Penyakit Inflamasi Yang Disebabkan Oleh Reaktivitas Humoral Abnormal Dan Penyakit Inflamasi Lainnya
50
BAGIAN 6 Penyakit Inflamasi yang Disebabkan oleh Reaktivitas Humoral Abnormal dan Penyakit Inflamasi Lainnya Bab 37 Imunitas Humoral dan Komplemen Lela A. Lee Sekilas Mengenai Imunitas Humoral dan Struktur Antibodi Imunitas humoral, yang difasilitasi oleh antibodi yang diproduksi oleh limfosit B, merupakan suatu bentuk imunitas spesifik yang ditujukan secara primer untuk berikatan dengan antigen-antigen ekstraseluler Molekul antibodi terbentuk dari dua rantai ringan (light chains) identik yang berikatan secara kovalen dengan dua rantai berat yang identik (heavy chains). Variable region yang dimiliki oleh setiap molekul antibodi yang berbeda bertanggungjawab dalam pembentukan ikatan yang terjadi, sedangkan constant region memediasi sebagian besar fungsi-fungsi efektor Terdapat 5 kelas antibodi, dimana masing-masing memiliki fungsi tertentu. Imunoglobulin (Ig) Mmerupakan kelas antibodi yang terlibat dalam
-
Upload
wildan-fahmy -
Category
Documents
-
view
28 -
download
0
description
inflamasi, humoral, imunitas
Transcript of Penyakit Inflamasi Yang Disebabkan Oleh Reaktivitas Humoral Abnormal Dan Penyakit Inflamasi Lainnya

BAGIAN 6
Penyakit Inflamasi yang Disebabkan oleh Reaktivitas Humoral
Abnormal dan Penyakit Inflamasi Lainnya
Bab 37 Imunitas Humoral dan Komplemen
Lela A. Lee
Sekilas Mengenai Imunitas Humoral dan Struktur Antibodi
Imunitas humoral, yang difasilitasi oleh antibodi yang diproduksi oleh
limfosit B, merupakan suatu bentuk imunitas spesifik yang ditujukan
secara primer untuk berikatan dengan antigen-antigen ekstraseluler
Molekul antibodi terbentuk dari dua rantai ringan (light chains) identik
yang berikatan secara kovalen dengan dua rantai berat yang identik (heavy
chains). Variable region yang dimiliki oleh setiap molekul antibodi yang
berbeda bertanggungjawab dalam pembentukan ikatan yang terjadi,
sedangkan constant region memediasi sebagian besar fungsi-fungsi
efektor
Terdapat 5 kelas antibodi, dimana masing-masing memiliki fungsi
tertentu. Imunoglobulin (Ig) Mmerupakan kelas antibodi yang terlibat
dalam respons antibodi, IgD merupakan reseptor antigen pada sel B naif
(naive B cells), IgA berperan dalam mekanisme imunitas mukosal, IgG
merupakan Ig mayor yang ditemukan dalam sirkulasi dan berperan penting
dalam respons antibodi sekunder, dan IgE memfasilitasi terjadinya
mekanisme imunitas terhadap parasit
Seorang individu dapat memproduksi jutaan antibodi yang diproduksi oleh
klona sel B melalui proses penyusunan ulang genetik (gene
rearrangement) dan diversitas jungsional (junctional diversity)
LIMFOSIT B

Selama evolusi, vertebrata berahang mengembangkan suatu kemampuan dalam
merespons invasi yang diakibatkan oleh sejumlah besar organisme asing.1
Imunitas spesifik ditandai dengan berbagai macam (diversitas) respons imunitas
yang terjadi paska paparan terhadap organisme-organisme tersebut.2 Sel-sel tubuh
yang dapat membedakan spesifitas substansi ataupun sel lain yang berasal dari
luar tubuh melalui reseptor-reseptornya yang berjumlah sangat banyak adalah
limfosit. Imunitas spesifik yang juga dikenal sebagai imunitas adaptif dimana
imunitas adaptif ini berkembang sebagai adaptasi terhadap infeksi yang terjadi,
dan dapat digolongkan ke dalam dua bentuk, yakni: imunitas humoral, yang
dimediasi oleh antibodi-antibodi yang diproduksi oleh limfosit B,dan imunitas
seluler, yang dimediasi oleh limfosit T. Dua bentuk imunitas spesifik tersebut
berkembang dan bertanggungjawab atas fungsi yang ebrbeda pula. Imunitas
humoral utamanya ditujukan untuk melawan antigen-antigen ekstraseluler, seperti
bakteri dan toksin yang bersirkulasi. Imunitas seluler utamanya ditujukan terhadap
berbagai antigen yang menginfeksi sel-sel inang (lihat Bab 10). Guna melawan
antigen-antigen ekstaseluler tersebut, agen imunitas harus dibentuk dalam jumlah
yang masif dan terdistribusi secara luas dalam tubuh, khususnya pada bagian-
bagian tubuh yang terpapar langsung dengan lingkungan. Antibodi-antibodi yang
dibentuk tubuh memenuhi berbagai karakteristik tersebut dan dapat disekresikan
dalam jumlah/kuantitas yang besar dari berbagai sel yang memproduksinya dan
dapat didistribusikan pada darah, mukosa, dan cairan interstisiel. Sebagai
tambahan, melalui reseptor Fc (Fc receptors, FcRs) antibodi dapat berikatan
dengan beberapa sel tertentu dalam sistem imunitas, seperti: sel mast, dan
memberikan spesifitas antigen terhadap sel-sel yang tidak memiliki reseptor
antigen spesifik yang diproduksi secara endogen (endogenously produced antigen-
spesific receptors). Disamping memerankan fungsi utamanya dalam imunitas
humoral sebagai produsen antibodi, limfosit B juga memainkan peranan penting
dalam presentasi antigen, regulasi subset-subset sel T dan sel dendritik, organisasi
jaringan limfoid, dan produksi sitokin dan kemokin.3,4
STRUKTUR ANTIBODI

Antibodi, atau imunoglobulin ((g) termasuk dalam keluarga glikoprotein yang
setiap kelasnya memiliki kesamaan struktur.2,5,6 Molekul antibodi memiliki bentuk
menyerupai huruf Y (Y-shape) yang terbentuk dari 2 rantai ringan yang identik,
dimana masing-masing memiliki berat molekul sebesar 24 kDa, dan membentuk
ikatan kovalen dengan 2 rantai berat yang identik, dimana masing-masing
memiliki berat molekul sebesar 55 atau 70 kDa, dan membentuk ikatan kovalen
satu sama lain. (Gambar 37-1). Baik dalam rantai berat maupun ringan terdapat
regio variabel dan konstan (variable and constant region). Fungsi mayor regio
variabel adalah untuk rekognisi/pengenalan antigen, sedangkan regio konstan
memfasilitasi fungsi-fungsi efektor. Rantai berat dan ringan terdiri dari
serangkaian ulangan, berupa unit homolog yang terbentuk dari 110 asam amino
yang diperkirakan memiliki struktur globuler dan dikenal sebagai domain Ig (Ig
domains). Motif domain Ig tersebut tidak hanya dapat ditemukan pada molekul
antibodi, tapi juga dapat ditemukan pada berbagai molekul lain yang tergolong
dalam Ig “superfamily”, termasuk reseptor sel T, the major histocompatibility
complex (MHC), CD4, CD8, intercellular adhesion molecule 1, dan beberapa
jenis molekul lain. Rantai ringan memiliki 2 domain mayor, (1) domain variabel
(VL) dan (domain konstan (Cl). Rantai berat memiliki 4 atau 5 domain mayor,
yakni: sebuah domain variabel (Vh) dan 3 domain konstan (pada IgA, IgD, dan
IgG) atau 4 domain konstan (pada IgM dan IgE) (CH1-4). Pada IgA, IgD, dan IgG,
terdapat sebuah regio persambungan/engsel (hinge region) antara CH1 dan CH2,
yang memberikan tambahan fleksibilitas bagi molekul antibodi. Domain-domain
variabel berada pada ujung N (N-terminus). Sedangkan pada ujung C (C-
terminus) dapat ditemukan domain-domain konstan dan pada ujung C rantai berat
dapat ditemukan domain-domain membran bound antibodies, transmembran, dan
sitoplasmik.
Dalam regio variabel dari rantai ringan dan berat terdapat 3 area yang
memiliki variabilitas yang besar, yang dikenal dengan nama regio hipervariabel
(hypervariable regions). Tiga regio tersebut yang notabene terletak berdekatan
pada struktur 3 dimensional antibodi, merupakan area yang paling

