
Working memory in the chick: parallel and lateralized mechanisms for encoding of object- and...
9
Behavioural Brain Research 157 (2005) 1–9 Research report Working memory in the chick: parallel and lateralized mechanisms for encoding of object- and position-specific information Lucia Regolin a,∗ , Beatrice Garzotto a , Rosa Rugani a , Piero Pagni b , Giorgio Vallortigara b,c,1 a Department of General Psychology, University of Padua, Via Venezia 8, 35131 Padova, Italy b Department of Psychology, University of Trieste, Via S. Anastasio 12, 34123 Trieste, Italy c BRAIN Center for Neuroscience, University of Trieste, Via S. Anastasio 12, 34123 Trieste, Italy Received 11 March 2004; received in revised form 31 May 2004; accepted 1 June 2004 Available online 30 July 2004 Abstract Working memory of chicks was probed in a delayed-response task, with either object- or position-specific information available in order to locate a goal that had disappeared behind one of two screens in a test arena. When the position of the correct screen was the only available cue (i.e., the two screens were identical) binocular and monocular chicks could easily locate the goal, and the same occurred when the visual characteristics of the screens (no matter what their spatial position) were the only relevant information available and when object- and position- specific cues were both available and in agreement (i.e., the two screens were different and maintained fixed spatial locations). In contrast, when object- and position-specific cues were available but were put in conflict (the two screens were different and were swapped during the delay time) left-eyed and binocular chicks went to the correct position, whereas right-eyed chicks seemed to choose both the correct spatial position and the correct object cue to the same degree. When during the delay the correct screen was substituted by an entirely novel screen, no left–right asymmetry associated with response to novelty was observed; monocular chicks preferentially approached the novel screen in the correct position, whereas binocular chicks did not show any clear choice. The results suggest that both object- and position-specific information is available to the two cerebral hemispheres in working memory tasks; however, when a conflict between cues arises, the right hemisphere preferentially attends to position-specific cues, whereas the left hemisphere tends to attend to object-specific cues. © 2004 Elsevier B.V. All rights reserved. Keywords: Working memory; Domestic chick; Brain lateralization 1. Introduction In recent years, birds have started to be used quite systematically as system models for the investigation of lateralized processes in spatial cognition (reviews in [14,18,19,22,28,33]). Some characteristics shared by the most commonly used avian species make them particularly suited for lateralization studies: given that the fibres of the avian optic nerve cross over completely and there is a rel- ∗ Corresponding author. Tel.: +39 049 8276950; fax: +39 049 8276600. E-mail addresses: [email protected] (L. Regolin), [email protected] (G. Vallortigara). 1 Tel.: +39 040 6762724; fax: +39 040 4528022. atively paucity of interhemispheric connections compared with mammals, each hemisphere of the avian brain can be tested separately using the method of temporary monocu- lar occlusion (reviews in [2,6,7,12,20,21]). Experiments us- ing this technique have revealed clear lateralization in birds. A well-demonstrated finding concerns the superiority of the right eye and left brain hemisphere in the visual discrimina- tion of food and non-food objects; this has been shown to occur identically in chicks [15], pigeons [11], zebra finches [1] and quails [27]. Evidence concerning a dominant role of the left eye (right hemisphere) in spatial cognition has also been obtained. Rashid and Andrew [17] first discovered more systematic and spatially-focused search for food when chicks used their 0166-4328/$ – see front matter © 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.bbr.2004.06.012
Transcript of Working memory in the chick: parallel and lateralized mechanisms for encoding of object- and...

Behavioural Brain Research 157 (2005) 1–9
Research report
Working memory in the chick: parallel and lateralized mechanismsfor encoding of object- and position-specific information
Lucia Regolina,∗, Beatrice Garzottoa, Rosa Rugania,Piero Pagnib, Giorgio Vallortigarab,c,1
a Department of General Psychology, University of Padua, Via Venezia 8, 35131 Padova, Italyb Department of Psychology, University of Trieste, Via S. Anastasio 12, 34123 Trieste, Italy
c BRAIN Center for Neuroscience, University of Trieste, Via S. Anastasio 12, 34123 Trieste, Italy
Received 11 March 2004; received in revised form 31 May 2004; accepted 1 June 2004Available online 30 July 2004
Abstract
Working memory of chicks was probed in a delayed-response task, with either object- or position-specific information available in ordert nly availablec n the visualc d position-s In contrast,w during thed rrect spatialp ovel screen,n ovel screeni ion-specifici s, the righth©
K
1
so[msa
v
redn beocu--irds.the
ina-n tos
ightined.tictheir
0d
o locate a goal that had disappeared behind one of two screens in a test arena. When the position of the correct screen was the oue (i.e., the two screens were identical) binocular and monocular chicks could easily locate the goal, and the same occurred wheharacteristics of the screens (no matter what their spatial position) were the only relevant information available and when object- anpecific cues were both available and in agreement (i.e., the two screens were different and maintained fixed spatial locations).hen object- and position-specific cues were available but were put in conflict (the two screens were different and were swappedelay time) left-eyed and binocular chicks went to the correct position, whereas right-eyed chicks seemed to choose both the coosition and the correct object cue to the same degree. When during the delay the correct screen was substituted by an entirely no left–right asymmetry associated with response to novelty was observed; monocular chicks preferentially approached the n
n the correct position, whereas binocular chicks did not show any clear choice. The results suggest that both object- and positnformation is available to the two cerebral hemispheres in working memory tasks; however, when a conflict between cues ariseemisphere preferentially attends to position-specific cues, whereas the left hemisphere tends to attend to object-specific cues.2004 Elsevier B.V. All rights reserved.
eywords:Working memory; Domestic chick; Brain lateralization
. Introduction
In recent years, birds have started to be used quiteystematically as system models for the investigationf lateralized processes in spatial cognition (reviews in
14,18,19,22,28,33]). Some characteristics shared by theost commonly used avian species make them particularly
uited for lateralization studies: given that the fibres of thevian optic nerve cross over completely and there is a rel-
∗ Corresponding author. Tel.: +39 049 8276950; fax: +39 049 8276600.E-mail addresses:[email protected] (L. Regolin),
[email protected] (G. Vallortigara).1 Tel.: +39 040 6762724; fax: +39 040 4528022.
atively paucity of interhemispheric connections compawith mammals, each hemisphere of the avian brain catested separately using the method of temporary monlar occlusion (reviews in[2,6,7,12,20,21]). Experiments using this technique have revealed clear lateralization in bA well-demonstrated finding concerns the superiority ofright eye and left brain hemisphere in the visual discrimtion of food and non-food objects; this has been showoccur identically in chicks[15], pigeons[11], zebra finche[1] and quails[27].
