







Bahasa
Halaman
Hukum


www.elsevier.com/locate/apsoil
Applied Soil Ecology 35 (2007) 340–355
Successional changes in soil, litter and macroinvertebrate
parameters following selective logging in a Mexican Cloud Forest
Simoneta Negrete-Yankelevich a,b,*, Carlos Fragoso b,Adrian C. Newton c, O. William Heal d
a School of Geosciences, University of Edinburgh, Crew Building, The King’s Buildings, West Mains Road, Edinburgh EH9 3JU, UKb Departamento de Biologıa de Suelos, Instituto de Ecologıa, A.C. Km. 25 Carretera Antigua a Coatepec, #351,
Congregacion El Haya, 91070 Xalapa, Veracruz, Mexicoc School of Conservation Sciences, Bournemouth University, Talbot Campus, Poole, Dorset BH12 5BB, UK
d Department of Biological Sciences, University of Durham, Durham DH1 3LE, UK
Received 19 October 2005; received in revised form 22 June 2006; accepted 24 July 2006
Abstract
The environmental and vegetation shifts associated with logging disturbance and secondary succession in Tropical Montane Cloud
Forests have been studied in detail, however little is known about the consequences that these changes have for the soil system. The
present study was undertaken to determine the impact of selective logging and subsequent secondary succession on soil micro-
environmental conditions, leaf litter quality and quantity, soil nutrient concentration and soil and litter macroinvertebrate community
composition. The study was carried out in three successional chronosequences, two recently logged sites and two pristine tropical
mountain cloud forest sites in Oaxaca, Mexico. Results showed that selective harvesting of Quercus spp. trees caused an increase in soil
temperature of ca. 4 8C that is not completely reversed after 100 years of succession. During 100 years of secondary succession litter
diversity increased and soil organic matter accumulated (16.4% increase in total C). The availability of cations (Ca, Mg, Na, and K) in
the topsoil decreased by more than 50% as a result of logging, and only Mg increased again between 75 and 100 years after disturbance.
Pristine cloud forests sustain a diverse litter and soil macroinvertebrate community, but its composition and diversity was
negatively affected by logging. The effect of Quercus harvesting activities on the litter community was apparent within 2 months of
disturbance (total abundance declined by ca. 65%, higher taxa richness by ca. 10% and diversity by ca. 35%). For the soil
community there was a time-lag in the effect of logging. Two months after disturbance there was no significant effect on the soil
community but 15 years after abandonment, total macroinvertebrate abundance in the soil was ca. 80% lower and higher taxa
richness ca. 30% lower compared to undisturbed sites. Full recovery of the macroinvertebrate community composition appeared to
take more than 100 years both in the litter and soil. Reduced abundances of Coleoptera and Enchytraeidae were apparent even after
100 years of succession. The endemic earthworm Ramiellona wilsoni was found almost exclusively in the pristine forests and
therefore its abundance could be used as a sensitive indicator of disturbance in these forests.
# 2006 Elsevier B.V. All rights reserved.
Keywords: Earthworms; Firewood extraction; Litter quality; Tropical montane forest; Soil macrofauna; Soil temperature
* Corresponding author at: Departamento de Biologıa de Suelos,
Instituto de Ecologıa, A.C. Km. 25 Carretera Antigua a Coatepec,
#351, Congregacion El Haya, 91070 Xalapa, Veracruz, Mexico.
Tel.: +52 55 228 842 18 50.
E-mail address: [email protected]
(S. Negrete-Yankelevich).
0929-1393/$ – see front matter # 2006 Elsevier B.V. All rights reserved.
doi:10.1016/j.apsoil.2006.07.006
1. Introduction
Tropical Mountain Cloud Forest (TMCF) is one of the
most endangered ecosystems of the world (Rzedowski,
1996; Bruijnzeel and Veneklaas, 1998). In many of these

S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355 341
forests in Mexico, logging for firewood extraction is
chronic, typically involving selective harvesting of
Quercus spp. trees (Ramırez-Marcial et al., 2001).
Despite being the most floristically diverse forest in
Mexico, the TMCF is one of the least studied forest
ecosystems in the country (Ramırez-Marcial et al., 2001;
Velazquez et al., 2003) and very little is known about the
impact of accelerated logging disturbance on the below-
ground subsystem, either in Mexico or in TMCF
elsewhere.
It has been widely recognised that forest disturbance
can affect nutrient cycling (Nilsson et al., 1995; Finer
et al., 2003) and soil biota (Davies et al., 1999; Lavelle,
2000; Brown et al., 2001; Pietikainen et al., 2003).
Logging disturbance can have short- and long-term
impacts on the below-ground subsystem. In the short-
term (within a few months), harvested plant residues
produce an above-ground flush of nutrient-rich organic
matter (Olsson et al., 1996a,b; Finer et al., 2003) and a
below-ground litter input from dead rooting systems.
The impact of logging activities may be particularly
important in forests growing on poor soils such as the
TMCF. In these ecosystems, plants are adapted to use
nutrients efficiently, decomposition is slow and most
nutrients are sequestered in plant and microbial biomass
(Waide et al., 1998; Hobbie, 1992; Vitousek, 1984;
Bruijnzeel and Proctor, 1995; Tanner et al., 1998). The
sudden input of nutrient-rich residues together with the
increase in radiation reaching the soil may bring a
temporary increase in the rate of decomposition and
increase nutrient availability (Butterfield, 1999; Siira-
Pietikainen et al., 2001; Finer et al., 2003), particularly
in the topsoil where most plant–soil biochemical
interactions occur (Gross et al., 1995). Below-ground
food-web responses to logging are poorly understood
(Bengtsson et al., 1997; Wardle et al., 1998). However,
the rise in availability of nutrient-rich organic matter
may also decrease the proportion of fungal-based over
bacterial-based food-webs (Wardle, 1992; Siira-Pieti-
kainen et al., 2001) and produce major changes in
macroinvertebrate community composition.
In the long-term (tens of years) the effects of logging
often include a delayed response of the soil system to
initial disturbance (Bengtsson et al., 1997; Zaitsev et al.,
2002) or an indirect consequence of successional changes
in the vegetation community composition after abandon-
ment (Switzer and Shelton, 1979; Gross et al., 1995).
Although the environmental and vegetation shifts
associated with post-logging secondary succession in
TMCF have been studied in detail (Gonzalez-Espinosa
et al., 1991; Quintana-Ascencio and Gonzalez-Espinosa,
1993; Romero-Najera, 2000; Blanco-Macias, 2001;
Galindo-Jaimes et al., 2002), little is known about the
consequences of these long-term changes on nutrient
budgets and faunal communities below-ground. In
Mexico, tree species of Holarctic phytogeographic origin
(such as Pinus spp.) are often pioneers in logged TMCFs
and are known to be more productive than hardwoods of
tropical origin (Williams-Linera and Toledo, 1996),
therefore they could potentially increase the rate of the
decomposition process and enhance nutrient availability.
Furthermore, after disturbance and in early succession,
the diversity of tree species diminishes relative to older
forests (Ramırez-Marcial et al., 2001), leading to a
decline in the diversity of resources available to the soil
system. This, together with the greater abundance of
resources, might be expected to lead to a more uniform
and less diverse soil macroinvertebrate fauna.
This paper presents a study of the litter composition,
topsoil nutrient concentration and soil macroinverte-
brate fauna in two pristine and two recently logged sites
as well as three successional chronosequences (15 to
100-year-old forests) of TMCF in Oaxaca, Mexico. The
objectives of the study were:
i. T
o determine whether changes happening above-ground as a result of logging and subsequent
secondary succession are coupled with changes in
microenvironmental conditions, litter quality and
quantity and soil nutrient concentration.
ii. T
o determine if the composition and diversity of thehigher taxa macroinvertebrate community reflect
these changes and can therefore serve as good
indicators of disturbance and degree of recovery of
the soil system.
2. Methods
2.1. Study sites
The research was carried out in an area known as El
Rincon (Villa Alta District), in the Sierra Norte of
Oaxaca, Mexico. The study sites were three secondary
successional chronosequences of Tropical Montane
Cloud Forest (TMCF) (Challenger, 1998) named here as
Tanetze, Juquila and Tarbis. Each of these chronose-
quences is formed by four sites of different successional
stage: approximately 15, 45, 75, and 100 years of age.
Additionally two un-logged forests (Pris and Pris II,
from here on named pristine) and two plots that were
selectively harvested for Quercus spp. trees 2 months
before sampling (Tar 0 and Tar 00) were examined. All
sites are located between 178180 and 178230N and
968150 and 968210W. The successional stages within

S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355342
each chronosequence are a short distance from each
other (less than 2000 m). Study sties are situated in the
following altitudes: Juquila 1700–1975 m, Tanetze
1700–1860 m, Tarbis 1700–1975 m, Tar 0 and Tar 00
1975 m and Pris and Pris II 2100 m. The mean annual
precipitation at the nearest meteorological station
(�16 km from the area) is 1719 mm year�1. The mean
annual temperature ranges between 20 and 22 8C(Anonymous, 1999).
Among the distinctive species in the original TMCF
in the area are Billia hyppocastanum, Cinnamomum
zapatea, Oreopanax flaccidus, Podocarpus matudae,
Quercus laurina, Symplocos coccinae, Ternstroemia
oocarpa and Beilschmedia ovails (Cordova and del
Castillo, 2001; Bautista-Cruz and del Castillo, 2005).
The approximate age of the forests in each of the studied
chronosequences was identified by del Castillo (1996)
and Bautista-Cruz and del Castillo (2005), based on the
age of the oldest Pinus chiapensis trees. Selective
logging (as well as clearance) of pristine cloud forests in
Mexico leads to the rapid colonisation by pine species
(Challenger, 1998), which permits a relatively precise
determination of the time since the last major
disturbance by calculating the age of the oldest pine
trees.
2.2. Field methods
Sampling was conducted in 30 m � 30 m grids
(with 49 intersections every 5 m) established in each
successional stage of all chronosequences, and in
recently logged and pristine sites. First an intensive
survey was carried out in the Juquila chronosequence
between 11 July and 17 August 2000. In this period
all 49 intersections of each of the four grids (15-, 45-,
75- and 100-year-old forests) were sampled. The
following year, between 25 June and 3 December
2001, all of the successional stages of Tanetze and
Tarbis, recently logged sites (Tar 0 and Tar 00) and
the pristine sites (Pris and Pris II) were sampled. On
this occasion, only seven random vertices in each grid
were selected for sampling. Both of these field
seasons were included in the rainiest half of the year.