bertanggungjawab atas fungsi pengikatan antigen. Hal ini disebabkan oleh regio
hipervariabel tersebut membentuk bentuk komplementer dari antigen, sehingga
regio hipervariabel juga dikenal sebagai complementarity-determining regions.
Area-area unik tersebut oleh regio-regio hipervariabel yang terdapat dalam
individu-individu tertentu yang tidak mampu membangkitkan self-tolerance.
Sehingga, sistem imunitas tidak mampu untuk membedakan porsi antibodi
tertentu sebagai bagian dari bagian tubuh dan akhirnya memproduksi antibodi
terhadap porsi/bagian antibodi tersebut. Area pada antibodi yang
bertanggungjawab dalam menimbulkan respons imunitas yang terjadi dikenal
sebagai idiotope, dan respons antibodi terhadap idiotope dapat menghasilkan
sebuah jejaring interaksi idiotipik-anti-idiotipik (a network of idiotypic-anti-
idiotypic interactions) yang dapat berperan dalam membantu meregulasi respons
imunitas humoral.7
Terdapat 2 tipe rantai ringan, Ƙ dan λ, masing-masingnya tersandikan
dalam kromosom-kromosom yang berbeda. Setiap molekul antibodi biasanya
memiliki 2 rantai Ƙ atau 2 rantai λ, atau tidak sama sekali. Diyakini, bila memang
benar ada, terdapat perbedaan fungsional antara rantai Ƙ dan λ, meskipun hingga
saat ini juga belum diketahui secara pasrti. Terdapat 5 macam rantai berat, (1) α,
(2) δ, (3) ε, (4) ϒ, dan (5) µ, yang secara berurutan berhubungan dengan kelas-
kelas antibodi IgA, IgD, IgD, IgE, dan IgM. Perbedaan kelas rantai berat yang
dimiliki masing-masing kelas antibodi secara signifikan menentukan berbagai
fungsi masing-masing kelas antibodi tersebut, sebagaimana yang didiskusikan
dalam Bagian “Kelas-Kelas Antibodi”. Kelas antibodi IgA dan IgG merupakan
subkelas yang berhubungan dekat, yang masing-masing terdiri dari IgA1 dan
IgA2 dan IgG1, IgG2, IgG3, dan IgG4 (Tabel 37-1).
Penguraian enzimatik dari molekul-molekul IgG oleh papain akan
menghasilkan 3 produk pecahan, berupa 2 fragmen identik yang tersusun atas
sebuah rantai ringan yang berikatan dengan regio VCH1 pada rantai berat dan
sebuah Fc portion yang tersusun dari dua CH2-CH3 pada rantai berat yang saling
berikatan satu sama lain. Nama Fab diturunkan berdasarkan fitur pengikatan

antigen yang dimilikinya, sedangkan nama Fc diturunkan dari fitur kristalisasi
yang dimilikinya. Ketika IgG mengalami pemecahan oleh pepsin, regio ujung C
akan terurai menjadi beberapa fragmen yang berukuran lebih kecil. Terdapat
produk sisa yang tersusun atas regio Fab dan hinge region. Fragmen-fragemen
Fab yang berikatan dengan hinge region tersebut dikenal dengan nama Fab’.
Ketika 2 fragmen Fab’ dalam sebuah molekul antibodi tetap terhubung, maka
fragmen tersebut disebut sebagai F(ab’)2.
KELAS-KELAS ANTIBODI
(lihat Tabel 37-1)
IMUNOGLOBULIN M
Dalam sejarah evolusinya, IgM merupakan kelas antibodi terdahulu dan
merupakan molekul antibodi pertama yang terekspresikan selama perkembangan
sel B.1 Bentuk sekretorik primernya berupa pentamer yang tersusun atas 5
molekul IgM yang bergabung pada masing-masing ujung C pada rantai ekor dan
distabilkan oleh sebuah molekul yang dikenal sebagai joining (J) chain.
Terbentuknya ikatan antara molekul-molekul IgM pada membran dengan antigen
dapat menimbulkan aktivasi naive B cells. IgM yang disekresikan dapat
mengenali berbagai antigen, umumnya melalui interaksi berafinitas rendah, yang
mana dapat mengativasikan aktivitas komplemen. IgM merupakan efektor mayor
dalam respons primer antibodi. Meskipun interaksi IgM umumnya memiliki
afinitas yang rendah, IgM dapat berperan efektif dalam merespons berbagai
antigen polivalen (seperti polisakarida yang memiliki epitop berulang), hal ini
dapat terjadi nakibat struktur prntamerik yang dimilikinya dapat memfasilitasi
terjadinya beberapa interaksi berafinitas rendah sekaligus, sehingga dihasilkan
interaksi yang kuat (high avidity interaction) (Avidity disini menunjukkan
kekuatan keseluruhan dari ikatan yang terjadi, sedangkan afinitas merupakan
kekuatan yang dimiliki oleh ikatan tunggal antara antigen dengan tempat ikatan).
IMUNOGLOBULIN D

Moleku IgD dapat dijumpai dalam bentuk berikatan dengan membran dan
merupakan kelas antibodi kedua yang diekspresikan selama perkembangan sel-sel
B. Fungsinya belum sepenuhnya dapat dimengerti, meskipun dalam bentuk ikatan
dengan membran tersebut, IgD dapat berperan sebagai reseptor antigen bagi sel-
sel B naif. IgD yang telah disekresikan dapat ditemukan pada permukaan basofil,
dimana pada tempat tersebut, IgD dapat menginduksi produksi substansi
antimikrobial, opsonisasi, inflamatorik, dan B-cell-stimulating factors.9
IMUNOGLOBULIN A
IgA merupakan molekul IgA yang paling melimpah keberadaannya dalam tubuh,
dapat ditemukan dalam kuantitas yang besar pada lapisan mukosa tubuh. IgA
bertanggungjawab dalam menyokong imunitas mukosal dan juga disekresikan
dalam air susu ibu, sehingga turut berkontribusi dalam imunitas neonatal. Dalam
bentuk sekresinya, IgA dapat ditemukan sebagai monomer, dimer, atau trimer,
dimana bentuk multimer dibentuk melalui interaksi-interaksi yang terjadi antara
ekor-ekor molekul dan distabilkan oleh J chain.Dalam transportasinya melewati
lapisan-lapisan epitelial, dimer IgA melekat pada satu tipe FcR yang dikenal
sebagai polymeric Ig receptor.10 Ketika proses transportasinya sudah lengkap,
dimer IgA akan tetap berikatan dengan bagian ekstraseluler dari reseptor tersebut,
yang dikenal sebagai secretory component, yang mana dapat menghindarkan IgA
dari proteolisis. Sel-sel dalam sistem imunitas yang memiliki reseptor terhadap
IgA diantaranya berupa neutrofil, eosinofil, dan monosit.
IMUNOGLOBULIN G
IgG merupakan molekul Ig yang paling melimpah keberadaannya dalam sirkulasi.
IgG disekresikan dalam bentuk monomer. IgG memainkan peranan penting dalam
respons antibodi sekunder, dan cenderung memiliki interaksi dengan afinitas yang
besar terhadap antigen, utamanya ketika sistem imunitas telah mengalami
maturasi. Sejumlah sel dalam sistem imunitas memiliki FcR untuk IgG,
diantaranya monosit, neutrofil, eosinofil, natural killer (NK) cells, dan B cells.
IgG akan mengopsonisasi (melapisi) antigen, sehingga antigen dapat difagositosis,

dan mengaktivasikan komplemen. Terdapat pengecualian bagi molekul IgG4 yang
tidak memiliki kemampuan dalam mengatifkan komplemen. IgG berperan penting
dalam imunitas neonatal, karena IgG merupakan satu-satunya kelas IgG yang
dapat melewati plasenta, dan juga disekresikan dalam air susu ibu. Interaksi yang
terjadi antara IgG dengan reseptor FcR terkait MHC kelas I (MHC-class I-related
receptor FcRs) terlibat dalam mekanisme lolosnya IgG dalam melewati plasenta
dan juga atas pemanjangan kadar IgG yang tinggi dalam sirkulasi.11 Waktu paruh
iGG dalam serum mencapai 23 hari, yang mana dipertimbangkan relatif lebih
lama dibandingkan dengan waktu paruh Ig kelas lain.
IMUNOGLOBULIN E
IgE ditemukan dalam jumlah yang sangat kecil dalam sirkulasi. Dijumpai
terdapatnya reseptor-reseptor berafinitas tinggi terhadap Fc portion dari IgE pada
permukaan sel mast, basofil, dan eosinofil, dan reseptor-reseptor berafinitas
rendah yang dijumpai pada sel-sel B dan sel-sel Langerhans. Pada sel mast dan
basofil, terbentuknya ikatan antara IgE dengan antigen akan mengaktifkan sel-sel
tersebut. IgE memediasi terjadinya hipersensitifitas, meskipun sebenarnya fungsi
utamnaya adalah melawan infeksi parasit-parasit.
MEKANISME TERBENTUKNYA DIVERSITAS ANTIBODI
Jumlah berbagai informasi genetik yang tersandikan dalam DNA masing-masing
individu terbatasi oleh keterbatasan berupa kapasitas pengemasan DNA dalam
ukuran sel yang terbatas. Ruang sel yang tersedia terlalu sempit untuk menyimpan
DNA yang menyandikan milyaran reseptor-reseptor limfosit yang berbeda apabila
gen-gen tersebut disandikan secara terpisah. Limfosit telah beradaptasi terhadap
keterbatasan tersebut melalui mekanisme khusus yang dapat ditingkatkan
intensitasnya.12 Setiap klona dari sel B memproduksi reseptor-reseptor antigen
yang identik (contoh: antibodi) dengan spesifitas yang khas. Diperkirakan bahwa
masing-masing individu memiliki sebanyak 107 klona sel B yang berbeda, yang
dapat menghasilkan sebanyak 107 antibodi yang berbeda pula. Mekanisme utama
yang melatarbelakangi terbentuknya diversitas dalam jumlah yang sangat besar ini