Evidence concerning a dominant role of the left eye (rhemisphere) in spatial cognition has also been obtaRashid and Andrew[17] first discovered more systemaand spatially-focused search for food when chicks used
166-4328/$ – see front matter © 2004 Elsevier B.V. All rights reserved.oi:10.1016/j.bbr.2004.06.012

2 L. Regolin et al. / Behavioural Brain Research 157 (2005) 1–9
left, rather than their right, eye. Somewhat more complicatedresults have been reported for spatial navigation in homingpigeons, that showed a right eye/left hemisphere superiorityin monocular tests[26]. Unilateral lesions to the hippocam-pal formation revealed, however, a more complex pattern.Although lesion studies identified the left, but not the righthippocampus as critical for navigational map learning[9],both hemispheres seemed to participate in learning to nav-igate over previously experienced terrain reliant on famil-iar landmarks[10]. Very recent work carried out by Kahnand Bingman[13] showed that the right and left hippocam-pus actually make different spatial memory contributions to-wards space representation: the left hippocampus seems tobe able to support a representation of goal locations relianton landmarks within bounded, experienced space, whereasthe right hippocampus is exclusively involved in represent-ing goal locations reliant on global spatial features (review in[3]). Somewhat similar results have been reported for chickstested in reference memory tasks with occlusion of one eye:chicks using only their right eye attended mainly to localobject-specific cues, whereas chicks using only their left eyeattended mainly to global, geometric cues[23,25,29,34,35].Unilateral lesions to the hippocampus in the chick also con-firmed this basic pattern[24].
A very limited amount of information is available for ther kingm ff hada as at in-t rrectf laceda singp andu righte h evi-d How-e orf ano lat-t merb nce.
icks,m em-o ever,V an“ top l (ani ento or theg icksa n oft ise ork-i d toc ory
tasks when a conflict arises between position-specific andobject-specific information.
2. Experiment
The basic procedure was similar to the delayed-responsetask developed by Vallortigara et al.[36]. Chicks were firstshown an imprinted object being hidden behind one of twodifferent opaque screens and were then allowed to search forthe goal after a 30 s delay. Several trials were administeredto each chick. In each trial, the position of the screen behindwhich the goal object was hidden was changed at randomand, during the delay, the left–right position of the screens wasswapped so that the ‘correct’ screen was located in the wrongposition and the ‘incorrect’ screen in the correct position. Theaim of the present research was to check whether right- andleft-eyed chicks would rely on different cues for the search,i.e. the position of the screen for left-eyed chicks and its visualcharacteristics for right-eyed chicks.
3. Materials and methods
3.1. Subjects
romto vi-c (PD)I al,c me-c gly,a( gesw ps( ge, ints t) ands thatt enti
3
cribedi di erec sd ced8 ed in ac ooru ena,a 0 cm× vend ning
ole of the two hemispheres of the avian brain during woremory tasks. Clayton and Krebs[4] tested the memory o
ood-storing and non-food-storing birds for feeders thattrial-unique location in an experimental room as well
rial-unique colour pattern. When, after a short retentionerval, birds were given dissociation tests in which the coeeder changed its position and a different feeder was pt the original location, all birds preferentially searched uosition-based cues when tested with only their left eyesing feeder-specific cues when tested with only theirye. These results seem to correspond quite closely witence obtained in reference memory tasks (see above).ver, Prior and Gunturkun [16] trained pigeons to search food in a maze in a spatial working memory task and inbject-specific working memory task and found that the
er task mainly involved the left hemisphere but in the foroth hemispheres contributed equally to spatial performa
No research has been conducted on this topic in chainly because no behavioural techniques for working mry tasks were available for this species. Recently, howallortigara et al.[36] developed such techniques usingunconventional” reward. Basically, chicks were trainederform a delayed response task. A very attractive goa
mprinted object) was hidden behind one of two differpaque screens and the chicks were allowed to search foal after different time delays. Results showed that chre exceptionally good at this task, retaining the locatio
he goal up to intervals of 60 s[36]. In this paper, we used thxperimental procedure to investigate lateralization of w
ng memory in chicks. In particular, we were interesteheck whether lateralization only occurs in working-mem
The subjects were 180 Hybro (a local variety derived fhe White Leghorn breed) domestic chicks (Gallus gallus),btained weekly from a local commercial hatchery (Aola Berlanda Edio and C. Snc, Carmignano di Brentataly) when they were only a few hours old. At their arrivhicks were immediately housed in standard metal hoages (28 cm× 32 cm large, 40 cm high) and reared sint a controlled temperature (about 28–31◦C) and humidity68%), with food and water available ad libitum. The caere constantly (24 h/day) lit by 36 W fluorescent lam
placed 15 cm above each cage). Within each rearing cahe centre, a small red plastic ball (4 cm× 3 cm× 3 cm) wasuspended by a fine thread (at about chicks’ head heigherved as an imprinting object (previous studies showedhis stimulus is very effective in producing social attachmn this strain of chicks; see[32].
