







Bahasa
Halaman
Hukum


Periphyton and phytoplankton associated with the tropical carnivorous
plant Utricularia foliosa
Jhon Dıaz-Olarte a, Vanessa Valoyes-Valois b, Castor Guisande c,*, Nestor Ned Torres d,Adriana Gonzalez-Bermudez e, Lizandro Sanabria-Aranda a, Ana M. Manjarres Hernandez f,
Santiago R. Duque d, Lili J. Marciales d, Marcela Nunez-Avellaneda g
a Facultad de Ciencias, Escuela de Biologıa, Universidad Industrial de Santander, 678 Bucaramanga, Colombiab Facultad de Ciencias Basicas, Universidad Tecnologica del Choco Diego Luis Cordoba, Quibdo, Colombia
c Facultad de Ciencias, Universidad de Vigo, Campus Lagoas-Marcosende, 36200 Vigo, Spaind Instituto Amazonico de Investigaciones-IMANI, Universidad Nacional de Colombia, km 2 vıa Tarapaca, Leticia, Colombia
e Facultad de Ciencias y Educacion, Universidad Distrital Francisco Jose de Caldas, 8668 Bogota, Colombiaf Grupo de Investigaciones en Cuencas y Humedales Tropicales, Universidad del Magdalena, 10004 Santa Marta, Colombia
g Instituto Amazonico de Investigaciones Cientıficas Sinchi, Leticia, Colombia
Received 20 February 2007; received in revised form 27 June 2007; accepted 29 June 2007
Available online 4 July 2007
Abstract
The abundance and taxonomic composition of periphyton attached to the bladders and phytoplankton associated with the aquatic carnivorous
plant Utricularia foliosa were quantified, to determine whether periphyton associated with U. foliosa would enhances predation success. Bladder
size, periphyton abundance and periphyton richness together explained 76% of the variation observed in the number of prey captured by the
bladders. The abundance and richness of periphyton followed the same pattern as phytoplankton, i.e., both increased as dissolved inorganic
phosphate concentration rose. This nutrient concentration explained 84 and 74% of the variation observed in richness and abundance of periphyton,
respectively. This suggests that abundance and richness of the periphyton associated with U. foliosa depend mostly on environmental conditions rather
than on facilitation mechanisms displayed by the plant. In conclusion, if periphyton affects U. folisosa negatively due to the competition for light or
nutrients, the plant is ‘‘fated to get along with the enemy’’ but apparently without the capacity to manipulate this ‘‘enemy’’ to its own advantage.
# 2007 Elsevier B.V. All rights reserved.
Keywords: Phytoplankton; Periphyton; Bladderwort; Aquatic carnivorous plant; Zooplankton; Nutrient
1. Introduction
The selective capture by Utricularia of prey from a wide
array of aquatic invertebrates is selective, since the percentage
of species captured by bladders is different from that in the
habitat (Harms, 1999; Jobson and Morris, 2001; Guisande et al.,
2000, 2004). Although this may be due to either the behavior of
some prey that swim close to the bladders looking for the food
attached to them or to the plant’s prey-attracting mechanisms,
the evolution of the latter is implied by the relative contribution
of prey nitrogen to total nitrogen content estimated at 51.8% in
Utricularia vulgaris (Friday and Quarmby, 1994).
* Corresponding author. Tel.: +34 986 812586; fax: +34 986 812556.
E-mail address: [email protected] (C. Guisande).
0304-3770/$ – see front matter # 2007 Elsevier B.V. All rights reserved.
doi:10.1016/j.aquabot.2007.06.010
However, the only strategy observed so far for luring prey is
the presence of antennae and bristles that probably lead
potential substratum-dwelling prey towards the valve used by
Utricularia spp. for trapping prey (Meyers and Strickler, 1979;
Sanabria-Aranda et al., 2006). Although it is suspected that
Utricularia spp. additionally employ one or more chemo-
attraction strategies to lure prey, so far none have been
satisfactorily demonstrated (Guisande et al., 2000, 2004;
Manjarres-Hernandez et al., 2006). Early hypotheses postulat-
ing the attraction of prey via the production of carbohydrates on
the valve (Cohn, 1875; Meyers, 1982; Luetzelburg, 1910) have
been rebutted by later studies (Meyers and Strickler, 1979;
Sanabria-Aranda et al., 2006).
An alternative chemical mechanism evolved to lure prey
may be to enhance periphyton quality or quantity, which might
attract grazing zooplankton and macroinvertebrates close to the

trap door. Since the composition of algal periphyton can vary
among different aquatic plant hosts (Blindow, 1987), the host
might manipulate the periphyton community to its own
advantage (Jones et al., 1999). Using surface texture or
chemical control, such as allelopathic substances influencing
algal growth (Gross et al., 2007), the host might enhance
periphyton abundance or favor highly edible and nutritious
periphyton species. Evidence that these periphyton biomass
enhancing mechanisms might be operative in Utricularia
include a model showing that the plant harnesses the production
of its own periphyton via intermediary grazers (Ulanowicz,
1995) and the greater abundance and diversity of periphyton
found on Utricularia relative to sympatric macrophytes
(Bosserman, 1983).
