
Variations in Accommodation and Convergence Responses in a Minimally Controlled Photorefractive...
14
ORIGINAL ARTICLE Variations in Accommodation and Convergence Responses in a Minimally Controlled Photorefractive Setting ANNA M. HORWOOD, MSc, DBO(T), JUDITH E. TURNER, PhD, SHEILA M. HOUSTON, BEd, and PATRICIA M. RIDDELL, DPhil Infant Vision Laboratory, Psychology Department, University of Reading, Reading, United Kingdom ABSTRACT:Purpose: A remote haploscopic photorefractor, designed for assessment of accommodation and conver- gence in infants and clinical groups, was used to determine heterophoria accommodative convergence/accommodation (AC/A) ratios in normal naïve adults. These were compared with conventional clinical measures. Methods: Twenty-one naïve subjects were used to compare occluded and unoccluded prism cover test responses with the remote haploscopic photorefractor using a text and picture target. Results: Although luminance was generally low for both targets, binocular vergences were appropriate for target demand in both studies. Binocular accommodation showed greater lag for the highest target accommodative demand and the less demanding target. Occlusion not only reduced vergence response, but also frequently caused a marked reduction in accommodation, especially to the picture target. Normal mean AC/A values were found, but with wide variations between individual subjects. Conclusions: Although mean accommodation, vergence, and AC/A values were comparable with published data, we suggest that in these conditions using naïve subjects, accommodation is frequently inaccurate, especially on occlusion, without concomitant loss of vergence, at least at low light levels. Accommodative convergence may play a less important part in, and other cues contribute more to, the near reflex than has been previously suggested. (Optom Vis Sci 2001;78:791–804) Key Words: accommodation, convergence, AC/A, photorefraction, CA/C A ppropriate accommodation, to overcome blur, and con- vergence, to overcome diplopia, are necessary for near fix- ation and rarely occur in isolation. Accommodation ac- companies vergence and vergence accompanies accommodation. These relationships are conventionally expressed as the accommo- dative convergence to accommodation (AC/A) and convergence accommodation to convergence (CA/C) ratios. The AC/A ratio in particular has been extensively researched in both laboratory and clinical studies. It is used clinically to try to explain differences between near and distance angles of alignment and, if abnormal, is generally considered to be causal in some types of strabismus. Care- ful control of the testing situation results in generally repeatable measures. 1 AC/A measurements are especially used clinically to provide accurate diagnosis, predict responses to lens treatments, and help surgeons plan surgery where surgical procedures can be specifically designed to change the ratio. Studies of the AC/A ratio generally stress that a lens gradient method, where accommodation is stimulated by lens-induced blur at a fixed fixation distance, is the most accurate way to measure the ratio because this eliminates proximal factors that can “contami- nate” the result. 2 The seminal references 1–4 suggest that a normal AC/A ratio is between 5:1 D (prism diopters of vergence to diopters of accommodation) and 3:1 D (approximately 0.5MA:1 D to 0.85 MA:1 D [meter angles of vergence to diopters of accom- modation]). This has passed into many influential texts on clinical practice. 5, 6 However, some recent studies, using the methods that are practical in clinical groups, i.e., prism cover test (PCT) meth- ods, have found that much lower ratios are common. 7, 8 The heterophoria method of Ogle and Martens, 2 where calcula- tions are based on interpupillary distance and near/distance compari- sons and the quantification of the Parks method by Ludwig et al., 9 where near/distance comparisons alone are used, have been shown to produce higher AC/A ratios than found using the lens gradient method AC/A, 3, 10 most probably due to proximal factors. Indeed, proximal convergence has been assumed to be equal to the difference between a lens gradient and heterophoria AC/A ratio. 11 In their early study, Fincham and Walton 12 alluded to idiosyn- cratic variations in how much subjects use accurate accommoda- 1040-5488/01/7811-0791/0 VOL. 78, NO. 11, PP. 791–804 OPTOMETRY AND VISION SCIENCE Copyright © 2001 American Academy of Optometry Optometry and Vision Science, Vol. 78, No. 11, November 2001
Transcript of Variations in Accommodation and Convergence Responses in a Minimally Controlled Photorefractive...

