
The role of the fusiform face area in social cognition: implications for the pathobiology of autism
14
doi: 10.1098/rstb.2002.1208 , 415-427 358 2003 Phil. Trans. R. Soc. Lond. B Marois and Pawel Skudlarski Robert T. Schultz, David J. Grelotti, Ami Klin, Jamie Kleinman, Christiaan Van der Gaag, René implications for the pathobiology of autism The role of the fusiform face area in social cognition: References http://rstb.royalsocietypublishing.org/content/358/1430/415#related-urls Article cited in: Email alerting service here top right-hand corner of the article or click Receive free email alerts when new articles cite this article - sign up in the box at the http://rstb.royalsocietypublishing.org/subscriptions go to: Phil. Trans. R. Soc. Lond. B To subscribe to This journal is © 2003 The Royal Society on May 24, 2011 rstb.royalsocietypublishing.org Downloaded from
Transcript of The role of the fusiform face area in social cognition: implications for the pathobiology of autism

doi 101098rstb20021208 415-427358 2003 Phil Trans R Soc Lond B
Marois and Pawel SkudlarskiRobert T Schultz David J Grelotti Ami Klin Jamie Kleinman Christiaan Van der Gaag Reneacute implications for the pathobiology of autismThe role of the fusiform face area in social cognition
Referenceshttprstbroyalsocietypublishingorgcontent3581430415related-urls
Article cited in
Email alerting service heretop right-hand corner of the article or click Receive free email alerts when new articles cite this article - sign up in the box at the
httprstbroyalsocietypublishingorgsubscriptions go to Phil Trans R Soc Lond BTo subscribe to
This journal is copy 2003 The Royal Society
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
Published online 21 January 2003
The role of the fusiform face area in social cognitionimplications for the pathobiology of autism
Robert T Schultz12 David J Grelotti1 Ami Klin1 Jamie Kleinman3Christiaan Van der Gaag4 Rene Marois5 and Pawel Skudlarski2
1Child Study Center Yale University School of Medicine 230 S Frontage Road New Haven CT 06520-7900 USA2Department of Diagnostic Radiology Yale University School of Medicine 333 Cedar Street New Haven CT 06510 USA
3Department of Psychology University of Connecticut Unit 1020 Storrs CT 06269-1020 USA4Academic Centre for Child and Adolescent Psychiatry PO Box 660 9700 AR Groningen The Netherlands
5Department of Psychology Vanderbilt University 111 21st Avenue Nashville TN 37203 USA
A region in the lateral aspect of the fusiform gyrus (FG) is more engaged by human faces than any othercategory of image It has come to be known as the lsquofusiform face arearsquo (FFA) The origin and extent ofthis specialization is currently a topic of great interest and debate This is of special relevance to autismbecause recent studies have shown that the FFA is hypoactive to faces in this disorder In two linkedfunctional magnetic resonance imaging (fMRI) studies of healthy young adults we show here that theFFA is engaged by a social attribution task (SAT) involving perception of human-like interactions amongthree simple geometric shapes The amygdala temporal pole medial prefrontal cortex inferolateral frontalcortex and superior temporal sulci were also significantly engaged Activation of the FFA to a task withoutfaces challenges the received view that the FFA is restricted in its activities to the perception of faces Wespeculate that abstract semantic information associated with faces is encoded in the FG region andretrieved for social computations From this perspective the literature on hypoactivation of the FFA inautism may be interpreted as a reflection of a core social cognitive mechanism underlying the disorder
Keywords amygdala autism fusiform face area medial prefrontal cortex social cognitionsuperior temporal sulcus
1 INTRODUCTION
For the first time the field of autism has a replicatedneurofunctional marker of the disordermdashhypoactivation ofthe FFA The FFA is that region of the middle aspect ofthe right FG that is selectively engaged by faces (whencontrasted with object perception tasks) (Puce et al 1995Kanwisher et al 1997 Kanwisher 2000) Anatomicallythe middle portion of the FG is split along its rostralndashcaudal extent by a shallow MFS In fMRI the centre ofactivation in face perception tasks is typically offsettowards the lateral aspect of the FG in the right hemi-sphere (Haxby et al 1999) Whereas individual subjectsmay or may not also show left FG activation during faceperception group composites always show right-side acti-vations to be larger At least five fMRI studies have shownthat older children adolescents and adults with autismspectrum disorders have reduced levels of activity toimages of the human face in this specialized face regionof the right hemisphere (Critchley et al 2000 Dierks etal 2001 Pierce et al 2001 Schultz et al 2000a 2001)These data are consistent with an older and more exten-sive psychology literature documenting performance defi-cits in face perception (Langdell 1978 Klin et al 1999)
Author for correspondence (robertschultzyaleedu)
One contribution of 14 to a Theme Issue lsquoAutism mind and brainrsquo
Phil Trans R Soc Lond B (2003) 358 415ndash427 415 Oacute 2003 The Royal SocietyDOI 101098rstb20021208
and facial expression recognition in autism (eg Hobsonet al 1988ab MacDonald et al 1989 Yirmiya et al1992) They seem to provide an important clue as to theneural ontogeny and pathobiology of autism
Whereas the consistency of these findings is encour-aging what it means to have an under-responsive FFAremains unclear Our initial interpretation of this findingfocused on the role of experience for shaping the visualcortices (Schultz et al 2000ab Grelotti et al 2001) It isknown that the ventral temporal visual areas are quiteplastic and can be moulded by early experiences (Gaffanet al 1988 Webster et al 1991 Fujita et al 1992 Lowel ampSinger 1992) Persons with autism pay much less attentionto the face (Osterling amp Dawson 1994 Klin et al 2002)and this may be why they fail to acquire normal perceptualskill in this domain Inadequate attention to faces duringcritical periods of cortical development should affect thematuration of these areas and presumably lead to unde-ractivation of the FFA during face perception
This interpretation fits nicely into the perceptual expert-ise model of the FFA first championed by Gauthier andcolleagues (Gauthier et al 1999 2000) Gauthier hasshown in two elegant fMRI studies that the FFAresponds preferentially to any class of object for which aperson is perceptually lsquoexpertrsquo For example she foundthat bird experts engage the FFA more strongly whenviewing birds than cars but the reverse is true for carexperts (Gauthier et al 2000) Moreover normal young
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
416 R T Schultz and others FFA and social cognition
adults can enhance their FFA activity to a class of novelobjects through extensive perceptual training (Gauthier etal 1999) Interpreting the hypoactivation of the FFA inautism from an expertise model however argues that thisfinding is an outcome of having autism rather than partof the cause In other words the hypoactivation of theFFA is merely a reflection of the social disability the cul-mination of a set of developmental experiences acrossmany years whereby the person has reduced interest inother people and pays inadequate attention to their facesIn this regard the under-responsiveness of the FFA is abiological marker Identifying an endophenotype such asthis is extremely important and takes the field one signifi-cant step closer to understanding the underlying biologicalmechanisms but it falls short of providing a snapshot ofthe brain mechanisms that actually cause autism
An alternative view would be that the FFA is a corecomponent of the lsquosocial brainrsquo Data emerging overrecent years from neuroimaging studies human lesionstudies and animal studies suggest a working model of thesocial brain that comprises a diverse set of frontal limbicand temporal lobe circuitry Select aspects of the orbitaland medial prefrontal cortices the amygdala and lateralaspects of the temporal cortex involving the STS haveeach been implicated in social functioning (Brothers 1990Fletcher et al 1995 Baron-Cohen et al 1999 Frith ampFrith 1999 Allison et al 2000 Castelli et al 2000 2002Schultz et al 2000b) The frontal and temporal corticeshave dense and often reciprocal connections to theamygdala (Carmichael amp Price 1995 Price et al 1996)The amygdala is centrally positioned and capable of mod-ulating and interpreting the emotional significance of dataprocessed in the perceptual cortices as well as assistingwith the integration of emotion and cognition for decisionmaking and action in the frontal cortices (Amaral et al1992 Schultz et al 2000b) Collectively this systemdefines a heuristic model of the social brain with the pre-cise functions of each node only understood in an as yetsuperficial manner
But is there a role for the FFA in this social circuitryWhereas the role of the FG in face perception is undis-puted only one prior study has implicated the FFA insocial cognition Castelli et al (2000) used social ani-mations involving interacting geometric shapes to probethe social brain These animations were based on theclassic study of Heider amp Simmel (1944) that showed howcertain movements by inanimate objects could stronglyand automatically suggest personal agency and that agroup of interacting geometric forms will naturally suggestsocial interactions All but one of the 34 female collegestudents in Heider and Simmelrsquos study described the ani-mations through a social lens and in human terms (egshapes chase one another fight entrap play get fright-ened elated etc) It seems that the contingent nature ofthe shape movements and the fact that their movementsviolate the rules of simple physics (ie the shapes seem tohave lsquoagencyrsquo or will) naturally invoke social cognitive andsocial perceptual ideation Using PET in six healthy youngadults Castelli et al showed that interpreting this type ofanimation engaged the medial prefrontal cortices the TPthe STS and the right FG Although the localization of thefusiform activations in their study is in the area generallyreported to be the FFA it is not clear whether this region
Phil Trans R Soc Lond B (2003)
of activation would have overlapped with the FFA in thesesubjects as location of the FFA can vary from person toperson Nevertheless engagement of the right FG by asocial cognitive task that does not involve images of theface suggests that this FG region may have a broadermore important set of functions extending beyond simpleface perception As such it provides a basis to argue thatthe hypoactivation of the FFA to faces in autism might beilluminating part of a causal mechanism as opposed to adevelopmental consequence of having autism
The current study also used an adaptation of the pro-cedure of Heider amp Simmel (1944) involving what Klin(2000) called the SAT Klin (2000) used the SAT to showhow persons with autism fail to spontaneously imposesocial meaning on these types of movements Other inves-tigators have also reported a paucity of theory of mindideation among persons with autism to other renditions ofthe HeiderndashSimmel procedure (Abell et al 2000Bowler amp Thommen 2000) Use of simple shapes to dis-play human social interactions without perceptual rep-resentations of real people strips the social event down tothe essential elements needed to convey social meaningsIn this regard the SAT is an ideal neuroimaging probefor assessing social cognitive and social perceptual pro-cesses in a way that is not confounded by perceptual pro-cesses that would be provoked if actual images of faces orpeople were used The original SAT from Klin (2000) isa 50 s silent film in which three moving geometric shapes(a circle a small triangle and a larger triangle) interactwith each other in a social manner Interestingly in Klinrsquosstudy some attributions by those with an autism spectrumdisorder were given in terms of physical meaning (egmagnetic forces) not social meaning We took note of thisobservation in developing the following fMRI experi-ments and created a control task for the SAT that involvesjudgements of object mass
In two fMRI studies of the same group of normal con-trol subjects we show that the FG is robustly engaged byan adaptation of the SAT suitable for a block design fMRIstudy Engagement of the right FG by non-face stimulisuggests that this region has functions beyond static faceperception To test the exact location of the fusiform acti-vations during the SAT nine of the 12 participants con-sented to return for a fMRI study of face perceptionResults from this second study found the location of theFFA to be highly overlapping with the FG activations tothe SAT Thus making social judgements on non-facegeometric figures and making identity judgements ongrey-scale pictures of human faces draws upon a similarneural substrate in the FG This result challenges thespecificity of the middle portion of the FG for faces andraises the possibility that the FFA is part of the primarycircuitry for social cognition As such it has importantimplications for the hypoactivation of the FFA in autismand more generally for specifying a distributed social net-work whose dysfunction might cause autism
2 METHODS
(a) ParticipantsTwelve participants were recruited for this study from the staff
and student populations at Yale University The sampleincluded six men (three left-handed) and six women (one left-
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 417
handed) ranging in age from 20 to 31 years (meanplusmn sd = 242 plusmn 31) Participants were screened for neurologicaland DSM IV Axis I psychiatric disorders Estimated full-scaleIQ as measured by four subtests of the Wechsler Adult Intelli-gence Scale 3rd edition (Wechsler 1997) averaged 1288(plusmn 104) (Information Vocabulary Picture Completion andBlock Design) All subjects scored in the normal range on theBenton Test of Facial Recognition (Benton 1994) (raw scorerange 41ndash50 mean plusmn sd = 468 plusmn 26) There were no signifi-cant differences between the sexes on any of these variables
Nine of the twelve agreed to participate in a follow-up fMRIstudy of face perception conducted so that localization of theFFA could be compared with SAT activations in the middle FGarea in the right hemisphere Five were male (one left-handed)and four were female (one left-handed) mean age was236 plusmn 26 years All subjects gave written informed consent forboth studies in accordance with procedures and protocolsapproved by the Institutional Review Board of the Yale Univer-sity School of Medicine
(b) Experimental tasksWe adapted the SAT for use in a fMRI block-design study by
programming 16 new SAT QuickTime film skits using Directorpublished by Macromedia (600 Townsend Ave San FranciscoCA wwwmacromediacom) From these a final set of eightwere chosen for use in the fMRI study on the basis of ratingsby project staff of the filmrsquos realism and ability to capture onersquossocial attention (these films can be downloaded from httpinfomedyaleeduchldstdyneuroimgsatFmovieshtm) Each filmlasted 151 s and was designed like the original SAT with move-ments intended to suggest a sense of personal agency andreciprocal and contingent interactions that were meant to be eas-ily interpreted as social Each film contained three types of whitegeometric figures (a triangle diamond and circle) that movedagainst a black background In common with the original SATthere was a box in the centre of the field with one wall thatopened as if on a hinge allowing the shapes to open and shutthe door and to enter chase or drag other shapes inside Eachfilm was scripted to follow a social story for example hide-and-seek a fight a love triangle etc The participants were asked todecide by pushing a button if all three of the shapes were lsquofri-endsrsquo or not Half of the films were intended to have lsquoall friendsrsquoas the correct answer (correct answer was determined by a con-sensus-rating process among the developers of the tasks witheach final film version judged to have a clear answer) The filmswere scripted such that any adversarial interactions occurred inthe final few seconds of the film to force the participant toattend throughout to derive the correct answer In creating ourcontrol task for contrast to the SAT we reasoned that each SATfilm requires three important processes
(i) monitoring the movements and physical interactionsbetween the shapes
(ii) pretending that the shapes represent something else iepeople and
(iii) an inferential social reasoning process based on the natureof the interactions (judging whether the movements rep-resent friendly or non-friendly interactions)
A lsquobumper carrsquo control task was created that contained all ofthe elements of the SAT films with the exception of the socialreasoning process This task also entailed eight 151 s filmsdepicting the same geometric shapes moving about a black field
Phil Trans R Soc Lond B (2003)
with the same centrally positioned white box The participantrsquostask was to pretend that these figures were lsquobumper carsrsquomdashsmallracing cars found at amusement parks that are encapsulated byrubber bumpers to allow safe playful collisions Participantsmonitored the carrsquos movements and interactions and decidedon the basis of the carrsquos trajectories and speed after each collisionif the three shapes were all equally lsquoheavyrsquo or not for exampleupon collision if one car shot off more rapidly than anotherthen the two were not equally heavy Key collisions that gaveaway the correct answer always occurred in the final seconds ofthe film Collisions early in the 151 s skit were often mere grazesthat failed to provide definitive information about relative massThus the control task contained the first two elements butinstead of a social decision participants were required to makea decision about a physical property The bumper car and SATfilms were designed to be equivalent with respect to movementquantity and location so that the comparison between the twotasks would reveal the location of brain processes that are dis-tinctly involved in social perceptual and social cognitive pro-cesses It is interesting to note that we piloted a version of thecontrol task that involved physical judgements on the SAT films(as opposed to the bumper car films) but participants reportedthat they were not able to consciously stop seeing the films associal stories Thus it did not seem possible to use the exactsame stimuli in both tasks as might otherwise be desirablebecause social perceptual and cognitive processes would prob-ably be engaged to a greater or lesser extent in both the experi-mental and control tasks Two other lower level controlconditions were also included in each experimental run in theblock design and were intended to further pull apart the threedistinct processes outlined above However the results of thesecontrasts were generally uninformative and thus are not reportedhere Between each film was a 12 s rest period with a blackscreen All participants underwent practice using films that didnot make the final group of eight in order to become completelyfamiliar with the tasks before fMRI scanning During the fMRIexperiment each film was preceded by a 3 s cue lsquoBUMPERCARS SAME WEIGHTrsquo or lsquoPEOPLE ALL FRIENDSrsquoSubjects responded by pressing a button upon completion ofeach film both as a measure of reaction time and accuracy andto ensure that the subjects watched the entire film
In the follow-up study (hereafter lsquoStudy 2rsquo) side-by-side grey-scale faces objects or patterns were presented in asamedifferent task in a block-design experiment to localize theFFA We have previously used this task to localize the FFA ina large group of normal controls (Schultz et al 2000b 2001)Image pairs were presented for 28 s with a 05 s inter-stimulusinterval The person identity task employed same-gender pairsof neutral (non expressive) faces on a black background Pic-tures were taken from standard sources and were edited toremove hair ears and shirt collars so as to force subjects tofocus on features of the face with central relevance to non-verbalsocial communication ie the eyes nose mouth and facegeometry Objects were pictures of spectacles taken from anonline retail catalogue that were contrast inverted to make thebackground black and the spectacles shades of grey to matchthe faces Patterns were distorted versions of the faces or spec-tacles Results from contrasts with patterns were not used tolocalize the FFA and thus are not reported here As is conven-tional the face versus subordinate-level object-discriminationcontrast defined the FFA Each task block lasted 165 s and wasseparated by a 105 s rest period during which cross hairs (1 1)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
418 R T Schultz and others FFA and social cognition
Figure 1 Composite t-map for 12 healthy controls contrasting the social attribution (yellowred) and the bumper car(bluepurple) tasks (p 00005) Right and left are reversed by convention Abbreviations BA Brodman area FG fusiformgyrus IFG inferior frontal gyrus MPFC medial prefrontal cortex STG superior temporal gyrus Y-coordinates are from thesystem of Talairach amp Tournoux (1988)
centred in the same position as the image pairs flashed with thesame presentation rate
(c) Data acquisitionSAT and face perception fMRI data were collected on differ-
ent occasions averaging approximately 25 months apart (range3 weeks to 6 months) Only after the original 12 subjects com-pleted the SAT fMRI study was the decision made to rescansubjects with the face-localization protocol Changes in BOLDcontrast were measured as subjects performed the SAT bumpercar face-discrimination and object-discrimination tasks Thestimuli were run as QuickTime films in Study 1 and as PICTimage files in Study 2 Studies were programmed in PsyScope
125 PPC (Carnegie Mellon University Pittsburgh PA USA)and run on a MacIntosh G3 computer Images were back-pro-jected onto a translucent screen mounted near the end of theMRI gantry and were viewed through a periscopic prism systemon the head coil Behavioural response data were collected witha fibre-optic button box with two response alternatives (Yes orNo for lsquoall friendsrsquo lsquosame weightrsquo lsquosame personrsquo and lsquosameobjectrsquo) The participantrsquos head was immobilized using foamwedges and tape across the forehead
T2 curren weighted images sensitive to BOLD contrast wereacquired on a GE Signa 15 Tesla scanner with a standard quad-
Phil Trans R Soc Lond B (2003)
rature head coil using a gradient echo single-shot echo planarsequence and a coronal orientation perpendicular to the planethrough the ACndashPC The pulse sequence for both studies wasTR = 1500 ms TE = 60 flip angle = 60 NEX = 1 in-planevoxel size = 3125 mm acute 3125 mm In the SAT study we col-lected 14 coronal slices 10 mm thick (skip 1 mm) starting at theanterior-most aspect of the frontal lobe and covering all of thebrain except the caudal-most aspect of the occipital lobe Datawere collected in four runs of an ABCD block design(block = one 151 s film) with blocks of each type presentedtwice per run in a pseudo-random order Across runs a total of80 echo planar images were collected per slice per task con-dition In the face-perception study we also collected 14 coronalslices perpendicular to the ACndashPC starting from the posterioraspect of the occipital cortex up through the rostral-most aspectof the cingulate gyrus Slice thickness was 9 mm (skip 1 mm) tobe compatible with a separate face-perception study ongoing atthat time Data were collected in a block design with a pseudo-random order across six separate runs with three blocks of eachtask per run for 180 echo planar images per slice per task con-dition Functional data in both studies were co-registered to T1-weighted structural images of the same thickness collected in thesame session (TR = 500 TE = 14 field of view = 200 mm256 mm acute 192 mm matrix 2 NEX)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 419
(d) Data analysisData were corrected for motion using SPM99 for three trans-
lation directions and for the three possible rotations (WellcomeDepartment of Cognitive Neurology London UK) Image vol-umes with centre of mass (x y or z) movement of more than15 mm within a run were discarded Image analyses and testsof statistical significance were done using locally developedsoftware (Skudlarski httpmrimedyaleedumembersFframedhtml) Motion corrected images were spatially smoothed with aGaussian filter with a full-width half-maximum value of625 mm The specific effects of each task were evaluated bycreating t-maps for each imaging series incorporating a correc-tion for linear drift (Skudlarski et al 1999) of specific task con-trasts social versus bumper car in Study 1 and face versusobject in Study 2 t-maps were averaged across imaging seriesand co-registered with the higher resolution anatomical imagesfor display and localization These maps were then transformedby in-plane registration and slice interpolation into a pro-portional three-dimensional grid defined by Talairach amp Tour-noux (1988) and averaged across all subjects to createcomposite t-maps with the acquired data in 14 slices interp-olated to 18 slices (NB 16 slices are shown in figure 1 as fMRIactivations on the first and last slice are corrupted by motioncorrection) The SAT versus bumper car maps are displayed inthe figures using a significance level of p 00005(uncorrected) Face versus object t-maps were created and dis-played at p 005 (uncorrected) with the a priori hypothesis thatthe right lateral FG would define the FFA No other brain areasare examined in the second study thus avoiding any multiplecomparison problem
ROI analyses were conducted in the SAT study by tracingsignificant pixels on the group composite activation map (figure1) in the following regions the right FG the right and left STSand STG the right TP the right amygdala and right and leftdorsal MPFC To more thoroughly assess activity in the FGmedial and lateral FG ROIs (and the combined whole FG) weredefined anatomically and traced across the two coronal sliceswhere there was significant SAT activation The activated SATROI for the FG is 33 of the size of the entire anatomicallydefined FG at those two slices Individual subject data wereinterrogated using the ROIs to obtain the mean per cent signalchange for each person for each region and Talairach centre ofmass coordinates (the centre of ROI activation weighted by theamplitude of activation across the region) Mean per cent signal-change data were used in correlational analyses to estimate theconsistency of conjoint activity between ROIs across subjects
3 RESULTS
(a) Behavioural performanceThere were no significant differences between the
social and bumper car tasks in performance accuracy(t1 2 2 = 163 p 010 social = 86 plusmn 16 correctbumper = 74 plusmn 16 correct) or reaction time (t1 22
= 062 p 060) There were no significant differences intask accuracy between males and females or left-handersand right-handers In addition there were no significantcorrelations between task performance and age Full ScaleIntelligence Quotient or Benton Face Recognition per-formance
Phil Trans R Soc Lond B (2003)
(b) Brain activity associated with the socialattribution task
As shown in figure 1 comparison of the SAT with thebumper car control condition resulted in a widely distrib-uted set of significant activations There was very little sig-nificant activation for the bumper car task with the oneregion shown clearly in figure 1 being bilateral activationof the dorsal bank of the intra-parietal sulcus The SATnetwork included a region within the right and left dorsalMPFC the right and left inferior frontal gyrus parsorbitalis and the lateral orbital gyrus the right TP theright amygdala the right and left STS and STG and theright FG It is important to note that at lower thresholds(eg p 001) there was also left amygdala activation anda ROI analysis of the per cent signal change failed to findsignificantly more right than left amygdala activation It isalso worth noting that the FG activations seem quitespecific to the SAT task in the sense that reducing thethreshold down to p 005 failed to show additional ven-tral pathway activation More widespread activationsmight have indicated a general SAT effect on arousal orattention that was manifested throughout the ventralstream but this was not the case
The largest areas of activation were the STG (especiallyon the right) and MPFC Direct comparison of the rightversus left MPFC mean per cent signal change failed tofind significant differences (paired t1 1 = 045 p 050)However the right STG was significantly more activatedthan the left STG (paired t1 1 = 264 p = 002) Table 1presents the Talairach coordinates for the centre of acti-vation mass for each ROI Table 2 presents a correlationalmatrix showing the consistency of conjoint activitybetween regions The strongest correlation is between theright amygdala and the ROI that defines the significantlyactivated region of the right FG (r = 071 p = 001) Inter-estingly this correlation is nominally larger than that ofthe entire FG and the smaller subcomponent FG ROIdefined by the SAT activated pixels (r = 069 p = 0013)Since some correlation would be expected between theseoverlapping ROIs especially since the data were spatiallysmoothed the robust correlation to the amygdala is evenmore impressive Other notable results from the corre-lation matrix include the lack of correlation between theMPFC and either the amygdala or temporal lobe ROIsWithin the temporal lobes however the right STG is sig-nificantly correlated with the left STG and with both defi-nitions of the right FG
Accuracy of performance on the SAT correlates withthe amount of activity in the anatomically defined rightFG (r = 065 p = 002) but not with any other node in theSAT network Females showed significantly more rightSTG activation than males (t1 0 = 234 p = 004) Maleshowever showed significantly more right TP activation(t1 0 = 253 p = 003) There were no other significant sexdifferences and no significant associations with hand-edness or age
(c) Comparison of activity in the right fusiformgyrus during social attribution task and face
perceptionAt the time Study 1 data were collected finding signifi-
cant right FG activation to the SAT was unexpected Toclarify whether the FG activation was in the precise
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
420 R T Schultz and others FFA and social cognition
Table 1 ROI centres of mass coordinates
mean Talairach coordinates (X Y Z )
ROI right hemisphere left hemisphere Brodmann areas
MPFC 49 349 436 274 361 434 6 8 9TP 463 138 2128 mdash 38amygdala 228 2109 2124 mdash mdashFG (SAT) 344 2468 290 mdash 37FG (face) 374 2480 2126 mdash 37STG 507 2571 152 2565 2608 194 22 39
Table 2 ROI correlation matrix(Correlations are based on mean per cent signal change from each ROI (see sect 2 for ROI procedures) SAT accuracy data representthe percentage of films that each participant correctly)
activated whole right right left left right rightright FG FG amygdala amygdala MPFC MPFC left STG STG right TP
whole right FG 069 curren curren curren
right amygdala 071 curren curren curren 032left amygdala 056 curren 019 070 curren curren curren
left MPFC 021 013 032 011right MPFC 2018 2008 021 019 048left STG 021 029 019 005 056 curren 018right STG 054 curren 069 curren curren curren 022 025 009 2016 060 curren curren
right TP 2007 2011 040 033 048 034 038 2010SAT accuracy 045 065 curren curren 2012 009 013 2017 201 022 2034
curren curren curren p lt 001 (r gt 069) curren curren p lt 005 (r gt 057) curren p lt 010
(a) SAT versus bumper car FFA activation
(
(c) enlargement and alignment of FFA
Figure 2 (a) Composite (n = 12) t-map at two slices showing significant (p 00005) activation for the SAT contrast(yellowred) with the bumper car control task (bluepurple) (b) Composite (n = 9) t-map at two slices showing significant(p 005) activation for the face (yellowred) versus object discrimination (bluepurple) This contrast defines the FFA (c)Subregions of composite t-maps shown in (a) and (b) are enlarged and aligned to demonstrate the overlap of activation in theFG for the SAT and face discrimination activations Subscripts 1 and 2 refer to the first (more anterior) and second coronalslices with significant activation
location of the FFA we compared it with the result of theface versus object discrimination contrast (the standardmeans of identifying the FFA in the literature) Both theFFA and the SAT activations of the FG were confined totwo coronal slices in highly overlapping locations Thesetwo sets of group composite maps are shown in figure 2
Phil Trans R Soc Lond B (2003)
The FG activations were stronger in the SAT versusbumper contrast than the face versus object contrast butthis may have had as much do with the baseline as theexperimental tasks The Talairach coordinates for theSAT and FFA activations show that their centre of massdiffers by less than one voxel The SAT activation is 3 mm
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 421
Figure 3 Scans of two individuals across three different occasions showing the reproducibility of FFA activations at two time-points and relationship to SAT activations (andashc) are from a 23-year-old male (dndashf ) are from a 24-year-old female Panels arearranged chronologically (ad) The first face versus object experiment (be) The second face versus object scan (cf ) SATversus bumper car contrast Both coronal slices are shown where there was FFA (t gt 15 in yellowred) or SAT activation(t gt 30 in yellowred) Arrows point to FG activity (right and left are reversed by convention) As in the group results (figure2) the SAT activation is centred slightly more medially along the MFS Left FG activation shown in these two subjects doesnot survive thresholding in the group composite (figure 2) Control tasks (object discrimination bumper car) are shown inpurpleblue
more medial than that of the FFA straddling the MFSthat delineates the lateral and medial aspects of the FGThe FFA on the other hand is clearly positioned in thelateral FG as expected (Haxby et al 1999) The SATactivation is also centred 36 mm more superior and about1 mm more anteriorly than the FFA A count of the over-lapping significant pixels showed that 50 of the SATactivation falls within the FFA This provides a goodapproximation of how these regions overlap Howeverthere is no definitive way to measure the percent overlapin this study because it would change with the use of dif-ferent control tasks in either condition or different signifi-cance levels for thresholding the t-maps
Two of the participants (one male one female) werealso part of a reproducibility study of the FFA and hadboth undergone the face discrimination protocol on twooccasions Figure 3 presents t-maps of their FG for eachface perception study and the SAT The female partici-pant shows reversed asymmetry as sometimes happenswith the left FG showing greater face activation than theright Nevertheless her SAT activations track her FFAand are more left-sided than typical These results showthat the FFA activation is reproducible so that the lessthan perfect overlap between the FFA and the SAT FGactivation is probably not a measurement or reliabilityissue
4 DISCUSSION
(a) The social brain networkThe current study required participants to observe the
movements of geometric figures and to interpret thesewith regard to a conceptual template about what consti-tutes a friendly or unfriendly social interaction It requiredclose attention to the contingent nature of a sequence ofmovements and inferences about mental states of eachcharacter to explain their actions Perception of the move-
Phil Trans R Soc Lond B (2003)
ments of these simple shapes as wilful seems to be auto-matic and effortless for healthy controls but not forpersons with autism (Klin 2000) As shown in figure 1the network engaged by the social attribution process (incontrast to the physical attribution control task) includednearly all of the brain areas implicated by past researchon the social brain (Brothers 1990) including cognitiveaspects such as theory of mind (Castelli et al 2000) aswell as perceptual aspects such as the perception of socialdisplays and biological movement (Allison et al 2000)We found significant activation of the bilateral MPFCsuperior STG and STS and inferior FG pars orbitalisextending into the lateral orbital gyri In addition therewere significant activations on the right side only for theamygdala TP and the FG The predominance of right-side activations is consistent with the notion that the righthemisphere is more concerned than the left with socialprocesses (Siegal et al 1996 Winner et al 2002) Ourresults differ from some past research by finding signifi-cant right FG activation and by failing to find orbito-MPFC activation
Several earlier neuroimaging studies have shown thatthe dorsal MPFC (ie that cortex anterior and superior tothe anterior cingulate gyrus) is a critical substrate for socialjudgements including empathizing and thinking aboutotherrsquos thoughts and intentions (Fletcher et al 1995 Goelet al 1995 Happe et al 1996 Castelli et al 2000 2002Gallagher et al 2000) Our findings are consistent withthese imaging studies and with non-human primate stud-ies that have documented social failures and loss of socialposition within the group following lesions to orbital pre-frontal cortices and MPFC (Butter et al 1969 Myers etal 1973 Bachevalier amp Mishkin 1986) The importantrole for the MPFC in social cognition is further suggestedby studies of autism spectrum disorders that find func-tional abnormalities in this area (Happe et al 1996 Ernstet al 1997 Haznedar et al 1997 Castelli et al 2002)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
422 R T Schultz and others FFA and social cognition
Gusnard et al (2001) suggest that the dorsal MPFC isinvolved in any kind of thought that uses the self as a refer-ent Thus the SAT activations in this area may have beendriven by theorizing about othersrsquo minds but with explicitreference to the participantrsquos own frame of reference as tohow they would feel in a similar situation
The orbital prefrontal cortex and to a lesser extent thedorsal MPFC have dense reciprocal connections withmedial temporal areas (Price et al 1996 Carmichael ampPrice 1995) providing the anatomical bases for a systemthat regulates emotional processes Damasio and col-leagues (Damasio et al 1990 Bechara et al 1996) haveargued that the orbito-MPFC have a primary function ofintegrating information about rewards and punishmentsto bias future behaviour (Rolls 1995 Dias et al 1996Hornak et al 1996 Lane et al 1997 Reiman et al 1997)A functional circuitry such as this would seem especiallyimportant in the development and acquisition of socialbehaviour However even acquired lesions to theseregions in adulthood can result in abnormalities of socialconduct (Damasio et al 1990) Brothers (1990) high-lighted the orbital prefrontal cortex as one of the threeprincipal brain regions involved in social cognition Weobserved poor signal in this region (these areas are notori-ously prone to fMRI signal drop-out and distortion) andthus we cannot know if the orbital prefrontal region wasengaged by the SAT or not However using a similarpsychological task but with PET where signal acquisitionin this region is not degraded Castelli et al (2000 2002)failed to find activation of the orbital prefrontal cortexCollectively these results call into question the impor-tance of the orbital prefrontal cortex in social cognitionand instead shift the focus on anterior cortices toward thedorsal MPFC
The SAT also generated robust activations of the rightamygdala and nearby cortex of the right TP The amyg-dala is often given a central role in theories of social per-ception and cognition (Brothers 1990 Bachevalier 1994Adolphs et al 1998 Baron-Cohen et al 2000 Schultz etal 2000b) The amygdala has a critical role in emotionalarousal assigning significance to environmental stimuliand mediating the formation of visual-reward associationsthat is lsquoemotionalrsquo learning (Gaffan et al 1988 LeDoux1996 Anderson amp Phelps 2001) It is reliably engagedduring judgements of personality characteristics from pic-tures of the face or part of the face (Adolphs et al 1998Baron-Cohen et al 1999 Winston et al 2002) Activationof the amygdala appears to be automatic and stimulusdriven as it can be engaged by images of facial expressionsin conscious awareness as well as by subliminal presen-tations of faces displaying affect (Morris et al 1998Whalen et al 1998 Critchley et al 2000) Thus theamygdalarsquos engagement by the SAT could stem from thegeneral emotional arousal evoked by the animations or itcould represent its computational role in some more spe-cific social perceptual process
The amygdala has dense reciprocal connections withthe ventral visual processing stream (Amaral amp Price1984) The strong correlation observed in this studybetween the right amygdala and the right FG could indi-cate that emotional inputs from the amygdala to the FGare necessary for engaging the social computational pro-cesses of the FG Brothers (1995) has speculated that the
Phil Trans R Soc Lond B (2003)
amygdala generates lsquosocial feelingsrsquo that are of importancein cutting through the complexities of social situations andguiding behaviour by simpler learned contingenciesSocial events are complex because their meaning oftencomes from specific combinations of features that do notadd up in a linear manner making it more difficult todecompose the events by cognitive analysis Effectivesocial engagement requires integration of context historicalrelationships and current socialndashemotional communi-cations expressed through prosodic tone as well as facialexpressions posture and gesture They are made morecomplex by the rapid pace of social transactions Thiswould necessitate some fast response system based ongeneral principles from past socialndashemotional experiencesa role for which the amygdala would seem ideally suitedIn other words the amygdala might drive intuition or lsquogutfeelingsrsquo to guide rapid non-verbal social interactionsinvolving facial expressions gesture etc Thus the strongamygdalandashFG correlation observed here could be inter-preted as the amygdala informing the FG of the relevanceof a social event and also of the outcome of its quick anddirty social perceptual analyses thereby guiding the FG(and other social nodes) in their finer grained compu-tations
Perhaps the largest and strongest activations to the SATwere those of the posterior aspects of the STS spreadinginto the adjacent STG This area has been implicated asa specific site for perception of biological motion (Bondaet al 1996 Allison et al 2000 Grezes et al 2001 Vainaet al 2001) Biological motion is a broad construct thatseems to encompass the perception of static images ofevents that could move or did move such as facialexpressions as well as actual movement by animateobjects This region of the STSndashSTG is also critical forthe decoding visual displays of social action or intention(eg gaze direction gesture and facial displays of emotion)(Critchley et al 2000 Hoffman amp Haxby 2000) In thisregard the SAT activations here are completely expectedand a testament to the effectiveness of these animationsin inducing the desired illusion of anthropomorphicaction The majority of the STSndashSTG activations wereanterior to location of the V5MT as defined in otherrecent fMRI studies (Culham et al 2001) but they alsoextended posteriorly into these more general movementsensitive cortices Unlike the amygdala some evidencesuggests that activity of the STS is mediated by explicitattention to social characteristics of the face and that STSengagement it is not automatic or stimulus driven(Winston et al 2002) We surmise therefore that acti-vation of the STS by the SAT was due to explicit taskinstructions to judge the social interactions
(b) Role of the fusiform face area in socialcognition
We also found significant SAT activation of the rightFG When compared directly in a subset of participants totheir FFA gleaned from a separate scanning session witha perceptual discrimination task of person versus objectidentity we found that both ROIs were confined to theright side on the same two coronal slices There wasabout 50 overlap between the SAT FG and FFA acti-vations and the centre of the two activations differed byless than 1 voxel The FFA activation was offset to the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 423
lateral prominence of the FG whereas the SAT activateda region of the FG that was closer to the centre of the FGbut still largely in the lateral aspect Because the SAT doesnot contain any face representations the FFA engagementwas unexpected as this region is thought of as selectivefor faces (Kanwisher et al 1997) or to other classes ofcomplex objects for which one is perceptually expert(Gauthier et al 2000)
These results have important implications for the poss-ible role of the fusiform in social brain circuitry andautism Interestingly the magnitude of the FG activationpredicted SAT task accuracy no other ROI correlated sig-nificantly with task accuracy This would seem to argueagainst any interpretation that the FFA activation is incon-sequential to the SAT that it is activated simply becauseit is well connected to other areas that are directly involvedin the social attribution process Using a task that is simi-lar to the SAT Castelli et al (2000) also reported signifi-cant right FG activation using PET However in theirfollow-up PET study comparing autism and normal con-trols (Castelli et al 2002) they failed to find significantFG activation in either group The failure of their secondstudy to find right FG activations may be due to the morestringent random effects model used in the data analysesHowever because we find right FG activation with fMRIwhich is more sensitive than PET and because Castelli etal (2000) found it with what amounts to a lower thres-hold it seems quite probable that the effect is real In factwe have preliminary evidence from an ongoing fMRIstudy that reproduces the SAT FG activations in healthycontrols (Schultz et al 2001) In addition there may besome task attributes that differ between our studies andCastelli et alrsquos that might impact into the strength withwhich FG computational processes are evoked
The key question then is why is the FG engaged bythe SAT and what role does the right FG have in socialcognition and perception Currently there are three com-peting theories of the functional organization of the FGand related ventral visual perceptual areas One modelput forth by Haxby Chao Martin and colleagues speci-fies that objects are encoded in a distributed fashion acrossa wide expanse of the ventral temporal-occipital cortex(Haxby et al 2001) They call this the lsquoobject formtypologyrsquo model (Ishai et al 1999) Their data indicatethat object category specificity is achieved by unique spa-tial patterns of activation across this extrastriate visual cor-tex They show that the pattern of activations across thiscortex is diagnostic of object category membership moreso than any localized activation maxima (Haxby et al2001) They also argue that object category perceptioninvolves retrieval of category-related information aboutspecific features and attributes of the object category(Chao et al 2002) They admit that the lsquonature of theinformation about objects that is represented in the ventraltemporal cortex is a great puzzlersquo (Haxby et al 2000 p4) but open the possibility that semantic information maybe important
A second view championed by Kanwisher et al holdsthat several select perceptual categories including facesbodies and spatial layout of places are encoded in highlyspecific locations in a modular fashion (Kanwisher et al1997 Epstein amp Kanwisher 1998 Downing et al 2001Spiridon amp Kanwisher 2002) The FFA is thought to be
Phil Trans R Soc Lond B (2003)
most concerned with discriminating among individualidentities and not with discriminating images at the categ-orical level In fact lesions to the FFA cause a specificdeficit in recognizing individual identities but not in reco-gnizing the general category of face versus non-face(Wada amp Yamamoto 2001) Spiridon amp Kanwisher (2002)provide a partial replication of Haxby et alrsquos (2001) objectform typology model They show that patterns of ventralvisual cortical activation can distinguish object categoriesNevertheless their data show category specificity for selectareas such as the FFA and they argue the ventral visualcortex is not equipotential
Gauthier et al however have argued that the ventraloccipital temporal pathway is organized by the nature ofthe perceptual computations and that these processingbiases are acquired through experience (Gauthier et al2000 Tarr amp Gauthier 2000) For example the parahip-pocampal place area is a function of a bias towards pro-cessing landscapes in terms of their spatial layout becausewe have learned through repeated experiences that this isvery useful information to extract when perceiving land-scapes and related visual images Similarly we learn quiteearly in life that it is important to discriminate faces on anindividual level for example discriminating mother fromothers This bias towards individual identification accord-ing to Gauthier accounts for the FFArsquos apparent modu-larity (Gauthier et al 1997 2000 Tarr amp Gauthier 2000)According to this third model the type of informationneeded and our cumulative experience in processing thatinformation organizes the ventral occipitalndashtemporal path-way into regional centres with preferred modes of pro-cessing
All three models seem to agree that the functionalorganization of the ventral visual cortices is driven by theneed to categorize perceptions into object classes Webelieve that our findings showing FFA activation by non-face objects are consistent with aspects of both the pro-cessing map model of Gauthier and the object formtypology model of Haxby et al First consistent withHaxby et al we suggest that semantic information isimportant to the ventral visual pathway for object categor-ization Second consistent with Gauthier et al we believethat repeated perceptual experience with faces biases thetype of information that FG finds important We proposethat the middle FG area encodes semantic attributes ofpeople because of repeated perceptual experiences withfaces that occur during social interactions In fact wewould guess that most perceptual experiences with facesoccur during social situations and these social situationsoften involve repeated social judgements Thus infor-mation about the social nature of people might be storedin the FG (though not exclusively)
The nature of the semantic information stored in theFG might be restricted to anything that would be helpfulin defining faces as a distinct category of object becausemaking such distinctions appears to be the primary chargeof the ventral visual pathway This would include knowl-edge of people as having personal agency and of havingthe capability to disturb each otherrsquos emotional homeo-stasis (eg to act friendly or unfriendly) By pretendingthat the three shapes in the SAT are people stored knowl-edge about people in social interactions might beretrieved causing the observed activations in the FG
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
424 R T Schultz and others FFA and social cognition
There would be a measure of efficiency from thisarrangement that is having representations and compu-tations of more abstract attributes of people inter-digitatedwith front-end perceptual processes about physical attri-butes Gauthierrsquos claim is that the same group of neuronscan be engaged by perceptually different categories ofobjects it is the type of processing rather than the visualdetails that are important Extrapolating from this wewould argue that the SAT engages a region of the FGoverlapping with the FFA because it demands compu-tational processes to classify the SAT geometric figures aspeople or person-like Chao et al (1999) reach a similarconclusion They argued that activity in the ventral path-way reflects stored information about an object categorynot just physical features They point out that thisarrangement could explain why lesion patients with cate-gory specific perceptual deficits also have trouble retriev-ing general information about that visual category Thuswe would add that the FFA must store general infor-mation about people or some meta-representation oflsquopeoplenessrsquo
(c) Implications for the pathobiology of autismThe SAT used in this current study appears to be an
effective neurobehavioural probe for engaging a distrib-uted network of brain regions involved in different aspectsof social perception and cognition It will be important touse this and similar procedures in persons with autism tobetter define the nature of brain functions in this disorderCastelli et al (2002) have already taken the first step inthis process and they describe a pattern of hypoactivationin the MPFC STS and TPs in autism In an ongoingstudy of autism spectrum disorders we have presentedpreliminary data using the SAT showing that we too findhypoactivation of these regions and in the amygdala andFG (Schultz et al 2001) Thus we predict that futurework in this area will show that the entire social brain net-work is underactive in autism during tasks requiring socialperceptual and social cognitive processing
There are already sufficient data to argue strongly for arole of the FG in the pathobiology of autism No less thanfive previous fMRI studies have shown the FFA region tobe significantly less engaged among persons with autismcompared with controls during face perception tasks(Critchley et al 2000 Dierks et al 2001 Pierce et al2001 Schultz et al 2000a 2001) Activation of the FGby the social judgements in the current study adds animportant piece of evidence in favour of a causal role forthe FG in the pathobiology of autism This conclusionhowever must remain tentative until additional studiesmore precisely define the factors leading to FG activationduring social attribution and gather better data to proveits computational role in social cognition In addition theeffect of FG lesions for social functioning must be clari-fied If the FG is involved in social computations onewould expect to find social cognitive deficits in personswith lesions to this area However social deficits in proso-pagnosics have not been widely reported It might be thatprosopagnosic patients have not been carefully tested onthis dimension In this regard using our behavioural ver-sion of the SAT (Klin 2000) to test social perceptionamong prosopagnosics would be quite interesting Theparallels however between autism and prosopagnosia are
Phil Trans R Soc Lond B (2003)
incomplete because autism is clearly a developmental dis-order whereas the research literature on prosopagnosia ismostly confined to lesions acquired in adolescence oradulthood Among the few cases of developmental proso-pagnosia reported in the literature there is indeed one thathighlights severe social impairments (Kracke 1994) Itmay be that the role of the FG in the development ofautistic symptoms is different from the role of the FG inthe maintenance of social cognitive functions after brainmaturity
It is also possible that the role of the FG in social pro-cesses is dependent on its functional relationships withother nodes in the social brain and that it is the collectiveaction and interaction of the network that is of primaryimportance for social behaviours For example we founda strong correlation across participants in the amount ofFG and amygdala activation A strictly modular view ofthese areas may be inappropriate as the functions of eachnode could be quite dependent on one another and whenonly considered in isolation quite insufficient to supportsocial processes Thus whereas a lesion to the FG mayimpair visual perception it might not have a large impacton social functioning if this depends on an extended net-work that is dynamic and capable of compensatory adjust-ments Dysfunction of the FG may be necessary but notsufficient to produce social deficits Indeed our relianceon modular models of brain functioning may be leadingus astray in our search for causal mechanisms in autismInstead it may be the collective action of a distributedsystem that is critical to the pathobiology of autism TheFG region may be a key partner in this distributed systembut nevertheless just one node and insufficient by itselfto support in any substantive manner social cognitive pro-cesses or to explain social cognitive deficits seen inautism
5 CONCLUSION AND FUTURE DIRECTIONS
Using the SAT we isolated a distributed network ofactivations that conform to the emerging model of thesocial brain Most important we found significant acti-vation of the central aspect of the FG thus adding thisregion to the expanding list of structures involved in socialprocesses The FG region activated by the SAT overlapsin its spatial extent with the FFA with centres of massdiffering by less than 1 voxel We speculate that these SATactivations represent computational processes associatedwith more abstract attributes of people
It is an open question as to whether the substantial over-lap between the FFA and SAT FG activation is due to anoverlapping or shared set of neuronal assemblies This willneed to be clarified by future work Electrophysiologicalstudies show that there are small face-specific patches inthe FG cortex (Allison et al 1999) In the current studysmall patches of face cells and SAT cells could be inter-mingled but distinct and not drawing on any of the sameneuronal assemblies The spatial resolution of fMRI haslimitations that may preclude any definitive answer to thisquestion but there are strategies that can be used toaddress the issue For example the current study designdid not contain SAT and face discriminations in the sameexperimental fMRI series Doing so would enable directcomparisons of the computational demands placed on the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 425
common area of the FFA It also would be informative iffollow-up studies superimposed faces within the geometricfigures of the SAT without changing the film scripts inany way We could then determine if the computations ofthe common area of the FFA increase in a predictablefashionmdashwould the activation be a linear summation ofthe original SAT plus face discrimination Any significantdeviation from an additive model would suggest that thereis some sharing of neuronal assemblies with the SAT andface discrimination tasks when presented alone
This work was supported by grants from the National Institutesof Child Health and Human Development (grants PO1 HD03008 and PO1 HDDC35482) and the Korczak FoundationThe authors thank the anonymous reviewers for their helpfulcomments Michael Lee for programming the fMRI tasksHedy Serofin and Terry Hickey for their assistance in acquiringthe fMRI data John Herrington for his valuable assistance indata analyses of preliminary studies and members of the YaleDevelopmental Neuroimaging Program for their helpful com-ments on this manuscript A special acknowledgement ofappreciation is given to Donald Cohen who passed away inOctober of 2001 for his outstanding mentorship in all aspectsof this program of work
REFERENCES
Abell F Happe F amp Frith U 2000 Do triangles play tricksAttribution of mental states to animated shapes in normaland abnormal development J Cogn Dev 15 1ndash20
Adolphs R Tranel D amp Damasio A R 1998 The humanamygdala in social judgement Nature 393 470ndash474
Allison T Puce A Spencer D D amp McCarthy G 1999Electrophysiological studies of human face perception Ipotentials generated in occipitotemporal cortex by face andnon-face stimuli Cerebr Cortex 9 415ndash430
Allison T Puce A amp McCarthy G 2000 Social perceptionfrom visual cues role of the STS region Trends Cogn Sci4 267ndash278
Amaral D G amp Price J L 1984 Amygdalo-cortical projec-tions in the monkey (Macaca fascicularis) J Comp Neurol230 465ndash496
Amaral D G Price J L Pitkanen A amp Carmichael S T1992 Anatomical organization of the primate amygdaloidcomplex In The amygdala neurobiological aspects of emotionmemory and mental dysfunction (ed J Aggleton) pp 1ndash66New York Wiley-Liss
Anderson A K amp Phelps E A 2001 Lesions of the humanamygdala impair enhanced perception of emotionally salientevents Nature 411 305ndash309
Bachevalier J 1994 Medial temporal lobe structures andautism a review of clinical and experimental findingsNeuropsychologia 32 627ndash648
Bachevalier J amp Mishkin M 1986 Visual recognition impair-ment follows ventromedial but not dorsolateral prefrontallesions in monkeys Behav Brain Res 20 249ndash261
Baron-Cohen S Ring H A Wheelwright S BullmoreE T Brammer M J Simmons A amp Williams S C R1999 Social intelligence in the normal and autistic brain anfMRI study Eur J Neurosci 11 1891ndash1898
Baron-Cohen S Ring H A Bullmore E T WheelwrightS Ashwina C amp Williams S C R 2000 The amygdalatheory of autism Neurosci Biobehav Rev 24 355ndash364
Bechara A Tranel D Damasio H amp Damasio A R 1996Failure to respond autonomically to anticipated future out-comes following damage to prefrontal cortex Cerebr Cortex6 215ndash225
Benton A 1994 Face recognition Los Angeles CA WesternPsychological Services
Phil Trans R Soc Lond B (2003)
Bonda E Petrides M Ostry D amp Evans A 1996 Specificinvolvement of human parietal systems and the amygdala inthe perception of biological motion J Neurosci 16 3737ndash3744
Bowler D M amp Thommen E 2000 Attribution of mechan-ical and social causality to animated displays by childrenwith autism Autism 4 147ndash171
Brothers L 1990 The social brain a project for integratingprimate behavior and neurophysiology in a new domainConcepts Neurosci 1 27ndash151
Brothers L 1995 Neurophysiology of the perception of inten-tion by primates In The cognitive neurosciences 2nd edn (edM S Gazzaniga) pp 1107ndash1115 Cambridge MA MITPress
Butter C M McDonald J A amp Snyder D R 1969 Oralitypreference behavior and reinforcement value of nonfoodobject in monkeys with orbital frontal lesions Science 1641306ndash1307
Carmichael S T amp Price J L 1995 Limbic connections ofthe orbital and medial prefrontal cortex in macaque monk-eys J Comp Neurol 363 615ndash641
Castelli F Happe F Frith U amp Frith C 2000 Movementand mind a functional imaging study of perception andinterpretation of complex intentional movement patternsNeuroImage 12 314ndash325
Castelli F Frith C Happe F amp Frith U 2002 AutismAsperger syndrome and brain mechanisms for the attri-bution of mental states to animated shapes Brain 1251839ndash1849
Chao L L Haxby J V amp Martin A 1999 Attribute-basedneural substrates in posterior temporal cortex for perceivingand knowing about objects Nat Neurosci 2 913ndash919
Chao L L Weisberg J amp Martin A 2002 Experience-dependent modulation of category-related cortical activityCerebr Cortex 12 545ndash551
Critchley H D (and 10 others) 2000 The functional neuroan-atomy of social behaviour changes in cerebral blood flowwhen people with autistic disorder process facial expressionsBrain 123 2203ndash2212
Culham J C Cavanagh P amp Kanwisher N G 2001 Atten-tion response functions characterizing brain areas usingfMRI activation during parametric variations of attentionalload Neuron 32 737ndash745
Damasio A R Tranel D amp Damasio H 1990 Individualswith sociopathic behavior caused by frontal damage fail torespond autonomically to social stimuli Behav Brain Res41 81ndash94
Dierks T Bolte S Huble D Lanfermannm H amp PoustkaF 2001 Alterations of face processing strategies in autism(a fMRI study) Presented at the 6th Annual Meeting of theOrganization for Human Brain Mapping Brighton UK 10ndash14 June 2001
Dias R Robbins T W amp Roberts A C 1996 Dissociationin prefrontal cortex of affective and attentional shifts Nature380 69ndash72
Downing P E Jiang Y Shuman M amp Kanwisher N 2001A cortical area selective for visual processing of the humanbody Science 293 2470ndash2473
Epstein R amp Kanwisher N 1998 A cortical representation ofthe local visual environment Nature 392 598ndash601
Ernst M Zametkin A J Matochik J A Pascualvaca D ampCohen R M 1997 Reduced medial prefrontal dopami-nergic activity in autistic children Lancet 350 638
Fletcher P C Happe F Frith U Baker S C DolanR J Frackowiak R S amp Frith C D 1995 Other minds inthe brain a functional imaging study of lsquotheory of mindrsquo instory comprehension Cognition 57 109ndash128
Frith C D amp Frith U 1999 Interacting minds a biologicalbasis Science 286 1692ndash1695
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
426 R T Schultz and others FFA and social cognition
Fujita I Tanaka K Ito M amp Cheng K 1992 Columns forvisual features of objects in monkey inferotemporal cortexNature 360 343ndash346
Gaffan E A Gaffan D amp Harrison S 1988 Disconnectionof the amygdala from visual association cortex impairs vis-ual-reward association learning in monkeys J Neurosci 83144ndash3150
Gallagher H L Happe F Brunswick N Fletcher P CFrith U amp Frith C D 2000 Reading the mind in cartoonsand stories an fMRI study of lsquotheory of mindrsquo in verbal andnonverbal tasks Neuropsychologia 38 11ndash21
Gauthier I Anderson A W Tarr M J Skudlarski P ampGore J C 1997 Levels of categorization in visual recog-nition studied using functional magnetic resonance imagingCurr Biol 7 645ndash651
Gauthier I Tarr M J Anderson A W Skudlarski P ampGore J C 1999 Activation of the middle fusiform face areaincreases with expertise in recognizing novel objects NatureNeurosci 2 568ndash573
Gauthier I Skudlarski P Gore J C amp Anderson A W2000 Expertise for cars and birds recruits brain areasinvolved in face recognition Nature Neurosci 3 191ndash197
Goel V Grafman J Sadato N amp Hallett M 1995 Mode-ling other minds Neuroreport 6 1741ndash1746
Grelotti D Gauthier I amp Schultz R T 2001 Social interestand the development of cortical face specialization whatautism teaches us about face processing Devl Psychobiol 40213ndash225
Grezes J Fonlupt P Bertenthal B amp Delon-Martin C2001 Does perception of biological motion rely on specificbrain regions NeuroImage 13 775ndash785
Gusnard D A Akbudak E Shulman G L amp RaichleM E 2001 Medial prefrontal cortex and self-referentialmental activity relation to a default mode of brain functionProc Natl Acad Sci USA 98 4259ndash4264
Happe F Ehlers S Fletcher P Frith U Johansson MGillberg C Dolan R Frackowiak R amp Frith C 1996lsquoTheory of mindrsquo in the brain Evidence from a PET scanstudy of Asperger syndrome Neuroreport 8 197ndash201
Haxby J V Ungerleider L G Clark V P Schouten J LHoffman E A amp Martin A 1999 The effect of face inver-sion on activity in human neural systems for face and objectperception Neuron 22 189ndash199
Haxby J V Isahai A Chao L L Ungerleider L G ampMartin A 2000 Object-form topology in the ventral tem-poral lobe response to I Gauthier (2000) Trends Cogn Sci4 3ndash4
Haxby J V Gobbini M I Furey M L Ishai ASchouten J L amp Pietrini P 2001 Distributed and overlap-ping representations of faces and objects in ventral temporalcortex Science 293 2425ndash2430
Haznedar M M Buchsbaum M S Metzger M Solim-ando A Spiegel-Cohen J amp Hollander E 1997 Anteriorcingulate gyrus volume and glucose metabolism in autisticdisorder Am J Psychiat 154 1047ndash1050
Heider F amp Simmel M 1944 An experimental study ofapparent behavior Am J Psychol 57 243ndash259
Hobson R P Ouston J amp Lee A 1988a Whatrsquos in a faceThe case of autism Br J Psychol 79 441ndash453
Hobson R P Ouston J amp Lee A 1988b Emotion recog-nition in autism coordinating faces and voices Psychol Med18 911ndash923
Hoffman E A amp Haxby J V 2000 Distinct representationsof eye gaze and identity in the distributed human neural sys-tem for face perception Nature Neurosci 3 80ndash84
Hornak J Rolls E T amp Wade D 1996 Face and voiceexpression identification in patients with emotional andbehavioural changes following ventral frontal lobe damageNeuropsychologia 34 247ndash261
Phil Trans R Soc Lond B (2003)
