
X-ray structures of the peridinin–chlorophyll-protein reconstituted with different chlorophylls
6
X-ray structures of the peridinin–chlorophyll-protein reconstituted with different chlorophylls Tim Schulte a , Roger G. Hiller b , Eckhard Hofmann a, * a Biophysics, Department of Biology and Biotechnology, Ruhr-University Bochum, D-44780 Bochum, Germany b Biology Department, Faculty of Science, Macquarie University, NSW 2109, Australia article info Article history: Received 12 January 2010 Accepted 21 January 2010 Available online 25 January 2010 Edited by Richard Cogdell Keywords: Light harvesting Peridinin Protein crystallography Refolding Photosynthesis abstract The peridinin–chlorophyll a-protein (PCP) from dinoflagellates is a soluble light harvesting antenna which gathers incoming photons mainly by the carotenoid peridinin. In PCPs reconstituted with dif- ferent chlorophylls, the peridinin to chlorophyll energy transfer rates are well predicted by a För- ster-like theory, but only if the pigment arrangements are identical in all PCPs. We have determined the X-ray structures of PCPs reconstituted with Chlorophyll-b (Chl-b), Chlorophyll-d (Chl-d) and Bacteriochlorophyll-a (BChl-a) to resolutions 62 Å. In all three cases the pigment arrangements are essentially the same as in native PCP. Hydrogen bonding is not responsible for preferential incorporation of ‘‘non-native” chlorophylls over Chl-a. Ó 2010 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. 1. Introduction Dinoflagellates contain a unique soluble light harvesting anten- na, the peridinin–chlorophyll a-protein (PCP), which differs from other antennas in using a carotenoid as main light harvesting pig- ment instead of chlorophylls (Chl) [1]. Elucidation of the structure of the main form of PCP (MFPCP) [2] has encouraged many spectro- scopic groups to investigate the energy transfer processes in PCP (reviewed in [3]) which are dominated by the unique properties of the carotenoid peridinin (Per) (summarized in [4]). In common with other polar carotenoids such as fucoxanthin and siphonaxanthin, peridinin has a conjugated carbonyl group (Fig. 1E) and shows an appreciable emission from the first excited singlet state S1 [5]. The peridinin S1 lifetime is dependent on the polarity of the environment which controls the level of an intramo- lecular charge transfer (ICT) state [3,4]. This ICT state has been shown by Zigmantas et al. [6] to be involved in the energy transfer from Per S1 to Chl Qy in the PCP complex. Polivka et al. have dem- onstrated that Förster theory can be applied to describe the Per S1/ ICT ? Chl Qy energy transfer rate [7] by reconstituting PCP [8] (called RFPCP) with Chls having different Qy energies. To interpret the transfer rates in terms of a Förster-like mechanism, it was as- sumed that the pigment arrangements in the different Chl-RFPCPs are indistinguishable from those in MFPCP [7]. We have recently shown that this assumption holds true for Chl-a RFPCP [9]. In this work we present the X-Ray structures of RFPCPs refolded with Chl-b, Chl-d and BChl-a (Chl structures shown in Fig. 1D). Their pigment arrangements and Chl binding modes are compared to the arrangement and binding mode of Chl-a in Chl-a RFPCP [9] (Fig. 1A–C). 2. Materials and methods 2.1. Protein expression and refolding of RFPCP RFPCP was prepared according to [9]. Details of the pigment purification are given in SI Text 1. 2.2. Crystallization of RFPCP Crystallization conditions for Chl-RFPCPs were the same as indi- cated in [9]. Chl-b RFPCP crystals only grew after addition of Chl-a RFPCP crystal seeds. Crystals were harvested according to [9]. 0014-5793/$36.00 Ó 2010 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.febslet.2010.01.041 Abbreviations: (B)Chl, (bacterio-)chlorophyll; ICT, intramolecular charge transfer; Per, peridinin; (RF/MF)PCP, (refolded/main form)peridinin–chlorophyll a-protein * Corresponding author. Address: Ruhr-Universität Bochum, AG Proteinkristal- lographie, Lehrstuhl für Biophysik, Gebäude ND 04/316, D-44780 Bochum. Fax: +49 (0)234 32 14762. E-mail address: [email protected] (E. Hofmann). FEBS Letters 584 (2010) 973–978 journal homepage: www.FEBSLetters.org
