







Bahasa
Halaman
Hukum


doi:10.1182/blood-2006-08-040238Prepublished online February 1, 2007;
Truong, Ioannis Theodorou, Francoise Barre-Sinoussi, Gianfranco Pancino and Pascale PaulSophie Ravet, Daniel Scott-Algara, Elodie Bonnet, Hung Khiem Tran, Ton Tran, Ngai Nguyen, Lien Xuan cell activation in HIV-exposed uninfected individualsDistinctive NK cell receptor repertoires sustain high-level constitutive NK
(4651 articles)Immunobiology � (3368 articles)Clinical Trials and Observations �
Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
digital object identifier (DOIs) and date of initial publication. theindexed by PubMed from initial publication. Citations to Advance online articles must include
final publication). Advance online articles are citable and establish publication priority; they areappeared in the paper journal (edited, typeset versions may be posted when available prior to Advance online articles have been peer reviewed and accepted for publication but have not yet
Copyright 2011 by The American Society of Hematology; all rights reserved.20036.the American Society of Hematology, 2021 L St, NW, Suite 900, Washington DC Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly by
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

1
Distinctive NK cell receptor repertoires sustain high-level constitutive NK cell activation
in HIV-exposed uninfected individuals
Short title: NK cells in HIV-exposed uninfected individuals
Sophie Ravet1, 2, Daniel Scott-Algara3, Elodie Bonnet1, Hung Khiem Tran4, Ton Tran4, 6,
Ngai Nguyen5, Lien Xuan Truong4, Ioannis Theodorou6, Françoise Barré-Sinoussi3,
Gianfranco Pancino3, Pascale Paul1, 2*
1- Laboratoire Exploration NK, Laboratoire d’Hématologie, CHU Conception,
Assistance Publique Hôpitaux de Marseille, 147 boulevard Baille, 13005 Marseille,
France
2- UMR-S 608 INSERM, URF de Pharmacie Université de la Méditerranée, 27 Bd Jean
Moulin, 13385 Marseille 5, 13005 Marseille
3- Unité de Régulation des Infections Rétrovirales, Institut Pasteur, Paris France
4- Pasteur Institute, Ho Chi Minh City, Vietnam
5- Binh Trieu Hospital, Ho Chi Minh City, Vietnam
6- INSERM U543, CHU Pitié-Salpêtrière, Paris, France
Corresponding author Pascale Paul Laboratoire Exploration NK, Hematology unit Hôpital de la Conception 147 Bd Baille 13005 Marseille, France Tel. 33 (0) 49 13 83 920 Fax 33(0) 49 13 83 012 Email: [email protected]
Word counts: Abstract : 199 Text: 4877 Scientific heading: Immunobiology
Blood First Edition Paper, prepublished online February 1, 2007; DOI 10.1182/blood-2006-08-040238
Copyright © 2007 American Society of Hematology
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

2
Abstract
We have previously associated high natural killer (NK) cell activity and protection
against HIV-1 infection in Vietnamese exposed uninfected intravenous drug users (EUs).
Considering that activating and inhibitory signals sensed by NK cell receptors regulate NK
cell activation, we performed phenotypic and qRT-PCR transcript analyses of the NK cell
receptor (NKR) repertoire in 25 EUs, 19 HIV+ intravenous drug users and 26 uninfected
blood donors. Although NK cell activation was not linked to a unique NKR repertoire in EUs,
various patterns consistent with NK cell activation were detected in EUs: high
KIR3DS1/KIR3DL1 ratio associated with down-regulated KIR3DL1 transcript levels,
KIR2DL3+ low affinity receptor expansion associated to group HLA-C1 ligand in 2DS2-
/2DL2- EUs, enhanced NKG2C/NKG2A ratio, and increased CD69 expression. Remarkably,
EUs exhibited high constitutive degranulation activity in the absence of exogenous
stimulation, as shown by the CD107a assay. Furthermore, CD161 expression was increased
within the CD107a+ NK cell compartment. Our results suggest that in response to viral
exposition, particular genetic and/or regulated features of the NKR repertoire of EUs
contribute to their high constitutive NK cell potential. This might allow NK cells to generate a
more rapid and effective immune response to HIV-1, thereby contributing to prevention
toward infection.
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

3
Introduction
Natural killer (NK) cells are key actors in innate host immune responses to a variety of
pathological challenges. NK cell activation results from integration of signals induced by the
interaction of a highly diversified repertoire of inhibitory and activating NK cell receptors
(NKRs) with constitutive or inducible ligands expressed on target cells, including virus-
infected cells1-5. Inhibitory receptors essentially recognize specific subsets of classical and
non-classical HLA class molecules, and include KIR-L, ILT2/CD85j, and heterodimeric
CD94/NKG2A NKRs. Inhibitory KIR2DL1 recognizes HLA-C group 2 alleles (HLA-C2:
Asn77Lys80), while KIR2DL2 and KIR2DL3 both recognize HLA-C group 1 alleles (HLA-
C1: Ser77Asn80) but with different affinity2,6. KIR3DL1 recognizes the HLA-Bw4 group of
alleles 7and KIR3DL2 HLA-A11/HLA-A3. Recognition of the relevant HLA class I molecule
by KIR-L receptors delivers NK inhibitory signals, whereas non recognition allow
stimulatory signals to activate NK cells. MHC-KIR interactions shape the NK cell repertoire
and calibrate NK cell potential to control infectious, tumoral or alloreactive challenges5,8,9 . In
contrast to KIR-L, activating KIR-S counterparts can deliver activating signals to NK cells,
but their physiological ligands remain mostly uncharacterized. Recent evidence suggest that
KIR2DS1 and KIR2DS2 respectively share the capacity to bind HLA-C2 or -C1 ligands with
their inhibitory counterparts in vitro, but with lower affinity10. Receptors delivering activating
signaling include NK-specific natural cytotoxicity receptors (NCRs), in addition to receptors
more broadly distributed among T lymphocyte subsets, such as activating KIR-S,
CD94/NKG2C, NKG2D, CD226/DNAM and CD161/NKRP1. Activated NK cells mediate
various effector mechanisms, including antibody-dependent and natural cytotoxicity, and
secretion of chemokines and cytokines with antiviral activity1,2,4,5. In addition to these effector
functions, crosstalk between NK cells and antigen-presenting cells, and cytokine secretion by
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

4
NK cells, influence the efficiency of adaptive immune responses, thereby constituting a major
link between innate and adaptive immunity6,7.
NK cells therefore have the potential to contribute to the prevention and control of
viral infections, including HIV-1 infection. The NK cell phenotype and functions are altered
in HIV+ patients7-14. Up-regulation of inhibitory receptors in viremic patients has been
proposed as a basis for the inhibition of NK cell activity observed in such subjects12,15,16.
Decreased expression of NKp30 and NKp46 NCR activating receptors has also been linked to
impaired cytolytic potential in viremic patients relative to HAART-treated aviremic subjects
and to healthy individuals11-13. Partial quantitative and functional reconstitution of NK cells
can be obtained during HAART, thus linking NK cell suppression to HIV replication14-17. A
role of NK cells in controlling HIV disease progression is suggested by the association
between delayed progression to AIDS and concomitant presence in the host genome of genes
that contribute to the HLA-Bw4 allelic group and activating KIR3DS1 NK receptor genes18.
The rare individuals who are naturally resistant to HIV-1 infection represent a unique model
in which to examine the role of innate immunity in the control of HIV-1, with potential
implications for preventive and therapeutic strategies. We have previously observed enhanced
NK cell cytotoxic activity and cytokine and β-chemokine production in Vietnamese
intravascular drug users who remained uninfected despite several years of high-risk exposure
to HIV-1, when compared with HIV+ drug users (IDU) and low-risk uninfected blood
donors19. In contrast, NK cell activity was low in some drug users who seroconverted during
the study19. These results suggested that NK cells contribute to the protection of these
exposed uninfected individuals (EUs). A recent study analyzing genomic HLA-KIR
combinations in African female sex workers suggests that resistance to HIV-1 infection
associates with inhibitory KIR in absence of their HLA ligand20. The present study was
designed to investigate whether specific patterns of NK cell receptor expression associate
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

