
A taphonomic study of the African wild dog (Lycaon pictus
14
1 23 Archaeological and Anthropological Sciences ISSN 1866-9557 Archaeol Anthropol Sci DOI 10.1007/s12520-013-0164-1 A taphonomic study of the African wild dog (Lycaon pictus) J. Yravedra, M. Andrés & M. Domínguez-Rodrigo
Transcript of A taphonomic study of the African wild dog (Lycaon pictus

1 23
Archaeological and AnthropologicalSciences ISSN 1866-9557 Archaeol Anthropol SciDOI 10.1007/s12520-013-0164-1
A taphonomic study of the African wild dog(Lycaon pictus)
J. Yravedra, M. Andrés &M. Domínguez-Rodrigo

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag Berlin Heidelberg. This e-offprint is
for personal use only and shall not be self-
archived in electronic repositories. If you wish
to self-archive your article, please use the
accepted manuscript version for posting on
your own website. You may further deposit
the accepted manuscript version in any
repository, provided it is only made publicly
available 12 months after official publication
or later and provided acknowledgement is
given to the original source of publication
and a link is inserted to the published article
on Springer's website. The link must be
accompanied by the following text: "The final
publication is available at link.springer.com”.

ORIGINAL PAPER
A taphonomic study of the African wild dog (Lycaon pictus )
J. Yravedra & M. Andrés & M. Domínguez-Rodrigo
Received: 3 November 2012 /Accepted: 29 October 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Carnivore taphonomy has been traditionally usedfor the interpretation of archaeological sites in order to dis-criminate human-generated or modified from non-anthropicbone assemblages. In most of this actualistic research, thefocus has mainly been placed on hyenas and felids, neglectingother carnivores. This paper analyzes the taphonomic impactof the African wild dog (Lycaon pictus) on equid bones andcompares it with the bone modification patterns produced byother canids, such as wolves (Canis lupus ) in order to com-pare medium-/large-sized canid variability on bone modifica-tion patterns and elaborate a referential framework whichcould be feasibly applied to the zooarchaeological record todetect canid intervention on archaeological assemblages in thepast.
Keywords Lycaon pictus . Carnivore . Taphonomy . Toothmarks . Bonemodification
Introduction
Taphonomic studies of the behavior of carnivores have sig-nificantly contributed to our understanding of Palaeolithic siteformation (e.g., Brain 1981; Pickering et al. 2004, 2005;Domínguez-Rodrigo et al. 2007a, b). The application of ref-erential experimental studies to the prehistoric archaeologicalrecord have shown that in some sites, carnivores rather thanhumans were responsible for either most of the bone accumu-lations found (e.g., Domínguez-Rodrigo et al. 2007a, b;
Egeland 2007, 2008; Yravedra 2007, 2010) or for at least partof them (e.g., Blasco 1995, 1997; Yravedra 2006). SinceBrain (1981) and Binford’s (1981, 1984) seminal works,many scholars have attempted to characterize the bone mod-ification patterns generated by different types of carnivores,aiming at empirically identifying their presence in archaeo-logical sites (Cruz Uribe 1991; Dominguez-Rodrigo 1999;Ruiter and Berger 2000; Selvaggio and Wilder 2001;Dominguez-Rodrigo and Piqueras 2003; Pickering et al.2004; Fosse et al. 2009; Domínguez-Rodrigo and Pickering2010; Yravedra et al. 2011).
A number of papers have provided a description of bonemodification patterns considered typical of specific carnivores,producing diagnostic models to identify if these carnivores hada first or secondary access to the carcasses studied(Blumenschine 1988, 1995; Capaldo 1997; Domínguez-Rodrigo 1997; Domínguez-Rodrigo and Barba 2005).However, most of these studies were focused on hyenas andfelids (Hughes 1954; Sutcliffe 1973; Brain 1981; Cruz Uribe1991; Dominguez-Rodrigo 1999; Ruiter and Berger 2000;Pickering et al. 2004, 2005; Domínguez-Rodrigo et al. 2007a,b; Fosse et al. 2009; Domínguez-Rodrigo and Pickering 2010;Stiner et al. 2012; Gidna et al. 2013), neglecting other predatorssuch as canids. Only Binford (1978, 1981), Haynes (1980a, b,1981, 1982, 1983), Andrea and Gotthardt (1984), Nadal(1996), Prucca (2003), Castel (2004), Fosse et al. (2004),Barja and Corona (2007), Campmas and Beauval (2008);Esteban et al. (2009, 2010), Yravedra et al. (2011, 2013), andEsteban (2012) have provided information on captive and wildwolves and their bone-modifying habits.
In Africa, lions, hyenas, and leopards have been favored intaphonomic studies as the main carnivores predating on her-bivores of more than 100 kg (Hughes 1954; Brain 1981;Blumenschine 1986; Dominguez-Rodrigo 1999; Ruiter andBerger 2000; Pickering et al. 2004, 2005; Domínguez-Rodrigo and Pickering (2010). However, the African wilddog, a gregarious canid which preys on a large array ofanimals, including greater kudu, hartebeest, waterbuck, orzebra (Estes and Goddard 1967; Pienaar 1969; Malcolm andVan-Lawick 1975; Fuller and Kat 1990; Pole et al. 2004;
J. Yravedra (*) :M. Andrés :M. Domínguez-RodrigoDepartment of Prehistory, Complutense University of Madrid,C/Profesor Aranguren, Ciudad Universitaria s/n, 28040 Madrid,Spaine-mail: [email protected]
J. Yravedrae-mail: [email protected]
M. Domínguez-RodrigoMuseo de los Orígenes, IDEA (Instituto de Evolución en África),Plaza de San Andrés 2, 28005 Madrid, Spain
Archaeol Anthropol SciDOI 10.1007/s12520-013-0164-1
Author's personal copy

Rhodes and Rhodes 2004; Hayward et al. 2006; Harstone-Rose 2008), is also present in the same ecosystems but has notbeen considered in previous taphonomic studies. The role ofthis medium-size canid in prehistoric assemblages in Africaremains thus elusive due to a lack of a proper analogical modelfor comparison.
This paper describes the bone-modifying behavior of theAfrican wild dog in order to provide this missing experimentaland comparative framework. Thus, the description of theirbone modification patterns will be presented and complemen-tarily compared with the models derived from new experi-mental data for captive wolves. The ultimate goal is to empir-ically assess Harstone-Rose (2008) claim that wild dogs’specific dental morphology, when compared to otherslightly larger canids such as wolves shows a lessdurophagous way of consuming carcasses, involvingless contact between the predator’s dentition and carcassbones during defleshing, as suggested by the reducedapical wear of the premolars and molars cusps of theseAfrican canids. The quantifications of occlusal radii-of-curvature of carnivore premolars as well as the study of thecorrelation of this morphologywith carcass-processing behav-iors initially indicates that wild dogs are more clearlyhypercarnivorous than wolves and hence inflict less damageto bones (Harstone-Rose 2008). The present study intends toexperimentally test this proposal.
Material and methods
The sample comprises an assemblage of horse long bonesmodified by two African wild dogs in the Cabárceno NaturalPark (Cantabria, Spain) during 2009 and 2010. The experi-mental set included 71 elements, consisting of 17 humerii, 20radii, 22 ulnae, six femora, five tibiae, and one metapodial.For a period of 35 weeks, a pair of bones was fed to these wilddogs on a weekly basis. Bones were exposed to them forweeks, since bone collection took place every 2 months. Theremains were usually individually left inside the enclosurewhere the animals lived. Sometimes, distal epiphyses ofhumerii and femora were articulated to the zygopodials. Allbones belong to adult individuals.