-
Upload
ruhr-uni-bochum -
Category
Documents
-
view
5 -
download
0
Transcript of X-ray structures of the peridinin–chlorophyll-protein reconstituted with different chlorophylls

FEBS Letters 584 (2010) 973–978
journal homepage: www.FEBSLetters .org
X-ray structures of the peridinin–chlorophyll-protein reconstitutedwith different chlorophylls
Tim Schulte a, Roger G. Hiller b, Eckhard Hofmann a,*
a Biophysics, Department of Biology and Biotechnology, Ruhr-University Bochum, D-44780 Bochum, Germanyb Biology Department, Faculty of Science, Macquarie University, NSW 2109, Australia
a r t i c l e i n f o
Article history:Received 12 January 2010Accepted 21 January 2010Available online 25 January 2010
Edited by Richard Cogdell
Keywords:Light harvestingPeridininProtein crystallographyRefoldingPhotosynthesis
0014-5793/$36.00 � 2010 Federation of European Biodoi:10.1016/j.febslet.2010.01.041
Abbreviations: (B)Chl, (bacterio-)chlorophyll;transfer; Per, peridinin; (RF/MF)PCP, (refolded/maina-protein
* Corresponding author. Address: Ruhr-Universitätlographie, Lehrstuhl für Biophysik, Gebäude ND 04/31(0)234 32 14762.
E-mail address: [email protected]
a b s t r a c t
The peridinin–chlorophyll a-protein (PCP) from dinoflagellates is a soluble light harvesting antennawhich gathers incoming photons mainly by the carotenoid peridinin. In PCPs reconstituted with dif-ferent chlorophylls, the peridinin to chlorophyll energy transfer rates are well predicted by a För-ster-like theory, but only if the pigment arrangements are identical in all PCPs. We havedetermined the X-ray structures of PCPs reconstituted with Chlorophyll-b (Chl-b), Chlorophyll-d(Chl-d) and Bacteriochlorophyll-a (BChl-a) to resolutions 62 Å. In all three cases the pigmentarrangements are essentially the same as in native PCP. Hydrogen bonding is not responsible forpreferential incorporation of ‘‘non-native” chlorophylls over Chl-a.� 2010 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
1. Introduction
Dinoflagellates contain a unique soluble light harvesting anten-na, the peridinin–chlorophyll a-protein (PCP), which differs fromother antennas in using a carotenoid as main light harvesting pig-ment instead of chlorophylls (Chl) [1]. Elucidation of the structureof the main form of PCP (MFPCP) [2] has encouraged many spectro-scopic groups to investigate the energy transfer processes in PCP(reviewed in [3]) which are dominated by the unique propertiesof the carotenoid peridinin (Per) (summarized in [4]).
In common with other polar carotenoids such as fucoxanthinand siphonaxanthin, peridinin has a conjugated carbonyl group(Fig. 1E) and shows an appreciable emission from the first excitedsinglet state S1 [5]. The peridinin S1 lifetime is dependent on thepolarity of the environment which controls the level of an intramo-lecular charge transfer (ICT) state [3,4]. This ICT state has beenshown by Zigmantas et al. [6] to be involved in the energy transferfrom Per S1 to Chl Qy in the PCP complex. Polivka et al. have dem-
chemical Societies. Published by E
ICT, intramolecular chargeform)peridinin–chlorophyll
Bochum, AG Proteinkristal-6, D-44780 Bochum. Fax: +49
ochum.de (E. Hofmann).
onstrated that Förster theory can be applied to describe the Per S1/ICT ? Chl Qy energy transfer rate [7] by reconstituting PCP [8](called RFPCP) with Chls having different Qy energies. To interpretthe transfer rates in terms of a Förster-like mechanism, it was as-sumed that the pigment arrangements in the different Chl-RFPCPsare indistinguishable from those in MFPCP [7].
We have recently shown that this assumption holds true forChl-a RFPCP [9]. In this work we present the X-Ray structures ofRFPCPs refolded with Chl-b, Chl-d and BChl-a (Chl structuresshown in Fig. 1D). Their pigment arrangements and Chl bindingmodes are compared to the arrangement and binding mode ofChl-a in Chl-a RFPCP [9] (Fig. 1A–C).
