完成其生长周期;(2)缺乏该元素植物会出现特有的症状,且必须施用该元
素后才能恢复;(3)该元素在植物营养中具有直接
作用而不是改善土壤或生长介质中不利的微生物或
化学条件[5]。人们对这些观点已广泛接受。1954 年,
氯也被确定为植物生长必需的营养元素[6]。到了近代,
关于植物必需营养元素的概念与界定出现了模糊,它
主要源自于 1960 年英国布里斯托尔大学(University of Bristol)Long Ashton 试验站 Nicholas 的一篇文章,
该文认为 Arnon 有关必需元素的定义第二条太严格或
太死板(too rigid),且有两个例子证明不符合事实,
一是固氮菌在固氮时需要钼,但钒(V)也有类似的
作用;二是氯是植物的必需元素,但其他卤族如溴也
可以替代。所以认为用更宽泛的概念可能更适合当前
的工作。“功能或代谢营养”(functional or metabolism nutrient)这个术语可能概括植物代谢中的功能,不必
考虑其专一性。所以,他认为除以上元素外,钠(Na)、钴(Co)、钒(V)也应是植物的必需营养元素[7]。
该观点也得到了很多人的认同,Tisdale 等[8]在其著的
《土壤肥力与肥料》专著第四版(1985)中将植物的
必须营养元素描述为 20 种,即碳(C)、氢(H)、
氧(O)、氮(N)、磷(P)、硫(S)、钙(Ca)、
镁(Mg)、钾(K)、铁(Fe)、锰(Mn)、钼(Mo)、
铜(Cu)、硼(B)、锌(Zn)、氯(Cl)、钠(Na)、
钴(Co)、钒(V)和硅(Si)。20 世纪 60 年代以后,
人们注意到了镍在植物营养方面的作用,1990 年,
Brown 等[9]证明缺镍大麦不能完成其生命周期,符合
Arnon 植物必需营养元素的条件。在 Lincoln Taiz 和Eduardo Zeiger 所著的《植物生理学》(第五版)中将
植物营养的必须元素列了 19 种,即碳(C)、氢(H)、
氧(O)、氮(N)、钾(K)、钙(Ca)、镁(Mg)、
磷(P)、硫(S)、硅(Si)、氯(Cl)、铁(Fe)、
硼(B)、锰(Mn)、钠(Na)、锌(Zn)、铜(Cu)、
镍(Ni)和钼(Mo)[10]。我国出版的高校教材《植物
生理学》引用了上述观点,也将上述 19 种元素列为了
植物的必需营养元素[11]。但是,Marschner[12]在其所著
的《高等植物矿质营养》一书中,把植物营养元素区
分为必需元素(essential mineral element)和有益元素
(beneficial element)。其中植物营养必需元素严格引
用了 Arnon 1939 年的植物必需营养元素标准,而有益
元素则定义为有益于植物生长,但不是必需或者是某
些植物种类必需或特殊条件下必需的元素。该书中将
植物的必需营养元素定义为 17 种,其中除碳(C)、
氢(H)和氧(O)外,大量元素 6 种,即氮(N)、
磷(P)、硫(S)、镁(Mg)、钙(Ca)、钾(K)。
微量元素 8 种,即:铁(Fe)、锰(Mn)、铜(Cu)、
锌(Zn)、镍(Ni)、钼(Mo)、硼(B)和氯(Cl)。
有益元素有钠(Na)、硅(Si)、钴(Co)、硒(Se)、
铝(Al)、碘(I)、钒(V)、钛(Ti)、镧(La)、铈(Ce)等。该书第二版第一次将镍(Ni)列为了植
物的必需营养元素。中国人教版的中学《生物》教材
中也将镍列为了植物生长必需的营养元素[13]。

17 期 白由路:植物营养与肥料研究的回顾与展望 3479
这里需要指出的是,由于对植物必需营养元素界
定的的标准不同,因此确定植物生长必需元素的数量
和种类也就不同。但是,随着科学技术的发展和分析
技术的提高,可能还会发现更多的植物必需营养元素。
1930 年以后,人们对植物必需元素的研究与发现主要
集中在微量元素和有益元素层面,特别是在地球环境
下,极痕量的元素可能对植物生长影响不大,但是,
随着太空技术的发展,若在太空环境下培养植物,或
许任何元素都可能影响到植物生长。 1.2 营养元素的吸收与转运
营养元素的吸收与转运过程可分为元素在植物体
外的传输过程、跨膜运输过程和植物体内转运过程。
营养元素在体外传输过程的研究主要为根际和肥际
养分的转化与运移。根际概念是 1904 年德国微生物
学家 L.Hiltner 提出的, 初主要是研究根际的微生
物效应。之后,人们开始注意到了根际土壤的微观
结构、土壤化学特征、养分的有效性及养分在根际
的运移等[14]。“肥际”一词源于鲁如坤先生的“肥际
微域”,是指“肥料施入土壤后,特别是集中施用时
都会在肥料和肥粒附近造成一个特殊的环境,其物理
性质、化学性质、生理化学性质和生物性质与整个土
体有巨大的不同”[15]。在之后的十多年间,许多学者
在肥际土壤的化肥养分转化迁移[16]、有机肥对肥际环
境的影响[17]、肥际土壤微生物特征[18]及脲酶等特性[19]
等方面进行了研究。 近年来,在生物技术的支持下,营养元素的高效
利用研究十分活跃。关于营养元素的高效利用,Moll等[20]1982 年提出了氮高效可分为两个部分,即氮吸收
高效和氮利用高效。Graham[21]1984 年定义了氮高效
基因型是在氮素限制条件下获得比其他基因型更高
产量的能力。以后的研究基本上是按这个定义进行
的[22-23]。 根据目前的研究资料,不同的植物都含有对某种
养分的高亲和运输系统(high affinity transport system)
和低亲和运输系统(low affinity transport system),当
环境中养分浓度不同时,相应亲和运输系统则发生作
用。不同的作物基因型所含的家族基因有一定的差别,
所以,不同基因型的植物会对环境中的营养元素浓度
产生不同的反应[24]。同时,针对植物体内的养分代谢,
人们也发现了不同的代谢反应受不同的酶和基因控
制[25],针对不同作物,人们对其进行了大量的研究。 1.3 营养元素的转移与再利用
1953 年 Greory[26]发现,在禾谷类作物生长进程
中,当营养体仅为生育期总干物重的 25%时,就吸收
了 90%以上的氮、磷元素。以后的研究表明,在低浓
度养分条件下,老叶中氮素的再利用率高,高、低养
分浓度条件下的氮素再利用率分别为 46%和 65%[27]。
有关植物营养元素的转移与再利用有很多表述,如再
吸收(resorption)、转移(retranslocation)、再分配
(redistribution)等[28]。如何对植物营养元素再利用的
类型进行区分,很多人进行了大量的研究。有结果表
明,植物营养元素再利用的数据在年际间变化很大,
且在不同土壤上表现也不一致,所以,植物营养元素
再利用的类型只能通过长期观察才能决定[29]。很多人
对叶片中的氮、磷比与植物营养元素的再利用进行了
研究,但也有人认为叶片中的氮、磷比不能很好地预
测植物营养元素的再利用状况[30]。 对多年生木本植物的营养物质再利用的研究较
多,实际上一年生的植物也存在营养物质的再利用,
如小麦成熟时叶片中营养物质的转移直接影响小麦的
产量[31-32]。烤烟叶片中的氮素代谢严重影响烤烟的质
量[33]。 关于植物体内营养元素再利用的机理及影响因素
目前还没能完全明白,可以证明,植物体内的营养元
素再利用对提高土壤和肥料中养分的利用效率具有十
分重要的意义[34]。 1.4 菌根在植物营养中的作用
植物与伸入根系的共生真菌组成的复合体叫菌
根,地球上 90%以上的维管束植物都有菌根[35]。自
1885 年德国的生物学家 Frank 发现菌根以来,人们对
菌根已进行了广泛的研究,菌根在农业生态系统中分
布广泛,包括内生菌根(Endomycorrhizae)和外生菌
根(Ectomycorrihizae)两大类[36]。 1.4.1 内生菌根 内生菌根是指生于根部皮层细
胞内和细胞间的共生真菌,包括兰科植物菌根、杜
鹃科植物菌根和泡囊-从枝菌根 3 类,目前研究较多
的是第 3 类。泡囊-从枝菌根(Vesicular-Arbuscular Mycorrihizae),2000 年前一般简称为 VA 菌根,但
是后来的研究发现,一些 VA 菌根并不是一定有泡囊
结构,而是在植物生长的某个阶段才有,丛枝结构
(Arbuscular Mycorrhizae)是普遍存在于植物根部的
结构。因此,统称为丛枝(AM)菌根[37]。
内生菌根对植物生长的作用早已被人们所认识[38]。
它在植物营养方面的作用机制是增加了植物根系的吸
收面积,促进植物对营养元素的吸收,特别是对磷的
吸收[39]。内生菌根还能增强植物抵抗力,保护植物免

3480 中 国 农 业 科 学 48 卷
受病原体、害虫和寄生虫等侵害[40]。内生菌根还能增
加豆科作物的固氮能力[41]。同时,内生菌根还能减轻
水分对植物胁迫作用等[42]。国内对内生菌根也进行了
大量的研究,结果表明内生菌根对小麦[43]、烟草[44]、
草莓[45]、番茄[46]、铁皮石斛[47]、蓝莓[48]以及多年生树
种[49]都有良好的影响。 1.4.2 外生菌根 外生菌根(Ectomycorrihizae)简
称 ECM。与内生菌不同,外生菌根在根周围形成一个
菌套,或者穿透植物的表皮细胞但仍保持在细胞间,
形成一个哈迪网[50]。据报道,全世界目前外生菌有
5 000—6 000 种,主要是担子菌(Basidiomycetes)和
子囊菌(Ascomycetes)[51]。外生菌根的主要作用是分
解土壤中有机物[52],增强外生菌根与寄主植物的碳、
水和矿质养分的交换[53-54]。同时,外生菌根还能降解
土壤的 DDT 等有害物质[55]。外生菌根能拮抗植物根
部病害病原体。研究表明,外生菌根红蜡蘑(Laccaria Laccatta)可抑制尖孢镰刀菌(Fusarium oxysporum)[56],
牛肝菌(Boletus sp.)等对多种植物病原体都有抑制作
用。他们能抑制病原体的生长速度,使重叠菌落死亡,
从而控制病原体繁殖体的形成[57-58]。
目前的研究表明,无论内生菌根菌还是外生菌根
菌对植物营养的作用均在土壤养分含量较低时才表
现,且林木较多。在养分充足条件下,菌根菌对植物
营养的作用研究较少。目前,由于菌根菌很难实现大
规模的人工培养,在生产上的应用还十分有限。 1.5 植物营养与植物抗病性
随着人们对生态环境的要求越来越高,如何减少
农药用量、保证足够粮食供应是人们面临的重要课题,
人们开始注意植物营养与植物抗病性的关系[59],尽管
这种关系还不能科学地解释清楚,但很多情况下人们
已经开始将这种关系应用于生产实践中。 在植物营养元素中,氮是 重要的元素之一,也
是目前报道影响植物病害 多的元素。一般认为,氮
素使用有增加植物病害的倾向。在以前研究的 61 例病
害中,氮素加重病害的有 21 例,减轻病害的 22 例,
有 18 例不能确定。使用铵态氮加重病害的 9 例,能减
轻病害的 18 例,34 例不能确定,使用硝态氮加重病
害的有 11 例,9 例为减轻病害,41 例不能确定[60]。
关于氮素与植物抗病性的机理可能是,当被感染组织
中游离氨基酸浓度增加时,它为病原体提供了养分,
所以,植物易感病;当被感染组织中游离氨基酸和多
肽减少时,它不能为病原体提供足够营养,所以,植
物表现为抗病[61]。
磷与植物病害的关系较为复杂。目前关于磷与植
物病害的关系已有大量研究,然而结果却大相径庭。
在目前 51 例植物病害与磷元素关系的研究案例中,有
28 例认为磷可减少作物病害,13 例认为磷加重植物病
害,10 例没有明确其关系。日本在水稻上的研究表明,
磷对稻瘟病几乎没有影响,当磷缺乏时,病害会减轻,
当磷适当时,再增加磷肥,则稻瘟病加重,磷增加病
害程度的加重仅出现在氮素处于高水平时[61]。有研究
表明,施磷肥后可增加作物根的体积从而提高作物对
线虫的抵抗能力。关于磷的抗线虫机理主要认为磷可
以增加植物体内维生素 C、植物油、酚类物质、过氧
化物酶等含量,这些物质不利于线虫繁殖[62]。 钾与植物病害的关系研究甚多。国际钾肥研究所
分析了 2 000 多篇研究报告指出,钾肥能减少植物病
害的蔓延,其中钾能抵抗真菌的占 70%,抵抗细菌病
害的占 69%,减少害虫的占 63%。但对病毒病的影响
结果差异很大,有 41%报道钾降低病毒病,而 51%则
报道施钾肥会加重病毒病发生[63]。另据报道,在 167例研究中,有 113 例表现为钾减轻病害,39 例表现为
钾可加重病害,8 例表现为钾对病害没有影响。钾对
线虫的影响方面,通过 10 例研究分析,钾减轻为害的
有 3 例,6 例表现为钾加重线虫为害,1 例表现为钾对
线虫病没有影响。有关钾对植物抗病性的影响机理还
不十分清楚,但钾对提高植物抗病性的主要机理可能
是钾可改变植物体内蛋白质和氨基酸的活性、降低植
物细胞的渗透性、防止植物组织软化等[64]。 钙是植物抗病性中 重要的元素,它能对 25 种作
物的 36 种病害起抵抗作用,其主要机理是钙可改变
pH,钙的丙酸盐可降低病原体的毒性,还可稳定细胞
壁和细胞膜等[65]。其他营养元素如镁、硫、铁、锰、
锌、铜、硅等都与植物病害有一定的关系[60]。
2 营养诊断
2.1 化学诊断
植物营养与肥料的发展离不开土壤养分的测试技
术,通过测定土壤养分了解土壤养分的供应状况是科
学施肥的基础。自 1840 年德国化学家 Liebig 提出植
物营养的矿物质营养学说以来,土壤养分测试技术的
研究从来没有间断过。 从严格意义上讲,土壤养分测定在 Liebig 的“矿
物质营养学说”之前就开始了,在 Liebig 著的《化学
在农业和生理学上的应用》一书中,就多次涉及对土
壤和肥料中的营养元素的分析,但所采用的分析方法

17 期 白由路:植物营养与肥料研究的回顾与展望 3481
不祥[2]。从这个意义上讲,土壤养分的测定是分析化
学研究的一个分支,即土壤养分测定是将分析化学的
技术和土壤植物营养学的原理相结合的产物。所以,
1840 年以后,在土壤养分测定方法研究的发展过程
中,土壤养分的测定可明显地分为两个方面,一方面
是土壤养分的提取技术,另一方面是提取液中养分的
分析技术。从学科上讲,前者无疑是土壤肥料学科的
研究内容,而后者更像是分析化学的内容,但两者又
密不可分,共同支撑了土壤养分测定技术的发展。 2.1.1 养分提取 1840 年以后的几十年间,土壤养
分测定方法并无显著进展。直到 19 世纪后期,土壤养
分测定方法的研究才有一些进展。进入 20 世纪后,土
壤养分的测定有了明显的发展,以下是土壤养分提取
方法研究的进程。 1883 年,Kjeldahl 提出凯氏定氮的方法测定全氮。
该方法被后人评价为“在分析化学历史上,没有一种
方法能像凯氏定氮法那样在如此短的时间内被全世界
广泛采用”,以后人们对其催化剂进行了一些改进[66],
该方法至今还被广泛使用。 1925 年,德国 Bechold 提出电超滤(EUF)法。
该方法可在一次测定中同时测得土壤中养分的强度、
容量、缓冲容量和固定能力等,并据此估算出肥料需
要量[67],有研究表明,这种方法在估计土壤钾长期供
应容量时,优于醋酸铵法和 M3 方法[68]。 1927 年,Schollenberger 提出 K2Cr2O7-H2SO4测定
土壤有机质;1934 年 Walkley 和 Black 在此基础上
提出了水合热法[69],目前水合热法是国际上常用的
方法[70]。中国目前多采用外加热法,即 1935 年 Turin提出的方法,也称丘林法[71]。由于 K2Cr2O7-H2SO4 测
定土壤有机质的方法成本高且费时,同时还存在铬和
强酸的环境风险,目前国际上开始趋向于采用烧失法
测定土壤有机质含量[72]。 1930 年,Troug 提出用 0.02N H2SO4 提取土壤
速效磷,该方法以后被修改为 H2SO4+(NH4)2SO4
浸提,磷钼蓝比色法[73],这种方法目前在日本还有
应用[74]。 1932 年,Morgan 提出 HOAc+NaOAc 为通用浸
提剂;1945 年,Bray 提出了 Bray1 土壤有效磷浸提剂;
1953 年 Mehlich 提出 Mehlich 1 土壤浸提剂;1954 年
Olsen 提出了土壤有效磷提取剂。目前在美国土壤速
效磷的浸提剂虽有 10 种,但使用较多的仅有 4 种,即
美国东北部州所采用的 Morgan 浸提剂、美国东南部
州采用的 Mehlich 1 土壤浸提剂、美国中北部州采用
的 Bray1 土壤有效磷浸提剂和美国西部州采用的
Olsen 土壤浸提剂。在土壤有效钾的提取方面,美国
有 5 种土壤浸提剂,但使用的主要有 3 种,即在美国
东北部州所采用的 Morgan 浸提剂、美国东南部州采
用的 Mehlich 1 土壤浸提剂、其他州则采用 1944 年
Bray 提出的中性 NH4OAc 浸提剂[75]。 20 世纪 70 年代以后,由于分析技术的提高,一
次性测定浸提液中的多种元素成为可能,特别是 ICP的出现,人们开始关注联合浸提剂,也称通用浸提剂
(Universal extractans),如 Mehlich3 浸提剂替代了
Mehlich1、Bray1 和中性醋酸铵用于黏重土壤,
Soltanpour 等改进了 1965 年 Lindsay 等提出用 DTPA浸提土壤微量元素,用 CaHCO3-DPTA(AB-DTPA)
替代了 Olsen 浸提剂,提取碱性土壤上的 P、K、Cu、Fe、Mn 和 Zn。Worlf 改进了 Morgan 浸提剂,将其扩
展到了浸提微量元素。Houba 等 1990 年建议用 0.01 mol·L-1 CaCl2 提取土壤中 P、K、Mg、Na、NO3-N、
NH4-N、OM-N、S、B、Fe、Cu、Mn、Zn 以及 Cd、Pb、Ni 等。美国 20 世纪 90 年代以后,又出现了很多
商业性的私人实验室,这些实验室所采用的方法主要
是区域土壤养分测试研究机构、推广机构或土壤植物
分析委员会推荐的方法。这样,土壤测试方法的重大
改变需要有足够的测试并由认证机构认可。所以,以
后有关土壤浸提方法的发展开始缓慢[75]。 1973 年,A. H. Hunter 在总结前人土壤测试工作
的基础上,吸收了美国北卡罗莱那州立大学的 D. Waugh、R. B. Cate 和 L. Nelson 的研究结果提出了评
价土壤养分状况的实验室化学分析方法[76]。该方法应
用联合浸提剂显著提高了测试效率,在中国-加拿大政
府间合作研究项目实施中引进中国[77],在测土推荐施
肥工作中成功应用。 从以上研究进程不难发现,在土壤养分测试的过
程中,由于全量养分的测试技术较为单一,所以,土
壤养分测试的研究比较侧重于土壤有效养分。在 20世纪 70 年代以前,土壤养分的浸提多以单元素为主,
90 年代以后,基本上以联合浸提为主,以单元素浸提
的方法基本上被固定下来,作为了土壤养分测定的基
准方法。在以往的研究中,由于土壤速效养分的不确
定性,所以,对土壤速效养分浸提方法的研究一直是
土壤养分研究的热点之一,但相对于土壤全量养分,
土壤速效养分浸提方法的研究则相对较少。 中国对土壤养分浸提剂的研究较少,目前基本上
还是采用 20 世纪 60 年代以前的测试方法,即土壤速

3482 中 国 农 业 科 学 48 卷
效磷采用 Olsen 方法,速效钾采用中性醋酸铵浸提方
法,这些方法的测定结果具有年代可比性,但分析速
度较慢,使得农化服务的效率大为下降。 2.1.2 养分测定 在土壤养分测定技术方面,除了将
养分从土壤中浸提到溶液中以外,定量分析溶液中的
养分也是研究的重要方面。但是,后者更多是借鉴分
析化学的研究成果,近观百年,土壤测试技术的发展,
可归纳为几个阶段。 20 世纪 50 年代以前,土壤浸提液中养分的分析
基本上以常规方法(重量法、容量法)为主[78]。 20 世纪 60 年代以后,光谱化学和分光光谱技术
应用到了土壤养分的测试中,集中表现在极谱仪、X光、荧光分析仪的应用[79-80]。
60 年代电化学分析应用于土壤养分分析,特别是
氢电极的应用在土壤 pH 测定中基本上取代了比色分
析法[81]。 70 年代以后,以发射光谱和吸收光谱为主的分析
方法应用于土壤养分分析中。表现为火焰光度计、原
子吸收分光光度计等的应用[82]。 70 年代以后的电感耦合等离子体光谱仪(ICP)
应用于土壤养分测定[83]。 80 年代以后,电子技术的发展,使仪器开始自动
化(电子天平、自动定氮仪)[8]。 90 年代以后的网络数据自动采集与传输应用于
土壤养分分析[77]。 以上分析启示我们必须要在充分利用现代科学成
就、掌握新技术的基础上,来革新土壤科学研究的面
貌,而创造和改进测试技术以及分析方法是十分重要
的方面 [85]。但是,土壤养分分析的发展与仪器分析也
不完全同步。由于土壤养分分析的特殊性,方便、快
捷、经济的测试方法一直是土壤科学工作者孜孜以求
的目标。 2.2 光谱诊断
光谱诊断是通过植物叶片对不同光的反射、透射
和吸收特征来反映植物对外部环境变化的内部生理反
应的一种方法,其基本假定是外界环境的部分胁迫
可引起植物光合作用的显著变化,这样的变化导致
植物叶片对光反射的显著变化。该方法具有快速,
且不破坏植株结构的特点。光谱诊断起源于 19 世纪
末和 20 世纪初[86],目前许多商业化的设备可成功检
测植物的非生物和生物胁迫,如干旱、营养、昆虫
等[87-88]。作物营养的光谱诊断是遥感技术与植物营
养学结合的产物。根据遥感平台,可分为卫星遥感
诊断[89]、航空遥感诊断[90]、低空遥感诊断[91]和地面
遥感诊断[92]等。在相同分辨率的情况下,遥感平台
越高,诊断的面积越大、精度越低。近年来,在植
物营养诊断方面,利用无人机的低空遥感诊断[93]和
利用高光谱技术的地面遥感诊断得到了迅速发展。
一些专门用于植物营养诊断的仪器也相继出现,如
SPAD[94]、Greenseeker[95]等。 作物营养元素含量是作物营养光谱诊断的重要方
面[96],作物营养光谱诊断研究 多的养分是氮素,其
主要原理是通过测定作物叶片中的叶绿素含量来推算
植物体的含氮量[97-98]。光谱诊断的方法一般是通过光
谱中不同谱段的计算,形成一个指数,通过不同的指
数换算成营养元素的含量, 常用的是归一化植被指
数(DNVI)[99],也有很多不同的指数被研究[100-101]。
植物反射光谱的红边位置与其叶绿素含量[102]和氮素
含量[103]也有密切关系。目前,光谱技术也用于了土壤
养分的预测[104]和其他作物特性[105],特别是作物的病
虫害的诊断[106]。
3 施肥技术
3.1 施肥模型
根据作物营养需求、生长环境和营养诊断的结果,
计算出作物的肥料需求数量及其在作物生育期中的分
配,即为施肥模型。从不同的角度可以将施肥模型分
为多种[107]。目前在生产实践中应用较多的有三大类。 3.1.1 养分分级模型 也称为土壤养分分级模型或
养分指标法,即把土壤养分或养分指数分为若干等级,
每一个等级对应一个作物的施肥量。这种方法简单明
了,便于操作,在等级划分时,可能将许多因素,如
土壤培肥、环境影响等考虑在内。目前在欧洲的施肥
指导手册中多采用这种方法[108]。
3.1.2 肥料效应函数模型 肥料效应函数模型是根
据 1909 年 Mitscherlich 提出的作物产量与土壤养分供
应量之间的关系发展而来的[109]。以后人们对此进行了
大量的研究,形成了 Mitscherlich-Bray 方程[110]。 由于这种方法有明确的施肥量与作物产量的关系曲线,
克服了养分分级模型中“等级内差异缩小化、等级间
差异扩大化”的弊端,是一种较为准确的施肥模型,
这种方法目前还在世界各地大量应用[111]。中国从2005年开始的测土配方施肥行动中[112]所规定的 3414 试
验,就是依据肥料效应函数模型提出的[113]。据此在全
国范围内的不同作物上都进行了大量的肥料效应函数
模型研究。

17 期 白由路:植物营养与肥料研究的回顾与展望 3483
3.1.3 养分平衡模型 养分平衡模型是基于作物吸
收与土壤养分供给和施用养分平衡的施肥量计算方
法。这个方法源于 Truog 1960 年第七次国际土壤学会
上做的“测土工作五十年”报告。本方法目前被称为
“目标产量法”。1967 年印度学者 Ramamoorthy 著文
推广应用该方法,也称 Turog-Ramamoorthy 法。1973年,美国学者 Stanford 提出了氮肥需用量公式[114],也
就是目前应用 广泛的作物目标产量法计算公式[115]。
该方法中关于土壤供肥量的确定一直是人们研究的热
点,特别是在氮素的供应方面[116-117]。 3.1.4 其他施肥模型 近年来,随着精准农业的发
展,人们需要快速确定作物的施肥量,一些养分传感
器技术应运而生。通过传感器对叶片中养分测定,直
接计算出施肥量。这种方法的代表是法国学者 Beaufil和南非学者 sumner 共同提出的 DRIS 法(diagnosis and recommendation integrated system)[118]。该方法主要是
利用植物叶片中营养元素的含量及其比值进行营养诊
断, 终确定施肥量的方法[119],适合于土壤养分测试
较困难的果树或林木的营养诊断及施肥[120]。
3.2 精准施肥
精准施肥起源于 20世纪 70年代中期和 80年代初
期,主要是基于地块内土壤养分的变异。同时,该时
期一些新技术的发展如微型计算机技术、地理信息系
统(GIS)、全球卫星定位系统(GPS)等,使得农田
空间信息的获取成为可能,并形成了一个新农业技术
—计算机控制与传感器。 初称为“根据土壤类型管
理(farming by soil type)”,以后又称为“精确点位
管理(site-specific management)”,现在统称为“精
准农业(precision agriculture)”[121]。精准农业是将
空间信息技术应用于农业以改进农业生产决策过程的
技术体系。在施肥方面,传统农业是根据每个地块的
土壤养分状况,在地块内均匀施肥;而精准施肥根据
土壤养分等特性将一个地块划分成若干区域,然后根
据每个区域的情况进行施肥,它能 大限度地发挥耕
地和肥料资源的作用[122]。精准施肥的理论依据是土壤
养分的空间变异(spatial variability),核心是变量技
术(variable-rate technology),技术支撑是信息技术
(information technology)。 目前,在精准施肥的实现方面,主要是通过两种
技术体系,一个是基于 3S(GIS 、GPS、RS)的技术
体系[123],另一个是基于传感器的技术体系[124]。基于
3S 技术的变量施肥是在地理信息系统(GIS)、全球
卫星定位系统(GPS)和遥感技术(RS)的支持下,
通过预先置入施肥机上的施肥图,在 GPS 的指导下,
调节施肥量的大小。对固体肥料,变量施肥机大部分
采用转动式,通过液压马达驱动排肥器,以达到变量
施肥的目的。对于液体肥料,一般采用流量倍速管组
合的方式达到变量施肥的控制。而基于传感器的变量
施肥则通过安装在施肥机的实时传感器,测定作物需
肥量的多少,然后通过控制变量机,达到变量施肥的
目的。 目前,依据不同技术体系的商用变量施肥机在世
界各大公司都有生产。美国的 AGCO 公司[125]、John Derre 公司[126]、CASE 公司[127]、Micro-Trak 公司[128]、
Mid-Tech 公司[129]、Trimble 公司、Agleader 公司[130]、
加拿大的 Agtron 公司[131]、欧洲 AMAZONE 公司[132]、
RDS 公司、AMASAT 公司、日本的 Hatsuta 公司等都
生产与变量施肥有关的整机或控制设备及信息技术设
备等[133]。 3.3 灌溉施肥
把肥料直接注入灌溉水中进行施肥的方法称为灌
溉施肥(fertigation)[134]。有关灌溉施肥研究始于 1958年[135]。20 世纪 60 年代初开始迅速发展[136]。该技术
可节水 40%以上,节肥 20%以上,节约土地 5%—7%,
比常规省工 90%,增产幅度达 30%—50%,有效保
护了生态环境[137]。这种技术被认为是一项高效的施
肥技术[138],目前该技术正在与信息技术结合,实现农
田水分-养分的全自动化管理[139]。
4 施肥与环境
随着人口的增长,粮食的需求越来越多,化学肥
料的用量也越来越大,2008—2013 年间,世界年均氮
肥用量增加了 2.2%、磷肥 3.8%、钾肥 5.3%。2013 年
世界化肥用量为 1.84 亿吨(纯养分)。然而由于不合
理的使用,导致在全世界范围内不同地区出现严重的
环境和经济问题[135]。 4.1 温室气体排放
肥料特别是化学肥料在生产、运输、使用过程中
都会产生温室气体。据统计,全世界每年因肥料制造
而产生的温室气体为 465 万吨 CO2当量,占全球温室
气体排放的 0.93%;肥料运输而生产的温室气体为 37万吨 CO2当量,占全球温室气体排放的 0.07%;肥料
使用中生产的 N2O 和尿素、碳酸氢铵中直接排放的
CO2 合计为 728.5 万吨 CO2 当量,占全球温室气体排
放的 1.5 %。总计因肥料引起的温室气体排放约占全
球温室气体排放的 2.5 %[140]。

3484 中 国 农 业 科 学 48 卷
4.2 肥料污染
大气 PM2.5 的化学组成中,铵盐占有相当数量,
其主要来源是农业养殖业[141]。美国切萨皮克湾氮磷污
染物中的一半来源于农业不合理的肥料使用[142],密西
西比河流域过量使用氮肥所造成的环境灾难形成了
6 000 平方英里的“死亡区”[143]。中国“第一次全国
污染源调查公报”指出:农业源氮排放占工农、生活
源总氮排放量的 57.2%,农业源磷排放量占工农、生
活源总磷排放的 67.4%[144]。 农业一方面为地球上的人口增长提供了必需的粮
食,同时也给环境造成了巨大的压力,现代农业离不
开化学肥料[145]。所以,解决这一问题的唯一可行的方
法就是改进肥料管理 [135]。
5 肥料产品
李比希提出的矿质营养学说为肥料特别是化学
肥料的生产奠定了理论基础,100 多年来,人们在
化学肥料的生产和改进上做了大量的工作。1909 年
德国科学家 Haber 提交了高压法生产合成氨的专
利,1912 年他与 Bosch 一起成功将该技术用于了工
业生产,奠定了现代化肥工业体系的基础[146],形成
了目前以化学肥料为主导的施肥技术体系,后人高
度评价了合成氨技术的作用,并于 1918 年获得了诺
贝尔化学奖,认为目前地球上约一半的人口是靠合
成氨技术来养活的[147]。 5.1 主要肥料品种
目前,在世界范围内,主要的大宗肥料分 3 类,
即尿素、磷铵和钾盐[148]。大部分的复合肥料都是以这
3 种肥料为原料进行二次加工。 以天然气和煤生产的尿素占 94%,中国主要以煤
为能源生产尿素,约占世界煤头尿素的 97%。全世界
用石脑油为能源生产的尿素仅占 2%,其中印度石脑
油尿素占全世界产量的 92%[140]。其他主要是以天然气
为能源生产尿素的,也称气头尿素,在中国,气头尿
素约占 12%。 目前,磷肥的主要品种为磷酸一铵(MAP)和
磷酸二铵(DAP),西方国家基本上采用是“磷酸
浓缩法工艺”,也称传统法,主要在生产过程中将
中间产品低浓度湿法磷酸蒸发浓缩,除去大部分水
后,再与氨中和反应,反应料浆直接造粒得到产品[149],
中国主要采用“中和料浆浓缩法磷铵工艺”,也称
“料浆法工艺”,它与传统法的区别在于先以氨中
和稀硫酸,制得的中和料浆再进行蒸发浓缩,从而
避开了磷酸浓缩的困难,也适合我国大部分的中、低
品位胶磷矿[150]。 世界钾盐产量的 93%用于钾肥生产,目前主要的
钾肥品种有氯化钾、硫酸钾和硫酸钾镁肥 3 个品种,
其中氯化钾占 90%以上。中国钾资源主要分布在西北
地区,至 2014 年,中国钾肥产量达 610.47 万吨(折
纯)[151]。 5.2 肥料资源
肥料产业是一个高度依赖资源和能源的产业。如
前所述,氮肥虽然从空气中制取,但需要能源,磷肥
的生产需要磷矿,钾肥生产需要钾矿。 在氮肥的能源消耗方面,在世界范围内约有 75%
—80%的合成氨由天然气制备,每吨合成氨需要约
1 230 m3的天然气,若全部合成氨由天然气生产,将
消耗全世界约 5%的天然气。天然气的价格会严重影
响肥料的价格,目前每年消耗的天然气为 3.2 万亿 m3,
按这种速度,世界天然气可消耗 50 年[152]。 目前磷矿主要用于磷肥的生产,世界范围内磷矿
贮量为 150 亿吨,基础贮量为 470 亿吨,主要分布在
摩洛哥和西撒哈拉、中国、美国和南非,约占世界贮
量的 80%以上,按目前的开采速度,高品位磷矿可用
90 年,加上低品位磷矿可用 290 年。中国磷矿可用 80年,加上低品位磷矿可用 210 年[152]。
钾盐是生产钾肥的基础矿物,全世界贮量约 83亿吨,基础贮量约为 180 亿吨,主要分布在加拿大、
俄罗斯、白俄罗斯、德国和巴西,约占世界总贮量的
95%以上,按目前的开采速度,可用 235 年。加上低
品位钾矿,可用 500 年[152]。 世界氮、磷、钾资源贮量在可预见的未来是丰富
的。但随易开采矿物的减少,肥料的价格可能会不断
上涨。总之,肥料资源属于不可再生资源,做好肥料
管理是保证农业可持续的关键。 5.3 肥料创新
在粮食生产和环境保护的双重压力下,如何在
保证粮食安全的同时,减轻肥料对环境造成的巨大
压力,提高肥料利用率,减少肥料的浪费成了肥料
研究的热点。在肥料方面,通过肥料技术的创新,
提高肥料的效果也是提高肥效的重要途径。肥料的
创新离不开 4 个方面,即肥料中的养分含量、养分
比例、养分形态和肥料助剂[145]。目前正在向着高浓
度、多形态方向发展。肥料助剂也从传统的以改善
肥料物理特性向促进作物养分吸收、调节植物生长
的方向发展。

17 期 白由路:植物营养与肥料研究的回顾与展望 3485
6 植物营养与肥料科学的展望
现代植物营养与肥料科学发展一个半世纪以来,
在保证人类的粮食和农产品供应方面起到了重要的支
撑作用。但是,如前所述,肥料也给环境造成了巨大
的压力,肥料的养分特别是氮肥的利用率低,成了世
界范围内的问题,这其中有许多理论问题,也有很多
技术问题,同时还有政策问题,都需要在今后的植物
营养与肥料研究中加以解决。 6.1 提高肥料利用率是解决肥料问题的关键
据报导,在世界范围内,60%的氮肥用于了谷物
生产,其利用率估计为 33%[153],磷肥的当季利用率在
15%以下,30 年的累积利用率也很难达到 50%[154]。
为了提高肥料利用率,一些组织开始针对施肥中的关
键问题,倡导 4R 技术,即正确的肥料(right source)、
正确的用量(right rate)、正确的时间(right time)和正确的位置(right place)[155],旨在提高肥料利用
率,减少肥料对环境的影响。众所周知,肥料的生产
率与利用率是一个问题两个方面,报酬递减率告诉我
们二者不能兼得。在一定的技术水平下,提高肥料用
量,肥料的生产率必然降低,它所带来的肥料利用率
必然也会下降。所以,只有改进技术措施,才能使肥
料的生产率与利用率同时提高。同时也必须认识到,
肥料的利用率在什么样的区间,对环境的影响降至人
类可接受的程度,可能是今后植物营养与肥料学科和
环境学科需要共同研究的问题。 6.2 解决蔬菜施肥过量需要理论与技术的支撑
施肥所引起的环境问题在蔬菜和果树生产上表现
尤为突出,人们目前都将此问题归结为蔬菜和果树施
肥的不合理。为什么这个问题长期得不到解决,其中
一个重要的原因是理论问题未能解决,即蔬菜根系养
分耗竭区的问题。蔬菜生长速度比大田作物快得多,
生物量在短时间内快速增加必然需要大量的营养元
素,这样使蔬菜根系周围的养分耗竭强度加大,耗竭
区相应增加,补充根系表面的养分,必须增大耗竭区
外的养分浓度,增加耗竭区外缘与根表的养分浓度梯
度,以使养分更多更快地移动到根表,供蔬菜根系吸
收。所以,在不提高养分移动速度的情况下,减少施
肥就降低了耗竭区外的养分浓度,蔬菜根系得不到养
分供应就会生长受阻。解决蔬菜施肥量过大问题的关
键是提高养分的移动速度,强化蔬菜根系的养分供应。 6.3 在正常养分条件下开发与利用养分高效基因
目前,人们对植物养分高效利用基因的研究较多,
但是还不能在生产上应用。原因是多数的研究都是在
土壤养分极低的情况下进行的,也可以认为这些所谓
的养分高效基因是在环境养分极低时才能表达。试
想,世界化肥使用有一百多年的历史,一般土壤中的
养分浓度都达到了相当的水平,这也是人们培肥地力
的目的。如果不能在常规养分浓度下表现出养分的高
效利用,作物的产量也不会达到人们的预期,其应用
必然会受到限制。所以,研究高浓度养分情况下的养
分高效利用基因,是未来提高作物养分利用率的重要
途径。 6.4 施肥新技术是提高肥效的重要措施
随着信息技术的发展,使施肥技术的精准化成
为了可能,传感器技术使施肥的自动化程度大为提
高。这些技术的应用,不仅可能减少人工对施肥的
干预,还大大提高了施肥的针对性。在目前条件下,
特别是在中国新型城镇化和农业的适度规模经营的
发展下,省工、省时、高效率、低成本的施肥技术
必将得到快速发展,这些技术包括精准施肥技术、
水肥一体化管理技术、简化施肥技术等,这些技术
需要新型肥料、施肥机械、信息技术等支撑,今后
施肥技术的发展会是多学科协同创新的结果。传统
的植物营养与肥料学科必须结合新技术才能有大的
跨跃式发展。 6.5 立足当前可用肥料资源发展肥料产业
肥料资源是不可再生的资源,人们对肥料的珍惜
和对肥料资源的忧虑都是可理解的。同时也要看到肥
料技术的创新。用传统的肥料生产技术对待肥料资源
短缺问题会有一定的局限性。对磷资源而言,高浓度
磷肥是现阶段磷肥发展的主流[156],对中低品位磷矿石
的利用需要选矿技术作为技术支撑。若干年后,当土
壤中的速效磷累积到一定程度,施用中、低浓度磷肥
的效果可能会更好。 中国钾肥产量不能满足需求,每年需进口大量的
钾肥。由于钾资源多集中在西北地区,近年来,人们
注意到了低浓度难溶性钾矿(钾长石)的开发利用[157]。
这种技术研究和贮备是必须的。但是,在全世界范围
内,现阶段钾肥的应用还是以水溶性固体钾盐资源为
主。 总之,植物营养与肥料学科的发展与人们所面临
的人口、资源和环境息息相关,肥料的施用既是保证
食物安全的需要,同时也给环境造成了巨大的压力,
科学的使用适量的肥料,获得 多的食物是今后植物
营养与肥料学科努力的方向。

3486 中 国 农 业 科 学 48 卷
References
[1] 胡厚宣. 殷代农作施肥说. 历史研究, 1955(1): 97-106.
Hu H X. Fertilization for crop in Yin dynasty. Historical Research,
1955(1): 97-106. (in Chinese)
[2] 尤·李比希著, 刘更另译. 化学在农业和生理学上的应用. 北京: 中
国农业出版社, 1983.
J.V.Liebig (writer), Liu G L (translator). Organic Chemistry in Its
Application to Agriculture and Physiology. Beijing: China Agriculture
Press, 1983. (in Chinese)
[3] Schreiner O. Essential plant food elements. American Potato Journal,
1930, 7(11): 307-321.
[4] McMurtrey J E. Distinctive plant symptoms caused by deficiency of
any one of the chemical elements essential for normal development.
The Botanical Review, 1938: 183-203.
[5] Arnon D I, Stout P R. The essentiality of certain elements in minute
quantity for plant with special reference to copper. Plant Physiology,
1939, 14: 371-375.
[6] Broyer T C, Carlton A B, Johnson C M, Stout P R. Chlorine-a
micronutrient element for higher plants. Plant Physiology, 1954, 29(6):
526-532.
[7] Nicholas D J D. Minor mineral elements. Annual Review of Plant
Physiology, 1961, 12: 63-90.
[8] Tisdal S L, Nelson W L, Beaton J D. Soil Fertility and Fertilizers, 4
edition. London: Macmillan Publish Company, 1985.
[9] Brown P H, Welch R M, Madison J T. Effect of nickel deficiency on
soluble anion, amino acid, and nitrogen levels in barley. Plant and Soil,
1990, 125(1): 19-27.
[10] Lincoln Taiz, Eduardo Zeiger. Plant Physiology (5th Edition),
Sunderland, Massachusetts: Sinauer Associates Inc.,Publishers, 2010.
[11] 潘瑞炽. 植物生理学. 北京: 高等教育出版社, 2004.
Pan R Z. Plant Physiology. Beijing: Higher Education Press, 2004. (in
Chinese)
[12] Marschner H. Mineral Nutrition of Higher Plant (2th Edition). London:
Academic Press, Company Publishers, 1995.
[13] 人民教育出版社生物室编著. 生物(第一册). 北京: 人民教育出
版社, 2003.
Editorial Department of Biology, People’s Education Press. Biology
(1th Edition). Beijing: People’s Education Press, 2003. (in Chinese)
[14] 苏德纯. 作物根际环境研究进展. 世界农业, 1994(1): 41-42.
Su D C. Review in crop rhizosphere environment. World Agriculture,
1994(1): 41-42. (in Chinese)
[15] 鲁如坤. “微域土壤学”―一个可能的土壤学的新分支. 土壤学报,
1999, 36(2): 287-288.
Lu R K. Microzone soli science-a possible new branch of soil science.
Acta Pedologica Sinica, 1999, 36(2): 287-288. (in Chinese)
[16] 杜振宇 , 周健民 . 钾在红壤肥际微域中的迁移 . 土壤学报 ,
2005,42(6): 1033-1039.
Du Z Y, Zhou J M. Movement of potassium in fertilizer microsites in
red earth. Acta Pedologica Sinica, 2005, 42(6): 1033-1039. (in
Chinese)
[17] 徐岩. 有机肥优化土壤微域环境的机制研究[D]. 吉林: 吉林农业
大学, 2003.
Xu Y. Studies on optimization soil microenvironment with organic
fertilizers[D]. Jilin: Jilin Agricultural University, 2003. (in Chinese)
[18] 喻田甜, 彭思利, 朱劲, 宝德俊, 袁玲. 施肥对潮土肥际微域微生
物数量分布的影响. 贵州农业科学, 2014, 42(3): 83-87.
Yu T T, Peng S L, Zhu J, Bao D J, Yuan L. Effect of fertilizations on
amount and distribution of soil microbes in fertilizer microsites of
fluvo-aquic soli. Guizhou Agricultural Sciences, 2014, 42(3): 83-87.
(in Chinese)
[19] 陈欢. 脲/酶硝化抑制剂的效果比较及 DMPP 对氮素肥际与小麦根
系微域分布的影响[D]. 南京: 南京农业大学, 2010.
Chen H. Comparison of urease/nitrification inhititors and influence of
3, 4-dimethylpyrazole phosphate (DMPP) on nitrogen patch and
wheat root micro-distribution[D]. Nanjing: Nanjing Agricultural
University, 2010. (in Chinese)
[20] Moll R H, Kamprath E J, Jackson W A. Analysis and interpretation of
factors which contribute to efficiency of nitrogen utilization.
Agronomy Journal, 1982, 74: 562-564.
[21] Graham R D. Breeding for nutritional characteristics in cereals.
Advances in Plant Nutrition, 1984, 1: 57-102.
[22] Gourley C J P, Allan D L, Russelle M P. Plant nutrient efficiency: A
comparison of definitions and suggested improvement. Plant and Soil,
1994, 158: 29-37.
[23] Sattelmacher B, Horst W J, Becker H C. Factors that contribute to
genetic variation for nutrient efficiency of crop plants. Journal of
Plant Nutrition and Soil Science, 1994, 157: 215-224.
[24] Gaur V S, Singh U S, Gupta A K, Kumar A. Understanding the
differential nitrogen sensing mechanism in rice genotypes through
expression analysis of high and low affinity ammonium transporter
genes. Molecular Biology Reports, 2012, 39(3): 2233-2242.
[25] 赵首萍, 赵学强, 施卫明. 高等植物氮素吸收分子机理研究进展.
土壤, 2007, 39(2): 173-180.
Zhao S P, Zhao X Q, Shi W M. Advance in research on molecular
mechanism for nitrogen absorption in higher plant. Soil, 2007, 39(2):
173-180. (in Chinese)

17 期 白由路:植物营养与肥料研究的回顾与展望 3487
[26] Greory F G. Report of the 13th international Horticultural Congress.
Royal horticultural Society, London, England, 1953, 573-584.
[27] William R F. Redistribution of mineral elements during development.
Annual Review of Plant Physiology, 1955, 6: 25-42.
[28] Francisco I, Pugnaire F, Stuart C. Controls over nutrient resorption
from leaves of evergreen mediterranean species. Ecology, 1993, 74(1):
124-129.
[29] 李志安, 王伯荪, 林永标, 曾友特. 植物营养转移研究进展. 武汉
植物学研究, 2000, 18(3): 229-236.
Li Z A, Wang B S, Lin Y B, Zeng Y T. A review of study on nutrient
resorption of plant. Journal of Wuhan Botanical Research, 2000, 18(3):
229-236. (in Chinese)
[30] Drenovsky R E, Koehler C E, Skelly K, Richards J H. Potential and
realized nutrient resorption in serpentine and non-serpentine chaparral
shrubs and trees. Oecologia, 2013, 171(1): 39-50.
[31] Ratnam J, Sankaran M, Hanan N P, Grant R C, Zambatis N. Nutrient
resorption patterns of plant functional groups in a tropical savanna:
Variation and functional significance. Oecologia, 2008, 157(1):
141-151.
[32] 陈冬梅, 马永安, 刘保华. 不同落黄类型小麦品种的籽粒灌浆及叶
片光合特性. 麦类作物学报, 2008, 28(6): 1063-1067.
Chen D M, Ma Y A, Liu B H. Grain filling and photosynthetic
charateristics of wheat varieties differing in senesced yellowing type.
Journal of Triticeae Crops, 2008, 28(6): 1063-1067. (in Chinese)
[33] 郜俊红, 赵建国. 不同落黄类型小麦品种灌浆特征研究. 农业科技
通讯, 2015(4): 80-82.
Gao J H, Zhao J G. Study on grain filling charateristics of wheat
varieties differing in senesced yellowing type. Bulletin of Agricultural
Science and Technology, 2015(4): 80-82. (in Chinese)
[34] 李明, 梁晓红, 熊德中. 不同肥力植烟土壤上施氮水平对烤烟氮代
谢的影响. 热带作物学报, 2014, 35(2): 271-276.
Li M, Liang X H, Xiong D Z. Effects of various nitrogen application
levels on nitrogen metabolism of flue-cured tobacco in different
fertility levels tobacco-planting soil. Chinese Journal of Tropical
Crops, 2014, 35(2): 271-276. (in Chinese)
[35] 王鸣岐. 菌根理论与研究进展. 世界科学, 1985(2): 13-16.
Wang M Q. Review in theory of mycorrhizae. World Science, 1985(2):
13-16. (in Chinese)
[36] 郝会军, 刘英, 洪波, 雪珍. VA 菌根与植物矿质营养的研究进展.
安徽农业科学, 2007, 35(4): 978-979.
Hao H J, Liu Y, Hong B, Xue Z. Research progress of VA-mycorrhizal
fungi and mineral nutrition of plant. Journal of Anhui Agricultural
Sciences, 2007, 35(4): 978-979. (in Chinese)
[37] http://zuoye.baidu.com/question/cad66e380e98c10afc58eded008373f5.
html
[38] Ames R N, Reid C P P, Porter L K. Hyphal uptake and transport of
nitrogen from two 15N-labelled sources by glomusmosseae, avesicu
lar-arbuscular mycorrhizal fungus. New Phytologist, 1983, 95(3):
381-396.
[39] Lambers H, Raven J A, Shaver G R, Smith S E. Plant
nutrient-acquisition strategies change with soil age. Trends in Ecology
Evolution, 2008, 23: 95-103.
[40] Jung S C, Martinez-Medina A, Lopez-Raez J A, Pozo M J.
Mycorrhiza-induced resistance and priming of plant defenses. Journal
of Chemical Ecology, 2012, 38: 651-664.
[41] Saia S, Amato G, Frenda A S, Giambalvo D, Ruisi P. Influence of
arbuscular mycorrhizae on biomass production and nitrogen fixation
of berseem clover plants subjected to water stress. PLoS One, 2014,
9(3): e90738.
[42] Augé R M. Water relations, drought and vesicular-arbuscular
mycorrhizal symbiosis. Mycorrhiza, 2001, 11: 3-42.
[43] 陈贤英. 内生菌根与小麦伤流液及产量的关系. 福建农业科技,
1984(5): 10-12.
Chen X Y. Relationship between endomycorrhizae, bleeding and yield
of wheat. Fujian Agricrltural Science and Technology, 1984(5): 10-12.
(in Chinese)
[44] 方宇澄, 刘延荣, 方榕. 烟草内生菌根真菌的分离鉴定. 真菌学报,
1986, 5(3): 185-190.
Fang Y C, Liu Y R, Fang R. Theisolation and identification of
endomycorrhizal fungi on tobacco. Acta Mycologica Sinica, 1986,
5(3): 185-190. (in Chinese)
[45] 郭鹏, 贺学礼. AM 真菌对草莓的接种效应研究. 河北农业大学学
报, 2006, 29(4): 53-56.
Guo P, He X L. Inoculation effect of arbuscular mycorrhizal fungi for
strawberry. Journal of Agricultural University of Hebei, 2006, 29(4):
53-56. (in Chinese)
[46] 洪淑梅, 李培香, 方宇澄. 番茄内生菌根研究初报. 山东农业大学
学报, 1987, 18(2): 48-52.
Hong S M, Li P X, Fang Y C. Preliminary report on the study of
endomycorrhizal fungi in tomato. Journal of Shandong Agricultural
University, 1987, 18(2): 48-52. (in Chinese)
[47] 曹斌, 廖月葵, 何开家, 周军, 司徒百兴, 赖其瑞. 广西人工栽培铁
皮石斛内生菌根的研究. 广西植物, 2010, 30(5): 626-628.
Cao B, Liao Y K, He K J, Zhou J, Situ B X, Lai Q R. Study on
endomycorrhiza of cultivated Dendrobium of ficanale. Guihaia, 2010,
30(5): 626-628. (in Chinese)

3488 中 国 农 业 科 学 48 卷
[48] 肖军, 陈珣, 肇莹, 王红, 龚娜, 王娜, 丑静, 杨涛, 杨镇. 内生菌根
菌对蓝莓抗氧化成分含量的影响. 湖北农业科学, 2013, 52(10):
2408-2410.
Xiao J, Chen X, Zhao Y, Wang H, Gong N, Wang N, Chou J, Yang T,
Yang Z. Effect of endomycorrhizae on antioxidant constituent of
Semen trigonellae. Hubei Agricultural Sciences, 2013, 52(10):
2408-2410. (in Chinese)
[49] 孙玥, 庄海峰, 贾淑霞, 谷加存, 王政权. 多年施用氮肥对水曲柳人
工林内生菌根真菌侵染及其根尖形态的影响. 林业科学, 2010,
46(9): 50-57.
Sun Y, Zhuang H F, Jia S X, Gu J C, Wang Z Q. Effects of nitrogen
fertilization on VA mycorrhizal fungal colonizationand root
morphology of the first order roots in Fraxinus mandshurica
plantation in response to the colonization. Scientia Silvae Sinicae,
2010, 46(9): 50-57. (in Chinese)
[50] Pierre-Emmanuel C, Marc B, Abdala G D, Pascale F K, Francois L T,
Francois R, Marie-Pierre T, Stephane U, Jean G. The role of
ectomycorrhizal communities in forest ecosystem processes: New
perspectives and emerging concepts. Soil Biology & Biochemistry,
2010, 42: 679-698.
[51] Diagne N J, Sanguin T H. Ectomycorrhizal diversity enhances growth
and nitrogen fixation of Acacia mangium seedlings. Soil Biology &
Biochemistry, 2013, 57: 468-476.
[52] Baldrian P. Ectomycorrhizal fungi and their enzymes in soils: Is there
enough evidence for their role as facultative soil saprotrophs?
Oecologia, 2009, 161(4): 657-660.
[53] Talbot J M, Bruns T D, Smith D P, Branco S, Glassman S I, Erlandson
S, Vilgalys R, Peay K G. Independent roles of ectomycorrhizal and
saprotrophic communities in soil organic matter decomposition. Soil
Biology & Biochemistry, 2013, 57: 282-291.
[54] Phillips R P, Brzostek E, Midgley M G. The mycorrhizal-associated
nutrient economy: A new framework for predicting carbon-nutrient
couplings in temperate forests. New Phytologist, 2013, 199: 41-51.
[55] Huang Y, Wang J. Degradation and mineralization of DDT by the
ectomycorrhizal fungi, Xerocomus chrysenteron. Chemosphere, 2013,
92: 760-764.
[56] Sylvia D M. Suppressive influence of Laccaria laccata on Fusarium
oxysporum and on douglas-fir seedlings. Phytopathology, 1983, 73(3):
384-392.
[57] 栾庆书. 几种外生菌根菌对土传病原菌的拮抗作用. 辽宁林业科
技, 1992(6): 45-49.
Luan Q S. Antagonistic effect of several extomycorrhizae on soil
borne pathogens. Liaoning Forestry Science and Technology,
1992(6): 45-49. (in Chinese)
[58] 吴小芹, 孙民琴, 高悦, 盛江梅, 叶建仁. 几种外生菌根菌对松苗
抗非根部病害的影响. 林业科学, 2007, 43(6): 88-93.
Wu X Q, Sun M Q, Gao Y, Sheng J M, Ye J R. Effects of some
ectomycorrhizas on pine seedlings to disease resistance. Scientia
Silvae Sinicae, 2007, 43(6): 88-93. (in Chinese)
[59] Huber D M, Watson R D, Steiner C W. Crop residues, nitrogen and
plant disease. Soil Science, 1965, 100: 302-308.
[60] Datnoff L E, Elmer W H, Huber D M. Mineral nutrition and plant
disease. Minnesota: The American Phytopathological Society Press,
Second printing, 2009.
[61] Huber D M, Keeler R R. Alteration of wheat peptidase activity after
infection with powdery mildew (Abstract). Proceedings of the
American Phytopathological Society, 1977, 4: 163.
[62] Anne S, Prabhu, Nand K, Fageria, Berni R F, Rodrigues F A.
Phosphorus and plant disease. In:mineral nutrition and plant disease.
45-55.
[63] Amtmann A, Troufflard S, Armengaud P. The effect of potassium
nutrition on pest and disease resistance in plants. Physiologia
Plantarum, 2008, 133: 682-691.
[64] Prabhu A S, Fageria N K, Huber D M. Potassium and plant disease.
In:mineral nutrition and plant disease. 57-78.
[65] Rahman M, Punia Z K. Calcium and Plant disease/mineral nutrition
and plant In:mineral nutrition and plant disease. 79-93.
[66] 严梅荣 . 一百余年来凯氏定氮法的进展 . 粮油仓储科技通讯 ,
1986(2): 49-51.
Yan M R. Progress in Kjeldah method of nitrogen determination for
hundreds. Liang You Cang Chu Ke Ji Tong Bao, 1986(2): 49-51. (in
Chinese)
[67] Németh K. Electro-ultrafiltration of aqueous soil suspension with
simultaneously varying temperature and voltage. Plant and Soil , 1982,
64(1): 7-23.
[68] Di Meo V, Michele A, Paola A. Availability of potassium, calcium,
magnesium, and sodium in “Bulk” and “Rhizosphere” soil of
field-grown corn determined by electro-ultrafiltration. Journal of
Plant Nutrition, 2003, 26(6): 1149-1168.
[69] Walkley A, Black I A. An examination of degtjareff method for
determining soil organic matter and a proposed modification of the
chromic acid titration method. Soil Science, 1934, 37: 29-37.
[70] Gelman F, Binstock R, Halicz L. Application of the Walkley-Black
titration for the organic carbon quantification in organic rich
sedimentary rocks. Fuel, 2012, 96: 608-610.
[71] 牛永绮, 陈兰生. 土壤有机质测定方法的进展. 干旱环境监测,

17 期 白由路:植物营养与肥料研究的回顾与展望 3489
1998, 12(2): 97-100.
Niu Y Q, Chen L S. Progress of soil organism determination. Arid
Environmental Nvironmental Monitoring, 1998, 12(2): 97-100. (in
Chinese)
[72] Salehi M H, Beni O H, Harchegani H B. Refining soil organic matter
determination by loss-on-ignition. Pedosphere, 2011, 21(4): 473-482.
[73] Murphy J, Riley J P. A modified single solution method for
determination of phosphate in natural waters. Analytica Chimica Acta,
1962, 27: 31-36.
[74] Rahman M H, Sugiyama S. Dynamics of microbial community in
Japanese andisol of apple orchard production systems. Communications
in Soil Science and Plant Analysis, 2008, 39: 1630-1657.
[75] Jones Jr J B. Soil test methods: Past, present, and furture use of soil
extractants. Communication in Soil Science and Plant Analysis, 1998,
29(11-14): 1543-1552.
[76] Portch S. A systematic approach to soil fertility evaluation and
improvement. http://www.agroservicesinternational.com/booklet.htm
[77] 金继运, 白由路, 杨俐苹. 高效土壤养分测试技术与设备. 北京:
中国农业出版社, 2006.
Jin J Y, Bai Y L, Yang L P. Technology and Equipment of Efficient Soil
Testing. Beijing: China Agriculture Press, 2006. (in Chinese)
[78] 中国科学院南京土壤研究所. 土壤理化分析. 上海: 上海科学技术
出版社, 1978.
Institute of Soil Science, Chinese Academy of Science. Soil Testing of
Physical and Chemical Properties. Shanghai: Shanghai Scientific and
Technical Publishers, 1978. (in Chinese)
[79] 陈志慧, 孙洛新, 钟莅湘, 王琰, 尚保忠, 肖抒. 快速催化极谱法
测定土壤中的有效态钼. 岩矿测试, 2014, 33(4): 584-588.
Chen Z H, Sun L X, Zhong L X, Wang Y, Shang B Z, Xiao S.
Determination of available molybdenum in soil by rapid catalytic
polarography. Rock and Mineral Analysis, 2014, 33(4): 584-588. (in
Chinese)
[80] 章炜, 张玉钧, 陈东, 刘晶, 王春龙, 张荣, 赵南京, 刘文清. 内标
法在土壤重金属镍元素X荧光分析中的应用研究. 光谱学与光谱
分析, 2012, 32(4): 1123-1126.
Zhang W, Zhang Y J, Chen D, Liu J, Wang C L, Zhang R, Zhao N J,
Liu W Q. Application of internal standard to analysis of the metal Ni
element in soils by X-ray fluorescen spectroscopy. Spectroscopy and
Spectral Analysis, 2012, 32(4): 1123-1126. (in Chinese)
[81] 中国科学院土壤研究所. 土壤 pH 值的测定. 土壤通报, 1965(2):
36-38.
Institute of Soil Science, Chinese Academy of Sciences. The test of
soil pH. Chinese Journal of Soil Science, 1965(2): 36-38. (in Chinese)
[82] 俞铭钦 . 应用火焰光度计的土壤全钾的简捷测定法 . 土壤通
报,1963(3): 55-59.
Yu M Q. Express method of soil potassium by flame photometer.
Chinese Journal of Soil Science, 1963(3): 55-59. (in Chinese)
[83] 赵帅群 , 刘刚, 欧全宏 , 徐娟 , 任静 , 郝建明 . 不同类型土壤
的 FTIR 和 ICP-MS 分析. 光谱学与光谱分析, 2014, 34(12):
3401-3405.
Zhao S Q, Liu G, Ou Q H, Xu J, Ren J, Hao J M. Analysis of different
types of soil by FTIR and ICP-MS. Spectroscopy and Spectral
Analysis, 2014, 34(12): 3401-3405. (in Chinese)
[84] 谢小玲, 李海锋, 李雪莹, 郑天晓, 黎汉强, 赖会珍, 郭翠婷. 土壤
全氮半微量定氮法与自动定氮仪定氮法的比较分析. 生态环境学
报, 2012, 21(6): 1071-1074.
Xie X L, Li H F, Li X Y, Zheng T X, Li H Q, Lai H Z, Guo C T.
Comparison of traditional method with automatic method in soil total
nitrogen testing. Ecology and Environmental Sciences, 2012, 21(6):
1071-1074. (in Chinese)
[85] 陈家坊. 近十年来土壤化学分析方法的进展. 土壤学报, 1963,
11(4): 444-451.
Chen J F. Review of soil chemical analysis in decade. Acta
Pedologica Sinica, 1963, 11(4): 444-451. (in Chinese)
[86] Carter G A, Knapp A K. Leaf optical properties in higher plants:
Linking spectral characteristics to stress and chlorophyll concentration.
American Journal of Botany, 2001, 88(4): 677-684.
[87] Jackson R D. Remote sensing of biotic and abiotic plant stress. Annual
Review of Phytopathology, 1986, 4: 65-87.
[88] 戴昌达. 病虫害的遥感探测. 自然灾害学报, 1992, 1(2): 40-46.
Dai C D. Detecting vegetation damage by remote sensing. Journal of
Natural Disasters, 1992, 1(2): 40-46. (in Chinese)
[89] Hosseini R, Newlands N K, Dean C B. Statistical modeling of soil
moisture, integrating satellite remote-sensing (SAR) and ground-based
data. Remote Sensing, 2015, 7: 2752-2780.
[90] 张桂英. 基于 Leica ADS80 系统的航空遥感影像成果质量控制研
究. 测绘通报, 2014(10): 89-91.
Zhang G Y. The study of quality control of aerial remote sensing
results based on Leica ADS80 system. Bulletin of Surveying and
Mapping, 2014(10): 89-91. (in Chinese)
[91] Zhang C, Walters D, Kovacs J M. Applications of low altitude remote
sensing in agriculture upon farmers' requests-a case study in
northeastern ontario, Canada. PLoS One, 2014, 9(11): e112894.
[92] Walsh O S, Klatt A R, Solie J B. Use of soil moisture data for refined
GreenSeeker sensor based nitrogen recommendations in winter wheat
(Triticum aestivum L.). Precision Agriculture, 2013, 14(3): 343-356.

