
Optical Biosensors for the Detection of Rheumatoid Arthritis ...
Upload
independentCategory
view
4download
0
Whole cell- and protein-based biosensors for the detection ofbioavailable heavy metals in environmental samples
Philippe Corbisiera,*, Daniel van der Leliea, Brigitte Borremansa, Ann Provoosta,Victor de Lorenzob, Nigel L. Brownc, Jonathan R. Lloydc, Jonathan L. Hobmanc,
Elisabeth CsoÈregid, Gillis Johanssond, Bo Mattiassond
aVlaamse Instelling voor Technologisch Onderzoek (VITO), Boeretang 200, B-2400 Mol, BelgiumbCentro Nacional de BiotecnologõÂa, Campus de Cantoblanco, Universidad AutoÂnoma, Spain
cThe University of Birmingham, School of Biological Sciences, Edgbaston, Birmingham B15 2TT, UKdDepartment of Biotechnology, Centre for Chemistry and Chemical Engineering, Lund University, PO Box 124, S-221 00 Lund, Sweden
Received 8 July 1998; received in revised form 9 October 1998; accepted 11 October 1998
Abstract
The principal goal of this work was to establish the feasibility of two biosensor technologies with enhanced speci®city and
selectivity for the detection of several bioavailable heavy metals in environmental samples. Two parallel strategies have been
followed. The ®rst approach was to construct whole cell bacterial biosensors that emit a bioluminescent or ¯uorescent signal
in the presence of a biologically available heavy metal. The molecular basis of s-54 promoters as sensing elements of
environmental pollutants has been determined and a number of metal-induced promoter regions have been identi®ed,
sequenced and cloned as promoter cassettes. The speci®city of the promoter cassettes has been determined using luxCDABE
reporter systems. Whole cell-biosensors containing metal-induced lux reporter systems have been incorporated into different
matrices for their later immobilisation on optic ®bres and characterised in terms of their sensitivity and storage capacity. The
second type of sensors was based on the direct interaction between metal-binding proteins and heavy metal ions. In this case,
the capacitance changes of the proteins, such as synechoccocal metallothionein (as a GST-SmtA fusion protein) and the
mercury regulatory protein, MerR, were detected in the presence of femtomolar to millimolar metal ion concentrations.
# 1999 Elsevier Science B.V. All rights reserved.
Keywords: Heavy metals; Bioavailability; Biosensors; Af®nity sensors; Capacitance measurements
1. Introduction
Most metal ions can be detected in environmental
samples using classical analytical methods such as
inductively coupled plasma atomic electron spectro-
metry (ICP/AES) or mass spectrometry (ICP/MS),
¯ow injection atomic absorption (FIAAS) or electro-
chemical methods that include ion selective electro-
des, polarography and other voltammetric electrodes.
The environmental samples need to be digested under
high temperature, pressure and acidic conditions to
free the metal ions in solution as a prerequisite for all
those methods. However, the total amount of metal
Analytica Chimica Acta 387 (1999) 235±244
*Corresponding author. Tel.: +32-14-335-112; fax: +32-14-580-
523; e-mail: [email protected]
0003-2670/99/$ ± see front matter # 1999 Elsevier Science B.V. All rights reserved.
P I I : S 0 0 0 3 - 2 6 7 0 ( 9 8 ) 0 0 7 2 5 - 9

detected after such an extraction may not always be
related to the toxicity of the samples because the
original biological availability of metal ions is not
taken into account. To quantify the biologically avail-
able fraction of metal in environmental samples,
different approaches have been followed. The ®rst
approach is based on the use of soil bacteria that
are genetically engineered so that a quanti®able signal
is produced when the bacteria are in contact with
bioavailable metal ions. A number of fundamental
aspects of the regulation of sensing elements have
been studied, and new metal-induced regulators have
been discovered and characterised. Soil bacteria able
to produce a bioluminescent signal in the presence of
speci®c metal ions have been constructed by genetic
manipulation and immobilised in solid matrices for
use with optic ®bres.
The second approach followed in this work was to
follow directly the interaction between metal ions and
speci®c puri®ed metal-binding proteins by recording
the capacitance changes on metal binding. Both tech-
niques are described below.
2. Experimental
2.1. Construction of heavy metal-induced promoter
cassettes
Two independent clones containing the copSRA
promoter region of the Alcaligenes eutrophus CH34
cop operon and the tetracycline marker (as an EcoRI
fragment from miniTn5-luxAB [1]) were constructed
in plasmid pUC18/S®I. An s®I cassette containing the
mercury responsive elements from Tn501 [2] was
constructed as follows: the E. coli merR gene and
mer promoter (PmerTPAD) of Tn501 was cloned as a
0.7 kb Xba-EcoRI fragment in pUC18/S®I and the
tetracycline resistance marker was cloned into the
HindIII site. An s®I cassette containing the chromate
responsive element from the A. eutrophus pMOL28
plasmid [3] was constructed by cloning the chr pro-
moter/operator region and the chrB gene of plasmid
pMOL28 as a 1.2 kb XbaI-PstI fragment in pUC18/
S®I together with the EcoRI fragment containing the
tetracycline resistance gene. The A. eutrophus pbrR
promoter/operator region and the partially deleted
pbrA gene were cloned as a 3.8 kb EcoRI fragment
in PUC18/S®I, and the tetracycline marker was cloned
into the HindIII site.