5
with higher NK cytotoxic and secretory activities in Vietnamese EU IDUs. We therefore
conducted comparative genomic, transcriptional and phenotypic analysis of the NK cell
repertoire, in association with the functional assay of the CD107a degranulation potential of
NK cells, in EUs, HIV+ IDUs, and uninfected low-risk blood donors.
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

6
Materials and Methods
Study population
Twenty-five exposed uninfected (EUs) intravascular drug users (IDU) from a previously
described cohort recruited in the Binh Trieu Hospital in Ho Chi Minh City19,21 were included
in this study. Risk factors for HIV exposure, the high seroprevalence of other blood-borne
viruses such HBV, HCV and HTLV, and the follow-up for this sub-study were previously
described19. ELISA confirmed the HIV-1 negative status of EUs and PCR as described21.
Nineteen HIV+ intravascular drug users were studied (HIV+ IDUs) and offered free diagnosis,
medical follow-up and treatment for opportunistic infections. Twenty-six low-risk uninfected
controls (C) were recruited from the Blood Bank of Ho Chi Minh City. Informed consent was
obtained from each participant at recruitment. All the subjects were males, and there was no
age difference among the study groups. The Ethics Committee of Binh Trieu Hospital, Ho
Chi Minh City, approved the protocol. Peripheral blood mononuclear cells (PBMC) were
recovered from blood samples after Ficoll-Hypaque gradient and cryopreserved in 90% FCS,
10% DMSO. When PBMC or NK cell counts were a limiting factor, in particular in HIV+
individuals, functional, phenotypic and transcriptional analysis of NK cells were privileged
over DNA extraction as a source of informative data.
HLA and KIR genotyping
HLA genotyping was performed with a reverse dot blot assay using a commercial kit and
according the manufacturer's instructions (Innogenetics Belgium). Due to the limited
availability of biological samples, KIR genotyping could only be performed on 14 EUs, 8
HIV+ IDUs, and 12 controls. A reverse hybridization assay read using Luminex technology
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

7
was used for KIR genotyping according to manufacturer’s instructions (One Lambda, Inc.,
CA USA).
RNA purification and real time RT-PCR analysis of KIR transcript levels
Total RNA was isolated from PBMC using RNeasy mini kits (QIAGEN Inc., Valencia, CA,
USA) including a DNase I digestion step removing genomic DNA. Total RNA (5µg) was
converted to cDNA using 200 U of Supercript II reverse transcriptase (Invitrogen). Real-time
PCR amplification was performed with the FastStart DNA MasterPLUS SYBR Green I kit as
recommended by the manufacturer, on a LightCycler instrument (Roche). Cycling conditions
were 10min at 95°C (hot-start PCR), followed by 40 cycles, 10s at 95°C (denaturation), 10s at
62°C (annealing) and 20s at 72°C (elongation). Melting curve analysis was performed to
check the specificity of amplification. Reported values are relative numbers of specific
transcripts detected per 106 GAPDH transcripts. KIR expression was analyzed with primers
specifically designed to discriminate KIR locus, as follows (forward and reverse sequences):
KIR2DL3 5’-CTCCTTCATCGCTGGTGCTG-3’ and 5’-
GGCAGGAGACAACTTTGGATCA-3’; KIR2DL4 5’-CTCCCAGAGCTCCTTTGACATC-
3’ and 5’-GAGCCGAAGCATCTGTAGGTCT-3’; KIR3DS1 5’-
CCAAGCTCCAAATCTGGTAACCT-3’ and 5’-TCAGAATCCTCGCTGTTCACTTCT-3’;
KIR3DL1 5’-GGACATCGTGGTCACAGGTCC-3’ and 5’-
CACTGAGGTCCCAATCAGAATG-3’; KIR3DL2 5’-CCAACTTCTCCATCGGTCCCT-3’
and 5’-GTTGACCTTGGGCACTGCAC-3’; GAPDH 5’-
GGTGGTCTCCTCTGACTTCAACA-3’ and 5’-GTTGCTGTAGCCAAATTCGTTGT-3’.
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

8
Phenotypic analysis
The percentage of CD3-CD56+CD16+ NK cells among total lymphocytes and the size of each
NKR+ NK cell subset were determined by three-color flow cytometry (Beckman-Coulter).
Eleven distinct NK cell subsets were defined as NK cells (CD3-CD56+) expressing specific
NKRs. The following antibodies were used: FITC-conjugated anti-CD3 (IgG1, UCHT1);
PC5-conjugated anti-CD56 (IgG1, N901); PE-conjugated: anti-CD158a/h (clone EB6, IgG1
recognizing KIR2DS1/2DL1), anti-CD158b1/b2/j (clone GL183, IgG1 recognizing
KIR2DS2/2DL2/2DL3), anti-CD158e1/e2 (clone Z27.3.7, IgG1, recognizing
KIR3DL1/KIR3DS1), anti-CD158i (clone FES172, IgG2a, recognizing KIR2DS4), anti-
CD25 (IgG2a, B.1.49.9), anti-CD161 (IgG1, 191B8), anti-NKp30 (IgG1, Z25), anti-NKp46
(IgG1, BAB281), anti-CD85j (IgG1, HP-F1), anti-CD159a (IgG2b, Z199), (all from Beckman
Coulter), anti-CD69 (IgG1, FN50, BD-Pharmingen), and anti-NKG2C (IgG1, 134591, R&D
Systems).
Flow cytometer analysis of NK function using the degranulation marker
CD107a/LAMP-1 (lysosome-associated membrane protein 1)
CD107a expression was used to measure NK cell degranulation, as described22,23. PBMC
were incubated with or without HLA class I -negative K562 target cells (ATCC) at an E: T
ratio of 2:1 for 3 hours. CD107a-PC5 antibody (IgG1, H4A3) and GolgiStop reagent diluted
at 1:150 (BD/Pharmingen) were added directly to the cultures. Cells were then stained with
anti-CD3-FITC (IgG1, UCHT1), CD56-PC7 (IgG1, N901) prior to FACS analysis. Additional
NKR-PE staining was performed with anti-CD85j, anti-CD159a, anti-CD161 (Beckman-
Coulter), anti-NKG2C (R&D System), and anti-CD69 (BD Pharmingen) antibodies. All
samples were analyzed by means of four-color flow cytometry and CXP software (Beckman-
Coulter).
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

9
Statistical analysis
Analyses were performed with Prism software (GraphPad 4.0b) implementing the
nonparametric Kruskall-Wallis test followed by Dunn’s post-test to compare three or more
continuous variables, the Mann-Whitney test to compare two unpaired groups, and the
Wilcoxon matched pairs test for the CD107a degranulation assay results.
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