3490 中 国 农 业 科 学 48 卷
[93] Salani E, Barrado C, Pastor E. UAV flight experiments applied to
the remote sensing of vegetated areas. Remote Sensing, 2014, 6:
11051-11081.
[94] Bullock D G, Anderson D S. Evaluation of the minolta SPAD-502
chlorophyll meter for nitrogen management in corn. Journal of Plant
Nutrition, 1998, 21(4): 741-755.
[95] Walsh O S, Klatt A R, Solie J B, Godsey C B, Raun W R. Use of soil
moisture data for refined GreenSeeker sensor based nitrogen
recommendations in winter wheat (Triticum aestivum L.). Precision
Agriculture, 2013, 14: 343-356.
[96] Ayala-Silva T, Beyl C A. Changes in spectral reflectance of wheat
leaves in response to specific macronutrient deficiency. Advances in
Space Research, 2005, 35(2): 305-317.
[97] Schepers J S, Blackmer T M, Wilhelm W W, Resende M.
Transmittance and reflectance measurements of corn leaves from
plants with different nitrogen and water supply. Journal of Plant
Physiology, 1996, 148: 523-529.
[98] Mariotti M, Ercoli L, Masoni A. Spectral properties of iron-deficient
corn and sunflower leaves. Remote Sensing of Environment, 1996, 58:
282-288.
[99] Schubert P, Lund M, Ström L, Eklundh L. Impact of nutrients on
peatland GPP estimations using MODIS time series data. Remote
Sensing of Environment, 2010, 114: 2137-2145.
[100] Bisson M, Fornaciai A, Coli A, Mazzarini F, Pareschi M T. The
vegetation resilience after fire (VRAF) index: Development,
implementation and an illustration from central Italy. International
Journal of Applied Earth Observation and Geoinformation, 2008, 10:
312-329.
[101] Fitzgerald G, Rodriguez D, O’Leary G. Measuring and predicting
canopy nitrogen nutrition in wheat using a spectral index-the canopy
chlorophyll content index (CCCI). Field Crops Research, 2010, 116:
318-324.
[102] Ju C H, Tian Y C, Yao X, Cao W, Zhu Y, Hannaway D. Estimating leaf
chlorophyll content using red edge parameters. Pedosphere, 2010,
20(5): 633-644.
[103] Kakani1 V G, Reddy K R. Mineral deficiency stress: Reflectance
properties, leaf photosynthesis and growth of nitrogen deficient big
bluestem (Andropogon gerardii). Journal of Agronomy and Crop
Science, 2010, 196: 379-390.
[104] Nawar S, Buddenbaum H, Hill J. Digital mapping of soil properties
using multivariate statistical analysis and ASTER data in an arid
region. Remote Sensing, 2015, 7: 1181-1205.
[105] Atzberger C. Advances in remote sensing of agriculture: Context
description, existing operational monitoring systems and major
information needs. Remote Sensing, 2013, 5: 949-981.
[106] Nansen C, Sidumo A J, Martini X, Stefanova K, Roberts J D.
Refectance-based assessment of spider mite “bio-response” to maize
leaves and plant potassium content in different irrigation regimes.
Computers and Electronics in Agriculture, 2013, 97: 21-26.
[107] 王兴仁, 陈新平, 张福锁, 毛达如. 施肥模型在我国推荐施肥中的
应用. 植物营养与肥料学报, 1998, 4(1): 67-74.
Wang X R, Chen X P, Zhang F S, Mao D R. Application of
fertilization model for fertilizer recommendation in China. Plant
Nutrition and Fertilizer Science, 1998, 4(1): 67-74. (in Chinese)
[108] Fertiliser Manual (RB209), 8th Edition. Department for Environment
Food and Rural Affairs. http://www.defra.gov.uk.
[109] 李仁岗. 肥料效应函数, 北京: 中国农业出版社, 1987.
Li R G. Fertilizer Effect Function. Beijing: China Agriculture Press,
1987. (in Chinese)
[110] Kashiath R S, Vipin P B. Application of Mitscherlich-Bray equation
for fertlizer use in wheat. Communication in Soil Science and Plant
Analysis, 2002, 33: 3241-3249.
[111] Afzala S, Islamb M, Obaid-Ur-Rehmanc. Application of mitscherlich-
bray equation for fertilizer use on groundnut. Communications in Soil
Science and Plant Analysis, 2014, 45: 1636-1645.
[112] 白由路, 杨俐苹. 我国农业中的测土配方施肥. 土壤肥料, 2006(2):
3-7.
Bai Y L, Yang L P. Soil testing and fertilizer recommendation in
Chinese agriculture. Soils and Fertilizers, 2006(2): 3-7. (in Chinese)
[113] 中华人民共和国农业部. 测土配方施肥技术规范(2011 年修订版).
http://www.moa.gov.cn/zwllm/tzgg/tz/201109/t20110922_2293389. htm
Ministry of Agriculture of the Peple’s Republic. Technical specification
of soil testing and fertilizer recommendation(2011 Revised edition).
http://www.moa.gov.cn/zwllm/tzgg/tz/201109/t20110922_2293389.
htm (in Chinese)
[114] 刘成祥, 周鸣铮. 对 Truog-Ramamoorthy测土施肥方法的研究与讨
论. 土壤学报, 1986, 23(3): 285-289.
Liu C X, Zhou M Z. Discussinon the principle and procedue of
fertilizer recommendation proposed by truog and ramamoorthy. Acta
Pedologica Sinica, 1986, 23(3): 285-289. (in Chinese)
[115] 白由路, 杨俐苹, 金继运. 测土配方施肥原理与实践. 北京: 中国
农业科技出版社, 2007.
Bai Y L, Yang L P, Jin J Y. Theory and Practice about Soil Testing and
Fertilizer Recommendation. Beijing: Chinese Agriculture Sci-Tech
Press, 2007. (in Chinese)
[116] Stanford G, Legg J O, Smith S J. Soil nitrogen availability evaluations

17 期 白由路:植物营养与肥料研究的回顾与展望 3491
based on nitrogen mineralization potentials of soil and uptake of
labeled and unlabeled nitrogen by plants. Plant and Soil, 1973, 39:
113-124.
[117] Miransari M, Mackenzie F. Development of a soil N test for fertilizer
requirement for wheat. Journal of Plant Nutrition, 2011, 34: 762-777.
[118] 耿增超, 张立新, 赵二龙, 张朝阳, 陈永欣. 陕西红富士苹果矿质
营养 DRIS 标准研究. 西北植物学报, 2003, 23(8): 1422-1428.
Geng Z C, Zhang L X, Zhao E L, Zhang C Y, Chen Y X. Study on the
diagnosis with DRIS norm of mineral elements in Fuji apple in Shanxi
province. Acta Botanica Boreali-occidentalia Sinica, 2003, 23(8):
1422-1428.
[119] Singh D, Singh K, Hundal H S. Diagnosis and recommendation
integrated system (dris) for evaluating nutrient status of cotton
(Gossipium hirsutum). Journal of Plant Nutrition, 2012, 35: 192-202.
[120] 田杰, 胡秋霞, 杨龙. 基于 DRIS 的养分诊断程序的实现. 电脑知
识与技术(学术交流) , 2007(20): 475-478.
Tian J, Hu Q X, Yang L. Design of nutrition diagnosis program based
on DRIS. Computer Knowledge and Technology(Academic Exchange),
2007(20): 475-478. (in Chinese)
[121] Robert P C. Precision agriculture: A challenge for crop nutrition
management. Plant and Soil, 2002, 247: 143-149.
[122] Morala F J, Terrónb J M, Rebollo F J. Site-specific management zones
based on the Rasch model and geostatistical techniques. Computers
and Electronics in Agriculture, 2011, 75: 223-230.
[123] Hummel J W, Gaultney L D, Sudduth K A. Soil property sensing for
site-specific crop management. Computers and Electronics in
Agriculture, 1996, 14: 121-136.
[124] Hassan S. Chattha A, Qamar U. Variable rate spreader for real-time
spot-application of granular fertilizer in wild blueberry. Computers
and Electronics in Agriculture, 2014, 100: 70-78.
[125] http://www.caseih.com.cn
[126] https://www.deere.com/en_US/products/equipment/nutrient_application/
nutrient_applicators/2510l/2510l.page
[127] http://www.caseih.com.cn
[128] http://www.micro-trak.com
[129] http://www.midtechserices.com/products.aspx,
[130] http://www.agleadercom.
[131] http://www.agtron.com/en/products/
[132] 宋毅, 殷华, 刘晓婧. 从法国 SIMA-SIMAGENA 展会看欧美农业
装备技术创新走势. 世界农业, 2013(4): 149-154.
Song Y, Yin H, Liu X J. The trend of agricultural equipment
innovation in European and American from SIMA-SIMAGENA fair
in France. World Agriculture, 2013(4):149-154. (in Chinese)
[133] 段洁利, 李君, 卢玉华. 变量施肥机械研究现状与发展对策. 农机
化研究, 2011(5) : 245-248.
Duan J L, Li J, Lu Y H. Research status and development
countermeasure of variable rate fertilization machinery. Journal of
Agricultural Mechanization Research, 2011(5): 245-248. (in Chinese)
[134] Hagin J, Lowengart A. Fertigation for minimizing environmental
pollution by fertilizers. Fertilizer Research, 1996, 43: 5-7.
[135] Ebrahimian H, Mohammad R, Playán K E. Surface fertigation: A
review, gaps and needs. Spanish Journal of Agricultural Research,
2014, 12(3): 820-837.
[136] Ayarsa J E, Pheneb C J, Hutmacher R B, Davis K R, Schoneman R A,
Vail S S, Mead R M. Subsurface drip irrigation of row crops: A review
of 15 years of research at the Water Management Research Laboratory.
Agricultural Water Management, 1999, 42: 1-27.
[137] 高祥照, 杜森, 钟永红, 吴勇, 张赓. 水肥一体化发展现状与展望.
中国农业信息, 2015, 173: 14-19.
Gao X Z, Du S, Zhong Y H, Wu Y, Zhang G. Review of fertigation.
China Agriculture Information, 2015, 173: 14-19. (in Chinese)
[138] Feigin A, Letey J, Jarrell W M. N utilization efficiency by drip
irrigated celery receiving preplant or water applied N fertilizer.
Agronomy Journal, 1982, 74: 978-983.
[139] Coates R W, Sahoo P K, Schwankl L J, Delwiche M J. Fertigation
techiques for use with multiple hydrozones in simultaneous operation.
Presicion Agriculture, 2012, 13: 219-235.
[140] IFA. Fertilizer, climate change and enhancing agricultural productivity
sustainably, 2009. http://www.fertilizer.org.
[141] Krupa S V. Effects of atmospheric ammonia (NH3) on terrestrial
vegetation: A review. Environment Pollution, 2003, 124: 179-221.
[142] Land L S. Chesapeake bay nutrient pollution: Contribution from the
land application of sewage sludge in Virginia. Marine Pollution
Bulletin, 2012, 64: 2305-2308.
[143] Paarlberg R. Food Politics, second edition. Oxford: Oxford University
Press, 2013.
[144] http://www.stats.gov.cn/tjsj/tjgb/qttjgb/qgqttjgb/201002/t20100211_
30641.html.
[145] 白由路. 我国肥料发展若干问题的思考. 中国农业信息, 2014(22):
5-9.
Bai Y L. Viewpoints on some problems of the development fertilizer
in China. China Agricultural Information, 2014(22): 5-9. (in Chinese)
[146] http://baike.baidu.com/link?url=AxjZsr4zqiVWqYmGHLmkCgbgE
4D7Pib6HHYqc-XuL73BkWqhA0FTTY3X-6KCeM2qC3hBARUoI
GjKtaCtMKSl4a
[147] Erisman J W, Sutton M A, Galloway J, Klimont Z, Winiwarter W.

3492 中 国 农 业 科 学 48 卷
How a century of ammonia synthesis changed the world. Nature
Geoscience, 2008, 1: 636-639.
[148] 汤炎, 张海林, 常志洲. 美国化肥使用概况. 磷肥与复肥, 2003,
18(6): 72-74.
Tang Y, Zhang H L, Chang Z Z. The general situation of fertilizer
application in USA. Phosphate & Compound Fertilizer, 2003, 18(6):
72-74. (in Chinese)
[149] 钟本和, 张允湘, 应建康. 料浆法磷铵工艺的发展和展望. 四川大
学学报: 工程科学版, 2003, 35(2): 1-5.
Zhong B H, Zhang Y X, Ying J K. Development and prospect of MAP
by ammoniated slurry concentrating process. Journal of Sichuan
University: Engineering Science Edition, 2003, 35(2): 1-5. (in Chinese)
[150] 钟本和. 料浆法磷铵生产工艺发展与五大磷肥工程改造. 中国工
程科学, 2004, 6(6): 22-25, 34.
Zhong B H. Development of ASCP for ammonium phosphate
technology and five projects of phosphate fertilizer in China. Engineering
Science, 2004, 6(6): 22-25, 34. (in Chinese)
[151] 2014 年中国钾肥产量数据统计分析. http://www.askci.com/chanye/
2015/02/08/17304d0u4. shtml
Analysis on the statistical output of potash in China in 2014. http://
www.askci.com/chanye/2015/02/08/17304d0u4. shtml. (in Chinese)
[152] Fixen P E, Johnston A M. World fertilizer nutrient reserves: A view to
the future. Journal of the Science of Food and Agriculture, 2012,
92(5): 1001-1005.
[153] Mohammend Y A, Jonathan K, Chim B K, Emily R, Kevin W,
Jeremiah M, Guilherme T, Kefyalew G D, William R. Nitrogen
fertilizer management for improved grain quality and yield in winter
wheat in Oklahoma. Journal of Plant Nutrition, 2013, 36: 749-761.
[154] Abbadi J, Gerendas J. Phosphorus use efficiency of safflower
(Carthamus tinctorius L.)and sunflower(Helianthus annuus L.).
Journal of Plant Nutrition, 2015, 38(7): 1121-1142
[155] http://www.nutrientstewardship.com/what-are-4rs
[156] 武希彦. 我国磷肥工业的现状及展望. 磷肥与复肥, 2001,16(1):
1-4.
Wu X Y. The present status & prospects of phosphate fertilizer
industry in China. Phosphate & Compound Fertilizer, 2001,16(1): 1-4.
(in Chinese)
[157] 夏举佩, 任雪娇, 李国斌, 苏毅, 晁静霞. 钾长石-硫酸钙-氧化钙
热反应制备可溶性钾机理研究. 北京工业大学学报, 2014, 40(11):
1735-1740.
Xia J P, Ren X J, Li G B, Su Y, Zhao J X. Mechanism study on the
preparation of soluble potassium from K-feldspar-CaSO4 -CaO
thermal system. Journal of Beijing University of Technology, 2014,
40(11): 1735-1740. (in Chinese)
(责任编辑 杨鑫浩)

中国农业科学 2015,48(17):3493-3503 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.015
收稿日期:2014-12-30;接受日期:2015-05-24 基金项目:中央高校基本科研业务费专项(2013XJ025)、“十三五”农业农村经济发展规划编制前期研究重大课题 联系方式:李保国,E-mail:[email protected]
蓝水和绿水视角下划定“中国农业用水红线”探索
李保国,黄 峰
(中国农业大学资源与环境学院土壤和水科学系/农业部华北耕地保育重点实验室,北京 100193)
摘要:水是制约中国粮食生产的最重要因素。实现中国粮食安全和口粮安全的战略目标必然要求粮食的持续
增产,而粮食增产对水资源需求的增长又是刚性的、不可减少的。在多部门用水需求加大的情况下,如何保证农
业用水量是亟待解决的科学和决策问题。文章首先回顾了国内外水资源管理领域的最新进展,总结了以“蓝水”
(耕地灌溉)和“绿水”(耕地降水)理念为核心的农业水管理分析范式的转变。在基于蓝水和绿水视角下的“广
义农业可用水资源”的概念和计算基础上,分析了 1998—2012 年中国农业(种植业中四大主要粮食作物)用水、
耗水和水分生产力,并基于此提出了“农业用水红线”。根据研究时段内 15 年的中国粮食水分生产力的变化趋
势,推导出 2020 年生产 5.5×108 t 粮食所需要的农业用水红线的蓝水分量的高、中、低值。并根据近 15 年来广
义农业可用水资源量中的绿水分量,推导出农业用水红线中绿水分量的高、中、低值。结果显示,到 2020 年,中
国保证粮食安全的灌溉水需求总体平均在 3 800×108 m
3左右,而农业用水红线中绿水分量平均在 4 500×10
8 m
3
左右。本研究最后提出了划定和实施农业用水红线的相关政策建议:从保护农业用水红线的角度,严格保护和实
施“18 亿亩耕地红线”;建立、健全完整、完善的蓝水和绿水测量和监督体系;加强以耕地土壤水为中心的农业
用水和耗水管理;利用各地水热资源特点,调整种植结构;继续提高灌溉的保证程度;加强耕地基础设施建设,
提高高标准农田和旱涝保收耕地面积和比例。本研究对回答农业部提出的治理农村环境污染“一控两减三基本”
的基本方针中的“一控”中的“控水的底线(即农业用水红线)是什么?”具有重要的理论和实践参考价值。
关键词:粮食安全;广义农业可用水资源;作物水分生产力;蓝水;绿水
Defining the Baselines for China Agricultural Water
Use in Green and Blue Water Approach LI Bao-guo, HUANG Feng
(Department of Soil and Water Science, College of Resources and Environment, China Agricultural University/Key Laboratory of
Arable Land Conservation (North China), Ministry of Agriculture, Beijing 100193)
Abstract: Water is the prime limiting factor in grain production in China. Agricultural water withdrawal has been under ever growing pressures in the context of population growth, economic development, urbanization process, diet change, and ecosystem preservation. However, to realize the national strategic goal of food security requires more grains to be produced, and consequently, does demand more water to be consumed. Hence, how to resolve the competing claims for water by various sectors while retaining adequate water for grain production will be the major challenge presenting to both researchers and decision makers. The paper firstly reviewed the state-of-the-arts ideas, concepts, and mindsets that had been emerged and developed in global water communities in the last 20 years, from which a paradigm shift in which green water and blue water lie at the center was identified and presented. Then, based on the “Broadly-defined Available Water for Agriculture (BAWA)” incorporating both green (i.e. cropland irrigation) and blue water (i.e. cropland precipitation) components, agricultural water use, consumption, and productivity were analyzed over a 15-year period from 1998 to 2012. Next the “Baselines for Agricultural Water Use” was proposed and proactively defined, in which blue
jia
文本框
返回目录

3494 中 国 农 业 科 学 48 卷
water component was derived by projecting historical crop water productivity onto 2020. The green water component was likewise determined by using projection of last 15-year BAWA. The results showed that the redlines for blue water withdrawal in terms of assuring 550 million tons of grains to the year 2020 was 380 km3, while the green water component of the redlines was 450 km3. The paper proposed a set of policies in implementing the redlines, including conserving arable land and improving arable land infrastructure, building a sound-developed system for measuring and monitoring blue and green water, developing a water-adapted farming system, improving crop water productivity, and adopting basin- and region-scale water saving practices. The conclusion of the paper will also have implications for recent policy directives issued by China Ministry of Agriculture to combat agriculture-derived pollution in rural areas, “one control, two abatements, and three basic requirements”, of which one control means the definition of baseline agricultural water withdrawals and water use efficiency.
Key words: food security; Broadly-defined Available water for Agriculture (BAWA); crop water productivity; green water; blue water 近年来,随着中国的人口增长、经济发展、城市
化进程以及收入水平提高,居民膳食结构改变,对优
质食物的需求增加。但是,尽管近 30 年的集约化农业
生产显著提高了农产品的产量和品质,但也带来耕地
质量退化[1]、农业面源污染[2]、集约化畜禽养殖对土壤
和水环境的破坏[3]等农业生产引发的生态和环境问
题。如何破解不断增长的食物需求和可持续发展之间
的矛盾成为中国面临的重大挑战。中国粮食安全是全
球关注的问题,相关研究都强调缺水是威胁中国粮食
安全的最主要因素[4]。水和粮是一个相互依存、相互
影响的问题。一方面,水资源的短缺威胁到灌溉农业
的有效灌溉量,从而影响到粮食产量和粮食安全;另
一方面,粮食生产又消耗大量的水,由于水分利用效
率的低下,在耗水同时造成农业用水的紧缺[5]。从这
个意义上说,水和粮的关系是一个互为因果的互动关
系。因此,解决粮食安全问题必须首先解决农业水资
源短缺的问题。 当前和未来,一些关键性的驱动力会对粮食安全、
水资源安全和生态安全产生重大影响。主要有:生物
物理驱动(气候变化、生态退化、环境污染);社会
经济驱动(经济发展、城市化、膳食结构改变);政
策驱动(粮食安全和口粮安全、水资源安全、生态安
全)。中国粮食安全近来出现了一些值得警惕的新动
向:国内粮食价格高于国际市场价格,谷物国内价格
比国际价格高 10%—15%,肉类价格是国际价格的两
倍;国内农业补贴空间日渐缩小;环境生态恶化的巨
大压力;农业劳动力持续减少;宏观经济环境变化的
不确定性[6]。这些因素都会影响中国粮食安全,但在
所有生物物理投入中,水依然是首要限制因子。 近 20 年来,全球水资源和农业用水领域出现了一
场分析和决策范式的转变(paradigm shift),涌现了
一批新的概念、理念和分析框架。这场范式转变主要
有以下概念组成:“蓝水”和“绿水”[7];水资源
收支解析[8];真实节水[9];净灌溉效率和有效灌溉效
率[10];水分生产力[11];虚拟水流动和贸易[12]。这些观
念和方法形成了一个相互关联而统一的有机整体,从
新的视角分析、解读农业用水和农业水资源管理,以
及和其他用水部门的关系。从更大的空间尺度分析、
解读农业用水和耗水管理,为实现粮食安全、水资源
安全、生态健康等多重目标的水资源综合利用提供了
有力分析工具。其中,“蓝水”和“绿水”是这一范
式转变中的核心理念。 与传统认识方式有所不同,笔者以“降水是所有
水资源的最终来源”的基本理念为出发点。因为传统
农业用水主要关注从地表(河流和湖泊)和地下水源
抽取出来用于灌溉的水。而实际上,降水可以被分成
流入河道的径流和暂时储存在土壤中的水分,而储存
于土壤中的水分则继续转化成土面蒸发和植物蒸腾。
用“蓝水”和“绿水”的概念可以描述上述复杂水循
环的来源和去向。“蓝水”是指在河流、地下含水层、
水库和湖泊中储存的水分,主要用于灌溉农业;“绿
水”则是由降水渗入土壤而产生、可以被植物吸收利
用的水分。绿水构成雨养农业的主要水源。但是在灌
溉农业中,蓝水在作物需水的关键期和干旱期补充绿
水的不足[5,13]。在农业实际生产中,雨养(旱作)和
灌溉农业之间存在一个连续的谱系,位于这个谱系上
的各农作体系之间的差别在于其对蓝水和绿水的不同
利用程度(图 1)。在中国,纯粹灌溉农业只出现于
某些干旱的西北绿洲农业类型中,因为这些农业完全
依赖地下水或高山融雪进行灌溉。而雨养(旱作)农
业为保证一定的产量,或多或少都会利用田间小型蓄
水设施(水窖)进行集雨补充灌水。因此,农业水管
理体系主要依赖于这样几种水源:降雨、地下水、地
表水和再利用的循环水。因此对农业用水的评价一定 要包括绿水和蓝水两大要素。
到 2030 年,中国人口预计达到 16 亿峰值,对食

17 期 李保国等:蓝水和绿水视角下划定“中国农业用水红线”初索 3495
物的需求也达到高峰。而食物(粮、蔬、果、肉、蛋、
奶)的生产需要消耗大量的水,其中既有蓝水,更有
绿水。2013 年国务院办公厅印发的《实行最严格水资
源管理制度考核办法的通知》中规定了各省、市、自
治区的“用水总量控制目标”、“用水效率控制目
标”和“重要江河湖泊水功能区水质达标率控制目
标”的“三条红线”。在“三条红线”中,用水总
量到 2020 年控制在 6 700×108 m3,到 2030 年控制
在 7 000×108 m3。在这条红线中,虽然规定了用水总
量,但并未涉及部门间配置。在经济快速发展、工业
化和城市化加快、生态环境需水量增长的大背景下,
农业用水受到了极大压力。到底需要给农业配置多少
水才能满足 2020 年和 2030 年的食物生产需求?这是
必须回答的重大科学和决策问题。因此,在基于蓝水
和绿水的“广义农业可用水资源”概念和计算的基础
上,笔者提出了蓝水和绿水视角下“农业用水红线”
的概念,并尝试划分其数值范围。其中,“农业用水
红线”中的蓝水部分是对“用水总量红线”的进一步
完善,而绿水部分则是本研究创新点以及需要进一步
研究的科学和决策问题。2015 年 4 月,农业部提出了
治理农村环境污染、改善农业生态环境的“一控、两
减、三基本”的政策,其中的一控就是要控制农业用
水的总量,并需要具体划定总量的红线和利用系数率
的红线。所以本研究对回答“一控”中的“控水的底
线(即农业用水红线)是什么?”也具有重要的理论
和实践参考价值。
图 1 农业用水管理的连续谱系[13]
Fig. 1 Continuous lineage of agricultural water management [13]
近几年,国际上对绿水和蓝水在农业生产,以及
绿水在其他主要陆地生态系统中的流动特征的研究日
趋活跃。Rockstrom 等[14]综合了全球农田水平衡数据
对 92 个发展中国家实现联合国千年发展目标(MDG)
的农田绿水潜力和作物水分生产力首次进行了评
估。很多同类研究都试图从全球[15-16]、大洲[17-18]、国
家[19-20]、区域[21-22]的宏观水平评价蓝水和绿水资源量,
回答农业中蓝水和绿水流的特征,以及提高蓝水和绿
水的水分生产力的发生机理和调控机制等科学问题。
全球作物平均虚拟水含量中绿水含量(1 100 m3·t-1)
和蓝水含量(291 m3·t-1)[23],绿水﹕蓝水为 8﹕2;全
球水稻产量中绿水、蓝水、灰水的贡献率分别是 48%、
44%、8%[24];全球农田生育期总耗水量 59 400×108
m3·a-1(其中 84%是绿水)、农田周年总耗水量为 73 230×108 m3·a-1(87%来自绿水)[25]。同时,相关模拟也
证明了蓝水的重要作用,如果没有“蓝水”灌溉,全
球谷物产量将减少 20%[24]。因此,综合提高农田蓝水
和绿水的消耗率及相应的作物水分生产力是当前和今
后一段时期内研究的方向和重点。国内有相关的农业
水资源阈值方面的研究[26-27]。 在综合国内外相关研究进展的基础上,笔者(1)
通过建立、计算和分析“广义农业可用水资源”,尝
试提出了基于蓝水和绿水的“中国农业用水红线”的
概念;(2)在广义农业可用水资源、粮食水分生产力、
粮食生产中蓝水和绿水贡献率的基础上计算了保证到
2020 年中国粮食总产达到 5.5×108 t 时所需要的灌溉
3496 中 国 农 业 科 学 48 卷
蓝水红线;基于历史和气候变化,以及耕地数量和质
量变化的分析基础上,计算了保证 5.5×108 t 粮所需要
的降水绿水红线;(3)对上述概念和结果进行了讨论,
提出了确保中国粮食安全的农业用水红线的研究、划
定和实施相关的对策建议。
1 蓝水和绿水视角下“广义农业可用
水资源”的概念、计算和分析
为解决从纯粹雨养农业到完全灌溉农业之间的连
续谱系中的农业生产用水问题,李保国和黄峰在全球
农业用水范式变迁的核心概念的基础上,建立了新的
中国农业用水分析综合框架[5,28],将上述概念和分析
方法融入其中,并初步建立了相应的计算和分析方
法[28-30]。该分析框架从蓝水和绿水概念出发,提出并
定义了“广义农业可用水资源”的概念,它由两个分
量组成,即耕地降水量(绿水分量)和耕地灌溉量(蓝
水分量)。耕地降水量是指天然降雨降落在耕地上的
有效降水量,即绿水分量;而耕地灌溉量则是耕地接
受的灌溉水量,即蓝水分量。该框架在水平衡分析的
基础上,进一步分析中国主要粮食作物(水稻、玉米、
小麦、大豆)农田耗水,即农作物经济产量形成所实
际消耗的水量,也即农作物全生育期的实际蒸散量。
如图 1 所示,在一般情况下,生产所消耗的实际蒸
散量中,一部分是来源于灌溉“蓝水”的蒸散量(蓝
水耗水量),另外一部分则来源于天然降水“绿水”
的蒸散量(绿水耗水量)。两个分量的总和即为形
成农作物经济产量的总耗水量。因此,蓝水耗水量
与总耗水量之比就定义为作物产量中的“蓝水(灌
溉)贡献率”。同理,降水耗水量与总耗水量之比
就定义为作物产量中的“绿水(降水)贡献率”。
因此,基于蓝水和绿水观点的农业水管理将农业用
水置于更广阔的范畴进行考察,具有重要的理论和
现实意义。 根据基于绿水和蓝水的中国广义农业可用水资源
的理论分析框架,在前期研究中,笔者应用水文模型
和统计数据,分析计算了全国、流域和省域水平的广
义农业可用水资源,及其绿水和蓝水组成分量。其中
绿水分量的计算主要根据区域降水和耕地空间分布,
以及基于水文模型的耕地土壤径流、入渗、深层渗漏
等水文过程。而蓝水的计算则除了根据上述水文过程
计算外,还根据水文-作物耦合建模中基于灌溉监测站
点的作物灌溉定额,最后与中国水资源公报中报告的
灌溉水量进行比较验证。该方法在全国和省域水平进
行了广泛的验证,证明了计算结果的可靠[5,28]。在此
框架的理论和计算基础上,笔者计算了中国近 15 年来
(1998—2012 年)的广义农业可用水资源量(表 1)、
和其中蓝水和绿水的比例(图 2)。
表 1 中国 1998—2012 年四大粮食作物(水稻、玉米、小麦、大豆)广义农业可用水资源量及其绿水(耕地降水)和蓝水
(耕地灌溉)分量(×108 m
3)的描述性统计特征值
Table 1 Broadly-defined Available Water for Agriculture (BAWA) for four major grain crops (rice, maize, wheat, soybean), and descriptive statistics on its blue water (cropland irrigation) and green water (cropland precipitation) components (1998-2012)
项目 Item 最大值 Max 最小值 Min 平均值 Mean 中值 Median 变异系数 CV
耕地降水 Cropland precipitation 4794 4223 4443 4390 0.042
耕地灌溉 Cropland irrigation 3619 3117 3367 3363 0.042
广义农业可用水资源 Broadly-defined Available Water for Agriculture (BAWA)
8226 7568 7810 7832 0.027
近 15 年来,中国广义农业可用水资源最大值为
8 226×108 m3,最小值 7 568×108 m3,平均值 7 810×108 m3(表 1)。在广义农业可用水资源的两个分量中,
耕地降水量和耕地灌溉量在 15 年间的变异系数基本
相等。两个分量的变异系数都高于总量的变异系数,
说明广义农业可用水资源比两个分量的稳定性高。 其中,耕地降水量,即绿水分量的比例最大值
58.8%,最小值 53.9%,平均值 56.9%;耕地灌溉量,
即蓝水分量的比例最大值 46.1%,最小值 41.2%,平
均值 43.1%(图 2)。耕地灌溉占比的变异系数为 0.037,而耕地降水占比的变异系数为 0.028,表明在广义农业
可用水资源中,绿水占比的稳定性高于蓝水占比。相
应地,中国 15 年来粮食产量中绿水贡献率和蓝水贡献
率的平均值分别为 59.8%和 40.2%,基本上是 6﹕4 的
关系(图 3),说明绿水在中国粮食生产中占有主要
的比例。而全球粮食生产中绿水﹕蓝水是 8﹕2[23],表
17 期 李保国等:蓝水和绿水视角下划定“中国农业用水红线”初索 3497
53.9 56.2 58.3 55.5 58.2 58.8 57.5 57.7 56.6 57.3 57.0 53.9 58.3 55.7 58.4
46.1 43.8 41.7 44.5 41.8 41.2 42.5 42.3 43.4 42.7 43.0 46.1 41.7 44.3 41.6
0
10
20
30
40
50
60
70
80
90
100
1998 1999 2000 2001 2002 2003 2004 2005 2006 2007 2008 2009 2010 2011 2012
年份 Year
绿水比例 Green water share 蓝水比例 Blue water share比
例 S
hare
(%)
图 2 中国 1998—2012 年四大粮食作物广义农业可用水资源量中蓝水和绿水的比例
Fig. 2 Share of green water and blue water in Broadly-defined Available Water for Agriculture (BAWA) of four major grain crops
(rice, wheat, maize, soybean) (1998-2012)
0
10
20
30
40
50
60
70
80
90
100
松辽Songliao
海河Hai
黄河Huang
淮河Huai
长江Chang
东南Southeast
珠江Zhu
水稻蓝水 Rice blue 水稻绿水 Rice green
松辽Songliao
海河Hai
黄河Huang
淮河Huai
长江Chang
东南Southeast
珠江Zhu
小麦蓝水 Wheat blue 小麦绿水 Wheat green
松辽Songliao
海河Hai
黄河Huang
淮河Huai
长江Chang
东南Southeast
珠江Zhu
玉米蓝水 Maize blue 玉米绿水 Maize green
松辽Songliao
海河Hai
黄河Huang
淮河Huai
长江Chang
东南Southeast
珠江Zhu
大豆蓝水 Soybean blue 大豆绿水 Soybean green
0
10
20
30
40
50
60
70
80
90
100
0
10
20
30
40
50
60
70
80
90
100
0
10
20
30
40
50
60
70
80
90
100
贡献率
Sha
re (%
)贡
献率
Sha
re (%
)
图 3 中国 1998—2012 年主要粮食作物(水稻、小麦、玉米、大豆)流域尺度绿水贡献率和蓝水贡献率
Fig. 3 Share of green and blue water of four major grain crops (rice, wheat, maize, soybean) at river basins (1998-2012)

3498 中 国 农 业 科 学 48 卷
明中国粮食生产中的蓝水占比远远高于世界水平。综
上所述,无论从绿水的绝对数量,还是从绿水的占比
及其在广义农业可用水资源中的稳定性来说,绿水都
在粮食生产中发挥了重要作用,因此,中国在确保粮
食安全的作物生产中,要同等重视绿水和蓝水。 中国主要粮食作物中,蓝水和绿水贡献率的区域
间差异较大。水稻在北方的松辽、黄、淮、海河流域
主要依靠蓝水,而南方的长江、东南和珠江流域水稻
的蓝水贡献率都低于 50%。在中国小麦主产区的海河、
黄河以及淮河流域部分地区,蓝水贡献率普遍超过
60%,黄河和海河都超过了 70%。而玉米的蓝水贡献
率在所有流域都未超过 40%,而大豆基本依靠绿水种
植(图 3)。在广义农业可用水资源中,蓝水比例(15年平均值)为 43.1%,绿水比例为 56.9%;而在最终
实际消耗在粮食生产中的蓝水和绿水比例分别是
40.2%和 59.8%。这表明绿水的实际利用率高于蓝水,
因此今后需要进一步提高蓝水的利用率。
2 基于水分生产力和广义农业可用水资
源的“农业用水红线”划定
“农业用水红线”是指为确保粮食安全和水安全的
目标,分配给农业(种植业、养殖业)的最低限度的
用水量。其中包括配置给农业生产的蓝水量(农田灌
溉;林、果、草地灌溉;鱼塘补水;牲畜用水),以
及耕地接受的天然降水的绿水量。 本文提出的农业用水红线主要根据四大粮食作物
(水稻、玉米、小麦、大豆)的用水和耗水特征。但
是在农田灌溉中,还包括蔬菜、棉花等其他作物。但
这四大粮食作物的耗水占作物耗水的主要部分,所以
先以这四大作物作为划定的基础。 其中,蓝水取水量根据水分生产力、蓝水贡献率
和蓝水耗水率等指标倒推。本文定义的水分生产力是
指粮食作物经济产量与作物生育期实际蒸散量之比
(kg·m-3),反映了作物单位实际蒸散耗水形成的经
济产量,是一系列生物物理(气候、品种、耕地)和
社会经济(结构、管理)因素的综合体现,所以依据
水分生产力倒推粮食生产的蓝水取水量具有很大程度
的合理性。 “用水红线”中的绿水分量是指耕地降水量,主要
受耕地本身性质和天然降水量的影响。耕地本身性质
包括:耕地数量、质量、空间位置、地质地貌类型、
地形,以及数量和质量的时空变异等因素。随着 18
亿亩耕地红线划定和更严格的执行,以及高标准农田
建设逐步推广,耕地吸纳保持天然降水的能力将基本
保持不变,并有可能适度提升。而天然降水量受到气
候模式和气候变化趋势的影响,具有一定的不确定性。
但是根据相关研究,未来气候变化趋势和降水变化以
及农业生产之间的关系可以定量判断[31-32]。因此,在
1.2 亿 hm2耕地红线下的“农业用水红线”的绿水分量
是可以相对准确地划定的。在划定农业用水红线的时
候,除了要考虑蓝水和绿水分量外,还要考虑不同降
水、水文和气象年型(干、湿、常)的影响,综合考
虑最终的水量。 2.1 农业用水红线之蓝水分量的划定——基于水分
生产力情景的高、中、低值
笔者采用 15 年来粮食作物水分生产力的现状预
测倒推 2020 年中国粮食作物生产的需水量。1998—2012 年间,中国粮食生产首先经历了为期 5 年的下滑
期,从 1998 年的历史最高产量持续下滑到 2003 年。
从 2004 年起,粮食生产实现“11 连增”。本研究计
算的水分生产力的变化趋势与粮食总产趋势相同,先
下滑至 2003 年,2004 年起持续提高。由于 1998—2003年的粮食生产下滑是不正常状态,所以,在分析近 15年的水分生产力情况用于未来粮食需水量预测时,本
文剔除了这 6 年的数据,而是根据 2004—2012 年水分
生产力的统计特征值(最大值、最小值、中值)来分
析预测未来的粮食生产需要的灌溉量(表 2)。 因此,2004—2012 年粮食作物水分生产力(作物
经济产量/作物实际蒸散量)的最大值为 1.030 kg·m-3,
相当于每 1 t 粮食需要耗水(蒸散量)971 m3;中值为
0.942 kg·m-3,相当于吨粮耗水 1 062 m3;最小值为
0.839 kg·m-3,相当于吨粮耗水 1 192 m3。假设到 2020年,中国粮食水分生产力就有可能分别达到这 3 个水
平,那么,要生产 5.5×108 t 粮食就需要分别消耗 5 340×108 m3,5 839×108 m3和 6 555×108 m3 的水;而在
这些耗水量中,来源于灌溉水(蓝水)的耗水量占
40.8%,即分别为2 179×108 m3,2 382×108 m3和2 675×108 m3;又由于灌溉水耗水率(实际消耗/毛灌溉水
量)为 0.63[33],则需要的灌溉水总量分别为 3 459×108 m3,3 782×108 m3和 4 245×108 m3, 平均为 3 819×108 m3。
近 15 年来(1998—2012 年),灌溉水占农业用
水的比例一直保持在 90%—95%之间,假设 2020 年灌
溉水占农业用水总量的 90%(乐观估计),那么农业
用水量的低、中、高值应该分别应该为 3 905×108 m3、

17 期 李保国等:蓝水和绿水视角下划定“中国农业用水红线”初索 3499
表 2 中国 1998—2012 年粮食作物水分生产力(kg·m-3)和满足 2020 年 5.5×108 t 粮食耗水的蓝水取水量(×108 m3)高、
中、低值预测
Table 2 Summary on crop water productivity (kg·m-3)from 1998-2010 in China, and projections on blue water withdrawal (×108 m3) based on high, medium and low scenarios to meet water requirements of 5.5×108 t grain production
项目 Item 高值 Max 中值 Median 低值 Min 平均值 Mean
粮食水分生产力 Crop water productivity (kg·m-3) 1.030 0.942 0.839 0.933
5.5×108 t 粮食耗水 Water consumption by 5.5×108 t grains (×108 m3) 5340 5840 6556 5897
蓝水取水量 Blue water withdrawal (×108 m3) 3459 3782 4245 3819
水分生产力的高、中、低值取的是 2004—2012 年间达到的水分生产力的最大值、中值和最小值。蓝水取水量的低、中、高值分别对应的是水分生产
力的高、中、低值 The max., median and min. of crop water productivity are the values obtained from 2004-2012. The min., median and max. of blue water withdrawals correspond to the max., median and min. of crop water productivity
4 270×108 m3 和 4 793×108 m3。因此,2020 年中国保
证粮食安全的灌溉水需求总体平均应该在 3 800×108 m3左右,而基本农业用水量应该在 4 300×108 m3左
右。综上所述,这就是保持中国粮食安全所必需的农
业用水红线中的蓝水分量。 2.2 农业用水红线之绿水分量的划定——基于近 15
年历史的高、中、低值
与农业用水红线的蓝水分量相比,确定红线中的
绿水分量具有更大的不确定性,因为要受到降水的时
空不确定性、耕地数量和质量及其时空分布的影响。
在本研究掌握的现有数据条件下,根据近 15 年来的广
义农业可用水资源广义农业可用水资源量中的绿水分
量(表 1),笔者建议的农业用水红线之绿水分量的
高、中、低值分别为 4 800×108、4 400×108和 4 200×108 m3,平均 4 470×108 m3左右。
3 讨论和建议
目前,国内有关确保中国粮食安全的水资源预测
普遍采用的是灌溉利用系数和灌溉定额的方法[34-36],
其他研究中对农业用水红线划定建议的主要根据是:
目前中国农业每年缺水 300×108 m3,1997 年以来农
业用水基本稳定在 3 700×108 m3左右,考虑到未来节
水潜能和水资源供给情况,粗略地估算农业用水红线为
3 700×108—3 900×108 m3左右,平均为 3 800×108 m3
左右[26-27]。而本文根据近 10 年来水分生产力的实际情
况,推断出确保中国粮食安全的蓝水红线在 3 800×108 m3左右,结论相似。
本研究还有许多不足和需要完善之处。在蓝水分
量中,对满足粮食耗水的灌溉取水量的计算较为可靠。
但由于蓝水分量中还包括其他蓝水取水,如林、果、
草地的灌溉、牲畜用水、鱼塘补水等项目,而目前采
用的是农田灌溉占包括上述用水在内的大致比例的方
法外推整个农业用水量,从而造成较大的不确定性。
因此,今后需要继续深入开展有关林、果、草水分生
产力水平和灌溉需求研究,并深入研究养殖业(畜禽
和水产)的用水和耗水规律,以获得更加准确的总体
蓝水分量。在绿水分量中,不确定性主要来源于耕地
本身的自然属性(数量、质量、空间、时间、土壤的
理、化、生性质等)和降水的年际变异性、年内变异
性、时空变异性等因素,特别是在气候变化情景下不
同年型(干、湿、平)的绿水分量。而本研究只是根据
近 15 年耕地水量的描述性统计数据大致估计了农业用
水红线中的绿水分量。今后,还需要在耕地和降水两方
面加强对红线中的绿水分量的准确测量和预测。 除此之外,还有一些问题需要特别关注,如:上
下游关系和区域尺度红线的划定、雨养农业、种植制
度等。除了在国家宏观层面上划定红线外,还需要在
区域(流域和行政单元)水平上进一步划定并落实用
水红线。灌溉和雨养农业之间并没有严格的界限,无
论灌溉还是雨养,都是农业水管理的连续谱系中的不
同型态,都涉及对蓝水和绿水的管理(图 1)。河流
上游雨养农业中的田间集雨设施(小型水窖),理论
上属绿水,但由于其拦截的是地表径流,如果这部分
径流不被拦截而流入河道中,就会成为蓝水资源。因
此,上游的局部集雨以提高雨养农业生产力的行为会
影响到下游用户可利用的蓝水资源量。并且,上游的
节水措施会减少流向下游的回归流,从而也影响下游
用户的可用蓝水量。这些都需要从流域角度去把握,
并且需要深入、彻底地了解流域尺度水文循环、用水、
耗水规律。从红线划定的水文水资源角度上讲,流域
或小流域是最为合适的空间单元。但由于水文单元和
行政单元大部分并不重合,因此,在划定和执行中就
会出现问题。同时,农业用水红线的划定还会受到种
植制度的影响。华南地区实行一年两熟与实行一年三

3500 中 国 农 业 科 学 48 卷
熟的蓝水需要量就大为不同。划定以及执行农业用水
红线,还需要更多、更深入的研究和实践,根据上述
计算和讨论,本研究提出下列相关建议: 3.1 牢固树立“土地利用决策就是水资源利用决策”
的观念,严格保护和实行“18 亿亩耕地红线”,
加强耕地基础设施建设,提高旱涝保收耕地面积
和比例 耕地是吸纳天然降水的重要载体,也是蓝水和
绿水转化的重要场所。耕地所吸纳的降水量在广义
农业可用水资源量中占有相当比例(58%),这也
从水资源的角度诠释了国家严格保护 1.2 亿 hm2 耕
地红线的重要意义。在 1.2 亿 hm2 耕地条件下,耕
地绿水应该在 4 300×108 m3 左右。保证 1.2 亿 hm2
耕地实际上是保证了农业用水红线中的绿水分量。
但由于降雨和作物生长发育期需水的不完全匹配,
灌溉蓝水是保证稳产高产的必要条件,所谓“有产
无产在绿水,稳产高产靠蓝水”。除东南(74.2%)
外,华北(66.3%)、东北(60.2%)、西南(64.9%)、
西北(57.4%)旱涝保收面积占总灌溉面积的百分
比均低于全国(67.0%)水平。尤其是在事关粮食
安全安危的华北和东北,要继续加大农田基础设施
投资,提高高标准农田和旱涝保收农田的面积和比
例。 3.2 加强以耕地土壤水为中心的农业用水和耗水管
理,继续重视并提高农艺节水措施在耕地土壤水
管理上的巨大作用 节水农业的最终目的是提高单位水资源的农业产
出效率,在农田尺度上就是提高作物对土壤水分的利
用效率,即消耗单位土壤水量而获得的经济产量。土
壤水是“四水”(降水、地表水、土壤水、地下水)
动态变化和转化的核心,也是“蓝水”和“绿水”最
终的交汇点[37]。加强土壤水管理的措施包括建立完善
农田土壤墒情监测;改善土壤理化生性质,提高有效
土壤水分含量;增加“生产性绿水流(植物蒸腾)”
在“总绿水流(实际蒸发蒸腾)”中的比例;减少“非
生产性绿水流(土面蒸发)”的比例;尽量将土面蒸
发转化为植物蒸腾。2007 年,东北农艺节水技术和措
施的采纳面积占全国的比例最高,达到 38.8%,华北
次之,为 23.9%;西南 17.5%;西北 12.3%;东南最低
7.5%[38]。总体上,西北和东南应该大力加强农艺节水
技术措施的应用力度。各个地区针对不同特点创造出
经济实用的节水技术,如西北和东北的膜下滴灌,水
肥同时施入,可以成倍地提高作物水分生产力,是值
得大力推广的技术。 3.3 利用当地水热资源特点,调整种植结构,“以水
定产”,发展“适水种植制度”
华北主要实行冬小麦-夏玉米种植制度,但是小
麦生长期严重缺水,需要大量灌溉,而且 50%以上
来源于地下水。但是玉米不仅水分生产力较高,而
且生长季可以充分利用自然降水。因此,在满足国
家粮食品种需求的前提下,如何充分利用主要粮食
作物的需水和耗水特性,合理优化作物蓝水的配置,
针对各地区水热资源特点,调整种植结构,建立水
资源高效利用的种植体系是今后需要加强的研究和
政策方向。 3.4 继续提高灌溉的保证程度
水库库容与耕地的匹配程度在一定程度上反映灌
溉保障状况。全国单位耕地的总库容匹配量为 5 203 m3·hm-2。分区来看,东南的水库潜在供水能力最高,
单位耕地的总库容匹配量达到 10 647 m3·hm-2,是全国
平均水平的 2 倍。其次是西北,3 925 m3·hm-2,是全
国平均水平的 76%;东北区,3 861 m3·hm-2,为全国
平均水平的 74%;华北区,3 345 m3·hm-2,为全国平
均水平的 64%;西南最少,只有 3 159 m3·hm-2,为全
国平均水平的 61%[39]。因此,需要进一步加强除了东
南地区以外所有地区的灌溉保障能力。 3.5 继续提高农业水分生产力,包括从种植业到养殖
业的整体的水分生产力
不仅要提高物质水分生产力(kg·m-3),还要提
高经济水分生产力(元/m3)。最近 15 年,中国粮
食作物水分生产力的提高主要原因有:单产提高、
种植结构调整(水分生产力较高的玉米播种面积占
比和产量双提高)、管理措施到位(品种、栽培、
节水、施肥、植保、耕作、田间管理措施)。但需
认识到水分生产力在单产较低的区间内提升潜力较
大,而在单产较高的情况下提升幅度有限[41]。而中
国粮食总体单产水平已经较高,因此需要更大的努
力提高水分生产力。 3.6 为了确保“农业用水红线”,必须建立、健全完
整、完善的广义水资源测量和监督体系
耕地的空间位置是固定的,“1.2 亿 hm2 耕地红
线”的测量和监督可以通过直接测量和统计的方法获
得。但是,水是运动的,所以对其准确测度更加困难。
一方面,国家需加大灌溉水计量的基础设施和技术设
备建设,加强对灌溉“蓝水”的用水度量。另一方面,
国家应加强在田间和区域尺度准确测度“绿水”的测

17 期 李保国等:蓝水和绿水视角下划定“中国农业用水红线”初索 3501
量技术和设备的理论和方法研究。只有能够相对准确
地测量耕地可利用的“蓝水”和“绿水”,才能够切
实保证“农业用水红线”落到实处。同时,加强对气
候变化对水资源和农业影响和适应性策略的基础、应
用和政策研究。
4 结论
笔者在对广义农业可用水资源的理论和计算分
析的基础上,初步提出了确保中国粮食安全的“农
业用水红线”,并计算了其中的蓝水分量和绿水分
量,同时还讨论了计算的不确定性和改进方向。与
相关研究比较,本研究划定的红线区间较为合理。
中国将实行最严格的水资源管理制度,到 2030 年,
用水总量红线将控制在 7 000×108 m3 左右,本文划
定的农业用水红线在 4 300×108 m3 左右,占 61%左
右,而目前农业用水占总用水量的比例为 62%。因
此,这条农业用水红线具有很大的现实可能性和实
现保障。农业用水红线中绿水分量的保证主要依靠
保证耕地红线和高标准农田建设,提高耕地截蓄、
吸纳、有效利用天然降水的能力。同时,继续全面
提升作物水分生产力,提高蓝水和绿水的利用率和
产出效率。“保蓝水、稳绿水、提效率”是保证未
来的中国粮食安全用水的重要战略举措。 最后,笔者提出的“农业用水红线”主要基于四
大粮食作物的耗水规律和水分生产力特征,因为这四
大粮食作物是作物耗水的主体。但今后还需进一步补
充蔬菜、棉花等作物的耗水和水分生产力特征,以及
果树、畜牧、渔业生产的用水和耗水特性,不断完善
“农业用水红线”的划定。
References
[1] 陈印军, 肖碧林, 方琳娜, 马宏岭, 杨瑞珍, 易小燕, 李倩倩. 中国
耕地质量状况分析. 中国农业科学, 2011, 44(17): 3557-3564.
Chen Y J, Xiao B L, Fang L N, Ma H L, Yang R Z, Yi X Y, Li Q Q.
The quality analysis of cultivated land in China. Scientia Agricultura
Sinica, 2011, 44(17): 3557-3564. (in Chinese)
[2] 张维理, 武淑霞, 冀宏杰, Kolbe H. 中国农业面源污染形势估计及
控制对策 I. 21 世纪初期中国农业面源污染的形势估计. 中国农业
科学, 2004, 37(7): 1008-1017.
Zhang W L, Wu S X, Ji H J, Kolbe H. Estimation of agricultural
non-point source pollution in China and the alleviating strategies I.
Estimation of agricultural non-point source pollution in China in early
21 Century. Scientia Agricultura Sinica, 2004, 37(7): 1008-1017. (in
Chinese)
[3] 杨飞, 吴根义, 诸云强, 付强. 中国各省区未来主要畜禽养殖量及
耕地氮载荷的预测. 水土保持研究, 2013, 3: 289-294.
Yang F, Wu G Y, Zhu Y Q, Fu Q. Forecast of livestock and poultry
amount and arable land nitrogen load of Chinese provinces. Research
of Soil and Water Conservation, 2013, 3: 289-294. (in Chinese)
[4] Ridoutt B G, Pfister S. A revised approach to water footprinting to
make transparent the impacts of consumption and production on
global freshwater scarcity. Global Environmental Change-Human and
Policy Dimensions, 2010, 20: 113-120.
[5] 李保国, 黄峰. 1998-2007 年中国农业用水分析. 水科学进展, 2010,
4: 575-583.
Li B G, Huang F. Trends in China’s agricultural water use during
recent decade using the green and blue water approach. Advances in
Water Science, 2010, 4: 575-583. (in Chinese)
[6] 陈锡文. 中国粮食安全面临五大历史性挑战. http://www.chinagrain.
cn/liangyou/2014/11/24/201411241485255566.shtml
Chen X W. Five major challenges presented to food security in China.
http://www.chinagrain.cn/liangyou/2014/11/24/201411241485255566.
shtml (in Chinese)
[7] Falkenmark M. Meeting water requirements of an expanding world
population. Philosophical Transaction of Royal Society of London,
Series B, 1997: 352, 929-936.
[8] Molden D J. Accounting for water use and productivity. SWIM Paper
1. System-wide initiative for water management. International Water
Management Institute, Colombo, Sri Lanka, 1997: 16.
[9] Seckler D. The new era of water resources management: From ‘dry’ to
‘wet’ water savings. Research Report 1. International Irrigation
Management Institute, Colombo, 1996.
[10] Keller A A, Keller J. Effective efficiency: A water use efficiency
concept for allocating freshwater resources. Discussion Paper 22.
Center for Economic Policy Studies, Winrock International, 1995: 19.
[11] Droogers P, Kite G. Water productivity from integrated basin modeling.
Irrigation and Drainage Systems, 1999, 13(3): 275-290.
[12] Allan J A. Virtual Water: A strategic resource global solutions to
regional deficits. Groundwater, 1998, 36: 545-546.
[13] Molden D, Faurès J M, Finlayson C M, Gitay H, Muylwijk J,
Schipper L, Vallée D, Coates D. Chapter 1. Setting the scene//Water
for Food, Water for Life–A Comprehensive Assessment of Water
Management in Agriculture. Earthscan, Routeledge. Taylor & Francis,
2007.
[14] Rockstrom J, Lannerstad M, Falkenmark M. Assessing the water
challenge of a new green revolution in developing countries//

