
Wolverine in Northern England at About 83,000 yr B.P.: Fauna1 Evidence for Climatic Change during...
14
QUATERNARY RESEARCH 24, 73-86 (1985) Wolverine in Northern England at About 83,000 yr B.P.: Fauna1 Evidence for Climatic Change during Isotope Stage 5 ANTONY J. SUTCLIFFE Department of Palaeontology, British Museum (Natural History). London SW7 5BD, United Kingdom THOMAS C. LORD Pigyard Museum, Settle, North Yorkshire, United Kingdom RUSSELL S. HARMON Department of Geological Sciences, Southern Methodist University+ Dallas, Texas 7.5275 MIRO IVANOVICH Uranium Series Disequilibrium Section, Nuclear Physics Division, AERE, Hat-well OX11 ORA. United Kingdom ANGELARAE School of Geography, Oxford University, Oxford OX1 3PR and Uranium Series Disequilibrium Section, Nuclear Physics Division% AERE, Harwell OXil ORA, United Kingdom AND JOHN W. HESS Water Resources Center, Desert Research Institute* Las Vegas, Nevada 89109 Received August 17, 1984 Cave sediments from Stump Cross Cave in northern England contain Pleistocene mammal re- mains. Uranium-series dating of calcium carbonate deposits closely associated with the fossilif- erous horizons has established an absolute age of 83,000 k 6000 yr B.P. for a fauna1 assemblage largely comprised of wolverines (Gulo gulo). This date lies firmly within the younger portion of oxygen-isotope stage 5. The occurrence of wolverines in the vicinity of Stump Cross Cave at ca. 83,000 yr B.P indicates a significant climatic deterioration from ca. 120,000 yr B.P., when an Ipswichian interglacial fauna with hippopotamus was present in this part of northern En- gland. 0 1985 University of Washington. INTRODUCTION The Quaternary has been characterized by episodic fluctuations in global climate. The physical processes accompanying these climatic changes have produced a wide variety of geological and geomorpho- logical features which various workers have studied in detail and used to reconstruct a paleoclimatic history of the period. How- ever, the Quaternary is sufficiently long that no single radiometric method has a dating range which covers the entire pe- riod. Uranium-series disequilibrium tech- niques provide a useful bridge between the 14C and K-Ar dating methods. In partic- ular, the 230Th/234U method has been ap- plied with particular success to speleo- thems, the calcium carbonate deposits formed in limestone caves. The dating of such pure calcite deposits, which often can be related stratigraphically to fossil verte- brates, provides a powerful tool for the ab- solute dating of specific events, the climatic 13 0033-5894/85 $3.00
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Wolverine in Northern England at About 83,000 yr B.P.: Fauna1 Evidence for Climatic Change during...

QUATERNARY RESEARCH 24, 73-86 (1985)
Wolverine in Northern England at About 83,000 yr B.P.: Fauna1 Evidence for Climatic Change during Isotope Stage 5
ANTONY J. SUTCLIFFE Department of Palaeontology, British Museum (Natural History). London SW7 5BD, United Kingdom
THOMAS C. LORD Pigyard Museum, Settle, North Yorkshire, United Kingdom
RUSSELL S. HARMON Department of Geological Sciences, Southern Methodist University+ Dallas, Texas 7.5275
MIRO IVANOVICH Uranium Series Disequilibrium Section, Nuclear Physics Division, AERE, Hat-well OX11 ORA.
United Kingdom
ANGELA RAE School of Geography, Oxford University, Oxford OX1 3PR and Uranium Series Disequilibrium Section,
Nuclear Physics Division% AERE, Harwell OXil ORA, United Kingdom
AND
JOHN W. HESS
Water Resources Center, Desert Research Institute* Las Vegas, Nevada 89109
Received August 17, 1984
Cave sediments from Stump Cross Cave in northern England contain Pleistocene mammal re- mains. Uranium-series dating of calcium carbonate deposits closely associated with the fossilif- erous horizons has established an absolute age of 83,000 k 6000 yr B.P. for a fauna1 assemblage largely comprised of wolverines (Gulo gulo). This date lies firmly within the younger portion of oxygen-isotope stage 5. The occurrence of wolverines in the vicinity of Stump Cross Cave at ca. 83,000 yr B.P indicates a significant climatic deterioration from ca. 120,000 yr B.P., when an Ipswichian interglacial fauna with hippopotamus was present in this part of northern En- gland. 0 1985 University of Washington.
INTRODUCTION
The Quaternary has been characterized by episodic fluctuations in global climate. The physical processes accompanying these climatic changes have produced a wide variety of geological and geomorpho- logical features which various workers have studied in detail and used to reconstruct a paleoclimatic history of the period. How- ever, the Quaternary is sufficiently long that no single radiometric method has a
dating range which covers the entire pe- riod. Uranium-series disequilibrium tech- niques provide a useful bridge between the 14C and K-Ar dating methods. In partic- ular, the 230Th/234U method has been ap- plied with particular success to speleo- thems, the calcium carbonate deposits formed in limestone caves. The dating of such pure calcite deposits, which often can be related stratigraphically to fossil verte- brates, provides a powerful tool for the ab- solute dating of specific events, the climatic
13 0033-5894/85 $3.00

74 SUTCLIFFE ET AL.
nature of which can be inferred from a knowledge of the habitat of a particular fauna1 association.
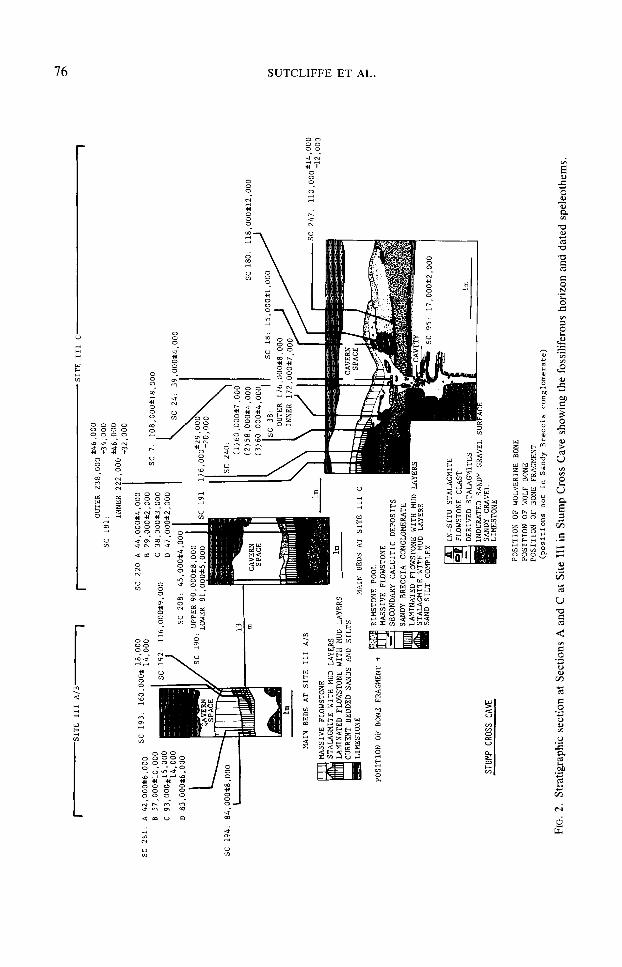
Stump Cross Cave, in north Yorkshire, England, has yielded a relatively large number of Pleistocene mammalian fossils in intimate association with dateable calcite speleothems. Four fossiliferous sites were found in the course of digging designed to improve access in parts of the cave open to tourists and to extend the wild cave. Three sites are situated at the same passage level in the cave system (Fig. 1). In 1955 incom- plete skeletons of four reindeer, Rurzgifer tmundus, were found at Site I (Collins, 1959). Parts of two wolverines were found at Site II in 1964 and parts of six wolverines and a variety of other mammals were re- covered during controlled excavations at Site III in 1980- 1981. The wolverine fossils from Sites II and III comprise the largest collection of remains of this animal yet
FIG. 1. Map showing location of fossiliferous Sites I, II, and III in Stump Cross Cave, north Yorkshire, En- gland.
