
A 16,000 14 C yr B.P. packrat midden series from the USA-Mexico Borderlands
11
A 16,000 14 C yr B.P. packrat midden series from the USA–Mexico Borderlands Camille A. Holmgren, a, * M. Cristina Pen ˜alba, b Kate Aasen Rylander, a and Julio L. Betancourt a a Desert Laboratory, University of Arizona & U.S. Geological Survey, 1675 W. Anklam Rd., Tucson, AZ 85745, USA b Universidad Nacional Auto ´noma de Me ´xico, Instituto de Ecologı ´a, A.P. 1354, Hermosillo 83000, Sonora, Mexico Received 26 February 2003 Abstract A new packrat midden chronology from Playas Valley, southwestern New Mexico, is the first installment of an ongoing effort to reconstruct paleovegetation and paleoclimate in the U.S.A.–Mexico Borderlands. Playas Valley and neighboring basins supported pluvial lakes during full and/or late glacial times. Plant macrofossil and pollen assemblages from nine middens in the Playas Valley allow comparisons of two time intervals: 16,000 –10,000 and 4000 – 0 14 C yr B.P. Vegetation along pluvial lake margins consisted of open pinyon–juniper communities dominated by Pinus edulis, Juniperus scopulorum, Juniperus cf. coahuilensis, and a rich understory of C 4 annuals and grasses. This summer-flowering understory is also characteristic of modern desert grassland in the Borderlands and indicates at least moderate summer precipitation. P. edulis and J. scopulorum disappeared or were rare in the midden record by 10,670 14 C yr B.P. The late Holocene is marked by the arrival of Chihuahuan desert scrub elements and few departures as the vegetation gradually became modern in character. Larrea tridentata appears as late as 2190 14 C yr B.P. based on macrofossils, but may have been present as early as 4095 14 C yr B.P. based on pollen. Fouquieria splendens, one of the dominant desert species present at the site today, makes its first appearance only in the last millennium. The midden pollen assemblages are difficult to interpret; they lack modern analogs in surface pollen assemblages from stock tanks at different elevations in the Borderlands. © 2003 University of Washington. Published by Elsevier Inc. All rights reserved. Keywords: Pleistocene; Holocene; pluvial lakes; vegetation history; packrat middens; pinyon–juniper woodland; desert grassland; pollen Introduction More than 2000 packrat middens have been dated and analyzed to reconstruct late Quaternary vegetation changes across western North America, but several key areas remain unexplored. One such gap is the U.S.A.–Mexico Border- lands, the region near the continental divide where the states of Arizona and New Mexico intersect with each other and the Mexican states of Chihuahua and Sonora. The Border- lands occupy the broad transition between the Chihuahuan and Sonoran Deserts, are a hotspot for plant and animal diversity, and are of special concern due to recent woody plant encroachment into desert grasslands (Bahre and Web- ster, 2001). The packrat midden record from the Borderlands has been previously overlooked for mostly physical reasons. The area is generally wetter and has fewer rock outcrops suitable for midden preservation than adjacent regions. Here we report on a series of nine middens from the eastern Playas Valley in southwestern New Mexico span- ning the past 16,000 14 C yr. This series represents the first effort to answer several key biogeographic and paleocli- matic questions about the Borderlands. For example, where did glacial woodlands dominated by two-needled pinyons to the east give way to those dominated by single-needled pinyons to the west? What was the importance of C 4 grasses and summer annuals in these glacial woodlands, and hence the extent of summer rainfall? And did creosote bush (Larrea tridentata) recently expand from populations long established, or is the current expansion associated with an * Corresponding author. Fax: 1-520-670-6806. E-mail address: [email protected] (C.A. Holmgren). R Available online at www.sciencedirect.com Quaternary Research 60 (2003) 319 –329 www.elsevier.com/locate/yqres 0033-5894/$ – see front matter © 2003 University of Washington. Published by Elsevier Inc. All rights reserved. doi:10.1016/j.yqres.2003.08.001
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A 16,000 14 C yr B.P. packrat midden series from the USA-Mexico Borderlands

A 16,00014C yr B.P. packrat midden seriesfrom the USA–Mexico Borderlands
Camille A. Holmgren,a,* M. Cristina Penalba,b Kate Aasen Rylander,a
and Julio L. Betancourta
a Desert Laboratory, University of Arizona & U.S. Geological Survey, 1675 W. Anklam Rd., Tucson, AZ 85745, USAb Universidad Nacional Auto´noma de Me´xico, Instituto de Ecologı´a, A.P. 1354, Hermosillo 83000, Sonora, Mexico
Received 26 February 2003
Abstract
A new packrat midden chronology from Playas Valley, southwestern New Mexico, is the first installment of an ongoing effort toreconstruct paleovegetation and paleoclimate in the U.S.A.–Mexico Borderlands. Playas Valley and neighboring basins supported pluviallakes during full and/or late glacial times. Plant macrofossil and pollen assemblages from nine middens in the Playas Valley allowcomparisons of two time intervals: 16,000–10,000 and 4000–014C yr B.P. Vegetation along pluvial lake margins consisted of openpinyon–juniper communities dominated byPinus edulis, Juniperus scopulorum, Juniperuscf. coahuilensis,and a rich understory of C4annuals and grasses. This summer-flowering understory is also characteristic of modern desert grassland in the Borderlands and indicatesat least moderate summer precipitation.P. edulisandJ. scopulorumdisappeared or were rare in the midden record by 10,67014C yr B.P.The late Holocene is marked by the arrival of Chihuahuan desert scrub elements and few departures as the vegetation gradually becamemodern in character.Larrea tridentataappears as late as 219014C yr B.P. based on macrofossils, but may have been present as early as4095 14C yr B.P. based on pollen.Fouquieria splendens,one of the dominant desert species present at the site today, makes its firstappearance only in the last millennium. The midden pollen assemblages are difficult to interpret; they lack modern analogs in surface pollenassemblages from stock tanks at different elevations in the Borderlands.© 2003 University of Washington. Published by Elsevier Inc. All rights reserved.
Keywords:Pleistocene; Holocene; pluvial lakes; vegetation history; packrat middens; pinyon–juniper woodland; desert grassland; pollen
Introduction
More than 2000 packrat middens have been dated andanalyzed to reconstruct late Quaternary vegetation changesacross western North America, but several key areas remainunexplored. One such gap is the U.S.A.–Mexico Border-lands, the region near the continental divide where the statesof Arizona and New Mexico intersect with each other andthe Mexican states of Chihuahua and Sonora. The Border-lands occupy the broad transition between the Chihuahuanand Sonoran Deserts, are a hotspot for plant and animaldiversity, and are of special concern due to recent woodyplant encroachment into desert grasslands (Bahre and Web-
ster, 2001). The packrat midden record from the Borderlandshas been previously overlooked for mostly physical reasons.The area is generally wetter and has fewer rock outcropssuitable for midden preservation than adjacent regions.