3502 中 国 农 业 科 学 48 卷
Proceedings of the National Academy of Sciences of the United States
of America, 2007, 104(15): 6253-6260.
[15] de Fraiture C. Integrated water and food analysis at the global and
basin level. An application of WATERSIM. Water Resources
Management, 2007, 21(1): 185-198.
[16] Rost S, Gerten D, Bondeau A, Lucht W, Rohwer J, Schaphoff S.
Agricultural green and blue water consumption and its influence on
the global water system. Water Resources Research, 2008, 44(9), doi:
10.1029/2007WR006331.
[17] Schuol J, Abbaspour K C, Srinivasan R, Yang H. Estimation of
freshwater availability in the West African sub-continent using the
SWAT hydrologic model. Journal of Hydrology, 2008, 352(1/2):
30-49.
[18] Schuol J, Abbaspour K C, Yang H, Srinivasan R, Zehnder A J B.
Modeling blue and green water availability in Africa. Water
Resources Research, 2008, 44(7), doi: 10.1029/2007wr006609
[19] Faramarzi M, Abbaspour K C, Schulin R, Yang H. Modelling blue
and green water resources availability in Iran. Hydrological Processes,
2009, 23(3): 486-501.
[20] Chiu Y W, Wu M. Assessing county-level water footprints of
different cellulosic-biofuel feedstock pathways. Environmental
Science & Technology, 2012, 46(16): 9155-9162.
[21] Sulser T B, Ringler C, Zhu T J, Msangi S, Bryan E, Rosegrant M W.
Green and blue water accounting in the Ganges and Nile basins:
Implications for food and agricultural policy. Journal of Hydrology,
2010, 384(3/4): 276-291.
[22] Antonelli M, Roson R, Sartori M. Systemic input-output computation
of green and blue virtual water ‘flows’ with an illustration for the
Mediterranean Region. Water Resources Management, 2012, 26(14):
4133-4146.
[23] Siebert S, Doll P. Quantifying blue and green virtual water contents in
global crop production as well as potential production losses without
irrigation. Journal of Hydrology, 2010, 384(3/4): 198-217.
[24] Chapagain A M, Hoekstra A Y. The blue, green and grey water
footprint of rice from production and consumption perspectives.
Ecological Economics, 2011, 70(4): 749-758.
[25] Liu J G, Yang H. Spatially explicit assessment of global consumptive
water uses in cropland: Green and blue water. Journal of Hydrology,
2010, 384(3/4): 187-197.
[26] 罗其友, 高明杰, 姜文来, 陶陶, 唐曲. 农业综合生产能力安全与
我国农业资源安全阈值测算. 农业现代化研究, 2010, 4: 392-396.
Luo Q Y, Gao M J, Jiang W L, Tao T, Tang Q. Study on resources
threshold values based on agricultural comprehensive production
capability security in China. Research on Agricultural Modernization.
2010, 4: 392-396. (in Chinese)
[27] 姜文来. 支撑粮食安全的农业水资源阈值研究与展望. 农业展望,
2010, 9: 23-25.
Jiang W L. Research and prospects on agricultural water use threshold
that is to support food security. Agriculture Prospects, 2010, 9: 23-25.
(in Chinese)
[28] Li B G, Huang F. Chapter 12, China’s food security and soil water
management-a green and blue water approach. Soil Water and
Agronomic Productivity//Lal R, Stewart B A. Advances in Soil
Science Vol.19. CRC Press. Taylor and Francis. 325-346.
[29] Huang F, Li B G. Assessing grain crop water productivity of China
using a hydro-model-coupled-statistics approach Part I: Method
development and validation. Agricultural Water Management, 2010,
97(7): 1077-1092.
[30] Huang F, Li B G. Assessing grain crop water productivity of China
using a hydro-model-coupled-statistics approach. Part II: Application
in breadbasket basins of China. Agricultural Water Management,
2010, 97(9): 1259-1268.
[31] 吴普特, 赵西宁. 气候变化对中国农业用水和粮食生产的影响. 农
业工程学报, 2010, 26(2): 1-6.
Wu P T, Zhao X N. Impact of climate change on agricultural water
use and grain production in China. Transactions of the Chinese
Society of Agricultural Engineering, 2010, 26(2): 1-6. (in Chinese)
[32] 王小军, 张建云, 王国庆, 贺瑞敏, 尚熳廷. 气候变化与农业用水
安全. 中国农村水利水电, 2012, 2: 23-25, 29.
Wang X J, Zhang J Y, Wang G Q, He R M, Shang M T. Climate
change and agricultural water security. Chinese Rural Water
Development and Hydraulic Electricity, 2012, 2: 23-25, 29. (in
Chinese)
[33] 水利部. 2012 年中国水资源公报. 北京: 中国水利水电出版社,
2013.
Ministry of Water Resources. Chinese Water Resources Bulletin 2012.
Beijing: Chinese Water and Hydraulics Electricity Press, 2013. (in
Chinese)
[34] 王玉宝, 吴普特, 赵西宁, 李甲林. 我国农业用水结构演变态势分
析. 中国生态农业学报, 2010, 18(2): 399-404.
Wang Y B, Wu P T, Zhao X N, Li J L. Development tendency of
agricultural water structure in China. Chinese Journal of Eco-
Agriculture, 2010, 18(2): 399-404. (in Chinese)
[35] 吴普特, 赵西宁, 冯浩, 王玉宝. 农业经济用水量与我国农业战略
节水潜力. 中国农业科技导报, 2007, 6: 13-17.
Wu P T, Zhao X N, Feng H, Wang Y B. Agricultural economic

17 期 李保国等:蓝水和绿水视角下划定“中国农业用水红线”初索 3503
rational water consumption and strategy analysis for water saving
potential in China. Journal of Agricultural Science and Technology,
2007, 6: 13-17. (in Chinese)
[36] 山仑, 张岁岐. 能否实现大量节约灌溉用水?——我国节水农业现
状与展望. 自然杂志, 2006, 2: 71-74.
Shan L, Zhang S Q. Is possible to save large irrigation water?-The
situation and prospect of water-saving agriculture in China. Chinese
Journal of Nature, 2006, 2: 71-74. (in Chinese)
[37] Li B G, Gong Y S, Zuo Q. Dynamic Modeling on Farmland Soil Water
and Its Applications. Beijing: Science Press, 2000. (in Chinese)
[38] 李保国, 彭士琪. 中国农业用水报告 1998-2007 年. 北京: 中国农
业出版社, 2009.
Li B G, Peng S Q. Chinese Agricultural Water Use Bulletin 1998-2007.
Beijing: Chinese Agricultural Press, 2009. (in Chinese)
[39] 全国农业技术推广服务中心. 中国农业用水报告 2008 年. 北京:
中国农业出版社, 2013.
National Agricultural Technology Extension Service. Chinese
Agricultural Water Use Bulletin 2008. Beijing: Chinese Agricultural
Press, 2013. (in Chinese)
[40] 全国农业技术推广服务中心.中国农业用水报告 2009 年. 北京: 中
国农业出版社, 2013.
National Agricultural Technology Extension Service. Chinese
Agricultural Water Use Bulletin 2009. Beijing: Chinese Agricultural
Press, 2013. (in Chinese)
(责任编辑 张晶,杨鑫浩)

中国农业科学 2015,48(17):3504-3514 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.016
收稿日期:2015-03-06;接受日期:2015-05-29 基金项目:国家自然科学基金创新研究群体项目(31221062) 联系方式:刘彬,E-mail:[email protected]。通信作者邓秀新,E-mail:[email protected]
基于文献计量的园艺学基础研究发展状况分析
刘 彬1,邓秀新
2
(1 华中农业大学科学技术发展研究院,武汉 430070;2 华中农业大学园艺林学学院/园艺植物生物学教育部重点实验室,武汉 430070)
摘要:【目的】了解基于 SCI 收录文献视角的园艺学基础研究发展状况,为相关科研工作者和管理决策者
提供参考。【方法】运用文献计量学的方法,通过 WoS 和 Citespace 对 2005-2014 年间以 60 余种常见园艺植物
为题目的 SCI 收录文献进行文献总量、年度分布、篇均他引次数、h 指数、研究机构、高影响因子期刊载文量、
文献研究方向及研究热点的分析。【结果】10 年间,以所选 60 余种常见园艺植物为题目的 SCI 收录文献数量
逐年上升,中国的文献数位居全球第二位,仅次于美国。2010-2014 年,中国园艺学文献数跃居全球第一。对
2005-2009 年与 2010-2014 年之间的文献进行比较,以相关果树为题目,中国的文献所占比例从 8.40%上升到
15.68%,文献数排名从第 3 位上升到第 2 位;在文献数排名前 10 位的国家中,中国文献的篇均他引次数从第 5
位上升到第 4 位,h 指数排名从第 3 位上升到第 2 位;文献数量全球最多的 25 个科研机构中,中国的科研机构
从 4 家增加至 7 家。以相关蔬菜(含西甜瓜)为题目,中国的文献所占比例从 8.79%上升到 16.90%;文献数两
个 5 年中排名均居第 2 位;在文献数排名前 10 位的国家中,中国文献的篇均他引次数两个 5 年中排名均居第 6
位,h指数从第 5位上升到第 2位;文献数量全球最多的 25 个科研机构中,中国的科研机构从 3 家增加至 5家。
以相关花卉和茶为题目,中国的文献所占比例从 14.12%上升到 26.14%,文献数排名从第 3位上升到第 1位;在文
献记数排名前 10 位的国家中,中国文献的篇均他引次数两个 5年均居第 7位,h指数排名从第 3位上升到第 2位;
文献数量全球最多的 25 个科研机构中,中国的科研机构从 3 家增加至 8 家。10 年间,生物学领域高影响因子杂
志刊载中国园艺学文献数排名从第 8位上升到第 5位;文献数量全球最多的 25 个科研机构中,中国的科研机构从
1 家增加至 3 家。研究方向分析显示,中国园艺学文献在生物化学与分子生物学方向比例较多,但在农学和昆虫
领域的文献比例较少。目前园艺学的研究热点主要集中在功能基因挖掘与组学研究、果实的发育与成熟以及多酚
类物质研究等 3 个领域。【结论】近 5 年,中国园艺学被 SCI 收录文献的数量在增加,产出已居世界第一,但质
量提升还未跟上;中国园艺学研究侧重于分子和组学等偏基础研究方向,与农业结合相对紧密的研究文献数量比
例偏少;未来应加强问题导向的研究,布局上应更加贴近产业各环节的问题,持之以恒,形成特色。
关键词:园艺学;基础研究;SCI;文献计量;发展状况;WoS;Citespace
Basic Research Development Status of Horticulture
Based on Bibliometric Analysis LIU Bin1, DENG Xiu-xin2
(1Office of Scientific Technology & Development, Huazhong Agricultural University, Wuhan 430070; 2College of Horticulture &
Forestry Sciences, Huazhong Agricultural University/Key Laboratory of Horticultural Plant Biology, Ministry of Education,
Wuhan 430070)
Abstract:【Objective】In order to provide data support for horticulture researchers and administrative decision makers, this study surveyed the basic research development status of horticulture based on papers published in SCI-indexed journals. 【Method】
jia
文本框
返回目录

17 期 刘彬等:基于文献计量的园艺学基础研究发展状况分析 3505
Bibliometric analysis was carried out to investigate the papers published in SCI-indexed journals during 2005-2014 with titles relevant to more than 60 kinds of common horticultural plants by using comparative analysis with WoS & citespace on annual distribution, average cited times without self-citations, h-index, countries (regions), institutions, publication in high-impact journals, research areas, hotspots, etc. 【Result】The study showed that the annual quantity of papers published in SCI-indexed journals had been increased year by year in the past decade and China had become the 2nd largest country in the number of papers after the USA. The number of Chinese horticultural papers became the largest in the world between 2010 and 2014. Comparing the papers with selected fruits as the theme between 2005-2009 and 2010-2014, it was found that the proportion of Chinese horticultural papers had risen from 8.40% to 15.68%, and the ranking had risen from 3rd to 2nd. The average cited times without self-citations of Chinese papers had risen from 5th to 4th, and the h-index had risen from 3rd to 2nd, in the top 10 countries of the paper records ranking. The number of Chinese institutions that ranked in the top 25 around the world had increased from 4 to 7. Comparing the papers with selected vegetables (including melon & watermelon) as the theme between 2005-2009 and 2010-2014, it was found that the proportion of Chinese papers had risen from 8.79% to 16.90%, and the ranking had remained at 2nd. The average cited times without self-citations of Chinese papers had remained 6th and the h-index ranking had risen from 5th to 2nd in the top 10 countries of the paper records ranking. The number of Chinese institutions that ranked in the top 25 around the world had increased from 3 to 5. Comparing the papers with selected flowers & tea as the theme between 2005-2009 and 2010-2014, it was found that the proportion of Chinese papers had risen from 14.12% to 26.14%, and the ranking had risen from 3rd to 1st. The average cited times without self-citations of Chinese papers had remained 7th and the h-index ranking had risen from 3rd to 2nd in the top 10 countries of the paper reorders ranking. The number of Chinese institutions that ranked in the top 25 around the world had increased from 3 to 8. The number of Chinese horticultural papers published in high-impact biology journals increased rapidly during the past decade with its ranking rising from 8th to 5th. The number of Chinese institutions that ranked in the top 25 around the world had increased from 1 to 3. The analysis of research area showed that the proportion of Chinese horticultural papers was large in biochemistry and molecular biology, but smaller in the fields of entomology and agriculture. The research hotspots were high quality draft genome, fruit development and maturation and phenolic compound. 【Conclusion】The number of Chinese horticultural papers published in SCI-indexed journals has been increased in the past five years and the output has ranked 1st in the world, but the quality has not kept up with the quantity. Compared with part of the development countries, the proportion of Chinese horticultural papers was larger in basic research direction, but smaller in the areas which are closely related to traditional agronomy. In the future, more attentions should be paid to the problem-oriented research and the connection and contribution to problems in various links of the industry.
Key words: horticulture; basic research; SCI; bibliometric; development status; WoS; citespace
0 引言
【研究意义】园艺植物一般认为包括果树、蔬菜、
花卉和茶。中国是世界园艺大国,果树、蔬菜、花卉
和茶的种植面积及产量均居世界首位[1-2]。园艺产业的
发展对中国农业乃至国计民生具有重要意义。近年来,
国内外以园艺植物为研究对象的相关基础研究不断深
入,取得了较好的成绩,相应的以园艺植物为题目的
SCI 收录文献也在逐年增加,开展以园艺植物为题目
的文献计量分析,可以从一个侧面深入了解园艺学基
础研究的发展概况。【前人研究进展】文献计量学是
对文献进行定量分析研究的科学[3]。科研论文是基础
研究成果的主要表现形式,其数量和质量是科研产出
能力及科研活动活跃程度的重要标志之一[4]。利用文
献计量学的方法对某一研究领域的科研论文进行定量
分析研究,是目前科研量化评价的重要方法之一,也
越来越受到科学界的重视[5]。文献计量学在农业领域
的应用越来越多,如邬亚文等[6]将文献计量学用于对
国内外水稻发展态势的研究;Josef 等[7]对 1950—2010年德国以苹果、梨、樱桃、李子、杏、桃等水果为题
目的相关文献进行了计量学分析;Tatry 等[8]对 2000—2009 年欧盟 27 国和美国主导的水果和蔬菜的相关
研究进行了文献计量研究;Leiser 等[9]对法国国家农业
研究院研究人员 2000—2006 年发表的相关蔬菜和
水果的文献进行了计量分析。国内外部分科研工作
者[10-20]近年以园艺学领域相关期刊刊载的文献或以相
关的园艺植物为主题的文献所做的计量分析,还涉及
椰枣、葡萄、蝴蝶兰、菊花等园艺植物。【本研究切
入点】园艺植物种类多样,分析基于以常见的 60 余种
园艺植物为题目的文献,以了解基于文献计量视角的
园艺学基础研究概况。统计分析所涉及的文献数量较
多、口径较广,目前尚未见相关报道。【拟解决的关

3506 中 国 农 业 科 学 48 卷
键问题】采用文献计量学的方法,通过对 2005—2014年 SCI 收录的以常见的 60 余种果树、蔬菜(含西甜
瓜)、花卉和茶等园艺植物为题目的 SCI 收录文献进
行计量分析,探索园艺学基础研究概况及其发展趋势,
为相关科研工作者和管理决策者提供参考。
1 数据来源及研究方法
1.1 数据来源与主要评价指标 研究所涉及的 SCI 收录文献数据均来源于 SCI-E
数据库中的检索结果。Web of Science(简称:WoS)是美国 Thomson Reuters 公司基于 Web 开发的产品,
是自然科学、社会科学、艺术和人文领域的权威学术
文献数据库[21],该库为文献计量的研究提供了方便,
SCI-E 是 WoS 三大核心库之一。 文献记录数:检索到的某一特定范围内的文献数
量。 篇均他引次数:检索到的某一特定范围内的所有
文献被他引次数除以文献记录数。 h 指数(h-index):是指一个科研工作者、科研
机构、学科、期刊或国家(地区)在一定期间内所发
表的论文至少有 n 篇的被引频次不低于 n 次,h 代表
“高引用次数”(high citations)。其目的是量化科研
人员作为独立个体的研究成果[22],因其综合考虑了生
产率(发表的论文总数)和质量(每篇论文的被引频
次)两个因素,而备受关注[23],目前 h 指数已被广泛
应用于评价科研工作者、科研机构、学科、期刊等在
一定时间内的学术成就[24-26]。 1.2 研究方法
通过 WoS 平台在 SCI-E 数据库中检索出 2005—2014 年以常见的果树、蔬菜(含西甜瓜)、花卉和茶
的英文名称为题目(表 1)的文献,用语种=English和文献类型=Article 进行精炼。编辑检索式时通过不
同的方式排除干扰文献:避免使用有多重含义的词汇
作为检索词,如 Rose 有多种含义,检索式使用 Rosa代替 Rose;通过布尔算法剔除可能产生的干扰文献,
如通过检索式 TI=(apple NOT(iphone OR ipad OR ipod OR mac OR macbook))剔除以苹果为题目的部
分干扰文献;部分园艺植物的英文名由两个单词组成,
如猕猴桃(kiwi fruit),在检索时将两个单词加英文
状态的引号,以避免检索到干扰文献等。检索时间:
2015 年 1 月 31 日。然后对文献记录数、国家(地区)、
研究机构、篇均他引次数、h 指数、研究方向和研究
热点等进行分析。 生物学领域高影响因子杂志刊载园艺学文献分
析:选取检索出的文献,通过国际知名的主流期刊名
称,如 Cell、Nature、Science、Nature Genetics、Nature Biotechnology、Nature Cell Biology、Pant Cell、Plant Physiology、Proceedings of the National Academy of Sciences of United States of America 、 Chemistry Biology、Molecular Cell Biology、New Phytologist、Journal of Cell Biology 、 PLoS Genetics 、 Gene Development、Current Biology、Trends in Plant Science、Genome Research 等进行精炼,然后进行计量分析。
研究方向分析:WoS 平台下的所有数据库都使用
了一套“研究方向”的分类方法,它实现了对数据库
中关于同一主题的文献进行识别、检索和分析。对检
索出的园艺学文献,选择相关的“研究方向”进行文
献计量分析。 研究热点分析:在检索出的园艺学文献中,选取
引用次数排名前 1%的文献,利用 CitespaceⅢ软件绘
制关键词共现和文献共被引科学知识图谱,时间阈值
选择:From 2005 To 2014,2 years per slice,节点阈值:
Top 50 per slice,通过 pathfinder 算法修剪网络,然后
通过图谱进行研究热点的识别和分析。
表 1 研究所选的常见园艺植物名称
Table 1 The name of common horticultural plants selected for studies 种类 Type 园艺植物名称 Name of horticultural plants
果树 Fruit tree
苹果 apple, 香蕉 banana, 蓝莓 blueberry, 柑橘 citrus, 樱桃 cherry, 葡萄 grape, 番石榴 guava, 桃 peach, 梨 pear, 柿 persimmon, 荔枝 litchi, 龙眼 longan, 枇杷 loquat, 芒果 mango, 木瓜 papaya, 李子 plum, 菠萝 pineapple, 草莓 strawberry, 火龙果 dragon fruit, 猕猴桃 kiwi fruit, 甜橙 sweet orange
蔬菜 Vegetable
西兰花 broccoli, 花椰菜 cauliflower, 卷心菜 cabbage, 胡萝卜 carrot, 芹菜 celery, 辣椒 chili, 豇豆 cowpea, 黄瓜 cucumber, 茄子 eggplant, 洋葱 onion, 马铃薯 potato, 萝卜 radish, 菠菜 spinach, 番茄 tomato, 生菜 lettuce, 韭菜 leek, 甜瓜 melon, 南瓜 pumpkin, 菜豆 kidney bean, 冬瓜 wax gourd, 西瓜 watermelon
花卉 Flower
红掌 anthurium, 杜鹃 azalea, 康乃馨 carnation, 兰花 cymbidium/orchid, 非洲菊 gerbera, 子花 gardenia, 唐菖蒲 gladiolus, 茉莉 jasmine, 百合 lily, 郁金香 tulip, 桂花 osmanthus, 夹竹桃 oleander, 三色堇 pansy, 牡丹 peony, 矮牵牛 petunia, 莲 lotus, 水仙 narcissus, 月季 rose, 紫薇 crape myrtle, 梅花 plum blossom
茶 Tea 茶 tea

17 期 刘彬等:基于文献计量的园艺学基础研究发展状况分析 3507
2 结果
2.1 园艺植物文献概况 2005—2014 年,以所选的 60 余种园艺植物为题
目的 SCI收录文献共计 69 497篇,文献数量逐年增加,
2014 年的文献数占总文献数的 12.05%(表 2)。SCI收录文献会晚于文献出版时间 1—2 个月上载入 WoS平台相应数据库,因检索时间为 2015 年 1 月 31 日,
部分 2014 年发表的文献还未完全载入数据库,这可能
是造成检索出的 2014 年文献数量比 2013 年略低的原
因。 从文献的国家(地区)分布来看,10 年间,以 60
余种园艺植物为题目被 SCI 收录的文献涉及的国家
(地区)共 179 个,其中,文献记录数 多的 10 个国
家中,美国的文献数量 多,中国的文献数位居第
2,与排名第 1 的美国差距较大,但比排名第 3 的日
本多了近 1 倍(表 3)。2010—2014 年间,中国园艺
学的文献数量跃居全球第一;同为发展中国家的印度、
表 2 2005-2014 年以所选园艺植物为题目的 SCI 收录文献年度分布
Table 2 The annual distribution of papers published in SCI-indexed journals with selected horticultural plants as the theme in 2005-2014
所选园艺植物 Selected horticultural plants
所选果树 Selected fruit trees
所选蔬菜(含西甜瓜) Selected vegetables(contain melon & watermelon)
所选花卉和茶 Selected flowers & tea
出版年 Publication year
记录数
Record count 比例
Percent (%) 记录数
Record count 比例
Percent (%)记录数
Record count 比例
Percent (%) 记录数
Record count比例
Percent (%)2014 8374 12.05 3512 12.33 3533 11.64 1417 12.51
2013 8561 12.32 3654 12.83 3550 11.69 1430 12.62
2012 8232 11.85 3396 11.92 3584 11.80 1349 11.91
2011 7909 11.38 3275 11.50 3413 11.24 1301 11.48
2010 7090 10.20 2923 10.26 3100 10.21 1136 10.03
2009 6664 9.59 2725 9.57 2937 9.67 1055 9.31
2008 6566 9.45 2616 9.19 2995 9.86 1020 9.00
2007 5885 8.47 2376 8.34 2635 8.68 924 8.16
2006 5334 7.68 2125 7.46 2375 7.82 892 7.87
2005 4882 7.03 1880 6.60 2240 7.38 806 7.11
2010—2014 40166 57.80 16760 58.84 17180 56.58 6633 58.54
2005—2009 29331 42.21 11722 41.16 13182 43.42 4697 41.46
表 3 2005-2014 年以所选园艺植物为题目的 SCI 收录文献数排名前 10 位的国家
Table 3 The top 10 most productive countries of papers published in SCI-indexed journals with selected horticultural plants as the theme in 2005-2014
2005—2014 2005—2009 2010—2014
国家/地区 Country/Region
记录数 Record count
比例 Percent (%)
国家/地区 Country/Region
记录数 Record count
比例 Percent (%)
国家/地区 Country/Region
记录数 Record count
比例 Percent (%)
1 美国 USA 13059 18.79 美国 USA 6168 21.03 中国 China 7187 17.89
2 中国 China 9975 14.35 中国 China 2788 9.51 美国 USA 6891 17.16
3 日本 Japan 5145 7.40 日本 Japan 2710 9.24 印度 India 2893 7.20
4 印度 India 5030 7.24 印度 India 2137 7.29 西班牙 Spain 2555 6.36
5 西班牙 Spain 4548 6.54 西班牙 Spain 1993 6.80 日本 Japan 2435 6.06
6 巴西 Brazil 3333 4.80 意大利 Italy 1385 4.72 巴西 Brazil 2182 5.43
7 意大利 Italy 3327 4.79 德国 Germany 1291 4.40 意大利 Italy 1942 4.84
8 德国 Germany 2724 3.92 加拿大 Canada 1167 3.98 韩国 Korea 1718 4.28
9 韩国 Korea 2711 3.90 巴西 Brazil 1151 3.92 德国 Germany 1433 3.57
10 法国 France 2320 3.34 法国 France 1066 3.63 土耳其Turkey 1308 3.26

3508 中 国 农 业 科 学 48 卷
巴西、土耳其文献数量也有较明显的增长,在国际上
的排名也有较明显的提高,这与世界园艺产业正向发
展中国家转移的趋势相一致。 2.2 果树学文献分析
从文献来源国家(地区)分布情况看,2010—2014年中国果树学的文献量增加迅速,并且在全球范围内
排名从 2005—2009 年的第 3 位上升到了 2010—2014年的第 2 位,文献所占比例也有很大的提升。文献
被引用次数常被用来作为文献学术水平和影响力的
评价指标[27],这是因为科研人员所发表文献的影响
力体现在同行的关注程度上,研究结果被同行在文
献中直接引用是其 重要的表现形式之一[28]。2005—2009 年,在果树学文献记录数排名前 10 位的国
家中,中国的文献篇均他引次数排名第 5,h 指数排
名第 3,2010—2014 年,篇均他引次数排名上升到
第 4 位,h 指数排名上升到第 2 位(表 4)。这说明
中国的果树学文献在数量增加的同时,质量也有了
一定幅度的提升。
表 4 2005—2014 年以所选果树为题目的 SCI 收录文献数排名前 10 位的国家
Table 4 The top 10 most productive countries of papers published in SCI-indexed journals with selected fruits as the theme in 2005-2014
2010—2014 2005—2009
国家/地区 Country/ Region
记录数 Record count
比例 Percent
(%)
篇均他引次数 Average cited times without self-citations
h 指数 h-index
国家/地区 Country/ Region
记录数 Record count
比例 Percent
(%)
篇均他引次数 Average cited times without self-citations
h 指数 h-index
1 美国 USA 2910 17.36 3.26 37 美国 USA 2602 22.20 16.08 69
2 中国 China 2628 15.68 3.49 33 西班牙 Spain 1134 9.67 19.81 58
3 西班牙 Spain 1431 8.54 4.55 27 中国 China 985 8.40 16.40 51
4 巴西 Brazil 1268 7.57 2.76 22 日本 Japan 829 7.07 13.72 44
5 印度 India 1136 6.78 2.50 22 印度 India 770 6.57 8.61 36
6 意大利 Italy 1025 6.12 5.24 26 意大利 Italy 749 6.39 18.04 50
7 日本 Japan 764 4.56 3.37 19 巴西 Brazil 640 5.46 10.05 33
8 法国 France 629 3.75 5.93 21 法国 France 529 4.51 20.60 47
9 土耳其 Turkey 610 3.64 2.54 17 加拿大 Canada 476 4.06 14.74 39
10 韩国 Korea 568 3.39 2.62 16 德国 Germany 429 3.66 19.68 43
对比两个 5 年,2005—2009 年,中国进入果树学
文献数量排名全球前 25 位的研究机构只有 4 家,分别
是中国科学院(排名第 10,188 篇),中国农业大学
(排名第 11,170 篇)、浙江大学(排名第 19,123篇)和华中农业大学(排名第 23,101 篇),而在 2010—2014 年,中国的研究机构达到了 7 家,分别是中国
农业大学(排名第 7,274 篇),中国科学院(排名第
8,242 篇),华中农业大学(排名第 9,221 篇),
西北农林科技大学(排名第 12,206 篇),华南农业
大学(排名第 16,160 篇),南京农业大学(排名第
19,154 篇)和浙江大学(排名第 23,142 篇)。 2.3 蔬菜学文献分析
以蔬菜(含西甜瓜)为题目,中国的文献记录数
排名虽然在两个 5 年内没有变化,均位居全球第 2 位,
但文献数所占比例有了大幅增加,从 2005—2009 年的
8.79%上升到了 2010—2014 年的 16.90%,接近两倍。
在文献数排名前 10 位的国家中,中国的文献篇均他引
次数排名保持稳定,位居第6位;而 h指数由2005—2009年的第 5 位上升到了 2010—2014 年的第 2 位(表 5)。
从蔬菜学文献记录数排名前 25 位的科研机构来
看,2005—2009 年间,中国有 3 个,分别是中国科学
院(排名第 14,181 篇),浙江大学(排名第 15,146篇)和中国农业大学(排名第 16,126 篇);2010—2014 年间,中国的科研机构增加到了 5 个,分别是中
国科学院(排名第 3,318 篇),中国农业大学(排名
第 8,260 篇),南京农业大学(排名第 11,237 篇),
浙江大学(排名第 12,225 篇)和华中农业大学(排
名第 25,131 篇)。与 2005—2009 年相比,2010—2014年中国的科研机构有了明显的增加,同时各科研机构
的排名也有了较明显的提升。
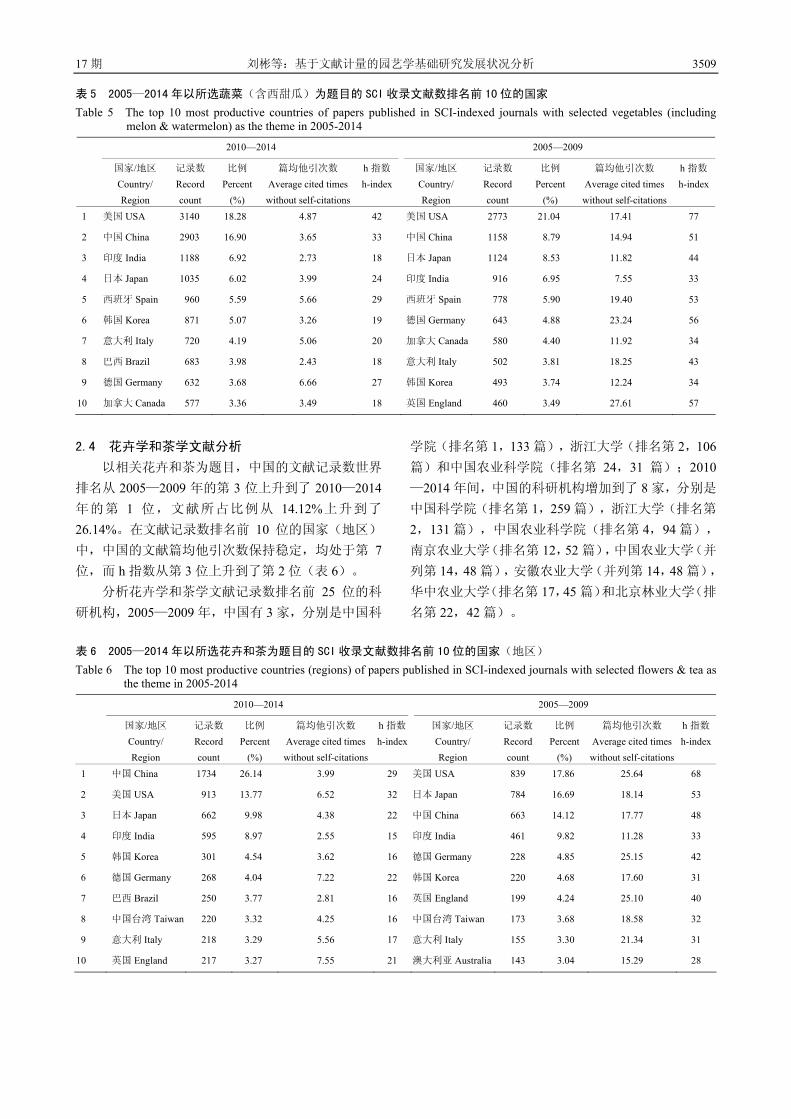
17 期 刘彬等:基于文献计量的园艺学基础研究发展状况分析 3509
表 5 2005—2014 年以所选蔬菜(含西甜瓜)为题目的 SCI 收录文献数排名前 10 位的国家
Table 5 The top 10 most productive countries of papers published in SCI-indexed journals with selected vegetables (including melon & watermelon) as the theme in 2005-2014
2010—2014 2005—2009
国家/地区 Country/ Region
记录数 Record count
比例 Percent
(%)
篇均他引次数 Average cited times without self-citations
h 指数 h-index
国家/地区 Country/ Region
记录数 Record count
比例 Percent
(%)
篇均他引次数 Average cited times without self-citations
h 指数
h-index
1 美国 USA 3140 18.28 4.87 42 美国 USA 2773 21.04 17.41 77
2 中国 China 2903 16.90 3.65 33 中国 China 1158 8.79 14.94 51
3 印度 India 1188 6.92 2.73 18 日本 Japan 1124 8.53 11.82 44
4 日本 Japan 1035 6.02 3.99 24 印度 India 916 6.95 7.55 33
5 西班牙 Spain 960 5.59 5.66 29 西班牙 Spain 778 5.90 19.40 53
6 韩国 Korea 871 5.07 3.26 19 德国 Germany 643 4.88 23.24 56
7 意大利 Italy 720 4.19 5.06 20 加拿大 Canada 580 4.40 11.92 34
8 巴西 Brazil 683 3.98 2.43 18 意大利 Italy 502 3.81 18.25 43
9 德国 Germany 632 3.68 6.66 27 韩国 Korea 493 3.74 12.24 34
10 加拿大 Canada 577 3.36 3.49 18 英国 England 460 3.49 27.61 57
2.4 花卉学和茶学文献分析
以相关花卉和茶为题目,中国的文献记录数世界
排名从 2005—2009 年的第 3 位上升到了 2010—2014年的第 1 位,文献所占比例从 14.12%上升到了
26.14%。在文献记录数排名前 10 位的国家(地区)
中,中国的文献篇均他引次数保持稳定,均处于第 7位,而 h 指数从第 3 位上升到了第 2 位(表 6)。
分析花卉学和茶学文献记录数排名前 25 位的科
研机构,2005—2009 年,中国有 3 家,分别是中国科
学院(排名第 1,133 篇),浙江大学(排名第 2,106篇)和中国农业科学院(排名第 24,31 篇);2010—2014 年间,中国的科研机构增加到了 8 家,分别是
中国科学院(排名第 1,259 篇),浙江大学(排名第
2,131 篇),中国农业科学院(排名第 4,94 篇),
南京农业大学(排名第 12,52 篇),中国农业大学(并
列第 14,48 篇),安徽农业大学(并列第 14,48 篇),
华中农业大学(排名第 17,45 篇)和北京林业大学(排
名第 22,42 篇)。
表 6 2005—2014 年以所选花卉和茶为题目的 SCI 收录文献数排名前 10 位的国家(地区)
Table 6 The top 10 most productive countries (regions) of papers published in SCI-indexed journals with selected flowers & tea as the theme in 2005-2014
2010—2014 2005—2009
国家/地区 Country/ Region
记录数 Record count
比例 Percent
(%)
篇均他引次数 Average cited times without self-citations
h 指数
h-index国家/地区 Country/ Region
记录数
Record count
比例 Percent
(%)
篇均他引次数 Average cited times without self-citations
h 指数
h-index
1 中国 China 1734 26.14 3.99 29 美国 USA 839 17.86 25.64 68
2 美国 USA 913 13.77 6.52 32 日本 Japan 784 16.69 18.14 53
3 日本 Japan 662 9.98 4.38 22 中国 China 663 14.12 17.77 48
4 印度 India 595 8.97 2.55 15 印度 India 461 9.82 11.28 33
5 韩国 Korea 301 4.54 3.62 16 德国 Germany 228 4.85 25.15 42
6 德国 Germany 268 4.04 7.22 22 韩国 Korea 220 4.68 17.60 31
7 巴西 Brazil 250 3.77 2.81 16 英国 England 199 4.24 25.10 40
8 中国台湾 Taiwan 220 3.32 4.25 16 中国台湾 Taiwan 173 3.68 18.58 32
9 意大利 Italy 218 3.29 5.56 17 意大利 Italy 155 3.30 21.34 31
10 英国 England 217 3.27 7.55 21 澳大利亚 Australia 143 3.04 15.29 28

3510 中 国 农 业 科 学 48 卷
2.5 生物学领域高影响因子杂志刊载文献分析 2005—2014 年被所选的高影响因子杂志收录的
以所选园艺植物为题目的文献共检索出 580 篇,其中
2005—2009 年 268 篇,2010—2014 年 312 篇,增幅
16.42%。来自中国的文献从 2005—2009 年的 23 篇增
加至 2010—2014 年的 42 篇,增幅 82.61%,远高于整
体增幅(表 7)。 文献数排名在前 10 位的国家及其文献记录数见
表 8。从 2005—2014 年、2005—2009 年及 2010—2014年 3 个窗口期来看,美国、德国、法国和英国的文献
记录数均稳居前 4 名。10 年间,中国的文献有 65 篇,
占总文献的 11.21%,排名位居第 5 位。将表 8 与表 3相比较,可以发现,虽然中国的文献总量在 2005—2009 年和 2010—2014 年两个 5 年窗口期内排名都比
较靠前,分别位居全球第 2 位和第 1 位,但在所选择
的生物学领域高影响因子杂志上所发表的文献数排
表 7 2005—2014 年以所选园艺植物为题目被生物学领域高影响因子杂志收录的 SCI 文献年度分布
Table 7 The annual distribution of papers published in high-impact biology journals with selected horticultural plants as the theme in 2005-2014
所有文献 Total 中国的文献 Paper of China 出版年 Publication year
记录数 Record count 比例 Percent (%) 记录数 Record count 比例 Percent (%)
2014 56 9.66 8 12.31
2013 72 12.41 14 21.54
2012 76 13.10 7 10.77
2011 57 9.83 9 13.85
2010 51 8.79 4 6.15
2009 63 10.86 9 13.85
2008 56 9.66 6 9.23
2007 54 9.31 2 3.08
2006 41 7.07 4 6.15
2005 54 9.31 2 3.08
2010-2014 312 53.79 42 64.62
2005-2009 268 46.21 23 35.39
表 8 2005—2014 年以所选园艺植物为题目被生物学领域高影响因子杂志收录的 SCI 文献数排名前 10 位的国家
Table 8 The top 10 most productive countries of papers published in high-impact biology journals with selected horticultural plants as the theme in 2005-2014
2005—2014 2005—2009 2010—2014
国家/地区 Country/Region
记录数 Record count
比例 Percent (%)
国家/地区 Country/Region
记录数 Record count
比例 Percent (%)
国家/地区 Country/Region
记录数 Record count
比例 Percent (%)
1 美国 USA 201 34.66 美国 USA 77 28.73 美国 USA 124 39.74
2 德国 Germany 140 24.14 德国 Germany 65 24.25 德国 Germany 75 24.04
3 法国 France 82 14.14 法国 France 33 12.31 法国 France 49 15.71
4 英国 England 75 12.93 英国 England 30 11.19 英国 England 45 14.43
5 中国 China 65 11.21 日本 Japan 30 11.19 中国 China 42 13.46
6 日本 Japan 60 10.35 荷兰 Netherlands 26 9.70 西班牙 Spain 39 12.50
7 西班牙 Spain 57 9.83 意大利 Italy 24 8.96 意大利 Italy 31 9.94
8 意大利 Italy 55 9.48 中国 China 23 8.58 日本 Japan 30 9.62
9 荷兰 Netherlands 54 9.31 西班牙 Spain 18 6.72 荷兰 Netherlands 28 8.97
10 以色列 Israel 41 7.07 澳大利亚 Australia 17 6.34 以色列 Israel 24 7.69
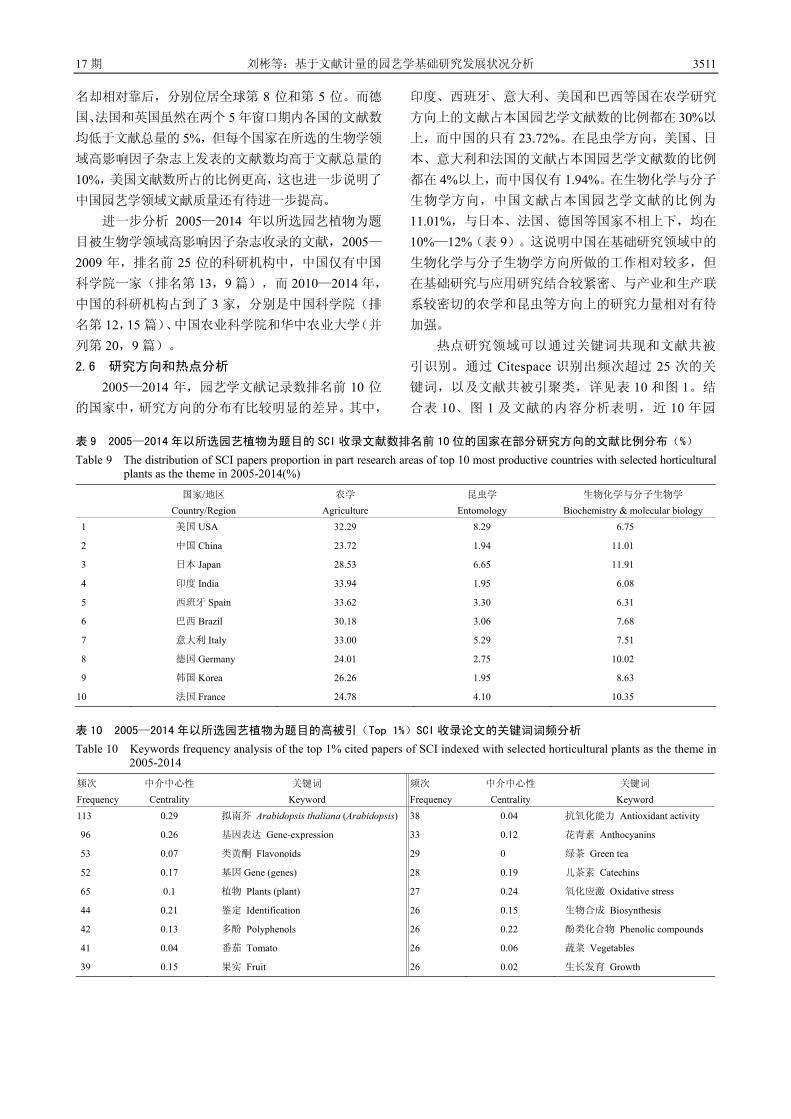
17 期 刘彬等:基于文献计量的园艺学基础研究发展状况分析 3511
名却相对靠后,分别位居全球第 8 位和第 5 位。而德
国、法国和英国虽然在两个 5 年窗口期内各国的文献数
均低于文献总量的 5%,但每个国家在所选的生物学领
域高影响因子杂志上发表的文献数均高于文献总量的
10%,美国文献数所占的比例更高,这也进一步说明了
中国园艺学领域文献质量还有待进一步提高。 进一步分析 2005—2014 年以所选园艺植物为题
目被生物学领域高影响因子杂志收录的文献,2005—2009 年,排名前 25 位的科研机构中,中国仅有中国
科学院一家(排名第 13,9 篇),而 2010—2014 年,
中国的科研机构占到了 3 家,分别是中国科学院(排
名第 12,15 篇)、中国农业科学院和华中农业大学(并
列第 20,9 篇)。 2.6 研究方向和热点分析
2005—2014 年,园艺学文献记录数排名前 10 位
的国家中,研究方向的分布有比较明显的差异。其中,
印度、西班牙、意大利、美国和巴西等国在农学研究
方向上的文献占本国园艺学文献数的比例都在 30%以
上,而中国的只有 23.72%。在昆虫学方向,美国、日
本、意大利和法国的文献占本国园艺学文献数的比例
都在 4%以上,而中国仅有 1.94%。在生物化学与分子
生物学方向,中国文献占本国园艺学文献的比例为
11.01%,与日本、法国、德国等国家不相上下,均在
10%—12%(表 9)。这说明中国在基础研究领域中的
生物化学与分子生物学方向所做的工作相对较多,但
在基础研究与应用研究结合较紧密、与产业和生产联
系较密切的农学和昆虫等方向上的研究力量相对有待
加强。 热点研究领域可以通过关键词共现和文献共被
引识别。通过 Citespace 识别出频次超过 25 次的关
键词,以及文献共被引聚类,详见表 10 和图 1。结
合表 10、图 1 及文献的内容分析表明,近 10 年园
表 9 2005—2014 年以所选园艺植物为题目的 SCI 收录文献数排名前 10 位的国家在部分研究方向的文献比例分布(%)
Table 9 The distribution of SCI papers proportion in part research areas of top 10 most productive countries with selected horticultural plants as the theme in 2005-2014(%)
国家/地区 Country/Region
农学 Agriculture
昆虫学 Entomology
生物化学与分子生物学 Biochemistry & molecular biology
1 美国 USA 32.29 8.29 6.75
2 中国 China 23.72 1.94 11.01
3 日本 Japan 28.53 6.65 11.91
4 印度 India 33.94 1.95 6.08
5 西班牙 Spain 33.62 3.30 6.31
6 巴西 Brazil 30.18 3.06 7.68
7 意大利 Italy 33.00 5.29 7.51
8 德国 Germany 24.01 2.75 10.02
9 韩国 Korea 26.26 1.95 8.63
10 法国 France 24.78 4.10 10.35
表 10 2005—2014 年以所选园艺植物为题目的高被引(Top 1%)SCI 收录论文的关键词词频分析
Table 10 Keywords frequency analysis of the top 1% cited papers of SCI indexed with selected horticultural plants as the theme in 2005-2014
频次 Frequency
中介中心性 Centrality
关键词 Keyword
频次 Frequency
中介中心性 Centrality
关键词 Keyword
113 0.29 拟南芥 Arabidopsis thaliana (Arabidopsis) 38 0.04 抗氧化能力 Antioxidant activity
96 0.26 基因表达 Gene-expression 33 0.12 花青素 Anthocyanins
53 0.07 类黄酮 Flavonoids 29 0 绿茶 Green tea
52 0.17 基因 Gene (genes) 28 0.19 儿茶素 Catechins
65 0.1 植物 Plants (plant) 27 0.24 氧化应激 Oxidative stress
44 0.21 鉴定 Identification 26 0.15 生物合成 Biosynthesis
42 0.13 多酚 Polyphenols 26 0.22 酚类化合物 Phenolic compounds
41 0.04 番茄 Tomato 26 0.06 蔬菜 Vegetables
39 0.15 果实 Fruit 26 0.02 生长发育 Growth

3512 中 国 农 业 科 学 48 卷
2005 2007 2009 2011 2013
酚类化合物Phenolic compounds
果实发育Fruit development
高质量基因组草图High quality genome draft
Cite Space, v. 3.8.R1 (32-bit)April 24, 2015 9:02:25 AM CSTF:/hotrtic/dataTimespan: 2005-2014 (Slice Length=2)Selection Criteria: Top 50 per sliceNetwork: N=220, E=503 (Density=0.0209)Pruning: PathfinderModul arity Q=0.808Mean Silhouette=0.5355
图 1 2005—2014 年以所选园艺植物为题目的高被引(Top 1%)SCI 收录论文的文献共被引图谱
Fig. 1 Mapping knowledge domains of the top 1% cited papers of SCI indexed with selected horticultural plants as the theme in
2005-2014
艺学研究热点领域主要集中在功能基因挖掘与组学
研究、果实发育与成熟研究、多酚类物质的研究等
方向。近年来,中国园艺学领域一系列基于基因组
测序和大数据分析的研究论文发表在较好的学术期
刊上,如以马铃薯、黄瓜、甜橙等为代表的文献发
表在 Nature 系列等期刊上。
3 讨论
3.1 更加重视科研成果的质量 近年来,中国的园艺学基础研究进入了快速发展
期,在中国建设创新型国家的大环境下,国家对科学
研究的投入越来越多,科研工作者们也开展了卓有成
效的工作,在国际上的活跃度逐渐增加,SCI 收录文
献的数量也逐年增加。近 10 年来,中国以常见的 60余种园艺植物为题目的 SCI 收录论文产出量已经距世
界第二,仅次于美国,特别是 2010—2014 年文献数量
跃居世界第一,但质量提升速度还未赶上数量的增长
速度,主要表现在高影响因子的“大文章”较少,小
文章或人们称为“豆腐块”的文章较多,论文的篇均
他引次数和 h 指数偏低,生物学领域高影响因子杂志
上所发表的文献数排名相对靠后。相对于论文的数
量,今后更应关注论文的质量。近年来,国内甚至出
现了学风问题,学术不端行为时有发生,国家自然科
学基金委曾两次举办新闻发布会,对国家自然科学基
金申报过程中的学术不端行为进行通报,应引起重视。 3.2 更加注重科研与生产实际的紧密结合
结合 2014 年 8 月在澳大利亚召开的第 29 届国际
园艺大会来看,国内无论是在研究方向的凝练、研究
的深度,还是在组建研究团队开展联合攻关方面,都
和发达国家有着不小的差距。发达国家的研究积累厚
重感强,理论与实践联系紧密。部分国家能围绕产业
问题全方位开展研究,例如美国、澳大利亚等国已将
分子技术成功应用于园艺植物育种和产业实践,开发
了标记辅助育种(marker-assisted breeding,MAB)。
从论文的学科分布来看,中国在与生产结合较紧密的
农艺和植保学科,与发达国家相比相对较弱,而在生
物化学与分子生物学等偏基础研究领域所做的研究相
对较多,但能应用于实践的还缺乏典型案例。研究缺

17 期 刘彬等:基于文献计量的园艺学基础研究发展状况分析 3513
乏创新性,研究方向分散、个别科研工作者不能坚持
有所为有所不为,喜欢“打一枪换一个地方”,研究
不成体系,以开展“解决科学问题为导向”的研究还
远远不够,而以项目(课题)为导向的研究和跟踪热 点的“跟风研究”相对较多。
目前,国际上高被引论文主要集中在组学和分子
生物学等研究领域。近年,高水平研究论文不少是在
做测序,体现了两个多:一是所花费的科研经费多,
二是论文的作者多。在中国目前财力较好、资助渠道
较多的时候,需更加注重全方位思考园艺产业的问题,
将研究力量分布得更加科学、合理,并做出特色和深
度。在利用现有的平台继续深入挖掘优异基因的同
时,产生的研究成果应不断向应用研究延伸,以指导
实践。从园艺学的国际研究趋势来看,今后应注重加
强栽培、病虫害防控等方面的研究;加强省力栽培技
术、机械化与农机农艺结合研究;积极拓展园艺研究
领域,扩展园艺产品的功能性成分分析和园艺治疗
(Horticultural Therapy)方面的研究。
4 结论
通过文献计量学的方法得到,近 5 年,中国园艺
学基础研究在论文产出数量上已居世界第一位,拥有
与园艺大国相称的地位,但质量提升还未跟上。在研
究方向上,中国园艺学文献侧重于分子水平和组学的
研究,与农业结合相对紧密的文献比例偏少。未来应
加强问题导向的研究,布局上应更加贴近产业各环节
的问题,并持之以恒的开展研究,形成特色,为经济
社会的发展和国家的重大战略需求提供长期的支撑。
References
[1] 中国科学技术协会. 园艺学学科发展报吿. 北京: 中国科学技术出
版社, 2012.
China Association for Science and Technology. Horticulture
Discipline Development Report. Beijing: China Science & Technology
Press, 2012. (in Chinese)
[2] 管曦 . 中国茶产业链纵向整合研究[D]. 南京 : 南京农业大学 ,
2012.
Guan X. Research on the vertical integration of Chinese tea industry
chain[D]. Nanjing: Nanjing Agricultural University, 2012. (in
Chinese)
[3] 高懋芳, 邱建军, 刘三超, 刘宏斌, 王立刚, 逄焕成. 基于文献计
量的农业面源污染研究发展态势分析. 中国农业科学, 2014, 47(6):
1140-1150.
Gao M F, Qiu J J, Liu S C, Liu H B, Wang L G, Pang H C. Status and
trends of agricultural diffuse pollution research based on bibliometrics.
Scientia Agricultura Sinica, 2014, 47(6): 1140-1150. (in Chinese)
[4] 黄宝晟. 文献计量法在基础研究评价中的问题分析. 科研与发展
管理, 2008, 20(6): 108-111.
Huang B S. Analysis on the application of literature metrology to
basic research evaluation. R&D Management, 2008, 20(6): 108-111.
(in Chinese)
[5] 陈静, 张保卫, 马克平, 蒋志刚. 中国保护生物学研究现状的文献
计量学分析. 生物多样性, 2009, 17(4): 423-429.
Chen J, Zhang B W, Ma K P, Jiang Z G. Bibliometric analysis of
status quo of conservation biology in China. Biodiversity Science,
2009, 17(4): 423-429. (in Chinese)
[6] 邬亚文, 夏小东, 职桂叶, 葛磊, 何建妹, 鄂志国, 李建, 王磊. 基
于文献的国内外水稻研究发展态势分析. 中国农业科学, 2011,
44(20): 4129-4141.
Wu Y W, Xia X D, Zhi G Y, Ge L, He J M, E Z G, Li J, Wang L.
Status and trends of rice science based on bibliometrics. Scientia
Agricultura Sinica, 2011, 44(20): 4129-4141. (in Chinese)
[7] Josef D V, Sanja B. Tree fruit growing—research and production in
Germany: a statistical and bibliometric analysis of the period
1950-2010. Erwerbs-Obstbau, 2012, 54(1): 11-30.
[8] Tatry M V, Dominique F, Benoıˆt J, Francoise D. EU27 and USA
leadership in fruit and vegetable research: a bibliometric study from
2000 to 2009. Scientometrics, 2014, 98(3): 2207-2222.
[9] Leiser H, Aventurier P, Fournier D, Dosba F, Jeannequin, B. Tools for
producing indicators from a bibliometric study of scientific production:
The case of fruit and vegetable publications by the French National
Institute for Agricultural Research (INRA). Fruits, 2009, 64(5):
305-312.
[10] Alhaider I, Ahmed M K K, Gupta B M. Global research output on
date palm (Pheonix dactylifera): a 12 years scientometric perspective.
Scientometrics, 2014, 98(1): 157-171.
[11] Glanzel W, Veugelers R. Science for wine: A bibliometric assessment
of wine and grape research for wine-producing and consuming
countries. American journal of enology and viticulture, 2006, 57(1):
23-32.
[12] 冯立娟, 尹燕雷, 招雪晴, 杨雪梅, 李英朋. 基于文献计量的世界
樱桃研究态势分析. 中国农业科技导报, 2014, 16(5): 175-181.
Feng L J, Yin Y L, Zhao X Q, Yang X M, Li Y P. Analysis of global
cherry research trend based on bibliometrics. Journal of Agricultural
Science and Technology, 2014, 16(5): 175-181. (in Chinese)
[13] Liu Y X, Rao I K R, Rousseau R. Empirical series of journal h-indices:

3514 中 国 农 业 科 学 48 卷
The JCR category Horticulture as a case study. Scientometrics, 2009,
80(1): 59-74.
[14] 张娟, 王宁, 张以民, 李云霞. 基于 Web of Science 的国际柑橘黄
龙病文献计量分析. 果树学报, 2014, 31(6): 1139-1146.
Zhang J, Wang N, Zhang Y M, Li Y X. Bibliometric analysis of the
research on citrus Huanblongbing based on web of science. Journal of
Fruit Science, 2014, 31(6): 1139- 1146. (in Chinese)
[15] 崔晓艳, 顾和平, 袁星星, 张红梅, 陈华涛, 陈新. 马铃薯 Y 病毒
属(Potyvirus)研究的文献计量分析. 微生物学通报, 2013, 40(5):
891-903.
Cui X Y, Gu H P, Yuan X X, Zhang H M, Chen H T, Chen X.
Research dynamics on Potyvirus based on bibliometric. Microbiology
China , 2013, 40(5): 891-903. (in Chinese)
[16] 晏志梅, 张春玲, 刘娟, 卢英琳, 陈全斌. 文献计量分析我国番木
瓜的研究现状与发展趋势. 广东农业科学, 2008, 26(2): 112-114.
Yan Z M, Zhang C L, Liu J, Lu Y L, Chen Q B. Bibliometric
analysis of research status and development trend of papaya in China.
Guangdong Agricultural Sciences, 2008, 26(2): 112-114. (in Chinese)
[17] 马秀文, 和艳会. 采用文献计量法分析我国蝴蝶兰研究现状与发
展趋势. 安徽农业科学, 2011, 39(6):3761-3762, 3765.
Ma X W, He Y H. Bibliometric analysis on research situation and
development trends of phalaenopsis. Journal of Anhui Agricultural
Sciences, 2011, 39(6):3761-3762, 3765. (in Chinese)
[18] 李金格, 王艳, 赵立强, 陈全斌. 我国菊花研究状况和发展趋势文
献计量分析. 现代农业科技, 2008, (2): 16-17, 20.
Li J G, Wang Y, Zhao L Q, Chen Q B. Bibliometric analysis research
status and development trend of chrysanthemum in China.Modern
Agricultural Science & Technology, 2008, (2): 16-17, 20. (in Chinese)
[19] 孙会军. 我国高校园艺学科SCI论文产出力与影响力比较研究. 北
方园艺, 2012(20): 194-197.
Sun H J. Comparative study on papers productivity and influence
based on SCI database of Chinese universities’ horticultural science.
Northern Horticulture, 2012(20): 194-197. (in Chinese)
[20] 孙会军. 中国与国际园艺学科SCI论文数量与质量的比较. 北方园
艺, 2013(16): 221-224.
Sun H J. Comparison of China and international SCI papers’ quantity
and quality in horticultural science. Northern Horticulture, 2013(16):
221-224. (in Chinese)
[21] 刘彬, 陈柳, 袁佩佩. 国家自然科学基金资助的 2009-2013 年 SCI
收录文献计量分析. 中国科学基金, 2014, 28(3): 214-218.
Liu B, Chen L, Yuan P P. Bibliometric analysis on SCI covered
papers funded by NSFC from 2009 to 2013. Bulletin of National
Natural Science Foundation of China, 2014, 28(3): 214-218. (in
Chinese)
[22] Hirsch J E. An index to quantify an individual’s scientific research
output. Proceedings of the National Academy of Sciences of the
United States of America, 2005, 102(46): 16569-16572.
[23] Van Haselen R. The h-index: a new way of assessing the scientific
impact of individual CAM authors. Complementary Therapies in
Medicine, 2007, 15(4): 225-227.
[24] Bornmann L, Daniel H D. Does the h-index for ranking of scientists
really work? Scientometrics, 2005, 65(3): 391-392.
[25] Ruane F, Tol R S J. Rational (successive) h-indices: An application to
economics in the Republic of Ireland. Scientometrics, 2008, 75(2):
395-405.
[26] Vanclay J K. Ranking forestry journals using the h-index. Journal of
Informetrics, 2008, 2(4): 326-334.
[27] 杨俊丽. 基于SCI的河南农业大学科技论文统计分析. 科技管理研
究, 2012, 32(11): 132-135.
Yang J L. The statistical analysis based on SCI papers of Henan
Agriculture University. Science & Technology Management Research,
2012, 32(11): 132-135. (in Chinese)
[28] 周辉, 张光红, 蔡晖, 郑英姿. 原创性研究成果的 SCI 引用分析.
中国科学基金, 2002, 16(2): 23-25.
Zhou H, Zhang G H, Cai H, Zheng Y Z. The SCI citation analysis of
the original innovation. Bulletin of National Natural Science
Foundation of China, 2002, 16(2): 23-25. (in Chinese)
(责任编辑 赵伶俐)