Ishai A Ungerleider L G Martin A Schouten J L ampHaxby J V 1999 Distributed representation of objects inthe human ventral visual pathway Proc Natl Acad Sci USA96 9379ndash9384
Kanwisher N 2000 Domain specificity in face perceptionNature Neurosci 3 759ndash763
Kanwisher N McDermott J amp Chun M M 1997 The fusi-form face area a module of extrastriate cortex specializedfor face perception J Neurosci 17 4302ndash4311
Klin A 2000 Attributing social meaning to ambiguous visualstimuli in higher functioning autism and Asperger syndromethe social attribution task J Child Psychol Psychiat 41831ndash846
Klin A Sparrow S S de Bildt A Cicchetti D V CohenD J amp Volkmar F R 1999 A normed study of face recog-nition in autism and related disorders J Autism Devl Dis-orders 29 499ndash508
Klin A Jones W Schultz R T Volkmar F amp Cohen D J2002 Visual fixation patterns during viewing of naturalisticsocial situations as predictors of social competence in indi-viduals with autism Arch Gen Psychiat 59 809ndash816
Kracke I 1994 Developmental prosopagnosia in Aspergersyndrome presentation and discussion of an individual caseDevl Med Child Neurol 36 873ndash886
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J 1997 Neuroanatomical correlates of happi-ness sadness and disgust Am J Psychiat 154 926ndash933
Langdell T 1978 Recognition of faces an approach for thestudy of autism J Child Psychol Psychiat 19 255ndash268
LeDoux J E 1996 The emotional brain New York Simon ampShuster
Lowel S amp Singer W 1992 Selection of intrinsic horizontalconnections in the visual cortex by correlated neuronalactivity Science 255 209ndash212
Macdonald H Rutter M Howlin P Rios P Le ConteurA Evered C amp Folstein S 1989 Recognition andexpression of emotional cues by autistic and normal adultsJ Child Psychol Psychiat 30 865ndash877
Morris J S Ohman A amp Dolan R J 1998 Conscious andunconscious emotional learning in the human amygdalaNature 393 467ndash470
Myers R E Swett C amp Miller M 1973 Loss of social groupaffinity following prefrontal lesions in free-rangingmacaques Brain Res 64 257ndash269
Osterling J amp Dawson G 1994 Early recognition of childrenwith autism a study of first birthday home video tapes JAutism Devl Disorders 24 247ndash257
Pierce K Muller R A Ambrose J Allen G amp Cour-chesne E 2001 Face processing occurs outside the fusiformlsquoface arearsquo in autism evidence from functional MRI Brain124 2059ndash2073
Price J L Carmichael S T amp Drevets W C 1996 Net-works related to the orbital and medial prefrontal cortex asubstrate for emotional behavior Prog Brain Res 107523ndash536
Puce A Allison T Gore J C amp McCarthy G 1995 Face-sensitive regions in human extrastriate cortex studied byfunctional MRI J Neurophysiol 74 1192ndash1199
Reiman E M Lane R D Ahern G L Schwartz G EDavidson R J Friston K J Yun L S amp Chen K 1997Neuroanatomical correlates of externally and internally gen-erated human emotion Am J Psychiat 154 918ndash925
Rolls E T 1995 A theory of emotion and consciousness andits application to understanding the neural basis of emotionIn The cognitive neurosciences (ed M S Gazzaniga) pp1091ndash1106 Cambridge MA MIT Press
Schultz R T Gauthier I Klin A Fulbright R K Ander-son A W Volkmar F Skudlarski P Lacadie C CohenD J amp Gore J C 2000a Abnormal ventral temporal
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from

Published online 21 January 2003
The role of the fusiform face area in social cognitionimplications for the pathobiology of autism
Robert T Schultz12 David J Grelotti1 Ami Klin1 Jamie Kleinman3Christiaan Van der Gaag4 Rene Marois5 and Pawel Skudlarski2
1Child Study Center Yale University School of Medicine 230 S Frontage Road New Haven CT 06520-7900 USA2Department of Diagnostic Radiology Yale University School of Medicine 333 Cedar Street New Haven CT 06510 USA
3Department of Psychology University of Connecticut Unit 1020 Storrs CT 06269-1020 USA4Academic Centre for Child and Adolescent Psychiatry PO Box 660 9700 AR Groningen The Netherlands
5Department of Psychology Vanderbilt University 111 21st Avenue Nashville TN 37203 USA
A region in the lateral aspect of the fusiform gyrus (FG) is more engaged by human faces than any othercategory of image It has come to be known as the lsquofusiform face arearsquo (FFA) The origin and extent ofthis specialization is currently a topic of great interest and debate This is of special relevance to autismbecause recent studies have shown that the FFA is hypoactive to faces in this disorder In two linkedfunctional magnetic resonance imaging (fMRI) studies of healthy young adults we show here that theFFA is engaged by a social attribution task (SAT) involving perception of human-like interactions amongthree simple geometric shapes The amygdala temporal pole medial prefrontal cortex inferolateral frontalcortex and superior temporal sulci were also significantly engaged Activation of the FFA to a task withoutfaces challenges the received view that the FFA is restricted in its activities to the perception of faces Wespeculate that abstract semantic information associated with faces is encoded in the FG region andretrieved for social computations From this perspective the literature on hypoactivation of the FFA inautism may be interpreted as a reflection of a core social cognitive mechanism underlying the disorder
Keywords amygdala autism fusiform face area medial prefrontal cortex social cognitionsuperior temporal sulcus
1 INTRODUCTION
For the first time the field of autism has a replicatedneurofunctional marker of the disordermdashhypoactivation ofthe FFA The FFA is that region of the middle aspect ofthe right FG that is selectively engaged by faces (whencontrasted with object perception tasks) (Puce et al 1995Kanwisher et al 1997 Kanwisher 2000) Anatomicallythe middle portion of the FG is split along its rostralndashcaudal extent by a shallow MFS In fMRI the centre ofactivation in face perception tasks is typically offsettowards the lateral aspect of the FG in the right hemi-sphere (Haxby et al 1999) Whereas individual subjectsmay or may not also show left FG activation during faceperception group composites always show right-side acti-vations to be larger At least five fMRI studies have shownthat older children adolescents and adults with autismspectrum disorders have reduced levels of activity toimages of the human face in this specialized face regionof the right hemisphere (Critchley et al 2000 Dierks etal 2001 Pierce et al 2001 Schultz et al 2000a 2001)These data are consistent with an older and more exten-sive psychology literature documenting performance defi-cits in face perception (Langdell 1978 Klin et al 1999)
Author for correspondence (robertschultzyaleedu)
One contribution of 14 to a Theme Issue lsquoAutism mind and brainrsquo
Phil Trans R Soc Lond B (2003) 358 415ndash427 415 Oacute 2003 The Royal SocietyDOI 101098rstb20021208
and facial expression recognition in autism (eg Hobsonet al 1988ab MacDonald et al 1989 Yirmiya et al1992) They seem to provide an important clue as to theneural ontogeny and pathobiology of autism
Whereas the consistency of these findings is encour-aging what it means to have an under-responsive FFAremains unclear Our initial interpretation of this findingfocused on the role of experience for shaping the visualcortices (Schultz et al 2000ab Grelotti et al 2001) It isknown that the ventral temporal visual areas are quiteplastic and can be moulded by early experiences (Gaffanet al 1988 Webster et al 1991 Fujita et al 1992 Lowel ampSinger 1992) Persons with autism pay much less attentionto the face (Osterling amp Dawson 1994 Klin et al 2002)and this may be why they fail to acquire normal perceptualskill in this domain Inadequate attention to faces duringcritical periods of cortical development should affect thematuration of these areas and presumably lead to unde-ractivation of the FFA during face perception
This interpretation fits nicely into the perceptual expert-ise model of the FFA first championed by Gauthier andcolleagues (Gauthier et al 1999 2000) Gauthier hasshown in two elegant fMRI studies that the FFAresponds preferentially to any class of object for which aperson is perceptually lsquoexpertrsquo For example she foundthat bird experts engage the FFA more strongly whenviewing birds than cars but the reverse is true for carexperts (Gauthier et al 2000) Moreover normal young
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
416 R T Schultz and others FFA and social cognition
adults can enhance their FFA activity to a class of novelobjects through extensive perceptual training (Gauthier etal 1999) Interpreting the hypoactivation of the FFA inautism from an expertise model however argues that thisfinding is an outcome of having autism rather than partof the cause In other words the hypoactivation of theFFA is merely a reflection of the social disability the cul-mination of a set of developmental experiences acrossmany years whereby the person has reduced interest inother people and pays inadequate attention to their facesIn this regard the under-responsiveness of the FFA is abiological marker Identifying an endophenotype such asthis is extremely important and takes the field one signifi-cant step closer to understanding the underlying biologicalmechanisms but it falls short of providing a snapshot ofthe brain mechanisms that actually cause autism
An alternative view would be that the FFA is a corecomponent of the lsquosocial brainrsquo Data emerging overrecent years from neuroimaging studies human lesionstudies and animal studies suggest a working model of thesocial brain that comprises a diverse set of frontal limbicand temporal lobe circuitry Select aspects of the orbitaland medial prefrontal cortices the amygdala and lateralaspects of the temporal cortex involving the STS haveeach been implicated in social functioning (Brothers 1990Fletcher et al 1995 Baron-Cohen et al 1999 Frith ampFrith 1999 Allison et al 2000 Castelli et al 2000 2002Schultz et al 2000b) The frontal and temporal corticeshave dense and often reciprocal connections to theamygdala (Carmichael amp Price 1995 Price et al 1996)The amygdala is centrally positioned and capable of mod-ulating and interpreting the emotional significance of dataprocessed in the perceptual cortices as well as assistingwith the integration of emotion and cognition for decisionmaking and action in the frontal cortices (Amaral et al1992 Schultz et al 2000b) Collectively this systemdefines a heuristic model of the social brain with the pre-cise functions of each node only understood in an as yetsuperficial manner
But is there a role for the FFA in this social circuitryWhereas the role of the FG in face perception is undis-puted only one prior study has implicated the FFA insocial cognition Castelli et al (2000) used social ani-mations involving interacting geometric shapes to probethe social brain These animations were based on theclassic study of Heider amp Simmel (1944) that showed howcertain movements by inanimate objects could stronglyand automatically suggest personal agency and that agroup of interacting geometric forms will naturally suggestsocial interactions All but one of the 34 female collegestudents in Heider and Simmelrsquos study described the ani-mations through a social lens and in human terms (egshapes chase one another fight entrap play get fright-ened elated etc) It seems that the contingent nature ofthe shape movements and the fact that their movementsviolate the rules of simple physics (ie the shapes seem tohave lsquoagencyrsquo or will) naturally invoke social cognitive andsocial perceptual ideation Using PET in six healthy youngadults Castelli et al showed that interpreting this type ofanimation engaged the medial prefrontal cortices the TPthe STS and the right FG Although the localization of thefusiform activations in their study is in the area generallyreported to be the FFA it is not clear whether this region
Phil Trans R Soc Lond B (2003)
of activation would have overlapped with the FFA in thesesubjects as location of the FFA can vary from person toperson Nevertheless engagement of the right FG by asocial cognitive task that does not involve images of theface suggests that this FG region may have a broadermore important set of functions extending beyond simpleface perception As such it provides a basis to argue thatthe hypoactivation of the FFA to faces in autism might beilluminating part of a causal mechanism as opposed to adevelopmental consequence of having autism
The current study also used an adaptation of the pro-cedure of Heider amp Simmel (1944) involving what Klin(2000) called the SAT Klin (2000) used the SAT to showhow persons with autism fail to spontaneously imposesocial meaning on these types of movements Other inves-tigators have also reported a paucity of theory of mindideation among persons with autism to other renditions ofthe HeiderndashSimmel procedure (Abell et al 2000Bowler amp Thommen 2000) Use of simple shapes to dis-play human social interactions without perceptual rep-resentations of real people strips the social event down tothe essential elements needed to convey social meaningsIn this regard the SAT is an ideal neuroimaging probefor assessing social cognitive and social perceptual pro-cesses in a way that is not confounded by perceptual pro-cesses that would be provoked if actual images of faces orpeople were used The original SAT from Klin (2000) isa 50 s silent film in which three moving geometric shapes(a circle a small triangle and a larger triangle) interactwith each other in a social manner Interestingly in Klinrsquosstudy some attributions by those with an autism spectrumdisorder were given in terms of physical meaning (egmagnetic forces) not social meaning We took note of thisobservation in developing the following fMRI experi-ments and created a control task for the SAT that involvesjudgements of object mass
In two fMRI studies of the same group of normal con-trol subjects we show that the FG is robustly engaged byan adaptation of the SAT suitable for a block design fMRIstudy Engagement of the right FG by non-face stimulisuggests that this region has functions beyond static faceperception To test the exact location of the fusiform acti-vations during the SAT nine of the 12 participants con-sented to return for a fMRI study of face perceptionResults from this second study found the location of theFFA to be highly overlapping with the FG activations tothe SAT Thus making social judgements on non-facegeometric figures and making identity judgements ongrey-scale pictures of human faces draws upon a similarneural substrate in the FG This result challenges thespecificity of the middle portion of the FG for faces andraises the possibility that the FFA is part of the primarycircuitry for social cognition As such it has importantimplications for the hypoactivation of the FFA in autismand more generally for specifying a distributed social net-work whose dysfunction might cause autism
2 METHODS
(a) ParticipantsTwelve participants were recruited for this study from the staff
and student populations at Yale University The sampleincluded six men (three left-handed) and six women (one left-
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 417
handed) ranging in age from 20 to 31 years (meanplusmn sd = 242 plusmn 31) Participants were screened for neurologicaland DSM IV Axis I psychiatric disorders Estimated full-scaleIQ as measured by four subtests of the Wechsler Adult Intelli-gence Scale 3rd edition (Wechsler 1997) averaged 1288(plusmn 104) (Information Vocabulary Picture Completion andBlock Design) All subjects scored in the normal range on theBenton Test of Facial Recognition (Benton 1994) (raw scorerange 41ndash50 mean plusmn sd = 468 plusmn 26) There were no signifi-cant differences between the sexes on any of these variables
Nine of the twelve agreed to participate in a follow-up fMRIstudy of face perception conducted so that localization of theFFA could be compared with SAT activations in the middle FGarea in the right hemisphere Five were male (one left-handed)and four were female (one left-handed) mean age was236 plusmn 26 years All subjects gave written informed consent forboth studies in accordance with procedures and protocolsapproved by the Institutional Review Board of the Yale Univer-sity School of Medicine
(b) Experimental tasksWe adapted the SAT for use in a fMRI block-design study by
programming 16 new SAT QuickTime film skits using Directorpublished by Macromedia (600 Townsend Ave San FranciscoCA wwwmacromediacom) From these a final set of eightwere chosen for use in the fMRI study on the basis of ratingsby project staff of the filmrsquos realism and ability to capture onersquossocial attention (these films can be downloaded from httpinfomedyaleeduchldstdyneuroimgsatFmovieshtm) Each filmlasted 151 s and was designed like the original SAT with move-ments intended to suggest a sense of personal agency andreciprocal and contingent interactions that were meant to be eas-ily interpreted as social Each film contained three types of whitegeometric figures (a triangle diamond and circle) that movedagainst a black background In common with the original SATthere was a box in the centre of the field with one wall thatopened as if on a hinge allowing the shapes to open and shutthe door and to enter chase or drag other shapes inside Eachfilm was scripted to follow a social story for example hide-and-seek a fight a love triangle etc The participants were asked todecide by pushing a button if all three of the shapes were lsquofri-endsrsquo or not Half of the films were intended to have lsquoall friendsrsquoas the correct answer (correct answer was determined by a con-sensus-rating process among the developers of the tasks witheach final film version judged to have a clear answer) The filmswere scripted such that any adversarial interactions occurred inthe final few seconds of the film to force the participant toattend throughout to derive the correct answer In creating ourcontrol task for contrast to the SAT we reasoned that each SATfilm requires three important processes
(i) monitoring the movements and physical interactionsbetween the shapes
(ii) pretending that the shapes represent something else iepeople and
(iii) an inferential social reasoning process based on the natureof the interactions (judging whether the movements rep-resent friendly or non-friendly interactions)
A lsquobumper carrsquo control task was created that contained all ofthe elements of the SAT films with the exception of the socialreasoning process This task also entailed eight 151 s filmsdepicting the same geometric shapes moving about a black field
Phil Trans R Soc Lond B (2003)
with the same centrally positioned white box The participantrsquostask was to pretend that these figures were lsquobumper carsrsquomdashsmallracing cars found at amusement parks that are encapsulated byrubber bumpers to allow safe playful collisions Participantsmonitored the carrsquos movements and interactions and decidedon the basis of the carrsquos trajectories and speed after each collisionif the three shapes were all equally lsquoheavyrsquo or not for exampleupon collision if one car shot off more rapidly than anotherthen the two were not equally heavy Key collisions that gaveaway the correct answer always occurred in the final seconds ofthe film Collisions early in the 151 s skit were often mere grazesthat failed to provide definitive information about relative massThus the control task contained the first two elements butinstead of a social decision participants were required to makea decision about a physical property The bumper car and SATfilms were designed to be equivalent with respect to movementquantity and location so that the comparison between the twotasks would reveal the location of brain processes that are dis-tinctly involved in social perceptual and social cognitive pro-cesses It is interesting to note that we piloted a version of thecontrol task that involved physical judgements on the SAT films(as opposed to the bumper car films) but participants reportedthat they were not able to consciously stop seeing the films associal stories Thus it did not seem possible to use the exactsame stimuli in both tasks as might otherwise be desirablebecause social perceptual and cognitive processes would prob-ably be engaged to a greater or lesser extent in both the experi-mental and control tasks Two other lower level controlconditions were also included in each experimental run in theblock design and were intended to further pull apart the threedistinct processes outlined above However the results of thesecontrasts were generally uninformative and thus are not reportedhere Between each film was a 12 s rest period with a blackscreen All participants underwent practice using films that didnot make the final group of eight in order to become completelyfamiliar with the tasks before fMRI scanning During the fMRIexperiment each film was preceded by a 3 s cue lsquoBUMPERCARS SAME WEIGHTrsquo or lsquoPEOPLE ALL FRIENDSrsquoSubjects responded by pressing a button upon completion ofeach film both as a measure of reaction time and accuracy andto ensure that the subjects watched the entire film
In the follow-up study (hereafter lsquoStudy 2rsquo) side-by-side grey-scale faces objects or patterns were presented in asamedifferent task in a block-design experiment to localize theFFA We have previously used this task to localize the FFA ina large group of normal controls (Schultz et al 2000b 2001)Image pairs were presented for 28 s with a 05 s inter-stimulusinterval The person identity task employed same-gender pairsof neutral (non expressive) faces on a black background Pic-tures were taken from standard sources and were edited toremove hair ears and shirt collars so as to force subjects tofocus on features of the face with central relevance to non-verbalsocial communication ie the eyes nose mouth and facegeometry Objects were pictures of spectacles taken from anonline retail catalogue that were contrast inverted to make thebackground black and the spectacles shades of grey to matchthe faces Patterns were distorted versions of the faces or spec-tacles Results from contrasts with patterns were not used tolocalize the FFA and thus are not reported here As is conven-tional the face versus subordinate-level object-discriminationcontrast defined the FFA Each task block lasted 165 s and wasseparated by a 105 s rest period during which cross hairs (1 1)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
418 R T Schultz and others FFA and social cognition
Figure 1 Composite t-map for 12 healthy controls contrasting the social attribution (yellowred) and the bumper car(bluepurple) tasks (p 00005) Right and left are reversed by convention Abbreviations BA Brodman area FG fusiformgyrus IFG inferior frontal gyrus MPFC medial prefrontal cortex STG superior temporal gyrus Y-coordinates are from thesystem of Talairach amp Tournoux (1988)
centred in the same position as the image pairs flashed with thesame presentation rate
(c) Data acquisitionSAT and face perception fMRI data were collected on differ-
ent occasions averaging approximately 25 months apart (range3 weeks to 6 months) Only after the original 12 subjects com-pleted the SAT fMRI study was the decision made to rescansubjects with the face-localization protocol Changes in BOLDcontrast were measured as subjects performed the SAT bumpercar face-discrimination and object-discrimination tasks Thestimuli were run as QuickTime films in Study 1 and as PICTimage files in Study 2 Studies were programmed in PsyScope
125 PPC (Carnegie Mellon University Pittsburgh PA USA)and run on a MacIntosh G3 computer Images were back-pro-jected onto a translucent screen mounted near the end of theMRI gantry and were viewed through a periscopic prism systemon the head coil Behavioural response data were collected witha fibre-optic button box with two response alternatives (Yes orNo for lsquoall friendsrsquo lsquosame weightrsquo lsquosame personrsquo and lsquosameobjectrsquo) The participantrsquos head was immobilized using foamwedges and tape across the forehead
T2 curren weighted images sensitive to BOLD contrast wereacquired on a GE Signa 15 Tesla scanner with a standard quad-
Phil Trans R Soc Lond B (2003)
rature head coil using a gradient echo single-shot echo planarsequence and a coronal orientation perpendicular to the planethrough the ACndashPC The pulse sequence for both studies wasTR = 1500 ms TE = 60 flip angle = 60 NEX = 1 in-planevoxel size = 3125 mm acute 3125 mm In the SAT study we col-lected 14 coronal slices 10 mm thick (skip 1 mm) starting at theanterior-most aspect of the frontal lobe and covering all of thebrain except the caudal-most aspect of the occipital lobe Datawere collected in four runs of an ABCD block design(block = one 151 s film) with blocks of each type presentedtwice per run in a pseudo-random order Across runs a total of80 echo planar images were collected per slice per task con-dition In the face-perception study we also collected 14 coronalslices perpendicular to the ACndashPC starting from the posterioraspect of the occipital cortex up through the rostral-most aspectof the cingulate gyrus Slice thickness was 9 mm (skip 1 mm) tobe compatible with a separate face-perception study ongoing atthat time Data were collected in a block design with a pseudo-random order across six separate runs with three blocks of eachtask per run for 180 echo planar images per slice per task con-dition Functional data in both studies were co-registered to T1-weighted structural images of the same thickness collected in thesame session (TR = 500 TE = 14 field of view = 200 mm256 mm acute 192 mm matrix 2 NEX)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 419
(d) Data analysisData were corrected for motion using SPM99 for three trans-
lation directions and for the three possible rotations (WellcomeDepartment of Cognitive Neurology London UK) Image vol-umes with centre of mass (x y or z) movement of more than15 mm within a run were discarded Image analyses and testsof statistical significance were done using locally developedsoftware (Skudlarski httpmrimedyaleedumembersFframedhtml) Motion corrected images were spatially smoothed with aGaussian filter with a full-width half-maximum value of625 mm The specific effects of each task were evaluated bycreating t-maps for each imaging series incorporating a correc-tion for linear drift (Skudlarski et al 1999) of specific task con-trasts social versus bumper car in Study 1 and face versusobject in Study 2 t-maps were averaged across imaging seriesand co-registered with the higher resolution anatomical imagesfor display and localization These maps were then transformedby in-plane registration and slice interpolation into a pro-portional three-dimensional grid defined by Talairach amp Tour-noux (1988) and averaged across all subjects to createcomposite t-maps with the acquired data in 14 slices interp-olated to 18 slices (NB 16 slices are shown in figure 1 as fMRIactivations on the first and last slice are corrupted by motioncorrection) The SAT versus bumper car maps are displayed inthe figures using a significance level of p 00005(uncorrected) Face versus object t-maps were created and dis-played at p 005 (uncorrected) with the a priori hypothesis thatthe right lateral FG would define the FFA No other brain areasare examined in the second study thus avoiding any multiplecomparison problem
ROI analyses were conducted in the SAT study by tracingsignificant pixels on the group composite activation map (figure1) in the following regions the right FG the right and left STSand STG the right TP the right amygdala and right and leftdorsal MPFC To more thoroughly assess activity in the FGmedial and lateral FG ROIs (and the combined whole FG) weredefined anatomically and traced across the two coronal sliceswhere there was significant SAT activation The activated SATROI for the FG is 33 of the size of the entire anatomicallydefined FG at those two slices Individual subject data wereinterrogated using the ROIs to obtain the mean per cent signalchange for each person for each region and Talairach centre ofmass coordinates (the centre of ROI activation weighted by theamplitude of activation across the region) Mean per cent signal-change data were used in correlational analyses to estimate theconsistency of conjoint activity between ROIs across subjects
3 RESULTS
(a) Behavioural performanceThere were no significant differences between the
social and bumper car tasks in performance accuracy(t1 2 2 = 163 p 010 social = 86 plusmn 16 correctbumper = 74 plusmn 16 correct) or reaction time (t1 22
= 062 p 060) There were no significant differences intask accuracy between males and females or left-handersand right-handers In addition there were no significantcorrelations between task performance and age Full ScaleIntelligence Quotient or Benton Face Recognition per-formance
Phil Trans R Soc Lond B (2003)
(b) Brain activity associated with the socialattribution task
As shown in figure 1 comparison of the SAT with thebumper car control condition resulted in a widely distrib-uted set of significant activations There was very little sig-nificant activation for the bumper car task with the oneregion shown clearly in figure 1 being bilateral activationof the dorsal bank of the intra-parietal sulcus The SATnetwork included a region within the right and left dorsalMPFC the right and left inferior frontal gyrus parsorbitalis and the lateral orbital gyrus the right TP theright amygdala the right and left STS and STG and theright FG It is important to note that at lower thresholds(eg p 001) there was also left amygdala activation anda ROI analysis of the per cent signal change failed to findsignificantly more right than left amygdala activation It isalso worth noting that the FG activations seem quitespecific to the SAT task in the sense that reducing thethreshold down to p 005 failed to show additional ven-tral pathway activation More widespread activationsmight have indicated a general SAT effect on arousal orattention that was manifested throughout the ventralstream but this was not the case
The largest areas of activation were the STG (especiallyon the right) and MPFC Direct comparison of the rightversus left MPFC mean per cent signal change failed tofind significant differences (paired t1 1 = 045 p 050)However the right STG was significantly more activatedthan the left STG (paired t1 1 = 264 p = 002) Table 1presents the Talairach coordinates for the centre of acti-vation mass for each ROI Table 2 presents a correlationalmatrix showing the consistency of conjoint activitybetween regions The strongest correlation is between theright amygdala and the ROI that defines the significantlyactivated region of the right FG (r = 071 p = 001) Inter-estingly this correlation is nominally larger than that ofthe entire FG and the smaller subcomponent FG ROIdefined by the SAT activated pixels (r = 069 p = 0013)Since some correlation would be expected between theseoverlapping ROIs especially since the data were spatiallysmoothed the robust correlation to the amygdala is evenmore impressive Other notable results from the corre-lation matrix include the lack of correlation between theMPFC and either the amygdala or temporal lobe ROIsWithin the temporal lobes however the right STG is sig-nificantly correlated with the left STG and with both defi-nitions of the right FG
Accuracy of performance on the SAT correlates withthe amount of activity in the anatomically defined rightFG (r = 065 p = 002) but not with any other node in theSAT network Females showed significantly more rightSTG activation than males (t1 0 = 234 p = 004) Maleshowever showed significantly more right TP activation(t1 0 = 253 p = 003) There were no other significant sexdifferences and no significant associations with hand-edness or age
(c) Comparison of activity in the right fusiformgyrus during social attribution task and face
perceptionAt the time Study 1 data were collected finding signifi-
cant right FG activation to the SAT was unexpected Toclarify whether the FG activation was in the precise
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
420 R T Schultz and others FFA and social cognition
Table 1 ROI centres of mass coordinates
mean Talairach coordinates (X Y Z )
ROI right hemisphere left hemisphere Brodmann areas
MPFC 49 349 436 274 361 434 6 8 9TP 463 138 2128 mdash 38amygdala 228 2109 2124 mdash mdashFG (SAT) 344 2468 290 mdash 37FG (face) 374 2480 2126 mdash 37STG 507 2571 152 2565 2608 194 22 39
Table 2 ROI correlation matrix(Correlations are based on mean per cent signal change from each ROI (see sect 2 for ROI procedures) SAT accuracy data representthe percentage of films that each participant correctly)
activated whole right right left left right rightright FG FG amygdala amygdala MPFC MPFC left STG STG right TP
whole right FG 069 curren curren curren
right amygdala 071 curren curren curren 032left amygdala 056 curren 019 070 curren curren curren
left MPFC 021 013 032 011right MPFC 2018 2008 021 019 048left STG 021 029 019 005 056 curren 018right STG 054 curren 069 curren curren curren 022 025 009 2016 060 curren curren
right TP 2007 2011 040 033 048 034 038 2010SAT accuracy 045 065 curren curren 2012 009 013 2017 201 022 2034
curren curren curren p lt 001 (r gt 069) curren curren p lt 005 (r gt 057) curren p lt 010
(a) SAT versus bumper car FFA activation
(
(c) enlargement and alignment of FFA
Figure 2 (a) Composite (n = 12) t-map at two slices showing significant (p 00005) activation for the SAT contrast(yellowred) with the bumper car control task (bluepurple) (b) Composite (n = 9) t-map at two slices showing significant(p 005) activation for the face (yellowred) versus object discrimination (bluepurple) This contrast defines the FFA (c)Subregions of composite t-maps shown in (a) and (b) are enlarged and aligned to demonstrate the overlap of activation in theFG for the SAT and face discrimination activations Subscripts 1 and 2 refer to the first (more anterior) and second coronalslices with significant activation
location of the FFA we compared it with the result of theface versus object discrimination contrast (the standardmeans of identifying the FFA in the literature) Both theFFA and the SAT activations of the FG were confined totwo coronal slices in highly overlapping locations Thesetwo sets of group composite maps are shown in figure 2
Phil Trans R Soc Lond B (2003)
The FG activations were stronger in the SAT versusbumper contrast than the face versus object contrast butthis may have had as much do with the baseline as theexperimental tasks The Talairach coordinates for theSAT and FFA activations show that their centre of massdiffers by less than one voxel The SAT activation is 3 mm
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 421
Figure 3 Scans of two individuals across three different occasions showing the reproducibility of FFA activations at two time-points and relationship to SAT activations (andashc) are from a 23-year-old male (dndashf ) are from a 24-year-old female Panels arearranged chronologically (ad) The first face versus object experiment (be) The second face versus object scan (cf ) SATversus bumper car contrast Both coronal slices are shown where there was FFA (t gt 15 in yellowred) or SAT activation(t gt 30 in yellowred) Arrows point to FG activity (right and left are reversed by convention) As in the group results (figure2) the SAT activation is centred slightly more medially along the MFS Left FG activation shown in these two subjects doesnot survive thresholding in the group composite (figure 2) Control tasks (object discrimination bumper car) are shown inpurpleblue
more medial than that of the FFA straddling the MFSthat delineates the lateral and medial aspects of the FGThe FFA on the other hand is clearly positioned in thelateral FG as expected (Haxby et al 1999) The SATactivation is also centred 36 mm more superior and about1 mm more anteriorly than the FFA A count of the over-lapping significant pixels showed that 50 of the SATactivation falls within the FFA This provides a goodapproximation of how these regions overlap Howeverthere is no definitive way to measure the percent overlapin this study because it would change with the use of dif-ferent control tasks in either condition or different signifi-cance levels for thresholding the t-maps
Two of the participants (one male one female) werealso part of a reproducibility study of the FFA and hadboth undergone the face discrimination protocol on twooccasions Figure 3 presents t-maps of their FG for eachface perception study and the SAT The female partici-pant shows reversed asymmetry as sometimes happenswith the left FG showing greater face activation than theright Nevertheless her SAT activations track her FFAand are more left-sided than typical These results showthat the FFA activation is reproducible so that the lessthan perfect overlap between the FFA and the SAT FGactivation is probably not a measurement or reliabilityissue
4 DISCUSSION
(a) The social brain networkThe current study required participants to observe the
movements of geometric figures and to interpret thesewith regard to a conceptual template about what consti-tutes a friendly or unfriendly social interaction It requiredclose attention to the contingent nature of a sequence ofmovements and inferences about mental states of eachcharacter to explain their actions Perception of the move-
Phil Trans R Soc Lond B (2003)
ments of these simple shapes as wilful seems to be auto-matic and effortless for healthy controls but not forpersons with autism (Klin 2000) As shown in figure 1the network engaged by the social attribution process (incontrast to the physical attribution control task) includednearly all of the brain areas implicated by past researchon the social brain (Brothers 1990) including cognitiveaspects such as theory of mind (Castelli et al 2000) aswell as perceptual aspects such as the perception of socialdisplays and biological movement (Allison et al 2000)We found significant activation of the bilateral MPFCsuperior STG and STS and inferior FG pars orbitalisextending into the lateral orbital gyri In addition therewere significant activations on the right side only for theamygdala TP and the FG The predominance of right-side activations is consistent with the notion that the righthemisphere is more concerned than the left with socialprocesses (Siegal et al 1996 Winner et al 2002) Ourresults differ from some past research by finding signifi-cant right FG activation and by failing to find orbito-MPFC activation
Several earlier neuroimaging studies have shown thatthe dorsal MPFC (ie that cortex anterior and superior tothe anterior cingulate gyrus) is a critical substrate for socialjudgements including empathizing and thinking aboutotherrsquos thoughts and intentions (Fletcher et al 1995 Goelet al 1995 Happe et al 1996 Castelli et al 2000 2002Gallagher et al 2000) Our findings are consistent withthese imaging studies and with non-human primate stud-ies that have documented social failures and loss of socialposition within the group following lesions to orbital pre-frontal cortices and MPFC (Butter et al 1969 Myers etal 1973 Bachevalier amp Mishkin 1986) The importantrole for the MPFC in social cognition is further suggestedby studies of autism spectrum disorders that find func-tional abnormalities in this area (Happe et al 1996 Ernstet al 1997 Haznedar et al 1997 Castelli et al 2002)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
422 R T Schultz and others FFA and social cognition
Gusnard et al (2001) suggest that the dorsal MPFC isinvolved in any kind of thought that uses the self as a refer-ent Thus the SAT activations in this area may have beendriven by theorizing about othersrsquo minds but with explicitreference to the participantrsquos own frame of reference as tohow they would feel in a similar situation
The orbital prefrontal cortex and to a lesser extent thedorsal MPFC have dense reciprocal connections withmedial temporal areas (Price et al 1996 Carmichael ampPrice 1995) providing the anatomical bases for a systemthat regulates emotional processes Damasio and col-leagues (Damasio et al 1990 Bechara et al 1996) haveargued that the orbito-MPFC have a primary function ofintegrating information about rewards and punishmentsto bias future behaviour (Rolls 1995 Dias et al 1996Hornak et al 1996 Lane et al 1997 Reiman et al 1997)A functional circuitry such as this would seem especiallyimportant in the development and acquisition of socialbehaviour However even acquired lesions to theseregions in adulthood can result in abnormalities of socialconduct (Damasio et al 1990) Brothers (1990) high-lighted the orbital prefrontal cortex as one of the threeprincipal brain regions involved in social cognition Weobserved poor signal in this region (these areas are notori-ously prone to fMRI signal drop-out and distortion) andthus we cannot know if the orbital prefrontal region wasengaged by the SAT or not However using a similarpsychological task but with PET where signal acquisitionin this region is not degraded Castelli et al (2000 2002)failed to find activation of the orbital prefrontal cortexCollectively these results call into question the impor-tance of the orbital prefrontal cortex in social cognitionand instead shift the focus on anterior cortices toward thedorsal MPFC
The SAT also generated robust activations of the rightamygdala and nearby cortex of the right TP The amyg-dala is often given a central role in theories of social per-ception and cognition (Brothers 1990 Bachevalier 1994Adolphs et al 1998 Baron-Cohen et al 2000 Schultz etal 2000b) The amygdala has a critical role in emotionalarousal assigning significance to environmental stimuliand mediating the formation of visual-reward associationsthat is lsquoemotionalrsquo learning (Gaffan et al 1988 LeDoux1996 Anderson amp Phelps 2001) It is reliably engagedduring judgements of personality characteristics from pic-tures of the face or part of the face (Adolphs et al 1998Baron-Cohen et al 1999 Winston et al 2002) Activationof the amygdala appears to be automatic and stimulusdriven as it can be engaged by images of facial expressionsin conscious awareness as well as by subliminal presen-tations of faces displaying affect (Morris et al 1998Whalen et al 1998 Critchley et al 2000) Thus theamygdalarsquos engagement by the SAT could stem from thegeneral emotional arousal evoked by the animations or itcould represent its computational role in some more spe-cific social perceptual process
The amygdala has dense reciprocal connections withthe ventral visual processing stream (Amaral amp Price1984) The strong correlation observed in this studybetween the right amygdala and the right FG could indi-cate that emotional inputs from the amygdala to the FGare necessary for engaging the social computational pro-cesses of the FG Brothers (1995) has speculated that the
Phil Trans R Soc Lond B (2003)
amygdala generates lsquosocial feelingsrsquo that are of importancein cutting through the complexities of social situations andguiding behaviour by simpler learned contingenciesSocial events are complex because their meaning oftencomes from specific combinations of features that do notadd up in a linear manner making it more difficult todecompose the events by cognitive analysis Effectivesocial engagement requires integration of context historicalrelationships and current socialndashemotional communi-cations expressed through prosodic tone as well as facialexpressions posture and gesture They are made morecomplex by the rapid pace of social transactions Thiswould necessitate some fast response system based ongeneral principles from past socialndashemotional experiencesa role for which the amygdala would seem ideally suitedIn other words the amygdala might drive intuition or lsquogutfeelingsrsquo to guide rapid non-verbal social interactionsinvolving facial expressions gesture etc Thus the strongamygdalandashFG correlation observed here could be inter-preted as the amygdala informing the FG of the relevanceof a social event and also of the outcome of its quick anddirty social perceptual analyses thereby guiding the FG(and other social nodes) in their finer grained compu-tations
Perhaps the largest and strongest activations to the SATwere those of the posterior aspects of the STS spreadinginto the adjacent STG This area has been implicated asa specific site for perception of biological motion (Bondaet al 1996 Allison et al 2000 Grezes et al 2001 Vainaet al 2001) Biological motion is a broad construct thatseems to encompass the perception of static images ofevents that could move or did move such as facialexpressions as well as actual movement by animateobjects This region of the STSndashSTG is also critical forthe decoding visual displays of social action or intention(eg gaze direction gesture and facial displays of emotion)(Critchley et al 2000 Hoffman amp Haxby 2000) In thisregard the SAT activations here are completely expectedand a testament to the effectiveness of these animationsin inducing the desired illusion of anthropomorphicaction The majority of the STSndashSTG activations wereanterior to location of the V5MT as defined in otherrecent fMRI studies (Culham et al 2001) but they alsoextended posteriorly into these more general movementsensitive cortices Unlike the amygdala some evidencesuggests that activity of the STS is mediated by explicitattention to social characteristics of the face and that STSengagement it is not automatic or stimulus driven(Winston et al 2002) We surmise therefore that acti-vation of the STS by the SAT was due to explicit taskinstructions to judge the social interactions
(b) Role of the fusiform face area in socialcognition
We also found significant SAT activation of the rightFG When compared directly in a subset of participants totheir FFA gleaned from a separate scanning session witha perceptual discrimination task of person versus objectidentity we found that both ROIs were confined to theright side on the same two coronal slices There wasabout 50 overlap between the SAT FG and FFA acti-vations and the centre of the two activations differed byless than 1 voxel The FFA activation was offset to the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 423
lateral prominence of the FG whereas the SAT activateda region of the FG that was closer to the centre of the FGbut still largely in the lateral aspect Because the SAT doesnot contain any face representations the FFA engagementwas unexpected as this region is thought of as selectivefor faces (Kanwisher et al 1997) or to other classes ofcomplex objects for which one is perceptually expert(Gauthier et al 2000)
These results have important implications for the poss-ible role of the fusiform in social brain circuitry andautism Interestingly the magnitude of the FG activationpredicted SAT task accuracy no other ROI correlated sig-nificantly with task accuracy This would seem to argueagainst any interpretation that the FFA activation is incon-sequential to the SAT that it is activated simply becauseit is well connected to other areas that are directly involvedin the social attribution process Using a task that is simi-lar to the SAT Castelli et al (2000) also reported signifi-cant right FG activation using PET However in theirfollow-up PET study comparing autism and normal con-trols (Castelli et al 2002) they failed to find significantFG activation in either group The failure of their secondstudy to find right FG activations may be due to the morestringent random effects model used in the data analysesHowever because we find right FG activation with fMRIwhich is more sensitive than PET and because Castelli etal (2000) found it with what amounts to a lower thres-hold it seems quite probable that the effect is real In factwe have preliminary evidence from an ongoing fMRIstudy that reproduces the SAT FG activations in healthycontrols (Schultz et al 2001) In addition there may besome task attributes that differ between our studies andCastelli et alrsquos that might impact into the strength withwhich FG computational processes are evoked
The key question then is why is the FG engaged bythe SAT and what role does the right FG have in socialcognition and perception Currently there are three com-peting theories of the functional organization of the FGand related ventral visual perceptual areas One modelput forth by Haxby Chao Martin and colleagues speci-fies that objects are encoded in a distributed fashion acrossa wide expanse of the ventral temporal-occipital cortex(Haxby et al 2001) They call this the lsquoobject formtypologyrsquo model (Ishai et al 1999) Their data indicatethat object category specificity is achieved by unique spa-tial patterns of activation across this extrastriate visual cor-tex They show that the pattern of activations across thiscortex is diagnostic of object category membership moreso than any localized activation maxima (Haxby et al2001) They also argue that object category perceptioninvolves retrieval of category-related information aboutspecific features and attributes of the object category(Chao et al 2002) They admit that the lsquonature of theinformation about objects that is represented in the ventraltemporal cortex is a great puzzlersquo (Haxby et al 2000 p4) but open the possibility that semantic information maybe important
A second view championed by Kanwisher et al holdsthat several select perceptual categories including facesbodies and spatial layout of places are encoded in highlyspecific locations in a modular fashion (Kanwisher et al1997 Epstein amp Kanwisher 1998 Downing et al 2001Spiridon amp Kanwisher 2002) The FFA is thought to be
Phil Trans R Soc Lond B (2003)
most concerned with discriminating among individualidentities and not with discriminating images at the categ-orical level In fact lesions to the FFA cause a specificdeficit in recognizing individual identities but not in reco-gnizing the general category of face versus non-face(Wada amp Yamamoto 2001) Spiridon amp Kanwisher (2002)provide a partial replication of Haxby et alrsquos (2001) objectform typology model They show that patterns of ventralvisual cortical activation can distinguish object categoriesNevertheless their data show category specificity for selectareas such as the FFA and they argue the ventral visualcortex is not equipotential
Gauthier et al however have argued that the ventraloccipital temporal pathway is organized by the nature ofthe perceptual computations and that these processingbiases are acquired through experience (Gauthier et al2000 Tarr amp Gauthier 2000) For example the parahip-pocampal place area is a function of a bias towards pro-cessing landscapes in terms of their spatial layout becausewe have learned through repeated experiences that this isvery useful information to extract when perceiving land-scapes and related visual images Similarly we learn quiteearly in life that it is important to discriminate faces on anindividual level for example discriminating mother fromothers This bias towards individual identification accord-ing to Gauthier accounts for the FFArsquos apparent modu-larity (Gauthier et al 1997 2000 Tarr amp Gauthier 2000)According to this third model the type of informationneeded and our cumulative experience in processing thatinformation organizes the ventral occipitalndashtemporal path-way into regional centres with preferred modes of pro-cessing
All three models seem to agree that the functionalorganization of the ventral visual cortices is driven by theneed to categorize perceptions into object classes Webelieve that our findings showing FFA activation by non-face objects are consistent with aspects of both the pro-cessing map model of Gauthier and the object formtypology model of Haxby et al First consistent withHaxby et al we suggest that semantic information isimportant to the ventral visual pathway for object categor-ization Second consistent with Gauthier et al we believethat repeated perceptual experience with faces biases thetype of information that FG finds important We proposethat the middle FG area encodes semantic attributes ofpeople because of repeated perceptual experiences withfaces that occur during social interactions In fact wewould guess that most perceptual experiences with facesoccur during social situations and these social situationsoften involve repeated social judgements Thus infor-mation about the social nature of people might be storedin the FG (though not exclusively)
The nature of the semantic information stored in theFG might be restricted to anything that would be helpfulin defining faces as a distinct category of object becausemaking such distinctions appears to be the primary chargeof the ventral visual pathway This would include knowl-edge of people as having personal agency and of havingthe capability to disturb each otherrsquos emotional homeo-stasis (eg to act friendly or unfriendly) By pretendingthat the three shapes in the SAT are people stored knowl-edge about people in social interactions might beretrieved causing the observed activations in the FG
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
424 R T Schultz and others FFA and social cognition
There would be a measure of efficiency from thisarrangement that is having representations and compu-tations of more abstract attributes of people inter-digitatedwith front-end perceptual processes about physical attri-butes Gauthierrsquos claim is that the same group of neuronscan be engaged by perceptually different categories ofobjects it is the type of processing rather than the visualdetails that are important Extrapolating from this wewould argue that the SAT engages a region of the FGoverlapping with the FFA because it demands compu-tational processes to classify the SAT geometric figures aspeople or person-like Chao et al (1999) reach a similarconclusion They argued that activity in the ventral path-way reflects stored information about an object categorynot just physical features They point out that thisarrangement could explain why lesion patients with cate-gory specific perceptual deficits also have trouble retriev-ing general information about that visual category Thuswe would add that the FFA must store general infor-mation about people or some meta-representation oflsquopeoplenessrsquo
(c) Implications for the pathobiology of autismThe SAT used in this current study appears to be an
effective neurobehavioural probe for engaging a distrib-uted network of brain regions involved in different aspectsof social perception and cognition It will be important touse this and similar procedures in persons with autism tobetter define the nature of brain functions in this disorderCastelli et al (2002) have already taken the first step inthis process and they describe a pattern of hypoactivationin the MPFC STS and TPs in autism In an ongoingstudy of autism spectrum disorders we have presentedpreliminary data using the SAT showing that we too findhypoactivation of these regions and in the amygdala andFG (Schultz et al 2001) Thus we predict that futurework in this area will show that the entire social brain net-work is underactive in autism during tasks requiring socialperceptual and social cognitive processing
There are already sufficient data to argue strongly for arole of the FG in the pathobiology of autism No less thanfive previous fMRI studies have shown the FFA region tobe significantly less engaged among persons with autismcompared with controls during face perception tasks(Critchley et al 2000 Dierks et al 2001 Pierce et al2001 Schultz et al 2000a 2001) Activation of the FGby the social judgements in the current study adds animportant piece of evidence in favour of a causal role forthe FG in the pathobiology of autism This conclusionhowever must remain tentative until additional studiesmore precisely define the factors leading to FG activationduring social attribution and gather better data to proveits computational role in social cognition In addition theeffect of FG lesions for social functioning must be clari-fied If the FG is involved in social computations onewould expect to find social cognitive deficits in personswith lesions to this area However social deficits in proso-pagnosics have not been widely reported It might be thatprosopagnosic patients have not been carefully tested onthis dimension In this regard using our behavioural ver-sion of the SAT (Klin 2000) to test social perceptionamong prosopagnosics would be quite interesting Theparallels however between autism and prosopagnosia are
Phil Trans R Soc Lond B (2003)
incomplete because autism is clearly a developmental dis-order whereas the research literature on prosopagnosia ismostly confined to lesions acquired in adolescence oradulthood Among the few cases of developmental proso-pagnosia reported in the literature there is indeed one thathighlights severe social impairments (Kracke 1994) Itmay be that the role of the FG in the development ofautistic symptoms is different from the role of the FG inthe maintenance of social cognitive functions after brainmaturity
It is also possible that the role of the FG in social pro-cesses is dependent on its functional relationships withother nodes in the social brain and that it is the collectiveaction and interaction of the network that is of primaryimportance for social behaviours For example we founda strong correlation across participants in the amount ofFG and amygdala activation A strictly modular view ofthese areas may be inappropriate as the functions of eachnode could be quite dependent on one another and whenonly considered in isolation quite insufficient to supportsocial processes Thus whereas a lesion to the FG mayimpair visual perception it might not have a large impacton social functioning if this depends on an extended net-work that is dynamic and capable of compensatory adjust-ments Dysfunction of the FG may be necessary but notsufficient to produce social deficits Indeed our relianceon modular models of brain functioning may be leadingus astray in our search for causal mechanisms in autismInstead it may be the collective action of a distributedsystem that is critical to the pathobiology of autism TheFG region may be a key partner in this distributed systembut nevertheless just one node and insufficient by itselfto support in any substantive manner social cognitive pro-cesses or to explain social cognitive deficits seen inautism
5 CONCLUSION AND FUTURE DIRECTIONS
Using the SAT we isolated a distributed network ofactivations that conform to the emerging model of thesocial brain Most important we found significant acti-vation of the central aspect of the FG thus adding thisregion to the expanding list of structures involved in socialprocesses The FG region activated by the SAT overlapsin its spatial extent with the FFA with centres of massdiffering by less than 1 voxel We speculate that these SATactivations represent computational processes associatedwith more abstract attributes of people
It is an open question as to whether the substantial over-lap between the FFA and SAT FG activation is due to anoverlapping or shared set of neuronal assemblies This willneed to be clarified by future work Electrophysiologicalstudies show that there are small face-specific patches inthe FG cortex (Allison et al 1999) In the current studysmall patches of face cells and SAT cells could be inter-mingled but distinct and not drawing on any of the sameneuronal assemblies The spatial resolution of fMRI haslimitations that may preclude any definitive answer to thisquestion but there are strategies that can be used toaddress the issue For example the current study designdid not contain SAT and face discriminations in the sameexperimental fMRI series Doing so would enable directcomparisons of the computational demands placed on the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 425
common area of the FFA It also would be informative iffollow-up studies superimposed faces within the geometricfigures of the SAT without changing the film scripts inany way We could then determine if the computations ofthe common area of the FFA increase in a predictablefashionmdashwould the activation be a linear summation ofthe original SAT plus face discrimination Any significantdeviation from an additive model would suggest that thereis some sharing of neuronal assemblies with the SAT andface discrimination tasks when presented alone
This work was supported by grants from the National Institutesof Child Health and Human Development (grants PO1 HD03008 and PO1 HDDC35482) and the Korczak FoundationThe authors thank the anonymous reviewers for their helpfulcomments Michael Lee for programming the fMRI tasksHedy Serofin and Terry Hickey for their assistance in acquiringthe fMRI data John Herrington for his valuable assistance indata analyses of preliminary studies and members of the YaleDevelopmental Neuroimaging Program for their helpful com-ments on this manuscript A special acknowledgement ofappreciation is given to Donald Cohen who passed away inOctober of 2001 for his outstanding mentorship in all aspectsof this program of work
REFERENCES
Abell F Happe F amp Frith U 2000 Do triangles play tricksAttribution of mental states to animated shapes in normaland abnormal development J Cogn Dev 15 1ndash20
Adolphs R Tranel D amp Damasio A R 1998 The humanamygdala in social judgement Nature 393 470ndash474
Allison T Puce A Spencer D D amp McCarthy G 1999Electrophysiological studies of human face perception Ipotentials generated in occipitotemporal cortex by face andnon-face stimuli Cerebr Cortex 9 415ndash430
Allison T Puce A amp McCarthy G 2000 Social perceptionfrom visual cues role of the STS region Trends Cogn Sci4 267ndash278
Amaral D G amp Price J L 1984 Amygdalo-cortical projec-tions in the monkey (Macaca fascicularis) J Comp Neurol230 465ndash496
Amaral D G Price J L Pitkanen A amp Carmichael S T1992 Anatomical organization of the primate amygdaloidcomplex In The amygdala neurobiological aspects of emotionmemory and mental dysfunction (ed J Aggleton) pp 1ndash66New York Wiley-Liss
Anderson A K amp Phelps E A 2001 Lesions of the humanamygdala impair enhanced perception of emotionally salientevents Nature 411 305ndash309
Bachevalier J 1994 Medial temporal lobe structures andautism a review of clinical and experimental findingsNeuropsychologia 32 627ndash648
Bachevalier J amp Mishkin M 1986 Visual recognition impair-ment follows ventromedial but not dorsolateral prefrontallesions in monkeys Behav Brain Res 20 249ndash261
Baron-Cohen S Ring H A Wheelwright S BullmoreE T Brammer M J Simmons A amp Williams S C R1999 Social intelligence in the normal and autistic brain anfMRI study Eur J Neurosci 11 1891ndash1898
Baron-Cohen S Ring H A Bullmore E T WheelwrightS Ashwina C amp Williams S C R 2000 The amygdalatheory of autism Neurosci Biobehav Rev 24 355ndash364
Bechara A Tranel D Damasio H amp Damasio A R 1996Failure to respond autonomically to anticipated future out-comes following damage to prefrontal cortex Cerebr Cortex6 215ndash225
Benton A 1994 Face recognition Los Angeles CA WesternPsychological Services
Phil Trans R Soc Lond B (2003)
Bonda E Petrides M Ostry D amp Evans A 1996 Specificinvolvement of human parietal systems and the amygdala inthe perception of biological motion J Neurosci 16 3737ndash3744
Bowler D M amp Thommen E 2000 Attribution of mechan-ical and social causality to animated displays by childrenwith autism Autism 4 147ndash171
Brothers L 1990 The social brain a project for integratingprimate behavior and neurophysiology in a new domainConcepts Neurosci 1 27ndash151
Brothers L 1995 Neurophysiology of the perception of inten-tion by primates In The cognitive neurosciences 2nd edn (edM S Gazzaniga) pp 1107ndash1115 Cambridge MA MITPress
Butter C M McDonald J A amp Snyder D R 1969 Oralitypreference behavior and reinforcement value of nonfoodobject in monkeys with orbital frontal lesions Science 1641306ndash1307
Carmichael S T amp Price J L 1995 Limbic connections ofthe orbital and medial prefrontal cortex in macaque monk-eys J Comp Neurol 363 615ndash641
Castelli F Happe F Frith U amp Frith C 2000 Movementand mind a functional imaging study of perception andinterpretation of complex intentional movement patternsNeuroImage 12 314ndash325
Castelli F Frith C Happe F amp Frith U 2002 AutismAsperger syndrome and brain mechanisms for the attri-bution of mental states to animated shapes Brain 1251839ndash1849
Chao L L Haxby J V amp Martin A 1999 Attribute-basedneural substrates in posterior temporal cortex for perceivingand knowing about objects Nat Neurosci 2 913ndash919
Chao L L Weisberg J amp Martin A 2002 Experience-dependent modulation of category-related cortical activityCerebr Cortex 12 545ndash551
Critchley H D (and 10 others) 2000 The functional neuroan-atomy of social behaviour changes in cerebral blood flowwhen people with autistic disorder process facial expressionsBrain 123 2203ndash2212
Culham J C Cavanagh P amp Kanwisher N G 2001 Atten-tion response functions characterizing brain areas usingfMRI activation during parametric variations of attentionalload Neuron 32 737ndash745
Damasio A R Tranel D amp Damasio H 1990 Individualswith sociopathic behavior caused by frontal damage fail torespond autonomically to social stimuli Behav Brain Res41 81ndash94
Dierks T Bolte S Huble D Lanfermannm H amp PoustkaF 2001 Alterations of face processing strategies in autism(a fMRI study) Presented at the 6th Annual Meeting of theOrganization for Human Brain Mapping Brighton UK 10ndash14 June 2001
Dias R Robbins T W amp Roberts A C 1996 Dissociationin prefrontal cortex of affective and attentional shifts Nature380 69ndash72
Downing P E Jiang Y Shuman M amp Kanwisher N 2001A cortical area selective for visual processing of the humanbody Science 293 2470ndash2473
Epstein R amp Kanwisher N 1998 A cortical representation ofthe local visual environment Nature 392 598ndash601
Ernst M Zametkin A J Matochik J A Pascualvaca D ampCohen R M 1997 Reduced medial prefrontal dopami-nergic activity in autistic children Lancet 350 638
Fletcher P C Happe F Frith U Baker S C DolanR J Frackowiak R S amp Frith C D 1995 Other minds inthe brain a functional imaging study of lsquotheory of mindrsquo instory comprehension Cognition 57 109ndash128
Frith C D amp Frith U 1999 Interacting minds a biologicalbasis Science 286 1692ndash1695
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
426 R T Schultz and others FFA and social cognition
Fujita I Tanaka K Ito M amp Cheng K 1992 Columns forvisual features of objects in monkey inferotemporal cortexNature 360 343ndash346
Gaffan E A Gaffan D amp Harrison S 1988 Disconnectionof the amygdala from visual association cortex impairs vis-ual-reward association learning in monkeys J Neurosci 83144ndash3150
Gallagher H L Happe F Brunswick N Fletcher P CFrith U amp Frith C D 2000 Reading the mind in cartoonsand stories an fMRI study of lsquotheory of mindrsquo in verbal andnonverbal tasks Neuropsychologia 38 11ndash21
Gauthier I Anderson A W Tarr M J Skudlarski P ampGore J C 1997 Levels of categorization in visual recog-nition studied using functional magnetic resonance imagingCurr Biol 7 645ndash651
Gauthier I Tarr M J Anderson A W Skudlarski P ampGore J C 1999 Activation of the middle fusiform face areaincreases with expertise in recognizing novel objects NatureNeurosci 2 568ndash573
Gauthier I Skudlarski P Gore J C amp Anderson A W2000 Expertise for cars and birds recruits brain areasinvolved in face recognition Nature Neurosci 3 191ndash197
Goel V Grafman J Sadato N amp Hallett M 1995 Mode-ling other minds Neuroreport 6 1741ndash1746
Grelotti D Gauthier I amp Schultz R T 2001 Social interestand the development of cortical face specialization whatautism teaches us about face processing Devl Psychobiol 40213ndash225
Grezes J Fonlupt P Bertenthal B amp Delon-Martin C2001 Does perception of biological motion rely on specificbrain regions NeuroImage 13 775ndash785
Gusnard D A Akbudak E Shulman G L amp RaichleM E 2001 Medial prefrontal cortex and self-referentialmental activity relation to a default mode of brain functionProc Natl Acad Sci USA 98 4259ndash4264