Information about the periphyton and phytoplankton
associated with aquatic carnivorous plants is scarce, although
in Utricularia foliosa desmids have been described as the most
abundant group, followed by Chlorococcales and Cyanophyta
(Lacoste de Diaz, 1981; Wagner and Mshigeni, 1986).
However, there are no studies on spatial or temporal changes
in abundance or composition of the periphyton associated with
bladders according to changes in the water quality, nor are there
studies to determine whether prey captured or prey abundance
around the plant is related to periphyton abundance and
composition associated with the bladders. In this study, we
examined these aspects with respect to U. foliosa located in
Yahuarcaca creek and lake (Colombian Amazon), with the aim
of determining whether the plants manipulate periphyton
community abundance or composition to their own advantage.
2. Materials and methods
Sampling was carried out five times from March 21 to
May 30 2005 from seven plants of U. foliosa growing in
seven different places along the Yahuarcaca creek and lake
in the Colombian Amazon, from closest to furthest from the
river: site 1 (4810012.200S, 69857038.200W), site 2 (489058.700S,
69857048.300W), site 3 (489036.800S, 6985800.0500W), site 4
(489029.100S, 6985800.0500W), site 5 (489016.200S, 69858010.700W),
site 6 (489018.900S, 6985800.0900W) and site 7 (48904.1.00S,
6985703800W). The sampling was carried out during the growing
season of the plant. This species occurs in the field (mainly from
March to May), just when the wet season begins and water from
the Amazon starts to enter the lakes and creeks.
Each sampling day we randomly collected three leaves of
each plant. Because each plant covers between 4 and 18 m2 in
surface and grows 1–4 m deep, the sampling was not detrimental
to the plant. The age of the leaf was not taken into account when
collecting the bladders. From each leaf, 50 bladders were also
randomly collected to determine the size distribution of bladders
with and without prey and the kind and number of prey captured
by the plant. Bladder size was quantified as the greatest length
from stalk end to the trap-door end.
Each sampling day, two quantitative samples of phyto-
plankton were taken, one at 0.5 m inside and the other 0.5 m
outside of each plant with a 2.5 L Van Dorn bottle. The samples
were preserved with Lugol’s solution. The abundance of each
species was determined by Utermohl’s method using 50 or
100 mL depending on the plankton abundance. Census was
halted when 100 units (cells, colonies or filaments) of the most
abundant species had been counted.
Periphyton associated with the bladders of each leaf was
analysed by five randomly collected bladders from different
size ranges: 800–1000, 1000–1200, 1200–1400, 1400–1600,
1600–1800 and 1800–2000 mm. The periphyton associated
with each set of five bladders were removed by scraping, and
later the bladders were gently broken using two needles and
preserved in an eppendorf vial with 1.5 mL of Lugol solution.
The periphyton was removed from surfaces of the bladders by
vigorous manual shaking in 1.5 mL Lugol solution during
3 min, which was confirmed by visual inspection of the
bladders. The abundance of each species was determined by
counting the entire 1.5 mL volume of Lugol under direct
microscope. In order to identify diatom species, samples were
treated to eliminate organic matter following the method
described by Hasle and Frixell (1970), using hot H2O2 before
mounting for observation. Observations were carried out with
an Olympus CH-2 microscope (400�).
Quantitative zooplankton samples were collected inside the
plant stand at the same places after collecting the phytoplankton
and periphyton samples. Samples were taken from 0.5 m depth
with a 20 L Schindler–Patalas plankton trap. The plant was
shaken strongly in the water before taking the zooplankton
sample inside the plant to collect both free-swimming and epi-
phytic zooplankton. The 20 L sample was concentrated through a
20 mm mesh net and preserved in 100 mL 4% formaldehyde
solution. Estimates of the abundance of zooplankton were made
by counting the quantitative samples using an inverted
microscope at 100� magnification. For each sample, at least
25% of the volume of the quantitative sample was examined.
At a depth of 0.5 m the oxygen concentration, pH,
conductivity, temperature, ammonia (NH4+), nitrate (NO3
�),
nitrite (NO2�), silicate (SiO4
�) and phosphate (PO43�) were
measured inside the plant and outside the plant for the seven
plants. Filtered water (0.45 mm) was used for measuring
nutrients with BRAN + LUEBBE AAIII autoanalyser.
The software used for the statistical analyses was SPSS (SPSS
Inc. 2003) and STATISTICA (Statsoft, 2004). Autocorrelation
among residuals and co-linearity among independent variables
was tested in the regression analyses (Guisande et al., 2006).
3. Results
3.1. Concentration of nutrients
The concentrations of both NO3� and PO4
3� were low in all
locations (Manjarres-Hernandez et al., 2006). The concentra-
tion of NO3�was lower in areas closer to the Amazon River and
higher in the areas more influenced by the creek, whereas the
concentration of PO43� showed an opposite pattern, being
higher in the areas closer to the Amazon River. The
concentration of SiO4� was high in all locations and hence,
diatoms were probably not limited by inorganic silicium
(Manjarres-Hernandez et al., 2006).