2.2. Immobilisation of the whole cell copper
biosensor
To immobilise the strain AE1239 in sodium algi-
nate (BDH Supplies Poole, Dorset, UK), 50 ml of
fresh bacterial culture was harvested by centrifugation
at 6000 rpm for 10 min in a JA-20 Beckman rotor. The
cell pellet was suspended in 20 ml 0.9% (w/w) NaCl,
and 20 ml 4% (w/w) sodium alginate was added to the
bacterial suspension. Next, a syringe with a needle (ID
1.2 mm) was used to create droplets added to 200 ml
0.2 M CaCl2 to form the alginate beads. The beads
were washed twice with 0.9% NaCl and kept in 0.9%
NaCl at 48C until use. To immobilise the whole cell
copper biosensor in seaplaque agarose, the same
procedure was followed but the cell pellet was sus-
pended in 9 ml 0.9% NaCl. Next, 2.22 g agarose was
dissolved in 100 ml 0.9% NaCl with heating, cooled to
308C in a water bath, and 5 ml bacterial suspension
was added. A 2 mm thick biogel was produced on
setting, and discs of 4 mm diameter were cut from
the gel. The biogel was kept at 48C in 0.9% NaCl
until use.
2.3. Bioluminescence measurements
For the copper whole cell-sensor AE1239, the
chromate whole cell-sensor AE2440, or the lead
whole cell-sensor AE2448, an overnight 25 ml LB
bacterial culture was centrifuged for 10 min at
10 000 rpm in a JA-20 rotor and resuspended in cool
cryoprotectants (1 ml/0.1 g wet wt). A volume of
150 ml cell suspension was dispensed in sterile lyo-
philisation glass vials and cooled in ice water before
being frozen at ÿ408C and lyophilised in a vacuum
chamber. The vacuum was released under nitrogen gas
and the vials were sealed. About 107±108 viable cells
were obtained after reconstitution in a reconstitution
medium (RM). This medium is similar to the Tris
medium described previously [4] but Tris buffer was
replaced by MOPS buffer and sodium phosphate by
sodium-b-glycerophosphate to minimise metal chela-
tion and precipitation. The RM medium contained
4.68 g of NaCl, 1.49 g of KCl, 1.07 g of NH4Cl,
430 mg of Na2SO4, 200 mg of MgCl2�6H20, 30 mg
236 P. Corbisier et al. / Analytica Chimica Acta 387 (1999) 235±244

of CaCl2�2H20, 294 mg of Na-b-glycerophosphate,
2 g of Na-acetate or 0.1 g of gluconate, 20 mM of
MOPS, pH 7.0 and one 1 ml of trace element solution
SL7 of Biebl and Pfennig [4] in 1 l of distilled water.
The metal salt solutions (20 ml) were added to 180 ml
cell suspension and the bioluminescence was recorded
over 5 h at 238C in a Lucy1 microtitre plate lumino-
meter.
2.4. Overexpression and purification of heavy-metal
binding proteins
The mercuric ion-binding regulatory protein, MerR
from transposon Tn501, has been puri®ed in large
amounts. The protein was overexpressed in E. coli and
an extraction protocol using sonication, salt-extrac-
tion, and liquid chromatography (LC), modi®ed from
[5], was used to purify 10 mg of MerR. The cloned
merR gene from Bacillus [6] was expressed from the
bacteriophage T7 promoter on plasmid pBS�, via
temperature induction (at 428C) of T7 RNA polymer-
ase from plasmid pGP1-2, co-transformed into the
E. coli host. After 3 h induction, the cells were
sonicated and the MerR protein puri®ed by LC af®nity
chromatography followed by size exclusion chroma-
tography. Yields were low (less than 100 mg l per
culture) but 0.6 mg puri®ed protein was produced
for immobilisation.
The fusion protein GST-SmtA, containing glu-
tathione-S-transferase linked to the synechococcal
metallothionein protein, was overexpressed in E. coli
from an expression vector pGEX3X (Pharmacia) con-
taining smtA, and puri®ed using glutathione sepharose
4B by published methods [7].
2.5. Protein immobilisation and capacitance
measurements
The fusion proteins GST-SmtA and MerR were
produced as described above and dissolved in phos-
phate buffered saline (70 mM NaCl, 1.3 mM KCl,
5 mM Na2HPO4, 0.9 mM KH2PO4, pH 7.3) contain-
ing 50% glycerol to a ®nal concentration of 1 mg/ml
protein. Thioctic acid and glutaraldehyde (GA) were
purchased from Sigma and 1-(3-dimethylaminopro-
pyl)-3ethyl-carbodiimide hydrochloride (EDC) was
obtained from Fluka. 1-Dodecanethiol and the gold
rods used for the electrodes came from Aldrich. Heavy
metal salts CuCl2�2H20, ZnCl2, HgCl2 and
Cd(NO3)2�4H20 were from Merck (Darmstadt, Ger-
many). Polyethyleneglycol diglycidyl ether
(PEGDGE) was obtained from Polysciences (USA).
All other reagents were of analytical grade.
The biosensors were prepared by immobilising
fusion proteins on the gold surface by EDC-mediated
coupling, PEGDGE entrapment or GA cooling. In all
cases 20 ml of the dissolved fusion proteins were
diluted with 480 ml 100 mM borate buffer, pH 8.75
and the solution was ®ltered through a micro-®lter
(Amicon, USA) with a molecular cut-off of 3000 D.