The samples for captive wolves were collected in the samearea than the ones for African wild dogs using the samemethodology, with a different number of individuals. Atotal of 36 bones were considered this time, correspondingto five femura, five radii, four ulnae, six humerii, six tibiae,and other bones such as scapulae, innominate, calcaneus, andtalus.
Bone modifications were analyzed considering the fre-quency of the diverse tooth mark types and their anatomicaldistribution, as well as tooth marks dimensions, and bonefracture type and pattern.
The variables considered in this study were:
I–II Quantification of NISP and MNE as described byYravedra and Domínguez-Rodrigo (2009).
III Size of the carcass, according to Bunn (1982).IV Stage of the predator: wild versus in captivity.V Survival rate for each anatomical part.VI Survival of each anatomical part, calculated by dividing
the number of bones given to the wild dogs by thenumber of bones present after consumption.
VII Percentages of tooth marked NISP/total NISP (TM/NISP). Marks were analyzed using ×10–20 hand lenses,as described by Blumenschine et al. (1996).
VIII Frequencies of tooth marks by anatomical sections oflong bones according to element and bone section type.The distribution of toothmarks on diaphyses and epiph-yses was also recorded.
IX Number of marks on bones: A: no marks, B: between 1and 5 marks, C: between 5 and 10marks, and D: between10 and 20 marks per specimen, E: >20 marks perspecimen.
X Number of tooth marks by type (pits and scores) sensuBinford (1981) on each bone and section.
Tooth pit size In order to determine tooth mark dimensions,the length and width of both pits and scores were measuredwith a digital calliper as described by Andrés et al. (2012).Marks were spotted with hand lenses (×10–20) and bothconspicuous and inconspicuous (i.e., marks that precise mag-nification to be properly identified) marks were measured.Measurements were made using hand lenses and an electroniccalliper directly on the marks. Length (maximum dimension)and breadth (maximum dimension transversal to length) oftooth pits were taken. Marks were measured including aperimeter enclosing any modification of the original corticalsurface. This includes mark edge flaking, since the control ofthe sample allowed the ascription of this modification tocarnivores rather than any non-biotic agent.
Furrowing is defined as the deletion of (preferably cancel-lous) tissue at the ends of the long bones. Furrowing canpotentially act as a good indicator of the type of carnivorewhich modified the bone. Furrowing was analyzed by scoringits presence or absence, and quantified regarding its anatom-ical location on both long bone ends.
Results
The bone sample studied comprises 72 horse remains belong-ing to 71 anatomical elements (Table 1). A total of 56 of theremains (77.7 %) yielded tooth marks; however, no destruc-tion (expressed in the deletion of a long bone portion) ofelements by the African wild dog was documented (Fig. 1a).
Archaeol Anthropol Sci
Author's personal copy

Thus, the bones fed to these animals were not fragmented,cylinders were not documented and the epiphyses were notdestroyed after their intervention (Figs. 1a and 2a), not evenwhen disarticulated. As a consequence, the frequency of
survival for both long bones and epiphyses constituted100 % of the sample.
Upper bones (humerus and femur) presented the highestnumber of tooth marks (83.4 %). Intermediate elements (tibia
Table 1 Main features of the taphonomic assemblage modified by African wild dogs in the Cabárceno Natural Park (Cantabria, Spain)
Features Altered African wild dog’s bones
I. NISP (number of identified specimens) NISP: 72
II. MNE (minimum number of elements) MNE 71
III. Size or carcass Equus ferus caballus size 3 (below Bunn, 1982)
IV. Stage of the predator Captivity
V. Survival of bones All the bones which were given to the African Wild Dogs were recovered
VI. Survival of each anatomical part 72/72:100 %
VI % of appendicular 72/72:100 %
VI % of epiph. 110/110:100 %
VII Percentages of tooth marks (TM) 56/72:77.7 %
VII Appendicular: complete bones 36/47:76.6 % (only tibia, radius, ulna)
VII Appendicular: non-complete bones 20/24. 83.3 %
VII All appendicular bones 56/72:77.7 %
VII Upper limbs (humerus+femur) 20/24. 83.4 %
VII Intermediate limbs (radius+tibia) 17/25:68 %
VII Intermediate limbs (radius+tibia+ulna) 36/47:76.6 %
VII Lower limbs (metapodials) 0/1:0 %
VIII: Frequencies of TM by anatomical sections
VIII Diaphyses in upper limbs 4/8:50 % humerus (1/2:50 %); femur (3/6:50 %)
VIII Proximal epiphyses in upper limbs 1/2:50 % humerus (1/1:100 %); femur (0/1:0 %)
VIII Distal epiphyses in upper limbs 18/23:78.2 % humerus (13/17:76.5 %); femur (5/6:83.3 %)
VIII Diaphyses in intermediate limbs 12/24:50 % radius (10/20:53.6 %); tibia (2/5:40 %)
VIII Proximal epiphyses in intermediate limbs 15/24:62.5 % radius (12/19:63.1 %); tibia (3/5:60 %)
VIII Distal epiphyses in intermediate limbs 3/24:12 % radius (3/20:15 %); tibia (0/5:0 %)
VIII Diaphyses in lower limbs 0/1:0 % no metacarpal; metatarsal (0/1:0 %)
VIII Proximal epiphyses in lower limbs 0/1:0 % no metacarpal; metatarsal (0/1:0 %)
VIII Distal epiphyses in lower limbs 0/1:0 % no metacarpal; metatarsal (0/1:0 %)
IX Number of tooth marks on bones Hum/Fem/Rad/Tib/Mc/MT Humerus and femur: C; more frequently radius: B; more frequently tibia:C; more frequently metatarsal: A
X Number of tooth marks on bones by type (pits and scores)