Here we report on a series of nine middens from theeastern Playas Valley in southwestern New Mexico span-ning the past 16,00014C yr. This series represents the firsteffort to answer several key biogeographic and paleocli-matic questions about the Borderlands. For example, wheredid glacial woodlands dominated by two-needled pinyons tothe east give way to those dominated by single-needledpinyons to the west? What was the importance of C4 grassesand summer annuals in these glacial woodlands, and hencethe extent of summer rainfall? And did creosote bush(Larrea tridentata) recently expand from populations longestablished, or is the current expansion associated with an
* Corresponding author. Fax:�1-520-670-6806.E-mail address:[email protected] (C.A. Holmgren).
R
Available online at www.sciencedirect.com
Quaternary Research 60 (2003) 319–329 www.elsevier.com/locate/yqres
0033-5894/$ – see front matter © 2003 University of Washington. Published by Elsevier Inc. All rights reserved.doi:10.1016/j.yqres.2003.08.001

ongoing late Holocene migration of creosote bush across theBorderlands?
Study area
The Borderlands, part of the Mexican Highland sectionof the Basin and Range Province, are characterized byseveral northwest–southeast trending and tilted fault-blockranges separated by closed topographic basins (bolsons).These basins now contain ephemeral playas, but held plu-vial lakes (Animas, Cloverdale, Cochise, Goodsight, Ha-chita, Palomas, Playas) during the Pleistocene. Playas Val-ley is just north of the Mexican border and �50 km south ofLordsburg, New Mexico (Fig. 1). Playas Valley is flankedby the Alamo Hueco Mountains and the Little and BigHatchet Mountains to the east and the San Luis and AnimasMountains on the west. Elevations range from 1300 m onthe valley floor to �2600 m in the Big Hatchet and AnimasMountains. In the late Pleistocene, Playas Valley held a lake�10 m deep and 65 km2 in area with the highstand at�1311 m elevation (Hawley et al., 2000; Schwennesen,1918). The packrat middens reported here were collectedfrom two prominent features, Sentinel Butte and CoyoteHills, at the southern and northern end of Playas Valley,respectively. Sentinel Butte is an isolated hill of Cretaceouslimestone just west of the Alamo Hueco Mountains. The
Coyote Hills are a low range consisting of Oligocene ash-flow tuff and volcaniclastic rocks.
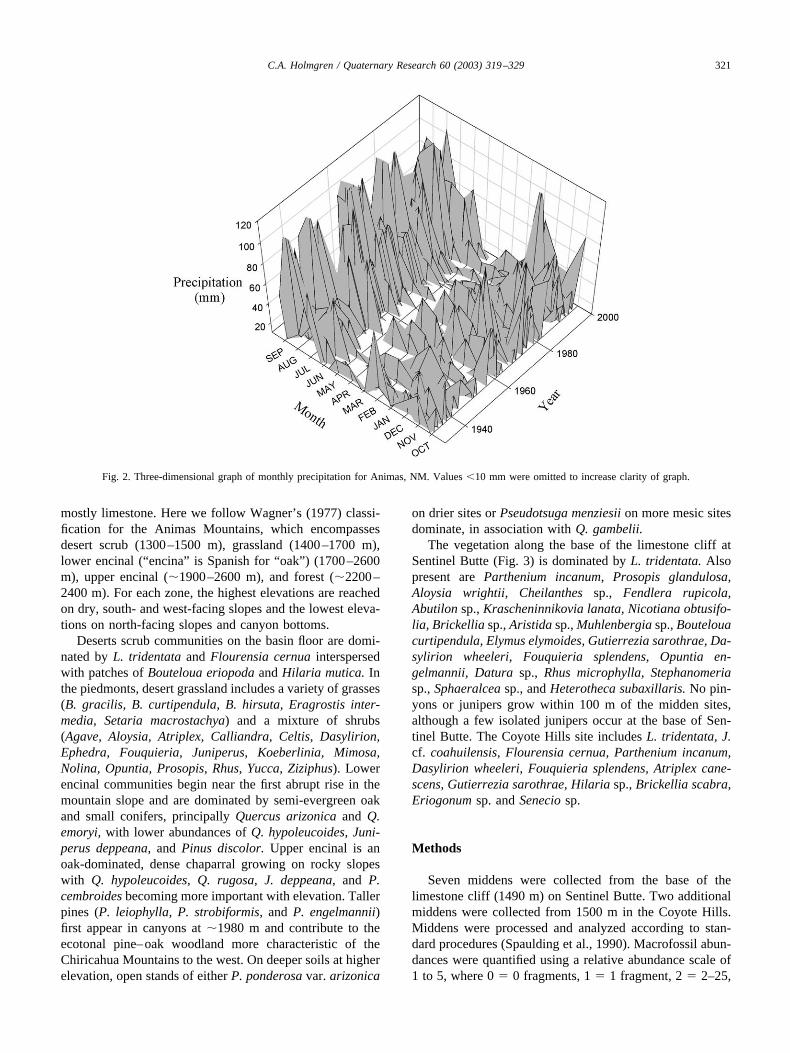
Average annual temperatures are �22.5°C in the low-lands, with daily highs/lows averaging 32°C/16°C in sum-mer (June–August) and 14°C/�4°C in winter (December–February). The valleys receive an annual average of 240–380 mm of precipitation, roughly half of which occursduring summer. Winter precipitation contributes �20% ofthe annual average precipitation. Seasonal and interannualprecipitation variability is evident in a three-dimensionalgraph of historical monthly precipitation for Animas, NewMexico near the northern end of Playas Valley (Fig. 2).Precipitation is more abundant and less variable from yearto year in summer than in winter. Winter precipitationresults from large Pacific frontal systems that migrate inlandfrom the west; the high year-to-year variability in winterand spring is modulated primarily by tropical sea surfacetemperature variations and associated teleconnections withthe subtropical jet and upper-air westerlies. Summer precip-itation variability is less well understood.
In the Playas Valley and surrounding mountains, vege-tation zonation with elevation varies widely with substrateand position on the landscape (i.e., slope aspect and degree,hillslope vs. canyon bottom). For example, oaks are bothdiverse and abundant, while Pinus discolor occurs sporad-ically in the igneous Animas and Alamo Hueco Mountains.The reverse is true in the Big Hatchet Mountains, which are
Fig. 1. Map of the Playas Basin and sites mentioned in text: (1) Tucson, AZ; (2) Willcox Playa/Paleolake Cochise; (3) Murray Springs; (4) Lehner Ranch;(5) Paleolake Cloverdale; (6) Sentinel Butte; (7) Coyote Hills; (8) Paleolake Animas; (9) Florida Mountains; (10) Bishop’s Cap; (11) Rough Canyon/OteroMesa; (12) Sacramento Mountains; (13) San Andreas Mountains; (14) Paleolake San Agustin; (15) Sevilleta LTER; (16) Paleolake Estancia; (17) HuecoMountains; (18) Guadalupe Mountains. Open squares correspond to pollen sites in Fig. 5: (A) 1220 m; (B) 1280 m; (C) 1260 m; (D) 1270 m; (E) 1380 m;(F) 1420 m; (G) 1460 m; (H) 1520 m; (I) 1580 m; (J) 1880 m.