.2. Apparatus and procedure
The apparatus and general procedure have been desn details elsewhere[36]. The apparatus (Fig. 1a) was locaten a nearby room, in which temperature and humidity wontrolled (respectively of 25◦C and 70%); the room waark, except for the light coming from a 40 W lamp pla0 cm above the apparatus. The test apparatus consistircular arena (95 cm in diameter, 30 cm height) with the flniformly covered by a white plastic sheet. Within the ardjacent to the outer wall, was positioned a small box (120 cm× 20 cm), where the chick was confined for a gi
elay period during the test phase. The side of the confi

L. Regolin et al. / Behavioural Brain Research 157 (2005) 1–9 3
Fig. 1. (a) The experimental apparatus consisted in a wide circular arena.At test the goal object was made to disappear behind one of two screensand the chicks were required to rejoin the object behind the correct screen.The screens are blue with a yellow “X” on them. The picture representsthe “position cue” condition, since the two screens are identical the onlyuseful information available to locate the goal object is the spatial positionof the correct screen. (b) The full set of screens employed in the differentexperimental conditions (the only missing screen is the one used as novelin the “novelty control” condition, such screen was uniformly dark yellowin colour, with no pattern present). A couple of identical screens was usedfor each type represented in the picture. The first screen from the left isblue with a yellow ‘X’, the second screen is white with a red “Florence lily”pattern, these two screens (first and second from the left) were used in all fiveexperimental conditions. Only for the condition “object cue” all six types ofscreens were alternatively used (see the text for more details). The coloursof the remaining screens represented are: the third screen from the left is redwith grey triangles on it, the fourth screen is yellow with black horizontalbars, the fifth screen is pink with black diagonal fine lines, and the last screenis green with pink rectangles on it.
box facing the centre of the arena consisted of a removableclear glass sheet (20 cm× 10 cm), the experimenter could liftit from above in order to release the test chick into the arena atthe end of the delay. An opaque partition was also used duringthe test, in order to prevent the chick from seeing the arenaduring the delay time (the opaque partition was superimposedon the glass sheet at the beginning of the delay and, at the endof the delay, the two partitions were simultaneously lifted bythe experimenter). Two opaque cardboard screens (16 cm×8 cm; with 3-cm sides bent back to prevent the chicks fromspotting the hidden ball) were positioned in the centre of thearena, symmetrically with respect to the confining box, 20 cmapart from each other and 35 cm away from the closest sideof the confining box. According to the different experimen-tal conditions, the screens could be identical to each other
or visually different in colour and patterning (two types ofscreens were mainly used: one was blue with a yellow ‘X’,and the other white with a red “Florence lily” pattern, morethan two types of screens were employed only for one of theexperimental conditions, see below andFig. 1b).
On day 4 of life, each chick underwent a preliminary train-ing. For the training, only one of the screens was used (in thecase of two different screens present at test, the blue one wasused for training half of the animals and the white one fortraining the other half), and positioned in front of, and 35 cmaway from the confining box. The chick, together with theimprinting red ball, was at first placed within the arena for acouple of minutes, free to move around and get acquainted tothe novel environment. The rearing ball, held from above (bya fine thread) by the experimenter (not visible to the chick),was slowly moved in the arena and then made to disappearbehind the training screen. This procedure was repeated a fewtimes, until the chick promptly responded by following andrejoining the ball behind the screen. Thereafter, the test chickwas confined in the box. The red ball, now outside of the box,was slowly moved by the experimenter until it disappearedbehind the screen; the experimenter took care that the chickwas watching in full the trajectory of the ball. The chick wasthen immediately set free in the arena by removing the trans-parent front partition, and every time the chick rejoined theb d tos rain-i creenw rall,d ouldl thei hisp bout2 er ofs
oths finedi ere itc f thet aquep ch aw s fora lassp thee ball.T chickw ehindw e wasc g. Ifa eenst ered.C wered f thec r the“ 15o rded
all behind the screen as a reinforcement it was allowepend a few seconds next to its artificial social partner. Tng ended when the chick reached the ball behind the sithin 10 s from release for six times consecutively. Oveepending on the chick’s behaviour, the training phase c
ast from 10 to 20 min (chicks that showed little interest inmprinted object, i.e. poor ball following behaviour, in thase, were discarded from the study, this occurred in a5% of cases, such chicks are not included in the numbubjects described below).
Testing took place on day 4, 2 h after the training. Bcreens were positioned in the arena. The chick was conn the small cage, behind the clear glass sheet, from whould see and track the ball disappearing behind one owo screens. After the disappearance of the ball, an opartition was located in front of the glass sheet, in suay of preventing the chick from seeing the two screentime delay of 30 s. After the delay, the opaque and g
artitions were both simultaneously lifted from above byxperimenter and the chick was allowed to search for thehe first screen approached and circled around by theas recorded, if such screen corresponded to the one bhich the ball had disappeared then the chick’s responsonsidered correct, otherwise it was considered wronfter 5 min the chick had not approached any of the scr
he response was considered null and not further considhicks with more than three consecutive null responsesiscarded from the study, this occurred in about 20% oases, no matter the experimental condition (except foobject cue” condition in which one third of the chicks —ut of 45 — were discarded for this reason); the disca

4 L. Regolin et al. / Behavioural Brain Research 157 (2005) 1–9
chicks are not included in the number of subjects describedin the procedure.
Each chick underwent 16 consecutive trials, the screenbehind which the ball disappeared was varied from trial totrial according to a semi-random sequence[8].
3.3. Testing conditions
There were five experimental conditions.In the first condition (“position cue”;N = 30 chicks, 15
males and 15 females) the two screens were physically iden-tical (i.e. two blue screens with a yellow ‘X’ for half of theanimals, and two white screens with a red “Florence lily”pattern for the other half) so that only the positional cue wasavailable to the chicks in order to identify the correct screen.
In the second condition (“object cue”;N = 30 chicks, 15males and 15 females) only non spatial cues could be used toidentify the correct screen. This was obtained by employingseveral screens of different colour and pattern. In each trialof the test the chick saw the ball disappearing behind onesingle, centrally located screen (much like what happenedduring the training phase), then, during the delay, while thechick’s sight was prevented by the opaque partition, a sec-ond screen of a different colour was introduced in the arena,and the two screens were positioned in the standard testingf eens,a ballh chickc romt ont re-s ensw o des uredi sentc ionso yed.O trialsa ct ans somet
3 wered eensu eree herw iona
ectc twos ver,t thes the rem , thea in thec , and
a screen that was in the wrong position but had the correctcolour and patterning. In this last condition, by convention,the choice of the correct position was considered correct.