adalah terjadinya penyusunan ulang gen (gene rearrangement), dimana segmen-
segmen DNA dalam limfosit mengalami rekombinasi somatik.13 Gen yang
menyandikan rantai-rantai pendek (light chain genes) memiliki 3 regio, (1) V
(variable),(2) J (joining), dan (3) C (constant), dan gen yang menyandikan rantai-
rantai berat (heavy chain genes) memiliki 4 regio, (1) V, (2) D (diversity), (3) J,
dan (4) C. Dalam masing-masing regio terdapat beberapa segmen gen yang
menentukan produk akhir antibodi manakah yang akan diproduksi, yang terjadi
melalui seleksi random satu segmen gen dari setiap regio. Peristiwa awitan yang
terjadi dalam pembentukan antibodi adalah penggabungan satu segmen D dan
satu segmen J dari heavy chain gene, yang diikuti delesi DNA diantara gabungan
kedua segmen. Selanjutnya, sebuah segmen V dipilih unruk digabungkan dengan
segmen DJ, dan segmen-segmen D yang bersisa didelsesi. Kompleks VDJ
bergabung dengan ujung 3’ dari segmen-segmen J yang tersisa plus regio C.
Segmen-segmen J yang tidak terpakai dihilangkan selama RNA processing.
Sebuah proses yang serupa terjadi dalam light chain loci; karena tidak dijumpai
keberadaan segmen-segmen D dalam light chain loci, sehingga akan terbantuk
kompleks VJ saja, bukan kompleks VDJ. (Khusus pada lokus K, rekombinasi VJ
dapat terjadi melalui mekanisme lain yang berbeda yang melibatkan inversi DNA
yang tanpa disertai delesi sekuens-sekuens yang tidak diperlukan, dimana
meskipun terdapat perbedaan struktural, keluaran fungsional yang dihasilkan tetap
sama).
Kemampuan untuk menyeleksi satu segmen dari beberapa segmen yang
tersedia dalam regio V, D, dan J menyumbang besarnya kemungkinan jenis
antibodi yang terbentuk. Tambahan atas terjadinya diversitas dalam jumlah yang
sangat besar ini juga diberikan oleh terjadinya proses yang dikenal dengan nama
the juxtaposition of a rearranged light chain to a rearranged heavy chain; juga
melalui terjadinya adisi, delesi, atau transposisi nukleotida-nukleotida yang berada
pada pertemuan (junctions) antara segmen V dengan D, segmen D dengan J, dan
segmen V dengan J – fenomena ini dikenal dengan nama junctional diversity, dan
melalui hipermutasi somatik paska stimulasi oleh antigen (lihat bawah).

MATURASI SEL B
Sekilas mengenai Maturasi Sel B
Pro-sel B (pro-B cell) mengekspresikan enzim-enzim yang diperlukan
dalam penyusunan ulang gen (gene rearrangement) dan junctional
diversity; meskipun demikian tidak terdapat rantai ringan maupun rantai
berat yang terekspresikan.
Pre sel B (pre-B cell) mensekresikan rantai berat µ (µ heavy chains) ke
dalam sitoplasma. Pada permukaan sel, rantai-rantai berattersebut
berikatan dengan rantai-rantai ringan yang sesuai (surrogate light chains)
untuk membentuk reseptor-reseptor pre-sel B.
Sel B imatur memproduksi rantai-rantai ringan dan dapat
mengekspresikan molekul-molekul antibodi pada permukaan sel. Apabila
terjadi paparan antigen pada fase ini, maka akan terjadi negative selection.
Selama fase transisional, sel-sel B secara gradual kehilangan sensitivitas
terhadap negative selection dan berubah menjadi kompeten secara
imunologis.
Sel B matur mengekspresikan baik IgM maupun IgD dan mampu
merespons terhadap paparan antigen yang terjadi.
Sel-sel yang digariskan menjadi sel-sel B matur akan mengalami berbagai
peristiwa dan progresi selama perkembangannya, sehingga dihasilkan pembentuan
rantai-rantai berat dan ringan yang sekuensial, dan keseluruhan molekul-molekul
antibodi, dimana dalam masa perkembangan tersebut terdapat beberapa
checkpoint guna menyeleksi sel-sel yang memproduksi gene rearrangement yang
tidak produktif atau antibodi-antibodi yang autoraktif (autoreactive antibody), dan
survival signals yang menyeleksi sel-sel yang dapat memproduksi antibodi-
antibodi yang memiliki kegunaan potensial. Proses perkembangan sel B terjadi
dalam beberapa tahap yang dapat dengan jelas diamati, dimana terdapat
karakteristik-karakteristik khusus berupa peristiwa-peristiwa spesifik dan dapat

diidentifikasikan melalui sejumlah penanda permukaan sel yang spesifik (spesific
cell surface markers) dan ekspresi gen Ig (Ig gene expression).
Sumsum tulang dan stem cell pada liver fetus yang bersifat pluripoten
dapat berkembang menjadi sel-sel B.2,14 Stem cell yang berkembang mengikuti
jalur limfositik pada awalnya akan berkembang menjadi common lymphoid
progenitors, yang berikutnya dapat berkembang menjadi sel B, sel T, atau NK
cells. Sel-sel B yang berkembang dari stem cell liver fetus utamanya berupa sel-
sel B1 (lihat Bagian “Aktivasi Sel B dan Fungsi Antibodi”), sedangkan sel-sel B
yang berasal dari sumsum tulang umumnya berupa sel-sel B folikuler. Sel-sel dan
molekul-meolekul ekstraseluler yang terdapat pada lingungan mikro stromal
menyediakan substansi dan pensinyalan yang diperlukan dalam proses diferensiasi
limfosit. Sekresi berbagai regulator transkripsional seperti EBF, E2A, dan Pax-5
mengakibatkan diproduksinya sejumlah protein yang memiliki peran krusial
dalam perkembangan sel B. Terjadinya Regulasi mRNA post-transkripsi
(posttranscriptional regulation of mRNA) oleh protein-protein pengikat RNA
(RNA-binding proteins) dan microRNAs memfasilitasi kontrol yang lebih lanjut
terhadap proses perkembangan dan diferensiasi sel B.15
Tahap selanjutnya dalam maturasi sel B dipegang oleh pre-B cell dan
ditandai dengan terjadinya sintesis rantai berat µ sitoplasmik (cytoplasmic µ heavy
chain). Karena rantai-rantai ringan belum diproduksi pada tahap ini, maka surface
Ig juga belum dapat diproduksi. Beberapa rantai berat µ berikatan dengan
invariant molecules yang dikenal sebagai surrogate light chains dan signal
transducing proteins Ig α dan Ig β untuk membentuk kompleks-kompleks yang
dikenal sebagai pre-B cell receptors. Sel-sel yang mensintesis rantai-rantai berat
dan telah dapat membentuk bagian-bagian pre-B cell receptor akan tetap bertahan
pada fase ini, karena pre-B cell receptors yang dibentuk memberikan sinyhal-
sinyal penting yang dapat menyokong survival, proliferasi, dan maturasi sel-sel
tersebut.

Pembentukan rantai-rantai ringan menandai terjadinya tahap selanjutnya
dalam maturasi sel B, yakni tahap sel B imatur (the immature B-cell stage).
Ketika rantai-rantai ringan bergabung dengan rantai-rantai berat µ, akan
dihasilkan sebuah molekul IgM yang akan diekspresikan pada permukaan sel.
Meskipun terbentuknya kompleks reseptor sel B memberikan kemampuan dalam
merekognisi antigen-antigen spesifik, meskipun demikian pada fase ini rekognisi
antigen yang terjadi belum dapat mengakibatkan terjadinya proliferasi atau
diferensiasi. Di lain pihak, sel-sel tersebut akan menagalmi negative selection
ketika terjadi paparan terhadap antigen. Sel-sel B imatur telah dapat mengenali
antigen sendiri (self-antigen) melalui terjadinya proses delesi,16 anergy, ataupun
receptor editing, yang merupakan sebuah proses berupa penyusunan ulang gen
sekunder yang dapat menghasilkan spesifitas yang baru terhadap antigen non-self
yang diperlukan dalam sistem imunitas.17
Perilisan sel-sel B imatur dari sumsum tulang menuju limpa menandai
awal terjadinya tahap selanjutnya, yakni berupa sel B-transisional (the transitional
B-cell stage).18 Sel-sel transisional secara gradual memperoleh ekspresi IgD,
CD21, dan CD23 pada permukaannya dan akan mejadi lebih kompeten secara
imunitas setelahnya. Alternative splicing of RNA yang terjadi menyebabkan
terjadinya ekspresi simultan dari IgM dan IgD. Pada awitan tahap ini, cross-link
antara reseptor-reseptor sel B mengakibatkan terjadinya negative selection.
Seiring berlanjutnya maturasi, sel-sel transisional menjadi responsif terhadap T-
cell helper dan mengalami hilangnya sensitifitas terhadap negative selection.
Sel B matur akan mengekspresikan IgM dan IgD dan telah kompeten
dalam merespons terhadap papran antigen. Sel-sel tersebut dipertimbangkan
masih bersifat naif (naive) apabila belum teraktivasikan oleh antigen. Mayoritas
sel B matur ditemukan bersirkulasi dalam jaringan limfoid periferal (splen, nodus
limfatikus, jaringan limfoid mukosal) dan dikenal sebagai sel B folikuler
(follicular B cells), atau recirculating B cells. Sel-sel B akan berkumpul menujun
folikel akibat pengaruh dari kemokin CXCL13, yang disekresikan oleh sel-sel
dendritik folikuler, dan bertahan dan hidup pada folikel dengan bantuan sitokin

