







Bahasa
Halaman
Hukum


J. Anat.
(2007)
210
, pp41–53 doi: 10.1111/j.1469-7580.2006.00662.x
© 2006 The Authors Journal compilation © 2006 Anatomical Society of Great Britain and Ireland
Blackwell Publishing Ltd
Assessing mechanical function of the zygomatic region in macaques: validation and sensitivity testing of finite element models
K. Kupczik,
1
C. A. Dobson,
2
M. J. Fagan,
2
R. H. Crompton,
3
C. E. Oxnard
1,4
and P. O’Higgins
1
1
Hull York Medical School, The University of York, UK
2
Centre for Medical Engineering and Technology, Department of Engineering, University of Hull, UK
3
Department of Human Anatomy & Cell Biology, University of Liverpool, UK
4
School of Anatomy and Human Biology, The University of Western Australia, Australia
Abstract
Crucial to the interpretation of the results of any finite element analysis of a skeletal system is a test of the validity
of the results and an assessment of the sensitivity of the model parameters. We have therefore developed finite
element models of two crania of
Macaca fascicularis
and investigated their sensitivity to variations in bone material
properties, the zygomatico-temporal suture and the loading regimen applied to the zygomatic arch. Maximum
principal strains were validated against data derived from
ex vivo
strain gauge experiments using non-physiological
loads applied to the macaque zygomatic arch. Elastic properties of the zygomatic arch bone and the zygomatico-
temporal suture obtained by nanoindentation resulted in a high degree of congruence between experimental and
simulated strains. The findings also indicated that the presence of a zygomatico-temporal suture in the model pro-
duced strains more similar to experimental values than a completely separated or fused arch. Strains were distinctly
higher when the load was applied through the modelled superficial masseter compared with loading an array of
nodes on the arch. This study demonstrates the importance of the accurate selection of the material properties
involved in predicting strains in a finite element model. Furthermore, our findings strongly highlight the influence
of the presence of craniofacial sutures on strains experienced in the face. This has implications when investigating
craniofacial growth and masticatory function but should generally be taken into account in functional analyses of
the craniofacial system of both extant and extinct species.
Key words
cranial sutures; elastic properties; experimental strain analysis; finite element analysis; primates; sensi-
tivity; skull; validation.
Introduction
Here we present a validation study of finite element
models (FEMs) of two macaque crania and assess the
sensitivity of these models to elastic properties of bone,
the presence or absence of a suture and the mode of
load application.
A major prerequisite of the study of the evolution of
the primate craniofacial complex is to develop an
understanding of how mechanical factors influence
facial growth and adult morphology (e.g. Moss, 1973;
Oyen et al. 1979; Dechow & Carlson, 1990). This issue
can be approached in several ways, but that adopted in
this paper is to develop FEMs of macaque faces at
different stages during growth and to explore how
stresses and strains experienced during static loading
are related to bone modelling. Finite element analysis
(FEA) is increasingly being used to test hypotheses per-
taining to the functional morphology of the primate
craniodental system (Spears & Crompton, 1996; Daegling
& Hylander, 1997, 2000; Chen & Chen, 1998; Spears &
Macho, 1998; Macho & Spears, 1999; McConnell &
Crompton, 2001; Witzel & Preuschoft, 2002; Preuschoft
& Witzel, 2004; Witzel et al. 2004; Macho et al. 2005;
Correspondence
Dr Kornelius Kupczik, Department of Human Evolution, Max Planck Institute for Evolutionary Anthropology, Deutscher Platz 6, 04103 Leipzig, Germany. T: +49 (341) 3550 756; F: +49 (341) 3550 399; E: [email protected]
Accepted for publication
18 September 2006

FEM of the macaque zygomatic region, K. Kupczik et al.
© 2006 The AuthorsJournal compilation © 2006 Anatomical Society of Great Britain and Ireland
42
Marinescu et al. 2005; Richmond et al. 2005; Ross et al.
2005; Strait et al. 2005). The reliability of models ought
to be tested against real-world data before they can be
employed confidently (Richmond et al. 2005). Several
authors have recognized this need; thus, Fagan et al.
(2002) carried out sensitivity studies of FEMs of inter-
vertebral discs, and Sellers & Crompton (2004) and
Wang et al. (2004) undertook both validation and
sensitivity studies to dynamic models of the masticatory
and locomotor systems. Those cranial FEA studies that
do take the need for verification and sensitivity into
account have done so, for example, by comparing the
FEM with known muscle physiological data (Ross et al.
2005) or by referring to material properties of bone
obtained by mechanical testing (Strait et al. 2005).
Additionally,
in vitro
experimental strain analyses have
enabled validation of FEMs (e.g. Marinescu et al. 2005).
The latter approach offers the particular advantage of
a better control of the loads and boundary conditions,
model geometry and elastic properties of the materials
defined in the model. Moreover, the strain gauge loca-
tions in the experimental specimens can be precisely
recorded, allowing an accurate comparison between
experimental and simulated strain results.
In this study we aim to validate and investigate the
sensitivity of FEMs of two crania of the crab-eating
macaque
Macaca fascicularis
focusing on the infraorbital
region and the zygomatic arch. We are particularly
concerned with the effects of (1) the variation of bone
material properties, (2) the complexity of the morphology
involved (i.e. the presence or absence of representation
of morphological features such as sutures in the FEM)
and (3) the nature of loading of the zygomatic arch on
the predicted strain magnitudes observed in the FEM
both locally within the zygomatic arch and the infra-
orbital region and globally throughout the cranium. The
results of the FEA are validated against data derived from
ex vivo
strain gauge experiments using non-physiological
loads applied to the macaque zygomatic arch.
It has been observed that the specification of the
material properties (i.e. magnitude, direction, spatial
variation) in the model has significant implications on
the results of an FEA (Marinescu et al. 2005; Richmond
et al. 2005; Strait et al. 2005). Although solving an FEM
with heterogeneous, orthotropic elastic properties pre-
dicts the most congruent strain results when compared
with experimental data (Marinescu et al. 2005; Strait
et al. 2005), for the sake of simplicity the materials
involved in the present FEA are modelled with homo-
geneous, isotropic and linear elastic properties. In order
to enable the validation of the FEA strain results, elastic
properties of selected regions in the bone of the zygo-
matic arch were obtained using a nanoindention tech-
nique (see below). Moreover, the effect of changes in
bone stiffness on the distribution of strains in the zygo-
matic arch and the circumorbital region are investigated
by sequential alteration in the FEA of the experiment-
ally determined Young’s modulus of elasticity (
E
, GPa)
of bone and these effects are compared with our strain
gauge data.
The zygomatic arch has been chosen as it represents
a discrete morphological region of the cranium of
importance in terms of investigating craniofacial bio-
logy and as a key component in the FEM (e.g. Hylander
& Johnson, 1997; Rafferty et al. 2000; Witzel et al.