Happe F Ehlers S Fletcher P Frith U Johansson MGillberg C Dolan R Frackowiak R amp Frith C 1996lsquoTheory of mindrsquo in the brain Evidence from a PET scanstudy of Asperger syndrome Neuroreport 8 197ndash201
Haxby J V Ungerleider L G Clark V P Schouten J LHoffman E A amp Martin A 1999 The effect of face inver-sion on activity in human neural systems for face and objectperception Neuron 22 189ndash199
Haxby J V Isahai A Chao L L Ungerleider L G ampMartin A 2000 Object-form topology in the ventral tem-poral lobe response to I Gauthier (2000) Trends Cogn Sci4 3ndash4
Haxby J V Gobbini M I Furey M L Ishai ASchouten J L amp Pietrini P 2001 Distributed and overlap-ping representations of faces and objects in ventral temporalcortex Science 293 2425ndash2430
Haznedar M M Buchsbaum M S Metzger M Solim-ando A Spiegel-Cohen J amp Hollander E 1997 Anteriorcingulate gyrus volume and glucose metabolism in autisticdisorder Am J Psychiat 154 1047ndash1050
Heider F amp Simmel M 1944 An experimental study ofapparent behavior Am J Psychol 57 243ndash259
Hobson R P Ouston J amp Lee A 1988a Whatrsquos in a faceThe case of autism Br J Psychol 79 441ndash453
Hobson R P Ouston J amp Lee A 1988b Emotion recog-nition in autism coordinating faces and voices Psychol Med18 911ndash923
Hoffman E A amp Haxby J V 2000 Distinct representationsof eye gaze and identity in the distributed human neural sys-tem for face perception Nature Neurosci 3 80ndash84
Hornak J Rolls E T amp Wade D 1996 Face and voiceexpression identification in patients with emotional andbehavioural changes following ventral frontal lobe damageNeuropsychologia 34 247ndash261
Phil Trans R Soc Lond B (2003)
Ishai A Ungerleider L G Martin A Schouten J L ampHaxby J V 1999 Distributed representation of objects inthe human ventral visual pathway Proc Natl Acad Sci USA96 9379ndash9384
Kanwisher N 2000 Domain specificity in face perceptionNature Neurosci 3 759ndash763
Kanwisher N McDermott J amp Chun M M 1997 The fusi-form face area a module of extrastriate cortex specializedfor face perception J Neurosci 17 4302ndash4311
Klin A 2000 Attributing social meaning to ambiguous visualstimuli in higher functioning autism and Asperger syndromethe social attribution task J Child Psychol Psychiat 41831ndash846
Klin A Sparrow S S de Bildt A Cicchetti D V CohenD J amp Volkmar F R 1999 A normed study of face recog-nition in autism and related disorders J Autism Devl Dis-orders 29 499ndash508
Klin A Jones W Schultz R T Volkmar F amp Cohen D J2002 Visual fixation patterns during viewing of naturalisticsocial situations as predictors of social competence in indi-viduals with autism Arch Gen Psychiat 59 809ndash816
Kracke I 1994 Developmental prosopagnosia in Aspergersyndrome presentation and discussion of an individual caseDevl Med Child Neurol 36 873ndash886
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J 1997 Neuroanatomical correlates of happi-ness sadness and disgust Am J Psychiat 154 926ndash933
Langdell T 1978 Recognition of faces an approach for thestudy of autism J Child Psychol Psychiat 19 255ndash268
LeDoux J E 1996 The emotional brain New York Simon ampShuster
Lowel S amp Singer W 1992 Selection of intrinsic horizontalconnections in the visual cortex by correlated neuronalactivity Science 255 209ndash212
Macdonald H Rutter M Howlin P Rios P Le ConteurA Evered C amp Folstein S 1989 Recognition andexpression of emotional cues by autistic and normal adultsJ Child Psychol Psychiat 30 865ndash877
Morris J S Ohman A amp Dolan R J 1998 Conscious andunconscious emotional learning in the human amygdalaNature 393 467ndash470
Myers R E Swett C amp Miller M 1973 Loss of social groupaffinity following prefrontal lesions in free-rangingmacaques Brain Res 64 257ndash269
Osterling J amp Dawson G 1994 Early recognition of childrenwith autism a study of first birthday home video tapes JAutism Devl Disorders 24 247ndash257
Pierce K Muller R A Ambrose J Allen G amp Cour-chesne E 2001 Face processing occurs outside the fusiformlsquoface arearsquo in autism evidence from functional MRI Brain124 2059ndash2073
Price J L Carmichael S T amp Drevets W C 1996 Net-works related to the orbital and medial prefrontal cortex asubstrate for emotional behavior Prog Brain Res 107523ndash536
Puce A Allison T Gore J C amp McCarthy G 1995 Face-sensitive regions in human extrastriate cortex studied byfunctional MRI J Neurophysiol 74 1192ndash1199
Reiman E M Lane R D Ahern G L Schwartz G EDavidson R J Friston K J Yun L S amp Chen K 1997Neuroanatomical correlates of externally and internally gen-erated human emotion Am J Psychiat 154 918ndash925
Rolls E T 1995 A theory of emotion and consciousness andits application to understanding the neural basis of emotionIn The cognitive neurosciences (ed M S Gazzaniga) pp1091ndash1106 Cambridge MA MIT Press
Schultz R T Gauthier I Klin A Fulbright R K Ander-son A W Volkmar F Skudlarski P Lacadie C CohenD J amp Gore J C 2000a Abnormal ventral temporal
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from

416 R T Schultz and others FFA and social cognition
adults can enhance their FFA activity to a class of novelobjects through extensive perceptual training (Gauthier etal 1999) Interpreting the hypoactivation of the FFA inautism from an expertise model however argues that thisfinding is an outcome of having autism rather than partof the cause In other words the hypoactivation of theFFA is merely a reflection of the social disability the cul-mination of a set of developmental experiences acrossmany years whereby the person has reduced interest inother people and pays inadequate attention to their facesIn this regard the under-responsiveness of the FFA is abiological marker Identifying an endophenotype such asthis is extremely important and takes the field one signifi-cant step closer to understanding the underlying biologicalmechanisms but it falls short of providing a snapshot ofthe brain mechanisms that actually cause autism
An alternative view would be that the FFA is a corecomponent of the lsquosocial brainrsquo Data emerging overrecent years from neuroimaging studies human lesionstudies and animal studies suggest a working model of thesocial brain that comprises a diverse set of frontal limbicand temporal lobe circuitry Select aspects of the orbitaland medial prefrontal cortices the amygdala and lateralaspects of the temporal cortex involving the STS haveeach been implicated in social functioning (Brothers 1990Fletcher et al 1995 Baron-Cohen et al 1999 Frith ampFrith 1999 Allison et al 2000 Castelli et al 2000 2002Schultz et al 2000b) The frontal and temporal corticeshave dense and often reciprocal connections to theamygdala (Carmichael amp Price 1995 Price et al 1996)The amygdala is centrally positioned and capable of mod-ulating and interpreting the emotional significance of dataprocessed in the perceptual cortices as well as assistingwith the integration of emotion and cognition for decisionmaking and action in the frontal cortices (Amaral et al1992 Schultz et al 2000b) Collectively this systemdefines a heuristic model of the social brain with the pre-cise functions of each node only understood in an as yetsuperficial manner
But is there a role for the FFA in this social circuitryWhereas the role of the FG in face perception is undis-puted only one prior study has implicated the FFA insocial cognition Castelli et al (2000) used social ani-mations involving interacting geometric shapes to probethe social brain These animations were based on theclassic study of Heider amp Simmel (1944) that showed howcertain movements by inanimate objects could stronglyand automatically suggest personal agency and that agroup of interacting geometric forms will naturally suggestsocial interactions All but one of the 34 female collegestudents in Heider and Simmelrsquos study described the ani-mations through a social lens and in human terms (egshapes chase one another fight entrap play get fright-ened elated etc) It seems that the contingent nature ofthe shape movements and the fact that their movementsviolate the rules of simple physics (ie the shapes seem tohave lsquoagencyrsquo or will) naturally invoke social cognitive andsocial perceptual ideation Using PET in six healthy youngadults Castelli et al showed that interpreting this type ofanimation engaged the medial prefrontal cortices the TPthe STS and the right FG Although the localization of thefusiform activations in their study is in the area generallyreported to be the FFA it is not clear whether this region
Phil Trans R Soc Lond B (2003)
of activation would have overlapped with the FFA in thesesubjects as location of the FFA can vary from person toperson Nevertheless engagement of the right FG by asocial cognitive task that does not involve images of theface suggests that this FG region may have a broadermore important set of functions extending beyond simpleface perception As such it provides a basis to argue thatthe hypoactivation of the FFA to faces in autism might beilluminating part of a causal mechanism as opposed to adevelopmental consequence of having autism
The current study also used an adaptation of the pro-cedure of Heider amp Simmel (1944) involving what Klin(2000) called the SAT Klin (2000) used the SAT to showhow persons with autism fail to spontaneously imposesocial meaning on these types of movements Other inves-tigators have also reported a paucity of theory of mindideation among persons with autism to other renditions ofthe HeiderndashSimmel procedure (Abell et al 2000Bowler amp Thommen 2000) Use of simple shapes to dis-play human social interactions without perceptual rep-resentations of real people strips the social event down tothe essential elements needed to convey social meaningsIn this regard the SAT is an ideal neuroimaging probefor assessing social cognitive and social perceptual pro-cesses in a way that is not confounded by perceptual pro-cesses that would be provoked if actual images of faces orpeople were used The original SAT from Klin (2000) isa 50 s silent film in which three moving geometric shapes(a circle a small triangle and a larger triangle) interactwith each other in a social manner Interestingly in Klinrsquosstudy some attributions by those with an autism spectrumdisorder were given in terms of physical meaning (egmagnetic forces) not social meaning We took note of thisobservation in developing the following fMRI experi-ments and created a control task for the SAT that involvesjudgements of object mass
In two fMRI studies of the same group of normal con-trol subjects we show that the FG is robustly engaged byan adaptation of the SAT suitable for a block design fMRIstudy Engagement of the right FG by non-face stimulisuggests that this region has functions beyond static faceperception To test the exact location of the fusiform acti-vations during the SAT nine of the 12 participants con-sented to return for a fMRI study of face perceptionResults from this second study found the location of theFFA to be highly overlapping with the FG activations tothe SAT Thus making social judgements on non-facegeometric figures and making identity judgements ongrey-scale pictures of human faces draws upon a similarneural substrate in the FG This result challenges thespecificity of the middle portion of the FG for faces andraises the possibility that the FFA is part of the primarycircuitry for social cognition As such it has importantimplications for the hypoactivation of the FFA in autismand more generally for specifying a distributed social net-work whose dysfunction might cause autism
2 METHODS
(a) ParticipantsTwelve participants were recruited for this study from the staff
and student populations at Yale University The sampleincluded six men (three left-handed) and six women (one left-
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 417
handed) ranging in age from 20 to 31 years (meanplusmn sd = 242 plusmn 31) Participants were screened for neurologicaland DSM IV Axis I psychiatric disorders Estimated full-scaleIQ as measured by four subtests of the Wechsler Adult Intelli-gence Scale 3rd edition (Wechsler 1997) averaged 1288(plusmn 104) (Information Vocabulary Picture Completion andBlock Design) All subjects scored in the normal range on theBenton Test of Facial Recognition (Benton 1994) (raw scorerange 41ndash50 mean plusmn sd = 468 plusmn 26) There were no signifi-cant differences between the sexes on any of these variables
Nine of the twelve agreed to participate in a follow-up fMRIstudy of face perception conducted so that localization of theFFA could be compared with SAT activations in the middle FGarea in the right hemisphere Five were male (one left-handed)and four were female (one left-handed) mean age was236 plusmn 26 years All subjects gave written informed consent forboth studies in accordance with procedures and protocolsapproved by the Institutional Review Board of the Yale Univer-sity School of Medicine
(b) Experimental tasksWe adapted the SAT for use in a fMRI block-design study by
programming 16 new SAT QuickTime film skits using Directorpublished by Macromedia (600 Townsend Ave San FranciscoCA wwwmacromediacom) From these a final set of eightwere chosen for use in the fMRI study on the basis of ratingsby project staff of the filmrsquos realism and ability to capture onersquossocial attention (these films can be downloaded from httpinfomedyaleeduchldstdyneuroimgsatFmovieshtm) Each filmlasted 151 s and was designed like the original SAT with move-ments intended to suggest a sense of personal agency andreciprocal and contingent interactions that were meant to be eas-ily interpreted as social Each film contained three types of whitegeometric figures (a triangle diamond and circle) that movedagainst a black background In common with the original SATthere was a box in the centre of the field with one wall thatopened as if on a hinge allowing the shapes to open and shutthe door and to enter chase or drag other shapes inside Eachfilm was scripted to follow a social story for example hide-and-seek a fight a love triangle etc The participants were asked todecide by pushing a button if all three of the shapes were lsquofri-endsrsquo or not Half of the films were intended to have lsquoall friendsrsquoas the correct answer (correct answer was determined by a con-sensus-rating process among the developers of the tasks witheach final film version judged to have a clear answer) The filmswere scripted such that any adversarial interactions occurred inthe final few seconds of the film to force the participant toattend throughout to derive the correct answer In creating ourcontrol task for contrast to the SAT we reasoned that each SATfilm requires three important processes
(i) monitoring the movements and physical interactionsbetween the shapes
(ii) pretending that the shapes represent something else iepeople and
(iii) an inferential social reasoning process based on the natureof the interactions (judging whether the movements rep-resent friendly or non-friendly interactions)
A lsquobumper carrsquo control task was created that contained all ofthe elements of the SAT films with the exception of the socialreasoning process This task also entailed eight 151 s filmsdepicting the same geometric shapes moving about a black field
Phil Trans R Soc Lond B (2003)
with the same centrally positioned white box The participantrsquostask was to pretend that these figures were lsquobumper carsrsquomdashsmallracing cars found at amusement parks that are encapsulated byrubber bumpers to allow safe playful collisions Participantsmonitored the carrsquos movements and interactions and decidedon the basis of the carrsquos trajectories and speed after each collisionif the three shapes were all equally lsquoheavyrsquo or not for exampleupon collision if one car shot off more rapidly than anotherthen the two were not equally heavy Key collisions that gaveaway the correct answer always occurred in the final seconds ofthe film Collisions early in the 151 s skit were often mere grazesthat failed to provide definitive information about relative massThus the control task contained the first two elements butinstead of a social decision participants were required to makea decision about a physical property The bumper car and SATfilms were designed to be equivalent with respect to movementquantity and location so that the comparison between the twotasks would reveal the location of brain processes that are dis-tinctly involved in social perceptual and social cognitive pro-cesses It is interesting to note that we piloted a version of thecontrol task that involved physical judgements on the SAT films(as opposed to the bumper car films) but participants reportedthat they were not able to consciously stop seeing the films associal stories Thus it did not seem possible to use the exactsame stimuli in both tasks as might otherwise be desirablebecause social perceptual and cognitive processes would prob-ably be engaged to a greater or lesser extent in both the experi-mental and control tasks Two other lower level controlconditions were also included in each experimental run in theblock design and were intended to further pull apart the threedistinct processes outlined above However the results of thesecontrasts were generally uninformative and thus are not reportedhere Between each film was a 12 s rest period with a blackscreen All participants underwent practice using films that didnot make the final group of eight in order to become completelyfamiliar with the tasks before fMRI scanning During the fMRIexperiment each film was preceded by a 3 s cue lsquoBUMPERCARS SAME WEIGHTrsquo or lsquoPEOPLE ALL FRIENDSrsquoSubjects responded by pressing a button upon completion ofeach film both as a measure of reaction time and accuracy andto ensure that the subjects watched the entire film
In the follow-up study (hereafter lsquoStudy 2rsquo) side-by-side grey-scale faces objects or patterns were presented in asamedifferent task in a block-design experiment to localize theFFA We have previously used this task to localize the FFA ina large group of normal controls (Schultz et al 2000b 2001)Image pairs were presented for 28 s with a 05 s inter-stimulusinterval The person identity task employed same-gender pairsof neutral (non expressive) faces on a black background Pic-tures were taken from standard sources and were edited toremove hair ears and shirt collars so as to force subjects tofocus on features of the face with central relevance to non-verbalsocial communication ie the eyes nose mouth and facegeometry Objects were pictures of spectacles taken from anonline retail catalogue that were contrast inverted to make thebackground black and the spectacles shades of grey to matchthe faces Patterns were distorted versions of the faces or spec-tacles Results from contrasts with patterns were not used tolocalize the FFA and thus are not reported here As is conven-tional the face versus subordinate-level object-discriminationcontrast defined the FFA Each task block lasted 165 s and wasseparated by a 105 s rest period during which cross hairs (1 1)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
418 R T Schultz and others FFA and social cognition
Figure 1 Composite t-map for 12 healthy controls contrasting the social attribution (yellowred) and the bumper car(bluepurple) tasks (p 00005) Right and left are reversed by convention Abbreviations BA Brodman area FG fusiformgyrus IFG inferior frontal gyrus MPFC medial prefrontal cortex STG superior temporal gyrus Y-coordinates are from thesystem of Talairach amp Tournoux (1988)
centred in the same position as the image pairs flashed with thesame presentation rate
(c) Data acquisitionSAT and face perception fMRI data were collected on differ-
ent occasions averaging approximately 25 months apart (range3 weeks to 6 months) Only after the original 12 subjects com-pleted the SAT fMRI study was the decision made to rescansubjects with the face-localization protocol Changes in BOLDcontrast were measured as subjects performed the SAT bumpercar face-discrimination and object-discrimination tasks Thestimuli were run as QuickTime films in Study 1 and as PICTimage files in Study 2 Studies were programmed in PsyScope
125 PPC (Carnegie Mellon University Pittsburgh PA USA)and run on a MacIntosh G3 computer Images were back-pro-jected onto a translucent screen mounted near the end of theMRI gantry and were viewed through a periscopic prism systemon the head coil Behavioural response data were collected witha fibre-optic button box with two response alternatives (Yes orNo for lsquoall friendsrsquo lsquosame weightrsquo lsquosame personrsquo and lsquosameobjectrsquo) The participantrsquos head was immobilized using foamwedges and tape across the forehead
T2 curren weighted images sensitive to BOLD contrast wereacquired on a GE Signa 15 Tesla scanner with a standard quad-
Phil Trans R Soc Lond B (2003)
rature head coil using a gradient echo single-shot echo planarsequence and a coronal orientation perpendicular to the planethrough the ACndashPC The pulse sequence for both studies wasTR = 1500 ms TE = 60 flip angle = 60 NEX = 1 in-planevoxel size = 3125 mm acute 3125 mm In the SAT study we col-lected 14 coronal slices 10 mm thick (skip 1 mm) starting at theanterior-most aspect of the frontal lobe and covering all of thebrain except the caudal-most aspect of the occipital lobe Datawere collected in four runs of an ABCD block design(block = one 151 s film) with blocks of each type presentedtwice per run in a pseudo-random order Across runs a total of80 echo planar images were collected per slice per task con-dition In the face-perception study we also collected 14 coronalslices perpendicular to the ACndashPC starting from the posterioraspect of the occipital cortex up through the rostral-most aspectof the cingulate gyrus Slice thickness was 9 mm (skip 1 mm) tobe compatible with a separate face-perception study ongoing atthat time Data were collected in a block design with a pseudo-random order across six separate runs with three blocks of eachtask per run for 180 echo planar images per slice per task con-dition Functional data in both studies were co-registered to T1-weighted structural images of the same thickness collected in thesame session (TR = 500 TE = 14 field of view = 200 mm256 mm acute 192 mm matrix 2 NEX)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 419
(d) Data analysisData were corrected for motion using SPM99 for three trans-
lation directions and for the three possible rotations (WellcomeDepartment of Cognitive Neurology London UK) Image vol-umes with centre of mass (x y or z) movement of more than15 mm within a run were discarded Image analyses and testsof statistical significance were done using locally developedsoftware (Skudlarski httpmrimedyaleedumembersFframedhtml) Motion corrected images were spatially smoothed with aGaussian filter with a full-width half-maximum value of625 mm The specific effects of each task were evaluated bycreating t-maps for each imaging series incorporating a correc-tion for linear drift (Skudlarski et al 1999) of specific task con-trasts social versus bumper car in Study 1 and face versusobject in Study 2 t-maps were averaged across imaging seriesand co-registered with the higher resolution anatomical imagesfor display and localization These maps were then transformedby in-plane registration and slice interpolation into a pro-portional three-dimensional grid defined by Talairach amp Tour-noux (1988) and averaged across all subjects to createcomposite t-maps with the acquired data in 14 slices interp-olated to 18 slices (NB 16 slices are shown in figure 1 as fMRIactivations on the first and last slice are corrupted by motioncorrection) The SAT versus bumper car maps are displayed inthe figures using a significance level of p 00005(uncorrected) Face versus object t-maps were created and dis-played at p 005 (uncorrected) with the a priori hypothesis thatthe right lateral FG would define the FFA No other brain areasare examined in the second study thus avoiding any multiplecomparison problem
ROI analyses were conducted in the SAT study by tracingsignificant pixels on the group composite activation map (figure1) in the following regions the right FG the right and left STSand STG the right TP the right amygdala and right and leftdorsal MPFC To more thoroughly assess activity in the FGmedial and lateral FG ROIs (and the combined whole FG) weredefined anatomically and traced across the two coronal sliceswhere there was significant SAT activation The activated SATROI for the FG is 33 of the size of the entire anatomicallydefined FG at those two slices Individual subject data wereinterrogated using the ROIs to obtain the mean per cent signalchange for each person for each region and Talairach centre ofmass coordinates (the centre of ROI activation weighted by theamplitude of activation across the region) Mean per cent signal-change data were used in correlational analyses to estimate theconsistency of conjoint activity between ROIs across subjects
3 RESULTS
(a) Behavioural performanceThere were no significant differences between the
social and bumper car tasks in performance accuracy(t1 2 2 = 163 p 010 social = 86 plusmn 16 correctbumper = 74 plusmn 16 correct) or reaction time (t1 22
= 062 p 060) There were no significant differences intask accuracy between males and females or left-handersand right-handers In addition there were no significantcorrelations between task performance and age Full ScaleIntelligence Quotient or Benton Face Recognition per-formance
Phil Trans R Soc Lond B (2003)
(b) Brain activity associated with the socialattribution task
As shown in figure 1 comparison of the SAT with thebumper car control condition resulted in a widely distrib-uted set of significant activations There was very little sig-nificant activation for the bumper car task with the oneregion shown clearly in figure 1 being bilateral activationof the dorsal bank of the intra-parietal sulcus The SATnetwork included a region within the right and left dorsalMPFC the right and left inferior frontal gyrus parsorbitalis and the lateral orbital gyrus the right TP theright amygdala the right and left STS and STG and theright FG It is important to note that at lower thresholds(eg p 001) there was also left amygdala activation anda ROI analysis of the per cent signal change failed to findsignificantly more right than left amygdala activation It isalso worth noting that the FG activations seem quitespecific to the SAT task in the sense that reducing thethreshold down to p 005 failed to show additional ven-tral pathway activation More widespread activationsmight have indicated a general SAT effect on arousal orattention that was manifested throughout the ventralstream but this was not the case
The largest areas of activation were the STG (especiallyon the right) and MPFC Direct comparison of the rightversus left MPFC mean per cent signal change failed tofind significant differences (paired t1 1 = 045 p 050)However the right STG was significantly more activatedthan the left STG (paired t1 1 = 264 p = 002) Table 1presents the Talairach coordinates for the centre of acti-vation mass for each ROI Table 2 presents a correlationalmatrix showing the consistency of conjoint activitybetween regions The strongest correlation is between theright amygdala and the ROI that defines the significantlyactivated region of the right FG (r = 071 p = 001) Inter-estingly this correlation is nominally larger than that ofthe entire FG and the smaller subcomponent FG ROIdefined by the SAT activated pixels (r = 069 p = 0013)Since some correlation would be expected between theseoverlapping ROIs especially since the data were spatiallysmoothed the robust correlation to the amygdala is evenmore impressive Other notable results from the corre-lation matrix include the lack of correlation between theMPFC and either the amygdala or temporal lobe ROIsWithin the temporal lobes however the right STG is sig-nificantly correlated with the left STG and with both defi-nitions of the right FG
Accuracy of performance on the SAT correlates withthe amount of activity in the anatomically defined rightFG (r = 065 p = 002) but not with any other node in theSAT network Females showed significantly more rightSTG activation than males (t1 0 = 234 p = 004) Maleshowever showed significantly more right TP activation(t1 0 = 253 p = 003) There were no other significant sexdifferences and no significant associations with hand-edness or age
(c) Comparison of activity in the right fusiformgyrus during social attribution task and face
perceptionAt the time Study 1 data were collected finding signifi-
cant right FG activation to the SAT was unexpected Toclarify whether the FG activation was in the precise
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
420 R T Schultz and others FFA and social cognition
Table 1 ROI centres of mass coordinates
mean Talairach coordinates (X Y Z )
ROI right hemisphere left hemisphere Brodmann areas
MPFC 49 349 436 274 361 434 6 8 9TP 463 138 2128 mdash 38amygdala 228 2109 2124 mdash mdashFG (SAT) 344 2468 290 mdash 37FG (face) 374 2480 2126 mdash 37STG 507 2571 152 2565 2608 194 22 39
Table 2 ROI correlation matrix(Correlations are based on mean per cent signal change from each ROI (see sect 2 for ROI procedures) SAT accuracy data representthe percentage of films that each participant correctly)
activated whole right right left left right rightright FG FG amygdala amygdala MPFC MPFC left STG STG right TP
whole right FG 069 curren curren curren
right amygdala 071 curren curren curren 032left amygdala 056 curren 019 070 curren curren curren
left MPFC 021 013 032 011right MPFC 2018 2008 021 019 048left STG 021 029 019 005 056 curren 018right STG 054 curren 069 curren curren curren 022 025 009 2016 060 curren curren
right TP 2007 2011 040 033 048 034 038 2010SAT accuracy 045 065 curren curren 2012 009 013 2017 201 022 2034
curren curren curren p lt 001 (r gt 069) curren curren p lt 005 (r gt 057) curren p lt 010
(a) SAT versus bumper car FFA activation
(
(c) enlargement and alignment of FFA
Figure 2 (a) Composite (n = 12) t-map at two slices showing significant (p 00005) activation for the SAT contrast(yellowred) with the bumper car control task (bluepurple) (b) Composite (n = 9) t-map at two slices showing significant(p 005) activation for the face (yellowred) versus object discrimination (bluepurple) This contrast defines the FFA (c)Subregions of composite t-maps shown in (a) and (b) are enlarged and aligned to demonstrate the overlap of activation in theFG for the SAT and face discrimination activations Subscripts 1 and 2 refer to the first (more anterior) and second coronalslices with significant activation
location of the FFA we compared it with the result of theface versus object discrimination contrast (the standardmeans of identifying the FFA in the literature) Both theFFA and the SAT activations of the FG were confined totwo coronal slices in highly overlapping locations Thesetwo sets of group composite maps are shown in figure 2
Phil Trans R Soc Lond B (2003)
The FG activations were stronger in the SAT versusbumper contrast than the face versus object contrast butthis may have had as much do with the baseline as theexperimental tasks The Talairach coordinates for theSAT and FFA activations show that their centre of massdiffers by less than one voxel The SAT activation is 3 mm
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 421
Figure 3 Scans of two individuals across three different occasions showing the reproducibility of FFA activations at two time-points and relationship to SAT activations (andashc) are from a 23-year-old male (dndashf ) are from a 24-year-old female Panels arearranged chronologically (ad) The first face versus object experiment (be) The second face versus object scan (cf ) SATversus bumper car contrast Both coronal slices are shown where there was FFA (t gt 15 in yellowred) or SAT activation(t gt 30 in yellowred) Arrows point to FG activity (right and left are reversed by convention) As in the group results (figure2) the SAT activation is centred slightly more medially along the MFS Left FG activation shown in these two subjects doesnot survive thresholding in the group composite (figure 2) Control tasks (object discrimination bumper car) are shown inpurpleblue
more medial than that of the FFA straddling the MFSthat delineates the lateral and medial aspects of the FGThe FFA on the other hand is clearly positioned in thelateral FG as expected (Haxby et al 1999) The SATactivation is also centred 36 mm more superior and about1 mm more anteriorly than the FFA A count of the over-lapping significant pixels showed that 50 of the SATactivation falls within the FFA This provides a goodapproximation of how these regions overlap Howeverthere is no definitive way to measure the percent overlapin this study because it would change with the use of dif-ferent control tasks in either condition or different signifi-cance levels for thresholding the t-maps
Two of the participants (one male one female) werealso part of a reproducibility study of the FFA and hadboth undergone the face discrimination protocol on twooccasions Figure 3 presents t-maps of their FG for eachface perception study and the SAT The female partici-pant shows reversed asymmetry as sometimes happenswith the left FG showing greater face activation than theright Nevertheless her SAT activations track her FFAand are more left-sided than typical These results showthat the FFA activation is reproducible so that the lessthan perfect overlap between the FFA and the SAT FGactivation is probably not a measurement or reliabilityissue
4 DISCUSSION
(a) The social brain networkThe current study required participants to observe the
movements of geometric figures and to interpret thesewith regard to a conceptual template about what consti-tutes a friendly or unfriendly social interaction It requiredclose attention to the contingent nature of a sequence ofmovements and inferences about mental states of eachcharacter to explain their actions Perception of the move-
Phil Trans R Soc Lond B (2003)
ments of these simple shapes as wilful seems to be auto-matic and effortless for healthy controls but not forpersons with autism (Klin 2000) As shown in figure 1the network engaged by the social attribution process (incontrast to the physical attribution control task) includednearly all of the brain areas implicated by past researchon the social brain (Brothers 1990) including cognitiveaspects such as theory of mind (Castelli et al 2000) aswell as perceptual aspects such as the perception of socialdisplays and biological movement (Allison et al 2000)We found significant activation of the bilateral MPFCsuperior STG and STS and inferior FG pars orbitalisextending into the lateral orbital gyri In addition therewere significant activations on the right side only for theamygdala TP and the FG The predominance of right-side activations is consistent with the notion that the righthemisphere is more concerned than the left with socialprocesses (Siegal et al 1996 Winner et al 2002) Ourresults differ from some past research by finding signifi-cant right FG activation and by failing to find orbito-MPFC activation
Several earlier neuroimaging studies have shown thatthe dorsal MPFC (ie that cortex anterior and superior tothe anterior cingulate gyrus) is a critical substrate for socialjudgements including empathizing and thinking aboutotherrsquos thoughts and intentions (Fletcher et al 1995 Goelet al 1995 Happe et al 1996 Castelli et al 2000 2002Gallagher et al 2000) Our findings are consistent withthese imaging studies and with non-human primate stud-ies that have documented social failures and loss of socialposition within the group following lesions to orbital pre-frontal cortices and MPFC (Butter et al 1969 Myers etal 1973 Bachevalier amp Mishkin 1986) The importantrole for the MPFC in social cognition is further suggestedby studies of autism spectrum disorders that find func-tional abnormalities in this area (Happe et al 1996 Ernstet al 1997 Haznedar et al 1997 Castelli et al 2002)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
422 R T Schultz and others FFA and social cognition
Gusnard et al (2001) suggest that the dorsal MPFC isinvolved in any kind of thought that uses the self as a refer-ent Thus the SAT activations in this area may have beendriven by theorizing about othersrsquo minds but with explicitreference to the participantrsquos own frame of reference as tohow they would feel in a similar situation
The orbital prefrontal cortex and to a lesser extent thedorsal MPFC have dense reciprocal connections withmedial temporal areas (Price et al 1996 Carmichael ampPrice 1995) providing the anatomical bases for a systemthat regulates emotional processes Damasio and col-leagues (Damasio et al 1990 Bechara et al 1996) haveargued that the orbito-MPFC have a primary function ofintegrating information about rewards and punishmentsto bias future behaviour (Rolls 1995 Dias et al 1996Hornak et al 1996 Lane et al 1997 Reiman et al 1997)A functional circuitry such as this would seem especiallyimportant in the development and acquisition of socialbehaviour However even acquired lesions to theseregions in adulthood can result in abnormalities of socialconduct (Damasio et al 1990) Brothers (1990) high-lighted the orbital prefrontal cortex as one of the threeprincipal brain regions involved in social cognition Weobserved poor signal in this region (these areas are notori-ously prone to fMRI signal drop-out and distortion) andthus we cannot know if the orbital prefrontal region wasengaged by the SAT or not However using a similarpsychological task but with PET where signal acquisitionin this region is not degraded Castelli et al (2000 2002)failed to find activation of the orbital prefrontal cortexCollectively these results call into question the impor-tance of the orbital prefrontal cortex in social cognitionand instead shift the focus on anterior cortices toward thedorsal MPFC
The SAT also generated robust activations of the rightamygdala and nearby cortex of the right TP The amyg-dala is often given a central role in theories of social per-ception and cognition (Brothers 1990 Bachevalier 1994Adolphs et al 1998 Baron-Cohen et al 2000 Schultz etal 2000b) The amygdala has a critical role in emotionalarousal assigning significance to environmental stimuliand mediating the formation of visual-reward associationsthat is lsquoemotionalrsquo learning (Gaffan et al 1988 LeDoux1996 Anderson amp Phelps 2001) It is reliably engagedduring judgements of personality characteristics from pic-tures of the face or part of the face (Adolphs et al 1998Baron-Cohen et al 1999 Winston et al 2002) Activationof the amygdala appears to be automatic and stimulusdriven as it can be engaged by images of facial expressionsin conscious awareness as well as by subliminal presen-tations of faces displaying affect (Morris et al 1998Whalen et al 1998 Critchley et al 2000) Thus theamygdalarsquos engagement by the SAT could stem from thegeneral emotional arousal evoked by the animations or itcould represent its computational role in some more spe-cific social perceptual process
The amygdala has dense reciprocal connections withthe ventral visual processing stream (Amaral amp Price1984) The strong correlation observed in this studybetween the right amygdala and the right FG could indi-cate that emotional inputs from the amygdala to the FGare necessary for engaging the social computational pro-cesses of the FG Brothers (1995) has speculated that the
Phil Trans R Soc Lond B (2003)
amygdala generates lsquosocial feelingsrsquo that are of importancein cutting through the complexities of social situations andguiding behaviour by simpler learned contingenciesSocial events are complex because their meaning oftencomes from specific combinations of features that do notadd up in a linear manner making it more difficult todecompose the events by cognitive analysis Effectivesocial engagement requires integration of context historicalrelationships and current socialndashemotional communi-cations expressed through prosodic tone as well as facialexpressions posture and gesture They are made morecomplex by the rapid pace of social transactions Thiswould necessitate some fast response system based ongeneral principles from past socialndashemotional experiencesa role for which the amygdala would seem ideally suitedIn other words the amygdala might drive intuition or lsquogutfeelingsrsquo to guide rapid non-verbal social interactionsinvolving facial expressions gesture etc Thus the strongamygdalandashFG correlation observed here could be inter-preted as the amygdala informing the FG of the relevanceof a social event and also of the outcome of its quick anddirty social perceptual analyses thereby guiding the FG(and other social nodes) in their finer grained compu-tations
Perhaps the largest and strongest activations to the SATwere those of the posterior aspects of the STS spreadinginto the adjacent STG This area has been implicated asa specific site for perception of biological motion (Bondaet al 1996 Allison et al 2000 Grezes et al 2001 Vainaet al 2001) Biological motion is a broad construct thatseems to encompass the perception of static images ofevents that could move or did move such as facialexpressions as well as actual movement by animateobjects This region of the STSndashSTG is also critical forthe decoding visual displays of social action or intention(eg gaze direction gesture and facial displays of emotion)(Critchley et al 2000 Hoffman amp Haxby 2000) In thisregard the SAT activations here are completely expectedand a testament to the effectiveness of these animationsin inducing the desired illusion of anthropomorphicaction The majority of the STSndashSTG activations wereanterior to location of the V5MT as defined in otherrecent fMRI studies (Culham et al 2001) but they alsoextended posteriorly into these more general movementsensitive cortices Unlike the amygdala some evidencesuggests that activity of the STS is mediated by explicitattention to social characteristics of the face and that STSengagement it is not automatic or stimulus driven(Winston et al 2002) We surmise therefore that acti-vation of the STS by the SAT was due to explicit taskinstructions to judge the social interactions
(b) Role of the fusiform face area in socialcognition
We also found significant SAT activation of the rightFG When compared directly in a subset of participants totheir FFA gleaned from a separate scanning session witha perceptual discrimination task of person versus objectidentity we found that both ROIs were confined to theright side on the same two coronal slices There wasabout 50 overlap between the SAT FG and FFA acti-vations and the centre of the two activations differed byless than 1 voxel The FFA activation was offset to the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 423
lateral prominence of the FG whereas the SAT activateda region of the FG that was closer to the centre of the FGbut still largely in the lateral aspect Because the SAT doesnot contain any face representations the FFA engagementwas unexpected as this region is thought of as selectivefor faces (Kanwisher et al 1997) or to other classes ofcomplex objects for which one is perceptually expert(Gauthier et al 2000)
These results have important implications for the poss-ible role of the fusiform in social brain circuitry andautism Interestingly the magnitude of the FG activationpredicted SAT task accuracy no other ROI correlated sig-nificantly with task accuracy This would seem to argueagainst any interpretation that the FFA activation is incon-sequential to the SAT that it is activated simply becauseit is well connected to other areas that are directly involvedin the social attribution process Using a task that is simi-lar to the SAT Castelli et al (2000) also reported signifi-cant right FG activation using PET However in theirfollow-up PET study comparing autism and normal con-trols (Castelli et al 2002) they failed to find significantFG activation in either group The failure of their secondstudy to find right FG activations may be due to the morestringent random effects model used in the data analysesHowever because we find right FG activation with fMRIwhich is more sensitive than PET and because Castelli etal (2000) found it with what amounts to a lower thres-hold it seems quite probable that the effect is real In factwe have preliminary evidence from an ongoing fMRIstudy that reproduces the SAT FG activations in healthycontrols (Schultz et al 2001) In addition there may besome task attributes that differ between our studies andCastelli et alrsquos that might impact into the strength withwhich FG computational processes are evoked
The key question then is why is the FG engaged bythe SAT and what role does the right FG have in socialcognition and perception Currently there are three com-peting theories of the functional organization of the FGand related ventral visual perceptual areas One modelput forth by Haxby Chao Martin and colleagues speci-fies that objects are encoded in a distributed fashion acrossa wide expanse of the ventral temporal-occipital cortex(Haxby et al 2001) They call this the lsquoobject formtypologyrsquo model (Ishai et al 1999) Their data indicatethat object category specificity is achieved by unique spa-tial patterns of activation across this extrastriate visual cor-tex They show that the pattern of activations across thiscortex is diagnostic of object category membership moreso than any localized activation maxima (Haxby et al2001) They also argue that object category perceptioninvolves retrieval of category-related information aboutspecific features and attributes of the object category(Chao et al 2002) They admit that the lsquonature of theinformation about objects that is represented in the ventraltemporal cortex is a great puzzlersquo (Haxby et al 2000 p4) but open the possibility that semantic information maybe important
A second view championed by Kanwisher et al holdsthat several select perceptual categories including facesbodies and spatial layout of places are encoded in highlyspecific locations in a modular fashion (Kanwisher et al1997 Epstein amp Kanwisher 1998 Downing et al 2001Spiridon amp Kanwisher 2002) The FFA is thought to be
Phil Trans R Soc Lond B (2003)
most concerned with discriminating among individualidentities and not with discriminating images at the categ-orical level In fact lesions to the FFA cause a specificdeficit in recognizing individual identities but not in reco-gnizing the general category of face versus non-face(Wada amp Yamamoto 2001) Spiridon amp Kanwisher (2002)provide a partial replication of Haxby et alrsquos (2001) objectform typology model They show that patterns of ventralvisual cortical activation can distinguish object categoriesNevertheless their data show category specificity for selectareas such as the FFA and they argue the ventral visualcortex is not equipotential
Gauthier et al however have argued that the ventraloccipital temporal pathway is organized by the nature ofthe perceptual computations and that these processingbiases are acquired through experience (Gauthier et al2000 Tarr amp Gauthier 2000) For example the parahip-pocampal place area is a function of a bias towards pro-cessing landscapes in terms of their spatial layout becausewe have learned through repeated experiences that this isvery useful information to extract when perceiving land-scapes and related visual images Similarly we learn quiteearly in life that it is important to discriminate faces on anindividual level for example discriminating mother fromothers This bias towards individual identification accord-ing to Gauthier accounts for the FFArsquos apparent modu-larity (Gauthier et al 1997 2000 Tarr amp Gauthier 2000)According to this third model the type of informationneeded and our cumulative experience in processing thatinformation organizes the ventral occipitalndashtemporal path-way into regional centres with preferred modes of pro-cessing
All three models seem to agree that the functionalorganization of the ventral visual cortices is driven by theneed to categorize perceptions into object classes Webelieve that our findings showing FFA activation by non-face objects are consistent with aspects of both the pro-cessing map model of Gauthier and the object formtypology model of Haxby et al First consistent withHaxby et al we suggest that semantic information isimportant to the ventral visual pathway for object categor-ization Second consistent with Gauthier et al we believethat repeated perceptual experience with faces biases thetype of information that FG finds important We proposethat the middle FG area encodes semantic attributes ofpeople because of repeated perceptual experiences withfaces that occur during social interactions In fact wewould guess that most perceptual experiences with facesoccur during social situations and these social situationsoften involve repeated social judgements Thus infor-mation about the social nature of people might be storedin the FG (though not exclusively)
The nature of the semantic information stored in theFG might be restricted to anything that would be helpfulin defining faces as a distinct category of object becausemaking such distinctions appears to be the primary chargeof the ventral visual pathway This would include knowl-edge of people as having personal agency and of havingthe capability to disturb each otherrsquos emotional homeo-stasis (eg to act friendly or unfriendly) By pretendingthat the three shapes in the SAT are people stored knowl-edge about people in social interactions might beretrieved causing the observed activations in the FG
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
424 R T Schultz and others FFA and social cognition
There would be a measure of efficiency from thisarrangement that is having representations and compu-tations of more abstract attributes of people inter-digitatedwith front-end perceptual processes about physical attri-butes Gauthierrsquos claim is that the same group of neuronscan be engaged by perceptually different categories ofobjects it is the type of processing rather than the visualdetails that are important Extrapolating from this wewould argue that the SAT engages a region of the FGoverlapping with the FFA because it demands compu-tational processes to classify the SAT geometric figures aspeople or person-like Chao et al (1999) reach a similarconclusion They argued that activity in the ventral path-way reflects stored information about an object categorynot just physical features They point out that thisarrangement could explain why lesion patients with cate-gory specific perceptual deficits also have trouble retriev-ing general information about that visual category Thuswe would add that the FFA must store general infor-mation about people or some meta-representation oflsquopeoplenessrsquo
(c) Implications for the pathobiology of autismThe SAT used in this current study appears to be an
effective neurobehavioural probe for engaging a distrib-uted network of brain regions involved in different aspectsof social perception and cognition It will be important touse this and similar procedures in persons with autism tobetter define the nature of brain functions in this disorderCastelli et al (2002) have already taken the first step inthis process and they describe a pattern of hypoactivationin the MPFC STS and TPs in autism In an ongoingstudy of autism spectrum disorders we have presentedpreliminary data using the SAT showing that we too findhypoactivation of these regions and in the amygdala andFG (Schultz et al 2001) Thus we predict that futurework in this area will show that the entire social brain net-work is underactive in autism during tasks requiring socialperceptual and social cognitive processing
There are already sufficient data to argue strongly for arole of the FG in the pathobiology of autism No less thanfive previous fMRI studies have shown the FFA region tobe significantly less engaged among persons with autismcompared with controls during face perception tasks(Critchley et al 2000 Dierks et al 2001 Pierce et al2001 Schultz et al 2000a 2001) Activation of the FGby the social judgements in the current study adds animportant piece of evidence in favour of a causal role forthe FG in the pathobiology of autism This conclusionhowever must remain tentative until additional studiesmore precisely define the factors leading to FG activationduring social attribution and gather better data to proveits computational role in social cognition In addition theeffect of FG lesions for social functioning must be clari-fied If the FG is involved in social computations onewould expect to find social cognitive deficits in personswith lesions to this area However social deficits in proso-pagnosics have not been widely reported It might be thatprosopagnosic patients have not been carefully tested onthis dimension In this regard using our behavioural ver-sion of the SAT (Klin 2000) to test social perceptionamong prosopagnosics would be quite interesting Theparallels however between autism and prosopagnosia are
Phil Trans R Soc Lond B (2003)
incomplete because autism is clearly a developmental dis-order whereas the research literature on prosopagnosia ismostly confined to lesions acquired in adolescence oradulthood Among the few cases of developmental proso-pagnosia reported in the literature there is indeed one thathighlights severe social impairments (Kracke 1994) Itmay be that the role of the FG in the development ofautistic symptoms is different from the role of the FG inthe maintenance of social cognitive functions after brainmaturity
It is also possible that the role of the FG in social pro-cesses is dependent on its functional relationships withother nodes in the social brain and that it is the collectiveaction and interaction of the network that is of primaryimportance for social behaviours For example we founda strong correlation across participants in the amount ofFG and amygdala activation A strictly modular view ofthese areas may be inappropriate as the functions of eachnode could be quite dependent on one another and whenonly considered in isolation quite insufficient to supportsocial processes Thus whereas a lesion to the FG mayimpair visual perception it might not have a large impacton social functioning if this depends on an extended net-work that is dynamic and capable of compensatory adjust-ments Dysfunction of the FG may be necessary but notsufficient to produce social deficits Indeed our relianceon modular models of brain functioning may be leadingus astray in our search for causal mechanisms in autismInstead it may be the collective action of a distributedsystem that is critical to the pathobiology of autism TheFG region may be a key partner in this distributed systembut nevertheless just one node and insufficient by itselfto support in any substantive manner social cognitive pro-cesses or to explain social cognitive deficits seen inautism
5 CONCLUSION AND FUTURE DIRECTIONS
Using the SAT we isolated a distributed network ofactivations that conform to the emerging model of thesocial brain Most important we found significant acti-vation of the central aspect of the FG thus adding thisregion to the expanding list of structures involved in socialprocesses The FG region activated by the SAT overlapsin its spatial extent with the FFA with centres of massdiffering by less than 1 voxel We speculate that these SATactivations represent computational processes associatedwith more abstract attributes of people
It is an open question as to whether the substantial over-lap between the FFA and SAT FG activation is due to anoverlapping or shared set of neuronal assemblies This willneed to be clarified by future work Electrophysiologicalstudies show that there are small face-specific patches inthe FG cortex (Allison et al 1999) In the current studysmall patches of face cells and SAT cells could be inter-mingled but distinct and not drawing on any of the sameneuronal assemblies The spatial resolution of fMRI haslimitations that may preclude any definitive answer to thisquestion but there are strategies that can be used toaddress the issue For example the current study designdid not contain SAT and face discriminations in the sameexperimental fMRI series Doing so would enable directcomparisons of the computational demands placed on the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 425
common area of the FFA It also would be informative iffollow-up studies superimposed faces within the geometricfigures of the SAT without changing the film scripts inany way We could then determine if the computations ofthe common area of the FFA increase in a predictablefashionmdashwould the activation be a linear summation ofthe original SAT plus face discrimination Any significantdeviation from an additive model would suggest that thereis some sharing of neuronal assemblies with the SAT andface discrimination tasks when presented alone
This work was supported by grants from the National Institutesof Child Health and Human Development (grants PO1 HD03008 and PO1 HDDC35482) and the Korczak FoundationThe authors thank the anonymous reviewers for their helpfulcomments Michael Lee for programming the fMRI tasksHedy Serofin and Terry Hickey for their assistance in acquiringthe fMRI data John Herrington for his valuable assistance indata analyses of preliminary studies and members of the YaleDevelopmental Neuroimaging Program for their helpful com-ments on this manuscript A special acknowledgement ofappreciation is given to Donald Cohen who passed away inOctober of 2001 for his outstanding mentorship in all aspectsof this program of work
REFERENCES
Abell F Happe F amp Frith U 2000 Do triangles play tricksAttribution of mental states to animated shapes in normaland abnormal development J Cogn Dev 15 1ndash20
Adolphs R Tranel D amp Damasio A R 1998 The humanamygdala in social judgement Nature 393 470ndash474
Allison T Puce A Spencer D D amp McCarthy G 1999Electrophysiological studies of human face perception Ipotentials generated in occipitotemporal cortex by face andnon-face stimuli Cerebr Cortex 9 415ndash430
Allison T Puce A amp McCarthy G 2000 Social perceptionfrom visual cues role of the STS region Trends Cogn Sci4 267ndash278
Amaral D G amp Price J L 1984 Amygdalo-cortical projec-tions in the monkey (Macaca fascicularis) J Comp Neurol230 465ndash496
Amaral D G Price J L Pitkanen A amp Carmichael S T1992 Anatomical organization of the primate amygdaloidcomplex In The amygdala neurobiological aspects of emotionmemory and mental dysfunction (ed J Aggleton) pp 1ndash66New York Wiley-Liss
Anderson A K amp Phelps E A 2001 Lesions of the humanamygdala impair enhanced perception of emotionally salientevents Nature 411 305ndash309
Bachevalier J 1994 Medial temporal lobe structures andautism a review of clinical and experimental findingsNeuropsychologia 32 627ndash648
Bachevalier J amp Mishkin M 1986 Visual recognition impair-ment follows ventromedial but not dorsolateral prefrontallesions in monkeys Behav Brain Res 20 249ndash261
Baron-Cohen S Ring H A Wheelwright S BullmoreE T Brammer M J Simmons A amp Williams S C R1999 Social intelligence in the normal and autistic brain anfMRI study Eur J Neurosci 11 1891ndash1898
Baron-Cohen S Ring H A Bullmore E T WheelwrightS Ashwina C amp Williams S C R 2000 The amygdalatheory of autism Neurosci Biobehav Rev 24 355ndash364
Bechara A Tranel D Damasio H amp Damasio A R 1996Failure to respond autonomically to anticipated future out-comes following damage to prefrontal cortex Cerebr Cortex6 215ndash225
Benton A 1994 Face recognition Los Angeles CA WesternPsychological Services
Phil Trans R Soc Lond B (2003)
Bonda E Petrides M Ostry D amp Evans A 1996 Specificinvolvement of human parietal systems and the amygdala inthe perception of biological motion J Neurosci 16 3737ndash3744
Bowler D M amp Thommen E 2000 Attribution of mechan-ical and social causality to animated displays by childrenwith autism Autism 4 147ndash171
Brothers L 1990 The social brain a project for integratingprimate behavior and neurophysiology in a new domainConcepts Neurosci 1 27ndash151
Brothers L 1995 Neurophysiology of the perception of inten-tion by primates In The cognitive neurosciences 2nd edn (edM S Gazzaniga) pp 1107ndash1115 Cambridge MA MITPress
Butter C M McDonald J A amp Snyder D R 1969 Oralitypreference behavior and reinforcement value of nonfoodobject in monkeys with orbital frontal lesions Science 1641306ndash1307
Carmichael S T amp Price J L 1995 Limbic connections ofthe orbital and medial prefrontal cortex in macaque monk-eys J Comp Neurol 363 615ndash641
Castelli F Happe F Frith U amp Frith C 2000 Movementand mind a functional imaging study of perception andinterpretation of complex intentional movement patternsNeuroImage 12 314ndash325
Castelli F Frith C Happe F amp Frith U 2002 AutismAsperger syndrome and brain mechanisms for the attri-bution of mental states to animated shapes Brain 1251839ndash1849
Chao L L Haxby J V amp Martin A 1999 Attribute-basedneural substrates in posterior temporal cortex for perceivingand knowing about objects Nat Neurosci 2 913ndash919
Chao L L Weisberg J amp Martin A 2002 Experience-dependent modulation of category-related cortical activityCerebr Cortex 12 545ndash551
Critchley H D (and 10 others) 2000 The functional neuroan-atomy of social behaviour changes in cerebral blood flowwhen people with autistic disorder process facial expressionsBrain 123 2203ndash2212
Culham J C Cavanagh P amp Kanwisher N G 2001 Atten-tion response functions characterizing brain areas usingfMRI activation during parametric variations of attentionalload Neuron 32 737ndash745
Damasio A R Tranel D amp Damasio H 1990 Individualswith sociopathic behavior caused by frontal damage fail torespond autonomically to social stimuli Behav Brain Res41 81ndash94
Dierks T Bolte S Huble D Lanfermannm H amp PoustkaF 2001 Alterations of face processing strategies in autism(a fMRI study) Presented at the 6th Annual Meeting of theOrganization for Human Brain Mapping Brighton UK 10ndash14 June 2001
Dias R Robbins T W amp Roberts A C 1996 Dissociationin prefrontal cortex of affective and attentional shifts Nature380 69ndash72
Downing P E Jiang Y Shuman M amp Kanwisher N 2001A cortical area selective for visual processing of the humanbody Science 293 2470ndash2473
Epstein R amp Kanwisher N 1998 A cortical representation ofthe local visual environment Nature 392 598ndash601
Ernst M Zametkin A J Matochik J A Pascualvaca D ampCohen R M 1997 Reduced medial prefrontal dopami-nergic activity in autistic children Lancet 350 638
Fletcher P C Happe F Frith U Baker S C DolanR J Frackowiak R S amp Frith C D 1995 Other minds inthe brain a functional imaging study of lsquotheory of mindrsquo instory comprehension Cognition 57 109ndash128
Frith C D amp Frith U 1999 Interacting minds a biologicalbasis Science 286 1692ndash1695
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
426 R T Schultz and others FFA and social cognition
Fujita I Tanaka K Ito M amp Cheng K 1992 Columns forvisual features of objects in monkey inferotemporal cortexNature 360 343ndash346
Gaffan E A Gaffan D amp Harrison S 1988 Disconnectionof the amygdala from visual association cortex impairs vis-ual-reward association learning in monkeys J Neurosci 83144ndash3150
Gallagher H L Happe F Brunswick N Fletcher P CFrith U amp Frith C D 2000 Reading the mind in cartoonsand stories an fMRI study of lsquotheory of mindrsquo in verbal andnonverbal tasks Neuropsychologia 38 11ndash21
Gauthier I Anderson A W Tarr M J Skudlarski P ampGore J C 1997 Levels of categorization in visual recog-nition studied using functional magnetic resonance imagingCurr Biol 7 645ndash651
Gauthier I Tarr M J Anderson A W Skudlarski P ampGore J C 1999 Activation of the middle fusiform face areaincreases with expertise in recognizing novel objects NatureNeurosci 2 568ndash573
Gauthier I Skudlarski P Gore J C amp Anderson A W2000 Expertise for cars and birds recruits brain areasinvolved in face recognition Nature Neurosci 3 191ndash197
Goel V Grafman J Sadato N amp Hallett M 1995 Mode-ling other minds Neuroreport 6 1741ndash1746
Grelotti D Gauthier I amp Schultz R T 2001 Social interestand the development of cortical face specialization whatautism teaches us about face processing Devl Psychobiol 40213ndash225
Grezes J Fonlupt P Bertenthal B amp Delon-Martin C2001 Does perception of biological motion rely on specificbrain regions NeuroImage 13 775ndash785
Gusnard D A Akbudak E Shulman G L amp RaichleM E 2001 Medial prefrontal cortex and self-referentialmental activity relation to a default mode of brain functionProc Natl Acad Sci USA 98 4259ndash4264
Happe F Ehlers S Fletcher P Frith U Johansson MGillberg C Dolan R Frackowiak R amp Frith C 1996lsquoTheory of mindrsquo in the brain Evidence from a PET scanstudy of Asperger syndrome Neuroreport 8 197ndash201
Haxby J V Ungerleider L G Clark V P Schouten J LHoffman E A amp Martin A 1999 The effect of face inver-sion on activity in human neural systems for face and objectperception Neuron 22 189ndash199
Haxby J V Isahai A Chao L L Ungerleider L G ampMartin A 2000 Object-form topology in the ventral tem-poral lobe response to I Gauthier (2000) Trends Cogn Sci4 3ndash4
Haxby J V Gobbini M I Furey M L Ishai ASchouten J L amp Pietrini P 2001 Distributed and overlap-ping representations of faces and objects in ventral temporalcortex Science 293 2425ndash2430
Haznedar M M Buchsbaum M S Metzger M Solim-ando A Spiegel-Cohen J amp Hollander E 1997 Anteriorcingulate gyrus volume and glucose metabolism in autisticdisorder Am J Psychiat 154 1047ndash1050
Heider F amp Simmel M 1944 An experimental study ofapparent behavior Am J Psychol 57 243ndash259
Hobson R P Ouston J amp Lee A 1988a Whatrsquos in a faceThe case of autism Br J Psychol 79 441ndash453
Hobson R P Ouston J amp Lee A 1988b Emotion recog-nition in autism coordinating faces and voices Psychol Med18 911ndash923
Hoffman E A amp Haxby J V 2000 Distinct representationsof eye gaze and identity in the distributed human neural sys-tem for face perception Nature Neurosci 3 80ndash84
Hornak J Rolls E T amp Wade D 1996 Face and voiceexpression identification in patients with emotional andbehavioural changes following ventral frontal lobe damageNeuropsychologia 34 247ndash261
Phil Trans R Soc Lond B (2003)
Ishai A Ungerleider L G Martin A Schouten J L ampHaxby J V 1999 Distributed representation of objects inthe human ventral visual pathway Proc Natl Acad Sci USA96 9379ndash9384
Kanwisher N 2000 Domain specificity in face perceptionNature Neurosci 3 759ndash763
Kanwisher N McDermott J amp Chun M M 1997 The fusi-form face area a module of extrastriate cortex specializedfor face perception J Neurosci 17 4302ndash4311
Klin A 2000 Attributing social meaning to ambiguous visualstimuli in higher functioning autism and Asperger syndromethe social attribution task J Child Psychol Psychiat 41831ndash846
Klin A Sparrow S S de Bildt A Cicchetti D V CohenD J amp Volkmar F R 1999 A normed study of face recog-nition in autism and related disorders J Autism Devl Dis-orders 29 499ndash508
Klin A Jones W Schultz R T Volkmar F amp Cohen D J2002 Visual fixation patterns during viewing of naturalisticsocial situations as predictors of social competence in indi-viduals with autism Arch Gen Psychiat 59 809ndash816
Kracke I 1994 Developmental prosopagnosia in Aspergersyndrome presentation and discussion of an individual caseDevl Med Child Neurol 36 873ndash886
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J 1997 Neuroanatomical correlates of happi-ness sadness and disgust Am J Psychiat 154 926ndash933
Langdell T 1978 Recognition of faces an approach for thestudy of autism J Child Psychol Psychiat 19 255ndash268
LeDoux J E 1996 The emotional brain New York Simon ampShuster
Lowel S amp Singer W 1992 Selection of intrinsic horizontalconnections in the visual cortex by correlated neuronalactivity Science 255 209ndash212
Macdonald H Rutter M Howlin P Rios P Le ConteurA Evered C amp Folstein S 1989 Recognition andexpression of emotional cues by autistic and normal adultsJ Child Psychol Psychiat 30 865ndash877
Morris J S Ohman A amp Dolan R J 1998 Conscious andunconscious emotional learning in the human amygdalaNature 393 467ndash470
