
Zinc is the molecular "switch" that controls the catalytic cycle of bacterial leucyl-tRNA synthetase
9
Zinc is the molecular “switch” that controls the catalytic cycle of bacterial leucyl-tRNA synthetase Manonmani Kumar b , Sathish A.P. Kumar b , Aleksandar Dimkovikj a , Layla N. Baykal a , Mallory J. Banton a , Maya M. Outlaw a , Kristen E. Polivka a , Rachel A. Hellmann-Whitaker a, ⁎ a Department of Chemistry and Physics, Coastal Carolina University, Smith Science Center, Room 216, 109 Chanticleer Dr. East, Conway, SC 29526, USA b Department of Computer Science and Information Systems, Coastal Carolina University, 301 Allied Drive, Conway, SC 29526, USA abstract article info Article history: Received 29 May 2014 Received in revised form 12 September 2014 Accepted 12 September 2014 Available online 22 September 2014 Keywords: Leucyl-tRNA synthetase ZN-1 domain Aminoacylation reaction The Escherichia coli (E. coli) leucyl-tRNA synthetase (LeuRS) enzyme is part of the aminoacyl-tRNA synthetase (aaRS) family. LeuRS is an essential enzyme that relies on specialized domains to facilitate the aminoacylation re- action. Herein, we have biochemically characterized a specialized zinc-binding domain 1 (ZN-1). We demon- strate that the ZN-1 domain plays a central role in the catalytic cycle of E. coli LeuRS. The ZN-1 domain, when associated with Zn 2+ , assumes a rigid architecture that is stabilized by thiol groups from the residues C159, C176 and C179. When LeuRS is in the aminoacylation complex, these cysteine residues form an equilateral planar triangular configuration with Zn 2+ , but when LeuRS transitions to the editing conformation, this geometric configuration breaks down. By generating a homology model of LeuRS while in the editing conformation, we con- clude that structural changes within the ZN-1 domain play a central role in LeuRS's catalytic cycle. Additionally, we have biochemically shown that C159, C176 and C179 coordinate Zn 2+ and that this interaction is essential for leucylation to occur, but is not essential for deacylation. Furthermore, calculated K d values indicate that the wild- type enzyme binds Zn 2+ to a greater extent than any of the mutant LeuRSs. Lastly, we have shown through secondary structural analysis of our LeuRS enzymes that Zn 2+ is an architectural cornerstone of the ZN-1 domain and that without its geometric coordination the domain collapses. We believe that future research on the ZN-1 domain may reveal a possible Zn 2+ dependent translocation mechanism for charged tRNA Leu . © 2014 Elsevier Inc. All rights reserved. 1. Introduction Aminoacyl-tRNA synthetases (aaRSs) are an ancient family of en- zymes that have evolved to ensure the fidelity of the first step of protein translation through the faithful production of correctly aminoacylated tRNAs [1]. There are twenty members of the aaRS family—one aaRS per amino acid [2]. The aaRSs are divided into two classes with each consisting of 10 enzymes [2–4]. These two classes have been partitioned according to structural homology and mechanistic similarities of the aminoacylation reaction [2–4]. The aminoacylation reaction is a two- step reaction that begins with the adenylation of an amino acid followed by the aminoacylation of tRNA [5]. Each enzyme within the family of aaRSs catalyzes the aminoacylation reaction. Surprisingly there is considerable diversity in domain insertions, placement and sequence homology within this family of enzymes [6,7]. Additional domain inser- tions within the core aminoacylation domain, such as the connective polypeptide (CP1) editing domain, were thought to be later evolutionary additions to aaRSs [8,9]. To support this hypothesis, mini- mal core aminoacylation domains have been constructed that function similar to the wild type enzyme [8–10]. These findings provide evidence that class I and class II synthetases may have evolved from a common ancestor and additionally that the CP1 domain can enhance the activity of the aaRS, but is not an absolute requirement for aminoacylation to occur [8,9,11,12]. Therefore, domain insertions, similar to the CP1 do- main, allowed for specialized charging of speci fic amino acids by each aaRS and allowed aaRSs to develop auxiliary biological func- tions [8,13,14]. This research effort focuses on a specific domain insertion, which is the zinc-binding domain. Many aaRSs bind zinc within specialized zinc-binding domains. Some zinc-binding domains are known to recog- nize and interact with tRNA [15,16]. Others are known to help facilitate the aminoacylation reaction [17]. These examples of different aaRS zinc- binding domains highlight the domain's functional diversification. Zinc-binding domain diversity also exists across speciation lines for leucyl-tRNA synthetase (LeuRS) enzymes. Two zinc-binding domains called ZN-1 and ZN-2 have been found in LeuRS enzymes. Based on the sequence alignment of bacterial, and eukaryote-mitochondrial LeuRSs, the ZN-1 domain is positioned immediately before the CP1 Journal of Inorganic Biochemistry 142 (2015) 59–67 ⁎ Corresponding author. Fax: 1 843-349-2841. E-mail address: [email protected] (R.A. Hellmann-Whitaker). http://dx.doi.org/10.1016/j.jinorgbio.2014.09.006 0162-0134/© 2014 Elsevier Inc. All rights reserved. Contents lists available at ScienceDirect Journal of Inorganic Biochemistry journal homepage: www.elsevier.com/locate/jinorgbio
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Zinc is the molecular "switch" that controls the catalytic cycle of bacterial leucyl-tRNA synthetase

Journal of Inorganic Biochemistry 142 (2015) 59–67
Contents lists available at ScienceDirect
Journal of Inorganic Biochemistry
j ourna l homepage: www.e lsev ie r .com/ locate / j inorgb io
Zinc is themolecular “switch” that controls the catalytic cycle of bacterialleucyl-tRNA synthetase
Manonmani Kumar b, Sathish A.P. Kumar b, Aleksandar Dimkovikj a, Layla N. Baykal a, Mallory J. Banton a,Maya M. Outlaw a, Kristen E. Polivka a, Rachel A. Hellmann-Whitaker a,⁎a Department of Chemistry and Physics, Coastal Carolina University, Smith Science Center, Room 216, 109 Chanticleer Dr. East, Conway, SC 29526, USAb Department of Computer Science and Information Systems, Coastal Carolina University, 301 Allied Drive, Conway, SC 29526, USA
⁎ Corresponding author. Fax: 1 843-349-2841.E-mail address: [email protected] (R.A. Hellmann
http://dx.doi.org/10.1016/j.jinorgbio.2014.09.0060162-0134/© 2014 Elsevier Inc. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 29 May 2014Received in revised form 12 September 2014Accepted 12 September 2014Available online 22 September 2014
Keywords:Leucyl-tRNA synthetaseZN-1 domainAminoacylation reaction
The Escherichia coli (E. coli) leucyl-tRNA synthetase (LeuRS) enzyme is part of the aminoacyl-tRNA synthetase(aaRS) family. LeuRS is an essential enzyme that relies on specialized domains to facilitate the aminoacylation re-action. Herein, we have biochemically characterized a specialized zinc-binding domain 1 (ZN-1). We demon-strate that the ZN-1 domain plays a central role in the catalytic cycle of E. coli LeuRS. The ZN-1 domain, whenassociated with Zn2+, assumes a rigid architecture that is stabilized by thiol groups from the residues C159,C176 and C179.When LeuRS is in the aminoacylation complex, these cysteine residues form an equilateral planartriangular configuration with Zn2+, but when LeuRS transitions to the editing conformation, this geometricconfiguration breaks down. By generating a homologymodel of LeuRSwhile in the editing conformation,we con-clude that structural changes within the ZN-1 domain play a central role in LeuRS's catalytic cycle. Additionally,we have biochemically shown that C159, C176 and C179 coordinate Zn2+ and that this interaction is essential forleucylation to occur, but is not essential for deacylation. Furthermore, calculated Kd values indicate that the wild-type enzyme binds Zn2+ to a greater extent than any of the mutant LeuRSs. Lastly, we have shown throughsecondary structural analysis of our LeuRS enzymes that Zn2+ is an architectural cornerstone of the ZN-1 domainand that without its geometric coordination the domain collapses. We believe that future research on the ZN-1domain may reveal a possible Zn2+ dependent translocation mechanism for charged tRNALeu.