After ultra®ltration, the fusion protein concentration
was adjusted to 0.04 mg/ml in borate buffer. Gold
electrodes were cleaned and pretreated with thioctic
acid, as described elsewhere [8].
The biosensor was arranged as the working elec-
trode in a three-electrode system connected to a fast
potentiostat. It was placed in a ¯ow cell with a dead
volume of 10 ml which was built in-house [8]. A
platinum foil served as the auxiliary and a platinum
wire as the reference electrode. An extra reference
electrode (Ag/AgCl) was placed in the outlet stream,
as the platinum does not have a de®ned potential. The
buffer solution was pumped by a peristaltic pump with
a ¯ow rate of 0.5 ml/min through the ¯ow cell. Sam-
ples were injected into the ¯ow via a 250 ml sample
loop. The buffer was 10 mM borate pH 8.75, ®ltered
through a 0.22 mm millipore ®lter and degassed before
use.
3. Results and discussion
3.1. Development of new genetic tools
A number of genetic tools and the background
knowledge necessary for the successful production
of metal-responsive strains have been developed. Sub-
stantial progress in understanding the molecular basis
of s-54 promoters as sensors of environmental pollu-
tants has been described earlier [9,10]. Understanding
the mechanism of activation of this regulator was
important in designing the gene-based sensors, which
are based on the activation of metal-induced promoter/
operator regions. The mechanisms of metalloregula-
tion of two promoters of E. coli based on the Fur
protein has also been dissected [11]. Finally, the
P. Corbisier et al. / Analytica Chimica Acta 387 (1999) 235±244 237
exploitation of the outer membrane protein LamB as
an anchor of heterologous metal-binding peptides
such as the yeast (CUP1) and mammalian (HMT-
1A) metallothioneins on the surface of Gram-negative
bacteria has been explored [12].
A. eutrophus was chosen as a reference organism
due to its ability to survive in harsh environments [13].
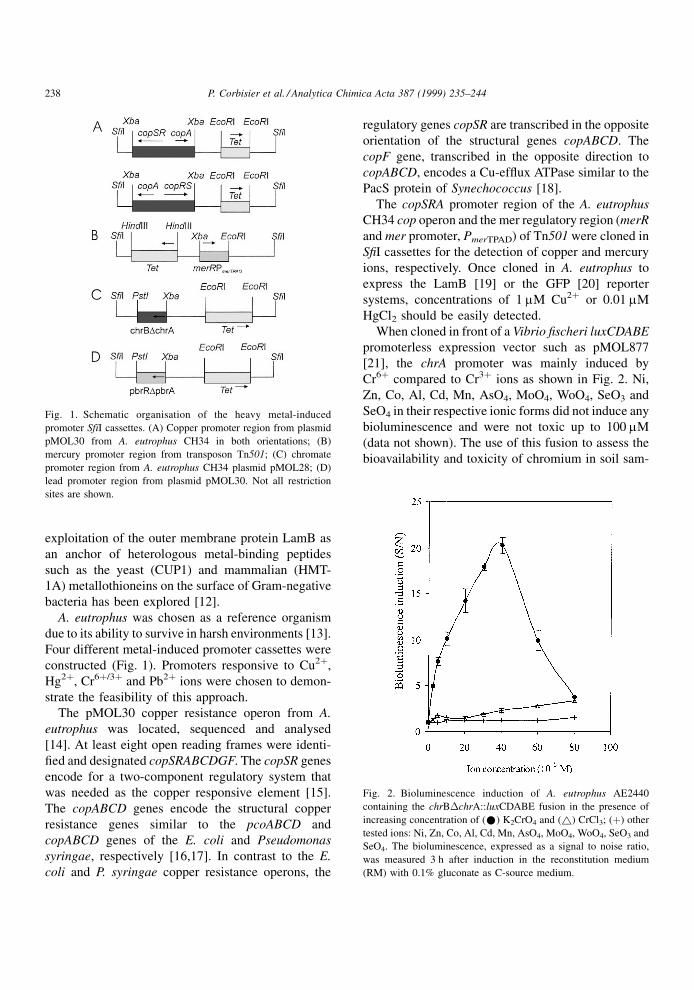
Four different metal-induced promoter cassettes were
constructed (Fig. 1). Promoters responsive to Cu2�,
Hg2�, Cr6�/3� and Pb2� ions were chosen to demon-
strate the feasibility of this approach.
The pMOL30 copper resistance operon from A.
eutrophus was located, sequenced and analysed
[14]. At least eight open reading frames were identi-
®ed and designated copSRABCDGF. The copSR genes
encode for a two-component regulatory system that
was needed as the copper responsive element [15].
The copABCD genes encode the structural copper
resistance genes similar to the pcoABCD and
copABCD genes of the E. coli and Pseudomonas
syringae, respectively [16,17]. In contrast to the E.
coli and P. syringae copper resistance operons, the
regulatory genes copSR are transcribed in the opposite
orientation of the structural genes copABCD. The
copF gene, transcribed in the opposite direction to
copABCD, encodes a Cu-ef¯ux ATPase similar to the
PacS protein of Synechococcus [18].