2. Materials and methods
2.1. Protein expression and refolding of RFPCP
RFPCP was prepared according to [9]. Details of the pigmentpurification are given in SI Text 1.
2.2. Crystallization of RFPCP
Crystallization conditions for Chl-RFPCPs were the same as indi-cated in [9]. Chl-b RFPCP crystals only grew after addition of Chl-aRFPCP crystal seeds. Crystals were harvested according to [9].
lsevier B.V. All rights reserved.

Fig. 1. (A) Structure of RFPCP [9]. (B) Chl-a is coordinated by a water molecule which is fixed by His-66. (C) Binding niche for Chl-a: the surface is created by surroundingresidue atoms which are located within 4 Å distance from the Chl-a macrocycle. (D) Structures of Chl-a, Chl-b, Chl-d and BChl-a. (E) Structure of peridinin.
974 T. Schulte et al. / FEBS Letters 584 (2010) 973–978
2.3. Data collection and structure determination
Oscillation data of the Chl-d RFPCP crystal were collected onthree different crystal positions at 100 K at the Swiss Light Source(Villigen, Switzerland) at beamline PXI using a MarCCD-225 detec-tor. The Chl-b RFPCP crystal was measured at the European Syn-chrotron Radiation Facility (ESRF) (Grenoble, France) at beamlineID23-2 using a MarCCD-225 detector. The BChl-a RFPCP crystalwas measured at the ESRF at beamline ID23-1 using a Q315r ADSCCCD detector. All data were processed and scaled using the XDSpackage [10]. Data statistics are listed in Table 1.
The Chl-RFPCP structures were solved by molecular replace-ment using Molrep [11] as implemented in CCP4 [12]. RFPCP (Pro-tein Data Bank code: 3IIS) was selected as search model. The modeland electron density maps were iteratively improved by refine-ment of the coordinates in Refmac [13] and interpretation of theresulting maps in Coot [14]. Data statistics including Rwork and Rfree
values as calculated by Phenix [15] and SFcheck [16] are given inTable 1. The lipid digalactosyldiacylglycerol (DGDG) was modeledwith two C18 acyl chains instead of one C18 and another C20 acylchain as in Chl-a RFPCP [9]. This is because different pigmentsources were used for the reconstitution: Chl-a RFPCP was recon-stituted with the whole MFPCP pigment extract from Amphidini-um carterae [9]. In contrast, Chl-RFPCPs reconstituted withpurified pigments could only be crystallized if DGDG extracted
from plant leaves was added to the reconstitution mixture (see SItext 1).
Structural figures were made with Pymol [17]. Chlorophyll omitelectron density maps were calculated as follows: (1) removal ofChl coordinates, (2) coordinate perturbation of the whole modelby an average of 0.5 Å using Moleman2 [18], (3) calculation of dif-ference maps using FFT in CCP4 [12].
2.4. Spectroscopy
Room-temperature steady-state absorption spectra of RFPCPand Chl-RFPCPs were measured with a Ultraspec 3000 pro Spec-trometer (Amersham Biosciences, Freiburg, Germany). Chl-RFPCPsconcentrations were made to Qy intensity of 60.25.
3. Results and discussion
The absorption spectra of the Chl-RFPCPs which were used forcrystallization are shown in Fig. 2. In these spectra the Chl Qypeaks are located at 648 nm (Chl-b), 670 nm (Chl-a), 698 nm(Chl-d) and 791 nm (BChl-a) which are comparable to recentlypublished data [7,8,19].
The rmsds of the superimposed Ca-atoms of the protein is inthe range of �0.1 Å (Table 2). Given this high structural consensusbetween the Ca atoms it is not surprising that the pigment

Table 1Summary of data collection and structure refinement statistics.