中国农业科学 2015,48(17):3515-3523 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.017
收稿日期:2015-01-20;接受日期:2015-05-20 基金项目:国家公益性行业(农业)科研专项(201203095,201203001)、国家大宗蔬菜产业技术体系(CARS-25-C-09) 联系方式:蒋卫杰,E-mail:[email protected]
设施园艺发展概况、存在问题与产业发展建议
蒋卫杰,邓 杰,余宏军
(中国农业科学院蔬菜花卉研究所/农业部园艺作物生物学与种质创制重点实验室,北京 100081)
摘要:近年来中国设施园艺产业发展迅速,在农产品供应、农民增收和农村经济繁荣方面发挥了重要作用,
在作物栽培管理技术、环境控制技术和温室工程技术及配套装备等方面也取得较迅速的发展。文章概述了国内外
设施园艺的发展概况与趋势,从设施装备、科技含量、连作障碍等方面分析了目前中国设施园艺存在的主要问题。
根据实际情况对中国设施园艺的持续发展提出了相关的重点研究方向:要加强温室结构装备和环境精确控制技术、
无土栽培技术、病虫害可持续控制技术、连作障碍克服技术、设施园艺专业品种培育技术等的研究。并从加强政
策宏观调控,加大科技创新投入力度,加强设施装备标准化发展,完善园艺产品供求预警及构建质量安全控制长
效机制等方面提出了产业化发展措施与建议。
关键词:设施园艺;日光温室;大棚;无土栽培;发展概况;产业特点
Development Situation, Problems and Suggestions on Industrial
Development of Protected Horticulture JIANG Wei-jie, DENG Jie, YU Hong-jun
(Institute of Vegetables & Flowers, Chinese Academy of Agricultural Sciences/Key Laboratory of Biology and Genetic Improvement
of Horticultural Crops, Ministry of Agriculture, Beijing 100081)
Abstract: Protected agriculture has developed rapidly in China and plays important roles in agricultural products supplying, farmers income and countryside economic prosperity. Meanwhile, high techniques have also been applied in protected horticulture, including crop management, environment control and greenhouse construction. This paper reviewed the update development characteristics of protected horticulture in China, summarized the world latest development trends and proposes. The main problems in the existing facilities in China were analyzed from the aspects of facility equipment, scientific and technological content and continuous cropping obstacles. It was pointed out that the structure of greenhouse equipment and environment precision control technology, soilless cultivation techniques, pest damage sustainable control technology, the technology for overcoming the obstacles of continuous cropping, and the protected horticulture varieties cultivation technology should be strengthened in research in the future. The suggestions in research fields and industry policy for sustainable development of protected horticulture industry in China include strengthening the macro-control policies, increasing investment in scientific and technological innovation, strengthening the standardization of facilities and equipment, improving the supply and demand of horticultural products and building a long-term effective mechanism for the construction of quality and safety control.
Key words: protected horticulture; solar greenhouse; plastic tunnel; soilless culture; development survey; industrial characteristics 设施园艺是通过现代农业工程、机械技术和管
理技术改善局部环境,以生产优质、高产、稳产农
产品的一种环控农业,是作物栽培技术、环境控制
技术与温室工程技术高度交叉融合的产业[1]。设施
jia
文本框
返回目录

3516 中 国 农 业 科 学 48 卷
园艺科技含量高,是一个国家或地区农业现代化水
平的重要标志之一,发展设施园艺是由传统农业向
现代化农业转变的重要组成部分[2]。目前,许多国
家发展了大规模的现代设施园艺,并成为其支柱产
业,从设施栽培专用品种、栽培方式、温室配套控
制技术及设备等方面推出标准化成套模式。欧美及
日本等发达国家在发展设施园艺过程中,不仅能高
水平地进行设施建设和能源利用,并且配套有先进
的栽培、管理技术,获得极高的经济效益[3]。目前,
荷兰已建成 1.1 万 hm2 玻璃温室,主要以文洛
(Venlo)型连栋温室为主,其中约 50%用于种植月
季、菊花、百合、小苍兰及兰花等观赏植物,每年
出口总额超过 60 亿美元,其余玻璃温室则主要用于
番茄、黄瓜和辣椒等蔬菜生产,其中番茄产量可达
800 t·hm-2、黄瓜产量可达 1 000 t·hm-2,达到了极高
的产量和效益水平。以色列设施园艺面积约 2.6 万
hm2,温室工程、新品种培育、栽培管理和节水灌溉
等方面研究和应用都处于世界领先水平,番茄产量
高达 480 t·hm-2,而温室种植玫瑰花 高产量达 320万株/hm2[4]。日本与韩国设施园艺也较为发达,在
设施环境调控、设施专用品种选育和无土栽培技术
等方面都处于亚洲领先地位。 中国发展设施园艺可有效提高土地产出率、资源
利用率和社会生产率,对保证农产品供给,促进农业
发展、农民增收,增强中国农业综合生产力都具有十
分重要的意义。
1 中国设施园艺发展现状
随着中国农业产业结构调整的不断深入,设施
园艺得到了迅猛发展,在农业中所占比重不断提
高,栽培面积从 1978 年不足 0.7 万 hm2 发展到 2013
年的 187.4 万 hm2,成为世界设施园艺第一生产大
国,栽培作物已从单一的蔬菜扩展到瓜果、食用菌、
中草药、花卉等多种经济作物领域。栽培地域也不
断扩大,目前中国所有省(区、市)都有设施园艺
生产,不同地区设施形态也各具特点。北方主要发
展高效节能日光温室[5],南方则以塑料大棚及简易
设施栽培为主。根据农业部农机化司统计[6-11],2008—2013 年,中国设施园艺(包括连栋温室、日光
温室、塑料大棚)栽培总面积从 81.3 万 hm2 增加
至 187.4 万 hm2(表 1),平均年增长率为 18.5%。
不同地区之间具有各自的区域特点,从设施类型上
看(截止 2013 年),设施园艺栽培面积中连栋温
室面积为 5.7 万 hm2,日光温室面积为 61.2 万 hm2,
塑料温室面积为 120.5 万 hm2(表 2),分别占设
施园艺栽培总面积的 3%、32.7%、64.3%。从产地
分布看,辽宁、山东、河北、江苏是中国设施园艺
发展 快的 4 个省,约占全国设施面积的 58.8%。
这些地区利用充足的光照资源发展节能日光温室,
实现越冬及早春果菜生产。长江中下游地区主要发
展塑料大棚等设施,实现冬季果菜、根菜、叶菜的
生产。
2 国外设施园艺主要进展
2.1 国外设施园艺概况
目前,设施园艺技术 发达的国家当属荷兰、以
色列、日本、美国和加拿大。根据相关资料统计,截
止 2010 年底,国外设施园艺总面积约为 56 万 hm2[4],
主要分布在日本、韩国及地中海各国(表 3)。荷兰
及北欧国家以玻璃温室为主,塑料大棚在日本、韩国
使用较为广泛。中国现代化设施园艺起步较晚,但发
展迅速,已成为世界第一生产大国[12]。
表 1 2008-2013 年全国不同结构类型设施园艺面积[6-11]
Table 1 Area of different protected horticulture types from 2008 to 2013 in China (hm2) [6-11]
年份 Year
连栋温室面积 Multi-span greenhouse area
日光温室面积 Solar greenhouse area
塑料大棚面积 Plastic tunnel area
设施园艺总面积 Total area
2008 17875.01 254131.79 541110.16 813116.96
2009 18156.66 241287.27 619300.3 878744.23
2010 19835.59 378928.94 701693.24 1100457.77
2011 20436.26 409636.05 801742.14 1231814.45
2012 26250.53 461227.72 918034.7 1405512.95
2013 56940.63 612338.97 1204907.19 1874236.79
17 期 蒋卫杰等:设施园艺发展概况、存在问题与产业发展建议 3517
表 2 2013 年中国设施园艺面积分布[11]
Table 2 Area of protected horticulture in China (hm2) [11]
省份 Province
连栋温室 Multi-span greenhouse
日光温室 Solar
greenhouse
塑料大棚
Plastic tunnel
温室总 面积
Total area
省份 Province
连栋温室
Multi-span greenhouse
日光温室 Solar
greenhouse
塑料大棚
Plastic tunnel温室总面积
Total area
北京 Beijing 715.27 8884.05 9597.99 19197.31 湖北 Hubei 4989.74 656.28 60331.68 65977.7
天津 Tianjing 1383.41 17808.47 15306.34 34498.22 湖南 Hunan 67.75 32.68 6301.71 6402.14
河北 Hebei 390.69 77251.35 127932.1 205574.18 广东 Guangdong 2041.59 90.95 8693.21 10825.75
山西 Shanxi 364.48 30240.65 26237.39 56842.52 广西 Guangxi 1.25 5.03 321.66 327.94
内蒙古 Inner Mongolia 2152.53 30989.22 21009.75 54151.50 海南 Hainan 1.06 15 2151.32 2167.38
辽宁 Liaoning 22995.29 228017.20 173013.4 424025.84 重庆 Chongqing 9.38 0.2 40838.63 40848.21
吉林 Jilin 194 5135.35 17657.01 22986.36 四川 Sichuan 741.57 2708.39 85544.37 88994.33
黑龙江 Heilongjiang 1591.85 851.13 21022.35 23465.33 贵州 Guizhou 10.23 2.98 409.65 422.86
上海 Shanghai 193.41 0 5512.59 5706 云南 Yunnan 276.26 240.61 23985.32 24502.19
江苏 Jiangsu 11509.16 18026.74 173397.6 202933.54 西藏 Xizang 0 207.38 731.09 938.47
浙江 Zhejiang 2332.65 38.56 41295.95 43667.16 陕西 Shaanxi 92.94 15653.29 46798.47 62544.70
安徽 Anhui 97 1915 16531 18543 甘肃 Gansu 43 21592.85 26637.95 48273.80
福建 Fujian 69.04 0.16 6303.21 6372.41 青海 Qinghai 101.79 1367.72 1548.29 3017.80
江西 Jiangxi 12.47 22.72 3116.15 3151.34 宁夏 Ningxia 33.81 10743.91 7984.67 18762.39
山东 Shandong 4213.67 96714.33 167297.6 268225.57 新疆 Xinjiang 81.97 19736.15 14873.68 34691.80
河南 Henan 221.73 21421.21 50548.42 72191.36 新疆兵团 Xinjiang production and Construction Corps
11.64 2019.43 1976.62 4007.69
全国 Total 56940.63 612389 1204907 1874236.8
表 3 国外设施园艺栽培面积[4]
Table 3 Aera of protected horticulture in foreign countries[4]
国家 Country 面积 Area (hm2) 国家 Country 面积 Area (hm2) 国家 Country 面积 Area (hm2)
意大利 Italy 72800 荷兰 Netherlands 10800 德国 Germany 3300
西班牙 Spain 71698 南斯拉夫 Yugoslavia 10185 葡萄牙 Portugal 2700
韩国 Korea 57440 突尼斯 Tunisia 8895 厄瓜多尔 Ecuador 2500
士耳其 Turkey 49746 罗马尼亚 Romania 7300 英国 England 2419
日本 Japan 49049 波兰 Poland 6500 加拿大 Canada 2300
埃及 Egypt 32800 阿尔及利亚 Algeria 6200 黑山 Montenegro 2250
美国 USA 27000 俄罗斯 Russia 6200 智利 Chile 1500
法国 France 26500 匈牙利 Hungary 5185 克罗地亚 Croatia 1410
以色列 Israel 26000 利比亚 Libya 5000 澳大利亚 Australia 1300
巴西 Brazil 17000 叙利亚 Syria 4422 塞浦路斯 Cyprus 733
希腊 Greece 14981 黎巴嫩 Lebanon 4000 总计 Total 555611
墨西哥 Mexico 11759 约旦 Jordan 3739
2.2 国外设施园艺发展的特点
2.2.1 先进工业技术广泛应用于设施园艺 设施园
艺要求高投入、高产出,当前先进的工业技术已不断
投入到设施园艺中并取得了显著的成效。国外发达国
家已将自动化技术应用到园艺作物的育苗、耕作、施
肥、病虫害防治及农产品的收获、分装、储藏、保鲜

3518 中 国 农 业 科 学 48 卷
等过程,节约了大量劳动力[13]。目前,荷兰、日本、
美国、以色列、韩国开发出的耕作、移栽、嫁接、施
肥、喷药、采摘、移栽、消毒等配套机器人装备,应
用于设施园艺生产过程中,不仅大幅节约了劳动力、
降低了工人劳动强度、提高了劳动效率、改善了设
施生产劳动环境,而且保证了生产过程中产品的均
一性[14]。荷兰开发的温室屋面自动清洗装置,用于清
洗温室玻璃灰尘,大幅度提高了温室的透光率[15]。在
园艺产品采收及后加工过程中,发达国家也广泛使用
自动化机器,在包装、产品转运、产品快速分级等过
程基本实现了机械化、自动化。 2.2.2 高新技术在设施园艺中大量应用 设施园艺
是多种高新技术集成的平台,包括计算机控制、物联
网、无土栽培、生物技术、物理化学技术、新材料及
新能源利用等,是多个学科交叉的集成体。现代化计
算机控制技术和传感器技术已使设施园艺向工厂化、
数字化、智能化发展,基于数字化的模型能够对温室
进行准确的管理。从育苗、定植、栽培、施肥、灌溉
等过程,发达国家设施园艺已基本实现智能化操作管
理,可根据作物生长特点自动调节温室内温度、湿度、
CO2 浓度等,为植物创造 适宜的生长环境。欧美一
些现代化温室采用物联网技术、逐步实现管理网络化。
日本、韩国在设施环境调控技术方面处于亚洲领先地
位[15-17]。节能减排也是欧美国家研究的重点领域,主
要包括提高温室覆盖材料的透光率、优化温室结构、
减少热能损失、热能多用途利用,大大降低了单位产
量的能耗。LED 作为补光光源也是目前国外节能领域
研究的热点之一,与传统钠灯相比,LED 光源节能效
率可达 50%以上,并且光源的使用寿命大幅延长[18-19]。 2.2.3 无土栽培技术的广泛应用 无土栽培以人工
制造的作物根系环境取代了土壤环境。可有效解决传
统土壤栽培中难以解决的水分、空气、养分的供应矛
盾,作物根系处于 适宜的环境条件,从而充分发挥
作物的增产潜力。目前无土栽培 发达的国家是荷兰,
所有温室蔬菜生产都采用无土栽培技术。商业性无土
栽培以基质培为主,荷兰基质栽培占无土栽培面积的
90%以上。无土栽培改变了传统的种植模式,可以避
免设施栽培出现土壤连作障碍,作物的产量大幅提高,
产品品质也有显著改善。 2.2.4 选育出大量设施园艺专用品种 品种是现代
农业的核心竞争力,现今设施园艺发达国家都非常注
重温室专用品种的培育,根据不同地区,有针对性地
进行设施园艺专用品种的研发。在品种的选育过程中,
细胞组织培养、体细胞杂交、原生质体融合、遗传标
记、转基因等技术也越来越多地应用于育种中,在茄
子、番茄、甜椒、黄瓜及叶菜类作物上培育出一大批
优良品种,欧美国家凭借其创新能力,使其设施园艺
作物种子在世界上处于垄断地位。 2.2.5 环境保护应用于设施园艺 设施园艺高产出
是以投入高,耗能高为前提,大量农药、化肥的使用
会导致环境恶化,发达国家已把环境保护技术和产品
应用于设施园艺。如通过释放害虫天敌已取得很好的
防治效果,蚜虫、白粉虱及斑潜蝇等主要害虫天敌已
商业化,可以通过“以虫治虫”的方式,来防治害虫,
大大降低了农药的用量。岩棉是一种比较好的无土栽
培基质,在欧美国家有大量应用,但由于岩棉在自然
界不易降解,大量使用过的旧岩棉会造成严重的环境
污染。加拿大、荷兰等国家已经研制出可以替代岩棉
的易降解的新型基质[20-21],以减轻对环境污染的压力。
荷兰等国采用熊蜂给番茄授粉,取代传统的震荡授粉
及喷施化学激素,既提高了番茄的产量和品质,又降
低了劳动力成本。
3 中国设施园艺发展特点
中国设施园艺经过 30 多年的发展,形成了不同地
域特色的设施类型。设施园艺生产按照优质、高产、
高效、生态、安全的要求,得到快速发展,已成为世
界设施面积 大的国家。在设施园艺规模不断扩大的
同时,国家和各级政府对设施园艺科技也极为重视。
针对中国设施园艺装备落后、设施自动化和智能化程
度低、能源短缺、设施专用品种缺乏等问题,开展多
学科联合攻关,均取得了一定的成果。 3.1 设施园艺增效明显
2008 年全国设施园艺总产值为 7 079.8 亿元,占
园艺产业总产值的 51.3%,利用 22%的播种面积,创
造了 36.8%的产量,63.1%的产值。2010 年设施园艺
总产值约 7 400 亿元。目前,全国设施园艺年产值已
超过 1 万亿元,部分省份设施园艺收入已超过当地农
民纯收入一半以上。与露地种植相比,设施园艺通过
对生产环境调控从而大幅度提高单产效益,产值则比
露地生产提高 3—5 倍[22]。 3.2 高新技术逐步应用于设施园艺
3.2.1 温室设施装备水平和环境控制技术不断提高
随着中国设施园艺产业化发展,设施装备水平也逐渐
提高。现代化连栋温室、日光温室的设计和建造水平
不断提高,温室的各项性能显著改善。同时,温室建

17 期 蒋卫杰等:设施园艺发展概况、存在问题与产业发展建议 3519
设、保养、维修的相关企业也快速增长,为设施园艺
发展提供了有力的保障。针对温室、大棚等特殊耕作
环境,研制生产了一些有多种作业功能的小型耕作机
械,小型整地起垄覆膜机、小型移栽机等。同时注重
开发适合中国不同区域设施栽培的新材料、新设备,
如温室降温保温材料、温室骨架复合材料、加温保温
设施、遮阳设施、灌溉设施、肥水一体化施肥机、机
械卷帘机等[23]。开发了一系列的温室采摘运输、穴
盘播种机,果蔬清洗分级机等装备系统,并应用于设
施园艺生产中,有力地提高了设施园艺的机械化生产
水平。 环境控制技术是现代设施园艺研究的重要内容,
中国已开发了一系列的设施园艺环境控制系统,如温
室环境数据采集控制系统、肥水一体化自动灌溉控制
系统,可以更加精确和高效地实现对温室环境参数的
自动控制;在栽培环境监测与控制方面,基于云技术、
无线传感器的物联网技术研究也取得显著进展。中国
设施园艺环境控制系统正朝着网络化、智能化和无线
化发展。 3.2.2 无土栽培技术和连作障碍调控技术已获广泛
应用 中国无土栽培系统主要包括水培和基质栽培两
大类,其中基质栽培占 95%以上。由中国农业科学院
蔬菜花卉研究所研发的有机生态型无土栽培是中国无
土栽培的主要形式,占全国无土栽培总面积 75%以上;
目前中国无土栽培技术主要用于设施园艺中番茄、黄
瓜、甜椒、甜瓜、生菜等蔬菜作物及花卉植物,在园
艺植物育苗上也有一定的应用[24-26]。近几年对无土栽
培的模式、基质配方和肥料配方、灌溉模式及产品品
质等[27-31]方面都做了深入研究。由于无土栽培在防止
肥料导致的环境污染、节约水资源、提高作物产量方
面的潜在优势,也越来越受到各地政府的重视。近年
来,中国设施园艺无土栽培面积不断扩大,已成为设
施作物生产的重要栽培方式。 设施园艺栽培由于土壤理化性质劣变、病原菌种
群数量激增及植物分泌的化感物质在土壤中聚集,造
成了不同程度的连作障碍,已成为制约中国设施园艺
持续发展的关键因素,土壤连作障碍调控技术是中国
设施园艺科研工作者多年的研究热点。通过合理的轮
作、间作伴生作物、种植填闲作物,可以改变土壤微
生态环境,减轻病害的发生,还可以改善土壤结构和
土壤肥力,提高作物产量和品质[32-33],在夏季利用太
阳光进行高温消毒、采用棉隆、威百亩和石灰氮进行
土壤消毒、增施有益菌(如“益生源”等)及选育抗
性品种等措施均能有效的缓解土壤连作障碍。 近几年,中国自主研发的设施蔬菜品种在长季节
栽培、外观品质、产量、耐贮性及抗病性等方面都有
了较快的提高,培育出一批番茄、辣椒、茄子、黄瓜
等专用品种,有效降低了国外品种在中国的市场占有
率。 3.2.3 资源利用率逐步提高,节能环保设施技术成为
设施园艺发展方向 中国设施园艺建设以合理利用当
地气候资源为原则,各个地区都研发了具有各自特点
的技术应用于日光温室及连栋温室,研究开发了多种
新型节能日光温室结构和新型高透光、防老化、防雾、
高保温的新型复合温室覆盖膜,研发和使用新型保温
材料、高效降温设备和相变蓄热墙体等,综合节能率
可达 11.6%—38.5%[12]。非耕地有效利用也取得了显著
进展,西部地区利用充足的光能资源和非耕地资源,
利用秸秆和沙为栽培基质进行主要设施蔬菜的生产,
实现了非耕地的有效利用[34-35]。膜下滴灌等微灌技术
获得广泛应用,肥料利用率提高 1 倍[36-37]。肥水一体
化使用技术和配套控制装备的研发也取得了长足的进
步,有效替代了国外类似产品的进口。 另外,中国设施作物病虫害防治技术,农药残
留控制技术等方面也取得了显著进步。通过选育抗
病、抗虫品种,推广应用了防虫网、性诱剂、黏虫
板、硫磺熏蒸、生物农药和有色薄膜等环境友好型
病虫害防治技术[38-39],减少了农药使用对环境的危
害。病虫害生态防治技术,对病虫害进行早期预测,
利用生物农药及天敌昆虫进行有效防治等都取得了
较好的效果。
4 中国设施园艺存在的主要问题
虽然近些年中国设施园艺发展迅速,设施栽培
面积与人均供应量已居于世界第一位。但由于中国
设施园艺的设施类型多样,相关科研工作开展时间
较短,在栽培技术、环境调控及专用品种等方面与
发达国家还有不小的差距。同时,目前中国设施园
艺仍以个体农户生产经营为主,产业化水平低,设
施园艺效益优势不能完全体现。目前中国设施面积
世界第一,其中 90%以上进行蔬菜生产,但中国温
室蔬菜平均年产量仅约为 90 000 kg·hm-2,设施蔬菜
的单位面积产量、质量、效益与劳动生产率与国外
相比还有较大的差距。 4.1 设施装备水平不高
中国设施园艺栽培面积已达到 187.4 万 hm2,但

3520 中 国 农 业 科 学 48 卷
90%以上为简易型设施,以个体农户小规模经营为主,
设施结构简单,性能差、空间小、结构不合理、抵御
不利气候条件能力较差、劳动强度大、劳动环境恶劣、
劳动生产率低等情况普遍存在。日光温室及塑料大棚
缺乏环境调控设备,对温室内温度、湿度、光照等调
控能力较低。目前中国还没有成套的设施结构及建造
的标准,个别标准之间存在重复和矛盾。机械化程度
较低,温室专用小型机械种类少而且没有普及,部分
地区基本依靠人力,导致劳动生产率低下。 4.2 科技含量相对较低
中国的设施园艺在环境控制、栽培管理技术、人
工智能技术、生物技术、物联网技术等方面与农业发
达国家还有很大的差距,导致设施农产品产量和质量
低下,水、肥资源浪费大,设施园艺效益差。中国无
土栽培面积仅有 1 000 余 hm2,只占设施栽培面积的
0.1%,远远低于发达国家 50%的水平。缺乏具有自主
知识产权的设施栽培专用品种,在商品品质、复合抗
病性、抗逆性等方面的育种水平与国外差距较大,温
室栽培蔬菜品种除华北型黄瓜外,其他品种尚未形成
主栽品种。温室作物生产技术都是零散性的经验、尤
其是大型温室生产,成套操作技术规范较少,缺少量
化指标。 4.3 土壤盐渍化、连作障碍、病虫害制约设施园艺发
展
由于越冬、早春设施栽培的低温弱光等原因,设
施栽培环境极易引起病虫害的发生,而且主要依赖化
学农药进行防治。中国设施栽培普遍依赖大肥大水提
高作物产量,养分利用率仅为 10%—20%,盲目超标
使用化肥和农药现象严重,导致产品产量低,品质下
降,有害物质超标,造成土壤盐渍化及环境和生态污
染,产品安全问题已成为中国社会普遍关心的问题。
中国的设施作物生产种植品种单一,连作条件下的病
虫害和连作障碍问题尤为严重,连作障碍和病虫害问
题成为制约设施蔬菜可持续生产的瓶颈,但中国的连
作障碍克服技术体系尚未成熟。
5 中国设施园艺发展方向与建议
国外设施园艺发展趋势是结构设备现代化、温室
管理自动化、经营规模大型化。总体来看,虽然中国
设施园艺近些年取得了快速的发展,但在先进技术、
设备研发、综合集成、推广等方面还远远不够。 5.1 中国设施园艺技术未来研究的重点领域
5.1.1 温室结构装备和环境精确控制技术 以低
碳节能设施为重点,研究具有蓄热—保温性能的轻
简装配式节能日光温室,提高温室抗逆能力和能源
利用效率;研究温室结构优化设计技术、开发适合
不同地区、气候条件的新型温室,尤其是新型日光
温室。研发温室新型覆盖材料、蓄热保温材料,研
究设施园艺生产过程机械技术和装备,提高机械化
作业水平。通过节能综合技术研究,优化设施结构,
增强保温性和光能利用率以及探索新型能源,降低
设施园艺能源消耗,提高经济效益和设施园艺可持
续发展能力。 研究温室智能环境控制及生产管理信息化系统和
温室控制专用传感器、物联网技术和云技术,实现对
温室内光、温、水、气、肥等诸多因子的优化精准控
制。 5.1.2 无土栽培技术研究 研究替代草炭和岩棉的
无土栽培生态型基质,开发生态型复合无土栽培基质,
栽培基质优化配制技术,研究无土栽培作物根区环境
的调控技术,营养液的消毒技术和配套设施设备。研
究营养液各种离子在线检测技术、基质无土栽培灌溉
自动控制技术和配套装备。开发低成本无土栽培模式、
无土栽培肥料、有机无土栽培技术。 5.1.3 病虫害可持续控制技术研究 研究主要设施
毁灭性病虫害粉虱、蓟马、甜菜夜蛾、根结线虫病、
灰霉病、晚疫病、枯萎病等的高效专一性天敌、生物
制剂,天敌与生物制剂规模化生产技术及其在不同设
施环境下的应用技术,开发设施病虫害的生态调控与
物理化学控制技术。 5.1.4 克服连作障碍技术研究 研究作物根际微生
态学变化,为研制减缓、调控连作障碍发生的功能性
产品与使用技术找出准确依据。研究功能性生物有机
肥、伴生和填闲作物种类来预防和治理连作障碍,研
制调控型生防制剂解决化学药剂使用量偏大、防治效
果不佳、药剂残留高等问题。 5.1.5 设施园艺专用品种 针对中国设施栽培环
境低温弱光、高温高湿、病虫害严重等特点,加大
蔬菜品种选育力度,选育适合不同栽培环境的专用
品种,摆脱跨国种业公司对中国高端蔬菜种子市场
的控制。 5.2 设施园艺产业化发展建议
设施园艺是中国农业生产中效益较高的支柱产
业,但产业化程度仍然较低,正处于全面提高的关键
期。政府对设施园艺产业化发展需加强宏观调控,采
取适当措施加速产业化进程。坚持优化布局,发挥区

17 期 蒋卫杰等:设施园艺发展概况、存在问题与产业发展建议 3521
域优势,选择设施园艺基础较好的区域,统筹育种、
栽培、装备、管理等多方面的力量,充分挖掘设施园
艺生产潜能,促进设施园艺持续发展。 5.2.1 加强政府宏观导向作用,加大惠农政策力度
设施园艺产业化发展应以市场为基础,以效益为中心。
各级地方政府应从本地区园艺产业发展实际情况出
发,按照各地的气候资源分布、市场需求、交通运输
情况及经济发展情况加强设施园艺发展的宏观引导,
因地制宜制定本地区设施园艺发展规划和发展目标,
有效地整合资金、技术、人才等资源,发挥比较优势,
突出地方特色,减少发展的盲目性,推进设施园艺产
业化进程。 落实中央一系列强农惠农富农政策,扶持鼓励
设施园艺发展。进一步扩大设施园艺装备纳入购机
补贴的范围,加大对农民和农民合作组织发展设施
园艺的扶持力度,加大对设施园艺财政、税费、信
贷和保险政策的支持,提高补贴与扶持的导向性与
效能。同时,加大基础设施建设投入,不断提高农
民发展设施园艺和抵御自然灾害的能力。带动和吸
引社会资金对设施园艺的投入,促进设施园艺快速
健康的发展。 5.2.2 加大设施园艺产业科技创新投入力度 加大
科技创新投入力度,支持设施园艺关键技术装备研发,
对从事设施园艺产业的科研、新技术和新品种的试验、
示范与推广工作给予资金的优先安排。国家及各级政
府应保障设施园艺科研、推广等经费,加强设施园艺
的职业技术教育资金投入,培养具有一定设施园艺理
论知识的种植者与管理者。 5.2.3 加强设施园艺装备标准化发展 虽然中国设
施园艺产业在温室建造、应用材料等方面取得了一定
的进展,但设施园艺相关的行业标准在温室设计、施
工、材料应用方面没有获得普遍的应用,在很大程度
上阻碍了设施园艺产业化发展。因此,各地区应根据
自身发展的需要加强产品标准和设计标准的制定和完
善,加强标准的执行,对农民自制的温室设施提出指
导建议,规范园艺设施装备生产建造。 5.2.4 完善园艺产品供求预警机制 园艺产品价格
波动主要是由产品供给波动造成的,为保证其供求平
衡和价格相对稳定,各级政府应完善风险防范机制。
在现有基础上加强设施园艺基础信息采集工作,分析
和预测产品供给情况及价格走势,提前进行市场预警,
使产品更加合理有效流通,提高应急能力。 5.2.5 构建质量安全控制长效机制 推广生态栽培
技术和高效低毒农药,严格控制肥料和农药等投入品
的施用,推进标准化生产和病虫害统防统治,大力推
广肥水一体化技术,构建质量安全控制长效机制。加
快科技成果转化应用,提高产业的整体技术水平,实
现产业不断升级。 References
[1] 喻景权. “十一五”我国设施蔬菜生产和科技进展及其展望. 中国
蔬菜, 2011(2): 11-23.
Yu J Q. Progress in protected vegetable production and research
during ‘the eleventh five-year plan’ in China. China Vegetable,
2011(2): 11-23. (in Chinese)
[2] 何芬, 马承伟. 我国设施农业发展现状与对策分析. 中国农学通报,
2007(3): 462-465.
He F, Ma C W. Development and strategy of facility agriculture in
China. Chinese Agricultural Science Bulletin, 2007(3): 462-465. (in
Chinese)
[3] Heuvelink E, Bakker M, Marcelis L F M.Climate and yield in a
closed greenhouse. Acta Horticulturae, 2008, 801:1083-1092.
[4] 郭世荣, 孙锦, 束胜, 李晶. 国外设施园艺发展概况、特点及趋势
分析. 南京农业大学学报, 2012, 35(5): 43-52.
Guo S R, Sun J, Shu S, Li J. General situations, characteristics and
trends of protected horticulture in foreign. Journal of Nanjing
Agricultural University, 2012, 35(5): 43-52. (in Chinese)
[5] Jiang W J. China’s Energy-Saving Greenhouse. Chronica Horticulturae,
2004(1): 15-17.
[6] 农业部农业机械化管理司 . 全国农业机械化统计年报 . 2008:
120-121.
Department of Agriculture Mechanization Management, Ministry of
Agriculture of the People’s Republic of China. The National
Agricultural Mechanization Statistical Yearbook. 2008: 120-121. (in
Chinese)
[7] 农业部农业机械化管理司. 全国农业机械化统计年报. 2009: 65.
Department of Agriculture Mechanization Management, Ministry of
Agriculture of the People’s Republic of China. The National
Agricultural Mechanization Statistical Yearbook. 2009: 65. (in
Chinese)
[8] 农业部农业机械化管理司. 全国农业机械化统计年报. 2010: 65.
Department of Agriculture Mechanization Management, Ministry of
Agriculture of the People’s Republic of China. The National
Agricultural Mechanization Statistical Yearbook. 2010: 65. (in
Chinese)
[9] 农业部农业机械化管理司. 全国农业机械化统计年报. 2011: 67.

3522 中 国 农 业 科 学 48 卷
Department of Agriculture Mechanization Management, Ministry of
Agriculture of the People’s Republic of China. The National
Agricultural Mechanization Statistical Yearbook. 2011: 67. (in
Chinese)
[10] 农业部农业机械化管理司. 全国农业机械化统计年报. 2012: 67.
Department of Agriculture Mechanization Management, Ministry of
Agriculture of the People’s Republic of China. The National
Agricultural Mechanization Statistical Yearbook. 2012: 67. (in
Chinese)
[11] 农业部农业机械化管理司. 全国农业机械化统计年报. 2013: 63.
Department of Agriculture Mechanization Management, Ministry of
Agriculture of the People’s Republic of China. The National
Agricultural Mechanization Statistical Yearbook. 2013: 63. (in
Chinese)
[12] 郭世荣, 孙锦, 束胜, 陆晓民, 田婧, 王军伟. 我国设施园艺概况
及发展趋势, 中国蔬菜, 2012(18): 1-14.
Guo S R, Sun J, Shu S, Lu X M, Tian J, Wang J W. Analysis of
general situation, characteristics, existing problems and development
trend of protected horticulture in China. China Vegetable, 2012(18):
1-14. (in Chinese)
[13] 金博, 乔晓军, 王成, 滕弘飞. 温室环境信息语音提示系统的设计
与实现. 农业工程学报, 2004, 20(4): 209-211.
Jing B, Qiao X, Wang C. Development of voice warning system for
the environmental information in greenhouse. Transactions of the
Chinese Society of Agricultural Engineering, 2004, 20(4): 209-211.
(in Chinese)
[14] Park D H, Cho S E, Park J W. The realization of greenhouse
monitoring and auto control system using wireless sensor network for
fungus propagation prevention in leaf of crop. Control and
Automation, 2009, 65: 28-34.
[15] van Straten G, van Willigenburg L G, van Henten E J. Optimal Control
of Greenhouse Cultivation. Boca Raton: CRC Press, 2011: 49-82 .
[16] Jiang W J, Qu D Y, Mu D, Wang L R. Protected cultivation of
horticultural crops in China. Horticulture. 2004, 30: 115-162.
[17] Wang J, Liu G. A design of greenhouse remote monitoring system
based on WSN and WEB. Computer and Computing Technologies in
Agriculture, 2012, 370: 247-256.
[18] Liu X Y, Guo S R.Xu Z G.Regulation of chloroplast ultrastrueture,
cross-section analomy of leaves, and morphology of tomato of cherry
tomato by different light irradiations of light-emitting siodes (LED).
HortScience, 2011, 46(2): 217-221.
[19] Kozai T, Ohyama K, Tong Y. Integrative environmental control using
heat pumps for reductions in energy consumption and CO2 gas
emission, humidity control and air circulation. Acta Horticuhurae,
2011, 893: 121-129.
[20] van der Lans C J M, Meijer R J M. A view of organic greenhouse
horticulture worldwide. Acta Horticulturae, 2011, 915: 1-22.
[21] Ghehsareh A M, Borji H, Jafarpour M. Effect of some culture
substrates (date-palm peat, cocopeat and perlitc) on some growing
indices and nutrient elements uptake in greenhouse tomato. African
Journal of Microbiology Research, 2011, 5(12): 1437-1442.
[22] 魏晓明, 齐飞, 丁小明, 鲍顺淑, 李中华, 何芬. 我国设施园艺取
得的主要成就. 农机化研究, 2010(12): 227-231.
Wei X M, Qi F, Ding X M, Bao S S, Li Z H, He F. Main achievements
in China facility horticulture. Journal of Agricultural Mechanization
Research, 2010(12): 227-231. (in Chinese)
[23] 齐飞, 周新群, 张跃峰, 李志. 世界现代化温室装备技术发展及对
中国的启示. 农业工程学报, 2008, 24(10): 279-284.
Qi F, Zhou X Q, Zhang Y F, Li Z. Development of world greenhouse
equipment and technology and some implications to China.
Transactions of the Chinese Society of Agricultural Engineering, 2008,
24(10): 279-284. (in Chinese)
[24] Jiang W J,Yu H J. Twenty years development of soilless culture in
mainland China. Acta Horticulturae, 2007, 759:181-193.
[25] 刘伟, 余宏军, 蒋卫杰. 我国蔬菜无土栽培基质研究与应用进展.
中国生态农业学报, 2006, 14(3): 4-7.
Liu W, Yu H J, Jiang W J. Review on research progress and
application of growing media for vegetable production in China.
Chinese Journal of Eco-Agriculture, 2006, 14(3): 4-7. (in Chinese)
[26] Jiang W J, Yu H J. Present situation and future development for
protected horticulture in Mainland China. Acta Horticulturae, 2008,
770: 29-35.
[27] Suo L N, Sun X Y Jiang W J. Effect of pretreatment on biomass
residue structure and the application of pyrolysed and composted
biomass residues in soilless culture. PLOS ONE, 2013,8(5): 1-9.
[28] 张晔, 余宏军, 杨学勇, 蒋卫杰. 棉秆作为无土栽培基质的适宜发
酵条件, 农业工程学报, 2013, 29(12): 210-217.
Zhang Y,Yu H J, Yang X Y, Jiang W J. Favorable conditions of cotton
straw composting using as soilless culture substrate. Transactions of
the Chinese Society of Agricultural Engineering, 2013, 29(12):
210-217. (in Chinese)
[29] 佟小刚, 蒋卫杰, 尹明安, 余宏军. 无土栽培基质中的微生物及其
对作物生长发育的影响. 园艺学报, 2005, 32(3): 544-550.
Tong X G, Jiang W J, Yin M A, Yu H J. Microorganism in soilless
substrates and its effects on crop growth. Acta Horticulturae Sinica,
2005, 32(3): 544-550. (in Chinese)

17 期 蒋卫杰等:设施园艺发展概况、存在问题与产业发展建议 3523
[30] Ahmed A F, Yu H J, Yang X Y, Jiang W J. Deficit irrigation affects
growth, yield, vitamin C content, and irrigation water use efficiency of
hot pepper grown in soilless culture. Hortiscience, 2014, 49(6):
722-728.
[31] 蒋卫杰, 余宏军, 朱德蔚, 杜永臣, 李红, 史振霞. 长季节无土栽
培番茄根部施用植物生长调节剂对根系活力的影响. 园艺学报,
2007, 34(3): 771-774.
Jiang W J, Yu H J, Zhu D W, Du Y C, L H, Shi Z X. Effect of plant
growth regulator on root activity of tomato in long-term soilless
cultivation. Acta Horticulturae Sinica, 2007, 34(3): 771-774. (in
Chinese)
[32] 苏世鸣, 任丽轩, 霍振华,杨兴明, 黄启为, 徐阳春, 周俊, 沈其荣.
西瓜与旱作水稻间作改善西瓜连作障碍及对土壤微生物区系的影
响. 中国农业科学, 2008, 41(3): 704-712.
Su S M, Ren L X, Huo Z H, Yang X M, Huang Q W, Xu Y C,
Zhou J, Shen Q R. Effects of intercropping watermelon with rain fed
rice on fusarium wilt and the microflora in the rhizosphere soil.
Scientia Agricultura Sinica, 2008, 41(3): 704-712. (in Chinese)
[33] 王玉彦, 吴凤芝, 周新刚. 不同间作模式对设施黄瓜生长及土壤环
境的影响. 中国蔬菜, 2009(16): 8-13.
Wang Y Y, Wu F Z, Zhou X G. Effects of different intercropping
patterns on the growth of cucumber in greenhouse and soil
environment. China Vegetables, 2009(16): 8-13. (in Chinese)
[34] 李建明, 王忠红, 邹志荣. 日光温室甜瓜沙化土有机配方研究. 西
北农林科技大学学报: 自然科学版, 2006, 34(6): 69-74.
Li J M, Wang Z H, Zou Z R. Study on sandy-soil organic prescription
for muskmelon in solar greenhouse. Journal of Northwest Sci-Tech
University of Agriculture and Forestry: Natural Science Edition, 2006,
34(6): 69-74. (in Chinese)
[35] 卜燕燕, 高艳明, 李建设, 李文甲. 甜椒沙培基质配方初选试验.
北方园艺, 2010(6): 82-84.
Bu Y Y, Gao Y M, Li W J. Primary election tests on different substrate
formulas of sweet pepper. Northern Horticulture, 2010(6): 82-84. (in
Chinese)
[36] 李战国. 不同灌溉施肥方式对樱桃番茄产量和品质的影响. 安徽
农业科学, 2008, 36(18): 7623-7624.
Li Z G. Effects of different treatments of irrigation and fertilization on
yield and quality of cherry tomato. Journal of Anhui Agricultural
Sciences, 2008, 36(18): 7623-7624. (in Chinese)
[37] 姬景红, 张玉龙, 张玉玲, 虞娜, 王伟承. 灌溉方法对保护地土壤
有机氮矿化特性的影响. 土壤学报, 2009(5): 869-877.
Ji J H, Zhang Y L, Zhang Y L, Yu N, Wang C W. Effect of irrigation
methods on mineral ization of soil organic nitrogen in protected field.
Acta Pedologica Sinica, 2009(5): 869-877. (in Chinese)
[38] 杨宇红, 刘俊平, 杨翠荣, 龚惠芝, 冯东昕, 谢丙炎. 无致病 hrp-突
变体防治茄科蔬菜青枯病. 植物保护学报, 2008, 35(5): 433-437.
Yang Y H, Liu J P, Yang C R, Gong H Z, Feng D X, Xie B Y. Control
of Solanaceae vegetable bacterial wilt with avirulent hrp-mutants.
Acta Phytophylacica Sinica, 2008, 35(5): 433-437. (in Chinese)
[39] 石延霞, 关爱民, 李宝聚. 瓜枝孢弱致病菌诱导黄瓜植保素的积累
及抑菌活性. 园艺学报, 2007, 34(2): 361-365.
Shi Y X, Guan A M, Li B J. Accumulation and activity of phytoalexins
in cucumber induced by pathogenicity-imparired strain of
Cladosporium cucumernium. Acta Horticulturae Sinica, 2007, 34(2):
361-365. (in Chinese)
(责任编辑 赵伶俐)

中国农业科学 2015,48(17):3524-3540 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.018
收稿日期:2015-02-12;接受日期:2015-06-18 基金项目:国家重点基础研究发展计划(2011CB100606)、国家公益性行业(农业)科研专项(201303093)、国家自然科学基金(31171932) 联系方式:陈学森,E-mail:[email protected]
主要果树果实品质遗传改良与提升实践
陈学森1,郭文武
2,徐 娟
2,丛佩华
3,王力荣
4,刘崇怀
4,李秀根
4,吴树敬
1,姚玉新
1,陈晓流
1
(1 山东农业大学园艺科学与工程学院/作物生物学国家重点实验室,山东泰安 271018;2华中农业大学园艺植物生物学教育部重点实验室/农业部
华中地区园艺作物生物学与种质创制重点实验室/柑橘研究所,武汉 430070;3 中国农业科学院果树研究所/农业部园艺作物种质资源利用重点实验室,
辽宁兴城 125100;4 中国农业科学院郑州果树研究所,郑州 450009)
摘要:全面总结分析国内外果树果实品质改良现状、中国果树品质改良与发达国家的差距与成因,在
此基础上系统阐述柑橘、苹果、梨、桃及葡萄等主要果树果实品质遗传改良的目标、技术途径与成果、品
质性状遗传倾向、芽变机理,并举例阐述柑橘原生质体融合与无核品种创制、功能型苹果及其育种技术体
系创建、早熟核果类果树品种和无核葡萄胚培育种、野生樱桃李资源品种化及转基因缩短童期新技术等方
面取得的突破性进展。最后针对中国果实品质改良提出几条建议:进一步加强野生果树资源评价及亲本利
用研究,努力拓宽栽培品种的遗传基础,协同提升果实品质;进一步开展细胞工程等生物技术以及芽变机
理和性状遗传变异规律研究,建立生物技术与常规技术结合、杂交育种与芽变选种优势互补的果树果实品
质高效改良技术体系;进一步加强新品系的国际品种区域栽培试验及国际品种权申请与保护,培育国际性
的果树“大品种”。
关键词:苹果;柑橘;梨;葡萄;桃;果实品质;改良育种
Genetic Improvement and Promotion of Fruit Quality of Main Fruit Trees
CHEN Xue-sen1, GUO Wen-wu2, XU Juan2, CONG Pei-hua3, WANG Li-rong4, LIU Chong-huai4, LI Xiu-gen4, WU Shu-jing1,YAO Yu-xin1, CHEN Xiao-liu1
(1College of Horticulture Science and Engineering, Shandong Agricultural University/State Key Laboratory of Crop Biology, Tai’an
271018, Shandong; 2Key Laboratory of Horticultural Plant Biology, Ministry of Education/Key Laboratory of Central China
Horticultural Crop Biology and Germplasm Enhancement, Ministry of Agriculture/Citrus Research Institute, Huazhong
Agricultural University, Wuhan 430070; 3 Research Institute of Pomology, Chinese Academy of Agricultural Sciences/Key
Laboratory of Fruit Germplasm Resourses Utilization, Ministry of Agriculture, Xingcheng 125100, Liaoning; 4 Zhengzhou
Fruit Research Institute, Chinese Academy of Agricultural Sciences, Zhengzhou 450009)
Abstract: The current situation of fruit quality improvement both at home and abroad, the gaps between China and the developed countries and the reasons causing these gaps were firstly summarized and analyzed. Based on the review, the aim of the genetic improvement for citrus, apple, pear, peach, grape and other main fruit trees, the techniques used in genetic improvement and the achievements obtained, fruit quality genetic predisposition and mechanisms of bud mutation sere systematically discussed. Meanwhile, the breakthroughs in citrus protoplast fusion and seedless variety creation, functional apple breeding system establishment, the embryo culture of early ripening stone fruit varieties, seedless grape, varieties developing from wild cherry plum, and juvenile shortening transgenic technology were illustrated through presenting the concrete successful cases. Lastly, three recommendations directed at Chinese fruit quality improvement were proposed: Further strengthening the research,
jia
文本框
返回目录

17 期 陈学森等:主要果树果实品质遗传改良与提升实践 3525
utilization and evaluation of wild fruit tree resources to broad the genetic basis of the fruit cultivars and enhance the fruit quality. Further deepening the research on biotechnological approaches such as cell engineering, the mechanism of bud mutation, and the principles of genetic variation of traits to establish a fruit improvement technique system with high efficiency combining modern biotechnology and conventional techniques and sexual cross breeding and bud mutation selection. Further strengthening the regional cultivation tests of new international varieties and applying for variety right international protection to culture international “Big variety”.
Key words: apple; citrus; pear; grape; peach; fruit quality; improved breeding
0 引言
果树产业的转型升级和果实品质提升与增效
是一项系统工程,要提高果实品质,首先要提高土
壤品质。土壤品质,即土壤综合肥力包括土壤有机
质含量及无机肥利用效率等 9 个因素,其中土壤有
机质含量的高低是评价土壤综合肥力的核心指标。
果实品质问题,大多数与土壤有关,例如,苹果果
实苦痘病的病因是土壤缺钙及土壤酸化等。清耕除
草、好气微生物对土壤有机质的降解(转化为 CO2
而逸出土壤)以及有机肥投入不足等,导致果园土
壤有机质含量呈持续下降及贫瘠化的态势。因此,
改革中国水果栽培模式及土壤管理制度,大力推广
现代“宽行高干集约高效”栽培模式及果园生草
培肥地力技术,是中国果树产业转型升级及提质增
效的关键[1-6]。 另外,良种是提质增效的遗传基础,良种与良法
配套能 大限度地发挥良种的基因效益。因此,优
质品种的选育与推广对水果产业的提质增效至关重
要。果实品质是由许多单位性状构成的综合性状,
其中许多单位性状是微效多基因或寡基因控制的数
量性状或数量—质量性状。依据研究或利用的角度
不同,可将果实品质分为外观品质、风味品质、营
养品质、贮运品质、鲜食品质、加工品质及遗传品
质等,构成因素包括大小、果形、色泽、质地、风
味、香气及功能成分等。例如,由果皮、果肉硬度
(firmness)及脆度(crispness)等因素构成的苹果
果实质地(fruit texture),不仅影响鲜食品质,而
且是决定贮运品质的关键因子。因此,硬脆多汁是
苹果品质育种的目标[7-8]。 近年,由于分子生物学技术的飞速发展,苹果、
柑橘、葡萄、梨及桃等主要果树果实品质形成机理
研究取得了较大进展[9-10]。本文仅对这些果树果实
品质遗传改良和提升的思路、方法及成果进行概述,
为中国果树品质育种提供借鉴或参考。
1 国内外果树果实品质改良现状及
水果产业发展趋势
1.1 发达国家果树果实品质改良现状、特点与趋势
近几年调研发现,美国、法国、新西兰及日本等
世界水果产业强国用工业化的思路、理念、措施来经
营、管理现代果业,全面实现了良种化、多样化、区
域化、集约化、规模化、标准化、机械化、设施化、
技术简化、生草化、水肥一体化、信息化、安全化、
组织化、一体化,取得了显著的经济、社会和生态效
益[11]。其中,与果实品质改良及品种有关的趋势与特
点主要有以下两点: (1)建立有效运行的水果新品种权保护、新品种
权转让及水果良种苗木繁育制度。 这不仅可规范水果
苗木市场,保障苗木质量,而且有效保护了育种者、
育苗者及栽培者的利益,调动育种者的积极性,加速
新品种的选育、推广与更新换代,保证了生产上采用
的品种及其品质能够满足市场及消费需求,实现了水
果产业的有序、有力和高效发展。 (2)良种的区域化、多样化和规模化。优良品
种高度的区域化:即每个地区有 佳的品种组成,
每个品种均种植在其 适宜的地区,以 大限度地发
挥优良品种的基因潜力;因消费习惯、产业定位及生
态条件的不同,世界各国水果生产的主栽品种比例可
能存在一定差异,但存在一个共同特点就是均采用多
样化的品种。例如,意大利主栽苹果品种比例分别为
‘金冠’23%、‘嘎拉’20%、‘元帅’16%、‘富士’
14%、‘布鲁本’12%、‘澳洲青苹’4%、‘粉红女
士’2%,其他品种不足 9%;欧美发达国家 70%的农
场主水果园经营面积在 3—12 hm2, 多超过 45 hm2,
实现了规模化,生产过程实现了标准化和机械化。因
此,虽然发达国家果园单位土地面积的利润率并不高,
但由于规模化和机械化,劳动生产率高、成本低,使
得农场主的收益率高。

3526 中 国 农 业 科 学 48 卷
1.2 中国主要果树果实品质遗传改良、资源保护与品
种推广中存在的主要问题
与世界水果生产先进国家相比,中国果树产业仍
存在较大差距,需要研究解决的问题依然很多。尽管
中国建立了相关产业技术体系,为育种专家提供了持
续稳定的经费支持,水果新品种(系)的数量和质量
稳步提高,果实品质遗传改良的成效显著,与国外发
达国家的差距在逐渐缩小。但在果实品质遗传改良、
品种选育与推广方面,依然存在如下 3 个主要问题: (1)野生资源评价及亲本利用的力度有待加强。
中国是柑橘、苹果、葡萄及桃等多种果树的起源演化
中心之一,野生资源极为丰富,是进行品质及抗逆育
种的珍贵基因库,但资源破坏严重,保护、评价及亲
本有效利用的力度尚待加强。 (2)中国育成的新品种(系)缺乏国际范围的大
区试。国内育成了许多综合经济性状优良的果树新品
种,均有可能成为世界性的“大品种”,但目前在国
际上的影响较小,主要原因是缺乏世界范围的“大区
试”及国际植物新品种权保护[11]。例如,郑州果树研
究所培育的苹果品种‘华冠’,经过在新西兰原苹果、
梨销售委员会下属的新西兰果树品种改良处(FIPIA)
的数十个国际苹果品种对比试验后被该国确认为是一
个具有国际推广价值的品种。据新西兰 ENZA Fruit Ltd.报告,2003 年由该公司在香港组织的苹果品种鉴
评会上,‘华冠’在与‘富士’‘嘎拉’和‘太平洋
玫瑰’(Pacific Rose)的较量中,位列第一[12]。 (3)果实品质遗传改良及新品种推广应用的体制
机制有待完善。依法保护育种者、育苗者及栽培者的
利益,是国外发达国家水果产业有序、有力、高效发
展的成功经验之一。中国新修改的《种子法》保护范
围虽然包括苹果等主要果树,但监管力度有待加强。
一是近几年中国授权的水果新品种权有 1 000 余例,
没有一例转让成功,育种者的利益得不到有效保护,
不利于新品种的选育和推广;二是苗木质量难以保障。
因此,必须依据《种子法》,进一步加强果树苗木繁
育市场的监管力度,而依法取得苗木生产许可证的公
司或单位,要严格按照苗木繁育有关技术标准生产优
质壮苗,建立苗木质量追溯制度[13]。
2 品质育种目标
2.1 苹果
世界大多数国家鲜食苹果品质育种的目标是肉
质脆、组织细、汁液多、酸甜味浓、有芳香、外观
好、色泽艳丽、耐贮藏、货架寿命长。当今 受欢
迎的品种是‘富士’‘嘎拉’‘司米斯’‘布瑞拜’
‘乔纳金’以及老品种‘金冠’和‘新红星’等,
这些品种虽然被世界大多数国家广泛栽培,但都存
在一些缺点,培育的新品种要以超过这些品种为标
准[12]。 苹果耐贮性好,供应周期长,果实含有较高比例
的、人体较容易吸收的游离多酚,具有很好的抗氧化、
抗肿瘤、预防心脑血管疾病及护肝等作用,营养保健
价值高,有“一天一苹果,医生远离我”的美誉,世
界上相当多的国家都将其列为主要消费果品而大力推
荐[11,14]。中国是世界上 大的苹果生产和消费国,其
中 2012 年生产苹果 3.95×107 t,主要用于鲜食,且近
70%是类黄酮含量较低的‘红富士’品种。因此,
为保证消费者从苹果中摄取足够的多酚和类黄酮,
陈学森等[15]提出了“功能型苹果”的概念(指在果实
中富含类黄酮、可鲜食或加工的保健型苹果)及其育
种思路,其中鲜食类的功能型苹果,应满足 3 个条件:
①具有较高的功能成分含量,类黄酮含量应是‘红富
士’苹果品种的 2 倍以上;②具有较好的鲜食品质,
包括果实硬度、脆度及可溶性固形物含量等;③具有
较好的外观和储运品质。 2.2 柑橘
世界鲜食柑橘品质育种的目标是无核、易剥皮、
风味浓和有香味。通过多年的育种实践,目前已推出
一些能满足这 4项要求的品种,例如摩洛哥培育的‘W.默科特’(W. Murcott,也称 Afourer),西班牙等国
培育出的几个克里曼丁橘(Clementine)品种,以及
美国育出的高糖系杂柑,日本育出的‘不知火’杂柑
等基本上可以满足以上要求,正在成为市场的新宠;
色泽作为重要的外观品质,同样受到重视,包括中国
在内的一些国家把果皮和果肉的色泽,特别是橙红或
者深红色作为育种目标。例如日本育出的‘天草’‘南
香’和‘山下红’均表现果皮色泽橙红鲜艳,十分吸
引消费者[16]。 2.3 桃
桃果实品质包括果实类型、大小、色泽、质地、
风味和其他一切影响消费者感官的因素,果实类型的
多样化是世界桃育种的方向,油桃、蟠桃、油蟠桃在
鲜食品种中的比例将大幅度提高,果实鲜红以及黄、
绿、白等纯色品种将更加吸引消费者,硬溶质或半不
溶质的鲜食品种将成为生产主流,风味浓郁、富含功
能性成分的品种将更受青睐[17]。