320 C.A. Holmgren / Quaternary Research 60 (2003) 319–329
mostly limestone. Here we follow Wagner’s (1977) classi-fication for the Animas Mountains, which encompassesdesert scrub (1300–1500 m), grassland (1400–1700 m),lower encinal (“encina” is Spanish for “oak” ) (1700–2600m), upper encinal (�1900–2600 m), and forest (�2200–2400 m). For each zone, the highest elevations are reachedon dry, south- and west-facing slopes and the lowest eleva-tions on north-facing slopes and canyon bottoms.
Deserts scrub communities on the basin floor are domi-nated by L. tridentata and Flourensia cernua interspersedwith patches of Bouteloua eriopoda and Hilaria mutica. Inthe piedmonts, desert grassland includes a variety of grasses(B. gracilis, B. curtipendula, B. hirsuta, Eragrostis inter-media, Setaria macrostachya) and a mixture of shrubs(Agave, Aloysia, Atriplex, Calliandra, Celtis, Dasylirion,Ephedra, Fouquieria, Juniperus, Koeberlinia, Mimosa,Nolina, Opuntia, Prosopis, Rhus, Yucca, Ziziphus). Lowerencinal communities begin near the first abrupt rise in themountain slope and are dominated by semi-evergreen oakand small conifers, principally Quercus arizonica and Q.emoryi, with lower abundances of Q. hypoleucoides, Juni-perus deppeana, and Pinus discolor. Upper encinal is anoak-dominated, dense chaparral growing on rocky slopeswith Q. hypoleucoides, Q. rugosa, J. deppeana, and P.cembroides becoming more important with elevation. Tallerpines (P. leiophylla, P. strobiformis, and P. engelmannii)first appear in canyons at �1980 m and contribute to theecotonal pine–oak woodland more characteristic of theChiricahua Mountains to the west. On deeper soils at higherelevation, open stands of either P. ponderosa var. arizonica
on drier sites or Pseudotsuga menziesii on more mesic sitesdominate, in association with Q. gambelii.
The vegetation along the base of the limestone cliff atSentinel Butte (Fig. 3) is dominated by L. tridentata. Alsopresent are Parthenium incanum, Prosopis glandulosa,Aloysia wrightii, Cheilanthes sp., Fendlera rupicola,Abutilon sp., Krascheninnikovia lanata, Nicotiana obtusifo-lia, Brickellia sp., Aristida sp., Muhlenbergia sp., Boutelouacurtipendula, Elymus elymoides, Gutierrezia sarothrae, Da-sylirion wheeleri, Fouquieria splendens, Opuntia en-gelmannii, Datura sp., Rhus microphylla, Stephanomeriasp., Sphaeralcea sp., and Heterotheca subaxillaris. No pin-yons or junipers grow within 100 m of the midden sites,although a few isolated junipers occur at the base of Sen-tinel Butte. The Coyote Hills site includes L. tridentata, J.cf. coahuilensis, Flourensia cernua, Parthenium incanum,Dasylirion wheeleri, Fouquieria splendens, Atriplex cane-scens, Gutierrezia sarothrae, Hilaria sp., Brickellia scabra,Eriogonum sp. and Senecio sp.
Methods
Seven middens were collected from the base of thelimestone cliff (1490 m) on Sentinel Butte. Two additionalmiddens were collected from 1500 m in the Coyote Hills.Middens were processed and analyzed according to stan-dard procedures (Spaulding et al., 1990). Macrofossil abun-dances were quantified using a relative abundance scale of1 to 5, where 0 � 0 fragments, 1 � 1 fragment, 2 � 2–25,
Fig. 2. Three-dimensional graph of monthly precipitation for Animas, NM. Values �10 mm were omitted to increase clarity of graph.
321C.A. Holmgren / Quaternary Research 60 (2003) 319–329

2.5 � 26–50, 3 � 51–75, 3.5 � 76–100, 4 � 101–150, 4.5� 151–200, and 5 � 200 fragments. Midden material wasscreened through both 2-mm (No. 10) and 1-mm (No. 18)sieves to facilitate sorting plant remains. The 1-mm portionof Sentinel Butte #2B was lost and is not included in theresults. The possibility of bias against taxa represented bysmaller macrofossils has been taken into account in theinterpretation of the record for this sample.
Pollen samples were extracted from all middens. A smallindurated subsample (�10 g) was soaked in distilled water,disaggregated, and screened through a 500 �m (No. 35)mesh screen. A 10% KOH solution was added to removehumic acids. HCl was used to remove carbonates and todissolve the matrix of Lycopodium spore tablets added toquantify differences in pollen concentration between mid-dens. The residue was treated with HF to eliminate silica,acetolyzed, and then mounted in glycerine. More than 300pollen grains were counted in all but one midden (mean �379 pollen grains/sample; range � 266 to 501). After 300grains, the rest of the slide was scanned for poorly repre-sented types. Spores (except Lycopodium) were included inthe pollen sum.
Results
Nine middens span the last �16,000 14C yr in the PlayasValley (Table 1). Two middens date from the Pleistocene
(�11,000 14C yr B.P.), two from the early Holocene(11,000–8000 yr B.P.), two from the boundary between themiddle and late Holocene (�4000 14C yr B.P.), and threefrom the late Holocene (�4000 14C yr B.P.). No middensdated between 10,630 and 4100 14C yr B.P. All dates arereported as corrected, uncalibrated radiocarbon ages. A listof plant taxa appears in Table 2.
Plant macrofossil assemblages are summarized in Fig. 4.Pleistocene middens are dominated by P. edulis and J. spp.The midden from Sentinel Butte (16,040 14C yr B.P.) con-tains a toothed juniper that we tentatively identify as J.coahuilensis based on stomate position and morphology.The identification of toothed junipers in southern New Mex-ico is problematical and is discussed elsewhere in detail(Betancourt et al., 2001). The midden from Coyote Hills(13,830 14C yr B.P.) contains J. scopulorum, a smooth-margined juniper, and a toothed juniper with unusualsunken stomates that we believe may be a separate morpho-type of J. coahuilensis. Berberis cf. haematocarpa andMirabilis oxybaphus, both typically found above �1500 m,are also found only in Pleistocene middens.
