
XB130, a tissue-specific adaptor protein that couples the RET/PTC oncogenic kinase to PI 3-kinase...
13
ORIGINAL ARTICLE XB130, a tissue-specific adaptor protein that couples the RET/PTC oncogenic kinase to PI 3-kinase pathway M Lodyga 1,4 , V De Falco 2,4 , X-h Bai 1 , A Kapus 3 , RM Melillo 2 , M Santoro 2 and M Liu 1,3 1 Division of Cellular and Molecular Biology, Toronto General Research Institute, University Health Network, Toronto, Ontario, Canada; 2 Dipartimento di Biologia e Patologia Cellulare e Molecolare, Instituto di Endocrinologia ed Oncologia Sperimentale del CNR ‘G. Salvatore’, Naples, Italy and 3 Department of Surgery, University of Toronto, Toronto, Ontario, Canada XB130 is a recently cloned 130kDa-adaptor protein and Src kinase substrate, structurally similar to actin-filament- associated protein. Here we show that XB130 is pre- dominantly expressed in the thyroid. Given that XB130 is a thyroid-specific tyrosine kinase substrate, we asked whether it is targeted by RET/PTC, a genetically rearranged, constitutively active, thyroid-specific tyrosine kinase that plays a pathogenic role in papillary thyroid cancer. RET/PTC induced robust tyrosine phosphory- lation of XB130, which promoted its subsequent associa- tion with the p85a subunit of phosphatidylinositol 3-kinase (PI 3-kinase). We identified tyrosine 54 of XB130 as the major target of RET/PTC-mediated phosphorylation and a critical binding site for the SH2 domains of p85a. Importantly, downregulation of XB130 in TPC1 papillary thyroid cancer cells, harboring the RET/PTC1 kinase, strongly reduced Akt activity without altering ERK1/2 phosphorylation, and concomitantly inhibited cell-cycle progression and survival in suspension. In conclusion, XB130 is a novel substrate of the RET/PTC kinase that links RET/PTC signaling to PI 3-kinase activation, and thereby plays an important role in sustaining proliferation and survival of thyroid tumor cells. Oncogene (2009) 28, 937–949; doi:10.1038/onc.2008.447; published online 8 December 2008 Keywords: adaptor protein; protein tyrosine phospho- rylation; thyroid cancer; RET/PTC; phosphatidylinositol 3-kinase; PKB/AKT Introduction During our recent studies aimed at the characterization of the actin filament-associated protein (AFAP; Flynn et al., 1993; Lodyga et al., 2002; Han et al., 2004), we cloned a novel 130 kDa protein, referred to as XB130 (Xu et al., 2007). The human xb130 gene is localized on chromosome 10q25.3, and encodes an 818 amino-acid protein that shares 35% similarity with AFAP. The overall structure of XB130 is highly suggestive of a complex adaptor protein as it is comprised of various protein–protein and lipid–protein interaction motifs. Specifically, the N-terminal region of XB130 includes several potential tyrosine phosphorylation sites and proline-rich sequences that might interact with SH2- and SH3-domain-containing proteins, respectively. The middle portion harbors two pleckstrin homology domains that may target proteins to cellular membranes through interactions with specific phospholipids (Haslam et al., 1993; Harlan et al., 1994), whereas the C-terminal region contains a coiled-coil domain, which might be involved in protein oligomerization and DNA binding. Our previous studies have indeed implicated XB130 as a likely substrate and regulator of tyrosine kinase-mediated signaling (Xu et al., 2007). Notably, XB130 was shown to associate with c-Src, and the overexpression of these proteins in COS-7 cells resulted in increased XB130 tyrosine phosphorylation, together with enhanced Src activity. We found that XB130 expression is restricted to thyroid follicular cells, whereas most other tissues (except the spleen) showed only marginal expression. We asked whether XB130 might be an adaptor particularly involved in thyroid-specific tyrosine kinase pathways. Dysregulation in tyrosine kinase signaling is an essential feature in the development of thyroid cancer. RET/PTC rearrangements are a genetic hall- mark of papillary thyroid cancer (PTC), the most prevalent type of thyroid cancer in human (Grieco et al., 1990). These rearrangements lead to the formation of chimeric and constitutively active versions of the proto-oncogene RET tyrosine kinase (Takahashi et al., 1988; Arighi et al., 2005). RET/PTC exhibits transform- ing ability, affecting differentiation, mitogenic and metastatic potential (Santoro et al., 1994b; Mai et al., 2001; Knauf et al., 2003). Indeed, transgenic mice with thyroid-targeted expression of RET/PTC1 develop carcinomas resembling those seen in human PTC (Santoro et al., 1996), suggesting that RET/PTC plays a disease-inducing role in thyroid cancer. Received 13 May 2008; revised 1 October 2008; accepted 1 November 2008; published online 8 December 2008 Correspondence: Dr M Liu, University Health Network Toronto General Hospital, Room: TMDT 2-814, 101 College Street, Toronto, Ontario, Canada M5G 1L7. E-mail: [email protected] or Dr M Santoro, Dipartimento di Biologia e Patologia Cellulare e Molecolare, Universita Federico II di Napoli, via S Pansini 5, 80131 Naples, Italy. E-mail: [email protected] 4 These authors contributed equally to this work. Oncogene (2009) 28, 937–949 & 2009 Macmillan Publishers Limited All rights reserved 0950-9232/09 $32.00 www.nature.com/onc
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of XB130, a tissue-specific adaptor protein that couples the RET/PTC oncogenic kinase to PI 3-kinase...

ORIGINAL ARTICLE
XB130, a tissue-specific adaptor protein that couples the RET/PTC
oncogenic kinase to PI 3-kinase pathway
M Lodyga1,4, V De Falco2,4, X-h Bai1, A Kapus3, RM Melillo2, M Santoro2 and M Liu1,3
1Division of Cellular and Molecular Biology, Toronto General Research Institute, University Health Network, Toronto, Ontario,Canada; 2Dipartimento di Biologia e Patologia Cellulare e Molecolare, Instituto di Endocrinologia ed Oncologia Sperimentale delCNR ‘G. Salvatore’, Naples, Italy and 3Department of Surgery, University of Toronto, Toronto, Ontario, Canada
XB130 is a recently cloned 130 kDa-adaptor protein andSrc kinase substrate, structurally similar to actin-filament-associated protein. Here we show that XB130 is pre-dominantly expressed in the thyroid. Given that XB130 is athyroid-specific tyrosine kinase substrate, we askedwhether it is targeted by RET/PTC, a geneticallyrearranged, constitutively active, thyroid-specific tyrosinekinase that plays a pathogenic role in papillary thyroidcancer. RET/PTC induced robust tyrosine phosphory-lation of XB130, which promoted its subsequent associa-tion with the p85a subunit of phosphatidylinositol 3-kinase(PI 3-kinase). We identified tyrosine 54 of XB130 as themajor target of RET/PTC-mediated phosphorylation anda critical binding site for the SH2 domains of p85a.Importantly, downregulation of XB130 in TPC1 papillarythyroid cancer cells, harboring the RET/PTC1 kinase,strongly reduced Akt activity without altering ERK1/2phosphorylation, and concomitantly inhibited cell-cycleprogression and survival in suspension. In conclusion,XB130 is a novel substrate of the RET/PTC kinase thatlinks RET/PTC signaling to PI 3-kinase activation, andthereby plays an important role in sustaining proliferationand survival of thyroid tumor cells.Oncogene (2009) 28, 937–949; doi:10.1038/onc.2008.447;published online 8 December 2008
Keywords: adaptor protein; protein tyrosine phospho-rylation; thyroid cancer; RET/PTC; phosphatidylinositol3-kinase; PKB/AKT
Introduction
During our recent studies aimed at the characterizationof the actin filament-associated protein (AFAP; Flynn
et al., 1993; Lodyga et al., 2002; Han et al., 2004), wecloned a novel 130 kDa protein, referred to as XB130(Xu et al., 2007). The human xb130 gene is localized onchromosome 10q25.3, and encodes an 818 amino-acidprotein that shares 35% similarity with AFAP. Theoverall structure of XB130 is highly suggestive of acomplex adaptor protein as it is comprised of variousprotein–protein and lipid–protein interaction motifs.Specifically, the N-terminal region of XB130 includesseveral potential tyrosine phosphorylation sites andproline-rich sequences that might interact with SH2-and SH3-domain-containing proteins, respectively. Themiddle portion harbors two pleckstrin homologydomains that may target proteins to cellular membranesthrough interactions with specific phospholipids(Haslam et al., 1993; Harlan et al., 1994), whereas theC-terminal region contains a coiled-coil domain, whichmight be involved in protein oligomerization and DNAbinding. Our previous studies have indeed implicatedXB130 as a likely substrate and regulator of tyrosinekinase-mediated signaling (Xu et al., 2007). Notably,XB130 was shown to associate with c-Src, and theoverexpression of these proteins in COS-7 cells resultedin increased XB130 tyrosine phosphorylation, togetherwith enhanced Src activity.