X %Pits on Epp/Epd/Diaph long bones Epp 12/16:75 %; Epd 17/21:81 %; Diaph 7/16:43.8 % ulna excluded
X %Scores on Epp/Epd/Diaph long bones Epp 8/16:50 %; Epd 13/21:62 %; Diaph 13/16:81.2 % ulna excluded
X %Pits on Hum/Fem/Rad/Tib/Mc/MT Epp Hu: 1/1:100 %; Fe: 0/0:0 %; Ra: 8/12:67 %; Ti: 3/3:100 %; Mt: 0/0:0 %
X %Pits on Hum/Fem/Rad/Tib/Mc/MT Epd Hu: 9/13:69 %; Fe: 5/5:100 %; Ra: 3/3:100 %; Ti: 0/0:0 %; Mt: 0/0:0 %
X % Pits on Hum/Fem/Rad/Tib/Mc/MT Diaph Hu: 1/1:100 %; Fe: 1/3:33.3 %; Ra: 5/10:50 %; Ti: 0/2:0 %; Mt: 0/0:0 %
X% scores on Hum/Fem/Rad/Tib/Mc/MT Epp Hu: 1/1:100 %; Fe: 0/0:0 %; Ra: 6/12:50 %; Ti: 1/3:33.3 %; Mt: 0/0:0 %
X % scores on Hum/Fem/Rad/Tib/Mc/MT Epd Hu: 11/13:85 %; Fe: 2/5:40 %; Ra: 0/3:0 %; Ti: 0/0:0 %; Mt: 0/0:0 %
X % scores on Hum/Fem/Rad/Tib/Mc/MT Diaph Hu: 1/1:100 %; Fe: 2/3:66.6 %; Ra: 8/10:80 %; Ti: 2/2:100 %; Mt: 0/0:0 %
Comments Bones were not highly altered except for some epiphyses which presentedmoderate-slight furrowing. African wild dogs seemed to produce fewalterations: they were rather meat-eaters instead
NISP with TM (tooth mark)/NISP. See variables in text
Epp and Epd proximal epiphyses and distal epiphyses, TM tooth mark, Epiph epiphyses,Upper Limbs humerus and femur, Intermediate Limbs radiusand tibia, Lower Limbs metapodial, Hum/Fem/Rad/Tib/Mc/Mt humerus/femur/radius/tibia/metacarpal/metatarsal, Epp/Epd/Diaph proximal epiphyses/distal epiphyses/diaphyses
Archaeol Anthropol Sci
Author's personal copy
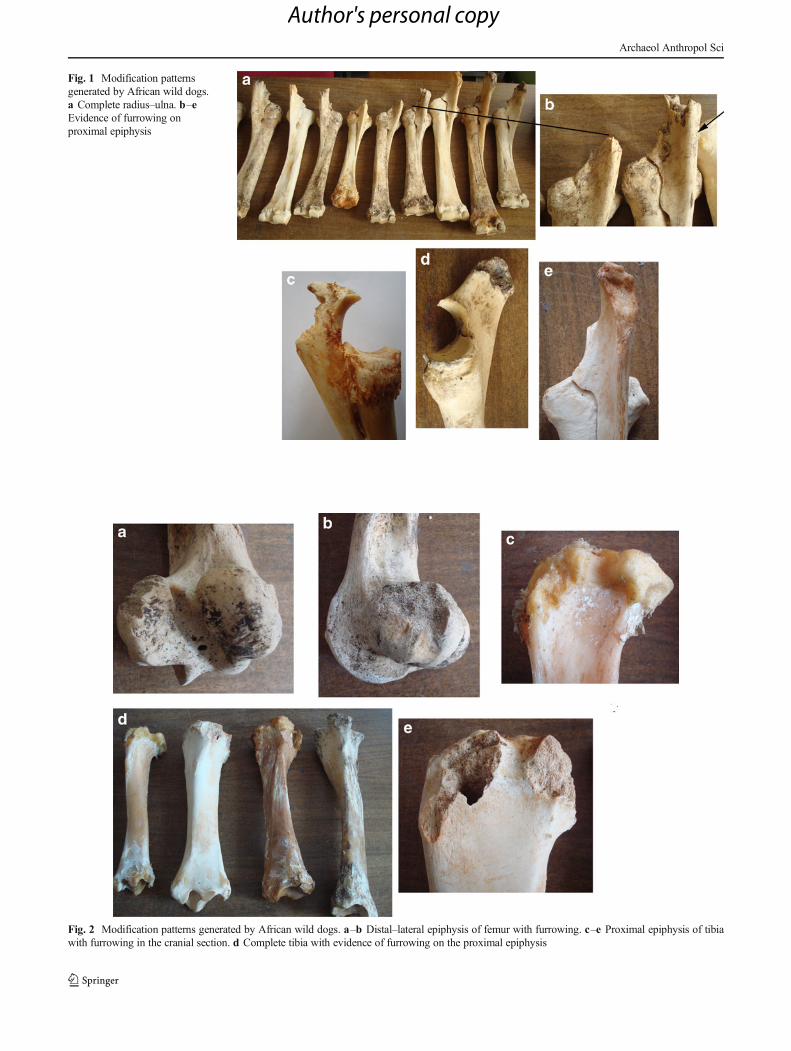
Fig. 1 Modification patternsgenerated by African wild dogs.a Complete radius–ulna. b–eEvidence of furrowing onproximal epiphysis
Fig. 2 Modification patterns generated by African wild dogs. a–b Distal–lateral epiphysis of femur with furrowing. c–e Proximal epiphysis of tibiawith furrowing in the cranial section. d Complete tibia with evidence of furrowing on the proximal epiphysis
Archaeol Anthropol Sci
Author's personal copy
and radius) showed 70% of tooth marked elements in the caseof radii (however, when ulna was included, this frequencyraised to 76.6%) whereas tibiae were tooth-marked in 60% ofcases. While not statistically relevant, the only metatarsalavailable did not exhibit any marks (Tables 1–3).
Regarding upper appendicular bones, distal epiphysesyielded a tooth marked frequency affecting 78.2 % of thecases (humerus, 76.5 %; femur, 83.3 %). In the case ofintermediate elements, proximal epiphyses showed thehighest frequency of tooth marks, with 62 % of the specimens(radius, 63.2 %; tibia, 60 %). The addition of the proximal
epiphyses of ulnae increased this value to 74 % in radii–ulnae(proximal epiphyses of ulna, 86.3 %). In this context, it wasremarkable that distal epiphyses, as well as the lower part ofthe diaphyses, yielded a rather low frequency of tooth marks,reaching only 12 % altogether (radius, 15 %; tibia, 0 %;Table 1).
Considering the number of marks per bone, frequencieswere often low, spanning on average less than 10 marks perspecimen (Tables 1–2). The tooth marks identified were bothpits and scores, distributed on all appendicular elements, bothon diaphysial and epiphysial sections (Tables 1–3), although
Table 2 Number of tooth marks per specimen, discriminating pits from scores
Distribution of pit and score by bone
Bone Section NISP NISPwith TM
No. of TM/specimen
Mean TM/specimen
No. of pits Mean pits/bone
No. ofscores
Mean score/bone
Humerus Prox. Eph. 1 1 8 8/1:8 6 6/1:6 2 2/1:2
Humerus Diaph. 2 1 54 54/1:54 15 15/1:15 39 39/1:39
Humerus Dist. Eph. 17 13 104 104/13:8 54 54/13:4.2 50 50/13:3.8
Radius Prox. Eph. 19 12 32 32/12:2.7 24 24/12:2 8 8/12:0.7
Radius Diaph. 19 10 31 31/10:3.1 4 4/10:0.4 27 27/10:2.7
Radius Dist. Eph. 20 3 12 12/3:4 10 10/3:3.3 2 2/3:0.7
Ulna Prox. Eph. 22 19 172 172/19:9.1 120 120/19:6.3 52 52/19:2.7
Ulna Diaph. 22 2 6 6/2:3 0 0/2:0 6 6/2:3
Femur Prox. Eph. 1 0 0 0/0:0 0 0/0:0 0 0/0:0
Femur Diaph. 6 3 3 3/3:1 2 2/3:0.7 1 1/3:0.3
Femur Dist. Eph. 6 5 65 65/5:13 48 48/5:9.6 17 17/5:3.4
Tibia Prox. Eph. 5 3 26 26/3:8.66 10 10/3:3.3 16 16/3:5.3
Tibia Diaph. 5 2 5 5/2:2.5 0 0/2:0 5 5/2:2.5
Tibia Dist. Eph. 5 0 0 0/0:0 0 0/0:0 0 0/0:0
Metatarsal Prox. Eph. 1 0 0 0/0:0 0 0/0:0 0 0/0:0
Metatarsal Diaph. 1 0 0 0/0:0 0 0/0:0 0 0/0:0
Metatarsal Dist. Eph. 1 0 0 0/0:0 0 0/0:0 0 0/0:0
Table 3 NISP, MNE, TM (tooth marks)
Bone with presence of pits and scores
Bone NISP MNE TM TM/NISP %TM Pits Pits/TM % pits Scores Scores/TM % scores
Humerus 18 17 15 83.3 % 11 73.3 % 12 80 %
Radius 20 20 14 70 % 9 64.3 % 12 85.7 %
Ulna 22 22 19 86.3 % 17 89.5 % 16 84.2 %
Femur 6 6 5 83.3 % 5 100 % 3 60 %
Tibia 5 5 3 60 % 3 100 % 2 66.7 %
Metatarsal 1 1 0 0 % 0 0.0 % 0 0.0 %
Total 72 71 56 77.7 % 46 82.1 % 45 80.4 %
Upper limbs 24 23 20 83.3 % 17 15 % 85 75.0 %
Intermediate limbs 25 25 17 68 % 12 14 % 71 82.4 %
Intermediate limbs 2 47 47 36 76.6 % 29 30 % 81 83.3 %
Pits-scores number of bones with tooth marks showing pits and scores, Upper limbs humerus+femur, Intermediate limbs radius+tibia, Intermediatelimbs 2 radius+ulna+tibia
Archaeol Anthropol Sci
Author's personal copy

epiphysis yielded pits more frequently and diaphysis pro-duced more scores (Tables 1–2). In any case, pits and scoresaffected 82.1 and 80.4 % of the bones with tooth marks,suggesting that both types of marks were frequently producedin all bones and sections, usually together (Table 3). The pit toscore ratio is 1.29, which shows that pits are more frequentthan scores as in other durophagous carnivores.