yang dikenal sebagai BAFF (B-cell activating factor), yang juga dikenal sebagai
BlyS (B lymphocyte stimulator). Sejumlah kecil persentase sel-sel B bersemayam
dalam zona marginal splen dan menetap di tempat tersebut.
Terpaparnya mature naive B cells pada antigen tertentu mengakibatkan
terjadinya aktivasi, proliferasi, dan diferensiasi sel B. (lihat Bagian “Aktivasi Sel
B dan Fungsi Antibodi”). Satu subsetsel B akan berkembang menjadi sekelompok
sel yang dikenal sebagai memory B cells, yang akan keberadaannya akan bertahan
dalam jangka waktu yang cukup lama meskipun tanpa adanya stimulasi oleh
antigen, dan dapat merespons kembali dengan cepat apabila dalam sirkulasi
dijumpai antigen serupa.19 Subset sel B yang lain berdiferensiasi menjadi sel-sel
yang mensekresikan IgG. Sel-sel B yang telah mencapai diferensiasi terminal
akan memproduksi dan mensekresikan sejumlah Ig, lini sel-sel tersebut dikenal
sebagai sel plasma yang umumnya memiliki retikulum endoplasma yang
melimpah, yang jelas konsisten dengan fungsi yang diperankannya sebagai pabrik
produksi antibodi.20
PENGIKATAN ANTIGEN OLEH SEL-SEL B
Sel-sel B mampu mengenali berbagai jenis makromolekul, termasuk protein, lipid,
karbohidrat, dan asam nukleat. Porsi/bagian molekul tertentu yang mampu
dikenali oleh antibodi disebut sebagai epitop atau determinan. Sel-sel B
merekognisi baik epitop linear (epitop yang dibentuk oleh beberapa asam amino
yang berdekatan) dan paling umum, dapat mengenali epitop konformasional
(epitop berupa pelipatan/folding dari makromolekul).21 Berbeda dengan sel-sel B,
respons imunitas yang dimiliki sel-sel T umumnya bersifat terbatas hanya mampu
mengenali epitop linear dari peptida-peptida tertentu saja.
Makromolekul-makromolekul, khususnya protein yang berukuran besar,
umumnya mengandung beberapa epitop yang berbeda, dan respons humoral
terhadap sebuah makromolekul tertentu umumnya difasilitasi oleh beberapa jenis
antibodi yang berbeda. Meskipun setiap antibodi bersifat spesifik terhadap
konfigurasi epitopik tertentu, kesamaan epitop dapat dijumpai pada antibodi

terhadap epitop tertentu dari makromolekuln tertentu, dan juga dapat ditemukan
terjadinya ikatan antibodi terhadap epitop yang berbeda dari makromeolekul yang
berbeda pula. Fenomena ini dikenal sebagai reaktivitas silang (cross-reactivity),
dan berperan penting dalam pembentukan/genesis respons antibodi autoimun.
Makromolekul-makromolekul yang memiliki epitop-epitop identik dalam
jumlah yang multipel diklasifikasikan sebagai polivalen atau multivalen. Reaksi
yang terjadi antara antibodi-antibodi terhadap berbagai makromolekul tersebut
atau agregat makromolekulernya dapat membentuk kompleks-kompleks yang
dikenal sebagai kompleks imun (immune complexes) dengan antigen. Dalam
konsentrasi tertentu, antibodi dan antigen berada pada zona ekuivalen (zone of
equivalence), akan terbentuk serangkaian jejaring yang tersusun dari antibodi-
antibodi dan antigen-antigen. Pada konsentrasi antibodi atau antigen yang lebih
rendah ataupun lebih tinggi, kompleks yang terbentuk umumnya berukuran lebih
kecil. Kompleks-kompleks imun, yang dibentuk dalam sirkulasi atau dalam
jaringan, diperkirakan bertanggungjawab terhadap terjadinya penyakit melalui
inisiasi respons inflamasi yang dibawanya.
AKTIVASI SEL B DAN FUNGSI ANTIBODi
SEKILAS MENGENAI AKTIVASI SEL B DAN FUNGSI ANTIBODI
Ketika reseptor sel B (antibodi permukaan) berikatan dengan antigen,
sebuah sinyal sekunder yang dirilis oleh ikatan yang terjadi antara C3d
dengan reseptor komplemen (complement receptor) 2 secara signifikan
menyebabkan dan memperkuat aktivasi sel B.
Respons sel B terhadap antigen protein umumnya memerlukan bantuan
sel T helper
Sel B teraktivasi dapat berkembang menjadi sel-sel plasma yang memiliki
waktu hidup yang pendek (short-lived plasma cells), sel-sel B memori
(memory B cells), atau sel-sel plasma yang memiliki periode hidup yang
panjang (long-lived plasma cells). Long-lived plasma cells akan
bermigrasi menuju sumsum tulang, dimana di tempat tersebut sel-sel

tersebut akan terakumulasi dan bertahan hidup dan menjadi sumber utama
yang memproduksi antibodi-antibodi spesifik antigen tertentu (antigen-
spesific antibodies) dalam sirkulasi.
Fungsi efektor dari antibodi berupa netralisasi antigen, aktivasi
komplemen, aktivasi sel, fagositosis dan antibody-dependent, cell-
mediated cytotoxicity. Sebagian besar fungsi tersebut diperantarai oleh
terbentuknya ikatan Ig dengan reseptor-reseptor Fc yang didalamnya
terkandung sebuah imunoreseptor, berupa motif aktivasi berbasis tirosin
(immunoreceptor tyrosine-based activation motif).
Saat terjadi cross-linking antara reseptor sel B matur dengan antigen, bentukan
kluster reseptor yang terjadi menginisiasi pensinyalan yang ditransduksikan oleh
Igα dan Igβ. Kaskade komplek sensinyalan (complex signaling cascade) tersebut
melibatkan terjadinya fosforilasi tirosin kinase, termasuk Lyn, Fyn, Btk, dan Syk,
yang mengakibatkan terjadinya ekspresi gen-gen yang terlibat dalam aktivasi sel
B.22 Aktivasi sel B difasilitasi oleh sinyal sekunder, yang ditransduksikan oleh
protein komplemen, C3d.23 Fragmen komplemen C3d terbentuk sebagai hasil
atas terjadinya aktivasi komplemen (lihat Bagian “Komplemen”). Pada
permukaan sel B dapat dijumpai sebuah kompleks ko-reseptor yang mengandung
complement receptor 2 (CR2), CD19, dan CD81 (yang juga dikenal sebagai
TAPA-1, atau target for antipro liferative antigen-1). Terjadinya pengikatan yang
simultan antara antigen dengan antibodi pada permukaan sel B dan C3d dengan
CR2 mengakibatkan terjadinya peningkatan aktivasi sel B. Aktivasi sel B dapat
juga terjadi melalui toll-like receptors yang dapat merekognisi/mengenali produk
mikrobial spesifik.24
Respons lanjutan yang terjadi akibat paparan antigen sering melibatkan
interaksi kompleks yang terjadi antara sel-sel B dan sel-sel T, dimana dari sini
akan dapat dibangkitkan respons imunitas yang berfungsi dengan baik.25
Rekognisi antigen baik oleh sel B maupun sel T menyebabkan terjadinya
peningkatan ekspresi protein-protein permukaan sel dan sitokin-sitokin yang
menyebabkan terjadinya migrasi sel-sel tersebut dan terjadinya interaksi secara

produktif diantaranya. Sel T mengenali peptide-class II MHC complexes yang
terdapat pada sel-sel dendritik dan menerima sinyal primer yang berasal dari
kompleks-kompleks tersebut dan sinyal sekumder yang berasal dari interaksi
kostimularik yang melibatkan pengikatan B7-1 dan B7-2 pada sel-sel dendritik
dengan CD28 pada sel T.26 Sel-sel T teraktivasi tersebut mengekspresikan CXCR5
yang merupakan ligan untuk CXCL13, sehingga menyebabkan terjadinya migrasi
sel T menuju folikel dan meningkatkan jumlahnya melebihi jumlah sel-sel B yang
ada. Respons yang timbul atas paparan antigen protein, sel B memfagositosis
(take-up) antigen, memprosesnya, dan mempresentasikan hasil olahan antigen
pada permukaan sel dalam kompleks MHCkelas II. Sel-sel B teraktivasi
mengekspresikan CXCR5 dalamjumlah yang lebih sedikit, sehingga
memungkinkan terjadinya migrasi sel-sel ini dari folikel menuju zona-zona sel T.
Pada perbatasan antara folikel dengan zona sel T,sel-sel T teraktivasi berinteraksi
dengan sel-sel B dan memberikan pensinyalan bagi sel-sel B melalui
pembentukan ikatan antara CD40 yang terdapat pada sel-sel B dengan ligan CD40
(CD154) pada sel-sel T dan melalui aktivitas sitokin-sitokin yang terjadi,
utamanya interleukin 2 (IL-2), IL-4, IL-21, BAFF, dan APRIL (a proliferation-
inducing ligand).2 Sinyal-sinyal tersebut berperan penting proses subsequent class
(heavy chain isotype) switching, afinitas maturasi, dan permbentukan dan
perkembangan sel B memori. Keseluruhan efek yang terjadi pada sel-sel B
menjadi stimulasi atas proliferasi dan diferensiasi yang terjadi selanjutnya.
Dalam fase ini, beberapa sel B teraktivasi berubah menjadi short-lived
plasma cells, dan juga menghasilkan respons inisial yang cukup terhadap antigen,
sedangkan sisanya bermigrasi kembali dari folikel di perifer dan berproliferasi
secara cepat serta membentuk germinal centers. Utamanya, pada germinal centers
inilah terjadi class switching, maturasi afinitas (affinity maturation), dan
berkembangnya sel B memori. Class switching antara IgM dengan IgA, IgE, atau
IgG terjadi akibat adanya interaksi yang terjadi antara sel-sel T dengan sel-sel
B.27,28 Penentuan kelas antibodi yang digunakan didasarkan pada tempat dimana
antigen ditangkap dan cytokine milieu. Sebagai contoh, respons sel B terhadap