ORIGINAL ARTICLE
Variations in Accommodation and ConvergenceResponses in a Minimally Controlled
Photorefractive Setting
ANNA M. HORWOOD, MSc, DBO(T), JUDITH E. TURNER, PhD, SHEILA M. HOUSTON, BEd,and PATRICIA M. RIDDELL, DPhil
Infant Vision Laboratory, Psychology Department, University of Reading, Reading, United Kingdom
ABSTRACT:Purpose: A remote haploscopic photorefractor, designed for assessment of accommodation and conver-gence in infants and clinical groups, was used to determine heterophoria accommodative convergence/accommodation(AC/A) ratios in normal naïve adults. These were compared with conventional clinical measures. Methods: Twenty-onenaïve subjects were used to compare occluded and unoccluded prism cover test responses with the remote haploscopicphotorefractor using a text and picture target. Results: Although luminance was generally low for both targets,binocular vergences were appropriate for target demand in both studies. Binocular accommodation showed greater lagfor the highest target accommodative demand and the less demanding target. Occlusion not only reduced vergenceresponse, but also frequently caused a marked reduction in accommodation, especially to the picture target. Normalmean AC/A values were found, but with wide variations between individual subjects. Conclusions: Although meanaccommodation, vergence, and AC/A values were comparable with published data, we suggest that in these conditionsusing naïve subjects, accommodation is frequently inaccurate, especially on occlusion, without concomitant loss ofvergence, at least at low light levels. Accommodative convergence may play a less important part in, and other cuescontribute more to, the near reflex than has been previously suggested. (Optom Vis Sci 2001;78:791–804)
Key Words: accommodation, convergence, AC/A, photorefraction, CA/C
Appropriate accommodation, to overcome blur, and con-vergence, to overcome diplopia, are necessary for near fix-ation and rarely occur in isolation. Accommodation ac-
companies vergence and vergence accompanies accommodation.These relationships are conventionally expressed as the accommo-dative convergence to accommodation (AC/A) and convergenceaccommodation to convergence (CA/C) ratios. The AC/A ratio inparticular has been extensively researched in both laboratory andclinical studies. It is used clinically to try to explain differencesbetween near and distance angles of alignment and, if abnormal, isgenerally considered to be causal in some types of strabismus. Care-ful control of the testing situation results in generally repeatablemeasures.1 AC/A measurements are especially used clinically toprovide accurate diagnosis, predict responses to lens treatments,and help surgeons plan surgery where surgical procedures can bespecifically designed to change the ratio.
Studies of the AC/A ratio generally stress that a lens gradientmethod, where accommodation is stimulated by lens-induced blurat a fixed fixation distance, is the most accurate way to measure the
ratio because this eliminates proximal factors that can “contami-nate” the result.2 The seminal references1–4 suggest that a normalAC/A ratio is between 5�:1 D (prism diopters of vergence todiopters of accommodation) and 3�:1 D (approximately 0.5MA:1D to 0.85 MA:1 D [meter angles of vergence to diopters of accom-modation]). This has passed into many influential texts on clinicalpractice.5, 6 However, some recent studies, using the methods thatare practical in clinical groups, i.e., prism cover test (PCT) meth-ods, have found that much lower ratios are common.7, 8
The heterophoria method of Ogle and Martens,2 where calcula-tions are based on interpupillary distance and near/distance compari-sons and the quantification of the Parks method by Ludwig et al.,9
where near/distance comparisons alone are used, have been shown toproduce higher AC/A ratios than found using the lens gradientmethod AC/A,3, 10 most probably due to proximal factors. Indeed,proximal convergence has been assumed to be equal to the differencebetween a lens gradient and heterophoria AC/A ratio.11
In their early study, Fincham and Walton12 alluded to idiosyn-cratic variations in how much subjects use accurate accommoda-
1040-5488/01/7811-0791/0 VOL. 78, NO. 11, PP. 791–804OPTOMETRY AND VISION SCIENCECopyright © 2001 American Academy of Optometry
Optometry and Vision Science, Vol. 78, No. 11, November 2001

tion in relation to vergence. They stressed that although subjectsreported the target as clear, the actual accommodation responsemay be inaccurate. They used this as their reason for emphasizingthat accommodative response rather than stimulus must be knownbefore assumptions can be made about accommodation/conver-gence linkages. They also suggested that there is an “elastic cou-pling” (p. 502) between the accommodative response and blur anddisparity cues to enable ametropic and presbyopic subjects to adaptto refractive correction. Some of their trained subjects were able toaccommodate by effort alone, possibly using voluntary conver-gence to induce accommodation via the CA/C link.
More recently published studies, using adult subjects, have con-centrated on the elimination of possible causes of variation to arriveat a measure of the precise relationship between accommodationand vergence, expressed by the AC/A and CA/C ratios. Most ofthese studies use highly non-naturalistic testing situations. In par-ticular, until recently, binocular measurement of the AC/A ratiohas not been possible due to limitations set by available measure-ment techniques, e.g., optometer-haploscopes. Many studies re-port on variations and idiosyncrasies of individual responses.2, 13–
17 Different instruction sets have been shown to produce differentaccommodative responses.18–20 Some authors have alluded to, orexplicitly stated, the necessity for finding a naturalistic method oftesting that has the ability to open accommodation or vergenceloops. For example, in his major review article on accommodation,Ciuffreda21 discusses the variability of responses found in manystudies and concludes that this variability is the reason for theequivocal responses found. He also comments on the need to in-vestigate fully the multiple inputs to the accommodative system“especially under naturalistic conditions” (p. 250).