Tooth mark dimensions were small, with pits on diaphysesmeasuring on average slightly more than 1 mm and some2.6 mm in the case of epiphyses (Table 4). Scores were alsosmall, about 5 mm in length, with some exceptional specimenslarger than 2 cm (Table 4). Lastly, marks related to the intensityof gnawing were negligible in the experimental sample(Table 5), as they had never completely collapsed the epiphysis(Figs. 1 and 2), not even on the ulna, the bone most seriouslyaffected by furrowing. Furthermore, with the only exception ofthe ulna, the proximal epiphysis of the tibia and distal epiphysisof the femur had furrowing frequencies higher than 50 %(Fig. 2); in the remaining cases, the frequencies were lowerthan 12 %, including the distal epiphysis of the humerus. Notonly the bones affected by furrowing were scarce (Table 5), butalso furrowing intensity was reduced, generally affecting thelateral and cranial parts of the epiphysis more than the medialones (Table 5, Fig. 2). Since wild dogs did not fracture or
collapse the epiphyses, producing only moderate furrowing,and large tooth marks were absent, they only produced areduced number of modifications per element. Then, it can begenerally argued that African wild dogs do not have a strongimpact on bone modification when medium-sized carcasses areconsidered, as opposed to wolves (Haynes 1980a, b, 1981,1983, Binford 1981; Nadal 1996; Prucca 2003; Castel 2004;Campmas and Beauval 2008; Yravedra et al. 2011).
Discussion
1. Comparison of tooth mark frequencies made by captiveAfrican wild dogs and wolves
For the comparison of bone modifications generatedby African wild dogs and those typically attributed towolves, tooth mark frequencies per element, as well asthe number of marks per specimen, were considered.Subsequently, tooth mark measurements, together withfurrowing on long bones were also discussed. The com-parison of the bones modified by African wild dogs withthe ones altered by captive wolves clearly showed thepreference of both canids for upper appendicular elements(Table 6). Thus, in all cases, tooth mark frequencies
Table 4 Dimensions of Africanwild dog’s tooth marks (in mm),considering pits and scores lengthand width depending on their lo-cation on either epiphyses ordiaphyses
No. Mean I.C. lower I.C. upper Standard deviation Min. Max.
Pit. Diaph. Length 21 1.1 0.85 1.35 0.56 0.3 2.27
Pit. Diaph. Width 21 0.76 0.6 0.92 0.35 0.3 1.56
Pit. Eph. Length 274 2.66 2.45 2.87 1.76 0.3 10.72
Pit. Eph. Width 274 2.02 1.85 2.19 1.41 0.3 8.78
Score. Diaph. Length 78 5.43 4.63 6.23 3.5 0.66 20.43
Score. Diaph. Width 78 0.42 0.32 0.51 0.42 0.14 3.69
Score. Eph. Length 147 5.1 4.55 5.54 3.16 0.75 17.37
Score. Eph. Width 147 0.7 0.58 0.82 0.68 0.16 4
Table 5 Furrowing produced by African wild dogs on horse appendicular bones
Bone Section NISP Furrow. boneswith epiph.
Furrow. bones/NISP %
Cranialpart
.% Caudalpart
% Lateralpart
% Medialpart
%
Humerus Prox. Eph. 1 0 0.0 0 0 0 0
Humerus Dist. Eph. 17 2 11.8 % 0 0 2 100 % 1 50 %
Radius Prox. Eph. 19 1 5.3 % 0 0 1 100 % 0
Radius Dist. Eph. 20 1 5.0 % 1 100 % 0 0 0
Ulna Prox. Eph. 22 22 100 % 20 91 % 21 95.5 % 21 95.5 % 21 95.5 %
Femur Prox. Eph. 1 0 0.0 0 0 0 0
Femur Dist. Eph. 6 3 50 % 3 100 % 1 33 % 3 100 % 1 33 %
Tibia Prox. Eph. 5 4 80 % 4 100 % 0 3 75 % 1 25 %
Tibia Dist. Eph. 5 0 0.0 0 0 0 0
Metapod. Prox. Eph. 1 0 0.0 0 0 0 0
Metapod. Dist. Eph. 1 0 0.0 0 0 0 0
Archaeol Anthropol Sci
Author's personal copy