In the case of Sentinel Butte 2C, temporal mixing ex-plains the presence of Prosopis glandulosa and a single L.tridentata leaf in the 16,040 14C yr B.P. sample containingabundant P. edulis and Juniperus. An AMS date of 17,02014C yr B.P. from a P. edulis needle falls within the 2� rangeof the original date (16,040 14C yr B.P.), while a date of4179 14C yr B.P. from Prosopis glandulosa indicates that itis indeed a younger contaminant. The Larrea leaf was toosmall to date even via AMS dating. However, given theabsence of L. tridentata from other northern ChihuahuanDesert middens before the late Holocene (Betancourt et al.,2001; Van Devender, 1990a; Van Devender and Toolin,1983; Van Devender et al., 1984), the fact that it is repre-sented by a solitary leaf, and the abundance of Larreasurrounding Sentinel Butte, we believe it is almost certainlya modern contaminant.
Some plants found in Pleistocene-aged middens werealso found in late Holocene middens. These include Arte-misia ludovicianal Asteraceae, Atriplex canescens, Boerha-via sp., Kallstroemia sp., Opuntia polyacantha–phaeacan-tha type, and the grasses Bothriochloa cf. barbinodis,Aristida sp., Bouteloua gracilis, Digitaria californica, Ely-mus elymoides/arizonicus, and Setaria sp. These species arepresent today and probably persisted at the site throughoutthe Holocene.
By the early Holocene, P. edulis had disappeared fromSentinel Butte (10,670 14C yr B.P.) and was rare at CoyoteHills (10,635 14C yr B.P.), where it is represented by asingle pinyon shell, possibly the result of long-distancedispersal by birds. Likewise, J. scopulorum had disappearedfrom Coyote Hills, although toothed junipers persisted atboth sites. The only record of Quercus in the Playas seriesis from the early Holocene at Coyote Hills.
Several plants characteristic of Chihuahuan desert scrub
Fig. 3. Photograph looking east toward Sentinel Butte.
322 C.A. Holmgren / Quaternary Research 60 (2003) 319–329

or desert grassland were present by 4000 14C yr B.P. Theseinclude Dasylirion wheeleri, Nicotiana obtusifolia, Parthe-nium incanum, Prosopis glandulosa, Bouteloua hirsuta, andViguiera sp. L. tridentata, a dominant species of Chihua-huan desert scrub, first appears in the macrofossils fromSentinel Butte at 2190 14C yr B.P. Fouquieria splendensand Senna sp. do not appear until 885 14C yr B.P.
A total of 49 different pollen types were identified in themiddens (Fig. 4). For Pinus, an attempt was made to dif-ferentiate the pollen grains at the species level, but distinc-tive characters could not be observed in most grains. Thefew pine pollen grains in which we could observe the distalmembrane indicate the presence of both P. edulis and P.ponderosa types at 16,040 14C yr B.P., P. ponderosa-type at10,635 and 220 14C yr B.P., and P. edulis-type at 10,670 and2190 14C yr B.P. The taxon Cupressaceae probably repre-sents Juniperus, since this is the only genus present in themacrofossil record.
In the 16,040 14C yr B.P. midden, Juniperus dominates(71.86%) with very low Asteraceae, Poaceae, and Chenopo-diaceae pollen. Pinus percentages are �10%, which con-trasts with the abundant P. edulis macrofossils in the sam-ple. However, the underrepresentation of pine pollen inmiddens, compared both to macrofossils (Thompson, 1985)and to the modern pollen rain (Anderson and Van Devender,1991; Martin, 1963), has been noted previously. Therefore,low pine pollen frequencies do not exclude the local pres-ence of Pinus. This may be especially true for P. edulis withpoorly dispersed pollen, while the well-dispersed P. pon-derosa pollen may come from distant populations. Quercusshows even lower pollen percentages although it producesabundant pollen. Low Quercus percentages were obtainedfrom modern pollen samples in open environments (Martin,1963, and see Fig. 5), suggesting that low percentages mayrepresent long-distance transport, in accordance with theabsence of macrofossils.
Late Pleistocene–early Holocene middens include the13,830, 10,670, and 10,635 14C yr B.P. samples. Main taxainclude Cupressaceae, Pinus, Quercus, Artemisia, Chenopo-diaceae, Poaceae, and Asteraceae Tubuliflorae (character-istic of the desert pollen spectra). The 10,635 14C yr B.P.
midden shows slightly higher Cupressaceae values, lowerPoaceae and Asteraceae values, and the only Rosaceae pol-len in the record. Traces of Potamogeton pollen are found inthe 13,830, 10,635, 4040, and 885 14C yr B.P. samples. Thisaquatic species lives in ponds and slow-moving rivers; itspresence in the middens may indicate that ephemeral pondsexisted during wet years or it may be derived from older,reworked Pleistocene playas.
The middens at 4095 and 4040 14C yr B.P. are charac-terized by high Asteraceae, Poaceae, and Chenopodiaceaeand very low Cupressaceae, Pinus, and Quercus pollenvalues. Prosopis, Mimosa, and Larrea have intermediatevalues. Sporadic pollen grains of Prosopis, Mimosa, andLarrea are seen before this time, but probably representcontamination. The 4040 14C yr B.P. midden also containsfive pollen grains that appear to be cereal based on theirlarge (�70-�m) diameter.
In the 2190 14C yr B.P. midden Prosopis, Mimosa,Larrea, Asteraceae, and Poaceae are dominant, suggestingessentially modern vegetation. Fouquieria splendens andOpuntia first occur in this zone, as well as the only pollengrain of Zea mays identified with certainty in the record. Z.mays had appeared at least a millennium earlier in NewMexico and Arizona (Shackley et al., 1999; Tagg, 1996;Upham et al., 1987; Wills, 1988). The 885 and 220 14C yrB.P. samples are characterized by very high pollen valuesfor Larrea and very low pollen values for Pinus, Quercusand other desert taxa.
Fig. 5 compares midden pollen and macrofossil assem-blages with surface pollen samples collected from stockponds at various elevations in the Borderlands (see Martin,1963, for original descriptions of the surface pollen data).Many of the differences between these types of assemblageswere to be expected from previous comparisons (Andersonand Van Devender, 1991; Davis and Anderson, 1987;Thompson, 1985). Midden pollen assemblages tend to bemore locally variable than surface pollen samples due to thediverse modes of pollen transport into middens and greaterheterogeneity in habitat and flora of cliffs compared to moreopen landscapes. Pollen may be incorporated into middensby wind or plant parts or via the packrat’s body, and locally
Table 1Site location and radiocarbon dates from Sentinel Butte and Coyote Hills packrat middens
Midden designation Latitude(°N)
Longitude(°W)
Elevation(m)
Materialdated
14C age(14C yr B.P.)
Standard deviation14C yr
�13C(‰)
Calibrated(2�) age range(cal yr B.P.)
Lab No.