Controlled experimental methods, designed to enhance accuracy,fail to address how naïve subjects behave. These studies frequently usesmall numbers of experienced subjects, prolonged training periods, orthe means of many trials. They also fail to demonstrate how, binocu-larly, subjects use a combination of accommodative vergence, dispar-ity-induced vergence, and proximal and looming clues together to fix areal near target. AC/A and CA/C ratios and vergence and accommo-dation gain have all been shown to be repeatable within subjects. It isnot clear, however, how they are weighted in a system where all areinvolved simultaneously in a complex feedback mechanism in a un-demanding binocular setting, or whether this weighting is similarwhen different targets are used.
Clinical studies fail to replicate laboratory studies. Recent at-tempts have used static linear models to understand discrepanciesthat are found in the AC/A ratio when tested using different clin-ical methods.22 These models consider not only the gain of thelinkage between accommodation and vergence, but also the intrin-sic gains of the accommodation and vergence feedback loops. Ref-erence to these components alone, however, has failed to explainthe differences found clinically. It is necessary to add the deadspaceoperators of depth of field and Panum’s fusional area to a nonlinearstatic model to explain the differences in measures of AC/A be-tween clinical methods satisfactorily. These deadspace operators,together with proximal influences mentioned earlier, combine toproduce the AC/A ratio. Thus, variability in any of these factorscan add to both interindividual and intermeasurement differences.It is perhaps surprising, given these sources of variability, that somestudies report such consistent values for the AC/A ratio. We were
interested in investigating whether this consistency would befound in a naïve adult population in conditions where both accom-modative vergence and proximal cues are available. Our setup alsoallowed us to measure both accommodation and vergence binoc-ularly, even with one eye occluded.
The remote haploscopic photorefractor (RHP) was designed byIsrael Abramov and Louise Hainline, Infant Study Center, Brook-lyn College of the City University of New York, for the objectiveinvestigation of vergence and accommodation. The system im-poses few subject constraints while measuring accommodation andvergence in both eyes, even in monocular conditions. In this ex-ploratory study, we used this equipment to investigate how accom-modation and vergences are used in less strictly controlled, butmore environmentally valid, circumstances. Under such condi-tions blur, disparity, and proximal cues do not exist in isolation butact simultaneously as stimuli to accommodation and vergence. Inparticular, we were interested in accommodative and vergence be-havior when disparity vergence cues are removed, leaving mainlyaccommodative and proximal cues. This information can then beused as a basis for studies looking at the ways in which cues com-bine to produce appropriate vergence and accommodative re-sponses in different contexts.
Two studies were carried out. Study 1 compared RHP responseand stimulus AC/A results with a range of clinical lens gradient andheterophoria AC/A ratios using a target comparable to that usedclinically. We predicted that similar results would be produced byall tests, but with a large range compatible with other clinicalstudies of the AC/A ratio.8, 10 Study 2 was carried out to furtherexplore the variations in responses obtained in study 1 but using aless demanding target. We were interested in ascertaining whethersimilar responses would be found to a less “accommodative” target.Apart from the intrinsic interest of this research, it has particularrelevance when comparing adult studies to infant work, whereinstructions to maintain a clear image cannot be given and targetsare larger than threshold and are often displayed in dim lightingconditions to enable undilated photorefraction.
STUDY 1
Study 1 was carried out to compare RHP responses with a rangeof clinical methods for measuring stimulus AC/A ratios using atarget that is comparable to that used clinically.
METHOD
Subjects
Ten subjects participated in study 1. The mean age was 16.7years (range, 9 to 25 years). All subjects were naïve to vision testingsituations and were school age or university undergraduates andpostgraduates with no vision science or optics background. Allsubjects had equal vision of at least 6/6 in either eye and did notwear glasses. None of the subjects had a refractive error greater thanspherical equivalent �0.75 DS or �0.25 DS or astigmatismgreater than �0.50 DC. Three subjects had non-symptom-pro-ducing near exophorias �6�, none more than 12�, of which theyhad been unaware. These were measured with a highly dissociativeprism cover test that reveals maximum angles of deviation.5 All
792 Accommodation and Convergence in a Natural Setting—Horwood et al.
Optometry and Vision Science, Vol. 78, No. 11, November 2001

reduced at 6 m to no more than 2�. It was decided not to excludesubjects with these small exophorias because they were all symp-tom-free and had been recruited from a nonselected population.Exophorias of this size found on prism cover test would not beconsidered pathological if convergence was normal, and it has longbeen accepted that there is no fixed angle of exophoria marking thedividing line between normal and abnormal amounts ofexophoria.23
All subjects had binocular convergence to a nearpoint of at least 7cm from the nose, measured by a RAF nearpoint gauge and were ableto overcome at least 40� base-out prism, indicating good motor fu-sion. All subjects had a subjective nearpoint of accommodation whentested with Times Roman N5 print to at least 6 cm uniocularly andbinocularly. All subjects had clinical subjective accommodative ampli-tudes measured with minus lenses of at least 4 D measured using adistance push-up method. None of the subjects complained of anyvisual symptoms after prolonged close work.