The copSRA promoter region of the A. eutrophus
CH34 cop operon and the mer regulatory region (merR
and mer promoter, PmerTPAD) of Tn501 were cloned in
S®I cassettes for the detection of copper and mercury
ions, respectively. Once cloned in A. eutrophus to
express the LamB [19] or the GFP [20] reporter
systems, concentrations of 1 mM Cu2� or 0.01 mM
HgCl2 should be easily detected.
When cloned in front of a Vibrio ®scheri luxCDABE
promoterless expression vector such as pMOL877
[21], the chrA promoter was mainly induced by
Cr6� compared to Cr3� ions as shown in Fig. 2. Ni,
Zn, Co, Al, Cd, Mn, AsO4, MoO4, WoO4, SeO3 and
SeO4 in their respective ionic forms did not induce any
bioluminescence and were not toxic up to 100 mM
(data not shown). The use of this fusion to assess the
bioavailability and toxicity of chromium in soil sam-
Fig. 1. Schematic organisation of the heavy metal-induced
promoter SfiI cassettes. (A) Copper promoter region from plasmid
pMOL30 from A. eutrophus CH34 in both orientations; (B)
mercury promoter region from transposon Tn501; (C) chromate
promoter region from A. eutrophus CH34 plasmid pMOL28; (D)
lead promoter region from plasmid pMOL30. Not all restriction
sites are shown.
Fig. 2. Bioluminescence induction of A. eutrophus AE2440
containing the chrB�chrA::luxCDABE fusion in the presence of
increasing concentration of (*) K2CrO4 and (~) CrCl3; (�) other
tested ions: Ni, Zn, Co, Al, Cd, Mn, AsO4, MoO4, WoO4, SeO3 and
SeO4. The bioluminescence, expressed as a signal to noise ratio,
was measured 3 h after induction in the reconstitution medium
(RM) with 0.1% gluconate as C-source medium.
238 P. Corbisier et al. / Analytica Chimica Acta 387 (1999) 235±244
ples has large potentials since it can easily differenti-
ate the trivalent from hexavalent chromium.
The lead resistance operon of A. eutrophus
pMOL30 plasmid has also been cloned and
sequenced. It contains two genes, pbrRA, required
for regulation and lead resistance, respectively. The
pbrR regulator was very similar to the MerR protein
(regulator of the mercury resistance operon). The
PbrA protein has all characteristics of an ef¯ux
ATPase. When cloned in into pMOL877, the pbrR
promoter appeared to be speci®cally induced by lead
ions as shown in Fig. 3. Those four different metal-
induced promoter cassettes can now be further
assembled in the LamB or GFP reporter systems.
The metal-induced bacterial biosensors [22] that
were already available have been further immobilised
in different solid matrices.
3.2. Immobilisation of the bacterial sensors
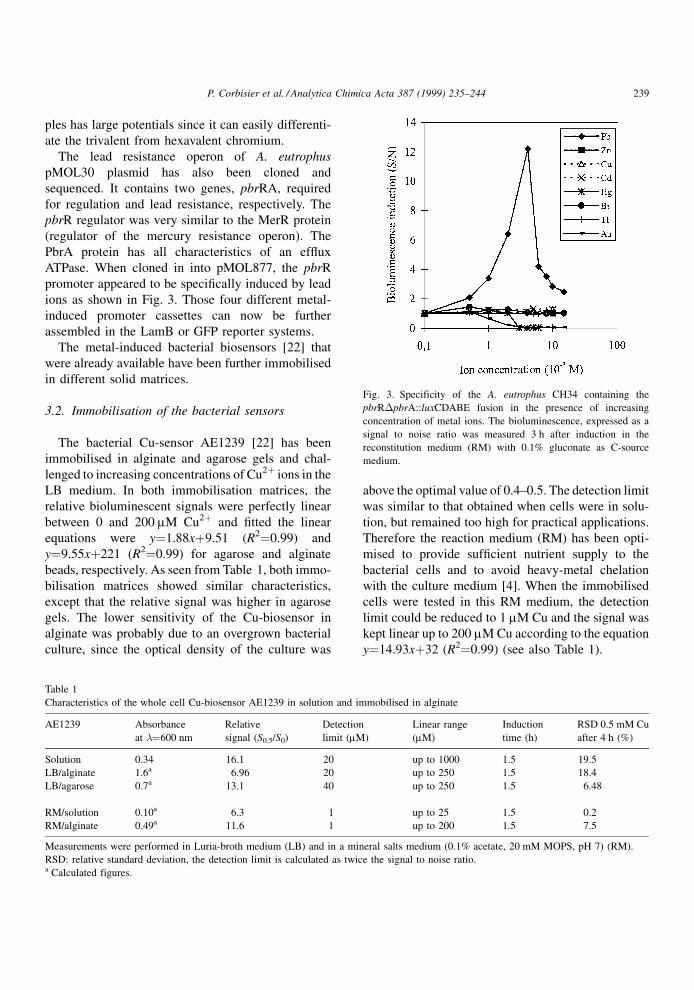
The bacterial Cu-sensor AE1239 [22] has been
immobilised in alginate and agarose gels and chal-
lenged to increasing concentrations of Cu2� ions in the
LB medium. In both immobilisation matrices, the
relative bioluminescent signals were perfectly linear
between 0 and 200 mM Cu2� and ®tted the linear
equations were y�1.88x�9.51 (R2�0.99) and
y�9.55x�221 (R2�0.99) for agarose and alginate
beads, respectively. As seen from Table 1, both immo-
bilisation matrices showed similar characteristics,
except that the relative signal was higher in agarose
gels. The lower sensitivity of the Cu-biosensor in
alginate was probably due to an overgrown bacterial
culture, since the optical density of the culture was
above the optimal value of 0.4±0.5. The detection limit
was similar to that obtained when cells were in solu-
tion, but remained too high for practical applications.