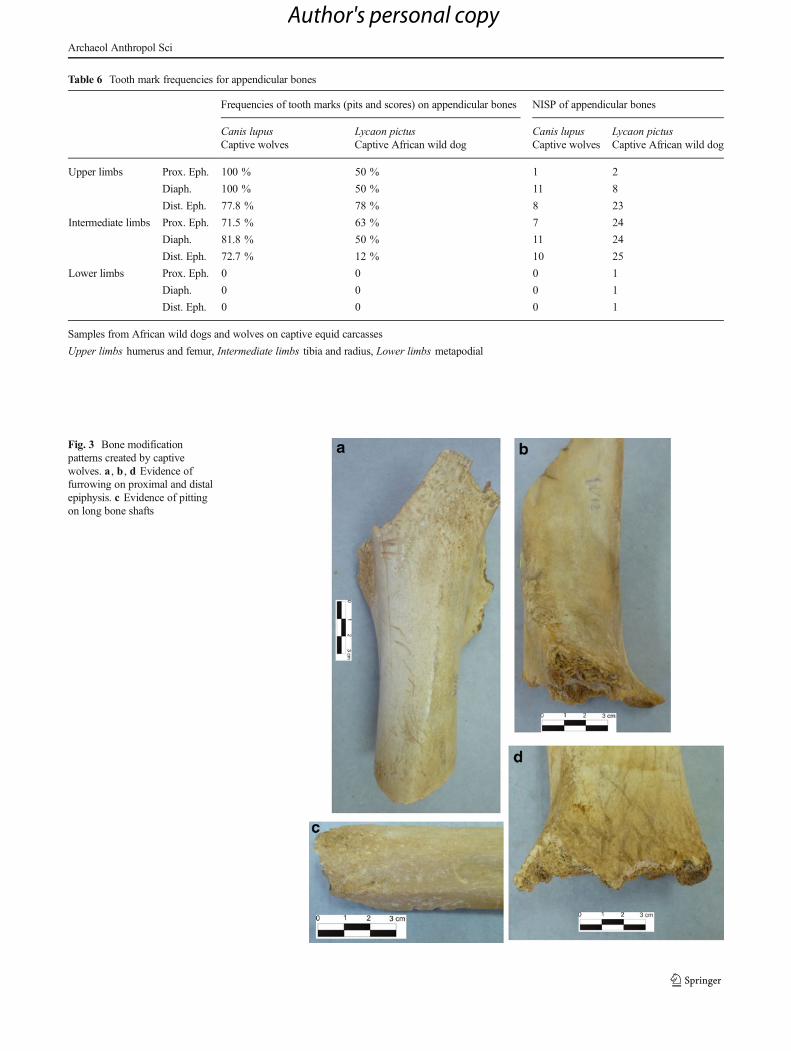
Table 6 Tooth mark frequencies for appendicular bones
Frequencies of tooth marks (pits and scores) on appendicular bones NISP of appendicular bones
Canis lupus Lycaon pictus Canis lupus Lycaon pictusCaptive wolves Captive African wild dog Captive wolves Captive African wild dog
Upper limbs Prox. Eph. 100 % 50 % 1 2
Diaph. 100 % 50 % 11 8
Dist. Eph. 77.8 % 78 % 8 23
Intermediate limbs Prox. Eph. 71.5 % 63 % 7 24
Diaph. 81.8 % 50 % 11 24
Dist. Eph. 72.7 % 12 % 10 25
Lower limbs Prox. Eph. 0 0 0 1
Diaph. 0 0 0 1
Dist. Eph. 0 0 0 1
Samples from African wild dogs and wolves on captive equid carcasses
Upper limbs humerus and femur, Intermediate limbs tibia and radius, Lower limbs metapodial
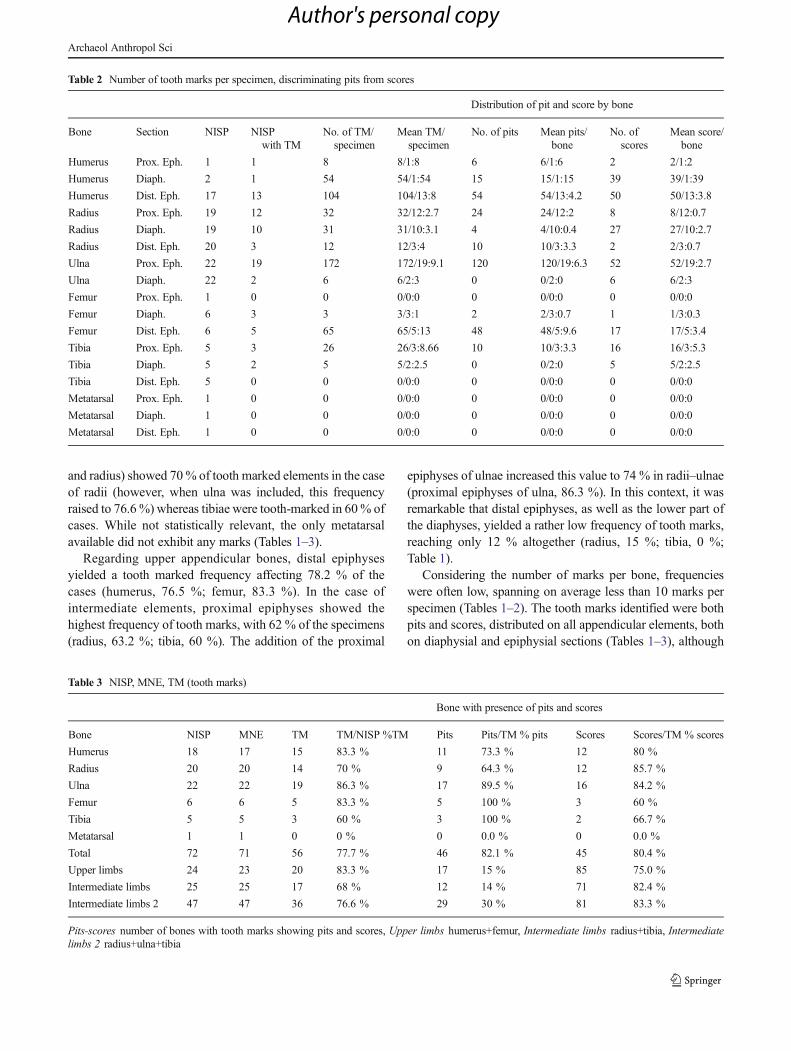
Fig. 3 Bone modificationpatterns created by captivewolves. a , b , d Evidence offurrowing on proximal and distalepiphysis. c Evidence of pittingon long bone shafts
Archaeol Anthropol Sci
Author's personal copy

reported on these bones were higher than 50 % on bothepiphyses and diaphyses.
For intermediate elements, tooth mark frequencieswere still high on the proximal portion of limbs, butreduced on the distal sections in the case of captiveAfrican wild dogs, not exceeding 12 % of the cases,whereas for captive wolves it yielded 72.7 % of toothmarks. From this perspective, the bone-modifying behav-ior of African wild dogs were dissimilar to the one report-ed for captive wolves, the former affecting those anatom-ical elements with higher meat yields and the latter beingfocused on the complete bone, including the distalepiphysis (Figs. 3 and 4).
When comparing average frequencies of tooth-markedelements per bone type, a non-parametricMann–Whitney–Wilcoxon test showed that wild dogs’ tooth mark frequen-cies on humerii (p =0.65), femora (p =0.63), radii–ulnae(p =0.64), and tibiae (p =0.64) were similar to the onesrecorded for wolves.
2. Number of marks per elementThe modification produced by African wild dogs on
bones can be analyzed from at least two complementaryapproaches: counting the number of marks per bone, andidentifying the degree of fragmentation and the type offurrowing present (see infra). In the first case, Africanwild dogs produced a reduced number of marks per bone:
Fig. 4 Modification patternsgenerated by captive wolves ontibiae, showing intensivefurrowing including collapse ofthe proximal epiphysis (a , b , c)and moderate furrowing on thedistal epiphysis (b , d)
Archaeol Anthropol Sci
Author's personal copy
between five and 10 marks/specimen on upper elements,and less than five marks/specimen on intermediate ones,although the tibia and the ulna may exceptionally yieldmore than five marks on their proximal epiphyses(Tables 1 and 2). On the other hand, for captive wolvesthe number of tooth marks per bone is high: between 30and 35marks/specimen on upper elements and between 15and 42 marks/specimen on intermediate bones. Captivewolves produced many pits and scores in all the sectionsof the bones, as well as occasional pitting, intensivefurrowing, and collapse of the epiphyses (Figs. 3 and 4).
The comparison presented above shows that Africanwilddogs do not produce considerable modifications on bones, afact also emphasized by the samples studied here. In thiscase, the long bones fed to African wild dogs in captivity fora week did not yield more than 10 marks per bone, asopposed to the patterns documented in captive wolves.