All subjects received minimal instruction to make results as life-like as possible. Because of the pilot and exploratory nature of thisstudy, a wide age range of young subjects was enrolled.
Remote Haploscopic Photorefractor
The remote haploscopic photorefractor (Fig. 1) consists of twooptical pathways, one for off-axis photorefraction and the other fortarget presentation so that binocular photorefraction can take placeindependent of target manipulation.
Target Pathway. The fixation target was an image on acomputer monitor mounted on a motorized beam presented to thesubject via an optical pathway that passed via two concave mirrorsand a beam splitter. Although the fixation targets were physicallyremote from the subject and the subjects viewed a virtual target, the
virtual targets were optically placed at the specified fixation dis-tances along the subject’s line of sight. The optical system was suchthat an image of the subject’s eyes was formed just before the upperconcave mirror (Fig. 1). If the upper image of one eye was occludedit obscured the subject’s view of the target in the same way as if anoccluder was placed directly before that eye, but both eyes were stillable to be photorefracted simultaneously.
The motor that moved the monitor was audible during monitormovement but was fixed 2.5 m from the subject. The sound mayhave alerted the subject to the occurrence of target movement butdid not in itself provide distance or direction clues. We were study-ing relatively uncontrolled conditions and, generally, looming/proximal/auditory cues are a part of near fixation, therefore we didnot make particular effort to exclude them in this study.
We were concerned that the virtual targets appeared real and atthe correct distance. The subjects were not aware of the true posi-tion of the computer monitor. During the calibration phase on aseparate calibration group and at the end of both studies, whenasked to touch the 0.3-m and 0.25-m targets, the adult subjectsreached for the image at the correct distance in midair and werefrequently surprised when it was not real. The adult subjects wereonly then shown the construction of the apparatus. Any awarenessof proximity came from the experimental target itself and not anyforeknowledge about the nature of the apparatus. After testing, thesubjects in this study were often unaware that they had been oc-cluded for part of the experiment and had not known that they hadnot been using both eyes together. This demonstrates that theoccluder was invisible to the subject.
The apparatus was enclosed in black-painted shuttering, androom lighting was dimmed so that light levels were low. The dimlighting was necessary to allow the pupils to dilate sufficiently for
FIGURE 1.Remote haploscopic photorefractor (RHP) and clown target. A: motorized beam; B: monitor; C: upperconcave mirror; D: lower concave mirror; E: image of subject’s eyes and position of occluder; F: beamsplitter; G: plain mirror; H: forehead rest; and J: digital camera.
Accommodation and Convergence in a Natural Setting—Horwood et al. 793
Optometry and Vision Science, Vol. 78, No. 11, November 2001

accurate photorefraction, but it did not result in significant darkadaptation. Previous studies of accommodative behavior in infantshave used similarly low light levels.24
Photorefraction Pathway. The conventional off-axis pho-torefraction optical pathway passed via a periscope so that thecenter of the camera lens was optically on the same axis as thefixation target. Apart from the periscopic arrangement, the pho-torefractor used was similar to that described by Abramov et al.25
Target. The target for study 1 was a patch of Times Roman textsubtending approximately 1.5 by 4.0° at each fixation distance. Tostimulate a level of accommodation necessary for demanding reading,the size of print was varied such that the letters subtended an equalvisual angle at each of the five fixation distances and were equivalent toletter size subtending approximately 7.5 min arc at each distance. Thisdemand compares well with other studies of accommodation/conver-gence relationships in clinical groups.26 The subjects were asked toread aloud a passage from a children’s book containing some unex-pected nonsense words. We chose text because we were interested inproviding as lifelike a detailed task as possible.
The text patch was against a black background and had a meanluminance of 0.50 cd/m2 and, thus, fell within the mesopic range.When looking into the apparatus, the shuttering and camera wereinvisible, and the target, set within the dim outline of the monitor,was all that was visible to the subject.
RHP Procedure. A minimum of a 2-min wait allowed pupildilation before photorefraction. The subjects then placed theirheads against a forehead rest and were instructed to look at thetarget. The targets were presented at 25 cm, 33 cm, 50 cm, 1 m,and 2 m, representing 4, 3, 2, 1, and 0.5 D accommodative de-mand presented in a pseudorandom order of 3, 0.5, 4, 1, and 2 D.A photograph was taken at each fixation distance, with sufficienttime given for initial after-images of the flash to subside betweenphotographs.