10
Results
Distinctive features of the KIR repertoire in exposed uninfected intravascular drug
users
The size of the NK cell compartment did not differ between EUs and controls (median 16%
and 16.4% respectively), whereas NK cell percentages were lower in HIV+ IDUs (HIV)
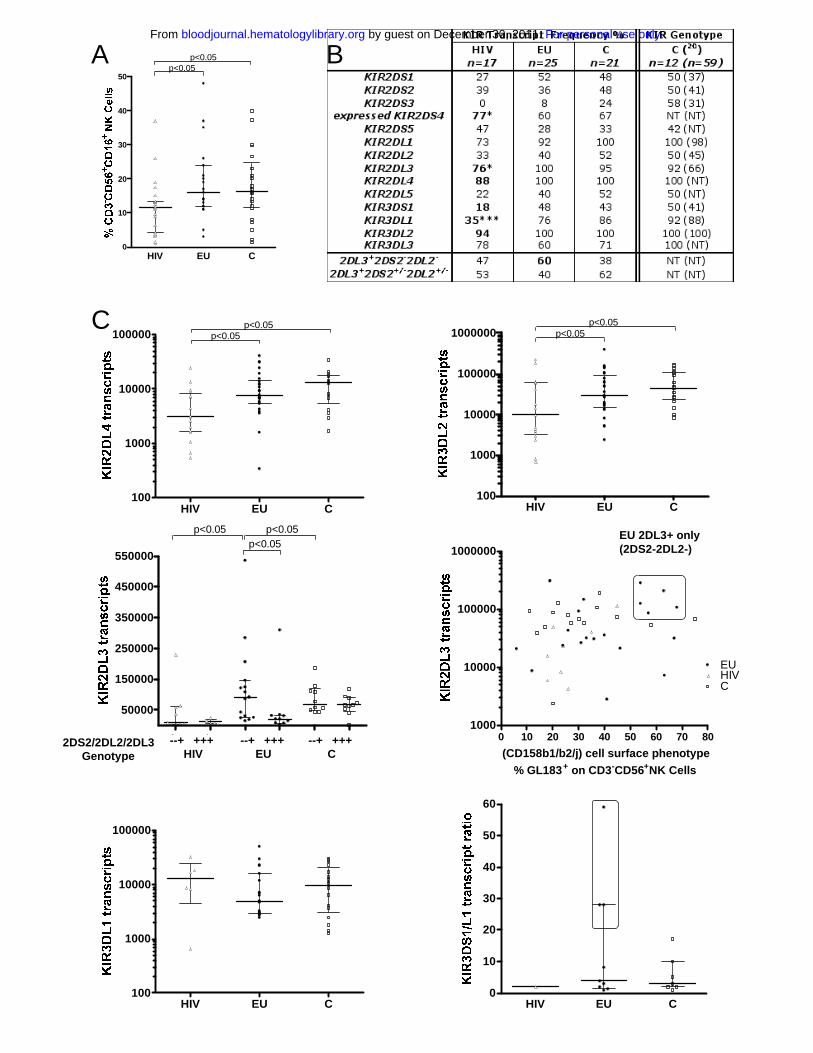
(median 11.6%, p<0.05, Figure 1A).
KIR+ NK cell subsets distribution was analyzed by flow cytometry in 25 EUs (Table 1), 19
HIV+ and 26 unexposed Vietnamese control donors. Four KIR+CD3-CD56+ NK cell subsets
were evaluated using antibodies EB6 (CD158a/h or KIR2DL1/2DS1), GL183 (CD158b1/b2/j
or KIR2DL2/2DL3/2DS2), Z27.3.7 (CD158e1/e2 or KIR3DL1/3DS1) and FES172 (CD158i
or KIR2DS4). As previously described in Caucasians24, high inter-individual heterogeneity
was observed in the distribution of each KIR+ NK cell subsets. Still, no significant differences
in the size of these KIR+ NK cell phenotype subsets were observed among the three
Vietnamese groups (data not shown).
As phenotypic analysis of the KIR repertoire with currently available mAbs cannot
discriminate between inhibitory and activating KIR-expressing NK cell subsets, we further
performed real-time PCR analysis of the KIR repertoire to quantify the mRNA transcripts
corresponding to each of the five activating KIR-S and six inhibitory KIR-L receptors. In
addition, HLA and KIR genotyping was also performed in 14 EUs, 8 HIV+ IDUs and 12
controls for which enough cells were available for DNA analysis (Table 1).
Overall, the KIR gene frequencies observed in our control group (Figure 1B) were
comparable to those previously reported in Vietnamese blood donors25. Moreover, we show
that KIR transcripts frequencies were mostly similar in EUs and controls (Figure 1B). In
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

11
contrast, transcript frequencies of given KIRs were selectively altered within the HIV+ group
(Figure 1B).
Remarkably, KIR2DL4 and KIR3DL2 transcripts, corresponding to core KIR genes found at
the genomic level in all individuals, were found in all EUs and controls, but were
undetectable in some HIV+ individuals (Figure 1B). Similarly, the core KIR3DL3 gene was
detected in 100% of controls but the corresponding transcripts were undetectable in 22% of
HIV+, 40% of EU and 29% of controls tested. The dissociation between KIR3DL3 gene
content and transcript frequency (p<0.05) was thus not specific of a given group (Figure 1B).
Transcript frequencies of other KIRs showed significant differences in HIV+ subjects in
comparison with EUs and controls (Figure 1B). First, the frequency of individuals with
detectable KIR3DL1 and KIR2DL3 transcripts were lowered in HIV+ IDUs (p=0.0007 and
p=0.0174 respectively). Similarly, frequency of individuals expressing KIR3DS1 transcripts
tended to be lower in HIV+ individuals (p=0.0992). Our transcriptional analysis included
design of specific primers that discriminate transcripts corresponding either to the KIR1D null
allele that is not expressed as an NK receptor or to the group of KIR2DS4 alleles that
contribute to cell surface expression of the KIR2DS4/CD158i receptor26. Using this
approach we showed that frequency of KIR2DS4 transcripts coding for KIR2DS4
receptor was higher in HIV+ IDUs than in EUs and controls (p=0.035, Figure 1B).
When allowed by cell or DNA availability, we further investigated whether the absence of
detectable KIR3DL1 or KIR3DS1 transcripts was associated to the absence of the gene or
whether it may correspond to lack of transcription in individuals who bear the gene. For
KIR3DS1, a good correlation between gene content and transcript detection was observed in
all groups. In particular, HIV+ IDUs in whom KIR3DS1 transcripts were undetectable were
also typed as KIR3DS1 negative at the gene level. In contrast, while the presence of the
KIR3DL1 gene was always associated to KIR3DL1 transcripts in controls, 3 out of 8 HIV+
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

12
IDUs and 3 out of 14 EUs genotyped were shown to be KIR3DL1-positive at the gene level
but lack the corresponding transcripts.
We further analyzed quantitative differences in the levels of KIR transcript expression in the
three groups (Figure 1C and results not shown). KIR2DL4 and KIR3DL2 transcripts levels
were lower in HIV+ subjects than in controls or EUs (p<0.05, Figure 1C). On the other hand,
KIR3DL3 transcript levels were also lower in EUs than in HIV+ and controls (p<0.05, data not
shown).
In contrast, in HIV+ subjects. KIR2DL3 transcript levels were lowered in reference to EUs and
controls (p<0.05 and p<0.001 respectively). Combined KIR2DS2-/2DL2-/2DL3+ transcripts
pattern tended to be more frequent in EUs than in HIV+ or controls (p=0.10, Fig 1B). We
further observed higher KIR2DL3 transcript levels in these KIR2DS2-/2DL2-/2DL3+ EU
individuals (p<0.05, Figure 1C). Notably, 5 EUs exhibiting more than 50% GL183+ stained
NK cells were further identified as KIR2DL3+and 2DL2-/2DS2-, both at the gene and
transcript levels (Figure 1C). KIR2DL3 was thus the only KIR gene associated to the GL183+
phenotype in these individuals.
Further analysis of HLA groups associated with the KIR repertoire in EUs showed that
groups HLA-C1/C2 and HLA-C1/C1 were equally represented, and that only 2 EUs were
homozygous HLA-C2/C2 (Table 1). The highly represented GL183+ NK cell subsets in
KIR2DL3+/2DL2-/2DS2- EUs was always associated with presence of the HLA-C1 ligand in
these individuals.
KIR3DL1 transcript levels also tended to be lower in EUs than in HIV+ subjects and controls
(Figure 1C), although statistic significance was not reached. Accordingly, and given the high
KIR3DS1 transcript levels in some EUs, the ratio between activating KIR3DS1 and inhibitory
KIR3DL1 transcript levels was higher in some EUs, and particularly elevated at values never
observed in tested controls in three EUs (Figure 1C; n° 6, 7, 16 in Table 1).
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