© 2014 Elsevier Inc. All rights reserved.
1. Introduction
Aminoacyl-tRNA synthetases (aaRSs) are an ancient family of en-zymes that have evolved to ensure the fidelity of the first step of proteintranslation through the faithful production of correctly aminoacylatedtRNAs [1]. There are twenty members of the aaRS family—one aaRSper amino acid [2]. The aaRSs are divided into two classes with eachconsisting of 10 enzymes [2–4]. These two classes have been partitionedaccording to structural homology and mechanistic similarities of theaminoacylation reaction [2–4]. The aminoacylation reaction is a two-step reaction that beginswith the adenylation of an amino acid followedby the aminoacylation of tRNA [5]. Each enzyme within the family ofaaRSs catalyzes the aminoacylation reaction. Surprisingly there isconsiderable diversity in domain insertions, placement and sequencehomology within this family of enzymes [6,7]. Additional domain inser-tions within the core aminoacylation domain, such as the connectivepolypeptide (CP1) editing domain, were thought to be later
-Whitaker).
evolutionary additions to aaRSs [8,9]. To support this hypothesis, mini-mal core aminoacylation domains have been constructed that functionsimilar to thewild type enzyme [8–10]. These findings provide evidencethat class I and class II synthetases may have evolved from a commonancestor and additionally that the CP1 domain can enhance the activityof the aaRS, but is not an absolute requirement for aminoacylation tooccur [8,9,11,12]. Therefore, domain insertions, similar to the CP1 do-main, allowed for specialized charging of specific amino acids byeach aaRS and allowed aaRSs to develop auxiliary biological func-tions [8,13,14].
This research effort focuses on a specific domain insertion, which isthe zinc-binding domain. Many aaRSs bind zinc within specializedzinc-binding domains. Some zinc-binding domains are known to recog-nize and interact with tRNA [15,16]. Others are known to help facilitatethe aminoacylation reaction [17]. These examples of different aaRS zinc-binding domains highlight the domain's functional diversification.
Zinc-binding domain diversity also exists across speciation lines forleucyl-tRNA synthetase (LeuRS) enzymes. Two zinc-binding domainscalled ZN-1 and ZN-2 have been found in LeuRS enzymes. Based onthe sequence alignment of bacterial, and eukaryote-mitochondrialLeuRSs, the ZN-1 domain is positioned immediately before the CP1

60 M. Kumar et al. / Journal of Inorganic Biochemistry 142 (2015) 59–67
domain, while ZN-2 divides the aminoacylation domain in approxi-mately half [18]. Certain subsets of LeuRSs are known to bind a zincatom in both the zinc-binding domains, while other subsets of LeuRSsbind one zinc atom or may not bind zinc at all although they may stillmaintain the domains [18]. There is also variability within the domains,in that some LeuRSsmay only have one zinc-binding domain,which canbe either ZN-1 or ZN-2 [18]. The domains also vary in size, ranging from30 to 60 residues [18]. Due to this diversity within the zinc-binding do-mains of LeuRSs, we set out to biochemically understand how the zinc-binding domain may affect Escherichia coli LeuRS catalysis, which hasnever been reported until now.
A crystal structure of E. coli LeuRS indicates that this enzymecontains a single zinc-binding domain 1 (ZN-1 domain) that binds to asingle zinc atom [12]. The crystal structure highlights the importanceof the ZN-1 domain in establishing the aminoacylation conformationand stabilizing the Leu-AMP adenylate [12]. However, the crystal struc-tures presented in Palencia et al. paper do not show E. coli LeuRS's ZN-1domain in the editing conformation, but shows Thermus thermophilusLeuRS's ZN-1 domain in the editing conformation. Palencia et al. compu-tationally built a partial ZN-1 domain for the E. coli LeuRS editing struc-ture using the T. thermophilus LeuRS editing structure as a reference,which provided helpful insight, but failed to capture the truly dynamicnature of the ZN-1 domain in its entirety [12]. Therefore, we computa-tionally generated a homology model of E. coli LeuRS's ZN-1 domain inthe editing conformation to better visualize how the ZN-1 domain's posi-tion and structuremay change as the enzymecompletes its catalytic cycle.We built our homologymodel using the T. thermophilus LeuRS's ZN-1 do-main in the editing conformation because the T. thermophilus LeuRS struc-ture is the only crystal structure in existence that visualizes the ZN-1domain in the editing conformation. Building a homology model of anyprotein requires a reference structure, since T. thermophilus shares morethan 40% sequence homology with E. coli LeuRS it was the most logicalchoice [19].