In order to preserve a balanced design, for compar-
isons including all chronosequences (Juquila, Tanetze
and Tarbis) and sites (Tar 0, Tar 00, Pris and Pris II),
only seven randomly selected samples were consid-
ered for each grid in the Juquila chronosequence. For
the detailed analysis performed exclusively in the
Juquila chronosequence (analysis of litter components
and soil nutrient content) all 49 samples were
considered.
In all of the field sites except Juquila, sampling at
each grid intersection consisted of the simultaneous
extraction of a soil monolith and the measurement of
microenvironmental conditions. In the Juquila chron-
osequence, microenvironmental conditions were mea-
sured in each successional stage on four different
sampling dates (11 and 18 July and 14 and 17 August
2000). The point measurements for each sampling
session were carried out within the same day and the
four repetitions for each sampling vertex were
averaged.
2.3. Soil microenvironment
Soil temperature, volumetric soil water content and
canopy cover (as a surrogate for solar radiation reaching
the soil) were considered as indicators of soil
microenvironmental conditions. For soil water content
and temperature a Delta-T-Theta Probe soil moisture
sensor attached to a Delta-T-HH1 meter (Delta-T
Devices Ltd., Cambridge, UK) and a Taylor soil digital
thermometer (model 9840; Taylor Precision Products
LP, Oak Brook, USA) were used, respectively. The final
calculation of volumetric soil water content was based
on a soil-specific calibration (Delta-T Devices Ltd.,
1999). Both the temperature and humidity sensor probes
were placed at a soil depth of 10 cm. Percent canopy
cover was measured with a Convex Spherical Crown
Densiometer (Forestry Suppliers Inc., Jackson, USA),
facing north and holding the densiometer at breast
height, above the sampling point.
2.4. Macroinvertebrate sampling
All monoliths were extracted with a box corer and
consisted of 25 cm � 25 cm � 5 cm depth of soil, plus
all the litter above it. The litter and soil sample was hand
sorted in situ for macroinvertebrates (defining macro-
invertebrates as all invertebrate animals larger than
2 mm in any of its dimensions). The macroinvertebrates
found in the monolith were preserved in 70% alcohol.
All macroinvertebrates collected were counted under
a dissecting microscope and classified into class, order
and groups of immature stages. Additionally all earth-
worms were identified to species following Fragoso and
Reynolds (1997). Although Enchytraeidae are currently
considered mesofauna, we have included this taxon in the
analysis because they were a particularly conspicuous
component of the faunal community and they have been
rarely studied in the temperate tropics (Rombke, 2003).
Two estimates of compositional diversity were calculated
for the macroinvertebrate community: the number of

S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355 343
elements (macroinvertebrate taxa) and the Shannon–
Weiner index (Magurran, 1996).
2.5. Vegetation census
Within the boundaries of the Juquila sampling grids,
as well as in a 5 m width frame around them, we
recorded the genus and diameter at breast height (DBH)
of all the live trees (with a DBH > 5 cm). Assuming a
cylindrical shape, the basal area per genus and per
successional stage, was calculated.
2.6. Litter resource availability and quality
In the laboratory the litter and soil samples were
oven dried at 80 8C to constant mass. The dry mass of
the litter in each monolith was recorded. The litter of all
the monoliths extracted from the Juquila chronose-
quence was sorted into six components: Pinus needles,
Quercus leaves, Lauraceae leaves including the three
genera present in all chronosequences – Persea, Ocotea
and Beilschmedia – woody and reproductive material,
leaves from other genera and unrecognisable leaf
material. The dry mass of these components was
recorded separately.
Recently fallen leaves of P. chiapensis, Oreopanax
xalapensis, Beilschmedia ovalis and Quercus laurina
were collected from the forest floor at Juquila, dried to
constant mass and analysed for litter quality. Leaf litter
quality determination consisted of the following
analyses: (1) total C concentration using the loss on
ignition method (Jackson, 1958); (2) total N concentra-
tion with sulphuric acid digestion followed by distilla-
tion and titration (Anderson and Ingram, 1993); (3) total
P concentration using sulphuric acid digestion followed
by colorimetric determination (Anderson and Ingram,
1993); (4) cation concentrations (Ca2+, Mg2+, K+ and
Na+) using a digestion with HNO3 and HClO4 followed
by atomic absorption spectroscopy (Allan, 1971); (5)
lignin content with a sequential procedure of neutral
detergent fibre (NDF) and acid detergent fibre (ADF)
extractions followed by acid detergent lignin extraction
with 72% sulphuric acid (Van Soest, 1994).
2.7. Soil nutrients
All soil samples from the Juquila chronosequence
were analysed for: (1) total C using the loss on ignition
method to determine total organic matter and dividing
by 1.742 to calculate total carbon concentration
(Jackson, 1958), (2) total P concentration using
sulphuric acid digestion followed by colorimetric
determination (Anderson and Ingram, 1993) and (3)
exchangeable cations (Ca2+, Mg2+, K+ and Na+) with
the ammonium acetate extraction method at pH 7
(Anderson and Ingram, 1993). Additionally, 10 ran-
domly chosen samples from the 49 intersections of each
successional stage were analysed for total N with acid
digestion followed by distillation and titration (Ander-
son and Ingram, 1993).
2.8. Data analysis
For microenvironment, litter mass, taxa diversity
indices, the individual abundances of dominant macro-
invertebrate taxa (those with a minimum mean
abundance of 6 ind m�2 in the soil or litter) and total
macroinvertebrate abundances, ANOVAs were used to
determine significant differences among forest types
(primary, recently logged and secondary) and among
secondary forest stages (15-, 45-, 75- and 100-year-old
forests). The spatial arrangement of some of the study
areas in chronosequences and others in separate
recently logged and pristine sites did not permit a
single analysis of variance that would test the effect of
site and forest age simultaneously for all studied areas.
For this reason, we divided the analyses into the
following steps. First a one-way ANOVA was
performed to determine whether there were differences
among the pristine (n = 2), recently logged (n = 2) and
secondary forests (n = 3). For this analysis we used the
means of the site samples as replicates. The three
chronosequences (Juquila, Tarbis and Tanetze) were
then analysed with a two-way ANOVA to determine the
effect of chronosequence and forest age (four levels,
n = 3). Measurements within a single grid are in this
context pseudoreplicates (Hurlbert, 1984) and were
averaged and considered a single replicate.
All variations of ANOVA, tests of their assumptions
and the Tukey’s honest significant difference post hoc
test (HSD; Legendre and Legendre, 1998) were
performed using the software STATISTICA v.5.0
(Statsoft Inc., Tulsa).
Differences among the Juquila successional stages in
the mass per sample of litter components and the
chemical composition of soils were tested with one-way
multivariate analyses of variance (MANOVA) with age
as a four level factor. The 49 samples extracted from the
intersections of each of the Juquila grids were
considered as single replicates. When the MANOVA
turned out to be significant, corresponding one-way
ANOVAs for each individual variable were then
performed. Wilk’s l and P-values are reported when
appropriate. The differences in the nutrient and lignin
S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355344
content among the litters of different tree species were
also tested with a one-way MANOVA. For this analysis
three replicates per leaf species were analysed. Each
replicate consisted of a 30 g sample of dry leaves
randomly drawn from the pool of leaves. If the
MANOVA was significant, the same procedure as
described above was followed.
MANOVA was only performed when the homo-
scedasticity (Box’s M-test) and multivariate normal
distribution assumptions were met. Multivariate nor-
mality was assumed to be achieved when all dependent
variables were themselves normally distributed (Kol-
mogorov–Smirnov test; Legendre and Legendre,
1998).
3. Results
3.1. Soil microenvironment
Recently logged sites had around 20% lower mean
canopy density than pristine sites and secondary forests,
Table 1
Means, coefficients of variation (in parentheses) and results of ANOVA fo
Among forest types
Forest type means
Primary Recent
Litter mass (g monolith�1) 68.1 (0.24) 67.4 (0
Soil temperature (8C) 11.9 (0.01) a 15.9 (0
Canopy cover (%) 94.7 (0.20) a 76.7 (0
Volume of soil water (cm3 cm�3) 0.2 (0.02) 0.2 (0
Soil taxa richness 10.6 (0.02) 10.1 (0
Soil taxa diversity (H0) 1.8 (0.03) 1.9 (0
Soil macroinvertebrate abundance 85.3 (0.38) 74.8 (0
Litter taxa richness 6.1 (0.15) a 10.1 (0
Litter taxa diversity (H0) 1.9 (0.02) a 1.4 (0
Litter macroinvertebrate abundance 16.8 (0.33) a 43.2 (<
Within secondary succession compa
Means per successional stage
15-Year-old 45-Year-old 7
Litter mass (g monolith�1) 83.3 (0.20) 95.9 (0.10) 1
Soil temperature (8C) 14.4 (0.02) a 13.6 (0.02) b
Canopy cover (%) 94.4 (0.02) 93.7 (0.01)
Volume of soil water (cm3 cm�3) 0.2 (0.36) 0.2 (0.36)
Soil taxa richness 7.0 (0.21) a 7.5 (0.33) ab
Soil taxa diversity (H0) 1.7 (0.17) 1.7 (0.14)
Soil macroinvertebrate abundance 16.6 (0.42) a 28.7 (0.76) ab
Litter taxa richness 6.9 (0.25) 7.8 (0.19)
Litter taxa diversity (H0) 1.7 (0.19) 1.8 (0.12)
Litter macroinvertebrate abundance 15.1 (0.33) 19.9 (0.45)
The first ANOVA compares forest types (pristine, recently logged and second
(15-, 45-, 75- and 100-year-old forests). Different letters denote significant
but there were no differences among secondary forests
of different age (Table 1). Mean soil temperature was
lowest in pristine sites (11.9 8C) and reached the
greatest value in recently logged sites (15.9 8C). Within
secondary chronosequences, 15-year-old forests had a
slightly higher soil temperature than other successional
stages (Table 1). High variability in volumetric soil
water content and no significant differences were found
among forest types or secondary successional stages
(Table 1).
3.2. Litter resource availability and quality
The number of tree genera (DBH > 5 cm) present in
each of the sampling grids of the Juquila chronose-
quence was 20 in the 15-year-old, 22 in the 45-year-old,
20 in the 75-year-old and 23 in the 100-year-old forests.