found at any British site. An associated skeleton of another wolverine was found in a deeper part of the cave system (Site IV) in 1981. Uranium-series dating of speleo- them deposits with clear stratigraphic re- lationships to the fossils has provided an absolute age for the fauna1 remains at Sites II and III and, by inference, information on the climatic conditions prevailing on the surface at the time.
LOCATION AND CAVE GEOMORPHOLOGY
Stump Cross Cave is a largely relict cave system, comprising about 5.5 km of ex- plored passages, which lies within the car- boniferous limestone outcrop of the Greenhow Anticline, 10 km east of Gras- sington, north Yorkshire (Dunham and Stubblefield, 1945; Waltham, 1975). The ar- tificial entrance, at an altitude of 360 m, penetrates about 1 m of till and l-2 m of limestone fault breccia before intersecting a narrow solution fissure which descends to about 16 m below the surface to the tourist-cave level (Fig. 1).
The limestone beds, which dip about 20°N, are locally highly fractured as the re- sult of tectonic activity. The cave system was initiated under phreatic conditions, with groundwater drainage trending against the direction of dip. Abandoned phreatic passages display pronounced horizontal de- velopment at the show-cave level and also at deeper levels down dip, implying that phreatic flow was successively lowered in response to falls in spring points resulting from surface erosion. Under current hy- drological conditions (artificially lowered by a nearby mine drain), there is an active cave stream about 30 m below the show- cave level, which may rise some 12 m at the highest flood stage following periods of heavy rain. Unsorted surface debris be- neath a joint chimney and a dip fissure at show-cave level demonstrates that subver- tical conduits were formerly open to the surface. The cave system had apparently been sealed by roof breakdown and fill until

16-AGE WOLVERINE 75
lead miners broke into it during the 19th century.
STRATIGRAPHY OF THE CAVE DEPOSITS
The stratigraphy of a 35m-long cutting at Site III was recorded in detail during ex- cavations 1980- 1981 (Fig. 2). Zn situ bone horizons were encountered at two separate places, represented here by Sections IIIA/ B and IIIC.
At Section IIIA/B a limestone boulder breccia is overlain by current-bedded sands, which are sealed beneath a lami- nated flowstone. This comprises a rhythmic deposit of alternating calcite and mud layers, most less than 3 mm thick, with small interbedded lenses of coarser mate- rial, including sand and broken straw sta- lactites. Bone fragments were recorded in situ from lenses at this horizon. The lami- nated flowstone is overlain by a massive cap flowstone up to 0.6 m thick. Unstrati- fied wolverine bones, a wolverine tooth, and a reindeer bone had previously been collected from the laminated flowstone in the vicinity of this section during the con- struction of the cutting.
Section IIIC is 13 m distant and at a lower elevation. It is situated at the lowest part of about 80 m of passage with a gentle U-shaped profile. The earliest deposit within the cave. resting directly on the eroded limestone floor, is a sandy water- laid gravel containing allochthonous peb- bles and a few limestone boulders. It is cut by a shallow channel, the floor of which is indurated by calcite. The indurated channel floor and the discrete stalagmites upon it are overlain by a laterally variable complex of sand and silt. Above the sand and silt is a laminated flowstone, similar to that at Section IIIA/B, but with lenses containing coarser elastic material, including frag- ments of broken calcite flowstone and de- tached stalagmites. Bones occur near the base of this deposit, which merges laterally into a fossiliferous variably cemented breccia/conglomerate. At one point this
overlies an Zn situ stalagmite standing on a limestone boulder; at another it extends un- conformably down through earlier deposits into a solution-enlarged joint in the lime- stone floor of the cave. A massive flow- stone with a rimstone pool facies caps the sequence and forms the present-day floor.
The current-bedded sands at Section IIIA/B and the sandy gravel at Section IIIC are interpreted as alluvial deposits. The cave must have drained sufficiently by the time of deposition of the overlying stalag- mites and flowstone to permit the precipi- tation of secondary calcium carbonate from vadose groundwater seepage at this level. Mud layers within the laminated flowstone deposits suggest that subsequently the pas- sage was repeatedly flooded by muddy water, which then drained away to permit the further precipitation of calcium car- bonate. We believe that the sand and silt complex within the shallow channel at Sec- tion IIIC was deposited during an episode of periodic stream flow. The sandy breccia conglomerate is interpreted as the infilling of a “pipe” that developed over a joint in the cave floor at the lowest part of the pas- sage and received water-transported mate- rials from both directions. It was subse- quently cemented by calcitic deposits.
Site II is no longer accessible, but we have examined the hitherto unpublished finds. These include a block of laminated flowstone, approximately 4 cm thick. which contains at its base an Zn site bone horizon with flowstone clasts.