The fifth condition was a control for the effects of novelty(“novelty”; N= 48 chicks, 24 males and 24 females). In fact,in the second and in the fourth conditions the chick’s choicecould have been affected by the changing in the appearance ofthe setting that occurred during the delay (i.e., in the secondcondition a second screen appeared, in the fourth conditionthe two screens were swapped). To check for any effect due tonovelty per se (rather than, for example, to a conflict betweenthe use of colour and position cues), in the fifth, control, con-dition two different screens were used and, during the delay,the correct screen was removed and a new screen was intro-duced instead of it (a completely yellow screen was used). Inthis way, although chicks were confronted with an entirelynovel screen, the other screen remained unchanged in bothcolour and position and thus no contrast between colour andposition occurred. (In a sense, contrast was still present, butlimited only to the positive screen, which was in the correctposition but different in its visual appearance; however, nocontrast between the characteristics of the two screens wasapparent as in condition four.)
For each of the five conditions, separate groups of chickswere tested in binocular and in monocular conditions ofv oft tickyt hema note dlinga bero
3
ei-t ls oft oices
TN isiona : lefte entalc
C
P
O
P
C 7
N
ashion described above. The chick faced now two scrnd was required to identify the one behind which thead disappeared, in order to succeed in such task theould only rely on the objectual (not positional) cues. Frial to trial the correct screen at test was never twicehe same side, but it alternated from left to right withpect to the chick’s position. A total of six different screere used: four new screens was used besides the twcribed for the previous condition (all screens are pictn Fig. 1b), such new screens were used only for the preondition, whereas in all the other experimental conditnly the two previously described screens were emplone same screen never appeared in two consecutivell screens appeared several times, some times as correome times as wrong ones, some times on the right andimes on the left of the chick’s starting position.
In the third condition (“position plus object cues”;N =0 chicks, 15 males and 15 females) the two screensifferent in colour and pattern. The same two different scrsed for different chicks in the “position cue” condition wmployed (i.e. one was blue with a yellow ’X’, and the othite with a red “Florence lily” pattern) so that both positnd colour might be used to identify the correct screen.
In the fourth condition (“contrasting position and objues”;N = 42 chicks, 21 males and 21 females) thecreens were different as in the third condition; howehis time, during the delay, while the chick could not seecreens, the experimenter swapped them, so that, afteroval of the opaque partition and the release of the chicknimal was forced to choose between a screen that wasorrect position but had the wrong colour and patterning
-
,d
-
ision. Monocular testing was carried out by meansemporary eye patches made of special, removable, sape, worn by the chicks 20 min before testing to get tcquainted to the new situation (binocular chicks wereye-patched but received an equivalent amount of hannd acquainting time as did monocular chicks). The numf chicks tested in the various conditions is shown inTable 1.
.4. Data analyses
The choice taken by each chick when circling aroundher a correct or a wrong screen in each of the 16 triahe test was considered. The percentage of correct ch
able 1umber of female and male chicks employed, for each condition of vt test (BIN: binocular; RE: right eye in use and left eye patched; LEye in use and right eye patched), and each of the five different experimonditions of testing (described in the text)
ondition Sex BIN RE LE
osition cue Male 5 5 5Female 5 5 5
bject cue Male 5 5 5Female 5 5 5
osition plus object cue Male 5 5 5Female 5 5 5
ontrasting position and object cues Male 7 7Female 7 7 7
ovelty control Male 8 8 8Female 8 8 8
L. Regolin et al. / Behavioural Brain Research 157 (2005) 1–9 5
was computed for each chick through the equation: numberof correct choices/total number of choices expressed by thechick (i.e. correct plus incorrect ones, the null trials are notconsidered)× 100. Each chick’s score could range from 0%(exclusive choice of the wrong screen) to 100% (the correctscreen is chosen every time); 50% represents no preferentialchoice for correct or incorrect screen.
Data were analysed through analysis of variance(ANOVA) separately run for each experimental condition,with sex (male and female) of the chicks and the eye (binoc-ular, BIN; left eye in use, LE, and right eye in use, RE) as be-tween subjects factors, and the percentage of correct choicesas the dependent variable. Significant departures from chancelevel (50%) were estimated by one-sample two-tailedt-tests.
4. Results
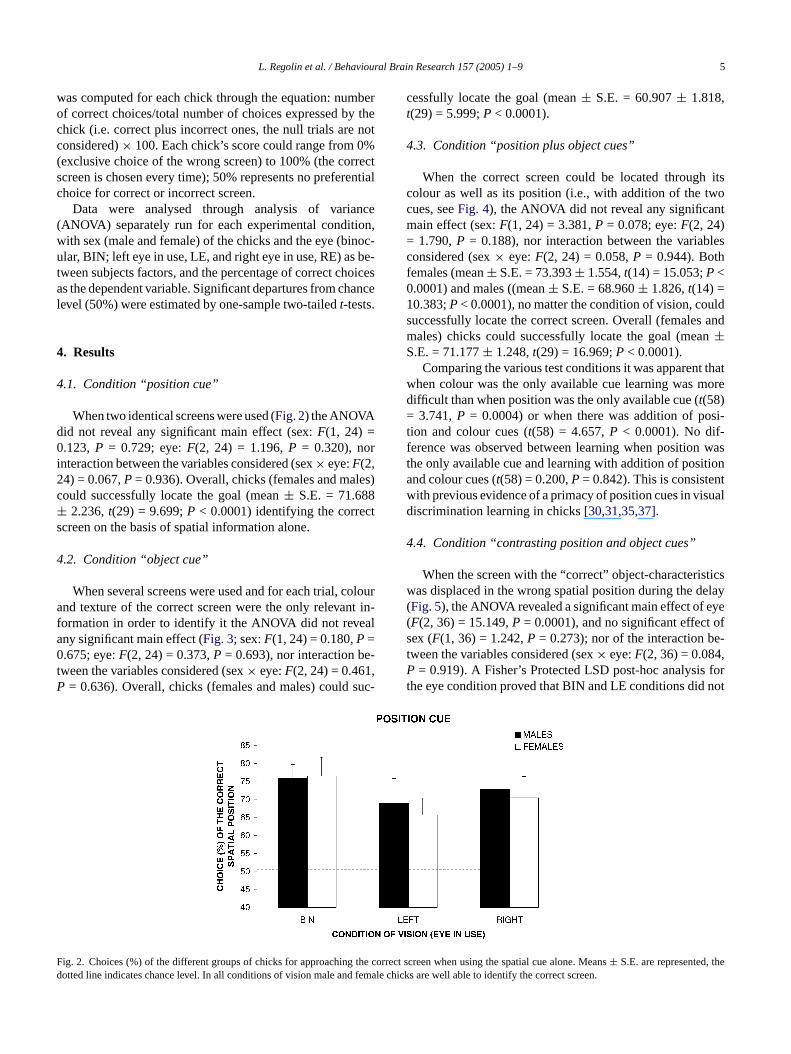
4.1. Condition “position cue”
When two identical screens were used (Fig. 2) the ANOVAdid not reveal any significant main effect (sex:F(1, 24) =0.123,P = 0.729; eye:F(2, 24) = 1.196,P = 0.320), norinteraction between the variables considered (sex× eye:F(2,24) = 0.067,P= 0.936). Overall, chicks (females and males)c± cts
4
oloura t in-f ala0 e-t ,P suc-
F the cor hed female
cessfully locate the goal (mean± S.E. = 60.907± 1.818,t(29) = 5.999;P < 0.0001).