The naïve subjects were told that the study was to investigatehow they looked at near targets and were told to “read the printaloud,” but were not asked to make any effort beyond that neces-sary for legibility. All subjects were able to read the print at allfixation distances, although some commented on more difficultywhen monocular. The screen was made blank between presenta-tions at different distances to minimize looming cues, although thedim screen edge still provided some cues to the near response. Theright eye was occluded at the level of the upper mirror in all sub-jects, and photographs were repeated.
Order of PCT or RHP testing and binocular or left occluded(right eye fixing) RHP conditions were counterbalanced acrosssubjects. The occluder was introduced for a full set of five photo-graphs, and occlusion had occurred for at least 1 min before thefirst photograph of a series was taken in the occluded condition.
The photographs were digitally stored. They were then magni-fied by approximately 3� for measurement using image-process-ing software (Adobe Photoshop). Each photograph was measuredby at least two scorers (with a third in cases of disagreement of�0.23 mm when crescent edges were indistinct).
Calculations. Plane of focus was measured from the size ofthe fundal reflex crescent. As target demand increases, the crescentin an emmetropic eye increases in size. The size of the visiblecrescent for a given accommodative response will also be depen-dent on pupil size. Calibration of plane of focus thus requires
information about known refractive state in conjunction with pu-pil and crescent size. The calibration procedure for plane of focuscompared accommodative response derived from dynamic retinos-copy with photographs taken by the RHP. Dynamic retinoscopyof a separate group of 39 subjects with a range of mild or norefractive errors was carried out by a skilled retinoscopist lying inthe optical path of the camera while the subjects fixated all fivetarget distances presented by the upper optical path. Thus, thesame target conditions existed for both photorefraction and reti-noscopy. Retinoscopy results (with appropriate correction forworking distance) were compared with crescent and pupil size datafrom the RHP photographs taken a few moments before. The datawere then processed using neural network techniques to produce asmooth two-dimensional surface that could be applied to futurephotorefraction data. This was done by using the raw data to trainto asymptote a simple feed-forward network using the conjugategradient learning algorithm with weight decay.27 The network wasfully connected with two input units (corresponding to pupil sizeand crescent size), 10 sigmoidal hidden units, and one linear out-put unit (corresponding to accommodative response). The outputsof the trained network gave the smoothed surface that was used tocalculate accommodation estimates from measurements of cornealreflection change and crescent size. Measurement accuracy for thismethod when interscorer reliability is taken into account is �0.3D of accommodation.
Eye position information is gained from analysis of the positionof the corneal reflection in relation to the pupil center. As conver-gence occurs, the pupil center moves nasally in relation to thecorneal reflection of the camera flash. A subset of 12 subjects mo-nocularly fixated a series of five targets arranged horizontally at 5°intervals. The eye rotation measurements for these subjects wereused to determine individual Hirschberg ratios. The mean Hirsch-berg ratio of this group was 12.14°/mm (SD � 1.51°/mm). Thismean was not significantly different from the published values ofaround 12.2°/mm found in infants by Riddell et al.28 and by otherauthors in adults.29–32 We therefore used the published average of12.2°/mm for all other subjects. Measurement accuracy for ourlaboratory is �1.1° (approximately 2�) of vergence change.
Different measurement units of convergence tend to be reportedin different types of studies. For ease of comparison, we have cho-sen to report convergence both in meter angles for easy comparisonwith accommodative plane in diopters and in prism diopters forcomparison with data from clinical studies.
Prism Cover Test
To enable comparisons with clinical methods of measuring theAC/A ratio, two prism cover test methods were used. These werecarried out by an experienced orthoptist (AH). Prism cover tests arecommonly used to assess clinical AC/A ratios in ophthalmology andorthoptic practice and provide strong dissociation. Although not assensitive as subjective vernier alignment tasks, they are accurate towithin at least 2�,5 thus comparing well with the sensitivity of mea-surements of angle using the RHP. Like the RHP method, they alsodo not rely on subjective responses.
Heterophoria Method Stimulus AC/A Ratio. The sub-jects were instructed to “look carefully at the target and read theletters” as is the clinical convention. Because a line of a Snellen