We found that XB130 expression is restricted tothyroid follicular cells, whereas most other tissues(except the spleen) showed only marginal expression.We asked whether XB130 might be an adaptorparticularly involved in thyroid-specific tyrosine kinasepathways. Dysregulation in tyrosine kinase signaling isan essential feature in the development of thyroidcancer. RET/PTC rearrangements are a genetic hall-mark of papillary thyroid cancer (PTC), the mostprevalent type of thyroid cancer in human (Griecoet al., 1990). These rearrangements lead to the formationof chimeric and constitutively active versions of theproto-oncogene RET tyrosine kinase (Takahashi et al.,1988; Arighi et al., 2005). RET/PTC exhibits transform-ing ability, affecting differentiation, mitogenic andmetastatic potential (Santoro et al., 1994b; Mai et al.,2001; Knauf et al., 2003). Indeed, transgenic mice withthyroid-targeted expression of RET/PTC1 developcarcinomas resembling those seen in human PTC(Santoro et al., 1996), suggesting that RET/PTC playsa disease-inducing role in thyroid cancer.
Received 13 May 2008; revised 1 October 2008; accepted 1 November2008; published online 8 December 2008
Correspondence: Dr M Liu, University Health Network TorontoGeneral Hospital, Room: TMDT 2-814, 101 College Street, Toronto,Ontario, Canada M5G 1L7.E-mail: [email protected] or Dr M Santoro, Dipartimento diBiologia e Patologia Cellulare e Molecolare, Universita Federico II diNapoli, via S Pansini 5, 80131 Naples, Italy.E-mail: [email protected] authors contributed equally to this work.
Oncogene (2009) 28, 937–949& 2009 Macmillan Publishers Limited All rights reserved 0950-9232/09 $32.00
www.nature.com/onc

Secondary to RET/PTC rearrangements, theC-terminal tyrosine kinase domain of RET fuses withthe end of one of several unrelated genes (Grieco et al.,1990). RET/PTC1 (the H4-RET fusion) and RET/PTC3(the NCOA4-RET fusion) are the most prevalentvariants. This rearrangement has three major conse-quences: the kinase becomes constitutively active as thefusion partner provides a domain capable of sponta-neous dimerization and autophosphorylation; the chi-mera loses its intramembrane localization and thusresides in the cytosol; and finally, the tissue expressionpattern changes, as the RET fusion partners areable to drive the expression of the rearranged RETkinase in the follicular epithelial cells of the thyroid(Arighi et al., 2005).
RET activation results in receptor oligomerization,autophosphorylation and the subsequent recruitment ofproteins containing phosphotyrosine-binding domains.Many of these (for example, Grb 7/10, Src, PLCg, Grb2)bind to individual phosphotyrosine residues, whereasother signaling molecules share tyrosine 1062 (Y1062) astheir common docking site (Hayashi et al., 2000; Arighiet al., 2005). When phosphorylated, Y1062 acts as thebinding site for several protein tyrosine binding (PTB)-containing proteins, including Shc, IRS1/2, FRS2 andDOK1/4/5. Binding to Shc and FRS2 mediates recruit-ment of Grb2/SOS complexes, which in turn stimulatethe Ras/RAF/ERK pathway. Y1062 also mediates RETbinding to Gab (Grb2-associated binding protein)family adaptors (Gab1 and Gab2) and IRS1/2, largedocking proteins recruiting multiple SH2 or SH3domain-containing proteins, including phosphatidylino-sitol 3-kinase (PI 3-kinase; Hayashi et al., 2000; Arighiet al., 2005). Both the RAS/RAF/MAPK and the PI 3-kinase/Akt pathways play a central role in RET/PTCsignaling and contribute to the ensuing transformation(Santoro et al., 1993; Miyagi et al., 2004; Jung et al.,2005; Melillo et al., 2005). Microarray studies in humanthyroid cancer indicate that RET/PTC rearrangement isassociated with expression of genes downstream of PI 3-kinase (Giordano et al., 2005). Further, in FRTL-5 ratthyroid cells overexpression of RET/PTC3 was sug-gested to preferentially activate the PI 3-kinase over theMAPK pathway (Miyagi et al., 2004). The p85regulatory subunit of PI 3-kinase contains two SH2domains (Escobedo et al., 1991), which recognizephosphotyrosine residues in the contexts of a pYXXMmotif. As RET does not have such sites, itsinteraction with PI 3-kinase is mediated by dockingproteins, of which Gab1 (recruited to RET in complexwith Grb2, Shc and ShcC; Besset et al., 2000; Maedaet al., 2004; De Falco et al., 2005), IRS-1 (Melillo et al.,2001) and IRS-2 (Miyagi et al., 2004) have beenimplicated.
Here we show that RET/PTC induces phosphoryla-tion of XB130, predominantly targeting tyrosine 54within its YXXM motif. The RET/PTC-induced phos-phorylation results in association between XB130 andp85 subunit of PI 3-kinase, and this interaction plays amajor role in the subsequent Akt activation, cellproliferation and survival.
Results
Thyroid specific expression of XB130We have demonstrated that XB130 qualifies asa multidomain adaptor protein (Figure 1a), and, ina heterologous expression system, can function as asubstrate and regulator of Src tyrosine kinase (Xu et al.,2007). We determined XB130’s tissue distribution usingnorthern blot analysis. We found a single transcript ofapproximately 4 kb, which was abundant in the thyroidand spleen, but was absent or barely detectable in othertissues (Figure 1b). Immunohistochemistry staining ofthyroid tissue with a specific monoclonal antibodyverified the presence of XB130 protein in thyroidfollicular cells (Figure 1c). No staining was detectedwhen the primary antibody was replaced with isotype-specific IgG2a (Figure 1d). The expression of XB130was also found in human papillary thyroid carcinoma;approximately half of PTC samples stained, containedlevels of expression comparable to that of normalthyroid tissue (Figure 1e), whereas the remainingcontained weaker staining (Figure 1f). These findingsestablish XB130 as a thyroid protein, highly expressed innormal follicular cells and in follicular-cell-derivedtumors.