4.3. Condition “position plus object cues”
When the correct screen could be located through itscolour as well as its position (i.e., with addition of the twocues, seeFig. 4), the ANOVA did not reveal any significantmain effect (sex:F(1, 24) = 3.381,P = 0.078; eye:F(2, 24)= 1.790,P = 0.188), nor interaction between the variablesconsidered (sex× eye:F(2, 24) = 0.058,P = 0.944). Bothfemales (mean± S.E. = 73.393± 1.554,t(14) = 15.053;P<0.0001) and males ((mean± S.E. = 68.960± 1.826,t(14) =10.383;P< 0.0001), no matter the condition of vision, couldsuccessfully locate the correct screen. Overall (females andmales) chicks could successfully locate the goal (mean±S.E. = 71.177± 1.248,t(29) = 16.969;P < 0.0001).
Comparing the various test conditions it was apparent thatwhen colour was the only available cue learning was moredifficult than when position was the only available cue (t(58)= 3.741,P = 0.0004) or when there was addition of posi-tion and colour cues (t(58) = 4.657,P < 0.0001). No dif-ference was observed between learning when position wasthe only available cue and learning with addition of positionand colour cues (t(58) = 0.200,P= 0.842). This is consistentw uald
4
ticsw elay( ye( ofs e-t ,P fort not
ould successfully locate the goal (mean± S.E. = 71.6882.236,t(29) = 9.699;P < 0.0001) identifying the corre
creen on the basis of spatial information alone.
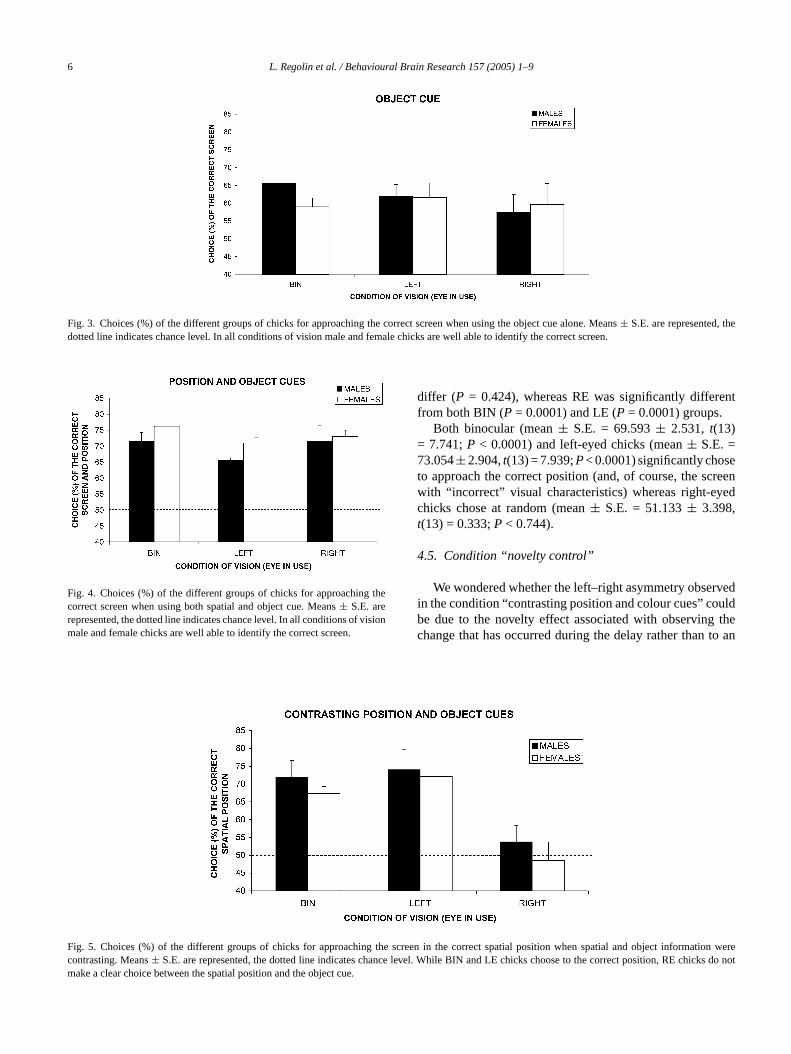
.2. Condition “object cue”
When several screens were used and for each trial, cnd texture of the correct screen were the only relevan
ormation in order to identify it the ANOVA did not reveny significant main effect (Fig. 3; sex:F(1, 24) = 0.180,P=.675; eye:F(2, 24) = 0.373,P = 0.693), nor interaction b
ween the variables considered (sex× eye:F(2, 24) = 0.461= 0.636). Overall, chicks (females and males) could
ig. 2. Choices (%) of the different groups of chicks for approachingotted line indicates chance level. In all conditions of vision male and
rect screen when using the spatial cue alone. Means± S.E. are represented, tchicks are well able to identify the correct screen.
ith previous evidence of a primacy of position cues in visiscrimination learning in chicks[30,31,35,37].