The lowest total basal area was found in the 100-year-
old forest (42 m2 ha�1) and the 15-year-old forest
(44 m2 ha�1). The greatest value of basal area was
recorded in the 45-year-old forest (84 m2 ha�1).
r various forest type parameters
ANOVA
Forest type effect
ly logged Secondary F(2,4) P
.19) 91.8 (0.08) 3.63 0.126
.02) b 14.4 (<0.01) ab 20.28 0.018
.38) b 93.5 (0.35) a 161.62 <0.001
.17) 0.2 (0.02) 0.77 0.520
.01) 9.3 (0.15) 3.58 0.128
.05) 1.9 (0.06) 0.53 0.625
.42) 37.9 (0.27) 3.1 0.154
.07) b 8.9 (<0.01) b 16.58 0.012
.02) b 1.7 (0.05) a 23.84 0.0060.01) b 27.2 (0.40) b 7.91 0.041
rison ANOVA
Age effect Chronose-
quence effect
5-Year-old 100-Year-old F(3,6) P F(2,6) P
03.0 (0.14) 85.1 (0.19) 1.26 0.370 1.06 0.404
13.7 (0.02) b 13.8 (0.02) b 23.81 0.014 191.36 0.00192.9 (0.02) 93.2 (0.01) 0.14 0.931 0.30 0.755
0.2 (0.31) 0.2 (0.53) 0.26 0.853 12.79 0.00711.1 (0.12) ab 10.8 (0.07) b 5.34 0.039 5.73 0.041
2.0 (0.07) 1.9 (0.03) 0.82 0.529 1.46 0.304
52.6 (0.16) ab 54.9 (0.08) b 11.32 0.007 5.46 0.0458.7 (0.06) 9.1 (0.04) 0.35 0.79 1.00 0.421
1.9 (0.05) 1.9 (0.07) 0.21 0.889 0.72 0.527
23.9 (0.23) 26.6 (0.63) 2.3 0.174 4.9 0.054
ary) and the second compares successional stages of secondary forests
differences by Tukey’s HSD paired comparisons.
S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355 345
The tree genera that dominated the basal area were
different in each successional stage. In early succes-
sional plots, there was a single genus that covered
around 40% of the total basal area recorded for the plot.
In the 15-year-old forest the dominant genus was
Clethra (38.4% of the total basal area) followed by
Pinus (15.7%); while in the 45-year-old forest Quercus
dominated (40.46%) followed again by Pinus (8.7%). In
later successional stages, the basal area was shared
among more genera. In the 75-year-old forest the three
most dominant genera were Quercus (26.04%),
Vaccinium (11.56%) and Temstroemia (11.04%) while
in the 100-year-old forest the three genera that
accounted for most basal area were Quetzalia
(19.05%), Persea (17.5%) and Oreopanax (15.09%).
There were no differences among forest types or
successional stages in terms of the total litter biomass
per monolith (Table 1). However, there were significant
differences among successional stages in the composi-
tion of the litter (Table 2). The 45- and 75-year-old
successional stages had more total unidentifiable
material than the 15- or 100-year-old forest. Leaves
that were recognisable formed the greatest proportion in
the 45-year-old forest (41%) and the lowest in the 100-
year-old forest (27%).
The leaves of the three taxa that were identified in the
litter (Quercus, Pinus and Lauraceae including Persea,
Belishcmiedia and Ocotea) comprised between 55 and
90% of the recognisable leaves and between 18 and
23% of the total litter mass in each successional stage.
The contribution of leaves from the dominant tree
genera to the standing biomass of leaf litter was not
Table 2
Results of one-way MANOVA for litter composition in the Juquila chronos
ANOVA comparing successional stages
Manova
Effect Fo
Wilks’ l 0.2
Rao’s R 22.
Degrees of freedom effect 18
Degrees of freedom error 529
P-value 0.0
Litter components Mean biomass per successional stage (g m�2)
15-Year-old 45-Year-old 75
Unidentifiable 94.2 144.6 18
Lauraceae 0.1 0.2
Pinus spp. 71.7 57.5
Quercus spp. 1.1 73.0 8
other species 60.3 15.2 2
w&r 95.1 124.8 11
w&r: woody and reproductive material.
proportional to their dominance in terms of basal area.
While Pinus only constituted circa 16% of the total
basal area in the 15-year-old forest, pine needles formed
the highest proportion (53.8%) of the recognisable
leaves in this plot. Independently of the basal area
dominance, oak leaves were the most abundant
recognisable leaf litter in the other three successional
stages (49.9–75.6%). Pine trees covered circa 9% of the
basal area in the 45-year-old forest and their needles
were the second-most abundant component in the litter
(39.4%). Pine trees and needles were absent from the
100-year-old forest. The leaves of other genera
constituted only 40.4% of the recognisable leaf litter
in the 100-year-old forest even if their basal area
reached 70% of the total. The presence of Lauraceae
leaves was negligible in the 15, 45 and 75 years
successional stages (0.05–0.7%).
Some of the main leaf components of the litter in
Juquila (Pinus, Quercus, Beilschmedia and Oreopanax)
differed significantly in their nutrient and lignin content
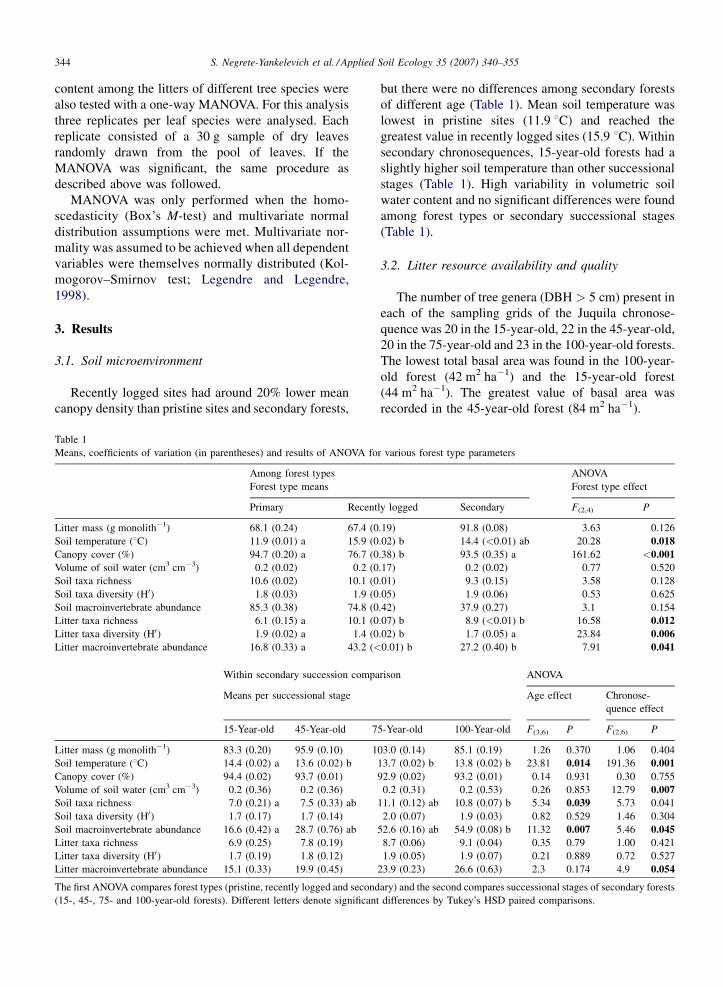
(Fig. 1). For example the mean concentration of
magnesium was lowest in Pinus needles (5.24 cmol
kg�1) and greatest in Oreopanax (35.05 cmol kg�1)
while phosphorus concentration was greatest in
Pinus (2.43 cmol kg�1) and lowest in Quercus
(0.95 cmol kg�1). Lignin content was greatest in
Beilschmedia (44.6%) and lowest in Oreopanax (29.2%).
3.3. Soil nutrients
All soil nutrients analysed differed significantly
among successional stages of the Juquila chronosequence
equence, mean biomass of individual litter components and results of
rest age
05
088
01
ANOVA for litter
components
-Year-old 100-Year-old F(3,192) P-value
9.0 92.4 13.80 <0.0010.9 5.8 40.32 <0.0012.9 0.0 26.01 <0.0017.2 50.8 51.30 <0.0014.4 38.3 15.35 <0.0017.9 122.6 1.58 0.20
S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355346
Fig. 1. Nutrient concentrations in the senescent leaves of four tree
species (n = 3). (a) Total carbon and lignin, (b) total phosphorus and
(c) magnesium and potassium. Wilk’s l < 0.0001 (P < 0.001). Dif-
ferent letters indicate significant differences (P < 0.05) in paired
comparisons between leaf species with Tukey’s HSD test.
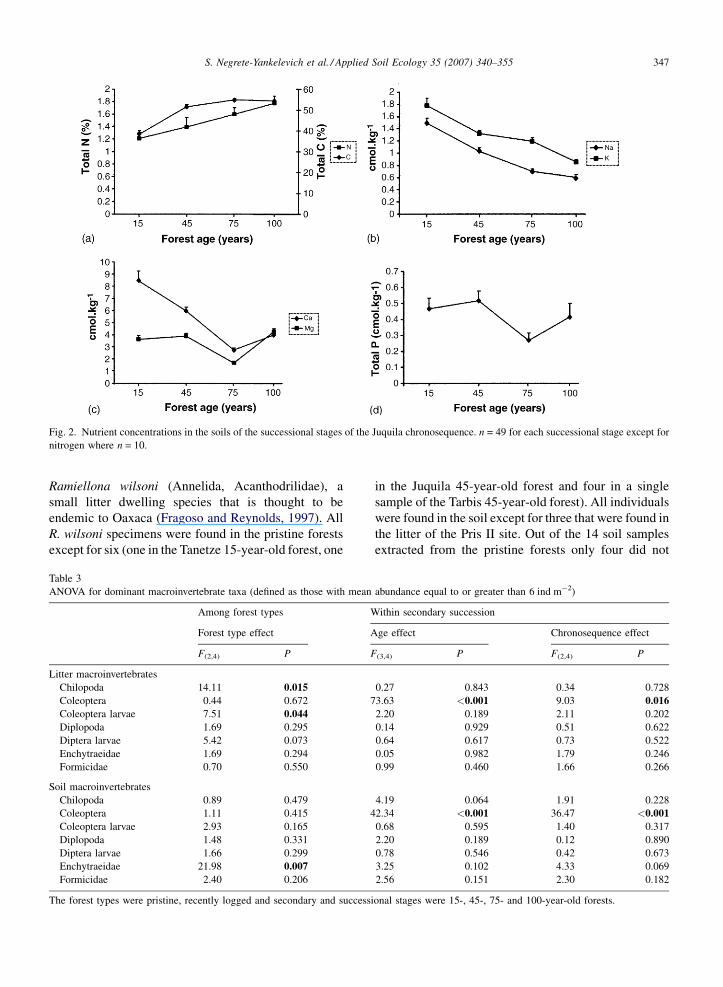
(Wilk’s l = 0.14, P < 0.001). The total concentration of
carbon was lower in the earliest successional stage
(35.4%) compared to the 45-, 75- and 100-year-old
forests (48.7–51.8%; F (3,192) = 44.87, P < 0.001; Fig. 2).