antigen-antigen yang ditangkap pada permukaan mukosa umumnya terjadi akibat
terdapatnya switching class menjadi IgA, dan transforming growth factor-β
merupakan salah satu sitokin penting yang turut berkontribusi didalamnya.IL-4
merupakan sinyal penting yang berperan dalam class switching menjadi IgE.
Interaksi sel T dengan sel B juga mengakibatkan terjadinya maturasi afinitas,
selain itu juga menyebabkan peningkatan afinitas antibodi terhadap antigen
secara progresif. Selama maturasi afinitas, hipermutasi somatik yang terjadi pada
gen-gen antibodi mengakibatkan dihasilkannya antibodi-antibodi yang memiliki
afinitas yang dapat lebih kuat sekaligus lebih lemah.29,30Antibodi-antibodi yang
memiliki afinitas lebih kuat umumnya memberikan manfaat survival bagi sel-sel
B yang memproduksinya. Secara progresif, populasi sel-sel B berevolusi guna
memproduksi antibodi-antibodi yang mememiliki afinitas antibodi terhadap
antigen yang lebih tinggi. Dalam prosesnya, baik class switching maupun
maturasi afinitas memerlukan keberadaan suatu enzim yang disebut activation-
induced cytosine deaminase (AID).31
Kulminasi aktivitas germinal center berupa pembentukan sel-sel B
memori dan long-lived plasma cells.32 Sejumlah regulator transkripsional terlibat
dalam perkembangan lanjut sel B (late B-cell development), diantaranya yakni
BLIMPI (B-lymphocyte maturation protein 1), IRF4 (interferon regulatory factor
4), dan XBP1 (X-box-binding protein 1). Sel plasma dapat saja berkembang dari
sel B memori atau dapat berkembang dari sel intermediat, plasmablast. Long-lived
plasma cells bermigrasi menuju dan hidup dalam sumsum tulang, dimana
ditempat tersebut itulah mereka akan bertahan. Sel plasma yang berada pada
sumsum tulang tersebut merupakan sumber utama dari antibodi spesfik antigen
(antigen-spesific antibody) yang beredar dalam sirkulasi.
Sebagaimana yang diuraikan dalam Bagian “Pengikatan Antigen oleh Sel
B”,respons sel T terbatasi hanya pada sebagian besar jenis peptida. Sehingga,
respons sel B terhadap antigen nonprotein tidak dapat membangkitkan reaksi sel T
helper melalui mekanisme-mekanisme yang telah diuraikan sebelumnya.33 Pada
beberapa kasus terpilih, T-cell independent nonprotein antigen dapat menginduksi

terjadinya class switching, tetapi umumnya, T-cell independent responses yang
terjadi ditandai oleh antibodi IgM dengan afinitas yang lebih rendah. Satu tipe T-
cell independent B-cell response menghasilkan satu substansi yang dikenal
sebagai natural antibodies – yakni, berupa antibodi IgM yang sebagian besarnya
berupa antibodi antikarbohidrat yang diproduksi tanpa adanya paparan terhadap
antigen.34 Natural antibodies ini diproduksi dalam jumlah yang terbatas dan
secara primer diproduksi oleh sel-sel peritoneal B1 atau secara spontan atau
sebagai respons terhadap bakteria yang hidup berkolonisasi pada saluran
pencernaan. Diperkirakan sel-sel zona marginal B yang terletak berdekatan
dengan sinus marginal pada splen juga memproduksi natural antibodies ini.
Melekatnya antigen pada tempat pengikatan antibodi (antibody-binding
sites) pada sel-sel B dapat menimbulkan fungsi fungsional, yang dikenal sebagai
fungsi efektor (effector functions). Dengan pengecualian berupa netralisasi
antigen direk melalui pengikatan antibodi (direct neutralization of antigen by
antibody binding), fungsi efektor biasanya difasilitasi melalui pengikatan Ig pada
FcR.35,36 FcR dapat dikategorikan ke dalam jenis yang dapat memicu aktivasi sel
dan jenis yang tidak memicu aktivasi sel. Jenis yang dapat memicu aktivasi
umumnya mengandung 1 atau lebih motif yang dikenal sebagai immunoreceptor
tyrosine-based inhibition motif. FcR yang tidak mengaktivasikan maupun
menginhibisi aktivasi sel terlibat dalam transport Ig melalui epitelia dan
prolongasi/pemanjangan waktu paruh IgG.
Fungsi efektor antibodi dalam mengeliminasi antigen diinisiasi oleh
respons imunitas dan juga di-downregulate oleh respons imunitas ketika aktivasi
tidak perlu dilanjutkan. Fungsi efektor
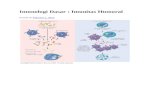
JALUR ALTERNATIF
Gambar 37-2

Langkah pertama dalam aktivasi jalur alternatif yaitu pengikatan C3b ke
permukaan sel seperti permukaan sel bakteri. C3 yang intak, merupakan molekul
inaktif, namun terdapat kadar yang rendah dari pembelahan spontan C3 yang
disebut dengan tickover, yang menyebabkan frgamen C3b selalu tersedioa. C3b
dapat berikatan dengan stabil pada permukaan sel melalui adanya interaksi dari
kelompok tioester C3b dan kelompok hidroksil dari permukaan sel. Pada C3 yang
intak, domain tioester ditutupi oleh residu hidrofobik yang mencegah hidrolisis
dari ikatan tioester. Domaian anafilaksis (ANA) dari C3 menstabilkan
konformasi inaktif tersebut. Ketika domain ANA membelah untuk mengeluarkan
C3a, kelompok tioester menjadi terpajan dan terjadi perubahan konformasional
yang menyebabkan munculnya kemampuan untuk berikatan pada permukaan
sel50,51. Apabila ikatan kimia tersebut tidak terjadi, kelompok tioester akan
dihidrolisasi dan C3b akan diinaktifkan.
Sekali saja pelekatan stabil dari C3b pada permukaan sel terjadi, protein
plasma yang disebut dengan Factor B akan berikatan pada C3b. Kemudian
dengan adanya faktor D, faktor B akan membelah menghasilkan Bb dan Ba.
Kompleks C3b dengan Bb distabilkan oleh protein plasma properidin, yang
merupakan jalur alternatif konvertase C3 (menyebabkan pembelahan C3 menjadi
C3a dan C3b). Hasil dari aktivitas C3 konvertase yaitu amplifikasi jalur melalui 2
atau 3 urutan. Penambahan C3b ke dalam kompleks tersebut akan membentuk
C3bBbC3b, yang mengubah jalur alternatif tersebut menjadi C5 konvertase.
Langkah yang lebih akhir dari aktivasi komplemen setelah pembelahan C5,
bersifat umum untuk ketiga jalur tersebut dan dijelaskan di dalam langkah akhir
dari aktivasi komplemen.
Oleh karena itu, kadar rendah dari C3b dalam plasma bertindak sebagai
mikronba sentinel. Sekali C3b berikatan pada permukaan sel, interaksi molekul
selanjutnya menghasilkan amplifikasi substansial dari jalur alternatif dan
pembelahan C5. Syarat untuk pengikatan C3b ke elemem struktural yaitu untuk
membatasi efek dari aktivasi komplemen hanya pada daerah dimana komplemen
diaktivasi. Jalur alternatif dari aktivasi komplemen tidak membutuhkan