.4. Condition “contrasting position and object cues”
When the screen with the “correct” object-characterisas displaced in the wrong spatial position during the d
Fig. 5), the ANOVA revealed a significant main effect of eF(2, 36) = 15.149,P = 0.0001), and no significant effectex (F(1, 36) = 1.242,P = 0.273); nor of the interaction bween the variables considered (sex× eye:F(2, 36) = 0.084
= 0.919). A Fisher’s Protected LSD post-hoc analysishe eye condition proved that BIN and LE conditions did
6 L. Regolin et al. / Behavioural Brain Research 157 (2005) 1–9
Fig. 3. Choices (%) of the different groups of chicks for approaching the correct screen when using the object cue alone. Means± S.E. are represented, thedotted line indicates chance level. In all conditions of vision male and female chicks are well able to identify the correct screen.
Fig. 4. Choices (%) of the different groups of chicks for approaching thecorrect screen when using both spatial and object cue. Means± S.E. arerepresented, the dotted line indicates chance level. In all conditions of visionmale and female chicks are well able to identify the correct screen.
Fig. 5. Choices (%) of the different groups of chicks for approaching thecontrasting. Means± S.E. are represented, the dotted line indicates chancmake a clear choice between the spatial position and the object cue.
differ (P = 0.424), whereas RE was significantly differentfrom both BIN (P = 0.0001) and LE (P = 0.0001) groups.
Both binocular (mean± S.E. = 69.593± 2.531, t(13)= 7.741;P < 0.0001) and left-eyed chicks (mean± S.E. =73.054±2.904,t(13) = 7.939;P< 0.0001) significantly choseto approach the correct position (and, of course, the screenwith “incorrect” visual characteristics) whereas right-eyedchicks chose at random (mean± S.E. = 51.133± 3.398,t(13) = 0.333;P < 0.744).
4.5. Condition “novelty control”
We wondered whether the left–right asymmetry observedin the condition “contrasting position and colour cues” couldbe due to the novelty effect associated with observing thechange that has occurred during the delay rather than to an
ation wecks do not
screen in the correct spatial position when spatial and object informree level. While BIN and LE chicks choose to the correct position, RE chi
L. Regolin et al. / Behavioural Brain Research 157 (2005) 1–9 7
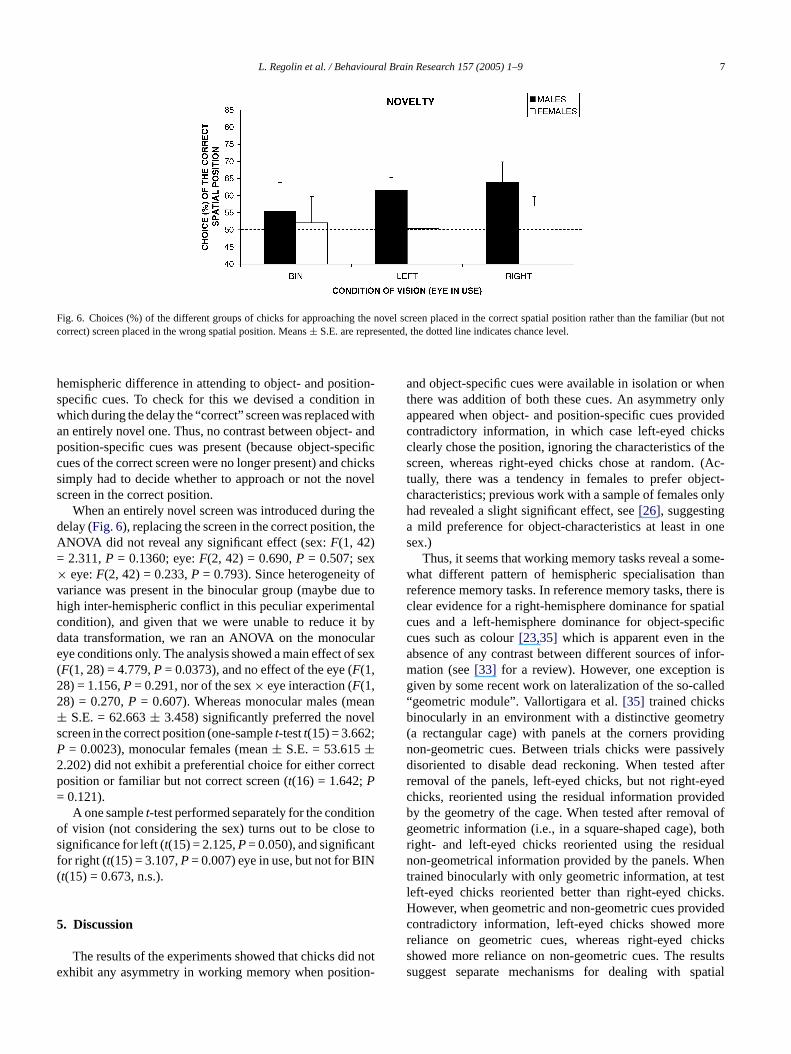
Fig. 6. Choices (%) of the different groups of chicks for approaching the novel screen placed in the correct spatial position rather than the familiar (but notcorrect) screen placed in the wrong spatial position. Means± S.E. are represented, the dotted line indicates chance level.
hemispheric difference in attending to object- and position-specific cues. To check for this we devised a condition inwhich during the delay the “correct” screen was replaced withan entirely novel one. Thus, no contrast between object- andposition-specific cues was present (because object-specificcues of the correct screen were no longer present) and chickssimply had to decide whether to approach or not the novelscreen in the correct position.