794 Accommodation and Convergence in a Natural Setting—Horwood et al.
Optometry and Vision Science, Vol. 78, No. 11, November 2001
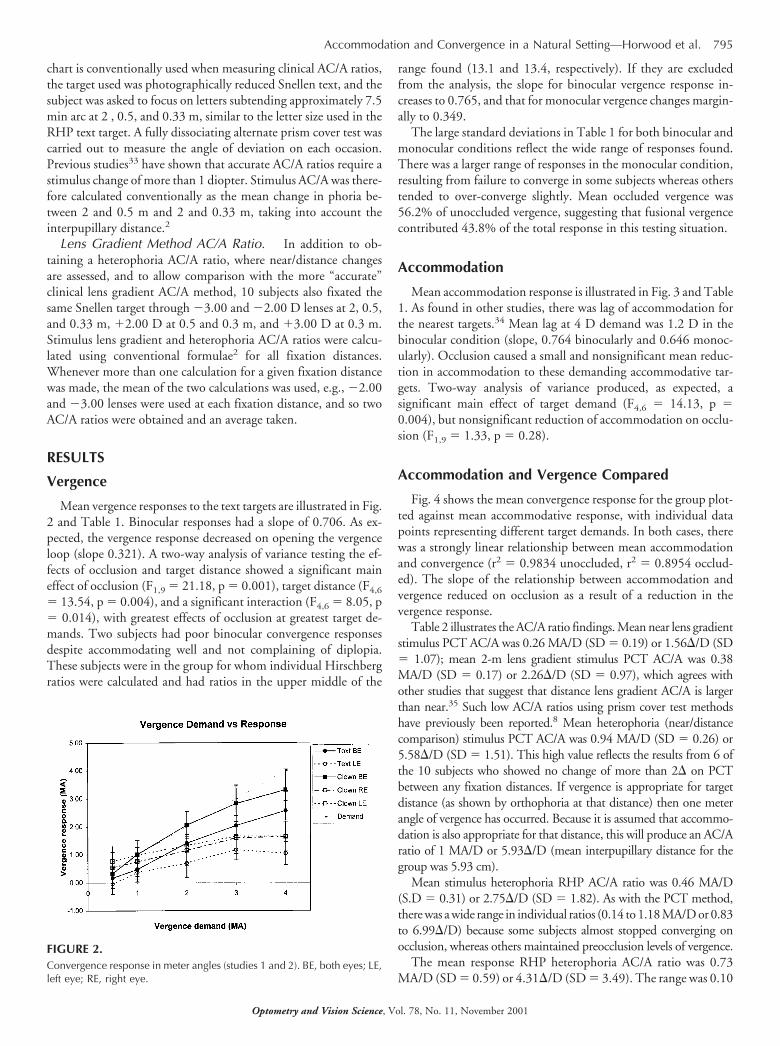
chart is conventionally used when measuring clinical AC/A ratios,the target used was photographically reduced Snellen text, and thesubject was asked to focus on letters subtending approximately 7.5min arc at 2 , 0.5, and 0.33 m, similar to the letter size used in theRHP text target. A fully dissociating alternate prism cover test wascarried out to measure the angle of deviation on each occasion.Previous studies33 have shown that accurate AC/A ratios require astimulus change of more than 1 diopter. Stimulus AC/A was there-fore calculated conventionally as the mean change in phoria be-tween 2 and 0.5 m and 2 and 0.33 m, taking into account theinterpupillary distance.2
Lens Gradient Method AC/A Ratio. In addition to ob-taining a heterophoria AC/A ratio, where near/distance changesare assessed, and to allow comparison with the more “accurate”clinical lens gradient AC/A method, 10 subjects also fixated thesame Snellen target through �3.00 and �2.00 D lenses at 2, 0.5,and 0.33 m, �2.00 D at 0.5 and 0.3 m, and �3.00 D at 0.3 m.Stimulus lens gradient and heterophoria AC/A ratios were calcu-lated using conventional formulae2 for all fixation distances.Whenever more than one calculation for a given fixation distancewas made, the mean of the two calculations was used, e.g., �2.00and �3.00 lenses were used at each fixation distance, and so twoAC/A ratios were obtained and an average taken.
RESULTS
Vergence
Mean vergence responses to the text targets are illustrated in Fig.2 and Table 1. Binocular responses had a slope of 0.706. As ex-pected, the vergence response decreased on opening the vergenceloop (slope 0.321). A two-way analysis of variance testing the ef-fects of occlusion and target distance showed a significant maineffect of occlusion (F1,9 � 21.18, p � 0.001), target distance (F4,6
� 13.54, p � 0.004), and a significant interaction (F4,6 � 8.05, p� 0.014), with greatest effects of occlusion at greatest target de-mands. Two subjects had poor binocular convergence responsesdespite accommodating well and not complaining of diplopia.These subjects were in the group for whom individual Hirschbergratios were calculated and had ratios in the upper middle of the
range found (13.1 and 13.4, respectively). If they are excludedfrom the analysis, the slope for binocular vergence response in-creases to 0.765, and that for monocular vergence changes margin-ally to 0.349.
The large standard deviations in Table 1 for both binocular andmonocular conditions reflect the wide range of responses found.There was a larger range of responses in the monocular condition,resulting from failure to converge in some subjects whereas otherstended to over-converge slightly. Mean occluded vergence was56.2% of unoccluded vergence, suggesting that fusional vergencecontributed 43.8% of the total response in this testing situation.