With the homology model, we were able to structurally and bio-chemically piece together the disjointed information that currentlyhas been published concerning the E. coli LeuRS ZN-1 domain. Weperformed alanine-scanning mutagenesis of the ZN-1 domain and bio-chemically determined that only C159, C176 and C179 are required tochelate Zn2+. Herein, we report the geometric changes that occur with-in this zinc chelating pocket of the ZN-1 domain as LeuRS transitions be-tween the aminoacylation and editing conformations. We report thenovel structural features of this ZN-1 domain and how these featurescontribute to the domain's inherent thermodynamic instability, whichmay enable the domain to adopt dramatically different structural statesbetween the aminoacylation and editing conformations. While LeuRS isin the aminoacylation conformation the ZN-1 domain packs against theactive site to stabilize Leu-AMP, but once the enzyme transitions to theediting conformation the ZN-1 domain appears to swing away 14° fromthe aminoacylation active site. We hypothesize that the movement ofthe ZN-1 domain away from the active site opens a structural path forcharged tRNALeu to effectively translocate to the CP1 domain since theZN-1 domain packs tightly against the aminoacylation active sitewhile the enzyme is in the aminoacylation conformation. Global struc-tural perturbations due tomolecular “hinge” sites within LeuRS in addi-tion to the “sling-shot” movement of the 3′-tRNALeu acceptor arm [20]may facilitate the movement of the ZN-1 domain away from theaminoacylation active site.
2. Experimental
2.1. Materials
T7 RNA polymerase (Invitrogen, Carlsbad, CA) was used to in vitrotranscribe tRNALeu via run-off transcription [21]. The tRNA productwas purified by urea-containing polyacrylamide gel electrophoresis[21] and quantitated based on absorbance at 260 nm using a molar
absorption coefficient of 840,700 M−1 cm−1 [22]. Purified tRNALeu
was denatured at 80 °C for 1 min, followed by an addition of 1 mMMgCl2 and quick-cooled on ice for re-folding.
2.2. Generating mutations
The wild type E. coli LeuRS gene is encoded within the plasmidpRWecLeuRS, which was generated by extracting E. coli genomic DNAand utilizing PCR to amplify and subsequently clone leuS (GeneBank Ac-cession #EG10532) into pET15b (Fisher Scientific, Pittsburg, PA) as pre-viously demonstrated by Härtlen and Madern [23]. To generatepRWecLeuRS we used synthesized oligonucleotides (Integrated DNATechnology (IDT), Coralville, IA). The forward primer 5′-CGCCATATGATGCAAGAGCAATACCGCCCG-3′ and reverse primer 5′-CGCGGATCCCGGTTGCTGGTCTAACTCCTC-3′, which contain the NdeI and BamHI restric-tion sites, respectively. The resulting pRWecLeuRS was used to insertmutations via PCR generating the plasmids: pRWC159A (C159A),pRWC176A (C176A), and pRWC179A (C179A). These mutations wereconstructed using synthesized complementary oligonucleotides (IDT,Coralville, IA), the forward primers are as follows: C159A, 5′-GCGGTCAACTGGGCTCCGAACGACCAG-3′; C176A, 5′-CGACGGCTGCGCTTGGCGCTGCG-3′; and C179A, 5′-CTGCTGGCGCGCTGATACCAAAG-3′. In addi-tion, we generated the T252Y mutant to purify the mischarged product[3H] Ile-tRNALeu (as described below) with the following forward prim-er: 5′-CCCGCCGGACTACTTTATGGGTTGTACC-3′ [24]. DNA sequencingfor all plasmids was carried out by Eurofins MWG Operon, Huntsville,AL.
2.3. Protein expression
To express the ZN-1 domain LeuRS mutants, the zinc-binding do-main mutant plasmids were used to transform E. coli BL21 (Stratagene,La Jolla, CA). A single colony was used to inoculate 5 ml of Luria-Bertanimedium containing 100 μg/ml ampicillin (LB-Amp) and grown over-night at 37 °C. The overnight culture was then transferred to 1 l ofLB-Amp and grown at 37 °C to an OD600 of 0.6–0.8. Expression ofLeuRS was induced with 1 mM IPTG for 3 h at 37 °C. Harvestedcells were lysed by sonication and each LeuRS, which contained anN-terminal six-histidine tag, was purified by affinity purificationusing HIS-Select HF nickel affinity resin as described before [25].The purified proteins were concentrated via Pierce concentrators50KMWCO (molecular weight cut-off) (Thermo Scientific, Rockford,IL) and quantitated using the Bio-Rad protein assay according to thecommercial protocol.
2.4. Aminoacylation assays
Leucylation reactions contained 60 mM Tris–HCl, pH 7.5, 10 mMMgCl2, 1 mM DTT, 4 μM folded tRNALeu, 21 μM [3H]-L-leucine(150 μCi/ml), and 50 nM enzyme. Initial velocities of leucylation weredetermined using 10 nMof enzyme. All leucylation reactionswere initi-atedwith 4mMATP. Aliquots of 10 μl were quenched on TCA-soaked fil-ter pads, washed, and quantitated via scintillation counting asdescribed before [25].
2.5. Mischarged tRNA preparation and enyzmatic deacylation
The [3H]-Ile-tRNALeu was prepared using an editing defective T252YLeuRSmutant in an aminoacylation reaction thatwas incubated at 30 °Cfor 3 h. Reactions were stopped using 0.18% acetic acid, and extractedusing two equal volumes of phenol/chloroform/isoamyl alcohol,pH 4.3 (125:24:1) [26]. A one-half volume of 4.6 M NH4OAc, pH 5.0,was added followed by an ethanol precipitation. The driedaminoacylated tRNA pellets were resuspended in 10 mM KH2PO4,pH 5.0. Hydrolytic editing assays were carried out at room temperaturein 60 mM Tris, pH 7.5, 10 mM MgCl2, and 0.8 μM [3H]-Ile-tRNALeu and

61M. Kumar et al. / Journal of Inorganic Biochemistry 142 (2015) 59–67
was initiated with 100 nM enzyme [27]. At specific time points, 5 μl al-iquots were spotted onto filter pads, washed, and quantitated.
2.6. Homology model construction
To generate a homology model of the ZN-1 domain of E. coli LeuRSwhile in the editing conformation, we employed in silico proteinstructure prediction techniques. The ZN-1 domain is a highly dynamicdomain and has only been crystallized while in the editing confor-mation for T. thermophilus LeuRS (1OBH) [28]. The sequence align-ment of T. thermophilus LeuRS and E. coli LeuRS indicates that thesetwo enzymes have greater than 40% sequence homology as hasbeen previously reported [19]. A computationally predicted modelof E. coli LeuRS while in the editing conformation was generatedusing Swiss Model [29] in the Alignment Mode. The quality of thestructure was determined by a QMEAN Z-score of 0.91. Further vali-dation of the predicted protein structure was done using ProQ [30].ProQ generates two quality measures: LGscore and MaxSub, whichare used to evaluate the accuracy of the predicted protein structure.The homology model of E. coli LeuRS was assigned a LGscore of 4.249and a MaxSub score of 0.373. Both values indicate a highly accuratestructure.