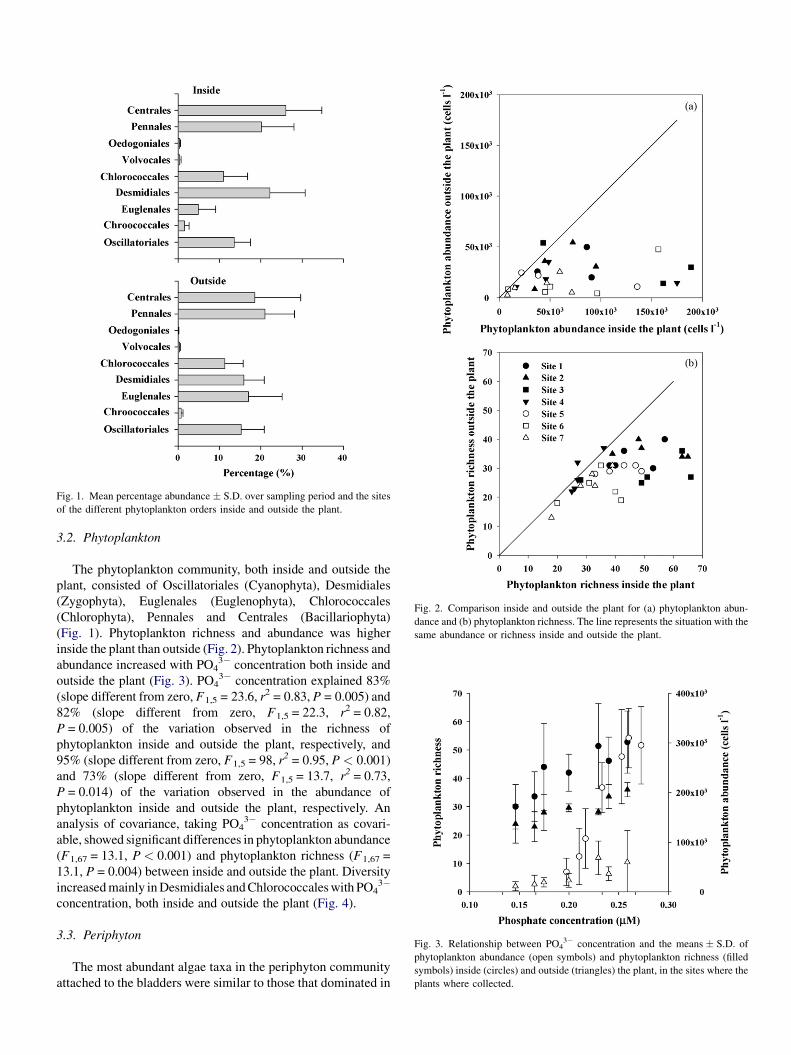
Fig. 1. Mean percentage abundance � S.D. over sampling period and the sites
of the different phytoplankton orders inside and outside the plant.
Fig. 2. Comparison inside and outside the plant for (a) phytoplankton abun-
dance and (b) phytoplankton richness. The line represents the situation with the
same abundance or richness inside and outside the plant.
Fig. 3. Relationship between PO43� concentration and the means � S.D. of
phytoplankton abundance (open symbols) and phytoplankton richness (filled
symbols) inside (circles) and outside (triangles) the plant, in the sites where the
plants where collected.
3.2. Phytoplankton
The phytoplankton community, both inside and outside the
plant, consisted of Oscillatoriales (Cyanophyta), Desmidiales
(Zygophyta), Euglenales (Euglenophyta), Chlorococcales
(Chlorophyta), Pennales and Centrales (Bacillariophyta)
(Fig. 1). Phytoplankton richness and abundance was higher
inside the plant than outside (Fig. 2). Phytoplankton richness and
abundance increased with PO43� concentration both inside and
outside the plant (Fig. 3). PO43� concentration explained 83%
(slope different from zero, F1,5 = 23.6, r2 = 0.83, P = 0.005) and
82% (slope different from zero, F1,5 = 22.3, r2 = 0.82,
P = 0.005) of the variation observed in the richness of
phytoplankton inside and outside the plant, respectively, and
95% (slope different from zero, F1,5 = 98, r2 = 0.95, P < 0.001)
and 73% (slope different from zero, F1,5 = 13.7, r2 = 0.73,
P = 0.014) of the variation observed in the abundance of
phytoplankton inside and outside the plant, respectively. An
analysis of covariance, taking PO43� concentration as covari-
able, showed significant differences in phytoplankton abundance
(F1,67 = 13.1, P < 0.001) and phytoplankton richness (F1,67 =
13.1, P = 0.004) between inside and outside the plant. Diversity
increased mainly in Desmidiales and Chlorococcales with PO43�
concentration, both inside and outside the plant (Fig. 4).