Chl-d Chl-b BChl-a
Data collectionSpace group C222(1) C222(1) C222(1)Unit cell parameters (a, b, c in Å; a, b, c in �) a = 68.6, b = 82.0, c = 75.2,
a = b = c = 90a = 68.6, b = 82.0, c = 75.4,a = b = c = 90
a = 68.5, b = 81.8, c = 75.0,a = b = c = 90
X-ray source SLS X06SA ESRF ID 23.2 ESRF ID23.1Temperature [K] 100 100 100Resolution limits [Å]� 43.11�1.8 (1.8�1.85) 43.15�1.95 (1.95�2.00) 43.01�1.75 (1.75�1.8)Wavelength [Å] 1.00789 0.8726 0.97625No. of observations 269 600 (16507) 62 249 (2982) 103 169 (7904)No. of unique reflections 20 022 (1470) 15 738 (1066) 21 555 (1724)Redundancy 13.5 (11.2) 4.00 (2.8) 4.8 (4.6)Completeness [%] 100 (100) 99.3 (93.3) 99.7 (99.8)I/r 16.6 (5.9) 13.8 (4.9) 14.6 (3.1)R(sym) [%] 15.0 (56.6) 9.4 (30.7) 8.6 (53.2)R(meas) [%]� 15.6 (59.2) 10.8 (36.6) 9.6 (59.9)R(mrgd-F) [%]� 7.1 (21.8) 9.9 (39.0) 11.7 (55.4)Structure refinementPDB Entry 2X1Z 2X20 2X21R(work) [%] Refmac (Phenix/SFcheck) 14.1 (14.4/16.9) 15.5 (15.7/17.5) 14.7 (15.1/16.3)R(free) [%] Refmac (Phenix/SFcheck) 18.3 (18.1/20.5) 19.6 (20.0/21.9) 18.6 (19.2/20.1)No. of refined atoms (protein/solvent/ligands/ions/
other)1758 (1200/214/323/14/7) 1689 (1149/196/324/13/7) 1725 (1138/237/323/20/7)
Mean isotropic B-value [Å2] 13.1 13.0 15.1Rmsd from ideal bond lengths [Å] 0.025 0.025 0.028Rmsd from ideal bond angle [�] 3.545 3.473 3.595MolprobityFavored [%] in Ramachandran plot 100 100 100Clash score (percentile) 6.93 (90th) 13.18 (64th) 12.79 (57th)Score (percentile) 1.38 (97th) 1.81 (84th) 1.62 (88th)
� Data in parantheses represent values in the highest-resolution bin.� From reference [10].
Table 2Comparison of pigment atom positions in Chl-a RFPCP with RFPCP with different Chls.
Chl-d Chl-b BChl-a
Protein 0.11 (151) 0.14 (151) 0.12 (151)PigmentsChl� 0.12 (44) 0.19 (45) 0.48 (45)Per� 0.13 (100) 0.13 (100) 0.15 (100)DGD 0.76 (66) 0.87 (66) 0.83 (66)RMSD ratioChl/protein 1.1 1.4 4.0
� Chl macrocycle without phytol tail.� Peridinin pi-conjugated system (C7-C6’) with epoxygroup (C6, O, C5) and methylsubstituents at C13 and C9’.
Fig. 2. Absorption spectra of RFPCP refolded with different Chls.
T. Schulte et al. / FEBS Letters 584 (2010) 973–978 975
arrangement is almost identical in all refolded Chl-RFPCPs (Table2). Several studies [19–21] have shown that the apparent bindingaffinities for the Chls with polar substituents at R3 and R7(Fig. 1D) are higher than for native Chl-a. These higher bindingaffinities were attributed to hydrogen bonds between the polarsubstituents of the Chls and polar groups of surrounding peridi-nins. Knowledge of the corresponding atom positions in the Chl-RFPCP X-ray structures enables us to estimate the H-bonding prob-abilities between these groups.
3.1. Chl-d RFPCP
In addition to the effects of polar R3 and R7 substitution on thebinding affinities discussed in the previous section, Di Valentin
et al. [22] have proposed a Chl-d porphyrin ring rotation as a pos-sible interpretation of their pulsed EPR experiments. However, inthe structures reported here, the rmsd between the Chl-d andChl-a atom positions is �0.1 Å which implies that the porphyrincycles are orientated in the same way in both complexes (Table 2).