pengenalan spesifik dari antigen dan oleh karena itu dianggap sebagai komponen
imunitas alami. Dan jika pengenalan spesifik tidak dibutuhkan, maka C3b dapat
berikatan pada sel manusia juga ke sel mikroba. Namun, aktivasi pada sel manusia
umumnya dicegah oleh adanya intervensi dari protein regulator yang berada pada
permukaan sel manusia yang melindungi sel tersebut dari serangan yang tidak
sesuai dan berbahaya52.
JALUR KLASIK
Gambar 37-3.
Langkah awal dalam aktivasi jalur klasik secara karakterisitik ditandai dengan
pengikatan dari bagian kompleks C1 yang disebut C1q pada antibodi IgG atau
IgM46. Molekul C1q terdiri dari 6 lengan identik yang melekat pada struktur
sentral. Akhir globular dari lengan tersebut, melekat pada regio pengikatan
komplemen dari sisi rantai berat dari kelas Ig tertentu. Agar C1q dapat diaktifkan,
maka zat tersebut harus berikatan secara terus menerus dengan setidaknya 2 buah
rantai berat Ig. Hal tersebut berarti bahwa untuk menimbulkan efek, Ig harus
sudah mengikat antigen. Pada kasus IgG, pengikatan dari epitop multipel oleh
antibodi menghasilkan proksimitas yang dekat dari antibodi dan oleh karena itu
dapat membentuk konfigurasi yang sesuai untuk aktivasi C1q (pengecualian untuk
IgG4). Karena IgM berada dalam bentuk pentamer, secara teoritis IgM tanpa
berikatan dengan antigen dapat mengaktifkan komplemen. Namun, ketika IgM
tidak berikatan dengan antigen maka tempat pengikatan C1q tidak dapat diakses.
Ketika antigen diikat, maka perubahan konformasional akan menghasilkan
pajanan dari tempat pengikatan C1q.
Komponen komplemen dari C1 merupakan kompleks dari C1q, C1r dan
C1s. Pengikatan dari 2 atau lebih dari kepala globular C1q akan mengakibatkan
aktivasi dari C1r. C1r yang teraktivasi merupakan protease yang menyebabkan
pembelahan dan aktivasi dari C1s, dan C1s teraktivasi, yang pada akhirnya akan
menyebabkan C4 membelah menjadi C4a dan C4b.

Penomoran protein komplemen berbeda dari posisinya dalam urutan
aktivasinya, karena komplemen ditemukan sebelum penemuan tentang kedudukan
mereka di dalam jalur aktivsinya. C4b sebagaimana C3b mengandung kelompok
tioester yang dapat membentuk ikatan yang stabil dengan kelompok hidroksil
dalam struktur tertentu. C4b yang terikat kemudian diikaat oleh C2, yang
kemudian membelah menjadi C2a dan C2b. Kompleks C4bC2b merupakan jalur
klasik C3 konvertase (Penambahan sisipan ‘a’ menandakan fragmen yang lebih
kecil dan sisipan ‘b’ menandakan fragmen yang lebih besar. Namun pengecualian
untuk C2, dimana C2a menunjukkan fragmen komplemen yang lebih kecil, dan
C2b mewakili fragmen yang lebih besar. Beberapa buku pada akhir-akhir ini
mengidentifikasi fragmen yang lebih kecil sebagai C2a dan fragmen yang lebih
besar sebagai C2b untuk mempertahankan konsistensi dengan nomenklatur
protein komplemen lainnya. Namun, banyak publikasi terbaru yang tetap
memakai nomenklatur yang lama).
Pembelahan C3 akan membentuk C3a dan C3b. Fragmen C3b kemudian
akan mengaktifkan komplemen melalui jalur alternatif, atau bertindak bersama
dengan C4bC2b untuk membentuk C4bC2bC3b, yang merupakan jalur klasik C5
konvertase.
Karakteristik utama dari jalur klasik sebagian besar mirip dengan jalur
alternatif. Aktivasi dari jalur tersebut membutuhkan adanya perlekatan dari
protein komplemen pada struktur seperti permukaan sel atau kompleks imun,
sehingga efek dari aktivasi komplemen menjadi terbatas. Langkah aktivasi awal
menyebabkan pembentukan C3 konvertase yang mebuat C3 membelah.
Pembelahan C3 mengakibatkan pembentukan C5 konvertase yang membuat C5
membelah menjadi C5a dan C5b. Perbedaan utamanya yaitu inisiasi dari jalur
klasik membutuhkan pengenalan spesifik dari antigen oleh antibodi. Hal tersebut
berkebalikan dengan jalur alternatif yang hanya membutuhkan pengenalan yang
bersifat kurang spesifik.
JALUR LEKTIN

Jalur lektin sangat mirip dengan jalur klasik, dengan perkecualian pada langkah
inisiasi. Langkah pertama merupakan pengikatan plasma lektin, mannose binding
protein (MBP) pada polisakarida di permukaan sel mikroba53. MBP, yang
merupakan anggota dari famili kolektin (kolagen lektin), secara struktural mirip
dengan C1q dan dapat berhubungan dengan C1r dan C1s. Perlekatan MBP pada
mikroba dapat memulai kaskade melalui aktivasi C1r dan C1s. MBP juga
berinteraksi dengan MBP-associatd-serine proteases (MASPs), yang merupakan
analog bagi C1r dan C1s. Pengikatan MBP pada permukaan sel mengakibatkan
pembelahan dari MBP-associatd-serine proteases (MASPs) dan pembelahan dari
C454. Pada kedua peristiwa tersebut akibat dari pembentukan C4b dan ikatannya
yang stabil pada permukaan sel. Sebagaimana kasus pada jalur alternatif, jalur
lektin juga merupakan komponen dari imunitas alami.
LANGKAH AKHIR DARI AKTIVASI KOMPLEMEN
Gambar 37-4
Jalur alternatif, klasik dan lektin mengakibatkan pembelahan dari C5.
Fragmen C5b tetap terikat pada permukaan sel. Langkah selanjutnya tidak
melibatkan pembelahan enzimatis namun merupakan pengikatan yang berurutan
dari C6, C7, dan C8 pada C5b. Kompleks C5b-8 secara stabil berikatan pada
membran sel yang kemudian menjadi membrane active complex (MAC) melalui
penambahan C9. C9 berpolimerisasi di sekitar kompleks dan membentuk pori-
pori pada membran sel. Pori-pori tersebut dapat mengakibatkan kematian melalui
ruptur osmotik terutama pada sel eritrosit tak berinti.
Sel berinti lebih resisten terhadap lisis, tapi tetap menampilkan efek yang
ditujukan pada pengikatan MAC55. Sangat mungkin bahwa perubahan non lisis
yang diinduksi oleh MAC bersifat lebih fungsional dan patologis dibandingkan
dengan lisis sel yang diinduksi langsung oleh MAC56. Efek non-lisis tersebut
dapat berbeda bergantung pada tipe sel dan karakteristiknya dan meskipun
terdapat efek yang sama akan menyebabkan hasil yang berbeda. Sebagai contoh
yaitu insersi MAC ke dalam membran sel fagosit dapat mengakibatkan produksi

mediator inflamasi seperti spesies oksigen reaktif dan prostaglandin, yang
menyebabkan aktivasi fagosit57. Sel epitel glomerulus juga dapat memicu produksi
mediator inflamasi, namun dalam konteks tersebut, mediator inflamasi dapat
menyebabklan kerusakan jaringan58. MAC juga telah dilaporkan menyebabkan
proliferasi sel tertentu dan dilaporkan juga bahwa MAC memiliki karakteristik
apoptosis maupun non-apoptosis55.
INISIATOR TAMBAHAN PADA AKTIVASI KOMPLEMEN
Selain karakteristik inisiator dari ketiga jalur yang telah disebutkan, struktur
tambahan tertentu dapat memicu aktivasi komplemen59. Hal tersebut termasuk: di
dalam jalur alternatif, yaitu kompleks imun IgA dan endotoksin; di dalam jalur
klasik yaitu C-reaktif protein, badan apoptosis, dan serum amiloid P; di dalam
jalur lektin yaitu serum fikolin (lektin yang berikatan pada N-acetylglucosamine).
FUNGSI PROTEIN KOMPLEMEN
Sebagaimana telah disebutkan di awal, fungsi paling awal dari sistem komplemen
yang ditemukan yaitu lisis bakteri. Pembunuhan mikroba melalui lisis langsung
dipeantarai oleh MAC, C5b-9. Mikroba juga dapat dihancurkan melalui proses
coating atau opsonisasi oleh komplemen dan fagositosis dari partikel
teropsonisasi oleh sel fagositik. Proses opsonisasi dan fagositosis juga merupakan
mekanisme untuk fungsi penting lainnya dari komplemen yang meliputi
pembersihan kompleks imun dan debris apoptosis dari sirkulasi.
Fungsi sekunder yang penting dari aktivasi komplemen yaitu sebagai
induksi inflamasi. Inflamasi yang ditandai dengan perubahan vaskular dan
aktivasi leukosist dan protein inflamasi menyebabkan augmentasi respon imun
lokal pada jaringan. Tiga mediator inflamasi yang muncul akibat aktivasi
komplemen yaitu fragmen komplemen C3a, C4a, dan C5a. Fragmen tersebut
disebut sebagai anafilatoksin karena kemampuannya dalam menginduksi
degranulasi sel mast, eosinofil, basofil, monosit , neutrofil, sel otot polos dan sel
epitel. Pengikatan dari C5a pada sel endotel mengakibatkan peningkatan
permeabilitas vaskular dan ekspresi dari P-selektin, dimana keduanya memicu