3.3. Periphyton
The most abundant algae taxa in the periphyton community
attached to the bladders were similar to those that dominated in
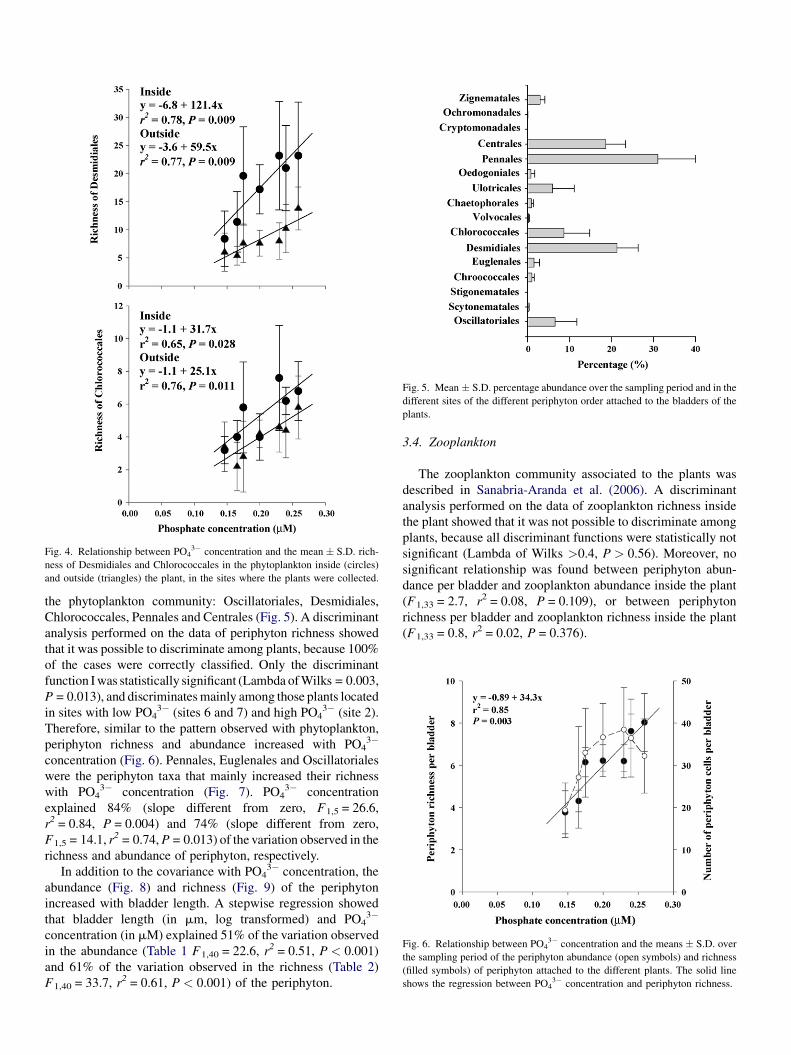
Fig. 4. Relationship between PO43� concentration and the mean � S.D. rich-
ness of Desmidiales and Chlorococcales in the phytoplankton inside (circles)
and outside (triangles) the plant, in the sites where the plants were collected.
Fig. 5. Mean � S.D. percentage abundance over the sampling period and in the
different sites of the different periphyton order attached to the bladders of the
plants.
Fig. 6. Relationship between PO43� concentration and the means � S.D. over
the sampling period of the periphyton abundance (open symbols) and richness
(filled symbols) of periphyton attached to the different plants. The solid line
shows the regression between PO43� concentration and periphyton richness.
the phytoplankton community: Oscillatoriales, Desmidiales,
Chlorococcales, Pennales and Centrales (Fig. 5). A discriminant
analysis performed on the data of periphyton richness showed
that it was possible to discriminate among plants, because 100%
of the cases were correctly classified. Only the discriminant
function I was statistically significant (Lambda of Wilks = 0.003,
P = 0.013), and discriminates mainly among those plants located
in sites with low PO43� (sites 6 and 7) and high PO4
3� (site 2).
Therefore, similar to the pattern observed with phytoplankton,
periphyton richness and abundance increased with PO43�
concentration (Fig. 6). Pennales, Euglenales and Oscillatoriales
were the periphyton taxa that mainly increased their richness
with PO43� concentration (Fig. 7). PO4
3� concentration
explained 84% (slope different from zero, F1,5 = 26.6,
r2 = 0.84, P = 0.004) and 74% (slope different from zero,
F1,5 = 14.1, r2 = 0.74, P = 0.013) of the variation observed in the
richness and abundance of periphyton, respectively.
In addition to the covariance with PO43� concentration, the
abundance (Fig. 8) and richness (Fig. 9) of the periphyton
increased with bladder length. A stepwise regression showed
that bladder length (in mm, log transformed) and PO43�
concentration (in mM) explained 51% of the variation observed
in the abundance (Table 1 F1,40 = 22.6, r2 = 0.51, P < 0.001)
and 61% of the variation observed in the richness (Table 2)
F1,40 = 33.7, r2 = 0.61, P < 0.001) of the periphyton.