The omit electron density map of Chl-d in RFPCP is shown inFig. 3. Chl-d differs from Chl-a in having a formyl (CHO) substituentat position R3 instead of a vinyl (C2H5) substituent. Althougheffective substitution of the carbon atom by an oxygen atom isnot directly detectable in the electron density map at the currentresolution, integration of Chl-d in exact replacement of Chl-a canbe deduced from analysis of alternate conformations. In Chl-aRFPCP, as well as in Chl-b RFPCP, the vinyl substituent on ring Ahas to be modeled in two conformations, whereas in Chl-d RFPCPthe formyl substituent is fixed (Fig. 3). The fixation of the formylgroup is probably caused by an H-bond (WHATIF 0-1GF = 0.7 (0–1 geometry factor: 0 = no hydrogen bond, 1 = optimal hydrogenbond) [23]) between the formyl group oxygen and the C10-hydro-xyl group of Per-612 (Fig. 4). Interestingly, Ilagan et al. [24] alsoproposed such a fixation of the formyl substituent. In the 10 Kabsorption spectra of Chl-d RFPCP, a split Chl-d Soret band was
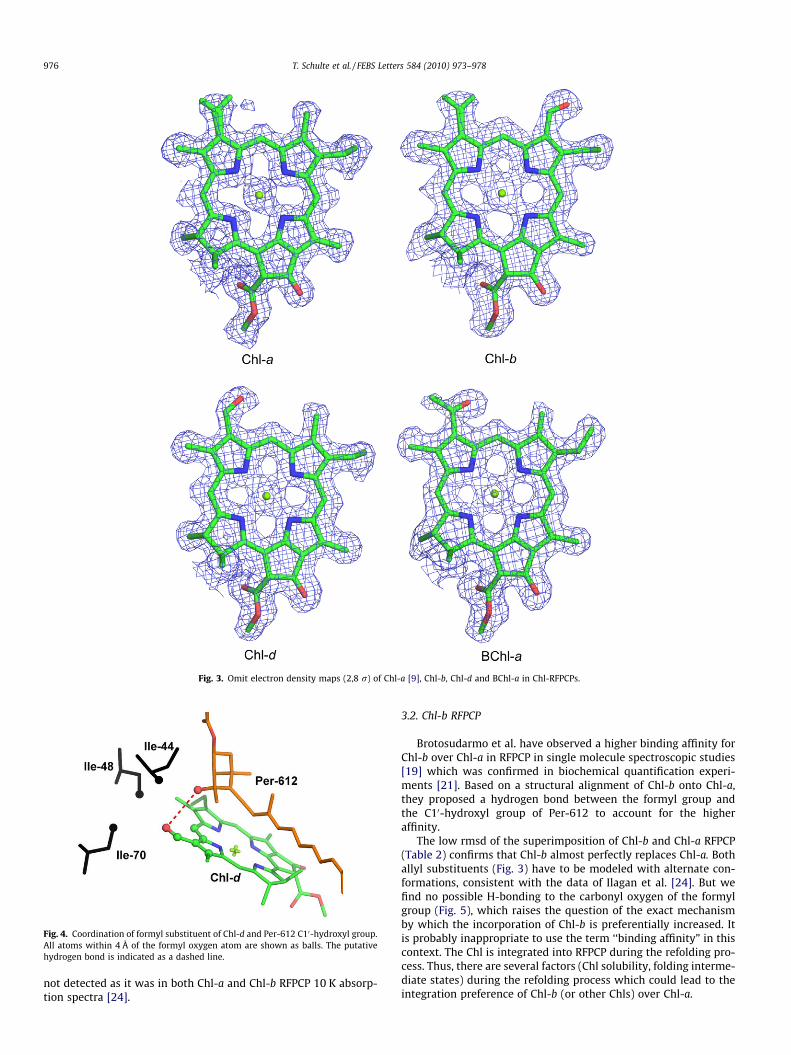
Fig. 3. Omit electron density maps (2,8 r) of Chl-a [9], Chl-b, Chl-d and BChl-a in Chl-RFPCPs.
Fig. 4. Coordination of formyl substituent of Chl-d and Per-612 C10-hydroxyl group.All atoms within 4 Å of the formyl oxygen atom are shown as balls. The putativehydrogen bond is indicated as a dashed line.
976 T. Schulte et al. / FEBS Letters 584 (2010) 973–978
not detected as it was in both Chl-a and Chl-b RFPCP 10 K absorp-tion spectra [24].