13
Increased expression of NK cell activation markers in EUs
The NK cell repertoire was further characterized by phenotypic analysis of NK cell subsets
expressing activating and inhibitory NKRs, CD25 and CD69 activation markers (Figure 2A,
B and data not shown). NKp30 expression was found on all NK cells and at comparable
levels in EUs and controls. The level of NKp30, assayed by mean fluorescence intensity of
NKp30 staining (mfi), was lower in HIV+ than in EUs and controls (p<0.05, Figure 2A). In
contrast, no significant difference in NKp46 (Figure 2A) nor NKG2D and DNAM expression
(data not shown) was observed among the three groups. Although the size of the CD161+ NK
cell subset was not different among the three groups, CD161 mean fluorescence in
CD161+CD3-CD56+ cells was significantly higher in EUs than in controls (p<0.05, Figure
2A). While the NK-cell surface expression of the CD25 activation marker was comparable in
all tested groups (data not shown), higher percentage of CD69+ NK cells was observed in EUs
and HIV+ as compared to controls (p<0.05, Figure 2A). Overall, CD85j, NKG2A and
NKG2C expression on NK cells did not differ significantly among the groups. Nevertheless,
expression of the activating NKG2C subunit of the CD94/NKG2 receptor was inversely
correlated to that of its inhibitory NKG2A counterpart in EUs, but not in the other two groups
(p<0.05, Figure 2A).
Strong constitutive activation of NK cells in EUs.
We then investigated the granule exocytosis pathway characterizing NK cell activation by
evaluating CD107a+ expression within CD3-CD56+ NK cells23, both without stimulation and
in the presence of stimulatory K562 cells (Figure 3).
Without stimulation, the size of the activated CD107a+ NK cell subset was much larger in
EUs (median: 34%, 25-75% interquartile ranges: 31-50%) than in HIV+ (10%, 5-19%,
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

14
p<0.01) and controls (8%, 1.7-12%, p<0.001) (Figure 3A, C). Kinetic analysis of CD107a
degranulation at 10 min, 1h, 2h and 3h showed that degranulation of unstimulated EU NK
cells occurred between 10 min and 1 hour, in contrast to 2 hours in cells from controls (data
not shown). Median percentage of CD107a+ cells among control CD3-CD56+ NK cells was
increased by K562 stimulation (from 8% to 17%, p<0.05), but remained unchanged in HIV+
individuals (Figure 3A).
Upon stimulation by K562 cells, median percentage of CD107a+ NK cells remain
significantly higher in EUs (39%, 24-46%) than in HIV+ (9.5%, 8-15%, p<0.001). and
stimulated controls (17%, 9-25%, p<0.01, Figure 3A).
We further analysed variations in intensity of CD107a expression, as assayed by mfi of
CD107 staining within the activated CD3-CD56+ NK cell subset. Stimulation with K562 cells
induced increased CD107a mean fluorescence staining intensity within CD107+ NK cells, in
controls as well as in EUs (Figure 3B). In contrast, the mfi of CD107a+ NK cells from HIV+
individuals was unchanged after K562 stimulation (Figure 3B).
Increased CD161 expression is associated with the CD107a+ activated NK compartment
Finally, we examined the NK cell surface expression of CD69, CD85j, NKG2A, NKG2C and
CD161 within the activated CD107+CD3-CD56+ NK cell compartment, both in the presence
and absence of stimulatory K562 target cells. Overall, the distribution of CD85j, NKG2A, and
NKG2C expression among CD107a+CD3-CD56+ NK cells did not differ significantly among
the three study groups (data not shown). Median percentages of CD69 expression among
CD107a+ NK cells tended to be higher in EUs (40%) than in HIV+ (20%) and controls (28%)
in the absence of K562 stimulation, and were induced to similar levels in the three groups
after K562 stimulation (data not shown).
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

15
In contrast, CD161 expression differed significantly between EUs and the other groups
(Figure 4). In the absence of K562 stimulation, the percentage of CD161+ cells among
CD107+CD3-CD56+ NK cells were higher in EUs than in controls (p<0.05) and HIV+ subjects
(p<0.01, Figure 4A, B). In addition, the intensity of CD161 staining in unstimulated
CD161+CD107+CD3-CD56+ NK cells (mfi) was comparable in EUs and controls and
significantly lowered in HIV+ (p<0.05, Figure 4B, C), while not affected by K562 stimulation
in any of the three groups (data not shown). Among CD161+ NK cells, intensity of CD161
staining (mfi) in EUs tended to be higher in the CD107a+ NK subset than in the CD107a- NK
subset (p=0.12, Figure 4D). In contrast, CD161 expression levels were similar in the CD107a+
and CD107- NK cell compartments in HIV+ and controls subjects (Figure 4D).
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

16
Discussion
The role of NK cells in controlling HIV infection has mainly been investigated in infected
individuals. Thus, while considerable information has been gained on NK cell dysfunctions
associated with HIV infection and viremia, the role of NK cells in protection against HIV
infection remains to be clarified. Natural resistance to infection among exposed uninfected
individuals may be due partly to efficient early innate antiviral responses. Indeed, we
previously showed that highly exposed uninfected Vietnamese IDUs have higher NK
cytokine/chemokine secretion capacities and lytic activity19. More recently, Montoya et al
reported increased interferon-γ (IFN-γ) production by NK cells from uninfected individuals
sexually exposed to HIV-127.
The present study was designed to further characterize the NK effector potential of
Vietnamese EU IDUs using a recently described degranulation functional assay22 and to
investigate whether increased levels of activation could be associated with peculiar features of
the NK cell receptor repertoire in EUs. In addition to the phenotypic analysis of KIR+ NK cell
subsets, our study is to our knowledge the first to provide a quantitative analysis
discriminating KIR transcripts in virally exposed IDU individuals in reference to low risk
control donors.
Using the CD107a NK functional assay22, we found that EU NK cells exhibit a high
constitutive degranulation potential. Indeed, degranulation occurred rapidly in vitro, without
the need for exogenous target cell stimulation. This high constitutive activity, which likely
reflects a strong in vivo activation status of the NK cell compartment in EUs, was not
observed in HIV+ or in low risk controls recruited in the same environment. In other studies,
high levels of CD107a expression on NK cells from HIV viremic subjects was associated
with a decreased number of CD3-CD56+CD161+ NK cells28. Up-regulation of CD107a has
been shown to correlate with NK cytokine secretion and cytolytic activity22,23,28. This is in
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

17
line with our previous observation of that EU exhibit a higher NK cytolytic potential cells
toward Daudi and K562 targets and an enhanced NK-mediated production of IFN-γ, TNF-α
and CCL3-519. The additional functional read out of NK cells provided in this study, therefore
stresses the fact that EU’s NK cells have a particularly high constitutive potential to display
immediate effector functions upon infectious challenge, that possibly contribute to the
protection of EUs against HIV infection.
The phenotypic analysis of the NK cell repertoire identified particular NK cell receptor
patterns that may sustain constitutive NK cell activation in EUs, such as enhanced CD69 and
CD161 expression. In particular, intensity of CD161 staining on NK cells was higher in EUs
than in HIV+ individuals and controls. Furthermore, following stimulation, CD161 was
preferentially expressed by the activated CD107a+ NK cell subset. Number of CD161+ NK
cells were shown to be reduced in HIV infected viremic subjects29, but no correlation with
NK functional defects observed in HIV+ patients has yet been established. CD161
engagement appears to have a complex role in the modulation of immune functions. Both
activating and inhibitory effects have been described in mice after CD161 receptor
engagement30. In humans, LLT1, an inhibitory ligand of human CD161, was shown to
modulate NK cell activity31,32. CD161 expression was also involved in the triggering of NK
cell cytotoxicity towards the Daudi tumor cell line33. Interestingly, we have previously shown
that EU NK cells display substantial cytotoxic activity against Daudi cells19. Although direct
evidence implicating CD161 in enhanced NK cell activation is still lacking, these
observations suggest that CD161 expression may contribute to the NK cell activation status,
at least in HIV-exposed uninfected subjects.
Although down-regulation of NKp30 was confirmed in HIV+ individuals12, no major
alteration of NCRs was found in EUs in comparison to low-risk controls, thus suggesting that
NK activation in EUs was not associated with increased expression of activating NKp30 or
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