3.4. Zooplankton
The zooplankton community associated to the plants was
described in Sanabria-Aranda et al. (2006). A discriminant
analysis performed on the data of zooplankton richness inside
the plant showed that it was not possible to discriminate among
plants, because all discriminant functions were statistically not
significant (Lambda of Wilks >0.4, P > 0.56). Moreover, no
significant relationship was found between periphyton abun-
dance per bladder and zooplankton abundance inside the plant
(F1,33 = 2.7, r2 = 0.08, P = 0.109), or between periphyton
richness per bladder and zooplankton richness inside the plant
(F1,33 = 0.8, r2 = 0.02, P = 0.376).
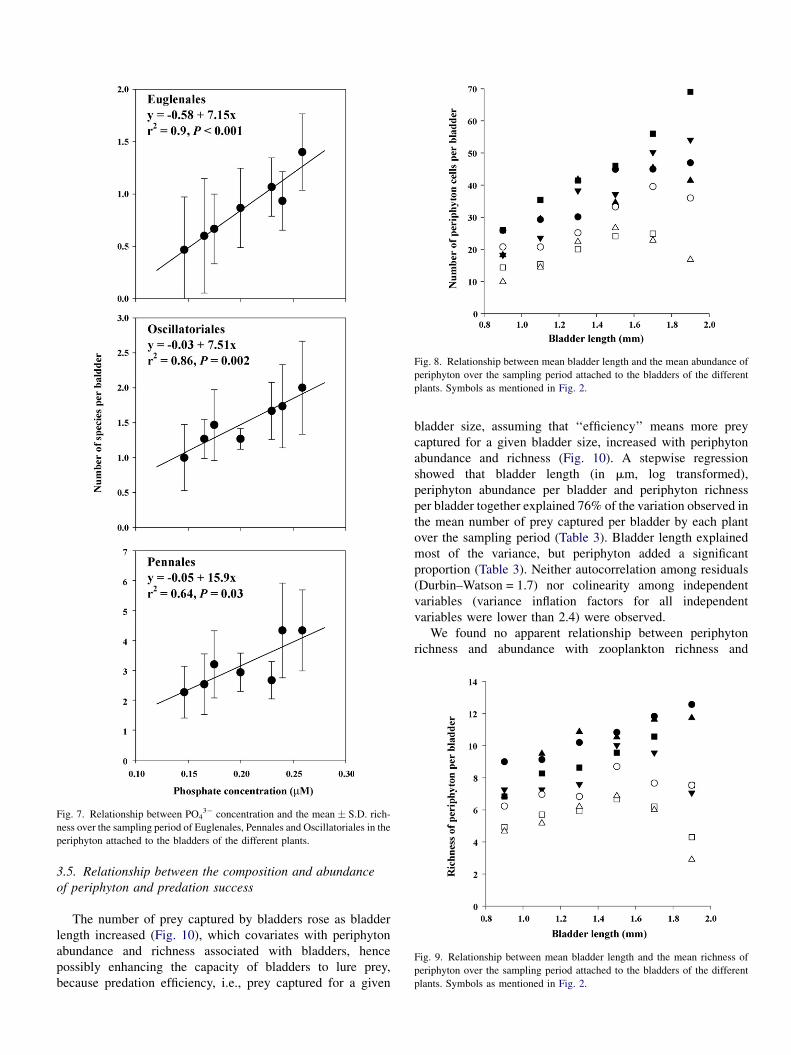
Fig. 7. Relationship between PO43� concentration and the mean � S.D. rich-
ness over the sampling period of Euglenales, Pennales and Oscillatoriales in the
periphyton attached to the bladders of the different plants.
Fig. 8. Relationship between mean bladder length and the mean abundance of
periphyton over the sampling period attached to the bladders of the different
plants. Symbols as mentioned in Fig. 2.
Fig. 9. Relationship between mean bladder length and the mean richness of
periphyton over the sampling period attached to the bladders of the different
plants. Symbols as mentioned in Fig. 2.
3.5. Relationship between the composition and abundance
of periphyton and predation success
The number of prey captured by bladders rose as bladder
length increased (Fig. 10), which covariates with periphyton
abundance and richness associated with bladders, hence
possibly enhancing the capacity of bladders to lure prey,
because predation efficiency, i.e., prey captured for a given
bladder size, assuming that ‘‘efficiency’’ means more prey
captured for a given bladder size, increased with periphyton
abundance and richness (Fig. 10). A stepwise regression
showed that bladder length (in mm, log transformed),
periphyton abundance per bladder and periphyton richness
per bladder together explained 76% of the variation observed in
the mean number of prey captured per bladder by each plant
over the sampling period (Table 3). Bladder length explained
most of the variance, but periphyton added a significant
proportion (Table 3). Neither autocorrelation among residuals
(Durbin–Watson = 1.7) nor colinearity among independent
variables (variance inflation factors for all independent
variables were lower than 2.4) were observed.