Therefore the reaction medium (RM) has been opti-
mised to provide suf®cient nutrient supply to the
bacterial cells and to avoid heavy-metal chelation
with the culture medium [4]. When the immobilised
cells were tested in this RM medium, the detection
limit could be reduced to 1 mM Cu and the signal was
kept linear up to 200 mM Cu according to the equation
y�14.93x�32 (R2�0.99) (see also Table 1).
Fig. 3. Specificity of the A. eutrophus CH34 containing the
pbrR�pbrA::luxCDABE fusion in the presence of increasing
concentration of metal ions. The bioluminescence, expressed as a
signal to noise ratio was measured 3 h after induction in the
reconstitution medium (RM) with 0.1% gluconate as C-source
medium.
Table 1
Characteristics of the whole cell Cu-biosensor AE1239 in solution and immobilised in alginate
AE1239 Absorbance
at ��600 nm
Relative
signal (S0.5/S0)
Detection
limit (mM)
Linear range
(mM)
Induction
time (h)
RSD 0.5 mM Cu
after 4 h (%)
Solution 0.34 16.1 20 up to 1000 1.5 19.5
LB/alginate 1.6a 6.96 20 up to 250 1.5 18.4
LB/agarose 0.7a 13.1 40 up to 250 1.5 6.48
RM/solution 0.10a 6.3 1 up to 25 1.5 0.2
RM/alginate 0.49a 11.6 1 up to 200 1.5 7.5
Measurements were performed in Luria-broth medium (LB) and in a mineral salts medium (0.1% acetate, 20 mM MOPS, pH 7) (RM).
RSD: relative standard deviation, the detection limit is calculated as twice the signal to noise ratio.a Calculated figures.
P. Corbisier et al. / Analytica Chimica Acta 387 (1999) 235±244 239
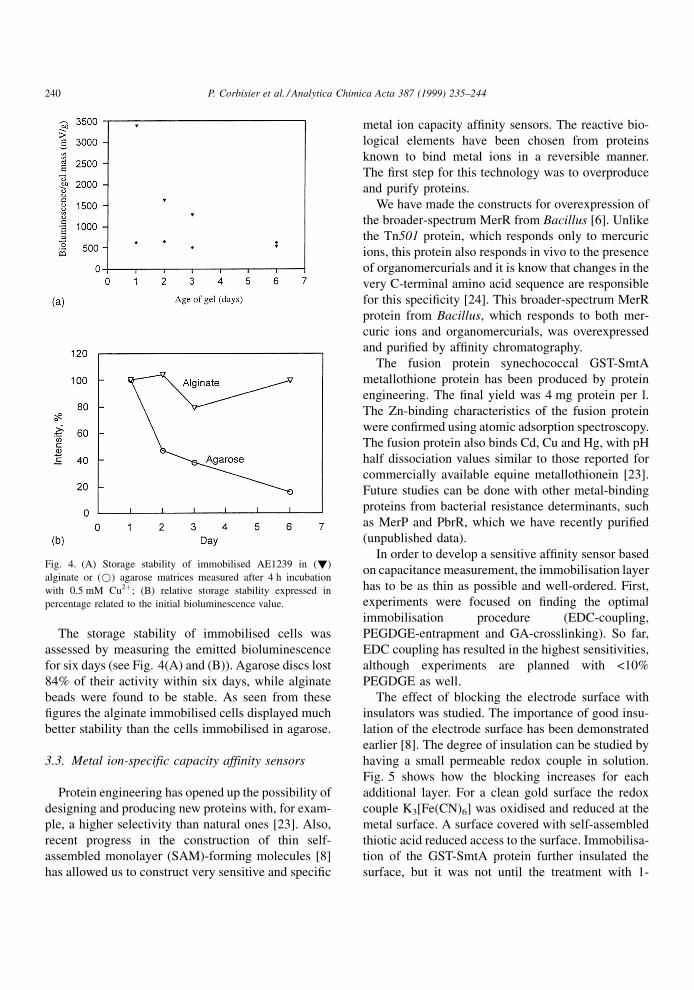
The storage stability of immobilised cells was
assessed by measuring the emitted bioluminescence
for six days (see Fig. 4(A) and (B)). Agarose discs lost
84% of their activity within six days, while alginate
beads were found to be stable. As seen from these
®gures the alginate immobilised cells displayed much
better stability than the cells immobilised in agarose.
3.3. Metal ion-specific capacity affinity sensors
Protein engineering has opened up the possibility of
designing and producing new proteins with, for exam-
ple, a higher selectivity than natural ones [23]. Also,
recent progress in the construction of thin self-
assembled monolayer (SAM)-forming molecules [8]
has allowed us to construct very sensitive and speci®c
metal ion capacity af®nity sensors. The reactive bio-
logical elements have been chosen from proteins
known to bind metal ions in a reversible manner.
The ®rst step for this technology was to overproduce
and purify proteins.