RADIOMETRIC DATING
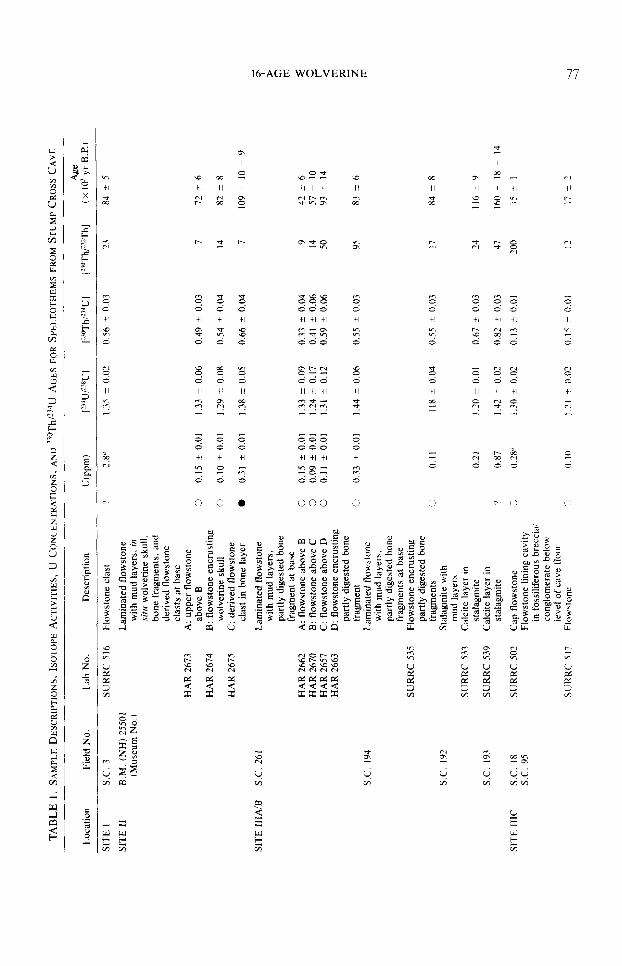
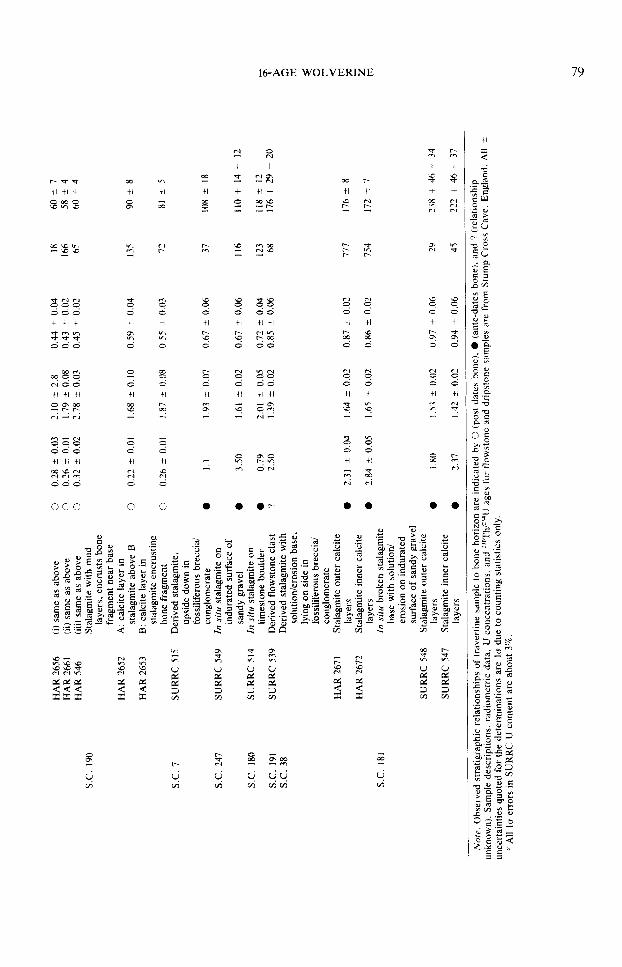
Uranium-series radiometric data ob- tained from the analysis of calcite deposits at Sites I, II, and III are presented in Table 1. Details of the isotope dilution and alpha spectrometry methods used have been de- scribed elsewhere (Harmon et ~1.. 1978: Ivanovich and Harmon, 1982). The equa- tions used to calculate the 23@Th/‘34U ages, together with the criteria for establishing the dateability of cave deposits by the U- series disequilibrium methods are dis-
I-SIT
E III
A/
B-]
I SI
TE
III
C
I
OU
TER
23
8,00
0 f;;
';;
; SC
18
1: IN
NER
2~
2,00
0 i4
6,O
OO
-3
2,00
0
SC 2
61:
A
42,0
00~6
,000
SC
19
3.
160,
OO
Oe
;;f;;
. '
SC
220
A
44,0
00*4
,000
B
29
,000
*2,0
00
I SC
7.
loa,
ooo*
lfJ,o
oo
I
SC
B 5
7,00
0~10
,000
D
83,O
OO
i6,0
00
0,.
>nn
UP
PE
R 9
O.O
OO
iB.0
00
>k.
L7"'
LOW
ER 8
1,00
0*5~
000
c 19
1.
17
3V,O
OO
i4,0
00
SC 2
40:
~1~6
0,00
0~7.
000
MAI
N
BE
DS
AT
SITE
III
A
/B
~~M
AS
SIV
E
FLO
WST
ON
E ST
ALAG
MIT
E W
lT"
M"D
LA
YF,R
S LA
MIN
ATED
FL
OW
STO
NE
WIT
H M
U
CU
RR
ENT
BED
DED
SA
ND
S A
ND
SIl
LIM
ESTO
NE
OU
TER
17
6,O
OO
i8,0
IN
NER
17
2 00
0*7,
0
lm
h L*
YER
s -
(2)5
8,00
0*4,
000
SC 1
80:
118,
000i
12,0
00
~3~6
0~00
0*4,
000
, SC
18:
15
,000
~1,0
00
.
.TS
MAI
N
BE
DS
AT
SITE
III
C
PI~>
LVP.
r,a
w~d
NE
SEC
ON
DAR
Y C
ALC
ITIC
D
EPO
SITS
S
AN
DY
BR
ECC
IA
CO
NG
LOM
ERAT
E LA
MIN
ATED
FL
OW
STO
NE
WIT
H M
"D
MY
ST
ALAG
MIT
E W
ITH
MU
D L
AY
ER
S
SAN
D S
ILT
CO
"PLE
X
IN-S
ITU
ST
ALAG
MIT
E FL
OW
STO
NE
CLA
ST
DER
IVED
ST
ALAG
NIT
ES
IND
UR
ATED
S
AN
DY
GR
AVEL
SIJ
S
AN
DY
GR
AVEL
LI
MES
TON
E ST
UNP
CRO
SS C
AVE
SC 9
5:
17,0
00*2
,000
247.
11
0.00
0~14
.000
-1
2,00
0
POSI
TIO
N
OF
WO
LVER
INE
BON
E PO
SITI
ON
O
F W
OLF
BO
NE
POSI
TIO
N
OF
BON
E FR
AGK?
.NT
(pos
ition
s no
t in
Sa
ndy
Brec
cia
cong
lom
erat
e)
FIG
. 2.
Stra
tigra
phic
sect
ion
at S
ectio
ns
A an
d C
at S
ite I
II in
Stum
p Cr
oss
Cave
sho
wing
th
e fo
ssilif
erou
s ho
rizon
an
d da
ted
spel
eoth
ems.
TABL
E 1.
SAM
PLE
DESC
WPT
IONS
. IS
OTOP
E AC
TWKI
ES,
U CO
NCEN
TRAT
IONS
, AN
D z3
0Th/z
s4U
AGES
FO
R SP
ELEO
THEM
S FK
OM
STUM
P CR
OSS
CAVE
Loca
tion
Field
No
. La
b No
. De
scrip
tion
LJtpp
rW
,34”,%
3~,
[=“fh/
zJ*U
] ,3”
-,-,,,X
Th,
fk!e
c x 1
Oj y
r B.