Myers R E Swett C amp Miller M 1973 Loss of social groupaffinity following prefrontal lesions in free-rangingmacaques Brain Res 64 257ndash269
Osterling J amp Dawson G 1994 Early recognition of childrenwith autism a study of first birthday home video tapes JAutism Devl Disorders 24 247ndash257
Pierce K Muller R A Ambrose J Allen G amp Cour-chesne E 2001 Face processing occurs outside the fusiformlsquoface arearsquo in autism evidence from functional MRI Brain124 2059ndash2073
Price J L Carmichael S T amp Drevets W C 1996 Net-works related to the orbital and medial prefrontal cortex asubstrate for emotional behavior Prog Brain Res 107523ndash536
Puce A Allison T Gore J C amp McCarthy G 1995 Face-sensitive regions in human extrastriate cortex studied byfunctional MRI J Neurophysiol 74 1192ndash1199
Reiman E M Lane R D Ahern G L Schwartz G EDavidson R J Friston K J Yun L S amp Chen K 1997Neuroanatomical correlates of externally and internally gen-erated human emotion Am J Psychiat 154 918ndash925
Rolls E T 1995 A theory of emotion and consciousness andits application to understanding the neural basis of emotionIn The cognitive neurosciences (ed M S Gazzaniga) pp1091ndash1106 Cambridge MA MIT Press
Schultz R T Gauthier I Klin A Fulbright R K Ander-son A W Volkmar F Skudlarski P Lacadie C CohenD J amp Gore J C 2000a Abnormal ventral temporal
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from

FFA and social cognition R T Schultz and others 417
handed) ranging in age from 20 to 31 years (meanplusmn sd = 242 plusmn 31) Participants were screened for neurologicaland DSM IV Axis I psychiatric disorders Estimated full-scaleIQ as measured by four subtests of the Wechsler Adult Intelli-gence Scale 3rd edition (Wechsler 1997) averaged 1288(plusmn 104) (Information Vocabulary Picture Completion andBlock Design) All subjects scored in the normal range on theBenton Test of Facial Recognition (Benton 1994) (raw scorerange 41ndash50 mean plusmn sd = 468 plusmn 26) There were no signifi-cant differences between the sexes on any of these variables
Nine of the twelve agreed to participate in a follow-up fMRIstudy of face perception conducted so that localization of theFFA could be compared with SAT activations in the middle FGarea in the right hemisphere Five were male (one left-handed)and four were female (one left-handed) mean age was236 plusmn 26 years All subjects gave written informed consent forboth studies in accordance with procedures and protocolsapproved by the Institutional Review Board of the Yale Univer-sity School of Medicine
(b) Experimental tasksWe adapted the SAT for use in a fMRI block-design study by
programming 16 new SAT QuickTime film skits using Directorpublished by Macromedia (600 Townsend Ave San FranciscoCA wwwmacromediacom) From these a final set of eightwere chosen for use in the fMRI study on the basis of ratingsby project staff of the filmrsquos realism and ability to capture onersquossocial attention (these films can be downloaded from httpinfomedyaleeduchldstdyneuroimgsatFmovieshtm) Each filmlasted 151 s and was designed like the original SAT with move-ments intended to suggest a sense of personal agency andreciprocal and contingent interactions that were meant to be eas-ily interpreted as social Each film contained three types of whitegeometric figures (a triangle diamond and circle) that movedagainst a black background In common with the original SATthere was a box in the centre of the field with one wall thatopened as if on a hinge allowing the shapes to open and shutthe door and to enter chase or drag other shapes inside Eachfilm was scripted to follow a social story for example hide-and-seek a fight a love triangle etc The participants were asked todecide by pushing a button if all three of the shapes were lsquofri-endsrsquo or not Half of the films were intended to have lsquoall friendsrsquoas the correct answer (correct answer was determined by a con-sensus-rating process among the developers of the tasks witheach final film version judged to have a clear answer) The filmswere scripted such that any adversarial interactions occurred inthe final few seconds of the film to force the participant toattend throughout to derive the correct answer In creating ourcontrol task for contrast to the SAT we reasoned that each SATfilm requires three important processes
(i) monitoring the movements and physical interactionsbetween the shapes
(ii) pretending that the shapes represent something else iepeople and
(iii) an inferential social reasoning process based on the natureof the interactions (judging whether the movements rep-resent friendly or non-friendly interactions)
A lsquobumper carrsquo control task was created that contained all ofthe elements of the SAT films with the exception of the socialreasoning process This task also entailed eight 151 s filmsdepicting the same geometric shapes moving about a black field
Phil Trans R Soc Lond B (2003)
with the same centrally positioned white box The participantrsquostask was to pretend that these figures were lsquobumper carsrsquomdashsmallracing cars found at amusement parks that are encapsulated byrubber bumpers to allow safe playful collisions Participantsmonitored the carrsquos movements and interactions and decidedon the basis of the carrsquos trajectories and speed after each collisionif the three shapes were all equally lsquoheavyrsquo or not for exampleupon collision if one car shot off more rapidly than anotherthen the two were not equally heavy Key collisions that gaveaway the correct answer always occurred in the final seconds ofthe film Collisions early in the 151 s skit were often mere grazesthat failed to provide definitive information about relative massThus the control task contained the first two elements butinstead of a social decision participants were required to makea decision about a physical property The bumper car and SATfilms were designed to be equivalent with respect to movementquantity and location so that the comparison between the twotasks would reveal the location of brain processes that are dis-tinctly involved in social perceptual and social cognitive pro-cesses It is interesting to note that we piloted a version of thecontrol task that involved physical judgements on the SAT films(as opposed to the bumper car films) but participants reportedthat they were not able to consciously stop seeing the films associal stories Thus it did not seem possible to use the exactsame stimuli in both tasks as might otherwise be desirablebecause social perceptual and cognitive processes would prob-ably be engaged to a greater or lesser extent in both the experi-mental and control tasks Two other lower level controlconditions were also included in each experimental run in theblock design and were intended to further pull apart the threedistinct processes outlined above However the results of thesecontrasts were generally uninformative and thus are not reportedhere Between each film was a 12 s rest period with a blackscreen All participants underwent practice using films that didnot make the final group of eight in order to become completelyfamiliar with the tasks before fMRI scanning During the fMRIexperiment each film was preceded by a 3 s cue lsquoBUMPERCARS SAME WEIGHTrsquo or lsquoPEOPLE ALL FRIENDSrsquoSubjects responded by pressing a button upon completion ofeach film both as a measure of reaction time and accuracy andto ensure that the subjects watched the entire film
In the follow-up study (hereafter lsquoStudy 2rsquo) side-by-side grey-scale faces objects or patterns were presented in asamedifferent task in a block-design experiment to localize theFFA We have previously used this task to localize the FFA ina large group of normal controls (Schultz et al 2000b 2001)Image pairs were presented for 28 s with a 05 s inter-stimulusinterval The person identity task employed same-gender pairsof neutral (non expressive) faces on a black background Pic-tures were taken from standard sources and were edited toremove hair ears and shirt collars so as to force subjects tofocus on features of the face with central relevance to non-verbalsocial communication ie the eyes nose mouth and facegeometry Objects were pictures of spectacles taken from anonline retail catalogue that were contrast inverted to make thebackground black and the spectacles shades of grey to matchthe faces Patterns were distorted versions of the faces or spec-tacles Results from contrasts with patterns were not used tolocalize the FFA and thus are not reported here As is conven-tional the face versus subordinate-level object-discriminationcontrast defined the FFA Each task block lasted 165 s and wasseparated by a 105 s rest period during which cross hairs (1 1)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
418 R T Schultz and others FFA and social cognition
Figure 1 Composite t-map for 12 healthy controls contrasting the social attribution (yellowred) and the bumper car(bluepurple) tasks (p 00005) Right and left are reversed by convention Abbreviations BA Brodman area FG fusiformgyrus IFG inferior frontal gyrus MPFC medial prefrontal cortex STG superior temporal gyrus Y-coordinates are from thesystem of Talairach amp Tournoux (1988)
centred in the same position as the image pairs flashed with thesame presentation rate
(c) Data acquisitionSAT and face perception fMRI data were collected on differ-
ent occasions averaging approximately 25 months apart (range3 weeks to 6 months) Only after the original 12 subjects com-pleted the SAT fMRI study was the decision made to rescansubjects with the face-localization protocol Changes in BOLDcontrast were measured as subjects performed the SAT bumpercar face-discrimination and object-discrimination tasks Thestimuli were run as QuickTime films in Study 1 and as PICTimage files in Study 2 Studies were programmed in PsyScope
125 PPC (Carnegie Mellon University Pittsburgh PA USA)and run on a MacIntosh G3 computer Images were back-pro-jected onto a translucent screen mounted near the end of theMRI gantry and were viewed through a periscopic prism systemon the head coil Behavioural response data were collected witha fibre-optic button box with two response alternatives (Yes orNo for lsquoall friendsrsquo lsquosame weightrsquo lsquosame personrsquo and lsquosameobjectrsquo) The participantrsquos head was immobilized using foamwedges and tape across the forehead
T2 curren weighted images sensitive to BOLD contrast wereacquired on a GE Signa 15 Tesla scanner with a standard quad-
Phil Trans R Soc Lond B (2003)
rature head coil using a gradient echo single-shot echo planarsequence and a coronal orientation perpendicular to the planethrough the ACndashPC The pulse sequence for both studies wasTR = 1500 ms TE = 60 flip angle = 60 NEX = 1 in-planevoxel size = 3125 mm acute 3125 mm In the SAT study we col-lected 14 coronal slices 10 mm thick (skip 1 mm) starting at theanterior-most aspect of the frontal lobe and covering all of thebrain except the caudal-most aspect of the occipital lobe Datawere collected in four runs of an ABCD block design(block = one 151 s film) with blocks of each type presentedtwice per run in a pseudo-random order Across runs a total of80 echo planar images were collected per slice per task con-dition In the face-perception study we also collected 14 coronalslices perpendicular to the ACndashPC starting from the posterioraspect of the occipital cortex up through the rostral-most aspectof the cingulate gyrus Slice thickness was 9 mm (skip 1 mm) tobe compatible with a separate face-perception study ongoing atthat time Data were collected in a block design with a pseudo-random order across six separate runs with three blocks of eachtask per run for 180 echo planar images per slice per task con-dition Functional data in both studies were co-registered to T1-weighted structural images of the same thickness collected in thesame session (TR = 500 TE = 14 field of view = 200 mm256 mm acute 192 mm matrix 2 NEX)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 419
(d) Data analysisData were corrected for motion using SPM99 for three trans-
lation directions and for the three possible rotations (WellcomeDepartment of Cognitive Neurology London UK) Image vol-umes with centre of mass (x y or z) movement of more than15 mm within a run were discarded Image analyses and testsof statistical significance were done using locally developedsoftware (Skudlarski httpmrimedyaleedumembersFframedhtml) Motion corrected images were spatially smoothed with aGaussian filter with a full-width half-maximum value of625 mm The specific effects of each task were evaluated bycreating t-maps for each imaging series incorporating a correc-tion for linear drift (Skudlarski et al 1999) of specific task con-trasts social versus bumper car in Study 1 and face versusobject in Study 2 t-maps were averaged across imaging seriesand co-registered with the higher resolution anatomical imagesfor display and localization These maps were then transformedby in-plane registration and slice interpolation into a pro-portional three-dimensional grid defined by Talairach amp Tour-noux (1988) and averaged across all subjects to createcomposite t-maps with the acquired data in 14 slices interp-olated to 18 slices (NB 16 slices are shown in figure 1 as fMRIactivations on the first and last slice are corrupted by motioncorrection) The SAT versus bumper car maps are displayed inthe figures using a significance level of p 00005(uncorrected) Face versus object t-maps were created and dis-played at p 005 (uncorrected) with the a priori hypothesis thatthe right lateral FG would define the FFA No other brain areasare examined in the second study thus avoiding any multiplecomparison problem
ROI analyses were conducted in the SAT study by tracingsignificant pixels on the group composite activation map (figure1) in the following regions the right FG the right and left STSand STG the right TP the right amygdala and right and leftdorsal MPFC To more thoroughly assess activity in the FGmedial and lateral FG ROIs (and the combined whole FG) weredefined anatomically and traced across the two coronal sliceswhere there was significant SAT activation The activated SATROI for the FG is 33 of the size of the entire anatomicallydefined FG at those two slices Individual subject data wereinterrogated using the ROIs to obtain the mean per cent signalchange for each person for each region and Talairach centre ofmass coordinates (the centre of ROI activation weighted by theamplitude of activation across the region) Mean per cent signal-change data were used in correlational analyses to estimate theconsistency of conjoint activity between ROIs across subjects
3 RESULTS
(a) Behavioural performanceThere were no significant differences between the
social and bumper car tasks in performance accuracy(t1 2 2 = 163 p 010 social = 86 plusmn 16 correctbumper = 74 plusmn 16 correct) or reaction time (t1 22
= 062 p 060) There were no significant differences intask accuracy between males and females or left-handersand right-handers In addition there were no significantcorrelations between task performance and age Full ScaleIntelligence Quotient or Benton Face Recognition per-formance
Phil Trans R Soc Lond B (2003)
(b) Brain activity associated with the socialattribution task
As shown in figure 1 comparison of the SAT with thebumper car control condition resulted in a widely distrib-uted set of significant activations There was very little sig-nificant activation for the bumper car task with the oneregion shown clearly in figure 1 being bilateral activationof the dorsal bank of the intra-parietal sulcus The SATnetwork included a region within the right and left dorsalMPFC the right and left inferior frontal gyrus parsorbitalis and the lateral orbital gyrus the right TP theright amygdala the right and left STS and STG and theright FG It is important to note that at lower thresholds(eg p 001) there was also left amygdala activation anda ROI analysis of the per cent signal change failed to findsignificantly more right than left amygdala activation It isalso worth noting that the FG activations seem quitespecific to the SAT task in the sense that reducing thethreshold down to p 005 failed to show additional ven-tral pathway activation More widespread activationsmight have indicated a general SAT effect on arousal orattention that was manifested throughout the ventralstream but this was not the case
The largest areas of activation were the STG (especiallyon the right) and MPFC Direct comparison of the rightversus left MPFC mean per cent signal change failed tofind significant differences (paired t1 1 = 045 p 050)However the right STG was significantly more activatedthan the left STG (paired t1 1 = 264 p = 002) Table 1presents the Talairach coordinates for the centre of acti-vation mass for each ROI Table 2 presents a correlationalmatrix showing the consistency of conjoint activitybetween regions The strongest correlation is between theright amygdala and the ROI that defines the significantlyactivated region of the right FG (r = 071 p = 001) Inter-estingly this correlation is nominally larger than that ofthe entire FG and the smaller subcomponent FG ROIdefined by the SAT activated pixels (r = 069 p = 0013)Since some correlation would be expected between theseoverlapping ROIs especially since the data were spatiallysmoothed the robust correlation to the amygdala is evenmore impressive Other notable results from the corre-lation matrix include the lack of correlation between theMPFC and either the amygdala or temporal lobe ROIsWithin the temporal lobes however the right STG is sig-nificantly correlated with the left STG and with both defi-nitions of the right FG
Accuracy of performance on the SAT correlates withthe amount of activity in the anatomically defined rightFG (r = 065 p = 002) but not with any other node in theSAT network Females showed significantly more rightSTG activation than males (t1 0 = 234 p = 004) Maleshowever showed significantly more right TP activation(t1 0 = 253 p = 003) There were no other significant sexdifferences and no significant associations with hand-edness or age
(c) Comparison of activity in the right fusiformgyrus during social attribution task and face
perceptionAt the time Study 1 data were collected finding signifi-
cant right FG activation to the SAT was unexpected Toclarify whether the FG activation was in the precise
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
420 R T Schultz and others FFA and social cognition
Table 1 ROI centres of mass coordinates
mean Talairach coordinates (X Y Z )
ROI right hemisphere left hemisphere Brodmann areas
MPFC 49 349 436 274 361 434 6 8 9TP 463 138 2128 mdash 38amygdala 228 2109 2124 mdash mdashFG (SAT) 344 2468 290 mdash 37FG (face) 374 2480 2126 mdash 37STG 507 2571 152 2565 2608 194 22 39
Table 2 ROI correlation matrix(Correlations are based on mean per cent signal change from each ROI (see sect 2 for ROI procedures) SAT accuracy data representthe percentage of films that each participant correctly)
activated whole right right left left right rightright FG FG amygdala amygdala MPFC MPFC left STG STG right TP
whole right FG 069 curren curren curren
right amygdala 071 curren curren curren 032left amygdala 056 curren 019 070 curren curren curren
left MPFC 021 013 032 011right MPFC 2018 2008 021 019 048left STG 021 029 019 005 056 curren 018right STG 054 curren 069 curren curren curren 022 025 009 2016 060 curren curren
right TP 2007 2011 040 033 048 034 038 2010SAT accuracy 045 065 curren curren 2012 009 013 2017 201 022 2034
curren curren curren p lt 001 (r gt 069) curren curren p lt 005 (r gt 057) curren p lt 010
(a) SAT versus bumper car FFA activation
(
(c) enlargement and alignment of FFA
Figure 2 (a) Composite (n = 12) t-map at two slices showing significant (p 00005) activation for the SAT contrast(yellowred) with the bumper car control task (bluepurple) (b) Composite (n = 9) t-map at two slices showing significant(p 005) activation for the face (yellowred) versus object discrimination (bluepurple) This contrast defines the FFA (c)Subregions of composite t-maps shown in (a) and (b) are enlarged and aligned to demonstrate the overlap of activation in theFG for the SAT and face discrimination activations Subscripts 1 and 2 refer to the first (more anterior) and second coronalslices with significant activation
location of the FFA we compared it with the result of theface versus object discrimination contrast (the standardmeans of identifying the FFA in the literature) Both theFFA and the SAT activations of the FG were confined totwo coronal slices in highly overlapping locations Thesetwo sets of group composite maps are shown in figure 2
Phil Trans R Soc Lond B (2003)
The FG activations were stronger in the SAT versusbumper contrast than the face versus object contrast butthis may have had as much do with the baseline as theexperimental tasks The Talairach coordinates for theSAT and FFA activations show that their centre of massdiffers by less than one voxel The SAT activation is 3 mm
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 421
Figure 3 Scans of two individuals across three different occasions showing the reproducibility of FFA activations at two time-points and relationship to SAT activations (andashc) are from a 23-year-old male (dndashf ) are from a 24-year-old female Panels arearranged chronologically (ad) The first face versus object experiment (be) The second face versus object scan (cf ) SATversus bumper car contrast Both coronal slices are shown where there was FFA (t gt 15 in yellowred) or SAT activation(t gt 30 in yellowred) Arrows point to FG activity (right and left are reversed by convention) As in the group results (figure2) the SAT activation is centred slightly more medially along the MFS Left FG activation shown in these two subjects doesnot survive thresholding in the group composite (figure 2) Control tasks (object discrimination bumper car) are shown inpurpleblue
more medial than that of the FFA straddling the MFSthat delineates the lateral and medial aspects of the FGThe FFA on the other hand is clearly positioned in thelateral FG as expected (Haxby et al 1999) The SATactivation is also centred 36 mm more superior and about1 mm more anteriorly than the FFA A count of the over-lapping significant pixels showed that 50 of the SATactivation falls within the FFA This provides a goodapproximation of how these regions overlap Howeverthere is no definitive way to measure the percent overlapin this study because it would change with the use of dif-ferent control tasks in either condition or different signifi-cance levels for thresholding the t-maps
Two of the participants (one male one female) werealso part of a reproducibility study of the FFA and hadboth undergone the face discrimination protocol on twooccasions Figure 3 presents t-maps of their FG for eachface perception study and the SAT The female partici-pant shows reversed asymmetry as sometimes happenswith the left FG showing greater face activation than theright Nevertheless her SAT activations track her FFAand are more left-sided than typical These results showthat the FFA activation is reproducible so that the lessthan perfect overlap between the FFA and the SAT FGactivation is probably not a measurement or reliabilityissue
4 DISCUSSION
(a) The social brain networkThe current study required participants to observe the
movements of geometric figures and to interpret thesewith regard to a conceptual template about what consti-tutes a friendly or unfriendly social interaction It requiredclose attention to the contingent nature of a sequence ofmovements and inferences about mental states of eachcharacter to explain their actions Perception of the move-
Phil Trans R Soc Lond B (2003)
ments of these simple shapes as wilful seems to be auto-matic and effortless for healthy controls but not forpersons with autism (Klin 2000) As shown in figure 1the network engaged by the social attribution process (incontrast to the physical attribution control task) includednearly all of the brain areas implicated by past researchon the social brain (Brothers 1990) including cognitiveaspects such as theory of mind (Castelli et al 2000) aswell as perceptual aspects such as the perception of socialdisplays and biological movement (Allison et al 2000)We found significant activation of the bilateral MPFCsuperior STG and STS and inferior FG pars orbitalisextending into the lateral orbital gyri In addition therewere significant activations on the right side only for theamygdala TP and the FG The predominance of right-side activations is consistent with the notion that the righthemisphere is more concerned than the left with socialprocesses (Siegal et al 1996 Winner et al 2002) Ourresults differ from some past research by finding signifi-cant right FG activation and by failing to find orbito-MPFC activation
Several earlier neuroimaging studies have shown thatthe dorsal MPFC (ie that cortex anterior and superior tothe anterior cingulate gyrus) is a critical substrate for socialjudgements including empathizing and thinking aboutotherrsquos thoughts and intentions (Fletcher et al 1995 Goelet al 1995 Happe et al 1996 Castelli et al 2000 2002Gallagher et al 2000) Our findings are consistent withthese imaging studies and with non-human primate stud-ies that have documented social failures and loss of socialposition within the group following lesions to orbital pre-frontal cortices and MPFC (Butter et al 1969 Myers etal 1973 Bachevalier amp Mishkin 1986) The importantrole for the MPFC in social cognition is further suggestedby studies of autism spectrum disorders that find func-tional abnormalities in this area (Happe et al 1996 Ernstet al 1997 Haznedar et al 1997 Castelli et al 2002)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
422 R T Schultz and others FFA and social cognition
Gusnard et al (2001) suggest that the dorsal MPFC isinvolved in any kind of thought that uses the self as a refer-ent Thus the SAT activations in this area may have beendriven by theorizing about othersrsquo minds but with explicitreference to the participantrsquos own frame of reference as tohow they would feel in a similar situation
The orbital prefrontal cortex and to a lesser extent thedorsal MPFC have dense reciprocal connections withmedial temporal areas (Price et al 1996 Carmichael ampPrice 1995) providing the anatomical bases for a systemthat regulates emotional processes Damasio and col-leagues (Damasio et al 1990 Bechara et al 1996) haveargued that the orbito-MPFC have a primary function ofintegrating information about rewards and punishmentsto bias future behaviour (Rolls 1995 Dias et al 1996Hornak et al 1996 Lane et al 1997 Reiman et al 1997)A functional circuitry such as this would seem especiallyimportant in the development and acquisition of socialbehaviour However even acquired lesions to theseregions in adulthood can result in abnormalities of socialconduct (Damasio et al 1990) Brothers (1990) high-lighted the orbital prefrontal cortex as one of the threeprincipal brain regions involved in social cognition Weobserved poor signal in this region (these areas are notori-ously prone to fMRI signal drop-out and distortion) andthus we cannot know if the orbital prefrontal region wasengaged by the SAT or not However using a similarpsychological task but with PET where signal acquisitionin this region is not degraded Castelli et al (2000 2002)failed to find activation of the orbital prefrontal cortexCollectively these results call into question the impor-tance of the orbital prefrontal cortex in social cognitionand instead shift the focus on anterior cortices toward thedorsal MPFC
The SAT also generated robust activations of the rightamygdala and nearby cortex of the right TP The amyg-dala is often given a central role in theories of social per-ception and cognition (Brothers 1990 Bachevalier 1994Adolphs et al 1998 Baron-Cohen et al 2000 Schultz etal 2000b) The amygdala has a critical role in emotionalarousal assigning significance to environmental stimuliand mediating the formation of visual-reward associationsthat is lsquoemotionalrsquo learning (Gaffan et al 1988 LeDoux1996 Anderson amp Phelps 2001) It is reliably engagedduring judgements of personality characteristics from pic-tures of the face or part of the face (Adolphs et al 1998Baron-Cohen et al 1999 Winston et al 2002) Activationof the amygdala appears to be automatic and stimulusdriven as it can be engaged by images of facial expressionsin conscious awareness as well as by subliminal presen-tations of faces displaying affect (Morris et al 1998Whalen et al 1998 Critchley et al 2000) Thus theamygdalarsquos engagement by the SAT could stem from thegeneral emotional arousal evoked by the animations or itcould represent its computational role in some more spe-cific social perceptual process
The amygdala has dense reciprocal connections withthe ventral visual processing stream (Amaral amp Price1984) The strong correlation observed in this studybetween the right amygdala and the right FG could indi-cate that emotional inputs from the amygdala to the FGare necessary for engaging the social computational pro-cesses of the FG Brothers (1995) has speculated that the
Phil Trans R Soc Lond B (2003)
amygdala generates lsquosocial feelingsrsquo that are of importancein cutting through the complexities of social situations andguiding behaviour by simpler learned contingenciesSocial events are complex because their meaning oftencomes from specific combinations of features that do notadd up in a linear manner making it more difficult todecompose the events by cognitive analysis Effectivesocial engagement requires integration of context historicalrelationships and current socialndashemotional communi-cations expressed through prosodic tone as well as facialexpressions posture and gesture They are made morecomplex by the rapid pace of social transactions Thiswould necessitate some fast response system based ongeneral principles from past socialndashemotional experiencesa role for which the amygdala would seem ideally suitedIn other words the amygdala might drive intuition or lsquogutfeelingsrsquo to guide rapid non-verbal social interactionsinvolving facial expressions gesture etc Thus the strongamygdalandashFG correlation observed here could be inter-preted as the amygdala informing the FG of the relevanceof a social event and also of the outcome of its quick anddirty social perceptual analyses thereby guiding the FG(and other social nodes) in their finer grained compu-tations
Perhaps the largest and strongest activations to the SATwere those of the posterior aspects of the STS spreadinginto the adjacent STG This area has been implicated asa specific site for perception of biological motion (Bondaet al 1996 Allison et al 2000 Grezes et al 2001 Vainaet al 2001) Biological motion is a broad construct thatseems to encompass the perception of static images ofevents that could move or did move such as facialexpressions as well as actual movement by animateobjects This region of the STSndashSTG is also critical forthe decoding visual displays of social action or intention(eg gaze direction gesture and facial displays of emotion)(Critchley et al 2000 Hoffman amp Haxby 2000) In thisregard the SAT activations here are completely expectedand a testament to the effectiveness of these animationsin inducing the desired illusion of anthropomorphicaction The majority of the STSndashSTG activations wereanterior to location of the V5MT as defined in otherrecent fMRI studies (Culham et al 2001) but they alsoextended posteriorly into these more general movementsensitive cortices Unlike the amygdala some evidencesuggests that activity of the STS is mediated by explicitattention to social characteristics of the face and that STSengagement it is not automatic or stimulus driven(Winston et al 2002) We surmise therefore that acti-vation of the STS by the SAT was due to explicit taskinstructions to judge the social interactions
(b) Role of the fusiform face area in socialcognition
We also found significant SAT activation of the rightFG When compared directly in a subset of participants totheir FFA gleaned from a separate scanning session witha perceptual discrimination task of person versus objectidentity we found that both ROIs were confined to theright side on the same two coronal slices There wasabout 50 overlap between the SAT FG and FFA acti-vations and the centre of the two activations differed byless than 1 voxel The FFA activation was offset to the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 423
lateral prominence of the FG whereas the SAT activateda region of the FG that was closer to the centre of the FGbut still largely in the lateral aspect Because the SAT doesnot contain any face representations the FFA engagementwas unexpected as this region is thought of as selectivefor faces (Kanwisher et al 1997) or to other classes ofcomplex objects for which one is perceptually expert(Gauthier et al 2000)
These results have important implications for the poss-ible role of the fusiform in social brain circuitry andautism Interestingly the magnitude of the FG activationpredicted SAT task accuracy no other ROI correlated sig-nificantly with task accuracy This would seem to argueagainst any interpretation that the FFA activation is incon-sequential to the SAT that it is activated simply becauseit is well connected to other areas that are directly involvedin the social attribution process Using a task that is simi-lar to the SAT Castelli et al (2000) also reported signifi-cant right FG activation using PET However in theirfollow-up PET study comparing autism and normal con-trols (Castelli et al 2002) they failed to find significantFG activation in either group The failure of their secondstudy to find right FG activations may be due to the morestringent random effects model used in the data analysesHowever because we find right FG activation with fMRIwhich is more sensitive than PET and because Castelli etal (2000) found it with what amounts to a lower thres-hold it seems quite probable that the effect is real In factwe have preliminary evidence from an ongoing fMRIstudy that reproduces the SAT FG activations in healthycontrols (Schultz et al 2001) In addition there may besome task attributes that differ between our studies andCastelli et alrsquos that might impact into the strength withwhich FG computational processes are evoked
The key question then is why is the FG engaged bythe SAT and what role does the right FG have in socialcognition and perception Currently there are three com-peting theories of the functional organization of the FGand related ventral visual perceptual areas One modelput forth by Haxby Chao Martin and colleagues speci-fies that objects are encoded in a distributed fashion acrossa wide expanse of the ventral temporal-occipital cortex(Haxby et al 2001) They call this the lsquoobject formtypologyrsquo model (Ishai et al 1999) Their data indicatethat object category specificity is achieved by unique spa-tial patterns of activation across this extrastriate visual cor-tex They show that the pattern of activations across thiscortex is diagnostic of object category membership moreso than any localized activation maxima (Haxby et al2001) They also argue that object category perceptioninvolves retrieval of category-related information aboutspecific features and attributes of the object category(Chao et al 2002) They admit that the lsquonature of theinformation about objects that is represented in the ventraltemporal cortex is a great puzzlersquo (Haxby et al 2000 p4) but open the possibility that semantic information maybe important
A second view championed by Kanwisher et al holdsthat several select perceptual categories including facesbodies and spatial layout of places are encoded in highlyspecific locations in a modular fashion (Kanwisher et al1997 Epstein amp Kanwisher 1998 Downing et al 2001Spiridon amp Kanwisher 2002) The FFA is thought to be
Phil Trans R Soc Lond B (2003)
most concerned with discriminating among individualidentities and not with discriminating images at the categ-orical level In fact lesions to the FFA cause a specificdeficit in recognizing individual identities but not in reco-gnizing the general category of face versus non-face(Wada amp Yamamoto 2001) Spiridon amp Kanwisher (2002)provide a partial replication of Haxby et alrsquos (2001) objectform typology model They show that patterns of ventralvisual cortical activation can distinguish object categoriesNevertheless their data show category specificity for selectareas such as the FFA and they argue the ventral visualcortex is not equipotential
Gauthier et al however have argued that the ventraloccipital temporal pathway is organized by the nature ofthe perceptual computations and that these processingbiases are acquired through experience (Gauthier et al2000 Tarr amp Gauthier 2000) For example the parahip-pocampal place area is a function of a bias towards pro-cessing landscapes in terms of their spatial layout becausewe have learned through repeated experiences that this isvery useful information to extract when perceiving land-scapes and related visual images Similarly we learn quiteearly in life that it is important to discriminate faces on anindividual level for example discriminating mother fromothers This bias towards individual identification accord-ing to Gauthier accounts for the FFArsquos apparent modu-larity (Gauthier et al 1997 2000 Tarr amp Gauthier 2000)According to this third model the type of informationneeded and our cumulative experience in processing thatinformation organizes the ventral occipitalndashtemporal path-way into regional centres with preferred modes of pro-cessing
All three models seem to agree that the functionalorganization of the ventral visual cortices is driven by theneed to categorize perceptions into object classes Webelieve that our findings showing FFA activation by non-face objects are consistent with aspects of both the pro-cessing map model of Gauthier and the object formtypology model of Haxby et al First consistent withHaxby et al we suggest that semantic information isimportant to the ventral visual pathway for object categor-ization Second consistent with Gauthier et al we believethat repeated perceptual experience with faces biases thetype of information that FG finds important We proposethat the middle FG area encodes semantic attributes ofpeople because of repeated perceptual experiences withfaces that occur during social interactions In fact wewould guess that most perceptual experiences with facesoccur during social situations and these social situationsoften involve repeated social judgements Thus infor-mation about the social nature of people might be storedin the FG (though not exclusively)
The nature of the semantic information stored in theFG might be restricted to anything that would be helpfulin defining faces as a distinct category of object becausemaking such distinctions appears to be the primary chargeof the ventral visual pathway This would include knowl-edge of people as having personal agency and of havingthe capability to disturb each otherrsquos emotional homeo-stasis (eg to act friendly or unfriendly) By pretendingthat the three shapes in the SAT are people stored knowl-edge about people in social interactions might beretrieved causing the observed activations in the FG
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
424 R T Schultz and others FFA and social cognition
There would be a measure of efficiency from thisarrangement that is having representations and compu-tations of more abstract attributes of people inter-digitatedwith front-end perceptual processes about physical attri-butes Gauthierrsquos claim is that the same group of neuronscan be engaged by perceptually different categories ofobjects it is the type of processing rather than the visualdetails that are important Extrapolating from this wewould argue that the SAT engages a region of the FGoverlapping with the FFA because it demands compu-tational processes to classify the SAT geometric figures aspeople or person-like Chao et al (1999) reach a similarconclusion They argued that activity in the ventral path-way reflects stored information about an object categorynot just physical features They point out that thisarrangement could explain why lesion patients with cate-gory specific perceptual deficits also have trouble retriev-ing general information about that visual category Thuswe would add that the FFA must store general infor-mation about people or some meta-representation oflsquopeoplenessrsquo
(c) Implications for the pathobiology of autismThe SAT used in this current study appears to be an
effective neurobehavioural probe for engaging a distrib-uted network of brain regions involved in different aspectsof social perception and cognition It will be important touse this and similar procedures in persons with autism tobetter define the nature of brain functions in this disorderCastelli et al (2002) have already taken the first step inthis process and they describe a pattern of hypoactivationin the MPFC STS and TPs in autism In an ongoingstudy of autism spectrum disorders we have presentedpreliminary data using the SAT showing that we too findhypoactivation of these regions and in the amygdala andFG (Schultz et al 2001) Thus we predict that futurework in this area will show that the entire social brain net-work is underactive in autism during tasks requiring socialperceptual and social cognitive processing
There are already sufficient data to argue strongly for arole of the FG in the pathobiology of autism No less thanfive previous fMRI studies have shown the FFA region tobe significantly less engaged among persons with autismcompared with controls during face perception tasks(Critchley et al 2000 Dierks et al 2001 Pierce et al2001 Schultz et al 2000a 2001) Activation of the FGby the social judgements in the current study adds animportant piece of evidence in favour of a causal role forthe FG in the pathobiology of autism This conclusionhowever must remain tentative until additional studiesmore precisely define the factors leading to FG activationduring social attribution and gather better data to proveits computational role in social cognition In addition theeffect of FG lesions for social functioning must be clari-fied If the FG is involved in social computations onewould expect to find social cognitive deficits in personswith lesions to this area However social deficits in proso-pagnosics have not been widely reported It might be thatprosopagnosic patients have not been carefully tested onthis dimension In this regard using our behavioural ver-sion of the SAT (Klin 2000) to test social perceptionamong prosopagnosics would be quite interesting Theparallels however between autism and prosopagnosia are
Phil Trans R Soc Lond B (2003)
incomplete because autism is clearly a developmental dis-order whereas the research literature on prosopagnosia ismostly confined to lesions acquired in adolescence oradulthood Among the few cases of developmental proso-pagnosia reported in the literature there is indeed one thathighlights severe social impairments (Kracke 1994) Itmay be that the role of the FG in the development ofautistic symptoms is different from the role of the FG inthe maintenance of social cognitive functions after brainmaturity
It is also possible that the role of the FG in social pro-cesses is dependent on its functional relationships withother nodes in the social brain and that it is the collectiveaction and interaction of the network that is of primaryimportance for social behaviours For example we founda strong correlation across participants in the amount ofFG and amygdala activation A strictly modular view ofthese areas may be inappropriate as the functions of eachnode could be quite dependent on one another and whenonly considered in isolation quite insufficient to supportsocial processes Thus whereas a lesion to the FG mayimpair visual perception it might not have a large impacton social functioning if this depends on an extended net-work that is dynamic and capable of compensatory adjust-ments Dysfunction of the FG may be necessary but notsufficient to produce social deficits Indeed our relianceon modular models of brain functioning may be leadingus astray in our search for causal mechanisms in autismInstead it may be the collective action of a distributedsystem that is critical to the pathobiology of autism TheFG region may be a key partner in this distributed systembut nevertheless just one node and insufficient by itselfto support in any substantive manner social cognitive pro-cesses or to explain social cognitive deficits seen inautism
5 CONCLUSION AND FUTURE DIRECTIONS
Using the SAT we isolated a distributed network ofactivations that conform to the emerging model of thesocial brain Most important we found significant acti-vation of the central aspect of the FG thus adding thisregion to the expanding list of structures involved in socialprocesses The FG region activated by the SAT overlapsin its spatial extent with the FFA with centres of massdiffering by less than 1 voxel We speculate that these SATactivations represent computational processes associatedwith more abstract attributes of people
It is an open question as to whether the substantial over-lap between the FFA and SAT FG activation is due to anoverlapping or shared set of neuronal assemblies This willneed to be clarified by future work Electrophysiologicalstudies show that there are small face-specific patches inthe FG cortex (Allison et al 1999) In the current studysmall patches of face cells and SAT cells could be inter-mingled but distinct and not drawing on any of the sameneuronal assemblies The spatial resolution of fMRI haslimitations that may preclude any definitive answer to thisquestion but there are strategies that can be used toaddress the issue For example the current study designdid not contain SAT and face discriminations in the sameexperimental fMRI series Doing so would enable directcomparisons of the computational demands placed on the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 425
common area of the FFA It also would be informative iffollow-up studies superimposed faces within the geometricfigures of the SAT without changing the film scripts inany way We could then determine if the computations ofthe common area of the FFA increase in a predictablefashionmdashwould the activation be a linear summation ofthe original SAT plus face discrimination Any significantdeviation from an additive model would suggest that thereis some sharing of neuronal assemblies with the SAT andface discrimination tasks when presented alone
This work was supported by grants from the National Institutesof Child Health and Human Development (grants PO1 HD03008 and PO1 HDDC35482) and the Korczak FoundationThe authors thank the anonymous reviewers for their helpfulcomments Michael Lee for programming the fMRI tasksHedy Serofin and Terry Hickey for their assistance in acquiringthe fMRI data John Herrington for his valuable assistance indata analyses of preliminary studies and members of the YaleDevelopmental Neuroimaging Program for their helpful com-ments on this manuscript A special acknowledgement ofappreciation is given to Donald Cohen who passed away inOctober of 2001 for his outstanding mentorship in all aspectsof this program of work
REFERENCES
Abell F Happe F amp Frith U 2000 Do triangles play tricksAttribution of mental states to animated shapes in normaland abnormal development J Cogn Dev 15 1ndash20
Adolphs R Tranel D amp Damasio A R 1998 The humanamygdala in social judgement Nature 393 470ndash474
Allison T Puce A Spencer D D amp McCarthy G 1999Electrophysiological studies of human face perception Ipotentials generated in occipitotemporal cortex by face andnon-face stimuli Cerebr Cortex 9 415ndash430
Allison T Puce A amp McCarthy G 2000 Social perceptionfrom visual cues role of the STS region Trends Cogn Sci4 267ndash278
Amaral D G amp Price J L 1984 Amygdalo-cortical projec-tions in the monkey (Macaca fascicularis) J Comp Neurol230 465ndash496
Amaral D G Price J L Pitkanen A amp Carmichael S T1992 Anatomical organization of the primate amygdaloidcomplex In The amygdala neurobiological aspects of emotionmemory and mental dysfunction (ed J Aggleton) pp 1ndash66New York Wiley-Liss
Anderson A K amp Phelps E A 2001 Lesions of the humanamygdala impair enhanced perception of emotionally salientevents Nature 411 305ndash309
Bachevalier J 1994 Medial temporal lobe structures andautism a review of clinical and experimental findingsNeuropsychologia 32 627ndash648
Bachevalier J amp Mishkin M 1986 Visual recognition impair-ment follows ventromedial but not dorsolateral prefrontallesions in monkeys Behav Brain Res 20 249ndash261
Baron-Cohen S Ring H A Wheelwright S BullmoreE T Brammer M J Simmons A amp Williams S C R1999 Social intelligence in the normal and autistic brain anfMRI study Eur J Neurosci 11 1891ndash1898
Baron-Cohen S Ring H A Bullmore E T WheelwrightS Ashwina C amp Williams S C R 2000 The amygdalatheory of autism Neurosci Biobehav Rev 24 355ndash364
Bechara A Tranel D Damasio H amp Damasio A R 1996Failure to respond autonomically to anticipated future out-comes following damage to prefrontal cortex Cerebr Cortex6 215ndash225
Benton A 1994 Face recognition Los Angeles CA WesternPsychological Services
Phil Trans R Soc Lond B (2003)
Bonda E Petrides M Ostry D amp Evans A 1996 Specificinvolvement of human parietal systems and the amygdala inthe perception of biological motion J Neurosci 16 3737ndash3744
Bowler D M amp Thommen E 2000 Attribution of mechan-ical and social causality to animated displays by childrenwith autism Autism 4 147ndash171
Brothers L 1990 The social brain a project for integratingprimate behavior and neurophysiology in a new domainConcepts Neurosci 1 27ndash151
Brothers L 1995 Neurophysiology of the perception of inten-tion by primates In The cognitive neurosciences 2nd edn (edM S Gazzaniga) pp 1107ndash1115 Cambridge MA MITPress
Butter C M McDonald J A amp Snyder D R 1969 Oralitypreference behavior and reinforcement value of nonfoodobject in monkeys with orbital frontal lesions Science 1641306ndash1307
Carmichael S T amp Price J L 1995 Limbic connections ofthe orbital and medial prefrontal cortex in macaque monk-eys J Comp Neurol 363 615ndash641
Castelli F Happe F Frith U amp Frith C 2000 Movementand mind a functional imaging study of perception andinterpretation of complex intentional movement patternsNeuroImage 12 314ndash325
Castelli F Frith C Happe F amp Frith U 2002 AutismAsperger syndrome and brain mechanisms for the attri-bution of mental states to animated shapes Brain 1251839ndash1849
Chao L L Haxby J V amp Martin A 1999 Attribute-basedneural substrates in posterior temporal cortex for perceivingand knowing about objects Nat Neurosci 2 913ndash919
Chao L L Weisberg J amp Martin A 2002 Experience-dependent modulation of category-related cortical activityCerebr Cortex 12 545ndash551
Critchley H D (and 10 others) 2000 The functional neuroan-atomy of social behaviour changes in cerebral blood flowwhen people with autistic disorder process facial expressionsBrain 123 2203ndash2212
Culham J C Cavanagh P amp Kanwisher N G 2001 Atten-tion response functions characterizing brain areas usingfMRI activation during parametric variations of attentionalload Neuron 32 737ndash745
Damasio A R Tranel D amp Damasio H 1990 Individualswith sociopathic behavior caused by frontal damage fail torespond autonomically to social stimuli Behav Brain Res41 81ndash94
Dierks T Bolte S Huble D Lanfermannm H amp PoustkaF 2001 Alterations of face processing strategies in autism(a fMRI study) Presented at the 6th Annual Meeting of theOrganization for Human Brain Mapping Brighton UK 10ndash14 June 2001
Dias R Robbins T W amp Roberts A C 1996 Dissociationin prefrontal cortex of affective and attentional shifts Nature380 69ndash72
Downing P E Jiang Y Shuman M amp Kanwisher N 2001A cortical area selective for visual processing of the humanbody Science 293 2470ndash2473
Epstein R amp Kanwisher N 1998 A cortical representation ofthe local visual environment Nature 392 598ndash601
Ernst M Zametkin A J Matochik J A Pascualvaca D ampCohen R M 1997 Reduced medial prefrontal dopami-nergic activity in autistic children Lancet 350 638
Fletcher P C Happe F Frith U Baker S C DolanR J Frackowiak R S amp Frith C D 1995 Other minds inthe brain a functional imaging study of lsquotheory of mindrsquo instory comprehension Cognition 57 109ndash128
Frith C D amp Frith U 1999 Interacting minds a biologicalbasis Science 286 1692ndash1695
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
426 R T Schultz and others FFA and social cognition
Fujita I Tanaka K Ito M amp Cheng K 1992 Columns forvisual features of objects in monkey inferotemporal cortexNature 360 343ndash346
Gaffan E A Gaffan D amp Harrison S 1988 Disconnectionof the amygdala from visual association cortex impairs vis-ual-reward association learning in monkeys J Neurosci 83144ndash3150
Gallagher H L Happe F Brunswick N Fletcher P CFrith U amp Frith C D 2000 Reading the mind in cartoonsand stories an fMRI study of lsquotheory of mindrsquo in verbal andnonverbal tasks Neuropsychologia 38 11ndash21
Gauthier I Anderson A W Tarr M J Skudlarski P ampGore J C 1997 Levels of categorization in visual recog-nition studied using functional magnetic resonance imagingCurr Biol 7 645ndash651
Gauthier I Tarr M J Anderson A W Skudlarski P ampGore J C 1999 Activation of the middle fusiform face areaincreases with expertise in recognizing novel objects NatureNeurosci 2 568ndash573
Gauthier I Skudlarski P Gore J C amp Anderson A W2000 Expertise for cars and birds recruits brain areasinvolved in face recognition Nature Neurosci 3 191ndash197
Goel V Grafman J Sadato N amp Hallett M 1995 Mode-ling other minds Neuroreport 6 1741ndash1746
Grelotti D Gauthier I amp Schultz R T 2001 Social interestand the development of cortical face specialization whatautism teaches us about face processing Devl Psychobiol 40213ndash225
Grezes J Fonlupt P Bertenthal B amp Delon-Martin C2001 Does perception of biological motion rely on specificbrain regions NeuroImage 13 775ndash785
Gusnard D A Akbudak E Shulman G L amp RaichleM E 2001 Medial prefrontal cortex and self-referentialmental activity relation to a default mode of brain functionProc Natl Acad Sci USA 98 4259ndash4264
Happe F Ehlers S Fletcher P Frith U Johansson MGillberg C Dolan R Frackowiak R amp Frith C 1996lsquoTheory of mindrsquo in the brain Evidence from a PET scanstudy of Asperger syndrome Neuroreport 8 197ndash201
Haxby J V Ungerleider L G Clark V P Schouten J LHoffman E A amp Martin A 1999 The effect of face inver-sion on activity in human neural systems for face and objectperception Neuron 22 189ndash199
Haxby J V Isahai A Chao L L Ungerleider L G ampMartin A 2000 Object-form topology in the ventral tem-poral lobe response to I Gauthier (2000) Trends Cogn Sci4 3ndash4
Haxby J V Gobbini M I Furey M L Ishai ASchouten J L amp Pietrini P 2001 Distributed and overlap-ping representations of faces and objects in ventral temporalcortex Science 293 2425ndash2430
Haznedar M M Buchsbaum M S Metzger M Solim-ando A Spiegel-Cohen J amp Hollander E 1997 Anteriorcingulate gyrus volume and glucose metabolism in autisticdisorder Am J Psychiat 154 1047ndash1050
Heider F amp Simmel M 1944 An experimental study ofapparent behavior Am J Psychol 57 243ndash259
Hobson R P Ouston J amp Lee A 1988a Whatrsquos in a faceThe case of autism Br J Psychol 79 441ndash453
Hobson R P Ouston J amp Lee A 1988b Emotion recog-nition in autism coordinating faces and voices Psychol Med18 911ndash923
Hoffman E A amp Haxby J V 2000 Distinct representationsof eye gaze and identity in the distributed human neural sys-tem for face perception Nature Neurosci 3 80ndash84
Hornak J Rolls E T amp Wade D 1996 Face and voiceexpression identification in patients with emotional andbehavioural changes following ventral frontal lobe damageNeuropsychologia 34 247ndash261
Phil Trans R Soc Lond B (2003)
Ishai A Ungerleider L G Martin A Schouten J L ampHaxby J V 1999 Distributed representation of objects inthe human ventral visual pathway Proc Natl Acad Sci USA96 9379ndash9384
Kanwisher N 2000 Domain specificity in face perceptionNature Neurosci 3 759ndash763
Kanwisher N McDermott J amp Chun M M 1997 The fusi-form face area a module of extrastriate cortex specializedfor face perception J Neurosci 17 4302ndash4311
Klin A 2000 Attributing social meaning to ambiguous visualstimuli in higher functioning autism and Asperger syndromethe social attribution task J Child Psychol Psychiat 41831ndash846
Klin A Sparrow S S de Bildt A Cicchetti D V CohenD J amp Volkmar F R 1999 A normed study of face recog-nition in autism and related disorders J Autism Devl Dis-orders 29 499ndash508
Klin A Jones W Schultz R T Volkmar F amp Cohen D J2002 Visual fixation patterns during viewing of naturalisticsocial situations as predictors of social competence in indi-viduals with autism Arch Gen Psychiat 59 809ndash816
Kracke I 1994 Developmental prosopagnosia in Aspergersyndrome presentation and discussion of an individual caseDevl Med Child Neurol 36 873ndash886
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J 1997 Neuroanatomical correlates of happi-ness sadness and disgust Am J Psychiat 154 926ndash933
Langdell T 1978 Recognition of faces an approach for thestudy of autism J Child Psychol Psychiat 19 255ndash268
LeDoux J E 1996 The emotional brain New York Simon ampShuster
Lowel S amp Singer W 1992 Selection of intrinsic horizontalconnections in the visual cortex by correlated neuronalactivity Science 255 209ndash212
Macdonald H Rutter M Howlin P Rios P Le ConteurA Evered C amp Folstein S 1989 Recognition andexpression of emotional cues by autistic and normal adultsJ Child Psychol Psychiat 30 865ndash877
Morris J S Ohman A amp Dolan R J 1998 Conscious andunconscious emotional learning in the human amygdalaNature 393 467ndash470
Myers R E Swett C amp Miller M 1973 Loss of social groupaffinity following prefrontal lesions in free-rangingmacaques Brain Res 64 257ndash269
Osterling J amp Dawson G 1994 Early recognition of childrenwith autism a study of first birthday home video tapes JAutism Devl Disorders 24 247ndash257
Pierce K Muller R A Ambrose J Allen G amp Cour-chesne E 2001 Face processing occurs outside the fusiformlsquoface arearsquo in autism evidence from functional MRI Brain124 2059ndash2073
Price J L Carmichael S T amp Drevets W C 1996 Net-works related to the orbital and medial prefrontal cortex asubstrate for emotional behavior Prog Brain Res 107523ndash536
Puce A Allison T Gore J C amp McCarthy G 1995 Face-sensitive regions in human extrastriate cortex studied byfunctional MRI J Neurophysiol 74 1192ndash1199
Reiman E M Lane R D Ahern G L Schwartz G EDavidson R J Friston K J Yun L S amp Chen K 1997Neuroanatomical correlates of externally and internally gen-erated human emotion Am J Psychiat 154 918ndash925
Rolls E T 1995 A theory of emotion and consciousness andits application to understanding the neural basis of emotionIn The cognitive neurosciences (ed M S Gazzaniga) pp1091ndash1106 Cambridge MA MIT Press
Schultz R T Gauthier I Klin A Fulbright R K Ander-son A W Volkmar F Skudlarski P Lacadie C CohenD J amp Gore J C 2000a Abnormal ventral temporal
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
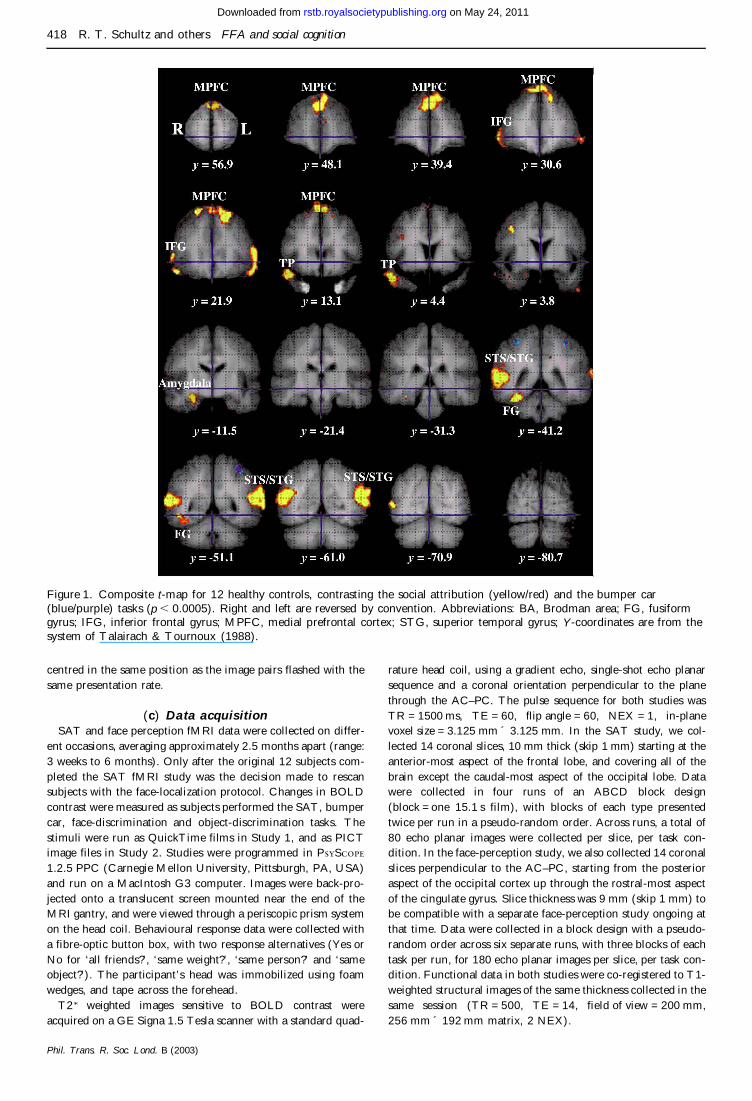
418 R T Schultz and others FFA and social cognition
Figure 1 Composite t-map for 12 healthy controls contrasting the social attribution (yellowred) and the bumper car(bluepurple) tasks (p 00005) Right and left are reversed by convention Abbreviations BA Brodman area FG fusiformgyrus IFG inferior frontal gyrus MPFC medial prefrontal cortex STG superior temporal gyrus Y-coordinates are from thesystem of Talairach amp Tournoux (1988)
centred in the same position as the image pairs flashed with thesame presentation rate
(c) Data acquisitionSAT and face perception fMRI data were collected on differ-
ent occasions averaging approximately 25 months apart (range3 weeks to 6 months) Only after the original 12 subjects com-pleted the SAT fMRI study was the decision made to rescansubjects with the face-localization protocol Changes in BOLDcontrast were measured as subjects performed the SAT bumpercar face-discrimination and object-discrimination tasks Thestimuli were run as QuickTime films in Study 1 and as PICTimage files in Study 2 Studies were programmed in PsyScope
125 PPC (Carnegie Mellon University Pittsburgh PA USA)and run on a MacIntosh G3 computer Images were back-pro-jected onto a translucent screen mounted near the end of theMRI gantry and were viewed through a periscopic prism systemon the head coil Behavioural response data were collected witha fibre-optic button box with two response alternatives (Yes orNo for lsquoall friendsrsquo lsquosame weightrsquo lsquosame personrsquo and lsquosameobjectrsquo) The participantrsquos head was immobilized using foamwedges and tape across the forehead
T2 curren weighted images sensitive to BOLD contrast wereacquired on a GE Signa 15 Tesla scanner with a standard quad-
Phil Trans R Soc Lond B (2003)
rature head coil using a gradient echo single-shot echo planarsequence and a coronal orientation perpendicular to the planethrough the ACndashPC The pulse sequence for both studies wasTR = 1500 ms TE = 60 flip angle = 60 NEX = 1 in-planevoxel size = 3125 mm acute 3125 mm In the SAT study we col-lected 14 coronal slices 10 mm thick (skip 1 mm) starting at theanterior-most aspect of the frontal lobe and covering all of thebrain except the caudal-most aspect of the occipital lobe Datawere collected in four runs of an ABCD block design(block = one 151 s film) with blocks of each type presentedtwice per run in a pseudo-random order Across runs a total of80 echo planar images were collected per slice per task con-dition In the face-perception study we also collected 14 coronalslices perpendicular to the ACndashPC starting from the posterioraspect of the occipital cortex up through the rostral-most aspectof the cingulate gyrus Slice thickness was 9 mm (skip 1 mm) tobe compatible with a separate face-perception study ongoing atthat time Data were collected in a block design with a pseudo-random order across six separate runs with three blocks of eachtask per run for 180 echo planar images per slice per task con-dition Functional data in both studies were co-registered to T1-weighted structural images of the same thickness collected in thesame session (TR = 500 TE = 14 field of view = 200 mm256 mm acute 192 mm matrix 2 NEX)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 419
(d) Data analysisData were corrected for motion using SPM99 for three trans-
lation directions and for the three possible rotations (WellcomeDepartment of Cognitive Neurology London UK) Image vol-umes with centre of mass (x y or z) movement of more than15 mm within a run were discarded Image analyses and testsof statistical significance were done using locally developedsoftware (Skudlarski httpmrimedyaleedumembersFframedhtml) Motion corrected images were spatially smoothed with aGaussian filter with a full-width half-maximum value of625 mm The specific effects of each task were evaluated bycreating t-maps for each imaging series incorporating a correc-tion for linear drift (Skudlarski et al 1999) of specific task con-trasts social versus bumper car in Study 1 and face versusobject in Study 2 t-maps were averaged across imaging seriesand co-registered with the higher resolution anatomical imagesfor display and localization These maps were then transformedby in-plane registration and slice interpolation into a pro-portional three-dimensional grid defined by Talairach amp Tour-noux (1988) and averaged across all subjects to createcomposite t-maps with the acquired data in 14 slices interp-olated to 18 slices (NB 16 slices are shown in figure 1 as fMRIactivations on the first and last slice are corrupted by motioncorrection) The SAT versus bumper car maps are displayed inthe figures using a significance level of p 00005(uncorrected) Face versus object t-maps were created and dis-played at p 005 (uncorrected) with the a priori hypothesis thatthe right lateral FG would define the FFA No other brain areasare examined in the second study thus avoiding any multiplecomparison problem
ROI analyses were conducted in the SAT study by tracingsignificant pixels on the group composite activation map (figure1) in the following regions the right FG the right and left STSand STG the right TP the right amygdala and right and leftdorsal MPFC To more thoroughly assess activity in the FGmedial and lateral FG ROIs (and the combined whole FG) weredefined anatomically and traced across the two coronal sliceswhere there was significant SAT activation The activated SATROI for the FG is 33 of the size of the entire anatomicallydefined FG at those two slices Individual subject data wereinterrogated using the ROIs to obtain the mean per cent signalchange for each person for each region and Talairach centre ofmass coordinates (the centre of ROI activation weighted by theamplitude of activation across the region) Mean per cent signal-change data were used in correlational analyses to estimate theconsistency of conjoint activity between ROIs across subjects
3 RESULTS
(a) Behavioural performanceThere were no significant differences between the
social and bumper car tasks in performance accuracy(t1 2 2 = 163 p 010 social = 86 plusmn 16 correctbumper = 74 plusmn 16 correct) or reaction time (t1 22
= 062 p 060) There were no significant differences intask accuracy between males and females or left-handersand right-handers In addition there were no significantcorrelations between task performance and age Full ScaleIntelligence Quotient or Benton Face Recognition per-formance
Phil Trans R Soc Lond B (2003)
(b) Brain activity associated with the socialattribution task
As shown in figure 1 comparison of the SAT with thebumper car control condition resulted in a widely distrib-uted set of significant activations There was very little sig-nificant activation for the bumper car task with the oneregion shown clearly in figure 1 being bilateral activationof the dorsal bank of the intra-parietal sulcus The SATnetwork included a region within the right and left dorsalMPFC the right and left inferior frontal gyrus parsorbitalis and the lateral orbital gyrus the right TP theright amygdala the right and left STS and STG and theright FG It is important to note that at lower thresholds(eg p 001) there was also left amygdala activation anda ROI analysis of the per cent signal change failed to findsignificantly more right than left amygdala activation It isalso worth noting that the FG activations seem quitespecific to the SAT task in the sense that reducing thethreshold down to p 005 failed to show additional ven-tral pathway activation More widespread activationsmight have indicated a general SAT effect on arousal orattention that was manifested throughout the ventralstream but this was not the case
The largest areas of activation were the STG (especiallyon the right) and MPFC Direct comparison of the rightversus left MPFC mean per cent signal change failed tofind significant differences (paired t1 1 = 045 p 050)However the right STG was significantly more activatedthan the left STG (paired t1 1 = 264 p = 002) Table 1presents the Talairach coordinates for the centre of acti-vation mass for each ROI Table 2 presents a correlationalmatrix showing the consistency of conjoint activitybetween regions The strongest correlation is between theright amygdala and the ROI that defines the significantlyactivated region of the right FG (r = 071 p = 001) Inter-estingly this correlation is nominally larger than that ofthe entire FG and the smaller subcomponent FG ROIdefined by the SAT activated pixels (r = 069 p = 0013)Since some correlation would be expected between theseoverlapping ROIs especially since the data were spatiallysmoothed the robust correlation to the amygdala is evenmore impressive Other notable results from the corre-lation matrix include the lack of correlation between theMPFC and either the amygdala or temporal lobe ROIsWithin the temporal lobes however the right STG is sig-nificantly correlated with the left STG and with both defi-nitions of the right FG