2.7. Generation of the Apo-LeuRS enzyme
Purified wild-type LeuRS (20 mg/ml) was dialyzed using Spectra/Por molecular porous membrane tubing MWCO 12–14,000 Da (Spec-trum Laboratories, Inc., Rancho Dominguez, CA) in 5 mM EDTA, pH 8.0with 50 mM Tris, pH 7.5 for 48 h at 4 °C [31].
2.8. Measurement of dissociation constants of Zn2+ from LeuRS enzymes
A constant LeuRS concentration of 6 μMwas used against an increas-ing concentration gradient of EDTA (0.5 μM–10 μM) to determine thedissociation constants of Zn2+ from wild-type and mutant LeuRS en-zymes [32]. To measure the dissociation constants we used a ThermoScientific Genesys 10S UV–Vis spectrophotometer (Waltham, MA).Scanning absorption spectra were taken of ZnCl2, which determinedits optimal absorptionwas 260 nm [33]. Spectral analysis was correctedfor LeuRS, EDTA and Mg2+ background signals at 260 nm. Data wasgraphed with GraphPad Prism 6 (GraphPad Software, Inc., La Jolla, CA)using the equation B = Ka[L] / 1 + Ka[L]. From this plot, we calculatedthe Kd values for Zn2+ from LeuRS enzymes.
2.9. Structural analysis of LeuRS enzymes using Fourier transform infraredspectroscopy (FTIR)
A Bruker Tensor 27 Confocheck FTIR system with an Aquaspecattachment was used to determine the secondary structural charac-teristics of wild-type and mutant LeuRS enzymes [34]. Preparationof the FTIR included a N2 flush for 2 h at 29 atm and temperature sta-bilization at 30 °C by continuous circulation of H2O through theAquaspec. 20 mg/ml of purified enzyme was equilibrated at 30 °Cfor 30 min prior to injecting the sample into the Aquaspec. The vi-brational patterns of the amide I and amide II bonds within thepolypeptide structure were analyzed with OPUS software and thepercentage of α-helix and β-sheet content was quantified fromthe resulting spectrum [34]. A spectrum of absorbance versus wave-number (cm−1) was generated.
3. Results
E. coli LeuRS is an essential multi-domain metalloenzyme thataminoacylates tRNALeu with leucine. There are two catalytic sites, theleucylation site housed within the aminoacylation domain and the hy-drolytic deacylation site housed within the CP1 editing domain
(Fig. 1). The ZN-1 domain is highlighted in blue in Fig. 1 and is knownto play an essential structural role in stabilizing the Leu-Amp adenylate[12].We are interested in biochemically understanding the physical pa-rameters that enable the ZN-1 domain to stabilize the adenylate as wellas structural changes that may occur within the ZN-1 domain as LeuRSundergoes its catalytic cycle.
The ZN-1 domain has never been crystallographically visualized forany one LeuRS in both the aminoacylation and editing conformations.Thus, the complete structural and biochemical impacts that the ZN-1domain may play in LeuRS catalysis have remained unknown. There-fore, we computationally generated a homology model of E. coli LeuRSwhile in the editing conformation based on the crystal structure ofT. thermophilus LeuRS. We chose to generate this homology model be-cause the ZN-1 domain is incredibly dynamic in LeuRS and therefore isdifficult to crystallize. A successful crystal structure of the ZN-1 domainwas achieved for T. thermophilus LeuRS while in the editing conforma-tion, but not in the aminoacylation conformation [12]. Additionally, asuccessful crystal structure of the ZN-1 domain was achieved for E. coliLeuRS while in the aminoacylation conformation but not in the editingconformation [12]. Therefore, we developed this homology structureto piece together the current disjointed empirical data that has beenpublished so far so that we can achieve a clear idea of how the ZN-1 do-main impacts the catalytic cycle of E. coli LeuRS.
Fig. 1 shows the crystal structure of E. coli LeuRS while in the editingconformation (purple) [12] overlaid with the homology model ofE. coli LeuRS while in the editing conformation (yellow) with theZN-1 domains presented in blue. Based on these overlaid structuresand on the computational parameters previously discussed in theMaterials and methods section, we believe that the homologymodel that we have generated is an accurate depiction of E. coliLeuRS while in the editing conformation. There are some minorstructural perturbations within the auxiliary domains when compar-ing these two structures, but according to our computationalanalysis these minor shifts are all within the probable workingframework for the enzyme.
Visualization of the ZN-1 domain (blue) in both the aminoacylation(green) and editing (yellow) conformations (Fig. 2A) indicates that thedomain undergoes a dramatic structural change as LeuRS proceedsthrough its catalytic cycle. We propose that the ZN-1 domain acts likea “cap” that seals the aminoacylation active site to maintain a hydro-phobic environment for leucylation to effectively occur [35]. In orderfor LeuRS to transition to the editing conformation, the ZN-1 domainhas to shift away from the aminoacylation active site into an “open”conformation. We illustrate the “open” and “closed” conformations ofthe ZN-1 domain with the cartoon depiction in Fig. 2B. We computa-tionally determined that the magnitude to which the ZN-1 domainhas shifted up is 14°, which is a substantial structural shift for any onedomain. In Fig. 2C, we computationally overlaid E. coli LeuRS while inthe aminoacylation conformation [12] with our homology model ofE. coli LeuRS while in the editing conformation to show the dramaticmovement of the ZN-1 domain away from the aminoacylation activesite.
