
Correlation of intrusive growth of cambial initials to rearrangement of rays in the vascular cambium

Annals of Botany 84 : 621–632, 1999Article No. anbo.1999.0957, available online at http:}}www.idealibrary.com on
Variations in the Lengths of Fusiform Cambial Cells and Vessel Elements
in Kalopanax pictus
P. KITIN*, R. FUNADA*† , Y. SANO*, H. BEECKMAN‡ and J. OHTANI*
*Department of Forest Science, Faculty of Agriculture, Hokkaido Uni�ersity, Sapporo 060-8589, Japan and
‡Laboratory for Wood Biology and Xylarium, Royal Museum for Central Africa, Ter�uren B-3080, Belgium
Received: 30 April 1999 Returned for revision: 2 June 1999 Accepted: 9 July 1999
Samples of a mature specimen of Kalopanax pictus, a ring-porous hardwood, were studied to compare the respectivelengths of fusiform cambial cells and vessel elements in the stem. The lengths of dormant and reactivated fusiformcambial cells were measured with a confocal laser scanning microscope in tissue that had been macerated by digestionwith pectinase and in thick tangential sections. The lengths of early wood and late wood vessel elements weremeasured in tissues that had been macerated by Franklin’s method. The vessel elements and fusiform cambial cellsvaried considerably in length within individual samples. The mean length of early wood vessel elements correspondedto that of fusiform cells in the dormant cambium but not in the reactivated cambium. Significant differences wereobserved between the mean lengths of dormant and reactivated fusiform cambial cells, between those of reactivatedfusiform cambial cells and early wood vessel elements, between those of reactivated fusiform cambial cells and latewood vessel elements, and between those of early wood and late wood vessel elements. The frequency distributionsof lengths of cambial cells were bimodal and differed from those of vessel elements, which more closely resembled anormal distribution. The proportion of shorter lengths was higher in the reactivated cambium than in the dormantcambium, the early wood and the late wood vessel elements. Our results do not suppot the hypothesis that the lengthsof early wood vessel elements in ring-porous hardwoods change during differentiation. The similar ranges of recordedlengths suggest that short and long vessel elements might be derived directly from short and long cambial cells,respectively. # 1999 Annals of Botany Company
Key words : Kalopanax pictus, cambium, vessel element, confocal laser scanning microscopy, maceration, cell length.
INTRODUCTION
Information on the lengths of cambial cells has beenrecognized as important in different aspects of botanicalresearch, in particular in plant taxonomy and phylogeny, aswell as in developmental and ecological anatomy (Metcalfeand Chalk, 1983). The dimensions of fusiform cambial cellsvary among and within species and they also varypositionally and seasonally within individual trees (Philip-son, Ward and Butterfield, 1971; Iqbal and Ghouse, 1990;Larson, 1994). Bailey (1920), who studied 13 species ofgymnosperm and 54 dicotyledonous species from 14families, provided extensive data on lengths of fusiformcambial cells. Extensive studies of tropical and subtropicalwoods have been performed in India (see Table andreferences in Larson, 1994). However, as Larson (1994)noted, the accumulated data on lengths of cambial cellswere based on a variety of sampling procedures and samplesizes. A universal method and a justified sample size forstudies of the lengths of cambial cells remain to beestablished.
In general, it is difficult to measure directly the lengths offusiform cambial cells. It is easier to measure the vessel
† For correspondence. Fax 81-11-736-1791, e-mail funada!for.agr.hokudai.ac.jp
elements and Bailey (1920) suggested that the lengths ofvessel elements might indicate the lengths of fusiformcambial cells. Since Bailey’s study, there have been nofurther detailed studies, to our knowledge, of the re-lationship between lengths of vessel elements and those offusiform cambial cells. However, some reports have in-dicated that the assumed approximate similarity in lengthbetween vessel elements and fusiform cambial cells shouldnot be accepted without further analysis. Lengths of vesselelements have been found to be equivalent to those offusiform cambial cells only in species with storied cambiumor diffuse-porous wood (Chalk and Chattaway, 1934, 1935;Butterfield, 1973; Rao, Rajput and Srinivas, 1996). How-ever, in ring-porous woods and in many tropical woods, themean length of vessel elements has usually been found to beshorter than that of fusiform cambial cells (Chalk andChattaway, 1934, 1935; Swamy, Parameswaran and Govin-darajulu, 1960; Su$ ss, 1967; Anand et al., 1981). Judgingfrom the lengths of axial parenchyma strands in wood,Chalk and Chattaway (1935) estimated that only the lengthsof late wood vessel elements in ring-porous woods could beconsidered equivalent to those of cambial initials. The widevessel elements of the early wood were found to be 13 to47% shorter than the narrow vessel elements of the latewood. Various hypotheses have been proposed to explaindifferences in length between vessel elements and fusiformcambial cells. Some authors have attributed the shortening
0305-7364}99}11062112 $30.00}0 # 1999 Annals of Botany Company

622 Kitin et al.—Cambial Cell Length in Kalopanax pictus
of vessel elements to a change in the position of the end wallduring differentiation, while others have suggested that theshorter vessel elements might be due to shortening of thecambial initials through anticlinal divisions (for references,see Larson, 1994).