2004). The attachment areas of the superficial and
deep heads of the masseter muscle are easily identifiable
and the arrangement and orientation of the fibres
makes this muscle easier to model than, for instance,
the temporalis muscle – attaching as it does to the
curved parietal bone. Moreover, the presence of the
zygomatico-temporal suture is of interest from both a
functional and a developmental point of view (Herring,
1972; Jaslow, 1990; Hylander & Johnson, 1997; Mao,
2002; Mao et al. 2003; Rafferty et al. 2003; Rayfield,
2005).
In vivo
strain gauge experiments have shown
that the zygomatic arch of
M. fascicularis
experiences a
strain gradient, the largest strains occurring at the
anterior end of the arch (Hylander & Johnson, 1997). As
the superficial masseter muscle attaches along the
anterior portion of the zygomatic arch anterior to the
zygomatico-temporal suture, the force will be concen-
trated anteriorly and hence strains are expected to be
larger in this region.
The zygomatic arch resembles to some extent a beam
with fixed ends, to which a uniform but off-centre load
has been applied (Hylander & Johnson, 1997). Under
this model bending moments are consequently largest
in the anterior portion of the arch, resulting in relatively
larger strains in this region and lower strains towards
the posterior arch (Hylander & Johnson, 1997). How-
ever, as these authors also noted such a beam model
would not take into account the zygomatico-temporal
suture connecting the zygomatic processes of the zygo-
matic and temporal bones. It is known from research
on miniature pigs that flexible cranial sutures are sub-
jected to large deformations and limit the strains that
can develop in the delicate bones of the face during

FEM of the macaque zygomatic region, K. Kupczik et al.
© 2006 The Authors Journal compilation © 2006 Anatomical Society of Great Britain and Ireland
43
dynamic loading (Herring & Teng, 2000; Rafferty et al.
2003). Flexible sutures are also said to act as shock-
absorbers, being able to absorb more impact energy
than bone (Buckland-Wright, 1978; Jaslow, 1990). More-
over, Sun et al. (2004) showed that the magnitude and
polarity of strains in some cranial sutures of pigs
change with increasing age. This is particularly relevant
to solving FEMs of juvenile vs. adult macaques. In terms
of bending moments and induced stresses and strains
in the zygomatic arch our model is like a beam with a
built-in (cantilevered) constraint at one end and a flex-
ible joint at the other. In immature individuals with a
patent suture the bending moment of the anterior
zygomatic arch will be higher than in older individuals
with more mineralized and thus stiffer sutures. It is thus
hypothesized that strains in the anterior zygomatic
arch in particular decrease with gradual obliteration of
the suture. This is tested by comparing experimental
strain values with those computed from FEMs with
zygomatic arches that are (1) disconnected, (2) fully
fused and (3) connected by a zygomatico-temporal
suture of varying elastic properties.
A further issue to be addressed here is the modelling
of the muscle forces involved. Commonly, force vectors
of a certain magnitude and direction are simply
applied to the surface nodes of the FEM. The selection
of these nodes should reflect the attachment site and
the distribution and the concentration of the muscle
fibres (Richmond et al. 2005; Ross et al. 2005). One
question of interest is how the strain field is influenced
by the distribution of the applied forces. It can be
hypothesized that the more focussed, centralised appli-
cation of loads, the more complex and potentially larger
the loading and induced stresses and strains experienced
by the bone will be. We investigate this by conducting
FEAs with varying arrangements of distributed load
while keeping the total force applied to the zygomatic
arch constant. An alternative approach is to incorpor-
ate the muscle itself into the FEM and load the zygo-
matic arch indirectly via the modelled muscle. This has
the advantage of capturing the exact geometry of the
muscle and might be expected to result in predicted
strains that are more congruent with experimental data.
Material and methods
We selected whole cadaveric heads of two adult male
M. fascicularis
for this study. The study animals (referred
to here as MAC-14 and MAC-17) had been used previ-
ously in dental caries experiments unrelated to the
present study (Smith & Beighton, 1986, 1987). The exact
history of preservation of the cadaveric heads is un-
certain, although it is known that they have been kept
in formaldehyde, 70% industrial methylated spirit and
other aqueous solutions at various times. MAC-14 is a
relatively small specimen with the third molars congenit-
ally missing, while MAC-17 is a large adult with a com-
plete dentition. The zygomatico-temporal suture is
clearly defined and unfused in MAC-14, whereas in the
more mature MAC-17 specimen the suture is less visible
and appears more mineralised.
Experimental strain analysis
In both specimens the skin and the underlying soft tissue
were removed to reveal the bone and the masticatory
muscles. The superficial masseter was dissected on one
side of each of the heads (MAC-14: left-hand side;
MAC-17: right-hand side) and detached from its inser-
tion site on the mandible sufficiently to permit in-
dependence when loads were applied to it. Dehydration
was minimized by applying a glycerine/water solution
onto the soft tissue and bone and by keeping the speci-
mens in a sealed container in a fridge when not under
experimental employment. Prior to the strain gauging
experiments the exposed bone was degreased with
isopropanol (M-Line GC-6 Degreaser, Vishay Measure-
ments Group, Basingstoke, UK) and abraded with pumice
powder. Six 120
±
0.5-
Ω
rosette strain gauges wired in
a three-wire quarter-bridge circuit (TML FRA-1–11,
Tokyo Sokki Kenkyujo, Tokyo, Japan) were bonded with
cyanoacrylate along the zygomatic arch of each speci-
men, from the infra-orbital region to the posterior
zygomatic arch (gauges IO and ZA1–ZA5), three gauges
being placed anterior to the suture and two posterior
to it (Fig. 1). The strain gauges were connected to a
Vishay 5100B Wheatstone bridge amplifier with an
excitation voltage of 0.5 V (Vishay Micro-Measurements).
The heads were then placed on a specially constructed
test rig and constrained by means of a rubber plug
fixing the foramen magnum. The second molars and
the first molars of MAC-17 and MAC-14, respectively,
rested on a rubber-coated bar. Each of the dissected
superficial masseters (working side) and the gonion
region of the mandible on the balancing side were per-
forated to allow passage and attachment of nylon-coated
strings (1 mm diameter, Berkley Outdoor Technologies
Group, Spirit Lake, IA, USA). We conducted the loading

FEM of the macaque zygomatic region, K. Kupczik et al.
© 2006 The AuthorsJournal compilation © 2006 Anatomical Society of Great Britain and Ireland
44
via the strings in line with the orientation of the super-
ficial masseters, by manually placing weights at the ends
of the strings up to a maximum of 15 N for the smaller
MAC-14 specimen and 20 N for the MAC-17 specimen.
Simultaneously, in each of the specimens an equal load
was applied to the string on the balancing side jaw also
in line with the orientation of the superficial masseter.
We recorded the orientation of the working side mas-
seter by taking three-dimensional (3D) coordinates with
a MicroScribe GTX digitizing system (Immersion Corpo-
ration, San Jose, CA, USA) in order to derive the force
vectors for the FEA. Maximum principal strains (expressed
as microstrain
µε
) under static loads were recorded as
the strain most readily comparable with the FE results.
Five consecutive strain readings at the strain peak were
taken per loading, and loadings were repeated four
times within 15 min of the experiment to minimize the
effects of dehydration and atmospheric change.