When an entirely novel screen was introduced during thedelay (Fig. 6), replacing the screen in the correct position, theANOVA did not reveal any significant effect (sex:F(1, 42)= 2.311,P = 0.1360; eye:F(2, 42) = 0.690,P = 0.507; sex× eye:F(2, 42) = 0.233,P = 0.793). Since heterogeneity ofvariance was present in the binocular group (maybe due tohigh inter-hemispheric conflict in this peculiar experimentalcondition), and given that we were unable to reduce it bydata transformation, we ran an ANOVA on the monoculareye conditions only. The analysis showed a main effect of sex(F(1, 28) = 4.779,P= 0.0373), and no effect of the eye (F(1,28) = 1.156,P= 0.291, nor of the sex× eye interaction (F(1,28) = 0.270,P = 0.607). Whereas monocular males (mean± S.E. = 62.663± 3.458) significantly preferred the novelscreen in the correct position (one-samplet-testt(15) = 3.662;P = 0.0023), monocular females (mean± S.E. = 53.615±2.202) did not exhibit a preferential choice for either correctp=
iono e tos tf N(
5
note on-
and object-specific cues were available in isolation or whenthere was addition of both these cues. An asymmetry onlyappeared when object- and position-specific cues providedcontradictory information, in which case left-eyed chicksclearly chose the position, ignoring the characteristics of thescreen, whereas right-eyed chicks chose at random. (Ac-tually, there was a tendency in females to prefer object-characteristics; previous work with a sample of females onlyhad revealed a slight significant effect, see[26], suggestinga mild preference for object-characteristics at least in onesex.)
Thus, it seems that working memory tasks reveal a some-what different pattern of hemispheric specialisation thanreference memory tasks. In reference memory tasks, there isclear evidence for a right-hemisphere dominance for spatialcues and a left-hemisphere dominance for object-specificcues such as colour[23,35] which is apparent even in theabsence of any contrast between different sources of infor-mation (see[33] for a review). However, one exception isgiven by some recent work on lateralization of the so-called“geometric module”. Vallortigara et al.[35] trained chicksbinocularly in an environment with a distinctive geometry(a rectangular cage) with panels at the corners providingnon-geometric cues. Between trials chicks were passivelydisoriented to disable dead reckoning. When tested afterr yedc idedb al ofg bothr dualn hent estl icks.H idedc orer ickss sultss atial
osition or familiar but not correct screen (t(16) = 1.642;P0.121).A one samplet-test performed separately for the condit
f vision (not considering the sex) turns out to be closignificance for left (t(15) = 2.125,P= 0.050), and significanor right (t(15) = 3.107,P= 0.007) eye in use, but not for BIt(15) = 0.673, n.s.).
. Discussion
The results of the experiments showed that chicks didxhibit any asymmetry in working memory when positi
emoval of the panels, left-eyed chicks, but not right-ehicks, reoriented using the residual information provy the geometry of the cage. When tested after removeometric information (i.e., in a square-shaped cage),ight- and left-eyed chicks reoriented using the resion-geometrical information provided by the panels. W
rained binocularly with only geometric information, at teft-eyed chicks reoriented better than right-eyed chowever, when geometric and non-geometric cues provontradictory information, left-eyed chicks showed meliance on geometric cues, whereas right-eyed chhowed more reliance on non-geometric cues. The reuggest separate mechanisms for dealing with sp

8 L. Regolin et al. / Behavioural Brain Research 157 (2005) 1–9
reorientation problems, with the right hemisphere takingcharge of large-scale geometry of the environment and withboth hemispheres taking charge of local, non-geometric cueswhen available in isolation, but with a predominance of theleft hemisphere when competition between geometric andnon-geometric information occurs (see also[29]).
Here, somewhat similar seems to occur as to the role ofthe right hemisphere, which does not reveal any dominancewhen the cues are available in isolation, but that shows cleardominance when competition between object- and position-specific cues occurs.
It is likely that under short intervals of retention informa-tion concerning the “what” and “where” is retained in bothhemispheres (in contrast, see[5] for evidence that integra-tion of what and where can be lateralized in reference mem-ory tasks). Lateralization, however, seems to arise even inworking memory when a decision about the use of availableinformation should be undertaken; apparently, the right hemi-sphere takes charge of control of behaviour in these cases (asdemonstrated by the fact that binocular chicks behave as left-eyed chicks) also revealing its intrinsic tendency to attend tospatial cues.
From a comparative perspective, it is unclear, judgingfrom the data with the most studied avian species whetherit is the nature of the task or species-differences that makesa herf othh emo e foro eresf dr and( ob-j jectc om-p ingt bjecc iewi ri-e (andt re incT comm wasp on-t area ltricias iffer-e hems (i.ei ject-s iffer-e e onp r ind basis
References
[1] Alonso Y. Lateralization of visual guided behaviour during feeding inzebra finches (Taeniopygia guttata). Behav Process 1998;43:257–63.
[2] Andrew RJ. The nature of behavioural lateralization in the chick. In:Andrew RJ, editor. Neural and behavioural plasticity: the use of thechick as a model. Oxford: Oxford University Press; 1991. p. 536–54.
[3] Bingman VP, Hough II GE, Kahn MC, Siegel JJ. The homing pigeonhippocampus and space: in search of adaptive specialization. BrainBehav Evol 2003;62:117–27.
[4] Clayton NS, Krebs JR. Memory for spatial and object-specific cuesin food storing and non-storing birds. J Compar Physiol A: SensoryNeural Behav Physiol 1994;174:371–9.
[5] Cozzutti C, Vallortigara G. Hemispheric memories for the contentand position of food caches in the domestic chick. Behav Neurosci2001;115:305–13.
[6] Deng C, Rogers LJ. Organisation of the tectorotundal and SP/IPS-rotundal projections in the chick. J Compar Neurol 1998;394:171–85.
[7] Deng C, Rogers LJ. Bilaterally projecting neurons in the two visualpathways of chicks. Brain Res 1998;794:281–90.
[8] Fellows BJ. Chance stimulus sequences for discrimination tasks. Psy-chol Bull 1967;67:87–92.
[9] Gagliardo A, Ioale P, Odetti F, Bingham UP, Vallortigara G. Hom-ing pigeons: differential role of left and right hippocampus for-mation in acquisition of the navigational maps. Eur J Neurosci2001;13:1617–24.
[10] Gagliardo A, Odetti F, Ioale P, Bingman VP, Tuttle S, VallortigaraG. Bilateral participation of the hippocampus in familiar landmarknavigation by homing pigeons. Behav Brain Res 2002;136:201–9.