P.)
SITE
I
S.C.
3
SURR
C 51
6
SITE
II
B.M.
(N
H)
2550
1 (M
useu
m No
.)
HAR
2673
HAR
2674
HAR
2675
SITE
III
A/B
S.C.
26
1
HAR
2662
HA
R 26
70
HAR
2657
HA
R 26
63
S.C.
lY
4
SURR
C 53
s
S.C.
lY
2
SURR
C 53
3
S.C.
lY
3 SU
RRC
539
SITE
IIIC
S.
C.
18
SURR
C SO
2 S.
C.
Y5
SLiR
RC
5!7
Flows
tone
clast
Lami
nate
d flo
wston
e wi
th
mud
layers
, in
.sh
wo
lverin
e sk
ull,
bone
fra
gmen
ts.
and
deriv
ed
flows
tone
clasts
at
base
A:
up
per
flows
tone
abov
e B
B: f
lowsto
ne
encru
sting
wo
lverin
e sk
ull
C: d
erive
d flo
wston
e cla
st in
bone
lay
er
Lami
nate
d flo
wst
one
with
mu
d lay
ers,
partly
dig
ested
bo
ne
fragm
ent
at ba
se
A: f
lowsto
ne
abov
e B
B: f
low
ston
e ab
ove
C C:
flow
stone
ab
ove
D D:
flo
wston
e en
crusti
ng
partly
dig
ested
bo
ne
fragm
ent
Lami
nate
d flo
wston
e wi
th
mud
layers
, pa
rtly
diges
ted
bone
fra
gmen
ts at
base
Flo
wston
e en
crusti
ng
partly
dig
ested
bo
ne
fragm
ents
Stala
gmite
wi
th
mud
layers
Ca
lcite
layer
in
stalag
mite
Ca
lcite
layer
in
s&lag
mite
Cap
flows
tone
Flows
tone
lining
ca
vity
in fos
silife
rous
br
eccia
co
nglom
erate
be
low
level
of ca
ve
floor
Flo
w<ton
c
2.8”
1.35
2 0.0
2 0.5
6 2
0.03
23
84 I
? 5
0.15
2 0.0
1 I.3
3 k
0.06
0.49
2 0.0
3
0.10
2 0.0
1 1.2
9 +I
0.08
0.54
2 0.0
4
0.31
* 0.0
1 I.3
8 k
0.05
0.66
2 0.0
4
7 12
k
6
14
82 2
8
7 10
9 +
10 -
9
0.15
? 0.0
1 0.0
9 5
0.01
0.11
* 0.0
1
0.33
5 0.0
1
1.33
2 0.0
9 0.3
3 2
0.04
1.24
2 0.1
7 0.4
1 2
0.06
1.31
5 0.1
2 0.5
9 2
0.06
1.44
k 0.0
6 0.5
5 It
0.03
9 42
?
6 14
57
2
10
SO
Y3 5
14
95
83 2
6
0.11
11x
z 0.0
4 o.s
5 2
0.03
17
84 2
8
0.21
I.20
2 0.0
1 0.6
7 k
0.03
24
II6
z 9
0.87
1.42
k 0.0
2 0.8
2 2
0.03
47
I60
t I8
14
0.28
‘ I.3
0 z
0.02
0.13
T 0.0
1 x0
15
k
I
‘-> n.
10
I.21
+ O.
02
0.15
- n.n
1
78 SUTCLIFFE ET AL.
HAR
2656
HA
R 26
61
HAR
546
SC.
190
HAR
2652
HAR
2653
SURR
C 51
s S.
C.
7
KC.
241
SURR
C 54
9
S.C.
18
0 SU
RRC
514
S.C.
19
1 SU
RRC
539
S.C.
38
HAR
2671
HAR
2612
S.C.
18
1
SURR
C 54
8
SURR
C 54
7
ci) s
ame
as a
bove
(ii)
sa
me
as a
bove
(iii
) sa
me
as a
bove
St
alagm
ite
with
mu
d lay
ers,
encru
sts
bone
fra
gmen
t ne
ar
base
A:
calc
ite
layer
in
stalag
mite
ab
ove
B B:
calc
ite
layer
in
stalag
mite
en
crusti
ng
bone
fra
gmen
t De
rived
sta
lagmi
te,
upsid
e do
wn
in fos
silife
rous
br
eccid
co
nglom
erate
In
zitu
sta
lagmi
te
on
indur
ated
surfa
ce
of sa
ndy
grav
el [P
I sift
sta
lagmi
te
on
limes
tone
bo
ulder
De
rived
flo
wston
e cla
st De
rived
sta
lagmi
te
with
so
lution
/eros
ion
base
. lyi
ng
on s
ide
in fos
silife
rous
br
eccia
/ co
nglom
erate
St
alagm
ite
outer
ca
lcite
layers
St
alagm
ite
inner
ca
lcite
layers
Zn
&IJ
brok
en
stzala
gmite
ba
se
with
so
lution
/ er
osion
on
ind
urate
d su
rface
of
sand
y gr
avel
Stala
gmite
ou
ter
calci
te lay
ers
Stala
gmite
inn
er
calci
te lay
ers
0 0.2
8 e
0.03
2.10
2 2.8
0.4
4 k
0.04
18
60 k
7
!2 0.2
6 ?
n.01
I.79
* 0.0
x 0.4
3 t
0.02
I66
58 k
4
0 0.3
2 k
0.02
2.78
z 0.0
3 0.4
5 5
0.02
65
60
+ 4
0.22
k 0.0
1
0.26
2 0.0
1
1.68
? 0.1
0
1.87
2 0.0
8
0.59
5 0.0
4
0.55
?z 0
.03
135
90 2
8
72
81 I
? 5
1.1
37
108
5 18
3.50
0.79
2.50
1.93
2 0.0
7 0.6
7 k
0.06
1.61
k 0.0
2 0.6
7 2
0.06
2.01
k 0.0
5 0.7
2 2
0.04
1.39
z 0.0
2 0.8
5 k
0.06
I16
110
+ 14
-
12
123
I18
k 12
68
17
6 +
29 -
20
2.31
? 0.0
4 1.6
4 e
0.02
0.87
k 0.0
2 77
7 17
6 2
x
2.84
5 0.0
5 1.6
5 2
0.02
0.86
z 0.0
2 7s
4 17
2 2
7
130
2.37
1.53
2 0.0
2
1.42
?z 0
.02
0.97
2 n.
lM
29
238
+ 46
-
34
0.94
2 0.0
6 45
22
2 +
46
- 37
Nore
. ob
serve
d str
atigr
aphic
re
lation
ships
of
Wav
ertin
e sa
mple
to bo
ne
horiz
on
are
indica
ted
by
9 (po
st-da
tes
bone
). 0
(ante
-date
s bo
ne),
and
? (re
lation
ship
unkn
own)
. Sa
mple
desc
riptio
ns.
radio
metric
da
ta,
IJ co
ncen
tratio
m.
and
~l”Th
/z’W
ag
e, fol
- flo
wston
c an
d dr
ipston
e sa
mples
ar
e fro
m St
ump
Cros
s Ca
ve.