Nanoindentation of zygomatic arch cortical bone and
zygomatico-temporal suture
Although the fixation history of the specimens is
unknown it is likely to have had an effect on mechanical
properties. Thus, to establish the elastic properties of
the cortical bone and the zygomatico-temporal suture
of the fixed specimens we investigated the mechanical
properties of the bone using an instrumented nano-
indenter. Following the strain gauge experiments, the
zygomatic arch from the balancing side of each specimen
was cut both at the anterior (zygomatic) end and at the
posterior (temporal) end of the arch. Subsequently, the
isolated bones were sectioned with an Isomet 5000
linear precision saw (Buehler Ltd, Lake Bluff, IL, USA) at
two locations along the arch of MAC-14 (anteriorly and
through the zygomatico-temporal suture) and three
locations along the arch of MAC-17 (anteriorly, through
the zygomatico-temporal suture and posteriorly). The
three samples were then embedded in epoxy cold-
mounting resin (Epothin, Buehler Ltd) before the
surfaces were ground (Phoenix Beta Vector, Buehler Ltd)
and polished (ChemoMat synthetic polishing pad and
Mastermet silica polishing suspension, Buehler Ltd).
Using a nano-hardness tester with a Berkovitch diamond
indenter (CSM Instruments S.A., Peseux, Switzerland),
we measured the Young’s elastic modulus at 16 loca-
tions along the perimeter of each cross-sectional sample,
using a load of 100 mN applied for 5 s. In addition, four
nanoindentation measurements were taken through
the zygomatico-temporal suture of the two specimens.
The mean of the
E
of bone and suture for each of the
sections of MAC-14 and MAC-17 are summarized in
Table 1. The grand mean for bone is 9.1
±
2.8 GPa
(MAC-14) and 11.0
±
3.0 GPa (MAC-17), while it is
1.9
±
1.7 and 7.7
±
3.4 GPa for the suture of MAC-14
and MAC-17, respectively.
CT scanning and 3D reconstruction
Coronal computed tomography (CT) scans were taken
of MAC-14 on a Philips Mx8000 scanner (helical mode,
Fig. 1 Three-dimensional reconstructions and finite element mesh of (A) MAC-14 with zygomatico-temporal suture (dashed line) and (B) MAC-17 with right superficial masseter muscle (dark grey). Nodes highlighted correspond to strain gauge locations on the zygomatic arch and the infraorbital region in the specimens used in the loading experiments (three nodes represent one location; see text for details). Numbers refer to strain gauge positions (1 = infraorbital region; 2–6 = zygomatic arch locations). Open arrows indicate load point on M1 (MAC-14) and M2 (MAC-17); thin arrows indicate line of action of muscle loaded.
Table 1 Mean, standard deviation (SD) and maximum and minimum Young’s elastic moduli (in GPa) for cortical bone of the zygomatic arch (MAC-14: two sections, MAC-17: three sections) and for the zygomatico-temporal suture in MAC-14 and MAC-17. The values are based on 16 nanoindentation measurements for each of the bone sections (anterior, suture and posterior) and four and five for the suture tissue of MAC-14 and MAC-17, respectively
Anterior section
Suture section
Posterior section
Grand mean
Zygomatico-temporal suture
MAC-14Mean 9.8 8.3 – 9.1 1.9SD 2.9 2.7 – 2.8 1.7Min. 4.0 1.0 – 0.8Max. 13.3 12.0 – 4.5
MAC-17Mean 11.1 11.5 10.3 11.0 7.7SD 2.5 2.4 4.1 3.0 3.4Min. 5.2 7.1 2.2 4.3Max. 16.8 15.7 14.9 12.1

FEM of the macaque zygomatic region, K. Kupczik et al.
© 2006 The Authors Journal compilation © 2006 Anatomical Society of Great Britain and Ireland
45
0.3 mm slice thickness, 120 kV, 228 mA). MAC-17 was
scanned on an X-Tek HMX 160 scanner (X-Tek Systems
Ltd, Tring, UK) at 123 kV, 87
µ
A and with a 0.2-mm Cu
filter. The voxel size was 0.23 mm. Prior to scanning,
the specimen MAC-17 was deliberately dehydrated for
14 days at room temperature in order to assist in the
visualization of the masticatory muscles. We sub-
sequently used the visualization software Amira (Mercury
Computer Systems, Inc., San Diego, CA, USA) to segment
the bone and teeth (treating enamel and dentine as a
single material) and, by extracting the surfaces, re-
constructed these materials in three dimensions. The
components of the periodontal ligament were not taken
into account as the limited resolution of the CT scans
did not allow for an accurate reconstruction. In MAC-
14 a clearly defined zygomatico-temporal suture was
also segmented as a discrete object, thus separating
the zygomatic from the temporal bone (Fig. 1A). In
MAC-17 this suture was partly fused and hence only
weakly indicated in the segmentation. Moreover, in
MAC-17 the superficial masseter was segmented and
reconstructed in three dimensions (Fig. 1B). Both surface
models were then converted into a volumetric tetrahedral
grid to be imported into ANSYS Mechanical (ANSYS,
Inc., Canonsburg, PA, USA).
Finite element modelling
The FEMs derived from the CT data used quadratic,
tetrahedral elements (ANSYS type SOLID 92). The MAC-14
model consists of 178 427 elements, while the larger
MAC-17 model including the superficial masseter com-
prises 232 564 elements. Given the size difference, this
resulted in a similar mesh density for the two samples.
All materials (i.e. bone, teeth, muscle and suture) were
modelled as linear elastic and isotropic (see
E
values for
each of the materials used below) with a Poisson’s ratio
of 0.3 in all cases.
In both models we modelled the boundary condi-
tions for the foramen magnum and occlusal surfaces of
the relevant molars by fully constraining the corre-
sponding nodes in these areas. In MAC-14, ten nodes
on the inferior border of the left zygomatic arch were
selected and a total load of 15 N applied. The loading
conditions of the zygomatic arch of MAC-17 are
described in detail below. In addition, the equivalent
surface nodes on the opposite zygomatic arch were
fully constrained. This idealization of the balancing
side forces was considered justifiable as it was remote
from the primary region of interest. Three series of
finite element analyses were conducted by varying:
(1) the
E
of the bone material (both models); (2) the
presence, absence and
E
of the zygomatico-temporal
suture (MAC-14 only); and (3) the method of load appli-
cation (MAC-17 only). The sensitivity of the FEMs to these
parameters was assessed and the parameters which
resulted in predicted strains concurring with those
measured experimentally were identified.
(1) Variation of bone elastic properties
We varied the Young’s modulus of the bone material
using values measured from the nanoindentation
study of the bone (see Table 1). A range of values were
noted and the highest, lowest and average value were
used for the modelling, with further fine adjustment to
identify the value that produced results similar to those
recorded experimentally. The teeth were assigned an
elastic modulus
E
of 70 GPa based on a mean of pub-
lished values for enamel and dentine (Cuy et al. 2002;
Kinney et al. 2003).
(2) Variations of the zygomatico-temporal suture
The defined zygomatico-temporal suture in the MAC-
14 specimen consisted of 420 tetrahedral elements.