Accuracy of performance on the SAT correlates withthe amount of activity in the anatomically defined rightFG (r = 065 p = 002) but not with any other node in theSAT network Females showed significantly more rightSTG activation than males (t1 0 = 234 p = 004) Maleshowever showed significantly more right TP activation(t1 0 = 253 p = 003) There were no other significant sexdifferences and no significant associations with hand-edness or age
(c) Comparison of activity in the right fusiformgyrus during social attribution task and face
perceptionAt the time Study 1 data were collected finding signifi-
cant right FG activation to the SAT was unexpected Toclarify whether the FG activation was in the precise
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
420 R T Schultz and others FFA and social cognition
Table 1 ROI centres of mass coordinates
mean Talairach coordinates (X Y Z )
ROI right hemisphere left hemisphere Brodmann areas
MPFC 49 349 436 274 361 434 6 8 9TP 463 138 2128 mdash 38amygdala 228 2109 2124 mdash mdashFG (SAT) 344 2468 290 mdash 37FG (face) 374 2480 2126 mdash 37STG 507 2571 152 2565 2608 194 22 39
Table 2 ROI correlation matrix(Correlations are based on mean per cent signal change from each ROI (see sect 2 for ROI procedures) SAT accuracy data representthe percentage of films that each participant correctly)
activated whole right right left left right rightright FG FG amygdala amygdala MPFC MPFC left STG STG right TP
whole right FG 069 curren curren curren
right amygdala 071 curren curren curren 032left amygdala 056 curren 019 070 curren curren curren
left MPFC 021 013 032 011right MPFC 2018 2008 021 019 048left STG 021 029 019 005 056 curren 018right STG 054 curren 069 curren curren curren 022 025 009 2016 060 curren curren
right TP 2007 2011 040 033 048 034 038 2010SAT accuracy 045 065 curren curren 2012 009 013 2017 201 022 2034
curren curren curren p lt 001 (r gt 069) curren curren p lt 005 (r gt 057) curren p lt 010
(a) SAT versus bumper car FFA activation
(
(c) enlargement and alignment of FFA
Figure 2 (a) Composite (n = 12) t-map at two slices showing significant (p 00005) activation for the SAT contrast(yellowred) with the bumper car control task (bluepurple) (b) Composite (n = 9) t-map at two slices showing significant(p 005) activation for the face (yellowred) versus object discrimination (bluepurple) This contrast defines the FFA (c)Subregions of composite t-maps shown in (a) and (b) are enlarged and aligned to demonstrate the overlap of activation in theFG for the SAT and face discrimination activations Subscripts 1 and 2 refer to the first (more anterior) and second coronalslices with significant activation
location of the FFA we compared it with the result of theface versus object discrimination contrast (the standardmeans of identifying the FFA in the literature) Both theFFA and the SAT activations of the FG were confined totwo coronal slices in highly overlapping locations Thesetwo sets of group composite maps are shown in figure 2
Phil Trans R Soc Lond B (2003)
The FG activations were stronger in the SAT versusbumper contrast than the face versus object contrast butthis may have had as much do with the baseline as theexperimental tasks The Talairach coordinates for theSAT and FFA activations show that their centre of massdiffers by less than one voxel The SAT activation is 3 mm
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 421
Figure 3 Scans of two individuals across three different occasions showing the reproducibility of FFA activations at two time-points and relationship to SAT activations (andashc) are from a 23-year-old male (dndashf ) are from a 24-year-old female Panels arearranged chronologically (ad) The first face versus object experiment (be) The second face versus object scan (cf ) SATversus bumper car contrast Both coronal slices are shown where there was FFA (t gt 15 in yellowred) or SAT activation(t gt 30 in yellowred) Arrows point to FG activity (right and left are reversed by convention) As in the group results (figure2) the SAT activation is centred slightly more medially along the MFS Left FG activation shown in these two subjects doesnot survive thresholding in the group composite (figure 2) Control tasks (object discrimination bumper car) are shown inpurpleblue
more medial than that of the FFA straddling the MFSthat delineates the lateral and medial aspects of the FGThe FFA on the other hand is clearly positioned in thelateral FG as expected (Haxby et al 1999) The SATactivation is also centred 36 mm more superior and about1 mm more anteriorly than the FFA A count of the over-lapping significant pixels showed that 50 of the SATactivation falls within the FFA This provides a goodapproximation of how these regions overlap Howeverthere is no definitive way to measure the percent overlapin this study because it would change with the use of dif-ferent control tasks in either condition or different signifi-cance levels for thresholding the t-maps
Two of the participants (one male one female) werealso part of a reproducibility study of the FFA and hadboth undergone the face discrimination protocol on twooccasions Figure 3 presents t-maps of their FG for eachface perception study and the SAT The female partici-pant shows reversed asymmetry as sometimes happenswith the left FG showing greater face activation than theright Nevertheless her SAT activations track her FFAand are more left-sided than typical These results showthat the FFA activation is reproducible so that the lessthan perfect overlap between the FFA and the SAT FGactivation is probably not a measurement or reliabilityissue
4 DISCUSSION
(a) The social brain networkThe current study required participants to observe the
movements of geometric figures and to interpret thesewith regard to a conceptual template about what consti-tutes a friendly or unfriendly social interaction It requiredclose attention to the contingent nature of a sequence ofmovements and inferences about mental states of eachcharacter to explain their actions Perception of the move-
Phil Trans R Soc Lond B (2003)
ments of these simple shapes as wilful seems to be auto-matic and effortless for healthy controls but not forpersons with autism (Klin 2000) As shown in figure 1the network engaged by the social attribution process (incontrast to the physical attribution control task) includednearly all of the brain areas implicated by past researchon the social brain (Brothers 1990) including cognitiveaspects such as theory of mind (Castelli et al 2000) aswell as perceptual aspects such as the perception of socialdisplays and biological movement (Allison et al 2000)We found significant activation of the bilateral MPFCsuperior STG and STS and inferior FG pars orbitalisextending into the lateral orbital gyri In addition therewere significant activations on the right side only for theamygdala TP and the FG The predominance of right-side activations is consistent with the notion that the righthemisphere is more concerned than the left with socialprocesses (Siegal et al 1996 Winner et al 2002) Ourresults differ from some past research by finding signifi-cant right FG activation and by failing to find orbito-MPFC activation
Several earlier neuroimaging studies have shown thatthe dorsal MPFC (ie that cortex anterior and superior tothe anterior cingulate gyrus) is a critical substrate for socialjudgements including empathizing and thinking aboutotherrsquos thoughts and intentions (Fletcher et al 1995 Goelet al 1995 Happe et al 1996 Castelli et al 2000 2002Gallagher et al 2000) Our findings are consistent withthese imaging studies and with non-human primate stud-ies that have documented social failures and loss of socialposition within the group following lesions to orbital pre-frontal cortices and MPFC (Butter et al 1969 Myers etal 1973 Bachevalier amp Mishkin 1986) The importantrole for the MPFC in social cognition is further suggestedby studies of autism spectrum disorders that find func-tional abnormalities in this area (Happe et al 1996 Ernstet al 1997 Haznedar et al 1997 Castelli et al 2002)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
422 R T Schultz and others FFA and social cognition
Gusnard et al (2001) suggest that the dorsal MPFC isinvolved in any kind of thought that uses the self as a refer-ent Thus the SAT activations in this area may have beendriven by theorizing about othersrsquo minds but with explicitreference to the participantrsquos own frame of reference as tohow they would feel in a similar situation
The orbital prefrontal cortex and to a lesser extent thedorsal MPFC have dense reciprocal connections withmedial temporal areas (Price et al 1996 Carmichael ampPrice 1995) providing the anatomical bases for a systemthat regulates emotional processes Damasio and col-leagues (Damasio et al 1990 Bechara et al 1996) haveargued that the orbito-MPFC have a primary function ofintegrating information about rewards and punishmentsto bias future behaviour (Rolls 1995 Dias et al 1996Hornak et al 1996 Lane et al 1997 Reiman et al 1997)A functional circuitry such as this would seem especiallyimportant in the development and acquisition of socialbehaviour However even acquired lesions to theseregions in adulthood can result in abnormalities of socialconduct (Damasio et al 1990) Brothers (1990) high-lighted the orbital prefrontal cortex as one of the threeprincipal brain regions involved in social cognition Weobserved poor signal in this region (these areas are notori-ously prone to fMRI signal drop-out and distortion) andthus we cannot know if the orbital prefrontal region wasengaged by the SAT or not However using a similarpsychological task but with PET where signal acquisitionin this region is not degraded Castelli et al (2000 2002)failed to find activation of the orbital prefrontal cortexCollectively these results call into question the impor-tance of the orbital prefrontal cortex in social cognitionand instead shift the focus on anterior cortices toward thedorsal MPFC
The SAT also generated robust activations of the rightamygdala and nearby cortex of the right TP The amyg-dala is often given a central role in theories of social per-ception and cognition (Brothers 1990 Bachevalier 1994Adolphs et al 1998 Baron-Cohen et al 2000 Schultz etal 2000b) The amygdala has a critical role in emotionalarousal assigning significance to environmental stimuliand mediating the formation of visual-reward associationsthat is lsquoemotionalrsquo learning (Gaffan et al 1988 LeDoux1996 Anderson amp Phelps 2001) It is reliably engagedduring judgements of personality characteristics from pic-tures of the face or part of the face (Adolphs et al 1998Baron-Cohen et al 1999 Winston et al 2002) Activationof the amygdala appears to be automatic and stimulusdriven as it can be engaged by images of facial expressionsin conscious awareness as well as by subliminal presen-tations of faces displaying affect (Morris et al 1998Whalen et al 1998 Critchley et al 2000) Thus theamygdalarsquos engagement by the SAT could stem from thegeneral emotional arousal evoked by the animations or itcould represent its computational role in some more spe-cific social perceptual process
The amygdala has dense reciprocal connections withthe ventral visual processing stream (Amaral amp Price1984) The strong correlation observed in this studybetween the right amygdala and the right FG could indi-cate that emotional inputs from the amygdala to the FGare necessary for engaging the social computational pro-cesses of the FG Brothers (1995) has speculated that the
Phil Trans R Soc Lond B (2003)
amygdala generates lsquosocial feelingsrsquo that are of importancein cutting through the complexities of social situations andguiding behaviour by simpler learned contingenciesSocial events are complex because their meaning oftencomes from specific combinations of features that do notadd up in a linear manner making it more difficult todecompose the events by cognitive analysis Effectivesocial engagement requires integration of context historicalrelationships and current socialndashemotional communi-cations expressed through prosodic tone as well as facialexpressions posture and gesture They are made morecomplex by the rapid pace of social transactions Thiswould necessitate some fast response system based ongeneral principles from past socialndashemotional experiencesa role for which the amygdala would seem ideally suitedIn other words the amygdala might drive intuition or lsquogutfeelingsrsquo to guide rapid non-verbal social interactionsinvolving facial expressions gesture etc Thus the strongamygdalandashFG correlation observed here could be inter-preted as the amygdala informing the FG of the relevanceof a social event and also of the outcome of its quick anddirty social perceptual analyses thereby guiding the FG(and other social nodes) in their finer grained compu-tations
Perhaps the largest and strongest activations to the SATwere those of the posterior aspects of the STS spreadinginto the adjacent STG This area has been implicated asa specific site for perception of biological motion (Bondaet al 1996 Allison et al 2000 Grezes et al 2001 Vainaet al 2001) Biological motion is a broad construct thatseems to encompass the perception of static images ofevents that could move or did move such as facialexpressions as well as actual movement by animateobjects This region of the STSndashSTG is also critical forthe decoding visual displays of social action or intention(eg gaze direction gesture and facial displays of emotion)(Critchley et al 2000 Hoffman amp Haxby 2000) In thisregard the SAT activations here are completely expectedand a testament to the effectiveness of these animationsin inducing the desired illusion of anthropomorphicaction The majority of the STSndashSTG activations wereanterior to location of the V5MT as defined in otherrecent fMRI studies (Culham et al 2001) but they alsoextended posteriorly into these more general movementsensitive cortices Unlike the amygdala some evidencesuggests that activity of the STS is mediated by explicitattention to social characteristics of the face and that STSengagement it is not automatic or stimulus driven(Winston et al 2002) We surmise therefore that acti-vation of the STS by the SAT was due to explicit taskinstructions to judge the social interactions
(b) Role of the fusiform face area in socialcognition
We also found significant SAT activation of the rightFG When compared directly in a subset of participants totheir FFA gleaned from a separate scanning session witha perceptual discrimination task of person versus objectidentity we found that both ROIs were confined to theright side on the same two coronal slices There wasabout 50 overlap between the SAT FG and FFA acti-vations and the centre of the two activations differed byless than 1 voxel The FFA activation was offset to the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 423
lateral prominence of the FG whereas the SAT activateda region of the FG that was closer to the centre of the FGbut still largely in the lateral aspect Because the SAT doesnot contain any face representations the FFA engagementwas unexpected as this region is thought of as selectivefor faces (Kanwisher et al 1997) or to other classes ofcomplex objects for which one is perceptually expert(Gauthier et al 2000)
These results have important implications for the poss-ible role of the fusiform in social brain circuitry andautism Interestingly the magnitude of the FG activationpredicted SAT task accuracy no other ROI correlated sig-nificantly with task accuracy This would seem to argueagainst any interpretation that the FFA activation is incon-sequential to the SAT that it is activated simply becauseit is well connected to other areas that are directly involvedin the social attribution process Using a task that is simi-lar to the SAT Castelli et al (2000) also reported signifi-cant right FG activation using PET However in theirfollow-up PET study comparing autism and normal con-trols (Castelli et al 2002) they failed to find significantFG activation in either group The failure of their secondstudy to find right FG activations may be due to the morestringent random effects model used in the data analysesHowever because we find right FG activation with fMRIwhich is more sensitive than PET and because Castelli etal (2000) found it with what amounts to a lower thres-hold it seems quite probable that the effect is real In factwe have preliminary evidence from an ongoing fMRIstudy that reproduces the SAT FG activations in healthycontrols (Schultz et al 2001) In addition there may besome task attributes that differ between our studies andCastelli et alrsquos that might impact into the strength withwhich FG computational processes are evoked
The key question then is why is the FG engaged bythe SAT and what role does the right FG have in socialcognition and perception Currently there are three com-peting theories of the functional organization of the FGand related ventral visual perceptual areas One modelput forth by Haxby Chao Martin and colleagues speci-fies that objects are encoded in a distributed fashion acrossa wide expanse of the ventral temporal-occipital cortex(Haxby et al 2001) They call this the lsquoobject formtypologyrsquo model (Ishai et al 1999) Their data indicatethat object category specificity is achieved by unique spa-tial patterns of activation across this extrastriate visual cor-tex They show that the pattern of activations across thiscortex is diagnostic of object category membership moreso than any localized activation maxima (Haxby et al2001) They also argue that object category perceptioninvolves retrieval of category-related information aboutspecific features and attributes of the object category(Chao et al 2002) They admit that the lsquonature of theinformation about objects that is represented in the ventraltemporal cortex is a great puzzlersquo (Haxby et al 2000 p4) but open the possibility that semantic information maybe important
A second view championed by Kanwisher et al holdsthat several select perceptual categories including facesbodies and spatial layout of places are encoded in highlyspecific locations in a modular fashion (Kanwisher et al1997 Epstein amp Kanwisher 1998 Downing et al 2001Spiridon amp Kanwisher 2002) The FFA is thought to be
Phil Trans R Soc Lond B (2003)
most concerned with discriminating among individualidentities and not with discriminating images at the categ-orical level In fact lesions to the FFA cause a specificdeficit in recognizing individual identities but not in reco-gnizing the general category of face versus non-face(Wada amp Yamamoto 2001) Spiridon amp Kanwisher (2002)provide a partial replication of Haxby et alrsquos (2001) objectform typology model They show that patterns of ventralvisual cortical activation can distinguish object categoriesNevertheless their data show category specificity for selectareas such as the FFA and they argue the ventral visualcortex is not equipotential
Gauthier et al however have argued that the ventraloccipital temporal pathway is organized by the nature ofthe perceptual computations and that these processingbiases are acquired through experience (Gauthier et al2000 Tarr amp Gauthier 2000) For example the parahip-pocampal place area is a function of a bias towards pro-cessing landscapes in terms of their spatial layout becausewe have learned through repeated experiences that this isvery useful information to extract when perceiving land-scapes and related visual images Similarly we learn quiteearly in life that it is important to discriminate faces on anindividual level for example discriminating mother fromothers This bias towards individual identification accord-ing to Gauthier accounts for the FFArsquos apparent modu-larity (Gauthier et al 1997 2000 Tarr amp Gauthier 2000)According to this third model the type of informationneeded and our cumulative experience in processing thatinformation organizes the ventral occipitalndashtemporal path-way into regional centres with preferred modes of pro-cessing
All three models seem to agree that the functionalorganization of the ventral visual cortices is driven by theneed to categorize perceptions into object classes Webelieve that our findings showing FFA activation by non-face objects are consistent with aspects of both the pro-cessing map model of Gauthier and the object formtypology model of Haxby et al First consistent withHaxby et al we suggest that semantic information isimportant to the ventral visual pathway for object categor-ization Second consistent with Gauthier et al we believethat repeated perceptual experience with faces biases thetype of information that FG finds important We proposethat the middle FG area encodes semantic attributes ofpeople because of repeated perceptual experiences withfaces that occur during social interactions In fact wewould guess that most perceptual experiences with facesoccur during social situations and these social situationsoften involve repeated social judgements Thus infor-mation about the social nature of people might be storedin the FG (though not exclusively)
The nature of the semantic information stored in theFG might be restricted to anything that would be helpfulin defining faces as a distinct category of object becausemaking such distinctions appears to be the primary chargeof the ventral visual pathway This would include knowl-edge of people as having personal agency and of havingthe capability to disturb each otherrsquos emotional homeo-stasis (eg to act friendly or unfriendly) By pretendingthat the three shapes in the SAT are people stored knowl-edge about people in social interactions might beretrieved causing the observed activations in the FG
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
424 R T Schultz and others FFA and social cognition
There would be a measure of efficiency from thisarrangement that is having representations and compu-tations of more abstract attributes of people inter-digitatedwith front-end perceptual processes about physical attri-butes Gauthierrsquos claim is that the same group of neuronscan be engaged by perceptually different categories ofobjects it is the type of processing rather than the visualdetails that are important Extrapolating from this wewould argue that the SAT engages a region of the FGoverlapping with the FFA because it demands compu-tational processes to classify the SAT geometric figures aspeople or person-like Chao et al (1999) reach a similarconclusion They argued that activity in the ventral path-way reflects stored information about an object categorynot just physical features They point out that thisarrangement could explain why lesion patients with cate-gory specific perceptual deficits also have trouble retriev-ing general information about that visual category Thuswe would add that the FFA must store general infor-mation about people or some meta-representation oflsquopeoplenessrsquo
(c) Implications for the pathobiology of autismThe SAT used in this current study appears to be an
effective neurobehavioural probe for engaging a distrib-uted network of brain regions involved in different aspectsof social perception and cognition It will be important touse this and similar procedures in persons with autism tobetter define the nature of brain functions in this disorderCastelli et al (2002) have already taken the first step inthis process and they describe a pattern of hypoactivationin the MPFC STS and TPs in autism In an ongoingstudy of autism spectrum disorders we have presentedpreliminary data using the SAT showing that we too findhypoactivation of these regions and in the amygdala andFG (Schultz et al 2001) Thus we predict that futurework in this area will show that the entire social brain net-work is underactive in autism during tasks requiring socialperceptual and social cognitive processing
There are already sufficient data to argue strongly for arole of the FG in the pathobiology of autism No less thanfive previous fMRI studies have shown the FFA region tobe significantly less engaged among persons with autismcompared with controls during face perception tasks(Critchley et al 2000 Dierks et al 2001 Pierce et al2001 Schultz et al 2000a 2001) Activation of the FGby the social judgements in the current study adds animportant piece of evidence in favour of a causal role forthe FG in the pathobiology of autism This conclusionhowever must remain tentative until additional studiesmore precisely define the factors leading to FG activationduring social attribution and gather better data to proveits computational role in social cognition In addition theeffect of FG lesions for social functioning must be clari-fied If the FG is involved in social computations onewould expect to find social cognitive deficits in personswith lesions to this area However social deficits in proso-pagnosics have not been widely reported It might be thatprosopagnosic patients have not been carefully tested onthis dimension In this regard using our behavioural ver-sion of the SAT (Klin 2000) to test social perceptionamong prosopagnosics would be quite interesting Theparallels however between autism and prosopagnosia are
Phil Trans R Soc Lond B (2003)
incomplete because autism is clearly a developmental dis-order whereas the research literature on prosopagnosia ismostly confined to lesions acquired in adolescence oradulthood Among the few cases of developmental proso-pagnosia reported in the literature there is indeed one thathighlights severe social impairments (Kracke 1994) Itmay be that the role of the FG in the development ofautistic symptoms is different from the role of the FG inthe maintenance of social cognitive functions after brainmaturity
It is also possible that the role of the FG in social pro-cesses is dependent on its functional relationships withother nodes in the social brain and that it is the collectiveaction and interaction of the network that is of primaryimportance for social behaviours For example we founda strong correlation across participants in the amount ofFG and amygdala activation A strictly modular view ofthese areas may be inappropriate as the functions of eachnode could be quite dependent on one another and whenonly considered in isolation quite insufficient to supportsocial processes Thus whereas a lesion to the FG mayimpair visual perception it might not have a large impacton social functioning if this depends on an extended net-work that is dynamic and capable of compensatory adjust-ments Dysfunction of the FG may be necessary but notsufficient to produce social deficits Indeed our relianceon modular models of brain functioning may be leadingus astray in our search for causal mechanisms in autismInstead it may be the collective action of a distributedsystem that is critical to the pathobiology of autism TheFG region may be a key partner in this distributed systembut nevertheless just one node and insufficient by itselfto support in any substantive manner social cognitive pro-cesses or to explain social cognitive deficits seen inautism
5 CONCLUSION AND FUTURE DIRECTIONS
Using the SAT we isolated a distributed network ofactivations that conform to the emerging model of thesocial brain Most important we found significant acti-vation of the central aspect of the FG thus adding thisregion to the expanding list of structures involved in socialprocesses The FG region activated by the SAT overlapsin its spatial extent with the FFA with centres of massdiffering by less than 1 voxel We speculate that these SATactivations represent computational processes associatedwith more abstract attributes of people
It is an open question as to whether the substantial over-lap between the FFA and SAT FG activation is due to anoverlapping or shared set of neuronal assemblies This willneed to be clarified by future work Electrophysiologicalstudies show that there are small face-specific patches inthe FG cortex (Allison et al 1999) In the current studysmall patches of face cells and SAT cells could be inter-mingled but distinct and not drawing on any of the sameneuronal assemblies The spatial resolution of fMRI haslimitations that may preclude any definitive answer to thisquestion but there are strategies that can be used toaddress the issue For example the current study designdid not contain SAT and face discriminations in the sameexperimental fMRI series Doing so would enable directcomparisons of the computational demands placed on the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 425
common area of the FFA It also would be informative iffollow-up studies superimposed faces within the geometricfigures of the SAT without changing the film scripts inany way We could then determine if the computations ofthe common area of the FFA increase in a predictablefashionmdashwould the activation be a linear summation ofthe original SAT plus face discrimination Any significantdeviation from an additive model would suggest that thereis some sharing of neuronal assemblies with the SAT andface discrimination tasks when presented alone
This work was supported by grants from the National Institutesof Child Health and Human Development (grants PO1 HD03008 and PO1 HDDC35482) and the Korczak FoundationThe authors thank the anonymous reviewers for their helpfulcomments Michael Lee for programming the fMRI tasksHedy Serofin and Terry Hickey for their assistance in acquiringthe fMRI data John Herrington for his valuable assistance indata analyses of preliminary studies and members of the YaleDevelopmental Neuroimaging Program for their helpful com-ments on this manuscript A special acknowledgement ofappreciation is given to Donald Cohen who passed away inOctober of 2001 for his outstanding mentorship in all aspectsof this program of work
REFERENCES
Abell F Happe F amp Frith U 2000 Do triangles play tricksAttribution of mental states to animated shapes in normaland abnormal development J Cogn Dev 15 1ndash20
Adolphs R Tranel D amp Damasio A R 1998 The humanamygdala in social judgement Nature 393 470ndash474
Allison T Puce A Spencer D D amp McCarthy G 1999Electrophysiological studies of human face perception Ipotentials generated in occipitotemporal cortex by face andnon-face stimuli Cerebr Cortex 9 415ndash430
Allison T Puce A amp McCarthy G 2000 Social perceptionfrom visual cues role of the STS region Trends Cogn Sci4 267ndash278
Amaral D G amp Price J L 1984 Amygdalo-cortical projec-tions in the monkey (Macaca fascicularis) J Comp Neurol230 465ndash496
Amaral D G Price J L Pitkanen A amp Carmichael S T1992 Anatomical organization of the primate amygdaloidcomplex In The amygdala neurobiological aspects of emotionmemory and mental dysfunction (ed J Aggleton) pp 1ndash66New York Wiley-Liss
Anderson A K amp Phelps E A 2001 Lesions of the humanamygdala impair enhanced perception of emotionally salientevents Nature 411 305ndash309
Bachevalier J 1994 Medial temporal lobe structures andautism a review of clinical and experimental findingsNeuropsychologia 32 627ndash648
Bachevalier J amp Mishkin M 1986 Visual recognition impair-ment follows ventromedial but not dorsolateral prefrontallesions in monkeys Behav Brain Res 20 249ndash261
Baron-Cohen S Ring H A Wheelwright S BullmoreE T Brammer M J Simmons A amp Williams S C R1999 Social intelligence in the normal and autistic brain anfMRI study Eur J Neurosci 11 1891ndash1898
Baron-Cohen S Ring H A Bullmore E T WheelwrightS Ashwina C amp Williams S C R 2000 The amygdalatheory of autism Neurosci Biobehav Rev 24 355ndash364
Bechara A Tranel D Damasio H amp Damasio A R 1996Failure to respond autonomically to anticipated future out-comes following damage to prefrontal cortex Cerebr Cortex6 215ndash225
Benton A 1994 Face recognition Los Angeles CA WesternPsychological Services
Phil Trans R Soc Lond B (2003)
Bonda E Petrides M Ostry D amp Evans A 1996 Specificinvolvement of human parietal systems and the amygdala inthe perception of biological motion J Neurosci 16 3737ndash3744
Bowler D M amp Thommen E 2000 Attribution of mechan-ical and social causality to animated displays by childrenwith autism Autism 4 147ndash171
Brothers L 1990 The social brain a project for integratingprimate behavior and neurophysiology in a new domainConcepts Neurosci 1 27ndash151
Brothers L 1995 Neurophysiology of the perception of inten-tion by primates In The cognitive neurosciences 2nd edn (edM S Gazzaniga) pp 1107ndash1115 Cambridge MA MITPress
Butter C M McDonald J A amp Snyder D R 1969 Oralitypreference behavior and reinforcement value of nonfoodobject in monkeys with orbital frontal lesions Science 1641306ndash1307
Carmichael S T amp Price J L 1995 Limbic connections ofthe orbital and medial prefrontal cortex in macaque monk-eys J Comp Neurol 363 615ndash641
Castelli F Happe F Frith U amp Frith C 2000 Movementand mind a functional imaging study of perception andinterpretation of complex intentional movement patternsNeuroImage 12 314ndash325
Castelli F Frith C Happe F amp Frith U 2002 AutismAsperger syndrome and brain mechanisms for the attri-bution of mental states to animated shapes Brain 1251839ndash1849
Chao L L Haxby J V amp Martin A 1999 Attribute-basedneural substrates in posterior temporal cortex for perceivingand knowing about objects Nat Neurosci 2 913ndash919
Chao L L Weisberg J amp Martin A 2002 Experience-dependent modulation of category-related cortical activityCerebr Cortex 12 545ndash551
Critchley H D (and 10 others) 2000 The functional neuroan-atomy of social behaviour changes in cerebral blood flowwhen people with autistic disorder process facial expressionsBrain 123 2203ndash2212
Culham J C Cavanagh P amp Kanwisher N G 2001 Atten-tion response functions characterizing brain areas usingfMRI activation during parametric variations of attentionalload Neuron 32 737ndash745
Damasio A R Tranel D amp Damasio H 1990 Individualswith sociopathic behavior caused by frontal damage fail torespond autonomically to social stimuli Behav Brain Res41 81ndash94
Dierks T Bolte S Huble D Lanfermannm H amp PoustkaF 2001 Alterations of face processing strategies in autism(a fMRI study) Presented at the 6th Annual Meeting of theOrganization for Human Brain Mapping Brighton UK 10ndash14 June 2001
Dias R Robbins T W amp Roberts A C 1996 Dissociationin prefrontal cortex of affective and attentional shifts Nature380 69ndash72
Downing P E Jiang Y Shuman M amp Kanwisher N 2001A cortical area selective for visual processing of the humanbody Science 293 2470ndash2473
Epstein R amp Kanwisher N 1998 A cortical representation ofthe local visual environment Nature 392 598ndash601
Ernst M Zametkin A J Matochik J A Pascualvaca D ampCohen R M 1997 Reduced medial prefrontal dopami-nergic activity in autistic children Lancet 350 638
Fletcher P C Happe F Frith U Baker S C DolanR J Frackowiak R S amp Frith C D 1995 Other minds inthe brain a functional imaging study of lsquotheory of mindrsquo instory comprehension Cognition 57 109ndash128
Frith C D amp Frith U 1999 Interacting minds a biologicalbasis Science 286 1692ndash1695
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
426 R T Schultz and others FFA and social cognition
Fujita I Tanaka K Ito M amp Cheng K 1992 Columns forvisual features of objects in monkey inferotemporal cortexNature 360 343ndash346
Gaffan E A Gaffan D amp Harrison S 1988 Disconnectionof the amygdala from visual association cortex impairs vis-ual-reward association learning in monkeys J Neurosci 83144ndash3150
Gallagher H L Happe F Brunswick N Fletcher P CFrith U amp Frith C D 2000 Reading the mind in cartoonsand stories an fMRI study of lsquotheory of mindrsquo in verbal andnonverbal tasks Neuropsychologia 38 11ndash21
Gauthier I Anderson A W Tarr M J Skudlarski P ampGore J C 1997 Levels of categorization in visual recog-nition studied using functional magnetic resonance imagingCurr Biol 7 645ndash651
Gauthier I Tarr M J Anderson A W Skudlarski P ampGore J C 1999 Activation of the middle fusiform face areaincreases with expertise in recognizing novel objects NatureNeurosci 2 568ndash573
Gauthier I Skudlarski P Gore J C amp Anderson A W2000 Expertise for cars and birds recruits brain areasinvolved in face recognition Nature Neurosci 3 191ndash197
Goel V Grafman J Sadato N amp Hallett M 1995 Mode-ling other minds Neuroreport 6 1741ndash1746
Grelotti D Gauthier I amp Schultz R T 2001 Social interestand the development of cortical face specialization whatautism teaches us about face processing Devl Psychobiol 40213ndash225
Grezes J Fonlupt P Bertenthal B amp Delon-Martin C2001 Does perception of biological motion rely on specificbrain regions NeuroImage 13 775ndash785
Gusnard D A Akbudak E Shulman G L amp RaichleM E 2001 Medial prefrontal cortex and self-referentialmental activity relation to a default mode of brain functionProc Natl Acad Sci USA 98 4259ndash4264
Happe F Ehlers S Fletcher P Frith U Johansson MGillberg C Dolan R Frackowiak R amp Frith C 1996lsquoTheory of mindrsquo in the brain Evidence from a PET scanstudy of Asperger syndrome Neuroreport 8 197ndash201
Haxby J V Ungerleider L G Clark V P Schouten J LHoffman E A amp Martin A 1999 The effect of face inver-sion on activity in human neural systems for face and objectperception Neuron 22 189ndash199
Haxby J V Isahai A Chao L L Ungerleider L G ampMartin A 2000 Object-form topology in the ventral tem-poral lobe response to I Gauthier (2000) Trends Cogn Sci4 3ndash4
Haxby J V Gobbini M I Furey M L Ishai ASchouten J L amp Pietrini P 2001 Distributed and overlap-ping representations of faces and objects in ventral temporalcortex Science 293 2425ndash2430
Haznedar M M Buchsbaum M S Metzger M Solim-ando A Spiegel-Cohen J amp Hollander E 1997 Anteriorcingulate gyrus volume and glucose metabolism in autisticdisorder Am J Psychiat 154 1047ndash1050
Heider F amp Simmel M 1944 An experimental study ofapparent behavior Am J Psychol 57 243ndash259
Hobson R P Ouston J amp Lee A 1988a Whatrsquos in a faceThe case of autism Br J Psychol 79 441ndash453
Hobson R P Ouston J amp Lee A 1988b Emotion recog-nition in autism coordinating faces and voices Psychol Med18 911ndash923
Hoffman E A amp Haxby J V 2000 Distinct representationsof eye gaze and identity in the distributed human neural sys-tem for face perception Nature Neurosci 3 80ndash84
Hornak J Rolls E T amp Wade D 1996 Face and voiceexpression identification in patients with emotional andbehavioural changes following ventral frontal lobe damageNeuropsychologia 34 247ndash261
Phil Trans R Soc Lond B (2003)
Ishai A Ungerleider L G Martin A Schouten J L ampHaxby J V 1999 Distributed representation of objects inthe human ventral visual pathway Proc Natl Acad Sci USA96 9379ndash9384
Kanwisher N 2000 Domain specificity in face perceptionNature Neurosci 3 759ndash763
Kanwisher N McDermott J amp Chun M M 1997 The fusi-form face area a module of extrastriate cortex specializedfor face perception J Neurosci 17 4302ndash4311
Klin A 2000 Attributing social meaning to ambiguous visualstimuli in higher functioning autism and Asperger syndromethe social attribution task J Child Psychol Psychiat 41831ndash846
Klin A Sparrow S S de Bildt A Cicchetti D V CohenD J amp Volkmar F R 1999 A normed study of face recog-nition in autism and related disorders J Autism Devl Dis-orders 29 499ndash508
Klin A Jones W Schultz R T Volkmar F amp Cohen D J2002 Visual fixation patterns during viewing of naturalisticsocial situations as predictors of social competence in indi-viduals with autism Arch Gen Psychiat 59 809ndash816
Kracke I 1994 Developmental prosopagnosia in Aspergersyndrome presentation and discussion of an individual caseDevl Med Child Neurol 36 873ndash886
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J 1997 Neuroanatomical correlates of happi-ness sadness and disgust Am J Psychiat 154 926ndash933
Langdell T 1978 Recognition of faces an approach for thestudy of autism J Child Psychol Psychiat 19 255ndash268
LeDoux J E 1996 The emotional brain New York Simon ampShuster
Lowel S amp Singer W 1992 Selection of intrinsic horizontalconnections in the visual cortex by correlated neuronalactivity Science 255 209ndash212
Macdonald H Rutter M Howlin P Rios P Le ConteurA Evered C amp Folstein S 1989 Recognition andexpression of emotional cues by autistic and normal adultsJ Child Psychol Psychiat 30 865ndash877
Morris J S Ohman A amp Dolan R J 1998 Conscious andunconscious emotional learning in the human amygdalaNature 393 467ndash470
Myers R E Swett C amp Miller M 1973 Loss of social groupaffinity following prefrontal lesions in free-rangingmacaques Brain Res 64 257ndash269
Osterling J amp Dawson G 1994 Early recognition of childrenwith autism a study of first birthday home video tapes JAutism Devl Disorders 24 247ndash257
Pierce K Muller R A Ambrose J Allen G amp Cour-chesne E 2001 Face processing occurs outside the fusiformlsquoface arearsquo in autism evidence from functional MRI Brain124 2059ndash2073
Price J L Carmichael S T amp Drevets W C 1996 Net-works related to the orbital and medial prefrontal cortex asubstrate for emotional behavior Prog Brain Res 107523ndash536
Puce A Allison T Gore J C amp McCarthy G 1995 Face-sensitive regions in human extrastriate cortex studied byfunctional MRI J Neurophysiol 74 1192ndash1199
Reiman E M Lane R D Ahern G L Schwartz G EDavidson R J Friston K J Yun L S amp Chen K 1997Neuroanatomical correlates of externally and internally gen-erated human emotion Am J Psychiat 154 918ndash925
Rolls E T 1995 A theory of emotion and consciousness andits application to understanding the neural basis of emotionIn The cognitive neurosciences (ed M S Gazzaniga) pp1091ndash1106 Cambridge MA MIT Press
Schultz R T Gauthier I Klin A Fulbright R K Ander-son A W Volkmar F Skudlarski P Lacadie C CohenD J amp Gore J C 2000a Abnormal ventral temporal
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from

FFA and social cognition R T Schultz and others 419
(d) Data analysisData were corrected for motion using SPM99 for three trans-
lation directions and for the three possible rotations (WellcomeDepartment of Cognitive Neurology London UK) Image vol-umes with centre of mass (x y or z) movement of more than15 mm within a run were discarded Image analyses and testsof statistical significance were done using locally developedsoftware (Skudlarski httpmrimedyaleedumembersFframedhtml) Motion corrected images were spatially smoothed with aGaussian filter with a full-width half-maximum value of625 mm The specific effects of each task were evaluated bycreating t-maps for each imaging series incorporating a correc-tion for linear drift (Skudlarski et al 1999) of specific task con-trasts social versus bumper car in Study 1 and face versusobject in Study 2 t-maps were averaged across imaging seriesand co-registered with the higher resolution anatomical imagesfor display and localization These maps were then transformedby in-plane registration and slice interpolation into a pro-portional three-dimensional grid defined by Talairach amp Tour-noux (1988) and averaged across all subjects to createcomposite t-maps with the acquired data in 14 slices interp-olated to 18 slices (NB 16 slices are shown in figure 1 as fMRIactivations on the first and last slice are corrupted by motioncorrection) The SAT versus bumper car maps are displayed inthe figures using a significance level of p 00005(uncorrected) Face versus object t-maps were created and dis-played at p 005 (uncorrected) with the a priori hypothesis thatthe right lateral FG would define the FFA No other brain areasare examined in the second study thus avoiding any multiplecomparison problem
ROI analyses were conducted in the SAT study by tracingsignificant pixels on the group composite activation map (figure1) in the following regions the right FG the right and left STSand STG the right TP the right amygdala and right and leftdorsal MPFC To more thoroughly assess activity in the FGmedial and lateral FG ROIs (and the combined whole FG) weredefined anatomically and traced across the two coronal sliceswhere there was significant SAT activation The activated SATROI for the FG is 33 of the size of the entire anatomicallydefined FG at those two slices Individual subject data wereinterrogated using the ROIs to obtain the mean per cent signalchange for each person for each region and Talairach centre ofmass coordinates (the centre of ROI activation weighted by theamplitude of activation across the region) Mean per cent signal-change data were used in correlational analyses to estimate theconsistency of conjoint activity between ROIs across subjects
3 RESULTS
(a) Behavioural performanceThere were no significant differences between the
social and bumper car tasks in performance accuracy(t1 2 2 = 163 p 010 social = 86 plusmn 16 correctbumper = 74 plusmn 16 correct) or reaction time (t1 22
= 062 p 060) There were no significant differences intask accuracy between males and females or left-handersand right-handers In addition there were no significantcorrelations between task performance and age Full ScaleIntelligence Quotient or Benton Face Recognition per-formance
Phil Trans R Soc Lond B (2003)
(b) Brain activity associated with the socialattribution task
As shown in figure 1 comparison of the SAT with thebumper car control condition resulted in a widely distrib-uted set of significant activations There was very little sig-nificant activation for the bumper car task with the oneregion shown clearly in figure 1 being bilateral activationof the dorsal bank of the intra-parietal sulcus The SATnetwork included a region within the right and left dorsalMPFC the right and left inferior frontal gyrus parsorbitalis and the lateral orbital gyrus the right TP theright amygdala the right and left STS and STG and theright FG It is important to note that at lower thresholds(eg p 001) there was also left amygdala activation anda ROI analysis of the per cent signal change failed to findsignificantly more right than left amygdala activation It isalso worth noting that the FG activations seem quitespecific to the SAT task in the sense that reducing thethreshold down to p 005 failed to show additional ven-tral pathway activation More widespread activationsmight have indicated a general SAT effect on arousal orattention that was manifested throughout the ventralstream but this was not the case
The largest areas of activation were the STG (especiallyon the right) and MPFC Direct comparison of the rightversus left MPFC mean per cent signal change failed tofind significant differences (paired t1 1 = 045 p 050)However the right STG was significantly more activatedthan the left STG (paired t1 1 = 264 p = 002) Table 1presents the Talairach coordinates for the centre of acti-vation mass for each ROI Table 2 presents a correlationalmatrix showing the consistency of conjoint activitybetween regions The strongest correlation is between theright amygdala and the ROI that defines the significantlyactivated region of the right FG (r = 071 p = 001) Inter-estingly this correlation is nominally larger than that ofthe entire FG and the smaller subcomponent FG ROIdefined by the SAT activated pixels (r = 069 p = 0013)Since some correlation would be expected between theseoverlapping ROIs especially since the data were spatiallysmoothed the robust correlation to the amygdala is evenmore impressive Other notable results from the corre-lation matrix include the lack of correlation between theMPFC and either the amygdala or temporal lobe ROIsWithin the temporal lobes however the right STG is sig-nificantly correlated with the left STG and with both defi-nitions of the right FG
Accuracy of performance on the SAT correlates withthe amount of activity in the anatomically defined rightFG (r = 065 p = 002) but not with any other node in theSAT network Females showed significantly more rightSTG activation than males (t1 0 = 234 p = 004) Maleshowever showed significantly more right TP activation(t1 0 = 253 p = 003) There were no other significant sexdifferences and no significant associations with hand-edness or age
(c) Comparison of activity in the right fusiformgyrus during social attribution task and face
perceptionAt the time Study 1 data were collected finding signifi-
cant right FG activation to the SAT was unexpected Toclarify whether the FG activation was in the precise
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
420 R T Schultz and others FFA and social cognition
Table 1 ROI centres of mass coordinates
mean Talairach coordinates (X Y Z )
ROI right hemisphere left hemisphere Brodmann areas
MPFC 49 349 436 274 361 434 6 8 9TP 463 138 2128 mdash 38amygdala 228 2109 2124 mdash mdashFG (SAT) 344 2468 290 mdash 37FG (face) 374 2480 2126 mdash 37STG 507 2571 152 2565 2608 194 22 39
Table 2 ROI correlation matrix(Correlations are based on mean per cent signal change from each ROI (see sect 2 for ROI procedures) SAT accuracy data representthe percentage of films that each participant correctly)
activated whole right right left left right rightright FG FG amygdala amygdala MPFC MPFC left STG STG right TP
whole right FG 069 curren curren curren
right amygdala 071 curren curren curren 032left amygdala 056 curren 019 070 curren curren curren
left MPFC 021 013 032 011right MPFC 2018 2008 021 019 048left STG 021 029 019 005 056 curren 018right STG 054 curren 069 curren curren curren 022 025 009 2016 060 curren curren
right TP 2007 2011 040 033 048 034 038 2010SAT accuracy 045 065 curren curren 2012 009 013 2017 201 022 2034
curren curren curren p lt 001 (r gt 069) curren curren p lt 005 (r gt 057) curren p lt 010
(a) SAT versus bumper car FFA activation
(
(c) enlargement and alignment of FFA
Figure 2 (a) Composite (n = 12) t-map at two slices showing significant (p 00005) activation for the SAT contrast(yellowred) with the bumper car control task (bluepurple) (b) Composite (n = 9) t-map at two slices showing significant(p 005) activation for the face (yellowred) versus object discrimination (bluepurple) This contrast defines the FFA (c)Subregions of composite t-maps shown in (a) and (b) are enlarged and aligned to demonstrate the overlap of activation in theFG for the SAT and face discrimination activations Subscripts 1 and 2 refer to the first (more anterior) and second coronalslices with significant activation
location of the FFA we compared it with the result of theface versus object discrimination contrast (the standardmeans of identifying the FFA in the literature) Both theFFA and the SAT activations of the FG were confined totwo coronal slices in highly overlapping locations Thesetwo sets of group composite maps are shown in figure 2
Phil Trans R Soc Lond B (2003)
The FG activations were stronger in the SAT versusbumper contrast than the face versus object contrast butthis may have had as much do with the baseline as theexperimental tasks The Talairach coordinates for theSAT and FFA activations show that their centre of massdiffers by less than one voxel The SAT activation is 3 mm
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 421
Figure 3 Scans of two individuals across three different occasions showing the reproducibility of FFA activations at two time-points and relationship to SAT activations (andashc) are from a 23-year-old male (dndashf ) are from a 24-year-old female Panels arearranged chronologically (ad) The first face versus object experiment (be) The second face versus object scan (cf ) SATversus bumper car contrast Both coronal slices are shown where there was FFA (t gt 15 in yellowred) or SAT activation(t gt 30 in yellowred) Arrows point to FG activity (right and left are reversed by convention) As in the group results (figure2) the SAT activation is centred slightly more medially along the MFS Left FG activation shown in these two subjects doesnot survive thresholding in the group composite (figure 2) Control tasks (object discrimination bumper car) are shown inpurpleblue
more medial than that of the FFA straddling the MFSthat delineates the lateral and medial aspects of the FGThe FFA on the other hand is clearly positioned in thelateral FG as expected (Haxby et al 1999) The SATactivation is also centred 36 mm more superior and about1 mm more anteriorly than the FFA A count of the over-lapping significant pixels showed that 50 of the SATactivation falls within the FFA This provides a goodapproximation of how these regions overlap Howeverthere is no definitive way to measure the percent overlapin this study because it would change with the use of dif-ferent control tasks in either condition or different signifi-cance levels for thresholding the t-maps
Two of the participants (one male one female) werealso part of a reproducibility study of the FFA and hadboth undergone the face discrimination protocol on twooccasions Figure 3 presents t-maps of their FG for eachface perception study and the SAT The female partici-pant shows reversed asymmetry as sometimes happenswith the left FG showing greater face activation than theright Nevertheless her SAT activations track her FFAand are more left-sided than typical These results showthat the FFA activation is reproducible so that the lessthan perfect overlap between the FFA and the SAT FGactivation is probably not a measurement or reliabilityissue
4 DISCUSSION
(a) The social brain networkThe current study required participants to observe the
movements of geometric figures and to interpret thesewith regard to a conceptual template about what consti-tutes a friendly or unfriendly social interaction It requiredclose attention to the contingent nature of a sequence ofmovements and inferences about mental states of eachcharacter to explain their actions Perception of the move-
Phil Trans R Soc Lond B (2003)
ments of these simple shapes as wilful seems to be auto-matic and effortless for healthy controls but not forpersons with autism (Klin 2000) As shown in figure 1the network engaged by the social attribution process (incontrast to the physical attribution control task) includednearly all of the brain areas implicated by past researchon the social brain (Brothers 1990) including cognitiveaspects such as theory of mind (Castelli et al 2000) aswell as perceptual aspects such as the perception of socialdisplays and biological movement (Allison et al 2000)We found significant activation of the bilateral MPFCsuperior STG and STS and inferior FG pars orbitalisextending into the lateral orbital gyri In addition therewere significant activations on the right side only for theamygdala TP and the FG The predominance of right-side activations is consistent with the notion that the righthemisphere is more concerned than the left with socialprocesses (Siegal et al 1996 Winner et al 2002) Ourresults differ from some past research by finding signifi-cant right FG activation and by failing to find orbito-MPFC activation
Several earlier neuroimaging studies have shown thatthe dorsal MPFC (ie that cortex anterior and superior tothe anterior cingulate gyrus) is a critical substrate for socialjudgements including empathizing and thinking aboutotherrsquos thoughts and intentions (Fletcher et al 1995 Goelet al 1995 Happe et al 1996 Castelli et al 2000 2002Gallagher et al 2000) Our findings are consistent withthese imaging studies and with non-human primate stud-ies that have documented social failures and loss of socialposition within the group following lesions to orbital pre-frontal cortices and MPFC (Butter et al 1969 Myers etal 1973 Bachevalier amp Mishkin 1986) The importantrole for the MPFC in social cognition is further suggestedby studies of autism spectrum disorders that find func-tional abnormalities in this area (Happe et al 1996 Ernstet al 1997 Haznedar et al 1997 Castelli et al 2002)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
422 R T Schultz and others FFA and social cognition
Gusnard et al (2001) suggest that the dorsal MPFC isinvolved in any kind of thought that uses the self as a refer-ent Thus the SAT activations in this area may have beendriven by theorizing about othersrsquo minds but with explicitreference to the participantrsquos own frame of reference as tohow they would feel in a similar situation
The orbital prefrontal cortex and to a lesser extent thedorsal MPFC have dense reciprocal connections withmedial temporal areas (Price et al 1996 Carmichael ampPrice 1995) providing the anatomical bases for a systemthat regulates emotional processes Damasio and col-leagues (Damasio et al 1990 Bechara et al 1996) haveargued that the orbito-MPFC have a primary function ofintegrating information about rewards and punishmentsto bias future behaviour (Rolls 1995 Dias et al 1996Hornak et al 1996 Lane et al 1997 Reiman et al 1997)A functional circuitry such as this would seem especiallyimportant in the development and acquisition of socialbehaviour However even acquired lesions to theseregions in adulthood can result in abnormalities of socialconduct (Damasio et al 1990) Brothers (1990) high-lighted the orbital prefrontal cortex as one of the threeprincipal brain regions involved in social cognition Weobserved poor signal in this region (these areas are notori-ously prone to fMRI signal drop-out and distortion) andthus we cannot know if the orbital prefrontal region wasengaged by the SAT or not However using a similarpsychological task but with PET where signal acquisitionin this region is not degraded Castelli et al (2000 2002)failed to find activation of the orbital prefrontal cortexCollectively these results call into question the impor-tance of the orbital prefrontal cortex in social cognitionand instead shift the focus on anterior cortices toward thedorsal MPFC
The SAT also generated robust activations of the rightamygdala and nearby cortex of the right TP The amyg-dala is often given a central role in theories of social per-ception and cognition (Brothers 1990 Bachevalier 1994Adolphs et al 1998 Baron-Cohen et al 2000 Schultz etal 2000b) The amygdala has a critical role in emotionalarousal assigning significance to environmental stimuliand mediating the formation of visual-reward associationsthat is lsquoemotionalrsquo learning (Gaffan et al 1988 LeDoux1996 Anderson amp Phelps 2001) It is reliably engagedduring judgements of personality characteristics from pic-tures of the face or part of the face (Adolphs et al 1998Baron-Cohen et al 1999 Winston et al 2002) Activationof the amygdala appears to be automatic and stimulusdriven as it can be engaged by images of facial expressionsin conscious awareness as well as by subliminal presen-tations of faces displaying affect (Morris et al 1998Whalen et al 1998 Critchley et al 2000) Thus theamygdalarsquos engagement by the SAT could stem from thegeneral emotional arousal evoked by the animations or itcould represent its computational role in some more spe-cific social perceptual process
The amygdala has dense reciprocal connections withthe ventral visual processing stream (Amaral amp Price1984) The strong correlation observed in this studybetween the right amygdala and the right FG could indi-cate that emotional inputs from the amygdala to the FGare necessary for engaging the social computational pro-cesses of the FG Brothers (1995) has speculated that the
Phil Trans R Soc Lond B (2003)
amygdala generates lsquosocial feelingsrsquo that are of importancein cutting through the complexities of social situations andguiding behaviour by simpler learned contingenciesSocial events are complex because their meaning oftencomes from specific combinations of features that do notadd up in a linear manner making it more difficult todecompose the events by cognitive analysis Effectivesocial engagement requires integration of context historicalrelationships and current socialndashemotional communi-cations expressed through prosodic tone as well as facialexpressions posture and gesture They are made morecomplex by the rapid pace of social transactions Thiswould necessitate some fast response system based ongeneral principles from past socialndashemotional experiencesa role for which the amygdala would seem ideally suitedIn other words the amygdala might drive intuition or lsquogutfeelingsrsquo to guide rapid non-verbal social interactionsinvolving facial expressions gesture etc Thus the strongamygdalandashFG correlation observed here could be inter-preted as the amygdala informing the FG of the relevanceof a social event and also of the outcome of its quick anddirty social perceptual analyses thereby guiding the FG(and other social nodes) in their finer grained compu-tations
Perhaps the largest and strongest activations to the SATwere those of the posterior aspects of the STS spreadinginto the adjacent STG This area has been implicated asa specific site for perception of biological motion (Bondaet al 1996 Allison et al 2000 Grezes et al 2001 Vainaet al 2001) Biological motion is a broad construct thatseems to encompass the perception of static images ofevents that could move or did move such as facialexpressions as well as actual movement by animateobjects This region of the STSndashSTG is also critical forthe decoding visual displays of social action or intention(eg gaze direction gesture and facial displays of emotion)(Critchley et al 2000 Hoffman amp Haxby 2000) In thisregard the SAT activations here are completely expectedand a testament to the effectiveness of these animationsin inducing the desired illusion of anthropomorphicaction The majority of the STSndashSTG activations wereanterior to location of the V5MT as defined in otherrecent fMRI studies (Culham et al 2001) but they alsoextended posteriorly into these more general movementsensitive cortices Unlike the amygdala some evidencesuggests that activity of the STS is mediated by explicitattention to social characteristics of the face and that STSengagement it is not automatic or stimulus driven(Winston et al 2002) We surmise therefore that acti-vation of the STS by the SAT was due to explicit taskinstructions to judge the social interactions
(b) Role of the fusiform face area in socialcognition
We also found significant SAT activation of the rightFG When compared directly in a subset of participants totheir FFA gleaned from a separate scanning session witha perceptual discrimination task of person versus objectidentity we found that both ROIs were confined to theright side on the same two coronal slices There wasabout 50 overlap between the SAT FG and FFA acti-vations and the centre of the two activations differed byless than 1 voxel The FFA activation was offset to the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 423
lateral prominence of the FG whereas the SAT activateda region of the FG that was closer to the centre of the FGbut still largely in the lateral aspect Because the SAT doesnot contain any face representations the FFA engagementwas unexpected as this region is thought of as selectivefor faces (Kanwisher et al 1997) or to other classes ofcomplex objects for which one is perceptually expert(Gauthier et al 2000)
These results have important implications for the poss-ible role of the fusiform in social brain circuitry andautism Interestingly the magnitude of the FG activationpredicted SAT task accuracy no other ROI correlated sig-nificantly with task accuracy This would seem to argueagainst any interpretation that the FFA activation is incon-sequential to the SAT that it is activated simply becauseit is well connected to other areas that are directly involvedin the social attribution process Using a task that is simi-lar to the SAT Castelli et al (2000) also reported signifi-cant right FG activation using PET However in theirfollow-up PET study comparing autism and normal con-trols (Castelli et al 2002) they failed to find significantFG activation in either group The failure of their secondstudy to find right FG activations may be due to the morestringent random effects model used in the data analysesHowever because we find right FG activation with fMRIwhich is more sensitive than PET and because Castelli etal (2000) found it with what amounts to a lower thres-hold it seems quite probable that the effect is real In factwe have preliminary evidence from an ongoing fMRIstudy that reproduces the SAT FG activations in healthycontrols (Schultz et al 2001) In addition there may besome task attributes that differ between our studies andCastelli et alrsquos that might impact into the strength withwhich FG computational processes are evoked
The key question then is why is the FG engaged bythe SAT and what role does the right FG have in socialcognition and perception Currently there are three com-peting theories of the functional organization of the FGand related ventral visual perceptual areas One modelput forth by Haxby Chao Martin and colleagues speci-fies that objects are encoded in a distributed fashion acrossa wide expanse of the ventral temporal-occipital cortex(Haxby et al 2001) They call this the lsquoobject formtypologyrsquo model (Ishai et al 1999) Their data indicatethat object category specificity is achieved by unique spa-tial patterns of activation across this extrastriate visual cor-tex They show that the pattern of activations across thiscortex is diagnostic of object category membership moreso than any localized activation maxima (Haxby et al2001) They also argue that object category perceptioninvolves retrieval of category-related information aboutspecific features and attributes of the object category(Chao et al 2002) They admit that the lsquonature of theinformation about objects that is represented in the ventraltemporal cortex is a great puzzlersquo (Haxby et al 2000 p4) but open the possibility that semantic information maybe important
A second view championed by Kanwisher et al holdsthat several select perceptual categories including facesbodies and spatial layout of places are encoded in highlyspecific locations in a modular fashion (Kanwisher et al1997 Epstein amp Kanwisher 1998 Downing et al 2001Spiridon amp Kanwisher 2002) The FFA is thought to be
Phil Trans R Soc Lond B (2003)
most concerned with discriminating among individualidentities and not with discriminating images at the categ-orical level In fact lesions to the FFA cause a specificdeficit in recognizing individual identities but not in reco-gnizing the general category of face versus non-face(Wada amp Yamamoto 2001) Spiridon amp Kanwisher (2002)provide a partial replication of Haxby et alrsquos (2001) objectform typology model They show that patterns of ventralvisual cortical activation can distinguish object categoriesNevertheless their data show category specificity for selectareas such as the FFA and they argue the ventral visualcortex is not equipotential
Gauthier et al however have argued that the ventraloccipital temporal pathway is organized by the nature ofthe perceptual computations and that these processingbiases are acquired through experience (Gauthier et al2000 Tarr amp Gauthier 2000) For example the parahip-pocampal place area is a function of a bias towards pro-cessing landscapes in terms of their spatial layout becausewe have learned through repeated experiences that this isvery useful information to extract when perceiving land-scapes and related visual images Similarly we learn quiteearly in life that it is important to discriminate faces on anindividual level for example discriminating mother fromothers This bias towards individual identification accord-ing to Gauthier accounts for the FFArsquos apparent modu-larity (Gauthier et al 1997 2000 Tarr amp Gauthier 2000)According to this third model the type of informationneeded and our cumulative experience in processing thatinformation organizes the ventral occipitalndashtemporal path-way into regional centres with preferred modes of pro-cessing