Accommodation
Mean accommodation response is illustrated in Fig. 3 and Table1. As found in other studies, there was lag of accommodation forthe nearest targets.34 Mean lag at 4 D demand was 1.2 D in thebinocular condition (slope, 0.764 binocularly and 0.646 monoc-ularly). Occlusion caused a small and nonsignificant mean reduc-tion in accommodation to these demanding accommodative tar-gets. Two-way analysis of variance produced, as expected, asignificant main effect of target demand (F4,6 � 14.13, p �0.004), but nonsignificant reduction of accommodation on occlu-sion (F1,9 � 1.33, p � 0.28).
Accommodation and Vergence Compared
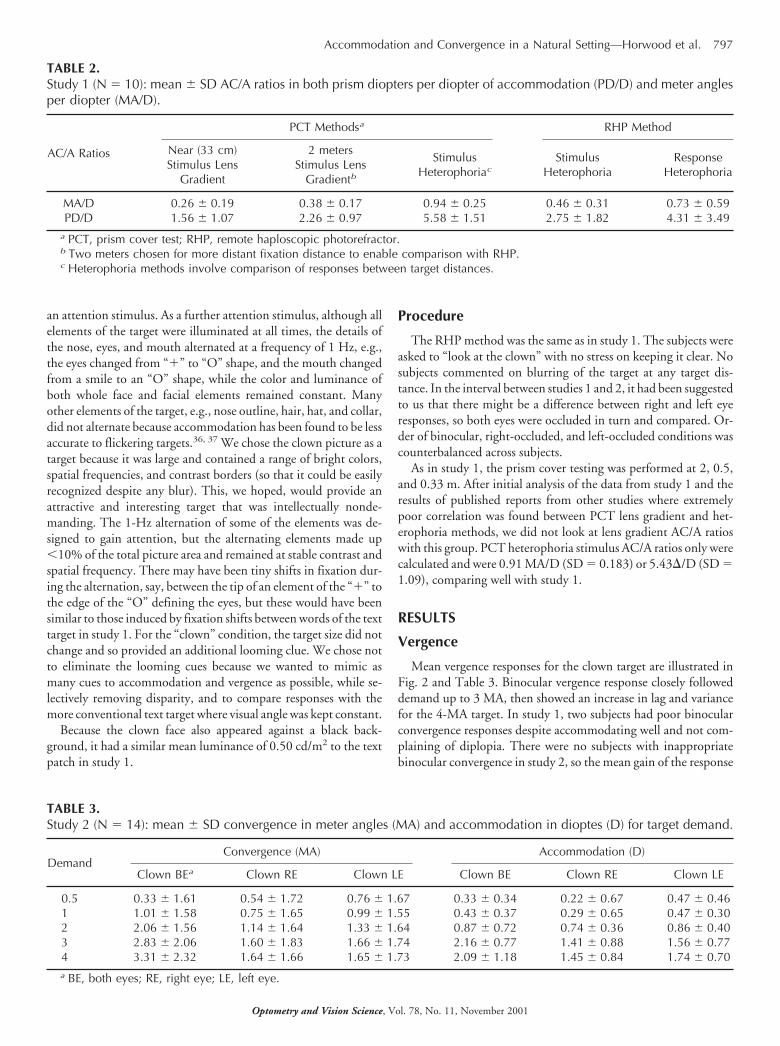
Fig. 4 shows the mean convergence response for the group plot-ted against mean accommodative response, with individual datapoints representing different target demands. In both cases, therewas a strongly linear relationship between mean accommodationand convergence (r2 � 0.9834 unoccluded, r2 � 0.8954 occlud-ed). The slope of the relationship between accommodation andvergence reduced on occlusion as a result of a reduction in thevergence response.
Table 2 illustrates the AC/A ratio findings. Mean near lens gradientstimulus PCT AC/A was 0.26 MA/D (SD � 0.19) or 1.56�/D (SD� 1.07); mean 2-m lens gradient stimulus PCT AC/A was 0.38MA/D (SD � 0.17) or 2.26�/D (SD � 0.97), which agrees withother studies that suggest that distance lens gradient AC/A is largerthan near.35 Such low AC/A ratios using prism cover test methodshave previously been reported.8 Mean heterophoria (near/distancecomparison) stimulus PCT AC/A was 0.94 MA/D (SD � 0.26) or5.58�/D (SD � 1.51). This high value reflects the results from 6 ofthe 10 subjects who showed no change of more than 2� on PCTbetween any fixation distances. If vergence is appropriate for targetdistance (as shown by orthophoria at that distance) then one meterangle of vergence has occurred. Because it is assumed that accommo-dation is also appropriate for that distance, this will produce an AC/Aratio of 1 MA/D or 5.93�/D (mean interpupillary distance for thegroup was 5.93 cm).
Mean stimulus heterophoria RHP AC/A ratio was 0.46 MA/D(S.D � 0.31) or 2.75�/D (SD � 1.82). As with the PCT method,there was a wide range in individual ratios (0.14 to 1.18 MA/D or 0.83to 6.99�/D) because some subjects almost stopped converging onocclusion, whereas others maintained preocclusion levels of vergence.