Sentinel Butte 1 31°26.460 108°37.080 1488 fecal pellets 2190 125 �23.5 2477–1873 GX21641Sentinel Butte 2A 31°26.460 108°37.080 1488 fecal pellets 4040 100 �22 4828–4280 GX21642Sentinel Butte 2B 31°26.460 108°37.080 1488 fecal pellets 10670 330 �22.2 13,201–11,547 GX21208Sentinel Butte 2C 31°26.460 108°37.080 1488 fecal pellets 16040 660 �22.7 20,771–17,540 GX21209Sentinel Butte 3A 31°26.460 108°37.080 1488 fecal pellets 885 115 �25.2 997–648 GX21643Sentinel Butte 3B 31°26.460 108°37.080 1488 fecal pellets 4095 145 �24.2 4888–4220 GX21644Sentinel Butte 4 31°26.460 108°37.080 1488 fecal pellets 220 105 �24.7 462–0 GX21645Coyote Hills 1A 32°00�10� 108°30�15� 1502 fecal pellets 10635 340 �24.4 13,190–11,333 GX19992Coyote Hills 1B 32°00�10� 108°30�15� 1502 fecal pellets 13830 165 �24.8 17,204–16,012 GX19993
323C.A. Holmgren / Quaternary Research 60 (2003) 319–329

occurring taxa tend to be better represented than in surfacepollen samples. Midden pollen assemblages frequently in-clude zoophilous species rarely recorded in surface samples,and some wind-pollinated species tend to be underrepre-sented, while others are overrepresented. For example,pines (Pinus) and grasses (Poaceae) tend to be underrepre-sented in midden pollen relative to midden macrofossils andsurface pollen samples, while juniper (Juniperus) pollen can
be overrepresented in middens due to the occasional incor-poration of male cones. Cheno-Am pollen, on the otherhand, is overrepresented in modern surface samples, prob-ably an artifact of chronic disturbances near stock tanks andovergrazed grasslands (Martin, 1963). Overall, we found noanalogs between the midden pollen assemblages at differenttimes and the surface pollen samples from different eleva-tions.
Table 2Scientific and common names of plant taxa mentioned in text
Scientific name Common name Scientific name Common name
Agavaceae NyctaginaceaeAgave spp. Agave Boerhavia sp. SpiderlingYucca spp. Yucca Mirabilis oxybaphus Four-o’ -clock
Anacardiaceae PinaceaeRhus microphylla Littleleaf sumac Pinus cembroides Mexican pinyon
Asteraceae Pinus discolor Border pinyonArtemisia ludoviciana Louisiana wormwood Pinus edulis Colorado pinyonBrickellia scabra Rough Brickelbush Pinus engelmannii Arizona longleaf pineFlourensia cernua Tarbush Pinus leiophylla Chihuahua pineGutierrezia sarothrae Broom snakeweed Pinus monophylla var. fallax Singleleaf pinyonHeterotheca subaxillaris Telegraph plant Pinus ponderosa var. arizonica Arizona yellow pineParthenium incanum Mariola Pinus ponderosa Ponderosa PineSenecio sp. Groundsel Pinus remota Papershell pinyonStephanomeria sp. Wire lettuce Pinus strobiformis Southwestern white pineViguiera sp. Goldeneye Pseudotsuga menziesii Douglas fir
Berberidaceae PoaceaeBerberis haematocarpa Red barberry Aristida sp. Threeawn
Cactaceae Bothriochloa barbinodis Cane beardgrassOpuntia engelmannii Engelman’s pricky pear Bouteloua curtipendula Sideoats gramaOpuntia phaecantha Variable prickly pear Bouteloua eripoda Black gramaOpuntia polycantha Plains prickly pear Bouteloua gracilis Blue grama
Chenopodiaceae Bouteloua hirsuta Hairy gramaAtriplex canescens Fourwing saltbush Digitaria californica Arizona cottontopKrascheninnikovia lanata Winterfat Elymus elymoides Squirreltail grass
Cupressaceae Eragrostis intermedia Plains lovegrassJuniperus scopulorum Rocky Mountain juniper Hilaria mutica Tobosa grassJuniperus deppeana Alligator bark juniper Muhlenbergia sp. MuhlyJuniperus coahuilensis Coahuila juniper Zea mays Corn
Ephedraceae Setaria macrostachya Plains bristlegrassEphedra sp. Joint fir Polygonaceae
Fabaceae Eriogonum sp. Wild buckwheatCalliandra sp. Fairy duster PotamogetonaceaeSenna sp. Senna Potamogeton sp. PondweedMimosa spp. Mimosa PteridaceaeProsopis glandulosa Honey mesquite Cheilanthes sp. Lip fern
Fagaceae RhamnaceaeQuercus arizonica Arizona white oak Celtis reticulata Netleaf hackberryQuercus emoryi Emory oak Ziziphus obtusifolia GraythornQuercus gambelii Gambel oak SaxifragaceaeQuercus hypoleucoides Silverleaf oak Fendlera rupicola Cliff FendlerbushQuercus rugosa Netleaf oak Solanaceae
Fouquieriaceae Datura sp. JimsonweedFouquieria splendens Ocotillo Nicotiana obtusifolia Desert tobacco
Koeberliniaceae VerbenaceaeKoeberlinia spinosa Allthorn Alyosia wrightii Spicebush
Malvaceae ZygophyllaceaeAbutilon sp. Indian mallow Kallstroemia sp. CaltropSpharalcea sp. Globemallow Larrea tridentata Creosotebush
NolinaceaeDasylirion wheeleri SotolNolina microcarpa Beargrass
324 C.A. Holmgren / Quaternary Research 60 (2003) 319–329

Fig. 4. Plant macrofossil abundance (top graph) and pollen percentage (bottom graph) through time for select species from Sentinel Butte and Coyote Hills, NM. Stippledarea indicates period from which no middens were found.
325C
.A.
Holm
gren/
Quaternary
Research
60(2003)
319–329

Discussion
The Borderlands midden record indicates that pinyon–juniper woodlands with a rich C4 understory bordered plu-vial lakes that formed in Playas Valley and neighboringbasins. The lake-level histories of these basins are not nec-essarily in phase, with some basins registering full-glacialhighstands and others late glacial ones. Lake Cloverdale insouthernmost Animas Valley filled between 20,000–18,00014C yr B.P. (Krider, 1998), similar to highstands in the SanAgustin Plains (Markgraf et al., 1984; Phillips et al., 1992)and Estancia Basin (Allen and Anderson, 2000) to the north,but fails to record the late glacial highstand seen in LakeCochise.
There are two lake chronologies for pluvial Lake Co-chise. The first was based on 14C dates from secondarycarbonates in two lacustrine green clay units that reached1290 m, above the prominent 1274-m shoreline above Will-cox Playa. Long (1966) proposed that lake levels weresignificantly higher than the 1274-m beach ridge from30,000 to 13,000 14C yr B.P., dropped between 13,000 and11,500 14C yr B.P., and rose again to near the 1274-m levelbetween 11,500 and 10,500 14C yr B.P. Later, Waters(1989) used a series of backhoe trenches to map and date theshoreline stratigraphy just below 1274 m. Radiocarbondates from charcoal in beach deposits overlying the greenclay suggest that lake levels were at 1274 m from at least13,750 to 13,400 14C yr B.P. (Waters, 1989). This impliesthat the highstand associated with the underlying greenclays formed prior to 13,750 14C yr B.P. and the carbonate
dates from the clays are too young. Dates from humates andorganic carbon also indicate that after a period of dessica-tion, Lake Cochise again filled to slightly below the 1274 mshoreline three times during the Holocene, once around orbefore 8900 14C yr B.P. and again at �5400 and �4000–3000 14C yr B.P. (Waters, 1989).