3.2. Chl-b RFPCP
Brotosudarmo et al. have observed a higher binding affinity forChl-b over Chl-a in RFPCP in single molecule spectroscopic studies[19] which was confirmed in biochemical quantification experi-ments [21]. Based on a structural alignment of Chl-b onto Chl-a,they proposed a hydrogen bond between the formyl group andthe C10-hydroxyl group of Per-612 to account for the higheraffinity.
The low rmsd of the superimposition of Chl-b and Chl-a RFPCP(Table 2) confirms that Chl-b almost perfectly replaces Chl-a. Bothallyl substituents (Fig. 3) have to be modeled with alternate con-formations, consistent with the data of Ilagan et al. [24]. But wefind no possible H-bonding to the carbonyl oxygen of the formylgroup (Fig. 5), which raises the question of the exact mechanismby which the incorporation of Chl-b is preferentially increased. Itis probably inappropriate to use the term ‘‘binding affinity” in thiscontext. The Chl is integrated into RFPCP during the refolding pro-cess. Thus, there are several factors (Chl solubility, folding interme-diate states) during the refolding process which could lead to theintegration preference of Chl-b (or other Chls) over Chl-a.

Fig. 5. Coordination of (A) Chl-b formyl and (B) BChl-a acetyl substituents which are highlighted by the black circles. All atoms within 4 Å of the formyl and acetyl oxygenatoms are shown as balls.
T. Schulte et al. / FEBS Letters 584 (2010) 973–978 977
3.3. BChl-a RFPCP
Polivka et al. observed a stochiometric ratio of 1:2 for Chl-a:BChl-a integration into RFPCP refolded with a 1:1 mixture ofthe Chls [20]. This observation is confirmed by the biochemicalquantification experiments of Brotosudarmo et al. who found thatacChl-a (difference between BChl-a and acChl-a is the saturation ofC7–C8 bond) is integrated twice as often as Chl-a [21]. Again, Bro-tosodarmu et al. proposed an H-bond to account for the observedpreference for acChl-a over Chl-a.
But BChl-a does not simply replace Chl-a in the binding niche.In contrast to the other Chls, the BChl-a atom positions of themacrocycle deviate remarkably from those of Chl-a by a rmsd of�0.5 Å (Table 2, Fig. 6). Regarding the proposed H-bond, the acetylsubstituent is shifted closer to the C10-hydroxyl group of Per-612,but at the same time rotated away from this putative hydrogenbond donor (Fig. 5). The orientation and distance of �4 Å betweenthe putative donor and acceptor atoms makes H-bond formationimprobable (Tested with WHATIF [23]).
Although the translational shift of BChl-a in the Chl bindingniche is not large (�0.4 Å), it changes the location of the porphyrinmacrocycle with regard to the peridinin molecules: BChl-a is clo-ser to Per-613 but further away from Per-611. Interestingly, Poli-vka et al. have reported a very fast time constant for the Per S1/ICT to BChl-a Qy energy transfer which was not predicted by För-ster theory (�3� faster than predicted), although the transfer ratesto Chl-b and Chl-d were predicted to within 15% [7]. Given the
Fig. 6. Superimposition of Pers-611, Pers-613 and the Chls from BChl-a RFPCP andChl-a RFPCP. The pigments of Chl-a RFPCP are in black.
knowledge of the geometry of the pigments in BChl-a-RFPCP andChl-a RFPCP it should be possible to recalculate the predictedtransfer rates using Förster theory and taking into account the ori-entation factor j [25]. But the dipole–dipole approximation forsuch a calculation is not valid for the pigment arrangement inPCP where the peridinins are in van der Waals contact with Chl[26]. A correct interpretation of the influence of the shifted BChl-a molecule on the energy transfer efficiency demands quantumchemical calculations using the exact description for the Coulombinteraction [27]. And although these structural changes mighthave only little effect on the rate compared to other spectral fea-tures of BChl-a [7], some effect seems to be present: the energytransfer from Per S1/ICT to acChl-a (same structure as BChl-a ex-cept for the saturated C7–C8 bond) Qy in acChl-a RFPCP is�1.5 times faster than predicted [7].