XB130 is a substrate of RET/PTCHaving verified that XB130 is a thyroid-specific adaptorand potential tyrosine kinase substrate expressed inpapillary thyroid carcinoma, we asked whether it mightbe targeted by RET/PTC (Figure 2a). We transientlytransfected HEK293 cells with myc-tagged XB130 alongwith the most common forms of the RET kinaserearrangement, RET/PTC1 (Pierotti et al., 1992) andRET/PTC3 (Santoro et al., 1994a). Immunoprecipita-tion of XB130 followed by anti-phosphotyrosine im-munoblotting revealed that each form of RET/PTCinduced strong tyrosine phosphorylation of XB130(Figure 2b). This effect was dependent on the kinaseactivity of RET/PTC as the cotransfection with akinase-dead (K�) mutant containing the K284M pointmutation (K758M in full-length RET; Melillo et al.,2001) in RET/PTC3 failed to invoke XB130 phospho-rylation (Figure 2c). Coexpression of XB130 andRET/PTC3 in HEK293 cells did not affect tyro-sine phosphorylation of RET/PTC3 (SupplementaryFigure 1).
We next asked whether autophosphorylation sites ofRET/PTC might be involved in RET/PTC-inducedXB130 tyrosine phosphorylation. We cotransfected aRET/PTC3 (4YF) mutant in which four identifiedautophosphorylation sites (corresponding to residuesY826, Y1015, Y1029, Y1062 in the wild-type RET) hadbeen mutated to phenylalanine. These substitutions didnot decrease the RET/PTC-triggered tyrosine phosphor-ylation of XB130 (Figure 2d). This observation impliesthat although these tyrosines are key docking sites for avariety of direct substrates or adaptors, they are notrequired for XB130 phosphorylation. To verify whetherRET/PTC and XB130 can form a protein complex, we
XB130, a novel adaptor in RET/PTC signalingM Lodyga et al
938
Oncogene

coexpressed myc-tagged RET/PTC3 with His-taggedXB130 in HEK293 cells. XB130 can coprecipitate withRET/PTC; moreover the interaction between these twoproteins critically depends on the kinase activity ofRET/PTC because the kinase-dead version fails tophosphorylate and form a complex with XB130(Figure 2e). The interaction was further confirmed bythe reverse immunoprecipitation of XB130 with anti-Hisantibody and immunoblotting for RET/PTC with anti-myc antibody (data not shown).
To determine the role of Src in the tyrosinephosphorylation of XB130 in the presence of RET/
PTC3, we coexpressed Src kinase dead mutant, as adominant negative inhibitor of Src PTK (Iavarone et al.,2006), along with XB130 and RET/PTC3 in HEK293cells. The presence of Src K� did not reduce RET/PTC3-induced tyrosine phosphorylation of XB130, nordid it affect signaling downstream of RET/PTC, Aktphosphorylation (Supplementary Figures 2a and b).Further XB130 tyrosine phosphorylation was detectedwhen we coexpressed XB130 along with RET/PTC3 inSYF mouse embryonic fibroblasts (SupplementaryFigure 3), where three major Src family members (Src,Yes and Fyn) have been genetically knocked out.
Figure 1 Expression of XB130 in thyroid and papillary thyroid cancer. (a) Schematic representation of the XB130 protein structure.(b) XB130 mRNA is highly expressed in human thyroid and spleen as detected by Northern blotting with XB130 cDNA fragmentencompassing nucleotides 811–3750. One major transcript of approximately 4 kb was observed. (c) XB130 protein is abundantlyexpressed in human thyroid follicular cells as revealed by immunohistostaining with an anti-XB130 monoclonal antibody. (d) MouseIgG2a was used as a negative control for staining. (e and f) Over 100 papillary thyroid carcinoma samples were stained for XB130expression. Approximately 50% of samples showed expression comparable to that of normal thyroid tissue, remaining samplespossessed relatively weaker staining in the tumor vs the surrounding thyroid tissue, as indicated by the arrow in panel f.
XB130, a novel adaptor in RET/PTC signalingM Lodyga et al
939
Oncogene

Tyrosine phosphorylation of XB130 was further en-hanced when Src was reintroduced into these cells.These observations imply that although Src may affect
the tyrosine phosphorylation of XB130 it is notnecessary for RET/PTC-mediated tyrosine phosphory-lation of XB130.
XB130, a novel adaptor in RET/PTC signalingM Lodyga et al
940
Oncogene

RET/PTC-induced phosphorylation targets Y54in XB130The N-terminal region of XB130 contains severalputative tyrosine phosphorylation sites, and our pre-vious studies have shown that this region serves as atarget for Src overexpression-induced phosphorylation.To determine whether the N terminus of XB130 isrequired for RET/PTC-dependent phosphorylation, andto identify critical target residues, we utilized anN-terminal deletion mutant XB130DN (deletion ofresidues 2–168), as well as several point mutations ofkey tyrosines present in this region. Specifically wesubstituted tyrosines 54 (DEEYIYMNKV), 124(PEGYYEEAEP) or 161 (SAPYQWPSPE) with phenylalanineto preclude phosphorylation of these sites (Figure 3a).HEK293 cells were transfected with full-length ormutant XB130 constructs either alone or in combinationwith RET/PTC. Cell lysates were immunoprecipitatedwith XB130 antibody followed by immunoblottingusing anti-phosphotyrosine and anti-XB130 antibodies.Figure 3b indicates that deletion of the N-terminalregion almost completely abolished the RET/PTC-provoked tyrosine phosphorylation of XB130. Experi-ments conducted to probe candidate tyrosine residueswithin this region revealed that point mutations Y124Fand Y161F did not reduce the level of RET/PTC-triggered XB130 tyrosine phosphorylation, as comparedto wild type. In contrast, the Y54F mutant displayed anapproximately 50% decrease in its tyrosine phosphor-ylation, indicating that Y54 is a major although notexclusive target of RET/PTC-mediated tyrosine phos-phorylation (Figure 3b).
XB130 associates with p85 in a RET/PTC-regulatedmannerThe phosphorylated YXXM motif is a specific bindingsite for the SH2 domain of the p85 regulatory subunit ofPI 3-kinase (Backer et al., 1992; Carpenter et al., 1993;Rordorf-Nikolic et al., 1995). As XB130 tyrosine 54resides in a context of YIYM sequence, which fits therequired profile, XB130 is a candidate-binding partnerof p85. To address this possibility we carried out bindingexperiments both in vivo and in vitro. HEK293 cells weretransiently transfected with myc-tagged XB130 alone orin combination with RET/PTC, XB130 was immuno-precipitated through the tag, and the precipitates were
probed for the presence of p85. A small amount of p85was coprecipitated with XB130 in HEK293 cells, andthe presence of RET/PTC robustly enhanced theassociation between p85 and XB130 (Figure 4a). p85contains two SH2 domains, either or both of them canserve as potential target(s) for XB130 recruitment. Togain insight into the domain requirement of thisassociation, we performed a pull-down assay, using N-terminal or C-terminal GST-SH2 fusion protein of p85as bait. The binding assay was carried out with cellextracts derived from HEK293 cells transiently trans-fected with myc-tagged XB130 alone or in combinationwith RET/PTC. Figure 4b shows that XB130 was ableto bind to both the N-terminal and the C-terminal SH2domain of p85. Importantly, the binding was greatlypotentiated in the presence of RET/PTC. To determinethe affinity of XB130 to the a- or the b-isoform of p85,we transfected HEK 293 cells with XB130 and/or RET/PTC3. Both a- and b-p85 were detected by directwestern blotting in cell lysates. However, XB130preferentially binds to p85a, but not to p85b (Supple-mentary Figure 4).