Pluvial Lake Animas experienced a high stand sometimebetween 24,000 and 8000 14C yr B.P. (Fleishhauer andStone, 1982), but inadequate dating precludes precise com-parison with other pluvial lakes. Suffice it to say that theclimatic conditions for development of pinyon–juniperwoodland at �1400–1500 m at 16,000 and 13,800 14C yrB.P. were also conducive to pluvial lake expansion in theBorderlands.
Late Pleistocene vegetation is marked by expansion ofwoodlands dominated by P. edulis and J. coahuilensis atSentinel Butte and P. edulis and J. coahuilensis plus J.scopulorum at Coyote Hills. This is consistent with otherregional records that indicate pinyon–juniper woodlands inwhat is currently Chihuahuan desert scrub (Betancourt etal., 2001; Lanner and Van Devender, 1981; Van Devender,1990a; Van Devender et al., 1984; Van Devender and Ever-itt, 1977; Van Devender and Toolin, 1983). Pinus pollenfrom Willcox Playa in southeastern Arizona reached 99%during the late Pleistocene (22,000 14C yr B.P.), indicatingexpansion of pines (Martin and Mehringer, 1965). Thesehigh pine percentages were previously attributed to pineforest expansion (P. ponderosa var. arizona, P. leiophylla,P. strobiformis, and P. engelmannii) (Martin, 1963). Thehigh pine pollen in lake sediments, however, could instead
Fig. 5. Comparison of midden macrofossils and pollen (top graph) with modern surface pollen samples (bottom graph). Open bars are for macrofossilabundances (1–5), closed bars for pollen percentages (1–100%). The modern surface pollen data are from Martin (1963, Table 1).
326 C.A. Holmgren / Quaternary Research 60 (2003) 319–329

represent the cumulative effect of pine pollen dispersal fromwidespread pinyon–juniper woodlands in the vast lowlandsupwind of Willcox Playa to the Gulf of California.
The Playas Valley middens add further detail to theglacial-age biogeography of pinyons. Dominant species in-cluded P. monophylla var. fallax in the Sonoran Desert, P.remota in the Chihuahuan Desert, and P. edulis in what arenow desert grasslands in New Mexico and eastern Arizona(Lanner and Van Devender, 1981). In the Catalina Moun-tains near Tucson, P. discolor was sympatric with a P.monophylla edulis hybrid (Van Devender, 1990a). The Pla-yas Valley middens indicate that P. edulis was the dominantpinyon across all of southern New Mexico. This begs thequestion of whether present populations of P. monophyllavar. fallax in the Florida Mountains northeast of PlayasValley represent Holocene dispersals instead of Pleistocenerelicts.
The presence of J. scopulorum at Coyote Hills indicatesit descended to �1500 m on igneous substrates during thelate Pleistocene, while its absence at Sentinel Butte at leastlocally may be due to more xeric limestone substrate. J.scopulorum was also common, however, in other Pleisto-cene middens at �1500 m from limestone in the Los Pinos,Guadalupe, Sacramento, and Hueco Mountains. An isolatedpopulation was also found at 975 m in Pleistocene middensfrom Organ Pipe National Monument along the Arizona–Sonora border (Van Devender, 1990b). The presence of J.scopulorum in the Playas Valley middens further suggeststhat J. deppeana, the dominant juniper at mid to high ele-vations in the Borderlands today, must have been displacedsouth into the Sierra Madre Occidental of northern Mexico(Betancourt et al., 2001).
Although both lake and pinyon–juniper expansion acrossthe lowlands have been attributed to greater winter precip-itation, the Pleistocene middens from Playas Valley containseveral summer rainfall indicators, including the midsum-mer annuals Kallstroemia and Boerhavia sp. and the C4
grasses Bouteloua cf. gracilis, Bothriochloa barbinodis, andDigitaria californica. Vertebrate remains from U-Bar Cave(1570 m), only a few km north of Sentinel Butte implywoodland with a well-developed grassy understory (Harris,1987). Carbon isotope values from soil carbonates else-where in the Borderlands (Buck and Monger, 1999; Liu etal., 1996; Monger et al., 1998) and tooth enamel frommegaherbivores at U-Bar Cave, also indicate C4 dominancein woodland understories and grasslands during the full tolate glacial (Connin et al., 1998).
Extirpation of pinyon–juniper woodlands must have oc-curred by 10,600 14C yr B.P., when P. edulis and J. scopu-lorum disappeared from the lowlands of the Playas Valley.This extirpation may have occurred abruptly during whatHaynes (1991) has termed the “Clovis-age Drought.” AtMurray Springs in the San Pedro Valley of southern Ari-zona, the water table began to decline after 14,000 14C yrB.P. and dropped to unusually low levels just before 10,90014C yr B.P., corresponding to the disappearance of Rancho-
labrean megafauna. Groundwater levels quickly reboundedby 10,800 14C yr B.P., depositing black organic mats(Haynes, 1991). New AMS dates bracket the black matswithin the Younger Dryas cold event from �11,000 to10,000 14C yr B.P. (Jull et al., 1998). At most sites in thenorthern Chihuahuan desert, the disappearance of pinyonand change to more xeric oak–juniper woodland also oc-curred around 11,000 14C yr B.P. (Van Devender, 1990a),although earlier departures are also reported (Betancourt etal., 2001). Pollen records from Lehner Ranch in the SanPedro (Mehringer and Haynes, 1965) and Sulfur Springsvalleys (Martin, 1963) also indicate that Pinus had de-creased drastically and desert grassland was established by11,000 14C yr B.P.
Although woody species changed dramatically in theearly Holocene, changes in understory vegetation were lessnoticeable. Faunal stratigraphy from Howell’s Ridge Cavein the Little Hatchet Mountains suggests well-developedgrasslands in the early Holocene (Van Devender, 1995; VanDevender and Worthington, 1977), while a shift from C4
grasses to C3 desert scrub is indicated by soil carbonates atlower, drier sites in the Southwest (Buck and Monger, 1999;Liu et al., 1996; Monger et al., 1998). The hillslope under-story seen in early Holocene Playas Valley middens wasremarkably stable with no noticeable changes in grass com-position or abundance.