Total soil nitrogen concentration increased along the
successional chronosequence (F (3,44) = 4.08, P = 0.01),
from a mean of 1.2% in the 15-year-old forest to a mean
of 1.8% in the 100-year-old forest. In the case of
phosphorus the 45- and 75-year-old forest had the
highest and lowest values, respectively (0.51 and
0.26 cmol kg�1; F (3,192) = 2.69, P = 0.05). The values
for magnesium were similar (means between 3.55 and
4.30 cmol kg�1) for all forests except for the 75-year-
old which had a significantly lower concentration (ca.
1.7 cmol kg�1; F (3,192) = 22.02, P < 0.001). The con-
centrations of sodium and potassium decreased more or
less steadily along the successional sequence
(F (3,192) = 49.75 and 26.84, respectively, P-values
<0.001). Similarly the concentration of calcium
decreases with succession up to the 75-year-old forest,
however it increased again in the 100-year-old forest
(F (3,192) = �29.09, P < 0.001; Fig. 2).
3.4. Macroinvertebrate community
Most macroinvertebrates collected in the survey
(65.7%) were found in the soil and consequently,
individual taxa often occurred in greater numbers in the
soil than in the litter. The most abundant taxa were
Enchytraeidae (18.1%), Formicidae (16.3%) and
Coleoptera (adults, 12.0%) in the soil and Coleoptera
(adults, 19.7%), Diplopoda (13.3%) and Formicidae
(9.9%) in the litter. Four taxa were relatively rare:
Dermaptera (2 individuals), Thysanura (8 individuals),
Uropygi (10 individuals) arid Blattaria (11 individuals)
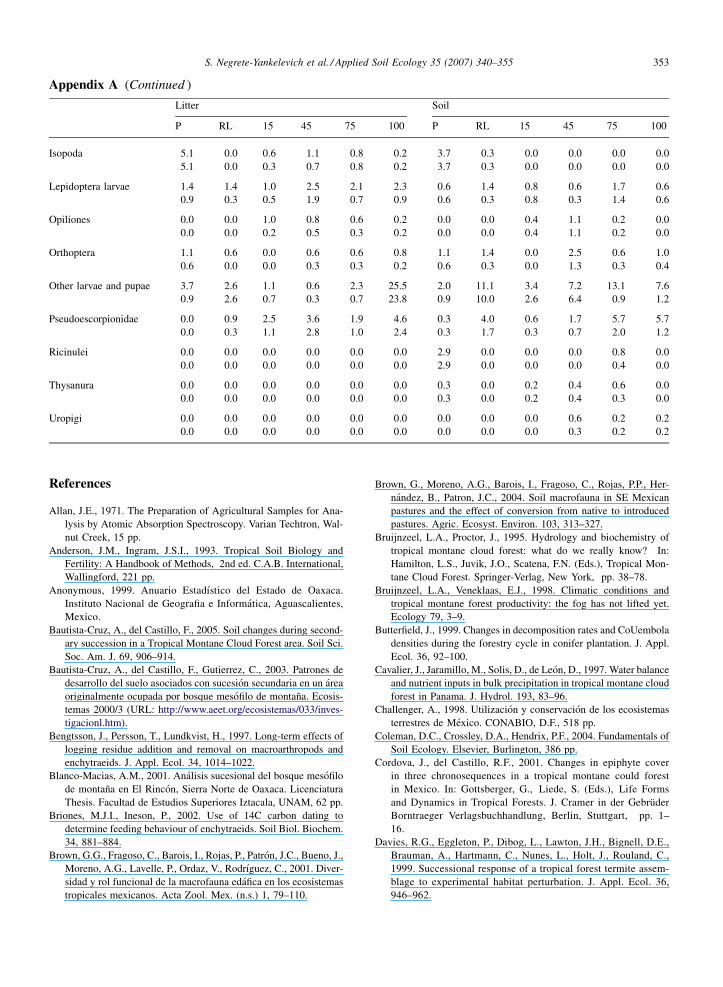
(see Appendix 1 for individual abundances). Seven
macroinvertebrate taxa or immature groups (Chilopoda,
Coleoptera, Coleoptera larvae, Diplopoda, Diptera
larvae, Enchytraeidae and Formicidae) were considered
dominant because they had a minimum total mean
abundance of 6 ind/m2 in the litter or soil. The
abundances of these dominant taxa were analysed
individually (Table 3).
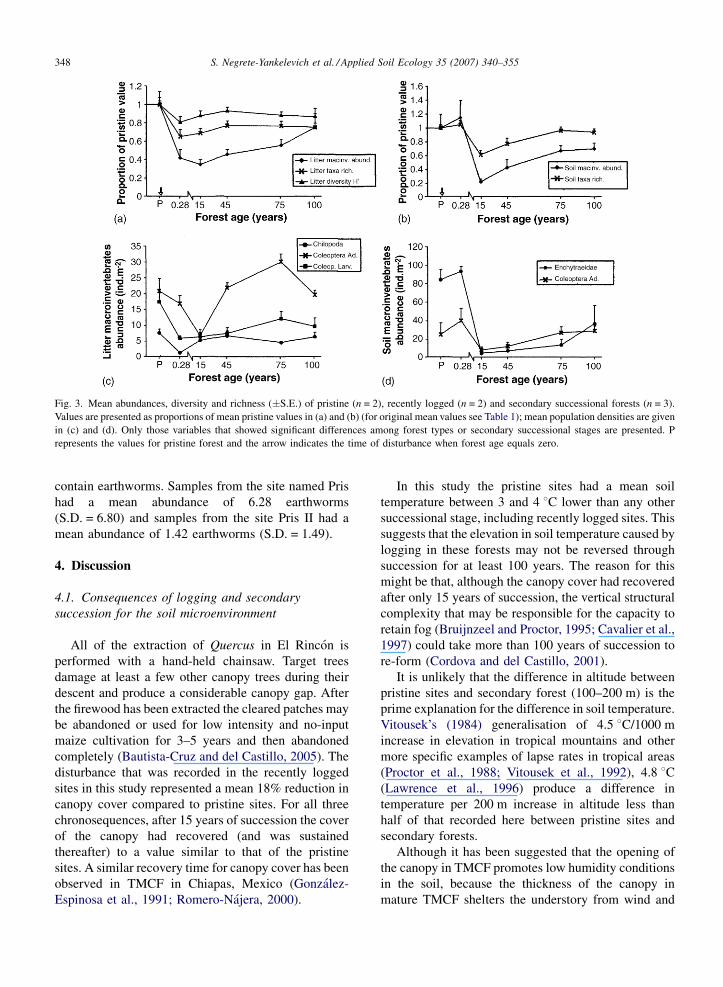
Total macroinvertebrate abundance, taxa richness,
Shannon’s diversity index and the individual mean
abundances of Chilopoda and Coleoptera larvae were
lower in the litter of recently logged sites than in pristine
sites (Fig. 3). In contrast, in the soil community, the
minimum values of all diversity variables and individual
abundances that were sensitive to disturbance were
reached in the 15-year-old forests, where total macro-
invertebrate abundance was ca. 80% lower and taxa
richness ca. 30% lower than in pristine sites. The
abundance of Enchytraeidae was ca. 94% lower and of
Coleoptera ca. 65% lower compared to undisturbed
sites (Fig. 3b and c).
Several diversity and abundance variables in the litter
and soil communities did not recover to the level of
pristine forests after 100 years of succession. This was the
case for richness and total abundance of litter taxa (both
ca. 25% lower in the 100-year-old forests), abundance of
litter Coleoptera larvae (44% lower) and abundance of
soil Enchytraeidae (56% lower). The total abundance and
taxa richness of soil macroinvertebrates increased with
succession but only taxa richness reached the level of
pristine sites in the 100-year-old forests. In contrast,
Shannon’s diversity and the abundance of Chilopoda in
the litter had already recovered close to pristine levels in
the 45-year-old forests (Fig. 3a and c).
A total of 60 earthworms (Order Lumbricina) were
found in the survey. All of them were identified as
S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355 347
Fig. 2. Nutrient concentrations in the soils of the successional stages of the Juquila chronosequence. n = 49 for each successional stage except for
nitrogen where n = 10.
Ramiellona wilsoni (Annelida, Acanthodrilidae), a
small litter dwelling species that is thought to be
endemic to Oaxaca (Fragoso and Reynolds, 1997). All
R. wilsoni specimens were found in the pristine forests
except for six (one in the Tanetze 15-year-old forest, one
Table 3
ANOVA for dominant macroinvertebrate taxa (defined as those with mean
Among forest types W
Forest type effect A
F(2,4) P F
Litter macroinvertebrates
Chilopoda 14.11 0.015Coleoptera 0.44 0.672 7
Coleoptera larvae 7.51 0.044Diplopoda 1.69 0.295
Diptera larvae 5.42 0.073
Enchytraeidae 1.69 0.294
Formicidae 0.70 0.550
Soil macroinvertebrates
Chilopoda 0.89 0.479
Coleoptera 1.11 0.415 4
Coleoptera larvae 2.93 0.165
Diplopoda 1.48 0.331
Diptera larvae 1.66 0.299
Enchytraeidae 21.98 0.007Formicidae 2.40 0.206
The forest types were pristine, recently logged and secondary and successi
in the Juquila 45-year-old forest and four in a single
sample of the Tarbis 45-year-old forest). All individuals
were found in the soil except for three that were found in
the litter of the Pris II site. Out of the 14 soil samples
extracted from the pristine forests only four did not
abundance equal to or greater than 6 ind m�2)
ithin secondary succession
ge effect Chronosequence effect
(3,4) P F(2,4) P
0.27 0.843 0.34 0.728
3.63 <0.001 9.03 0.0162.20 0.189 2.11 0.202
0.14 0.929 0.51 0.622
0.64 0.617 0.73 0.522
0.05 0.982 1.79 0.246
0.99 0.460 1.66 0.266
4.19 0.064 1.91 0.228
2.34 <0.001 36.47 <0.0010.68 0.595 1.40 0.317
2.20 0.189 0.12 0.890
0.78 0.546 0.42 0.673
3.25 0.102 4.33 0.069
2.56 0.151 2.30 0.182
onal stages were 15-, 45-, 75- and 100-year-old forests.