Next we checked whether the XB130–p85 interactionis indeed dependent on the phosphorylation status ofXB130. We performed a pull-down assay with SH2domains of p85 and cell extracts derived from HEK293cells transiently cotransfected with RET/PTC andXB130 mutants (DN, Y54F, Y124F, Y161F). As shownin Figure 4c, the N terminus of XB130 is essential for theinteraction between XB130 and p85, as in the absence ofthis region the interaction is almost completely lost. Inagreement with our results showing a 50% decrease inRET/PTC-mediated tyrosine phosphorylation of theY54F XB130 mutant compared to wild type, thismutant exhibited significant reduction in the interactionwith both the N- and C-terminal SH2 domain of p85 inRET/PTC expressing cells. Mutations of Y124F andY161F did not noticeably impact the interaction(Figure 4c). To substantiate the importance of Y54 inthe XB130–p85 interaction, we synthesized two17-amino-acid peptides based on the sequence surround-ing Y54: one harbors the nonphosphorylated tyrosineresidue (p54), whereas the other contains a correspond-ing pY (pp54), mimicking the Y54-phosphorylatedmotif within XB130. As shown in Figure 4d, thephosphopeptide (pp54) inhibited the interactionbetween the SH2 domain of p85 and XB130 in
Figure 2 XB130 is a substrate of RET/PTC kinase. (a) Schematic representation of RET and RET/PTC proteins. SP, signal peptide;EC, extracellular ligand-binding domain; Cys, cysteine-rich region; TM, transmembrane domain; JX, juxtamembrane domain; TK,tyrosine kinase domain. The RET/PTC oncoproteins contain tyrosine kinase domain (TK) of RET and a heterologous domain (X).The catalytic lysine (K758) and major RET autophosphorylation sites (Y826, Y1015, Y1029, Y1062) are indicated on the wild-typeversion of RET. (b) Tyrosine phosphorylation of XB130 is induced by RET/PTC1 or RET/PTC3. HEK293 cells were cotransfectedwith cDNAs encoding myc-tagged XB130 and RET/PTC1 or RET/PTC3. Immunoprecipitation of XB130 using a myc antibody wasperformed followed by western blotting for XB130 with anti-myc and anti-pY antibodies. Total lysates were developed with RETantibody to monitor the transfection level of the RET/PTC (RET/PTC1: 50 kDa; RET/PTC3: 70 kDa). (c) Tyrosine phosphorylationof XB130 depends on RET/PTC kinase activity. XB130 was expressed alone, or coexpressed with RET/PTC3 or a kinase dead (K�)mutant. (d) Tyrosine phosphorylation of XB130 does not depend on RET/PTC autophosphorylation sites (Y352, Y541, Y555, Y588corresponding to Y826, Y1015, Y1029 and Y1062 in full-length RET, respectively). (e) XB130 and RET/PTC3 can form a proteincomplex. HEK293 cells were transfected with cDNAs encoding His-tagged XB130 alone or in combination with myc-tagged RET/PTC3 (or its K� mutant). Cell lysates were immunoprecipitated with anti-myc antibody and blotted for anti-His, anti-pY or anti-mycantibodies. The fast migration of PTC3K� is most likely due to less tyrosine phosphorylation of the protein.
XB130, a novel adaptor in RET/PTC signalingM Lodyga et al
941
Oncogene

a concentration-dependent manner, fully abolishing thebinding at 10 mM. In contrast the nonphosphorylated(p54) peptide failed to affect the association even at50 mM. Collectively these data demonstrates that RET/PTC mediates phosphorylation of XB130 at Y54, andthis phosphorylation is required for the efficient inter-action between XB130 and the p85 subunit of PI3-kinase.
Next we examined whether the RET/PTC-dependentphosphorylation of XB130 impacts the potential inter-action between XB130 and p85 in a natural setting, thatis, where these proteins are endogenously expressed. We
used TPC1 thyroid papillary carcinoma cells carryingthe RET/PTC1 rearrangement, and treated them with aspecific RET kinase inhibitor, ZD6474 (Carlomagnoet al., 2002), to suppress intrinsic RET/PTC activity.Although in untreated TPC1 cells, p85 was constitu-tively associated with XB130, ZD6474-treated cellsfailed to associate with endogenous XB130, tested bycoimmunoprecipitation (Supplementary Figure 5a), oreither the N-terminal or C-terminal SH2 fusion proteinsof p85, as verified by GST pull-down assay (Supple-mentary Figure 5b). Following 24 h of ZD6474 treat-ment we observed reduced level of XB130 protein in
Figure 3 Identification of RET/PTC-mediated XB130 phosphorylation site(s). (a) Schematic representation of the XB130 mutations.DN, lacking N-terminal amino-acid residues 2–168; Y54F, Y124F and Y161F, carrying the tyrosine (Y) to phenylalanine (F) changesof the indicated tyrosines. (b) Full-length XB130 or its mutants described above were transfected in HEK293 in the presence or absenceof RET/PTC3. Cell lysates were immunoprecipitated with anti-XB130 antibody and immunoblotted with anti-phosphotyrosineantibody. Total protein lysates were probed with anti-XB130 and anti-RET antibodies.
XB130, a novel adaptor in RET/PTC signalingM Lodyga et al
942
Oncogene

TPC1 cells (data not shown). To avoid any confoundingfactors that might arise from changes in the endogenousXB130 levels, we expressed the myc-tagged XB130construct in TPC1 cells before the RET kinase inhibitortreatment. The levels of XB130 protein phosphorylationand its binding to p85 were measured at 3, 6, 12 and 24 hafter treatment with the inhibitor. The efficiency ofRET/PTC inhibition by ZD6474 was followed at thesame time points by measuring the autophosphorylationof RET/PTC on residue Y905, which is essentialfor its kinase activity (Iwashita et al., 1996). The resultsshow that XB130 achieved high basal phosphorylationwhen introduced into the TPC1 cells. Pretreatment ofthe cells with ZD6474 caused a large decrease in thephosphorylation of XB130, an effect which correlatedwith the inhibition of RET/PTC phosphorylation(Supplementary Figure 5c). In addition, ZD6474 treat-ment reduced the interaction of XB130 with p85 byapproximately 45%. Collectively, these data demon-
strate that RET/PTC-mediated XB130 tyrosinephosphorylation generates binding sites that promotethe interaction between XB130 and p85 in thyroidtumor cells.
Akt is a downstream target of the RET/PTC-XB130signaling pathwayTo assess the functional significance of XB130 in theRET/PTC-PI 3-kinase pathway, we evaluated theactivation of the serine-threonine kinase Akt, a down-stream target of PI 3-kinase. We measured Aktphosphorylation on Ser 473, which is a sensitiveindicator of Akt activity (Alessi et al., 1996). InHEK293 cells, XB130 or RET/PTC alone led to amoderate increase in Akt phosphorylation, whereascoexpression of these two proteins exerted a strongsynergy with regards to Akt phosphorylation. Impor-tantly, this induction of Akt phosphorylation wascritically dependent on intact, phosphorylatable
Figure 4 RET/PTC3 enhanced XB130 and p85 interaction. (a) Myc-tagged XB130 was transfected in the presence or absence of RET/PTC3 in HEK293 cells. Cell lysates were immunoprecipitated with anti-myc and immunoblotted with anti-myc or anti-p85 antibody.(b) Cell lysates collected from HEK293 cells transfected with myc-tagged XB130 and/or RET/PTC3 were incubated with glutathione-Sepharose-immobilized GST-p85 N-terminal or C-terminal SH2 fusion protein. The binding of XB130 to the SH2 domains of p85 wasassessed by western blotting using anti-myc antibody. XB130 protein levels in total cell lysates are shown for normalization.(c) Full-length or mutant XB130 cDNA were expressed in the presence or absence of RET/PTC3 in HEK293 cells. Cell lysates wereincubated with glutathione-Sepharose-immobilized GST-p85 N-terminal or C-terminal SH2 fusion protein. The samples collected werethen processed by western blotting using anti-XB130 antibody. Total cell lysates were analysed for levels of expression of XB130 andRET/PTC3. (d) Cell lysates from HEK293 cells transfected with XB130 and RET/PTC3 were incubated with glutathione-Sepharose-immobilized GST-N-terminal SH2 domain of p85, in the presence of increasing amounts of synthesized peptides designed surroundingTyrosine 54 of XB130. The phosphorylated peptide (ppY54), but not the nonphosphorylated peptide (p54) reduced binding of XB130to p85.