We have clearly shown that the ZN-1 domain undergoes dramaticstructural changes as LeuRS transitions from the aminoacylation to theediting conformations. The question we raised next was: How is thisstructural change achieved within this domain? Based on crystallo-graphic data, the ZN-1 domain within E. coli LeuRS has been shown tobind to one Zn2+ ion [12]which no one has biochemically characterizeduntil this study. We conducted an alanine scan of the ZN-1 domain toidentify residues that may play a role in chelating and stabilizing theZn2+ ion. We identified C159, C176 and C179 as the cysteine clusterthat makes-up the Zn2+ chelating pocket. The Zn2+ chelating pocketwithin E. coli LeuRS is unique when compared to other ZN-1 domainswithin the aaRS family. In Fig. 3A and B we show the Zn2+ chelatingpockets for E. coli LeuRS [12] and E. coli isoleucyl-tRNA synthetase(IleRS) [36], respectively. LeuRS chelates Zn2+ with only three cysteine
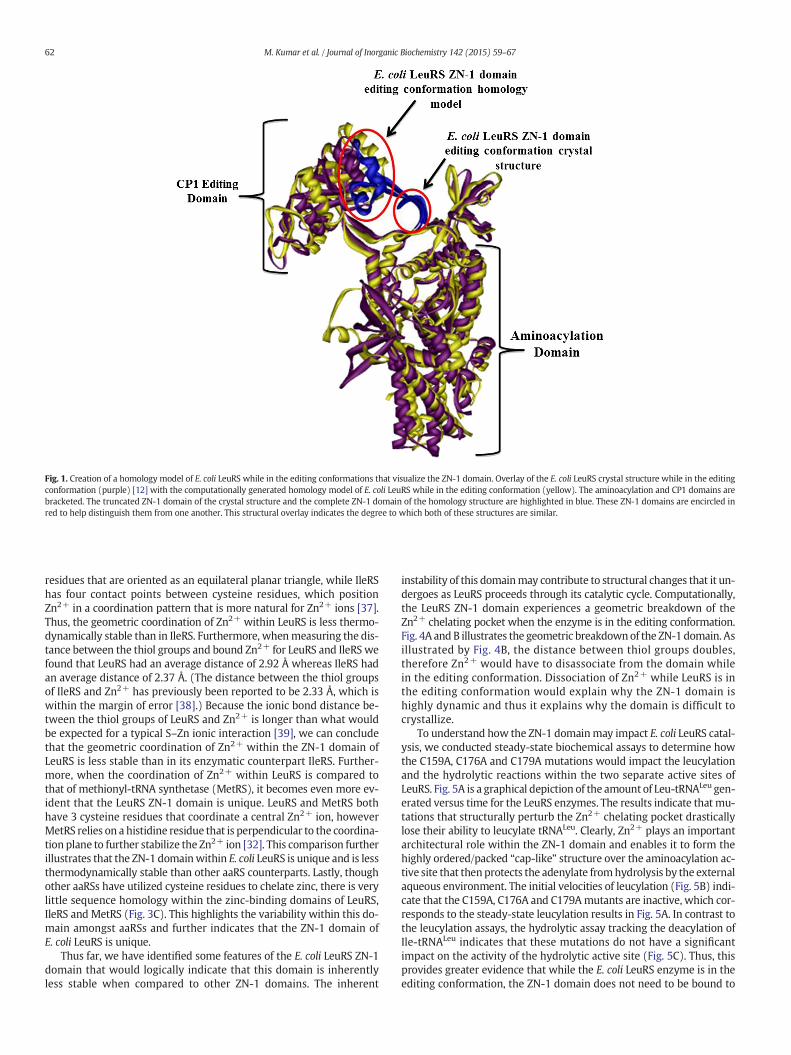
Fig. 1. Creation of a homology model of E. coli LeuRS while in the editing conformations that visualize the ZN-1 domain. Overlay of the E. coli LeuRS crystal structure while in the editingconformation (purple) [12] with the computationally generated homology model of E. coli LeuRS while in the editing conformation (yellow). The aminoacylation and CP1 domains arebracketed. The truncated ZN-1 domain of the crystal structure and the complete ZN-1 domain of the homology structure are highlighted in blue. These ZN-1 domains are encircled inred to help distinguish them from one another. This structural overlay indicates the degree to which both of these structures are similar.
62 M. Kumar et al. / Journal of Inorganic Biochemistry 142 (2015) 59–67
residues that are oriented as an equilateral planar triangle, while IleRShas four contact points between cysteine residues, which positionZn2+ in a coordination pattern that is more natural for Zn2+ ions [37].Thus, the geometric coordination of Zn2+ within LeuRS is less thermo-dynamically stable than in IleRS. Furthermore, whenmeasuring the dis-tance between the thiol groups and bound Zn2+ for LeuRS and IleRS wefound that LeuRS had an average distance of 2.92 Å whereas IleRS hadan average distance of 2.37 Å. (The distance between the thiol groupsof IleRS and Zn2+ has previously been reported to be 2.33 Å, which iswithin the margin of error [38].) Because the ionic bond distance be-tween the thiol groups of LeuRS and Zn2+ is longer than what wouldbe expected for a typical S–Zn ionic interaction [39], we can concludethat the geometric coordination of Zn2+ within the ZN-1 domain ofLeuRS is less stable than in its enzymatic counterpart IleRS. Further-more, when the coordination of Zn2+ within LeuRS is compared tothat of methionyl-tRNA synthetase (MetRS), it becomes even more ev-ident that the LeuRS ZN-1 domain is unique. LeuRS and MetRS bothhave 3 cysteine residues that coordinate a central Zn2+ ion, howeverMetRS relies on a histidine residue that is perpendicular to the coordina-tion plane to further stabilize the Zn2+ ion [32]. This comparison furtherillustrates that the ZN-1 domainwithin E. coli LeuRS is unique and is lessthermodynamically stable than other aaRS counterparts. Lastly, thoughother aaRSs have utilized cysteine residues to chelate zinc, there is verylittle sequence homology within the zinc-binding domains of LeuRS,IleRS and MetRS (Fig. 3C). This highlights the variability within this do-main amongst aaRSs and further indicates that the ZN-1 domain ofE. coli LeuRS is unique.
Thus far, we have identified some features of the E. coli LeuRS ZN-1domain that would logically indicate that this domain is inherentlyless stable when compared to other ZN-1 domains. The inherent
instability of this domainmay contribute to structural changes that it un-dergoes as LeuRS proceeds through its catalytic cycle. Computationally,the LeuRS ZN-1 domain experiences a geometric breakdown of theZn2+ chelating pocket when the enzyme is in the editing conformation.Fig. 4A and B illustrates the geometric breakdownof the ZN-1 domain. Asillustrated by Fig. 4B, the distance between thiol groups doubles,therefore Zn2+ would have to disassociate from the domain whilein the editing conformation. Dissociation of Zn2+ while LeuRS is inthe editing conformation would explain why the ZN-1 domain ishighly dynamic and thus it explains why the domain is difficult tocrystallize.