All three models seem to agree that the functionalorganization of the ventral visual cortices is driven by theneed to categorize perceptions into object classes Webelieve that our findings showing FFA activation by non-face objects are consistent with aspects of both the pro-cessing map model of Gauthier and the object formtypology model of Haxby et al First consistent withHaxby et al we suggest that semantic information isimportant to the ventral visual pathway for object categor-ization Second consistent with Gauthier et al we believethat repeated perceptual experience with faces biases thetype of information that FG finds important We proposethat the middle FG area encodes semantic attributes ofpeople because of repeated perceptual experiences withfaces that occur during social interactions In fact wewould guess that most perceptual experiences with facesoccur during social situations and these social situationsoften involve repeated social judgements Thus infor-mation about the social nature of people might be storedin the FG (though not exclusively)
The nature of the semantic information stored in theFG might be restricted to anything that would be helpfulin defining faces as a distinct category of object becausemaking such distinctions appears to be the primary chargeof the ventral visual pathway This would include knowl-edge of people as having personal agency and of havingthe capability to disturb each otherrsquos emotional homeo-stasis (eg to act friendly or unfriendly) By pretendingthat the three shapes in the SAT are people stored knowl-edge about people in social interactions might beretrieved causing the observed activations in the FG
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
424 R T Schultz and others FFA and social cognition
There would be a measure of efficiency from thisarrangement that is having representations and compu-tations of more abstract attributes of people inter-digitatedwith front-end perceptual processes about physical attri-butes Gauthierrsquos claim is that the same group of neuronscan be engaged by perceptually different categories ofobjects it is the type of processing rather than the visualdetails that are important Extrapolating from this wewould argue that the SAT engages a region of the FGoverlapping with the FFA because it demands compu-tational processes to classify the SAT geometric figures aspeople or person-like Chao et al (1999) reach a similarconclusion They argued that activity in the ventral path-way reflects stored information about an object categorynot just physical features They point out that thisarrangement could explain why lesion patients with cate-gory specific perceptual deficits also have trouble retriev-ing general information about that visual category Thuswe would add that the FFA must store general infor-mation about people or some meta-representation oflsquopeoplenessrsquo
(c) Implications for the pathobiology of autismThe SAT used in this current study appears to be an
effective neurobehavioural probe for engaging a distrib-uted network of brain regions involved in different aspectsof social perception and cognition It will be important touse this and similar procedures in persons with autism tobetter define the nature of brain functions in this disorderCastelli et al (2002) have already taken the first step inthis process and they describe a pattern of hypoactivationin the MPFC STS and TPs in autism In an ongoingstudy of autism spectrum disorders we have presentedpreliminary data using the SAT showing that we too findhypoactivation of these regions and in the amygdala andFG (Schultz et al 2001) Thus we predict that futurework in this area will show that the entire social brain net-work is underactive in autism during tasks requiring socialperceptual and social cognitive processing
There are already sufficient data to argue strongly for arole of the FG in the pathobiology of autism No less thanfive previous fMRI studies have shown the FFA region tobe significantly less engaged among persons with autismcompared with controls during face perception tasks(Critchley et al 2000 Dierks et al 2001 Pierce et al2001 Schultz et al 2000a 2001) Activation of the FGby the social judgements in the current study adds animportant piece of evidence in favour of a causal role forthe FG in the pathobiology of autism This conclusionhowever must remain tentative until additional studiesmore precisely define the factors leading to FG activationduring social attribution and gather better data to proveits computational role in social cognition In addition theeffect of FG lesions for social functioning must be clari-fied If the FG is involved in social computations onewould expect to find social cognitive deficits in personswith lesions to this area However social deficits in proso-pagnosics have not been widely reported It might be thatprosopagnosic patients have not been carefully tested onthis dimension In this regard using our behavioural ver-sion of the SAT (Klin 2000) to test social perceptionamong prosopagnosics would be quite interesting Theparallels however between autism and prosopagnosia are
Phil Trans R Soc Lond B (2003)
incomplete because autism is clearly a developmental dis-order whereas the research literature on prosopagnosia ismostly confined to lesions acquired in adolescence oradulthood Among the few cases of developmental proso-pagnosia reported in the literature there is indeed one thathighlights severe social impairments (Kracke 1994) Itmay be that the role of the FG in the development ofautistic symptoms is different from the role of the FG inthe maintenance of social cognitive functions after brainmaturity
It is also possible that the role of the FG in social pro-cesses is dependent on its functional relationships withother nodes in the social brain and that it is the collectiveaction and interaction of the network that is of primaryimportance for social behaviours For example we founda strong correlation across participants in the amount ofFG and amygdala activation A strictly modular view ofthese areas may be inappropriate as the functions of eachnode could be quite dependent on one another and whenonly considered in isolation quite insufficient to supportsocial processes Thus whereas a lesion to the FG mayimpair visual perception it might not have a large impacton social functioning if this depends on an extended net-work that is dynamic and capable of compensatory adjust-ments Dysfunction of the FG may be necessary but notsufficient to produce social deficits Indeed our relianceon modular models of brain functioning may be leadingus astray in our search for causal mechanisms in autismInstead it may be the collective action of a distributedsystem that is critical to the pathobiology of autism TheFG region may be a key partner in this distributed systembut nevertheless just one node and insufficient by itselfto support in any substantive manner social cognitive pro-cesses or to explain social cognitive deficits seen inautism
5 CONCLUSION AND FUTURE DIRECTIONS
Using the SAT we isolated a distributed network ofactivations that conform to the emerging model of thesocial brain Most important we found significant acti-vation of the central aspect of the FG thus adding thisregion to the expanding list of structures involved in socialprocesses The FG region activated by the SAT overlapsin its spatial extent with the FFA with centres of massdiffering by less than 1 voxel We speculate that these SATactivations represent computational processes associatedwith more abstract attributes of people
It is an open question as to whether the substantial over-lap between the FFA and SAT FG activation is due to anoverlapping or shared set of neuronal assemblies This willneed to be clarified by future work Electrophysiologicalstudies show that there are small face-specific patches inthe FG cortex (Allison et al 1999) In the current studysmall patches of face cells and SAT cells could be inter-mingled but distinct and not drawing on any of the sameneuronal assemblies The spatial resolution of fMRI haslimitations that may preclude any definitive answer to thisquestion but there are strategies that can be used toaddress the issue For example the current study designdid not contain SAT and face discriminations in the sameexperimental fMRI series Doing so would enable directcomparisons of the computational demands placed on the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 425
common area of the FFA It also would be informative iffollow-up studies superimposed faces within the geometricfigures of the SAT without changing the film scripts inany way We could then determine if the computations ofthe common area of the FFA increase in a predictablefashionmdashwould the activation be a linear summation ofthe original SAT plus face discrimination Any significantdeviation from an additive model would suggest that thereis some sharing of neuronal assemblies with the SAT andface discrimination tasks when presented alone
This work was supported by grants from the National Institutesof Child Health and Human Development (grants PO1 HD03008 and PO1 HDDC35482) and the Korczak FoundationThe authors thank the anonymous reviewers for their helpfulcomments Michael Lee for programming the fMRI tasksHedy Serofin and Terry Hickey for their assistance in acquiringthe fMRI data John Herrington for his valuable assistance indata analyses of preliminary studies and members of the YaleDevelopmental Neuroimaging Program for their helpful com-ments on this manuscript A special acknowledgement ofappreciation is given to Donald Cohen who passed away inOctober of 2001 for his outstanding mentorship in all aspectsof this program of work
REFERENCES
Abell F Happe F amp Frith U 2000 Do triangles play tricksAttribution of mental states to animated shapes in normaland abnormal development J Cogn Dev 15 1ndash20
Adolphs R Tranel D amp Damasio A R 1998 The humanamygdala in social judgement Nature 393 470ndash474
Allison T Puce A Spencer D D amp McCarthy G 1999Electrophysiological studies of human face perception Ipotentials generated in occipitotemporal cortex by face andnon-face stimuli Cerebr Cortex 9 415ndash430
Allison T Puce A amp McCarthy G 2000 Social perceptionfrom visual cues role of the STS region Trends Cogn Sci4 267ndash278
Amaral D G amp Price J L 1984 Amygdalo-cortical projec-tions in the monkey (Macaca fascicularis) J Comp Neurol230 465ndash496
Amaral D G Price J L Pitkanen A amp Carmichael S T1992 Anatomical organization of the primate amygdaloidcomplex In The amygdala neurobiological aspects of emotionmemory and mental dysfunction (ed J Aggleton) pp 1ndash66New York Wiley-Liss
Anderson A K amp Phelps E A 2001 Lesions of the humanamygdala impair enhanced perception of emotionally salientevents Nature 411 305ndash309
Bachevalier J 1994 Medial temporal lobe structures andautism a review of clinical and experimental findingsNeuropsychologia 32 627ndash648
Bachevalier J amp Mishkin M 1986 Visual recognition impair-ment follows ventromedial but not dorsolateral prefrontallesions in monkeys Behav Brain Res 20 249ndash261
Baron-Cohen S Ring H A Wheelwright S BullmoreE T Brammer M J Simmons A amp Williams S C R1999 Social intelligence in the normal and autistic brain anfMRI study Eur J Neurosci 11 1891ndash1898
Baron-Cohen S Ring H A Bullmore E T WheelwrightS Ashwina C amp Williams S C R 2000 The amygdalatheory of autism Neurosci Biobehav Rev 24 355ndash364
Bechara A Tranel D Damasio H amp Damasio A R 1996Failure to respond autonomically to anticipated future out-comes following damage to prefrontal cortex Cerebr Cortex6 215ndash225
Benton A 1994 Face recognition Los Angeles CA WesternPsychological Services
Phil Trans R Soc Lond B (2003)
Bonda E Petrides M Ostry D amp Evans A 1996 Specificinvolvement of human parietal systems and the amygdala inthe perception of biological motion J Neurosci 16 3737ndash3744
Bowler D M amp Thommen E 2000 Attribution of mechan-ical and social causality to animated displays by childrenwith autism Autism 4 147ndash171
Brothers L 1990 The social brain a project for integratingprimate behavior and neurophysiology in a new domainConcepts Neurosci 1 27ndash151
Brothers L 1995 Neurophysiology of the perception of inten-tion by primates In The cognitive neurosciences 2nd edn (edM S Gazzaniga) pp 1107ndash1115 Cambridge MA MITPress
Butter C M McDonald J A amp Snyder D R 1969 Oralitypreference behavior and reinforcement value of nonfoodobject in monkeys with orbital frontal lesions Science 1641306ndash1307
Carmichael S T amp Price J L 1995 Limbic connections ofthe orbital and medial prefrontal cortex in macaque monk-eys J Comp Neurol 363 615ndash641
Castelli F Happe F Frith U amp Frith C 2000 Movementand mind a functional imaging study of perception andinterpretation of complex intentional movement patternsNeuroImage 12 314ndash325
Castelli F Frith C Happe F amp Frith U 2002 AutismAsperger syndrome and brain mechanisms for the attri-bution of mental states to animated shapes Brain 1251839ndash1849
Chao L L Haxby J V amp Martin A 1999 Attribute-basedneural substrates in posterior temporal cortex for perceivingand knowing about objects Nat Neurosci 2 913ndash919
Chao L L Weisberg J amp Martin A 2002 Experience-dependent modulation of category-related cortical activityCerebr Cortex 12 545ndash551
Critchley H D (and 10 others) 2000 The functional neuroan-atomy of social behaviour changes in cerebral blood flowwhen people with autistic disorder process facial expressionsBrain 123 2203ndash2212
Culham J C Cavanagh P amp Kanwisher N G 2001 Atten-tion response functions characterizing brain areas usingfMRI activation during parametric variations of attentionalload Neuron 32 737ndash745
Damasio A R Tranel D amp Damasio H 1990 Individualswith sociopathic behavior caused by frontal damage fail torespond autonomically to social stimuli Behav Brain Res41 81ndash94
Dierks T Bolte S Huble D Lanfermannm H amp PoustkaF 2001 Alterations of face processing strategies in autism(a fMRI study) Presented at the 6th Annual Meeting of theOrganization for Human Brain Mapping Brighton UK 10ndash14 June 2001
Dias R Robbins T W amp Roberts A C 1996 Dissociationin prefrontal cortex of affective and attentional shifts Nature380 69ndash72
Downing P E Jiang Y Shuman M amp Kanwisher N 2001A cortical area selective for visual processing of the humanbody Science 293 2470ndash2473
Epstein R amp Kanwisher N 1998 A cortical representation ofthe local visual environment Nature 392 598ndash601
Ernst M Zametkin A J Matochik J A Pascualvaca D ampCohen R M 1997 Reduced medial prefrontal dopami-nergic activity in autistic children Lancet 350 638
Fletcher P C Happe F Frith U Baker S C DolanR J Frackowiak R S amp Frith C D 1995 Other minds inthe brain a functional imaging study of lsquotheory of mindrsquo instory comprehension Cognition 57 109ndash128
Frith C D amp Frith U 1999 Interacting minds a biologicalbasis Science 286 1692ndash1695
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
426 R T Schultz and others FFA and social cognition
Fujita I Tanaka K Ito M amp Cheng K 1992 Columns forvisual features of objects in monkey inferotemporal cortexNature 360 343ndash346
Gaffan E A Gaffan D amp Harrison S 1988 Disconnectionof the amygdala from visual association cortex impairs vis-ual-reward association learning in monkeys J Neurosci 83144ndash3150
Gallagher H L Happe F Brunswick N Fletcher P CFrith U amp Frith C D 2000 Reading the mind in cartoonsand stories an fMRI study of lsquotheory of mindrsquo in verbal andnonverbal tasks Neuropsychologia 38 11ndash21
Gauthier I Anderson A W Tarr M J Skudlarski P ampGore J C 1997 Levels of categorization in visual recog-nition studied using functional magnetic resonance imagingCurr Biol 7 645ndash651
Gauthier I Tarr M J Anderson A W Skudlarski P ampGore J C 1999 Activation of the middle fusiform face areaincreases with expertise in recognizing novel objects NatureNeurosci 2 568ndash573
Gauthier I Skudlarski P Gore J C amp Anderson A W2000 Expertise for cars and birds recruits brain areasinvolved in face recognition Nature Neurosci 3 191ndash197
Goel V Grafman J Sadato N amp Hallett M 1995 Mode-ling other minds Neuroreport 6 1741ndash1746
Grelotti D Gauthier I amp Schultz R T 2001 Social interestand the development of cortical face specialization whatautism teaches us about face processing Devl Psychobiol 40213ndash225
Grezes J Fonlupt P Bertenthal B amp Delon-Martin C2001 Does perception of biological motion rely on specificbrain regions NeuroImage 13 775ndash785
Gusnard D A Akbudak E Shulman G L amp RaichleM E 2001 Medial prefrontal cortex and self-referentialmental activity relation to a default mode of brain functionProc Natl Acad Sci USA 98 4259ndash4264
Happe F Ehlers S Fletcher P Frith U Johansson MGillberg C Dolan R Frackowiak R amp Frith C 1996lsquoTheory of mindrsquo in the brain Evidence from a PET scanstudy of Asperger syndrome Neuroreport 8 197ndash201
Haxby J V Ungerleider L G Clark V P Schouten J LHoffman E A amp Martin A 1999 The effect of face inver-sion on activity in human neural systems for face and objectperception Neuron 22 189ndash199
Haxby J V Isahai A Chao L L Ungerleider L G ampMartin A 2000 Object-form topology in the ventral tem-poral lobe response to I Gauthier (2000) Trends Cogn Sci4 3ndash4
Haxby J V Gobbini M I Furey M L Ishai ASchouten J L amp Pietrini P 2001 Distributed and overlap-ping representations of faces and objects in ventral temporalcortex Science 293 2425ndash2430
Haznedar M M Buchsbaum M S Metzger M Solim-ando A Spiegel-Cohen J amp Hollander E 1997 Anteriorcingulate gyrus volume and glucose metabolism in autisticdisorder Am J Psychiat 154 1047ndash1050
Heider F amp Simmel M 1944 An experimental study ofapparent behavior Am J Psychol 57 243ndash259
Hobson R P Ouston J amp Lee A 1988a Whatrsquos in a faceThe case of autism Br J Psychol 79 441ndash453
Hobson R P Ouston J amp Lee A 1988b Emotion recog-nition in autism coordinating faces and voices Psychol Med18 911ndash923
Hoffman E A amp Haxby J V 2000 Distinct representationsof eye gaze and identity in the distributed human neural sys-tem for face perception Nature Neurosci 3 80ndash84
Hornak J Rolls E T amp Wade D 1996 Face and voiceexpression identification in patients with emotional andbehavioural changes following ventral frontal lobe damageNeuropsychologia 34 247ndash261
Phil Trans R Soc Lond B (2003)
Ishai A Ungerleider L G Martin A Schouten J L ampHaxby J V 1999 Distributed representation of objects inthe human ventral visual pathway Proc Natl Acad Sci USA96 9379ndash9384
Kanwisher N 2000 Domain specificity in face perceptionNature Neurosci 3 759ndash763
Kanwisher N McDermott J amp Chun M M 1997 The fusi-form face area a module of extrastriate cortex specializedfor face perception J Neurosci 17 4302ndash4311
Klin A 2000 Attributing social meaning to ambiguous visualstimuli in higher functioning autism and Asperger syndromethe social attribution task J Child Psychol Psychiat 41831ndash846
Klin A Sparrow S S de Bildt A Cicchetti D V CohenD J amp Volkmar F R 1999 A normed study of face recog-nition in autism and related disorders J Autism Devl Dis-orders 29 499ndash508
Klin A Jones W Schultz R T Volkmar F amp Cohen D J2002 Visual fixation patterns during viewing of naturalisticsocial situations as predictors of social competence in indi-viduals with autism Arch Gen Psychiat 59 809ndash816
Kracke I 1994 Developmental prosopagnosia in Aspergersyndrome presentation and discussion of an individual caseDevl Med Child Neurol 36 873ndash886
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J 1997 Neuroanatomical correlates of happi-ness sadness and disgust Am J Psychiat 154 926ndash933
Langdell T 1978 Recognition of faces an approach for thestudy of autism J Child Psychol Psychiat 19 255ndash268
LeDoux J E 1996 The emotional brain New York Simon ampShuster
Lowel S amp Singer W 1992 Selection of intrinsic horizontalconnections in the visual cortex by correlated neuronalactivity Science 255 209ndash212
Macdonald H Rutter M Howlin P Rios P Le ConteurA Evered C amp Folstein S 1989 Recognition andexpression of emotional cues by autistic and normal adultsJ Child Psychol Psychiat 30 865ndash877
Morris J S Ohman A amp Dolan R J 1998 Conscious andunconscious emotional learning in the human amygdalaNature 393 467ndash470
Myers R E Swett C amp Miller M 1973 Loss of social groupaffinity following prefrontal lesions in free-rangingmacaques Brain Res 64 257ndash269
Osterling J amp Dawson G 1994 Early recognition of childrenwith autism a study of first birthday home video tapes JAutism Devl Disorders 24 247ndash257
Pierce K Muller R A Ambrose J Allen G amp Cour-chesne E 2001 Face processing occurs outside the fusiformlsquoface arearsquo in autism evidence from functional MRI Brain124 2059ndash2073
Price J L Carmichael S T amp Drevets W C 1996 Net-works related to the orbital and medial prefrontal cortex asubstrate for emotional behavior Prog Brain Res 107523ndash536
Puce A Allison T Gore J C amp McCarthy G 1995 Face-sensitive regions in human extrastriate cortex studied byfunctional MRI J Neurophysiol 74 1192ndash1199
Reiman E M Lane R D Ahern G L Schwartz G EDavidson R J Friston K J Yun L S amp Chen K 1997Neuroanatomical correlates of externally and internally gen-erated human emotion Am J Psychiat 154 918ndash925
Rolls E T 1995 A theory of emotion and consciousness andits application to understanding the neural basis of emotionIn The cognitive neurosciences (ed M S Gazzaniga) pp1091ndash1106 Cambridge MA MIT Press
Schultz R T Gauthier I Klin A Fulbright R K Ander-son A W Volkmar F Skudlarski P Lacadie C CohenD J amp Gore J C 2000a Abnormal ventral temporal
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
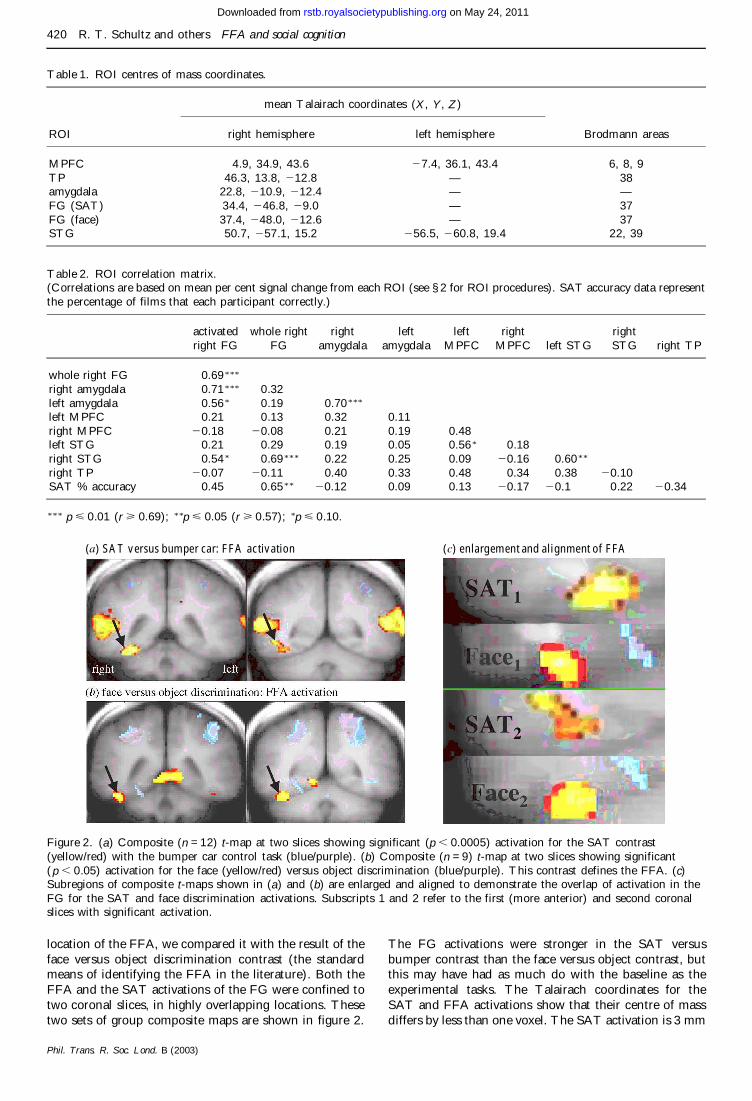
420 R T Schultz and others FFA and social cognition
Table 1 ROI centres of mass coordinates
mean Talairach coordinates (X Y Z )
ROI right hemisphere left hemisphere Brodmann areas
MPFC 49 349 436 274 361 434 6 8 9TP 463 138 2128 mdash 38amygdala 228 2109 2124 mdash mdashFG (SAT) 344 2468 290 mdash 37FG (face) 374 2480 2126 mdash 37STG 507 2571 152 2565 2608 194 22 39
Table 2 ROI correlation matrix(Correlations are based on mean per cent signal change from each ROI (see sect 2 for ROI procedures) SAT accuracy data representthe percentage of films that each participant correctly)
activated whole right right left left right rightright FG FG amygdala amygdala MPFC MPFC left STG STG right TP
whole right FG 069 curren curren curren
right amygdala 071 curren curren curren 032left amygdala 056 curren 019 070 curren curren curren
left MPFC 021 013 032 011right MPFC 2018 2008 021 019 048left STG 021 029 019 005 056 curren 018right STG 054 curren 069 curren curren curren 022 025 009 2016 060 curren curren
right TP 2007 2011 040 033 048 034 038 2010SAT accuracy 045 065 curren curren 2012 009 013 2017 201 022 2034
curren curren curren p lt 001 (r gt 069) curren curren p lt 005 (r gt 057) curren p lt 010
(a) SAT versus bumper car FFA activation
(
(c) enlargement and alignment of FFA
Figure 2 (a) Composite (n = 12) t-map at two slices showing significant (p 00005) activation for the SAT contrast(yellowred) with the bumper car control task (bluepurple) (b) Composite (n = 9) t-map at two slices showing significant(p 005) activation for the face (yellowred) versus object discrimination (bluepurple) This contrast defines the FFA (c)Subregions of composite t-maps shown in (a) and (b) are enlarged and aligned to demonstrate the overlap of activation in theFG for the SAT and face discrimination activations Subscripts 1 and 2 refer to the first (more anterior) and second coronalslices with significant activation
location of the FFA we compared it with the result of theface versus object discrimination contrast (the standardmeans of identifying the FFA in the literature) Both theFFA and the SAT activations of the FG were confined totwo coronal slices in highly overlapping locations Thesetwo sets of group composite maps are shown in figure 2
Phil Trans R Soc Lond B (2003)
The FG activations were stronger in the SAT versusbumper contrast than the face versus object contrast butthis may have had as much do with the baseline as theexperimental tasks The Talairach coordinates for theSAT and FFA activations show that their centre of massdiffers by less than one voxel The SAT activation is 3 mm
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 421
Figure 3 Scans of two individuals across three different occasions showing the reproducibility of FFA activations at two time-points and relationship to SAT activations (andashc) are from a 23-year-old male (dndashf ) are from a 24-year-old female Panels arearranged chronologically (ad) The first face versus object experiment (be) The second face versus object scan (cf ) SATversus bumper car contrast Both coronal slices are shown where there was FFA (t gt 15 in yellowred) or SAT activation(t gt 30 in yellowred) Arrows point to FG activity (right and left are reversed by convention) As in the group results (figure2) the SAT activation is centred slightly more medially along the MFS Left FG activation shown in these two subjects doesnot survive thresholding in the group composite (figure 2) Control tasks (object discrimination bumper car) are shown inpurpleblue
more medial than that of the FFA straddling the MFSthat delineates the lateral and medial aspects of the FGThe FFA on the other hand is clearly positioned in thelateral FG as expected (Haxby et al 1999) The SATactivation is also centred 36 mm more superior and about1 mm more anteriorly than the FFA A count of the over-lapping significant pixels showed that 50 of the SATactivation falls within the FFA This provides a goodapproximation of how these regions overlap Howeverthere is no definitive way to measure the percent overlapin this study because it would change with the use of dif-ferent control tasks in either condition or different signifi-cance levels for thresholding the t-maps
Two of the participants (one male one female) werealso part of a reproducibility study of the FFA and hadboth undergone the face discrimination protocol on twooccasions Figure 3 presents t-maps of their FG for eachface perception study and the SAT The female partici-pant shows reversed asymmetry as sometimes happenswith the left FG showing greater face activation than theright Nevertheless her SAT activations track her FFAand are more left-sided than typical These results showthat the FFA activation is reproducible so that the lessthan perfect overlap between the FFA and the SAT FGactivation is probably not a measurement or reliabilityissue
4 DISCUSSION
(a) The social brain networkThe current study required participants to observe the
movements of geometric figures and to interpret thesewith regard to a conceptual template about what consti-tutes a friendly or unfriendly social interaction It requiredclose attention to the contingent nature of a sequence ofmovements and inferences about mental states of eachcharacter to explain their actions Perception of the move-
Phil Trans R Soc Lond B (2003)
ments of these simple shapes as wilful seems to be auto-matic and effortless for healthy controls but not forpersons with autism (Klin 2000) As shown in figure 1the network engaged by the social attribution process (incontrast to the physical attribution control task) includednearly all of the brain areas implicated by past researchon the social brain (Brothers 1990) including cognitiveaspects such as theory of mind (Castelli et al 2000) aswell as perceptual aspects such as the perception of socialdisplays and biological movement (Allison et al 2000)We found significant activation of the bilateral MPFCsuperior STG and STS and inferior FG pars orbitalisextending into the lateral orbital gyri In addition therewere significant activations on the right side only for theamygdala TP and the FG The predominance of right-side activations is consistent with the notion that the righthemisphere is more concerned than the left with socialprocesses (Siegal et al 1996 Winner et al 2002) Ourresults differ from some past research by finding signifi-cant right FG activation and by failing to find orbito-MPFC activation
Several earlier neuroimaging studies have shown thatthe dorsal MPFC (ie that cortex anterior and superior tothe anterior cingulate gyrus) is a critical substrate for socialjudgements including empathizing and thinking aboutotherrsquos thoughts and intentions (Fletcher et al 1995 Goelet al 1995 Happe et al 1996 Castelli et al 2000 2002Gallagher et al 2000) Our findings are consistent withthese imaging studies and with non-human primate stud-ies that have documented social failures and loss of socialposition within the group following lesions to orbital pre-frontal cortices and MPFC (Butter et al 1969 Myers etal 1973 Bachevalier amp Mishkin 1986) The importantrole for the MPFC in social cognition is further suggestedby studies of autism spectrum disorders that find func-tional abnormalities in this area (Happe et al 1996 Ernstet al 1997 Haznedar et al 1997 Castelli et al 2002)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
422 R T Schultz and others FFA and social cognition
Gusnard et al (2001) suggest that the dorsal MPFC isinvolved in any kind of thought that uses the self as a refer-ent Thus the SAT activations in this area may have beendriven by theorizing about othersrsquo minds but with explicitreference to the participantrsquos own frame of reference as tohow they would feel in a similar situation
The orbital prefrontal cortex and to a lesser extent thedorsal MPFC have dense reciprocal connections withmedial temporal areas (Price et al 1996 Carmichael ampPrice 1995) providing the anatomical bases for a systemthat regulates emotional processes Damasio and col-leagues (Damasio et al 1990 Bechara et al 1996) haveargued that the orbito-MPFC have a primary function ofintegrating information about rewards and punishmentsto bias future behaviour (Rolls 1995 Dias et al 1996Hornak et al 1996 Lane et al 1997 Reiman et al 1997)A functional circuitry such as this would seem especiallyimportant in the development and acquisition of socialbehaviour However even acquired lesions to theseregions in adulthood can result in abnormalities of socialconduct (Damasio et al 1990) Brothers (1990) high-lighted the orbital prefrontal cortex as one of the threeprincipal brain regions involved in social cognition Weobserved poor signal in this region (these areas are notori-ously prone to fMRI signal drop-out and distortion) andthus we cannot know if the orbital prefrontal region wasengaged by the SAT or not However using a similarpsychological task but with PET where signal acquisitionin this region is not degraded Castelli et al (2000 2002)failed to find activation of the orbital prefrontal cortexCollectively these results call into question the impor-tance of the orbital prefrontal cortex in social cognitionand instead shift the focus on anterior cortices toward thedorsal MPFC
The SAT also generated robust activations of the rightamygdala and nearby cortex of the right TP The amyg-dala is often given a central role in theories of social per-ception and cognition (Brothers 1990 Bachevalier 1994Adolphs et al 1998 Baron-Cohen et al 2000 Schultz etal 2000b) The amygdala has a critical role in emotionalarousal assigning significance to environmental stimuliand mediating the formation of visual-reward associationsthat is lsquoemotionalrsquo learning (Gaffan et al 1988 LeDoux1996 Anderson amp Phelps 2001) It is reliably engagedduring judgements of personality characteristics from pic-tures of the face or part of the face (Adolphs et al 1998Baron-Cohen et al 1999 Winston et al 2002) Activationof the amygdala appears to be automatic and stimulusdriven as it can be engaged by images of facial expressionsin conscious awareness as well as by subliminal presen-tations of faces displaying affect (Morris et al 1998Whalen et al 1998 Critchley et al 2000) Thus theamygdalarsquos engagement by the SAT could stem from thegeneral emotional arousal evoked by the animations or itcould represent its computational role in some more spe-cific social perceptual process
The amygdala has dense reciprocal connections withthe ventral visual processing stream (Amaral amp Price1984) The strong correlation observed in this studybetween the right amygdala and the right FG could indi-cate that emotional inputs from the amygdala to the FGare necessary for engaging the social computational pro-cesses of the FG Brothers (1995) has speculated that the
Phil Trans R Soc Lond B (2003)
amygdala generates lsquosocial feelingsrsquo that are of importancein cutting through the complexities of social situations andguiding behaviour by simpler learned contingenciesSocial events are complex because their meaning oftencomes from specific combinations of features that do notadd up in a linear manner making it more difficult todecompose the events by cognitive analysis Effectivesocial engagement requires integration of context historicalrelationships and current socialndashemotional communi-cations expressed through prosodic tone as well as facialexpressions posture and gesture They are made morecomplex by the rapid pace of social transactions Thiswould necessitate some fast response system based ongeneral principles from past socialndashemotional experiencesa role for which the amygdala would seem ideally suitedIn other words the amygdala might drive intuition or lsquogutfeelingsrsquo to guide rapid non-verbal social interactionsinvolving facial expressions gesture etc Thus the strongamygdalandashFG correlation observed here could be inter-preted as the amygdala informing the FG of the relevanceof a social event and also of the outcome of its quick anddirty social perceptual analyses thereby guiding the FG(and other social nodes) in their finer grained compu-tations
Perhaps the largest and strongest activations to the SATwere those of the posterior aspects of the STS spreadinginto the adjacent STG This area has been implicated asa specific site for perception of biological motion (Bondaet al 1996 Allison et al 2000 Grezes et al 2001 Vainaet al 2001) Biological motion is a broad construct thatseems to encompass the perception of static images ofevents that could move or did move such as facialexpressions as well as actual movement by animateobjects This region of the STSndashSTG is also critical forthe decoding visual displays of social action or intention(eg gaze direction gesture and facial displays of emotion)(Critchley et al 2000 Hoffman amp Haxby 2000) In thisregard the SAT activations here are completely expectedand a testament to the effectiveness of these animationsin inducing the desired illusion of anthropomorphicaction The majority of the STSndashSTG activations wereanterior to location of the V5MT as defined in otherrecent fMRI studies (Culham et al 2001) but they alsoextended posteriorly into these more general movementsensitive cortices Unlike the amygdala some evidencesuggests that activity of the STS is mediated by explicitattention to social characteristics of the face and that STSengagement it is not automatic or stimulus driven(Winston et al 2002) We surmise therefore that acti-vation of the STS by the SAT was due to explicit taskinstructions to judge the social interactions
(b) Role of the fusiform face area in socialcognition
We also found significant SAT activation of the rightFG When compared directly in a subset of participants totheir FFA gleaned from a separate scanning session witha perceptual discrimination task of person versus objectidentity we found that both ROIs were confined to theright side on the same two coronal slices There wasabout 50 overlap between the SAT FG and FFA acti-vations and the centre of the two activations differed byless than 1 voxel The FFA activation was offset to the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 423
lateral prominence of the FG whereas the SAT activateda region of the FG that was closer to the centre of the FGbut still largely in the lateral aspect Because the SAT doesnot contain any face representations the FFA engagementwas unexpected as this region is thought of as selectivefor faces (Kanwisher et al 1997) or to other classes ofcomplex objects for which one is perceptually expert(Gauthier et al 2000)
These results have important implications for the poss-ible role of the fusiform in social brain circuitry andautism Interestingly the magnitude of the FG activationpredicted SAT task accuracy no other ROI correlated sig-nificantly with task accuracy This would seem to argueagainst any interpretation that the FFA activation is incon-sequential to the SAT that it is activated simply becauseit is well connected to other areas that are directly involvedin the social attribution process Using a task that is simi-lar to the SAT Castelli et al (2000) also reported signifi-cant right FG activation using PET However in theirfollow-up PET study comparing autism and normal con-trols (Castelli et al 2002) they failed to find significantFG activation in either group The failure of their secondstudy to find right FG activations may be due to the morestringent random effects model used in the data analysesHowever because we find right FG activation with fMRIwhich is more sensitive than PET and because Castelli etal (2000) found it with what amounts to a lower thres-hold it seems quite probable that the effect is real In factwe have preliminary evidence from an ongoing fMRIstudy that reproduces the SAT FG activations in healthycontrols (Schultz et al 2001) In addition there may besome task attributes that differ between our studies andCastelli et alrsquos that might impact into the strength withwhich FG computational processes are evoked
The key question then is why is the FG engaged bythe SAT and what role does the right FG have in socialcognition and perception Currently there are three com-peting theories of the functional organization of the FGand related ventral visual perceptual areas One modelput forth by Haxby Chao Martin and colleagues speci-fies that objects are encoded in a distributed fashion acrossa wide expanse of the ventral temporal-occipital cortex(Haxby et al 2001) They call this the lsquoobject formtypologyrsquo model (Ishai et al 1999) Their data indicatethat object category specificity is achieved by unique spa-tial patterns of activation across this extrastriate visual cor-tex They show that the pattern of activations across thiscortex is diagnostic of object category membership moreso than any localized activation maxima (Haxby et al2001) They also argue that object category perceptioninvolves retrieval of category-related information aboutspecific features and attributes of the object category(Chao et al 2002) They admit that the lsquonature of theinformation about objects that is represented in the ventraltemporal cortex is a great puzzlersquo (Haxby et al 2000 p4) but open the possibility that semantic information maybe important
A second view championed by Kanwisher et al holdsthat several select perceptual categories including facesbodies and spatial layout of places are encoded in highlyspecific locations in a modular fashion (Kanwisher et al1997 Epstein amp Kanwisher 1998 Downing et al 2001Spiridon amp Kanwisher 2002) The FFA is thought to be
Phil Trans R Soc Lond B (2003)
most concerned with discriminating among individualidentities and not with discriminating images at the categ-orical level In fact lesions to the FFA cause a specificdeficit in recognizing individual identities but not in reco-gnizing the general category of face versus non-face(Wada amp Yamamoto 2001) Spiridon amp Kanwisher (2002)provide a partial replication of Haxby et alrsquos (2001) objectform typology model They show that patterns of ventralvisual cortical activation can distinguish object categoriesNevertheless their data show category specificity for selectareas such as the FFA and they argue the ventral visualcortex is not equipotential
Gauthier et al however have argued that the ventraloccipital temporal pathway is organized by the nature ofthe perceptual computations and that these processingbiases are acquired through experience (Gauthier et al2000 Tarr amp Gauthier 2000) For example the parahip-pocampal place area is a function of a bias towards pro-cessing landscapes in terms of their spatial layout becausewe have learned through repeated experiences that this isvery useful information to extract when perceiving land-scapes and related visual images Similarly we learn quiteearly in life that it is important to discriminate faces on anindividual level for example discriminating mother fromothers This bias towards individual identification accord-ing to Gauthier accounts for the FFArsquos apparent modu-larity (Gauthier et al 1997 2000 Tarr amp Gauthier 2000)According to this third model the type of informationneeded and our cumulative experience in processing thatinformation organizes the ventral occipitalndashtemporal path-way into regional centres with preferred modes of pro-cessing
All three models seem to agree that the functionalorganization of the ventral visual cortices is driven by theneed to categorize perceptions into object classes Webelieve that our findings showing FFA activation by non-face objects are consistent with aspects of both the pro-cessing map model of Gauthier and the object formtypology model of Haxby et al First consistent withHaxby et al we suggest that semantic information isimportant to the ventral visual pathway for object categor-ization Second consistent with Gauthier et al we believethat repeated perceptual experience with faces biases thetype of information that FG finds important We proposethat the middle FG area encodes semantic attributes ofpeople because of repeated perceptual experiences withfaces that occur during social interactions In fact wewould guess that most perceptual experiences with facesoccur during social situations and these social situationsoften involve repeated social judgements Thus infor-mation about the social nature of people might be storedin the FG (though not exclusively)
The nature of the semantic information stored in theFG might be restricted to anything that would be helpfulin defining faces as a distinct category of object becausemaking such distinctions appears to be the primary chargeof the ventral visual pathway This would include knowl-edge of people as having personal agency and of havingthe capability to disturb each otherrsquos emotional homeo-stasis (eg to act friendly or unfriendly) By pretendingthat the three shapes in the SAT are people stored knowl-edge about people in social interactions might beretrieved causing the observed activations in the FG
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
424 R T Schultz and others FFA and social cognition
There would be a measure of efficiency from thisarrangement that is having representations and compu-tations of more abstract attributes of people inter-digitatedwith front-end perceptual processes about physical attri-butes Gauthierrsquos claim is that the same group of neuronscan be engaged by perceptually different categories ofobjects it is the type of processing rather than the visualdetails that are important Extrapolating from this wewould argue that the SAT engages a region of the FGoverlapping with the FFA because it demands compu-tational processes to classify the SAT geometric figures aspeople or person-like Chao et al (1999) reach a similarconclusion They argued that activity in the ventral path-way reflects stored information about an object categorynot just physical features They point out that thisarrangement could explain why lesion patients with cate-gory specific perceptual deficits also have trouble retriev-ing general information about that visual category Thuswe would add that the FFA must store general infor-mation about people or some meta-representation oflsquopeoplenessrsquo
(c) Implications for the pathobiology of autismThe SAT used in this current study appears to be an
effective neurobehavioural probe for engaging a distrib-uted network of brain regions involved in different aspectsof social perception and cognition It will be important touse this and similar procedures in persons with autism tobetter define the nature of brain functions in this disorderCastelli et al (2002) have already taken the first step inthis process and they describe a pattern of hypoactivationin the MPFC STS and TPs in autism In an ongoingstudy of autism spectrum disorders we have presentedpreliminary data using the SAT showing that we too findhypoactivation of these regions and in the amygdala andFG (Schultz et al 2001) Thus we predict that futurework in this area will show that the entire social brain net-work is underactive in autism during tasks requiring socialperceptual and social cognitive processing
There are already sufficient data to argue strongly for arole of the FG in the pathobiology of autism No less thanfive previous fMRI studies have shown the FFA region tobe significantly less engaged among persons with autismcompared with controls during face perception tasks(Critchley et al 2000 Dierks et al 2001 Pierce et al2001 Schultz et al 2000a 2001) Activation of the FGby the social judgements in the current study adds animportant piece of evidence in favour of a causal role forthe FG in the pathobiology of autism This conclusionhowever must remain tentative until additional studiesmore precisely define the factors leading to FG activationduring social attribution and gather better data to proveits computational role in social cognition In addition theeffect of FG lesions for social functioning must be clari-fied If the FG is involved in social computations onewould expect to find social cognitive deficits in personswith lesions to this area However social deficits in proso-pagnosics have not been widely reported It might be thatprosopagnosic patients have not been carefully tested onthis dimension In this regard using our behavioural ver-sion of the SAT (Klin 2000) to test social perceptionamong prosopagnosics would be quite interesting Theparallels however between autism and prosopagnosia are
Phil Trans R Soc Lond B (2003)
incomplete because autism is clearly a developmental dis-order whereas the research literature on prosopagnosia ismostly confined to lesions acquired in adolescence oradulthood Among the few cases of developmental proso-pagnosia reported in the literature there is indeed one thathighlights severe social impairments (Kracke 1994) Itmay be that the role of the FG in the development ofautistic symptoms is different from the role of the FG inthe maintenance of social cognitive functions after brainmaturity
It is also possible that the role of the FG in social pro-cesses is dependent on its functional relationships withother nodes in the social brain and that it is the collectiveaction and interaction of the network that is of primaryimportance for social behaviours For example we founda strong correlation across participants in the amount ofFG and amygdala activation A strictly modular view ofthese areas may be inappropriate as the functions of eachnode could be quite dependent on one another and whenonly considered in isolation quite insufficient to supportsocial processes Thus whereas a lesion to the FG mayimpair visual perception it might not have a large impacton social functioning if this depends on an extended net-work that is dynamic and capable of compensatory adjust-ments Dysfunction of the FG may be necessary but notsufficient to produce social deficits Indeed our relianceon modular models of brain functioning may be leadingus astray in our search for causal mechanisms in autismInstead it may be the collective action of a distributedsystem that is critical to the pathobiology of autism TheFG region may be a key partner in this distributed systembut nevertheless just one node and insufficient by itselfto support in any substantive manner social cognitive pro-cesses or to explain social cognitive deficits seen inautism
5 CONCLUSION AND FUTURE DIRECTIONS
Using the SAT we isolated a distributed network ofactivations that conform to the emerging model of thesocial brain Most important we found significant acti-vation of the central aspect of the FG thus adding thisregion to the expanding list of structures involved in socialprocesses The FG region activated by the SAT overlapsin its spatial extent with the FFA with centres of massdiffering by less than 1 voxel We speculate that these SATactivations represent computational processes associatedwith more abstract attributes of people
It is an open question as to whether the substantial over-lap between the FFA and SAT FG activation is due to anoverlapping or shared set of neuronal assemblies This willneed to be clarified by future work Electrophysiologicalstudies show that there are small face-specific patches inthe FG cortex (Allison et al 1999) In the current studysmall patches of face cells and SAT cells could be inter-mingled but distinct and not drawing on any of the sameneuronal assemblies The spatial resolution of fMRI haslimitations that may preclude any definitive answer to thisquestion but there are strategies that can be used toaddress the issue For example the current study designdid not contain SAT and face discriminations in the sameexperimental fMRI series Doing so would enable directcomparisons of the computational demands placed on the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 425
common area of the FFA It also would be informative iffollow-up studies superimposed faces within the geometricfigures of the SAT without changing the film scripts inany way We could then determine if the computations ofthe common area of the FFA increase in a predictablefashionmdashwould the activation be a linear summation ofthe original SAT plus face discrimination Any significantdeviation from an additive model would suggest that thereis some sharing of neuronal assemblies with the SAT andface discrimination tasks when presented alone
This work was supported by grants from the National Institutesof Child Health and Human Development (grants PO1 HD03008 and PO1 HDDC35482) and the Korczak FoundationThe authors thank the anonymous reviewers for their helpfulcomments Michael Lee for programming the fMRI tasksHedy Serofin and Terry Hickey for their assistance in acquiringthe fMRI data John Herrington for his valuable assistance indata analyses of preliminary studies and members of the YaleDevelopmental Neuroimaging Program for their helpful com-ments on this manuscript A special acknowledgement ofappreciation is given to Donald Cohen who passed away inOctober of 2001 for his outstanding mentorship in all aspectsof this program of work
REFERENCES
Abell F Happe F amp Frith U 2000 Do triangles play tricksAttribution of mental states to animated shapes in normaland abnormal development J Cogn Dev 15 1ndash20
Adolphs R Tranel D amp Damasio A R 1998 The humanamygdala in social judgement Nature 393 470ndash474
Allison T Puce A Spencer D D amp McCarthy G 1999Electrophysiological studies of human face perception Ipotentials generated in occipitotemporal cortex by face andnon-face stimuli Cerebr Cortex 9 415ndash430
Allison T Puce A amp McCarthy G 2000 Social perceptionfrom visual cues role of the STS region Trends Cogn Sci4 267ndash278
Amaral D G amp Price J L 1984 Amygdalo-cortical projec-tions in the monkey (Macaca fascicularis) J Comp Neurol230 465ndash496
Amaral D G Price J L Pitkanen A amp Carmichael S T1992 Anatomical organization of the primate amygdaloidcomplex In The amygdala neurobiological aspects of emotionmemory and mental dysfunction (ed J Aggleton) pp 1ndash66New York Wiley-Liss
Anderson A K amp Phelps E A 2001 Lesions of the humanamygdala impair enhanced perception of emotionally salientevents Nature 411 305ndash309
Bachevalier J 1994 Medial temporal lobe structures andautism a review of clinical and experimental findingsNeuropsychologia 32 627ndash648
Bachevalier J amp Mishkin M 1986 Visual recognition impair-ment follows ventromedial but not dorsolateral prefrontallesions in monkeys Behav Brain Res 20 249ndash261
Baron-Cohen S Ring H A Wheelwright S BullmoreE T Brammer M J Simmons A amp Williams S C R1999 Social intelligence in the normal and autistic brain anfMRI study Eur J Neurosci 11 1891ndash1898
Baron-Cohen S Ring H A Bullmore E T WheelwrightS Ashwina C amp Williams S C R 2000 The amygdalatheory of autism Neurosci Biobehav Rev 24 355ndash364
Bechara A Tranel D Damasio H amp Damasio A R 1996Failure to respond autonomically to anticipated future out-comes following damage to prefrontal cortex Cerebr Cortex6 215ndash225
Benton A 1994 Face recognition Los Angeles CA WesternPsychological Services
Phil Trans R Soc Lond B (2003)
Bonda E Petrides M Ostry D amp Evans A 1996 Specificinvolvement of human parietal systems and the amygdala inthe perception of biological motion J Neurosci 16 3737ndash3744
Bowler D M amp Thommen E 2000 Attribution of mechan-ical and social causality to animated displays by childrenwith autism Autism 4 147ndash171
Brothers L 1990 The social brain a project for integratingprimate behavior and neurophysiology in a new domainConcepts Neurosci 1 27ndash151
Brothers L 1995 Neurophysiology of the perception of inten-tion by primates In The cognitive neurosciences 2nd edn (edM S Gazzaniga) pp 1107ndash1115 Cambridge MA MITPress
Butter C M McDonald J A amp Snyder D R 1969 Oralitypreference behavior and reinforcement value of nonfoodobject in monkeys with orbital frontal lesions Science 1641306ndash1307
Carmichael S T amp Price J L 1995 Limbic connections ofthe orbital and medial prefrontal cortex in macaque monk-eys J Comp Neurol 363 615ndash641
Castelli F Happe F Frith U amp Frith C 2000 Movementand mind a functional imaging study of perception andinterpretation of complex intentional movement patternsNeuroImage 12 314ndash325
Castelli F Frith C Happe F amp Frith U 2002 AutismAsperger syndrome and brain mechanisms for the attri-bution of mental states to animated shapes Brain 1251839ndash1849
Chao L L Haxby J V amp Martin A 1999 Attribute-basedneural substrates in posterior temporal cortex for perceivingand knowing about objects Nat Neurosci 2 913ndash919
Chao L L Weisberg J amp Martin A 2002 Experience-dependent modulation of category-related cortical activityCerebr Cortex 12 545ndash551
Critchley H D (and 10 others) 2000 The functional neuroan-atomy of social behaviour changes in cerebral blood flowwhen people with autistic disorder process facial expressionsBrain 123 2203ndash2212
Culham J C Cavanagh P amp Kanwisher N G 2001 Atten-tion response functions characterizing brain areas usingfMRI activation during parametric variations of attentionalload Neuron 32 737ndash745
Damasio A R Tranel D amp Damasio H 1990 Individualswith sociopathic behavior caused by frontal damage fail torespond autonomically to social stimuli Behav Brain Res41 81ndash94
Dierks T Bolte S Huble D Lanfermannm H amp PoustkaF 2001 Alterations of face processing strategies in autism(a fMRI study) Presented at the 6th Annual Meeting of theOrganization for Human Brain Mapping Brighton UK 10ndash14 June 2001
Dias R Robbins T W amp Roberts A C 1996 Dissociationin prefrontal cortex of affective and attentional shifts Nature380 69ndash72
Downing P E Jiang Y Shuman M amp Kanwisher N 2001A cortical area selective for visual processing of the humanbody Science 293 2470ndash2473
Epstein R amp Kanwisher N 1998 A cortical representation ofthe local visual environment Nature 392 598ndash601
Ernst M Zametkin A J Matochik J A Pascualvaca D ampCohen R M 1997 Reduced medial prefrontal dopami-nergic activity in autistic children Lancet 350 638
Fletcher P C Happe F Frith U Baker S C DolanR J Frackowiak R S amp Frith C D 1995 Other minds inthe brain a functional imaging study of lsquotheory of mindrsquo instory comprehension Cognition 57 109ndash128
Frith C D amp Frith U 1999 Interacting minds a biologicalbasis Science 286 1692ndash1695
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
426 R T Schultz and others FFA and social cognition
Fujita I Tanaka K Ito M amp Cheng K 1992 Columns forvisual features of objects in monkey inferotemporal cortexNature 360 343ndash346
Gaffan E A Gaffan D amp Harrison S 1988 Disconnectionof the amygdala from visual association cortex impairs vis-ual-reward association learning in monkeys J Neurosci 83144ndash3150
Gallagher H L Happe F Brunswick N Fletcher P CFrith U amp Frith C D 2000 Reading the mind in cartoonsand stories an fMRI study of lsquotheory of mindrsquo in verbal andnonverbal tasks Neuropsychologia 38 11ndash21
Gauthier I Anderson A W Tarr M J Skudlarski P ampGore J C 1997 Levels of categorization in visual recog-nition studied using functional magnetic resonance imagingCurr Biol 7 645ndash651
Gauthier I Tarr M J Anderson A W Skudlarski P ampGore J C 1999 Activation of the middle fusiform face areaincreases with expertise in recognizing novel objects NatureNeurosci 2 568ndash573
Gauthier I Skudlarski P Gore J C amp Anderson A W2000 Expertise for cars and birds recruits brain areasinvolved in face recognition Nature Neurosci 3 191ndash197
Goel V Grafman J Sadato N amp Hallett M 1995 Mode-ling other minds Neuroreport 6 1741ndash1746
Grelotti D Gauthier I amp Schultz R T 2001 Social interestand the development of cortical face specialization whatautism teaches us about face processing Devl Psychobiol 40213ndash225
Grezes J Fonlupt P Bertenthal B amp Delon-Martin C2001 Does perception of biological motion rely on specificbrain regions NeuroImage 13 775ndash785
Gusnard D A Akbudak E Shulman G L amp RaichleM E 2001 Medial prefrontal cortex and self-referentialmental activity relation to a default mode of brain functionProc Natl Acad Sci USA 98 4259ndash4264
Happe F Ehlers S Fletcher P Frith U Johansson MGillberg C Dolan R Frackowiak R amp Frith C 1996lsquoTheory of mindrsquo in the brain Evidence from a PET scanstudy of Asperger syndrome Neuroreport 8 197ndash201
Haxby J V Ungerleider L G Clark V P Schouten J LHoffman E A amp Martin A 1999 The effect of face inver-sion on activity in human neural systems for face and objectperception Neuron 22 189ndash199
Haxby J V Isahai A Chao L L Ungerleider L G ampMartin A 2000 Object-form topology in the ventral tem-poral lobe response to I Gauthier (2000) Trends Cogn Sci4 3ndash4
Haxby J V Gobbini M I Furey M L Ishai ASchouten J L amp Pietrini P 2001 Distributed and overlap-ping representations of faces and objects in ventral temporalcortex Science 293 2425ndash2430
Haznedar M M Buchsbaum M S Metzger M Solim-ando A Spiegel-Cohen J amp Hollander E 1997 Anteriorcingulate gyrus volume and glucose metabolism in autisticdisorder Am J Psychiat 154 1047ndash1050
Heider F amp Simmel M 1944 An experimental study ofapparent behavior Am J Psychol 57 243ndash259
Hobson R P Ouston J amp Lee A 1988a Whatrsquos in a faceThe case of autism Br J Psychol 79 441ndash453
Hobson R P Ouston J amp Lee A 1988b Emotion recog-nition in autism coordinating faces and voices Psychol Med18 911ndash923
Hoffman E A amp Haxby J V 2000 Distinct representationsof eye gaze and identity in the distributed human neural sys-tem for face perception Nature Neurosci 3 80ndash84
Hornak J Rolls E T amp Wade D 1996 Face and voiceexpression identification in patients with emotional andbehavioural changes following ventral frontal lobe damageNeuropsychologia 34 247ndash261
Phil Trans R Soc Lond B (2003)
Ishai A Ungerleider L G Martin A Schouten J L ampHaxby J V 1999 Distributed representation of objects inthe human ventral visual pathway Proc Natl Acad Sci USA96 9379ndash9384
Kanwisher N 2000 Domain specificity in face perceptionNature Neurosci 3 759ndash763
Kanwisher N McDermott J amp Chun M M 1997 The fusi-form face area a module of extrastriate cortex specializedfor face perception J Neurosci 17 4302ndash4311
Klin A 2000 Attributing social meaning to ambiguous visualstimuli in higher functioning autism and Asperger syndromethe social attribution task J Child Psychol Psychiat 41831ndash846
Klin A Sparrow S S de Bildt A Cicchetti D V CohenD J amp Volkmar F R 1999 A normed study of face recog-nition in autism and related disorders J Autism Devl Dis-orders 29 499ndash508
Klin A Jones W Schultz R T Volkmar F amp Cohen D J2002 Visual fixation patterns during viewing of naturalisticsocial situations as predictors of social competence in indi-viduals with autism Arch Gen Psychiat 59 809ndash816
Kracke I 1994 Developmental prosopagnosia in Aspergersyndrome presentation and discussion of an individual caseDevl Med Child Neurol 36 873ndash886
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J 1997 Neuroanatomical correlates of happi-ness sadness and disgust Am J Psychiat 154 926ndash933
Langdell T 1978 Recognition of faces an approach for thestudy of autism J Child Psychol Psychiat 19 255ndash268
LeDoux J E 1996 The emotional brain New York Simon ampShuster
Lowel S amp Singer W 1992 Selection of intrinsic horizontalconnections in the visual cortex by correlated neuronalactivity Science 255 209ndash212
Macdonald H Rutter M Howlin P Rios P Le ConteurA Evered C amp Folstein S 1989 Recognition andexpression of emotional cues by autistic and normal adultsJ Child Psychol Psychiat 30 865ndash877
Morris J S Ohman A amp Dolan R J 1998 Conscious andunconscious emotional learning in the human amygdalaNature 393 467ndash470
Myers R E Swett C amp Miller M 1973 Loss of social groupaffinity following prefrontal lesions in free-rangingmacaques Brain Res 64 257ndash269
Osterling J amp Dawson G 1994 Early recognition of childrenwith autism a study of first birthday home video tapes JAutism Devl Disorders 24 247ndash257
Pierce K Muller R A Ambrose J Allen G amp Cour-chesne E 2001 Face processing occurs outside the fusiformlsquoface arearsquo in autism evidence from functional MRI Brain124 2059ndash2073
Price J L Carmichael S T amp Drevets W C 1996 Net-works related to the orbital and medial prefrontal cortex asubstrate for emotional behavior Prog Brain Res 107523ndash536
Puce A Allison T Gore J C amp McCarthy G 1995 Face-sensitive regions in human extrastriate cortex studied byfunctional MRI J Neurophysiol 74 1192ndash1199
Reiman E M Lane R D Ahern G L Schwartz G EDavidson R J Friston K J Yun L S amp Chen K 1997Neuroanatomical correlates of externally and internally gen-erated human emotion Am J Psychiat 154 918ndash925
Rolls E T 1995 A theory of emotion and consciousness andits application to understanding the neural basis of emotionIn The cognitive neurosciences (ed M S Gazzaniga) pp1091ndash1106 Cambridge MA MIT Press
Schultz R T Gauthier I Klin A Fulbright R K Ander-son A W Volkmar F Skudlarski P Lacadie C CohenD J amp Gore J C 2000a Abnormal ventral temporal
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from

FFA and social cognition R T Schultz and others 421
Figure 3 Scans of two individuals across three different occasions showing the reproducibility of FFA activations at two time-points and relationship to SAT activations (andashc) are from a 23-year-old male (dndashf ) are from a 24-year-old female Panels arearranged chronologically (ad) The first face versus object experiment (be) The second face versus object scan (cf ) SATversus bumper car contrast Both coronal slices are shown where there was FFA (t gt 15 in yellowred) or SAT activation(t gt 30 in yellowred) Arrows point to FG activity (right and left are reversed by convention) As in the group results (figure2) the SAT activation is centred slightly more medially along the MFS Left FG activation shown in these two subjects doesnot survive thresholding in the group composite (figure 2) Control tasks (object discrimination bumper car) are shown inpurpleblue
more medial than that of the FFA straddling the MFSthat delineates the lateral and medial aspects of the FGThe FFA on the other hand is clearly positioned in thelateral FG as expected (Haxby et al 1999) The SATactivation is also centred 36 mm more superior and about1 mm more anteriorly than the FFA A count of the over-lapping significant pixels showed that 50 of the SATactivation falls within the FFA This provides a goodapproximation of how these regions overlap Howeverthere is no definitive way to measure the percent overlapin this study because it would change with the use of dif-ferent control tasks in either condition or different signifi-cance levels for thresholding the t-maps
Two of the participants (one male one female) werealso part of a reproducibility study of the FFA and hadboth undergone the face discrimination protocol on twooccasions Figure 3 presents t-maps of their FG for eachface perception study and the SAT The female partici-pant shows reversed asymmetry as sometimes happenswith the left FG showing greater face activation than theright Nevertheless her SAT activations track her FFAand are more left-sided than typical These results showthat the FFA activation is reproducible so that the lessthan perfect overlap between the FFA and the SAT FGactivation is probably not a measurement or reliabilityissue
4 DISCUSSION
(a) The social brain networkThe current study required participants to observe the
movements of geometric figures and to interpret thesewith regard to a conceptual template about what consti-tutes a friendly or unfriendly social interaction It requiredclose attention to the contingent nature of a sequence ofmovements and inferences about mental states of eachcharacter to explain their actions Perception of the move-
Phil Trans R Soc Lond B (2003)
ments of these simple shapes as wilful seems to be auto-matic and effortless for healthy controls but not forpersons with autism (Klin 2000) As shown in figure 1the network engaged by the social attribution process (incontrast to the physical attribution control task) includednearly all of the brain areas implicated by past researchon the social brain (Brothers 1990) including cognitiveaspects such as theory of mind (Castelli et al 2000) aswell as perceptual aspects such as the perception of socialdisplays and biological movement (Allison et al 2000)We found significant activation of the bilateral MPFCsuperior STG and STS and inferior FG pars orbitalisextending into the lateral orbital gyri In addition therewere significant activations on the right side only for theamygdala TP and the FG The predominance of right-side activations is consistent with the notion that the righthemisphere is more concerned than the left with socialprocesses (Siegal et al 1996 Winner et al 2002) Ourresults differ from some past research by finding signifi-cant right FG activation and by failing to find orbito-MPFC activation
Several earlier neuroimaging studies have shown thatthe dorsal MPFC (ie that cortex anterior and superior tothe anterior cingulate gyrus) is a critical substrate for socialjudgements including empathizing and thinking aboutotherrsquos thoughts and intentions (Fletcher et al 1995 Goelet al 1995 Happe et al 1996 Castelli et al 2000 2002Gallagher et al 2000) Our findings are consistent withthese imaging studies and with non-human primate stud-ies that have documented social failures and loss of socialposition within the group following lesions to orbital pre-frontal cortices and MPFC (Butter et al 1969 Myers etal 1973 Bachevalier amp Mishkin 1986) The importantrole for the MPFC in social cognition is further suggestedby studies of autism spectrum disorders that find func-tional abnormalities in this area (Happe et al 1996 Ernstet al 1997 Haznedar et al 1997 Castelli et al 2002)
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
422 R T Schultz and others FFA and social cognition
Gusnard et al (2001) suggest that the dorsal MPFC isinvolved in any kind of thought that uses the self as a refer-ent Thus the SAT activations in this area may have beendriven by theorizing about othersrsquo minds but with explicitreference to the participantrsquos own frame of reference as tohow they would feel in a similar situation
The orbital prefrontal cortex and to a lesser extent thedorsal MPFC have dense reciprocal connections withmedial temporal areas (Price et al 1996 Carmichael ampPrice 1995) providing the anatomical bases for a systemthat regulates emotional processes Damasio and col-leagues (Damasio et al 1990 Bechara et al 1996) haveargued that the orbito-MPFC have a primary function ofintegrating information about rewards and punishmentsto bias future behaviour (Rolls 1995 Dias et al 1996Hornak et al 1996 Lane et al 1997 Reiman et al 1997)A functional circuitry such as this would seem especiallyimportant in the development and acquisition of socialbehaviour However even acquired lesions to theseregions in adulthood can result in abnormalities of socialconduct (Damasio et al 1990) Brothers (1990) high-lighted the orbital prefrontal cortex as one of the threeprincipal brain regions involved in social cognition Weobserved poor signal in this region (these areas are notori-ously prone to fMRI signal drop-out and distortion) andthus we cannot know if the orbital prefrontal region wasengaged by the SAT or not However using a similarpsychological task but with PET where signal acquisitionin this region is not degraded Castelli et al (2000 2002)failed to find activation of the orbital prefrontal cortexCollectively these results call into question the impor-tance of the orbital prefrontal cortex in social cognitionand instead shift the focus on anterior cortices toward thedorsal MPFC