We have made the constructs for overexpression of
the broader-spectrum MerR from Bacillus [6]. Unlike
the Tn501 protein, which responds only to mercuric
ions, this protein also responds in vivo to the presence
of organomercurials and it is know that changes in the
very C-terminal amino acid sequence are responsible
for this speci®city [24]. This broader-spectrum MerR
protein from Bacillus, which responds to both mer-
curic ions and organomercurials, was overexpressed
and puri®ed by af®nity chromatography.
The fusion protein synechococcal GST-SmtA
metallothione protein has been produced by protein
engineering. The ®nal yield was 4 mg protein per l.
The Zn-binding characteristics of the fusion protein
were con®rmed using atomic adsorption spectroscopy.
The fusion protein also binds Cd, Cu and Hg, with pH
half dissociation values similar to those reported for
commercially available equine metallothionein [23].
Future studies can be done with other metal-binding
proteins from bacterial resistance determinants, such
as MerP and PbrR, which we have recently puri®ed
(unpublished data).
In order to develop a sensitive af®nity sensor based
on capacitance measurement, the immobilisation layer
has to be as thin as possible and well-ordered. First,
experiments were focused on ®nding the optimal
immobilisation procedure (EDC-coupling,
PEGDGE-entrapment and GA-crosslinking). So far,
EDC coupling has resulted in the highest sensitivities,
although experiments are planned with <10%
PEGDGE as well.
The effect of blocking the electrode surface with
insulators was studied. The importance of good insu-
lation of the electrode surface has been demonstrated
earlier [8]. The degree of insulation can be studied by
having a small permeable redox couple in solution.
Fig. 5 shows how the blocking increases for each
additional layer. For a clean gold surface the redox
couple K3[Fe(CN)6] was oxidised and reduced at the
metal surface. A surface covered with self-assembled
thiotic acid reduced access to the surface. Immobilisa-
tion of the GST-SmtA protein further insulated the
surface, but it was not until the treatment with 1-
Fig. 4. (A) Storage stability of immobilised AE1239 in (!)
alginate or (*) agarose matrices measured after 4 h incubation
with 0.5 mM Cu2�; (B) relative storage stability expressed in
percentage related to the initial bioluminescence value.
240 P. Corbisier et al. / Analytica Chimica Acta 387 (1999) 235±244
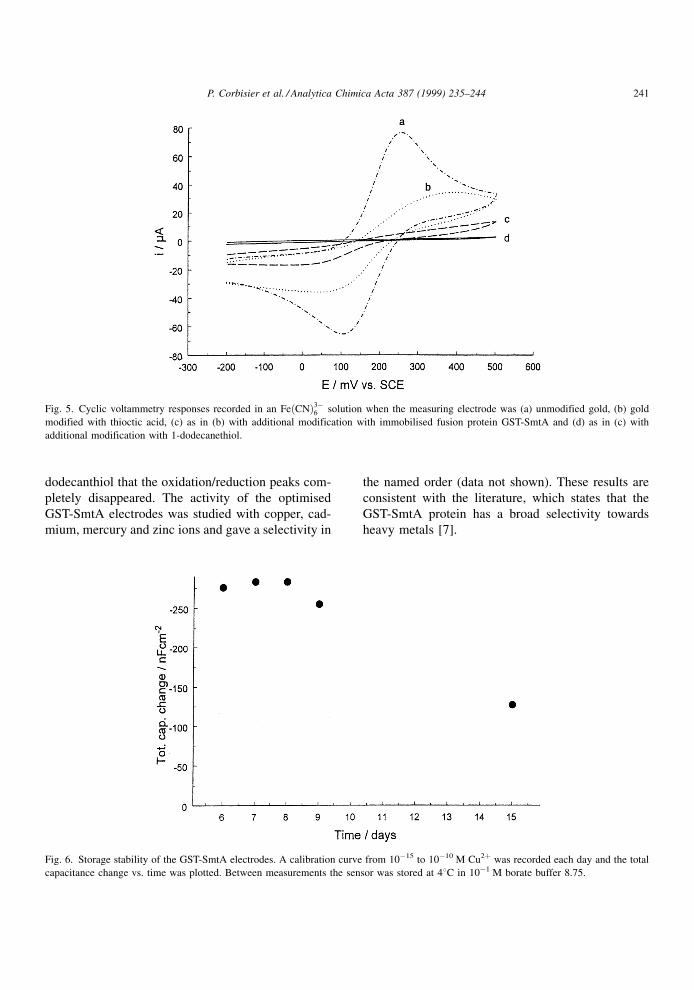
dodecanthiol that the oxidation/reduction peaks com-
pletely disappeared. The activity of the optimised
GST-SmtA electrodes was studied with copper, cad-
mium, mercury and zinc ions and gave a selectivity in
the named order (data not shown). These results are
consistent with the literature, which states that the
GST-SmtA protein has a broad selectivity towards
heavy metals [7].
Fig. 5. Cyclic voltammetry responses recorded in an Fe�CN�3ÿ6 solution when the measuring electrode was (a) unmodified gold, (b) gold
modified with thioctic acid, (c) as in (b) with additional modification with immobilised fusion protein GST-SmtA and (d) as in (c) with
additional modification with 1-dodecanethiol.