There are two basic methods for determining thedimensions of cells : measurements can be made ontangential sections or on macerated tissue. As Larson (1994)pointed out, a difference between the mean length of vesselelements and that of fusiform cambial cells might be theresult of comparing measurements of cells in maceratedxylem with measurements of cells in tangential sections ofthe cambium. It is easier to measure isolated cells than tomeasure cells in intact tissue, and it is accepted as a standardprocedure that vessel elements are measured in maceratedsamples because the tips are more visible after maceration.However, in samples of cambium, the maceration oftencauses breakage, shrinkage or contraction of cells. Inaddition, the cambium is a thin layer and, therefore, it isdifficult to obtain pure macerated cambial cells withoutcontamination by adjacent tissues. We have found noreports that include lengths or photographs of maceratedcambial cells. The reported lengths of fusiform cambial cellsin dicotyledonous species are all based on measurements ofcells in tangential sections (Bailey, 1920; Butterfield, 1973;Anand et al., 1981; Iqbal and Ghouse, 1985; Ajmal andIqbal, 1987; Siddiqi, 1991; Rao et al., 1996) or on indirectestimations made from cambial derivatives (Chalk andChattaway, 1935; Su% ss, 1967; Cumbie, 1967, 1983). Themethod used for measurements of cells on tangential sectionsis associated with certain problems. The fusiform cambialcells are several hundreds or thousands of micrometres longand only a few micrometres wide. Thus, the full length of afusiform cambial cell in a tangential section often cannot beviewed on one field of view under a conventional lightmicroscope.
Another reason for the difference between mean lengthsof cambial cells and vessel elements might be that seasonalvariations occur in the dimensions of fusiform cambial cells(Bannan, 1967, 1970; Iqbal and Ghouse, 1985; Ajmal andIqbal, 1987; Siddiqi, 1991). To date, annual fluctuations inlengths of cambial cells have not been taken into con-sideration in comparisons of mean lengths of fusiformcambial cells and vessel elements.
The present study was designed to compare the lengths offusiform cambial cells with those of vessel elements in thestem of Kalopanax pictus Nakai (Araliaceae), a ring-porouswood species. The vessel elements in the secondary xylem ofK. pictus exhibit wide variations in lengths and, therefore,we tried to determine whether corresponding variations inlengths of fusiform cambial cells might also occur. Weattempted to evaluate the hypothesis that the length ofvessel elements can be used to estimate the length offusiform cambial cells. We also attempted to obtain reliabledata on cell length by improving the method used formeasurements. The lengths of cambial cells were determinedboth in macerated tissue and on tangential sections. Weused confocal laser scanning microscopy for serial opticalsectioning of the cambial zone. This method allowed us tostudy relatively thick tangential sections (60–80 µm) and to
determine the precise lengths of fusiform cambial cells onimages of serial optical sections.
MATERIALS AND METHODS
Sampling and selection of material for obser�ation
A Kalopanax pictus tree (diameter at breast height, 76 cm),growing on the campus of Hokkaido University, Sapporo,was used for the study. Small blocks containing cambiumand the adjacent phloem and xylem were taken from thestem at breast height in late autumn (7 Nov. 1996) and earlyspring (3 Apr. 1997). They were fixed in FAA (50%ethanol :acetic acid:formaldehyde at 18:1:1, v}v) or in anaqueous 4% solution of glutaraldehyde. Specimens thathad been fixed in glutaraldehyde were post-fixed in 1%osmium tetroxide and embedded in epoxy resin. Semi-thintransverse sections (1 µm) were cut with a glass knife usingan ultramicrotome (Ultracut ; Reichert-Jung, Vienna,Austria) and were then double stained with safranin andgentian violet in a 1% aqueous solution. The cambium wasobserved under a light microscope (BH-2; Olympus, Tokyo,Japan) in order to identify stages of activity on the twosampling dates.
We attempted to ensure that measurements were madewithin cambial zones composed of cambial initials andphloem and xylem mother cells, and we tried to excludedifferentiating phloem and xylem elements as far as possible.To define the cambial zone morphologically, we embeddedsamples that had been fixed in FAA in celloidin (Sass, 1951)and cut serial tangential and radial sections on a slidingmicrotome. The sections were stained with gentian violetand examined under the light microscope. The term ‘cambialcells ’ in this study refers both to cambial initials and toxylem and phloem mother cells at the early stages ofdevelopment, before any evidence of enlargement andformation of the secondary wall can be detected.
We examined the morphology of vessel elements intangential and radial sections by conventional light mi-croscopy and in resin casts prepared as described by Fujii(1993). The resin casts of the secondary xylem were fixed onspecimen stubs with carbon paint, coated with carbon andgold by vacuum evaporation, and observed under a fieldemission-scanning electron microscope (FE-SEM; JSM6301F; JEOL, Tokyo, Japan) operated at an acceleratingvoltage of 2 kV.
Preparation and measurements of macerated material
Samples that had been fixed in FAA were macerated bydigestion with pectinase (Pectinase from mould, 0±01 Umg−" ; Fluka, Buchs, Switzerland). The powdered pectinasewas dissolved in ice-cold So% rensen phosphate buffer (pH 6±5)to prepare a stock solution of 50 mg ml−". The sampleblocks were washed in tap water and then cut in thelongitudinal and radial direction with a razor blade to yieldslabs that were 2–3 mm thick and 2 cm long and containedcambium and the adjacent xylem and phloem. These slabswere treated with pectinase at 2 mg ml−" in an incubator at35 °C for 2 h and then shaken for 5 h on a laboratory shaker

Kitin et al.—Cambial Cell Length in Kalopanax pictus 623
F. 1. Transverse sections showing cambium and the adjacent xylemand phloem in the stem of Kalopanax pictus. A, Dormant cambium (7Nov. 1996). No division plates are visible in the cambial cells and thecells have denser cytoplasm than that in the phloem elements. B,Reactivated cambium (3 Apr. 1997). A vessel mother cell (asterisk) islocated in the second layer of cambial cells next to the xylem. Divisionplates are visible (arrows) in the reactivated cambial cells. Bar¯ 50 µm.xy, Xylem; ca, cambium; dph, undifferentiated phloem elements ;
ph, phloem.
at room temperature. The reaction was stopped by replacingthe solution of enzyme with a mixture of 10% acetic acidand 25% ethanol (1:1, v}v) and the samples were storedtemporarily in a refrigerator prior to further preparation forobservation.