XB130, a novel adaptor in RET/PTC signalingM Lodyga et al
943
Oncogene

XB130, as it was absent on introduction of theXB130DN mutant (Figure 5a). To substantiate thatthe XB130-RET/PTC promoted phosphorylation ofAkt was indeed mediated by PI 3-kinase, we pretreatedXB130-RET/PTC3 cotransfected HEK293 cells withLY294002, a PI 3-kinase inhibitor, which abolishedRET/PTC3-XB130-facilitated Akt phosphorylation(Figure 5b).
Tyrosine 1062 is a crucial RET binding site for theactivation of various signaling pathways (Hayashi et al.,2000; Arighi et al., 2005). However, RET/PTC3 4YFmutation, which includes Y1062F (Figure 2a) was aseffective as the intact RET/PTC3 at the phosphorylation
of XB130 (Figure 2d). In agreement with this result,4YF mutant was equally proficient at promotingbinding between XB130 and either SH2 domain of p85(Figure 5c). Accordingly, 4YF mutant was as effectiveas the intact RET/PTC3 at promoting the RET/PTC3-XB130-enhanced Akt phosphorylation, however it wasincapable of inducing ERK phosphorylation(Figure 5d). This indicates that XB130 represents analternative route available to RET/PTC to stimulate PI3-kinase/Akt in a Y1062-indpendent manner. Moreover,as XB130 related signaling affects Akt phosphorylationbut not ERK, the XB130-dependent pathway is selectivein terms of downstream consequences.
Figure 5 Akt activation is downstream of RET/PTC-XB130 signaling pathway via a mechanism independent of residue Y1062 ofRET/PTC3. (a) HEK293 cells were transfected with XB130, XB130DN, RET/PTC3 alone or in combination. Cell lysates wereanalysed for phosphorylated Akt (pAKT S473) and total Akt with appropriate antibodies. (b) HEK293 cells were cotransfected withXB130 and RET/PTC3 and incubated either in the absence or presence of 25mM LY294002. Cell lysates were immunoblotted for totaland phosphorylated form of Akt. (c) HEK293 cells were cotransfected with XB130, or XB130 plus RET/PTC3 or 4YF mutant. Celllysates were incubated with GST-p85 N-terminal or C-terminal SH2 fusion protein. The binding of XB130 to the SH2 domains wasdetermined by western blotting using anti-myc antibody. (d) HEK293 cells were transfected as indicated. Western blot was performedto detect the impact of XB130 on Akt and ERK phosphorylation in the presence of the intact RET/PTC3, or its 4YF or the kinasedead mutant. The changes in phosphorylation were detected with specific Akt and ERK antibodies.
XB130, a novel adaptor in RET/PTC signalingM Lodyga et al
944
Oncogene

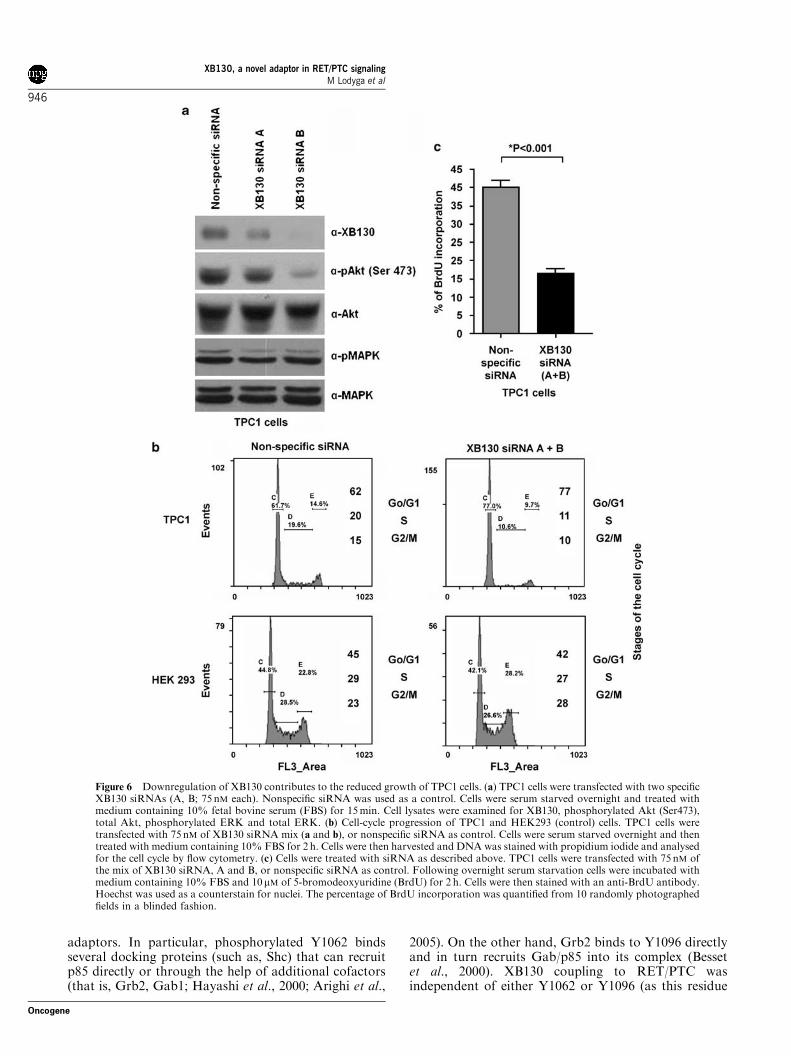
To corroborate this effect on natural XB130 expres-sors, TPC1 cells were treated for 48 h with two XB130siRNAs (A and B) designed against independentsegments of XB130. These tumor cells showed sub-stantial Akt phosphorylation (on Ser 473) under basalconditions. Treatment with either XB130 siRNAstrongly reduced this basal Akt phosphorylation, ascompared to nonspecific siRNA, whereas the total Aktlevels remained unaffected (Figure 6a). The secondsiRNA (B) proved to be a more potent inhibitor of Aktphosphorylation, in agreement with the observation thatit caused a stronger (near complete) reduction in XB130expression. In addition, this inhibition was specific tothe Akt pathway as ERK phosphorylation was notaffected. Together these observations indicate thatXB130 can function as a specific mediator in the RET/PTC/PI 3-kinase/Akt pathway.
Downregulation of XB130 in thyroid papillary carcinomacells inhibits cell-cycle progression and survivalPI 3-kinase/Akt pathway plays a central role in RET/PTC signaling and is involved in cell proliferation andsurvival. To determine the role of XB130 in cellproliferation, TPC1 and HEK293 cells were transfectedwith nonspecific siRNA or XB130 siRNA mix (AþB).Cells were harvested 48 h after transfection, stained withpropidium iodide for DNA content and processedthrough flow cytometry. Figure 6b shows that XB130siRNA increased the proportion of TPC1 cells accumu-lated in the G0/G1 phase of the cell cycle. However inHEK293 cells, which are devoid of detectable endogen-ous XB130 expression, such effect was not observed. Tosubstantiate this growth-arresting effect, and to directlyassess the impact of XB130 on the proliferative potentialof these tumor cells, we performed 5-bromodeoxyur-idine (BrdU) incorporation experiments. Transfectionwith the XB130 siRNAs significantly decreased BrdUuptake in TPC1 cells (Figure 6c). Treatment of TPC1cells with siRNA did not induce obvious morphologicalchanges (Supplementary Figure 6). As TPC1 cells carryRET/PTC, these data suggest that XB130 is animportant mediator of RET/PTC-enhanced cell prolif-eration.