Fig. 6. Storage stability of the GST-SmtA electrodes. A calibration curve from 10ÿ15 to 10ÿ10 M Cu2� was recorded each day and the total
capacitance change vs. time was plotted. Between measurements the sensor was stored at 48C in 10ÿ1 M borate buffer 8.75.
P. Corbisier et al. / Analytica Chimica Acta 387 (1999) 235±244 241
The GST-SmtA electrodes could be regenerated by
injection of 1 mM EDTA. It was found that if EDTA
was injected just before storage, the biosensor lost
activity on overnight storage. However, if the elec-
trode was stored in the presence of heavy metal ions
and regenerated immediately before taking measure-
ments on the following day, no activity loss was
observed. This was probably due to the protein being
protected from denaturation (possibly oxidation) by
binding of the heavy metal to cysteine sulphydryl
groups. The stability of these electrodes over 16 days
is shown in Fig. 6.
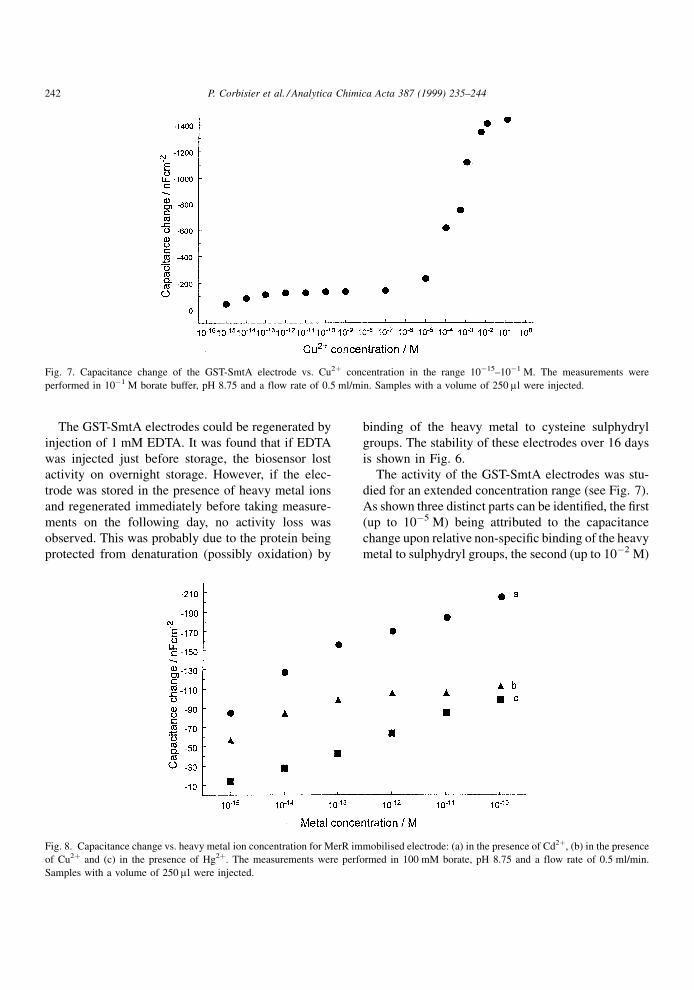
The activity of the GST-SmtA electrodes was stu-
died for an extended concentration range (see Fig. 7).
As shown three distinct parts can be identi®ed, the ®rst
(up to 10ÿ5 M) being attributed to the capacitance
change upon relative non-speci®c binding of the heavy
metal to sulphydryl groups, the second (up to 10ÿ2 M)
Fig. 7. Capacitance change of the GST-SmtA electrode vs. Cu2� concentration in the range 10ÿ15±10ÿ1 M. The measurements were
performed in 10ÿ1 M borate buffer, pH 8.75 and a flow rate of 0.5 ml/min. Samples with a volume of 250 ml were injected.
Fig. 8. Capacitance change vs. heavy metal ion concentration for MerR immobilised electrode: (a) in the presence of Cd2�, (b) in the presence
of Cu2� and (c) in the presence of Hg2�. The measurements were performed in 100 mM borate, pH 8.75 and a flow rate of 0.5 ml/min.
Samples with a volume of 250 ml were injected.
242 P. Corbisier et al. / Analytica Chimica Acta 387 (1999) 235±244

probably due to the formation of a closed metallothio-
nein `cage' containing the metal ions, and the third due
to saturation (levelling off). Preliminary experiments
were also carried out with an MerR electrode, immo-
bilised by the same method found to be optimal for
GST-SmtA protein. This protein is known to be highly
speci®c towards Hg2� ions in vivo and in vitro [23].
The protein was immobilised on the sensor surface by
the EDC method. The sensitivity was studied for the
three different heavy metals; mercury, copper and
cadmium. It was found, as expected, that the sensi-
tivity was the highest for mercury, and that this
electrode showed higher selectivity than the GST-
SmtA electrode at low metal ion concentrations
(Fig. 8).
4. Conclusions
This work has shown the feasibility of technologies
for the detection of heavy metal ions based on whole
cell-biosensors and on protein-based sensors. A num-
ber of metal-induced promoter regions have been
identi®ed and arranged in cassettes that can be easily
used to activated reporter system such as the lux or
GFP reporter genes or the expression of outer mem-
brane epitopes that can be easily detected by immu-
nochemistry. The high speci®city of such an induced
gene expression has been shown for the lead and
chromate ions. Preliminary results for immobilised
whole cell-sensors have been obtained and demon-
strated the applicability of this technology.