[11] Gunturkun O, Kesh S. Visual lateralization during feeding in pigeons.
rt
rn-No.eu-
ndSci
the
-and
to-gia
tor.del.
lop-Mol
ken.
s inand
am-
arkin.
ro-rma-
nimals showing sometimes dominance of one hemispor a particular cue and sometimes participation of bemispheres for another cue. For instance, in working mry tasks pigeons showed left-hemispheric dominancbject-specific cues and participation of both hemisph
or position-specific cues[16]. In contrast, chicks showeight-hemispheric dominance for position-specific cueslargely) bilateral participation of both hemispheres forect specific cues (condition “contrasting position and obues”). In a similar condition, parids showed complete clementarity of function, with the right hemisphere attend
o position cues and the left hemisphere attending to oues[4]. In the chick and pigeon, there is evidence (revn [18–20]) that in the natural condition the embryo is onted in the egg so that the right eye is exposed to light
he left eye is occluded) and this puts the left hemispheharge of certain visually-guided patterns of behaviour[21].he chicks used in the present experiments came from aercial hatchery, in which no light exposure to the eggsrovided (though, obviously, there were not the strict c
rol conditions of a laboratory). Note, also, that chicksprecocial species, whereas parids and pigeons are a
pecies, and this may also contribute to behavioural dnces. Nonetheless, the fact that the basic pattern ofpheric specialisation is the same in the various speciesn no case a dominance of the right hemisphere for obpecific cues has been observed) makes it likely that dnt species may possess intrinsic tendencies to makarticular hemisphere in charge of control of behaviouifferent settings, though each hemisphere retains itspecialization.
e
-
t
-
l
i-.,
e
c
Behav Neurosci 1987;101:433–5.[12] Gunturkun O. Avian visual lateralization. A review. NeuroRepo
1997;8:3–4.[13] Kahn MC, Bingman VP. Lateralization of landmark spatial lea
ing strategies in the hippocampus of homing pigeons. Program585.16. Abstract viewer/Itinerary planner. Washington, DC: Soc Nrosci, 2002 [CD-ROM].
[14] Matsushima T, Izawa EI, Aoki N, Yanagihara S. The mithrough chick eyes: memory, cognition and anticipation. Zool2003;20:395–408.
[15] Mench J, Andrew RJ. Lateralization of a food search task indomestic chick. Behav Neural Biol 1986;46:107–14.
[16] Prior H, Gunturkun O. Parallel working memory for spatial location and food-related object cues in foraging pigeons: binocularlateralized monocular performance. Learn Mem 2001;8:44–51.
[17] Rashid NY, Andrew RJ. Right hemisphere advantage forpographical orientation in the domestic chick. Neuropsycholo1989;27:937–48.
[18] Rogers LJ. Development of lateralization. In: Andrew RJ, ediNeural and behavioural plasticity: the use of the chick as a moOxford: Oxford University Press; 1991. p. 507–35.
[19] Rogers LJ. The molecular neurobiology of early learning, devement, and sensitive periods, with emphasis on the avian brain.Neurobiol 1993;7:161–87.
[20] Rogers LJ. The development of brain and behaviour in the chicWallingford, UK: CAB International; 1995.
[21] Rogers LJ. Behavioral, structural and neurochemical asymmetriethe avian brain: a model system for studying visual developmentprocessing. Neurosci Biobeh Rev 1996;20:487–503.
[22] Rogers LJ, Andrew RJ. Comparative vertebrate lateralization. Cbridge, UK: Cambridge University Press; 2002.
[23] Tommasi L, Vallortigara G. Encoding of geometric and landminformation in the left and right hemispheres of the avian braBehav Neurosci 2001;115:602–13.
[24] Tommasi L, Gagliardo A, Andrew RJ, Vallortigara G. Separate pcessing mechanisms for encoding geometric and landmark infotion in the avian brain. Eur J Neurosci 2003;17:1695–702.

L. Regolin et al. / Behavioural Brain Research 157 (2005) 1–9 9
[25] Tommasi L, Andrew RJ, Vallortigara G. Eye use in search is deter-mined by the nature of the task in the domestic chick. Behav BrainRes 2000;112:119–26.
[26] Ulrich C, Prior H, Duka T, Leschchins’ka I, Valenti P, GunturkunO, et al. Left hemispheric superiority for visuospatial orientation inhoming pigeons. Behav Brain Res 1999;104:169–78.
[27] Valenti A, Sovrano VA, Vallortigara G. Visual lateralization in quails.Laterality 2003;8:67–78.
[28] Vallortigara G. Comparative neuropsychology of the dual brain: astroll through left and right animals’ perceptual worlds. Brain Lang2000;73:189–219.
[29] Vallortigara G. Visual cognition and representation in birds and pri-mates. In: Rogers, L.J., Kaplan, G., editors. Vertebrate comparativecognition: are primates superior to non-primates? Kluwer Academic/Plenum Publishers; 2004. p. 57–94.
[30] Vallortigara G, Zanforlin M. Position learning in chicks. Behav Pro-cess 1986;12:23–32.
[31] Vallortigara G, Zanforlin M. Place and object learning in chicks(Gallus gallus domesticus). J Compar Psychol 1989;103:201–9.
[32] Vallortigara G, Andrew RJ. Lateralization of response by chicks tochange in a model partner. Anim Behav 1991;41:187–94.
[33] Vallortigara G, Regolin L. Facing an obstacle: lateralization of ob-ject and spatial cognition. In: Andrew RJ, Rogers LJ, editors. Com-parative vertebrate lateralization. Cambridge: Cambridge UniversityPress; 2002. p. 383–444.
[34] Vallortigara G, Rogers LJ, Bisazza A. Possible evolutionary originsof cognitive brain lateralization. Brain Res Rev 1999;30:164–75.
[35] Vallortigara G, Pagni P, Sovrano VA. Separate geometric and non-geometric modules for spatial reorientation: evidence from a lopsidedanimal brain. J Cogn Neurosci 2004;16:390–400.
[36] Vallortigara G, Regolin L, Rigoni M, Zanforlin M. Delayed searchfor a concealed imprinted object in the domestic chick. Anim Cogn1998;1:17–24.
[37] Zanforlin M, Vallortigara G. Form preferences and stimulus gener-alization in domestic chicks. Boll Zool 1985;52:231–8.




