To understand how the ZN-1 domainmay impact E. coli LeuRS catal-ysis, we conducted steady-state biochemical assays to determine howthe C159A, C176A and C179A mutations would impact the leucylationand the hydrolytic reactions within the two separate active sites ofLeuRS. Fig. 5A is a graphical depiction of the amount of Leu-tRNALeu gen-erated versus time for the LeuRS enzymes. The results indicate that mu-tations that structurally perturb the Zn2+ chelating pocket drasticallylose their ability to leucylate tRNALeu. Clearly, Zn2+ plays an importantarchitectural role within the ZN-1 domain and enables it to form thehighly ordered/packed “cap-like” structure over the aminoacylation ac-tive site that thenprotects the adenylate fromhydrolysis by the externalaqueous environment. The initial velocities of leucylation (Fig. 5B) indi-cate that the C159A, C176A and C179Amutants are inactive, which cor-responds to the steady-state leucylation results in Fig. 5A. In contrast tothe leucylation assays, the hydrolytic assay tracking the deacylation ofIle-tRNALeu indicates that these mutations do not have a significantimpact on the activity of the hydrolytic active site (Fig. 5C). Thus, thisprovides greater evidence that while the E. coli LeuRS enzyme is in theediting conformation, the ZN-1 domain does not need to be bound to

Fig. 2. ZN-1 domain configuration changes through the catalytic cycle of E. coli LeuRS. A. E. coli LeuRS crystal structure in the aminoacylation complex (green) [12] with the ZN-1 domain(blue) positioned directly over the aminoacylation active site. An arrow points to the homologymodel of E. coli LeuRS in the editing conformation (yellow) with the ZN-1 domain in blue[28]. B. Cartoonmodels representing the position of the ZN-1 domain (blue)while E. coli LeuRS is in the aminoacylation (green) and editing (yellow) configurations, respectively. C. Struc-tural overlay of E. coli LeuRS crystal structure while in the aminoacylation conformation (green) with the E. coli LeuRS homology model (yellow) in the editing conformation. The ZN-1domains for each structure are highlighted in blue. The ZN-1 domain is encircled in red within the LeuRS aminoacylation complex and is indicated to swing up 14° by a red arrow onceLeuRS assumes the editing conformation.
63M. Kumar et al. / Journal of Inorganic Biochemistry 142 (2015) 59–67
Zn2+. The Apo-LeuRS enzyme has diminished hydrolytic activity, whichmay be due to the global loss of structure in the absence of both Mg2+
[35] and Zn2+.To quantitatively determine the loss of structure when LeuRS is no
longer able to bind to Zn2+, we analyzed the secondary structural char-acteristics of LeuRS using FTIR. One of the hypotheses thatwe have beenalluding to throughout this entire research project is that the Zn2+ ionplays an important architectural role within the ZN-1 domain and thatin its absence the ZN-1 domain cannot form a “cap” over theaminoacylation active site. The FTIR spectrum shows (Fig. 6) thatwild-type LeuRS has a strong Amide I peak signal, which quantitativelycorresponds to 38.3% α-helix content and 19.52% β-sheet content.These values are consistent with previously reported crystallographicdata [40]. When analyzing the structural features of the mutant LeuRSs,we consistently saw an overall drop of 1.0–2.0% in bothα-helical and β-sheet content for C159A, C176A and C179A. These values correspond tothe crystallographic data that indicates that the ZN-1 domain comprises5.47% of the total structural content of LeuRS and that the domain itselfhas two smallα-helical elements and small β-sheet stretches, thereforeseeing a dramatic drop in secondary structural features for the mutantLeuRS enzymes would be unrealistic [40]. However, as indicated bythe spectrum (Fig. 6) and the quantitative analysis we just outlined,we consistently saw a drop in secondary structural elements that is con-sistent with the size and secondary structural make-up of the ZN-1domain.
We further biochemically analyzed the Zn2+ chelating pocket ofE. coli LeuRS by determining the dissociation constant values of Zn2+
for each of the LeuRS enzymes (Fig. 7). We used 6 μM of LeuRS againstan increasing concentration gradient of EDTA and determined howmuch Zn2+ was present based on absorption signals. Both of the[LeuRS-Zn2+] and [Zn2+-EDTA] complexes have a 1:1 stoichiometricratio, which was subtracted from the background spectral signal thusenabling Kd values to be calculated. From the graph, it is clear thatC159A, C176A and C179A disassociate from Zn2+ much more readilythan wild-type LeuRS. Based on the reciprocal relationship between Ka
and Kd, we calculated the Kd values for wild-type, C159A, C176A andC179A to be: 1.3 × 10−9 M, 8.8 × 10−7 M, 9.4 × 10−7 M and 9.2 ×10−7 M, respectively. Based on these Kd values, it is clear that wild-type LeuRS binds more tightly to Zn2+ than does C159A, C176A orC179A.
4. Discussion
The E. coli LeuRS enzyme is an essential enzyme and is the modelLeuRS enzyme for bacterial systems. This enzyme has been the success-ful target of anti-microbial compounds and therefore plays an impor-tant role in future drug development research projects [40,41]. It isessential thatwe continue to conduct the basic scientific researchneces-sary to understand the catalytic cycle of this enzyme.With this researchproject we set out to structurally and biochemically analyze the ZN-1

Fig. 3. E. coli LeuRS's ZN-1 domain is inherently less stablewhen compared to its enzymatic counterparts. A. Coordination of Zn2+ by C159, C176 andC179within the ZN-1domainof LeuRS[12]. B. Coordination of Zn2+ by C181, C184, C389 and C392 within the ZN-1 domain of IleRS [36]. C. Sequence alignment of E. coli LeuRS, E. coli IleRS, E. coliMetRS, T. thermophilus LeuRS,T. thermophilus IleRS, T. thermophilusMetRS. There are no amino acids in the sequence alignment having 6–10 grades in the Unconserved–Conserved scale.
64 M. Kumar et al. / Journal of Inorganic Biochemistry 142 (2015) 59–67
domain as it pertains to LeuRS catalysis. The ZN-1 domain has structur-ally been shown to help stabilize Leu-AMP while the enzyme is in theaminoacylation conformation [12]. We set-out to determine how theZN-1 domain is able to stabilize Leu-AMP and in addition how thisdomain may structurally change in order to accommodate the editingconformation of LeuRS. The ZN-1 domain has never been visualizedfor any one LeuRS enzyme in both the aminoacylation and editing con-formations. Therefore, it has remained unclear how this domain impactsLeuRS catalysis.
Using computational tools and known crystal structures, we were,for the first time, able to visualize the ZN-1 domain of E. coli LeuRSthroughout its catalytic cycle. We computationally determined thatthe ZN-1 domain toggles 14° between the aminoacylation and editingconformations. We have shown that when the ZN-1 domain is in theaminoacylation conformation, the domain is packed tightly against theaminoacylation active site and has a rigid geometric coordination ofZn2+. However, when the ZN-1 domain is in the editing conformationthe geometric coordination of Zn2+ breaks down, which results in thedissociation of Zn2+ and a highly dynamic domain that cannot be effec-tively crystallized. We have demonstrated that the ZN-1 domain ofE. coli LeuRS is inherently less thermodynamically stable (due to lessstructural support elements) than ZN-1 domains within IleRS orMetRS. This inherent instability within the ZN-1 domain may allow for
the easier geometric breakdown of the Zn2+ chelating pocket as the en-zyme assumes the editing conformation.
We supported these claims with biochemical data that indicatedthat an intact ZN-1 domain is essential to maintain the functionality ofthe aminoacylation active site. However, if Zn2+ is disassociated fromthe ZN-1 domain, the enzyme can still carry out hydrolytic deacylationactivities. These biochemical findings support our argument that theZN-1 domain's architecture can change between the aminoacylationand editing states of the enzyme without affecting either active site'sfunctionality.