The SAT also generated robust activations of the rightamygdala and nearby cortex of the right TP The amyg-dala is often given a central role in theories of social per-ception and cognition (Brothers 1990 Bachevalier 1994Adolphs et al 1998 Baron-Cohen et al 2000 Schultz etal 2000b) The amygdala has a critical role in emotionalarousal assigning significance to environmental stimuliand mediating the formation of visual-reward associationsthat is lsquoemotionalrsquo learning (Gaffan et al 1988 LeDoux1996 Anderson amp Phelps 2001) It is reliably engagedduring judgements of personality characteristics from pic-tures of the face or part of the face (Adolphs et al 1998Baron-Cohen et al 1999 Winston et al 2002) Activationof the amygdala appears to be automatic and stimulusdriven as it can be engaged by images of facial expressionsin conscious awareness as well as by subliminal presen-tations of faces displaying affect (Morris et al 1998Whalen et al 1998 Critchley et al 2000) Thus theamygdalarsquos engagement by the SAT could stem from thegeneral emotional arousal evoked by the animations or itcould represent its computational role in some more spe-cific social perceptual process
The amygdala has dense reciprocal connections withthe ventral visual processing stream (Amaral amp Price1984) The strong correlation observed in this studybetween the right amygdala and the right FG could indi-cate that emotional inputs from the amygdala to the FGare necessary for engaging the social computational pro-cesses of the FG Brothers (1995) has speculated that the
Phil Trans R Soc Lond B (2003)
amygdala generates lsquosocial feelingsrsquo that are of importancein cutting through the complexities of social situations andguiding behaviour by simpler learned contingenciesSocial events are complex because their meaning oftencomes from specific combinations of features that do notadd up in a linear manner making it more difficult todecompose the events by cognitive analysis Effectivesocial engagement requires integration of context historicalrelationships and current socialndashemotional communi-cations expressed through prosodic tone as well as facialexpressions posture and gesture They are made morecomplex by the rapid pace of social transactions Thiswould necessitate some fast response system based ongeneral principles from past socialndashemotional experiencesa role for which the amygdala would seem ideally suitedIn other words the amygdala might drive intuition or lsquogutfeelingsrsquo to guide rapid non-verbal social interactionsinvolving facial expressions gesture etc Thus the strongamygdalandashFG correlation observed here could be inter-preted as the amygdala informing the FG of the relevanceof a social event and also of the outcome of its quick anddirty social perceptual analyses thereby guiding the FG(and other social nodes) in their finer grained compu-tations
Perhaps the largest and strongest activations to the SATwere those of the posterior aspects of the STS spreadinginto the adjacent STG This area has been implicated asa specific site for perception of biological motion (Bondaet al 1996 Allison et al 2000 Grezes et al 2001 Vainaet al 2001) Biological motion is a broad construct thatseems to encompass the perception of static images ofevents that could move or did move such as facialexpressions as well as actual movement by animateobjects This region of the STSndashSTG is also critical forthe decoding visual displays of social action or intention(eg gaze direction gesture and facial displays of emotion)(Critchley et al 2000 Hoffman amp Haxby 2000) In thisregard the SAT activations here are completely expectedand a testament to the effectiveness of these animationsin inducing the desired illusion of anthropomorphicaction The majority of the STSndashSTG activations wereanterior to location of the V5MT as defined in otherrecent fMRI studies (Culham et al 2001) but they alsoextended posteriorly into these more general movementsensitive cortices Unlike the amygdala some evidencesuggests that activity of the STS is mediated by explicitattention to social characteristics of the face and that STSengagement it is not automatic or stimulus driven(Winston et al 2002) We surmise therefore that acti-vation of the STS by the SAT was due to explicit taskinstructions to judge the social interactions
(b) Role of the fusiform face area in socialcognition
We also found significant SAT activation of the rightFG When compared directly in a subset of participants totheir FFA gleaned from a separate scanning session witha perceptual discrimination task of person versus objectidentity we found that both ROIs were confined to theright side on the same two coronal slices There wasabout 50 overlap between the SAT FG and FFA acti-vations and the centre of the two activations differed byless than 1 voxel The FFA activation was offset to the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 423
lateral prominence of the FG whereas the SAT activateda region of the FG that was closer to the centre of the FGbut still largely in the lateral aspect Because the SAT doesnot contain any face representations the FFA engagementwas unexpected as this region is thought of as selectivefor faces (Kanwisher et al 1997) or to other classes ofcomplex objects for which one is perceptually expert(Gauthier et al 2000)
These results have important implications for the poss-ible role of the fusiform in social brain circuitry andautism Interestingly the magnitude of the FG activationpredicted SAT task accuracy no other ROI correlated sig-nificantly with task accuracy This would seem to argueagainst any interpretation that the FFA activation is incon-sequential to the SAT that it is activated simply becauseit is well connected to other areas that are directly involvedin the social attribution process Using a task that is simi-lar to the SAT Castelli et al (2000) also reported signifi-cant right FG activation using PET However in theirfollow-up PET study comparing autism and normal con-trols (Castelli et al 2002) they failed to find significantFG activation in either group The failure of their secondstudy to find right FG activations may be due to the morestringent random effects model used in the data analysesHowever because we find right FG activation with fMRIwhich is more sensitive than PET and because Castelli etal (2000) found it with what amounts to a lower thres-hold it seems quite probable that the effect is real In factwe have preliminary evidence from an ongoing fMRIstudy that reproduces the SAT FG activations in healthycontrols (Schultz et al 2001) In addition there may besome task attributes that differ between our studies andCastelli et alrsquos that might impact into the strength withwhich FG computational processes are evoked
The key question then is why is the FG engaged bythe SAT and what role does the right FG have in socialcognition and perception Currently there are three com-peting theories of the functional organization of the FGand related ventral visual perceptual areas One modelput forth by Haxby Chao Martin and colleagues speci-fies that objects are encoded in a distributed fashion acrossa wide expanse of the ventral temporal-occipital cortex(Haxby et al 2001) They call this the lsquoobject formtypologyrsquo model (Ishai et al 1999) Their data indicatethat object category specificity is achieved by unique spa-tial patterns of activation across this extrastriate visual cor-tex They show that the pattern of activations across thiscortex is diagnostic of object category membership moreso than any localized activation maxima (Haxby et al2001) They also argue that object category perceptioninvolves retrieval of category-related information aboutspecific features and attributes of the object category(Chao et al 2002) They admit that the lsquonature of theinformation about objects that is represented in the ventraltemporal cortex is a great puzzlersquo (Haxby et al 2000 p4) but open the possibility that semantic information maybe important
A second view championed by Kanwisher et al holdsthat several select perceptual categories including facesbodies and spatial layout of places are encoded in highlyspecific locations in a modular fashion (Kanwisher et al1997 Epstein amp Kanwisher 1998 Downing et al 2001Spiridon amp Kanwisher 2002) The FFA is thought to be
Phil Trans R Soc Lond B (2003)
most concerned with discriminating among individualidentities and not with discriminating images at the categ-orical level In fact lesions to the FFA cause a specificdeficit in recognizing individual identities but not in reco-gnizing the general category of face versus non-face(Wada amp Yamamoto 2001) Spiridon amp Kanwisher (2002)provide a partial replication of Haxby et alrsquos (2001) objectform typology model They show that patterns of ventralvisual cortical activation can distinguish object categoriesNevertheless their data show category specificity for selectareas such as the FFA and they argue the ventral visualcortex is not equipotential
Gauthier et al however have argued that the ventraloccipital temporal pathway is organized by the nature ofthe perceptual computations and that these processingbiases are acquired through experience (Gauthier et al2000 Tarr amp Gauthier 2000) For example the parahip-pocampal place area is a function of a bias towards pro-cessing landscapes in terms of their spatial layout becausewe have learned through repeated experiences that this isvery useful information to extract when perceiving land-scapes and related visual images Similarly we learn quiteearly in life that it is important to discriminate faces on anindividual level for example discriminating mother fromothers This bias towards individual identification accord-ing to Gauthier accounts for the FFArsquos apparent modu-larity (Gauthier et al 1997 2000 Tarr amp Gauthier 2000)According to this third model the type of informationneeded and our cumulative experience in processing thatinformation organizes the ventral occipitalndashtemporal path-way into regional centres with preferred modes of pro-cessing
All three models seem to agree that the functionalorganization of the ventral visual cortices is driven by theneed to categorize perceptions into object classes Webelieve that our findings showing FFA activation by non-face objects are consistent with aspects of both the pro-cessing map model of Gauthier and the object formtypology model of Haxby et al First consistent withHaxby et al we suggest that semantic information isimportant to the ventral visual pathway for object categor-ization Second consistent with Gauthier et al we believethat repeated perceptual experience with faces biases thetype of information that FG finds important We proposethat the middle FG area encodes semantic attributes ofpeople because of repeated perceptual experiences withfaces that occur during social interactions In fact wewould guess that most perceptual experiences with facesoccur during social situations and these social situationsoften involve repeated social judgements Thus infor-mation about the social nature of people might be storedin the FG (though not exclusively)
The nature of the semantic information stored in theFG might be restricted to anything that would be helpfulin defining faces as a distinct category of object becausemaking such distinctions appears to be the primary chargeof the ventral visual pathway This would include knowl-edge of people as having personal agency and of havingthe capability to disturb each otherrsquos emotional homeo-stasis (eg to act friendly or unfriendly) By pretendingthat the three shapes in the SAT are people stored knowl-edge about people in social interactions might beretrieved causing the observed activations in the FG
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
424 R T Schultz and others FFA and social cognition
There would be a measure of efficiency from thisarrangement that is having representations and compu-tations of more abstract attributes of people inter-digitatedwith front-end perceptual processes about physical attri-butes Gauthierrsquos claim is that the same group of neuronscan be engaged by perceptually different categories ofobjects it is the type of processing rather than the visualdetails that are important Extrapolating from this wewould argue that the SAT engages a region of the FGoverlapping with the FFA because it demands compu-tational processes to classify the SAT geometric figures aspeople or person-like Chao et al (1999) reach a similarconclusion They argued that activity in the ventral path-way reflects stored information about an object categorynot just physical features They point out that thisarrangement could explain why lesion patients with cate-gory specific perceptual deficits also have trouble retriev-ing general information about that visual category Thuswe would add that the FFA must store general infor-mation about people or some meta-representation oflsquopeoplenessrsquo
(c) Implications for the pathobiology of autismThe SAT used in this current study appears to be an
effective neurobehavioural probe for engaging a distrib-uted network of brain regions involved in different aspectsof social perception and cognition It will be important touse this and similar procedures in persons with autism tobetter define the nature of brain functions in this disorderCastelli et al (2002) have already taken the first step inthis process and they describe a pattern of hypoactivationin the MPFC STS and TPs in autism In an ongoingstudy of autism spectrum disorders we have presentedpreliminary data using the SAT showing that we too findhypoactivation of these regions and in the amygdala andFG (Schultz et al 2001) Thus we predict that futurework in this area will show that the entire social brain net-work is underactive in autism during tasks requiring socialperceptual and social cognitive processing
There are already sufficient data to argue strongly for arole of the FG in the pathobiology of autism No less thanfive previous fMRI studies have shown the FFA region tobe significantly less engaged among persons with autismcompared with controls during face perception tasks(Critchley et al 2000 Dierks et al 2001 Pierce et al2001 Schultz et al 2000a 2001) Activation of the FGby the social judgements in the current study adds animportant piece of evidence in favour of a causal role forthe FG in the pathobiology of autism This conclusionhowever must remain tentative until additional studiesmore precisely define the factors leading to FG activationduring social attribution and gather better data to proveits computational role in social cognition In addition theeffect of FG lesions for social functioning must be clari-fied If the FG is involved in social computations onewould expect to find social cognitive deficits in personswith lesions to this area However social deficits in proso-pagnosics have not been widely reported It might be thatprosopagnosic patients have not been carefully tested onthis dimension In this regard using our behavioural ver-sion of the SAT (Klin 2000) to test social perceptionamong prosopagnosics would be quite interesting Theparallels however between autism and prosopagnosia are
Phil Trans R Soc Lond B (2003)
incomplete because autism is clearly a developmental dis-order whereas the research literature on prosopagnosia ismostly confined to lesions acquired in adolescence oradulthood Among the few cases of developmental proso-pagnosia reported in the literature there is indeed one thathighlights severe social impairments (Kracke 1994) Itmay be that the role of the FG in the development ofautistic symptoms is different from the role of the FG inthe maintenance of social cognitive functions after brainmaturity
It is also possible that the role of the FG in social pro-cesses is dependent on its functional relationships withother nodes in the social brain and that it is the collectiveaction and interaction of the network that is of primaryimportance for social behaviours For example we founda strong correlation across participants in the amount ofFG and amygdala activation A strictly modular view ofthese areas may be inappropriate as the functions of eachnode could be quite dependent on one another and whenonly considered in isolation quite insufficient to supportsocial processes Thus whereas a lesion to the FG mayimpair visual perception it might not have a large impacton social functioning if this depends on an extended net-work that is dynamic and capable of compensatory adjust-ments Dysfunction of the FG may be necessary but notsufficient to produce social deficits Indeed our relianceon modular models of brain functioning may be leadingus astray in our search for causal mechanisms in autismInstead it may be the collective action of a distributedsystem that is critical to the pathobiology of autism TheFG region may be a key partner in this distributed systembut nevertheless just one node and insufficient by itselfto support in any substantive manner social cognitive pro-cesses or to explain social cognitive deficits seen inautism
5 CONCLUSION AND FUTURE DIRECTIONS
Using the SAT we isolated a distributed network ofactivations that conform to the emerging model of thesocial brain Most important we found significant acti-vation of the central aspect of the FG thus adding thisregion to the expanding list of structures involved in socialprocesses The FG region activated by the SAT overlapsin its spatial extent with the FFA with centres of massdiffering by less than 1 voxel We speculate that these SATactivations represent computational processes associatedwith more abstract attributes of people
It is an open question as to whether the substantial over-lap between the FFA and SAT FG activation is due to anoverlapping or shared set of neuronal assemblies This willneed to be clarified by future work Electrophysiologicalstudies show that there are small face-specific patches inthe FG cortex (Allison et al 1999) In the current studysmall patches of face cells and SAT cells could be inter-mingled but distinct and not drawing on any of the sameneuronal assemblies The spatial resolution of fMRI haslimitations that may preclude any definitive answer to thisquestion but there are strategies that can be used toaddress the issue For example the current study designdid not contain SAT and face discriminations in the sameexperimental fMRI series Doing so would enable directcomparisons of the computational demands placed on the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 425
common area of the FFA It also would be informative iffollow-up studies superimposed faces within the geometricfigures of the SAT without changing the film scripts inany way We could then determine if the computations ofthe common area of the FFA increase in a predictablefashionmdashwould the activation be a linear summation ofthe original SAT plus face discrimination Any significantdeviation from an additive model would suggest that thereis some sharing of neuronal assemblies with the SAT andface discrimination tasks when presented alone
This work was supported by grants from the National Institutesof Child Health and Human Development (grants PO1 HD03008 and PO1 HDDC35482) and the Korczak FoundationThe authors thank the anonymous reviewers for their helpfulcomments Michael Lee for programming the fMRI tasksHedy Serofin and Terry Hickey for their assistance in acquiringthe fMRI data John Herrington for his valuable assistance indata analyses of preliminary studies and members of the YaleDevelopmental Neuroimaging Program for their helpful com-ments on this manuscript A special acknowledgement ofappreciation is given to Donald Cohen who passed away inOctober of 2001 for his outstanding mentorship in all aspectsof this program of work
REFERENCES
Abell F Happe F amp Frith U 2000 Do triangles play tricksAttribution of mental states to animated shapes in normaland abnormal development J Cogn Dev 15 1ndash20
Adolphs R Tranel D amp Damasio A R 1998 The humanamygdala in social judgement Nature 393 470ndash474
Allison T Puce A Spencer D D amp McCarthy G 1999Electrophysiological studies of human face perception Ipotentials generated in occipitotemporal cortex by face andnon-face stimuli Cerebr Cortex 9 415ndash430
Allison T Puce A amp McCarthy G 2000 Social perceptionfrom visual cues role of the STS region Trends Cogn Sci4 267ndash278
Amaral D G amp Price J L 1984 Amygdalo-cortical projec-tions in the monkey (Macaca fascicularis) J Comp Neurol230 465ndash496
Amaral D G Price J L Pitkanen A amp Carmichael S T1992 Anatomical organization of the primate amygdaloidcomplex In The amygdala neurobiological aspects of emotionmemory and mental dysfunction (ed J Aggleton) pp 1ndash66New York Wiley-Liss
Anderson A K amp Phelps E A 2001 Lesions of the humanamygdala impair enhanced perception of emotionally salientevents Nature 411 305ndash309
Bachevalier J 1994 Medial temporal lobe structures andautism a review of clinical and experimental findingsNeuropsychologia 32 627ndash648
Bachevalier J amp Mishkin M 1986 Visual recognition impair-ment follows ventromedial but not dorsolateral prefrontallesions in monkeys Behav Brain Res 20 249ndash261
Baron-Cohen S Ring H A Wheelwright S BullmoreE T Brammer M J Simmons A amp Williams S C R1999 Social intelligence in the normal and autistic brain anfMRI study Eur J Neurosci 11 1891ndash1898
Baron-Cohen S Ring H A Bullmore E T WheelwrightS Ashwina C amp Williams S C R 2000 The amygdalatheory of autism Neurosci Biobehav Rev 24 355ndash364
Bechara A Tranel D Damasio H amp Damasio A R 1996Failure to respond autonomically to anticipated future out-comes following damage to prefrontal cortex Cerebr Cortex6 215ndash225
Benton A 1994 Face recognition Los Angeles CA WesternPsychological Services
Phil Trans R Soc Lond B (2003)
Bonda E Petrides M Ostry D amp Evans A 1996 Specificinvolvement of human parietal systems and the amygdala inthe perception of biological motion J Neurosci 16 3737ndash3744
Bowler D M amp Thommen E 2000 Attribution of mechan-ical and social causality to animated displays by childrenwith autism Autism 4 147ndash171
Brothers L 1990 The social brain a project for integratingprimate behavior and neurophysiology in a new domainConcepts Neurosci 1 27ndash151
Brothers L 1995 Neurophysiology of the perception of inten-tion by primates In The cognitive neurosciences 2nd edn (edM S Gazzaniga) pp 1107ndash1115 Cambridge MA MITPress
Butter C M McDonald J A amp Snyder D R 1969 Oralitypreference behavior and reinforcement value of nonfoodobject in monkeys with orbital frontal lesions Science 1641306ndash1307
Carmichael S T amp Price J L 1995 Limbic connections ofthe orbital and medial prefrontal cortex in macaque monk-eys J Comp Neurol 363 615ndash641
Castelli F Happe F Frith U amp Frith C 2000 Movementand mind a functional imaging study of perception andinterpretation of complex intentional movement patternsNeuroImage 12 314ndash325
Castelli F Frith C Happe F amp Frith U 2002 AutismAsperger syndrome and brain mechanisms for the attri-bution of mental states to animated shapes Brain 1251839ndash1849
Chao L L Haxby J V amp Martin A 1999 Attribute-basedneural substrates in posterior temporal cortex for perceivingand knowing about objects Nat Neurosci 2 913ndash919
Chao L L Weisberg J amp Martin A 2002 Experience-dependent modulation of category-related cortical activityCerebr Cortex 12 545ndash551
Critchley H D (and 10 others) 2000 The functional neuroan-atomy of social behaviour changes in cerebral blood flowwhen people with autistic disorder process facial expressionsBrain 123 2203ndash2212
Culham J C Cavanagh P amp Kanwisher N G 2001 Atten-tion response functions characterizing brain areas usingfMRI activation during parametric variations of attentionalload Neuron 32 737ndash745
Damasio A R Tranel D amp Damasio H 1990 Individualswith sociopathic behavior caused by frontal damage fail torespond autonomically to social stimuli Behav Brain Res41 81ndash94
Dierks T Bolte S Huble D Lanfermannm H amp PoustkaF 2001 Alterations of face processing strategies in autism(a fMRI study) Presented at the 6th Annual Meeting of theOrganization for Human Brain Mapping Brighton UK 10ndash14 June 2001
Dias R Robbins T W amp Roberts A C 1996 Dissociationin prefrontal cortex of affective and attentional shifts Nature380 69ndash72
Downing P E Jiang Y Shuman M amp Kanwisher N 2001A cortical area selective for visual processing of the humanbody Science 293 2470ndash2473
Epstein R amp Kanwisher N 1998 A cortical representation ofthe local visual environment Nature 392 598ndash601
Ernst M Zametkin A J Matochik J A Pascualvaca D ampCohen R M 1997 Reduced medial prefrontal dopami-nergic activity in autistic children Lancet 350 638
Fletcher P C Happe F Frith U Baker S C DolanR J Frackowiak R S amp Frith C D 1995 Other minds inthe brain a functional imaging study of lsquotheory of mindrsquo instory comprehension Cognition 57 109ndash128
Frith C D amp Frith U 1999 Interacting minds a biologicalbasis Science 286 1692ndash1695
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
426 R T Schultz and others FFA and social cognition
Fujita I Tanaka K Ito M amp Cheng K 1992 Columns forvisual features of objects in monkey inferotemporal cortexNature 360 343ndash346
Gaffan E A Gaffan D amp Harrison S 1988 Disconnectionof the amygdala from visual association cortex impairs vis-ual-reward association learning in monkeys J Neurosci 83144ndash3150
Gallagher H L Happe F Brunswick N Fletcher P CFrith U amp Frith C D 2000 Reading the mind in cartoonsand stories an fMRI study of lsquotheory of mindrsquo in verbal andnonverbal tasks Neuropsychologia 38 11ndash21
Gauthier I Anderson A W Tarr M J Skudlarski P ampGore J C 1997 Levels of categorization in visual recog-nition studied using functional magnetic resonance imagingCurr Biol 7 645ndash651
Gauthier I Tarr M J Anderson A W Skudlarski P ampGore J C 1999 Activation of the middle fusiform face areaincreases with expertise in recognizing novel objects NatureNeurosci 2 568ndash573
Gauthier I Skudlarski P Gore J C amp Anderson A W2000 Expertise for cars and birds recruits brain areasinvolved in face recognition Nature Neurosci 3 191ndash197
Goel V Grafman J Sadato N amp Hallett M 1995 Mode-ling other minds Neuroreport 6 1741ndash1746
Grelotti D Gauthier I amp Schultz R T 2001 Social interestand the development of cortical face specialization whatautism teaches us about face processing Devl Psychobiol 40213ndash225
Grezes J Fonlupt P Bertenthal B amp Delon-Martin C2001 Does perception of biological motion rely on specificbrain regions NeuroImage 13 775ndash785
Gusnard D A Akbudak E Shulman G L amp RaichleM E 2001 Medial prefrontal cortex and self-referentialmental activity relation to a default mode of brain functionProc Natl Acad Sci USA 98 4259ndash4264
Happe F Ehlers S Fletcher P Frith U Johansson MGillberg C Dolan R Frackowiak R amp Frith C 1996lsquoTheory of mindrsquo in the brain Evidence from a PET scanstudy of Asperger syndrome Neuroreport 8 197ndash201
Haxby J V Ungerleider L G Clark V P Schouten J LHoffman E A amp Martin A 1999 The effect of face inver-sion on activity in human neural systems for face and objectperception Neuron 22 189ndash199
Haxby J V Isahai A Chao L L Ungerleider L G ampMartin A 2000 Object-form topology in the ventral tem-poral lobe response to I Gauthier (2000) Trends Cogn Sci4 3ndash4
Haxby J V Gobbini M I Furey M L Ishai ASchouten J L amp Pietrini P 2001 Distributed and overlap-ping representations of faces and objects in ventral temporalcortex Science 293 2425ndash2430
Haznedar M M Buchsbaum M S Metzger M Solim-ando A Spiegel-Cohen J amp Hollander E 1997 Anteriorcingulate gyrus volume and glucose metabolism in autisticdisorder Am J Psychiat 154 1047ndash1050
Heider F amp Simmel M 1944 An experimental study ofapparent behavior Am J Psychol 57 243ndash259
Hobson R P Ouston J amp Lee A 1988a Whatrsquos in a faceThe case of autism Br J Psychol 79 441ndash453
Hobson R P Ouston J amp Lee A 1988b Emotion recog-nition in autism coordinating faces and voices Psychol Med18 911ndash923
Hoffman E A amp Haxby J V 2000 Distinct representationsof eye gaze and identity in the distributed human neural sys-tem for face perception Nature Neurosci 3 80ndash84
Hornak J Rolls E T amp Wade D 1996 Face and voiceexpression identification in patients with emotional andbehavioural changes following ventral frontal lobe damageNeuropsychologia 34 247ndash261
Phil Trans R Soc Lond B (2003)
Ishai A Ungerleider L G Martin A Schouten J L ampHaxby J V 1999 Distributed representation of objects inthe human ventral visual pathway Proc Natl Acad Sci USA96 9379ndash9384
Kanwisher N 2000 Domain specificity in face perceptionNature Neurosci 3 759ndash763
Kanwisher N McDermott J amp Chun M M 1997 The fusi-form face area a module of extrastriate cortex specializedfor face perception J Neurosci 17 4302ndash4311
Klin A 2000 Attributing social meaning to ambiguous visualstimuli in higher functioning autism and Asperger syndromethe social attribution task J Child Psychol Psychiat 41831ndash846
Klin A Sparrow S S de Bildt A Cicchetti D V CohenD J amp Volkmar F R 1999 A normed study of face recog-nition in autism and related disorders J Autism Devl Dis-orders 29 499ndash508
Klin A Jones W Schultz R T Volkmar F amp Cohen D J2002 Visual fixation patterns during viewing of naturalisticsocial situations as predictors of social competence in indi-viduals with autism Arch Gen Psychiat 59 809ndash816
Kracke I 1994 Developmental prosopagnosia in Aspergersyndrome presentation and discussion of an individual caseDevl Med Child Neurol 36 873ndash886
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J 1997 Neuroanatomical correlates of happi-ness sadness and disgust Am J Psychiat 154 926ndash933
Langdell T 1978 Recognition of faces an approach for thestudy of autism J Child Psychol Psychiat 19 255ndash268
LeDoux J E 1996 The emotional brain New York Simon ampShuster
Lowel S amp Singer W 1992 Selection of intrinsic horizontalconnections in the visual cortex by correlated neuronalactivity Science 255 209ndash212
Macdonald H Rutter M Howlin P Rios P Le ConteurA Evered C amp Folstein S 1989 Recognition andexpression of emotional cues by autistic and normal adultsJ Child Psychol Psychiat 30 865ndash877
Morris J S Ohman A amp Dolan R J 1998 Conscious andunconscious emotional learning in the human amygdalaNature 393 467ndash470
Myers R E Swett C amp Miller M 1973 Loss of social groupaffinity following prefrontal lesions in free-rangingmacaques Brain Res 64 257ndash269
Osterling J amp Dawson G 1994 Early recognition of childrenwith autism a study of first birthday home video tapes JAutism Devl Disorders 24 247ndash257
Pierce K Muller R A Ambrose J Allen G amp Cour-chesne E 2001 Face processing occurs outside the fusiformlsquoface arearsquo in autism evidence from functional MRI Brain124 2059ndash2073
Price J L Carmichael S T amp Drevets W C 1996 Net-works related to the orbital and medial prefrontal cortex asubstrate for emotional behavior Prog Brain Res 107523ndash536
Puce A Allison T Gore J C amp McCarthy G 1995 Face-sensitive regions in human extrastriate cortex studied byfunctional MRI J Neurophysiol 74 1192ndash1199
Reiman E M Lane R D Ahern G L Schwartz G EDavidson R J Friston K J Yun L S amp Chen K 1997Neuroanatomical correlates of externally and internally gen-erated human emotion Am J Psychiat 154 918ndash925
Rolls E T 1995 A theory of emotion and consciousness andits application to understanding the neural basis of emotionIn The cognitive neurosciences (ed M S Gazzaniga) pp1091ndash1106 Cambridge MA MIT Press
Schultz R T Gauthier I Klin A Fulbright R K Ander-son A W Volkmar F Skudlarski P Lacadie C CohenD J amp Gore J C 2000a Abnormal ventral temporal
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from

422 R T Schultz and others FFA and social cognition
Gusnard et al (2001) suggest that the dorsal MPFC isinvolved in any kind of thought that uses the self as a refer-ent Thus the SAT activations in this area may have beendriven by theorizing about othersrsquo minds but with explicitreference to the participantrsquos own frame of reference as tohow they would feel in a similar situation
The orbital prefrontal cortex and to a lesser extent thedorsal MPFC have dense reciprocal connections withmedial temporal areas (Price et al 1996 Carmichael ampPrice 1995) providing the anatomical bases for a systemthat regulates emotional processes Damasio and col-leagues (Damasio et al 1990 Bechara et al 1996) haveargued that the orbito-MPFC have a primary function ofintegrating information about rewards and punishmentsto bias future behaviour (Rolls 1995 Dias et al 1996Hornak et al 1996 Lane et al 1997 Reiman et al 1997)A functional circuitry such as this would seem especiallyimportant in the development and acquisition of socialbehaviour However even acquired lesions to theseregions in adulthood can result in abnormalities of socialconduct (Damasio et al 1990) Brothers (1990) high-lighted the orbital prefrontal cortex as one of the threeprincipal brain regions involved in social cognition Weobserved poor signal in this region (these areas are notori-ously prone to fMRI signal drop-out and distortion) andthus we cannot know if the orbital prefrontal region wasengaged by the SAT or not However using a similarpsychological task but with PET where signal acquisitionin this region is not degraded Castelli et al (2000 2002)failed to find activation of the orbital prefrontal cortexCollectively these results call into question the impor-tance of the orbital prefrontal cortex in social cognitionand instead shift the focus on anterior cortices toward thedorsal MPFC
The SAT also generated robust activations of the rightamygdala and nearby cortex of the right TP The amyg-dala is often given a central role in theories of social per-ception and cognition (Brothers 1990 Bachevalier 1994Adolphs et al 1998 Baron-Cohen et al 2000 Schultz etal 2000b) The amygdala has a critical role in emotionalarousal assigning significance to environmental stimuliand mediating the formation of visual-reward associationsthat is lsquoemotionalrsquo learning (Gaffan et al 1988 LeDoux1996 Anderson amp Phelps 2001) It is reliably engagedduring judgements of personality characteristics from pic-tures of the face or part of the face (Adolphs et al 1998Baron-Cohen et al 1999 Winston et al 2002) Activationof the amygdala appears to be automatic and stimulusdriven as it can be engaged by images of facial expressionsin conscious awareness as well as by subliminal presen-tations of faces displaying affect (Morris et al 1998Whalen et al 1998 Critchley et al 2000) Thus theamygdalarsquos engagement by the SAT could stem from thegeneral emotional arousal evoked by the animations or itcould represent its computational role in some more spe-cific social perceptual process
The amygdala has dense reciprocal connections withthe ventral visual processing stream (Amaral amp Price1984) The strong correlation observed in this studybetween the right amygdala and the right FG could indi-cate that emotional inputs from the amygdala to the FGare necessary for engaging the social computational pro-cesses of the FG Brothers (1995) has speculated that the
Phil Trans R Soc Lond B (2003)
amygdala generates lsquosocial feelingsrsquo that are of importancein cutting through the complexities of social situations andguiding behaviour by simpler learned contingenciesSocial events are complex because their meaning oftencomes from specific combinations of features that do notadd up in a linear manner making it more difficult todecompose the events by cognitive analysis Effectivesocial engagement requires integration of context historicalrelationships and current socialndashemotional communi-cations expressed through prosodic tone as well as facialexpressions posture and gesture They are made morecomplex by the rapid pace of social transactions Thiswould necessitate some fast response system based ongeneral principles from past socialndashemotional experiencesa role for which the amygdala would seem ideally suitedIn other words the amygdala might drive intuition or lsquogutfeelingsrsquo to guide rapid non-verbal social interactionsinvolving facial expressions gesture etc Thus the strongamygdalandashFG correlation observed here could be inter-preted as the amygdala informing the FG of the relevanceof a social event and also of the outcome of its quick anddirty social perceptual analyses thereby guiding the FG(and other social nodes) in their finer grained compu-tations
Perhaps the largest and strongest activations to the SATwere those of the posterior aspects of the STS spreadinginto the adjacent STG This area has been implicated asa specific site for perception of biological motion (Bondaet al 1996 Allison et al 2000 Grezes et al 2001 Vainaet al 2001) Biological motion is a broad construct thatseems to encompass the perception of static images ofevents that could move or did move such as facialexpressions as well as actual movement by animateobjects This region of the STSndashSTG is also critical forthe decoding visual displays of social action or intention(eg gaze direction gesture and facial displays of emotion)(Critchley et al 2000 Hoffman amp Haxby 2000) In thisregard the SAT activations here are completely expectedand a testament to the effectiveness of these animationsin inducing the desired illusion of anthropomorphicaction The majority of the STSndashSTG activations wereanterior to location of the V5MT as defined in otherrecent fMRI studies (Culham et al 2001) but they alsoextended posteriorly into these more general movementsensitive cortices Unlike the amygdala some evidencesuggests that activity of the STS is mediated by explicitattention to social characteristics of the face and that STSengagement it is not automatic or stimulus driven(Winston et al 2002) We surmise therefore that acti-vation of the STS by the SAT was due to explicit taskinstructions to judge the social interactions
(b) Role of the fusiform face area in socialcognition
We also found significant SAT activation of the rightFG When compared directly in a subset of participants totheir FFA gleaned from a separate scanning session witha perceptual discrimination task of person versus objectidentity we found that both ROIs were confined to theright side on the same two coronal slices There wasabout 50 overlap between the SAT FG and FFA acti-vations and the centre of the two activations differed byless than 1 voxel The FFA activation was offset to the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 423
lateral prominence of the FG whereas the SAT activateda region of the FG that was closer to the centre of the FGbut still largely in the lateral aspect Because the SAT doesnot contain any face representations the FFA engagementwas unexpected as this region is thought of as selectivefor faces (Kanwisher et al 1997) or to other classes ofcomplex objects for which one is perceptually expert(Gauthier et al 2000)
These results have important implications for the poss-ible role of the fusiform in social brain circuitry andautism Interestingly the magnitude of the FG activationpredicted SAT task accuracy no other ROI correlated sig-nificantly with task accuracy This would seem to argueagainst any interpretation that the FFA activation is incon-sequential to the SAT that it is activated simply becauseit is well connected to other areas that are directly involvedin the social attribution process Using a task that is simi-lar to the SAT Castelli et al (2000) also reported signifi-cant right FG activation using PET However in theirfollow-up PET study comparing autism and normal con-trols (Castelli et al 2002) they failed to find significantFG activation in either group The failure of their secondstudy to find right FG activations may be due to the morestringent random effects model used in the data analysesHowever because we find right FG activation with fMRIwhich is more sensitive than PET and because Castelli etal (2000) found it with what amounts to a lower thres-hold it seems quite probable that the effect is real In factwe have preliminary evidence from an ongoing fMRIstudy that reproduces the SAT FG activations in healthycontrols (Schultz et al 2001) In addition there may besome task attributes that differ between our studies andCastelli et alrsquos that might impact into the strength withwhich FG computational processes are evoked
The key question then is why is the FG engaged bythe SAT and what role does the right FG have in socialcognition and perception Currently there are three com-peting theories of the functional organization of the FGand related ventral visual perceptual areas One modelput forth by Haxby Chao Martin and colleagues speci-fies that objects are encoded in a distributed fashion acrossa wide expanse of the ventral temporal-occipital cortex(Haxby et al 2001) They call this the lsquoobject formtypologyrsquo model (Ishai et al 1999) Their data indicatethat object category specificity is achieved by unique spa-tial patterns of activation across this extrastriate visual cor-tex They show that the pattern of activations across thiscortex is diagnostic of object category membership moreso than any localized activation maxima (Haxby et al2001) They also argue that object category perceptioninvolves retrieval of category-related information aboutspecific features and attributes of the object category(Chao et al 2002) They admit that the lsquonature of theinformation about objects that is represented in the ventraltemporal cortex is a great puzzlersquo (Haxby et al 2000 p4) but open the possibility that semantic information maybe important
A second view championed by Kanwisher et al holdsthat several select perceptual categories including facesbodies and spatial layout of places are encoded in highlyspecific locations in a modular fashion (Kanwisher et al1997 Epstein amp Kanwisher 1998 Downing et al 2001Spiridon amp Kanwisher 2002) The FFA is thought to be
Phil Trans R Soc Lond B (2003)
most concerned with discriminating among individualidentities and not with discriminating images at the categ-orical level In fact lesions to the FFA cause a specificdeficit in recognizing individual identities but not in reco-gnizing the general category of face versus non-face(Wada amp Yamamoto 2001) Spiridon amp Kanwisher (2002)provide a partial replication of Haxby et alrsquos (2001) objectform typology model They show that patterns of ventralvisual cortical activation can distinguish object categoriesNevertheless their data show category specificity for selectareas such as the FFA and they argue the ventral visualcortex is not equipotential
Gauthier et al however have argued that the ventraloccipital temporal pathway is organized by the nature ofthe perceptual computations and that these processingbiases are acquired through experience (Gauthier et al2000 Tarr amp Gauthier 2000) For example the parahip-pocampal place area is a function of a bias towards pro-cessing landscapes in terms of their spatial layout becausewe have learned through repeated experiences that this isvery useful information to extract when perceiving land-scapes and related visual images Similarly we learn quiteearly in life that it is important to discriminate faces on anindividual level for example discriminating mother fromothers This bias towards individual identification accord-ing to Gauthier accounts for the FFArsquos apparent modu-larity (Gauthier et al 1997 2000 Tarr amp Gauthier 2000)According to this third model the type of informationneeded and our cumulative experience in processing thatinformation organizes the ventral occipitalndashtemporal path-way into regional centres with preferred modes of pro-cessing
All three models seem to agree that the functionalorganization of the ventral visual cortices is driven by theneed to categorize perceptions into object classes Webelieve that our findings showing FFA activation by non-face objects are consistent with aspects of both the pro-cessing map model of Gauthier and the object formtypology model of Haxby et al First consistent withHaxby et al we suggest that semantic information isimportant to the ventral visual pathway for object categor-ization Second consistent with Gauthier et al we believethat repeated perceptual experience with faces biases thetype of information that FG finds important We proposethat the middle FG area encodes semantic attributes ofpeople because of repeated perceptual experiences withfaces that occur during social interactions In fact wewould guess that most perceptual experiences with facesoccur during social situations and these social situationsoften involve repeated social judgements Thus infor-mation about the social nature of people might be storedin the FG (though not exclusively)
The nature of the semantic information stored in theFG might be restricted to anything that would be helpfulin defining faces as a distinct category of object becausemaking such distinctions appears to be the primary chargeof the ventral visual pathway This would include knowl-edge of people as having personal agency and of havingthe capability to disturb each otherrsquos emotional homeo-stasis (eg to act friendly or unfriendly) By pretendingthat the three shapes in the SAT are people stored knowl-edge about people in social interactions might beretrieved causing the observed activations in the FG
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
424 R T Schultz and others FFA and social cognition
There would be a measure of efficiency from thisarrangement that is having representations and compu-tations of more abstract attributes of people inter-digitatedwith front-end perceptual processes about physical attri-butes Gauthierrsquos claim is that the same group of neuronscan be engaged by perceptually different categories ofobjects it is the type of processing rather than the visualdetails that are important Extrapolating from this wewould argue that the SAT engages a region of the FGoverlapping with the FFA because it demands compu-tational processes to classify the SAT geometric figures aspeople or person-like Chao et al (1999) reach a similarconclusion They argued that activity in the ventral path-way reflects stored information about an object categorynot just physical features They point out that thisarrangement could explain why lesion patients with cate-gory specific perceptual deficits also have trouble retriev-ing general information about that visual category Thuswe would add that the FFA must store general infor-mation about people or some meta-representation oflsquopeoplenessrsquo
(c) Implications for the pathobiology of autismThe SAT used in this current study appears to be an
effective neurobehavioural probe for engaging a distrib-uted network of brain regions involved in different aspectsof social perception and cognition It will be important touse this and similar procedures in persons with autism tobetter define the nature of brain functions in this disorderCastelli et al (2002) have already taken the first step inthis process and they describe a pattern of hypoactivationin the MPFC STS and TPs in autism In an ongoingstudy of autism spectrum disorders we have presentedpreliminary data using the SAT showing that we too findhypoactivation of these regions and in the amygdala andFG (Schultz et al 2001) Thus we predict that futurework in this area will show that the entire social brain net-work is underactive in autism during tasks requiring socialperceptual and social cognitive processing
There are already sufficient data to argue strongly for arole of the FG in the pathobiology of autism No less thanfive previous fMRI studies have shown the FFA region tobe significantly less engaged among persons with autismcompared with controls during face perception tasks(Critchley et al 2000 Dierks et al 2001 Pierce et al2001 Schultz et al 2000a 2001) Activation of the FGby the social judgements in the current study adds animportant piece of evidence in favour of a causal role forthe FG in the pathobiology of autism This conclusionhowever must remain tentative until additional studiesmore precisely define the factors leading to FG activationduring social attribution and gather better data to proveits computational role in social cognition In addition theeffect of FG lesions for social functioning must be clari-fied If the FG is involved in social computations onewould expect to find social cognitive deficits in personswith lesions to this area However social deficits in proso-pagnosics have not been widely reported It might be thatprosopagnosic patients have not been carefully tested onthis dimension In this regard using our behavioural ver-sion of the SAT (Klin 2000) to test social perceptionamong prosopagnosics would be quite interesting Theparallels however between autism and prosopagnosia are
Phil Trans R Soc Lond B (2003)
incomplete because autism is clearly a developmental dis-order whereas the research literature on prosopagnosia ismostly confined to lesions acquired in adolescence oradulthood Among the few cases of developmental proso-pagnosia reported in the literature there is indeed one thathighlights severe social impairments (Kracke 1994) Itmay be that the role of the FG in the development ofautistic symptoms is different from the role of the FG inthe maintenance of social cognitive functions after brainmaturity
It is also possible that the role of the FG in social pro-cesses is dependent on its functional relationships withother nodes in the social brain and that it is the collectiveaction and interaction of the network that is of primaryimportance for social behaviours For example we founda strong correlation across participants in the amount ofFG and amygdala activation A strictly modular view ofthese areas may be inappropriate as the functions of eachnode could be quite dependent on one another and whenonly considered in isolation quite insufficient to supportsocial processes Thus whereas a lesion to the FG mayimpair visual perception it might not have a large impacton social functioning if this depends on an extended net-work that is dynamic and capable of compensatory adjust-ments Dysfunction of the FG may be necessary but notsufficient to produce social deficits Indeed our relianceon modular models of brain functioning may be leadingus astray in our search for causal mechanisms in autismInstead it may be the collective action of a distributedsystem that is critical to the pathobiology of autism TheFG region may be a key partner in this distributed systembut nevertheless just one node and insufficient by itselfto support in any substantive manner social cognitive pro-cesses or to explain social cognitive deficits seen inautism
5 CONCLUSION AND FUTURE DIRECTIONS
Using the SAT we isolated a distributed network ofactivations that conform to the emerging model of thesocial brain Most important we found significant acti-vation of the central aspect of the FG thus adding thisregion to the expanding list of structures involved in socialprocesses The FG region activated by the SAT overlapsin its spatial extent with the FFA with centres of massdiffering by less than 1 voxel We speculate that these SATactivations represent computational processes associatedwith more abstract attributes of people
It is an open question as to whether the substantial over-lap between the FFA and SAT FG activation is due to anoverlapping or shared set of neuronal assemblies This willneed to be clarified by future work Electrophysiologicalstudies show that there are small face-specific patches inthe FG cortex (Allison et al 1999) In the current studysmall patches of face cells and SAT cells could be inter-mingled but distinct and not drawing on any of the sameneuronal assemblies The spatial resolution of fMRI haslimitations that may preclude any definitive answer to thisquestion but there are strategies that can be used toaddress the issue For example the current study designdid not contain SAT and face discriminations in the sameexperimental fMRI series Doing so would enable directcomparisons of the computational demands placed on the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 425
common area of the FFA It also would be informative iffollow-up studies superimposed faces within the geometricfigures of the SAT without changing the film scripts inany way We could then determine if the computations ofthe common area of the FFA increase in a predictablefashionmdashwould the activation be a linear summation ofthe original SAT plus face discrimination Any significantdeviation from an additive model would suggest that thereis some sharing of neuronal assemblies with the SAT andface discrimination tasks when presented alone
This work was supported by grants from the National Institutesof Child Health and Human Development (grants PO1 HD03008 and PO1 HDDC35482) and the Korczak FoundationThe authors thank the anonymous reviewers for their helpfulcomments Michael Lee for programming the fMRI tasksHedy Serofin and Terry Hickey for their assistance in acquiringthe fMRI data John Herrington for his valuable assistance indata analyses of preliminary studies and members of the YaleDevelopmental Neuroimaging Program for their helpful com-ments on this manuscript A special acknowledgement ofappreciation is given to Donald Cohen who passed away inOctober of 2001 for his outstanding mentorship in all aspectsof this program of work
REFERENCES
Abell F Happe F amp Frith U 2000 Do triangles play tricksAttribution of mental states to animated shapes in normaland abnormal development J Cogn Dev 15 1ndash20
Adolphs R Tranel D amp Damasio A R 1998 The humanamygdala in social judgement Nature 393 470ndash474
Allison T Puce A Spencer D D amp McCarthy G 1999Electrophysiological studies of human face perception Ipotentials generated in occipitotemporal cortex by face andnon-face stimuli Cerebr Cortex 9 415ndash430
Allison T Puce A amp McCarthy G 2000 Social perceptionfrom visual cues role of the STS region Trends Cogn Sci4 267ndash278
Amaral D G amp Price J L 1984 Amygdalo-cortical projec-tions in the monkey (Macaca fascicularis) J Comp Neurol230 465ndash496
Amaral D G Price J L Pitkanen A amp Carmichael S T1992 Anatomical organization of the primate amygdaloidcomplex In The amygdala neurobiological aspects of emotionmemory and mental dysfunction (ed J Aggleton) pp 1ndash66New York Wiley-Liss
Anderson A K amp Phelps E A 2001 Lesions of the humanamygdala impair enhanced perception of emotionally salientevents Nature 411 305ndash309
Bachevalier J 1994 Medial temporal lobe structures andautism a review of clinical and experimental findingsNeuropsychologia 32 627ndash648
Bachevalier J amp Mishkin M 1986 Visual recognition impair-ment follows ventromedial but not dorsolateral prefrontallesions in monkeys Behav Brain Res 20 249ndash261
Baron-Cohen S Ring H A Wheelwright S BullmoreE T Brammer M J Simmons A amp Williams S C R1999 Social intelligence in the normal and autistic brain anfMRI study Eur J Neurosci 11 1891ndash1898
Baron-Cohen S Ring H A Bullmore E T WheelwrightS Ashwina C amp Williams S C R 2000 The amygdalatheory of autism Neurosci Biobehav Rev 24 355ndash364
Bechara A Tranel D Damasio H amp Damasio A R 1996Failure to respond autonomically to anticipated future out-comes following damage to prefrontal cortex Cerebr Cortex6 215ndash225
Benton A 1994 Face recognition Los Angeles CA WesternPsychological Services
Phil Trans R Soc Lond B (2003)
Bonda E Petrides M Ostry D amp Evans A 1996 Specificinvolvement of human parietal systems and the amygdala inthe perception of biological motion J Neurosci 16 3737ndash3744
Bowler D M amp Thommen E 2000 Attribution of mechan-ical and social causality to animated displays by childrenwith autism Autism 4 147ndash171
Brothers L 1990 The social brain a project for integratingprimate behavior and neurophysiology in a new domainConcepts Neurosci 1 27ndash151
Brothers L 1995 Neurophysiology of the perception of inten-tion by primates In The cognitive neurosciences 2nd edn (edM S Gazzaniga) pp 1107ndash1115 Cambridge MA MITPress
Butter C M McDonald J A amp Snyder D R 1969 Oralitypreference behavior and reinforcement value of nonfoodobject in monkeys with orbital frontal lesions Science 1641306ndash1307
Carmichael S T amp Price J L 1995 Limbic connections ofthe orbital and medial prefrontal cortex in macaque monk-eys J Comp Neurol 363 615ndash641
Castelli F Happe F Frith U amp Frith C 2000 Movementand mind a functional imaging study of perception andinterpretation of complex intentional movement patternsNeuroImage 12 314ndash325
Castelli F Frith C Happe F amp Frith U 2002 AutismAsperger syndrome and brain mechanisms for the attri-bution of mental states to animated shapes Brain 1251839ndash1849
Chao L L Haxby J V amp Martin A 1999 Attribute-basedneural substrates in posterior temporal cortex for perceivingand knowing about objects Nat Neurosci 2 913ndash919
Chao L L Weisberg J amp Martin A 2002 Experience-dependent modulation of category-related cortical activityCerebr Cortex 12 545ndash551
Critchley H D (and 10 others) 2000 The functional neuroan-atomy of social behaviour changes in cerebral blood flowwhen people with autistic disorder process facial expressionsBrain 123 2203ndash2212
Culham J C Cavanagh P amp Kanwisher N G 2001 Atten-tion response functions characterizing brain areas usingfMRI activation during parametric variations of attentionalload Neuron 32 737ndash745
Damasio A R Tranel D amp Damasio H 1990 Individualswith sociopathic behavior caused by frontal damage fail torespond autonomically to social stimuli Behav Brain Res41 81ndash94
Dierks T Bolte S Huble D Lanfermannm H amp PoustkaF 2001 Alterations of face processing strategies in autism(a fMRI study) Presented at the 6th Annual Meeting of theOrganization for Human Brain Mapping Brighton UK 10ndash14 June 2001
Dias R Robbins T W amp Roberts A C 1996 Dissociationin prefrontal cortex of affective and attentional shifts Nature380 69ndash72
Downing P E Jiang Y Shuman M amp Kanwisher N 2001A cortical area selective for visual processing of the humanbody Science 293 2470ndash2473
Epstein R amp Kanwisher N 1998 A cortical representation ofthe local visual environment Nature 392 598ndash601
Ernst M Zametkin A J Matochik J A Pascualvaca D ampCohen R M 1997 Reduced medial prefrontal dopami-nergic activity in autistic children Lancet 350 638
Fletcher P C Happe F Frith U Baker S C DolanR J Frackowiak R S amp Frith C D 1995 Other minds inthe brain a functional imaging study of lsquotheory of mindrsquo instory comprehension Cognition 57 109ndash128
Frith C D amp Frith U 1999 Interacting minds a biologicalbasis Science 286 1692ndash1695
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
426 R T Schultz and others FFA and social cognition
Fujita I Tanaka K Ito M amp Cheng K 1992 Columns forvisual features of objects in monkey inferotemporal cortexNature 360 343ndash346
Gaffan E A Gaffan D amp Harrison S 1988 Disconnectionof the amygdala from visual association cortex impairs vis-ual-reward association learning in monkeys J Neurosci 83144ndash3150
Gallagher H L Happe F Brunswick N Fletcher P CFrith U amp Frith C D 2000 Reading the mind in cartoonsand stories an fMRI study of lsquotheory of mindrsquo in verbal andnonverbal tasks Neuropsychologia 38 11ndash21
Gauthier I Anderson A W Tarr M J Skudlarski P ampGore J C 1997 Levels of categorization in visual recog-nition studied using functional magnetic resonance imagingCurr Biol 7 645ndash651
Gauthier I Tarr M J Anderson A W Skudlarski P ampGore J C 1999 Activation of the middle fusiform face areaincreases with expertise in recognizing novel objects NatureNeurosci 2 568ndash573
Gauthier I Skudlarski P Gore J C amp Anderson A W2000 Expertise for cars and birds recruits brain areasinvolved in face recognition Nature Neurosci 3 191ndash197
Goel V Grafman J Sadato N amp Hallett M 1995 Mode-ling other minds Neuroreport 6 1741ndash1746
Grelotti D Gauthier I amp Schultz R T 2001 Social interestand the development of cortical face specialization whatautism teaches us about face processing Devl Psychobiol 40213ndash225
Grezes J Fonlupt P Bertenthal B amp Delon-Martin C2001 Does perception of biological motion rely on specificbrain regions NeuroImage 13 775ndash785
Gusnard D A Akbudak E Shulman G L amp RaichleM E 2001 Medial prefrontal cortex and self-referentialmental activity relation to a default mode of brain functionProc Natl Acad Sci USA 98 4259ndash4264
Happe F Ehlers S Fletcher P Frith U Johansson MGillberg C Dolan R Frackowiak R amp Frith C 1996lsquoTheory of mindrsquo in the brain Evidence from a PET scanstudy of Asperger syndrome Neuroreport 8 197ndash201
Haxby J V Ungerleider L G Clark V P Schouten J LHoffman E A amp Martin A 1999 The effect of face inver-sion on activity in human neural systems for face and objectperception Neuron 22 189ndash199
Haxby J V Isahai A Chao L L Ungerleider L G ampMartin A 2000 Object-form topology in the ventral tem-poral lobe response to I Gauthier (2000) Trends Cogn Sci4 3ndash4
Haxby J V Gobbini M I Furey M L Ishai ASchouten J L amp Pietrini P 2001 Distributed and overlap-ping representations of faces and objects in ventral temporalcortex Science 293 2425ndash2430
Haznedar M M Buchsbaum M S Metzger M Solim-ando A Spiegel-Cohen J amp Hollander E 1997 Anteriorcingulate gyrus volume and glucose metabolism in autisticdisorder Am J Psychiat 154 1047ndash1050
Heider F amp Simmel M 1944 An experimental study ofapparent behavior Am J Psychol 57 243ndash259
Hobson R P Ouston J amp Lee A 1988a Whatrsquos in a faceThe case of autism Br J Psychol 79 441ndash453
Hobson R P Ouston J amp Lee A 1988b Emotion recog-nition in autism coordinating faces and voices Psychol Med18 911ndash923
Hoffman E A amp Haxby J V 2000 Distinct representationsof eye gaze and identity in the distributed human neural sys-tem for face perception Nature Neurosci 3 80ndash84
Hornak J Rolls E T amp Wade D 1996 Face and voiceexpression identification in patients with emotional andbehavioural changes following ventral frontal lobe damageNeuropsychologia 34 247ndash261
Phil Trans R Soc Lond B (2003)
Ishai A Ungerleider L G Martin A Schouten J L ampHaxby J V 1999 Distributed representation of objects inthe human ventral visual pathway Proc Natl Acad Sci USA96 9379ndash9384
Kanwisher N 2000 Domain specificity in face perceptionNature Neurosci 3 759ndash763
Kanwisher N McDermott J amp Chun M M 1997 The fusi-form face area a module of extrastriate cortex specializedfor face perception J Neurosci 17 4302ndash4311
Klin A 2000 Attributing social meaning to ambiguous visualstimuli in higher functioning autism and Asperger syndromethe social attribution task J Child Psychol Psychiat 41831ndash846
Klin A Sparrow S S de Bildt A Cicchetti D V CohenD J amp Volkmar F R 1999 A normed study of face recog-nition in autism and related disorders J Autism Devl Dis-orders 29 499ndash508
Klin A Jones W Schultz R T Volkmar F amp Cohen D J2002 Visual fixation patterns during viewing of naturalisticsocial situations as predictors of social competence in indi-viduals with autism Arch Gen Psychiat 59 809ndash816
Kracke I 1994 Developmental prosopagnosia in Aspergersyndrome presentation and discussion of an individual caseDevl Med Child Neurol 36 873ndash886
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J 1997 Neuroanatomical correlates of happi-ness sadness and disgust Am J Psychiat 154 926ndash933
Langdell T 1978 Recognition of faces an approach for thestudy of autism J Child Psychol Psychiat 19 255ndash268
LeDoux J E 1996 The emotional brain New York Simon ampShuster
Lowel S amp Singer W 1992 Selection of intrinsic horizontalconnections in the visual cortex by correlated neuronalactivity Science 255 209ndash212
Macdonald H Rutter M Howlin P Rios P Le ConteurA Evered C amp Folstein S 1989 Recognition andexpression of emotional cues by autistic and normal adultsJ Child Psychol Psychiat 30 865ndash877
Morris J S Ohman A amp Dolan R J 1998 Conscious andunconscious emotional learning in the human amygdalaNature 393 467ndash470
Myers R E Swett C amp Miller M 1973 Loss of social groupaffinity following prefrontal lesions in free-rangingmacaques Brain Res 64 257ndash269
Osterling J amp Dawson G 1994 Early recognition of childrenwith autism a study of first birthday home video tapes JAutism Devl Disorders 24 247ndash257
Pierce K Muller R A Ambrose J Allen G amp Cour-chesne E 2001 Face processing occurs outside the fusiformlsquoface arearsquo in autism evidence from functional MRI Brain124 2059ndash2073
Price J L Carmichael S T amp Drevets W C 1996 Net-works related to the orbital and medial prefrontal cortex asubstrate for emotional behavior Prog Brain Res 107523ndash536
Puce A Allison T Gore J C amp McCarthy G 1995 Face-sensitive regions in human extrastriate cortex studied byfunctional MRI J Neurophysiol 74 1192ndash1199
Reiman E M Lane R D Ahern G L Schwartz G EDavidson R J Friston K J Yun L S amp Chen K 1997Neuroanatomical correlates of externally and internally gen-erated human emotion Am J Psychiat 154 918ndash925
Rolls E T 1995 A theory of emotion and consciousness andits application to understanding the neural basis of emotionIn The cognitive neurosciences (ed M S Gazzaniga) pp1091ndash1106 Cambridge MA MIT Press
Schultz R T Gauthier I Klin A Fulbright R K Ander-son A W Volkmar F Skudlarski P Lacadie C CohenD J amp Gore J C 2000a Abnormal ventral temporal
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from

FFA and social cognition R T Schultz and others 423
lateral prominence of the FG whereas the SAT activateda region of the FG that was closer to the centre of the FGbut still largely in the lateral aspect Because the SAT doesnot contain any face representations the FFA engagementwas unexpected as this region is thought of as selectivefor faces (Kanwisher et al 1997) or to other classes ofcomplex objects for which one is perceptually expert(Gauthier et al 2000)
These results have important implications for the poss-ible role of the fusiform in social brain circuitry andautism Interestingly the magnitude of the FG activationpredicted SAT task accuracy no other ROI correlated sig-nificantly with task accuracy This would seem to argueagainst any interpretation that the FFA activation is incon-sequential to the SAT that it is activated simply becauseit is well connected to other areas that are directly involvedin the social attribution process Using a task that is simi-lar to the SAT Castelli et al (2000) also reported signifi-cant right FG activation using PET However in theirfollow-up PET study comparing autism and normal con-trols (Castelli et al 2002) they failed to find significantFG activation in either group The failure of their secondstudy to find right FG activations may be due to the morestringent random effects model used in the data analysesHowever because we find right FG activation with fMRIwhich is more sensitive than PET and because Castelli etal (2000) found it with what amounts to a lower thres-hold it seems quite probable that the effect is real In factwe have preliminary evidence from an ongoing fMRIstudy that reproduces the SAT FG activations in healthycontrols (Schultz et al 2001) In addition there may besome task attributes that differ between our studies andCastelli et alrsquos that might impact into the strength withwhich FG computational processes are evoked