akumulasi leukosit pada jaringan. Pengikatan pada neutrofil menyebabkan
peningkatan motilitas neutrofil, adhesi pada sel endotel dan produksi spesies
oksigen reaktif. Hasil keseluruhan yaitu akumulasi sel inflamasi pada daerah lokal
dalam jaringan dimana sel tersebut dapat memfagosit mikroba secara efisien.
Aktivasi komplemen juga menghasilkan stimulasi sistem imun humoral
melalui produksi C3d. Sel B yang antibodi permukaan selnya mengenali antigen
yang terikat pada komplemen, mengalami peningkatan ekspresi oleh pengikatan
lanjut dari C3d pada CR2 di permukaan sel B. Opsonisasi oleh komplemen juga
memfasilitasi terjadinya presentasi antigen pada sel B oleh sel dendritik folikular.
RESEPTOR KOMPLEMEN
SEKILAS TENTANG RESEPTOR KOMPLEMEN DAN PROTEIN
REGULATOR
Beberapa efek penting dari komplemen diperantarai oleh pengikatan
protein komplemen pada reseptor komplemen. CR1 berfungsi dalam
proses fagositosis, pembersihan kompleks imun dan penurunan ekspresi
reseptor. CR2 penting dalam stimulasi imunitas humoral. CR3 dan CR4
memicu timbulnya fagositosis.
Regulasi aktivasi komplemen dilakukan oleh serum tertentu dan protein
permukaan sel. Kebanyakan protein regulator permukaan sel diekspresikan
pada sel manusia namun tidak diekspresikan pada mikroba. Hal tersebut
untuk melindungi sel manusia dari kerusakan komplemen.
Beberapa dari protein regulator menurunkan aktivasi komplemen melalui
penggantian komponen pada langkah awal dari kaskade aktivasi. Hal
tersebut termasuk inhibitor C1,faktor H, C4 binding inhibitor, faktor
penghancur akselerator, protein kofaktor membran dan CR1.
Terdapat beberapa protein yang berinteraksi dengan komplemen dan
menjadi pengatur dari fungsi komplemen. Reseptor α yang merupakan anggota
dari seven-transmembrane α helical G protein yang disebut dengan regulators of
complement activationein coupled receptor family yang telah disebutkan pada

bagian sebelumnya. Beberapa reseptor yang telah berhasil dijelaskan dengan baik
yaitu CR1-CR4. Reseptor komplemen tipe 1, CR1 (CD35), merupakan anggota
famili protein yang disebut sebagai regulator of complement activation (RCA),
yang berbagi struktur umum yang terdiri dari complement control protein
repeats52. CR mengikat C3b atau C4b dan diekspresikan pada sel darah perifer
termasuk monosit, limfosit T dan B, neutrofil, eosinofil, dan eritrosit; pada sel
dendritik folikular dan pada keratinosit62. Pada sel fagositik, pengikatan CR1 pada
C3b atau C4b menyebabkan fagositosis dari partikel yang teropsonisasi oleh
fragmen komplemen, serta aktivasi mekanisme mikrobisidal dalam sel fagositik.
Pada eritrosit, pengikatan CR1 pada C3b atau C4b yang dilapisi oleh kompleks
imun menyebabkan transpor kompleks tersebut menuju limpa dan hati, dimana
kompleks tersebut akan mengalami pembersihan dari sirkulasi oleh sel fagositk.
Oleh karena itu, CR1 memiliki peran penting sebagai mediator dari fungsi
komplemen. CR1 dapat menurunkan aktivasi komplemen, karena terlibat dalam
disosiasi dari kompleks C5 konvertase.
Reseptor komplemen tipe 2 yaitu CR2 (CD21) juga merupakan anggota
famili RCA59. CR2 mengikat fragmen C3 yaitu C3b, C3dg, dan C3d serta Epstein
Barr virus, interferon alfa dan protein imunoregulator yaitu CD23. CR2
diekspresikan pada subset limfosit T dan B, basofil, sel mast, sel dendritik
folikular, serta beberapa sel epitel dan keratinosit. Pada sel B, CR2 bertindak
sebagai koreseptor untuk aktivasi sel B. Ketika CR2 diikat oleh C3d, tingkat
aktivasi sel B akan naik sesuai dengan urutan ukurannya63. Pada sel sendritik,
pengikatan CR2 menyebabkan terperangkapnya kompleks imun pada pusat
germinal. CR2 juga berperan dalam presentasi antigen pada sel T.
Reseptor komplemen tipe 3, yaitu CR3 (CD11b/CD18, Mac-1) merupakan
molekul permukaan sel integrin yang diekspresikan pada monosit, neutrofil, sel
NK, dan sel mast64. Reseptor tersebut berfungsi untuk memicu fagositosis
mikroba melalui pengikatan pada iC3b (C3b inaktif) dan melalui ikatan langsung
pada mikroba. Reseptor tersebut berinteraksi dengan molekul adhesi intraseluler 1
yang diekspresikan secara endogen pada sel endotel untuk menstabilkan adhesi

leukosit pada endotelium yang pada akhirnya memfasilitasi perekrutan leukosit
dari sirkulasi ke dalam jaringan.
Reseptor komplemen tipe 4 , yaitu CR4 (CD11c/CD18), juga merupakan
molekul permukaan sel integrin. Reseptor tersebut diekspresikan pada monosit,
neutrofil, sel NK, dan sel dendritik serta memiliki fungsi yang mirip dengan CR3.
Diantara reseptor komplemen yang baru-baru ini dideskripsikan, yaitu
SIGN-RI,65 yang berikatan pada C1qq dan diekspresikan pada makrofag limpa
zona marginal, dan CR1g (reseptor komplemen dari famili imunoglobulin)66, yang
berikatan pada C3b dan iC3b serta diekspresikan pada subset jaringan makrofag
pendatang.
REGULASI AKTIVASI KOMPLEMEN
Molekul yang terlibat dalam regulasi aktivasi komplemen berfungsi untuk
menurunkan ekspresi respon imun saat respon imun sudah tidak lagi dibutuhkan
dan untuk membatasi respon imun hanya pada daerah dimana C1 berada dalam
bentuk kompleks C1q dengan sebuah tetramer yang terdiri dari 2 buah fragmen
C1r dan 2 buah C1s. Ketika C1q mengikat antibodi, C1 INH dapat bertindak
untuk membatasi aktivasi komplemen dengan berikatan pada tetramer C1rC1s
sehingga membuatnya berdisosiasi dari C1q dan mencegah aktivasi selanjutnya
dari jalur komplemen tersebut.
Poin utama lain yang penting dalam interaksi dari protein regulator yaitu
adanya pengikatan dengan C3b atau C4b52. Seperti yang telah dijelaskan di atas,
kelompok tioester yang belum berikatan dengan C3b atau C4b akan dihidrolisasi
dengan cepat sehingga membuatnya menjadi inaktif. Untuk ikatan permukaan
dengan C3b atau C4b, proses inaktivasi terjadi melalui pemindahan komponen
jalur alternatif atau jalur klasik C3 konvertase dari C3b atau C4b. Pada jalur
alternatif, inaktivasi dari kompleks C3 konvertase yaitu C3bBb, dapat terjadi
melalui pemindahan Bb dari dari C3 oleh faktor plasma protein H atau protein
permukaan sel bernama decay accelerating factor (DAF, CD55), membrane
cofactor protein (MCP,CD46) dan CR1. Ketiga protein permukaan sel tersebut

merupakan anggota dari famili RCA, yang diekspresikan pada sel manusia,
namun tidak diekspresikan pada mikroba, dan oleh karenanya perlindungan
terhadap kerusakan sel manusia akibat aktivasi komplemen dapat tetap dilakukan.
Sementara itu proses pembunuhan sel mikroba juga berlangsung. Faktor H lebih
suka untuk berikatan pada permukaan sel dengan kadar asam sialit yang tinggi
pada sel manusia, namun tidak pada mikroba, dan selanjutnya akan menurunkan
aktivitas komplemen pada sel manusia. Dalam jalur klasik dan lektin, C3
konvertase yang berperan yaitu C4bC2b, DAF, MCP, dan CR1 yang
menggantikan C2b dari C4b, serta plasma protein C4 binding (C4BP). Oleh
karena itu, protein sel permukaan yaitu DAF, MCP dan CR1 dapat mendisosiasi
C3 konvertase baik dari jalur alternatif, klasik maupun lektin, sedangkan faktor
protein plasma H dan C4BP bersifat spesifik untuk jalur alternatif atau
klasik/lektin. Proteolisis dari C3b atau C4b diperantarai oleh faktor I yang
merupakan protein plasma yang memerlukan kofaktor untuk aktivitasnya. MCP,
CR1, faktor H dan C4BP dapat bertindak sebagai kofaktor untuk faktor I.
Regulasi dari komplemen pada langkah akhir diperantarai oleh CD59,
yang merupakan protein sel permukaan yang diekspresikan pada sel manusia,
namun tidak diekspresikan pada mikroba. Senyawa tersebut berikatan pada
kompleks C5b8 dan menghambat penambahan C9, sehingga memblok
pembentukan MAC. Protein plasma S berikatan pada kompleks C5b7 dan
memblok insersi ke dalam membran sel serta menghambat polimerisasi C967.
MACs intak dapat dihilangkan dari sel melalui pemutusan pada transpor sel
membran atau melalui internalisasi dan degradasi55.
Karboksipeptidase N dapat menghilangkan arginin terminal dari C3a, C4a,
dan C5a dan disebut dengan istilah ANA inactivator68. Karboksipeptidase R juga
telah ditunjukkan dapat menghilangkan arginin terminal dari C3a dan C5a69.
KOMPLEMEN DAN PENYAKIT
ABNORMALITAS GENETIK DARI SISTEM KOMPLEMEN