We found no apparent relationship between periphyton
richness and abundance with zooplankton richness and
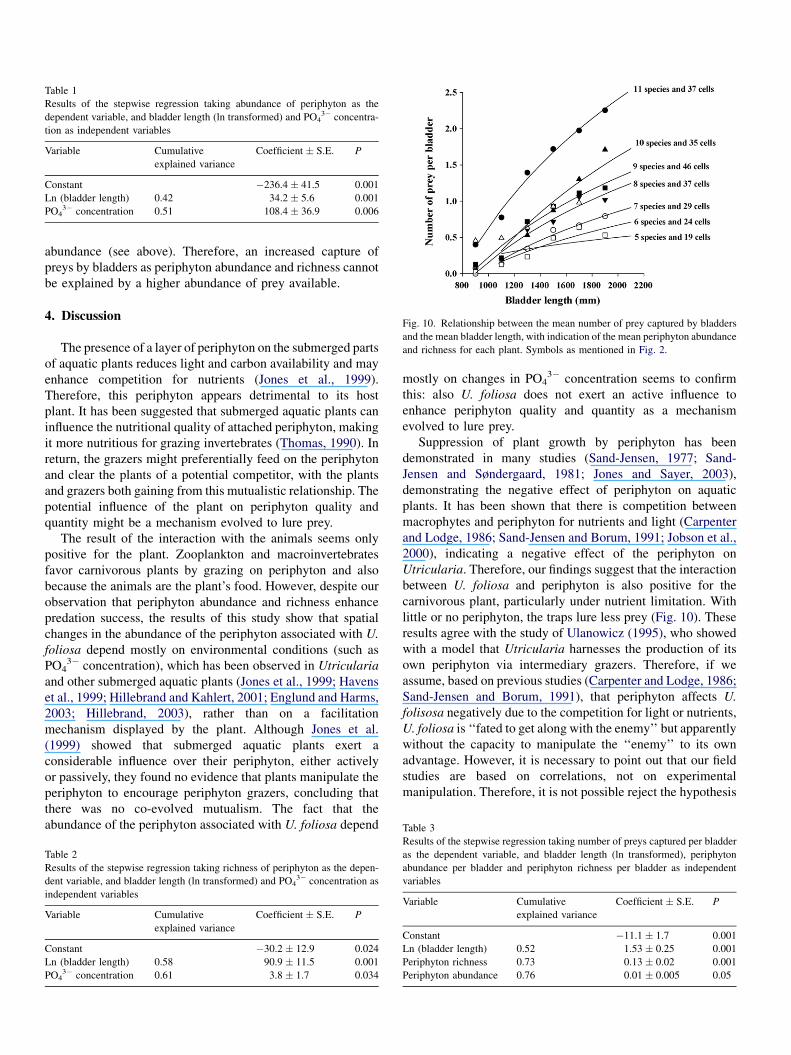
Table 1
Results of the stepwise regression taking abundance of periphyton as the
dependent variable, and bladder length (ln transformed) and PO43� concentra-
tion as independent variables
Variable Cumulative
explained variance
Coefficient � S.E. P
Constant �236.4 � 41.5 0.001
Ln (bladder length) 0.42 34.2 � 5.6 0.001
PO43� concentration 0.51 108.4 � 36.9 0.006
Fig. 10. Relationship between the mean number of prey captured by bladders
and the mean bladder length, with indication of the mean periphyton abundance
and richness for each plant. Symbols as mentioned in Fig. 2.
Table 3
abundance (see above). Therefore, an increased capture of
preys by bladders as periphyton abundance and richness cannot
be explained by a higher abundance of prey available.
4. Discussion
The presence of a layer of periphyton on the submerged parts
of aquatic plants reduces light and carbon availability and may
enhance competition for nutrients (Jones et al., 1999).
Therefore, this periphyton appears detrimental to its host
plant. It has been suggested that submerged aquatic plants can
influence the nutritional quality of attached periphyton, making
it more nutritious for grazing invertebrates (Thomas, 1990). In
return, the grazers might preferentially feed on the periphyton
and clear the plants of a potential competitor, with the plants
and grazers both gaining from this mutualistic relationship. The
potential influence of the plant on periphyton quality and
quantity might be a mechanism evolved to lure prey.
The result of the interaction with the animals seems only
positive for the plant. Zooplankton and macroinvertebrates
favor carnivorous plants by grazing on periphyton and also
because the animals are the plant’s food. However, despite our
observation that periphyton abundance and richness enhance
predation success, the results of this study show that spatial
changes in the abundance of the periphyton associated with U.
foliosa depend mostly on environmental conditions (such as
PO43� concentration), which has been observed in Utricularia
and other submerged aquatic plants (Jones et al., 1999; Havens
et al., 1999; Hillebrand and Kahlert, 2001; Englund and Harms,
2003; Hillebrand, 2003), rather than on a facilitation
mechanism displayed by the plant. Although Jones et al.