Engla
nd.
All
2 un
certa
inties
qu
oted
for
the
de
termi
natio
ns
are
lo du
e to
coun
ting
statis
tics
only.
” A
ll lu
errors
in
SURR
C U
conte
nt ar
e ab
out
3%.

80 SUTCLIFFE ET AL.
cussed in Ivanovich and Harmon (1982) and references therein. Although there is a large variation in speleothem uranium content throughout the cave (0.09 to 3.50 ppm), the ages given in Table 1 can be accepted with confidence because variations in uranium content within individual units are small, 23’?h/232Th activity ratios are high (~20 in all but four cases), and the ages for samples from individual travertine units or subsam- ples from single stalagmites decrease up- ward in a systematic manner (within the analytical uncertainty quoted) in correct lithostratigraphic succession.
The main process governing the deposi- tion of CaC03 speleothems is the degassing of CO2 from vadose groundwaters. Such waters will have equilibrated with dissolved high levels of CO2 during their passage through the soil zone or while in contact with decaying organic matter prior to their entrance into carbonate bedrock above a cave. Subsequent dissolution of carbonate rock by vadose percolation will cause CaC03 to be deposited when the waters enter a cave in which the partial pressure of CO2 in the water is greater than that in the cave atmosphere. During periods of cold climate, groundwater percolation may be interrupted by glacier-ice cover or by periglacial conditions at the surface above the cave, or it may be greatly retarded by lack of surface vegetation and biogenic CO2 production, a feature characteristic of al- pine areas at present (Harmon et ul., 1977).
Three important points are evident from Table 1. First, by ca. 230,000 yr BP the passages at the tourist-cave level were drained sufficiently to permit speleothem deposition on the surface of the basal alluvial deposits at Site IIIC. Second, the dates, when considered collectively or in- dividually, appear to fall into four major groups: 238,000 to 160,000 yr BP, 118,000 to 72,000 yr B.P., 60,000 to 29,000 yr, B.P., and <17,000 yr B-P. Similar grouping of speleotherm ages, at times of interglacial or interstadial climate, has also been observed at other cave sites (Atkinson et al., 1978;
Gascoyne et al., 1981) and appears to be an intrinsic feature of the speleothem paleo- climate record in Britain. Finally, the U- series ages establish upper and lower time limits for the bone layer at Sites II and IIIC and an upper limit for Site IIIA/B. Only one fossiliferous episode is apparent from the radiometric evidence; it occurred within the interval 108,000 & 18,000 to 81,000 ? 5000 yr B.P. Significantly, the dates for Iive speleothem samples stratigraphically below the bone deposits fall within the interval 118,000 ? 12,000 to 108,000 ? 18,000 yr BP., whereas speleothem calcite encrusting bone deposits yields ages of 84,000 ? 8000, 83,000 & 6000, 82,000 & 8000, and 81,000 ? 5000 yr B.P. Thus, the episode of bone deposition was both preceded and followed by episodes of speleothem deposition, which we consider to be times of intergla- cial or interstadial climatic conditions in northern Britain. This interpretation is sup- ported by the correlation of the two groups of U-series ages with the lower and upper substages of stage 5 of the marine oxygen- isotope record (Emiliani, 1955; Shackleton, 1969; Shackleton and Opdyke, 1973).
The age of the reindeer bones from Site I was not determined. An attempt by R. Burleigh (British Museum) to obtain a ra- diocarbon date for fossils from Site IIIA/B was unsuccessful as the bones examined contained insufficient collagen.
THE MAMMALIAN FAUNA
The identifiable skeletal parts of the five species of mammalian fossils recovered from Sites II, IIIA/B, and IIIC are listed in Table 2. The homogeneous character of these fauna is consistent with the lithostra- tigraphic and radiometric evidence indi- cating only one event of bone deposition. We interpret the mammalian remains to represent a single fauna1 assemblage com- prised of the following animals: G& gulo L. (wolverine), Canis lupus L. (wolf), Vulpes vulpes L. or Alopex lagopus L. (red or arctic fox), Rung$er turundus L. (rein-
16-AGE WOLVERINE 81
TABLE 2. PROPORTIONS OFDETERMINABLE SKELETAL PARTS FROM SITES Il,IIlA/B ANII MC (STUMPCROSSCAVE,ENGLAND)
~~- Mandibular Isolated
Skulls rami teeth Vertebrae Scapulae Ribs Humeri Radii ---~.
Site II Gulo gulo L. 1 2 I I I I
Site IIIA/B Gulo g&o L. I 2 3 Canis hipus L. Rangifer
tarandus L. I
Site IIIC Gulo gulo L. 3 6 6 5 1 12 I 5 Canis lufms L. 2 3 3 6 I I
Fox I cf. Bison I
Min. No. of Ulnae Pelves Femora Tibiae Podials Metapodials Phalanges individuals
Site II Guh gulo L. I I I 1 I 3 1 2
Site IIIA/B Gulo guh L. 2 3 2
Canis lupus L. 2 2 Rangifer
tarandus L. I
Site IIIC Gulo g/h L. 2 2 I 6 16 II 6
Canis hpus L. 2 I 2 I 4 I Fox I I
cf. Bison I - -~.-.
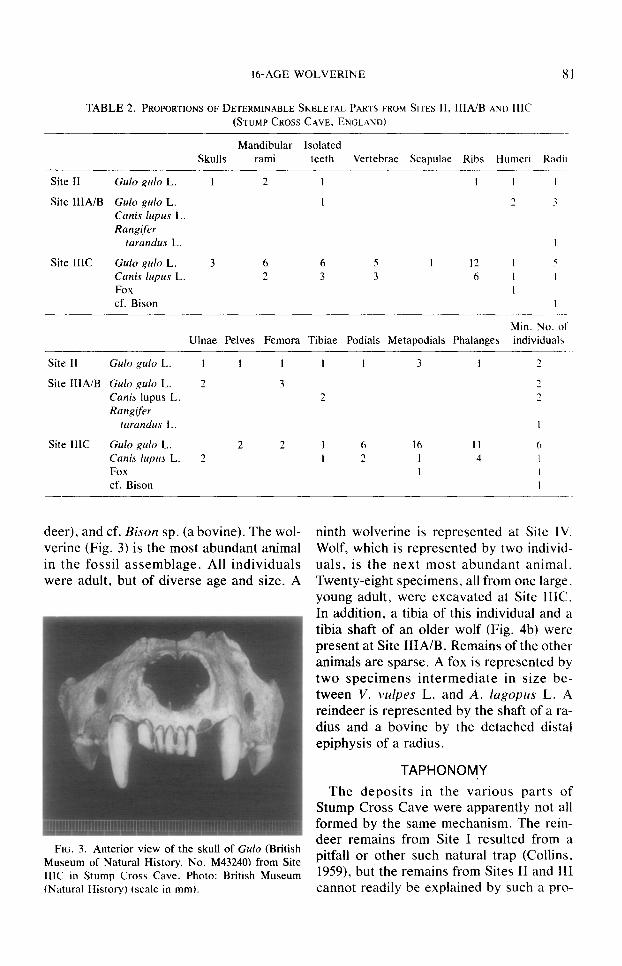
deer), and cf. Bison sp. (a bovine). The wol- verine (Fig. 3) is the most abundant animal in the fossil assemblage. All individuals were adult, but of diverse age and size. A
FIG. 3. Anterior view of the skull of Gulo (British Museum of Natural History, No. M43240) from Site IIIC in Stump Cross Cave. Photo: British Museum (Natural History) CscaIe in mm).