Since the cambium consists of a very thin layer of cells, itcan easily become mixed with the adjacent phloem and, inparticular, with undifferentiated phloem elements in macer-ated samples. Therefore, appropriate isolation of thecambium is important in the preparation of maceratedsamples. We achieved this goal using the following simpleprocedure. First, the tissue blocks were treated with themaceration solution as described above. Then, under a
stereoscopic microscope, the phloem was gently separatedfrom the xylem with dissecting needles. Forcing the phloemapart resulted in tangential splitting of the cambium, afterwhich several layers of cambial cells remained attached tothe xylem. To identify the cells that remained on the xylemside, we processed some specimens for scanning electronmicroscopy. After removal of the phloem, the blocks weredehydrated in a graded ethanol series, rinsed in isoamylacetate, and dried with carbon dioxide in a critical-pointdryer (HCP-2; Hitachi, Tokyo, Japan). Then the blockswere mounted on specimen stubs, coated with carbon andgold by vacuum evaporation and observed with the FE-SEM at an accelerating voltage of 2 to 3 kV. Observationswith the FE-SEM revealed that the samples contained onlyxylem and one to four layers of cambial cells at the xylemboundary (data not shown). The macerated cambial cellswere scraped gently with a dissecting needle from theexposed cambial zone onto a slide glass and mounted in amixture of equal volumes of water and glycerin. Thus, thesamples on slides from the pectinase-macerated specimenscontained only cambial cells from cell layers at the xylemboundary.
Vessel elements were macerated by the method describedoriginally by Franklin (1945). Portions of early wood andlate wood were separately soaked in a mixture of equalvolumes of glacial acetic acid and hydrogen peroxide andheated at 60 °C for 1 or 2 d.
The lengths of macerated cambial cells and vessel elementswere measured on transmitted light images obtained withNomarski optics of a confocal laser scanning microscope(LSM-310; Carl Zeiss, Oberkochen, Germany). The fulllengths of vessel elements were measured, including thetails. Only straight parts of cells were measured. Thus, thelengths of cambial cells were recorded in segments whensuch cells were curved or banded. Cells, which werepresumably precursors to axial parenchyma strands, werenot measured.
Measurements of cell length on tangential sections byconfocal laser scanning microscopy (CLSM )
FAA-fixed materials were frozen on a freezing stage(MA-101; Komatsu Electronics, Tokyo, Japan), which wasattached to a sliding microtome, and thick tangentialsections (60–80 µm) that included portions of cambium andxylem were cut in a frozen state to prevent deformation ofthe thin-walled cambial cells during sectioning. They werestained with an aqueous solution of acridine orange (1 mgml−") and then washed in phosphate-buffered saline (PBS)six times for 5 min each, and mounted in 50% glycerin inPBS on glass slides. Excitation from a helium neon ion laser(wavelength, 543 nm) was used for observations by CLSM.Consecutive confocal images of tangential sections wereobtained at 2 µm intervals starting from the cambium andcontinuing as far as the xylem. The images were storeddigitally on the computer of the CLSM system. The lengthof each fusiform cambial cell was measured as the distancebetween the cell tips using the image-analysis software of theCLSM system. In some cases, both tips of a single cell werenot visible on a single confocal image but their positions

624 Kitin et al.—Cambial Cell Length in Kalopanax pictus
could be determined from a series of confocal sections. Thefarthest ends of each cell, which could be observed onseveral consecutive confocal images, were taken as the celltips. Only the lengths of fusiform cambial cells located closeto the xylem were measured. Images were printed with adigital colour printer (UP-D8000; Sony, Tokyo, Japan) asdescribed previously (Furusawa et al., 1998).
Statistical methods
Differences between samples and between methods ofmeasurement were examined by one-way analysis ofvariance (ANOVA) with cell length as the dependentvariable. To homogenize variances, the data were trans-formed by taking square roots. The Bonferroni method wasused to identify post hoc significant differences betweenpairs of means. All calculations were performed with theSYSTAT 7.0.1 software package (SYSTAT 7.0.1; SPSSInc. ; Chicago, USA).
RESULTS
Stage of cambial acti�ity and identification of fusiformcambial cells
The cambium was dormant on 7 November and it consistedof ten to 12 rows of cells with a total radial width of about
F. 2. Radial section showing xylem, cambium and phloem. A, Light micrograph. Crystals (arrows) are visible in some parenchyma cells in theregion of the undifferentiated phloem. B, Polarized light image of the section in A, showing birefringence in the xylem and in the crystals.
Bar¯ 50 µm. For abbreviations, see legend to Fig. 1.
80 µm (Fig. 1A). On 3 April, cambial activity had resumedand division plates were observed in many of the cells on thephloem side (Fig. 1B). Division plates were not seen in thetwo or three layers of cambial cells adjacent to the xylemboundary but slight increases in the radial diameter of cellswere often detectable. Enlarging cells, which were pre-sumably precursors to vessel elements, were present withinthese two or three layers of the cambium (Fig. 1B). Thesamples obtained on 7 November and 3 April wererepresentative of the dormancy and reactivation stages ofthe cambium, respectively.