(1999) showed that submerged aquatic plants exert a
considerable influence over their periphyton, either actively
or passively, they found no evidence that plants manipulate the
periphyton to encourage periphyton grazers, concluding that
there was no co-evolved mutualism. The fact that the
abundance of the periphyton associated with U. foliosa depend
Table 2
Results of the stepwise regression taking richness of periphyton as the depen-
dent variable, and bladder length (ln transformed) and PO43� concentration as
independent variables
Variable Cumulative
explained variance
Coefficient � S.E. P
Constant �30.2 � 12.9 0.024
Ln (bladder length) 0.58 90.9 � 11.5 0.001
PO43� concentration 0.61 3.8 � 1.7 0.034
mostly on changes in PO43� concentration seems to confirm
this: also U. foliosa does not exert an active influence to
enhance periphyton quality and quantity as a mechanism
evolved to lure prey.
Suppression of plant growth by periphyton has been
demonstrated in many studies (Sand-Jensen, 1977; Sand-
Jensen and Søndergaard, 1981; Jones and Sayer, 2003),
demonstrating the negative effect of periphyton on aquatic
plants. It has been shown that there is competition between
macrophytes and periphyton for nutrients and light (Carpenter
and Lodge, 1986; Sand-Jensen and Borum, 1991; Jobson et al.,
2000), indicating a negative effect of the periphyton on
Utricularia. Therefore, our findings suggest that the interaction
between U. foliosa and periphyton is also positive for the
carnivorous plant, particularly under nutrient limitation. With
little or no periphyton, the traps lure less prey (Fig. 10). These
results agree with the study of Ulanowicz (1995), who showed
with a model that Utricularia harnesses the production of its
own periphyton via intermediary grazers. Therefore, if we
assume, based on previous studies (Carpenter and Lodge, 1986;
Sand-Jensen and Borum, 1991), that periphyton affects U.
folisosa negatively due to the competition for light or nutrients,
U. foliosa is ‘‘fated to get along with the enemy’’ but apparently
without the capacity to manipulate the ‘‘enemy’’ to its own
advantage. However, it is necessary to point out that our field
studies are based on correlations, not on experimental
manipulation. Therefore, it is not possible reject the hypothesis
Results of the stepwise regression taking number of preys captured per bladder
as the dependent variable, and bladder length (ln transformed), periphyton
abundance per bladder and periphyton richness per bladder as independent
variables
Variable Cumulative
explained variance
Coefficient � S.E. P
Constant �11.1 � 1.7 0.001
Ln (bladder length) 0.52 1.53 � 0.25 0.001
Periphyton richness 0.73 0.13 � 0.02 0.001
Periphyton abundance 0.76 0.01 � 0.005 0.05

that the bladders with more periphyton have more prey, not
because of the periphyton, but because of something associated
with more periphyton. Experimental manipulation would be
necessary to corroborate these correlations observed in the
field.
This study supports the need of a tri-trophic perspective,
rather than bi-trophic perspective based on either plant–
herbivore or herbivore–carnivore interactions alone (Singer and
Stireman, 2005).
Acknowledgements
We thank the Universidad Nacional de Colombia Sede
Amazonia, XUNTA de GALICIA and the University of Vigo
for financial support, and to Allan Wood for useful comments
on the manuscript.
References
Blindow, I., 1987. The composition and density of epiphyton on several species
of submerged macrophytes—the neutral substrate hypothesis tested. Aquat.
Bot. 29, 157–168.
Bosserman, R.W., 1983. Elemental composition of Utricularia-periphyton
ecosystems from Okefenokee Swamp. Ecology 64, 1637–1645.
Carpenter, S.R., Lodge, D.M., 1986. Effects of submersed macrophytes on
ecosystem processes. Aquat. Bot. 26, 341–370.
Cohn, F., 1875. Uber die Funktion der Blasen von Aldrovanda und Utricularia.
Beitr. Biol. Pflanzen 1, 71–92.
Englund, G., Harms, S., 2003. Effects of light and microcrustacean prey on
growth and investment in carnivory in Utricularia vulgaris. Freshwater
Biol. 48, 786–794.
Friday, L., Quarmby, C., 1994. Uptake and translocation of prey-derived 15N
and 32P in Utricularia vulgaris L. New Phytol. 121, 439–445.
Gross, E.M., Hilt (nee Korner), S., Lombardo, P., Mulderij, G., 2007.
Searching for allelopathic effects of submerged macrophytes on phyto-
plankton-state of the art and open questions. Hydrobiologia 584, 77–
88.
Guisande, C., Andrade-Sossa, C., Granado-Lorencio, C., Duque, S.R.,
Nunez-Avellaneda, M., 2000. Effects of zooplankton and conductivity on
tropical Utricularia foliosa investment in carnivory. Aquat. Ecol. 34, 137–
142.
Guisande, C., Aranguren, N., Andrade-Sossa, C., Prat, N., Granado-Lorencio,
C., Barrios, M.L., Bolivar, A., Nunez-Avellaneda, M., Duque, S.R., 2004.