18
NKp46 receptors. Other features of the NK cell repertoire that senses virally induced
modification of HLA class I expression may sustain EU’s NK cell activation. Cell activity
against HIV-1-infected blasts was shown to be partly dependent on NK cell recognition of
HIV-modulated HLA-E expression34. This recognition is mediated by the heterodimeric
receptor CD94, in association with either its inhibitory NKG2A subunit or its activating
NKG2C subunit. In some EUs the stimulatory NKG2C to inhibitory NKG2A ratio was
inverted, in favour of the activating subunit. While no significant change in the relative size of
the CD94+ NK subset was found in EUs, this suggests that HLA-E interaction with NK cells
expressing the CD94/NKG2C activating receptor may be dominant in some EUs. Such a
switch from inhibitory to activating NKG2 subunit expression of the CD94/NKG2 receptor
for HLA-E has been linked to concomitant infections in HIV+ individuals and may be driven
in EUs by exposure to viral infections other than HIV35.
Although the phenotypic distribution of NK cells expressing KIR and CD85j MHC-class I
receptors was not profoundly affected in EU, further discrimination of the activating and
inhibitory KIR genes that contribute to the phenotype of KIR+ NK cell subsets, by locus-
specific real-time PCR transcript quantification, revealed qualitative and/or quantitative
differences between EUs and the other two groups of subjects. In contrast to the other two
study groups, high percentages of GL183-positive NK cells in EUs correlate with the
expansion of KIR2DL3 transcripts in the absence of KIR2DS2 and KIR2DL2 transcripts. In
addition, this expansion of KIR2DL3 transcripts was observed in EUs displaying either HLA-
C1/C1 or C1/C2 allelic specificities. As the inhibitory KIR2DL3 receptor binds to its ligand
HLA-C1 with lower affinity than KIR2DL2, KIR2DL3 expression in the absence of
KIR2DL2 expression may favour NK cell activation in EUs, by a mechanism similar to that
proposed for KIR2DL3-HLA-C1 in the resolution of HCV infection36. Indeed, NK cell
interactions involving KIR2DL3-HLA-C1 have been shown to directly influence the
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

19
resolution of hepatitis C virus (HCV) infection in Caucasians and African Americans36. This
led to the concept that not only activation but also lowered NK inhibitory responses may be
an advantage in the control of viral infection. KIR2DL2/2DL3 heterozygosity in absence of
HLA-C1 ligand has been recently reported to be associated to the seronegative status of
exposed African sex workers20. In our study, all KIR2DL2/2DL3 heterozygous EUs were
homozygous HLA-C1/C1 or heterozygous HLA-C1/C2. Moreover, the two C2/C2 EUs were
typed as homozygous for KIR2DL3. Therefore, although our sample is clearly too limited to
establish any firm statistics on HLA-KIR combinations in EUs, the KIR2DL2/2DL3
heterozygosity in absence of HLA-C1 does not appear to contribute to resistance in
Vietnamese EU IDUs.
Resistance to HIV-1 infection among African sex workers was also associated with KIR3DL1
homozygosity in the absence of its cognate HLA-Bw4 ligand20. No such trend was identified
in Vietnamese EUs, as homozygous HLA-Bw6 and Bw4/Bw6 were equally represented in
KIR3DL1 homozygous EUs (Table 1). Interestingly, in some EUs, concomitant expression of
lowered inhibitory KIR3DL1 transcript levels and high activating KIR3DS1 levels resulted in
a high KIR3DS1/KIR3DL1 ratio that may confer an enhanced activating NK cell repertoire
profile to these EUs. Indeed, very recent data suggest that KIR3DS1 associates with DAP12
and triggers both cytolysis and IFN-γ production37. Previous genetic studies have linked
KIR3DS1 and HLA-Bw4 expression to slow progression toward AIDS18,38. However, in our
study, strongest KIR3DS1 expansion in EUs occurred in HLA-Bw6 individuals rather than in
homozygous HLA-Bw4 EUs who were not found in our study population (Table 1).
Furthermore, it was recently suggested that activating KIR3DS1 receptor does not recognize
HLA-Bw437,39.
Although the number of individuals analyzed was too small to draw statistically powerful
conclusions, combined transcriptional and genomic analysis suggested that KIR3DL1 and
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

20
core KIR2DL4 and KIR3DL2 transcription might be specifically abolished in some HIV-
exposed individuals. Indeed, KIR3DL1 transcripts reflected the KIR3DL1 gene content in
controls, whereas the absence of KIR3DL1 transcripts was observed in presence of the
KIR3DL1 gene in some HIV+ and EU IDUs. Although the possibility that KIR3DL1 gene
polymorphisms specific to HIV-exposed Vietnamese were not detected by our RT-PCR is not
excluded, since PCR primers designed to specifically detect KIR3DL1 transcripts did not
allow their use for gene amplification, this is unlikely considering that KIR3DL1 genotype
was confirmed in these individuals by commercial available kits. Our transcriptional approach
of the KIR repertoire, although limited by the number of individuals studied, thus suggests
that genomic KIR content is not always associated to transcript detection in HIV-exposed
populations, and that mechanisms regulating KIR transcripts may occur in exposed
individuals. Representation of KIR transcripts might not only represent a down- or up-
regulation but a selection at infection. Interestingly, the frequency of individuals that exhibit
KIR2DS4 transcripts encoding the KIR2DS4 receptor was significantly increased in HIV+
IDUs, suggesting that the KIR2DS4 bearing haplotype, often associated to lack of other
activating KIR-S gene content8, might be more prevalent in HIV+ individuals. A challenging
issue to be addressed in wider population studies would be to investigate whether KIR
regulation is selected at infection and to what extent regulatory mechanisms can be
specifically related to HIV or to concomitant co-infections occurring in exposed individuals.
In conclusion, our results and data from other studies of HIV+ patients suggest that the NK
cell receptor repertoires in EUs may be quite different from those of HIV-1-infected
individuals, whether they are viremic or non progressors13,18,28,29,40. Several mechanisms,
including cellular restriction of HIV replication, T cell activity and specific IgA responses
have been proposed to explain resistance to HIV infection41, but it is more likely that complex
genetic and/or selected mechanisms operate independently or in synergy to confer individual
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

21
resistance to HIV-1. HIV-exposed uninfected individuals are a heterogeneous population, and
it is therefore not surprising that the high level of NK cell activation that characterizes the
Vietnamese EUs studied here was associated with diverse NK receptor repertoires. Yet, in
some EUs, no particular pattern of NK receptors could be identified. The constitutive highly
activated status of NK cells from EUs may also be dependent on the HLA-NK receptor
genetic background and/or on selective induction of NK receptors that endow NK cells with
the capacity to mount an effective defence against HIV-1. The increased NK cell activity that
we found in Vietnamese EUs may reflect more general immune activation resulting from
several factors including exposure to multiple viruses42. However, HIV-1-infected IDUs
exposed to similar environmental conditions and to multiple viral infections19,43 have different
NK cell receptor patterns. Therefore, our data strongly suggest that the increased NK cell
activity in virally exposed EUs is linked to particular genetic backgrounds that adapt the
regulation of diverse NK cell receptors upon viral challenge, thereby shaping a constitutively
highly activated NK cell status that may contribute to protection of EUs toward HIV-1
infection.
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

22
Acknowledgments
This work was supported by grants from Agence Nationale de Recherche sur le Sida,
Assistance publique Hôpitaux de Marseille (AP-HM), INSERM (action thématique concertée
ATC Biothérapies), Ministère de la Recherche Réseau Genhomme. Authors' contributions:
PP, FB-S, DS-A and GP conceived the study. PP, DS-A and GP contributed to the
experimental design and coordination of the study. LXT and NN performed the recruitment of
the study subjects and coordinated the study in Vietnam. SR, EB and HKT performed the
transcriptional, phenotypic and functional analyses. IT and TT contributed the HLA and KIR
genotyping. PP, SR, GP and DS-A wrote the manuscript, IT to its critical correction. All
authors read and approved the final manuscript. We thank all the physicians, family
practitioners and medical staff from the Binh Trieu Hospital and the Pasteur Institute of Hô
Chi Minh City who took part in this study. We thank Denis Reviron, Christophe Picard and
Coralie Frassati for their helpful advice in KIR genotyping and members of the Hematology
unit and AP-HM for support and availability of flow cytometer facilities and Françoise
Dignat-George for providing helpful comments on the manuscript.
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