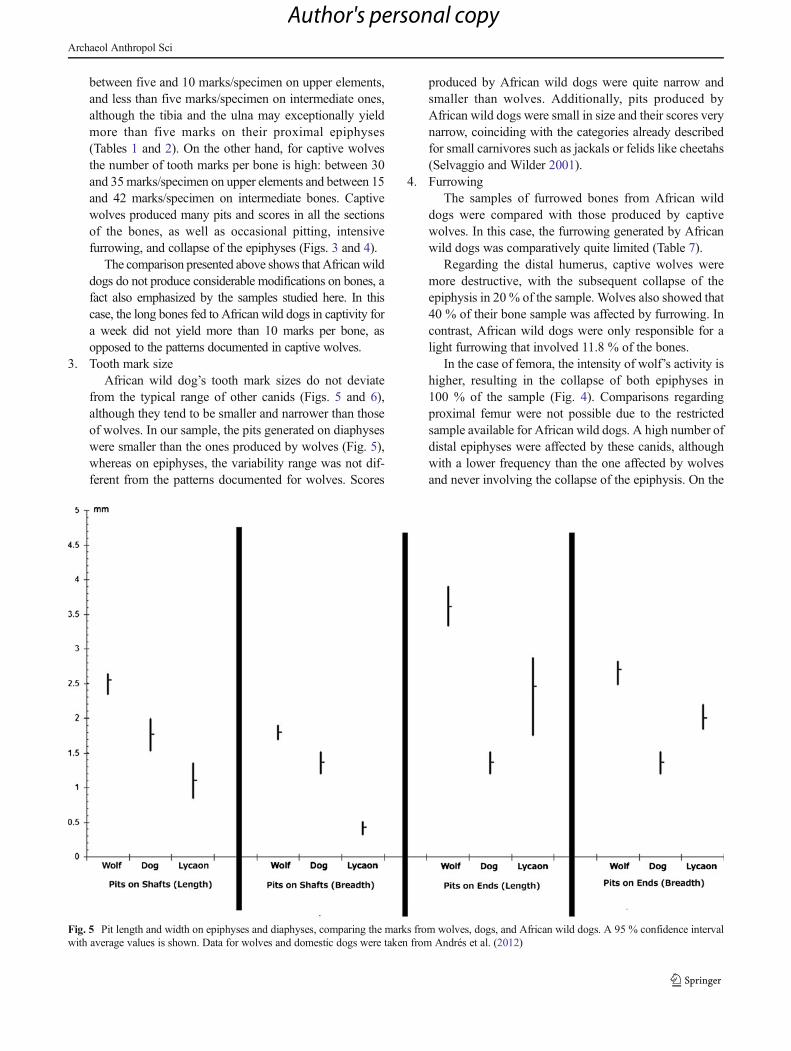
3. Tooth mark sizeAfrican wild dog’s tooth mark sizes do not deviate
from the typical range of other canids (Figs. 5 and 6),although they tend to be smaller and narrower than thoseof wolves. In our sample, the pits generated on diaphyseswere smaller than the ones produced by wolves (Fig. 5),whereas on epiphyses, the variability range was not dif-ferent from the patterns documented for wolves. Scores
produced by African wild dogs were quite narrow andsmaller than wolves. Additionally, pits produced byAfrican wild dogs were small in size and their scores verynarrow, coinciding with the categories already describedfor small carnivores such as jackals or felids like cheetahs(Selvaggio and Wilder 2001).
4. FurrowingThe samples of furrowed bones from African wild
dogs were compared with those produced by captivewolves. In this case, the furrowing generated by Africanwild dogs was comparatively quite limited (Table 7).
Regarding the distal humerus, captive wolves weremore destructive, with the subsequent collapse of theepiphysis in 20 % of the sample. Wolves also showed that40 % of their bone sample was affected by furrowing. Incontrast, African wild dogs were only responsible for alight furrowing that involved 11.8 % of the bones.
In the case of femora, the intensity of wolf’s activity ishigher, resulting in the collapse of both epiphyses in100 % of the sample (Fig. 4). Comparisons regardingproximal femur were not possible due to the restrictedsample available for African wild dogs. A high number ofdistal epiphyses were affected by these canids, althoughwith a lower frequency than the one affected by wolvesand never involving the collapse of the epiphysis. On the
Fig. 5 Pit length and width on epiphyses and diaphyses, comparing the marks from wolves, dogs, and African wild dogs. A 95 % confidence intervalwith average values is shown. Data for wolves and domestic dogs were taken from Andrés et al. (2012)
Archaeol Anthropol Sci
Author's personal copy
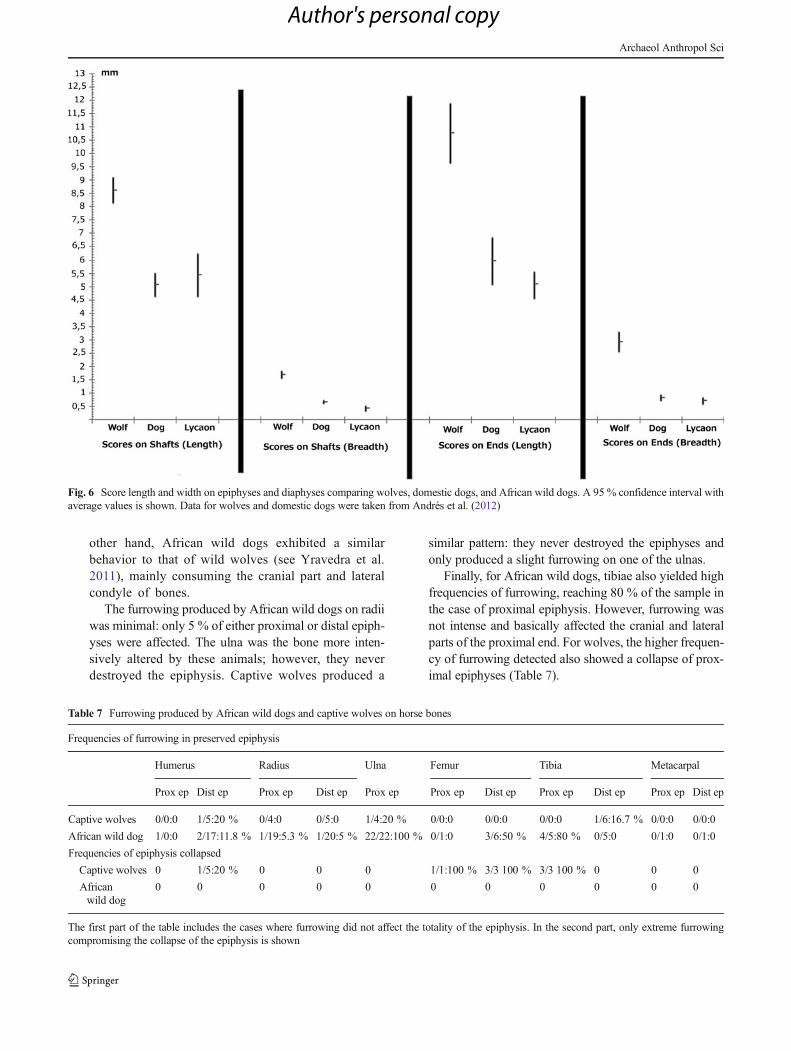
other hand, African wild dogs exhibited a similarbehavior to that of wild wolves (see Yravedra et al.2011), mainly consuming the cranial part and lateralcondyle of bones.
The furrowing produced by African wild dogs on radiiwas minimal: only 5 % of either proximal or distal epiph-yses were affected. The ulna was the bone more inten-sively altered by these animals; however, they neverdestroyed the epiphysis. Captive wolves produced a
similar pattern: they never destroyed the epiphyses andonly produced a slight furrowing on one of the ulnas.
Finally, for African wild dogs, tibiae also yielded highfrequencies of furrowing, reaching 80 % of the sample inthe case of proximal epiphysis. However, furrowing wasnot intense and basically affected the cranial and lateralparts of the proximal end. For wolves, the higher frequen-cy of furrowing detected also showed a collapse of prox-imal epiphyses (Table 7).