Modelling was initially conducted to represent the two
extremes of fusion and complete disconnection. We
modelled the suture either with the same properties as
the surrounding bone (
E
= 9.1GPa) or omitted it from
the model completely, thus separating the zygomatic
and temporal portions of the zygomatic arch. We sub-
sequently modelled the suture with
E
-values derived
from the literature (1.20
±
0.20 MPa in 8-week-old
rabbits; Radhakrishnan & Mao, 2004) and also from the
nanoindentation results (see Table 1), with further
variation conducted to provide agreement with the
experimental results.
(3) Variation of load application
In MAC-17 we modelled the zygomatic arch loading
either by direct application of the load vectors to nodes
along the zygomatic arch or by application of the load
vectors to the modelled superficial masseter muscle.
Where direct loading was used, the load of 20 N was
uniformly distributed between 10, 20 and 41 nodes
along the superficial masseter bone scar on the zygomatic

FEM of the macaque zygomatic region, K. Kupczik et al.
© 2006 The AuthorsJournal compilation © 2006 Anatomical Society of Great Britain and Ireland
46
arch (Fig. 2). These loading conditions represented a
linear distribution (ten nodes) to an area distribution
covering the full attachment site of the muscle (41
nodes).
In the case of the muscle loading in the MAC-17
model, the load was applied to attempt to replicate the
experimental load conditions in a number of ways. In
the absence of a literature value for the bulk elastic
modulus of the superficial masseter, we experimented
by varying
E
and Poisson’s ratio. A value for
E
of 1 GPa
and Poisson’s ratio of 0.3 was found to deviate least
from the experimentally determined values. Thus, in
loading scenarios 1–3 the load of 20 N was distributed
between all the nodes of a cross-section of the muscle
through inferior, central and superior parts of the mus-
cle (Fig. 3). In loading scenario 4, a closer replication of
experimental conditions was attempted by loading a
line of nodes through the depth of the muscle at the
approximate location where the loading string pene-
trated the muscle (Fig. 3).
We compared the results from the finite element
modelling in each case to the experimental results. This
was done by calculating the mean maximum principal
strain of three superficial nodes in the model coincid-
ing with each of the six strain gauge locations on the
skulls (see Fig. 1). The overall difference between the
strains obtained experimentally at all six locations and
those obtained in the FEA was expressed as the Eucli-
dean distance calculated for each of the models. This
was computed as the square root of the sum of squared
differences between experiment and model. The
minimum Euclidean distance values obtained identi-
fied the modelling parameters which best fit the experi-
mental results. No statistical treatment of sensitivity
beyond this was feasible given the small number of
models solved.
Results
Variation of bone elastic properties
Figure 4(A,B) compares the mean maximum principal
strains recorded for five strain gauges (MAC-14 – with
the suture modelled using optimum
E
from the studies
of Fig. 5, see below) and six strain gauges (MAC-17) in
the loading experiments with the results of the FE
modelling while taking into account a range of bone
E
values. The strain gauge ZAL3 for MAC-14 turned out
to be faulty, and hence the results were excluded from
the subsequent analyses. In MAC-14, at all bone stiff-
ness levels the largest maximum principal strains occur
in the anterior zygomatic arch (ZAL1) with rapidly
decreasing strains posteriorly (Fig. 4A; except ZAL4,
E
= 4.0 GPa). Both the experimental and the modelled
strains in MAC-17 are more uniformly distributed and
peak in the middle of the anterior zygomatic arch for
E
= 11–17 GPa (ZAR-2, Fig. 4B).
Fig. 2 Variation of load application. Basal view of right zygomatic arch of MAC-17 with (A) 10, (B) 20 and (C) 41 nodes selected for loading. The area covered by the selected nodes reflects the attachment site of the superficial masseter (grey shaded area).
Fig. 3 Variation of loading of the right superficial masseter in MAC-17. 1, 2, 3: inferior, central and superior part of muscle loaded; 4: line of nodes loaded through the depth of the muscle (see text for details). Arrow indicates load point on M2.

FEM of the macaque zygomatic region, K. Kupczik et al.
© 2006 The Authors Journal compilation © 2006 Anatomical Society of Great Britain and Ireland
47
In general, the strains decrease with an increase in the
stiffness of bone in both models. The Euclidean distances
for MAC-14 indicate that the difference between experi-
mental and FE strains is lowest when modelled with the
mean
E
of bone derived from nanoindentation (9.1 GPa).
By contrast, for MAC-17 it was found that
E
= 14 GPa yields
a lower Euclidean distance than the average value obtained
by nanoindentation (11.0 GPa). The largest discrepancy
between the strains in the FEM and in the experiments
in MAC-17 is found for location IOR (
c
. 75
µε
see Fig. 4B).
Fig. 4 Effect of variation of bone material properties on maximum principal strains in FEA compared with experimental strain values (Exp, mean and standard deviation). (A) MAC-14; 15 N load application (please note that no experimental strain value is given for ZAL 3 due to gauge malfunction). Ebone ranges from 4 to 13.3 GPa with a mean of 9.1 GPa with Esuture = 0.0025 GPa constant (Poisson’s ratio is 0.3 throughout). (B) MAC-17; a load of 20 N was uniformly distributed between ten nodes on the zygomatic arch. Ebone ranges from 2.2 to 13 GPa with a mean of 11 GPa. ED = Euclidean distances (expressing the difference between the experimental and simulated strains; see text for details). IOL/R = infraorbital, left/right; ZAL/R1 to ZAL/R5 = zygomatic arch, left/right.
Fig. 5 Presence, absence and elastic properties variation of zygomatico-temporal suture in MAC-14 (Ebone = 9.1 GPa, Poisson’s ratio = 0.3). (A) Maximum principal strains along zygomatic arch with arch completely separated, fused (Esuture = Ebone) and suture with E = 0.0025 GPa (Poisson’s ratio = 0.3). (B) Effects of varying elastic properties of zygomatico-temporal suture on maximum principal strains in zygomatic arch. Esuture ranges from 0.001 to 1.9 GPa. Abbreviations as in Fig. 4.
FEM of the macaque zygomatic region, K. Kupczik et al.
© 2006 The AuthorsJournal compilation © 2006 Anatomical Society of Great Britain and Ireland
48
Variations of the zygomatico-temporal suture
Figure 5(A) illustrates the effects on the maximum prin-
cipal strains in MAC-14 when the zygomatic arch is fully
fused (i.e. the
E
of the zygomatico-temporal suture
equals that of bone), completely disconnected and
connected by a zygomatico-temporal suture. The bone
elastic modulus is kept constant in all three models
(
E
= 9.1 GPa). If the arch is separated, the strains are
markedly higher in the infraorbital region and on the
anterior portion of the zygomatic arch, whilst a decrease
is observed at these sites when the arch is completely
fused. The Euclidean distance shows that modelling the
zygomatic arch with a suture with
E
= 0.0025 GPa results
in strain values that are most similar to the experimen-
tal ones. If the stiffness of the suture is either increased
(
E
= 1.9 GPa; the mean obtained by our nanoindentation
study) or decreased (
E
= 0.0012 GPa; Radhakrishnan &
Mao, 2004) the differences in strain magnitude between
the model and the experiments become larger (Fig. 5B).