23
References 1. Bottino C, Moretta L, Moretta A. NK cell activating receptors and tumor recognition in humans. Curr Top Microbiol Immunol. 2006;298:175-182. 2. Stewart CA, Vivier E, Colonna M. Strategies of natural killer cell recognition and signaling. Curr Top Microbiol Immunol. 2006;298:1-21. 3. Lodoen MB, Lanier LL. Natural killer cells as an initial defense against pathogens. Curr Opin Immunol. 2006. 4. Lanier LL. Natural killer cells: roundup. Immunol Rev. 2006;214:5-8. 5. Parham P. Taking license with natural killer cell maturation and repertoire development. Immunol Rev. 2006;214:155-160. 6. Yawata M, Yawata N, Draghi M, Little AM, Partheniou F, Parham P. Roles for HLA and KIR polymorphisms in natural killer cell repertoire selection and modulation of effector function. J Exp Med. 2006;203:633-645. 7. Thananchai H, Gillespie G, Martin MP, et al. Cutting Edge: Allele-Specific and Peptide-Dependent Interactions between KIR3DL1 and HLA-A and HLA-B. J Immunol. 2007;178:33-37. 8. Khakoo SI, Carrington M. KIR and disease: a model system or system of models? Immunol Rev. 2006;214:186-201. 9. Anfossi N, Andre P, Guia S, et al. Human NK cell education by inhibitory receptors for MHC class I. Immunity. 2006;25:331-342. 10. Stewart CA, Laugier-Anfossi F, Vely F, et al. Recognition of peptide-MHC class I complexes by activating killer immunoglobulin-like receptors. Proc Natl Acad Sci U S A. 2005;102:13224-13229. 11. Fogli M, Costa P, Murdaca G, et al. Significant NK cell activation associated with decreased cytolytic function in peripheral blood of HIV-1-infected patients. Eur J Immunol. 2004;34:2313-2321. 12. De Maria A, Fogli M, Costa P, et al. The impaired NK cell cytolytic function in viremic HIV-1 infection is associated with a reduced surface expression of natural cytotoxicity receptors (NKp46, NKp30 and NKp44). Eur J Immunol. 2003;33:2410-2418. 13. Mavilio D, Benjamin J, Daucher M, et al. Natural killer cells in HIV-1 infection: dichotomous effects of viremia on inhibitory and activating receptors and their functional correlates. Proc Natl Acad Sci U S A. 2003;100:15011-15016. 14. Weber K, Meyer D, Grosse V, Stoll M, Schmidt RE, Heiken H. Reconstitution of NK cell activity in HIV-1 infected individuals receiving antiretroviral therapy. Immunobiology. 2000;202:172-178. 15. Sondergaard SR, Aladdin H, Ullum H, Gerstoft J, Skinhoj P, Pedersen BK. Immune function and phenotype before and after highly active antiretroviral therapy. J Acquir Immune Defic Syndr. 1999;21:376-383. 16. Sirianni MC, Ensoli F, Alario C, et al. Distribution of the natural killer-related receptor for HLA-C during highly active antiretroviral therapy for human immunodeficiency virus infection. Hum Immunol. 2001;62:1328-1334. 17. Mendila M, Heiken H, Becker S, et al. Immunologic and virologic studies in long-term nonprogressors with HIV-1 infection. Eur J Med Res. 1999;4:417-424. 18. Martin MP, Gao X, Lee JH, et al. Epistatic interaction between KIR3DS1 and HLA-B delays the progression to AIDS. Nat Genet. 2002;31:429-434. 19. Scott-Algara D, Truong LX, Versmisse P, et al. Cutting edge: increased NK cell activity in HIV-1-exposed but uninfected Vietnamese intravascular drug users. J Immunol. 2003;171:5663-5667.
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

24
20. Jennes W, Verheyden S, Demanet C, et al. Cutting Edge: Resistance to HIV-1 Infection among African Female Sex Workers Is Associated with Inhibitory KIR in the Absence of Their HLA Ligands. J Immunol. 2006;177:6588-6592. 21. Truong LX, Luong TT, Scott-Algara D, et al. CD4 cell and CD8 cell-mediated resistance to HIV-1 infection in exposed uninfected intravascular drug users in Vietnam. Aids. 2003;17:1425-1434. 22. Alter G, Malenfant JM, Altfeld M. CD107a as a functional marker for the identification of natural killer cell activity. J Immunol Methods. 2004;294:15-22. 23. Uhrberg M. The CD107 mobilization assay: viable isolation and immunotherapeutic potential of tumor-cytolytic NK cells. Leukemia. 2005;19:707-709. 24. Pascal V, Schleinitz N, Brunet C, et al. Comparative analysis of NK cell subset distribution in normal and lymphoproliferative disease of granular lymphocyte conditions. Eur J Immunol. 2004;34:2930-2940. 25. Toneva M, Lepage V, Lafay G, et al. Genomic diversity of natural killer cell receptor genes in three populations. Tissue Antigens. 2001;57:358-362. 26. Hsu KC, Liu XR, Selvakumar A, Mickelson E, O'Reilly RJ, Dupont B. Killer Ig-like receptor haplotype analysis by gene content: evidence for genomic diversity with a minimum of six basic framework haplotypes, each with multiple subsets. J Immunol. 2002;169:5118-5129. 27. Montoya CJ, Velilla PA, Chougnet C, Landay AL, Rugeles MT. Increased IFN-gamma production by NK and CD3+/CD56+ cells in sexually HIV-1-exposed but uninfected individuals. Clin Immunol. 2006. 28. Alter G, Malenfant JM, Delabre RM, et al. Increased natural killer cell activity in viremic HIV-1 infection. J Immunol. 2004;173:5305-5311. 29. Jacobs R, Weber K, Wendt K, Heiken H, Schmidt RE. Altered coexpression of lectin-like receptors CD94 and CD161 on NK and T cells in HIV patients. J Clin Immunol. 2004;24:281-286. 30. Ljutic B, Carlyle JR, Filipp D, Nakagawa R, Julius M, Zuniga-Pflucker JC. Functional requirements for signaling through the stimulatory and inhibitory mouse NKR-P1 (CD161) NK cell receptors. J Immunol. 2005;174:4789-4796. 31. Rosen DB, Bettadapura J, Alsharifi M, Mathew PA, Warren HS, Lanier LL. Cutting Edge: Lectin-Like Transcript-1 Is a Ligand for the Inhibitory Human NKR-P1A Receptor. J Immunol. 2005;175:7796-7799. 32. Aldemir H, Prod'homme V, Dumaurier MJ, et al. Cutting edge: lectin-like transcript 1 is a ligand for the CD161 receptor. J Immunol. 2005;175:7791-7795. 33. Azzoni L, Zatsepina O, Abebe B, Bennett IM, Kanakaraj P, Perussia B. Differential transcriptional regulation of CD161 and a novel gene, 197/15a, by IL-2, IL-15, and IL-12 in NK and T cells. J Immunol. 1998;161:3493-3500. 34. Bonaparte MI, Barker E. Killing of human immunodeficiency virus-infected primary T-cell blasts by autologous natural killer cells is dependent on the ability of the virus to alter the expression of major histocompatibility complex class I molecules. Blood. 2004;104:2087-2094. 35. Mela CM, Burton CT, Imami N, et al. Switch from inhibitory to activating NKG2 receptor expression in HIV-1 infection: lack of reversion with highly active antiretroviral therapy. Aids. 2005;19:1761-1769. 36. Khakoo SI, Thio CL, Martin MP, et al. HLA and NK cell inhibitory receptor genes in resolving hepatitis C virus infection. Science. 2004;305:872-874. 37. Carr WH, Rosen DB, Arase H, Nixon DF, Michaelsson J, Lanier LL. Cutting Edge: KIR3DS1, a Gene Implicated in Resistance to Progression to AIDS, Encodes a DAP12-
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