Fig. 6 Score length and width on epiphyses and diaphyses comparing wolves, domestic dogs, and African wild dogs. A 95 % confidence interval withaverage values is shown. Data for wolves and domestic dogs were taken from Andrés et al. (2012)
Table 7 Furrowing produced by African wild dogs and captive wolves on horse bones
Frequencies of furrowing in preserved epiphysis
Humerus Radius Ulna Femur Tibia Metacarpal
Prox ep Dist ep Prox ep Dist ep Prox ep Prox ep Dist ep Prox ep Dist ep Prox ep Dist ep
Captive wolves 0/0:0 1/5:20 % 0/4:0 0/5:0 1/4:20 % 0/0:0 0/0:0 0/0:0 1/6:16.7 % 0/0:0 0/0:0
African wild dog 1/0:0 2/17:11.8 % 1/19:5.3 % 1/20:5 % 22/22:100 % 0/1:0 3/6:50 % 4/5:80 % 0/5:0 0/1:0 0/1:0
Frequencies of epiphysis collapsed
Captive wolves 0 1/5:20 % 0 0 0 1/1:100 % 3/3 100 % 3/3 100 % 0 0 0
Africanwild dog
0 0 0 0 0 0 0 0 0 0 0
The first part of the table includes the cases where furrowing did not affect the totality of the epiphysis. In the second part, only extreme furrowingcompromising the collapse of the epiphysis is shown
Archaeol Anthropol Sci
Author's personal copy

Summing up, African wild dogs produced furrowingmainly on proximal ulnas, distal femora, proximal tibiae,and distal humerii. Except for the ulnas, the remainingbones only showed moderate furrowing (rather gnawing)which did not end in the collapse of the epiphysis. Incontrast, wolves were responsible for a more consistentfurrowing on distal and proximal femora and proximaltibiae (Table 7, Figs. 3 and 4).
Conclusions
As already discussed, taphonomic studies on carnivores havetraditionally been focused on hyenas and felids (Hughes 1954;Sutcliffe 1973; Brain 1981; Cruz Uribe 1991; Dominguez-Rodrigo 1999; Ruiter and Berger 2000; Pickering et al. 2004,2005; Domínguez-Rodrigo et al. 2007a, b; Fosse et al. 2009;Domínguez-Rodrigo and Pickering 2010). However, othercarnivores such as the African wild dog can also producespecific bone modification patterns that require furtherconsideration.
The African wild dog, unlike other canids, is unique ingenerating moderate modification on bones. The tooth markfrequencies they produce on appendicular elements are higherthan 50 %, both on upper bones (humerus and femur) andintermediate elements (tibia and radius). As opposed to othercanids such as wolves, the number of tooth marks yielded byeach individual bone is lower than 10 marks. This shows thatthe damage inflicted by these animals on bones is definitelynot intense. This interpretation is also supported by the mod-erate furrowing recorded in the sample, the lack of bonebreakage and cylinders, and the collapse of epiphyses.Regarding tooth marks, the scores produced by wild dogsare narrow and the pits, small, similar to the ones generatedby small carnivores such as felids or exclusive flesh-eaters likecheetahs or leopards. Taking into account the tooth mark sizerange and morphology created by African wild dogs as well asthe moderate impact they generate on bones, it is possible toadvance that their carcass-processing behavior seems to beoriented towards meat rather than bone consumption. Thishypothesis can be further tested by the tooth wear analysiscarried out by Harstone-Rose (2008) on African wild dogs’dental morphology. Nevertheless, the ideas presented in thispaper are intended to be further put to the test in an on-goingstudy of the taphonomic impact of the African wild dog onsize one to two preys (sensu Bunn 1982) and carcasses huntedand consumed in the wild.
Acknowledgments Many thanks to Mr. Santiago Borragán, ChiefVeterinary Officer and the staff of the felid section at Cabárceno NaturalPark (Cantabria), for kindly facilitating research onAfrican wild dogs andfor collaborating with us. They were most helpful with the feeding ofthese canids and the recovery of the bones. Finally, we thank Aixa Vidalfor the translation.
References
Andrea AC, Gotthardt RM (1984) Predator and scavenger modificationof recent equid skeletal assemblages. Artic 36:276–283
Andrés M, Gidna AO, Yravedra J, Domínguez-Rodrigo M (2012) Astudy of dimensional differences of tooth marks (pits and scores)on bones modified by small and large carnivores. ArchaeolAnthropol Sci 4:209–219
Barja I, Corona E (2007) El análisis de excretas desde la Etología y laArqueozoología: el caso del lobo ibérico. BAR S1627 .Archaeopress, Oxford
Binford LR (1978) Nunamiut ethnoarchaeology. Academic, New YorkBinford LR (1981) Bones: ancient men and modern myths. Academic,
New YorkBinford LR (1984) Faunal remains from Klasies River Moupth.
Academic, New YorkBlasco MF (1995) Hombres y Fieras. Estudio Zooarqueológico y
Tafonómico del Yacimiento del Paleolítico Medio de la Cueva deGabasa 1 (Huesca). Monografías de la Universidad de Zaragoza,Zaragoza
Blasco MF (1997) Cave site of Gabasa in the Spanish Pyrinees. JAntropol Res 53:177–218
Blumenschine R (1986) Early hominid scavenging opportunities.Implications of carcass availability in the Serengeti and Ngrongoroecosystems. B.A.R. International Series 283. Archaeopress, Oxford
Blumenschine R (1988) An experimental model of the timing of hominidand carnivore influence on archaeological bone assemblages. JArchaeol Scie 15:483–502
Blumenschine R (1995) Percussion marks, tooth marks and the experi-mental determinations of the timing of hominid and carnivore accessto long bones at FLK Zinjanthropus, Olduvai Gorge, Tanzania. JHum Evol 29:21–51
Blumenschine RJ, Marean CW, Capaldo SD (1996) Blind tests of inter-analyst correspondence and accuracy in the identification of cutmarks, percussion marks, and carnivore tooth marks on bone sur-faces. J Archaeol Sci 23:493–507
Brain CK (1981) The hunters or the hunted? University of Chicago Press,Chicago
Bunn HT (1982) Meat-eating and human evolution: studies on the dietand subsistence patterns of Plio-Pleistocene hominins in East Africa.Ph. Dissertation. University of California, Berkeley
Campmas E, Beauval C (2008) Consommation osseuse des carnivores :résultats de l’étude de l’exploitation de carcasses de bœufs (Bostaurus) par des loups captifs. Annales de Paléontologie 94:167–186
Capaldo SD (1997) Experimental determinations of carcass proceedingby Plio-Pleistocene hominids and carnivores at FLK 22(Zinjanthropus), Olduvai Gorge, Tanzania. J Hum Evol 33:555–598
Castel JC (2004) L'influence des canidés sur la formation des ensemblesarchéologiques. Caractérisation des desrtructions dues au loup.Revue de Paléobilogie 23:675–693
Cruz Uribe K (1991) Distinguishing hyena from hominid bone accumu-lations. J Field Archaeol 18:467–486
Domínguez-Rodrigo M (1997) Meat eating by early hominids at FLKZinj 22 site, Olduvay Gorge Tanzania: an experimental approachusing cut-mark data. J Hum Evol 33:669–690
Dominguez-Rodrigo M (1999) Flesh availability and bone modificationin carcasses consumed by lions. Paleogeogeography, Paleoclimate,Paleoecology 149:373–388
Domínguez-Rodrigo M, Barba R (2005) A study of cut marks on small-sized carcasses and its application to the study of cut marked bonesfrom small mammals at the FLK Zinj site. J Taphon 3:121–134
Domínguez-Rodrigo M, Pickering TR (2010) A multivariate approachfor discriminating bone accumulations created by spotted hyenasand leopards: harnessing actualistic data from East and SouthernAfrica. J Taphon 8:155–179
Archaeol Anthropol Sci
Author's personal copy

Dominguez-Rodrigo M, Piqueras A (2003) The use of tooth pits toidentify carnivore taxa in tooth-marked archaeofaunas and theirrelevance to reconstruct hominid carcass processing behaviours. JArchaeol Sci 30:1385–1391
Domínguez-Rodrigo M, Barba M, Egeland CP (2007a) DeconstructingOlduvai: a taphonomic study of the Bed I sites. Springer, Dordrecht
Domínguez-Rodrigo M, Egeland CP, Pickering TR (2007b) Equifinalityin carnivore tooth marks and the extended concept of archaeologicalpalimpsests: implications for models of passive scavenging of earlyhominids. In: Pickering TR, Schick K, Toth N (eds) Breathing lifeinto fossils: taphonomic studies in honor of C.K. (Bob) Brain. StoneAge Institute, Bloomington, pp 255–288
Egeland C (2007) Zooarchaeological and taphonomic perspectives onhominid and carnivore interactions at Olduvai Gorge. Ph.Dissertation. University of Indiana, Tanzania
Egeland C (2008) Patterns of early hominid site use at Olduvai Gorge.Mitteilungen der Gesellschaft für Urgeschichte 17:9–37
Esteban M (2012) Can archaeozoology and taphonomy contribute toknowledge of the feeding habits of the Iberian wolf? J ArchaeolSci 39:3208–3216
Esteban M, Cáceres I, Tarazona C (2009) Experimentando con lobos;secuencia de acceso, consumo y dispersión de una carcasa de équidoen la Sierra de la Culebra, Zamora (Península Ibérica). Bloque 3,Capítulo XLII:351–356.