We examined the structure of the cambial zone and theadjacent tissues by conventional light microscopy and byCLSM in order to identify the true cambium (Figs 2–5). Thedormant cambium was bordered by axial parenchyma cellsand undifferentiated phloem elements on the phloem side.These adjacent undifferentiated cells appeared to have lesscytoplasm than the cambial cells (Figs 1A and 2A). In free-hand transverse sections it was found that substances, whichstained positively with Sudan IV but not with I
#-KI, were
present in the dormant cambial cells but not in theundifferentiated phloem elements (data not shown). Thehistochemical reactions suggest that these substances werelipid droplets (Jensen, 1962). The shapes of cambial cells ontangential sections resembled those of the undifferentiatedphloem elements. However, the region of the undifferen-

Kitin et al.—Cambial Cell Length in Kalopanax pictus 625
F. 3. Tangential sections showing morphological differences between the cambium and the differentiating phloem. A, Oblique tangential sectionshowing the last-formed xylem and cambium. No crystals are visible around the rays. B, Polarized light image of the section in A, showing strongbirefringence in the xylem. Slight birefringence is also evident in the ray initials. C, Tangential section through the differentiating phloem. Crystalsare visible in some axial and ray parenchyma cells (arrows). D, Polarized light image of the section in C, showing the strong birefringence of thecrystals. The birefringence of the walls of the cells is slightly greater than that in the region of the cambium shown in B. Bar¯ 50 µm. For
abbreviations, see legend to Fig. 1.
626 Kitin et al.—Cambial Cell Length in Kalopanax pictus
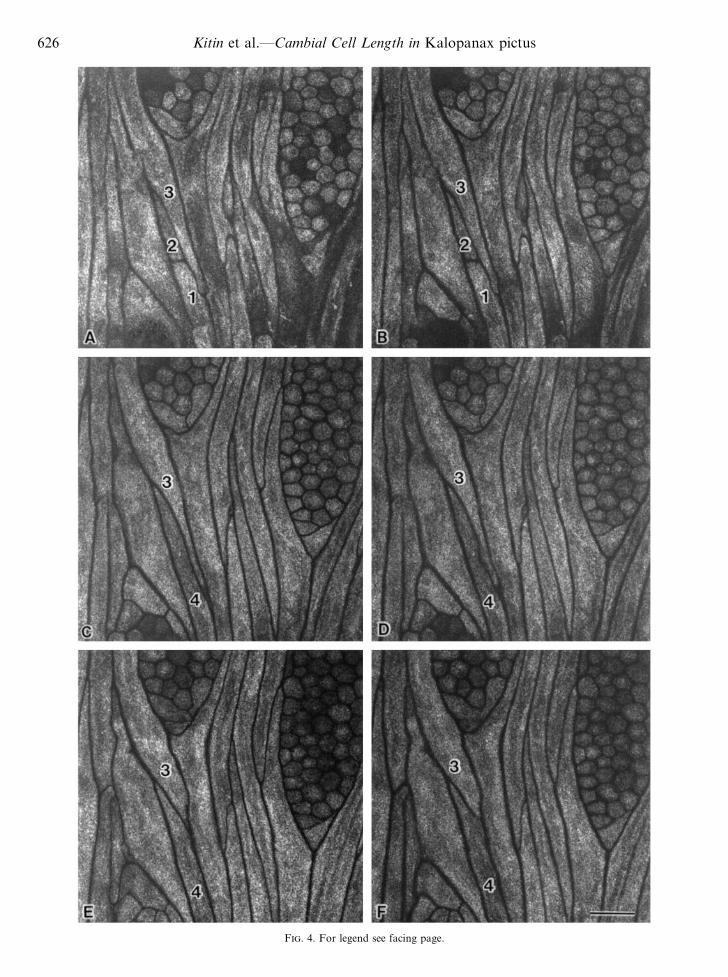
F. 4. For legend see facing page.

Kitin et al.—Cambial Cell Length in Kalopanax pictus 627
F. 5. Light micrograph of a tangential section (20 µm) of thecambium. The apparent tips of the fusiform cells may not be the actualtips (cf. Fig. 4) ; study of serial sections would be needed to establish
this. Bar¯ 50 µm.
tiated phloem was characterized by the presence of crystalsin some axial and ray parenchyma cells (Figs 2, 3C and D).
Methods used to measure lengths of fusiform cambial cells
Excitation of acridine orange-stained specimens with thehelium neon ion laser yielded negative fluorescence imagesof the cambium with dark cell walls and bright protoplasm(Fig. 4). The fluorescence from unstained specimens wasmuch weaker (data not shown). In general, it was difficult toview entire cambial cells in one focal plane with theconventional light microscope (Fig. 5) or on a singleconfocal image obtained by CLSM. However, serial opticalsectioning by CLSM allowed us to identify the tips of eachcell. The tips of fusiform cambial cells had often grownintrusively between adjacent cells. In the consecutiveconfocal images of cell 3 in Fig. 4, one of the tips can be seenin Fig. 4C, in which the cell appears the longest. Anexamination of successive cell images on serial tangentialsections also revealed the occurrence of anticlinal divisions.Cells 1 and 2 (Fig. 4A and B) are adjacent to cell 4 (Fig.4C–F) in a radial file of cells that originated from the same
F. 4. Serial confocal images at 2 µm intervals (A–F) from an approx. 60 µm tangential section of dormant cambium. Excitation by thehelium neon laser of the acridine orange-stained specimen yielded negative fluorescence images of the cambium, with dark cell walls and brightlumens. Note the change in the apparent length of cell 3 on sequential optical sections. The tip of cell 3 can be seen only in panel C. Cells 1 and
2 are produced by anticlinal division. They are adjacent to cell 4 in the same tier of fusiform cambial cells. Bar¯ 50 µm.
F. 6. Fusiform cambial cells after maceration by pectinase, viewedwith Nomarski differential interference contrast optics. Note thedifferent lengths of the cells (A–D). The isodiametric and the brick-likecells on the right in Fig. 6A are ray initials and initials of axial
parenchyma, respectively. Bar¯ 100 µm.
initial. Cells 1 and 2 appear to have formed as the result ofthe anticlinal division of a fusiform cell.