4. Conclusion
The Chl-RFPCP structures are almost identical to RFPCP re-folded with ‘‘native” Chl-a. Alternate conformations of the allylsubstituents at position R3 of Chl-a and Chl-b can account forthe split Chl Soret bands observed in 10 K absorption spectra[24]. The observed preference [19–21] of BChl-a/Chl-b over Chl-a for integration into RFPCP is not induced by postulated stabiliz-ing H-bonds to the polar substituents of BChl-a/Chl-b, but byparameters that determine the refolding process. Interpretationof recent spectroscopic [7,20] results in terms of a Förster-liketheory is valid, as the overall pigment arrangement is not chan-ged for the different Chls. Only the BChl-a macrocycle shows aremarkable rmsd of 0.5 Å, which might account to some small ex-tent for the discrepancy between measured and calculated energytransfer times [7].
Acknowledgements
We thank Dr. Tomas Polivka for helpful discussions and Dr. MinChen (Sydney University) for the gift of Acaryochloris cells. Wethank the beamline staff at ID23.1 and ID23.2 at the European Syn-chrotron Radiation Facility (ESRF) and at PXI at the Swiss LightSource (SLS) for assistance during data collection. This work wassupported by Sonderforschungsbereich 480 from the DeutscheForschungsgemeinschaft (Teilprojekt C6) and by a grant from theHelmhotz Society in the framework of the Virtual Institute for Bio-logical Structure Research (VH-VI-157) (to E.H and T.S.), the Re-search School of the Ruhr-University Bochum (T.S.) and theMacquarie University Research Development Office (R.G.H.).

978 T. Schulte et al. / FEBS Letters 584 (2010) 973–978
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.febslet.2010.01.041.
References
[1] Macpherson, A.N. and Hiller, R.G. (2003) Light-harvesting systems inchlorophyll c-containing algae. in: Advances in photosynthesis andrespiration. Vol. 13: Light harvesting antennas in Photosynthesis. (Green,B.R. and Parson, W.W., Eds.), pp. 323–352. Kluwer academic publishers,Dordrecht.
[2] Hofmann, E., Wrench, P.M., Sharples, F.P., Hiller, R.G., Welte, W. andDiederichs, K. (1996) Structural basis of light harvesting by carotenoids:peridinin–chlorophyll-protein from Amphidinium carterae. Science 272, 1788–1791.
[3] Polívka, T., Hiller, R.G. and Frank, H.A. (2007) Spectroscopy of the peridinin–chlorophyll-a protein: insight into light-harvesting strategy of marine algae.Arch. Biochem. Biophys. 458, 111–120.
[4] Polívka, T. and Sundström, V. (2004) Ultrafast dynamics of carotenoid excitedStates-from solution to natural and artificial systems. Chem. Rev. 104, 2021–2071.
[5] Mimuro, M., Nagashima, U., Takaichi, S., Nishimura, Y., Yamazaki, I. and Katoh,T. (1992) Molecular-structure and optical-properties of carotenoids for theinvivo energy-transfer function in the algal photosynthetic pigment system.Biochim. Biophys. Acta 1098, 271–274.
[6] Zigmantas, D., Hiller, R.G., Sundstrom, V. and Polivka, T. (2002) Carotenoid tochlorophyll energy transfer in the peridinin–chlorophyll-a-protein complexinvolves an intramolecular charge transfer state. Proc. Natl. Acad. Sci. USA 99,16760–16765.
[7] Polívka, T., Pascher, T., Sundström, V. and Hiller, R.G. (2005) Tuning energytransfer in the peridinin–chlorophyll complex by reconstitution with differentchlorophylls. Photosynth. Res. 86, 217–227.
[8] Miller, D.J., Catmull, J., Puskeiler, R., Tweedale, H., Sharples, F.P. and Hiller, R.G.(2005) Reconstitution of the peridinin––chlorophyll a protein (PCP): evidencefor functional flexibility in chlorophyll binding. Photosynth. Res. 86, 229–240.
[9] Schulte, T., Niedzwiedzki, D.M., Birge, R.R., Hiller, R.G., Polívka, T., Hofmann, E.and Frank, H.A. (2009) Identification of a single peridinin sensing Chl-aexcitation in reconstituted PCP by crystallography and spectroscopy. Proc.Natl. Acad. Sci. USA 106, 20764–20769.
[10] Kabsch, W. (1993) Automatic processing of rotation diffraction data fromcrystals of initially unknown symmetry and cell constants. J. Appl. Crystallogr.26, 795–800.