It is worth noting that the difference in Euclidean
distance between complete arch fusion (
E
= 9.1 GPa,
Fig. 5A),
E
= 1.9 GPa and
E
= 0.8 GPa (minimum value
from nanoindentation) is negligible.
Variation of load application to the zygomatic arch
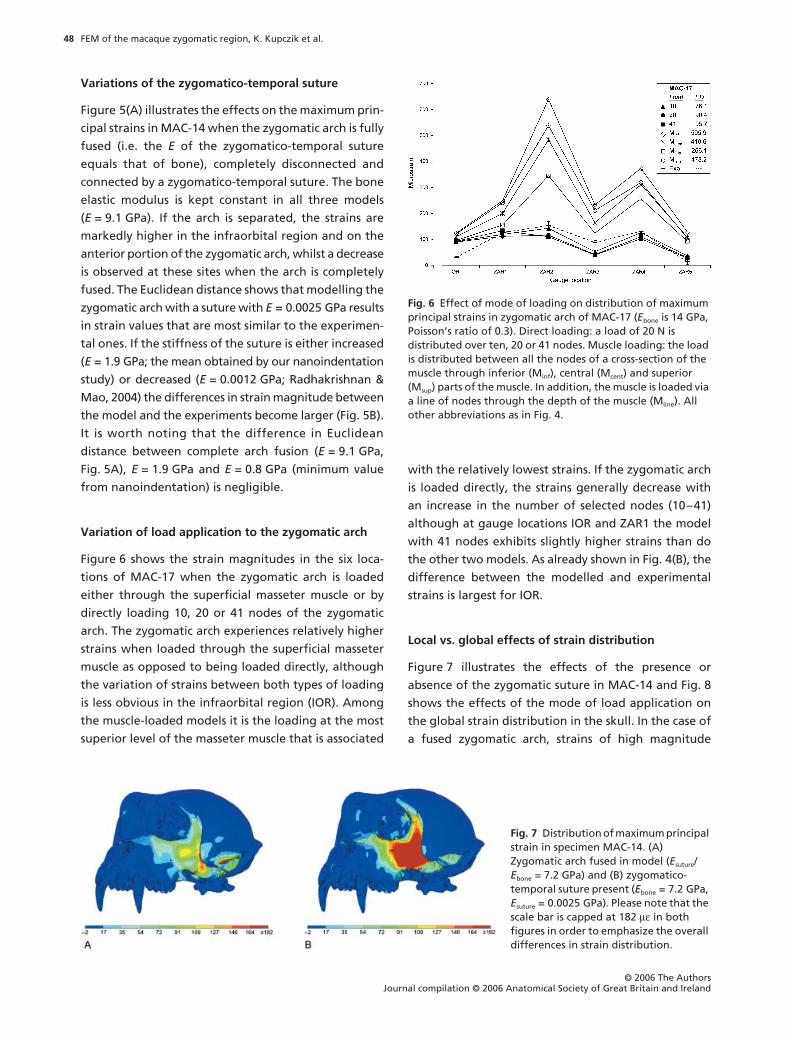
Figure 6 shows the strain magnitudes in the six loca-
tions of MAC-17 when the zygomatic arch is loaded
either through the superficial masseter muscle or by
directly loading 10, 20 or 41 nodes of the zygomatic
arch. The zygomatic arch experiences relatively higher
strains when loaded through the superficial masseter
muscle as opposed to being loaded directly, although
the variation of strains between both types of loading
is less obvious in the infraorbital region (IOR). Among
the muscle-loaded models it is the loading at the most
superior level of the masseter muscle that is associated
with the relatively lowest strains. If the zygomatic arch
is loaded directly, the strains generally decrease with
an increase in the number of selected nodes (10–41)
although at gauge locations IOR and ZAR1 the model
with 41 nodes exhibits slightly higher strains than do
the other two models. As already shown in Fig. 4(B), the
difference between the modelled and experimental
strains is largest for IOR.
Local vs. global effects of strain distribution
Figure 7 illustrates the effects of the presence or
absence of the zygomatic suture in MAC-14 and Fig. 8
shows the effects of the mode of load application on
the global strain distribution in the skull. In the case of
a fused zygomatic arch, strains of high magnitude
Fig. 7 Distribution of maximum principal strain in specimen MAC-14. (A) Zygomatic arch fused in model (Esuture/Ebone = 7.2 GPa) and (B) zygomatico-temporal suture present (Ebone = 7.2 GPa, Esuture = 0.0025 GPa). Please note that the scale bar is capped at 182 µε in both figures in order to emphasize the overall differences in strain distribution.
Fig. 6 Effect of mode of loading on distribution of maximum principal strains in zygomatic arch of MAC-17 (Ebone is 14 GPa, Poisson’s ratio of 0.3). Direct loading: a load of 20 N is distributed over ten, 20 or 41 nodes. Muscle loading: the load is distributed between all the nodes of a cross-section of the muscle through inferior (Minf), central (Mcent) and superior (Msup) parts of the muscle. In addition, the muscle is loaded via a line of nodes through the depth of the muscle (Mline). All other abbreviations as in Fig. 4.

FEM of the macaque zygomatic region, K. Kupczik et al.
© 2006 The Authors Journal compilation © 2006 Anatomical Society of Great Britain and Ireland
49
(i.e. > 164
µε
) are only present in the anterior end of
the zygomatic process and at the base of the zygomatic
process of the temporal (Fig. 7A). By contrast, if the
zygomatic arch is modelled with the suture present,
the relatively highest maximum principal strains are
observed in the zygoma covering parts of the infra-
orbital region and extending into the lateral orbital
margin (Fig. 7B). In MAC-17, the variation of the loading
conditions has less of an impact on regions in the face
adjacent to the zygomatic arch (Fig. 8A,B). The relatively
highest strains (> 273
µε
) are limited to the anterior
portion of the zygomatic process when the arch is
loaded directly (Fig. 8A). By contrast, when the arch is
loaded via the superficial masseter, these peak strains
occur both in the zygomatic arch and in the root of the
zygomatic process (Fig. 8B).
Discussion
The main objective of this study is to validate and assess
the sensitivity of FEMs of the macaque cranium to
variations in bone material properties, the presence or
absence of the zygomatico-temporal suture and the
loading regimen applied to the zygomatic arch.
To this end we carried out three studies to assess the
sensitivity of the principal strain distribution to varia-
tions in these parameters. The first examined the effects
of varying the material properties of the bone and was
carried out in conjunction with direct measurement of
the material properties of the experimental specimens.