S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355348
Fig. 3. Mean abundances, diversity and richness (�S.E.) of pristine (n = 2), recently logged (n = 2) and secondary successional forests (n = 3).
Values are presented as proportions of mean pristine values in (a) and (b) (for original mean values see Table 1); mean population densities are given
in (c) and (d). Only those variables that showed significant differences among forest types or secondary successional stages are presented. P
represents the values for pristine forest and the arrow indicates the time of disturbance when forest age equals zero.
contain earthworms. Samples from the site named Pris
had a mean abundance of 6.28 earthworms
(S.D. = 6.80) and samples from the site Pris II had a
mean abundance of 1.42 earthworms (S.D. = 1.49).
4. Discussion
4.1. Consequences of logging and secondary
succession for the soil microenvironment
All of the extraction of Quercus in El Rincon is
performed with a hand-held chainsaw. Target trees
damage at least a few other canopy trees during their
descent and produce a considerable canopy gap. After
the firewood has been extracted the cleared patches may
be abandoned or used for low intensity and no-input
maize cultivation for 3–5 years and then abandoned
completely (Bautista-Cruz and del Castillo, 2005). The
disturbance that was recorded in the recently logged
sites in this study represented a mean 18% reduction in
canopy cover compared to pristine sites. For all three
chronosequences, after 15 years of succession the cover
of the canopy had recovered (and was sustained
thereafter) to a value similar to that of the pristine
sites. A similar recovery time for canopy cover has been
observed in TMCF in Chiapas, Mexico (Gonzalez-
Espinosa et al., 1991; Romero-Najera, 2000).
In this study the pristine sites had a mean soil
temperature between 3 and 4 8C lower than any other
successional stage, including recently logged sites. This
suggests that the elevation in soil temperature caused by
logging in these forests may not be reversed through
succession for at least 100 years. The reason for this
might be that, although the canopy cover had recovered
after only 15 years of succession, the vertical structural
complexity that may be responsible for the capacity to
retain fog (Bruijnzeel and Proctor, 1995; Cavalier et al.,
1997) could take more than 100 years of succession to
re-form (Cordova and del Castillo, 2001).
It is unlikely that the difference in altitude between
pristine sites and secondary forest (100–200 m) is the
prime explanation for the difference in soil temperature.
Vitousek’s (1984) generalisation of 4.5 8C/1000 m
increase in elevation in tropical mountains and other
more specific examples of lapse rates in tropical areas
(Proctor et al., 1988; Vitousek et al., 1992), 4.8 8C(Lawrence et al., 1996) produce a difference in
temperature per 200 m increase in altitude less than
half of that recorded here between pristine sites and
secondary forests.
Although it has been suggested that the opening of
the canopy in TMCF promotes low humidity conditions
in the soil, because the thickness of the canopy in
mature TMCF shelters the understory from wind and

S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355 349
solar radiation (Olander et al., 1998), our results did not
show any significant difference between pristine,
recently logged and secondary forests. This could be
a result of the water content in the soil of these forests
being greatly influenced by seasonal changes (Negrete-
Yankelevich, 2004), variability of humus structure
(Bautista-Cruz and del Castillo, 2005) and/or variability
of soil texture (Bautista-Cruz et al., 2003). The complex
structure of the mature TMCF canopy may play a
significant role in buffering extreme changes in soil
temperatures and humidity in the soil (Romero-Najera,
2000), therefore during the dry season it is expected that
significant differences might occur.
4.2. Consequences of logging and secondary
succession for quality and availability of litter
resources
Once TMCF in Mexico is abandoned after a major
disturbance, different plant species become associated
with different successional stages. The vegetation
succession described for our sites in the Juquila
chronosequence generally agrees with the patterns
described formerly for Mexican TMCF (Quintana-
Ascencio and Gonzalez-Espinosa, 1993; Blanco-
Macias, 2001; Galindo-Jaimes et al., 2002). Initially,
shade-intolerant genera of holarctic origin, mainly
Pinus, colonise recently disturbed sites. Under the
incomplete canopy of pine-dominated stands, Quercus
spp. are able to regenerate and are commonly found to
dominate mid-successional canopies. In late-succession
stands, genera of holarctic origin are replaced by
hardwoods of pantropical affinity (such as Persea,
Beilschmedia and Oreopanax), or genera with Andean-
tropical (such as Clethra) and East Asian–North
American (such as Temstroemia) origins. An exception
to this trend was the dominance of the genus Clethra
over Pinus in our 15-year-old plot. In other tropical
montane forests, species of Clethra act also as pioneers
after disturbance (Newton and Healey, 1989; Blanco-
Macias, 2001). Even if the genus Clethra dominated the
basal area of our 15-year-old plot, in terms of
contribution to the mass of standing litter, pine needles
still dominated over other genera in this plot.
If mature TMCF occurs on nutrient-poor soils and
under low solar radiation conditions, it is expected that
the dominant tree species are slow-growing and highly
efficient in nutrient use (Tanner et al., 1998). Often, the
efficient use of resources implies production of nutrient-
poor litter (Vitousek, 1984) that is shed infrequently
(Hobbie, 1992), and therefore promotes further scarcity
of nutrients in the soil. We hypothesised that this positive
feedback cycle may be disrupted initially by the flush of
litter caused by logging and the penetration of solar
radiation to the forest floor, and later by the increase in
dominance of more productive genera in early succes-
sion. We did not find any evidence of difference in the
mass of standing litter on the forest floor between
recently logged sites and pristine forests and among
successional stages. However, some evidence of greater
litter input by dominant trees in younger forests was
found when the different litter components were
compared between successional stages.
The mass of standing crop of leaf litter in all sites and
chronosequences (2.0–4.2 Mg ha�1) was similar to
values found in other TMCFs (Tanner, 1980; Proctor
et al., 1989; McDonald and Healey, 2000). Except for
the unusually low mean K concentration in oak leaves
and of Mg in pine needles, the mean concentrations of
elements in recently fallen leaves were also within the
range of those reported in other TMCF of similar
altitude (Bruijnzeel and Proctor, 1995; McDonald and
Healey, 2000; Vitousek et al., 1992). This indicates that
the forests studied here are as nutrient limited as other
TMCF of high altitude around the world.
Pirns dominance may affect organic matter cycling
in early succession not only through its high litter
productivity but also through the relatively high
phosphorus and low magnesium concentration in its
needles. Although pine litter has been reported to have
large amounts of phenolic compounds and lignin that
slow its decomposition rate (Scholes and Nowicki,
1998), in these forests we found that compared to three
other canopy species, P. chiapensis needles had only
more lignin than O. xalapensis. Higher concentrations
of P in the litter of young secondary TMCFs compared
to mature forests have also been observed in TMCF in
the Blue Mountains (McDonald and Healey, 2000).
4.3. Consequences of succession for soil chemistry
In this study we evaluated only the first 5 cm of soil,
because in forests the topsoil is where most plant–soil
biochemical interactions occur (Gross et al., 1995) and
where most macroinvertebrates are found (Lavelle and
Pashanasi, 1989). In general, and independently of the
successional stage, the mean concentrations of nutrients
in the soils were as low as those reported for other
TMCF of similar altitude (Bruijnzeel and Proctor, 1995;
McDonald and Healey, 2000; Romero-Najera, 2000;
Wilcke et al., 2003). An exception was the particularly
low mean concentration of total P in the soils of all
successional stages (133.8–205.5 mg kg�1), which was
well below the 208–500 mg kg�1 recorded in other

S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355350
studies (Bruijnzeel and Proctor, 1995; McDonald and
Healey, 2000; Romero-Najera, 2000; Wilcke et al.,
2003).
The relatively high concentration of cations found in
the 15-year-old forest in this study, could be related to
the stem extraction of Quercus spp. trees. Soon after
logging, the increase in high quality organic matter in
harvesting residues added to the decrease in nutrient
uptake due to removal of canopy trees and the higher
radiation entering via the canopy opening, are thought
to speed decomposition and cation release (Finer et al.,
2003). Even if evidence was found of nutrients being
more available in the 15-year-old forest, not all of them
were found to be associated with equivalently high
concentrations in the dominant litter.
The increase in total carbon and nitrogen in the topsoil
through succession indicates an accumulation of organic
matter. Because the decomposition rate in Juquila has
been found to be generally low with no differences
among successional stages (Negrete-Yankelevich, 2004),
the accumulation of litter from productive species (such
as oak and pine) in early succession is likely to be the
origin of high concentrations of semi-decomposed
organic matter in late successional stages. This increase
is also reflected in the greater thickness of the O horizon
through succession reported by Bautista-Cruz and del
Castillo (2005) in these forests. Therefore, our results
suggest that, even if there is an initial pulse of nutrients
available in the soil within the first few years after forest
clearance, nutrients become sequestered once more
during succession, when the canopy closes over. Semi-
decomposed organic matter accumulates in the topsoil,
promoting nutrient immobilisation in the soil. In a similar
TMCF in Chiapas, the dominance of pine over oak in the
canopy was found to be negatively correlated with the
content of organic carbon, cation exchange capacity, total
nitrogen content and acidity in the soil (Romero-Najera,
2000; Galindo-Jaimes et al., 2002), all indicators of an
accumulation of organic matter. The availability of
phosphorus may be playing a particularly important role
in nutrient cycling in these forests. P was less limiting in
mid-successional stages than in mature forests. This was
probably a result of logging residues and pine litter in
early succession that might have released some of the P
locked up in the mature forest and made its cycling
temporarily less efficient.
4.4. Macroinvertebrate communities after logging
and through succession
The community composition of the soil and litter
fauna has been recognised as an important indicator of
soil disturbance and health (Ferris et al., 2001; Brown
et al., 2001; Ruf et al., 2003; Coleman et al., 2004). In
the short-term, logging activities can disturb soil faunal
communities by physically altering their habitat and
also by changing microenvironmental conditions due to
increased radiation after canopy opening. In this study
we found that the total abundance, number of taxa and
diversity index of macroinvertebrates, as well as the
individual mean abundances of Chilopoda and Coleop-
tera larvae in the litter (but no faunal parameter in the
soil) seem to be affected by logging in the short-term (2
months).