17 期 陈学森等:主要果树果实品质遗传改良与提升实践 3527
2.4 葡萄和梨
鲜食葡萄育种的目标是大粒、无核、抗病、高含
糖量、具有玫瑰香味、耐贮运、货架期长、适应不同
生态区[18]。世界梨品种改良的目标主要为内在品质
好、果型较大、红皮、耐贮藏、栽培省力、抗黑星病
和腐烂病,中国梨以鲜食为主,果实内在品质和外观
品质提升是梨遗传改良的主要目标[19]。
3 果实品质育种的技术途径
3.1 野生资源利用
农业野生植物的遗传多样性不仅是现代农业可持
续发展的物质基础,也是维护生态安全的重要屏障。
因此,保护和利用农业野生植物的遗传多样性是现代
农业的重要特征之一,符合保护自然、利用自然、回
归自然的发展理念[20]。野生果树不仅遗传多样性丰
富,而且在长期的自然选择与生存抗争的过程中,逐
渐形成了一些优良特性,是品质与抗逆育种的珍贵基
因库,但因农田开垦等因素而导致野生果树资源的遗
传多样性正遭到严重破坏,濒临灭绝[21-22]。因此,研
究建立包括原生境保护、异地建圃保存、离体器官建
库保存及利用保存等多层次的种质资源保护、保存技
术体系,对野生果树种质资源的科学保护与持续高效
利用具有重要意义[23-28]。通过野生资源品种化及资源
亲本利用等方式,选育出能产业应用的新品种,达到
种质保存的目的[29]。因此,野生资源品种化是利用
野生资源进行果实品质改良的重要方式之一。例如,
樱桃李(Prunus cerasifera Ehrh.)仅分布在新疆伊犁,
是核果类果树 原始的一个种,其果实的总酚含量和抗
氧化能力分别为 1.3—6.1 g·kg-1 FW 和 11.2—44.8 μmol Trolox·g-1 FW,其中紫果的抗氧化能力和保健价值高于蓝
莓,但濒临灭绝[30-33]。因此,山东农业大学从新疆伊犁
引进 2.8 万株新疆野生樱桃李自然杂交实生苗,在青岛胶
州建立了迁地种植圃;由于水、温匹配,青岛胶州的单
果质量变异系数是伊犁霍城的 7.7 倍,优异性状得以充分
表达,从中选育出鲜食型‘森果佳人’和鲜食与加工兼
用型‘森果红露’2 个大果型新品种,实现了野生樱桃李
资源的品种化[34-36]。 野生资源亲本利用也是果实品质改良的重要方式
之一。遗传多样性评价及优良性状功能基因挖掘是亲
本利用的基础,近几年来,中国在野生稻及野生大豆
等农业野生植物资源遗传多样性评价及优良性状功
能基因挖掘等方面的研究已取得了许多突破性的进
展[37-39]。在野生果树方面,安萌萌等[40-41]研究表明,
野生秋子梨(Pyrus ussuriensis Maxim)在 AFLP 标记、
果实形态、糖酸组分及香气成分上存在广泛的遗传变
异,参试的 35 个实生株系间差异明显,遗传多样性极
为丰富,并且有酸类和烃衍生物类 2 类特有化合物和
111 种特有成分,进一步挖掘利用的潜力巨大。在新
疆野苹果方面,山东农业大学陈学森课题组研究发现,
新疆野苹果的果实形状、大小、颜色、挥发性化合物
总含量、各类挥发性化合物种类数及其含量、主要挥
发性化合物分离比率与含量以及糖、酸及酚类物质组
分等表型性状的变异系数均在 10%以上,表现出丰富
的遗传多样性[21,42-45]。为达到亲本利用的目的,采用
以“扩大遗传基础”为核心、以“多组合、大群体”
为保障的品质育种策略,自 2009 年以来,先后构建了
‘红富士’苹果品种×新疆红肉苹果的 F1及回交一代
(BC1)杂种分离群体 40 个,定植杂种实生苗 4 万株,
并选育出‘红心 7 号’等新品系 10 个。 3.2 常规育种
根据新品种遗传变异的来源,果树常规育种主要
包括两类:一是利用基因重组进行新品种培育或创
制,包括营养性杂交和远缘杂交育种(人工杂交)及
实生选种(自然杂交);二是利用基因突变对现有品
种进行修饰或改良,主要指芽变选种。近 30 年来,
广大果树育种工作者根据生产、市场和消费者的需
求,综合采用杂交、实生和芽变选种技术,取长补短,
优势互补,不断选育出新品种,使柑橘、苹果、葡萄、
梨及桃等主要果树果实外观、风味、营养、贮运及鲜
食品质均得到有效改良,有力推动了水果产业的高效
发展[16-19,46-48]。 对柑橘、苹果、葡萄、梨、桃及杏等主要果树育
成品种的育种方法或途径的统计表明(表 1),不同
树种及同一树种不同品种间存在明显差异。苹果、柑
橘、葡萄、梨、桃及杏等树种共调查 3 674 个品种,
芽变品种 600 个,占 16.3%,其中以苹果和柑橘芽变
品种的比例 高,分别是 36.2%和 75.4%,其次是葡
萄芽变品种的比例(17.1%)与所调查各树种的平均
值(16.3%)较接近,梨和桃芽变品种的比例均不足
5%,而杏至今没有发现可利用的芽变。除了芽变的频
率外,芽变的幅度或范围亦存在较大差异,以柑橘的
变异幅度 大,从极早熟到极晚熟、从果皮颜色到果
肉颜色等许多性状均有大量的芽变品种,例如芽变品
种‘奉晚脐橙’比原来的‘奉节 72-1’晚 2 个月采
收,而梨仅有频率较低的早熟、倍性、大果及果皮
颜色的芽变[16,46,50]。已有的研究结果表明,苹果和柑
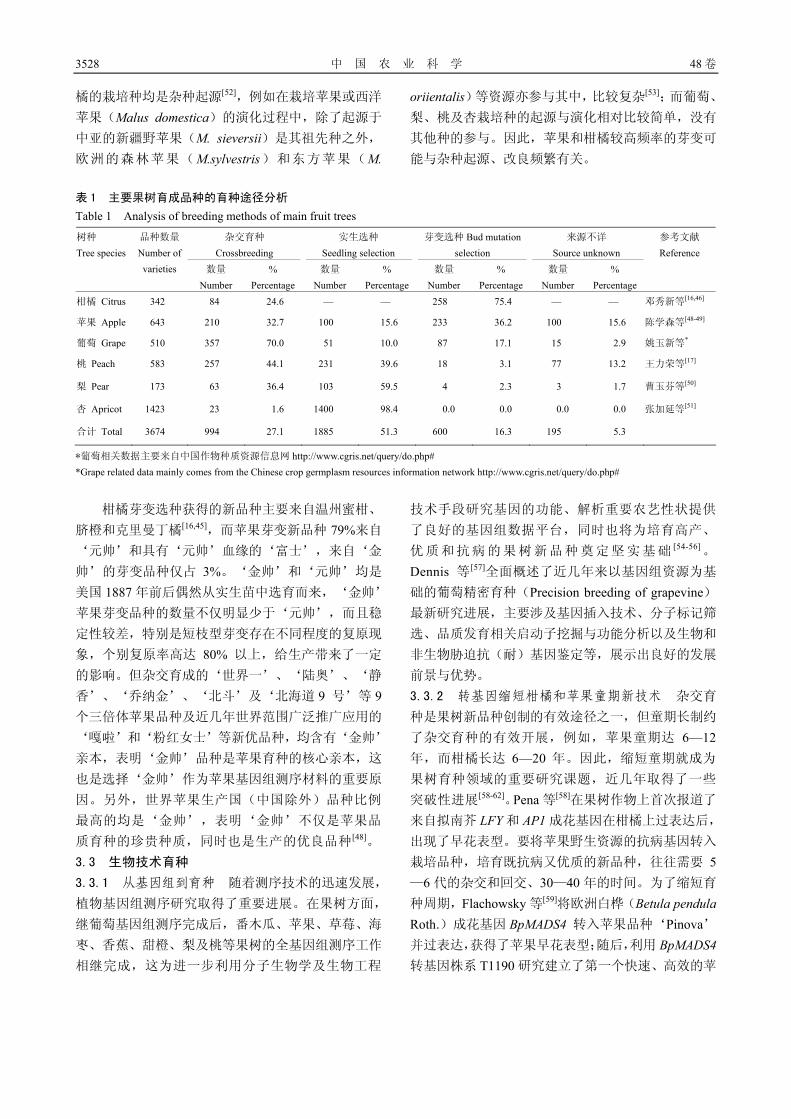
3528 中 国 农 业 科 学 48 卷
橘的栽培种均是杂种起源[52],例如在栽培苹果或西洋
苹果(Malus domestica)的演化过程中,除了起源于
中亚的新疆野苹果(M. sieversii)是其祖先种之外,
欧洲的森林苹果(M.sylvestris)和东方苹果(M.
oriientalis)等资源亦参与其中,比较复杂[53];而葡萄、
梨、桃及杏栽培种的起源与演化相对比较简单,没有
其他种的参与。因此,苹果和柑橘较高频率的芽变可
能与杂种起源、改良频繁有关。
表 1 主要果树育成品种的育种途径分析
Table 1 Analysis of breeding methods of main fruit trees
杂交育种 Crossbreeding
实生选种 Seedling selection
芽变选种 Bud mutation selection
来源不详 Source unknown
树种 Tree species
品种数量 Number of varieties 数量
Number %
Percentage 数量
Number %
Percentage数量
Number %
Percentage数量
Number %
Percentage
参考文献 Reference
柑橘 Citrus 342 84 24.6 — — 258 75.4 — — 邓秀新等[16,46]
苹果 Apple 643 210 32.7 100 15.6 233 36.2 100 15.6 陈学森等[48-49]
葡萄 Grape 510 357 70.0 51 10.0 87 17.1 15 2.9 姚玉新等*
桃 Peach 583 257 44.1 231 39.6 18 3.1 77 13.2 王力荣等[17]
梨 Pear 173 63 36.4 103 59.5 4 2.3 3 1.7 曹玉芬等[50]
杏 Apricot 1423 23 1.6 1400 98.4 0.0 0.0 0.0 0.0 张加延等[51]
合计 Total 3674 994 27.1 1885 51.3 600 16.3 195 5.3
*葡萄相关数据主要来自中国作物种质资源信息网 http://www.cgris.net/query/do.php# *Grape related data mainly comes from the Chinese crop germplasm resources information network http://www.cgris.net/query/do.php#
柑橘芽变选种获得的新品种主要来自温州蜜柑、
脐橙和克里曼丁橘[16,45],而苹果芽变新品种 79%来自
‘元帅’和具有‘元帅’血缘的‘富士’,来自‘金
帅’的芽变品种仅占 3%。‘金帅’和‘元帅’均是
美国 1887 年前后偶然从实生苗中选育而来,‘金帅’
苹果芽变品种的数量不仅明显少于‘元帅’,而且稳
定性较差,特别是短枝型芽变存在不同程度的复原现
象,个别复原率高达 80% 以上,给生产带来了一定
的影响。但杂交育成的‘世界一’、‘陆奥’、‘静
香’、‘乔纳金’、‘北斗’及‘北海道 9 号’等 9个三倍体苹果品种及近几年世界范围广泛推广应用的
‘嘎啦’和‘粉红女士’等新优品种,均含有‘金帅’
亲本,表明‘金帅’品种是苹果育种的核心亲本,这
也是选择‘金帅’作为苹果基因组测序材料的重要原
因。另外,世界苹果生产国(中国除外)品种比例
高的均是‘金帅’,表明‘金帅’不仅是苹果品
质育种的珍贵种质,同时也是生产的优良品种[48]。 3.3 生物技术育种
3.3.1 从基因组到育种 随着测序技术的迅速发展,
植物基因组测序研究取得了重要进展。在果树方面,
继葡萄基因组测序完成后,番木瓜、苹果、草莓、海
枣、香蕉、甜橙、梨及桃等果树的全基因组测序工作
相继完成,这为进一步利用分子生物学及生物工程
技术手段研究基因的功能、解析重要农艺性状提供
了良好的基因组数据平台,同时也将为培育高产、
优质和抗病的果树新品种奠定坚实基础 [54-56]。
Dennis 等[57]全面概述了近几年来以基因组资源为基
础的葡萄精密育种(Precision breeding of grapevine)新研究进展,主要涉及基因插入技术、分子标记筛
选、品质发育相关启动子挖掘与功能分析以及生物和
非生物胁迫抗(耐)基因鉴定等,展示出良好的发展
前景与优势。
3.3.2 转基因缩短柑橘和苹果童期新技术 杂交育
种是果树新品种创制的有效途径之一,但童期长制约
了杂交育种的有效开展,例如,苹果童期达 6—12年,而柑橘长达 6—20 年。因此,缩短童期就成为
果树育种领域的重要研究课题,近几年取得了一些
突破性进展[58-62]。Pena 等[58]在果树作物上首次报道了
来自拟南芥 LFY 和 AP1 成花基因在柑橘上过表达后,
出现了早花表型。要将苹果野生资源的抗病基因转入
栽培品种,培育既抗病又优质的新品种,往往需要 5—6 代的杂交和回交、30—40 年的时间。为了缩短育
种周期,Flachowsky 等[59]将欧洲白桦(Betula pendula Roth.)成花基因 BpMADS4 转入苹果品种‘Pinova’并过表达,获得了苹果早花表型;随后,利用 BpMADS4转基因株系 T1190 研究建立了第一个快速、高效的苹

17 期 陈学森等:主要果树果实品质遗传改良与提升实践 3529
果抗病育种技术体系,可将育种周期由 30—40 年缩短
至几年时间[60]。为了进一步改进快速育种体系的应用
效果,Kathleen 等[61]已将成花基因 BpMADS4 转入到
了 4 个苹果品种的不同连锁群。 近,Yamagishi 等[62]研发了一种方法,即通
过基因修饰,同时将能过表达的拟南芥成花基因
(AtFT)和沉默的苹果终端花基因(MdTFL1-1)与
苹果潜隐球形病毒(ALSV)构建表达载体(ALSV- AtFT/MdTFL1-1),苹果实生苗子叶接种这种病毒
载体后,绝大多数实生幼苗在当年就能正常开花结
实并产生种子,且潜隐病毒不能通过种子传播,对
下一代无任何影响。因此,这种方法作为育种新技
术,对缩短果树杂交育种周期、提高育种效率具有
重要意义。 3.3.3 柑橘原生质体融合技术 柑橘为全球第一大
水果,也是中国南方 重要的果树。中国柑橘资源丰
富,为柑橘品质改良和新品种培育奠定了种质基础。
中国柑橘以鲜食为主,果实品质提升是中国柑橘遗传
改良的主要目标。近年,通过常规育种途径,全世界
已获得一批新种质和新品种[16]。但是,柑橘常规育种
存在童期长、珠心胚干扰、性器官败育、自交不亲和
及遗传上高度杂合等问题,使得通过常规手段进行柑
橘遗传改良的进展缓慢、效率不高。为此,华中农业
大学围绕柑橘细胞工程开展了 20 余年的系统研究,有
效克服了上述生殖障碍,成功应用于柑橘品质改良,
特别是无核育种取得了诸多突破性进展,分述如下: (1)创制异源四倍体体细胞杂种 与常规有性杂交相比,原生质体融合可以打破生
殖隔离,为柑橘不同种、属间遗传重组提供了可能;
可以实现有益性状的定向转移,对柑橘品质遗传改良
具有重要意义。目前全世界范围内已获得近 500 例不
同种、属间的体细胞杂种[63]。四倍体体细胞杂种鲜有
作为鲜食品种直接推广的,其主要目标是为无核三倍
体育种提供优良的育种中间材料,多数组合均是针对
该目标设计的。但也有少数组合,如‘Nova 橘柚
+Succari 甜橙’、‘Pink Marsh 葡萄柚+Murcott 橘橙’
体细胞杂种表现果实品质优良、易剥皮、具香气、果
实少籽甚至无籽等特点[64]。因此,培育四倍体体细胞
杂种可作为柑橘品质改良的一条重要途径。另外,四
倍体体细胞杂种的花粉还可用于柚瘪籽果实生产,授
粉后的果实多数为瘪籽和种痕,且果实品质(糖含量)
还提高了,这是植物体细胞杂种直接用于生产实践的 先例,也是柚无核改良的一条有效途径。
(2)倍性杂交培育无核三倍体 三倍体一般减数分裂异常,为天然的无核类型。
除了无核外,三倍体育种还可实现其他性状的改良,
如三倍体可以解决无核突变体由于种子数减少而导致
的果实变小问题。因此,三倍体育种在柑橘品质遗传
改良中应用潜力巨大。目前,世界上包括美国、西班
牙、中国、意大利、日本等国家均开展了柑橘三倍体
育种研究[16],尤其是近年来细胞融合技术的快速发
展,大大推动了柑橘三倍体育种的进程。美国佛罗里
达大学柑橘研究与教育中心采用倍性杂交已获得约
12 000 株三倍体杂种,其中很大一部分是体细胞杂种
做父本与二倍体杂交获得的,部分三倍体已渡过童期
并开花结果,表现果实无籽的性状[63]。除了无核性
状,美国还选育出了几个呋喃香豆素含量很低的葡萄
柚三倍体杂种[65]。华中农业大学也很早开始了三倍
体育种研究,伊华林和邓秀新[66]以体细胞杂种‘哈
姆林甜橙+粗柠檬’与单胚性柚及单多胚混合品种华
农本地早橘杂交,获得了一大批三倍体植株,为中国
柑橘三倍体育种技术路线的建立打下了坚实的基础。
近年来,华中农业大学以新开花的品质优良的体细胞
杂种为父本与中国一些品质优良的二倍体地方品种
杂交,共配置近 40 个组合,经幼胚离体培育及流式
细胞仪倍性分析,共获得三倍体 1 400 余株,四倍体
150 余株[67-69],这些三倍体及四倍体育种材料为中国
柑橘无核品种选育及未来三倍体育种奠定了坚实的
材料基础。 柑橘同源四倍体在三倍体育种方面也有不可替代
的价值,利用化学诱导的同源四倍体进行倍性杂交培
育柑橘三倍体取得了良好的进展[16]。如西班牙从 4400余株三倍体中选出一个果品品质优良、香气浓郁的三
倍体品种‘IVIA-600’[70];意大利也成功选育出‘Tacle’‘Clara’‘Camel’和‘Reale’等三倍体新品种[71]。
同样,日本也从约 1 000 株三倍体中获得了一个无核、
晚熟、易剥皮的三倍体新品种‘Kiho’。中国近年也
开展了柑橘同源四倍体的发掘研究,华中农业大学发
掘有 20 余个品种的同源四倍体以及其他类型的多倍
体,其中‘鄂柑 1 号’椪柑等品种的同源四倍体已开
花结果,正用于无核三倍体培育(郭文武,未发表资
料)。值得注意的是,由于 2n 配子的存在,柑橘二倍
体间杂交也可以获得三倍体。西班牙通过此方式获得
了 4 000 余株三倍体,并从中选育出了两个品质优良
的三倍体品种‘Sofar’和‘Garbi’[72]。 (3)创制胞质杂种转移 CMS 性状

3530 中 国 农 业 科 学 48 卷
由于倍性增加,柑橘多倍体也表现出一些不良的
园艺学性状,如枝刺粗长、果皮变厚等。因此,二倍
体水平上的柑橘无核育种意义重大。在柑橘细胞融合
再生的植株中,除了预期的四倍体体细胞杂种外,还
获得了非亲本类型的二倍体植株,这类植株包含叶肉
原生质体亲本的细胞核与另一亲本的细胞质基因组,
被称为胞质杂种[73]。胞质杂种的产生为在二倍体水平
上的柑橘无核育种提供了一条新的思路,即通过对称
融合将温州蜜柑(作为愈伤亲本)的线粒体基因组
(CMS 类型)转移到二倍体有籽品种中,达到无核改
良的目的[74],从而保证实现无核的同时, 大限度地
保留原有品种的优良性状。目前,世界范围内已获得
了多例转移了温州蜜柑 CMS 性状的胞质杂种[73,75],对
其核基因组和胞质基因组来源分析表明,所有胞质杂
种的核基因组来自叶肉亲本,线粒体基因组来自愈伤
亲本,叶绿体基因组则随机遗传,证明其均为转移了
温州蜜柑线粒体基因组类型的胞质杂种。胞质杂种‘国
庆 1 号温州蜜柑+HB 柚’(‘华柚 2 号’)已连续几
年开花结果,表型上其雄蕊完全退化、雌蕊正常,在
隔离种植的条件下,果实完全无籽[76]。 线粒体基因组为母系遗传,以雄性不育胞质杂种
为母本与有籽品种为父本的杂交也可能实现柑橘在二
倍体水平上的无核改良。日本‘清见’橘橙系以特罗
维塔甜橙与温州蜜柑杂交育成,其品质优良,为 CMS特性的雄性不育类型。由于其单胚性,‘清见’橘橙
作为杂交母本被广泛用于日本的柑橘无核品种培育,
并从中选育了‘Tsunokaori’和‘Shiranui’等优质、
无核的新品种[77]。‘华柚 2 号’兼具有 CMS 特性及
单胚性,可作为育种亲本用于柑橘有籽品种的无核改
良,华中农业大学正以‘华柚 2 号’为母本,以各种
有籽的柚品种为父本,进行有性杂交,以期培育系列
无核柚新品种,目前已获得近千株有性后代植株。 经过几十年的发展,中国在柑橘果实品质遗传改
良方面已取得一定进展,尤其是近年来生物技术的快
速发展和应用,加快了中国柑橘品种品质改良的步
伐。得益于细胞融合技术以及分子标记技术的应用,
提出了细胞工程用于柑橘无核品质改良的新思路,并
已获得一大批优良的柑橘育种材料,为柑橘品质改
良,尤其是无核品质改良奠定了坚实的材料基础,对
中国未来一个时期的柑橘育种和种业发展将产生重
要影响。 3.3.4 胚抢救技术 高等植物中,胚是一个具有全能
性的多细胞结构,有两种来源,一是有性来源,为合
子胚;另一个是无性来源,为体细胞胚或不定胚。在
正常情况和适宜的条件下,胚能发育成熟,并且可
以直接播种发育成完整植株。但是,在果树育种中
一些常用的育种方法如远缘杂交、二倍体与四倍体
杂交、以早熟品种作杂交母本及柑橘育种中,获得
的合子胚往往在发育的早期阶段就败育或退化。胚
早期退化对果树栽培来说是一个优良特性,能够获
得无籽果实;但对于果树育种来说,是一个不良的
特性,使育种效率降低。组织培养技术的发展,有
效地克服了这个障碍。通过胚抢救,可能败育或退
化的胚,经过适宜的培养,获得再生[78]。目前,胚
抢救在核果类早熟新品种与种质创制、无核葡萄等
方面的研究均取得了很好的进展,育成了一批新品
种,创造了一批新种质[79-83]。 (1)核果类早熟新品种及种质创制 核果类果树的早熟品种果实发育期短,胚发育不
成熟,导致常规层积播种很难成苗,胚抢救技术则能
够有效提高早熟品种的萌芽率和成苗率,为核果类早
熟以及特早熟品种育种工作的顺利开展提供了条件。
迄今为止,国内外已通过胚培技术培育出了许多毛桃、
油桃及蟠桃的优良早熟品种或品系,其中国内先后培
育出了‘京早 3 号’、‘春蕾’、‘春花’、‘早花
露’、‘早霞露’、‘玫瑰露’、‘丹墨’、‘早红
霞’、‘春秀’、‘早美’和‘云露’等品种,使中
国桃幼胚培养的研究处于国际领先水平[79,81]。 此外,以早熟品种为母本进行有性杂交,对发
育成熟的杂种胚进行胚培养是特早熟杏育种的技术
关键[83-88]。为提高杂交育种效率,陈学森等[89-90]研究
提出了果树杂交育种的“连被去雄法”,杂交效率比传
统方法提高 20 倍以上,且在杏花期发生-4℃寒流伤害
的情况下,采用连被去雄授粉后,坐果良好,已在苹
果、梨、桃、李及杏[81]等广泛应用。远缘杂交是种质
创新的有效途径之一,针对远缘杂交的不亲和性及远
缘杂种的不育性,山东农业大学陈学森课题组发明了
以胚抢救技术为核心的“利用远缘杂交创造核果类果
树新种质的三级放大法”,提高了远缘杂交育种效率,
成功实现了桃×杏及甜樱桃×欧李等核果类果树种
属间的远缘杂交,并获得 32 份远缘杂种新种质,进
一步丰富了核果类果树资源的遗传多样性[92-95]。 (2)无核葡萄育种 无核化是葡萄鲜食和制干葡萄的重要品质性状之
一,杂交育种是无核葡萄品种培育的有效途径。但传
统杂交育种中只能采用有核葡萄作母本,无核葡萄作

17 期 陈学森等:主要果树果实品质遗传改良与提升实践 3531
父本进行杂交,这种有性杂交方式选育无核葡萄的效
率很低,杂种后代中仅有 0—15.9%的无核概率[96]。
1982 年美国葡萄育种家 Ramming 等改变了传统的育
种模式,创立了无核葡萄胚挽救(Embryo rescue)技
术,以无核葡萄作母本进行杂交,极大提高了杂种后
代中的无核概率[97]。随着胚挽救技术的研究和应用,
突破了传统的只能以有核品种作母本与无核品种杂交
的育种方式的限制,使无核品种间杂交、无核品种
作母本与有核品种杂交、远缘杂交改良品种等育种
方式成为可能,极大地拓宽了葡萄育种的范围和效
率,丰富了杂交组合方式,更缩短了育种周期,推
动了无核葡萄育种的进程,国内外先后培育出了一系
列无核品种,如美国育成了‘Princess’、‘Thomcord’、‘Sweet Scarlet’和‘Scarlet Royal’等,上海农科院
育出了无核葡萄新品种‘沪培 1 号’、‘沪培 2 号’
及‘申爱’[98-104]。幼胚培养研究为其他果树如枣的杂
交育种技术体系的创建也提供了技术支撑[82,105-107]。
4 品质性状遗传倾向及芽变机理
4.1 品质性状遗传变异特点
杂交育种是果树新品种创制的有效途径,遗传研
究是育种的重要基础。由于果树作物多数自交不亲和、
杂合程度高及童期长等特点,设计专门的试验开展遗
传研究较困难。因此,根据育种记录总结性状遗传变
异趋势或倾向是果树性状遗传研究的重要特点[15]。已
有的研究表明,除苹果含酸量由一对不完全显性的主
效基因和多基因两种遗传系统共同控制外,梨、桃及
杏等果树杂交后代果实大小、颜色、糖酸、香味和酚
类物质含量等果实品质性状多数为多基因控制的数量
性状遗传[108-114]。尽管不同果树及其性状的广义遗传
力等遗传参数存在明显差异,但与 1、2 年生的大田作
物相比,果树作物遗传变异有以下 3 个共同特点:①
营养系品种间杂种一代(F1)都发生复杂的多样性分
离,而且变异幅度胜于大田作物的 F2。②苹果、梨、
柑橘等果树由显性、超显性、上位性效应在产量、果
实大小、品质等方面表现的杂种优势可以通过嫁接等
营养繁殖方式长期保持,每一次育种都选择杂种优势
强的基因型繁殖成新的营养系品种。因此,果树营
养系品种都是非加性效应比重很大的基因型。③营养
系品种杂交,由于非加性效应解体,导致果实大小等
单向选择的经济性状普遍退化(即杂种平均值都显著
低于亲中值),而果实成熟期等双向选择的经济性状
趋中变异[115]。
4.2 芽变选种
芽变选种是优中选优,通过芽变选种,对现有果
树品种着色、成熟期及株型等个别品质和农艺性状进
行了有效改良,推动了果树品种结构的优化调整与升
级换代。因此,进一步探讨芽变形成机理对果树品种
品质改良及产业发展具有重要意义。 4.2.1 苹果 现有苹果品种中,约有 30%左右来自
芽变选种,全世界苹果总产量中大约有 1/2 左右源于
芽变品种,而其中绝大多数又是红色或短枝浓红型芽
变[116-117]。陈学森等[118]、宋杨等[119]研究结果表明,短
枝浓红型苹果芽变新品种‘龙富’花后 80 d 的枝条赤
霉素含量及节间长度为 110 ng·g-1 FW 和 2.0 cm,分别
是亲本对照‘长富 2 号’的 44.2%和 76.9%,因而表
现早果性强的突出特点。进一步研究发现,两试材赤
霉素合成途径中关键酶基因 GA20-氧化酶和贝壳杉烯
氧化酶基因 cDNA 序列相同,无碱基突变、插入或
缺失现象。其相对表达量的变化趋势与赤霉素含量
的变化趋势相似。结果表明,GA20-氧化酶和贝壳
杉烯氧化酶基因在两试材中的表达量差异,是造成
其赤霉素含量变化的原因,其调控机理有待进一步
研究[120-123]。 ‘国光’苹果因其酸甜适口风味浓、类黄酮等功能
成分含量高等特点,倍受消费者的青睐与欢迎,曾是
苹果生产的主栽品种,但因果个小、着色差而影响其
进一步发展。从‘国光’苹果选育出的芽变新品种‘山
农红’果个大、着色好,将为满足消费市场的多样化
需求发挥重要作用[124]。进一步研究发现,MdMYB1表达量的升高导致花青苷合成结构基因表达水平协
同上调,进而提高了花青苷的积累,MdMYB1 启动
子区甲基化水平的差异可能是其表达上升的主要原
因[125-126]。 4.2.2 柑橘 柑橘生产中 重要的品种,如华盛顿脐
橙、伏令夏橙、葡萄柚、温州蜜柑和克里曼丁橘等系
列品种,大多数起源于芽变。柑橘为多年生果树,遗
传上高度杂合,人们对其芽变分子机理了解还有限,
现有研究主要集中在反转录转座子插入、DNA 甲基
化、基因结构和表达差异等方面[46]。近年来,柑橘中
发现了相当数量的可转座因子的存在,为其容易出现
芽变的现象提供了部分解释。转座子(transposable element)在基因组中的转移,可造成基因失活或改
变附近基因的表达活性,产生原有基因的新等位基
因[127]。Asins 等[128]研究表明,柑橘容易产生变异的
类型,如温州蜜柑和脐橙均比芽变较少的类型在反转

3532 中 国 农 业 科 学 48 卷
座子胞嘧啶甲基化程度上有差异;而且柑橘基因组
中,有很多 copia-like 反转录转座子的存在。利用
copia-like 反转录酶(RT)探针进行 Southern 杂交表
明由于反转录转座子的插入,导致从普通甜橙产生脐
橙类芽变。该研究还发现,柑橘比桃含有较多的反转
录转座子拷贝,这在某种程度上解释了柑橘芽变比桃
要多的现象。
5 育成品种亲本溯源分析
杂交育种中,亲本选择与选配是决定品质改良成
效大小的关键。因此,认真总结分析果树育成品种的
亲本溯源或遗传背景,对今后主要果树果实品质改良
意义重大。郑州果树研究所培育的‘华冠’苹果,之
所以综合品质性状优良、被新西兰确认为是一个具有
国际推广价值的品种,其亲本组合是‘金冠’ב富
士’(‘国光’ב元帅’)。新西兰育成的品种‘太
平洋玫瑰’(Pacific Rose)是综合性状优良的中晚熟
苹果品种,被列为“21 世纪世界五大品种之一”,其
亲本组合为‘嘎拉’ב华丽’,而‘嘎啦’的亲本
为‘红基橙’(‘元帅’ב桔苹’)ב金冠’,
‘华丽’的亲本为‘金冠’ב惠’(‘国光’ב红
玉’)。因此,‘太平洋玫瑰’具有了‘金冠’‘国
光’‘红玉’‘元帅’和‘桔苹’5 个品种的血统,
遗传背景宽[12]。新西兰为了拓宽苹果品种的遗传基
础,从 1990 年后采用开放授粉系统下的实生选种方法
代替有控制的杂交育种,其供选群体是来自遗传背景
差别大、具有多个不同血统的 350 个品种或类型(其
中包括来自哈萨克斯坦的新疆野苹果 Malus sieversii 株系 55 个)自然授粉的实生后代[129]。陈学森等[15]提
出并实施的功能型苹果育种计划,其主要目的之一也
是为了拓宽栽培苹果品种的遗传基础。 ‘莱阳慈’梨和‘ 山酥’梨是中国白梨系统
(Pyrus bretschneideri Rehd.)的 2 个著名传统地方
名产品种,但均存在一定的缺陷或不足,有待进一步
改良。为此,近期育成并暂定名为‘新慈香’梨(‘新
梨 7 号’ב莱阳慈’梨)和‘山农酥’梨(‘新梨
7 号’ב 山酥梨’)的 2 个新品系,综合经济或
品质性状优良,很有发展前景。其中,‘新梨 7 号’
亲本组合是‘库尔勒香梨’(新疆梨,P. Sinkiangensis Yu)ב早酥梨’,‘早酥梨’亲本是‘苹果梨’(沙
梨,P. pyrifolia Burm.)ב身不知’,经考证,‘身
不知’是西洋梨(P. communis L.)与白梨(P. bretschneideri Rehd.)的杂种后代。因此,‘新慈香’
梨和‘山农酥’梨均含有梨属(Pyrus)新疆梨、沙梨、
西洋梨和白梨等 4 个种的血缘(数据未提供)。上述
研究结果表明,复杂的遗传背景和丰富的遗传多样性
是杂交亲本选择进行品质育种的关键。 中国桃育种取得了重要进展,育成品种外观和内
在品质显著提高,类型趋于多样化,果实成熟期显著
提前,栽培适宜区域有效扩大。但中国桃育种整体水
平落后世界桃育种强国约 20 年,面临诸多严峻挑战,
其中中国育成品种的亲本品种是‘白花’‘大久保’
‘早生黄金’和‘兴津油桃’,而这 4 个品种均来自
上海水蜜,遗传背景非常狭窄,这正是造成中国桃育
种不能取得有效突破的根本因素之一[17]。因此,扩大
遗传基础或背景应该成为水果果实品质改良的一个共
识。
6 结束语
100 多年来,中国苹果、柑橘、桃、梨及葡萄等
主要果树果实品质改良取得了巨大成就,为推动果树
产业高效发展和农民持续增收提供了优质品种支撑。
但与世界果树果实品质改良先进国家相比,仍存在较
大差距。为此,提出 4 点建议: (1)必须依据《种子法》,进一步加强果树苗木
繁育市场的监管力度,彻底改变目前果树苗木乱繁乱
育的现状,建立有效运行的品种权保护与品种权转让
制度,依法保护育种者、育苗者及栽培者的利益,推
动中国果树产业的有序、有力和高效发展。 (2)进一步加强野生果树资源评价及亲本利用研
究,努力拓宽栽培品种的遗传基础,协同提升品质与
抗性。 (3)深入开展原生质体融合及胚抢救等生物技术
以及芽变机理和性状遗传变异规律研究,建立生物技
术与常规技术结合、杂交育种与芽变选种优势互补的
果树果实品质高效改良技术体系。 (4)加强新品系的国际品种区域栽培试验及国际
品种权申请与保护,培育国际性的果树“大品种”。
References
[1] 吴玉森, 张艳敏, 冀晓昊, 张芮, 刘大亮, 张宗营, 李文燕, 陈学
森. 自然生草对黄河三角洲梨园土壤养分、酶活性及果实品质的影
响. 中国农业科学, 2013, 46(1): 99-108.
Wu Y S, Zhang Y M, Ji X H, Zhang R, Liu D L, Zhang Z Y, Li W Y,
Chen X S. Effects of natural grass on soil nutrient, enzyme activity
and fruit quality of pear orchard in Yellow River delta. Scientia

17 期 陈学森等:主要果树果实品质遗传改良与提升实践 3533
Agricultura Sinica, 2013, 46(1): 99-108. (in Chinese)
[2] 王艳廷, 冀晓昊, 张艳敏, 吴玉森, 安萌萌, 张芮, 王立霞, 张晶,
刘文, 李敏, 李文燕, 陈学森. 自然生草对黄河三角洲梨园土壤物
理性状及微生物多样性影响. 生态学报, 2015, 35(16): 1-13.
Wang Y T, Ji X H, Zhang Y M, Wu Y S, An M M, Zhang R, Wang L X,
Zhang J, Liu W, Li M, Li W Y, Chen X S. Effects of self-sown grass
on soil physical properties and microbial diversity of pear rchard in
Yellow River delta. Acta Ecologica Sinica, 2015, 35(16): 1-13. (in
Chinese)
[3] 王艳廷, 冀晓昊, 吴玉森, 张艳敏, 安萌萌, 毛志泉, 姜远茂, 彭福
田, 王志强, 陈学森. 我国果园生草研究进展与建议. 应用生态学
报, 2015, 26(6): 1892-1900.
Wang Y T, Ji X H, Wu Y S, Zhang Y M, An M M, Mao Z Q, Jiang Y
M, Peng F T, Wang Z Q, Chen X S. Research progress and
suggestions of cover crop in Chinese orchard. Chinese Journal of
Applied Ecology, 2015, 26(6): 1892-1900. (in Chinese)
[4] 陈学森, 高东升, 辛力, 原永兵, 刘成连, 李文燕. 给果园小草一
点阳光, 苹果更灿烂——苹果果实品质提升的途径与关键技术.
落叶果树, 2012, 44(3): 1-4.
Chen X S, Gao D S, Xin L, Yuan Y B, Liu C L, Li W Y. If given some
sunshine to the grass of orchard, apple is more brilliant-the ways and
key technologies to enhance the fruit quality of apple. Deciduous
Fruits, 2012, 44(3): 1-4. (in Chinese)
[5] 陈学森, 苏桂林, 姜远茂, 毛志泉. 可持续发展果园的经营与管理
—再谈果园生草培肥地力及其配套技术. 落叶果树, 2013, 45(1):
1-3.
Chen X S, Su G L, Jiang Y M, Mao Z Q. Management and
Administration of the Sustainable development orchard-talk about the
orchard grass soil fertility and its supporting technology. Deciduous
Fruits, 2013, 45(1): 1-3. (in Chinese)
[6] 陈学森 , 王艳廷 . 力推果园生草 , 建设生态文明 . 烟台果树 ,
2014(1): 1-4.
Chen X S, Wang Y T. Promoting orchard grass, building ecological
civilization. Yantai Fruits, 2014(1): 1-4. (in Chinese)
[7] King G J, Maliepaard C, Lynn J R, Alston F H, Durel C E, Evans K M,
Griffon B, Laurens F, Manganaris A G, Schrevens E, Tartarini S,
Verhaegh J. Quantitative genetic analysis and comparison of
physicaland sensory descriptors relating to fruit flesh firmness in
apple (Malus pumila Mill.). Theoretical and Applied Genetics, 2000,
100: 1074-1084.
[8] 陈学森, 宋君, 高利平, 冀晓昊, 张宗营, 毛志泉, 张艳敏, 刘大亮,
张芮, 李敏. ‘乔纳金’苹果及其脆肉芽变果实质地发育机理. 中国
农业科学, 2014, 47(4): 727-735.
Chen X S, Song J, Gao L P, Ji X H, Zhang Z Y, Mao Z Q, Zhang Y M,
Liu D L, Zhang R, Li M. Developing mechanism of fruits texture in
‘Jonagold’ apple and its crisp flesh sport. Scientia Agricultura Sinica,
2014, 47(4): 727-735. (in Chinese)
[9] Kayesh E, Shangguan L F, Korir N K, Sun Xi, Bilkish N, Zhang Y P,
Han J, Song C N, Cheng Z M, Fang J G. Fruit skin color and the role
of anthocyanin. Acta Physiology of Plant, 2013, 35: 2879-2890.
[10] Czemmel S, Heppel S C, Bogs J. R2R3 MYB transcription factors:
key regulators of the flavonoid biosynthetic pathway in grapevine.
Protoplasma, 2012, 249(Suppl. 2): S109-S118.
[11] 陈学森, 韩明玉, 苏桂林, 刘凤之, 过国南, 姜远茂, 毛志泉, 彭福
田, 束怀瑞. 当今世界苹果产业发展趋势及我国苹果产业优质高
效发展意见. 果树学报, 2010, 27(4): 598-604.
Chen X S, Han M Y, Su G L, Liu F Z, Guo G N, Jiang Y M, Mao Z Q,
Peng F T, Shu H R. Discussion on today’s world apple industry trends
and the suggestions on sustainable and efficient development of apple
industry in China. Journal of Fruit Science, 2010, 27(4): 598-604. (in
Chinese)
[12] 王宇霖. 关于我国苹果育种研究工作的几点想法. 果树学报, 2008,
25(3): 559-565.
Wang Y L. Some ideas about the apple br eeding in China. Journal of
Fruit Science, 2008, 25(3): 559-565. (in Chinese)
[13] 陈学森, 毛志泉, 姜远茂, 王志刚. 山东苹果产业转型升级的建议
—一靠政策,二靠科技,创新驱动,转型升级,提质增效. 落叶果树,
2015, 47(1): 2-4.
Chen X S, Mao Z Q, Jiang Y M, Wang Z G. Suggestions about apple's
industrial transformation and upgrading of Shandong-one by the
policy, and on science and technology, innovation-driven, transformation
and upgrading, quality and efficiency. Deciduous Fruits, 2015, 47(1):
2-4. (in Chinese)
[14] Boyer J, Liu R H. Apple phytochemicals and their health benefits.
Nutrition Journal, 2004, 3: 1-15.
[15] 陈学森, 张晶, 刘大亮, 冀晓昊, 张宗营, 张芮, 毛志泉, 张艳敏,
王立霞, 李敏. 新疆红肉苹果杂种一代的遗传变异及功能型苹果
优株评价. 中国农业科学, 2014, 47(11): 2193-2204.
Chen X S, Zhang J, Liu D L, Ji X H, Zhang Z Y, Zhang R, Mao Z Q,
Zhang Y M, Wang L X, Li M. Genetic variation of F1 population
between Malus sieversii f. neidzwetzkyana and apple varieties and
evaluation on fruit characters of functional apple excellent strains.
Scientia Agricultura Sinica, 2014, 47(11): 2193-2204. (in Chinese)
[16] 邓秀新 . 世界柑橘品种改良的进展 . 园艺学报 , 2005, 32(6):
1140-1146.
Deng X X. Advances in worldwide citrus breeding. Acta Horticulturae

3534 中 国 农 业 科 学 48 卷
Sinica, 2005, 32(6): 1140-1146. (in Chinese)
[17] 王力荣, 朱更瑞, 方伟超. 中国桃遗传资源. 北京: 中国农业出版
社, 2012.
Wang L R, Zhu G R, Fang W C. Peach Genetic Resources in China.
Beijing: China Agriculture Press, 2012. (in Chinese)
[18] 樊秀彩, 张颖, 姜建福, 孙海生, 李民, 刘崇怀. 近 20 年来国外鲜
食葡萄品种选育进展. 中外葡萄与葡萄酒, 2012(2): 53-59.
Fan X C, Zhang Y, Jiang J F, Sun H S, Li M, Liu C H. Foreign grape
varieties breeding In the recent 20 years. Sino-Overseas Grapevine &
Wine, 2012(2): 53-59. (in Chinese)
[19] 李秀根, 杨健, 王龙, 王苏珂. 我国近 30a 梨育种研究进展与今后
工作建议. 果树学报, 2010, 27(6): 987-994.
Li X G, Yang J, Wang L, Wang S K. Progress and proposal in research
of pear breeding in China. Journal of Fruit Science, 2010, 27(6):
987-994. (in Chinese)
[20] 杨庆文, 秦文斌, 张万霞, 乔卫华, 于寿娜, 郭青. 中国农业野生
植物原生境保护实践与未来研究方向. 植物遗传资源学报, 2013,
14(1): 1-7.
Yang Q W, Qin W B, Zhang W X, Qiao Wei H, Yu S N, Guo Q. In-situ
conservation practices and future development of wild relatives of
crops in China. Journal of Plant Genetic Resources, 2013, 14(1): 1-7.
(in Chinese)
[21] 冯涛, 张红, 陈学森, 张艳敏, 何天明, 冯建荣, 许正. 新疆野苹果
果实形态与矿质元素含量多样性以及特异性状单株. 植物遗传资
源学报, 2006, 7(3): 270-276.
Feng T, Zhang H, Chen X S, Zhang Y M, He T M, Feng J R, Xu Z.
Genetic diversity of fruit morphological traits and content of mineral
element in Malus sieversii (Ldb.) Roem. and its elite seedlings.
Journal of Plant Genet Research, 2006, 7(3): 270-276. (in Chinese)
[22] 冯涛, 张艳敏, 陈学森. 新疆野苹果居群年龄结构及郁闭度研究.
果树学报, 2007, 24(5): 571-573.
Feng T, Zhang Y M, Chen X S. Study on the age structure and density
of the wild apple forest of Malus sieversii. Journal of Fruit Science,
2007, 24(5): 571-573. (in Chinese)
[23] 张春雨, 陈学森, 林群, 苑兆和, 张红, 张小燕, 刘崇祺, 吴传金.
新疆野苹果群体遗传结构和遗传多样性的 SRAP 分析. 园艺学报,
2009, 36(1): 7-14.
Zhang C Y, Chen X S, Lin Q, Yuan Z H , Zhang H, Zhang X Y, Liu C
Q, Wu C J.SRAP Markers for Popula tion Genetic Structure and
Genetic Diversity in Malus sieversii from Xinjiang, China.Acta
Horticulturae Sinica, 2009, 36(1): 7-14. (in Chinese)
[24] Zhang C Y, Chen X S, He T M, Liu X L, Feng T, Yuan Z H. Genetic
structure of Malus sieversii population from Xinjiang, China, revealed
by SSR markers. Journal of Genetics and Genomics, 2007, 34(10):
947-955.
[25] 吴传金, 陈学森, 曾继吾, 易干军, 刘崇琪, 张大海. 新疆野苹果
(Malus sieversii)超低温保存与植株再生. 植物遗传资源学报, 2008,
8(2): 243-247.
Wu C J, Chen X S, Zeng J W, Yi G J, Liu C Q, Zhang D H.
Cryopreservation of in vitro shoot tips of Malus sieversii by
vitrification and its regeneration. Journal of Plant Genetic Resources,
2008, 8(2): 243-247. (in Chinese)
[26] 刘遵春, 张春雨, 张艳敏, 张小燕, 吴传金, 王海波, 石俊, 陈学森.
利用数量性状构建新疆野苹果核心种质方法研究. 中国农业科学,
2010, 43(2): 358-370.
Liu Z C, Zhang C Y, Zhang Y M, Zhang X Y, Wu C J, Wang H P, Shi J,
Chen X S. Study on method of constructing core collection of Malus
sieversii based on quantitative traits. Scientia Agricultura Sinica, 2010,
43(2): 358-370. (in Chinese)
[27] 刘遵春, 刘大亮, 崔美, 李敏, 焦其庆, 高利平, 陈学森. 整合农艺
性状和分子标记数据构建新疆野苹果核心种质. 园艺学报, 2012,
39(6): 1045-1054.
Liu Z C, Liu D L, Cui M, Li M, Jiao Q Q, Gao L P, Chen X S.
Combining agronomic traits and molecular marker data for constructing
Malus sieversii core collection. Acta Horticulturae Sinica, 2012, 39(6):
1045-1054. (in Chinese)
[28] 张春雨, 陈学森, 张艳敏, 苑兆和, 刘遵春, 王延龄. 采用分子标
记构建新疆野苹果核心种质的方法. 中国农业科学, 2009, 42(2):
597-604.
Zhang C Y, Chen X S, Zhang Y M, Yuan Z H, Liu Z C, Wang Y L. A
method for constructing core collection of Malus sieversii using
molecular markers. Scientia Agricultura Sinica, 2009, 42(2): 597-604.
(in Chinese)
[29] 王立霞, 冀晓昊, 安萌萌, 张宗营, 王艳廷, 王传增, 吴玉森, 吴树
敬, 陈学森. 几个功能型苹果优株果实风味品质评价. 果树学报,
2014, 31(5): 753-759.
Wang L X, Ji X H, An M M, Zhang Z Y, Wang Y T, Wang C Z, Wu Y
S, Wu S J, Chen X S. The evaluation on apple fruit flavor quality of
several functional superior apple strains. Journal of Fruit Science,
2014, 31(5): 753-759. (in Chinese)
[30] 刘崇琪, 陈学森, 王金政, 陈晓流, 王海波, 田长平, 吴传金. 新疆
野生樱桃李(Prunus cerasifera Ehrh.)部分表型性状的遗传多样性.
园艺学报, 2008, 35(9): 1261-1268.
Liu C Q, Chen X S, Wang J Z, Chen X L, Wang H B, Tian C P, Wu C
J. Studies on genetic diversity of phenotypic traits in wild myrobalan
plum (Prunus cerasifera Ehrh.). Acta Horticulturae Sinica, 2008,

17 期 陈学森等:主要果树果实品质遗传改良与提升实践 3535
35(9): 1261-1268. (in Chinese)
[31] 刘崇琪, 陈晓流, 张艳敏, 林群, 王金政, 张红, 张春雨, 陈学森.
新疆野生樱桃李 S-RNase 基因分离与鉴定的初步研究. 园艺学报,
2009, 36(3): 333-340.
Liu C Q, Chen X L, Zhang Y M, Lin Q, Wang J Z, Zhang H, Zhang C
Y, Chen X S. Primary study on identification of S-RNase genes in
wild myrobalan plum (Prunuscerasifera Ehrh.). Acta Horticulturae
Sinica, 2009, 36(3): 333-340.
[32] Wang Y, Chen X L, Zhang Y M, Chen X S, Antioxidant activities and
major anthocyanins of myrobalan plum (Prunus cerasifera Ehrh.).
Journal of Food Science, 2012, 77(4): 388-393.
[33] 周龙, 廖康, 许正, 王磊. 新疆野生樱桃李种质资源研究进展. 经
济林研究, 2010, 28(2): 142-145.
Zhou L, Liao K, Xu Z, Wang L. Advance in research on germplasm
resources in Prunus cerasifera Ehrh. Nonwood Forest Research. 2010,
28(2): 142-145. (in Chinese)
[34] 冀晓昊, 毛志泉, 张芮, 匡林光, 王燕, 张艳敏, 陈学森. 樱桃李新
品种‘森果红露’. 园艺学报, 2012, 39(4): 797-798.
Ji X H, Mao Z Q, Zhang R, Kuang L G, Wang Y, Zhang Y M, Chen X
S. A new myrobalan plum cultivar ‘Senguo Honglu’. Acta
Horticulturae Sinica, 2012, 39(4): 797-798. (in Chinese)
[35] 张芮, 毛志泉, 冀晓昊, 匡林光, 鹿明芳, 王燕, 陈学森. 樱桃李鲜
食新品种‘森果佳人’. 园艺学报, 2012, 39(5): 999-1000.
Zhang R, Mao Z Q, Ji X H, Kuang L G, Lu M F, Wang Y, Chen X S. A
new myrobalan plum cultivar ‘Senguo Jiaren’. Acta Horticulturae
Sinica, 2012, 39(5): 999-1000. (in Chinese)
[36] 冀晓昊, 张芮, 毛志泉, 匡林光, 鹿明芳, 王燕, 张艳敏, 陈学森.
新疆野生樱桃李实生后代遗传变异及优异种质挖掘. 园艺学报,
2012, 39(8): 1551-1558.
Ji X H, Zhang R, Mao Z Q, Kuang L G, Lu M F, Wang Y, Zhang Y M,
Chen X S. The analysis of characteristic variations of the seedlings of
Xinjiang wild myrobalan plum and excavation of the excellent
germplasm resources. Acta Horticulturae Sinica, 2012, 39(8):
1551-1558. (in Chinese)
[37] Huang X H, Kurata N, Wei X H, Wang Z X, Wang A H, Zhao Q, Zhao
Y, Liu K Y, Lu H Y, Li W J, Guo Y L, Lu Y Q, Zhou C C, Fan D L,
Weng Q J, Zhu C R, Huang T, Zhang L,Wang Y C, Feng L, Furuumi
H, Kubo T, Miyabayashi T, Yuan X P, Xu Q, Dong G J, Zhan Q L, Li
CY, Fujiyama A, Toyoda A, Lu T T, Feng Q, Qian Q, Li J Y, Han B f.
A map of rice genome variation reveals the origin of cultivated rice.
Nature, 2012, 490: 497-501.
[38] 杨庆文, 黄娟. 中国普通野生稻遗传多样性研究进展. 作物学报,
2013, 39(4): 580-588.
Yang Q W, Huang J. Research progress on genetic diversity of oryza
rufipogon in China. Acta Agricultura Sinica, 2013, 39(4): 580-588. (in
Chinese)
[39] 范虎, 赵团结, 丁艳来, 邢光南, 盖钧镒. 中国野生大豆群体特征
和地理分化的遗传分析. 中国农业科学, 2012, 45(3): 414-425.
Fan H, Zhao T J, Ding Y L, Xing G N, Gai J Y. Genetic analysis of the
characteristics and geographic differentiation of Chinese wild soybean
population. Scientia Agricultura Sinica, 2012, 45(3): 414-425. (in
Chinese)
[40] 安萌萌, 刘畅, 何天明, 张艳敏, 冀晓昊, 王艳廷, 李敏, 王传增,
陈学森. 黑龙江省野生秋子梨群体遗传结构的荧光 AFLP 分析. 植
物遗传资源学报, 2014, 15(4): 728-733.
An M M, Liu C, He T M, Zhang Y M, Ji X H, Wang Y T, Li M, Wang
C Z, Chen X S. Genetic structure analyses based on fluorescent-AFLP
markers of wild populations of Pyrus ussuriensis Maxim in
Heilongjiang province. Journal of Plant Genetic Resources 2014,
15(4): 728-733. (in Chinese)
[41] 安萌萌, 王艳廷, 宋杨, 冀晓昊, 刘畅, 王楠, 吴玉森, 刘文, 曹玉
芬, 冯守千, 陈学森. 野生秋子梨(Pyrus ussuriensis Maxim)果实
性状的遗传多样性. 中国农业科学, 2014, 47(15): 3034-3043.
An M M, Wang Y T, Song Y, Ji X H, Liu C, Wang N,Wu Y S, Liu W,
Cao Y F, Feng S Q, Chen X S. Genetic diversity of fruit phenotypic
traits of wild Pyrus ussuriensis Maxim. Scientia Agricultura Sinica,
2014, 47(15): 3034-3043. (in Chinese)
[42] 张小燕, 陈学森, 彭勇, 王海波, 石俊, 张红. 新疆野苹果酚类物
质组分的遗传多样性. 园艺学报, 2008, 35(9): 1351-1356.
Zhang X Y, Chen X S, Peng Y, Wang H B, Shi J, Zhang H. Genetic
diversity of phenolic compounds in Malus sieversii. Acta
Horticulturae Sinica, 2008, 35(9): 1351-1356. (in Chinese)
[43] 张小燕, 陈学森, 彭勇, 王海波, 石俊, 张红. 新疆野苹果矿质元
素与糖酸组分的遗传多样性. 园艺学报, 2008, 35(2): 277-280.
Zhang X Y, Chen X S, Peng Y, Wang H B, Shi J, Zhang H. Genetic
diversity of mineral elements, sugar and acid components in Malus
sieversii (Ldb.) Roem. Acta Horticulturae Sinica, 2008, 35(2):
277-280. (in Chinese)
[44] 刘遵春, 苗卫东, 刘大亮, 陈学森. 新疆野苹果分离群体的构建和
评价. 果树学报, 2012, 29(5): 722-728.
Liu Z C, Miao W D, Liu D L, Chen X S. Construction and evaluation
of the segregation population in Malus sieversii. Journal of Fruit
Science, 2012, 29(5): 722-728. ( in Chinese)
[45] 高利平, 冀晓昊, 张艳敏, 宋君, 李敏, 刘大亮, 张芮, 陈学森. 新
疆野苹果杂种 F1代部分株系绵/脆肉发育机理的初步研究. 园艺学
报, 2013, 40(6): 1153-1158.