The second examined the effects of presence or absence
and varying material properties of the zygomatico-
temporal suture. This aimed to test the hypothesis that
strains in the zygomatic arch decrease with gradual
obliteration of the suture. The third investigated the
effects of varying the ways in which loads are applied
to the zygomatic arch. The influence of the distribution
of the applied force on the predicted strain magnitudes
in the model was ascertained. It was hypothesised that a
more focussed, centralised application of loads on the
zygomatic arch would result in a larger bending moment
and hence maximum principal strains. Additionally, we
experimented with applying loads through a simplified
(i.e. isotropic, linear elastic) model of the superficial
masseter in order to see whether this led to strains that
are more congruent with experimental data. Finally,
we plotted strain maps in order to evaluate local vs.
global effects of the above experiments. The validity of
the different models was evaluated by examining
maximum principal strains in the infraorbital region and
the zygomatic arch and comparing values measured by
experimental strain gauge analysis with those obtained
through FEA simulation.
Variation of bone elastic properties
The results of Fig. 4 indicate that variations in
E
have
marked effects on the observed magnitudes of the
maximum principal strain as measured at each strain
gauge site. Unfortunately, our experiments suffered
from a strain gauge malfunction (ZAL3, MAC-14). This
gave extraordinarily high strain readings, which we
were able to demonstrate were erroneous in sub-
sequent experiments with a replacement gauge that gave
much better agreement with the modelling results.
However, because of uncertainties about alterations to
bone material properties due to dehydration in the
intervening period we have omitted the data obtained
for this site.
The Euclidean distances between successive experi-
ments indicate that using values of
E
close to experi-
mentally determined values results in a high degree of
congruence between experiment and model. The result
in MAC-14 was optimal when using the experimentally
obtained value (
E
= 9.1 GPa) while that in MAC-17 was
very close (using the experimentally determined value
of 11 GPa), but an iteratively determined value of
E
=
14 GPa provided an even closer match. In MAC-17 very
Fig. 8 Distribution of maximum principal strain in specimen MAC-17. (A) Zygomatic arch loaded directly through 20 nodes (Ebone = 11.3 GPa) and (B) zygomatic arch loaded via superficial masseter (Ebone = 11.3 GPa, Emasseter = 1 GPa; muscle not shown here). Please note that the scale bar is capped at 303 µε in both figures in order to emphasize the overall difference in the strain distribution.

FEM of the macaque zygomatic region, K. Kupczik et al.
© 2006 The AuthorsJournal compilation © 2006 Anatomical Society of Great Britain and Ireland
50
close agreement between experimental and modelled
strains is obtained at this value for E at all locations
except IOR (c. 75 µε discrepancy; see Fig. 4B). In this
specimen this gauge was placed on the maxilla with the
zygomatico-maxillary suture between it and the load
while in MAC-14 the equivalent gauge was on the
zygomatic. Thus, in MAC-17 it is highly likely that the
omission of this suture from the model underlies this
result (see below).
Irrespective of the bone Young’s modulus used, maxi-
mum principal strains peak in the anterior portion of
the zygomatic arch and then fall off sharply towards
the posterior end (MAC-14; Fig. 4A). These findings
confirm results of in vivo experiments of adult macaques
by Hylander & Johnson (1997), who also described a
steep strain gradient in the zygomatic arch. The situa-
tion is somewhat different in both the experimental
and the modelling results of MAC-17 where such a
strain gradient is less pronounced (Fig. 4B). This can
probably be attributed to the fact that the zygomatico-
temporal suture was more fused in this specimen and
hence not clearly defined in our FEM (see also below).
The mean E values of the zygomatic arch obtained by
the nanoindentation study do not compare well with
those used in previous FE studies of the macaque cra-
nium. Strait et al. (2005, their table 2) used an E value
of 12.5 GPa for the posterior zygomatic arch (based on
unpublished data for Macaca mulatta by Wang &
Dechow), whilst E = 20.8 GPa was used for the anterior
zygomatic arch. This lack of correspondence between
our measurements of E (see Table 1) and those of other
workers is little surprising given the uncertain preser-
vation history of our material. Several studies have sug-
gested that formalin fixation changes, to some extent,
the material properties of bone (Reilly & Burstein,
1974; Zioupos et al. 2000). In particular, it was shown
that fixation results in a decrease in bone stiffness of
pigs after 2 weeks of preservation (Dechow & Huynh,
1994), although Currey et al. (1995) showed that fixing
bone from an 18-month-old bovine in a 10% formalin
mixture for 3 h and subsequent buffering for 3 days
has only a limited effect on the elastic modulus in ten-
sion. While FEA should be accompanied by mechanical
testing of the materials involved, the anisotropic and
heterogeneous behaviour of bone as well as the type
(cortical or cancellous), the provenance (i.e. species and
anatomical region) and the preservation should ideally
be considered as well (Dechow & Hylander, 2000; Zioupos
et al. 2000; Peterson & Dechow, 2003; Schwartz-Dabney
& Dechow, 2003; Marinescu et al. 2005; Strait et al.
2005). For example, Peterson & Dechow (2003) found
that the zygomatic process of the temporal bone has
the highest anisotropy of all sites. We did not take this
into account in this study because of the complexity
and size of the model and yet obtained a remarkably
close match between experimental and modelling
results. It will be of interest in future studies further to
explore the sensitivity of FEMs to variations in the
direction and spatial variation of material properties
over a more substantial anatomical area.
Variations of the zygomatico-temporal suture
As noted in the introduction, the zygomatic arch has
been likened to a beam with fixed ends that is sub-
jected to an off-centre but uniform load, although the
zygomatico-temporal suture is likely to modulate the
ways in which loads are borne (Hylander & Johnson,
1997). Consequently, in our FEMs we introduced the
zygomatico-temporal suture such that the zygomatic arch
possessed a built-in constraint at one end and a flexible
joint at the other. The effects on the maximum principal
strains in MAC-14 of variations in the zygomatico-
temporal suture were therefore examined in order to
investigate the role of this suture in the FEM.
In these experiments the zygomatic arch is either
fully fused, disconnected or connected by a suture (with
E = 0.0025 GPa). Our results (Fig. 5A) indicate that when
the arch is fused the strains are markedly lower in the
infraorbital region (IOL) and on the anterior portion of
the zygomatic arch (ZAL1), whereas strains increase
when the arch is completely disconnected. By contrast,
introducing a suture into the model results in predicted
strains most similar to experimental values.
A second set of experiments examined the effects of
varying the stiffness of the suture (between E = 1.9 GPa
and E = 0.0012 GPa). The results (Fig. 5B) indicate that
a value of 0.0025 GPa, close to that reported in the
literature, gives the best overall fit to the experimentally
determined zygomatic and facial strains. This is not
consistent with the estimate of E = 1.9 GPa from
nanoindentation and serves to illustrate the difficulties
we experienced in sectioning the bone and suture
while maintaining low temperatures to avoid drying
out of the material.
Our study supports the view that sutures are decisive
for the accuracy of skeletal FEMs, at least with respect
to the cranium (Rayfield, 2005; Wang et al. 2006) and

FEM of the macaque zygomatic region, K. Kupczik et al.
© 2006 The Authors Journal compilation © 2006 Anatomical Society of Great Britain and Ireland
51
that the elastic properties of the suture have profound
effects on estimated strain magnitudes. This is consist-
ent with experimental findings, which show that the
strains in facial bones are limited while the interposed
sutures experience large deformations (Herring et al.