Few studies have found a detectable difference in the
macroinvertebrate community composition in the short-
term after disturbance (Okwakol, 1994; Zaitsev et al.,
2002). Many studies have found that the short-term
impact on the invertebrate community is imperceptible if
the intensity of extraction is low as in our recently logged
sites (Davies et al., 1999; termites, Siira-Pietikainen
et al., 2001; macroarthropods). Furthermore, some
studies have not found a short-term effect, even after
clear-cutting, on the macroinvertebrate community
(Theenhaus and Schaefer, 1995). This discrepancy
may be explained by differences in the factors limiting
productivity in the system. In forests that are not as
nutrient- or energy-limited as TMCFs, subtle changes in
these factors caused by selective logging may not alter
prevailing conditions to an extent that results in an impact
on the composition of the macroinvertebrate community.
In the topsoil, there seems to be a delayed response
of the macroinvertebrate community to logging
disturbance or successional changes. Macroinvertebrate
taxa richness and total abundance, as well as the
abundances of some dominant taxa in the soil reached
their lowest values in the 15-year-old forests. A similar
delayed response was observed by Zaitsev et al. (2002)
for oribatid mites. Differences in the response of the
litter and soil community to early succession could be
explained by the litter community being driven by
present environmental conditions, while the soil
community could have been experiencing a delayed
response to logging disturbance. After logging, the
substantial input of high quality residues and rise in
temperature may have increased decomposition rate
(Zaitsev et al., 2002) and consumed the accumulated
semi-decomposed organic matter that sustains the soil
macroinvertebrate community in mature TMCF.
Although we did not find a univariate macroinverte-
brate community indicator that could accurately
distinguish pristine, recently logged and secondary
successional stages, litter taxa richness was the most
sensitive variable to distinguish primary from recently

S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355 351
logged or secondary sites. In contrast, the number of
higher taxa and the total macroinvertebrate abundance
in the soil were the best univariate discriminators of
secondary successional stages, but failed to distinguish
primary from recently logged sites.
Little is known about the importance of limited
nutrient availability in cloud forests for soil fauna
(Waide et al., 1998); it is known, however, that nutrient
limitation may be associated with bottom-up control of
food-web complexity (Scheu and Schaffer, 1998; Halaj
and Wise, 2002; Neilson et al., 2002). A relevant result
in this context is the significant decline in the abundance
of litter Chilopoda after logging which only appears to
recover after 45 years of succession. Chilopoda, as a
purely predator group, lie at the top of the food chain
and a decrease in their population may indicate changes
in the bottom-up controls as a result of disturbance.
Compared to other dominant taxa, enchytraeid
densities showed the most dramatic effect of logging
disturbance. Their density dropped to 6% of the pristine
value in recently disturbed soils and after 100 years of
succession they only appear to have recovered to 40% of
the pristine densities. These results are consistent with
the known sensitivity of enchytraeid populations to
disturbance (Hanel, 2001; Uhıa and Briones, 2002),
temperature regimes (Briones and Ineson, 2002;
Lavelle and Spain, 2001) and to the quantity (Bengtsson
et al., 1997) and type of available litter resources
(Schlaghamersky, 2002), all factors that change after
logging and during secondary succession. Even if
enchytraeids are known to exist in particularly low
numbers in the tropics, ranging from a few hundred to a
few thousand individuals per square metre (Lavelle and
Spain, 2001), the densities recorded in this study (14–
93 ind m�2) may still be an underestimation. This
underestimation was probably an artifact of the hand
sorting, which is not an efficient extraction technique
for small or fast macroinivertebrates. For this reason
abundance estimates for taxa like Enchytraeidae or
Chilopoda should be considered very tentative.
Earthworms have been singled out as one of the most
important group of soil animals in terms of their feeding
upon detritus and their effect on soil structure (Lavelle,
2000). In this study both in litter and soil, the only
species of earthworm found (R. wilsoni) was almost
exclusive to the pristine forests. This native litter
dweller, has been found previously in this region
(Fragoso and Reynolds, 1997). The almost complete
absence of R. wilsoni from secondary forests even after
100 years of succession may be the result of low
dispersal and re-colonisation rates (Fragoso et al., 1997)
or be caused by very specific niche or resource
requirements that have not recovered even after this
time interval. The disappearance of earthworms as a
result of forest clearing has also been observed in the
humid tropics (Tian, 1998) where their absence is
thought to have important functional consequences such
as slower decomposition rates in severely disturbed
habitats. Further studies are needed to understand the
effects of human disturbance on R. wilsoni as it appears
to be a sensitive indicator of disturbance in these forests.
In the long-term, the opening of new niches with the
continuous disturbance of native ecosystems often
creates favourable conditions for the invasion of the
more adaptive exotic earthworm species (Brown et al.,
2004). The disappearance of R. wilsoni after logging of
the TMCF of Oaxaca and its absence from secondary
forests may be posing a risk of invasion from the exotic
species that have been introduced in neighbouring
coffee and maize fields (personal observation).
Often studies on the impact of human activities on the
relationship between the above-ground and below-
ground communities are performed simulating distur-
bance in artificial conditions and/or are only able to track
impacts for a relatively short period of time (Wardle et al.,
1997; Hooper and Vitousek, 1997; Mikola et al., 2005).
In this study, we examined a forest system where the
impacts of selective logging of a canopy tree (Quercus
sp.) on the below-ground system would be assessed for a
period of up to 100 years. This was possible because the
study area of El Rincon has a mosaic of forests that have
been logged at different times during the past 100 years,
providing a rare opportunity to perform a replicated study
of chronosequences. All our sites, by being located
within an area of approximately 100 km2, are similar in
terms of climate, soil and vegetation, reducing the risk of
confounding factors that could mask the effect of
logging.
Our results demonstrate that in a TMCF, logging
disturbance has profound short-term and long-lasting
effects on the below-ground system. Selective harvest-
ing of Quercus spp. trees causes an increase in soil
temperature of ca. 4 8C that is not completely reversed
after 100 years of succession. During 100 years of
secondary succession litter diversity increases and soil
organic matter accumulates. The availability of cations
in the topsoil decreases by more than 50% as a result of
logging, and only Mg recovers after 75 years of
succession. The effect of Quercus harvesting on the
litter macroinvertebrate community is apparent within 2
months (total abundance declined by ca. 65% and
diversity by ca. 35%). For the topsoil community there
is a time-lag in the effect of logging. Fifteen years after
abandonment, total macroinvertebrate abundance in the
S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355352
soil was ca. 80% lower and higher taxa richness ca. 30%
lower compared to undisturbed sites. Full recovery of
the macroinvertebrate community composition appears
to take more than 100 years both in the litter and soil.
Acknowledgments
This research was funded by a postgraduate scholar-
ship provided by the Mexican CONACYT (Num. Reg.
Appendix A
Macroinvertebrate densities (ind m�2) in pristine (P), recen
100-year-old forests). Means and standard errors (below) are
averaging the values of all grid intersections. Sample size (n)
for the pristine and recently logged sites
Litter
P RL 15 45 75
Aranea 12.3 4.0 9.7 8.4 8.8
2.6 3.4 3.3 4.8 4.2
Blattaria 0.0 0.0 0.4 0.2 0.0
0.0 0.0 0.4 0.2 0.0
Chilopoda 7.7 1.4 5.3 6.9 4.8
1.4 0.3 1.7 2.6 0.2
Coleoptera 20.9 16.9 6.9 21.9 30.1
4.9 2.6 2.0 1.6 2.5
Coleoptera larvae 17.4 6.0 6.5 7.6 12.2
3.7 0.9 1.3 0.8 2.2
Dermaptera 0.0 0.0 0.0 0.0 0.2
0.0 0.0 0.0 0.0 0.2
Diplopoda 32.9 2.9 8.4 12.0 13.0
23.7 1.1 2.8 4.7 6.3
Diptera 2.9 1.1 0.4 0.6 0.8
1.7 1.1 0.4 0.6 0.4
Diptera larvae 19.4 4.6 5.0 2.5 2.5
8.0 0.6 2.2 1.1 0.5
Diplura 8.3 3.1 0.8 2.3 3.8
3.1 2.6 0.2 1.7 2.3
Enchytraeidae 29.1 7.1 1.3 1.1 1.0
24.0 2.0 1.3 0.9 0.5
Formicidae 0.0 10.9 2.7 2.7 8.2
0.0 4.0 0.8 1.2 4.5
Gastropoda 2.9 0.9 0.2 0.6 0.2
0.6 0.9 0.2 0.3 0.2
Hemiptera 7.4 2.3 6.3 2.3 2.1
2.6 0.9 0.6 0.8 0.2
Hymenoptera 0.3 0.3 0.8 1.3 0.4
0.3 0.3 0.8 0.4 0.4
131536) and by supplementary support of The UK
Darwin Initiative. We are indebted to Graham Russell, Jo
Anderson, David Wardle, Vinicio Sosa and Rafael del
Castillo for invaluable suggestions during the develop-
ment of this research. We are particularly grateful for
field assistance beyond the call of duty of Raul Rivera and
dedicated figure edition by Rafael Ruiz. We also thank
two anonymous reviewers that provided valuable
suggestions to improve the manuscript.