3536 中 国 农 业 科 学 48 卷
Gao L P, Ji X H, Zhang Y M, Song J, Li M, Liu D L, Zhang R, Chen
X S. The preliminary study on the enzymes activity related to fruit
texture of the fruit of soft/crisp strains from the cross progenies of
‘Fuji’ and Malus sieversii. Acta Horticulturae Sinica, 2013, 40(6):
1153-1158. (in Chinese)
[46] 张敏, 邓秀新. 柑橘芽变选种以及芽变性状形成机理研究进展. 果
树学报, 2006, 23(6): 871-876.
Zhang M, Deng X X. Advances in r esear ch of citrus cultivar s
selected by bud mutation and the mechanism of formation of mutated
char acter istics. Journal of Fruit Science, 2006, 23(6): 871-876. (in
Chinese)
[47] 过国南, 阎振立, 张顺妮. 我国建国以来苹果品种选育研究的回顾
及今后育种的发展方向. 果树学报, 2003, 20(2): 127-134.
Guo G B, Yan Z L, Zhang S N. Review and outlook of apple breeding
in China. Journal of Fruit Science, 2003, 20(2): 127-134. (in Chinese)
[48] 陈学森, 辛培刚, 杜欣阁, 杨传友, 温吉华. 元帅和金帅在苹果新
品种选育中的作用. 山东农业大学学报, 1994, 25(2): 236-248.
Chen X S, Xin P G, Du X G, Yang C Y, Wen J H. Effect of delicious
and golden delicious on the breeding and selection of new apple
varieties. Journal of Shandong Agricultural University, 1994, 25(2):
236-248. (in Chinese)
[49] 河北省农林科学院昌黎果树研究所. 河北苹果志. 北京: 中国农业
出版社, 1986.
Changli Institute of Fruit Science, the Hebei Academy of agricultural
and Forestry Sciences. Apples of Hebei. Beijing: China Agriculture
Press, 1986. (in Chinese)
[50] 曹玉芬. 中国梨品种. 北京: 中国农业出版社, 2014.
Cao Y F. Pear Varieties in China. Beijing: China Agriculture Press,
2014. (in Chinese)
[51] 张加延, 张钊. 中国果树志·杏卷. 北京: 中国林业出版社, 2003.
Zhang J Y, Zhang Z. China Fruit Zhi·Volume of Apricot. Beijing:
China Forestry Press, 2003. (in Chinese)
[52] Chen X S, Feng T, Zhang Y M, He T M, Feng J R, Zhang C Y. Genetic
diversity of volatile components in Xinjiang wild apple (Malus
sieversii). Journal of Genetics and Genomics, 2007, 34(2): 171-179.
[53] 张艳敏, 冯涛, 张春雨, 何天明, 张小燕, 刘遵春, 王艳玲, 束怀瑞,
陈学森. 新疆野苹果研究进展. 园艺学报, 2009, 36(3): 447-452.
Zhang Y M, Feng T, Zhang C Y, He T M, Zhang X Y, Liu Z C, Wang
Y L, Shu H R, Chen X S. Advances in research of the Malus sieversii
(Lebed.) Roem. Acta Horticulturae Sinica, 2009, 36(3): 447-452. (in
Chinese)
[54] 乔鑫, 李梦, 殷豪, 李雷廷, 吴俊, 张绍铃. 果树全基因组测序研
究进展, 园艺学报, 2014, 41(1): 165-177.
Qiao X, Li M, Yin H, Li L T, Wu J, Zhang S L. Advances on whole
genome sequencing in fruit trees. Acta Horticulturae Sinica, 2014,
41(1): 165-177. (in Chinese)
[55] Troggio M, Gleave A, Salvi S, Chagné D, Cestaro A, Kumar S,
Crowhurst R N, Gardiner S E. Apple, from genome to breeding. Tree
Genetics & Genomes, 2012, 8: 509-529.
[56] Iwata H, Hayashi T, Terakami S, Takada N, Sawamura Y, Yamamoto T.
Potential assessment of genome-wide association study and genomic
selection in Japanese pear Pyrus pyrifolia. Breeding Science, 2013, 63:
125-140.
[57] Dennis J Graya, Zhijian T Li , Sadanand A Dhekney. Precision
breeding of grapevine (Vitis vinifera L.) for improved traits. Plant
Science, 2014, 228: 3-10.
[58] Pena L, Martin-Trillo M, Juarez J, Pina J A, Navarro L, Martinez-
Zapater J M. Constitutive expression of Arabidopsis LEAFY or
APETALA1 genes in citrus reduces their generation time. Nature
Biotechnology, 2001, 19: 263-267.
[59] Flachowsky H, Peil A, Sopanen T, Elo A, Hanke V. Overexpression
of BpMADS4 from silver birch (Betula pendula Roth.) induces
early-flowering in apple (Malus domestica Borkh.). Plant Breed, 2007,
126: 137-145.
[60] Flachowsky H, Le Roux P M, Peil A, Patocchi A, Richter K, Hanke
M V. Application of a high-speed breeding technology to apple
(Malus domestica) based on transgenic early flowering plants and
marker-assisted selection. New Phytology, 2011, 192: 364-377.
[61] Weig K, Wenze S, Flachowsky H, Peil A, Hanke M V. Integration of
BpMADS4 on various linkage groups improves the utilization of the
rapid cycle breeding system in apple. Plant Biotechnology Journal,
2015, 13: 246-258.
[62] Yamagishi N, Kishigami R, Yoshikawa N. Reduced generation time
of apple seedlings to within ayear by means of a plant virus vector: a
new plant-breeding technique with no transmission of geneticmodification
to the next generation. Plant Biotechnology Journal, 2014, 12: 60-68.
[63] Grosser J W, Gmitter F G Jr. Protoplast fusion for production of
tetraploids and triploids: applications for scion and rootstock breeding
in citrus. Plant Cell, Tissue and Organ Culture, 2011, 104(3):
343-357.
[64] Grosser J W, Ollitrault P, Olivares-Fuster O. Invited review: Somatic
hybridization in citrus: an effective tool to facilitate variety
improvement. In Vitro Cellular & Developmental Biology of Plant,
2000, 36(6): 434-449.
[65] Chen C X, Cancalon P, Haun C, Gmitter F G J. Characterization of
furanocoumarin profile and inheritance toward selection of low

17 期 陈学森等:主要果树果实品质遗传改良与提升实践 3537
furanocoumarin seedless grapefruit cultivars. Journal of the American
Society for Horticultural Science, 2011, 136(5): 358-363.
[66] 伊华林, 邓秀新. 培养三倍体柑橘植株的研究. 果树科学, 1998,
15(3): 212-216.
Yi H L, Deng X X. A study of culture of citrus triploid plantlets.
Journal of Fruit Science, 1998, 15(3): 212-216. (in Chinese)
[67] 宋健坤, 郭文武, 伊华林, 刘继红, 陈春丽, 邓秀新. 以异源四倍
体体细胞杂种为父本与二倍体杂交创造柑橘三倍体的研究. 园艺
学报, 2005, 30(4): 594-598.
Song J K, Guo W W, Yi H L, Liu J H, Chen C L, Deng X X. Creation
of triploid citrus plants by crossing elite allotetraploid somatic hybrid
pollen parents with diploid cultivars. Acta Horticulturae Sinica, 2005,
30(4): 594-598. (in Chinese)
[68] 解凯东 , 王惠芹 , 王晓培 , 梁武军 , 谢宗周 , 伊华林 , 邓秀新 ,
Grosser J W, 郭文武. 单胚性二倍体为母本与异源四倍体杂交大
规模创制柑橘三倍体. 中国农业科学, 2013, 46(21): 4550-4557.
Xie K D, Wang H Q, Wang X P, Liang W J, Xie Z Z, Yi H L, Deng X
X, Grosser J W, Guo W W. Extensive citrus triploid breeding by
crossing monoembryonic diploid females with allotetraploid male
parents. Scientia Agricultura Sinica, 2013, 46(21): 4550-4557. (in
Chinese)
[69] 解凯东, 王晓培, 王惠芹, 梁武军, 谢宗周, 郭大勇, 伊华林, 邓秀
新, Grosser J W, 郭文武. 以柑橘多胚性二倍体母本倍性杂交培育
三倍体, 园艺学报, 2014, 41(4): 613-620.
Xie K D, Wang X P, Wang H Q, Liang W J, Xie Z Z, Guo D Y, Yi H L,
Deng X X, Grosser J W, Guo W W. High efficient and extensive
production of triploid citrus plants by crossing polyembryonic
diploids with tetraploids. Acta Horticulturae Sinica, 2014, 41(4):
613-620. (in Chinese)
[70] Aleza P, Juárez J, Cuenca J, Ollitrault P, Navarro L. Extensive citrus
triploid hybrid production by 2x × 4x sexual hybridizations and
parent-effect on the length of the juvenile phase. Plant Cell Reports,
2012, 31: 1723-1735.
[71] Recupero G R, Russo G, Recupero S. New promising citrus triploid
hybrids selected from crosses between monoembryonic diploid female
and tetraploid male parents. HortScience, 2005, 40(3): 516-520.
[72] Aleza P, Juárez J, Cuenca J, Ollitrault P, Navarro L. Recovery of citrus
triploid hybrids by embryo rescue and flow cytometry from 2x × 2x
sexual hybridisation and its application to extensive breeding
programs. Plant Cell Reports, 2010, 29(9): 1023-1034.
[73] Guo W W, Xiao S X, Deng X X. Somatic cybrid production via
protoplast fusion for citrus improvement. Scientia Horticulturae, 2013,
163: 20-26.
[74] Guo W W, Prasad D, Cheng Y J, Serrano P, Deng X X, Grosser J W.
Targeted cybridization in citrus: transfer to Satsuma cytoplasm to
seedy cultivars for potential seedlessness. Plant Cell Reports, 2004,
22: 752-758.
[75] Xiao S X, Biswas M K, Li M Y, Deng X X, Xu Q, Guo W W.
Production and molecular characterization of diploid and tetraploid
somatic cybrid plants between male sterile Satsuma mandarin and
seedy sweet orange cultivars. Plant Cell Tissue and Organ Culture,
2014, 116: 81-88.
[76] Zheng B B, Wu X M, Ge X X, Deng X X, Grosser J W, Guo W W.
Comparative transcript profiling of a male sterile cybrid pummel and
its fertile type revealed altered gene expression related to flower
development. Plos One, 7(8): e43758.
[77] Yoshida T. Breeding of citrus cultivar ‘Kiyomi’ with high quality and
monoembryony. Breeding Research (Japan), 2003, 5: 103-107.
[78] 伊华林, 邓秀新, 付春华. 胚抢救技术在果树上的应用. 果树学报,
2001, 18(4): 224-228.
Yi H L, Deng X X, Fu C H. Application of embryo rescue techniques
in fruit crops. Journal of Fruit Science, 2001, 18(4): 224-228. (in
Chinese)
[79] 庄恩及, 徐祝英, 吴鈺良, 蔡奚平. 特早熟桃的育种. 园艺学报,
1982, 9(3): 1-7.
Zhuang E J, Xu Z Y, Wu Y L, Cai X P. The breeding of very early
ripening peaches. Acta Horticulturae Sinica, 1982, 9(3): 1-7. (in
Chinese)
[80] 王玉柱, 孙浩元, 杨丽. 核果类果树胚培养研究进展和育种成效.
果树学报, 2004, 21(1): 59-63.
Wang Y Z, Sun H Y, Yang L. Progress of Embryo Culture Research in
Stone Fruit Breding. Journal of Fruit Science, 2004, 21(1): 59-63. (in
Chinese)
[81] 梁青, 陈学森, 刘文, 吴燕. 胚抢救在果树育种上的研究及应用.
园艺学报, 2006, 33(2): 445-452.
Liang Q, Chen X S, Liu W, Wu Y. Research and application of embryo
rescue techniques in fruit tree breeding. Acta Horticulturae Sinica,
2006, 33(2): 445-452. (in Chinese)
[82] 任海燕, 杜学梅, 王玉国, 王永康, 薛晓芳, 赵爱玲, 隋串玲, 李登
科, 赵武娟. 枣胚培养研究进展. 果树学报, 2014, 31(5): 967-972.
Ren H Y, Du X M, Wang Y G, Wang Y K, Xue X F, Zhao A L, Sui C L,
Li D K, Zhao W J. Advances in research on embryo culture of
Chinese jujube. Journal of Fruit Science, 2014, 31(5): 967-972. (in
Chinese)
[83] 陈学森, 张艳敏, 张连忠, 杜欣阁, 郭延奎. 特早熟杏的育种. 园
艺学报, 1996, 23(1): 27-31.

3538 中 国 农 业 科 学 48 卷
Chen X S, Zhang Y M, Zhang L Z, Du X G, Guo Y K. Breeding of
very early ripening apricot. Acta Horticulturae Sinica, 1996, 23(1):
27-31. (in Chinese)
[84] 陈学森, 高东升, 李宪利, 张艳敏, 张连忠. 胚培早熟杏新品种—
新世纪. 园艺学报, 2001, 28(5): 475.
Chen X S, Gao D S, Li X L, Zhang Y M, Zhang L Z. ‘Xinshiji’—a
new early ripening apricot variety obtained by embryo culture. Acta
Horticulturae Sinica, 2001, 28(5): 475. (in Chinese)
[85] 陈学森, 高东升, 李宪利, 张艳敏, 张连忠. 胚培早熟杏新品种—
红丰. 园艺学报, 2001, 28(6): 575.
Chen X S, Gao D S, Li X L, Zhang Y M, Zhang L Z. ‘Hongfeng’-a
new early ripening apricot variety obtained by embryo culture. Acta
Horticulturae Sinica, 2001, 28(6): 575. (in Chinese)
[86] 陈学森, 束怀瑞, 李宪利, 高东升, 张艳敏, 沈向, 陈晓流, 何天明.
胚培杏新品种—山农凯新 1 号. 园艺学报, 2005, 32(1): 176.
Chen X S, Shu H R, Li X L, Gao D S, Zhang Y M, Shen X, Chen X L,
He T M. Shannongkaixin 1—a new apricot variety obtained by
embryo culture. Acta Horticulturae Sinica, 2005, 32(1): 176. (in
Chinese)
[87] 陈学森, 束怀瑞, 李宪利, 高东升, 张艳敏, 沈向, 陈晓流, 何天明.
胚培杏新品种—山农凯新 2 号. 园艺学报, 2005, 32(2): 368.
Chen X S, Shu H R, Li X L, Gao D S, Zhang Y M, Shen X, Chen X L,
He T M. Shannongkaixin 2—a new apricot variety obtained by embryo
culture. Acta Horticulturae Sinica, 2005, 32(2): 368. (in Chinese)
[88] 张艳敏, 陈晓流, 毛志泉, 张芮, 冀晓昊, 冯守千, 陈学森. 极早熟
杏新品种‘极早红’. 园艺学报, 2012, 39(3): 595-596.
Zhang Y M, Chen X L, Mao Z Q, Zhang R, Ji X H, Feng S Q, Chen X
S. A new very early-maturing apricot cultivar ‘Jizaohong’. Acta
Horticulturae Sinica, 2012, 39(3): 595-596. (in Chinese)
[89] 陈学森, 王志刚, 周荣永, 姜广仲. 果树杂交去雄方法的研究. 山
东农业大学学报: 自然科学版, 1989, 20(3): 21-26.
Chen X S, Wang Z G, Zhou R Y, Jiang G W. Studies on the
emasculation method for fruit cross. Journal of Shandong Agricultural
University: Natural Science Edition, 1989, 20(3): 21-26. (in Chinese)
[90] 陈学森, 李宪利, 张艳敏, 吴树敬, 沈洪波, 束怀瑞. 杏种质资源
评价及遗传育种研究进展. 果树学报, 2001, 18(3): 178-181.
Chen X S, Li X L, Zhang Y M, Wu S J, Shen H B, Shu H R. Advances
in apricot germplasm resources evaluation and genetic breeding.
Journal of Fruit Science, 2001, 18(3): 178-181. (in Chinese)
[91] 柴菊华, 赵习平. 我国极早熟杏育种研究进展. 园艺学报, 2002,
29(增刊): 635-638.
Chai J H, Zhao X P. Advances in breeding of very early ripening
apricot in China. Acta Horticulturae Sinica, 2002, 29(Suppl.):
635-638. (in Chinese)
[92] 杨红花, 陈学森, 李玉晖, 冯宝春, 穆秀家. 利用远缘杂交创造核
果类果树新种质的研究,Ⅰ—核果类果树远缘杂交不亲和性的调控
研究. 中国农业科学, 2004, 37(7): 1034-1038.
Yang H H, Chen X S, Li Y H, Feng B C, Mu X J. Creating new
germplasm by distant hybridization in stone fruits . effects of Ⅰ
different treatments on cross-compatibility of distant. Scientia
Agricultura Sinica, 2004, 37(7): 1034-1038. (in Chinese)
[93] 杨红花, 陈学森, 冯宝春, 刘焕芳, 郑洲. 利用远缘杂交创造核果
类果树新种质的研究.Ⅱ—李杏远缘杂种的胚抢救与杂种鉴定. 中
国农业科学, 2004, 37(8): 1203-1207.
Yang H H, Chen X S, Feng B C, Liu H F, Zheng Z. Creating new
germ plasm by distant hybridization in stone fruits Ⅱ. embyro reseue
and hybrid identification between plum and aprieot. Scientia
Agricultura Sinica, 2004, 37(8): 1203-1207. (in Chinese)
[94] 刘焕芳, 陈学森, 段成国, 杨红花, 冯宝春. 甜樱桃与中国樱桃杂
种的胚抢救及杂种鉴定. 园艺学报, 2004, 31(3): 303-308.
Liu H F, Chen X S , Duan C G, Yang H H, Feng B C. Embryo rescue
and identification of hybrids between sweet cherry and Chinese cherry.
Acta Horticulturae Sinica, 2004, 31(3): 303-308. (in Chinese)
[95] 刘文, 陈学森, 梁青, 刘冠军, 吴燕. 桃远缘杂种的获得及杂种胚
抢救技术的建立. 园艺学报, 2007, 34(1): 29-34.
Liu W, Chen X S, Liu G J, Wu Y. Interspecific hybridization of prunus
persica with P. armeniaca and P. salicina using embryo rescue. Acta
Horticulturae Sinica, 2007, 34(1): 29-34. (in Chinese)
[96] 张剑侠, 牛茹萱. 无核葡萄胚挽救技术的研究现状与展望. 园艺学
报, 2013, 40(9): 1645-1655.
Zhang J X, Niu R X. The present dituation and prospect of embryo
rescue technique research in seedless grape breeding. Acta
Horticulturae Sinica, 2013, 40(9): 1645-1655. (in Chinese)
[97] Ramming D W, Emershad R L. In-ovule embryo culture of seeded and
seedless Vitis vinifera L. Hortscience, 1982, 17(3): 4871.
[98] Ramming D W, Emershad R, Tarailo R. A stenosper mocarpic,
seedless Vitis vinifera × Vitis rotundifolia hybrid developed by embryo
rescue. Hortscience, 2000, 35(4): 732-734. (in Chinese)
[99] Shima M, Tamura S, Inabe Y, et al. New grape [Vitis vinifera]
cultivar ‘Ruby Roman’. Bulletin of the Ishikawa Agriculture
Research Center, 2006(27): 33-36.
[100] 郭海江, 王跃进, 张剑侠, 潘学军, 唐冬梅, 田丽丽. 葡萄抗病无
核胚挽救育种及分子标记辅助选择. 西北植物学报, 2005, 25(12):
2395-2401.
Guo H J, Wang Y J, Zhang J X, Pan X J, Tang D M, Tian L L.
Development of resitant and seedless grape germplasms by embryo

17 期 陈学森等:主要果树果实品质遗传改良与提升实践 3539
rescue and marker-assisted. Acta Botanica Boreali-0ccidentalia Sinica,
2005, 25(12): 2395-2401. (in Chinese)
[101] 蒋爱丽, 李世诚, 金佩芳, 杨天仪, 骆军. 胚培无核葡萄新品种-
沪培 1 号. 果树学报, 2007, 24(3): 402-403.
Jiang A L, Li S C, Jin P F, Yang T Y, Luo J. Hupei 1-a new triploid
seedless grape cultivar obtained by embryo culture. Journal of Fruit
Science, 2007, 24(3): 402-403. (in Chinese)
[102] 蒋爱丽, 李世诚, 杨天仪, 骆军, 张朝轩, 金佩芳. 无核葡萄新品
种-沪培 2 号的选育. 果树学报, 2008, 25(4): 618-619.
Jiang A L, Li S C, Yiang T Y, Luo J, Zhang C X, Jin P F. A new
seedless grape cultivar—Hupei No.2. Journal of Fruit Science, 2008,
25(4): 618-619. (in Chinese)
[103] 蒋爱丽, 奚晓军, 程杰山, 田益华. 早熟葡萄新品种-‘申爱’的选
育. 果树学报, 2014, 32(2): 335-336.
Jiang A L, Xi X J, Cheng J S, Tian Y H.‘Shen’ai’. a new early grape
cultivar. Journal of Fruit Science, 2014, 32(2): 335-336. (in Chinese)
[104] Li G R, Ji W, Wang G, Zhang J X, Wang Y J. An improved
embryo-rescue protocol for hybrid progenyfrom seedless Vitis vinifera
grapes× wild Chinese vitis species. In Vitro Cellular & Developmental
Biology of Plant, 2014, 50(1): 110-120.
[105] 梁春莉, 赵锦, 刘孟军. 冬枣极早期幼胚培养成苗技术研究. 园艺
学报, 2014, 41(10): 2115-2124.
Liang C L, Zhao J, Liu M J. A high-efficiency regeneration system for
immature embryo of Ziziphus jujube ‘Dongzao’ at very early stage.
Acta Horticulturae Sinica, 2014, 41(10): 2115-2124. (in Chinese)
[106] 祁业凤, 刘孟军. 枣的胚败育及幼胚培养研究. 园艺学报, 2004,
31(1): 78-80.
Qi Y F, Liu M J. Embryo abortion and young embryo culture of
Chinese jujube. Acta Horticulturae Sinica, 2004, 31(1): 78-80. (in
Chinese)
[107] 刘孟军, 王玖瑞, 刘平, 林敏娟, 肖京, 刘志国, 孙学超. 枣树免去
雄杂交育种的设计与实践. 园艺学报, 2014, 41(7): 1495-1502.
Liu M J, Wang J R, Liu P, Lin M J, Xiao J, Liu Z G, Sun X C. Design
and practice of emasculation-free cross breeding in Chinese jujube.
Acta Horticulturae Sinica, 2014, 41(7): 1495-1502. (in Chinese)
[108] 李宝江, 景士西, 丁玉英, 张景娥. 苹果糖酸遗传和选择研究. 遗
传学报, 1994, 21(2): 147-154.
Li B J, Jing S X, Ding Y Y, Zhang J E. The research of apple suger
and acid genetic and choice. Acta Genetica Sinica, 1994, 21(2):
147-154. (in Chinese)
[109] Chen X S, Wu Y, Chen M X, He T M, Feng J R, Liang Q, Liu W, Yang
H H, Zhang L J. Inheritance and correlation of self-compatibility and
other yield components in the apricot hybrid F1 populations.
Euphytica, 2006, 150: 69-74.
[110] 吴燕, 陈学森, 冯建荣, 陈晓流. 杏杂种一代群体 S 基因的遗传研
究. 园艺学报, 2005, 32(3): 397-402.
Wu Y, Chen X S, Feng J R, Chen X L. Inheritance of S-gene among
the F1 progenies in apricot. Acta Horticulturae Sinica, 2005, 32(3):
397-402. (in Chinese)
[111] 陈美霞, 陈学森, 王新国, 慈志娟. 凯特与新世纪杏杂种后代风味
物质遗传的初步研究. 园艺学报, 2006, 33(5): 942-946.
Chen M X, Chen X S, Wang X G, Ci Z J. Inheritance of flavor
constituents in F1 progenies of Katy and Xinshiji apricot. Acta
Horticulturae Sinica, 2006, 33(5): 942-946. (in Chinese)
[112] 崔艳波, 陈慧, 乐文全, 张树军, 伍涛, 陶书田, 张绍铃. ‘京白梨’
与‘鸭梨’正反交后代果实性状遗传倾向研究. 园艺学报, 2011,
38(2): 215-224.
Cui Y B, Chen H, Yue W Q, Zhang S J, Wu T, Tao S T, Zhang S L.
Studies on genetic tendency of fruit characters in reciprocal crosses
generation between ‘Jingbaili’ and ‘Yali’ pear cultivars. Acta
Horticulturae Sinica, 2011, 38(2): 215-224. (in Chinese)
[113] Celia M. Cantin,Yolanda Gogorcena, María Á ngeles Moreno.
Phenotypic diversity and relationships of fruit quality traitsin peach
and nectarine [Prunus persica (L.) Batsch] breeding progenies.
Euphytica, 2010, 171: 211-226.
[114] Wu B H, Zhao J B, Chen J, Xi H F, Jiang Q, Li S H. Maternal
inheritance of sugars and acids in peach (P. persica (L.) Batsch) fruit.
Euphytica, 2013, 189: 351-364.
[115] 景士西, 吴录平, 李宝江. 果树遗传变异的特点初探. 遗传, 1995,
17(1): 40-44.
Jing S X, Wu L P, Li B J. Preliminary study on genetic characteristics
of fruit planst. Hereditas, 1995, 17(1): 40-44. (in Chinese)
[116] 伊凯, 闰忠业, 刘志, 王冬梅, 杨峰, 张景娥. 苹果芽变选种鉴定
及应用研究. 果树学报, 2006, 23(5): 745-749.
Yi K, Yan Z Y, Liu Z, Wang D M ,Yang F, Zhang J E. Review on
identification and utilization of apple sport selection. Journal of Fruit
Science, 2006, 23(5): 745-749. (in Chinese)
[117] 王传增, 张艳敏, 徐玉亭, 董飞, 宋杨, 刘美艳, 刘金, 陈学森. 苹
果红色芽变香气组分及脂肪酸代谢相关酶活性分析. 园艺学报,
2012, 39(12): 2447-2456.
Wang C Z, Zhang Y M, Xu Y T, Dong F, Song Y, Liu M Y, Liu J,
Chen X S. Analysis of aroma components and related enzymes of fatty
acid metabolism of red bud sports. Acta Horticulturae Sinica, 2012,
39(12): 2447-2456. (in Chinese)
[118] 陈学森, 王恩琪, 毛志泉, 张艳敏, 吴树敬. 短枝型苹果新品种‘龙
富’. 园艺学报, 2013, 40(9): 1851-1852.

3540 中 国 农 业 科 学 48 卷
Chen X S, Wang E Q, Mao Z Q, Zhang Y M, Wu S J. A new spur-type
apple cultivar ‘Longfu’. Acta Horticulturae Sinica, 2013, 40(9):
1851-1852. (in Chinese)
[119] 宋杨, 张艳敏, 刘金, 王传增, 刘美艳, 冯守千, 陈学森. GA 含量
与其合成酶基因在‘长富 2 号’苹果及其短枝型芽变品种之间的比
较分析. 中国农业科学, 2012, 45(13): 2668-2675.
Song Y, Zhang Y M, Liu J, Wang C Z, Liu M Y, Feng S Q, Chen X S.
Comparison of gibberellin acid content and the genes relatived to GA
biosynthesis between ‘Changfu 2’ apple (Malus domestica Borkh.)
and its spur sport. Scientia Agricultura Sinica, 2012, 45(13):
2668-2675. (in Chinese)
[120] 宋杨, 张艳敏, 王传增, 刘美艳, 刘金, 王延玲, 陈学森. 苹果光敏
色素作用因子基因 PIF 的克隆和分析. 园艺学报, 2012, 39(4):
743-748.
Song Y, Zhang Y M, Wang C Z, Liu M Y, Liu J, Wang Y L, Chen X S.
Clone and expression analyzing of phytochrome-interacting factor
gene PIF of spur type apple. Acta Horticulturae Sinica, 2012, 39(4):
743-748. (in Chinese)
[121] 宋杨, 张艳敏, 刘美艳, 王传增, 刘金, 冯守千, 王延玲, 陈学森.
短枝型苹果 MdRGL 基因的克隆及原核表达分析. 中国农业科学,
2012, 45(7): 1347-1354.
Song Y, Zhang Y M, Liu M Y, Wang C Z, Liu J, Feng S Q,Wang Y L,
Chen X S. Cloning and prokaryotic expression of MdRGL gene from
spur-type apple (Malus domestica Borkh.). Scientia Agricultura Sinica,
2012, 45(7): 1347-1354. (in Chinese)
[122] 宋杨, 吴树敬, 张艳敏, 陈学森. 短枝型苹果 SSH 文库构建及相关
基因表达分析. 中国农业科学, 2013, 46(20): 4301-4309.
Song Y, Wu S J, Zhang Y M, Chen X S. Construction of the
suppression subtractive hybridization library and analysis of related
genes of spur-type apple (Malus domestica Borkh.). Scientia
Agricultura Sinica, 2013, 46(20): 4301-4309. (in Chinese)
[123] 宋杨, 张艳敏, 吴树敬, 冯守千, 王传增, 陈学森. 短枝型苹果
MdGID1a 基因的克隆、表达及启动子序列分析. 园艺学报, 2013,
40(11): 2237-2244.
Song Y, Zhang Y M, Wu S J, Feng S Q, Wang C Z, Chen X S. Cloning,
expression analysis of MdGID1a gene and promoter from spur-type
apple. Acta Horticulturae Sinica, 2013, 40(11): 2237-2244. (in Chinese)
[124] 冀晓昊, 毛志泉, 张艳敏, 吴树敬, 陈晓流, 陈为一, 陈学森. 晚熟
苹果新品种‘山农红’. 园艺学报, 2014, 41(7): 1503-1504.
Ji X H, Mao Z Q, Zhang Y M, Wu S J, Chen X L, Chen W Y, Chen X
S. A new late ripening apple cultivar ‘Shannonghong’. Acta
Horticulturae Sinica, 2014, 41(7): 1503-1504. (in Chinese)
[125] Xu Y T, Feng S Q, Jiao Q Q, Liu C C, Zhang W W, Chen W Y, Chen
X S. Comparison of MdMYB1 sequences and expression of
anthocyanin biosynthetic and regulatory genes between Malus
domestica Borkh. Cultivar ‘Ralls’ and its blushed sport. Euphytica,
2012, 185: 157-170.
[126] 刘晓静, 冯宝春, 冯守千, 王海波, 石俊, 王娜, 陈为一, 陈学森.
‘国光’苹果及其红色芽变花青苷合成与相关酶活性的研究. 园艺
学报, 2009, 36(9): 1249-1254.
Liu X J, Feng B C, Feng S Q, Wang H B, Shi J, Wang N, Chen W Y,
Chen X S. Studies on anthocyanin biosynthesisand activities of related
enzymes of ’Ralls’ and its bud mutation. Acta Horticulturae Sinica,
2009, 36(9): 1249-1254. (in Chinese)
[127] Xu Q, Chen LL, Ruan X, Chen D, Zhu A, Chen C, Bertrand D, Jiao
W B, Hao B H, Lyon M P, Chen J, Gao S, Xing F, Lan H, Chang J W,
Ge X, Lei Y, Hu Q, Miao Y, Wang L, Xiao S, Biswas M K, Zeng W,
Guo F, Cao H, Yang X, Xu X W, Cheng Y J, Xu J, Liu J H, Luo O J,
Tang Z, Guo W W, Kuang H, Zhang H Y, Roose M L, Nagarajan N,
Deng X X, Ruan Y. The draft genome of sweet orange (Citrus
sinensis). Nature Genetics, 2013, 45: 59-66.
[128] Asins M J, Monforte A J, Mestre P F, Carbonell E A. Citrus and
Prunus copia-like retrotransposons. Theoretical and Applied Genetics,
1999, 99: 503-510.
[129] Kumar S, Volz R K, Alspach P A, Bus V G. Development of a
recurrent apple breeding programme in New Zealand: a synthesis of
results, and a proposed revised breeding strategy, Euphytica, 2010,
173: 207-222.
(责任编辑 赵伶俐)

中国农业科学 2015,48(17):3541-3546 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.019
收稿日期:2015-01-20;接受日期:2015-06-17 联系方式:姚惠源,E-mail:[email protected]
中国粮食加工科技与产业的发展现状与趋势
姚惠源
(江南大学食品学院,江苏无锡 214122)
摘要:中国是世界粮食生产和粮食加工大国,米面加工能力和产量均居世界之首。米面产品结构不断优化、
总体质量水平明显提升,加工企业规模和生产集中度显著提高,工业化生产和加工装备水平提升。但在产业规模
化、产业链建设、粮食资源高效利用及科技自主创新能力方面与发达国家尚有一定差距。对中国粮食加工科技与
产业发展中尚存在的问题及成因进行分析,主要问题是发展方式比较粗放、初加工产能相对过剩、粮食加工产品
质量安全和保障体系不够完善、主食品工业化水平不高;主要成因为基础研究薄弱,技术创新跟不上产业发展的
需求。同时展望了中国粮食加工业的发展前景,要更加推进产业结构调整与升级,提高科技成果转化率,提升关
键装备自主化率水平,加速提高产品质量安全水平,加强全谷物食品的研发与推广。未来 10 年,中国粮食加工的
区域布局和产业结构将更加合理、产品将更加丰富多元、质量更加绿色安全。
关键饲:粮食加工;科技;产业结构;前景
The Present Development Status and Tendency of Grain
Processing Technology and Industry in China YAO Hui-yuan
(School of Food Science and Technology, Jiangnan University, Wuxi 214122, Jiangsu)
Abstract: This paper discussed the development of grain processing technology and industry in China. Rice processing capacity and output in China rank the first in the world. The product structure of rice and flour is optimized continuously and the overall level of quality has been improved significantly. The processing enterprise production scale and concentration degree have been greatly improved, the industrial production and processing equipment have been upgraded. Being a huge nation of grain production and processing in the world, China still has some gaps in industrial-scale, industry chains, efficient use of grain resources, and the innovation ability in science & technology as compared with the developed countries. The existing problems and causes of grain processing technology in China and industrial development were analyzed. The main problems are extensive development mode, relative overproduction of primary processing products, imperfection of quality safety of grain products and security system, and low level of the industrialization of the staple food. The primary causes are weak basal researches, insufficient support degree of processing technology research to the development of industry. This paper also addressed the prospect of grain processing technology and industry of China. The paper was suggested that the industrial restructuring and upgrading should be further promoted, the scientific and technological achievements conversion rate and the level of key equipment autonomy should be improved, the improvement of product quality and safety level should be accelerated, and the development and promotion of whole grain food should be strengthen in the future. In the next 10 years, the regional layout and industrial structure of grain processing in China will be more reasonable, the product will be richer, quality will be greener and more security.
Key words: grain processing; technology; industrial structure; prospect
jia
文本框
返回目录

3542 中 国 农 业 科 学 48 卷
0 引言
中国是全球粮食生产大国,2014 年中国粮食总产
量达到 6.07 亿 t,实现半个世纪以来首次连续 11 年增
产,创历史新高,创造了用仅占世界 8.06%耕地面积,
生产了占全球 25%粮食的奇迹,为保障世界粮食安全
做出了重要贡献[1]。中国也是世界粮食加工大国,粮
食加工涵盖的面很广,包括稻谷、小麦、玉米三大主
粮及其延伸的米、面食品加工,多品种杂粮及薯类加
工也属于粮食加工的范围。粮食的初级加工是农作物
加工的基础产业,在食品工业中处于支柱地位。加工
技术是粮食加工产业发展的重要科技支撑,近 10 多年
来,中国在稻米、小麦加工工艺、生产技术、加工装
备等领域取得了数以百计的重大科研成果,成果水平
已达到或接近世界先进水平,有力支撑了中国粮食
加工产业的迅猛发展[2]。但当前中国粮食加工科技
和产业与世界发达国家相比尚有一定差距,主要表
现在粮食加工产业规模化生产、集约化经营有待提
高;粮食加工产业链建设有待完善、粮食资源高效
利用技术有待突破;产能过剩和节能减耗有待解决;
科技自主创新能力需加强等。国家粮食主管部门和
粮食加工业内已看到中国与发达国家的这些差距,
近年来,国家工业信息化部、国家粮食局和农业部
在大量调查研究的基础上联合制定了中国粮食加工
产业未来 10 年的发展规划,要用近 10 年时间大力
提高粮食加工的技术实力,使之达到国际先进水平。
因此,中国粮食加工科技及其产业将迎来重要的快
速发展机遇。
1 以稻米和小麦为主体的中国粮食加
工科技与产业发展现状
改革开放以来,尤其是进入新世纪以来,中国以
稻谷和小麦产量占世界第一的粮食资源优势和自主创
新的粮食加工科技,使中国稻米和小麦加工产业取得
了突飞猛进的发展,其发展速度、发展规模和发展质
量,在中国历史上、乃至世界历史上都是前所未有的。
目前中国有大米加工企业 8 500 多个、小麦粉加工企
业 3 000 多个,年处理稻谷和小麦能力为 4.03 亿 t,均居世界首位[3]。中国大米、小麦面粉加工的科技水平、
装备水平、主要经济技术指标以及产品品种和质量等
诸多方面已经达到或接近世界先进水平,当然距离粮
食加工科技与产业强国尚有一定差距。
1.1 大米、面粉加工科技取得重大研究成果
在粮食生产和加工快速发展的形势下,中国粮食
加工科技近 10 年来已取得许多重大科研成果,中国粮
食加工技术和设备工艺的持续提高,使粮食加工业在
现代化道路上迈出了坚实的一步。 稻米与副产品深加工高效增值创新技术在国内取
得突破,该成果已经获国家科学技术进步二等奖,这
项成果在稻谷加工技术方面取得了突破性成果,如生
物酶、分子修饰、挤压、超细粉碎等,并在国内首次
研究并建立了稻米与副产品深加工高效循环增值系
统。 高品质蒸谷米加工关键技术和产业化取得突破,
该技术攻克了利用早籼稻生产高品质蒸谷米的关键技
术,解决了目前普遍存在的早籼稻食用率、利用率低
下的问题,通过研究早籼稻的加工特性和营养特点,
重点解决了影响蒸谷米品种的 5 个关键生产技术,包
括变压分级浸泡、双温变速干燥、通风缓苏、辅助碾
白与产品质量评价。这项成果的应用,使出米率提高
了 18%以上(最高达 71%),生产效率与国外相比提
高了 1.5 倍,而成本较国外下降 20%。同时,营养品
质有了大幅度提升,其中 VB1、VB2 和铁含量分别增
加了 267%、67%和 135%。 稻米主食生产关键技术及产业化研究取得成功,
研制与开发了有机糙米粥口感好、且减少营养物质流
失的生产工艺与技术;研究成功蒸煮特性和口感好的
营养复合米生产工艺及关键设备。 米制品加工基础理论和关键生产技术研究取得新
突破,研究了稻米淀粉组分、米淀粉糊化、米淀粉老
化与凝胶化等基础理论和米制食品生产关键技术,为
中国袋装、灌装等各类包装的熟制方便米饭及糙米糊、
米粉等米制食品的快速发展提供了技术支撑。 高效节能与清洁安全小麦加工新技术研究与应用
取得突破性进展,研发了多项独具特色的小麦湿法、
干法加工工艺,并且开发出磨撞均衡出粉等制粉新技
术,使中国制粉工艺处于国际先进水平,该成果获国
家科学技术进步二等奖。 小麦专用粉和传统面制品工业化生产技术与装备
开发取得新突破,研制了面条、馒头、饼干等 30 种食
品专用小麦粉,开发了方便面、挂面、馒头、水饺自
动生产线,提高了产品得率,降低了能耗,优化了产
品结构。 小麦硬度测定技术及其设备的开发取得突破,解
决了小麦收购、储运及加工环节由于缺乏量化的小麦

17 期 姚惠源:中国粮食加工科技与产业的发展现状与趋势 3543
硬度测试技术而导致无法对小麦科学定等、合理调整
制粉工艺以及影响小麦粉出率等技术难题。 1.2 大米、面粉的加工能力和产量均居世界之首
2013 年中国大米总产量达 1 亿 t、面粉总产量
为 9 000 多万 t,创历史新高。稻谷及小麦加工企业数
量和加工能力已经跃居世界首位。生产工艺和装备技
术的不断提高,实现了中国大米、小麦加工业的现代
化,接近国际先进水平。粮食加工产业不仅能够充分
满足城乡居民基本生活需求,并且在产品结构上也得
到了优化。大米加工能力和产量 5 年中平均年增长
12.9%和 32.7%;小麦粉加工能力和产量 5 年中平均年
增长 13.7%和 24.1%[3]。 大米、面粉年产量须以《国家粮食安全中长期发
展规划纲要》(2008—2020 年)和国家粮食局《2009年度全社会粮食供需平衡调查报告》有关规划和数据
为依据,保证供需平衡。根据中国粮食消费需求预测,
2015—2020 年中国粮食消费总量为 5 725 亿 kg(5.725亿 t)。其中口粮 2 550 亿 kg,饲料粮 2 162.5 亿 kg,工业用粮 810 亿 kg,种子粮 115 亿 kg。口粮中稻谷
1 500 亿 kg、小麦 1 050 亿 kg,分别折合大米 9 925万 t、小麦面粉 7 500 万 t。因此,中国每年生产大米
1 亿 t 左右,面粉 8 000 万 t 左右,能够达到供需平衡,
保障国民粮食供应安全[3]。 1.3 大米、面粉产品结构不断优化
2013 年大米总产量中 80%是特等大米和标准一
等大米;小麦粉产品中,特制一等粉、特制二等粉和
专用粉的产量为面粉总量的 79.27%,分别比 5 年前增
加 137%和 127.2%[3],说明中国大米、面粉以优等产
品为主体的产品结构己经形成。 1.4 大米、面粉企业规模增大和生产集中度显著提高
近年来规模以上企业的产品产能、产量占比有所
提高,规模化集约化水平进一步提高。其中稻谷、小
麦加工业规模效应不断显现,日处理稻谷和小麦 400 t以上大型企业的产能占比分别为 23.0%及 53.9%,产
量占比分别为 33.3%及 63.4%[3],表明中国大米、面粉
加工企业正在向规模化生产、集约化经营迈进。 1.5 大米、面粉产品总体质量水平明显提升
根据 2009 年以来国家质检总局公布的大米、面粉
产品实物质量抽样检查结果,大米抽样合格率保持在
98%左右,小麦粉抽样合格率保持在 99%左右。在中
国名牌产品评选中,分别有 28 家大米加工企业、29家小麦粉加工企业生产的大米、面粉被誉为中国名牌
产品。
1.6 大米、面粉主食品的工业化生产快速发展
随着中国国民经济的快速发展和人民生活水平的
迅速提高,近 10 年来中国主食工业化生产取得了快速
发展,特别在面制主食方面,如面条、馒头、饺子、
方便米粉及八宝粥等已经进行工业化生产并初具规
模。速冻主食工业化生产也得到了快速发展。中国五
千年的饮食文明造就了品种多样、花色繁多的传统主
食品,其中,以馒头、面条、水饺等为代表的主食品
种消费量已占面制食品总量的 85%。 馒头是中国北方地区的主食,在中国已有几千年
的历史。以前馒头生产一直以手工制作为主,但近几
年来中国大型馒头生产线已投入使用,最大特点是该
设备模拟手工馒头制作全过程,口感与手工制作馒头
相差无几,为馒头产业化生产提供了设备保证。 面条也是中国传统主食,主要包括挂面和方便面
两大品种。随着加工技术的改进,大型面辊机组研发
成功,挂面产量明显提升,降低了生产成本。目前中
国挂面年产量稳定在 250 万 t,年面粉用量大约为 300万 t。年产 5 000 t 以上的企业有 100 多家,10 000 t以上的有 30 家,排名前十的企业销量占总量的 5%,
说明中国挂面生产已经步入规模化、集约化阶段。
中国方便面产业经过 30 年的发展和整合,生产方便
面企业已由以前的 800 多家整合至 80 家,产业已向
规模化、聚集化发展。2013 年中国方便面产量达 500多亿包,总产量占到全球的 51%,销售额超 500 亿
元,已成为世界第一大方便面生产国。但笔者认为
方便面目前仍不属于健康食品,未来应在“健康”
二字上多加改进。 米饭是中国百姓的重要主食,2/3 的人以米饭为主
食,但由于家庭生活中米饭的蒸煮较为简单快捷,因
此,米饭的工业化生产比馒头明线缓慢。近 10 年来,
米制品的工业化生产主要以米线为主,普通米饭的流
水线制作主要集中在学校和工厂等集中用餐的单位。
保鲜米饭、冷冻米饭、速煮米饭等方便米饭及保鲜米
线、方便米粉(米线)等米制主食和调理方便米制品
已进行工业化生产,也有快速发展势头。 此外,全谷物食品在中国悄然兴起。全谷物食品
是世界公认的健康食品,美国临床化学协会将全谷物
定义为:完整、碾碎、破碎或压片的颖果,其基本组
成包括淀粉胚乳、胚芽与麸皮,各组成部分的相对比
例与完整颖果一样。美国食品药品管理局对全谷物定
义与以上表述几乎相同。科学研究证实,食用全谷物
食品可以减少心脏病[4]、II 型糖尿病[5]、消化道癌症[5]、

3544 中 国 农 业 科 学 48 卷
CVA 脑血管病[6]等慢性疾病的危害[7]。在欧美全谷物
(Whole Grain)营养健康食品快速发展的引领下,中
国全谷物食品也得到了快速发展[8]。 1.7 粮食加工装备水平迅速提升
随着科技水平的整体进步,通过自主创新和引进
消化吸收,中国粮食加工设备和工艺已经达到国际先
进水平,并且已经走向国外市场。通过自主创新和引
进消化吸收,并应用计算机辅助设计和制造、精密铸
造、数控加工中心、数控折弯冲压、激光切割、机电
一体化和自动化控制等先进技术和手段,新产品不断
涌现,质量和性能不断提高,大型粮食加工成套设备
制造技术提升较快,日处理稻谷 300 t、小麦 1 000 t的成套设备与工艺达到国际先进水平,不仅能满足国
内粮食加工业发展需要,同时也远销国外。 稻米加工装备在谷糙分离机、色选机、厚度分级
机、大米抛光机等设备研发,小麦加工装备在磨粉机、
撞击机、平筛、清粉机等主要设备的研发等诸多领域
已达到或接近世界先进水平。
2 中国粮食加工科技与产业发展中存
在的问题及成因分析
改革开放 30 多年来,中国粮食加工科技和产业虽
取得了飞跃发展,但离国际先进水平尚有一定差距,
粮食加工业总体上还属于粗放型产业。中国目前粮食
加工的机械化水平和生产效率与美国、欧洲等发达国
家差距较大,粮食加工主体主要是遍布城乡的小型企
业,加工的粮食数量约占粮食加工总量的 70%。全美
国面粉加工企业劳动生产率为中国面粉加工企业的 2倍以上,全美国只有约 100 家面粉加工企业,日加工
能力超过 1 000 t 的面粉厂却占总生产能力的 50%以
上。另外,集约化程度高,有的企业生产几十个品种
的面粉,却只有 100 余员工。中国粮食加工业对米、
面加工片面注重外观的白度和亮度,致使粮食过度加
工,既使粮食中的营养素大量流失,又大量浪费宝贵
的粮食资源的问题突出。还有米、面质量标准尚停留
在传统的物理指标,缺乏营养和卫生标准,食品质量
安全追溯体系尚待建立。另外由于过度加工造成能耗
高、环境污染、资源浪费。 2.1 粮食加工业总体上是属于粗放型产业
中国粮食产业结构不够合理,发展方式仍较粗放,
深加工水平较低,粮食食用率只有 65%—70%。过度
加工现象较普遍,小麦加工精制面粉出品率约 70%,
稻米加工精制米出品率约 65%,剩余部分浪费严重。
稻壳、麸皮、米糠等具有的营养价值和能源价值没有
被充分地开发利用。 目前发达国家正在掀起全谷物食品的热潮,因为
全谷物食物保留了谷物皮层中丰富的 VB1和 VB2等 B族维生素、矿物元素和膳食纤维,有利于防止多种慢
性疾病。而目前中国粮食资源的高效、科学利用及其
基础理论和应用技术研究也较薄弱,致使大量的粮食
资源被浪费。 2.2 粮食加工产品质量安全和保障体系不够完善
中国大米、面粉产品的质量标准尚停留在一些
物理指标,如水分、杂质、灰分、碎米含量及色泽
等感观指标,缺少营养和卫生指标,特别质量安全
的追溯制度尚未建立。20 世纪 90 年代开始,许多
发达国家通过建立追溯制度来推进粮食产品质量安
全管理。中国目前粮食产量已经能够满足人民需要,
迫切需要借鉴美国、欧盟和日本等较早开展食品追
溯标准化工作的经验,尽快建立起健全的法律法规
和执行机构。全面形成以预防、控制和追溯为特征
的食品质量安全监管体系,保障人民对食品安全日
益增长的需求[10]。 2.3 粮食加工全产业链条较短
粮食加工对稻米和小麦粮食资源的副产物综合利
用率较低,缺乏深度开发利用,产品附加值低。中国
粮食加工副产品资源丰富,稻谷加工产生的稻壳年总
量达 4 000 万 t 左右,米糠 1 000 多万 t,小麦加工面
粉所产生的麸皮 2 000 多万 t,还有营养丰富的稻米、
小麦胚芽等副产物,但由于缺乏有效的深度开发与利
用研究,产业链短或不完善,致使副产物综合利用率
低,副产品附加值低[11]。稻壳用于发电和锅炉直接燃
烧的不到 33%,米糠用于制油和深加工的不足 10%,
小麦加工副产物更缺乏有效利用,充分说明中国粮食
加工产业链与发达国家的差距,抓紧建设粮食加工产
业链已经迫在眉捷。 2.4 粮食主食品工业化研究水平不高
中国几千传承下来的传统面、米主食品种样式繁
多,制作方法多为蒸、煮,且具有丰富多彩的文化底
蕴,为现代米、面食品工业的发展提供了丰富的资源
库。但传统主食品的开发需要创造性和高科技,其并
非简单的规模化生产与自动化改造。国外如肯德基、
麦当劳汉堡包,方便面等就是成功开发的范例。它既
包括了对产品从营销学角度的定位和设计,也需要运
用现代营养学、加工学、工程学等知识和技术生产出

17 期 姚惠源:中国粮食加工科技与产业的发展现状与趋势 3545
受市场欢迎的新产品。中国本土的馒头、饺子等,其
发酵工艺、老化控制、风味和营养增强等加工工艺方
面的深入研究尚需加强,在标准化、规格化等方面与
商品性要求尚有一定差距。 2.5 粮食加工科技基础理论研究和自主创新能力较
薄弱
很长一段时间以来,中国对粮食加工科技研究的
投入不足,远低于发达国家 2%—3%的平均水平。基
础研究薄弱,技术支撑力度不够,制约了粮食加工业
的产业结构调整与升级。国家工程中心、工程实验室
等创新平台建设滞后,产、学、研结合不够紧密,创
新人才和开拓型经营管理人才不足,关键技术装备的
开发多处于仿制阶段,产品技术含量不高,集中反映
出自主创新能力较为薄弱[2]。
3 中国粮食加工科技与产业发展趋势
为了使中国粮食加工科技与产业由世界大国转
变为世界强国,中国国家工业信息化部、国家粮食
局和农业部在大量调查研究的基础上,联合制订并
于 2012 年 2 月 24 日发布了《粮食加工业发展规划
2011—2020 年》[12]。规划中明确了今后 10 年中国
粮食加工科技和产业的发展目标、重点任务、产业
布局、发展方向和重点工程等,这为中国粮食加工
产业发展指明了道路,也是粮食加工产业发展的趋
势。 3.1 中国粮食加工业发展目标
至 2020 年,要形成安全营养、优质高效、绿色
生态、布局合理、结构优化、协调发展的现代粮食
加工产业体系。要合力谋发展,做大做强,形成一
批销售收入 100 亿元以上的大型粮食加工企业集
团,建成一批粮食加工产业园区,培育一批知名品
牌[12]。 3.2 中国粮食加工科技与产业发展重点任务
推进产业结构调整与升级。加快推动主食品工业
现代化、产业化和健康化。加强对中国传统食品进行
全面系统地调查、整理、发掘和工业化改造的研究。
重点推广高效节能小麦和稻米加工高端设备。强化物
料分级与磨撞均衡出粉技术、小麦剥皮制粉等工艺;
推广稻谷低温干燥、产地脱壳、糙米调质、低温升碾
米先进实用技术。 提高科技成果转化率和提升关键装备自主化率水
平。粮食加工业技术进步贡献率分别达到 40%和 45%,
加工关键设备自主化率达到 60%—80%。加速建设技
术创新服务平台,建成若干个国家工程实验室或工程
技术研究中心[12]。 加速研究粮食食品现代技术。研究主食食品生物
技术、现代高效分离技术、非热杀菌技术、现代食品
干燥技术和淀粉物理改性技术等;着力推广副产品高
效增值深加工技术;推广稻壳发电及热能利用等技术
装备,加强麸皮、米糠加工膳食纤维新技术以及谷物
胚芽综合利用新技术研究。 加速提高产品质量安全水平。修订和制订相关标
准 1 000 项以上,产品质量合格率提高 2%—3%。使
制定的标准与国际接轨,要求大米、小麦粉总体合格
率分别达 98%和 99.5%以上,同时要尽快建立产品质
量安全追溯监管体系。 加大节能减排力度。当前中国稻米和小麦面粉加
工的单位能耗较高,由于过度加工致使每吨米、粉电
耗高达 65—70 kW,因此,节能减排成为未来 10 年粮
食加工行业的重要任务,到 2020 年单位产值能耗比
2010 年降低 20%以上,单位产值二氧化碳排放比降低
至少 20%,逐步实现绿色发展。 要做大、做强中国粮食加工科技和产业,需借鉴
国际上总销售额超过 2 000 亿美元(世界 500 强)的
A、B、C、D 四大粮食集团公司,即美国 ADM、BUNGE、CARGILL 和法国的 LOUIS DREYTUS 的“科技、人
才领先,产、学、研联合,科工贸为一体的发展经验”。
值得庆贺的是中国中粮集团公司 2014 年的销售额已
突破 3 000 亿人民币,居世界 500 强前 100 位,因此,
现在世界粮食巨商应加上中国的 COFCO(中粮)。
总体来说,粮食科技与产业发展关系国计民生,中国
的粮食产业发展之路任重道远。
References
[1] 姚惠源. 粮食加工企业的产业定位和产业规划. 粮食与食品工业,
2014, 21(6): 3-5.
Yao H Y. Industrial positioning and industrial planning of grain
processing enterprises. Cereal & Food Industry, 2014, 21(6): 3-5. (in
Chinese)
[2] 中国粮油学会. 粮油科学与技术学科发展报告(2010-2011). 北京:
中国科学技术出版社, 2011: 60-76.
China agri Society. Report on Advances in Cereal and Oil Science and
Technology (2010-2011). Beijing: China Science and Technology
Press, 2011: 60-76. (in Chinese)
[3] 姚惠源. 中国粮食加工产业转型升级的思考. 粮食与食品工业,
2013, 20(6): 1-2.