Structurally, we showed that mutations within the ZN-1 domainthat aberrantly affect the chelation of Zn2+ cause the entire ZN-1domain to collapse. Therefore, Zn2+ plays a key structural role andwe hypothesize that it acts as a molecular “switch” that allows theZN-1 domain to transition between the aminoacylation and editingconformations. We further demonstrated the importance of Zn2+
as a structural cornerstone of the ZN-1 domain by calculating theKd values of our various LeuRS enzymes. The Kd values indicate thatthere is a drastic change in the dissociation constant values whencomparing the wild-type and mutant LeuRS enzymes. This meansthat when the Zn2+ chelating pocket is disturbed, the LeuRS enzymecan easy disassociate from Zn2+. We believe that the wild-typeLeuRS can disassociate from Zn2+ under natural circumstances

Fig. 4. Geometric breakdown of the zinc chelating pocket within the ZN-1 domain of E. coli LeuRS. A. The geometric coordination of Zn2+ (purple) while LeuRS is in the aminoacylationconformation [12]. B. The geometric breakdown of the zinc chelating pocket within the ZN-1 domain of LeuRS. (The Zn2+ ion (purple) was artificially put in as a point of reference.)(For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
65M. Kumar et al. / Journal of Inorganic Biochemistry 142 (2015) 59–67
because of global structural changes that occur within the enzymedue to internal molecular “hinge” sites, in addition to the “sling-shot” movement of the tRNALeu 3′-acceptor stem during its translo-cation between the two active sites of LeuRS [20]. Our hypothesis isthat under the pressure of these global structural changes, the ZN-1domain breaks down the geometric coordination of the Zn2+
Fig. 5. Coordination of Zn2+within the ZN-1 domain is essential for leucylation, but is not essenZN-1 domain mutant LeuRSs. The reaction was carried out with 50 nM LeuRS, 4 μM tRNALeu trwild type and ZN-1 domain mutant LeuRSs. The reaction was carried out with 10 nM LeuRS, 4E. coli Ile-tRNALeu by wild type and mutant LeuRSs. Reaction conditions include 1.5 μM [3H]-Ile-(□); C179A (ο) and Apo-LeuRS, (◊). Error bars are present for all points and represent reaction
chelating pocket. This perturbation of the LeuRS structure allowsthe ZN-1 domain to move away from the aminoacylation active siteto allow the tRNALeu 3′-acceptor stem to gain access to the editing hy-drolytic active site. Once the Leu-tRNALeu product has disassociatedfrom LeuRS, the ZN-1 domain swings down to the aminoacylationactive site as it reassociates with Zn2+.
tial for deacylation ofmischarged tRNALeu products. A. Leucylation activity ofwild type andanscript, and 22 μM [3H]-leucine (167 μCi/ml). B. Initial velocities of leucylation activity ofμM tRNALeu transcript, and 22 μM [3H]-leucine (167 μCi/ml). C. Deacylation of mischargedtRNALeu and 5 nM enzyme. Symbols represent the following:WT, (■); C159A, (▲); C176,s that were repeated in triplicate.

Fig. 6. Zn2+ serves as the architectural cornerstone for the ZN-1 domain. Secondary structural analysis of wild-type andmutant LeuRSs using a Bruker Confocheck FTIR with an Aquaspecattachment. FTIR spectrum indicating a strong amide I peak that quantitatively corresponds to theα-helix and β-sheet content for each enzyme. Arrows point to the amide I peak of eachenzyme to identify their respective spectrums. Spectrum indicates the loss of secondary structural features for the C159A, C176A, C179A and Apo-LeuRS enzymes when compared to thewild-type enzyme.
66 M. Kumar et al. / Journal of Inorganic Biochemistry 142 (2015) 59–67
In conclusion, the ZN-1 domain of E. coli LeuRS is a unique domainthat is less thermodynamically stable than other ZN-1 domains fromclosely related aaRS enzymes. We have computationally shown thatthe ZN-1 domain experiences drastic structural changes that appear tobe dependent on the presence or absence of Zn2+. Biochemically, wehave supported this claim by demonstrating that Zn2+ is vital for theaminoacylation reaction to occur but is not needed for the deacylationreaction. These findings indicate that Zn2+ can act as a molecular“switch” that can associate or disassociate from the ZN-1 domain tohelp move forward the catalytic cycle of LeuRS. Furthermore, we haveshown that Zn2+ plays an important structural role within the ZN-1domain, which when present, the ZN-1 domain can assume theaminoacylation conformation but when absent the domain loses its ar-chitectural integrity and becomes highly dynamic.We hypothesize thatthe geometric breakdown of the ZN-1 domain allows charged tRNALeu
to translocate between the two active sites of LeuRS. Therefore, we be-lieve that further ZN-1 domain studiesmay reveal possible Zn2+ depen-dent tRNALeu translocation mechanisms.
Fig. 7. Zn2+more readily disassociates fromC159A, C176A andC179A LeuRSmutants thanit does from the wild-type LeuRS enzyme. Dissociation curve of Zn2+ from 6 μM of eachLeuRS enzyme against an increasing concentration gradient of EDTA. Absorption readingswere taken at 260 nm and background signals produced by LeuRS, EDTA and Mg2+
were subtracted from absorption readings. Kd values for Zn2+ were calculated fromthis dissociation curve, they are: wild-type, 1.3 × 10−9 M; C159A, 8.8 × 10−7 M; C176A,9.4 × 10−7 M and C179A, 9.2 × 10−7 M. Symbols represent the following: WT, (■);C159A, (▲); C176, (□); C179A (ο) and Apo-LeuRS, (◊). Error bars are present for all pointsand represent reactions that were repeated in triplicate.
Abbreviations
aaRS aminoacyl-tRNA synthetaseCP1 domain connective polypeptide 1 domainIleRS isoleucyl-tRNA synthetaseLeuRS leucyl-tRNA synthetaseMetRS methionyl-tRNA synthetaseLeu-AMP leucylated adenylateZN-1 domainzinc-binding domain 1ZN-2 domainzinc-binding domain 2
Acknowledgments
This work was supported by the Coastal Carolina University Collegeof Science. We thank Dr. Valdislav Gulis at Coastal Carolina Universityfor generously providing 3H-amino acid aswell as access to his scintilla-tion counter. We acknowledge Dr. Susan Martinis at the University ofIllinois, U–C, with whom a different but related project was initiated.