tly logged (RL) and secondary successional stages (15–
presented. The value per replicate plot was calculated
is therefore 3 for the 15–100-year-old forests and n = 2
Soil
100 P RL 15 45 75 100
6.7 6.3 9.1 5.1 4.4 4.2 9.1
0.2 1.7 5.7 2.2 1.7 1.6 2.8
0.4 0.0 0.0 0.2 0.0 0.0 0.0
0.4 0.0 0.0 0.2 0.0 0.0 0.0
6.5 27.4 14.6 6.5 15.6 27.0 19.8
1.4 13.7 0.3 1.9 4.1 2.2 7.8
19.6 25.1 40.3 8.6 12.4 27.2 29.0
1.5 15.4 13.4 3.2 4.4 6.4 5.3
9.7 27.4 24.3 7.8 11.2 14.1 15.6
2.7 9.7 0.3 2.9 5.0 5.7 3.4
0.0 0.0 0.0 0.0 0.0 0.0 0.0
0.0 0.0 0.0 0.0 0.0 0.0 0.0
12.6 50.9 18.6 5.1 10.7 17.5 17.7
6.1 34.3 9.4 1.0 3.6 3.9 4.8
1.0 2.9 2.9 0.0 0.0 0.2 1.0
0.2 1.7 1.7 0.0 0.0 0.2 0.4
2.7 34.9 17.1 1.9 9.1 5.9 17.9
1.5 21.7 2.9 0.4 4.0 1.0 13.6
0.6 18.9 10.9 4.6 7.8 10.3 16.4
0.0 9.1 2.3 1.5 1.0 2.9 7.2
1.0 84.3 93.4 5.3 7.6 13.9 37.0
0.4 13.4 5.4 3.4 6.2 6.1 19.6
31.2 6.0 82.9 13.5 20.2 49.1 22.1
29.2 0.3 50.3 4.6 17.9 12.3 4.3
0.2 0.3 0.3 0.0 0.0 0.2 0.0
0.2 0.3 0.3 0.0 0.0 0.2 0.0
2.3 2.9 7.7 2.3 0.8 0.8 1.1
1.6 0.6 2.9 1.2 0.5 0.5 0.8
1.5 1.1 0.9 0.2 0.8 1.5 1.0
0.2 0.0 0.9 0.2 0.2 0.8 0.2
S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355 353
Appendix A (Continued )
Litter Soil
P RL 15 45 75 100 P RL 15 45 75 100
Isopoda 5.1 0.0 0.6 1.1 0.8 0.2 3.7 0.3 0.0 0.0 0.0 0.0
5.1 0.0 0.3 0.7 0.8 0.2 3.7 0.3 0.0 0.0 0.0 0.0
Lepidoptera larvae 1.4 1.4 1.0 2.5 2.1 2.3 0.6 1.4 0.8 0.6 1.7 0.6
0.9 0.3 0.5 1.9 0.7 0.9 0.6 0.3 0.8 0.3 1.4 0.6
Opiliones 0.0 0.0 1.0 0.8 0.6 0.2 0.0 0.0 0.4 1.1 0.2 0.0
0.0 0.0 0.2 0.5 0.3 0.2 0.0 0.0 0.4 1.1 0.2 0.0
Orthoptera 1.1 0.6 0.0 0.6 0.6 0.8 1.1 1.4 0.0 2.5 0.6 1.0
0.6 0.0 0.0 0.3 0.3 0.2 0.6 0.3 0.0 1.3 0.3 0.4
Other larvae and pupae 3.7 2.6 1.1 0.6 2.3 25.5 2.0 11.1 3.4 7.2 13.1 7.6
0.9 2.6 0.7 0.3 0.7 23.8 0.9 10.0 2.6 6.4 0.9 1.2
Pseudoescorpionidae 0.0 0.9 2.5 3.6 1.9 4.6 0.3 4.0 0.6 1.7 5.7 5.7
0.0 0.3 1.1 2.8 1.0 2.4 0.3 1.7 0.3 0.7 2.0 1.2
Ricinulei 0.0 0.0 0.0 0.0 0.0 0.0 2.9 0.0 0.0 0.0 0.8 0.0
0.0 0.0 0.0 0.0 0.0 0.0 2.9 0.0 0.0 0.0 0.4 0.0
Thysanura 0.0 0.0 0.0 0.0 0.0 0.0 0.3 0.0 0.2 0.4 0.6 0.0
0.0 0.0 0.0 0.0 0.0 0.0 0.3 0.0 0.2 0.4 0.3 0.0
Uropigi 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.6 0.2 0.2
0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.3 0.2 0.2
References
Allan, J.E., 1971. The Preparation of Agricultural Samples for Ana-
lysis by Atomic Absorption Spectroscopy. Varian Techtron, Wal-
nut Creek, 15 pp.
Anderson, J.M., Ingram, J.S.I., 1993. Tropical Soil Biology and
Fertility: A Handbook of Methods, 2nd ed. C.A.B. International,
Wallingford, 221 pp.
Anonymous, 1999. Anuario Estadıstico del Estado de Oaxaca.
Instituto Nacional de Geografia e Informatica, Aguascalientes,
Mexico.
Bautista-Cruz, A., del Castillo, F., 2005. Soil changes during second-
ary succession in a Tropical Montane Cloud Forest area. Soil Sci.
Soc. Am. J. 69, 906–914.
Bautista-Cruz, A., del Castillo, F., Gutierrez, C., 2003. Patrones de
desarrollo del suelo asociados con sucesion secundaria en un area
originalmente ocupada por bosque mesofilo de montana. Ecosis-
temas 2000/3 (URL: http://www.aeet.org/ecosistemas/033/inves-
tigacionl.htm).
Bengtsson, J., Persson, T., Lundkvist, H., 1997. Long-term effects of
logging residue addition and removal on macroarthropods and
enchytraeids. J. Appl. Ecol. 34, 1014–1022.
Blanco-Macias, A.M., 2001. Analisis sucesional del bosque mesofilo
de montana en El Rincon, Sierra Norte de Oaxaca. Licenciatura
Thesis. Facultad de Estudios Superiores Iztacala, UNAM, 62 pp.
Briones, M.J.I., Ineson, P., 2002. Use of 14C carbon dating to
determine feeding behaviour of enchytraeids. Soil Biol. Biochem.
34, 881–884.
Brown, G.G., Fragoso, C., Barois, I., Rojas, P., Patron, J.C., Bueno, J.,
Moreno, A.G., Lavelle, P., Ordaz, V., Rodrıguez, C., 2001. Diver-
sidad y rol funcional de la macrofauna edafica en los ecosistemas
tropicales mexicanos. Acta Zool. Mex. (n.s.) 1, 79–110.
Brown, G., Moreno, A.G., Barois, I., Fragoso, C., Rojas, P.P., Her-
nandez, B., Patron, J.C., 2004. Soil macrofauna in SE Mexican
pastures and the effect of conversion from native to introduced
pastures. Agric. Ecosyst. Environ. 103, 313–327.
Bruijnzeel, L.A., Proctor, J., 1995. Hydrology and biochemistry of
tropical montane cloud forest: what do we really know? In:
Hamilton, L.S., Juvik, J.O., Scatena, F.N. (Eds.), Tropical Mon-
tane Cloud Forest. Springer-Verlag, New York, pp. 38–78.
Bruijnzeel, L.A., Veneklaas, E.J., 1998. Climatic conditions and
tropical montane forest productivity: the fog has not lifted yet.
Ecology 79, 3–9.
Butterfield, J., 1999. Changes in decomposition rates and CoUembola
densities during the forestry cycle in conifer plantation. J. Appl.
Ecol. 36, 92–100.
Cavalier, J., Jaramillo, M., Solis, D., de Leon, D., 1997. Water balance
and nutrient inputs in bulk precipitation in tropical montane cloud
forest in Panama. J. Hydrol. 193, 83–96.
Challenger, A., 1998. Utilizacion y conservacion de los ecosistemas
terrestres de Mexico. CONABIO, D.F., 518 pp.
Coleman, D.C., Crossley, D.A., Hendrix, P.F., 2004. Fundamentals of
Soil Ecology. Elsevier, Burlington, 386 pp.
Cordova, J., del Castillo, R.F., 2001. Changes in epiphyte cover
in three chronosequences in a tropical montane could forest
in Mexico. In: Gottsberger, G., Liede, S. (Eds.), Life Forms
and Dynamics in Tropical Forests. J. Cramer in der Gebruder
Borntraeger Verlagsbuchhandlung, Berlin, Stuttgart, pp. 1–
16.
Davies, R.G., Eggleton, P., Dibog, L., Lawton, J.H., Bignell, D.E.,
Brauman, A., Hartmann, C., Nunes, L., Holt, J., Rouland, C.,
1999. Successional response of a tropical forest termite assem-
blage to experimental habitat perturbation. J. Appl. Ecol. 36,
946–962.

S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355354
del Castillo, R.F., 1996. Aspectos autoecologicos de Pinus chiapensis.
In: Garduno, L.L., Chavarria, G.V., Magdaleno, P.L., Perez, I.M.
(Eds.), Memorias del 2do. Coloquio Regional de Investigacion,
Ciencias Exactas y Naturales. Universidad Autonoma del Estado
de Mexico, Toluca, pp. 63–68.
Devices-T Devices Ltd., 1999. Theta Probe Soil Moisture Sensor (type
ML2x) User Manual. Delta-T Devices Ltd., Cambridge, 22 pp.
Ferris, H., Bongers, T., de Goede, R.G.M., 2001. A framework for soil
foodweb diagnostics: extension of the nematode faunal analysis
concept. Appl. Soil Ecol. 18, 13–29.
Finer, L., Mannerkoski, H., Piirainen, S., Starr, M., 2003. Carbon and
nitrogen pools in an old growth Norway spruce mixed forest in
eastern Finland and changes associated with clear-cutting. For.
Ecol. Manage. 174, 51–63.
Fragoso, C., Brown, G.G., Patron, J.C., Blanchart, E., Lavelle, P.,
Pashanasi, B., Senapati, B., Kumar, T., 1997. Agricultural inten-
sification, soil biodiversity and agroecosystem function in the
tropics: the role of earthworms. Appl. Soil Ecol. 6, 17–35.
Fragoso, C., Reynolds, J.W., 1997. On some earthworms from central
and southern Mexican mountains, including two new species of
the genus Dichogaster (Dichogastrini). Megadrilogica 7, 9–19.
Galindo-Jaimes, L., Gonzalez-Espinosa, M., Quintana-Ascencio, P.,
Garcıa-Barrios, L., 2002. Tree composition and structure in dis-
turbed stands with varying dominance by Pinus spp. in the high-
lands of Chiapas, Mexico. Plant Ecol. 162, 259–272.
Gonzalez-Espinosa, M., Quintana-Ascencio, P., Ramırez-Marcial, N.,
Gaytan-Guzman, P., 1991. Secondary succession in disturbed
Pinus–Quercus forests in the highlands of Chiapas, Mexico. J.
Veg. Sci. 2, 351–360.
Gross, K.L., Pregitzer, K.S., Burton, J.A., 1995. Spatial variation in
nitrogen availability in three successional plant communities. J.
Ecol. 83, 357–367.
Halaj, J., Wise, D.H., 2002. Impact of a detrital subsidy on the trophic
cascade in a terrestrial grazing food web. Ecology 83, 3141–3151.