ninth wolverine is represented at Site IV. Wolf, which is represented by two individ- uals, is the next most abundant animal. Twenty-eight specimens, all from one large, young adult, were excavated at Site IIIC. In addition, a tibia of this individual and a tibia shaft of an older wolf (Fig. 4b) were present at Site IIINB. Remains of the other animals are sparse. A fox is represented by two specimens intermediate in size be- tween V. vdpes L. and A. lagopus L. A reindeer is represented by the shaft of a ra- dius and a bovine by the detached distal epiphysis of a radius.
TAPHONOMY
The deposits in the various parts of Stump Cross Cave were apparently not all formed by the same mechanism. The rein- deer remains from Site I resulted from a pitfall or other such natural trap (Collins, 1959), but the remains from Sites II and III cannot readily be explained by such a pro-
82 SUTCLIFFE ET AL.
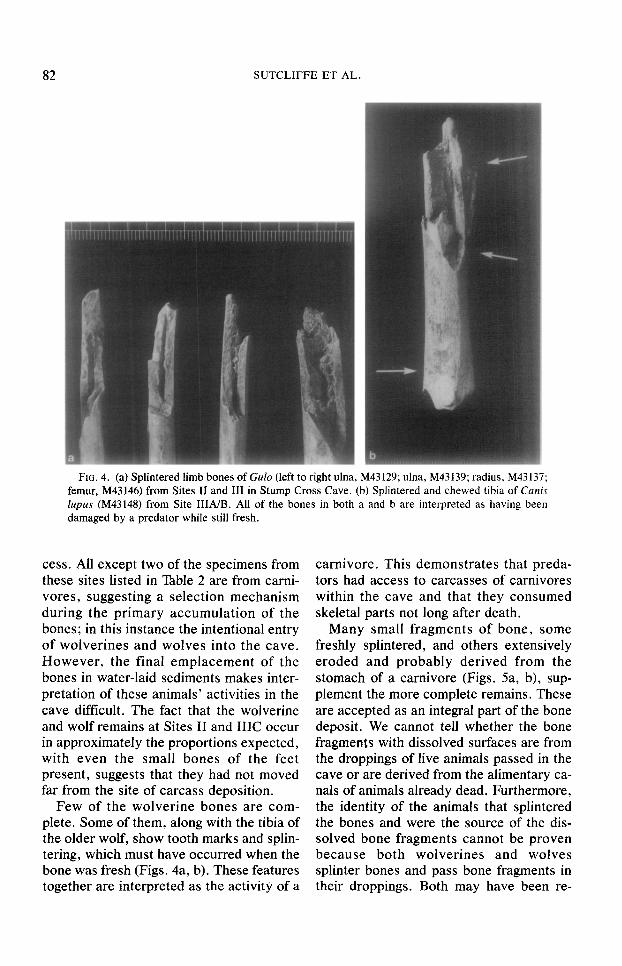
FIG. 4. (a) Splintered limb bones of Gu/o (left to right ulna, M43129; ulna, M43139; radius, M43137; femur, M43146) from Sites II and III in Stump Cross Cave. (b) Splintered and chewed tibia of Cunis lupus (M43148) from Site 1&4/B. All of the bones in both a and b are interpreted as having been damaged by a predator while still fresh.
cess. All except two of the specimens from these sites listed in Table 2 are from carni- vores, suggesting a selection mechanism during the primary accumulation of the bones; in this instance the intentional entry of wolverines and wolves into the cave. However, the final emplacement of the bones in water-laid sediments makes inter- pretation of these animals’ activities in the cave difficult. The fact that the wolverine and wolf remains at Sites II and IIIC occur in approximately the proportions expected, with even the small bones of the feet present, suggests that they had not moved far from the site of carcass deposition.
Few of the wolverine bones are com- plete. Some of them, along with the tibia of the older wolf, show tooth marks and splin- tering, which must have occurred when the bone was fresh (Figs. 4a, b). These features together are interpreted as the activity of a
carnivore. This demonstrates that preda- tors had access to carcasses of carnivores within the cave and that they consumed skeletal parts not long after death.
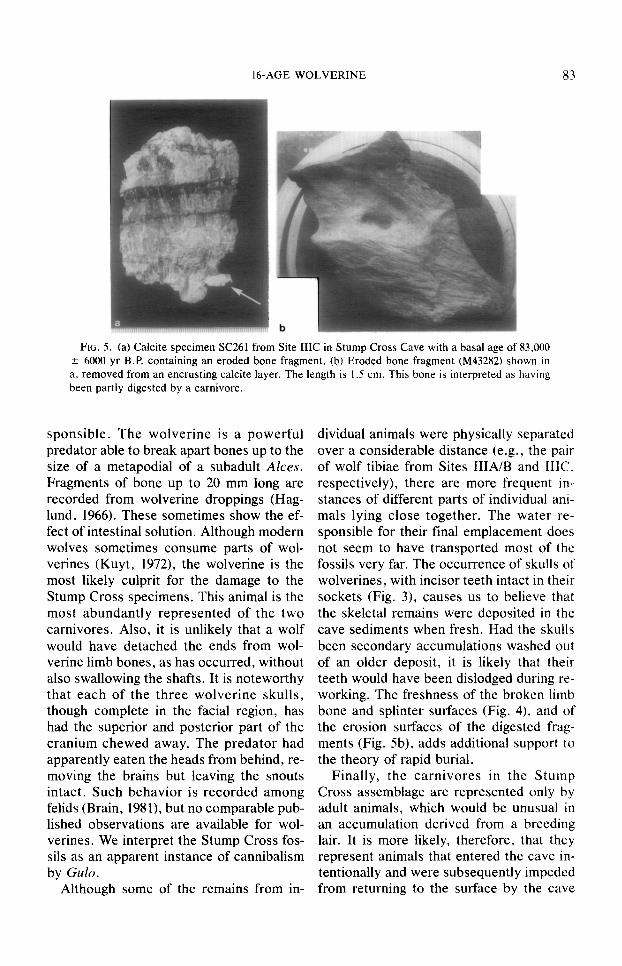
Many small fragments of bone, some freshly splintered, and others extensively eroded and probably derived from the stomach of a carnivore (Figs. 5a, b), sup- plement the more complete remains. These are accepted as an integral part of the bone deposit. We cannot tell whether the bone fragments with dissolved surfaces are from the droppings of live animals passed in the cave or are derived from the alimentary ca- nals of animals already dead. Furthermore, the identity of the animals that splintered the bones and were the source of the dis- solved bone fragments cannot be proven because both wolverines and wolves splinter bones and pass bone fragments in their droppings. Both may have been re-
16-AGE WOLVERINE 83
FIG. 5. (a) Calcite specimen SC261 from Site IIIC in Stump Cross Cave with a basal age of 83,000 k 6000 yr B.P. containing an eroded bone fragment. (b) Eroded bone fragment (M43282) shown in a. removed from an encrusting calcite layer. The length is 1.5 cm. This bone is interpreted as having been partly digested by a carnivore.