3546 中 国 农 业 科 学 48 卷
Yao H Y. Thinking on the transformation and upgrading of the grain
processing industry in China. Cereal & Food Industry, 2011, 4: 60-76.
(in Chinese)
[4] Anderson J W, Hanna T J, Peng X. Whole grain foods and heart
disease risk. Journal of the American College of Nutrition, 2000(3):
291-299.
[5] Slavin J L, Jacobs D, Marquart L. Grain processing and nutrition.
Critical Reviews in Biotechnology, 2001, 21(1): 49-66.
[6] Mohsen M. Vitamin E and atherosclerosis beyond prevention of LDL
oxidation. Journal of Nutrition, 2001, 131(2): 366s-368s.
[7] Jacobs D R Jr, Meyer K A, Kushi L H, Folsom A R. Whole-grain
intake may reduce the risk of ischemic heart disease death in
postmenopausal women: the Iowa women’s health study. American
Journal of Clinical Nutrition, 1998, 68(2): 248-257.
[8] 姚惠源. 全谷物健康食品发展趋势. 粮食与食品工业, 2012, 19(1):
1-3.
Yao H Y. Development trend of whole grain healthy food. Cereal &
Food Industry, 2012, 19(1): 1-3. (in Chinese)
[9] 谭斌, 谭洪卓, 刘明, 田晓红, 李爱科, 林家永. 粮食(全谷物)的营
养与健康. 中国粮油学报, 2010, 25(4): 100-107.
Tan B, Tan H Z, Liu M, Tian X H, Li A K, Lin J Y. The grain, the
wholegrain: nutrition and health benefits. Journal of the Chinese
Cereals and Oils Association, 2010, 25(4): 100-107. (in Chinese)
[10] 王瑞元. 中国粮食加工业的发展趋势. 粮食与食品工业, 2011,
18(5): 1-6.
Wang R Y. The developing trend of Chinese grain processing industry.
Cereal & Food Industry, 2011, 18(5): 1-6. (in Chinese)
[11] 王瑞元. 米制食品和米粉产业的发展. 粮油加工, 2009(2): 16-18,
23.
Wang R Y. Development of food and rice noodles industry. Cereals
and Oils Processing, 2009(2): 16-18, 23. (in Chinese)
[12] 工业和信息化部. 粮食加工业发展规划(2011-2020 年). 农产品市
场周刊, 2012(8): 24.
Ministry of Industry and Information Technology. Development plan
of grain processing industry (2011-2020). Market Weekly, 2012(8): 24.
(in Chinese)
(责任编辑 赵伶俐)

中国农业科学 2015,48(17):3547-3564 Scientia Agricultura Sinica doi: 10.3864/j.issn.0578-1752.2015.17.020
收稿日期:2015-01-20;接受日期:2015-04-27 基金项目:国家科技支撑计划(2013BAD12B00)(2013-2017)、国家公益性行业(农业)专项(201103008)(2011-2015) 联系方式:刘在新,Tel/Fax:0931-8342587;E-mail:[email protected]
全球口蹄疫防控技术及病原特性研究概观
刘在新
(中国农业科学院兰州兽医研究所/家畜疫病病原生物学国家重点实验室 OIE/中国国家口蹄疫参考实验室,兰州 730046)
摘要:口蹄疫是危害猪牛羊等主要家畜畜种的疫病,可造成巨大的经济损失,引发严重的负面社会影响,被
中国政府列在一类动物疫病的首位。中国猪、牛、羊养殖量最大、邻国众多,防控口蹄疫不仅对本国农牧业发展
起关键作用,而且对全球口蹄疫防控有重要意义。目前,除一些发达国家消灭了口蹄疫并保持着无疫状态外,口
蹄疫仍在众多发展中国家流行或散发。中国周边口蹄疫疫情不断,对中国造成高压威胁,近年流行的
O/ME-SA/PanAsia、O/SEA/Mya-98 和 A/ASIA/Sea-97 病毒均是传入的。从周边流行的病毒、流行的频率和循环的
区域位置判断,O/PanAsia-2、A/Iran-05、Asia1/Sindh-08 和 O/Ind-2001 病毒未来威胁依然较大。几十年来,
中国不断完善各级兽医服务体系,充分发挥国家口蹄疫参考实验室科研带动、技术指导、疫情监控的职能,采取
疫苗强制免疫为主的防控政策,卓有成效。依照世界动物卫生组织(OIE)推荐的口蹄疫渐进控制路线图(PCP-FMD),
中国目前处于阶段 3。推进口蹄疫防控效果进入更高阶段,关键在诊断检测技术和疫苗。自主研发的液相阻断
ELISA、非结构蛋白 3ABC 抗体检测 ELISA、多重 RT-PCR 等已达世界先进水平,有力支撑了中国口蹄疫诊断检测工
作,并在朝鲜、越南等国应用。新一代测序、胶体金和纳米示踪材料标记免疫层析和生物反应效应分子检测等更
精准便捷的新技术,未来也有望在口蹄疫诊断中实现应用。全病毒灭活疫苗免疫效力最优,是目前应用的主要疫
苗,但存在干扰鉴别诊断等缺陷。随着免疫学理论和基因工程技术的进步,发展和应用标记疫苗、活载体疫苗、
表位蛋白疫苗和病毒颗粒样疫苗等新型疫苗是未来的趋势。近十余年来,反向遗传操作凭借其定向改造基因组等
强大技术优势,不仅促进了标记疫苗等新型基因工程疫苗的研发,而且推动了口蹄疫病毒宿主嗜性、复制机制、
受体利用和先天性免疫应答等方面的基础研究,取得的研究成果必将有力推动防控应用型研究的进步。
关键词:口蹄疫;流行现状;防控新技术;病原特性
Progress and Prospect of the Technologies to Control Foot-and-
Mouth Disease and Its Pathogen Characteristics Worldwide LIU Zai-xin
(Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences/State Key Laboratory of Veterinary Etiological
Biology, OIE/China National Foot-and-Mouth Disease Reference Laboratory, Lanzhou 730046)
Abstract: Foot-and-mouth disease (FMD) is a highly infectious disease of head livestock, e.g. pigs, cattle and sheep, and may
cause enormous economic losses and serious negative social problems. It is ranked first in the A list of infectious diseases of animals in China. As a largest country for pigs, cattle, sheep and goats farming and a country with lots of neighboring countries, FMD prevention and control is not only crucial for the development of animal husbandry in China, but also of significance for the global FMD prevention and control. Nowadays, except some developed countries where FMD was eradicated and the FMD-free status has been maintained, prevalence and outbreaks of FMD happen in most of developing countries. China suffers a serious threat from the continued outbreak of FMD in neighboring countries. O/ME-SA/PanAsia, O/SEA/Mya-98 and A/ASIA/Sea-97, FMDV lineages epidemic in China in recent years, were came from neighboring countries. Based on the analyses of the epidemic lineages in
jia
文本框
返回目录

3548 中 国 农 业 科 学 48 卷
surrounding countries, frequency of outbreak and locations of circulation, O/PanAsia-2, A/Iran-05, Asia1/Sindh-08 and O/Ind-2001 are the epidemic lineages threatening China in the future. For decades, veterinary service systems at all levels have been constantly improved in China, the functions (e.g. scientific research, technical guidance, epidemic monitoring) of National FMD Reference Laboratory have been given full exerting and compulsory immunization and prevention-oriented policy have been proven to be highly effective. Based on the progressive control pathway for FMD (PCP-FMD) recommended by OIE (Office International des Epizootics), the effect of prevention and control in China is in the third stage of PCP-FMD. To promote the effect of FMD prevention and control into a higher level, the keys are the diagnostic testing technology and vaccines. The liquid-phase blocking enzyme linked immunosorbent assay (ELISA), non-structural protein 3ABC antibody detecting ELISA, multiplex reverse transcription-polymerase chain reaction (RT-PCR) and other independent research and development products have reached the world advanced level, strongly supporting the FMD diagnostic testing in China and also applied in North Korea, Vietnam and other countries. New generation of sequencing technology, colloidal gold and nanometre tracer tag immune chromatography, biological response effect of molecular detection and other more punctual and more convenient technologies are expected to achieve application in FMD diagnosis in the future. As the most widely used vaccine, FMD inactivated vaccine has the optimal immunization effect, apart from interference to differentiate between FMDV infected and vaccinated animals. Along with the progress of the immunology theory and genetic engineering technology, marker vaccine, live carrier vaccine, protein epitope vaccine, virus-like particles vaccine and other new type vaccines will be developed and applied in the future. In the last ten years, with its powerful technical advantages, such as directional changes in FMDV genome, reverse genetics promoted not only the research and development of marker vaccine and other new type of genetic engineering vaccines, but also FMDV basic research, such as host tropism, replication mechanism, receptor usage and innate immune response, and the basic research will vigorously promote the applied research of FMD prevention and control.
Key words: foot-and-mouth disease; epidemic status; new technologies for disease prevention and control; virus characteristics
0 引言
口蹄疫(foot-and-mouth disease, FMD)是由口蹄
疫病毒引起的急性、热性、高度接触性传染的动物疫
病。该病可快速远距离传播,侵染对象为猪、牛、羊
等主要畜种及其它偶蹄动物,易感者多达 70 余种。发
病动物的特征症状是口、鼻、蹄和母畜乳头等部位发
生水泡,或水泡破损后形成的溃疡或斑痂,表现流涎、
跛行和卧地,由此导致生产力大幅下降。口蹄疫传染性
强,发病率 100%,可造成巨额经济损失和社会政治负
面影响,素有“政治经济病”之称,国际动物卫生组织
(OIE)将该病列在法定上报的动物传染病之首,是国
际活畜及畜产品通关贸易必检的一类疫病,中国政府也
将其排在一类动物传染病的首位,充分显示了因该病危
害引起的各国重视程度[1]。 口蹄疫的病原为口蹄疫病毒( foot-and-mouth
disease virus, FMDV),有 A、O、C、Asia1、SAT1、SAT2 和 SAT3(South Africa type, SAT)共 7 个血清
型。虽然各型病毒引起的发病征候相同,但型间无交
叉免疫,病后康复或免疫动物仍可感染其它血清型病
毒而发病。因此,同一个地区可同时流行多个血清型
的口蹄疫。近 5 年来,全球除 C 型口蹄疫未见流行外,
其他 6 个血清型都在流行。A 型和 O 型流行面 广,
其次为 Asia1 型和 SAT2 型。A 型和 O 型口蹄疫遍布
亚洲、中东和非洲地区,散发于南美地区;Asia1 型
口蹄疫常驻于亚洲和中东大部分地区;SAT2 型口蹄
疫主要在非洲流行,也见于埃及和巴林等中东地区。
总的来看,亚洲、中东和非洲的大多数地区是口蹄疫
的重疫区。口蹄疫现状可概括为:有经济势力的发达
国家消灭了口蹄疫,享受无疫的惠利地位;经济状况
良好的国家能有效控制口蹄疫,正逐步建立无疫区,
企望获得无疫惠利;欠发达或社会秩序不稳定的国家,
口蹄疫频发,仍在遭受严重的危害。 口蹄疫病毒的血清型多,持续感染无症状带毒严
重,准确诊断血清型和精准监测隐性带毒,是防控该病
的首要环节。与生命领域的诊断监测技术发展同步,口
蹄疫诊断监测技术的发展历程可分为 3 个阶段,即早期
的补体结合试验和病毒中和试验,随后以酶联免疫吸附
反应(ELISA)为核心的病原和抗体检测方法,现今的
以聚合酶链反应(PCR)为基础的多种分子诊断技术。
3 个阶段相互衔接,技术互相补充和应证,形成了多方
位多层面的诊断监测手段。然而,随着生物技术的发展,
已有方法表现出了这样或那样的不足,或烦琐费时,或
没有达到十分精准的程度。未来诊断技术的发展是以轻
简化或精准为目标的新型定性定量技术。 疫苗免疫是防控口蹄疫的主要手段。早期使用的
疫苗主要是弱毒疫苗,因其存在与流行毒重组和更替
畜主返强的潜在风险,以及活毒的持续感染妨碍疫源

17 期 刘在新:全球口蹄疫防控技术及病原特性研究概观 3549
净化等缺陷,20 世纪 70 年代后逐渐被灭活疫苗取代。
口蹄疫灭活疫苗是目前使用的主要疫苗,在防控中发
挥了重要作用。欧洲经历了十多年的灭活疫苗接种后,
疫情得到了控制,多数国家达到了无疫标准。在此情
况下,欧共体虽于 1991 年停止了疫苗的使用,可疫苗
库中始终留存战备疫苗。南美也有一些国家使用灭活
疫苗,有效控制了疫情,取得了无口蹄疫国家地位。
但是,灭活疫苗生产时需要增殖大量的活病毒,残留或
多或少的病毒非结构蛋白。这种情况既不利于以检测非
结构蛋白抗体为金标准的感染和免疫鉴别诊断,也存在
病毒逸出车间的散毒风险。因此,未来疫苗的发展是研
制标记灭活疫苗和不动用活病毒的新型疫苗。 标记灭活疫苗的种毒可采用反向遗传操作技术构
建。在获得口蹄疫病毒全基因组 cDNA 片段的基础上,
利用基因操作缺失、插入、替换目标基因后,按基因
组的组成结构拼接 cDNA 片段,加上 T7 或真核的启
动子,构建病毒基因组的全长感染性 cDNA(即感染
性克隆),转染真核细胞, 经复制翻译,装配出完整
的病毒粒子。这一技术既可用于标记疫苗研究,也可
用于研究病原的生物学特性。可以说口蹄疫病毒反向
遗传学操作技术,是研究病原特性的里程碑技术。众
所周知,口蹄疫病毒是 RNA 病毒,很易发生变异,
衍化出了众多的变异株,出现了同型病毒交叉免疫程
度很低甚至无交叉免疫抗病现象;一部分病毒倾向感
染牛,而其变异株又倾向感染猪;一些病毒偏好结合
整联蛋白(intergrin)受体,而另一些病毒易与硫酸乙
酰肝素(heparan sulphate,HS)受体结合。诸如此类
的生物学特性揭示,反向遗传学技术显示了不可替代
的作用,与新一代测序技术结合,将在未来的口蹄疫
病原特性研究中发挥重要作用。 亚洲地区口蹄疫流行频繁,中国也难逃其害,近
年来不断有 O 型和 A 型传入和流行。中国是世界上
大的猪、牛、羊养殖国,与众多国家的陆地和海洋接
壤,拥有长且多样的边境线。在全球贸易频繁的当今,
中国口蹄疫的防控不仅对发展农牧业起关键作用,而
且对全球口蹄疫的控制具有重要的意义。本文就以上
主要内容展开叙述。
1 口蹄疫流行态势
世界动物卫生组织(office international des epizootics, OIE)根据地域和流行毒的分子特征,把全
球口蹄疫流行区域划分为 7 个流行循环圈(Pool)。
亚洲和中东,包括欧亚接壤地区位于 Pool1、Pool2 和
Pool3;非洲包括亚非接壤地区位于 Pool4、Pool5 和
Pool6,;南美洲位于 Pool7。中国大部分中东南部区
域属于 Pool1,西部区域属于 Pool2 和 Pool3。一个流
行循环圈内的同型流行毒,具有明显的遗传关系,且
时常在该区域内循环流行,把这一属性称为拓扑型
(topotype),以地域或罗马数字命名(如 ME-SA为中东-南亚拓扑型)。拓扑型中的流行毒,若遗传
脉络紧密,核酸碱基构成上相近,在聚类分析绘制的
遗传树上归于同一个末端分支,则属于一个谱系。每
个拓扑型中包括多个病毒谱系,命名流行毒常用法则
为:血清型/拓扑型/谱系/年代。划分流行毒的拓扑型
和谱系,可用于口蹄疫传播路线调查和疫源追踪,研
究病毒的遗传变异等特性,为该病的防治提供科学依
据[2-3]。 1.1 全球口蹄疫现状
近 5 年来,北美洲、大洋洲和大部分欧洲继续保
持无口蹄疫状态,亚洲和非洲口蹄疫流行频繁,部分国
家或地区疫情严重,南美洲一些地区取得了无疫地位,
而另一些地区呈散发态势,使得无疫地区的状态处于动
荡之中。2011 年无疫的英国暴发了口蹄疫,不过已在
短期内扑灭了疫情。总的格局是发达国家继续保持无口
蹄疫地位,发展中国家仍在遭受口蹄疫的危害。 1.1.1 欧洲 除俄罗斯外,欧洲近 5 年有保加利亚和
英国发生过口蹄疫。保加利亚与土耳其邻接,2009—2013 年发生过由 O/ME-SA/PanAsia-2 引起的疫情,
2011 年疫情严重,2013—2014 年未见报告疫情。2011年英国暴发了口蹄疫,但引发疫情的毒株不清,曾有
说法是疫苗车间泄露的病毒,因措施得当,很快扑灭
了疫情。之前的 2007 年在英国发生过由 O/EURO- SA/O1 引起的口蹄疫。 1.1.2 南美 呈散发,为 O 型和 A 型口蹄疫。2010年厄瓜多尔发生了由 O/EURO-SA 引起的疫情,A 型
流行毒出现了南美的新变异株,如引发巴西 2011—2012 年的口蹄疫。2014 年多巴哥岛暴发了口蹄疫,毒
株和血清型不清。 1.1.3 非洲 口蹄疫重疫区。因经济欠发达,社会秩
序不稳定,口蹄疫控制处于无序状态,疫情频繁严重。
野生动物,尤其是非洲水牛隐性感染多,带毒严重且
时间长,是 SAT1、SAT2 和 SAT3 型病毒的保藏者和
传播者,推测也是南非三型的适应者和发展地。邻近
地中海和红海地区,即东北非地区,疫情严重,主要
为O型和A型口蹄疫;西南非地区主要为A型和SAT2型口蹄疫,详细状况列于表 1。

3550 中 国 农 业 科 学 48 卷
表 1 2009-2014 年非洲地区口蹄疫疫情状态和病原血清型
Table 1 Outbreak status and virus serotype of foot and mouth disease in Africa during 2009-2014
国家或地区 Country or region
血清型 Serotype
病毒拓扑型 Virus topotype
病毒谱系 Virus lineage
疫情状况 Epidemic situation
博茈瓦纳 Botswana
SAT1 SAT2
SAT1/III SAT2/I, SAT2/II, SAT2/III
SAT1/III/WZ 连年流行。2012 年疫情严重,主要由 SAT2/III 病毒所致 Being endemic without intermission. The reason of large-scale outbreaks in 2012 was mostly infected by SAT2/III virus
喀麦隆 Cameroon
A SAT2
A/AFRICA SAT2/VII
连年流行。2014 年由 SAT2 引起了严重的疫情,至少有 40 余起。
2006 年之前也有 O 型(WEST AFRICA)流行 Being continuous endemic. SAT2 virus resulted in frequent endemic in 2014, at least 40 outbreaks. Serotype O (WEST AFRICA) was also endemic before 2006
刚果 Congo
A O
A/AFRICA O/EA-2
疫情不断 Being endemic without intermission
埃及 Egypt
O A SAT2
O/ME-SA,O/EA-3 A/AFRICA,A/ASIA SAT2/VII
2014 年 O、A 和 SAT2 型均有发生,2009 年以来,疫情很严重,
每年有约 40 起 Serotypes O, A and SAT2 were endemic in 2014. From 2009, frequent endemic, about 40 outbreaks every year
厄里特比亚 Eritrea
A O
A/AFRICA O/EA-3
疫情严重 Being continuous endemic
埃塞俄比亚 Ethiopia
O A SAT2
O/EA-3 A/AFRICA SAT2/XIII
疫情十分严重,2014 年 O、A 和 SAT2 型同时流行。2009 年前曾
有 SAT1 和 C 型 High-frequent outbreaks. Serotypes O, A and SAT2 were simultaneous endemic in 2014. Serotypes SAT1 and C were endemic before 2009
肯尼亚 Kenya
O A SAT1 SAT2
O/EA-1,O/EA-2 A/AFRICA SAT1/I SAT2/IV
疫情十分严重,由 SAT1/I 引起的疫情 为严重。2009 年前曾有 C型发生 Being high-frequent outbreaks. SAT1/I virus was mostly widely infected. Serotype C was endemic before 2009
利比亚 Liberia
O A SAT2
O/ME-SA,O/EA-3 A/ASIA SAT2/IV
O/ME-SA/ PanAsia-2
连年来,疫情特别严重 Being high-frequent outbreaks during last 5 years
尼日利亚 Nigeria
O A SAT2
O/EA-3 A/AFRICA SAT2/VII
多年的疫情严重 Being continuous endemic during many years
坦桑尼亚 Tanzania
O A SAT1 SAT2
O/EA-2 A/AFRICA SAT1/I SAT2/IV
疫情不断 Being endemic without intermission
赞比亚 Zambia
O SAT1 SAT2
O/EA-2 SAT1/I SAT2/III
多年的疫情严重 Being continuous endemic during many years
津巴布韦 Zimbabwe
SAT1 SAT2 SAT3
SAT1,SAT2,SAT3 不时有疫情出现。非洲水牛带毒常见 Being often endemic. The carriers of three SAT types virus always exist in the African buffalo population
1.1.4 中东 口蹄疫重疫区。因经济发展不平衡,社
会动荡,口蹄疫没有得到有效控制,疫情发生频繁,
流行时间长,病原衍化的谱系多。近 5 年来主要流行
A 型和 O 型口蹄疫,其次为 Asia1 型,与非洲邻接地
区有 SAT2 型流行,详细状况列于表 2。 1.1.5 亚洲 口蹄疫重疫区。由于国家和地区经济发
展不平衡,社会秩序不稳定,宗教信仰等原因,部分
地区动物卫生管理水平不高,口蹄疫流行严重且频繁,
流行时间长,病原衍化的谱系多。在贸易不断发展,
尤其是价值链驱动下,活畜及畜产品移动广泛,疫源
波及了无疫或疫情极少的国家。1997—2000 年亚洲发
生大规模的 O 型口蹄疫,打破了台湾地区 68 年、日
本 92 年、韩国 66 年无疫状态,冲击了中国 27 年内陆
偶发单一流行毒和单一血清型疫情的格局。近 5 年来,
东亚、东南亚和邻接中东地区,主要流行 A 型和 O 型
口蹄疫,流行毒谱系多样,详细状况列于表 3—5。

17 期 刘在新:全球口蹄疫防控技术及病原特性研究概观 3551
表 2 2009-2014 年中东地区口蹄疫疫情状态和病原血清型
Table 2 Outbreak status and virus serotype of foot and mouth disease in the Middle East during 2009-2014
国家或地区 Country or region
血清型 Serotype
病毒拓扑型 Virus topotype
病毒谱系 Virus lineage
疫情状况 Epidemic situation
巴林 Bahrain
A O Asia1 SAT2
A/ASIA O/ME-SA Asia1/ASIA SAT2/IV
A/ASIA/Iran0-5 O/ME-SA/PanAsia-2Asia1/ASIA/Sindh-08SAT2/IV/Ken-09
连年流行,其中 2012 年疫情频繁 Being continuous endemic, high-frequent outbreaks in 2012
埃及 Egypt
O A SAT2
O/ME-SA, O/EA-3 A/AFRICA, A/ASIA SAT2/VII
O/ME-SA/Sharqia-72A/AFRICA/G-VII A/ASIA/Iran-05 SAT2/VII/Ghb-12 SAT2/VII/Alx-12
疫情不断。2014 年疫情严重,O 型、A 型和 SAT2 型均在流行,O 型
由 O/EA-3 病毒、A 型由 A/AFRICA/G-VII、SAT2 型由 SAT2/VII/Alx-12病毒所致 Being endemic without intermission. Serotypes O, A and SAT2, infected by O/EA-3、A/AFRICA/G-VII、SAT2/VII/Alx-12 respectively, were simultaneous high-frequent outbreaks in 2014
伊朗 Iran
A O Asia1
A/ASIA O/ME-SA Asia1/ASIA
A/ASIA/Iran-05 O/ME-SA/PanAsia-2Asia1/ASIA/Sindh-08
疫情连年不断。2013-2014 年度 A 型、O 型和 Asia1 型在同时流行 Being endemic without intermission. Serotypes A, O and Asia1 were simultaneous endemic during 2013-2014
伊拉克 Iraq
A O Asia1
A/ASIA O/ME-SA Asia1/ASIA
A/ASIA/Iran-05 O/ME-SA/PanAsia-2Asia1/ASIA/Sindh-08
主发 A 型。2013-2014 年度 A 型和 Asia1 型在同时流行 Serotype A was mostly endemic. Serotypes A and Asia1 were simultaneous endemic during 2013-2014
以色列 Israel
A O
A/ASIA O/ME-SA
A/ASIA/Iran-05 O/ME-SA/PanAsia-2
流行 A 型和 O 型。2010 年后未见 A 型流行,2011 年疫情严重 Serotypes A and O were endemic. Serotype A was not outbreaks after 2010, but O type was frequent outbreaks in2011
约旦 Jordan
A O
A/ASIA O/ME-SA
A/ASIA/Iran-05 O/ME-SA/PanAsia-2
流行 A 型和 O 型,2011 年疫情较严重 Serotypes A and O were endemic, and. higher frequent outbreaks in 2014
黎巴嫩 Lebabon
A O
A/ASIA O/ME-SA
A/ASIA/Iran-05 O/ME-SA/PanAsia-2
流行 A 型和 O 型,2011 年疫情较严重 Serotypes A and O were endemic, and. higher frequent outbreaks in 2011
科威特 Kuwait
A O
A/ASIA O/ME-SA
A/ASIA/Iran-05 O/ME-SA/PanAsia-2
流行 A 型和 O 型,2011 年疫情较严重 Serotypes A and O were endemic, and. higher frequent outbreaks in 2011
巴勒斯坦 Palestinian
A O
A/ASIA O/ME-SA
A/ASIA/Iran-05 O/ME-SA/PanAsia-2
流行 A 型和 O 型,2011 年疫情较严重 Serotypes A and O were endemic, and. higher frequent outbreaks in 2011
沙特阿拉伯 Saudi Arabia
O O/ME-SA O/ME-SA/PanAsia-2O/ME-SA/Ind-2001d
近 5 年流行 O 型,未见其它型疫情报道 Serotype O was endemic during last 5 years, and no reporting other serotypes endemic
土耳其 Turkey
A O Asia1
A/ASIA O/ME-SA Asia1/ASIA
A/ASIA/Iran-05 O/ME-SA/PanAsia-2 Asia1/ASIA/Sindh-08
疫情严重。2014 年 A 型、O 型和 Asia1 型在同时流行 Being continuous endemic. Serotypes A, O and Asia1 were simultaneous endemic in 2014
阿拉伯联合 酋长国 United Arab Emirate
O O/ME-SA O/ME-SA/PanAsia-2O/ME-SA/Ind-2001
近 5 年只流行 O 型,由中东-南亚(ME-SA)拓扑型的 2 个谱系毒引发
Serotype O was only endemic during last 5 years, infected by 2 virus lineages of the ME-SA topotype
1.2 中国未来口蹄疫威胁
口蹄疫传播途径广泛,方式多样,国家和地区间
因动物跨界移动或贸易流通传递病原。中国边境线长
达 2 万多公里,与朝鲜、俄罗斯联邦、蒙古、哈萨克
斯坦、吉尔吉斯斯坦、塔吉克斯坦、阿富汗、巴基斯
坦、印度、尼泊尔、不丹、缅甸、老挝、越南等 14国陆路相通,与韩国、日本、菲律宾、文莱、马来西
亚及印度尼西亚等 6 国隔海相望。周边国家或地区,
口蹄疫疫情不断,造成了渗透式入侵和高压式威胁。
近年来中国流行的 O/ME-SA/PanAsia、O/SEA/Mya-98
和 A/ASIA/Sea-97 病毒均是传入的。从周遍国家或地
区流行的病毒、流行的频率和循环的区域位置判断, O/PanAsia-2、A/Iran-05、Asia1/Sindh-08 和 O/Ind-2001病毒可列入“高危病毒”,是未来中国针对境外口蹄
疫防控的重点。 O/PanAsia-2 病毒在 20 世纪 90 年代后期,逐渐由
PanAsia 毒株衍变而来,源头在印度。2003 年,该毒
株在尼泊尔和不丹发现,后来马来西亚也检测到该毒。
2007 年,在阿富汗、不丹、伊朗、以色列、哈萨克斯
坦和吉尔吉斯斯坦等国,相继由该毒引发疫情。同年,

3552 中 国 农 业 科 学 48 卷
表 3 2009-2014 年东亚地区口蹄疫疫情状态和病原血清型
Table 3 Outbreak status and virus serotype of foot and mouth disease in the East Asia during 2009-2014
国家或地区 Country or region
血清型 Serotype
病毒拓扑型 Virus topotype
病毒谱系 Virus lineage
疫情状况 Epidemic situation
中国 China
O A
O/ME-SA O/SEA A/ASIA
O/ME-SA/PanAsiaO/SEA/Mya-98 A/ASIA/Sea-97
近年来流行 O 型和 A 型。O/PanAsia 病毒引起多起疫情,2010-2014 年 O/Mya-98引发疫情,2013-2014 年又由 A/ Sea-97 引发疫情。2009 年前曾流行过 Asia1 型,
之后再未见病例 Serotypes A and O were endemic during last 5 years. O/PanAsia virus resulted in a lot of outbreaks. O/Mya-98 virus incursion and spread arose outbreaks during 2010-2014,and also A/ Sea-97 virus during 2013-2014. Serotype Asia1 was endemic before 2009, and no case thereafter
中国香港 Hong Kong
O O/CATHAY O/SEA
O/SEA/Mya-98 近年来发生 O 型口蹄疫,2012 年后为 O/CATHAY 病毒所致 Serotype O was endemic during last 5 years, but epidemic virus belonged to O/CATHAY topotype after 2012
日本 Japan
O O/ME-SA O/SEA
O/ME-SA/PanAsiaO/SEA/Mya-98
2000 年由 O/PanAsia 病毒引发疫情,2010 年由 O/Mya-98 病毒引起流行,2010年后再未出现疫情 O/PanAsia virus resulted in outbreaks in 2000. O/Mya-98 virus arose outbreaks in 2010, and no case thereafter
蒙古 Mongolia
O A
O/SEA O/ME-SA A/ASIA
O/SEA/Mya-98 O/ME-SA/PanAsiaA/ASIA/Sea-97
2013 年前只流行 O 型,之后又出现了 A 型。2014 年 O 型疫情由 O /PanAsia 病
毒所致 Serotype O was only endemic before 2013, and Serotype A thereafter. O/PanAsia virus resulted in outbreaks in 2014
朝鲜 North Korea
O A
O/SEA O/ME-SA
O/SEA/Mya-98 O/ME-SA/PanAsia
近年流行 O 型,未见 A 型的报告 Serotype O was endemic during last 5 years. Serotype A was no report
俄罗斯 Russian
O A
O/SEA O/ME-SA A/ASIA
O/SEA/Mya-98 O/ME-SA/PanAsiaA/ASIA/Iran-05 A/ASIA/Sea-97
近年来流行 O 型和 A 型,流行毒谱系多样,疫情严重 Serotypes A and O were endemic during last 5 years. Epidemic virus strains were different lineages, and arose a lot of outbreaks
韩国 South Korea
O A
O/SEA A/ASIA
O/SEA/Mya-98 A/ASIA/Sea-97
2010 年出现 A 型疫情,2011-2014 年的疫情均由 O/SEA/Mya-98 引起 Serotype A was endemic in 2010. Serotype O was endemic during 2011-2014, and infected by O/SEA/Mya-98
表 4 2009-2014 年东南亚地区口蹄疫疫情状态和病原血清型
Table 4 Outbreak status and virus serotype of foot and mouth disease in the South East Asia during 2009-2014
国家或地区 Country or region
血清型 Serotype
病毒拓扑型 Virus topotype
病毒谱系 Virus lineage
疫情状况 Epidemic situation
柬埔寨 Cambodia
O A
O/ME-SA A/ASIA
O/ME-SA/PanAsia A/ASIA/Sea-97
疫情不断。2010 前流行 A 型和 O 型,2010 年后基本为 O 型 Being continuous endemic. Serotypes A and O were endemic before 2010, and only serotype O thereafter
老挝 Laos
O A
O/ME-SA O/SEA A/ASIA
O/ME-SA/PanAsia O/SEA/Mya-98 A/ASIA/Sea-97
连年流行 A 型和 O 型 O/Mya-98 和 O/PanAsia 间隔年度交替出现,2014 年 O 型由 O/Mya-98所致 Serotypes A and O were continuous endemic. Serotype O was alternatelyinfected by O/Mya-98 and O/PanAsia at interval year, and by O/Mya-98 in 2014
马来西亚 Malaysia
O A
O/ME-SA O/SEA A/ASIA
O/ME-SA/PanAsia-2 O/SEA/Mya-98 A/ASIA/Sea-97
近 5 年流行 A 型和 O 型。2009 年发生过 O/ PanAsia-2 病毒的疫情,
2014 年 O 型和 A 均在流行 Serotypes A and O were endemic during last 5 years. One of epidemic O type virus strains was O/ PanAsia-2 before 2009. Serotypes A and O were simultaneous endemic in2014
缅甸 Myanmer
O A
O/SEA A/ASIA
O/SEA/Mya-98 A/ASIA/Sea-97
常年多发 A 型和 O 型。2009 年以后未见 Asia1 型报告 Serotypes A and O were always endemic. Serotype Asia1 was no report after 2009
泰国 Thailand
O A
O/SEA A/ASIA
O/SEA/Mya-98 A/ASIA/Sea-97
疫情多发。2014 年 O 型和 A 型均有发生,相比之下,A 型疫情较多
Being continuous endemic. Serotypes A and O were simultaneous endemic in 2014, but more serotype A
越南 Vietnam
O A
O/ME-SA O/SEA A/ASIA
O/ME-SA/PanAsia O/SEA/Mya-98 A/ASIA/Sea-97
疫情多发。2011-2014 年由 O/ME-SA/PanAsia 病毒引发疫情多见 Being continuous endemic. Serotypes O was more infected by O/ME-SA/PanAsia during 2011-2014
印度尼西亚、新加坡、菲律宾和文莱等无口蹄疫 Indonesia, Singapor and Brunei are FMD-free status

17 期 刘在新:全球口蹄疫防控技术及病原特性研究概观 3553
表 5 2009-2014 年亚洲其他地区口蹄疫疫情状态和病原血清型
Table 5 Outbreak status and virus serotype of foot and mouth disease in the other regions of Asia during 2009-2014
国家或地区 Country or region
血清型 Serotype
病毒拓扑型 Virus topotype
病毒谱系 Virus lineage
疫情状况 Epidemic situation
阿富汗 Afghanistan
O A Asia1
O/ME-SA A/ASIA Asia1/ASIA
O/ME-SA/PanAsia-2A/ASIA/Iran-05 Asia1/ASIA/Sindh-08
2009-2011 年疫情 为严重,有数百起,主要由 O/PanAsia-2 所致,
其次是 A/ Iran-05。2014 年主要流行 A 型,Asia1 型见于 2012 年 Being continuous endemic during 2009-1011, a lot of hundred outbreaks mostly arose by O/PanAsia-2 strain, following A/ Iran-05 strain. Serotype A was endemic in 2014, and Serotype Asia1 in 2012
孟加拉 Bengladesh
O
O/ME-SA
O/ME-SA/Ind-2001 2009 年流行 O 型,2010 年后流行情况和毒株不明 Serotype O was endemic in 2009, and epidemic status and virus strains no report after 2010
不丹 Bhutan
O
O/ME-SA
O/ME-SA/PanAsia-2O/ME-SA/Ind-2001
2009 年以来流行的疫情多由 O/ME-SA/Ind-2001 毒所致引起 FMD outbreaks mostly arose by O/ME-SA/Ind-2001 strain after 2009
哈萨克斯坦 Kazakhstan
O A
O/ME-SA A/ASIA
O/ME-SA/PanAsia O/ME-SA/PanAsia-2A/ASIA/Iran-05 A/ASIA/Sea-97
2009-2012 年由 O 型两谱系病毒引发疫情,2012 年后流行 A 型口蹄
疫,病原两谱系均有 Serotype O outbreaks mostly arose by two O type virus lineages during 2009-2012, and Serotype A outbreaks by two A type virus lineages after 2012
吉尔吉斯斯坦 Kyrgyzstan
O A
O/ME-SA A/ASIA
O/ME-SA/PanAsia-2A/ASIA/Iran-05
2010 年前流行 O 型,2011 年流行 A 型,之后未见报道 Serotype O was endemic in 2010, and Serotype A in 2011, and no reports thereafter
尼泊尔 Nepal
O O/ME-SA O/ME-SA/Ind-2001 疫情由 O/ Ind-2001 引起,未见 A 型的报告 Serotype O outbreaks arose by O/ Ind-2001 strain, and serotype A no report
巴基斯坦 Pakistan
O A Asia1
O/ME-SA A/ASIA Asia1/ASIA
O/ME-SA/PanAsia-2A/ASIA/Iran-05 Asia1/ASIA/Sindh-08
流行毒株多,疫情频繁。在发病动物中可同时检测到 O 型和 Asia1型病毒或 A 型和 Asia1 型病毒。2009 年之前广泛流行过 O/PanAsia病毒疫情,2014 年 A 型、Asia1 型 和 O 型均在流行 Being continuous endemic with widely infecting a lot of virus lineages. O type and Asia1 type virus could be detected, or A type and Asia1 type virus. Serotype O was wide endemic by infecting O/PanAsia strain before 2009. Serotypes A, Asia1 and O were simultaneous endemic in 2014
斯里兰卡 Srilanka
O O/ME-SA O/ME-SA/PanAsia-2O/ME-SA/Ind-2001/
多发 O 型。2014 年疫情较多,均由 O/Ind-2001 病毒引起 Serotype O was wide endemic. The more serotype O outbreaks mostly arose by O/Ind-2001 strain in 2014
印度,因社会欠发达和宗教信仰,使口蹄疫难以控制,O、A 和 Asia1 型均有流行,但未见详细的疫情报道。从周遍地区疫情状况推测,该地区流
行毒株多,疫情频繁 In India, because of not enough developed and religion, controlling FMD strategies could not carry out. Serotypes A, O and Asia1 were always endemic, all but no any reports. Presuming FMD outbreak infected by a lot of virus lineages often occurred being based on regions around
经土耳其引起色雷斯(巴尔干半岛东南部地区)的疫
情,并通过约旦、黎巴嫩、以色列、巴勒斯坦进入埃
及,疫情形势进一步升级。2008—2010 年,巴林、科
威特、巴基斯坦、巴勒斯坦、土耳其、沙特及印度等
国报道流行。2011 年初,经土耳其传入保加利亚,直
逼欧洲大陆。O/PanAsia-2 病毒似有定居中东地区之
势,近 5 年(2009—2014 年)在几乎所有中东国家
引发了频繁的疫情。从该毒起源、变异、传播路径、
传播速度和防控效果等方面综合分析,该毒株具有较
强的流行潜力。因此,O/PanAsia-2 病毒对中国的威
胁很大。
A/Iran-05 病毒于 2003 年在伊朗首次发现,2005年扩散至沙特阿拉伯,2006 年传播到巴基斯坦、土耳
其和约旦,其中土耳其和伊朗为严重流行地区。先后
在巴林(2008)、伊朗(2009)、巴基斯坦(2009)、
黎巴嫩(2009)、 伊拉克(2009)、科威特(2009)、
利比亚 (2009)和巴勒斯坦(2009)等国家监测到该
谱系病毒,且已成为 2009—2014 年引起中东及周遍地
区 A 型疫情连发的主要病原。虽然上述国家和地区在
积极免疫,但从流行现状来看,效果并不明显。主要
原因是国家、区域经济发展不平衡,军事争端常发,
疫病防控体系不健全,缺少互动联防机制,病原持续

3554 中 国 农 业 科 学 48 卷
存在,流行势头未减,且衍化出了 4 个亚系病毒。巴
基斯坦、阿富汗与中国新疆接壤,贸易往来频繁,活
畜和野生动物流动性较大,给 A/Iran-05 病原扩散提供
了便利,传入风险很大。 Asia1/Sindh-08病毒在2010年底至2011年初被引
起重视,先后在巴基斯坦巴哈瓦尔布尔市、旁遮普省
(与印度、伊斯兰地区交界)、卡拉奇市、信德省等
地区连续引发多起 Asia1 型口蹄疫疫情。2011 年 2 月,
巴林地区也检测到该病毒。2011 年 5 月,该毒进一步
扩散至伊朗和阿富汗,引起了欧洲国家的恐慌。2012年初正式命名为 Asia1/Sindh-08 毒株,2014 年在巴林、
伊朗、伊拉克、土耳其、阿富汗和巴基斯坦广泛流行,
是未来威胁中国的主要病原之一。 O/Ind-2001 病毒属 ME-SA 拓扑型,主要在南亚、
中东地区引发疫情。该毒起源于印度,在尼泊尔、不
丹和孟加拉等周围地区引起了广泛的流行,随后扩散
到中东的阿联酋等地区,至今在尼泊尔、不丹和孟加
拉一直存在,是引发这些地区 2009—2014 年疫情的主
要病原。印度、尼泊尔和不丹与中国陆地接壤,边贸
活动频繁,O/Ind-2001 病毒传入风险极大。
2 口蹄疫防控技术研究
一个多世纪以来,世界各国在与口蹄疫的斗争中,
总结出了行之有效的防控措施。其内容主要包括扑杀
患病及感染动物、疫苗免疫易感动物、限制动物及染
毒物品移动、消毒灭源、流行病学监测和预警风险分
析。根据疫情状态和经济实力,可采取有所侧重的策
略。归纳为扑灭根除策略、免疫控制策略和预防传入
策略。2007 年和 2010 年英国暴发口蹄疫后,采取了
扑灭根除策略,很快消除了疫情,短期内恢复了无疫
状态;以巴西为代表的南美几个国家,采取免疫控制
策略,取得了无疫区地位。中国近十几年采取的疫苗
强制免疫为主的控制政策,也卓有成效。2009 年后全
国范围内再无 Asia1 型疫情,海南岛和吉林永吉地区
达到了免疫无疫状态。 中国口蹄疫防控成绩的取得,是不断完善兽医服
务体系和制度的结果。近十几年来,逐渐健全了兽医
服务体系和透明的疫情公布制度;注重流行病学调查
和预报制度,并具备了快速鉴定疫情能力;设立了网
络式运行的国家参考实验室,并于 2009 年取得了 OIE口蹄疫参考实验室资质;以参考实验室为主体,系统
开展了疫苗、诊断和流行病学研究,并与各省市诊断
实验室畅通交流和沟通,指导其工作;制定了口蹄疫
防控应急预案,并实施了应急演练;建立了区域性养
殖企业登记和动物标识体系,以便监管动物移动和追
溯。这些体系和制度的内容,与 FAO/OIE 推荐的口蹄
疫防控措施实施的必要条件一致。相比而言,中国在
口蹄疫防控的条件建设、能力建设和防控成绩上,走
在了重疫区非洲和亚洲其它国家的前列。 作为 OIE 成员国,未来中国将参照 OIE 推荐的
PCP-FMD 路线图(图 1)推进口蹄疫防控。PCP-FMD是 FAO 和 OIE 推荐的指导原则,应用于有口蹄疫国
家或地区制定控制计划,也是 FAO/OIE 实施全球控制
口蹄疫策略的准则。PCP-FMD 不规定具体的控制过程
和实施方案,主要是评估防控结果,承认在不同的国
家和地区取得关键结果的 有效途径存在差异。依据
PCP-FMD 内容的评估规定,中国目前处于阶段 3。 推进口蹄疫防控效果进入更高阶段,核心技术是
诊断监测技术和疫苗。诊断监测技术和疫苗既是所有
防控措施实施的基础和支撑,又是评估防控效果不可
或缺的手段。从前述可以看出,全球口蹄疫频发,流
行态势仍很严峻,控制乃至消灭口蹄疫还需相当长的
过程,未来需要更加精准的诊断监测技术和安全有效
的疫苗。 2.1 诊断监测技术
改革开放以来,尤其是进入 21 世纪以来,中国的
口蹄疫诊断监测技术研发取得了可喜的成绩。以国家
参考实验室为龙头,研制了液相阻断 ELISA、非结构
蛋白 3ABC 抗体检测 ELISA、定型 ELISA、感染抗体
检测试纸条、多重 RT-PCR、定型 RT-PCR 和荧光定
量 RT-PCR 等多项技术[4-6],已在全国广泛应用,成为
中国口蹄疫诊断和疫情监测的支撑技术,也在朝鲜和
越南等国应用。这些技术的自主研发,弥补了缺项,
达到了欧美先进参考实验室的水平。随着生物技术的
发展,世界多个实验室正在研发或应用新一代测序技
术、胶体金和纳米示踪材料标记免疫层析技术和生物
反应效应分子检测等新技术,未来几年有望实现应用。 2.1.1 新一代测序技术 病原的遗传特性是唯一性
的,在疫病的诊断监测中具有中心作用,测定核酸序
列就可对疫病做出诊断。测序技术已发展到第 3 代,
也称为新一代测序技术。第一代测序技术平台是利用
双脱氧核糖核酸链末端终止法(Sanger 法)原理,于
20 世纪 90 年代,基于阵列毛细管电泳、荧光标记和
自动成像系统搭建的。进入 21 世纪后,基于基因芯片
的焦磷酸标记或合成法建立了第二代测序技术平台,
其原理是在 DNA 两端加上通用接头构建文库,每条

17 期 刘在新:全球口蹄疫防控技术及病原特性研究概观 3555
阶段Stage
0
阶段Stage
1
阶段Stage
2
阶段Stage
3
阶段Stage
4
阶段Stage
5
非免疫无口蹄疫Free withoutvaccination
从4到5申请官方认可 (OIE):非免疫无口蹄疫From 4 to 5Apply for officialstatus (OIE):free without vaccination
从3到4畜群中无地方性口蹄疫
流行From 3 to 4No endemic FMD indomestic livestock从2到3
实施有效的根除口蹄疫战略From 2 to 3Develop aggressive strategy to eliminate FMD
从1到2基于风险的口蹄疫控制计划From 1 to 2Risk-based FMD controlplan
从0到1按计划全面了解口蹄疫
流行病学From 0 to 1Comprehensive studyof FMD epidemiologyplanned
口蹄疫风险未被控制
无可靠数据FMD risk not controlledNo reliable information
风险认知与控制方案Identify risk and control options
实现基于风险的控制措施Implementrisk-based control
实行根除流行的控制措施
减少病毒循环Implement control strategyto eliminate circulation
维持无病毒循环和入侵Maintain zero circulation &Incursions
确保无病毒循环和入侵
不实施免疫Maintain zero circulation &Incursions; withdrawvaccination
口蹄疫防控状态分为 5 个渐进的阶段,用于国家或地区评价防控计划实施的成效 PCP describes a set of activities, divided into different stages, which can be used to evaluate the stage of advancement of a country or region in their FMD control and eradication programs
图 1 口蹄疫控制渐进路线图(引自 OIE 网页)
Fig. 1 The progressive control pathway for foot-and-mouth disease
文库片段再经桥式 PCR 扩增形成一簇,采用边合成边
测序反应,即在碱基延伸过程中,每个循环反应仅延
伸一个互补的碱基,经多个循环后,读取完整的核酸
序列。2010 年起,发展了第三代测序技术,该技术是
基于纳米微孔设备的合成法单分子测序技术。采用电
泳技术,驱动单分子逐一通过纳米孔,纳米孔的直径
非常小,仅允许单个分子通过,而 A、T、C、G 单个
碱基的带电性质不同,分辨电信号差异就可检出碱基
类别,实现测序。 第一代和第二代测序技术耗时,需要靶基因组知
识以扩增目标 DNA 片段,制备 DNA 文库,加接头,
这些过程会掺入错配的碱基,造成测序不准。第三代
测序技术快速,超高通量,不需要克隆扩增模板,排
除了碱基错配,可实现盲目的单分子实时测序(single
DNA molecules in real time, known as SMRT sequencing),2011 年平台开始上市,已用于基因组
学、转录组学、代谢组学和蛋白组学等系统生物学研
究。该技术测序速度快(前两代的万倍),读取序列
长(几千个碱基),精度高(达 99.9999%),可直接
测定 RNA 和甲基化的 DNA 序列,也可用于免疫学研
究,揭示免疫信号传导网络和 MicroRNA 调控等机理,
助于疫苗研发,将成为疫病防控的新手段。 新一代测序技术可用于口蹄疫的病原学、基因组
学、遗传进化和流行病学,以及宿主与病原互作研究,
为防控提供新的视觉和概念。近 5 年,已在该病的传
播路线追踪、遗传变异特征、疫源起因和暴发监控(疫
情传播走向)有所应用。口蹄疫病毒 RNA 复制因无
校对功能,碱基错配多,加之复制速度快,复制率高,

3556 中 国 农 业 科 学 48 卷
单一宿主内形成的是不同基因组的复杂混合体,即
“准种”(quasispecies)[7]。新一代测序技术能提供
大量的序列数据,分析不同类型样本中的一步变化
(step change),可做到检测单个畜体内病毒的移行
路线,不同畜体之间病毒感染的顺序,不同养殖场病
毒到达的先后,不同疫点病毒传播的次序,不同国家
或地区间病毒传入的时序[8-10],从时空上追踪和圈点
疫情走向,揭示病原变异规律。但该技术检测每个流
行病学单元(个体或群体)的小样本可能会出现抽样
偏差,分析全基因组数据时的软件算法和参考数据会
出现误差。相信软件的改进、化学的进步和选样策略
的优化会降低误差[11]。新一代测序技术在逐渐完善,
费用在降低,操作程序在简化,设备界面逐步友好,
兽医诊断实验室可用这一革命性的工具,预测口蹄疫
暴发的路径,采取灵活措施应对突发病情和不可预测
的病毒变异株。 2.1.2 免疫层析标记技术 免疫层析标记技术是以
微孔膜为固相载体,定位包被已知抗体或抗原,加入
待检样品后,因毛细管虹吸和微孔膜的渗滤作用,驱
使样品中抗原或抗体迁移,与膜上包被的抗体或抗原
相遇后结合,生成抗原抗体复合物,再经标记物与该
复合物反应,形成可见颜色结果,达到检测的目的。
该技术的应用以金标试制条为代表,与其它诊断技术
相比,操作简单快速,不需要特殊仪器设备,结果判
断直观,是典型的轻简化傻瓜式诊断方法,应用广泛。 免疫层析金标诊断技术起于 20 世纪 80 年代末,
国内外相继研制了口蹄疫定型试纸条和抗体检测试纸
条[12-15]。但金标层析技术仅限于定性或半定量检测,
眼观判断结果欠精准,存在灵敏度不高等缺陷,难以
满足定量化要求,研究者结合新示踪材料和新技术,
发展了新型免疫层析技术。新技术不存在胶体金标记
物需大量聚集才能显色的特点,而是以光、电、磁信
号放大系统提高检测灵敏度,减少样品的本底干扰,
有传统标记物无可比拟的优势。免疫层析标记技术正
朝着定量、高灵敏度、多标记物的方向发展。应用的
示踪材料包括稀土元素、荧光乳胶、荧光微球、量子
点、磁珠等。根据膜上包被的标记物不同,可分为上
转换纳米材料发光层析技术、荧光标记层析技术、量
子点层析技术和磁珠层析技术[16]。 上转换纳米材料发光层析技术是以上转换磷光材
料(upconverting phosphor,UCP)作为标记物(图 2)。UCP 是由 2 种稀土金属元素掺入氧化硫等惰性材料
中,构成能上转发光的纳米级示踪材料。因天然生物
材料不具上转发光特性,检测的发光信号不受环境的
(A) 检测抗原的上转换发光层析试验。一抗 IgG 和二抗 IgG 分别包被在层析膜上,为检测带和对照带。结合了上转换发光材料标记一抗 IgG 的抗原
能被检测带一抗 IgG 捕获实现检测 (B) 检测抗体的上转换发光层析试验。抗原和二抗 IgG 分别包被在层析膜上,为检测带和对照带。结合了上转换发光材料标记抗原的抗体能被检测
带的抗原捕获实现检测 (A) Upconverting phosphors (UCPs)-based lateral-flow( LF) assay for antigen detection. The first anti-antigen IgG and the second anti-antigen IgG were coated on the membrane as the test and control line, respectively. The presence of the target antigen that was captured by the first anti-antigen IgG was detected with UCP-conjugated to the same first anti-antigen IgG. (B) UCPs-based LF assay for antibody detection. The specific antigen and the anti-antigen IgG were coated on the membrane as the test and control line, respectively. Presence of the target antibody that was captured by the antigen coated on the test line was detected with UCPs conjugated covalently with the same antigen coated on the test line
图 2 上转换发光层析试纸条构造和反应原理(引自文献 17, Guo H C 等)
Fig. 2 Upconverting phosphor strip conformation and reaction elements

17 期 刘在新:全球口蹄疫防控技术及病原特性研究概观 3557
影响,故样品本底低而灵敏度高,很适合定量检测。
镧系稀土元素标记物,具有灵敏度高、操作简便、示
踪物稳定、无放射性污染、标记物储存时间长等优点,
利用其可建立不同模式的层析技术[17],用于微量定量
检测,发展前景广。荧光标记层析技术是以荧光乳胶
或荧光微球作为标记物,受激发光源刺激后发出荧光,
可有效排除背景色的干扰,经荧光仪检测荧光信号强
弱,实现定量检测,比传统金标层析技术敏感。为识
别不同待测分子,采用标记不同颜色荧光素,实现多
标记检测的目的。量子点(quantum dots,QDs)层析
技术是以主族Ⅱ—Ⅵ(如 CdSe)、Ⅲ—Ⅴ(如 InP、InAs、GaSe)、副族化合物和 Si 等元素组成的纳米颗
粒为标记物,相对于其他传统的有机荧光染料,QDs可发出不同颜色的荧光,分辨率高,荧光寿命长,适
合生物大分子的多标记、高通量定量检测。免疫磁珠
(immunomagnetic beads,IMB)层析技术是以磁性微
球为标记物,经标记蛋白捕获目标分子后,在振荡磁
场内测定磁信号大小,得出定量结果。磁性微球表面
积大,能结合多量的蛋白分子,可实现高敏感性和准
确性的多标记定量检测。但免疫磁珠制备工艺烦琐,
可调参数多,有待进一步发展。 未来新型免疫层析技术在口蹄疫诊断监测中将有
广泛的用途。可用表位肽、亚单位和病毒样粒子
(virus-like particles, VLPs)等抗原,以及单链抗体、
单特异性抗体和多抗互检,也可用标记的核酸适配体
(筛选或设计的 20—40nt 寡核苷酸)检测病毒核酸。 2.1.3 效应分子(标示物)监测技术 口蹄疫亚临床
感染是疫情发生的潜在风险,常用的监测方法是刮取
反刍动物的咽部分泌物(oro-pharyngeal fluid,OPF)和采集非反刍动物的淋巴结,用 RT-PCR 检测病毒核
酸。该法采集样品操作困难,PCR 易污染,需要新法
替换。感染口蹄疫病毒时,动物唾液中存在特异性
IgA, IgA 和 OPF 中持续存在的病毒或病毒基因相关,
非纯化疫苗的使用不影响 IgA 的检测,且易收样品,
用ELISA技术检测动物唾液中的 IgA可作为病毒存在
的效应分子[18]。但唾液样中高含量的脱落细胞、蛋白
酶和组织碎片,封闭抗原,表现非特异性反应。未来
可采用微量 ELISA 检测系统(micro-ELISA system),
提高检测特异性。该系统配置热激光显微探头和微流
通道,用包被微珠取代 96 孔板,提高了敏感性和特异
性[19]。也可采用纳米磁分离技术,降低非特异性反应。 2.1.4 生物传感器技术 生物传感器(biosensor)是利用生物物质(如酶、抗体、抗原、细胞器和核酸
适配体等)作为识别元件,把生化反应转变成可定量
的物理或化学信号,检测目标生物分子的系统。常采
用生化反应转换为电信号,放大或模数转换后,检测
目标分子及其浓度,其核心部分由生物分子识别元件
和信号转换元件组成。这类传感器是集生物化学、生
物技术、化学、物理和电子工程技术等学科的交叉系
统,研发精细度高。 细胞传感器(cell-based sensors)属生物传感器研
究的热点之一。细胞有许多分子识别的元件,包括受
体、离子通道、酶等,当有外界刺激时,能按照固有
的机制发生相应的生理活动,分析信号变化,达到检
测目的[20]。一种由细胞内钙离子和水母荧光蛋白(一
种钙离子敏感蛋白)组成信号转换元件的 B 淋巴细胞
传感器,代替了电极和光导纤维等电子元件转换器,
打破了经典传感器跨学科的限制,可开发能检测病原
体的 B 细胞传感器[21]。中国国内已有学者涉足检测口
蹄疫病毒的细胞传感器研究。 2.2 疫苗
口蹄疫疫苗免疫效力有赖于抗原的完整性和抗原
谱的广度,抗原可分为合成肽、表位蛋白、病毒样颗
粒和完整病毒 4 类。合成肽是由病毒结构蛋白 VP1 上
的一段主要抗原位点残基构成,可命名为“一维抗
原”。因化学合成技术所限,难以合成理想的结构性
长肽,因此,合成肽疫苗的抗原谱窄,免疫力有限。
表位蛋白是由病毒多个结构蛋白的抗原表位组成,可
看作“二维抗原”。参照多谱系病毒序列设计复合表
位蛋白基因,经原核或真核系统表达生成,免疫力优
于合成肽。病毒样颗粒是病毒的全部结构蛋白按自然
病毒结构组装而成,代表了病毒粒子的所有抗原位点
和构像,具有三维立体特征,暂名为“三维抗原”。
可参照多谱系病毒序列,修饰结构蛋白基因,经原核
或真核系统表达后组装而成,能模拟灭活的完整病毒
诱导免疫力,但限于目前技术水平,组装效率不高。
完整病毒抗原是经细胞培养增殖的完整病毒粒子,不
仅具有抗原位点完整性和构像性,而且包含了趋变的
“准种”抗原,命名为“四维抗原”。完整病毒抗原
经灭活后制备疫苗,灭活疫苗免疫效力 优,是目前
应用的主要疫苗,但存在干扰鉴别诊断的缺陷。随着
免疫学理论和基因工程技术进步,发展和应用标记疫
苗、活载体疫苗、表位蛋白疫苗和病毒颗粒样疫苗等
新型疫苗是未来的趋势。 2.2.1 标记疫苗 口蹄疫标记疫苗,或称之为 DIVA(differentiating infected from vaccinated animals)疫

3558 中 国 农 业 科 学 48 卷
苗,是缺失了病毒某段致病相关基因或优势表位基因
的弱毒苗或灭活疫苗。该类疫苗保持了弱毒苗或灭活
疫苗免疫效力 优的特性,且消除了常规疫苗多次免
疫干扰鉴别诊断的缺陷。利用反向遗传操作技术,即
可有目的缺失基因,构建标记病毒,也可同时修饰抗
原表位基因,拓展病毒抗原谱[22]。从这一点来看,标
记疫苗不仅是为了鉴别诊断,还可改善疫苗的免疫力
和抗原谱。 国外近 5 年开始研究口蹄疫标记疫苗。2010 年,
Fowler 等首先报道了缺失病毒 VP1 结构蛋白 G-H 环
的标记疫苗,虽然免疫牛能保护强毒攻击,且易鉴别
免疫畜群中的感染者[23]。但 G-H 环是重要的抗原表
位,缺失会损失疫苗的免疫原性[24];时隔 2 年,美国
梅岛动物疫病研究中心的Uddowla等发表了非结构蛋
白 3B 缺失和/或 3D 修饰的弱毒标记疫苗[25],能满足
免疫保护和鉴别诊断的目的,但弱毒有与流行毒重组
和毒力返强的风险,所以不主张使用[26]。中国研究口
蹄疫标记灭活疫苗与国外同期或早于国外,只是报道
见于 2014 年[27]。目前,中国农业科学院兰州兽医研
究所已完成了口蹄疫 O 型标记灭活疫苗的实验室研
究,进入了临床试验阶段,走在了世界的前列。 未来标记疫苗的发展以病毒抗原位点的解析为基
础,提高标记病毒免疫力,拓展抗原谱,建立配套的
精准鉴别诊断技术,研制多价标记疫苗和储备标记疫
苗。 2.2.2 活载体重组疫苗 口蹄疫活载体重组疫苗伴
随着基因工程产生。以某一病毒为载体,在其基因组
的非必须区插入口蹄疫病毒的抗原基因,构建成活的
重组病毒疫苗,接种动物后,目的抗原基因随载体病
毒基因表达,产生抗原,诱导特异性免疫应答。已报
道的病毒载体有痘病毒( poxvirus )、腺病毒
(adenovirus)和脊髓灰质炎病毒(poliovirus)等[28-32]。
活载体疫苗的特点是对靶动物细胞的感染效率高,进
入细胞的效率优于 DNA 疫苗,免疫效果较好;可载
荷几种不同的抗原基因,形成多价疫苗;生产时不需
要动用载体原始毒和口蹄疫病毒,无生物安全风险。
但活载体疫苗多次免疫,机体会产生针对载体抗原的
免疫反应,克服这一缺点可采用不同的免疫程序,例
如,初次免疫采用载体疫苗,再次免疫可用 DNA 疫
苗或亚单位疫苗,或者使用不同的病毒载体。 早报道的口蹄疫活载体疫苗研究是痘病毒作载
体[33],到目前为止免疫效果 好的是腺病毒活载体疫
苗[34-35]。20 世纪 90 年代中期开始,国外利用痘病毒
载体研究口蹄疫活载体疫苗,表达口蹄疫病毒完整结
构蛋白 P1 的重组痘苗,免疫小鼠后可产生高滴度的
中和抗体[28],但未见免疫靶动物的效果评价,推测其
免疫效果有限。到 90 年代后期,以美国梅岛动物疫病
研究中心为主,连续开展了腺病毒(Ad5)为载体的
口蹄疫活载体疫苗研究,历经十多年,到 2012 年,完
成了疫苗研制,通过了美国 FDA 认证,可作为应急疫
苗[34-35]。该疫苗是美国历史上通过注册的第一个口蹄
疫疫苗,OIE 推荐可在无疫国家或地区紧急使用。期
间,中国学者于 2008 年也报道了口蹄疫重组腺病毒活
载体疫苗的研究成果[36]。 口蹄疫活载体疫苗未来仍有较大的研发空间。表
达干扰素的重组腺病毒,可在免疫后 1 周内诱发早期
抗病毒保护[37],与口蹄疫重组腺病毒活载体疫苗配合
应用,或构建成一个载体疫苗,能降低早期感染率,
减慢疫病的传播速度。此外,在有效抗原结构研究的
基础上,设计精练的抗原基因,缩短插入基因片段,
可提高载体的表达量和免疫效果。利用在田间应用的
其它疫病弱毒疫苗株,研制多价活载体疫苗,实现多
重免疫,也是未来研究的方向之一。 2.2.3 表位蛋白疫苗 表位蛋白疫苗也称鸡尾酒式
疫苗,是病毒多个结构蛋白抗原表位及辅助性表位的
疫苗。依据病毒的抗原位点,设计多个抗原表位(包
括多谱系病毒抗原表位)基因序列,加上淋巴细胞辅
助性表位序列,构建表达框,经原核或真核表达蛋白
而成。与传统灭活疫苗相比,表位蛋白疫苗有多个优
点:生产过程不涉及活病毒,不存在散毒的危险;既
可随时根据流行毒株的变化调整表位序列,也可使用
不同的抗原序列,增加抗原的广谱性;配以适当佐剂,
经鼻内、口腔和皮肤等方式接种,产生黏膜免疫和全
身性免疫应答[38],易于感染和免疫鉴别诊断。 国外研究口蹄疫表位疫苗始于 20 世纪 90 年代
末[39],中国稍晚于国外。近年来,该疫苗逐渐成为研
究的热点。2008 年 Cubillos 等设计了一种树状肽,含
有 1 个拷贝的 T 细胞表位(3A 蛋白 21—35 位残基)
和 4 个拷贝的 B 细胞表位(VP1 蛋白 136—154 位残
基),两次免疫猪后,效果良好[40]。中国学者 2011年报道了一种嵌合表位蛋白疫苗,免疫猪后产生了接
近灭活疫苗的免疫效力[41],这一结果显示了复合表位
疫苗的良好前景。随后曹轶梅等报道了 3 个不同谱系
O 型病毒 B 细胞表位和通用性 T 细胞表位串联表达,
加入聚肌胞佐剂,免疫猪上调了细胞介导的免疫应答,
显著提高了保护不同谱系强毒攻击的水平[42-43]。