Growing cells on polyHEMA-coated surface rendersanchorage-dependent cells unable to attach to the plates,and consequently causing them to enter an apoptoticpathway, Anoikis. To determine the role of XB130 incell survival, TPC1 cells transfected with nonspecific orXB130 siRNA were harvested, counted and grown onpolyHEMA for 24 h. Cells were collected, doublestained with Annexin-V-flourescein and propidiumiodide, then processed by flow cytometry. The cellviability is defined as: viable cells (Annexin-V and PI-negative), early apoptosis (Annexin-V-positive, PI-ne-gative), late apoptosis (double positive for Annexin-Vand PI; Figure 7a). Although the overall apoptosis(early plus late) did not yield significant differencebetween XB130 and nonspecific siRNA-treated group,there was a significantly lower percentage of cells in theearly phase and higher in the late phase of apoptosis in
XB130 siRNA treated group (Figure 7b), indicating thatdownregulation of XB130 accelerates the apoptoticprocess.
Discussion
Here we show that XB130 is preferentially expressed inthe thyroid tissue, moreover, its expression has beendetected in papillary thyroid carcinoma, and in TPC1cells that carry the RET/PTC1 rearrangement. We alsoshow that XB130 is a novel substrate of the thyroid-specific RET/PTC tyrosine kinase. Accordingly, coex-pression of XB130 and active (but not kinase dead)RET/PTC in HEK293 cells leads to significant XB130phosphorylation. Further, inhibitor blockade of RETkinase abolishes XB130 phosphorylation in TPC1 cells,in which both RET/PTC1 and XB130 are endogenouslyexpressed.
Although the kinase activity of RET/PTC is clearlyindispensable for the RET/PTC-mediated XB130 phos-phorylation, the exact mechanism of this processremains elusive. RET/PTC tyrosines (Y826, Y1015,Y1029 and Y1062), which are known to dock severalintracellular mediators, are not essential for XB130phosphorylation. XB130 forms a detectable complexwith RET/PTC. The interaction between RET/PTC andits established binding partners has been proposed to bemediated by either the SH2 or PTB domains. As XB130does not contain such domains, the interaction betweenXB130 and RET/PTC is likely mediated by analternative mechanism and/or possibly by intermediatebinding partners.
We found that Y54 in XB130 is a major targetthat accounts for approximately 50% of RET/PTC-dependent phosphorylation. The sequence surroundingY54 corresponds to a binding site for the SH2 domainsof the p85 regulatory subunit of PI 3-kinase (Backeret al., 1992; Carpenter et al., 1993; Rordorf-Nikolicet al., 1995). Accordingly, both the N- and the C-terminal SH2 domains of p85 bind XB130 in pull-downassays, and this binding is greatly enhanced by RET/PTC. Moreover, pharmacological inhibition of RET/PTC disrupts the binding between XB130 and p85 inRET/PTC1-positive TPC1 cells. Cotransfection ofXB130 with RET/PTC increases the basal Akt phos-phorylation on Ser473 in HEK293 cells, whereas siRNAdownregulation of XB130 in TPC1 cells reduces theirconstitutively increased Akt activity. Together theseobservations indicate that XB130 can function as aspecific mediator in the RET/PTC/PI 3-kinase/Aktpathway.
Several mechanisms have been elucidated that serve asa bridge between the activated native RET receptor orthe constitutively activated RET/PTC and the P13-kinase pathway. As the p85 regulatory subunit of PI 3-kinase does not bind to RET directly, adaptor proteinserve as links that relay the downstream signalingcascade. RET residues Y1062 (common to both RET-9 and RET-51 splicing variants) and Y1096 (specific forthe RET-51 form) provide binding sites for these
XB130, a novel adaptor in RET/PTC signalingM Lodyga et al
945
Oncogene
adaptors. In particular, phosphorylated Y1062 bindsseveral docking proteins (such as, Shc) that can recruitp85 directly or through the help of additional cofactors(that is, Grb2, Gab1; Hayashi et al., 2000; Arighi et al.,
2005). On the other hand, Grb2 binds to Y1096 directlyand in turn recruits Gab/p85 into its complex (Bessetet al., 2000). XB130 coupling to RET/PTC wasindependent of either Y1062 or Y1096 (as this residue
Figure 6 Downregulation of XB130 contributes to the reduced growth of TPC1 cells. (a) TPC1 cells were transfected with two specificXB130 siRNAs (A, B; 75 nM each). Nonspecific siRNA was used as a control. Cells were serum starved overnight and treated withmedium containing 10% fetal bovine serum (FBS) for 15min. Cell lysates were examined for XB130, phosphorylated Akt (Ser473),total Akt, phosphorylated ERK and total ERK. (b) Cell-cycle progression of TPC1 and HEK293 (control) cells. TPC1 cells weretransfected with 75 nM of XB130 siRNA mix (a and b), or nonspecific siRNA as control. Cells were serum starved overnight and thentreated with medium containing 10% FBS for 2 h. Cells were then harvested and DNA was stained with propidium iodide and analysedfor the cell cycle by flow cytometry. (c) Cells were treated with siRNA as described above. TPC1 cells were transfected with 75 nM ofthe mix of XB130 siRNA, A and B, or nonspecific siRNA as control. Following overnight serum starvation cells were incubated withmedium containing 10% FBS and 10mM of 5-bromodeoxyuridine (BrdU) for 2 h. Cells were then stained with an anti-BrdU antibody.Hoechst was used as a counterstain for nuclei. The percentage of BrdU incorporation was quantified from 10 randomly photographedfields in a blinded fashion.
XB130, a novel adaptor in RET/PTC signalingM Lodyga et al
946
Oncogene

was lacking from the RET/PTC constructs used in thisstudy). Accordingly, cotransfection of XB130 with theRET/PTC 4YF mutant was able to selectively rescue theactivation of the Akt pathway, without affecting theERK pathway. Thus, our findings imply that XB130may represent a new mechanism alternative to Y1062and Y1096 that couples RET/PTC to PI 3-kinase andselectively operates in thyrocytes. Such redundancy inthe mechanism of PI 3-kinase activation might reflectfine-tuning and versatility in the regulation, because theassembly of particular signaling complexes may deter-mine the activation of one or several signaling cascades,and may influence the intensity and duration of theresponse, as well as the specificity for the activation ofone pathway over the other.
The PI 3-kinase/Akt pathway plays a central role inmediating cell growth and survival (Hemmings, 1997;Kaplan and Miller, 1997; Marte and Downward, 1997).Our data provide direct evidence that XB130 is involved
in these critical functions. Downregulation of XB130leads to the accumulation of TPC1 cells in the G0 phaseof the cell cycle, and reduces proliferation as revealed bythe lower BrdU incorporation. The proliferative poten-tial of TPC1 cells has been attributed to the RET/PTCrearrangement. In fact, chemical inhibition of RET/PTC1 (Carlomagno et al., 2002; Mariggio et al., 2007) orRET/PTC1 silencing by RNAi (de Martimprey et al.,2008) blunted TPC1 cell proliferation. Moreover, ourdata show that anoikis challenged TPC1 cells are morepredisposed to cell death on XB130 downregulation.Thus, XB130 could be an important intermediate in thepropagation of PI 3-kinase signaling, which contributesto abnormal cell proliferation and survival in thyroidcancer cells sustained by RET/PTC rearrangement.Although XB130 is highly expressed in thyroid andthyroid tumors, it has been found in other cell types(data not shown), hence the interaction between XB130and p85 subunit of PI 3-kinase may also contribute to
Figure 7 XB130 downregulation accelerates aniokis-induced apoptosis in TPC1 cells. (a) TPC1 cells treated with nonspecific orXB130 siRNA were plated on poly-HEMA coated 60mm dishes and incubated in the adhesion compromised environment for 24 h,followed by Annexin-V-flourescein/PI double staining. Apoptosis was evaluated by flow cytometry, and representative 2D plot imagesare shown. (b) Statistical analysis of experiment described in (a). Results are summarized from three independent experiments.