25
Associated Receptor Expressed on NK Cells That Triggers NK Cell Activation. J Immunol. 2007;178:647-651. 38. Gaudieri S, DeSantis D, McKinnon E, et al. Killer immunoglobulin-like receptors and HLA act both independently and synergistically to modify HIV disease progression. Genes Immun. 2005;6:683-690. 39. O'Connor G M, Guinan KJ, Cunningham RT, Middleton D, Parham P, Gardiner CM. Functional Polymorphism of the KIR3DL1/S1 Receptor on Human NK Cells. J Immunol. 2007;178:235-241. 40. Andre P, Brunet C, Guia S, et al. Differential regulation of killer cell Ig-like receptors and CD94 lectin-like dimers on NK and T lymphocytes from HIV-1-infected individuals. Eur J Immunol. 1999;29:1076-1085. 41. Fauci AS, Mavilio D, Kottilil S. NK cells in HIV infection: paradigm for protection or targets for ambush. Nat Rev Immunol. 2005;5:835-843. 42. Tran HK, Chartier L, Troung LX, et al. Systemic immune activation in HIV-1-exposed uninfected Vietnamese intravascular drug users. AIDS Res Hum Retroviruses. 2006;22:255-261. 43. Follezou JY, Lan NY, Lien TX, et al. Clinical and biological characteristics of human immunodeficiency virus-infected and uninfected intravascular drug users in Ho Chi Minh City, Vietnam. Am J Trop Med Hyg. 1999;61:420-424.
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

26
Table 1: Genomic, transcriptional and phenotypic analysis of the KIR repertoire in EUs. HLA-B and -C typing was performed and grouped as HLA-C1 (Asn80), HLA-C2 (Lys80) and Bw4/Bw6 groups, according to KIR recognition pattern. Real-time PCR evaluation of each KIR transcript levels was performed in 25 EUs and associated to KIR genotyping in 14 EUs When KIR transcripts were detected (+), the ratio between activating and inhibitory counterparts was indicated. Phenotypic expression of KIR receptors is indicated as the percentage of KIR+ cells among CD3-CD56+ NK cells using EB6+, GL183+, Z27.3.7+, and FES172 mAbs. Data from exposed uninfected individuals (EU) were analyzed in reference to median values and 25th-75th percentiles in low-risk Vietnamese unexposed control blood donors (C) and HIV-infected individuals (HIV, lower panel)
Genotype 2DS1/2DL1
Transcripts 2DS1/2DL1 ratio
Phenotype % EB6+
Genotype 2DS2/2DL2/2DL3
Transcripts 2DS2/2DL2/2DL3
Phenotype % GL183+
Genotype 3DS1/3DL1
Transcripts 3DS1/3DL1 ratio
Phenotype % Z27.3.7+
Genotype 2DS4
Transcripts 2DS4/1D
Phenotype % FES172+
1 C1 Bw4 Bw6 -/+ -/+ 15 -/-/+ -/-/+ 24 -/+ -/+ 30 + +/- 532 C1 Bw4 Bw6 -/+ -/+ 21 -/-/+ -/-/+ 57 -/+ -/+ 7 + +/- 813 C1 Bw4 Bw6 nd -/+ nd nd -/-/+ nd nd -/+ nd nd +/- nd4 C1 Bw4 Bw6 -/+ -/+ 6 +/+/+ +/+/+ 63 -/+ -/+ 13 + -/+ 385 C1 Bw6 -/+ -/+ 32 -/-/+ -/-/+ 68 -/+ -/- 0 + -/- 06 C1 Bw6 +/+ +/+ 2.8 29 -/-/+ -/-/+ 26 +/+ +/+ 28 6 + -/+ 07 C1 Bw6 +/+ +/+ 61 4 +/+/+ +/+/+ nd +/+ +/+ 59 1 + -/+ 08 C1 Bw6 nd +/+ 0.9 8 nd +/+/+ 33 nd -/+ 22 nd +/+ 459 C1 Bw6 nd +/+ 3.7 nd nd -/-/+ nd nd +/+ 1.3 nd nd -/+ nd
10 C1 C2 Bw4 Bw6 +/+ +/- 13 +/+/+ +/+/+ 67 +/+ +/- 6 + +/+ 2411 C1 C2 Bw4 Bw6 nd +/+ 4.0 50 nd -/-/+ 54 nd +/+ 3.0 27 nd +/+ 4412 C1 C2 Bw4 Bw6 +/+ +/+ 1.0 83 +/+/+ +/+/+ 6 +/+ +/+ 1.9 6 + +/- 3713 C1 C2 Bw4 Bw6 +/+ +/+ 2.3 21 -/-/+ -/-/+ 36 +/+ +/+ 0.9 26 + -/+ 014 C1 C2 Bw4 Bw6 nd -/+ nd nd -/-/+ nd nd -/- nd nd +/+ nd15 C1 C2 Bw6 -/+ -/- 10 +/+/+ -/+/+ 41 +/+ +/- nd + +/+ 1616 C1 C2 Bw6 +/+ +/+ 1.0 38 -/-/+ -/-/+ 32 +/+ +/+ 28 8 + -/+ 017 C1 C2 Bw6 -/+ -/+ 9 +/+/+ +/+/+ 12 -/+ -/+ 5 + +/- 3018 C1 C2 Bw6 +/+ +/+ 0.1 47 -/-/+ -/-/+ 63 -/+ -/+ 17 + +/+ 2219 C2 Bw4 Bw6 nd -/+ nd nd -/-/+ 31 nd -/+ nd nd +/- nd20 C2 Bw6 -/+ -/+ nd -/-/+ -/-/+ nd -/+ -/+ nd + -/+ nd21 nd nd nd +/+ 1.0 25 nd -/-/+ 54 nd +/+ 8.1 10 nd +/- 4022 nd nd nd +/+ 3.45 31 nd +/+/+ 46 nd +/+ 3.8 21 nd +/- 4523 nd nd nd -/+ 5 nd +/+/+ 19 nd -/+ 1 nd -/+ 024 nd nd nd +/+ 47 20 nd +/+/+ 40 nd +/ni 8 nd -/- nd25 nd nd nd -/+ nd nd -/-/+ 30 nd -/+ nd nd -/+ nd
HIV - - -1.4
(1.4-1.4)11.3
(7.6-22.5) - -
28.4 (19.5-44.1)
-2.1
(2.1-2.1)11.3
(6.3-15.7) - -
33.7 (18.1-37.7)
EU - - -1.7
(0.5-3.6)15
(9-31) - -
36 (23-56.7)
-4.7
(1.5-28.0)8.6
(6.9-17.5) - -
30.5 (20.9-42.0)
C - - -4.1
(3.1-4.8)14.5
(7-26) - -
35.2 (20-48.3)
-3.0
(2.0-10.0)12
(8.8-21.5) - -
32.6 (21.7-65.0)
KIR2DS2/2DL2/2DL3 KIR3DS1/KIR3DL1 KIR2DS4EU HLA-C HLA-B
KIR2DS1/KIR2DL1
F
or personal use only. by guest on D
ecember 30, 2011.
bloodjournal.hematologylibrary.org
From