1996; Herring & Teng, 2000). Clearly this issue is most
significant when dealing with subadult material as
sutures tend to fuse gradually with age in larger mam-
mals (Sun et al. 2004). Under parasagittal bending the
infant/juvenile zygomatic arch with a relatively un-
mineralized suture will initially experience a relatively steep
anterior–posterior strain gradient due to large bending
moments at the anterior end (cf. Fig. 5B; E = 0.001 GPa).
With a gradual fusion of the suture the arch will resemble
more of a beam with two built-in ends and consequently
the bending moments and hence strains decrease ante-
riorly (cf. Fig. 5B; E = 1.9 GPa).
The material properties of sutures are highly variable
within a single cranium and between species. Thus, the
cranial sutures of neonate and young rats and pigs have
Young’s moduli ranging between 0.64 and 171.5 MPa
(Margulies & Thibault, 2000; McLaughlin et al. 2000;
Tanaka et al. 2000). This might have implications for
primate FEMs if the FE model uses sutural elastic pro-
perties taken from non-primate material. However, the
estimation of the material properties of sutures is not
straightforward as our attempt has shown. Therefore,
it might be reasonable where experimental data are
available to estimate the Young’s modulus iteratively
from the FE model – as indeed we have done in this
study. This approach is, however, only applicable in
limited situations and would not be possible for fossils.
In any case our findings underline the importance of
sutures and their mechanical properties in building
FEMs of crania.
Variation of load application to the zygomatic arch
A further experiment examined the effects of different
approaches to loading of the zygomatic arch. In the
first approach we applied loads to the arch directly,
varying the distribution of the loads however by select-
ing varying numbers of model nodes for their applica-
tion. In the second approach loads were applied via a
simplified model of the superficial masseter. The results
indicate that direct loadings consistently produce results
that are closer to the experimentally determined values.
Interestingly, loading the arch through a linear array of
ten nodes shows the best agreement with experimental
strains, although differences compared with the models
with a higher number of selected nodes are relatively
small. The findings also corroborate the hypothesis that
a more centralized load application results in relatively
larger maximum principal strains in the zygomatic arch
(Fig. 6).
The effect on the model’s results of loading the zygo-
matic arch through the masseter muscle is very surpris-
ing, particularly as the magnitude of the predicted
strains is significantly higher (Fig. 6) when the muscle is
included. It is not clear why this is the case. Possibly the
muscle is providing an additional torsional load to the
arch and it might also add leverage if the force vectors
are not exactly in line with the long axis of the muscle,
as suggested by an anonymous reviewer. Alternatively,
it may indicate that the superficial masseter transfers
forces to the zygomatic arch via a limited number of
tendinous connections, most like the arrangement of
loads in Fig. 2(A). The failure of the muscle loading simu-
lations to achieve results close to experimental values
likewise indicates the inadequacy of modelling muscle
as an isotropic linear elastic material. In future models,
if muscles are to be used to apply loads it will be necessary
to include more accurate information on the muscle’s
elasticity as well as anisotropies and non-linearities to
account for true muscle architecture.
Local vs. global effects of strain distribution
Finally we examined the full FEM of each macaque
cranium under varying loadings and conditions as in
the preceding experiments. FEA has the advantage of
allowing strain maps of the whole to be estimated and
visualized although, as we have shown here, there are
many variables that can undermine the accuracy of the
FEM. In producing these maps our aim is to evaluate
local vs. global effects of the above experiments.
From Figs 7 and 8 it is clear that the effects we
describe above in relation to the areas local to strain
gauges obtain more generally. Thus, the presence of a
zygomatico-temporal suture affects the strains experi-
enced over a wide area (Fig. 7A vs. Fig. 7B) including
the anterior part of the zygomatic and infraorbital
regions. The effect of direct vs. ‘muscle’ loading of the
zygomatic arch (Fig. 8A vs. Fig. 8B) is more confined to
within the zygomatic region. However, within this
region the effects are substantial.
In detail, the effects of varying these diverse para-
meters are significant in that estimates of absolute

FEM of the macaque zygomatic region, K. Kupczik et al.
© 2006 The AuthorsJournal compilation © 2006 Anatomical Society of Great Britain and Ireland
52
values of strain at specific loci are markedly affected.
However, in general all loading regimens show that
the zygomatic arch manifests relatively large strains,
most concentrated anteriorly and falling away in mag-
nitude with distance. Whether this level of generality is
acceptable is entirely dependent on the question at
hand; but our data indicate that reasonable approxi-
mations of how loads are handled in the cranium might
be obtained for regions not amenable to strain gauging
in living species and in fossil material as long as atten-
tion is paid to the modelling of sutures and muscle
attachments. If accuracy is required then the details of
suture morphology, muscle attachment, applied loads
and mechanical properties become of too great signi-
ficance to be ignored.
Acknowledgements
We gratefully acknowledge the financial support of
the Leverhulme Trust (F/00224/I). We also thank Sue
Taft, Neil Curtis, Lee Page, Andrea Cardini, Sam Cobb,
Callum Ross, Fred Spoor, Chris Dean and staff at Hull
Royal Infirmary for their advice and assistance. We are
also grateful to Daniel Lieberman and three anony-
mous reviewers for their helpful comments during the
preparation of the manuscript.
References
Buckland-Wright JC (1978) Bone structure and the patterns offorce transmission in the cat skull (Felis catus). J Morph 155,35–62.
Chen XB, Chen H (1998) The influence of alveolar structures onthe torsional strain field in a gorilla corporeal cross-section.J Hum Evol 35, 611–633.
Currey JD, Brear K, Zioupos P, Reilly GC (1995) Effect offormaldehyde fixation on some mechanical properties ofbovine bone. Biomaterials 16, 1267–1271.
Cuy JL, Mann AB, Livi KJ, Teaford MF, Weihs TP (2002) Nano-indentation mapping of the mechanical properties of humanmolar tooth enamel. Arch Oral Biol 47, 281–291.
Daegling DJ, Hylander WL (1997) Occlusal forces and mandibularbone strain: is the primate jaw ‘overdesigned’? J Hum Evol33, 705–717.
Daegling DJ, Hylander WL (2000) Experimental observation,theoretical models, and biomechanical inference in the studyof mandibular form. Am J Phys Anthropol 112, 541–551.
Dechow PC, Carlson DS (1990) Occlusal force and craniofacialbiomechanics during growth in rhesus monkeys. Am J PhysAnthropol 83, 219–237.
Dechow PC, Huynh T (1994) Elastic properties and biomechanicsof the baboon mandible [abstract]. Am J Phys AnthropolSuppl 22, 94–95.
Dechow PC, Hylander WL (2000) Elastic properties and masti-catory bone stress in the macaque mandible. Am J PhysAnthropol 112, 553–574.
Fagan MJ, Julian S, Siddall DJ, Mohsen A (2002) Patient-specificspine models – Part 1: finite element analysis of the lumbarintervertebral disc – a material sensitivity study. Proc IME HJ Eng Med 216, 299–314.