XB130, a novel adaptor in RET/PTC signalingM Lodyga et al
947
Oncogene

signal transduction in cells containing endogenousXB130 expression.
Materials and methods
Portion of the Materials and methods is presented asSupplementary materials, including northern blot analysis,immunohistochemical staining, cell lines, protein studies,siRNA transfection, flow cytometry and BrdU incorporation.
PlasmidsRET constructs used in this study were cloned into pCDNA3.1(Invitrogen, San Diego, CA, USA) containing either a myc tagor pBABE (no tag) vectors. They encode the short RET splicedform (RET-9) as described elsewhere (Castellone et al., 2003).RET/PTC1 and RET/PTC3 constructs encode the H4-RETand RFG (Ele1/ARA70/Ncoa4)-RET chimeric oncogenes,respectively. RET/PTC3(K�) is a kinase-dead mutant, carry-ing the substitution of the catalytic lysine (residue 758 in full-length RET) with a methionine. RET/PTC3(4YF), in whichtyrosines corresponding to Y826, Y1015, Y1029 and Y1062 infull-length RET were mutated to phenylalanine residues, wasgenerated by site-directed mutagenesis using a QuikChangemutagenesis kit (Stratagene, La Jolla, CA, USA; Melillo et al.,2001). Expression vectors for XB130 and its mutants(XB130DN, 2–168 amino-acid deletion), XB130(Y124F) andXB130(Y161F) were cloned into a pCDNA3 backbonecontaining a His tag as described previously (Xu et al.,2007). In addition, full-length XB130 was subcloned intopCDNA3.1 vector containing a myc tag. XB130(Y54F) pointmutation was generated by site-directed mutagenesis using theQuikChange mutagenesis kit (Stratagene). The mutations wereconfirmed by DNA sequencing.
Binding assaysGST-N-SH2 p85 and GST-C-SH2 p85 domains (Pelicci et al.,2002) were a kind gift of G Pelicci (Milan, Italy). Recombinantproteins were produced in Escherichia coli and purified onglutathione-conjugated sepharose by standard procedure (GEHealthcare Bio-Sciences Corp., Piscataway, NJ, USA). Forpull-down assays, 20mg of the recombinant protein bound tosepharose were incubated with cell lysates for 2 h at 4 1C. After
washing the beads, proteins were separated by SDS–polyacrylamide gel electrophoresis and probed with indicatedantibodies.
Two peptides were synthesized based on XB130 amino-acid sequence surrounding Tyrosine 54 (YXXM motif):KSSSSDEEYIYMNKVTI (p54) and KSSSSDEEpYIYMNKVTI (pp54), by Neosystem Laboratoire (Strasbourg,France). GST-pull-down experiment was performed asdescribed above.
Anoikis assayApoptosis in TPC1 cells treated with control or XB130 siRNAwas induced by inhibition of contact with the extracellularmatrix. Briefly, 60mm dishes were coated with 1 ml of 10 mg/ml of poly-HEMA (poly(2-hydroxyethylmethacrylate)) in 95%ethanol and allowed to dry. This process was then repeated.The dishes were washed with phophate-suffered saline andstored dry until further use. TPC1 cells grown for 24 h in thepresence of siRNA were detached using trypsin/EDTAsolution and plated onto the polyHEMA-coated wells at1� 106 cells per dish. After 24 h cells were harvested frompolyHEMA-coated wells, centrifuged and resuspended. Apop-totic cells were evaluated with Cytomix FC500 flow cytometrysystem, using the Annexin-V-fluorescein and propidium iodine(PI) double staining method according to manufacturer’sinstructions (Roche Molecular Diagnostics, Basel, Switzer-land). Cells were gated according to the staining, viable cells(negative for both Annexin-V and PI), Early Apoptosis(Annexin-V-positive only), and Late Apoptosis (doublepositive for Annexin-V and PI).
Acknowledgements
We are grateful to Dr B Han and Dr D Winer for technicalassistance. We thank AJ Ryan, AstraZeneca for the ZD6474inhibitor. We also thank Dr S Asa and JM Hershman forhuman thyroid cell lines, Dr G Pelicci for GST-fusion proteinsand Dr M Chiariello for the Src mutant. This work wassupported by operating grants (MOP-13270, MOP-42546)from Canadian Institutes of Health Research, a grant from theItalian Association for Cancer Research, MIUR, Alleanzacontro iL Cancro and the European Union Contract FP6-36495 (GENRISK-T).
References
Alessi DR, Andjelkovic M, Caudwell B, Cron P, Morrice N, Cohen Pet al. (1996). Mechanism of activation of protein kinase B by insulinand IGF-1. EMBO J 15: 6541–6551.
Arighi E, Borrello MG, Sariola H. (2005). RET tyrosine kinasesignaling in development and cancer. Cytokine Growth Factor Rev
16: 441–467.Backer JM, Myers Jr MG, Shoelson SE, Chin DJ, Sun XJ, Miralpeix
M et al. (1992). Phosphatidylinositol 30-kinase is activated byassociation with IRS-1 during insulin stimulation. EMBO J 11:3469–3479.
Besset V, Scott RP, Ibanez CF. (2000). Signaling complexes andprotein-protein interactions involved in the activation of the Rasand phosphatidylinositol 3-kinase pathways by the c-Ret receptortyrosine kinase. J Biol Chem 275: 39159–39166.
Carlomagno F, Vitagliano D, Guida T, Ciardiello F, Tortora G,Vecchio G et al. (2002). ZD6474, an orally available inhibitor ofKDR tyrosine kinase activity, efficiently blocks oncogenic RETkinases. Cancer Res 62: 7284–7290.
Carpenter CL, Auger KR, Chanudhuri M, Yoakim M, SchaffhausenB, Shoelson S et al. (1993). Phosphoinositide 3-kinase is activated by
phosphopeptides that bind to the SH2 domains of the 85-kDasubunit. J Biol Chem 268: 9478–9483.
Castellone MD, Cirafici AM, De Vita G, De Falco V, Malorni L, Tallini Get al. (2003). Ras-mediated apoptosis of PC CL 3 rat thyroid cellsinduced by RET/PTC oncogenes. Oncogene 22: 246–255.
De Falco V, Guarino V, Malorni L, Cirafici AM, Troglio F, Erreni Met al. (2005). RAI(ShcC/N-Shc)-dependent recruitment of GAB 1 toRET oncoproteins potentiates PI 3-K signalling in thyroid tumors.Oncogene 24: 6303–6313.
de Martimprey H, Bertrand JR, Fusco A, Santoro M, Couvreur P,Vauthier C et al. (2008). siRNA nanoformulation against the ret/PTC1 junction oncogene is efficient in an in vivo model of papillarythyroid carcinoma. Nucleic Acids Res 36: e2.
Escobedo JA, Navankasattusas S, Kavanaugh WM, Milfay D,Fried VA, Williams LT. (1991). cDNA cloning of a novel 85 kdprotein that has SH2 domains and regulates binding of PI3-kinase tothe PDGF beta-receptor. Cell 65: 75–82.
Flynn DC, Leu TH, Reynolds AB, Parsons JT. (1993). Identificationand sequence analysis of cDNAs encoding a 110-kilodalton actinfilament-associated pp60src substrate. Mol Cell Biol 13: 7892–7900.