17 期 刘在新:全球口蹄疫防控技术及病原特性研究概观 3559
口蹄疫表位蛋白疫苗弥补了常规灭活疫苗的诸多
不足,成为口蹄疫疫苗发展的主要趋势之一。目前,
表位蛋白疫苗的研究多集中于线性表位,而缺乏对构
象表位的研究和筛选;如何选择高效安全的佐剂也是
未来的研究内容;此前过多的表位研究主要集中于 B 细胞表位和 Th 细胞表位,而缺乏对 CTL 细胞表位的
研究。这些问题的解决,将突现表位蛋白疫苗的优势,
作为一种安全、无毒和稳定的新型疫苗,可以预见今
后的发展前景广阔。 2.2.4 病毒样颗粒(VLPs)疫苗 病毒样颗粒疫苗
是由病毒的多个结构蛋白组装成与自然病毒粒子相似
的空心颗粒制备的疫苗。经原核或真核表达的病毒结
构蛋白,体外或胞内自组装成病毒样颗粒,不含病毒
核酸,因此没有感染和传播风险。病毒样颗粒与天然
病毒外部结构相似,如同天然病毒一样与 B 细胞受
体交联,诱导强烈的免疫应答[44]。VLPs 能整体进入
树突状细胞(DC)胞质内,被树突状细胞有效加工,以
交叉提呈或胞内直接加载的方式提呈给细胞毒性 T淋巴细胞(cytotoxic T lymphocytes, CTLs)[45],部分
VLPs还能直接诱导 DCs表型和功能分化成熟。因此,
VLPs 可诱导 CTL 反应[46]。组装 VLPs 需要几种蛋白,
表达不同蛋白的重组载体进入同一细胞的概率小,降
低了各蛋白之间的相互作用机会;构建多基因共表
达载体难度大,且同一载体上各基因的表达比例不
易控制;某些蛋白可能对表达细胞有毒性,不利于
表达[47]。这些因素降低了 VLPs 组装。虽然,已用多
种表达系统成功构建了 VLPs,但提高 VLPs 组装效
率仍面临许多挑战。 VLPs 疫苗是亚单位疫苗的进一步发展。自 1981
年 Kleid 等首开口蹄疫病毒基因工程亚单位疫苗研究
后[48],人们一直在寻找合适的抗原基因和表达系统,
以提高亚单位疫苗的免疫效果。1995 年,在总结早期
亚单位疫苗的效果不理想,可能是原核表达和抗原基
因少所致,Abrams 等用病毒结构蛋白(P1)和非结构
蛋白基因(2A-3C),经重组牛痘病毒载体,在真核
系统中正确组装了病毒样颗粒[49]。进入本世纪,病毒
样颗粒疫苗成为研究的热点,多用昆虫细胞组装病毒
样颗粒,但该系统表达组装的病毒样颗粒量少,成本
高。近几年,依据泛素化(SUMO)修饰能够维持蛋
白的水溶性,阻止蛋白无序聚集的理论,在原核系统
中实现了 VLPs 的正确组装[50],为利用原核表达系统
简单、方便和快捷的特性,研制可用的病毒样颗粒疫
苗奠定了基础。中国于 2005 年开始研究口蹄疫病毒样
颗粒疫苗[51-52],近 3 年取得了长足进步,尤其是利用
泛素化原核系统,研制的疫苗免疫效力与灭活疫苗相
当,达到了 OIE 和中国的口蹄疫疫苗要求标准[53]。未
来应解决的问题是如何有效放大生产工艺,提高 VLPs组装效率,降低成本等。
3 口蹄疫病原特性研究
口蹄疫病原特性研究的进步受益于反向遗传学技
术。口蹄疫反向遗传学操作允许改造基因组,加快了
试验速度。拯救得到的低代病毒来自单克隆,基因组
单一,相对于具有“准种”特性的自然分离毒,便于
阐释基因型与表型之间的关系。改造的病毒经体内(in vivo)途径直接观察生命周期中的病毒表型,比体外
表达蛋白、体外转录 RNA 和借助兔网织红细胞裂解
系统翻译蛋白等体外(in vitro)途径真实。利用反向
遗传学技术研究口蹄疫病原特性,为防治技术研发提
供理论基础。 3.1 口蹄疫病毒反向遗传操作技术发展历史
从1990年德国科学家Zibert首次报道口蹄疫病毒
反向遗传操作技术起,迄今走过了 25 年的发展历程。
根据该技术的成熟水平和应用状态,大致可分成 3 个
阶段。因各国研究水平存在差距,所以这 3 个阶段没
有精确的时间界限。 第一阶段 各种拯救病毒策略尝试期。第一株口
蹄疫病毒感染性 cDNA 克隆构建遇到了很大阻力,限
于当年的探针扫描分析序列和 Klenow 聚合酶扩增
DNA 片段难度大,不象今天常规测序和 PCR 技术成
熟。另一个阻碍感染性克隆构建的因素是,口蹄疫病
毒基因组有长的多聚胞嘧啶(poly(C))区(50—250nt),当时合成困难。 终,Zibert 借助酵母系统获得了含
poly(C)32 的感染性 cDNA 克隆,并拯救到活病毒,但
含 poly(C)25 的克隆没有拯救到活病毒[54]。可是,时
隔 3 年,Rieder 等用 poly(C)2 的感染性克隆拯救了病
毒[55]。这些现象表明,poly(C)的长度与拯救病毒有关,
但也与病毒株基因组骨架等因素有关。为简化操作,
提高拯救病毒效率,各国研究人员又尝试了多种拯救
策略。选用不同的 RNA 聚合酶,采取体外转录或体
内转录,用脂质体转染或电转染,“转录”是各种拯救
策略的焦点。现大多采用 T7 RNA 聚合酶及其启动子,
既可体内转录也可体外转录。脂质体转染效率相对较
低,电转染虽然转染效率较高,但对细胞的损伤大。
除了转染细胞拯救病毒外,Baranowski 等将体外转录
的全长 RNA 接种小鼠拯救了病毒,借助小鼠体内拯

3560 中 国 农 业 科 学 48 卷
救方法绕过了细胞的约束,使这株无法在细胞上成活
的突变毒,在小鼠上拯救成功[56],拓宽了拯救病毒的
方式。 第二阶段:反向遗传操作技术平台建立期。这一
时期的第一个特点是,构建病毒感染性 cDNA 克隆的
技术成熟,技巧多样。用 cDNA 末端快速扩增法(rapid amplification of cDNA end, RACE)获得病毒基因组 5′端准确序列,扩增病毒基因组 5′端的上游引物中加入
启动子序列,3′端的下游引物中引入 poly(A)序列,采
用融合 PCR 得到含 poly(C)的片段,在 T7 启动子和基
因组序列之间添加锤头状核酶( hammer head ribozyme)序列,以准确切除转录本 5′端的外源 RNA序列。另一个特点是,各血清型经典毒株的感染性克
隆平台建立成功。不同国家之间互赠感染性克隆质粒
等试验材料,在保证生物安全的前提下促进了平台的
建立。德国构建的第一个口蹄疫病毒 O 型感染性克隆
赠予英国、西班牙,英、西两国在此基础上,又有所
发展,并于 2010 年建成了 C 型病毒的感染性克隆;
美国梅岛动物疫病研究中心建立了A型病毒的反向遗
传操作技术平台,2004 年南非的 van Rensburg 等在美
国借助已建的平台,构建了 SAT2 型病毒的感染性克
隆。目前,除 SAT1 和 SAT3 两种血清型外,其它 5个血清型病毒的反向遗传操作技术平台均已建成。
2004 年,中国科研人员成功构建了口蹄疫 O 型病毒的
感染性克隆[57],成为中国第一株、世界第三株口蹄疫
病毒感染性克隆。目前,已建立了反向遗传操作技术
平台,可操作 A 型、O 型和 Asia1 型病毒[58-59]。此外,
阿根廷、印度等国也相继发展了本国流行毒的感染性
克隆。 第三阶段:反向遗传操作技术平台应用期。无论
是研究病毒遗传变异、与宿主互作、致病机理的基础
研究,还是发展候选疫苗株,口蹄疫病毒反向遗传操
作技术平台发挥了重要作用。具体内容在下一部分叙
述。 3.2 利用口蹄疫病毒反向遗传操作技术研究病原特
性
口蹄疫病毒感染性克隆建立后 初十余年,研究
集中于致弱、嵌合基因工程疫苗候选株的构建和评价。
2004 年后,兴起反向遗传学的病原特性研究。中国在
2004 年建成第一株自有病毒的感染性克隆,赶上了反
向遗传学研究的步伐。近十年间,该技术在口蹄疫病
毒宿主嗜性、复制机制、受体利用和先天性免疫应答
等方面做出了突出成绩。
利用反向遗传操作技术的第一个基础课题是病毒
的宿主嗜性问题。1997 年,由 Cathay 拓扑型病毒引
发中国台湾地区的口蹄疫疫情,几乎摧毁了养猪业,
但是,牛却在疫情中幸免[60]。这一明显的宿主嗜性需
要科学的解释。美国梅岛利用 A 型病毒感染性克隆得
到了答案,原因是引发台湾猪疫情的病毒 O/Taiwan/ 1997,其非结构蛋白 3A 缺失 93—102 位氨基酸[61,62]。
2013 年,考虑到之前的研究过程过于复杂,致使对结
果的证明有不确实的地方,梅岛中心又以 O1/Campos/ Brazil/1958 的感染性克隆为骨架,比较了 3A 蛋白
87—106 位缺失后宿主嗜性的变化,确实地证明了 3A蛋白的缺失降低了病毒对牛的致病力[63]。鉴于 Cathay拓扑型病毒 3A 蛋白存在自然缺失的特点,2012 年
中国率先构建成功了该型病毒的感染性克隆[64],拯
救的基因工程病毒 3A 区 93—102 位的缺失十分稳定,
对猪的致病力比牛高至少 1 000 倍。2010—2011 年,
O 型 Mya-98 谱系病毒肆虐东亚,中国猪牛养殖业也
遭重创[65]。笔者所在实验室紧急响应,在国际上率先
构建成功了 2 株 Mya-98 病毒的感染性克隆平台。利
用该平台,进一步证实了 3A 蛋白 93—102 位缺失是
一种对牛致弱性变异,并且第一次明确提出:3A 蛋白
93—102 位的缺失并非完全决定口蹄疫病毒的宿主嗜
性,该位置以外的区域也起作用[66]。这些决定宿主嗜性
的区域是如何发挥作用的?2012 年,西班牙同行证实
Q44 和 D32 两个关键氨基酸影响 3A 蛋白的二聚化,
终影响病毒嗜性[67];2013 年,美国梅岛中心发现
3A 蛋白与宿主蛋白 DCTN3 结合,促进病毒复制,而
突变的 3A 蛋白不能与 DCTN3 结合,病毒复制能力降
低,对牛致病力减弱[68]。 利用反向遗传操作技术的第二个基础课题是病毒
与受体的结合问题。整联蛋白(Integrin)和硫酸乙酰
肝素(HS)是口蹄疫病毒的常用受体[69],此外,还存
在第三类受体或第三种病毒侵入细胞的机制。整联蛋
白与病毒 VP1 的 RGD 基序相互特异识别,曾是口蹄
疫病毒入侵宿主时的公认法则。2011 年,笔者实验室
发现 Asia1 型病毒经不同宿主传代,RGD 可衍化为
RDD 和 RSD。利用 Asia1/CHN/2005 毒株感染性克隆,
证明这些基序对于侵染 BHK-21 细胞以及对猪、牛致
病力没有明显差异[58]。该成果拓宽了认知口蹄疫病毒
利用整联蛋白受体的知识。1996 年,英国 Pirbright 研究所首次明确指出 HS 是口蹄疫病毒的第二类受体[70]。
2010 年和 2014 年,白兴文等利用反向遗传操作技术
平台,鉴定了 Q2080 和 K1083 是决定 PanAsia(泛亚)

17 期 刘在新:全球口蹄疫防控技术及病原特性研究概观 3561
谱系病毒利用 HS 的关键氨基酸位点[59]。除已知的两
类受体外,2003 年赵启祖等以 A 型和 O 型病毒的感
染性克隆为工具,阐明病毒外壳上发生从带负电氨基
酸到带正电氨基酸的变异(如 E1083K),能增强与
HS 结合的能力,但是能与 HS 结合并不一定利用 HS受体,口蹄疫病毒侵染细胞和动物时可能存在新的机
制[71]。由此可见,口蹄疫病毒入侵宿主的机制仍有未
解之迷,功能性新受体到底是哪种分子,病毒变换受
体利用的变异规律是怎样的。这些问题的揭示,反向
遗传操作技术将发挥关键作用。 除嗜性、受体外,口蹄疫病毒准种的复制机制也
是对病原特性研究的重要课题。一直以来,西班牙
CSIC-UAM 实验室引领着这一领域的发展,但是近期
中国也取得了不错的成果。于力等借助 Asia1 型病毒
感染性克隆,用利巴韦林筛选鉴定了病毒 RNA 聚合
酶(RNA dependent RNA polymerase, RdRp)的保真性
变异,保真性提高后病毒准种多样性减少,适应能力
下降,表现为致病力减弱[72]。这一研究成果反向揭示
了口蹄疫病毒高变异率对其生存能力的正面价值。 宿主对病毒的免疫应答机制一直是科研人员热衷
的研究课题。借助口蹄疫病毒感染性克隆,各国同行
发展了多种基因工程疫苗候选株,并对它们诱发的宿
主免疫应答做了评价。但是,宿主抗病毒先天性免疫
方面的工作相对较少。2014 年,中国的一项研究在这
方面做出了积极探索。利用 O/Tibet/CHN/1999 的感染
性克隆,杜以军等聚焦 JAK-STST信号通路,发现 3Cpro
通过降解核转运受体蛋白 KPNA1 ,进而阻止
STAT1/STAT2 入核, 终拮抗 IFN-I 的产生[73]。该结
果为宿主应答口蹄疫病毒免疫信号网络途径研究提供
了数据。 口蹄疫病毒反向遗传技术平台应用越来越广。一
项技术在实验室的常规化正说明它将发挥日益重要的
作用,利用该技术平台提供的病毒体内(in vivo)活动
重现,以鲜活的方式揭示病毒生命活动规律,阐明病毒
入侵宿主机制、病毒复制与嗜性关系、病毒感染与宿主
免疫应答调控机理,为口蹄疫防控提供科学依据。
致谢:本文在完成过程中,孙世琪、郭慧琛、何继军、
张强、卢曾军、曹轶梅、李平花、付元芳和姜韶东等同仁提
供了宝贵的素材,在此谢忱。
References
[1] 谢庆阁. 口蹄疫. 北京:中国农业出版社, 2004: 1-15.
Xie Q G. Foot-and-Mouth Disease. Beijing: China Agriculture Press,
2004:1-15. (in Chinese)
[2] WRLFMD: The foot-and-mouth disease home page. http://www.wrlfmd.org
[3] OIE: World animal health information system. http://www.oie.int/wahis
[4] Lu Z J, Cao Y M, Bao H F, Qi S Y, Guo J H, Shang Y J, Jiang T, Ma J
W, Liu Z X, Yin H. Techniques development in China for
foot-and-mouth disease diagnosis. Transboundary and Emerging
Disease, 2008, 55: 196-199.
[5] Fu Y F, Cao Y M, Sun P, Bao H F, Bai X W, Li P H, Li D, Lu Z J, Liu
Z X. Development of a dot immunoblot method for differentiation of
animals infected with foot-and-mouth disease virus vaccinated
animals using non-structural preteins expressed prokaryotically.
Journal of Virological Methods, 2011, 171: 234-240.
[6] Fu Y F, Lu Z J, Li P H, Cao Y M, Sun P, Tian M N, Wang N, Bao H F,
Bai X W, Li D, Chen Y L, Liu Z X. Development of a blocking
ELISA based on a monoclonal antibody against a predominant epitope
in non-Structural protein 3B2 of foot-and-mouth disease virus for
differentiating infected from vaccinated animals. PLoS One, 2014,
9(11): e111737.
[7] Lauring A S, Andino R. Quasispecies theory and the behavior of RNA
viruses. PLoS Pathogens, 2010, 6(7): e1001005.
[8] Wright C F, Morelli M J, Thébaud G, Knowles N J, Herzyk P, Paton D
J, Haydon D T, King D P. Beyond the consensus: dissecting
within-host viral population diversity of foot -and-mouth disease virus
by using next generation genome sequencing. Journal of Virology,
2011, 85(5): 2266-2275.
[9] Morelli M J, Wright C F, Knowles N J, Juleff N, Paton D J, King D P,
Haydon D T. Evolution of foot-and-mouth disease virus intra-sample
sequence diversity during serial transmission in bovine hosts.
Veterinary Research, 2013, 44: 12.
[10] Logan G, Freimanis G L, King D J, Valdazo-González B, Bachanek-
Bankowska K, Sanderson N D, Knowles N J, King D P, Cottam E M.
A universal protocol to generate consensus level genome sequences
for foot-and-mouth disease virus and other positive-sense
polyadenylated RNA viruses using the Illumina MiSeq. BMC
Genomics, 2014, 15: 828.
[11] Van Borm S l, Belák S, Freimanis G. Veterinary Infection Biology:
Molecular Diagnostics and High-Throughput Strategies. Methods in
Molecular Biology, 2015, 1247: 415-436.
[12] Reid S M, Ferris N P, Brüning A, Hutchings G H, Kowalska Z,
Akerblom L. Development of a rapid chromatographic strip test for
the pen-side detection of foot and mouth disease virus antigen.
Journal of Virological Methods, 2001, 96(2): 189-202.

3562 中 国 农 业 科 学 48 卷
[13] Sugimura T, Suzuki T, Chatchawanchonteera A, Sinuwonkwar P,
Tsuda T, Murakami Y. Application of latex beads agglutination test for
the detection of the antibody against virus-infection-associated (VIA)
antigen of Foot and Mouth Disease (FMD) virus. Journal of
Veterinary Medical Science, 2000, 62 (7): 805-807.
[14] 蒋韬, 梁仲, 陈涓, 何继军, 吕律, 马维民, 刘在新, 刘湘涛. 口蹄
疫病毒 O、A、AsiaⅠ型定型诊断胶体金免疫层析方法的建立. 中
国农业科学, 2008,41(11): 3801-3808.
Jiang T, Liang Z, Chen J, He J J, Lü L, Ma W M, Liu Z X, Liu X T.
Development of a rapid gold immunochromatographic strip test for
the diagnosis of foot-and-mouth disease virus O, A and Asia I type.
Scientia Agricultura Sinica, 2008, 41(11): 3801-3808. (in Chinese)
[15] Wu L, Jiang T, Lu Z J, Yang Y M, Sun P, Liang Z, Li D, Fu Y F, Cao Y
M, Liu X T, Liu Z X. Development and validation of a prokaryotically
expressed foot-and-mouth disease virus nonstructural protein
2C’3AB-based immunochromatographic strip to differentiate between
infected and vaccinated animals. Virology Journal, 2011, 8: 186.
[16] 任志奇, 吴英松, 刘天才. 新型免疫层析技术的研究进展. 广东医
学, 2013, 34(2): 312-314.
Ren Z Q, Wu Y S, Liu T C. Progress of new immunochromatographic
technologies. Guangdong Medical Journal, 2013, 34(2): 312-314. (in
Chinese)
[17] Guo H C, Sun S Q. Lanthanide-doped upconverting phosphors for
bioassay and therapy. Nanoscale, 2012, 4: 6692-6706.
[18] Parida S, Anderson J, Cox S J, Barnett P V, Paton D J. Secretory IgA
as an indicator of oro-pharyngeal foot-and-mouth disease virus
replication and as a tool for post vaccination surveillance. Vaccine,
2006, 24(8): 1107-1116.
[19] Dong Y Y, Xu Y, Liu Z X, Fu Y F, Ohashi T, Tanaka Y, Mawatari K,
Kitamori T. Rapid screening swine foot-and-mouth disease virus using
micro-ELISA system. Lab on a Chip, 2011, 11(13): 2153-2155.
[20] McFadden P. Tech. Sight. Biosensors. Broadband biodetection:
Holmes on a chip. Science, 2002, 297(5589): 2075-2076.
[21] Rider T H, Petrovick M S, Nargi F E, Harper J D, Schwoebei E D,
Mathews R H, Blanchard D J, Bortolin L T, Young A M, Chen J,
Hollis. A B cell-based sensor for rapid identification of pathogens.
Science, 2003, 301(5630): 213-215.
[22] Li P H, Bai X W, Sun P, Li D, Lu Z J, Cao Y M, Fu Y F, Bao H F,
Chen Y L, Xie B X, Liu Z X. Evaluation of a genetically modified
foot-and-mouth disease virus vaccine candidate generated by reverse
genetics. BMC Veterinary Research, 2012, 8: 57.
[23] Fowler V L, Knowles N J, Paton D J, Barnett P V. Marker vaccine
potential of a foot-and-mouth disease virus with a partial VP1 G-H
loop deletion. Vaccine, 2010, 28: 3428-3434.
[24] Grubman M J, Baxt B. Foot-and-mouth disease. Clinical Microbiology
Reviews, 2004, 17(2): 465-493.
[25] Uddowla S, Hollister J, Pacheco J M, Rodriguez L L, Rieder E. A safe
foot-and-mouth disease vaccine platform with two negative markers
for differentiating infected from vaccinated animals. Journal of
Virology, 2012, 86: 11675-11685.
[26] Martin W B, Edwards L T. A field trial in South Africa of an
attenuated vaccine against foot-and-mouth disease. Research in
Veterinary Science, 1965, 6: 196-201.
[27] Li P H, Lu Z J, Bai X W, Li D, Sun P, Bao H F, Fu Y F, Cao Y M,
Chen Y L, Xie B X, Liu Z X. Evaluation of a 3A-truncated
foot-and-mouth disease virus in pigs for its potential as a marker
vaccine. Veterinary Research, 2014, 45: 51.
[28] Berinstein A, Tami C, Taboga O, Smitsaart E, Carrillo E. Protective
immunity against foot-and-mouth disease virus induced by a
recombinant vaccinia virus. Vaccine, 2000, 18: 2231-2238.
[29] Moraes M P, Mayr G A, Mason P W, Grubman M J. Early protection
against homologous challenge after a single dose of replication-
defective human adenovirus type 5 expressing capsid proteins of
foot-and-mouth disease virus(FMDV) strain A24. Vaccine, 2002,
20(11-12): 1631-1639.
[30] Kitson J D A, Burke K L, Pullen L A, Belsham G J, Almond J W.
Chimeric polioviruses that include sequences derived from two
independent antigenic sites of foot-and-mouth diseases virus (FMDV)
induce neutralizing antibodies against FMDV in guinea pigs. Journal
of Virology, 1991, 65(6): 3068-3075.
[31] 童武, 徐彦召, 周艳君, 姜一峰, 张善瑞, 王亚欣, 朱建平, 虞凌
雪, 孙晶, 陈焕春, 童光志. 表达 O 型口蹄疫病毒保护性抗原表
位重组 PRRSV 的构建及其鉴定. 生物工程学报, 2012, 28(12):
1431-1440.
Tong W, Xu Y Z, Zhou Y J, Jiang Y F, Zhang S R, Wang Y X, Zhu J P,
Yu L X, Sun J, Chen H C, Tong G Z. Construction and identification
of a recombinant PRRSV expressing protective antigens of type O
foot-and-mouth disease virus. Sheng Wu Gong Cheng Xue Bao, 2012,
28(12): 1431-1440. (in Chinese)
[32] Huang L, Zhang F, Tang Q, Wei Y, Wu H, Guo L, Fu Y, Liu C. A
recombinant porcine circovirus type 2 expressing the VP1 epitope of
the type O foot-and-mouth disease virus is infectious and induce both
PCV2 and VP1 epitope antibodies. Applied Microbiology and
Biotechnology, 2014, 98(22): 9339-9350.
[33] Abram C C, King A M Q, Belsham G J. Assembly of foot-and-mouth
disease virus empty capsid synthesized by a vaccinia virus expression

17 期 刘在新:全球口蹄疫防控技术及病原特性研究概观 3563
system. Journal of General Virology, 1995, 76: 3089-3098.
[34] Pacheco J M, Brum M C, Moraes M P, Golde W T, Grubman M J.
Rapid protection of cattle from direct challenge with foot-and-mouth
disease virus (FMDV) by a single inoculation with an adenovirus-
vectored FMDV subunit vaccine. Virology, 2005, 337(2): 205-209.
[35] Brake D A, McIlhaney M, Miller T, Christianson K, Keene A, Lohnas
G, Purcell C, Neilan J, Schutta C, Barrera J, Burrage T, Brough DE,
Butman B T. Human adenovirus-vectored foot-and-mouth disease
vaccines: establishment of a vaccine product profile through in vitro
testing. Developments in Biologicals, 2012, 134: 123-133.
[36] Lu Z J, Bao H F, Cao Y M, Sun P, Guo J H, Li P H, Bai X W, Chen Y
L, Xie B X, Li D, Liu Z X, Xie Q G. Protection of guinea pigs and
swine by a recombinant adenovirus expressing O serotype of
foot-and-mouth disease virus whole capsid and 3C protease. Vaccine,
2008, 26S: G48-G53.
[37] Kim S M, Kim S K, Park J H, Lee K N, Ko Y J, Lee H S, Seo M G,
Shin Y K, Kim B. A recombinant adenovirus bicistronically
expressing porcine interferon-α and interferon-γ enhances antiviral
effects against foot-and-mouth disease virus. Antiviral Research, 2014,
104:52-58.
[38] Zhang Z W, Pan L, Ding Y Z, Zhou P, Lv J L, Chen H T, Fang Y Z,
Liu X S, Chang H Y, Zhang J, Shao J J, Lin T, Zhao F R, Zhang Y G,
Wang Y L. Efficacy of synthetic peptide candidate vaccines against
serotype-A foot-and-mouth disease virus in cattle. Applied Microbiology
and Biotechnology, 2014, 09 November, published online.
[39] Volpina O M, Surovoy A Y, Zhmak M N, Kuprianova M A, Koroev D
O, Chepurkin A V, Toloknov A S, Ivanov V T. A peptide construct
containing B-cell and T-cell epitopes from the foot-and-mouth disease
viral VP1 protein induces efficient antiviral protection. Vaccine, 1999,
17(6): 577-584.
[40] Cubillos C, de la Torre B G, Jakab A, Clementi G, Borrás E, Bárcena J,
Andreu D, Sobrino F, Bianco E. Enhanced mucosal immunoglobulin
A response and solid protection against foot-and-mouth disease virus
challenge induced by a novel dendrimeric peptide. Journal of Virology,
2008, 82(14): 7223-7230.
[41] Shao J J, Wong C K, Lin T, Lee S K, Cong G Z, Sin F W, Du J Z, Gao
S D, Liu X T, Cai X P, Chang H Y, Liu J X. Promising
Multiple-epitope recombinant vaccine against foot-and-mouth disease
virus type O in swine. Clinical and Vaccine Immunology, 2011, 18(1):
143-149.
[42] Cao Y M, Lu Z J, Li Y L, Sun P, Li D, Li P H, Bai X W, Fu Y F, Bao
H F, Zhou C X, Xie B X, Chen Y L, Liu Z X. Poly (I: C) combined
with multi-epitope protein vaccine completely protects against
virulent foot-and-mouth disease virus challenge in pigs. Antiviral
Research, 2013, 97: 145-153.
[43] Cao Y M, Lu Z J, Li D, Fan P J, Sun P, Bao H F, Fu Y F, Li P H, Bai X
W, Chen Y L, Xie B X, Liu Z X. Evaluation of cross-protection
against three topotypes of serotype O foot-and-mouth disease in pigs
vaccinated with multi-epitope protein vaccine incorporated with
poly(I:C). Veterinary Microbiology, 2014, 168: 294-301.
[44] Chackerian B. Virus-like particles: flexible platforms for vaccine
development. Expert Review of Vaccines, 2007, 6(3): 381-390.
[45] Ludwig C, Wagner R. Virus-like particles-universal molecular tool
boxes. Current Opinion in Biotechnology, 2007, 18(6): 537-545.
[46] Gamvrellis A, Leong D, Hanley J C, Xiang S D, Mottram P, Piebanski
M. Vaccines that facilitate antigen entry into dendritic cells.
Immunology and Cell Biology, 2004, 82(5): 506-516.
[47] Dong H, Guo H C, Sun S Q. Virus-like particles in picornavirus
vaccine development. Applied of Microbiology and Biotechnology,
2014, 98: 4321-4329.
[48] Kleid D G, Yansura D, Small B, Dowbenko D, Moore D M, Grubman
M J, McKercher P D, Morgan D O, Robertson B H, Bachrach H L.
Cloned viral protein vaccine for foot-and-mouth disease: responses in
cattle and swine. Science, 1981, 214(4525):1125-1129.
[49] Abrams C C, King A M, Belsham G J. Assembly of foot-and-mouth
disease virus empty capsids synthesized by a vaccinia virus expression
system. Journal of General Virology, 1995, 76(12): 3089-3098.
[50] Lee C D, Yan Y P, Liang S M, Wang T F. Production of FMDV
virus-like particles by a SUMO fusion protein approach in Escherichia
coli. Journal of Biomedical Science, 2009, 16: 69.
[51] Li Z Y, Yi Y Z, Yin X P, Zhang Z F, Liu J X. Expression of
foot-and-mouth disease virus capsid proteins in silkworm-baculovirus
expression system and its utilization as a subunit vaccine. PLoS One,
2008, 3(5): e2273.
[52] Cao Y M, Sun P, Fu Y F, Bai X W, Tian F P, Liu X T, Lu Z J, Liu Z X.
Formation of virus-like particles from O-type foot-and-mouth disease
virus in insect cells using codon-optimized synthetic genes.
Biotechnology Letters, 2010, 32(9): 1223-1229.
[53] Guo H C, Sun S Q, Jin Y, Yang S L, Wei Y Q, Sun D H, Yin S H, Ma J
W, Liu Z X, Guo J H, Luo J X, Yin H, Liu X T, Liu D X.
Foot-and-mouth disease virus-like particles produced by a SUMO
fusion protein system in Escherichia coli induce potent protective
immune responses in guinea pigs, swine and cattle. Veterinary
Research, 2013, 44: 48.
[54] Zibert A, Maass K, Strebel K, Falk M M, Beck E. Infectious
foot-and-mouth disease virus derived from a cloned full-length cDNA.

3564 中 国 农 业 科 学 48 卷
Journal of Virology, 1990, 64(6): 2467-2473.
[55] Rieder E, Bunch T, Brown F, Mason P W. Genetically engineered
foot-and-mouth disease viruses with poly(C) tracts of two nucleotides
are virulent in mice. Journal of Virology, 1993, 67(9): 5139-5145.
[56] Baranowski E, Molina N, Nunez J I, Sobrino F, Sáiz M. Recovery of
infectious foot-and-mouth disease virus from sucking mice after direct
inoculation with in vitro-transcribed RNA. Journal of Virology, 2003,
77(20): 11290-11295.
[57] Liu G, Liu Z, Xie Q, Chen Y L, Bao H F, Chang H Y, Liu X T.
Generation of an infectious cDNA clone of an FMDV strain isolated
from swine. Virus Research, 2004, 104: 157-164.
[58] Bai X, Bao H, Li P, Wei W, Zhang M, Sun P, Cao Y M, Lu Z J, Fu Y F,
Xie B X, Chen Y L, Liu Z X. Effect of two amino acid substitutions in
the capsid proteins on the interaction of two cell-adapted PanAsia-1
strains of foot-and-mouth disease virus serotype O with heparan
sulfate receptor. Virology Journal, 2014, 11: 132.
[59] Li P H, Lu Z J, Bao H F, Li D, King D P, Sun P, Bai X W, Cao Y M,
Gubbins S, Chen Y L, Xie B X, Guo J H, Yin H, Liu Z X. In-vitro and
in-vivo phenotype of type Asia 1 foot-and-mouth disease viruses
utilizing two non-RGD receptor recognition sites. BMC Microbiology,
2011, 11: 154.
[60] Dunn C S, Donaldson A I. Natural adaptation to pigs of a Taiwanese
isolate of foot-and-mouth disease virus. Veterinary Record, 1997, 141:
174-175.
[61] Beard C W, Mason P W. Genetic determinants of altered virulence of
Taiwanese foot-and-mouth disease virus. Journal of Virology, 2000,
74(2): 987-991.
[62] Pacheco J M, Henry T M, O’Donnell V K, Gregory J B, Mason P W.
Role of nonstructural proteins 3A and 3B in host range and
pathogenicity of foot-and-mouth disease virus. Journal of Virology,
2003, 77(24): 13017-13027.
[63] Pacheco J M, Gladue D P, Holinka L G, Arzt J, Bishop E, Smoliga G,
Pauszek S J, Bracht A J, O’Donnell V, Fernandez-Sainz I, Fletcher P,
Piccone M E, Rodriguez L L, Borca M V. A partial deletion in
non-structural protein 3A can attenuate foot-and-mouth disease virus
in cattle. Virology, 2013, 446: 260-267.
[64] Li P H, Bai X W, Sun P, Li D, Lu Z J, Cao Y M, Fu Y F, Bao H F,
Chen Y L, Xie B X, Liu Z X. Evaluation of a genetically modified
foot-and-mouth disease virus vaccine candidate generated by reverse
genetics. BMC Veterinary Research, 2012, 8: 57.
[65] Valdazo-Gonzalez B, Timina A, Scherbakov A, Abdul-Hamid N F,
Knowles N J, King D P. Multiple introductions of serotype O
foot-and-mouth disease viruses into East Asia in 2010-2011.
Veterinary Research, 2013, 44: 76.
[66] Ma X Q, Li P H, Bai X W, Sun P, Bao H F, Lu Z J, Cao Y M, Li D,
Chen Y L, Qiao Z L, Liu Z X. Sequences outside that of residues
93-102 of 3A protein can contribute to the ability of foot-and-mouth
disease virus (FMDV) to replicate in bovine-derived cells. Virus
Research, 2014, 191: 161-171.
[67] Gonzalez-Magaldi M, Postigo R, Beatriz G, et al. Mutations that
hamper dimerization of foot-and-mouth disease virus 3A protein are
detrimental for infectivity. Journal of Virology, 2012, 86(20):
11013-11023.
[68] Gladue D P, O’Donnell V, Baker-Bransetter R, Pacheco J M, Holinka
L G, Arzt J, Pauszek S, Fernandez-Sainz I, Flerher P, Brocchi E, Lu Z,
Rodriguez L L, Borca M V. Interaction of foot-and-mouth disease
virus non-structural protein 3A with host protein DCTN3 is important
for viral virulence in cattle. Journal of Virology, 2013, 88(5):
2737-2747.
[69] Ruiz-Saenz J, Goez Y, Tabares W, Lopez-Herrera A. Cellular receptors
for foot-and-mouth disease virus. Intervirology, 2009, 52(4): 201-212.
[70] Jackson T, Ellard F A, Ghazaleh R A, Brookes S M, Blakemore W E,
Corteyn A H, Stuart D I, Newman J W I, King A M Q. Efficient
infection of cells in culture by type O foot-and-mouth disease virus
requires binding to cell surface heparin sulfate. Journal of Viroplogy,
1996, 70(8): 5282-5287.
[71] Zhao Q, Pacheco J, Mason P. Evalution of genetically engineered
derivatives of a Chinese strain of foot-and-mouth disease virus reveals
a novel cell-binding site which functions in cell culture and in animals.
Journal of Virology, 2003, 77(5): 3269-3280.
[72] Zeng J, Wang H, Xie X, Li Chen, Zhou G H, Yang D C, Yu L.
Ribavirin-restricted variants of foot-and-mouth disease virus: the
effect of restriced quasispecies diversity on viral virulence. Journal of
Virology, 2014, 88(8): 4008-4020.
[73] Du Y J, Bi J S, Liu J Y, Liu X, Wu X J, Jiang P, Yoo D W, Zhang Y G,
Wu J Q, Wan R Z, Zhao X M, Guo L H, Sun W B, Cong X Y, Chen L,
Wang J B. 3Cpro of foot-and-mouth disease virus antagonizes the
interferon signaling pathway by blocking STAT1/STAT2 nuclear
translocation. Journal of Virology, 2014, 88(9): 4908-4920.
(责任编辑 林鉴非)

17 期 《中国农业科学》创刊以来被引次数居前 100 的论文 3565
《中国农业科学》创刊以来被引次数居前 100 的论文
(依据 CNKI 数据库,检索日期:2015.8.20)
1. 贾继增. 分子标记种质资源鉴定和分子标记育种. 1996, 29(4):1-10.(1801 次) 2. 张维理, 武淑霞, 冀宏杰, Kolbe H. 中国农业面源污染形势估计及控制对策 I.21世纪初期中国农业面源污染
的形势估计. 2004, 37(7): 1008-1017.(1150 次) 3. 彭少兵, 黄见良, 钟旭华, 杨建昌, 王光火, 邹应斌, 张福锁, 朱庆森, Roland Buresh, Christian Witt. 提高中
国稻田氮肥利用率的研究策略. 2002, 35(9): 1095-1103.(884 次) 4. 凌启鸿, 张洪程, 蔡建中, 苏祖芳, 凌励. 水稻高产群体质量及其优化控制探讨. 1993, 26(6): 1-11.(645 次) 5. 刘录祥, 孙其信, 王士芸. 灰色系统理论应用于作物新品种综合评估初探. 1989, 22(3): 22-27.(614 次) 6. 李博. 中国北方草地退化及其防治对策. 1997, 30(6): 1-9.(567 次) 7. 任天志. 持续农业中的土壤生物指标研究. 2000, 33(1): 68-75.(533 次) 8. 莫惠栋. 我国稻米品质的改良. 1993, 26(4): 8-14.(501 次) 9. 郭三堆, 崔洪志, 夏兰芹, 武东亮, 倪万潮, 张震林, 张保龙, 徐英俊. 双价抗虫转基因棉花研究. 1999, 32(3):
1-7.(461 次) 10. 曾三省. 中国玉米杂交种的种质基础. 1990, 23(4): 1-9.(456 次) 11. 周光宇, 翁坚, 龚蓁蓁, 曾以申, 杨晚霞, 沈慰芳, 王自芬, 陶全洲, 黄骏麒, 钱思颖, 刘桂玲, 应苗成, 薛达
元, 洪爱华, 徐英俊, 陈善葆, 段晓岚. 农业分子育种 授粉后外源 DNA 导入植物的技术. 1988, 21(3): 1-6.(440 次)
12. 赵聚宝, 梅旭荣, 薛军红, 钟兆站, 张天佑. 秸秆覆盖对旱地作物水分利用效率的影响. 1996, 29(2): 59-66.(425 次)
13. 舒庆尧, 吴殿星, 夏英武, 高明尉, Anna McClung. 稻米淀粉 RVA 谱特征与食用品质的关系. 1998, 31(3): 26-29.(418 次)
14. 吴景锋. 我国主要玉米杂交种种质基础评述. 1983, 16(2): 1-8(409 次) 15. 石明松. 对光照长度敏感的隐性雄性不育水稻的发现与初步研究. 1985, 18(2): 44-48.(387 次) 16. 王月福, 姜东, 于振文, 曹卫星. 氮素水平对小麦籽粒产量和蛋白质含量的影响及其生理基础. 2003, 36(5):
513-520.(369 次) 17. 闫湘, 金继运, 何萍, 梁鸣早. 提高肥料利用率技术研究进展. 2008, 41(2): 450-459.(368 次) 18. 王懿波, 王振华, 王永普, 张新, 陆利行. 中国玉米主要种质杂交优势利用模式研究. 1997, 30(4): 16-24.(360
次) 19. 蒋柏藩, 顾益初. 石灰性土壤无机磷分级体系的研究. 1989, 22(3): 58-66.(356 次) 20. 刘新芝, 彭泽斌, 傅骏骅, 李连城, 黄长玲. RAPD 在玉米类群划分研究中的应用. 1997, 30(3): 44-51.(356
次) 21. 袁隆平. 两系法杂交水稻研究的进展. 1990, 23(3): 1-6.(349 次) 22. 李新海, 傅骏骅, 张世煌, 袁力行, 李明顺. 利用 SSR 标记研究玉米自交系的遗传变异. 2000, 33(2): 1-9.(346
次) 23. 杨守仁, 张龙步, 王进民. 水稻理想株形育种的理论和方法初论. 1984, 17(3): 6-13.(345 次) 24. 凌启鸿, 张洪程, 戴其根,丁艳锋, 凌励, 苏祖芳, 徐茂, 阙金华, 王绍华. 水稻精确定量施氮研究. 2005,
38(12): 2457-2467.(322 次)
jia
文本框
返回目录

3566 中 国 农 业 科 学 48 卷
25. 高云超, 朱文珊, 陈文新. 秸秆覆盖免耕土壤微生物生物量与养分转化的研究. 1994, 27(6): 41-49.(316 次) 26. 姚大年, 李保云, 朱金宝, 梁荣奇, 刘广田. 小麦品种主要淀粉性状及面条品质预测指标的研究. 1999, 32(6):
84-88.(303 次) 27. 张维理, 徐爱国, 冀宏杰, Kolbe H. 中国农业面源污染形势估计及控制对策 Ⅲ.中国农业面源污染控制中存
在问题分析. 2004, 37(7): 1026-1033.(300 次) 28. 贾士荣. 转基因植物食品中标记基因的安全性评价. 1997, 30(2): 1-15.(298 次) 29. 张明生, 谢波, 谈锋, 张启堂. 甘薯可溶性蛋白、叶绿素及 ATP 含量变化与品种抗旱性关系的研究. 2003,
36(1):13-16.(295 次) 30. 马德华, 庞金安, 霍振荣, 李淑菊. 黄瓜对不同温度逆境的抗性研究. 1999, 32(5): 28-35.(294 次) 31. 张维理, 冀宏杰, Kolbe H., 徐爱国. 中国农业面源污染形势估计及控制对策 Ⅱ.欧美国家农业面源污染状况
及控制. 2004, 37(7): 1018-1025.(293 次) 32. 王允白, 王宝华, 郭承芳, 王凤龙, 周健. 影响烤烟评吸质量的主要化学成分研究. 1998, 31(1): 89-91.(288
次) 33. 王月福, 于振文, 李尚霞, 余松烈. 施氮量对小麦籽粒蛋白质组分含量及加工品质的影响. 2002, 35(9):
1071-1078.(281 次) 34. 江立庚, 曹卫星, 甘秀芹, 韦善清, 徐建云, 董登峰, 陈念平, 陆福勇, 秦华东. 不同施氮水平对南方早稻氮
素吸收利用及其产量和品质的影响. 2004, 37(4): 490-496.(276 次) 35. 文雁成, 王汉中, 沈金雄, 刘贵华, 张书芬. 用 SRAP 标记分析中国甘蓝型油菜品种的遗传多样性和遗传基
础. 2006, 39(2): 246-256.(274 次) 36. 刘立军, 桑大志, 刘翠莲, 王志琴, 杨建昌, 朱庆森. 实时实地氮肥管理对水稻产量和氮素利用率的影响.
2003, 36(12): 1456-1461.(273 次) 37. 李秀英, 赵秉强, 李絮花, 李燕婷, 孙瑞莲, 朱鲁生, 徐晶, 王丽霞, 李小平, 张夫道. 不同施肥制度对土壤
微生物的影响及其与土壤肥力的关系. 2005, 38(8): 1591-1599.(271 次) 38. 段晓岚, 陈善葆. 外源 DNA 导入水稻引起性状变异. 1985, 18(3): 6-11.(269 次) 39. 郭三堆. 植物 Bt 抗虫基因工程研究进展. 1995, 28(5): 8-13.(268 次) 40. 凌启鸿, 杨建昌. 水稻群体“粒叶比”与高产栽培途径的研究. 1986, 19(3): 1-8.(266 次) 41. 魏国强, 朱祝军, 方学智, 李娟, 程俊. NaCl 胁迫对不同品种黄瓜幼苗生长、叶绿素荧光特性和活性氧代谢的
影响. 2004, 37(11): 1754-1759.(265 次) 42. 何萍, 金继运, 林葆. 氮肥用量对春玉米叶片衰老的影响及其机理研究. 1998, 31(3): 66-71.(260 次) 43. 李新海, 袁力行, 李晓辉, 张世煌, 李明顺, 李文华. 利用 SSR 标记划分 70 份我国玉米自交系的杂种优势群.
2003, 36(6): 622-627.(256 次) 44. 张学勇, 杨欣明, 董玉琛. 醇溶蛋白电泳在小麦种质资源遗传分析中的应用. 1995, 28(4): 25-32.(255 次) 45. 朱金宝, 刘广田, 张树榛, 孙辉. 小麦籽粒高、低分子量谷蛋白亚基及其与品质关系的研究. 1996, 29(1):
34-39.(254 次) 46. 吕卫光, 张春兰, 袁飞, 彭宇. 化感物质抑制连作黄瓜生长的作用机理. 2002, 35(1): 106-109.(253 次) 47. 鲁如坤, 时正元, 施建平. 我国南方 6 省农田养分平衡现状评价和动态变化研究. 2000, 33(2): 63-67.(249 次) 48. 朱庆森, 张祖建, 杨建昌, 曹显祖, 郎有忠, 王增春. 亚种间杂交稻产量源库特征. 1997, 30(3): 52-59.(247次) 49. 巨晓棠, 刘学军, 邹国元, 王朝辉, 张福锁. 冬小麦/夏玉米轮作体系中氮素的损失途径分析. 2002, 35(12):
1493-1499.(246 次) 50. 张春庆, 李晴祺. 影响普通小麦加工馒头质量的主要品质性状的研究. 1993, 26(2): 39-46.(244 次) 51. 杨建昌, 王志琴, 朱庆森. 不同土壤水分状况下氮素营养对水稻产量的影响及其生理机制的研究. 1996,
29(4): 58-66.(243 次) 52. 梁永超, 胡锋, 杨茂成, 朱遐亮, 王广平, 王永乐. 水稻覆膜旱作高产节水机理研究. 1999, 32(1): 26-32.(241

17 期 《中国农业科学》创刊以来被引次数居前 100 的论文 3567
次) 53. 姜东, 于振文, 李永庚, 余松烈. 施氮水平对高产小麦蔗糖含量和光合产物分配及籽粒淀粉积累的影响.
2002 , 35( 2): 157-162.(239 次) 54. 周广生, 梅方竹, 周竹青, 朱旭彤. 小麦不同品种耐湿性生理指标综合评价及其预测. 2003, 36(11): 1378-1382.
(236 次) 55. 聂华堂, 陈竹生, 计玉. 水分胁迫下柑桔的生理变化与抗旱性的关系. 1991, 24(4): 14-18.(234 次) 56. 宋海星, 李生秀. 玉米生长量、养分吸收量及氮肥利用率的动态变化. 2003, 36 (1): 71- 76.(231 次) 57. 周光宇. 从生物化学的角度探讨远缘杂交的理论. 1978(2): 16-20.(230 次) 58. 李娟, 赵秉强, 李秀英, Hwat Bing So. 长期有机无机肥料配施对土壤微生物学特性及土壤肥力的影响. 2008,
41(1): 144-152.(224 次) 59. 李宗智, 孙馥亭, 张彩英, 金亚辰. 不同小麦品种品质特性及其相关性的初步研究. 1990, 23(6): 35-41.(223
次) 60. 倪万潮, 张震林, 郭三堆. 转基因抗虫棉的培育. 1998, 31(2): 8-13.(223 次) 61. 徐凯, 郭延平, 张上隆. 不同光质对草莓叶片光合作用和叶绿素荧光的影响. 2005, 38(2): 369-375.(222 次) 62. 张学勇, 董玉琛, 游光侠, 王兰芬, 李培, 贾继增. 中国小麦大面积推广品种及骨干亲本的高分子量谷蛋白
亚基组成分析. 2001, 34(4): 355-362.(221 次) 63. 岳寿松, 于振文, 余松烈, 许玉敏. 不同生育时期施氮对冬小麦旗叶衰老和粒重的影响. 1997, 30(2): 42-46.
(221 次) 64. 王立秋, 靳占忠, 曹敬山, 王占宇. 水肥因子对小麦籽粒及面包烘烤品质的影响. 1997, 30(3): 67-73.(217 次) 65. 张洪程, 王秀芹, 戴其根, 霍中洋, 许轲. 施氮量对杂交稻两优培九产量、品质及吸氮特性的影响. 2003,
36(7): 800-806.(216 次) 66. 何中虎, 林作楫, 王龙俊, 肖志敏, 万富世, 庄巧生. 中国小麦品质区划的研究. 2002, 35(4): 359-364.(212次) 67. 刘宏斌, 李志宏, 张云贵, 张维理, 林葆. 北京市农田土壤硝态氮的分布与累积特征. 2004, 37(5): 692-698.
(208 次) 68. 阎俊, 张勇, 何中虎. 小麦品种糊化特性研究. 2001, 34(1): 9-13.(206 次) 69. 朱新开, 郭文善, 周君良, 胡宏, 张影, 李春燕, 封超年, 彭永欣. 氮素对不同类型专用小麦营养和加工品质
调控效应. 2003, 36 (6): 640- 645.(204 次) 70. 袁力行, 傅骏骅, 刘新芝, 彭泽斌, 张世煌, 李新海, 李连城. 利用分子标记预测玉米杂种优势的研究. 2000,
33(6): 6-12.(203 次) 71. 朱冰, 黄大年, 杨炜, 薛锐, 肖晗, 田文忠, 李良才, 戴顺洪. 利用基因枪法获得可遗传的抗除草剂转基因水
稻植株. 1996, 29(6): 15-20.(203 次) 72. 杨建昌, 杜永, 吴长付, 刘立军, 王志琴, 朱庆森. 超高产粳型水稻生长发育特性的研究. 2006, 39(7):
1336-1345.(202 次) 73. 白由路, 金继运, 杨俐苹, 梁鸣早. 基于 GIS 的土壤养分分区管理模型研究. 2001, 34(1): 1-4.(199 次) 74. 高吉寅, 胡荣海, 路漳, 杨国良. 水稻等品种苗期抗旱生理指标的探讨. 1984, 17(4): 41-45.(198 次) 75. 汪炳良, 徐敏, 史庆华, 曹家树. 高温胁迫对早熟花椰菜叶片抗氧化系统和叶绿素及其荧光参数的影响.
2004, 37(8): 1245-1250.(197 次) 76. 凌启鸿, 凌励. 水稻不同层次根系的功能及对产量形成作用的研究. 1984, 17(5): 3-11.(196 次) 77. 王惠聪, 黄旭明, 胡桂兵, 黄辉白. 荔枝果皮花青苷合成与相关酶的关系研究. 2004, 37(12): 2028-2032.(194
次) 78. 孙旭初. 水稻茎秆抗倒性的研究. 1987, 20(4): 32-37.(191 次) 79. 庄恒扬, 刘世平, 沈新平, 陈后庆, 陆建飞. 长期少免耕对稻麦产量及土壤有机质与容重的影响. 1999, 32(4):
39-44.(190 次)

3568 中 国 农 业 科 学 48 卷
80. 程宪国, 汪德水, 张美荣, 周涌, 金轲, 郭世昌, 王自力, 王书子. 不同土壤水分条件对冬小麦生长及养分吸
收的影响. 1996, 29(4): 67-74.(189 次) 81. 刘国栋, 刘更另. 论缓解我国钾源短缺问题的新对策. 1995, 287(1): 25-32.(188 次) 82. 叶兴国, Shirley Sato, 徐惠君, 杜丽璞, Tom Clemente. 小麦农杆菌介导转基因植株的稳定获得和检测. 2001,
34( 5): 465-468.(185 次) 83. 巨晓棠, 刘学军, 张福锁. 冬小麦与夏玉米轮作体系中氮肥效应及氮素平衡研究. 2002, 35(11): 1361-1368.(185
次) 84. 刘铮. 我国土壤中锌含量的分布规律. 1994, 27(1): 30-37.(185 次) 85. 杨建昌, 朱庆森, 曹显祖. 水稻群体冠层结构与光合特性对产量形成作用的研究. 1992, 25(4): 7-14.(183 次) 86. 卜玉山 , 苗果园 , 周乃健 , 邵海林 , 王建程 . 地膜和秸秆覆盖土壤肥力效应分析与比较 . 2006, 39(5):
1069-1075.(183 次) 87. 杨建昌, 王志琴, 朱庆森. 水稻品种的抗旱性及其生理特性的研究. 1995, 28(5): 65-72.(183 次) 88. 马存, 简桂良, 孙文姬. 我国棉花抗黄萎病育种现状、问题及对策. 1997, 30(2): 58-64.(183 次) 89. 赵先贵, 肖玲, 兰叶霞, 王书转, 马彩虹. 陕西省生态足迹和生态承载力动态研究. 2005, 38(4): 746-753.(182
次) 90. 杨建昌, 苏宝林, 王志琴, 郎有忠, 朱庆森. 亚种间杂交稻籽粒灌浆特性及其生理的研究. 1998, 31(1): 7-14.
(180 次) 91. 徐正进, 陈温福, 张龙步, 杨守仁. 水稻不同穗型群体冠层光分布的比较研究. 1990, 23(4): 10-16.(177 次) 92. 陈文新, 汪恩涛, 陈文峰. 根瘤菌-豆科植物共生多样性与地理环境的关系. 2004, 37(1): 81-86.(177 次) 93. 吴凤芝, 王学征. 设施黄瓜连作和轮作中土壤微生物群落多样性的变化及其与产量品质的关系. 2007, 40(10):
2274-2280.(177 次) 94.隋炯明, 李欣, 严松, 严长杰, 张蓉, 汤述翥, 陆驹飞, 陈宗祥, 顾铭洪. 稻米淀粉 RVA 谱特征与品质性状相
关性研究. 2005, 38(4): 657-663.(176 次) 95. 宋福平, 张杰, 谢天健, 杨自文, 戴莲韵, 李国勋. 苏云金芽孢杆菌cry基因PCR-RFLP鉴定体系的建立. 1998,
31(3): 13-18.(176 次) 96. 吴凤芝, 黄彩红, 赵凤艳. 酚酸类物质对黄瓜幼苗生长及保护酶活性的影响. 2002, 35(7): 821 -825.(174 次) 97. 林鸿宣, 闵绍楷, 熊振民, 钱惠荣, 庄杰云, 陆军, 郑康乐, 黄宁. 应用 RFLP 图谱定位分析籼稻粒形数量性
状基因座位. 1995, 28(4): 1-7.(174 次) 98. 金继运, 何萍. 氮钾营养对春玉米后期碳氮代谢与粒重形成的影响. 1999, 32(4): 56-62.(174 次) 99. 李凤民, 鄢珣, 王俊, 李世清, 王同朝. 地膜覆盖导致春小麦产量下降的机理. 2001, 34(3): 330-333.(172 次) 100. 李自超, 张洪亮, 曾亚文, 杨忠义, 申时全, 孙传清, 王象坤. 云南地方稻种资源核心种质取样方案研究.
2000, 33(5): 1-7.(171 次)
(接封四,续《中国农业科学》编委会)
Clinton C.Shock,USA
Song Kwan Jeong,Korea
Wolfgang Spielmeyer,Australia
Tatsuhiko shiraiwa,Japan
T.C.Tso,USA
Herman van Keulen,Netherlands
Virginia Walbot, USA
Colin White,Australia
Yunbi Xu,CIMMYT
Wengui Yan,USA
Yang Haishun,USA
Yoshio Inoue, Japan
Ralf-Udo Ehlers,Germany
M.D.Gale,UK
Geng Shu,USA
Bikram Gill,USA
Jude Grosser,USA
David Hannaway,USA
Jerry L.Hatfield,USA
Hiroyuki T.Kawashima,Japan
Gerrit Hoogenboom,USA
Irish V F,USA
Kshirod Kumar Jena,Korea
Harry K,Kaya,USA
G.S.Khush,USA
Xingen Lei,USA
Li Yi,USA
Lin Xi,USA
Gabor L.Lovei,Denmark
Lu Jiang,USA
Greg McMaster,USA
T.W.Mew,IRRI
Shaobing Peng,IRRI
Pichersky E,USA
David Powlson,UK
Avi sadka,Isreali
Mike Sadowsky,USA
Sanjaya Rajaram,ICARDA Zhang Hailin,USA
(以下按姓氏笔画排序)
万方浩 万建民 于振文 于康震 才学鹏 马峙英 马瑞昆 孔宪刚 文 杰 方智远 王 韧
王 强 王 蒂 王小佳 王小虎 王才林 王汉中 王立贤 王有年 王纪华 王秀峰 王志强
王坤波 王金宝 王海波 王笑梅 王艳玲 计 成 邓秀新 冯双庆 冯书堂 包满珠 叶志华
田文儒 白由路 石庆华 任光俊 刘 旭 刘 波 刘大群 刘永志 刘树生 吕 杰 吕加平
孙 寰 孙占祥 孙启忠 孙其信 孙宗修 孙武学 朱士恩 朱化彬 朱立煌 朱庆森 朱有勇
毕 阳 许世卫 严少华 贡锡峰 何中虎 何晨阳 吴孔明 吴爱忠 张 沅 张天真 张世煌
张玉龙 张志芳 张改平 张和平 张国政 张学勇 张洪程 张陆彪 张爱民 张献龙 张新友
张福锁 张端品 李 宁 李天来 李世清 李生秀 李立会 李庆章 李向林 李寿山 李杨瑞
李建生 李金泉 李保国 李春俭 李培武 李增智 李潮海 杜永臣 杜立新 杜德忠 杨 宁
杨汉春 杨怀文 杨建昌 辛志勇 邹 琦 陆文龙 陆承平 陈光宇 陈同斌 陈志强 陈怀满
陈昆松 周光宏 周常勇 屈冬玉 岳德荣 庞鸿宾 林 伟 林 敏 林而达 林矫矫 罗 军
罗玉柱 罗奇祥 罗振锋 罗绪刚 郑小波 郑有良 郑康乐 金继运 侯向阳 侯喜林 姚惠源
柳纪省 段爱旺 荣威恒 赵 明 赵开军 赵玉民 赵春江 骆世明 唐华俊 徐云碧 徐明岗
殷 宏 贾士荣 贾继增 郭三堆 郭予元 郭天财 郭文善 钱 前 钱克明 陶承光 高希武
高旺盛 常有宏 常汝镇 康绍忠 康振生 曹卫星 曹宏鑫 曹家树 梁 劬 梅旭荣 章力建
黄大昉 黄兴奇 黄季焜 喻大昭 喻树迅 喻景权 彭于发 彭友良 彭抒昂 程 序 程式华
童光志 董树亭 董常生 蒋宗勇 谢从华 谢关林 谢庆阁 谢建坤 谢笔钧 韩振海 路文如
路铁刚 翟虎渠 蔡贵信 蔡辉益 雒秋江 樊小林 黎志康 薛飞群 戴小枫 魏钦平 魏益民

Michael T. Clegg
M. Alley, USA David A. Andow, USA R. Appels, Australia Jill Shore Auburn, USA Bing Yang, USA Bas Bouman, IRRI
Bouzayen M, France John Bower, South Africa Kenneth G. Cassman, USA Chen Xian-ming, USA Chen Z X, USA Michael T. Clegg, USA
Matthew J. W. Cock, Switzerland Nick Costa, Australia Thomas Crenshaw, USA Zhanao Deng, USA C. Robert Dove, USA Lester E. Ehler, USA
Laurie Burkitt
Scientia Agricultura Sinica 1960 (Semimonthly, Started in 1960)
2015 9 1 48 17 Vol.48 No.17 September 1, 2015 Superintended by Ministry of Agriculture, P. R. China Sponsored by Chinese Academy of Agricultural Sciences
Chinese Association of Agricultural Science Societies Published by Editorial Department of Scientia Agricultura Sinica
12 Address No.12 South Street, Zhongguancun, Beijing 100081 Postcode 100081
010-82109808 82106281 Telephone 010-82109808 82106281 010-82106247 Fax 010-82106247 http://www.ChinaAgriSci.com Website http://www.ChinaAgriSci.com
[email protected] E-mail [email protected] Issued by Beijing Newspapers and Periodicals Distribution OfficeSubscription Domestic Post Office Issued Abroad by International Book Trading Corporation,
P.O.Box 399, Beijing, P.R. China 399 Abroad Issued No.BM43
49.50 / 1188.00 / Price US$ 49.50 / Issue US$ 1188.00 / year
ISSN 0578-1752 2-138
CN 11-1328/S BM43 0178
ISSN 0578-1752CODEN CKNYAR
SCIENTIA AGRICULTURA SINICA
Special Issue for 55th Anniversary
中国农业科学2015 2015.9.2, 2:49 PM17




