No middens in our series dated from the middle Holo-cene (8000–4000 14C yr B.P.), a period during which mid-dens are scarce across the Southwest. One possible expla-nation for this scarcity is that a pervasive winter droughtduring the middle Holocene led to a decline in woodyperennials, which predominantly depend on winter mois-ture, and in packrat populations and midden production(Betancourt, 1993; Spaulding, 1991). Because our seriesconsists of only nine samples, however, the lack of middensduring this period may be due to small sample size. Studiesusing larger numbers of middens should help resolve thisissue.
The Playas Valley middens record progressive arrivals ofdesert scrub species from �4000 14C yr B.P. onwards. Thefirst occurrence of L. tridentata macrofossils at 2190 14C yrB.P. is later than in other regional midden records, althougha small peak in Larrea pollen at 4095 14C yr B.P. couldindicate an earlier arrival. Larrea is found in ChihuahuanDesert middens from Otero Mesa by 4750 14C yr B.P., fromthe San Andres Mountains by 4340 14C yr B.P., and fromthe Sacramento Mountains by 3300 (Betancourt et al., 2001;Van Devender, 1990a; Van Devender et al., 1984; VanDevender and Toolin, 1983). Earlier records of Larrea fromthe Sonoran desert suggest that dispersal from the north andwest was also possible. The closest record in the Sonorandesert is from the Waterman Mountains west of Tucson,Arizona where Larrea was present in middens by 6195 14Cyr B.P (Anderson and Van Devender, 1991; Van Devender,1990b). Although the diploid race (2n � 26) of Larreaoccurs primarily in the Chihuahuan desert and the tetraploid
327C.A. Holmgren / Quaternary Research 60 (2003) 319–329

race (2n � 52) in the Sonoran desert, ploidy level cannot beused to determine the source of the diploid macrofossilspresent at Sentinel Butte. Diploid isolates are found inregions of the Sonoran desert dominated by tetraploids,including the Waterman Mountains, where the Larrea mac-rofossils identified in 6195 14C yr B.P. and 1320 14C yr B.P.middens are diploid (Hunter et al., 2001). Thus, the diploidLarrea at Sentinel Butte could be derived from diploidpopulations in either the Chihuahuan or Sonoran Desert.
The arrival of Fouquieria splendens was also later thanin surrounding areas. Fouquieria was present at RoughCanyon, New Mexico by 6295 14C yr B.P. (Betancourt etal., 2001) and in the Waterman Mountains, Arizona by 260014C yr B.P. (Anderson and Van Devender, 1991). The latearrival of Fouquieria splendens at 885 14C yr B.P. is similarto that at the Sevilleta LTER site south of Albuquerque(Betancourt et al., in preparation) and suggests that vegeta-tion was essentially modern by this time.
The midden sequence indicates that pinyon–juniperwoodland with a C4 understory expanded downslope to themargins of pluvial lakes in Playas Valley and surroundingbasins during the late Pleistocene, suggesting increased ef-fective moisture with at least moderate summer rainfall. Thetiming of pinyon–juniper woodland extirpation conforms toother regional records, although little change is seen in theunderstory. By about 4000 14C yr B.P., desert scrub ele-ments begin to appear, marking the transition to present-dayvegetation. Development of more extensive midden seriesfrom the Borderlands could help resolve lingering questionsregarding precipitation seasonality, understory dynamics,mid-Holocene climate, and the exact timing of desert scrubarrivals in the area.
Acknowledgments
We thank R.S. Barclay, M.L. Betancourt, and R.M.Turner for field assistance, L. Rivero Montes for laboratoryassistance, William Hurt for access to Sentinel Butte, andthe Geochronology Laboratory for 14C dates. We thank S.Pearson and P. Griffiths for assistance with figures and R.S.Anderson, P.S. Martin, and T.R. Van Devender for thought-ful reviews. This study was supported by the USGS GlobalChange Program.
References
Allen, B.D., Anderson, R.Y., 2000. A continuous record of late Pleistoceneclimate variability from the Estancia Basin, New Mexico. GeologicalSociety of America Bulletin 112, 1444–1458.
Anderson, R.S., Van Devender, T.R., 1991. Comparison of pollen andmacrofossils in packrat middens: A chronological sequence from theWaterman Mountains of southern Arizona, USA. Review of Paleo-botany and Palynology 68, 1–28.
Bahre, C.J., Webster, G.L., Eds. 2001. Changing Plant Life of La Frontera:Observations of Vegetation in the United States/Mexico Borderlands.University of New Mexico Press, Albuquerque, pp. 84–95.
Betancourt, J.L., Pierson, E.A., Aasen-Rylander, K., Fairchild-Parks, J.A.,Dean, J.S., 1993. Influence of history and climate on New Mexicopinyon–juniper woodlands, in: Aldon, E.F., Shaw, D.W. (Eds.), Man-aging Pinyon–Juniper Ecosystems for Sustainability and Social Needs.Proceedings of the Symposium April 26–30, Santa Fe, New Mexico.Gen. Tech. Rep. RM-236. USDA Forest Service, Rocky MountainForest and Range Exp. Station, Ft. Collins, CO, pp. 42–62.
Betancourt, J.L., Rylander, K.A., Penalba, C., McVickar, J.L., 2001. LateQuaternary vegetation history of Rough Canyon, south-central NewMexico, USA. Palaeogeography, Palaeoclimatology, Palaeoecology165, 71–95.
Buck, B.J., Monger, H.C., 1999. Stable isotopes and soil-geomorphologyas indicators of Holocene climate change, northern Chihuahuan Desert.Journal of Arid Environments 43, 357–373.
Connin, S.L., Betancourt, J.L., Quade, J., 1998. Late Pleistocene C4 plantdominance and summer rainfall in the southwestern United States fromisotopic study of herbivore teeth. Quaternary Research 50, 179–193.
Davis, O.K., Anderson, R.S., 1987. Pollen in packrat (Neotoma) middens:pollen transport and the relationship of pollen to vegetation. Palynology11, 185–198.
Fleischhauer, H.L., Jr., Stone, W.J., (1982). Quaternary Geology of LakeAnimas, Hidalgo Country, New Mexico, New Mexico Bureau of Minesand Mineral Resources Circular 174. Socorro, NM.
Harris, A.H., 1987. Reconstruction of Mid-Wisconsin environments insouthern New Mexico. National Geographic Research 3, 142–151.
Hawley, J.W., Hibbs, H.J., Kennedy, J.F., Creel, B.J., Remmenga, M.D.,Johnson, M., Lee, M., Dinterman, P., (2000). Trans-InternationalBoundary Aquifers in Southwestern New Mexico. Technical Comple-tion Report, Interagency Contract No. X-996-350-01-3. New MexicoWater Resources Research Institute. Available at: http://wrri.nmsu.edu/publish/otherrpt/swnm/.
Haynes, C.V., 1991. Geoarchaeological and paleohydrological evidencefor a Clovis–age drought in North America and its bearing on extinc-tion. Quaternary Research 35, 438–450.