The cambial cells that had been macerated with pectinasewere sufficiently well preserved to allow measurements oflength to be made (Fig. 6).
Variations in the morphology and the lengths of fusiformcambial cells and �essel elements
An examination of both dormant and reactivated cam-bium in tangential sections showed that the fusiform cambialcells had variable dimensions, in particular around the raycambial cells (Figs 4 and 5). As observed in tangentialsections, the fusiform cambial cells in the macerated tissuevaried in terms of both shape and size (Fig. 6).
The morphology of vessel elements might provideimportant information about the pattern of expansion offusiform cambial cells. The vessel elements in the secondaryxylem of K. pictus varied in both size and shape (Fig. 7).Chalk and Chattaway (1934) defined the body of a vesselelement as the cell portion between the perforations.According to these same authors, the tails represent thatpart of the tapering end wall of the fusiform cambial cellthat has remained unoccupied by perforations and has notexpanded during differentiation. However, in K. pictus, thetails varied in width and length and perforations seemed tohave occurred not only at the tapering end walls but also atvarious sites on the body of the fusiform cambial cells (Fig.7). As suggested by Chalk and Chattaway (1934), the fulllength of a vessel element, including its body and tails, is

628 Kitin et al.—Cambial Cell Length in Kalopanax pictus
F. 7. Early wood vessel elements from tissue that had been macerated by Franklin’s method. Note the variations in size and structure (A–F).Bar¯ 100 µm.
F. 8. FE-SEM micrograph of a resin cast of the secondary xylem ofKalopanax pictus, showing an early wood vessel composed of elements
(arrows) of variable length. Bar¯ 500 µm.
representative of the length of the fusiform initial. If vesselelements are not macerated, their tails are not easily visible(Fig. 8). Figure 8 suggests variations in the lengths of thecells that form a single vessel but the precise length of eachvessel element cannot be measured.
700
0DCM
Sample
Len
gth
(µm
)
RCTDCT RCM
200
600
500
400
300
100
VE VL
65
586
92
631
56
524
50
526
70
597
150
640
F. 9. Box plots of cell lengths obtained from all samples examined.DCM, Cells of dormant cambium measured after maceration. DCT,Cells of dormant cambium measured on tangential sections. RCM,Cells of reactivated cambium measured after maceration. RCT, Cells ofreactivated cambium measured on tangential sections. VE, Early woodvessel elements. VL, Late wood vessel elements. Numbers at the endsof bars indicate the longest and the shortest lengths recorded.Horizontal bars within boxes show median values. The length of eachbox is the interquartile range of the length distribution (50% of caseshave values within the box). A larger spread of cell lengths is seen in thecase of reactivated cambium (RCM and RCT). By contrast, the celllengths of the vessel elements (VE and VL) are mostly concentrated
around the respective median values.
A summary of the distributions of cell lengths is displayedas box plots in Fig. 9 and histograms in Fig. 10. The averagelengths and standard deviations are shown in the histogramsin Fig. 10. In all samples, we found wide variations in celllength. The early wood vessel elements yielded severaloutlying values (Fig. 9). As seen in the box plots in Fig. 9,the median length of cells in the reactivated cambium was

Kitin et al.—Cambial Cell Length in Kalopanax pictus 629
30
00–50
Length (µm)
Fre
quen
cy (
%)
10
25
20
15
5
VE
>100 >150 >200 >250 >300 >350 >400 >450 >500 >550 >600 >650
n = 209s.d. = 107Mean = 355
30
0
Length (µm)
10
25
20
15
5
VL
n = 209s.d. = 90Mean = 400
30
00–50
Fre
quen
cy (
%)
10
25
20
15
5
RCM
>100 >150 >200 >250 >300 >350 >400 >450 >500 >550 >600 >650
n = 226s.d. = 137Mean = 280
30
0
10
25
20
15
5
RCT
n = 272s.d. = 127Mean = 248
30
00–50
Fre
quen
cy (
%)
10
25
20
15
5
DCM
>100 >150 >200 >250 >300 >350 >400 >450 >500 >550 >600 >650
n = 517s.d. = 120Mean = 324
30
0
10
25
20
15
5
DCT
n = 176s.d. = 144Mean = 392
0–50 >100 >150 >200 >250 >300 >350 >400 >450 >500 >550 >600 >650
0–50 >100 >150 >200 >250 >300 >350 >400 >450 >500 >550 >600 >650
0–50 >100 >150 >200 >250 >300 >350 >400 >450 >500 >550 >600 >650
F. 10. Frequency distributions, means and statistical summaries of cell lengths. DCM, Cells of dormant cambium measured after maceration.DCT, Cells of dormant cambium measured on tangential sections. RCM, Cells of reactivated cambium measured after maceration. RCT, Cellsof reactivated cambium measured on tangential sections. VE, Early wood vessel elements. VL, Late wood vessel elements. Mean, mean value;
s.d., standard deviation; n, number of cells measured.
shorter than that of cells in the dormant cambium and thevessel elements. The spread of lengths (expressed by thelength of a box) was widest in the reactivated cambium,narrower in the dormant cambium and narrowest in thevessel elements. The distributions of lengths of both dormantand reactivated cambium, measured in macerated tissue,were negatively skewed. The skew of data from the othersamples was much more limited (Fig. 9). The lengthsmeasured in the reactivated cambium had a bimodaldistribution, with peaks at 351–400 µm and around 100 µm(Fig. 10). Lengths in the dormant cambium also tended
towards bimodality but this tendency was less pronounced,with peaks at 351–400 µm and around 150 µm (Fig. 10). Thedistributions of lengths of samples from vessel elementswere close to a normal distribution with a peak at351–400 µm (Fig. 10). The proportion of shorter lengthswas higher in the reactivated cambium than in othersamples.