sponsible. The wolverine is a powerful predator able to break apart bones up to the size of a metapodial of a subadult Alces. Fragments of bone up to 20 mm long are recorded from wolverine droppings (Hag- lurid, 1966). These sometimes show the ef- fect of intestinal solution. Although modern wolves sometimes consume parts of wol- verines (Kuyt, 1972), the wolverine is the most likely culprit for the damage to the Stump Cross specimens. This animal is the most abundantly represented of the two carnivores. Also, it is unlikely that a wolf would have detached the ends from wol- verine limb bones, as has occurred, without also swallowing the shafts. It is noteworthy that each of the three wolverine skulls, though complete in the facial region, has had the superior and posterior part of the cranium chewed away. The predator had apparently eaten the heads from behind, re- moving the brains but leaving the snouts intact. Such behavior is recorded among felids (Brain, 198 l), but no comparable pub- lished observations are available for wol- verines. We interpret the Stump Cross fos- sils as an apparent instance of cannibalism by Gulo.
Although some of the remains from in-
dividual animals were physically separated over a considerable distance (e.g., the pair of wolf tibiae from Sites IIIA/B and IIIC. respectively), there are more frequent in- stances of different parts of individual ani- mals lying close together. The water re- sponsible for their final emplacement does not seem to have transported most of the fossils very far. The occurrence of skulls of wolverines, with incisor teeth intact in their sockets (Fig. 3), causes us to believe that the skeletal remains were deposited in the cave sediments when fresh. Had the skulls been secondary accumulations washed out of an older deposit, it is likely that their teeth would have been dislodged during re- working. The freshness of the broken limb bone and splinter s&aces (Fig. 4). and of the erosion surfaces of the digested frag- ments (Fig. 5b), adds additional support to the theory of rapid burial.
Finally, the carnivores in the Stump Cross assemblage are represented only by adult animals, which would be unusual in an accumulation derived from a breeding lair. It is more likely, therefore, that they represent animals that entered the cave in- tentionally and were subsequently impeded from returning to the surface by the cave

84 SUTCLIFFE ET AL.
topography. Feeding on animals which had already died would have provided an in- terim means of combatting starvation. That animals followed each other into the cave in relatively rapid succession is supported by the availability of fresh remains for con- sumption.
PALEOECOLOGY
All available evidence suggests that the mammalian remains from Sites II and III in Stump Cross Cave constitute a contempo- raneous assemblage. The behavior and hab- itat of the living wolverine has been de- scribed by several writers, among them Krott (1959), Haglund (1966), and Se- menov-Tian-Shanskii (1982). At present this animal has a circumboreal distribution in Scandinavia, the Soviet Union, and North America. Typically, it is an inhab- itant of the forests and treeless highlands of the taiga but also penetrates onto the tundra. Wolverines occur widely within these regions and are not restricted to any single biotope. With a long coarse coat and dense underfur in winter (even the soles of the feet are furred), it is well adapted to living under conditions of snow and ex- treme cold. The wolverine feeds mainly by scavenging, but during the winter, when the snow conditions are unfavorable for its prey, it is also a capable hunter able to kill Rung$er and even subadult Alces. The present distribution of the wolverine closely coincides with that of Rangifer tar- andus, which comprises the major part of its diet. The wolverine also coexists with the wolf. In Alaska, for example, these two species occur together, even in the same valley. They nevertheless occupy slightly different ecological niches, the lighter wol- verine being able to frequent areas of deeper snow in winter and boggier places in summer. The wolverine is usually a lone walker with no regular shelter, although it digs dens for breeding, commonly in hard snow drifts connected to heaps of rocks. It has the habit of storing caches of food in
snow and among rocks, sometimes carried from as far alield as 10 km.
Too close an analogy should not be drawn to any modern biome and those which may have existed in the past because there is some evidence to suggest that biomes ex- isted during the Pleistocene which were quite unlike any at present (Hopkins er uf., 1982). However, the presence of wolverine in the mammalian fossil assemblage in Stump Cross Cave is sufficiently diagnostic in the upper Pleistocene in Britain to indi- cate conditions of cold climate.
DISCUSSION AND CONCLUSIONS
The unweathered condition of the Stump Cross mammalian remains, with some in- stances of intact incisor tooth rows, indi- cates that the interval between their final deposition and subsequent burial by calcite precipitation was not great. The radio- metric age of a flowstone encrusting the wolverine skull at Site II is 82,000 2 8000 yr B.P., two ages of a flowstone encrusting bone fragments at Site IIIA/B are 83,000 ? 6000 and 84,000 ? 8000 yr B.P., and the age for a calcite layer encrusting a bone fragment at IIIC is 81,000 ? 5000 yr B.P. We believe that this tight grouping of U- series dates from the three fossil sites places the age of the fauna shortly before the mean age of 83,000 & 6000 yr BP. at the 95% confidence level.
The radiometrically dated remains of wolverine from Stump Cross Cave, York- shire, are important because they supple- ment the sparse stratigraphic record of this animal in Britain during the Pleistocene. Wolverine was present in the British Isles on at least five different occasions: in the Cromerian (sense &o) Forest Bed de- posits of Mundesley (Newton, 1880), in the cave deposits at Westbury-sub-Mendip, Somerset, which are probably slightly youn- ger (Bishop, 1982), in the pre-Ipswich- ian (>120,000 yr B.P.) Glutton Stratum of Tornewton Cave, Devon (Sutcliffe and Zeuner, 1962), in Stump Cross Cave shortly

16-AGE WOLVERINE 85
before ca. 83.000 vr B.P. (this studv), and represented by the single fragmentary specimen from d < ,
in the upper Devensian at Bacon Hole, Site III, J. Brugal for examining the bovid specimen.
south Wales (Currant et u/., 1984), and in A. P. Currant for valuable discussion, R. Burleigh for
Wetton Mill rock shelter, Staffordshire attempted radiocarbon dating, P. Bonvoisin for skilled
(I+-amwell, 19761. During the Iate DeVen- laboratory work on the mammalian remains, D. Clauaher for stereoscan studies. H. Tavior for nro-
Sian, Gulo was widespread in Europe, viding Fig. 3, and J. N. Downs for Fig. 4. Part of the where it was portrayed- by French Paleo- lithic artists. Its fossil remains have been found as far south as the Pyrenees in northern Spain (Clot, 1983) and northern Italy. All the British discoveries, except that from the Mundesley site, have been from caves.
toria Cave has been dated at ca. 120.000 vr
The presence of wolverine in Stump Cross Cave at ca. 83,000 yr B .P. has further significance. Victoria Cave, near Settle in Yorkshire, is only 23 km away and is at ap- proximately the same altitude. This fortui- tous situation makes possible a fauna1 com- parison, uncomplicated by major geograph- ical factors. The Ipswichian (X~SU micto) hippopotamus interglacial fauna from Vic-
U-series dating work described here was conducted in the Isotope Geology Unit of the Scottish Universitie\ Research and Reactor Centre, (East Kitbride. Scot- land) and part at the Atomic Energy Research Estab lishment (Harwell. England).