Hunter, K.L., Betancourt, J.L., Riddle, B.R., Van Devender, T.R., Cole,K.L., Spaulding, W.G., 2001. Ploidy race distributions since the LastGlacial Maximum in the North American desert shrub, Larrea triden-tata. Global Ecology and Biogeography 10, 521–533.
Jull, A.J., Haynes, C.V., Donahue, D.J., Burr, G.S., Beck, J.W., (1998).Radiocarbon ages at Murray Springs, Arizona and the influence ofclimate change on Clovis Man. Revue d’ Archaeometrie Supp 339–343.
Krider, P.R., 1998. Paleoclimatic significance of late Quaternary lacustrineand alluvial stratigraphy, Animas Valley, New Mexico. QuaternaryResearch 50, 283–289.
Lanner, R.M., Van Devender, T.R., 1981. Late Pleistocene pinon pines inthe Chihuahuan Desert. Quaternary Research 15, 278–280.
Liu, B., Phillips, F.M., Campbell, A.R., 1996. Stable carbon and oxygenisotopes of pedogenic carbonates, Ajo Mountains, southern Arizona:implications for paleoenvironmental change. Palaeogeography, Palaeo-climatology, Palaeoecology 124, 233–246.
Long, A., 1966. Late Pleistocene and Recent Chronologies of Playa Lakesin Arizona and New Mexico, unpublished Ph.D. dissertation. Univer-sity of Arizona, Tucson.
Markgraf, V., Bradbury, J.P., Forester, R.M., Singh, G., Sternberg, R.S.,1984. San Agustin Plains, New Mexico: age and paleoenvironmentalpotential reassessed. Quaternary Research 22, 336–343.
Martin, P.S., 1963. The last 10,000 years. Univ. of Arizona Press, Tucson.Martin, P.S., Mehringer Jr., P.J., 1965. Pleistocene pollen analysis and
biogeography of the southwest, in: Wright Jr., H.E., Frey, D.G. (Eds.),The Quaternary of the United States, Princeton University Press,Princeton, NJ, pp. 433–451.
Mehringer, P.J., Haynes, C.V., 1965. The pollen evidence for the environ-ment of early man and extinct mammals of the Lehner mammoth site,southeastern Arizona. American Antiquity 32, 17–23.
Monger, H.C., Cole, D.R., Gish, J.W., Giordano, T.H., 1998. Stable carbonand oxygen isotopes in Quaternary soil carbonates as indicators of
328 C.A. Holmgren / Quaternary Research 60 (2003) 319–329

ecogeomorphic changes in the northern Chihuahuan Desert, USA.Geoderma 82, 137–172.
Phillips, F.M., Campbell, A.R., Kruger, C., Johnson, P., Roberts, R.,Keyes, E., (1992). A Reconstruction of the Water Balance in WesternUnited States Lake Basins to Climatic Change, New Mexico WaterResources Research Institute Report 269.
Schwennesen, A.T., (1918). Ground Water in the Animas, Playas, Hachitaand San Luis Basins, New Mexico, United States Geological SurveyWater-Supply Paper 422.
Shackley, M.S., Huckell, B.B., Huckell, L.W., 1999. McEuen Cave inEarly Maize in the Greater Southwest. Archaeology Southwest 13, 12.
Spaulding, W.G., 1991. A middle Holocene vegetation record from theMojave Desert of North America and its paleoclimatic significance.Quaternary Research 35, 427–437.
Spaulding, W.G., Betancourt, J.L., Croft, L.K., Cole, K.L., 1990. Packratmiddens: their composition and methods of analysis, in: Betancourt,J.L., Van Devender, T.R., Martin, P.S. (Eds.), Packrat Middens: TheLast 40,000 Years of Biotic Change, University of Arizona Press,Tucson, pp. 59–84.
Tagg, M.D., 1996. Early cultigens from Fresnal Shelter, southeastern NewMexico. American Antiquity 62, 311–324.
Thompson, R.S., 1985. Palynology and Neotoma middens. American As-sociation of Stratigraphic Palynologists Contributions Series 16, 89–112.
Upham, S., MacNeish, R.S., Galinat, W.C., Stevenson, C.M., 1987. Evi-dence concerning the origin of Maiz de Ocho. American Anthropolo-gist 89, 410–419.
Van Devender, T.R., 1990a. Late Quaternary vegetation and climate of theChihuahuan Desert, United States and Mexico, in: Betancourt, J.L.,Van Devender, T.R., Martin, P.S. (Eds.), Packrat Middens: The Last40,000 Years of Biotic Change, University of Arizona Press, Tucson,pp. 104–133.
Van Devender, T.R., 1990b. Late Quaternary vegetation and climate of the
Sonoran Desert, United States and Mexico, in: Betancourt, J.L., VanDevender, T.R., Martin, P.S. (Eds.), University of Arizona Press, Tuc-son, pp. 134–165.
Van Devender, T.R., 1995. Desert grassland history: changing climates,evolution, biogeography, and community dynamics, in: McClaran,M.P., Van Devender, T.R. (Eds.), The Desert Grassland, University ofArizona Press, Tucson, pp. 68–95.
Van Devender, T.R., Betancourt, J.L., Wimberley, M., 1984. Biogeo-graphic implications of a packrat midden sequence from the Sacra-mento Mountains, south-central New Mexico. Quaternary Research 22,344–360.
Van Devender, T.R., Everitt, B.L., 1977. The latest Pleistocene and Recentvegetation of Bishop’s Cap, south-central New Mexico. Southwest.Nature 23, 289–302.
Van Devender, T.R., Toolin, L.J., 1983. Late Quaternary vegetation of theSan Andres Mountains, Sierra County, New Mexico, in: Eidenbach,P.L. (Ed.), The Prehistory of Rhodes Canyon: Survey and Mitigation,Report to Holloman Air Force Base, Human Systems Research, Tylo-role, NM, pp. 33–54.
Van Devender, T.R., Worthington, R.D., 1977. The herpetofauna of How-ell’s Ridge Cave and the paleoecology of the northwestern ChihuahuanDesert, in: Wauer, R.H., Riskind, D.H. (Eds.), Transactions of theSymposium on the Biological Resources of the Chihuahuan DesertRegion, United States and Mexico, Transactions and Proceedings Se-ries Number 13. U.S. Department of Interior National Park Service,Washington, DC, pp. 85–106.
Wagner, W.L., 1977. Floristic affinities of Animas Mountains, southwest-ern New Mexico, M.S. thesis. University of New Mexico, Albuquerque.
Waters, M.R., 1989. Late Quaternary lacustrine history and paleoclimaticsignificance of pluvial Lake Cochise, southeastern Arizona. QuaternaryResearch 32, 1–11.
Wills, W.H., 1988. Early Prehistoric Agriculture in the American South-west. School of American Research Press, Santa Fe.
329C.A. Holmgren / Quaternary Research 60 (2003) 319–329




