References
[1] C.R. Woese, G.J. Olsen, M. Ibba, D. Söll, Microbiol. Mol. Biol. Rev. 64 (2000) 202–236.[2] S. Cusack, Curr. Opin. Struct. Biol. 7 (1997) 881–889.[3] J.J. Burbaum, P. Schimmel, J. Biol. Chem. 266 (1991) 16965–16968.[4] G. Eriani, M. Delarue, O. Poch, J. Gangloff, D. Moras, Nature 347 (1990) 203–206.[5] T. Hendrickson, P. Schimmel, J. Lapointe, L. Brakier-Gingras, Translation Mecha-
nisms, Eurekah.com and Kluwer Academic/Plenum Publishers, Georgetown, TXand New York, NY, 2003. 34–64.
[6] P. Schimmel, Protein science: a publication of the protein society, Protein Sci. 17(2008) 1643–1652.
[7] W. Yan, M. Tan, G. Eriani, E.D. Wang, Nucleic Acids Res. 41 (2013) 4988–4998.[8] M. Jasin, L. Regan, P. Schimmel, Nature 306 (1983) 441–447.[9] Y. Pham, L. Li, A. Kim, O. Erdogan, V. Weinreb, G.L. Butterfoss, B. Kuhlman, C.W.
Carter Jr., Mol. Cell 25 (2007) 851–862.[10] M. Tan, B. Zhu, R.J. Liu, X. Chen, X.L. Zhou, E.D. Wang, Biochem. J. 449 (2013)
123–131.[11] N. Cvetesic, J.J. Perona, I. Gruic-Sovulj, J. Biol. Chem. 287 (2012) 25381–25394.[12] A. Palencia, T. Crepin, M.T. Vu, T.L. Lincecum Jr., S.A. Martinis, S. Cusack, Nat. Struct.
Mol. Biol. 19 (2012) 677–684.[13] J.M. Han, M.J. Lee, S.G. Park, S.H. Lee, E. Razin, E.C. Choi, S. Kim, J. Biol. Chem. 281
(2006) 38663–38667.[14] S.G. Park, K.L. Ewalt, S. Kim, Trends Biochem. Sci. 30 (2005) 569–574.[15] J. Liu, Y. Gagnon, J. Gauthier, L. Furenlid, P.J. L'Heureux, M. Auger, O. Nureki, S.
Yokoyama, J. Lapointe, J. Biol. Chem. 270 (1995) 15162–15169.[16] E. Glasfeld, J.A. Landro, P. Schimmel, Biochemistry 35 (1996) 4139–4145.

67M. Kumar et al. / Journal of Inorganic Biochemistry 142 (2015) 59–67
[17] S. Bilokapic, T. Maier, D. Ahel, I. Gruic-Sovulj, D. Söll, I. Weygand-Durasevic, N. Ban,EMBO J. 25 (2006) 2498–2509.
[18] S. Cusack, A. Yaremchuk, M. Tukalo, EMBO J. 19 (2000) 2351–2361.[19] K.W. Lee, J.M. Briggs, Proteins 54 (2004) 693–704.[20] R.A. Hellmann, S.A. Martinis, J. Biol. Chem. 284 (2009) 11478–11484.[21] J.R. Sampson, O.C. Uhlenbeck, Proc. Natl. Acad. Sci. U. S. A. 85 (1988) 1033–1037.[22] J.D. Puglisi, I. Tinoco Jr., Methods Enzymol. 180 (1989) 304–325.[23] M. Hartlein, D. Madern, Nucleic Acids Res. 15 (1987) 10199–10210.[24] R.S. Mursinna, S.A. Martinis, J. Am. Chem. Soc. 124 (2002) 7286–7287.[25] A.P. Mascarenhas, S.A. Martinis, Biochemistry 47 (2008) 4808–4816.[26] A.A. Schreier, P.R. Schimmel, Biochemistry 11 (1972) 1582–1589.[27] R.S. Mursinna, T.L. Lincecum Jr., S.A. Martinis, Biochemistry 40 (2001) 5376–5381.[28] M. Tukalo, A. Yaremchuk, R. Fukunaga, S. Yokoyama, S. Cusack, Nat. Struct. Mol. Biol.
12 (2005) 923–930.[29] K. Arnold, L. Bordoli, J. Kopp, T. Schwede, Bioinformatics 22 (2006) 195–201.[30] B. Wallner, A. Elofsson, Protein science: a publication of the protein society, Protein
Sci. 12 (2003) 1073–1086.[31] J.Y. Li, L.L. Chen, Y.M. Cui, Q.L. Luo, J. Li, F.J. Nan, Q.Z. Ye, Biochem. Biophys. Res.
Commun. 307 (2003) 172–179.
[32] O. Nureki, T. Kohno, K. Sakamoto, T. Miyazawa, S. Yokoyama, J. Biol. Chem. 268(1993) 15368–15373.
[33] H. Fredholm, Sven. Kem. Tidskr. 44 (1932) 44–48.[34] J. Kong, S. Yu, Acta Biochim. Biophys. Sin. 39 (2007) 549–559.[35] J.G. Arnez, D. Moras, Trends Biochem. Sci. 22 (1997) 211–216.[36] T. Nakama, O. Nureki, S. Yokoyama, J. Biol. Chem. 276 (2001) 47387–47393.[37] S. De, S.M. Ali, A. Ali, V.G. Gaikar, Phys. Chem. Chem. Phys. 11 (2009) 8285–8294.[38] J.A. Landro, E. Schmidt, P. Schimmel, D.L. Tierney, J.E. Penner-Hahn, Biochemistry 33
(1994) 14213–14220.[39] P.L. Hayes, B.L. Lytle, B.F. Volkman, F.C. Peterson, Protein Sci. 17 (2008) 571–576.[40] V. Hernandez, T. Crepin, A. Palencia, S. Cusack, T. Akama, S.J. Baker, W. Bu, L. Feng, Y.R.
Freund, L. Liu, M. Meewan, M. Mohan, W. Mao, F.L. Rock, H. Sexton, A. Sheoran, Y.Zhang, Y.K. Zhang, Y. Zhou, J.A. Nieman, M.R. Anugula, M. Keramane el, K. Savariraj,D.S. Reddy, R. Sharma, R. Subedi, R. Singh, A. O'Leary, N.L. Simon, P.L. De Marsh, S.Mushtaq, M. Warner, D.M. Livermore, M.R. Alley, J.J. Plattner, Antimicrob. AgentsChemother. 57 (2013) 1394–1403.
[41] F.L. Rock, W. Mao, A. Yaremchuk, M. Tukalo, T. Crepin, H. Zhou, Y.K. Zhang, V.Hernandez, T. Akama, S.J. Baker, J.J. Plattner, L. Shapiro, S.A. Martinis, S.J. Benkovic,S. Cusack, M.R. Alley, Science 316 (2007) 1759–1761.




