REFERENCES
Atkinson, T. C.. Harmon, R. S., Smart, P. L., and Waltham, A. C. (1978). Palaeochmatic and geo-
Mill rock shelter. Z~I “The Excavation of Wetton
morphic implications of the Z30Th/z34U dates on spe- leothems for Britain. Nu&re (Lornfdn) 272, 24-28,
Bishop, M. J. (1982). The mammal fauna of the early Middle Pleistocene cavern infill site of Westbury- Sub-Mendip, Somerset. Fu~ueontolo~ica~ Society ($ London ,Speciu/ Paper 28, I - 108.
Brain, C. K. (1981). “The Hunters or the Hunted’?.’ Univ. of Chicago Press, Chicago.
Bramwell, D. (1976). The vertebrate fauna at Wetton ,
indicates that substantial climatic deterio- ration had occurred in northern England
B.P. and correlated with marine isotopic
between these dates. The age of the Stump
substage 5e (Gascoyne et al., 1981). The
Cross fauna lies firmly within the youngest portion of marine isotope stage 5. If we ac-
presence of a fauna with wolverine in
cept the current view that the Eemian- Weichselian/Ipswichian-Devensian bound-
Stump Cross Cave, at ca. 83,000 yr B.P.,
ary should be drawn between isotope stages 5e and 5d (Mangerud et al., 1979; Woillard, 1982: Stuart, 1983), then the Stump Cross Cave fauna is of early Deven- Sian age.
Bulletin de 1~ Soci~5tk d*Hisioire Nutwelle Tou/o~se
Mill Rock Shelter. Maniford Valley, Staffordshire’. (J. H. Kelly, Ed.), pp. 40-51. City Museum and Art Gallery. Stoke-on-Trent. Staffordshire.
Clot, A. (1983). Le glouton (Gr&> &cl (L.1, Muste- hdae, Carnivora) de Labastide (Hautes Pyrenees).
118, lOl-110. Collins, E. R. (1959). The discovery of reindeer bones
in Stump Cross Caverns. Greenhow Hill. y~~r~.s/~& Archaeological Journul40, l60- 162.
Currant. A. P., Stringer? C. B., and Collcutt. S. N. (1984). Bacon Hole. 1/r “Handbook, South Wdlcs Field Meeting. April 1984.’ (D. Q. Bowen, Ed.), pp. 38-44. Quaternary Research Association, Abervs- twyth.
Dunham, K. C.. and Stubblefield, C. J. (194s). The stratigraphy% structure and mineralization of the Greenhow mining area. Yorkshire. Quurtvr/y Jourm/ of the Geo/ogicu/ Society <?f Lmdon 100. 209-268.
ACKNOWLEDGMENTS Analytical work was supported by a NERC grant
(GR3/4243) and by an EMR contract from AERE- Harweh. We are grateful to the late Ci. Gill, former proprietor of Stump Cross, and B. Gill and G. Hanley, the present proprietors, for allowing our investigation, for their help and encouragement, and for presenting specimens to the British Museum (Natural History). We thank A. Lister for determining the species of deer
Emiliani, C. (19%). Pleistocene temperatures. Jo~uxu/ of Geo/og~~ 63, 538-578.
Gascoyne, M., Currant. A. P., and Lord, T. ( I981 1. lpswichian fauna of Victoria Cave and the marine palaeoclimatic record. Nutwe CLondon) 294, 65?- 654.
Haglund, B. (1966). Winter habits of the lynx (L~~Ix !~H.x L.) and wolverine (Gu/o g~t/o L.) as revealed by trdcking in the snow. Vi/trevy 4, 81-3 IO.
Harmon, R. S., Ford, D. C., and Schwartz. H. P.

86 SUTCLIFFE ET AL.
(1977). Interglacial chronology of the Rocky and Mackenzie mountains based upon 23@fh/234U dating of calcite speleothems. Canadian Journal of Earth Sciences 14, 2543-2552.
Harmon, R. S., Schwartz, H. P., and Ford, D. C. (1978). Late Pleistocene sea-level history of Ber- muda. Quaternary Research 9, 20.5-218.
Hopkins, D. M., Matthews, J. V., Schweger, C. E., and Young, S. B. (1982). “Plaeoecology of Ber- ingia.” Academic Press, New York.
Ivanovich, M., and Harmon, R. S. (1982). “Uranium Series Disequilibrium: Applications to Environ- mental Problems.” Oxford Univ. Press (Clarendon), London/New York.
Krott, P. R. (1959). Der Vielfrasz. Moaographien der Wildstiugetiere, 13.
Kuyt, E. (1972). Food habits of wolves on barren- ground caribou range. Canadian Wildlife Service Report Series 21, l-36.
Mangerud, J., Sonstegaard, E., and Sejrup, H. (1979). Correlation of the Eemian (interglacial) Stage and the deep-sea oxygen-isotope stratigraphy. Nature (Londou) 277, 189- 192.
Newton, E. T. (1880). On the occurrence of the
glutton, Gulo luscus, in the “Forest Bed” of Mundesley, Norfolk. Geological Magazine 2, 424- 427.
Semenov-Tian-Shanskii, 0. I. (1982). “Animals of the Murmansk Region” Murmanskoe Knijnoe Izda- telstvo. [in Russian]
Shackleton, N. J. (1969). The last interglacial in the marine and terrestrial records. Proceedings of the Royal Society of London 174, 135-154.
Shackleton, N. J., and Opdyke, N. D. (1973). Oxygen isotope and palaeomagnetic stratigraphy of equato- rial core V28-238. Quaternary Research 3, 39-55.
Stuart, A. (1983). Pleistocene bone caves in Britain and Ireland. Studies in Speleology 4, 9-36.
Sutchffe, A. J., and Zeuner, F. E. (1962). Excavations in the Torbryan Caves, Devonshire. 1. Tornewton Cave. Proceedings of the Devon Archaelogical Ex- ploration Society 5, 127-145.
Waltham, A. C. (1975). “Limestones and Caves of North-West England.” David and Charles, Newton Abbot.
Woillard, G. M. (1982). Carbon-14 dates at Grande Pile: Correlation of land and sea chronologies. Sci- ence (Washington, D.C.) 215, 159-161.




