Af®nity sensors based on proteins were also demon-
strated as suitable for monitoring heavy metal ions at
trace levels. The metal ion-speci®c capacitance sen-
sors have an exceptional sensitivity and a wide oper-
ating range. They are also versatile systems because
different kinds of recognition elements can be immo-
bilised directly in a self-assembling monolayer com-
pletely covering the surface of the measuring noble
metal electrode. The electrode then becomes selective
to those metal ions in the solution that show af®nity to
the recognition element on the surface. Compared to
previously described electrochemical sensors, the pro-
tein-based sensor shows many orders of magnitude
better sensitivity [25].
The whole cell-sensor and the protein-based sensor
now need to be tested on real environmental samples.
The main advantage of the whole cell-sensors will
remain their ability to react only to the bioavailable
fraction of metal ions, whereas the protein-based
sensor's potential application remains in its high
sensitivity towards metals ions.
Acknowledgements
We thank Professor Nigel Robinson (Newcastle) for
supplying the pGEX3X-smtA fusion plasmid. NB
thanks Kenneth J. Jakeman for technical assistance.
PC thanks S. Leth, S. Maltoni and A. Bossus for
technical assistance and L. Diels and M. Mergeay
for the constructive discussions. This work was sup-
ported by the European Commission as part of the
contract ENV4-CT95-0141.
References
[1] M. Herrero, V. de Lorenzo, K.N. Timmis, J. Bacteriol. 172
(1990) 6557.
[2] P.A. Lund, N.L. Brown, J. Mol. Biol. 205 (1989) 343.
[3] A. Nies, D.H. Nies, S. Silver, J. Biol. Chem. 265 (1990) 5648.
[4] P. Corbisier, E. Thiry, A. Masolijn, L. Diels, in: A.K.
Campbell, L.J. Kricka, P.E. Stanley (Eds.), Bioluminescence
and Chemiluminescence: Fundamentals and Applied Aspects,
Wiley, New York, 1994, pp. 151±155.
[5] J. Parkhill, A.Z. Ansari, J.G. Wright, N.L. Brown, T.V.
O'Halloran, EMBO J. 12 (1993) 413.
[6] J.D. Helmann, Y. Wang, I. Mahler, C.T. Walsh, J. Bacteriol.
171 (1989) 222.
[7] A.M. Tommey, J. Shi, W.P. Lindsay, P.E. Urwin, N.J.
Robinson, FEBS Lett. 292 (1991) 48.
[8] C. Berggren, G. Johansson, Anal. Chem. 69 (1997) 3651.
[9] J. PeÂrez-MartõÂn, V. de Lorenzo, J. Mol. Biol. 258 (1996) 562.
[10] J. PeÂrez-MartõÂn, V. de Lorenzo, J. Mol. Biol. 258 (1996) 575.
[11] L. Escolar, V. de Lorenzo, J. PeÂrez-MartõÂn, Mol. Microbiol.
26 (1997) 799.
[12] C. Sousa, A. Cebolla, V. de Lorenzo, Nature Biotechnol. 14
(1996) 1017.
[13] M. Mergeay, D. Nies, H.G. Schlegel, J. Gerits, P. Charles, F.J.
VanGijsegem, Bacteriology 162 (1985) 328.
[14] B. Borremans, A. Provoost, P. Corbisier, M. Mergeay, J.L.
Hobman, N.L. Brown, D. van der Lelie, Proceedings of the VI
International Congress on Pseudomonas: Molecular Biology
and Biotechnology, 1997, p. IX.
[15] D.A. Rouch, N.L. Brown, Microbiology 143 (1997) 1191.
[16] N.L. Brown, S.R. Barrett, J. Camakaris, B.T.O. Lee, D.A.
Rouch, Mol. Microbiol. 17 (1995) 1153.
[17] S.D. Molls, C.-K. Lim, D.A. Cooksey, Mol. Gen. Genet. 244
(1994) 341.
P. Corbisier et al. / Analytica Chimica Acta 387 (1999) 235±244 243

[18] K. Kanamaru, S. Kashiwagi, T. Mizuno, Mol. Microbiol. 13
(1994) 369.
[19] A. Charbit, J. Ronco, V. Michel, C. Werts, M. Hofnung, J.
Bacteriol. 173 (1991) 262.
[20] M. Chalfie, Y. Tu, G. Euskircher, W. Ward, D.C. Prasher,
Science 263 (1994) 802.
[21] D. van der Lelie, T. Schwuchow, U. Schidetzky, S. Wuertz,
W. Baeyens, M. Mergeay, D. Nies, Mol. Microbiol. 23 (1997)
493±503.
[22] P. Corbisier, E. Thiry, L. Diels, Environ. Toxicol. Water
Quality 11 (1996) 171.
[23] B. Frantz, T.V. O'Halloran, Biochemistry 29 (1990) 4747.
[24] G. Nucifora, L. Chu, S. Silver, T.K. Misra, J. Bacteriol. 171
(1989) 4241.
[25] Metal Ion Specific Affinity Sensors, Swedish Patent no.
9703315-3.
244 P. Corbisier et al. / Analytica Chimica Acta 387 (1999) 235±244
Copyright © 2022 FDOKUMEN




