Figure Legends
Figure 1: Genomic and real time PCR transcriptional analysis of the KIR repertoire.
A. The size of the NK cell compartment was analyzed by flow cytometry (% CD3-
CD56+CD16+ cells) in exposed HIV-infected IDU (HIV �) exposed HIV-uninfected IDU
(EU �) and unexposed control blood donors (C �).
B. The frequency of KIR transcript detection was analyzed in 17 HIV-infected, 25 EUs and 21
unexposed controls. Analysis of genomic KIR frequencies was conducted on a subgroup of 12
unexposed controls (and compared to genomic KIR frequencies reported by Toneva et al in
59 Vietnamese subjects25). HIV+ individuals show lower KIR transcript frequencies for
KIR3DS1 (p<0.10) and KIR3DL1 (p<0.001). Genomic analysis confirmed the weak KIR3DS1
frequency in HIV+ (18%). Dissociation between KIR3DL1 gene and transcript detection was
detected in 3 out of 14 EU tested (21.4%) and 3 out of 8 HIV (37.5%) individuals, but was
never observed in tested controls (n=12). Expressed KIR2DS4 transcript frequencies refer to
transcription alleles encoding the expressed KIR2DS4 receptor (exclusive of individuals only
expressing the KIR1D null allele). Level of significance is indicated as follow: * p<0.05, **
p<0.01, *** p<0.001.
C. Real time PCR analysis of altered KIR transcript patterns in HIV and EUs. KIR2DL4,
KIR3DL2, KIR2DL3 and KIR3DL1 mRNA transcript levels are represented as relative copy
numbers per 106 GAPDH transcripts. KIR2DL3 transcript levels where examined according to
the alternative KIR2DL3+KIR2DS2-KIR2DL2- or KIR2DL3+KIR2DS2+KIR2DL2+ genomic
gene combinations (middle left) and also plotted according to the size of the phenotypically
defined GL183+ NK cell subset (middle right). High percentage of GL183+ NK cells (> 50%
of GL183+ NK cells within CD3-CD56+ NK cells) identified a specific subgroup of
KIR2DL3+KIR2DS2-KIR2DL2- EU individuals who exhibit higher KIR2DL3 transcript levels
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

in the absence of KIR2DS2/KIR2DL2 transcription. Activating KIR3DS1/inhibitory KIR3DL1
transcript ratio is represented (bottom right).
Figure 2: Flow cytometer analysis of NKR expression on CD3-CD56+ NK cells.
A. The size of NKR+ NK cell subsets expressing NKp30, NKp46, CD161, NKG2A and
NKG2C, CD69, CD85j were evaluated after gating of positive CD3-CD56+ NK cells in
exposed HIV-infected IDU (HIV �), exposed HIV-uninfected IDU (EU �), and unexposed
control blood donors (C �, 25th percentile, median, 75th percentile). NKp30 and NKp46 were
expressed on all NK cells (100% CD3-CD56+), and comparison of expression levels where
thus analysed by analysis of mean fluorescence intensity (mfi). NK cell surface expression of
NKG2A inhibitory subunit of the CD94/NKG2A heterodimeric receptor was inversely
correlated to that of the stimulatory NKG2C subunit among CD3-CD56+ NK cells in EU
subjects (Spearman’s correlation test), while correlation was non significant in C and HIV
groups.
B. Representative flow cytometer analyses of NKR staining in exposed HIV-infected IDU
(HIV), exposed uninfected IDU (EU) and unexposed controls (C) (white: isotypic control
staining, black, NKR specific staining).
Figure 3: Constitutive and inducible activation of NK cells is revealed by the CD107
degranulation assay.
A. Comparative analysis of CD107a expression on CD3-CD56+ NK cells was performed in
the three study groups, in the absence of stimulation (unstim) and upon K562 target cells
stimulation (K562). Boxes represent CD3-CD56+ cells expressing CD107a in exposed HIV-
infected IDU (HIV), exposed HIV-uninfected IDU (EU), and unexposed control donors (C), y
axis refer to 25th percentile, median, 75th percentile values.
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

B. Comparative analysis of the mean fluorescence intensity (mfi) of CD107a staining among
CD107+CD3-CD56+ NK cells in exposed HIV-infected IDU (HIV), exposed uninfected IDU
(EU) and controls (C) (white: isotypic control staining, black, NKR specific staining). In the
absence of stimulation (unstim) and upon stimulation with K562 target cells (K562). Boxes
represent the median mfi of CD107a staining among CD3-CD56+ NK cells (25th percentile,
median, 75th percentile).
C. Examples of flow cytometer analyses of CD107a staining on CD3-CD56+ NK cells in the
absence of stimulation (white: isotypic control staining, black, NKR specific staining).
Figure 4: Analysis of CD161 expression by the activated CD107a+CD3-CD56+ NK cell
subset.
A. Cell surface expression of CD161 was evaluated by four-color flow cytometry after gating
on CD107a+CD3-CD56+ NK cells. The percentage of CD161+ stained cells was analyzed
within CD107a+CD3-CD56+ NK cells in absence of stimulation (unstim) and after 3 hours of
K562 cell stimulation (K562) in exposed HIV-infected IDUs (HIV), exposed uninfected IDUs
(EU) and unexposed controls (C). Y-axis values boxed represent 25th percentile, median and
75th percentile.
B. Representative flow cytometer analysis of % and mean fluorescence intensity (mfi) of
CD161 staining among the CD107a+CD3-CD56+ NK cell subset before exogenous stimulation
(white: isotypic control staining, black, CD161 specific staining).
C. Mean fluorescence intensity (mfi) of CD161 staining among the CD161+ unstimulated
CD107a+CD3-CD56+ NK cell subset. Y-axis values boxed represent 25th percentile, median
and 75th percentile.
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

D. Evaluation of mean fluorescence intensity of CD161 expression within the CD107apositive
and CD107anegative CD3-CD56+NK cell subsets after K562 cell stimulation. Y-axis values
boxed represent 25th percentile, median and 75th percentile.
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom
HIV EU C0
10
20
30
40
50p<0.05
p<0.05
HIV EU C100
1000
10000
100000p<0.05
p<0.05
HIV EU C100
1000
10000
100000
1000000p<0.05
p<0.05
50000
150000
250000
350000
450000
550000p<0.05
2DS2/2DL2/2DL3Genotype
--+ +++HIV
--+ +++EU
--+ +++C
p<0.05 p<0.05
A B
C
0 10 20 30 40 50 60 70 801000
10000
100000
1000000
HIVEU
C
EU 2DL3+ only(2DS2-2DL2-)
(CD158b1/b2/j) cell surface phenotype% GL183+ on CD3-CD56+NK Cells
HIV EU C100
1000
10000
100000
HIV EU C0
10
20
30
40
50
60
For personal use only. by guest on December 30, 2011. bloodjournal.hematologylibrary.orgFrom

B
84%4.24 mfi
NKp30
NKp46
HIV EU C
95%6.95mfi
38%1.55 mfi
mfi0.47
mfi1.29
mfi1.50
mfi0.70
mfi1.31
mfi0.61
NKG2A
NKG2C
30%4.25 mfi
17%8.0 mfi
65%8.4mfi
10%2.08 mfi
50%2.61mfi
15%2.48 mfi
CD69
101 102 10100 101 102 10100101 102 103100 101 102 103100101 102 103100 101 102 103100
CD85j
42%2.84 mfi
41%2.16mfi
32 %1.96 mfi
10%2.26 mfi
18%1.72mfi
1.4%1.71 mfi
CD161HIV EU C0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
p<0.05
p<0.05
HIV EU C0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
HIV EU C0.00.51.01.52.02.53.03.54.04.55.0 p<0.05
HIV EU C0
102030405060708090
100
p<0.05
p<0.05
HIV EU C0
102030405060708090
100
HIV EU C0
102030405060708090
100
HIV EU C0
102030405060708090
100
0 10 20 30 40 50 60 70 80 90 1000
10
20
30
40
50
60
70
80
90
100
p<0.05r = - 0.59
EU
% NKG2A+
on CD3-CD56+ cells
A
F
or personal use only. by guest on D
ecember 30, 2011.
bloodjournal.hematologylibrary.org
From

unstim K562 unstim K562 unstim K562 0
10
20
30
40
50
60
70
p<0.001
p<0.001p<0.01 p<0.05
HIV CEU
p<0.01
unstim K562 unstim K562 unstim K5620
2
4
6
8
10
12
14
16
18
20
HIV CEU
p<0.05
p<0.01
p<0.05
p<0.05A B
C
F
or personal use only. by guest on D
ecember 30, 2011.
bloodjournal.hematologylibrary.org
From

HIV EU C0
2
4
6
8
10
12p<0.05
p<0.05
unstim K562 unstim K562 unstim K5620
10
20
30
40
50
60
70
80
90
100p<0.05p<0.01
p<0.01
HIV CEU
CD107a + CD107a - CD107a + CD107a - CD107a + CD107a -0
2
4
6
8
10
12 p<0.001
p<0.01
HIV CEU
p<0.05
A B
C D
F
or personal use only. by guest on D
ecember 30, 2011.
bloodjournal.hematologylibrary.org
From
Top Related
Copyright © 2022 FDOKUMEN