Hanel, L., 2001. Succession of soil nematodes in pine forests on coal-
mining sands near Cottbus, Germany. Appl. Soil Ecol. 23–34.
Hobbie, S.E., 1992. Effects of plant species on nutrient cycling.
Trends Ecol. Evol. 7, 336–339.
Hooper, D.U., Vitousek, P.M., 1997. The effect of plant composition
and diversity on ecosystem processes. Science 277, 1302–1305.
Hurlbert, S.H., 1984. Pseudoreplication and the design of ecological
field experiments. Ecol. Monog. 54, 187–211.
Jackson, M.L., 1958. Soil Chemical Analysis. Prentice-Hall, Engle-
wood Cliffs, 498 pp.
Lavelle, P., 2000. Ecological challenges for soil science. J. Soil Sci.
165, 73–86.
Lavelle, P., Pashanasi, B., 1989. Soil macrofauna and land manage-
ment in Peruvian Amazonia (Yurimaguas, Loreto). Pedobiologia
33, 283–291.
Lavelle, P., Spain, A., 2001. Soil Ecology. Kluwer Academic Publish-
ers, Dordrecht, 654 pp.
Lawrence, R., Walker, L.R., Zimmerman, J.K., Lodge, D.J., Guzman-
Grajales, S., 1996. An altitudinal comparison of growth and
species composition in hurricane-damaged forests in Puerto Rico.
J. Ecol. 84, 877–889.
Legendre, P., Legendre, L., 1998. Numerical Ecology, 2nd ed. Else-
vier, New York, 853 pp.
Magurran, A.E., 1996. Ecological Diversity and its Measurements.
University Press, Cambridge, 179 pp.
McDonald, M.A., Healey, J.R., 2000. Nutrient cycling in secondary
forests in the Blue Mountains Jamaica. For. Ecol. Manage. 139,
257–278.
Mikola, J., Nieminen, M., Ilmarinen, K., Vestberg, M., 2005. Below-
ground responses by AM fungi and animal trophic groups to
repeated defoliation in an experimental grassland community. Soil
Biol. Biochem. 37, 1630–1639.
Negrete-Yankelevich, S., 2004. Integrating soil macroinvertebrate
diversity, litter decomposition and secondary succession in a
tropical montane cloud forest in Mexico. Ph.D. Thesis. The
University of Edinburgh, UK, 345 pp. http://www.era.lib.ed.a-
c.uk/handle/1842/592.
Neilson, R., Robinson, D., Marriott, C.A., Scrimgeour, C.M., Hamil-
ton, D., Wishart, J., Boag, B., Handley, L.L., 2002. Above-ground
grazing affects floristic composition and modifies soil trophic
interactions. Soil Biol. Biochem. 34, 1507–1512.
Newton, A.C., Healey, J.R., 1989. Establishment of Clethra occiden-
talis on stems of the tree-fern Cyathea pubescens in a Jamaican
montane rain forest. J. Trop. Ecol. 5, 441–445.
Nilsson, L.O., Huttl, R.F., Johansson, U.T., 1995. Nutrient Uptake and
Cycling in Forest Ecosystems. Kluwer Academic Publishers,
Dordrecht, 696 pp.
Okwakol, M.J.N., 1994. The effect of change in land use on soil
macrofauna communities in Mabira Forest, Uganda. Afr. J. Ecol.
32, 273–282.
Olander, L.P., Scatena, F.N., Silver, L., 1998. Impacts of disturbance
initiated by road construction in a subtropical cloud forest in the
Luquillo Experimental Forest, Puerto Rico. For. Ecol. Manage.
109, 33–49.
Olsson, B.A., Bengtsson, J., Lundkvist, H., 1996a. Effect of different
forest harvest intensities on the pools of exchangeable cations in
coniferous forest soils. For. Ecol. Manage. 84, 135–147.
Olsson, B.A., Staaf, H., Lundkvist, H., Bengtsson, J., Rosen, K.,
1996b. Carbon and nitrogen in coniferous forests soils after clear-
felling and harvests of different intensity. For. Ecol. Manage. 82,
19–32.
Pietikainen, J., Haimi, J., Siitonen, J., 2003. Short-term responses of
soil macroarthropod community to clear felling and alternative
forest regeneration methods. For. Ecol. Manage. 172, 339–353.
Proctor, J., Lee, Y.F., Langley, A.M., Munro, W.R.C., Nelson, T.,
1988. Ecological studies on Gunung Silam, a small ultrabasic
mountain in Sabah, Malaysia. I. Environment, forest structure and
floristics. J. Ecol. 76, 320–340.
Proctor, J., Phillipps, C., Duff, G.K., Heaney, A., Robertson, F.M.,
1989. Ecological studies on Gunung Silam, a small ultrabasic
mountain in Sabah, Malaysia. II. Some forest processes. J. Ecol.
77, 317–331.
Quintana-Ascencio, P., Gonzalez-Espinosa, M., 1993. Afinidad fito-
geografica y papel sucesional de la flora lenosa de los bosques de
pino-encino de los altos de chiapas, Mexcio. Act. Bot. Mex. 21,
43–57.
Ramırez-Marcial, N., Gonzalez-Espinosa, M., Williams-Linera, G.,
2001. Anthropogenic disturbance and tree diversity in Montane
Rain Forest in Chiapas, Mexico. For. Ecol. Manage. 154,
311–326.
Rombke, J., 2003. The role of Gilberto Rhighi in the development of
tropical Microdrile taxonomy. Pedobiologia 47, 405–412.
Romero-Najera, I., 2000. Estructura y condiciones microambien-
tales en bosques perturbados de los altos de Chiapas, Mexico.
Undergraduate Thesis. Universidad Autonoma de Mexico, D.F.,
72 pp.
Ruf, A., Beck, L., Dreher, P., Hund-Rinke, K., Rombke, I., Spelda, J.,
2003. A biological classification concept for the assessment of soil
quality: ‘‘biological soil classification scheme’’ (BBSK). Agric.
Ecos. Environ. 98, 263–271.

S. Negrete-Yankelevich et al. / Applied Soil Ecology 35 (2007) 340–355 355
Rzedowski, J., 1996. Analisis preliminar de la flora vascular de los
bosques mesofilos de montana de Mexico. Act. Bot. Mex. 35,
25–44.
Scheu, S., Schaffer, M., 1998. Bottom-up control of the soil macro-
fauna community in a beechwood limestone: manipulation of a
food resource. Ecology 79, 1573–1585.
Schlaghamersky, J., 2002. The Enchytraeidae of spruce forest plots of
different exposure and acid deposition in a German mountain
range. Eur. J. Soil Biol. 38, 305–309.
Scholes, M.C., Nowicki, T.E., 1998. Effects of pines on soil properties
and processes. In: Richardson, D.M. (Ed.), Ecology and Biogeo-
graphy of Pinus. Cambridge University Press, Cambridge, pp.
341–353.
Siira-Pietikainen, A., Pietikainen, J., Fritze, H., Haimi, J., 2001. Short-
term responses of soil decomposer communities to forest manage-
ment: clear felling versus alternative forest harvesting. Can. J. For.
Res. 31, 88–99.
Switzer, G.L., Shelton, M.G., 1979. Successional development of the
forest floor and soil surface on upland sites of the east gulf coastal
plain. Ecology 60, 1162–1171.
Tanner, E.V.J., 1980. Litterfall in montane rain forest of Jamaica and
its relation to climate. J. Ecol. 68, 833–848.
Tanner, E.V.J., Vitousek, P.M., Cuevas, E., 1998. Experimental inves-
tigation of nutrient limitation of forest growth on wet tropical
mountains. Ecology 79, 10–22.
Theenhaus, A., Schaefer, M., 1995. The effect of clear-cutting and
liming on the soil macrofauna of a beech forest. For. Ecol.
Manage. 77, 35–51.
Tian, G., 1998. Effect of soil degradation on leaf decomposition and
nutrient release under humid tropical conditions. Soil Sci. 163,
897–906.
Uhıa, E., Briones, M.J.I., 2002. Population dynamics and vertical
distribution of enchytraeids and tardigrades in response to defor-
estation. Acta Oecol. 23, 349–359.
Van Soest, P.J., 1994. Fiber and physicochemical properties of feeds.
In: Van Soest, P.J. (Ed.), Nutritional Ecology of the Rumiant.
Cornell University Press, Ithaca, pp. 140–160.
Velazquez, A., Duran, E., Ramırez, I., Mas, J.-F., Bocco, G., Ramırez,
G., Palacio, J.-L., 2003. Land use-cover change processes in
highly biodiverse areas: the case of Oaxaca, Mexico. Glob.
Environ. Change 13, 175–184.
Vitousek, P.M., 1984. Litterfall, nutrient cycling, and nutrient limita-
tion in tropical forests. Ecology 65, 285–298.
Vitousek, P.M., Aplet, G., Turner, D.P., Lockwood, J.J., 1992. The
Mauna Loa environmental matrix: foliar and soil nutrients. Oeco-
logia 89, 372–382.
Waide, R.B., Zimmerman, J.K., Scatena, F.N., 1998. Controls of
primary productivity: lessons from the Luquillo Mountains in
Puerto Rico. Ecology 79, 31–37.
Wardle, D.A., 1992. A compartative assessment of factors which
influence microbial biomass carbon and nitrogen levels in the
soil. Biol. Rev. 67, 321–358.
Wardle, D.A., Bonner, K.I., Nicholson, K.S., 1997. Biodiversity and
plant litter: experimental evidence which does not support the view
that enhanced species richness improves ecosystem function.
Oikos 79, 247–258.
Wardle, D.A., Verhoef, H.A., Clarholm, M., 1998. Trophic relation-
ships in the soil microfood-web: predicting the responses to a
changing global environment. Glob. Change Biol. 4, 713–727.
Wilcke, W., Valladarez, H., Stoyan, R., Yasin, S., Valarezo, C., Zech,
W., 2003. Soil properties in a chronosequence of landslides in
montane rain forest, Ecuador. Catena 53, 79–95.
Williams-Linera, G., Toledo, J., 1996. Litterfall, temperate and tro-
pical dominant trees, and climate in a Mexican Lower Montane
Forest. Biotropica 28, 649–656.
Zaitsev, A.S., Chauvat, M., Pflug, A., Wolters, V., 2002. Oribatid mite
diversity and community dynamics in a spruce chronosequence.
Soil Biol. Biochem. 35, 1919–1927.
Top Related
Copyright © 2022 FDOKUMEN