Analysis of variance revealed significant differences inmean cell length that depended on cell type, sampling date,and the method used for measurement (1603 d.f.,F¯ 56±688, P! 0±0005). A multiple comparison test by

630 Kitin et al.—Cambial Cell Length in Kalopanax pictus
T 1. Matrix of pairwise comparison probabilities
Sample DCM DCT RCM RCT VE VL
DCM —DCT 0±000 —RCM 0±000 0±000 —RCT 0±000 0±000 0±063 —VE 0±027 0±544 0±000 0±000 —VL 0±000 1±000 0±000 0±000 0±008 —
Post hoc test using the least-squares means of squared lengths(Bonferroni method; MSE¯ 13±889; d.f.¯ 1603).
DCM, Cells of dormant cambium measured after maceration. DCT,Cells of dormant cambium measured on tangential sections. RCM,Cells of reactivated cambium measured after maceration. RCT, Cells ofreactivated cambium measured on tangential sections. VE, Early woodvessel elements. VL, Late wood vessel elements. P¯ 0±000 means thatthe value is less than 0±0005.
Bonferroni’s method (Table 1) revealed a significantdifference (P! 0±01) in mean length between dormant andreactivated cambial cells and between reactivated cambialcells, early wood vessel elements and late wood vesselelements. There was a significant difference between themean length of cells of the dormant cambium whenmeasured in macerated tissue and when measured ontangential sections.
DISCUSSION
The cells within several layers of the cambial zone areidentical in terms of morphological and ultrastructuralcharacteristics and, thus, it is difficult to distinguish betweencambial initials and their adjacent derivatives (Catesson,1974, 1990, 1994; Barnett, 1981). In general, in earlierstudies, the lengths of any fusiform cells in the cambial zonehave been used to represent the lengths of fusiform initials(Bailey, 1920; Esau and Cheadle, 1955; Catesson, 1964;Ghouse and Yunus, 1974; Yunus, Yunus and Iqbal, 1978;Anand et al., 1981; Iqbal and Ghouse, 1985; Ajmal, Khanand Iqbal, 1986; Ajmal and Iqbal, 1987; Rao, Menon andDave, 1989; Siddiqi, 1991; Rao et al., 1996). It has beenassumed that as long as these cells remain within thecambial zone during their ontogenesis, measurements oflengths should be fairly representative of those of trueinitials (Larson, 1994). Our first goal was to define thecambial zone and to distinguish the cambial cells fromdifferentiating thin-walled cells. The differentiating phloemcould be identified by the presence of crystals in some axialand ray parenchyma cells (Fig. 3). Similarly, Catesson(1994) used oxalate-containing cells to identify the borderbetween cambial initials and undifferentiated phloem cellsin the dormant cambium of Fraxinus excelsior. In Aesculushippocastanum and F. excelsior, it has been suggested thatthe dormant cambial cells adjacent to the xylem (boundary-layer cells) differentiate into xylem (Barnett, 1992; Catesson,1994). The cells adjacent to the xylem boundary layer of thecambium have been identified as true initials, and all othercambial cells in the direction of the phloem have beenassumed to be committed to phloem formation (Barnett,1992; Catesson, 1994). Imagawa and Ishida (1972) observedthat the first vessel elements of the growing season in K.
pictus were derived from cambial cells adjacent to the xylem.Our observations also suggested that the early wood vesselelements in K. pictus might have been derived from cellslocated close to the xylem boundary. Therefore, in our serialtangential sections, we measured lengths of cambial cellsonly in layers close to the xylem. Macerated samples wereprepared from the first one to four layers of cells in thecambium adjacent to the xylem. In this way, we attemptedto confine our measurements to cambial initials and xylemmother cells at the early stage of development.
Many earlier studies of cambial cell length involvedspecies with storied cambium, in which cells are easier tomeasure in tangential sections (Butterfield, 1973; Ghouseand Yunus, 1974; Yunus et al., 1978; Ajmal et al., 1986;Ajmal and Iqbal, 1987; Rao et al., 1989, 1996; Siddiqi,1991). However, measurements of cell length in non-storiedcambium can be difficult. The fusiform cells of non-storiedcambium are usually longer than those of storied cambium(Metcalfe and Chalk, 1983) ; they overlap at their radial ortangential walls and can possibly be shifted to some extentfrom an axial orientation. Thus, long fusiform cells oftencannot be viewed from tip to tip in one focal plane (Fig. 5).One of the best methods for determining the dimensions andshapes of cambial cells is reconstruction from serialtransverse thin sections (Fujita, Tohyama and Harada,1984; Fujita, 1993). This method is, however, both laboriousand time-consuming. CLSM appears to be a useful new toolwith which to measure the lengths of cambial cells. Themethod involves serial optical sectioning through thecambial zone and reconstruction of the three-dimensionalstructure of cells. Results are obtained much faster thanwhen mechanical sections are used for the same purpose.
Our measurements of lengths of cells in the reactivatedcambium gave similar values with no significant difference(P" 0±05) between results from macerated tissue and fromtangential sections. However, there was a significantdifference between the mean lengths obtained from our twotypes of sample in the case of the dormant cambium. Thedistribution of lengths of cells in both dormant andreactivated cambium, measured after maceration, wasnegatively skewed compared to lengths measured ontangential sections. This bias suggests that mean cell lengthson tangential sections might be overestimated because offailure to detect short fusiform cells. The entire length offusiform cambial cells can be viewed at low magnification.Thus, short cells are better visualized after maceration andcan easily be overlooked in tangential sections. Werecommend maceration of samples as the more appropriatemethod for precise measurements of cambial cell length,provided that the cambial cells can be clearly identified.