The key question then is why is the FG engaged bythe SAT and what role does the right FG have in socialcognition and perception Currently there are three com-peting theories of the functional organization of the FGand related ventral visual perceptual areas One modelput forth by Haxby Chao Martin and colleagues speci-fies that objects are encoded in a distributed fashion acrossa wide expanse of the ventral temporal-occipital cortex(Haxby et al 2001) They call this the lsquoobject formtypologyrsquo model (Ishai et al 1999) Their data indicatethat object category specificity is achieved by unique spa-tial patterns of activation across this extrastriate visual cor-tex They show that the pattern of activations across thiscortex is diagnostic of object category membership moreso than any localized activation maxima (Haxby et al2001) They also argue that object category perceptioninvolves retrieval of category-related information aboutspecific features and attributes of the object category(Chao et al 2002) They admit that the lsquonature of theinformation about objects that is represented in the ventraltemporal cortex is a great puzzlersquo (Haxby et al 2000 p4) but open the possibility that semantic information maybe important
A second view championed by Kanwisher et al holdsthat several select perceptual categories including facesbodies and spatial layout of places are encoded in highlyspecific locations in a modular fashion (Kanwisher et al1997 Epstein amp Kanwisher 1998 Downing et al 2001Spiridon amp Kanwisher 2002) The FFA is thought to be
Phil Trans R Soc Lond B (2003)
most concerned with discriminating among individualidentities and not with discriminating images at the categ-orical level In fact lesions to the FFA cause a specificdeficit in recognizing individual identities but not in reco-gnizing the general category of face versus non-face(Wada amp Yamamoto 2001) Spiridon amp Kanwisher (2002)provide a partial replication of Haxby et alrsquos (2001) objectform typology model They show that patterns of ventralvisual cortical activation can distinguish object categoriesNevertheless their data show category specificity for selectareas such as the FFA and they argue the ventral visualcortex is not equipotential
Gauthier et al however have argued that the ventraloccipital temporal pathway is organized by the nature ofthe perceptual computations and that these processingbiases are acquired through experience (Gauthier et al2000 Tarr amp Gauthier 2000) For example the parahip-pocampal place area is a function of a bias towards pro-cessing landscapes in terms of their spatial layout becausewe have learned through repeated experiences that this isvery useful information to extract when perceiving land-scapes and related visual images Similarly we learn quiteearly in life that it is important to discriminate faces on anindividual level for example discriminating mother fromothers This bias towards individual identification accord-ing to Gauthier accounts for the FFArsquos apparent modu-larity (Gauthier et al 1997 2000 Tarr amp Gauthier 2000)According to this third model the type of informationneeded and our cumulative experience in processing thatinformation organizes the ventral occipitalndashtemporal path-way into regional centres with preferred modes of pro-cessing
All three models seem to agree that the functionalorganization of the ventral visual cortices is driven by theneed to categorize perceptions into object classes Webelieve that our findings showing FFA activation by non-face objects are consistent with aspects of both the pro-cessing map model of Gauthier and the object formtypology model of Haxby et al First consistent withHaxby et al we suggest that semantic information isimportant to the ventral visual pathway for object categor-ization Second consistent with Gauthier et al we believethat repeated perceptual experience with faces biases thetype of information that FG finds important We proposethat the middle FG area encodes semantic attributes ofpeople because of repeated perceptual experiences withfaces that occur during social interactions In fact wewould guess that most perceptual experiences with facesoccur during social situations and these social situationsoften involve repeated social judgements Thus infor-mation about the social nature of people might be storedin the FG (though not exclusively)
The nature of the semantic information stored in theFG might be restricted to anything that would be helpfulin defining faces as a distinct category of object becausemaking such distinctions appears to be the primary chargeof the ventral visual pathway This would include knowl-edge of people as having personal agency and of havingthe capability to disturb each otherrsquos emotional homeo-stasis (eg to act friendly or unfriendly) By pretendingthat the three shapes in the SAT are people stored knowl-edge about people in social interactions might beretrieved causing the observed activations in the FG
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
424 R T Schultz and others FFA and social cognition
There would be a measure of efficiency from thisarrangement that is having representations and compu-tations of more abstract attributes of people inter-digitatedwith front-end perceptual processes about physical attri-butes Gauthierrsquos claim is that the same group of neuronscan be engaged by perceptually different categories ofobjects it is the type of processing rather than the visualdetails that are important Extrapolating from this wewould argue that the SAT engages a region of the FGoverlapping with the FFA because it demands compu-tational processes to classify the SAT geometric figures aspeople or person-like Chao et al (1999) reach a similarconclusion They argued that activity in the ventral path-way reflects stored information about an object categorynot just physical features They point out that thisarrangement could explain why lesion patients with cate-gory specific perceptual deficits also have trouble retriev-ing general information about that visual category Thuswe would add that the FFA must store general infor-mation about people or some meta-representation oflsquopeoplenessrsquo
(c) Implications for the pathobiology of autismThe SAT used in this current study appears to be an
effective neurobehavioural probe for engaging a distrib-uted network of brain regions involved in different aspectsof social perception and cognition It will be important touse this and similar procedures in persons with autism tobetter define the nature of brain functions in this disorderCastelli et al (2002) have already taken the first step inthis process and they describe a pattern of hypoactivationin the MPFC STS and TPs in autism In an ongoingstudy of autism spectrum disorders we have presentedpreliminary data using the SAT showing that we too findhypoactivation of these regions and in the amygdala andFG (Schultz et al 2001) Thus we predict that futurework in this area will show that the entire social brain net-work is underactive in autism during tasks requiring socialperceptual and social cognitive processing
There are already sufficient data to argue strongly for arole of the FG in the pathobiology of autism No less thanfive previous fMRI studies have shown the FFA region tobe significantly less engaged among persons with autismcompared with controls during face perception tasks(Critchley et al 2000 Dierks et al 2001 Pierce et al2001 Schultz et al 2000a 2001) Activation of the FGby the social judgements in the current study adds animportant piece of evidence in favour of a causal role forthe FG in the pathobiology of autism This conclusionhowever must remain tentative until additional studiesmore precisely define the factors leading to FG activationduring social attribution and gather better data to proveits computational role in social cognition In addition theeffect of FG lesions for social functioning must be clari-fied If the FG is involved in social computations onewould expect to find social cognitive deficits in personswith lesions to this area However social deficits in proso-pagnosics have not been widely reported It might be thatprosopagnosic patients have not been carefully tested onthis dimension In this regard using our behavioural ver-sion of the SAT (Klin 2000) to test social perceptionamong prosopagnosics would be quite interesting Theparallels however between autism and prosopagnosia are
Phil Trans R Soc Lond B (2003)
incomplete because autism is clearly a developmental dis-order whereas the research literature on prosopagnosia ismostly confined to lesions acquired in adolescence oradulthood Among the few cases of developmental proso-pagnosia reported in the literature there is indeed one thathighlights severe social impairments (Kracke 1994) Itmay be that the role of the FG in the development ofautistic symptoms is different from the role of the FG inthe maintenance of social cognitive functions after brainmaturity
It is also possible that the role of the FG in social pro-cesses is dependent on its functional relationships withother nodes in the social brain and that it is the collectiveaction and interaction of the network that is of primaryimportance for social behaviours For example we founda strong correlation across participants in the amount ofFG and amygdala activation A strictly modular view ofthese areas may be inappropriate as the functions of eachnode could be quite dependent on one another and whenonly considered in isolation quite insufficient to supportsocial processes Thus whereas a lesion to the FG mayimpair visual perception it might not have a large impacton social functioning if this depends on an extended net-work that is dynamic and capable of compensatory adjust-ments Dysfunction of the FG may be necessary but notsufficient to produce social deficits Indeed our relianceon modular models of brain functioning may be leadingus astray in our search for causal mechanisms in autismInstead it may be the collective action of a distributedsystem that is critical to the pathobiology of autism TheFG region may be a key partner in this distributed systembut nevertheless just one node and insufficient by itselfto support in any substantive manner social cognitive pro-cesses or to explain social cognitive deficits seen inautism
5 CONCLUSION AND FUTURE DIRECTIONS
Using the SAT we isolated a distributed network ofactivations that conform to the emerging model of thesocial brain Most important we found significant acti-vation of the central aspect of the FG thus adding thisregion to the expanding list of structures involved in socialprocesses The FG region activated by the SAT overlapsin its spatial extent with the FFA with centres of massdiffering by less than 1 voxel We speculate that these SATactivations represent computational processes associatedwith more abstract attributes of people
It is an open question as to whether the substantial over-lap between the FFA and SAT FG activation is due to anoverlapping or shared set of neuronal assemblies This willneed to be clarified by future work Electrophysiologicalstudies show that there are small face-specific patches inthe FG cortex (Allison et al 1999) In the current studysmall patches of face cells and SAT cells could be inter-mingled but distinct and not drawing on any of the sameneuronal assemblies The spatial resolution of fMRI haslimitations that may preclude any definitive answer to thisquestion but there are strategies that can be used toaddress the issue For example the current study designdid not contain SAT and face discriminations in the sameexperimental fMRI series Doing so would enable directcomparisons of the computational demands placed on the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 425
common area of the FFA It also would be informative iffollow-up studies superimposed faces within the geometricfigures of the SAT without changing the film scripts inany way We could then determine if the computations ofthe common area of the FFA increase in a predictablefashionmdashwould the activation be a linear summation ofthe original SAT plus face discrimination Any significantdeviation from an additive model would suggest that thereis some sharing of neuronal assemblies with the SAT andface discrimination tasks when presented alone
This work was supported by grants from the National Institutesof Child Health and Human Development (grants PO1 HD03008 and PO1 HDDC35482) and the Korczak FoundationThe authors thank the anonymous reviewers for their helpfulcomments Michael Lee for programming the fMRI tasksHedy Serofin and Terry Hickey for their assistance in acquiringthe fMRI data John Herrington for his valuable assistance indata analyses of preliminary studies and members of the YaleDevelopmental Neuroimaging Program for their helpful com-ments on this manuscript A special acknowledgement ofappreciation is given to Donald Cohen who passed away inOctober of 2001 for his outstanding mentorship in all aspectsof this program of work
REFERENCES
Abell F Happe F amp Frith U 2000 Do triangles play tricksAttribution of mental states to animated shapes in normaland abnormal development J Cogn Dev 15 1ndash20
Adolphs R Tranel D amp Damasio A R 1998 The humanamygdala in social judgement Nature 393 470ndash474
Allison T Puce A Spencer D D amp McCarthy G 1999Electrophysiological studies of human face perception Ipotentials generated in occipitotemporal cortex by face andnon-face stimuli Cerebr Cortex 9 415ndash430
Allison T Puce A amp McCarthy G 2000 Social perceptionfrom visual cues role of the STS region Trends Cogn Sci4 267ndash278
Amaral D G amp Price J L 1984 Amygdalo-cortical projec-tions in the monkey (Macaca fascicularis) J Comp Neurol230 465ndash496
Amaral D G Price J L Pitkanen A amp Carmichael S T1992 Anatomical organization of the primate amygdaloidcomplex In The amygdala neurobiological aspects of emotionmemory and mental dysfunction (ed J Aggleton) pp 1ndash66New York Wiley-Liss
Anderson A K amp Phelps E A 2001 Lesions of the humanamygdala impair enhanced perception of emotionally salientevents Nature 411 305ndash309
Bachevalier J 1994 Medial temporal lobe structures andautism a review of clinical and experimental findingsNeuropsychologia 32 627ndash648
Bachevalier J amp Mishkin M 1986 Visual recognition impair-ment follows ventromedial but not dorsolateral prefrontallesions in monkeys Behav Brain Res 20 249ndash261
Baron-Cohen S Ring H A Wheelwright S BullmoreE T Brammer M J Simmons A amp Williams S C R1999 Social intelligence in the normal and autistic brain anfMRI study Eur J Neurosci 11 1891ndash1898
Baron-Cohen S Ring H A Bullmore E T WheelwrightS Ashwina C amp Williams S C R 2000 The amygdalatheory of autism Neurosci Biobehav Rev 24 355ndash364
Bechara A Tranel D Damasio H amp Damasio A R 1996Failure to respond autonomically to anticipated future out-comes following damage to prefrontal cortex Cerebr Cortex6 215ndash225
Benton A 1994 Face recognition Los Angeles CA WesternPsychological Services
Phil Trans R Soc Lond B (2003)
Bonda E Petrides M Ostry D amp Evans A 1996 Specificinvolvement of human parietal systems and the amygdala inthe perception of biological motion J Neurosci 16 3737ndash3744
Bowler D M amp Thommen E 2000 Attribution of mechan-ical and social causality to animated displays by childrenwith autism Autism 4 147ndash171
Brothers L 1990 The social brain a project for integratingprimate behavior and neurophysiology in a new domainConcepts Neurosci 1 27ndash151
Brothers L 1995 Neurophysiology of the perception of inten-tion by primates In The cognitive neurosciences 2nd edn (edM S Gazzaniga) pp 1107ndash1115 Cambridge MA MITPress
Butter C M McDonald J A amp Snyder D R 1969 Oralitypreference behavior and reinforcement value of nonfoodobject in monkeys with orbital frontal lesions Science 1641306ndash1307
Carmichael S T amp Price J L 1995 Limbic connections ofthe orbital and medial prefrontal cortex in macaque monk-eys J Comp Neurol 363 615ndash641
Castelli F Happe F Frith U amp Frith C 2000 Movementand mind a functional imaging study of perception andinterpretation of complex intentional movement patternsNeuroImage 12 314ndash325
Castelli F Frith C Happe F amp Frith U 2002 AutismAsperger syndrome and brain mechanisms for the attri-bution of mental states to animated shapes Brain 1251839ndash1849
Chao L L Haxby J V amp Martin A 1999 Attribute-basedneural substrates in posterior temporal cortex for perceivingand knowing about objects Nat Neurosci 2 913ndash919
Chao L L Weisberg J amp Martin A 2002 Experience-dependent modulation of category-related cortical activityCerebr Cortex 12 545ndash551
Critchley H D (and 10 others) 2000 The functional neuroan-atomy of social behaviour changes in cerebral blood flowwhen people with autistic disorder process facial expressionsBrain 123 2203ndash2212
Culham J C Cavanagh P amp Kanwisher N G 2001 Atten-tion response functions characterizing brain areas usingfMRI activation during parametric variations of attentionalload Neuron 32 737ndash745
Damasio A R Tranel D amp Damasio H 1990 Individualswith sociopathic behavior caused by frontal damage fail torespond autonomically to social stimuli Behav Brain Res41 81ndash94
Dierks T Bolte S Huble D Lanfermannm H amp PoustkaF 2001 Alterations of face processing strategies in autism(a fMRI study) Presented at the 6th Annual Meeting of theOrganization for Human Brain Mapping Brighton UK 10ndash14 June 2001
Dias R Robbins T W amp Roberts A C 1996 Dissociationin prefrontal cortex of affective and attentional shifts Nature380 69ndash72
Downing P E Jiang Y Shuman M amp Kanwisher N 2001A cortical area selective for visual processing of the humanbody Science 293 2470ndash2473
Epstein R amp Kanwisher N 1998 A cortical representation ofthe local visual environment Nature 392 598ndash601
Ernst M Zametkin A J Matochik J A Pascualvaca D ampCohen R M 1997 Reduced medial prefrontal dopami-nergic activity in autistic children Lancet 350 638
Fletcher P C Happe F Frith U Baker S C DolanR J Frackowiak R S amp Frith C D 1995 Other minds inthe brain a functional imaging study of lsquotheory of mindrsquo instory comprehension Cognition 57 109ndash128
Frith C D amp Frith U 1999 Interacting minds a biologicalbasis Science 286 1692ndash1695
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
426 R T Schultz and others FFA and social cognition
Fujita I Tanaka K Ito M amp Cheng K 1992 Columns forvisual features of objects in monkey inferotemporal cortexNature 360 343ndash346
Gaffan E A Gaffan D amp Harrison S 1988 Disconnectionof the amygdala from visual association cortex impairs vis-ual-reward association learning in monkeys J Neurosci 83144ndash3150
Gallagher H L Happe F Brunswick N Fletcher P CFrith U amp Frith C D 2000 Reading the mind in cartoonsand stories an fMRI study of lsquotheory of mindrsquo in verbal andnonverbal tasks Neuropsychologia 38 11ndash21
Gauthier I Anderson A W Tarr M J Skudlarski P ampGore J C 1997 Levels of categorization in visual recog-nition studied using functional magnetic resonance imagingCurr Biol 7 645ndash651
Gauthier I Tarr M J Anderson A W Skudlarski P ampGore J C 1999 Activation of the middle fusiform face areaincreases with expertise in recognizing novel objects NatureNeurosci 2 568ndash573
Gauthier I Skudlarski P Gore J C amp Anderson A W2000 Expertise for cars and birds recruits brain areasinvolved in face recognition Nature Neurosci 3 191ndash197
Goel V Grafman J Sadato N amp Hallett M 1995 Mode-ling other minds Neuroreport 6 1741ndash1746
Grelotti D Gauthier I amp Schultz R T 2001 Social interestand the development of cortical face specialization whatautism teaches us about face processing Devl Psychobiol 40213ndash225
Grezes J Fonlupt P Bertenthal B amp Delon-Martin C2001 Does perception of biological motion rely on specificbrain regions NeuroImage 13 775ndash785
Gusnard D A Akbudak E Shulman G L amp RaichleM E 2001 Medial prefrontal cortex and self-referentialmental activity relation to a default mode of brain functionProc Natl Acad Sci USA 98 4259ndash4264
Happe F Ehlers S Fletcher P Frith U Johansson MGillberg C Dolan R Frackowiak R amp Frith C 1996lsquoTheory of mindrsquo in the brain Evidence from a PET scanstudy of Asperger syndrome Neuroreport 8 197ndash201
Haxby J V Ungerleider L G Clark V P Schouten J LHoffman E A amp Martin A 1999 The effect of face inver-sion on activity in human neural systems for face and objectperception Neuron 22 189ndash199
Haxby J V Isahai A Chao L L Ungerleider L G ampMartin A 2000 Object-form topology in the ventral tem-poral lobe response to I Gauthier (2000) Trends Cogn Sci4 3ndash4
Haxby J V Gobbini M I Furey M L Ishai ASchouten J L amp Pietrini P 2001 Distributed and overlap-ping representations of faces and objects in ventral temporalcortex Science 293 2425ndash2430
Haznedar M M Buchsbaum M S Metzger M Solim-ando A Spiegel-Cohen J amp Hollander E 1997 Anteriorcingulate gyrus volume and glucose metabolism in autisticdisorder Am J Psychiat 154 1047ndash1050
Heider F amp Simmel M 1944 An experimental study ofapparent behavior Am J Psychol 57 243ndash259
Hobson R P Ouston J amp Lee A 1988a Whatrsquos in a faceThe case of autism Br J Psychol 79 441ndash453
Hobson R P Ouston J amp Lee A 1988b Emotion recog-nition in autism coordinating faces and voices Psychol Med18 911ndash923
Hoffman E A amp Haxby J V 2000 Distinct representationsof eye gaze and identity in the distributed human neural sys-tem for face perception Nature Neurosci 3 80ndash84
Hornak J Rolls E T amp Wade D 1996 Face and voiceexpression identification in patients with emotional andbehavioural changes following ventral frontal lobe damageNeuropsychologia 34 247ndash261
Phil Trans R Soc Lond B (2003)
Ishai A Ungerleider L G Martin A Schouten J L ampHaxby J V 1999 Distributed representation of objects inthe human ventral visual pathway Proc Natl Acad Sci USA96 9379ndash9384
Kanwisher N 2000 Domain specificity in face perceptionNature Neurosci 3 759ndash763
Kanwisher N McDermott J amp Chun M M 1997 The fusi-form face area a module of extrastriate cortex specializedfor face perception J Neurosci 17 4302ndash4311
Klin A 2000 Attributing social meaning to ambiguous visualstimuli in higher functioning autism and Asperger syndromethe social attribution task J Child Psychol Psychiat 41831ndash846
Klin A Sparrow S S de Bildt A Cicchetti D V CohenD J amp Volkmar F R 1999 A normed study of face recog-nition in autism and related disorders J Autism Devl Dis-orders 29 499ndash508
Klin A Jones W Schultz R T Volkmar F amp Cohen D J2002 Visual fixation patterns during viewing of naturalisticsocial situations as predictors of social competence in indi-viduals with autism Arch Gen Psychiat 59 809ndash816
Kracke I 1994 Developmental prosopagnosia in Aspergersyndrome presentation and discussion of an individual caseDevl Med Child Neurol 36 873ndash886
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J 1997 Neuroanatomical correlates of happi-ness sadness and disgust Am J Psychiat 154 926ndash933
Langdell T 1978 Recognition of faces an approach for thestudy of autism J Child Psychol Psychiat 19 255ndash268
LeDoux J E 1996 The emotional brain New York Simon ampShuster
Lowel S amp Singer W 1992 Selection of intrinsic horizontalconnections in the visual cortex by correlated neuronalactivity Science 255 209ndash212
Macdonald H Rutter M Howlin P Rios P Le ConteurA Evered C amp Folstein S 1989 Recognition andexpression of emotional cues by autistic and normal adultsJ Child Psychol Psychiat 30 865ndash877
Morris J S Ohman A amp Dolan R J 1998 Conscious andunconscious emotional learning in the human amygdalaNature 393 467ndash470
Myers R E Swett C amp Miller M 1973 Loss of social groupaffinity following prefrontal lesions in free-rangingmacaques Brain Res 64 257ndash269
Osterling J amp Dawson G 1994 Early recognition of childrenwith autism a study of first birthday home video tapes JAutism Devl Disorders 24 247ndash257
Pierce K Muller R A Ambrose J Allen G amp Cour-chesne E 2001 Face processing occurs outside the fusiformlsquoface arearsquo in autism evidence from functional MRI Brain124 2059ndash2073
Price J L Carmichael S T amp Drevets W C 1996 Net-works related to the orbital and medial prefrontal cortex asubstrate for emotional behavior Prog Brain Res 107523ndash536
Puce A Allison T Gore J C amp McCarthy G 1995 Face-sensitive regions in human extrastriate cortex studied byfunctional MRI J Neurophysiol 74 1192ndash1199
Reiman E M Lane R D Ahern G L Schwartz G EDavidson R J Friston K J Yun L S amp Chen K 1997Neuroanatomical correlates of externally and internally gen-erated human emotion Am J Psychiat 154 918ndash925
Rolls E T 1995 A theory of emotion and consciousness andits application to understanding the neural basis of emotionIn The cognitive neurosciences (ed M S Gazzaniga) pp1091ndash1106 Cambridge MA MIT Press
Schultz R T Gauthier I Klin A Fulbright R K Ander-son A W Volkmar F Skudlarski P Lacadie C CohenD J amp Gore J C 2000a Abnormal ventral temporal
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from

424 R T Schultz and others FFA and social cognition
There would be a measure of efficiency from thisarrangement that is having representations and compu-tations of more abstract attributes of people inter-digitatedwith front-end perceptual processes about physical attri-butes Gauthierrsquos claim is that the same group of neuronscan be engaged by perceptually different categories ofobjects it is the type of processing rather than the visualdetails that are important Extrapolating from this wewould argue that the SAT engages a region of the FGoverlapping with the FFA because it demands compu-tational processes to classify the SAT geometric figures aspeople or person-like Chao et al (1999) reach a similarconclusion They argued that activity in the ventral path-way reflects stored information about an object categorynot just physical features They point out that thisarrangement could explain why lesion patients with cate-gory specific perceptual deficits also have trouble retriev-ing general information about that visual category Thuswe would add that the FFA must store general infor-mation about people or some meta-representation oflsquopeoplenessrsquo
(c) Implications for the pathobiology of autismThe SAT used in this current study appears to be an
effective neurobehavioural probe for engaging a distrib-uted network of brain regions involved in different aspectsof social perception and cognition It will be important touse this and similar procedures in persons with autism tobetter define the nature of brain functions in this disorderCastelli et al (2002) have already taken the first step inthis process and they describe a pattern of hypoactivationin the MPFC STS and TPs in autism In an ongoingstudy of autism spectrum disorders we have presentedpreliminary data using the SAT showing that we too findhypoactivation of these regions and in the amygdala andFG (Schultz et al 2001) Thus we predict that futurework in this area will show that the entire social brain net-work is underactive in autism during tasks requiring socialperceptual and social cognitive processing
There are already sufficient data to argue strongly for arole of the FG in the pathobiology of autism No less thanfive previous fMRI studies have shown the FFA region tobe significantly less engaged among persons with autismcompared with controls during face perception tasks(Critchley et al 2000 Dierks et al 2001 Pierce et al2001 Schultz et al 2000a 2001) Activation of the FGby the social judgements in the current study adds animportant piece of evidence in favour of a causal role forthe FG in the pathobiology of autism This conclusionhowever must remain tentative until additional studiesmore precisely define the factors leading to FG activationduring social attribution and gather better data to proveits computational role in social cognition In addition theeffect of FG lesions for social functioning must be clari-fied If the FG is involved in social computations onewould expect to find social cognitive deficits in personswith lesions to this area However social deficits in proso-pagnosics have not been widely reported It might be thatprosopagnosic patients have not been carefully tested onthis dimension In this regard using our behavioural ver-sion of the SAT (Klin 2000) to test social perceptionamong prosopagnosics would be quite interesting Theparallels however between autism and prosopagnosia are
Phil Trans R Soc Lond B (2003)
incomplete because autism is clearly a developmental dis-order whereas the research literature on prosopagnosia ismostly confined to lesions acquired in adolescence oradulthood Among the few cases of developmental proso-pagnosia reported in the literature there is indeed one thathighlights severe social impairments (Kracke 1994) Itmay be that the role of the FG in the development ofautistic symptoms is different from the role of the FG inthe maintenance of social cognitive functions after brainmaturity
It is also possible that the role of the FG in social pro-cesses is dependent on its functional relationships withother nodes in the social brain and that it is the collectiveaction and interaction of the network that is of primaryimportance for social behaviours For example we founda strong correlation across participants in the amount ofFG and amygdala activation A strictly modular view ofthese areas may be inappropriate as the functions of eachnode could be quite dependent on one another and whenonly considered in isolation quite insufficient to supportsocial processes Thus whereas a lesion to the FG mayimpair visual perception it might not have a large impacton social functioning if this depends on an extended net-work that is dynamic and capable of compensatory adjust-ments Dysfunction of the FG may be necessary but notsufficient to produce social deficits Indeed our relianceon modular models of brain functioning may be leadingus astray in our search for causal mechanisms in autismInstead it may be the collective action of a distributedsystem that is critical to the pathobiology of autism TheFG region may be a key partner in this distributed systembut nevertheless just one node and insufficient by itselfto support in any substantive manner social cognitive pro-cesses or to explain social cognitive deficits seen inautism
5 CONCLUSION AND FUTURE DIRECTIONS
Using the SAT we isolated a distributed network ofactivations that conform to the emerging model of thesocial brain Most important we found significant acti-vation of the central aspect of the FG thus adding thisregion to the expanding list of structures involved in socialprocesses The FG region activated by the SAT overlapsin its spatial extent with the FFA with centres of massdiffering by less than 1 voxel We speculate that these SATactivations represent computational processes associatedwith more abstract attributes of people
It is an open question as to whether the substantial over-lap between the FFA and SAT FG activation is due to anoverlapping or shared set of neuronal assemblies This willneed to be clarified by future work Electrophysiologicalstudies show that there are small face-specific patches inthe FG cortex (Allison et al 1999) In the current studysmall patches of face cells and SAT cells could be inter-mingled but distinct and not drawing on any of the sameneuronal assemblies The spatial resolution of fMRI haslimitations that may preclude any definitive answer to thisquestion but there are strategies that can be used toaddress the issue For example the current study designdid not contain SAT and face discriminations in the sameexperimental fMRI series Doing so would enable directcomparisons of the computational demands placed on the
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 425
common area of the FFA It also would be informative iffollow-up studies superimposed faces within the geometricfigures of the SAT without changing the film scripts inany way We could then determine if the computations ofthe common area of the FFA increase in a predictablefashionmdashwould the activation be a linear summation ofthe original SAT plus face discrimination Any significantdeviation from an additive model would suggest that thereis some sharing of neuronal assemblies with the SAT andface discrimination tasks when presented alone
This work was supported by grants from the National Institutesof Child Health and Human Development (grants PO1 HD03008 and PO1 HDDC35482) and the Korczak FoundationThe authors thank the anonymous reviewers for their helpfulcomments Michael Lee for programming the fMRI tasksHedy Serofin and Terry Hickey for their assistance in acquiringthe fMRI data John Herrington for his valuable assistance indata analyses of preliminary studies and members of the YaleDevelopmental Neuroimaging Program for their helpful com-ments on this manuscript A special acknowledgement ofappreciation is given to Donald Cohen who passed away inOctober of 2001 for his outstanding mentorship in all aspectsof this program of work
REFERENCES
Abell F Happe F amp Frith U 2000 Do triangles play tricksAttribution of mental states to animated shapes in normaland abnormal development J Cogn Dev 15 1ndash20
Adolphs R Tranel D amp Damasio A R 1998 The humanamygdala in social judgement Nature 393 470ndash474
Allison T Puce A Spencer D D amp McCarthy G 1999Electrophysiological studies of human face perception Ipotentials generated in occipitotemporal cortex by face andnon-face stimuli Cerebr Cortex 9 415ndash430
Allison T Puce A amp McCarthy G 2000 Social perceptionfrom visual cues role of the STS region Trends Cogn Sci4 267ndash278
Amaral D G amp Price J L 1984 Amygdalo-cortical projec-tions in the monkey (Macaca fascicularis) J Comp Neurol230 465ndash496
Amaral D G Price J L Pitkanen A amp Carmichael S T1992 Anatomical organization of the primate amygdaloidcomplex In The amygdala neurobiological aspects of emotionmemory and mental dysfunction (ed J Aggleton) pp 1ndash66New York Wiley-Liss
Anderson A K amp Phelps E A 2001 Lesions of the humanamygdala impair enhanced perception of emotionally salientevents Nature 411 305ndash309
Bachevalier J 1994 Medial temporal lobe structures andautism a review of clinical and experimental findingsNeuropsychologia 32 627ndash648
Bachevalier J amp Mishkin M 1986 Visual recognition impair-ment follows ventromedial but not dorsolateral prefrontallesions in monkeys Behav Brain Res 20 249ndash261
Baron-Cohen S Ring H A Wheelwright S BullmoreE T Brammer M J Simmons A amp Williams S C R1999 Social intelligence in the normal and autistic brain anfMRI study Eur J Neurosci 11 1891ndash1898
Baron-Cohen S Ring H A Bullmore E T WheelwrightS Ashwina C amp Williams S C R 2000 The amygdalatheory of autism Neurosci Biobehav Rev 24 355ndash364
Bechara A Tranel D Damasio H amp Damasio A R 1996Failure to respond autonomically to anticipated future out-comes following damage to prefrontal cortex Cerebr Cortex6 215ndash225
Benton A 1994 Face recognition Los Angeles CA WesternPsychological Services
Phil Trans R Soc Lond B (2003)
Bonda E Petrides M Ostry D amp Evans A 1996 Specificinvolvement of human parietal systems and the amygdala inthe perception of biological motion J Neurosci 16 3737ndash3744
Bowler D M amp Thommen E 2000 Attribution of mechan-ical and social causality to animated displays by childrenwith autism Autism 4 147ndash171
Brothers L 1990 The social brain a project for integratingprimate behavior and neurophysiology in a new domainConcepts Neurosci 1 27ndash151
Brothers L 1995 Neurophysiology of the perception of inten-tion by primates In The cognitive neurosciences 2nd edn (edM S Gazzaniga) pp 1107ndash1115 Cambridge MA MITPress
Butter C M McDonald J A amp Snyder D R 1969 Oralitypreference behavior and reinforcement value of nonfoodobject in monkeys with orbital frontal lesions Science 1641306ndash1307
Carmichael S T amp Price J L 1995 Limbic connections ofthe orbital and medial prefrontal cortex in macaque monk-eys J Comp Neurol 363 615ndash641
Castelli F Happe F Frith U amp Frith C 2000 Movementand mind a functional imaging study of perception andinterpretation of complex intentional movement patternsNeuroImage 12 314ndash325
Castelli F Frith C Happe F amp Frith U 2002 AutismAsperger syndrome and brain mechanisms for the attri-bution of mental states to animated shapes Brain 1251839ndash1849
Chao L L Haxby J V amp Martin A 1999 Attribute-basedneural substrates in posterior temporal cortex for perceivingand knowing about objects Nat Neurosci 2 913ndash919
Chao L L Weisberg J amp Martin A 2002 Experience-dependent modulation of category-related cortical activityCerebr Cortex 12 545ndash551
Critchley H D (and 10 others) 2000 The functional neuroan-atomy of social behaviour changes in cerebral blood flowwhen people with autistic disorder process facial expressionsBrain 123 2203ndash2212
Culham J C Cavanagh P amp Kanwisher N G 2001 Atten-tion response functions characterizing brain areas usingfMRI activation during parametric variations of attentionalload Neuron 32 737ndash745
Damasio A R Tranel D amp Damasio H 1990 Individualswith sociopathic behavior caused by frontal damage fail torespond autonomically to social stimuli Behav Brain Res41 81ndash94
Dierks T Bolte S Huble D Lanfermannm H amp PoustkaF 2001 Alterations of face processing strategies in autism(a fMRI study) Presented at the 6th Annual Meeting of theOrganization for Human Brain Mapping Brighton UK 10ndash14 June 2001
Dias R Robbins T W amp Roberts A C 1996 Dissociationin prefrontal cortex of affective and attentional shifts Nature380 69ndash72
Downing P E Jiang Y Shuman M amp Kanwisher N 2001A cortical area selective for visual processing of the humanbody Science 293 2470ndash2473
Epstein R amp Kanwisher N 1998 A cortical representation ofthe local visual environment Nature 392 598ndash601
Ernst M Zametkin A J Matochik J A Pascualvaca D ampCohen R M 1997 Reduced medial prefrontal dopami-nergic activity in autistic children Lancet 350 638
Fletcher P C Happe F Frith U Baker S C DolanR J Frackowiak R S amp Frith C D 1995 Other minds inthe brain a functional imaging study of lsquotheory of mindrsquo instory comprehension Cognition 57 109ndash128
Frith C D amp Frith U 1999 Interacting minds a biologicalbasis Science 286 1692ndash1695
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
426 R T Schultz and others FFA and social cognition
Fujita I Tanaka K Ito M amp Cheng K 1992 Columns forvisual features of objects in monkey inferotemporal cortexNature 360 343ndash346
Gaffan E A Gaffan D amp Harrison S 1988 Disconnectionof the amygdala from visual association cortex impairs vis-ual-reward association learning in monkeys J Neurosci 83144ndash3150
Gallagher H L Happe F Brunswick N Fletcher P CFrith U amp Frith C D 2000 Reading the mind in cartoonsand stories an fMRI study of lsquotheory of mindrsquo in verbal andnonverbal tasks Neuropsychologia 38 11ndash21
Gauthier I Anderson A W Tarr M J Skudlarski P ampGore J C 1997 Levels of categorization in visual recog-nition studied using functional magnetic resonance imagingCurr Biol 7 645ndash651
Gauthier I Tarr M J Anderson A W Skudlarski P ampGore J C 1999 Activation of the middle fusiform face areaincreases with expertise in recognizing novel objects NatureNeurosci 2 568ndash573
Gauthier I Skudlarski P Gore J C amp Anderson A W2000 Expertise for cars and birds recruits brain areasinvolved in face recognition Nature Neurosci 3 191ndash197
Goel V Grafman J Sadato N amp Hallett M 1995 Mode-ling other minds Neuroreport 6 1741ndash1746
Grelotti D Gauthier I amp Schultz R T 2001 Social interestand the development of cortical face specialization whatautism teaches us about face processing Devl Psychobiol 40213ndash225
Grezes J Fonlupt P Bertenthal B amp Delon-Martin C2001 Does perception of biological motion rely on specificbrain regions NeuroImage 13 775ndash785
Gusnard D A Akbudak E Shulman G L amp RaichleM E 2001 Medial prefrontal cortex and self-referentialmental activity relation to a default mode of brain functionProc Natl Acad Sci USA 98 4259ndash4264
Happe F Ehlers S Fletcher P Frith U Johansson MGillberg C Dolan R Frackowiak R amp Frith C 1996lsquoTheory of mindrsquo in the brain Evidence from a PET scanstudy of Asperger syndrome Neuroreport 8 197ndash201
Haxby J V Ungerleider L G Clark V P Schouten J LHoffman E A amp Martin A 1999 The effect of face inver-sion on activity in human neural systems for face and objectperception Neuron 22 189ndash199
Haxby J V Isahai A Chao L L Ungerleider L G ampMartin A 2000 Object-form topology in the ventral tem-poral lobe response to I Gauthier (2000) Trends Cogn Sci4 3ndash4
Haxby J V Gobbini M I Furey M L Ishai ASchouten J L amp Pietrini P 2001 Distributed and overlap-ping representations of faces and objects in ventral temporalcortex Science 293 2425ndash2430
Haznedar M M Buchsbaum M S Metzger M Solim-ando A Spiegel-Cohen J amp Hollander E 1997 Anteriorcingulate gyrus volume and glucose metabolism in autisticdisorder Am J Psychiat 154 1047ndash1050
Heider F amp Simmel M 1944 An experimental study ofapparent behavior Am J Psychol 57 243ndash259
Hobson R P Ouston J amp Lee A 1988a Whatrsquos in a faceThe case of autism Br J Psychol 79 441ndash453
Hobson R P Ouston J amp Lee A 1988b Emotion recog-nition in autism coordinating faces and voices Psychol Med18 911ndash923
Hoffman E A amp Haxby J V 2000 Distinct representationsof eye gaze and identity in the distributed human neural sys-tem for face perception Nature Neurosci 3 80ndash84
Hornak J Rolls E T amp Wade D 1996 Face and voiceexpression identification in patients with emotional andbehavioural changes following ventral frontal lobe damageNeuropsychologia 34 247ndash261
Phil Trans R Soc Lond B (2003)
Ishai A Ungerleider L G Martin A Schouten J L ampHaxby J V 1999 Distributed representation of objects inthe human ventral visual pathway Proc Natl Acad Sci USA96 9379ndash9384
Kanwisher N 2000 Domain specificity in face perceptionNature Neurosci 3 759ndash763
Kanwisher N McDermott J amp Chun M M 1997 The fusi-form face area a module of extrastriate cortex specializedfor face perception J Neurosci 17 4302ndash4311
Klin A 2000 Attributing social meaning to ambiguous visualstimuli in higher functioning autism and Asperger syndromethe social attribution task J Child Psychol Psychiat 41831ndash846
Klin A Sparrow S S de Bildt A Cicchetti D V CohenD J amp Volkmar F R 1999 A normed study of face recog-nition in autism and related disorders J Autism Devl Dis-orders 29 499ndash508
Klin A Jones W Schultz R T Volkmar F amp Cohen D J2002 Visual fixation patterns during viewing of naturalisticsocial situations as predictors of social competence in indi-viduals with autism Arch Gen Psychiat 59 809ndash816
Kracke I 1994 Developmental prosopagnosia in Aspergersyndrome presentation and discussion of an individual caseDevl Med Child Neurol 36 873ndash886
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J 1997 Neuroanatomical correlates of happi-ness sadness and disgust Am J Psychiat 154 926ndash933
Langdell T 1978 Recognition of faces an approach for thestudy of autism J Child Psychol Psychiat 19 255ndash268
LeDoux J E 1996 The emotional brain New York Simon ampShuster
Lowel S amp Singer W 1992 Selection of intrinsic horizontalconnections in the visual cortex by correlated neuronalactivity Science 255 209ndash212
Macdonald H Rutter M Howlin P Rios P Le ConteurA Evered C amp Folstein S 1989 Recognition andexpression of emotional cues by autistic and normal adultsJ Child Psychol Psychiat 30 865ndash877
Morris J S Ohman A amp Dolan R J 1998 Conscious andunconscious emotional learning in the human amygdalaNature 393 467ndash470
Myers R E Swett C amp Miller M 1973 Loss of social groupaffinity following prefrontal lesions in free-rangingmacaques Brain Res 64 257ndash269
Osterling J amp Dawson G 1994 Early recognition of childrenwith autism a study of first birthday home video tapes JAutism Devl Disorders 24 247ndash257
Pierce K Muller R A Ambrose J Allen G amp Cour-chesne E 2001 Face processing occurs outside the fusiformlsquoface arearsquo in autism evidence from functional MRI Brain124 2059ndash2073
Price J L Carmichael S T amp Drevets W C 1996 Net-works related to the orbital and medial prefrontal cortex asubstrate for emotional behavior Prog Brain Res 107523ndash536
Puce A Allison T Gore J C amp McCarthy G 1995 Face-sensitive regions in human extrastriate cortex studied byfunctional MRI J Neurophysiol 74 1192ndash1199
Reiman E M Lane R D Ahern G L Schwartz G EDavidson R J Friston K J Yun L S amp Chen K 1997Neuroanatomical correlates of externally and internally gen-erated human emotion Am J Psychiat 154 918ndash925
Rolls E T 1995 A theory of emotion and consciousness andits application to understanding the neural basis of emotionIn The cognitive neurosciences (ed M S Gazzaniga) pp1091ndash1106 Cambridge MA MIT Press
Schultz R T Gauthier I Klin A Fulbright R K Ander-son A W Volkmar F Skudlarski P Lacadie C CohenD J amp Gore J C 2000a Abnormal ventral temporal
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from

FFA and social cognition R T Schultz and others 425
common area of the FFA It also would be informative iffollow-up studies superimposed faces within the geometricfigures of the SAT without changing the film scripts inany way We could then determine if the computations ofthe common area of the FFA increase in a predictablefashionmdashwould the activation be a linear summation ofthe original SAT plus face discrimination Any significantdeviation from an additive model would suggest that thereis some sharing of neuronal assemblies with the SAT andface discrimination tasks when presented alone
This work was supported by grants from the National Institutesof Child Health and Human Development (grants PO1 HD03008 and PO1 HDDC35482) and the Korczak FoundationThe authors thank the anonymous reviewers for their helpfulcomments Michael Lee for programming the fMRI tasksHedy Serofin and Terry Hickey for their assistance in acquiringthe fMRI data John Herrington for his valuable assistance indata analyses of preliminary studies and members of the YaleDevelopmental Neuroimaging Program for their helpful com-ments on this manuscript A special acknowledgement ofappreciation is given to Donald Cohen who passed away inOctober of 2001 for his outstanding mentorship in all aspectsof this program of work
REFERENCES
Abell F Happe F amp Frith U 2000 Do triangles play tricksAttribution of mental states to animated shapes in normaland abnormal development J Cogn Dev 15 1ndash20
Adolphs R Tranel D amp Damasio A R 1998 The humanamygdala in social judgement Nature 393 470ndash474
Allison T Puce A Spencer D D amp McCarthy G 1999Electrophysiological studies of human face perception Ipotentials generated in occipitotemporal cortex by face andnon-face stimuli Cerebr Cortex 9 415ndash430
Allison T Puce A amp McCarthy G 2000 Social perceptionfrom visual cues role of the STS region Trends Cogn Sci4 267ndash278
Amaral D G amp Price J L 1984 Amygdalo-cortical projec-tions in the monkey (Macaca fascicularis) J Comp Neurol230 465ndash496
Amaral D G Price J L Pitkanen A amp Carmichael S T1992 Anatomical organization of the primate amygdaloidcomplex In The amygdala neurobiological aspects of emotionmemory and mental dysfunction (ed J Aggleton) pp 1ndash66New York Wiley-Liss
Anderson A K amp Phelps E A 2001 Lesions of the humanamygdala impair enhanced perception of emotionally salientevents Nature 411 305ndash309
Bachevalier J 1994 Medial temporal lobe structures andautism a review of clinical and experimental findingsNeuropsychologia 32 627ndash648
Bachevalier J amp Mishkin M 1986 Visual recognition impair-ment follows ventromedial but not dorsolateral prefrontallesions in monkeys Behav Brain Res 20 249ndash261
Baron-Cohen S Ring H A Wheelwright S BullmoreE T Brammer M J Simmons A amp Williams S C R1999 Social intelligence in the normal and autistic brain anfMRI study Eur J Neurosci 11 1891ndash1898
Baron-Cohen S Ring H A Bullmore E T WheelwrightS Ashwina C amp Williams S C R 2000 The amygdalatheory of autism Neurosci Biobehav Rev 24 355ndash364
Bechara A Tranel D Damasio H amp Damasio A R 1996Failure to respond autonomically to anticipated future out-comes following damage to prefrontal cortex Cerebr Cortex6 215ndash225
Benton A 1994 Face recognition Los Angeles CA WesternPsychological Services
Phil Trans R Soc Lond B (2003)
Bonda E Petrides M Ostry D amp Evans A 1996 Specificinvolvement of human parietal systems and the amygdala inthe perception of biological motion J Neurosci 16 3737ndash3744
Bowler D M amp Thommen E 2000 Attribution of mechan-ical and social causality to animated displays by childrenwith autism Autism 4 147ndash171
Brothers L 1990 The social brain a project for integratingprimate behavior and neurophysiology in a new domainConcepts Neurosci 1 27ndash151
Brothers L 1995 Neurophysiology of the perception of inten-tion by primates In The cognitive neurosciences 2nd edn (edM S Gazzaniga) pp 1107ndash1115 Cambridge MA MITPress
Butter C M McDonald J A amp Snyder D R 1969 Oralitypreference behavior and reinforcement value of nonfoodobject in monkeys with orbital frontal lesions Science 1641306ndash1307
Carmichael S T amp Price J L 1995 Limbic connections ofthe orbital and medial prefrontal cortex in macaque monk-eys J Comp Neurol 363 615ndash641
Castelli F Happe F Frith U amp Frith C 2000 Movementand mind a functional imaging study of perception andinterpretation of complex intentional movement patternsNeuroImage 12 314ndash325
Castelli F Frith C Happe F amp Frith U 2002 AutismAsperger syndrome and brain mechanisms for the attri-bution of mental states to animated shapes Brain 1251839ndash1849
Chao L L Haxby J V amp Martin A 1999 Attribute-basedneural substrates in posterior temporal cortex for perceivingand knowing about objects Nat Neurosci 2 913ndash919
Chao L L Weisberg J amp Martin A 2002 Experience-dependent modulation of category-related cortical activityCerebr Cortex 12 545ndash551
Critchley H D (and 10 others) 2000 The functional neuroan-atomy of social behaviour changes in cerebral blood flowwhen people with autistic disorder process facial expressionsBrain 123 2203ndash2212
Culham J C Cavanagh P amp Kanwisher N G 2001 Atten-tion response functions characterizing brain areas usingfMRI activation during parametric variations of attentionalload Neuron 32 737ndash745
Damasio A R Tranel D amp Damasio H 1990 Individualswith sociopathic behavior caused by frontal damage fail torespond autonomically to social stimuli Behav Brain Res41 81ndash94
Dierks T Bolte S Huble D Lanfermannm H amp PoustkaF 2001 Alterations of face processing strategies in autism(a fMRI study) Presented at the 6th Annual Meeting of theOrganization for Human Brain Mapping Brighton UK 10ndash14 June 2001
Dias R Robbins T W amp Roberts A C 1996 Dissociationin prefrontal cortex of affective and attentional shifts Nature380 69ndash72
Downing P E Jiang Y Shuman M amp Kanwisher N 2001A cortical area selective for visual processing of the humanbody Science 293 2470ndash2473
Epstein R amp Kanwisher N 1998 A cortical representation ofthe local visual environment Nature 392 598ndash601
Ernst M Zametkin A J Matochik J A Pascualvaca D ampCohen R M 1997 Reduced medial prefrontal dopami-nergic activity in autistic children Lancet 350 638
Fletcher P C Happe F Frith U Baker S C DolanR J Frackowiak R S amp Frith C D 1995 Other minds inthe brain a functional imaging study of lsquotheory of mindrsquo instory comprehension Cognition 57 109ndash128
Frith C D amp Frith U 1999 Interacting minds a biologicalbasis Science 286 1692ndash1695
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
426 R T Schultz and others FFA and social cognition
Fujita I Tanaka K Ito M amp Cheng K 1992 Columns forvisual features of objects in monkey inferotemporal cortexNature 360 343ndash346
Gaffan E A Gaffan D amp Harrison S 1988 Disconnectionof the amygdala from visual association cortex impairs vis-ual-reward association learning in monkeys J Neurosci 83144ndash3150
Gallagher H L Happe F Brunswick N Fletcher P CFrith U amp Frith C D 2000 Reading the mind in cartoonsand stories an fMRI study of lsquotheory of mindrsquo in verbal andnonverbal tasks Neuropsychologia 38 11ndash21
Gauthier I Anderson A W Tarr M J Skudlarski P ampGore J C 1997 Levels of categorization in visual recog-nition studied using functional magnetic resonance imagingCurr Biol 7 645ndash651
Gauthier I Tarr M J Anderson A W Skudlarski P ampGore J C 1999 Activation of the middle fusiform face areaincreases with expertise in recognizing novel objects NatureNeurosci 2 568ndash573
Gauthier I Skudlarski P Gore J C amp Anderson A W2000 Expertise for cars and birds recruits brain areasinvolved in face recognition Nature Neurosci 3 191ndash197
Goel V Grafman J Sadato N amp Hallett M 1995 Mode-ling other minds Neuroreport 6 1741ndash1746
Grelotti D Gauthier I amp Schultz R T 2001 Social interestand the development of cortical face specialization whatautism teaches us about face processing Devl Psychobiol 40213ndash225
Grezes J Fonlupt P Bertenthal B amp Delon-Martin C2001 Does perception of biological motion rely on specificbrain regions NeuroImage 13 775ndash785
Gusnard D A Akbudak E Shulman G L amp RaichleM E 2001 Medial prefrontal cortex and self-referentialmental activity relation to a default mode of brain functionProc Natl Acad Sci USA 98 4259ndash4264
Happe F Ehlers S Fletcher P Frith U Johansson MGillberg C Dolan R Frackowiak R amp Frith C 1996lsquoTheory of mindrsquo in the brain Evidence from a PET scanstudy of Asperger syndrome Neuroreport 8 197ndash201
Haxby J V Ungerleider L G Clark V P Schouten J LHoffman E A amp Martin A 1999 The effect of face inver-sion on activity in human neural systems for face and objectperception Neuron 22 189ndash199
Haxby J V Isahai A Chao L L Ungerleider L G ampMartin A 2000 Object-form topology in the ventral tem-poral lobe response to I Gauthier (2000) Trends Cogn Sci4 3ndash4
Haxby J V Gobbini M I Furey M L Ishai ASchouten J L amp Pietrini P 2001 Distributed and overlap-ping representations of faces and objects in ventral temporalcortex Science 293 2425ndash2430
Haznedar M M Buchsbaum M S Metzger M Solim-ando A Spiegel-Cohen J amp Hollander E 1997 Anteriorcingulate gyrus volume and glucose metabolism in autisticdisorder Am J Psychiat 154 1047ndash1050
Heider F amp Simmel M 1944 An experimental study ofapparent behavior Am J Psychol 57 243ndash259
Hobson R P Ouston J amp Lee A 1988a Whatrsquos in a faceThe case of autism Br J Psychol 79 441ndash453
Hobson R P Ouston J amp Lee A 1988b Emotion recog-nition in autism coordinating faces and voices Psychol Med18 911ndash923
Hoffman E A amp Haxby J V 2000 Distinct representationsof eye gaze and identity in the distributed human neural sys-tem for face perception Nature Neurosci 3 80ndash84
Hornak J Rolls E T amp Wade D 1996 Face and voiceexpression identification in patients with emotional andbehavioural changes following ventral frontal lobe damageNeuropsychologia 34 247ndash261
Phil Trans R Soc Lond B (2003)
Ishai A Ungerleider L G Martin A Schouten J L ampHaxby J V 1999 Distributed representation of objects inthe human ventral visual pathway Proc Natl Acad Sci USA96 9379ndash9384
Kanwisher N 2000 Domain specificity in face perceptionNature Neurosci 3 759ndash763
Kanwisher N McDermott J amp Chun M M 1997 The fusi-form face area a module of extrastriate cortex specializedfor face perception J Neurosci 17 4302ndash4311
Klin A 2000 Attributing social meaning to ambiguous visualstimuli in higher functioning autism and Asperger syndromethe social attribution task J Child Psychol Psychiat 41831ndash846
Klin A Sparrow S S de Bildt A Cicchetti D V CohenD J amp Volkmar F R 1999 A normed study of face recog-nition in autism and related disorders J Autism Devl Dis-orders 29 499ndash508
Klin A Jones W Schultz R T Volkmar F amp Cohen D J2002 Visual fixation patterns during viewing of naturalisticsocial situations as predictors of social competence in indi-viduals with autism Arch Gen Psychiat 59 809ndash816
Kracke I 1994 Developmental prosopagnosia in Aspergersyndrome presentation and discussion of an individual caseDevl Med Child Neurol 36 873ndash886
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J 1997 Neuroanatomical correlates of happi-ness sadness and disgust Am J Psychiat 154 926ndash933
Langdell T 1978 Recognition of faces an approach for thestudy of autism J Child Psychol Psychiat 19 255ndash268
LeDoux J E 1996 The emotional brain New York Simon ampShuster
Lowel S amp Singer W 1992 Selection of intrinsic horizontalconnections in the visual cortex by correlated neuronalactivity Science 255 209ndash212
Macdonald H Rutter M Howlin P Rios P Le ConteurA Evered C amp Folstein S 1989 Recognition andexpression of emotional cues by autistic and normal adultsJ Child Psychol Psychiat 30 865ndash877
Morris J S Ohman A amp Dolan R J 1998 Conscious andunconscious emotional learning in the human amygdalaNature 393 467ndash470
Myers R E Swett C amp Miller M 1973 Loss of social groupaffinity following prefrontal lesions in free-rangingmacaques Brain Res 64 257ndash269
Osterling J amp Dawson G 1994 Early recognition of childrenwith autism a study of first birthday home video tapes JAutism Devl Disorders 24 247ndash257
Pierce K Muller R A Ambrose J Allen G amp Cour-chesne E 2001 Face processing occurs outside the fusiformlsquoface arearsquo in autism evidence from functional MRI Brain124 2059ndash2073
Price J L Carmichael S T amp Drevets W C 1996 Net-works related to the orbital and medial prefrontal cortex asubstrate for emotional behavior Prog Brain Res 107523ndash536
Puce A Allison T Gore J C amp McCarthy G 1995 Face-sensitive regions in human extrastriate cortex studied byfunctional MRI J Neurophysiol 74 1192ndash1199
Reiman E M Lane R D Ahern G L Schwartz G EDavidson R J Friston K J Yun L S amp Chen K 1997Neuroanatomical correlates of externally and internally gen-erated human emotion Am J Psychiat 154 918ndash925
Rolls E T 1995 A theory of emotion and consciousness andits application to understanding the neural basis of emotionIn The cognitive neurosciences (ed M S Gazzaniga) pp1091ndash1106 Cambridge MA MIT Press
Schultz R T Gauthier I Klin A Fulbright R K Ander-son A W Volkmar F Skudlarski P Lacadie C CohenD J amp Gore J C 2000a Abnormal ventral temporal
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from

426 R T Schultz and others FFA and social cognition
Fujita I Tanaka K Ito M amp Cheng K 1992 Columns forvisual features of objects in monkey inferotemporal cortexNature 360 343ndash346
Gaffan E A Gaffan D amp Harrison S 1988 Disconnectionof the amygdala from visual association cortex impairs vis-ual-reward association learning in monkeys J Neurosci 83144ndash3150
Gallagher H L Happe F Brunswick N Fletcher P CFrith U amp Frith C D 2000 Reading the mind in cartoonsand stories an fMRI study of lsquotheory of mindrsquo in verbal andnonverbal tasks Neuropsychologia 38 11ndash21
Gauthier I Anderson A W Tarr M J Skudlarski P ampGore J C 1997 Levels of categorization in visual recog-nition studied using functional magnetic resonance imagingCurr Biol 7 645ndash651
Gauthier I Tarr M J Anderson A W Skudlarski P ampGore J C 1999 Activation of the middle fusiform face areaincreases with expertise in recognizing novel objects NatureNeurosci 2 568ndash573
Gauthier I Skudlarski P Gore J C amp Anderson A W2000 Expertise for cars and birds recruits brain areasinvolved in face recognition Nature Neurosci 3 191ndash197
Goel V Grafman J Sadato N amp Hallett M 1995 Mode-ling other minds Neuroreport 6 1741ndash1746
Grelotti D Gauthier I amp Schultz R T 2001 Social interestand the development of cortical face specialization whatautism teaches us about face processing Devl Psychobiol 40213ndash225
Grezes J Fonlupt P Bertenthal B amp Delon-Martin C2001 Does perception of biological motion rely on specificbrain regions NeuroImage 13 775ndash785
Gusnard D A Akbudak E Shulman G L amp RaichleM E 2001 Medial prefrontal cortex and self-referentialmental activity relation to a default mode of brain functionProc Natl Acad Sci USA 98 4259ndash4264
Happe F Ehlers S Fletcher P Frith U Johansson MGillberg C Dolan R Frackowiak R amp Frith C 1996lsquoTheory of mindrsquo in the brain Evidence from a PET scanstudy of Asperger syndrome Neuroreport 8 197ndash201
Haxby J V Ungerleider L G Clark V P Schouten J LHoffman E A amp Martin A 1999 The effect of face inver-sion on activity in human neural systems for face and objectperception Neuron 22 189ndash199
Haxby J V Isahai A Chao L L Ungerleider L G ampMartin A 2000 Object-form topology in the ventral tem-poral lobe response to I Gauthier (2000) Trends Cogn Sci4 3ndash4
Haxby J V Gobbini M I Furey M L Ishai ASchouten J L amp Pietrini P 2001 Distributed and overlap-ping representations of faces and objects in ventral temporalcortex Science 293 2425ndash2430
Haznedar M M Buchsbaum M S Metzger M Solim-ando A Spiegel-Cohen J amp Hollander E 1997 Anteriorcingulate gyrus volume and glucose metabolism in autisticdisorder Am J Psychiat 154 1047ndash1050
Heider F amp Simmel M 1944 An experimental study ofapparent behavior Am J Psychol 57 243ndash259
Hobson R P Ouston J amp Lee A 1988a Whatrsquos in a faceThe case of autism Br J Psychol 79 441ndash453
Hobson R P Ouston J amp Lee A 1988b Emotion recog-nition in autism coordinating faces and voices Psychol Med18 911ndash923
Hoffman E A amp Haxby J V 2000 Distinct representationsof eye gaze and identity in the distributed human neural sys-tem for face perception Nature Neurosci 3 80ndash84
Hornak J Rolls E T amp Wade D 1996 Face and voiceexpression identification in patients with emotional andbehavioural changes following ventral frontal lobe damageNeuropsychologia 34 247ndash261
Phil Trans R Soc Lond B (2003)
Ishai A Ungerleider L G Martin A Schouten J L ampHaxby J V 1999 Distributed representation of objects inthe human ventral visual pathway Proc Natl Acad Sci USA96 9379ndash9384
Kanwisher N 2000 Domain specificity in face perceptionNature Neurosci 3 759ndash763
Kanwisher N McDermott J amp Chun M M 1997 The fusi-form face area a module of extrastriate cortex specializedfor face perception J Neurosci 17 4302ndash4311
Klin A 2000 Attributing social meaning to ambiguous visualstimuli in higher functioning autism and Asperger syndromethe social attribution task J Child Psychol Psychiat 41831ndash846
Klin A Sparrow S S de Bildt A Cicchetti D V CohenD J amp Volkmar F R 1999 A normed study of face recog-nition in autism and related disorders J Autism Devl Dis-orders 29 499ndash508
Klin A Jones W Schultz R T Volkmar F amp Cohen D J2002 Visual fixation patterns during viewing of naturalisticsocial situations as predictors of social competence in indi-viduals with autism Arch Gen Psychiat 59 809ndash816
Kracke I 1994 Developmental prosopagnosia in Aspergersyndrome presentation and discussion of an individual caseDevl Med Child Neurol 36 873ndash886
Lane R D Reiman E M Ahern G L Schwartz G E ampDavidson R J 1997 Neuroanatomical correlates of happi-ness sadness and disgust Am J Psychiat 154 926ndash933
Langdell T 1978 Recognition of faces an approach for thestudy of autism J Child Psychol Psychiat 19 255ndash268
LeDoux J E 1996 The emotional brain New York Simon ampShuster
Lowel S amp Singer W 1992 Selection of intrinsic horizontalconnections in the visual cortex by correlated neuronalactivity Science 255 209ndash212
Macdonald H Rutter M Howlin P Rios P Le ConteurA Evered C amp Folstein S 1989 Recognition andexpression of emotional cues by autistic and normal adultsJ Child Psychol Psychiat 30 865ndash877
Morris J S Ohman A amp Dolan R J 1998 Conscious andunconscious emotional learning in the human amygdalaNature 393 467ndash470
Myers R E Swett C amp Miller M 1973 Loss of social groupaffinity following prefrontal lesions in free-rangingmacaques Brain Res 64 257ndash269
Osterling J amp Dawson G 1994 Early recognition of childrenwith autism a study of first birthday home video tapes JAutism Devl Disorders 24 247ndash257
Pierce K Muller R A Ambrose J Allen G amp Cour-chesne E 2001 Face processing occurs outside the fusiformlsquoface arearsquo in autism evidence from functional MRI Brain124 2059ndash2073
Price J L Carmichael S T amp Drevets W C 1996 Net-works related to the orbital and medial prefrontal cortex asubstrate for emotional behavior Prog Brain Res 107523ndash536
Puce A Allison T Gore J C amp McCarthy G 1995 Face-sensitive regions in human extrastriate cortex studied byfunctional MRI J Neurophysiol 74 1192ndash1199
Reiman E M Lane R D Ahern G L Schwartz G EDavidson R J Friston K J Yun L S amp Chen K 1997Neuroanatomical correlates of externally and internally gen-erated human emotion Am J Psychiat 154 918ndash925
Rolls E T 1995 A theory of emotion and consciousness andits application to understanding the neural basis of emotionIn The cognitive neurosciences (ed M S Gazzaniga) pp1091ndash1106 Cambridge MA MIT Press
Schultz R T Gauthier I Klin A Fulbright R K Ander-son A W Volkmar F Skudlarski P Lacadie C CohenD J amp Gore J C 2000a Abnormal ventral temporal
on May 24 2011rstbroyalsocietypublishingorgDownloaded from
FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from

FFA and social cognition R T Schultz and others 427
cortical activity during face discriminations among individ-uals with autism and Asperger syndrome Arch Gen Psych-iat 37 331ndash340
Schultz R T Romanski L M amp Tsatsanis K D 2000bNeurofunctional models of autistic disorder and Aspergersyndrome In Asperger syndrome (ed A Klin F R Volk-mar amp S S Sparrow) pp 172ndash209 New York The Guil-ford Press
Schultz R T Grelotti D J Klin A Levitan E CanteyT Skudlarski P Gore J C Volkmar F amp Cohen DJ 2001 An fMRI study of face recognition facial expressiondetection and social judgment in autism spectrumconditions Presented at the International Meeting forAutism Research San Diego CA 9ndash10 November2001
Siegal M Carrington J amp Radel M 1996 Theory of mindand pragmatic understanding following right hemispheredamage Brain Lang 53 40ndash50
Skudlarski P Constable R T amp Gore J C 1999 ROCanalysis of statistical methods used in functional MRI indi-vidual subjects Neuroimage 9 311ndash329
Spiridon M amp Kanwisher N 2002 How distributed is visualcategory information in human occipito-temporal cortex AnfMRI study Neuron 35 1157ndash1165
Talairach J amp Tournoux P 1988 Co-planar stereotaxic atlasof the human brain 3-Dimensional proportional system anapproach to cerebral imaging New York Thieme
Tarr M J amp Gauthier I 2000 FFA a flexible fusiform areafor subordinate level visual processing automatized byexpertise Nature Neurosci 3 764ndash769
Vaina L M Solomoni J Chowdhury S Sinha P amp Belli-veau J W 2001 Functional neuroanatomy of biologicalmotion perception in humans Proc Natl Acad Sci 9811 656ndash11 661
Wada Y amp Yamamoto T 2001 Selective impairment of facialrecognition due to a haematoma restricted to the right fusi-form and lateral occipital region J Neurol Neurosurg Psych-iat 71 254ndash257
Webster M J Ungerleider L G amp Bachlevalier J 1991
Phil Trans R Soc Lond B (2003)
Lesions of the inferior temporal area TE in infant monkeysalter cortico-amygdalar projections Neuroreport 2 769ndash772
Wechsler D 1997 Wechsler Adult Intelligence Scale 3rd ednSan Antonio TX Psychological Corp
Whalen P J Rauch S L Etcoff N L McInerney S CLee M B amp Jenike M A 1998 Masked presentations ofemotional facial expressions modulate amygdala activitywithout explicit knowledge J Neurosci 18 411ndash418
Winner E Brownell H Happe F Blum A amp Pincus D2002 Distinguishing lies from jokes theory of mind deficitsand discourse interpretation in right hemisphere brain-dam-aged patients Brain Lang 62 89ndash106
Winston J S Strange B A OrsquoDoherty J amp Dolan R J2002 Automatic and intentional brain responses duringevaluation of trustworthiness of faces Nature Neurosci 5277ndash283
Yirmiya N Sigman M D Kasari C amp Mundy P 1992Empathy and cognition in high-functioning children withautism Child Dev 63 150ndash160
GLOSSARY
ACndashPC anterior commissurendashposterior commissureBOLD blood oxygen level dependentFFA fusiform face areaFG fusiform gyrusfMRI functional magnetic resonance imagingIQ intelligence quotientMFS mid-fusiform sulcusMPFC medial prefrontal cortexNEX number of excitationsPET positron emission tomographyROI region of interestSAT social attribution taskSTG superior temporal gyrusSTS superior temporal sulcusTE echo timeTP temporal poleTR repetition time
on May 24 2011rstbroyalsocietypublishingorgDownloaded from




