Esteban M, Cáceres I, Fosse P (2010) Characterization of a currentcoprogenic sample originated by Canis lupus as a tool for identify-ing a taphonomic agent. J Archaeol Sci 37:2959–2970
Estes RD, Goddard J (1967) Prey selection and hunting behavior of theAfrican wild dog. J Wildl Manag 31:52–69
Fosse P, Laudet F, Selva N, Wajrak A (2004) Premières observationsnéotaphonomiques sur des assemblages osseux deBialowieza (N. E.Pologne): Intérêts pour les gisements Pleistocenes d'Europe. Paléo16:91–116
Fosse P, Avery G, Fourvel JB, Lesur-Gebremariam J, Monchot H, BrugalJP, Kolska Horwitz L (2009) The modern hyena dens: a criticalsurvey of their taphonomic characterization from new excavatedsites (Republic of Djibouti, South Africa) and from the literature.Zona Arqueológica 13:108–117
Fuller TK, Kat PW (1990) Movements, activity, and prey relationships ofAfrican wild dogs (Lycaon pictus ) near Aitong, southwesternKenya. Afr J Ecol 28:330–350
Gidna A, Yravedra J, Domínguez-Rodrigo M (2013) A cautionary noteon the use of captive carnivores to model wild predator behavior: acomparison of bone modification patterns on long bones by captiveand wild lions. J Archaeol Sci 40:1903–1910
Harstone-Rose A (2008) Evaluating the hominin scavenging niche throughanalysis of the carcass-processing abilities of the carnivore guild. Ph.Dissertation. Duke University, Graduate School, Department ofBiological Anthroppology and Anatomy, Durham, NC
Haynes G (1980a) Prey bones and predators. Potential ecologic informa-tion from analysis bones of bone site. OSSA 7:75–97
Haynes G (1980b) Evidence of carnivore gnawing on Pleistocene andrecent mammalian bones. Paleobiology 6:341–351
Haynes G (1981) Bone modification and skeletal disturbances by naturalagencies: studies in North America. University MicrofilmsInternational. The Catholic University of America. Ph D
Haynes G (1982) Utilization and skeletal disturbances of North Americanprey carcasses. Artic 35:266–281
Haynes G (1983) A guide for differentiating mammalian carnivore taxaresponsible for gnaw damage to herbivore limb bones. Paleobiology9:164–172
Hayward MW, O’Brian J, Hofmeyr M, Kerley GIH (2006) Prey prefer-ences of the african wild dog Lycaon pictus (Canidae: Carnivora):
ecological requirements for conservation. J Mammal 87(6):1122–1131
Hughes A (1954) Hyaenas versus australopithecines as agents of boneaccumulation. Am J Phys Anthropol 12:467–486
Malcolm JR, Van-Lawick HV (1975) Notes onwild dogs (Lycaon pictus)hunting zebras. Mammalia 39:231–240
Nadal J (1996) Patrones de desmembración en herbívoros consumidospor lobos (Canis lupus). In:Meléndez G, BlascoMF, Pérez I (eds) IIReunión de Tafonomía y Fosilización. Institución Fernando elCatólico, Zaragoza, pp 259–263
Pickering TR, Dominguez-Rodrigo M, Egeland CP, Brain CK (2004)Beyond leopards: tooth marks and the contribution of multiplecarnivore taxa to the accumulation of the Swartkans Member 3fossil assemblage. J Hum Evol 46:595–604
Pickering TR, Domínguez-Rodrigo M, Egeland C, Brain CK (2005) Thecontributions of limb bone fracture patterns to reconstructing earlyHominid behaviour at Swartkrans Cave (South Africa): archaeolog-ical application of a new analytical method. Int J Osteoarchaeol 15:247–256
Pienaar UDV (1969) Predator–prey relationships amongst the largermammals of the Kruger National Park. Koedoe 12:108–176
Pole A, Gordon IJ, Gorman ML, Macaskill M (2004) Prey selection byAfrican wild dogs (Lycaon pictus) in southern Zimbabwe. J Zool262:207–215
Prucca A (2003) Caracterisaion de l’impact des loups sur desossements d’herbivores (cerfs de Virginie, origneaux, Bisons):Etude des modifications inliges par des loups captifs et sauvagesNord-americains. DEA prehistorie, Archeologie, Histoire et civ-ilisations de l’antiquité et du moyen age. Aix Es Provence.MMHH.
Rhodes R, Rhodes G (2004) Prey selection and use of natural and man-made barriers by African wild dogs while hunting. S Afr JWildl Res34:135–142
Ruiter JD, Berger LR (2000) Leopard as a taphonomic agents indolomitic caves. Implications for bone accumulations in thehominid bearning deposits of South Africa. J Archaeol Sci27:665–684
Selvaggio MM, Wilder J (2001) Identifing the involvement of multiplecarnivore taxon with archaeological bone assemblages. J ArchaeolSci 28:465–470
Stiner M, Munro N, Sanz M (2012) Carcass damage and digested bonefrom mountain lions (Felis concolor): implications for carcass per-sistence on landscapes as a function of prey age. J Archaeol Sci 39:896–907
Sutcliffe AJ (1973) Caves of the east African Rift Valley. Trans Cave ResGroup G B 15:41–65
Yravedra J (2006) Aportes naturales y antrópicos en la Cueva delEsquilleu (Cantabria, España). Zona Arqueológica 7:280–289
Yravedra J (2007) Nuevas contribuciones en el comportamientocinegético de la Cueva de Amalda. Munibe 58:43–88
Yravedra J (2010) A taphonomic perspective on the origins of the faunalremains from Amalda Cave (Spain). J Tapho 8:301–334
Yravedra J, Domínguez-Rodrigo M (2009) The shaft-based method-ological approach to the quantification of long limb bones andits relevance to understanding hominid subsistence in thePleistocene: application to four Palaeolithic sites. J Quaternary Sci24:85–96
Yravedra J, Lagos L, Bárcena F (2011) A taphonomic study of wild wolf(Canis lupus) modification of horse bones in Northwestern Spain. JTaphon 9:37–67
Yravedra J, Lagos L, Bárcena F (2013) The wild wolf (Canis lupus) as adispersal agent of animal carcasses in northwestern Spain. Journal ofTaphonomy
Archaeol Anthropol Sci
Author's personal copy




