Cheadle and Esau (1964) reported a range of lengths offusiform cambial cells from 205 to 678 µm in Liriodendrontulipifera (for reference, see Larson, 1994). Evert (1961,1963) reported a range of lengths from 252 to 711 µm inPyrus communis and from 268 to 717 µm in Pyrus malus.The lengths of fusiform cambial cells in K. pictus were morevariable within individual samples than the reported lengthsin other dicotyledonous species.
Our results reveal the occurrence of annual fluctuations inthe lengths of fusiform cambial cells in K. pictus. The mean

Kitin et al.—Cambial Cell Length in Kalopanax pictus 631
length of cambial cells after reactivation was about 20%less than that of dormant cambial cells and vessel elements.It has been suggested that the annual fluctuations in themean length of fusiform cambial cells might be due tochanges in the frequency of anticlinal divisions (Swamy etal., 1960; Bannan, 1967, 1970; Ajmal and Iqbal, 1987).
The vessel elements in early wood were significantlyshorter than those in late wood. Similar results have beenreported for other ring-porous hardwoods and it has beenproposed that shortening occurs during the differentiationof early wood vessel elements (Chalk and Chattaway, 1935;Su% ss, 1967). It seems more likely, however, that the differencebetween the mean lengths of vessel elements in early woodand late wood might be due to annual fluctuations in thelengths of fusiform cambial cells i.e. there is probably somedisparity among lengths of mother cells of vessel elementsthat are formed at different times of year. Our study showsthat the difference between the mean lengths of the fusiformcambial cells and the vessel elements depends on the time ofthe year at which the cambium is sampled, at least in K.pictus.
Comparisons of mean length cannot reveal the truerelationship between vessel elements and fusiform cambialcells. The average length might not be informative withrespect to the most frequent length, in particular if thedistribution of length is skewed. For example, in the case ofthe reactivated cambium of K. pictus, when cells weremeasured in macerated samples, the average length was280 µm, while the mode was within 351 to 400 µm. Oneinteresting issue worth discussing here is the tendencytowards bimodality of the distribution of cell lengths. Thistendency was strong in the case of samples from thereactivated cambium and weaker in the dormant cambium.The fusiform cambial cells most probably divide approx-imately into two halves by anticlinal division, which wouldresult in a bimodal distribution of lengths. However, thepattern of distribution of the lengths of vessel elements wasdifferent even though some evidence of bimodality was alsoevident. The difference between the patterns of lengthdistribution suggests that the relationship between vesselelements and fusiform cambial cells might not be as direct aspreviously assumed. It has not been possible to obtainsamples from cambium that contain only precursors tovessel elements and, thus, the mean length of fusiformcambial cells cannot be related directly to that of vesselelements.
In K. pictus, a ring-porous hardwood, the mean length ofearly wood vessel elements was indicative of the meanlength of fusiform cells in the dormant cambium but not ofthose in the reactivated cambium. The early wood vesselelements most probably originate from over-wintering cells,as suggested by Imagawa and Ishida (1972). The similarranges of lengths of dormant fusiform cambial cells andearly wood vessel elements suggest that short and longvessel elements might evolve from short and long fusiformcambial cells, respectively, without any change in length.
The length of fusiform cambial cells and vessel elementscan vary several-fold within a single sample. This variationnecessitates a high number of measurements to reveal truemean lengths. A minimum of 25 measurements, as required
by the IAWA Committee (1989), would not ensure reliableresults in the case of the mean lengths of fusiform cambialcells and vessel elements in K. pictus, a species with non-storied cambium.
ACKNOWLEDGEMENTS
This work was supported in part by Grants-in-Aid forScientific Research from the Japan Society for the Pro-motion of Science (no. JSPS-RFTF 96L00605).
LITERATURE CITED
Ajmal S, Iqbal M. 1987. Annual rhythm of cambial activity in Streblusasper. IAWA Bulletin (new series) 8 : 275–283.
Ajmal S, Khan R, Iqbal M. 1986. Cambial structure of Holopteleaintegrifolia Planch. in relation to age. Flora 178 : 197–202.
Anand SK, Singh DP, Paliwal SP, Paliwal GS. 1981. Size correlationsamong cambial initials and their derivatives in Jacarandamimosaefolia D. Don and Putranji�a roxburghii Wall. IndianJournal of Forestry 4 : 179–185.
Bailey IW. 1920. The cambium and its derivative tissues II. Sizevariations of cambial initials in gymnosperms and angiosperms.American Journal of Botany 7 : 355–367.
Bannan MW. 1967. Sequential changes in rate of anticlinal division,cambial cell length, and ring width in the growth of coniferoustrees. Canadian Journal of Botany 45 : 1359–1369.
Bannan MW. 1970. A survey of cell length and frequency ofmultiplicative divisions in the cambium of conifers. CanadianJournal of Botany 48 : 1585–1589.
Barnett JR. 1981. Secondary xylem cell development. In: Barnett JR,ed. Xylem cell de�elopment. Tunbridge Wells : Castle HousePublications, 47–95.
Barnett JR. 1992. Reactivation of the cambium in Aesculus hippo-castanum L.: a transmission electron microscope study. Annals ofBotany 70 : 169–177.
Butterfield BG. 1973. Variation in size of fusiform cambial initials andvessel members in Hoheria angustifolia Raoul. New ZealandJournal of Botany 11 : 391–410.
Catesson AM. 1964. Origine, fonctionnement et variations cytologiquessaisonnie' res du camb ium de l’Acer pseudoplatanus L. (Ace! race! es).Annales des Sciences Naturelles, Botanique (1e[ me seU rie) 5 : 229–498.