Relative balance of the cost and benefit associated with carnivory in the
tropical Utricularia foliosa. Aquat. Bot. 80, 271–282.
Guisande, C., Barreiro, A., Maneiro, I., Riveiro, I., Vergara, A., Vaamonde, A.,
2006. Tratamiento de Datos. Dıaz de Santos, Madrid, Spain.
Harms, S., 1999. Prey selection in three species of the carnivorous aquatic plant
Utricularia (bladderwort). Arch. Hydrobiol. 146, 449–470.
Hasle, G.R., Frixell, G.A., 1970. Diatoms: cleaning and mounting for light and
electron microscopy. Trans. Am. Microsc. Soc. 89, 469–474.
Havens, K.E., East, T.L., Rodusky, A.J., Sharfstein, B., 1999. Littoral periph-
yton responses to nitrogen and phosphorus: an experimental study in a
subtropical lake. Aquat. Bot. 63, 267–290.
Hillebrand, H., 2003. Opposing effects of grazing and nutrients on diversity.
Oikos 100, 592–600.
Hillebrand, H., Kahlert, M., 2001. Effect of grazing and nutrient supply on
periphyton biomass and nutrient stoichiometry in habitats of different
productivity. Limnol. Oceanogr. 46, 1881–1898.
Jobson, R.W., Morris, E.C., 2001. Feeding ecology of a carnivorous bladder-
wort (Utricularia uliginosa Lentibulariaceae). Aust. Ecol. 26, 680–691.
Jobson, R.W., Morris, E.C., Burgin, S., 2000. Carnivory and nitrogen supply
affect that growth of the bladderwort Utricularia uliginosa. Aust. J. Bot. 48,
549–560.
Jones, J.I., Sayer, C.D., 2003. Does the fish-invertebrate-periphyton cascade
precipitate plant loss in shallow lakes? Ecology 84, 2155–2167.
Jones, J.I., Young, J.O., Haynes, G.M., Moss, B., Eaton, J.W., Hardwick, K.J.,
1999. Do submerged aquatic plants influence their periphyton to enhance
the growth and reproduction of invertebrate mutualists? Oecologia 120,
463–474.
Lacoste de Diaz, E.L., 1981. Desmidiaceae en Utricularia foliosa L. Lilloa
XXXV 3, 67–83.
Luetzelburg, P., 1910. Beitrage zur Kenntniss der Utricularien. Flora Bot. Zeit.
100, 145–212.
Manjarres-Hernandez, A., Guisande, C., Torres, N.N., Valoyes-Valois, V.,
Gonzalez-Bermudez, A., Dıaz-Olarte, J., Sanabria-Aranda, L., Duque,
S.R., 2006. Temporal and spatial change of the investment in carnivory
of the tropical Utricularia foliosa. Aquat. Bot. 85, 212–218.
Meyers, D.G., 1982. Darwin‘s investigation of carnivorous aquatic plants of the
genus Utricularia: misconception, contribution, and controversy. Proc.
Natl. Acad. Sci. Philadelphia 134, 1–11.
Meyers, D.G., Strickler, J.R., 1979. Capture enhancement in a carnivorous
aquatic plant: function of antennae and bristles in Utricularia vulgaris.
Science 203, 1022–1025.
Sanabria-Aranda, L., Gonzalez-Bermudez, A., Torres, N., Guisande, C., Man-
jarres-Hernandez, A., Valoyes-Valois, V., Dıaz-Olarte, J.J., Andrade-Sossa,
C., Duque, S.R., 2006. Predation by the tropical plant Utricularia foliosa.
Freshwater Biol. 51, 1999–2008.
Sand-Jensen, K., 1977. Effects of epiphytes on eelgrass photosynthesis. Aquat.
Bot. 3, 55–63.
Sand-Jensen, K., Borum, J., 1991. Interactions among phytoplankton, periph-
yton and macrophytes in temperate freshwaters and estuaries. Aquat. Bot.
41, 137–175.
Sand-Jensen, K., Søndergaard, M., 1981. Phytoplankton and epiphyte devel-
opment and their shading effect on submerged macrophytes in lakes of
different nutrient status. Int. Rev. Gesamten Hydrobiol. 66, 529–552.
Singer, M.S., Stireman, J.O., 2005. The tri-trophic niche concept and adaptive
radiation of phytophagous insects. Ecol. Lett. 8, 1247–1255.
SPSS, 2003. Copyright 1989–2003# SPSS Inc.
STATISTICA 7, 2004. Copyright 1984–2006# Statsoft Inc.
Thomas, J.D., 1990. Mutualistic interactions in freshwater modular systems
with molluscan components. Adv. Ecol. Res. 20, 125–178.
Ulanowicz, R.E., 1995. Utricularias secrets the advantage of positive feedback
in oligotrophic environments. Ecol. Model. 79, 49–57.
Wagner, G.M., Mshigeni, K.E., 1986. The Utricularia-Cyanophyta association
and its nitrogen-fixing capacity. Hydrobiologia 141, 255–261.
Top Related
Copyright © 2022 FDOKUMEN