Herring SW (1972) Sutures – a tool in functional cranial analysis.Acta Anat 83, 222–247.
Herring SW, Teng SY (2000) Strain in the braincase and itssutures during function. Am J Phys Anthropol 112, 575–593.
Herring SW, Teng SY, Huang XF, Mucci RJ, Freeman J (1996)Patterns of bone strain in the zygomatic arch. Anat Rec 246,446–457.
Hylander WL, Johnson KR (1997) In vivo bone strain patternsin the zygomatic arch of macaques and the significance ofthese patterns for functional interpretations of craniofacialform. Am J Phys Anthropol 102, 203–232.
Jaslow CR (1990) Mechanical properties of cranial sutures. JBiomech 23, 313–321.
Kinney JH, Marshall SJ, Marshall GW (2003) The mechanicalproperties of human dentin: a critical review and re-evaluationof the dental literature. Crit Rev Oral Biol M 14, 13–29.
Macho GA, Spears IR (1999) Effects of loading on the bio-mechanical behavior of molars of Homo, Pan, and Pongo.Am J Phys Anthropol 109, 211–227.
Macho GA, Shimizu D, Jiang Y, Spears LR (2005) Austra-lopithecus anamensis: a finite element approach to studyingthe functional adaptations of extinct hominins. Anat RecPart A 283A, 310–318.
Mao JJ (2002) Mechanobiology of craniofacial sutures. J DentRes 81, 810–816.
Mao JJ, Wang X, Kopher RA (2003) Biomechanics of craniofacialsutures: orthopedic implications. Angle Orthod 73, 128–135.
Margulies SS, Thibault KH (2000) Infant skull and suture pro-perties: measurements and implications for mechanismsof pediatric brain injury. J Biomech Eng-T Asme 122, 364–371.
Marinescu R, Daegling DJ, Rapoff AJ (2005) Finite-elementmodeling of the anthropoid mandible: the effects of alteredboundary conditions. Anat Rec Part A 283A, 300–309.
McConnell CJ, Crompton RH (2001) Finite element stressanalysis of food–tooth interactions in primates. In ComputerMethods in Biomechanics and Biomedical Engineering 3(eds Middleton J, Jones ML, Shrive NG, Pande GN), pp. 673–678. Amsterdam: Gordon and Breach.
McLaughlin E, Zhang Y, Pashley D, Borke J, Yu J (2000) Theload-displacement characteristics of neonatal rat cranialsutures. Cleft Palate-Cran J 37, 590–595.
Moss ML (1973) A functional cranial analysis of primatecraniofacial growth. In Symposium IVth International Con-gress on Primatology: Craniofacial Biology of the Primates 3(ed. Zingeser MR), pp. 191–208. Basel: Karger.
Oyen OJ, Walker AC, Rice RW (1979) Craniofacial growth inolive baboons (Papio cynocephalus): browridge formation.Growth 43, 174–187.
Peterson J, Dechow PC (2003) Material properties of thehuman cranial vault and zygoma. Anat Rec 274A, 785–797.
Preuschoft H, Witzel U (2004) Functional structure of the skullin hominoidea. Folia Primatol 75, 219–252.

FEM of the macaque zygomatic region, K. Kupczik et al.
© 2006 The Authors Journal compilation © 2006 Anatomical Society of Great Britain and Ireland
53
Radhakrishnan P, Mao JJ (2004) Nanomechanical properties offacial sutures and sutural mineralization front. J Dent Res83, 470–475.
Rafferty KL, Herring SW, Artese F (2000) Three-dimensionalloading and growth of the zygomatic arch. J Exp Biol 203,2093–2104.
Rafferty KL, Herring SW, Marshall CD (2003) Biomechanics ofthe rostrum and the role of facial sutures. J Morph 257, 33–44.
Rayfield EJ (2005) Using finite-element analysis to investigatesuture morphology: a case study using large carnivorousdinosaurs. Anat Rec Part A 283A, 349–365.
Reilly DT, Burstein AH (1974) The mechanical properties ofcortical bone. J Bone Joint Surg 56A, 1001–1022.
Richmond BG, Wright BW, Grosse L, et al. (2005) Finite elementanalysis in functional morphology. Anat Rec Part A 283A,259–274.
Ross CF, Patel BA, Slice DE, et al. (2005) Modeling masticatorymuscle force in finite element analysis: Sensitivity analysis usingprincipal coordinates analysis. Anat Rec Part A 283A, 288–299.
Schwartz-Dabney CL, Dechow PC (2003) Variations in corticalmaterial properties throughout the human dentate mandi-ble. Am J Phys Anthropol 120, 252–277.
Sellers WI, Crompton RH (2004) Using sensitivity analysis tovalidate the predictions of a biomechanical model of biteforces. Ann Anat 186, 89–95.
Smith K, Beighton D (1986) The effects of the availability ofdiet on the levels of exoglycosidases in the supragingivalplaque of macaque monkeys. J Dent Res 65, 1349–1352.
Smith K, Beighton D (1987) Proteolytic activities in the suprag-ingival plaque of monkeys (Macaca fascicularis). Arch OralBiol 32, 473–476.
Spears IR, Crompton RH (1996) The mechanical significance ofthe occlusal geometry of great ape molars in food break-down. J Hum Evol 31, 517–535.
Spears IR, Macho GA (1998) Biomechanical behaviour ofmodern human molars: implications for interpreting thefossil record. Am J Phys Anthropol 106, 467–482.
Strait DS, Wang Q, Dechow PC, et al. (2005) Modeling elasticproperties in finite element analysis: how much precision isneeded to produce an accurate model? Anat Rec Part A283A, 275–287.
Sun ZY, Lee E, Herring SW (2004) Cranial sutures and bones:growth and fusion in relation to masticatory strain. AnatRec Part A 276A, 150–161.
Tanaka E, Miyawaki Y, del Pozo R, Tanne K (2000) Changes inthe biomechanical properties of the rat interparietal sutureincident to continuous tensile force application. Arch OralBiol 45, 1059–1064.
Wang Q, Dechow PC, Richmond B, et al. (2006) Fusion ofcraniofacial sutures in monkey skulls with special referenceto the Finite Element Analysis. Am J Phys Anthrop Suppl129, 184–185.
Wang WJ, Crompton RH, Carey TS, et al. (2004) Comparisonof inverse-dynamics musculo-skeletal models of AL 288-1Australopithecus afarensis and KNM-WT 15000 Homoergaster to modern humans, with implications for theevolution of bipedalism. J Hum Evol 47, 453–478.
Witzel U, Preuschoft H (2002) Function-dependent shapecharacteristics of the human skull. Anthropol Anz 60,113–135.
Witzel U, Preuschoft H, Sick H (2004) The role of the zygomaticarch in the statics of the skull and its adaptive shape. FoliaPrimatol 75, 202–218.
Zioupos P, Smith CW, An YH (2000) Factors affecting mechan-ical properties of bone. In Mechanical Testing of Bone andthe Bone–Implant Interface (eds An YH, Draughn RA),pp. 65–85. New York: CRC Press.
Top Related
Copyright © 2022 FDOKUMEN