Catesson AM. 1974. Cambial cells. In: Robards AW, ed. Dynamicaspects of plant ultrastructure. New York: McGraw Hill, 358–390.
Catesson AM. 1990. Cambial cytology and biochemistry. In: Iqbal M,ed. The �ascular cambium. Taunton, Somerset : Research StudiesPress, 63–112.
Catesson AM. 1994. Cambial ultrastructure and biochemistry: changesin relation to vascular tissue differentiation and the seasonal cycle.International Journal of Plant Sciences 155 : 251–261.
Chalk L, Chattaway MM. 1934. Measuring the length of vesselmembers. Tropical Woods 40 : 19–26.
Chalk L, Chattaway MM. 1935. Factors affecting dimensionalvariations of vessel members. Tropical Woods 41 : 17–37.
Cheadle VI, Esau K. 1964. Secondary phloem of Liriodendron tulipifera.Uni�ersity of California Publications in Botany 36 : 143–252.
Cumbie BG. 1967. Developmental changes in the vascular cambium ofLeitneria floridana. American Journal of Botany 54 : 414–424.
Cumbie BG. 1983. Developmental changes in the wood of Bocconia�ulcanica Donn. Smith. IAWA Bulletin (new series) 4 : 131–140.
Esau K, Cheadle VI. 1955. Significance of cell divisions in differentiatingsecondary phloem. Acta Botanica Neerlandica 4 : 346–357.
Evert RF. 1961. Some aspects of cambial development in Pyruscommunis. American Journal of Botany 48 : 479–488.
Evert RF. 1963. Ontogeny and structure of the secondary phloem inPyrus malus. American Journal of Botany 50 : 8–37.
Franklin GL. 1945. Preparation of thin sections of synthetic resins andwood-resin composites, and a new macerating method for wood.Nature 155 : 51.
Fujii T. 1993. Application of a resin casting method to wood anatomyof some Japanese Fagaceae species. IAWA Journal 14 : 273–288.

632 Kitin et al.—Cambial Cell Length in Kalopanax pictus
Fujita M. 1993. Three-dimensional analyses of cambial activity andxylem differentiation. Fifteenth International Botanical Congress,Yokohama, Japan, August 28–September 3, 1993. Abstracts, 88.
Fujita M, Tohyama M, Harada H. 1984. Three-dimensional analyses ofthe cambium and the derivatives in Robinia pseudoacacia L. SudoS, ed. Proceedings of Pacific Regional Wood Anatomy Conference,October 1–7, 1984, Ibaraki, Japan. Tsukuba: Forestry and ForestProducts Research Institute, 91–93.
Furusawa O, Funada R, Murakami Y, Ohtani J. 1998. Arrangement ofcortical microtubules in compression wood tracheids of Taxuscuspidata visualized by confocal laser microscopy. Journal ofWood Science 44 : 230–233.
Ghouse AKM, Yunus M. 1974. Cambial structure in Dalbergia.Phytomorphology 24 : 152–158.
IAWA Committee 1989. IAWA list of microscopic features forhardwood identification. IAWA Bulletin (new series) 10 : 219–332.
Imagawa H, Ishida S. 1972. Study on the wood formation in trees.Report III. Occurrence of over-wintering cells in cambial zone inseveral ring-porous trees. Research Bulletin of the College Ex-perimental Forests. College of Agriculture, Hokkaido Uni�ersity 39 :207–221.
Iqbal M, Ghouse AKM. 1985. Impact of climatic variation on thestructure and activity of vascular cambium in Prosopis spicigera.Flora 177 : 147–156.
Iqbal M, Ghouse AKM. 1990. Cambial concept and organization. In:Iqbal M, ed. The �ascular cambium. Taunton, Somerset : ResearchStudies Press, 1–36.
Jensen AW. 1962. Botanical histochemistry. Principles and practice.San Francisco and London: W. H. Freeman and Company.
Larson PR. 1994. The �ascular cambium. De�elopment and structure.Berlin: Springer-Verlag.
Metcalfe CR, Chalk L. 1983. Anatomy of the dicotyledons. Vol. II 2ndedn. Oxford: Oxford University Press.
Philipson WR, Ward JM, Butterfield BG. 1971. The �ascular cambium:its de�elopment and acti�ity. London: Chapman & Hall.
Rao KS, Menon ARS, Dave Y. 1989. Developmental and dimensionalchanges in the cambial fusiform initials of Holoptelea integrifolia(Rohb.) Planch. Biologia Plantarum 31 : 126–132.
Rao KS, Rajput KS, Srinivas T. 1996. Comparative structure ofvascular cambium and its derivatives in some species of Sterculia.IAWA Journal 17 : 311–318.
Sass JE. 1951. Botanical microtechnique. Iowa: The Iowa State CollegePress.
Siddiqi TO. 1991. Impact of seasonal variation on the structure andactivity of vascular cambium in Ficus religiosa. IAWA Bulletin(new series) 12 : 177–185.
Su% ss H. 1967. U% ber die La$ ngena$ nderungen der Parenchymstra$ nge,Holzfasern und Gefa$ ssglieder von Laubho$ lzern im Verlauf einerZuwachsperiode. Holz als Roh- und Werkstoff 25 : 369–377.
Swamy BGL, Parameswaran N, Govindarajulu E. 1960. Variation invessel length within one growth ring of certain arborescentdicotyledons. Indian Botanical Society Journal 39 : 163–170.
Yunus M, Yunus D, Iqbal M. 1978. On the cambial structure of someindustrially important tropical trees. Flora 167 : 159–163.
Copyright © 2022 FDOKUMEN




