[11] Vagin, A. and Teplyakov, A. (1997) MOLREP: an automated program formolecular replacement. J. Appl. Crystallogr. 30, 1022–1025.
[12] The CCP4 suite: programs for protein crystallography (1994) Acta. Crystallogr.D Biol. Crystallogr. 50, 760–763.
[13] Murshudov, G.N., Vagin, A.A. and Dodson, E.J. (1997) Refinement ofmacromolecular structures by the maximum-likelihood method. ActaCrystallogr. D Biol. Crystallogr. 53, 240–255.
[14] Emsley, P. and Cowtan, K. (2004) Coot: model-building tools for moleculargraphics. Acta Crystallogr. D Biol. Crystallogr. 60, 2126–2132.
[15] Adams, P.D., Grosse-Kunstleve, R.W., Hung, L.W., Ioerger, T.R., McCoy, A.J.,Moriarty, N.W., Read, R.J., Sacchettini, J.C., Sauter, N.K. and Terwilliger, T.C.(2002) PHENIX: building new software for automated crystallographicstructure determination. Acta Crystallogr. D Biol. Crystallogr. 58, 1948–1954.
[16] Vaguine, A.A., Richelle, J. and Wodak, S.J. (1999) SFCHECK: a unified set ofprocedures for evaluating the quality of macromolecular structure-factor dataand their agreement with the atomic model. Acta Crystallogr. D Biol.Crystallogr. 55, 191–205.
[17] DeLano, W.L. (2002) The PyMOL User’s Manual, DeLano Scientific, Palo Alto,CA, USA.
[18] Kleywegt, G.J. and Jones, T.A. (1997) Model building and refinement practice.Methods Enzymol. 277, 208–230.
[19] Brotosudarmo, T.H.P., Hofmann, E., Hiller, R.G., Wörmke, S., Mackowski, S.,Zumbusch, A., Bräuchle, C. and Scheer, H. (2006) Peridinin–chlorophyll-protein reconstituted with chlorophyll mixtures: preparation, bulk and singlemolecule spectroscopy. FEBS Lett. 580, 5257–5262.
[20] Polívka, T., Pascher, T. and Hiller, R.G. (2008) Energy transfer in the peridinin–chlorophyll protein complex reconstituted with mixed chlorophyll sites.Biophys. J. 94, 3198–3207.
[21] Brotosudarmo, T.H.P., Mackowski, S., Hofmann, E., Hiller, R.G., Bräuchle, C. andScheer, H. (2008) Relative binding affinities of chlorophylls in peridinin–chlorophyll-protein reconstituted with heterochlorophyllous mixtures.Photosynth. Res. 95, 247–252.
[22] Di Valentin, M., Agostini, G., Salvadori, E., Ceola, S., Giacometti, G.M., Hiller,R.G. and Carbonera, D. (2009) Triplet-triplet energy transfer in peridinin–chlorophyll a-protein reconstituted with Chl a and Chl d as revealed byoptically detected magnetic resonance and pulse EPR: Comparison with thenative PCP complex from Amphidinium carterae. Biochim. Biophys. Acta 1787,168–175.
[23] Vriend, G. (1990) WHAT IF: a molecular modeling and drug design program. J.Mol. Graph. 8, 52–56. 29.
[24] Ilagan, R.P., Chapp, T.W., Hiller, R.G., Sharples, F.P., Polívka, T. and Frank, H.A.(2006) Optical spectroscopic studies of light-harvesting by pigment-reconstituted peridinin–chlorophyll-proteins at cryogenic temperatures.Photosynth. Res. 90, 5–15.
[25] Scholes, G. (2003) Long-range resonance energy transfer in molecular systems.Annu. Rev. Phys. Chem. 54, 87. 57.
[26] Damjanovic, A., Ritz, T. and Schulten, K. (2000) Excitation transfer in theperidinin–chlorophyll-protein of Amphidinium carterae. Biophys. J. 79, 1695–1705.
[27] Hu, X., Ritz, T., Damjanovic, A., Autenrieth, F. and Schulten, K. (2002)Photosynthetic apparatus of purple bacteria. Q. Rev. Biophys. 35, 1–62.




