XB130, a novel adaptor in RET/PTC signalingM Lodyga et al
948
Oncogene

Giordano TJ, Kuick R, Thomas DG, Misek DE, Vinco M, Sanders Det al. (2005). Molecular classification of papillary thyroid carcinoma:distinct BRAF, RAS, and RET/PTC mutation-specific gene expres-sion profiles discovered by DNA microarray analysis. Oncogene 24:6646–6656.
Grieco M, Santoro M, Berlingieri MT, Melillo RM, Donghi R,Bongarzone I et al. (1990). PTC is a novel rearranged form of the retproto-oncogene and is frequently detected in vivo in human thyroidpapillary carcinomas. Cell 60: 557–563.
Han B, Bai XH, Lodyga M, Xu J, Yang BB, Keshavjee S et al. (2004).Conversion of mechanical force into biochemical signaling. J Biol
Chem 279: 54793–54801.Harlan JE, Hajduk PJ, Yoon HS, Fesik SW. (1994). Pleckstrin
homology domains bind to phosphatidylinositol-4,5-bisphosphate.Nature 371: 168–170.
Haslam RJ, Koide HB, Hemmings BA. (1993). Pleckstrin domainhomology. Nature 363: 309–310.
Hayashi H, Ichihara M, Iwashita T, Murakami H, Shimono Y, Kawai Ket al. (2000). Characterization of intracellular signals via tyrosine1062 in RET activated by glial cell line-derived neurotrophic factor.Oncogene 19: 4469–4475.
Hemmings BA. (1997). PtdIns(3,4,5)P3 gets its message across. Science
277: 534.Iavarone C, Acunzo M, Carlomagno F, Catania A, Melillo RM,
Carlomagno SM et al. (2006). Activation of the Erk8 mitogen-activated protein (MAP) kinase by RET/PTC3, a constitutivelyactive form of the RET proto-oncogene. J Biol Chem 281:10567–10576.
Iwashita T, Asai N, Murakami H, Matsuyama M, Takahashi M.(1996). Identification of tyrosine residues that are essential fortransforming activity of the ret proto-oncogene with MEN2A orMEN2B mutation. Oncogene 12: 481–487.
Jung HS, Kim DW, Jo YS, Chung HK, Song JH, Park JS et al. (2005).Regulation of protein kinase B tyrosine phosphorylation bythyroid-specific oncogenic RET/PTC kinases. Mol Endocrinol 19:2748–2759.
Kaplan DR, Miller FD. (1997). Signal transduction by the neuro-trophin receptors. Curr Opin Cell Biol 9: 213–221.
Knauf JA, Kuroda H, Basu S, Fagin JA. (2003). RET/PTC-induceddedifferentiation of thyroid cells is mediated through Y1062signaling through SHC-RAS-MAP kinase. Oncogene 22: 4406–4412.
Lodyga M, Bai XH, Mourgeon E, Han B, Keshavjee S, Liu M. (2002).Molecular cloning of actin filament-associated protein: a putativeadaptor in stretch-induced Src activation. Am J Physiol Lung Cell
Mol Physiol 283: L265–L274.Maeda K, Murakami H, Yoshida R, Ichihara M, Abe A, Hirai Met al. (2004). Biochemical and biological responses induced bycoupling of Gab1 to phosphatidylinositol 3-kinase in RET-expres-sing cells. Biochem Biophys Res Commun 323: 345–354.
Mai KT, Vaccani JP, Thomas J, Odell PF. (2001). Immunostaining forret oncogene proteins in papillary thyroid carcinoma: a correlationof ret immunoreactivity and potential of lymph node metastasis.Thyroid 11: 859–863.
Mariggio S, Filippi BM, Iurisci C, Dragani LK, De Falco V, SantoroM et al. (2007). Cytosolic phospholipase A2 alpha regulates cell
growth in RET/PTC-transformed thyroid cells. Cancer Res 67:11769–11778.
Marte BM, Downward J. (1997). PKB/Akt: connecting phosphoinosi-tide 3-kinase to cell survival and beyond. Trends Biochem Sci 22:355–358.
Melillo RM, Carlomagno F, De Vita G, Formisano P, Vecchio G,Fusco A et al. (2001). The insulin receptor substrate (IRS)-1 recruitsphosphatidylinositol 3-kinase to Ret: evidence for a competitionbetween Shc and IRS-1 for the binding to Ret. Oncogene 20:209–218.
Melillo RM, Castellone MD, Guarino V, De Falco V, Cirafici AM,Salvatore G et al. (2005). The RET/PTC-RAS-BRAF linearsignaling cascade mediates the motile and mitogenic phenotype ofthyroid cancer cells. J Clin Invest 115: 1068–1081.
Miyagi E, Braga-Basaria M, Hardy E, Vasko V, Burman KD, Jhiang Set al. (2004). Chronic expression of RET/PTC 3 enhances basal andinsulin-stimulated PI3 kinase/AKT signaling and increases IRS-2expression in FRTL-5 thyroid cells. Mol Carcinog 41: 98–107.
Pelicci G, Troglio F, Bodini A, Melillo RM, Pettirossi V, Coda L et al.(2002). The neuron-specific Rai (ShcC) adaptor protein inhibitsapoptosis by coupling Ret to the phosphatidylinositol 3-kinase/Aktsignaling pathway. Mol Cell Biol 22: 7351–7363.
Pierotti MA, Santoro M, Jenkins RB, Sozzi G, Bongarzone I, Grieco Met al. (1992). Characterization of an inversion on the long arm ofchromosome 10 juxtaposing D10S170 and RET and creatingthe oncogenic sequence RET/PTC. Proc Natl Acad Sci USA 89:1616–1620.
Rordorf-Nikolic T, Van Horn DJ, Chen D, White MF, Backer JM.(1995). Regulation of phosphatidylinositol 30-kinase by tyrosylphosphoproteins. Full activation requires occupancy of bothSH2 domains in the 85-kDa regulatory subunit. J Biol Chem 270:3662–3666.
Santoro M, Chiappetta G, Cerrato A, Salvatore D, Zhang L,Manzo G et al. (1996). Development of thyroid papillarycarcinomas secondary to tissue-specific expression of the RET/PTC1 oncogene in transgenic mice. Oncogene 12: 1821–1826.
Santoro M, Dathan NA, Berlingieri MT, Bongarzone I, Paulin C,Grieco M et al. (1994a). Molecular characterization of RET/PTC3;a novel rearranged version of the RETproto-oncogene in a humanthyroid papillary carcinoma. Oncogene 9: 509–516.
Santoro M, Melillo RM, Grieco M, Berlingieri MT, Vecchio G, FuscoA. (1993). The TRK and RET tyrosine kinase oncogenes cooperatewith ras in the neoplastic transformation of a rat thyroid epithelialcell line. Cell Growth Differ 4: 77–84.
Santoro M, Wong WT, Aroca P, Santos E, Matoskova B, Grieco Met al. (1994b). An epidermal growth factor receptor/ret chimeragenerates mitogenic and transforming signals: evidence for a ret-specific signaling pathway. Mol Cell Biol 14: 663–675.
Takahashi M, Buma Y, Iwamoto T, Inaguma Y, Ikeda H, Hiai H.(1988). Cloning and expression of the ret proto-oncogene encoding atyrosine kinase with two potential transmembrane domains.Oncogene 3: 571–578.
Xu J, Bai XH, Lodyga M, Han B, Xiao H, Keshavjee S et al. (2007).XB130, a novel adaptor protein for signal transduction. J Biol Chem
282: 16401–16412.
Supplementary Information accompanies the paper on the Oncogene website (http://www.nature.com/onc)
XB130, a novel adaptor in RET/PTC signalingM Lodyga et al
949
Oncogene




