Defisiensi protein yang terlibat dalam kaskade komplemen, reseptor komplemen
atau protein regulator komplemen dapat mengakibatkan berkembangnya berbagai
penyakit70. Defisiensi genetik dari komplemen telah dikaitkan dengan peningkatan
resiko infeksi dan autoimunitas. Sebagai contoh, defisiensi dari berbagai
komponen komplemen terutama komponen komplemen tahap awal yaitu C1-C4,
telah dikaitkan dengan kejadian systemic lupus eritematosus (SLE), sementara itu
defisiensi C3 telah dikaitkan dengan infeksi piogenik yang mengancam nyawa,
dan defisiensi C5-C9 telah dikaitkan dengan infeksi Neisseria. Defisiensi genetik
dari mannose-binding lectin bersifat umum, dengan kadar rendah yang terjadi
dalam jumlah sekitar 10% pada ras Kaukasian, dan berkaitan dnegan peningkatan
resiko infeksi serta autoimunitas termasuk SLE71,72.
Perubahan ekspresi dari CR3 (CD11b/CD18) dan CR4 (CD11c/CD18)
terjadi pada kondisi defisiensi-1 leukosit, yaitu sebuah kelainan kongenital akibat
mutasi pada gen yang mengkode CD18. Mutasi pada CD18 juga mempengaruhi
ekspresi CD11a/ CD18 (leukocyte function associated antigen 1). Pasien dengan
defisiensi-1 adhesi leukosit, akan menunjukkan abnormalitas yang signifikan dari
adesi leukosit dan akan mengalami infeksi yang berulang. Defisiensi dari protein
regulator komplemen C1 INH akan menyebabkan angioneurotik edema sebagai
akibat dari aktivasi jalur klasik yang kurang teratur dan kelebihan bradikinin
akibat aksi kalikrein dan faktor XII. Angioedema juga dapat terjadi jika seseorang
memiliki kadar inaktivator ANA yang sangat kurang yaitu karboksipeptidase N73.
Defisiensi protein yang diperlukan untuk ekspresi yang sesuai dari DAF
dan CD59 pada permukaan sel berkaitan dengan paroxismal nocturnal
hemoglobinuria, suatu penyakit yang ditandai dengan lisis eritrosis yang
diperantarai oleh komplemen74. Sindrom uremik hemolitik juga dikaitkan dengan
mutasi pada MCP, faktor H dan faktor I75. Defisiensi faktor H juga telah dikaitkan
dengan membranoproliferatif glomeruonefritis.

Resiko untuk mengalami degenerasi makula akibat penuaan juga
dipengaruhi oleh adanya polimorfisme gen tertentu dari sistem komplemen,
terutama komplemen faktor H dan juga faktor B dan C276.
Defisiensi genetik dari komponen komplemen telah dikaitkan teruatama
dengan kerentanan terkena penyakit infeksi atau autoimunitas. Defisiensi
genetik dari protein regulator komplemen dapat mengakibatkan
perpanjangan aktivasi komplemen seperti yang terjadi pada kasus
defisiensi inhibitor C1.
Komplemen berkaitan dengan SLE melalui beberapa mekanisme yang
mungkin. Hal tersebut termasuk peningkatan resiko autoimunitas yang
terjadi akibat defisiensi komplemen tertentu, kerusakan jaringan yang
disebabkan oleh aktivasi komplemen yang diinduksi autoantibodi,
pembersihan yang tidak efektif dari debris apoptosis yang memicu
autoimunitas dan kegagalan dalam mengeliminasi sel reactive B cells.
Aktivasi komplemen dianggap terlibat dalam patogenesis aterosklerosis,
reperfusi kerusakan setelah infark miokard akut, penyakit diabetik
mikrovaskular, dan infark serebral pada stroke iskemia.
Agen infeksius tertentu menunjukkan mekanisme penghindaran dari
destruksi oleh komplemen, dan beberapa menggunakan reseptor
komplemen atau protein regulator untuk mendapatkan jalan masuk ke
dalam sel.
KOMPLEMEN, SYSTEMIC LUPUS ERYTHEMATOSUS (SLE) DAN
AUTOANTIBODI
Komplemen diduga kuat berhubungan dengan SLE, melalui jalur yang tampaknya
masih paradoks77,78. Defisiensi genetik dari komponen komplemen berhubungan
dengan SLE, namun beberapa dari kerusakan jaringan yang terlihat pada SLE
tampaknya diperantarai sebagian oleh aktivasi komplemen. Oleh karena itu,
komplemen tampaknya bersifat protektif dan mendelesi secara terus-menerus.
Pengamatan tersebut menggarisbawahi peran protean dari komplemen dalam

sistem imun. Aktivasi komplemen berpotensi untuk mengakibatkan kerusakan
jaringan, namun komponen komplemen mungkin berguna dalam pembersihan
kompleks imun dan debris apoptosis. Badan apoptosis yang tidak dibersihkan
secara efektif, akan memicu autoimunitas melalui presentasi autoantigen yang
telah mengalami sekuesterisasi ke dalam sistem imun. Komplemen juga dianggap
berpartisipasi dalam mengeliminasi self-reactive immune B cell, yang menjadi
mekanisme dasar dalam menimbulkan efek protektif komplemen pada SLE78.
Perubahan ekspresi dari CR1 maupun CR2 telah diamati pada pasien
dengan SLE79. Dalam model murin lupus, tikus dengan kekurangan ekspresi CR1
dan CR2 (yang terletak pada gen yang sama dalam tikus dan diproduksi melalui
pemecahan alternatif) menunjukkan peningkatan autoimunitas . Penemuan
tersebut menunjukkan bahwa interaksi antara CR2 dengan C3d, berperan penting
dalam respon sel B terhadap antigen, yang merupakan faktor penentu lain
terhadap kerentanan terkena SLE78. Autoantibodi terhadap C1q bersifat umum
pada SLE dan telah dikaitkan dengan beberapa penyakit renal berat, kemungkinan
melalui sebuah reaksi efek samping dalam proses pembersihan kompleks imun
atau badan apoptosis81. Faktor nefritik C3 merupakan autoantibodi terhadap C3
konvertase, yaitu C3bBb, yang bertindak untuk menstabilkan kompleks tersebut.
Signifikasnsi klinis tersebut yaitu hubungannya dengan membranoproliferatif
glomerulonefritis tipe II dan lipodistrofi parsial.
KOMPLEMEN DAN PENYAKIT VASKULAR
Aktivasi komplemen telah terbukti terlibat dalam patogenesis aterosklerosis,
reperfusi kerusakan setelah iskemia miokardia82,83. Pada penyakit mikrovaskular
proliferatif yang berhubungan dengan diabetes, glikation, dan inaktivasi dari
CD59 dapat mengakibatkan proliferasi seluler akibat efek proliferatif non lisis
dari MAC84. Aktivasi komplemen juga telah terbukti berperan dalam penolakan
hiperakut dari xenotransplan akibat adanya antibodi alami terhadap komponen
dari sel endotel dari organ transplan, yang ditandai dengan adanya aktivasi
komplemen, kerusakan sel endotel, dan koagulasi intravaskular.

MEKANISME PENGHINDARAN KOMPLEMEN OLEH MIKROBA
Pengamatan pada individu dengan defisiensi komponen tahap akhir dari aktivasi
komplemen menunjukkan bahwa individu tersebut beresiko tinggi untuk terkena
serangkaian infeksi yang terbatas. Fenomena tersebut membuktikan bahwa agen
infeksius mengadakan mekanisme penghindaran terhadap destruksi komplemen85.
Bakteri gram positif memiliki dinding yang tebal yang sulit untuk ditembus oleh
MAC46. Protein M Streptococcus grup A yang mengikat faktor H, yang
menurunkan aktivasi komplemen dan patogen lainnya, memiliki mekanisme
penghindaran untuk menarik faktor H melalui ekspresi asam sialik pada
permukaannya. Staphylococcuc aureus mengekspresikan beberapa protein yang
menghambat aktivasi C3. Sebuah protein vaccinia virus (VCP-1, vaccinia virus
complement control protein-1) bertindak sebagai kofaktor untuk faktor I, yang
mengakibatkan terjadinya proteolisis dari C3b dan C4b. Pada infeksi human
immunodeficiency virus (HIV), inklusi dari molekul penurun aktivasi sistem
komplemen ke dalam virus atau membran sel host menyebabkan HIV
menghindari destruksi yang diperantarai oleh komplemen86. DAF dan CD59 dapat
dimasukkan ke dalam membran virus HIV karena pembentukan tunas dari sel
manusia yang terinfeksi, dan faktor H yang dapat berikatan pada glikoprotein
permukaan HIV di dalam sel manusia yang terinfeksi.
Sistem komplemen telah direduksi oleh agen infeksius tertentu agar dapat
masuk ke dalam sel. Epstein-Barr virus berpenetrasi ke dalam sel B melalui
pengikatan terhadap CR2 pada permukaan sel B46. Virus campak berikatan pada
sel melalui MCP. Mycobacteria membuat molekul mirip C4 yang berikatan
dengan C2b dan kemudian membelah menjadi C3. Deposisi dari C3b pada
membran sel mikobakterium yang mengakibatkan influks ke dalam makrofag,
yang berada dalam bentuk parasit intraseluler. Pengetahuan mengenai strategi
penghindaran patogen tersebut dapat berguna dalam merancang vaksin dan target
terapi85.