The mean response RHP heterophoria AC/A ratio was 0.73MA/D (SD � 0.59) or 4.31�/D (SD � 3.49). The range was 0.10
FIGURE 2.Convergence response in meter angles (studies 1 and 2). BE, both eyes; LE,left eye; RE, right eye.
Accommodation and Convergence in a Natural Setting—Horwood et al. 795
Optometry and Vision Science, Vol. 78, No. 11, November 2001
to 1.91 MA/D or 0.59 to 11.32�/D. Not only were vergenceresponses varied, but additional variations also occurred in theresponse to accommodative demand. For example, reduction inaccommodation on occlusion with no concomitant reduction inconvergence results in a high AC/A ratio, whereas loss of conver-gence on occlusion without concomitant loss of accommodationproduces a low AC/A ratio.
No method of AC/A ratio correlated significantly with anotherwithin subjects. This failure can be attributed to the variability ofthe RHP data, the low PCT lens gradient AC/A ratios, and highPCT heterophoria AC/A ratios (produced by the small changes inangle found under the different PCT fixation conditions). Corre-
lation coefficients (Pearson’s r) were �0.049 between 2- and0.3-m PCT lens gradient methods, �0.329 between 2-m PCTlens gradient and PCT heterophoria method, �0.027 betweenPCT and RHP stimulus heterophoria methods, and �0.07 be-tween PCT heterophoria method and the RHP response hetero-phoria method. There was closer correlation between stimulus andresponse RHP AC/A ratios within subjects (r � 0.72).
STUDY 2
Study 2 was carried out to compare RHP stimulus and responseAC/A ratios using different target conditions. Ratios were measuredusing a less demanding target for comparison with ratios measuredwith the detailed targets conventionally used in adults. We wanted tosee whether significant differences in responses occurred when accom-modation was not critical to target identification.
METHOD
Fourteen subjects participated in study 2. The mean age was16.7 years (range, 9 to 28 years). Three subjects had non-symp-tom-producing near exophorias �6�, none more than 12�, ofwhich they had been unaware and similar normal accommodationand convergence to nearpoint. The three youngest subjects hadparticipated in study 1 nine months earlier, but had not beenshown the construction of the apparatus at that time.
Target
The second target was chosen specifically for the infant workalso being carried out in our lab and was designed to be attractive toinfants with a range of visual and intellectual capabilities. At theoutset of these studies, we were interested in the adult response tosuch targets so that future comparisons could be made. But, inaddition, the majority of objects viewed in real-life situations are ofthis nature, with detail being available but not critical to recogni-tion or attention. Little work has been done in this field. Thesecond target (Fig. 1) was a brightly colored picture of a clown(with a white face, red eyes, hair, and mouth, a blue hat with ayellow pom-pom, and a yellow collar against a black background)containing a range of spatial frequencies and measuring 12 by 8 cmsubtending 4.2 by 2.8° at 2 m (i.e., larger than study 1). All colorborders had brightness as well as color contrast. The target had around nose made up of concentric red and black rings, with theinner rings narrower than the outer, which were designed to act as
TABLE 1.Study 1 (N � 10): mean � SD convergence in meter angles (MA) and accommodation in dioptes (D) for target demand.
DemandConvergence (MA) Accommodation (D)
Text BEa Text LE Text BE Text LE
0.5 0.17 � 1.16 �0.05 � 1.14 0.41 � 0.20 0.44 � 0.231 0.48 � 1.16 0.37 � 1.07 0.50 � 0.26 0.62 � 0.252 1.41 � 0.95 0.70 � 1.66 1.57 � 0.76 1.30 � 0.733 2.04 � 1.15 1.17 � 1.16 2.53 � 0.97 2.25 � 0.804 2.57 � 1.09 1.04 � 1.33 2.82 � 0.87 2.52 � 1.15a BE, both eyes; LE, left eye.
FIGURE 3.Accommodation response in diopters (studies 1 and 2). BE, both eyes; LE,left eye; RE, right eye.
FIGURE 4.Study 1. Mean binocular (left chart) and occluded (right chart) conver-gence (MA) plotted against accommodation (D). Individual points repre-sent different target demands. Accom, accomodation; Conv, convergence;Resp, response.
796 Accommodation and Convergence in a Natural Setting—Horwood et al.
Optometry and Vision Science, Vol. 78, No. 11, November 2001
an attention stimulus. As a further attention stimulus, although allelements of the target were illuminated at all times, the details ofthe nose, eyes, and mouth alternated at a frequency of 1 Hz, e.g.,the eyes changed from “�” to “O” shape, and the mouth changedfrom a smile to an “O” shape, while the color and luminance ofboth whole face and facial elements remained constant. Manyother elements of the target, e.g., nose outline, hair, hat, and collar,did not alternate because accommodation has been found to be lessaccurate to flickering targets.36, 37 We chose the clown picture as atarget because it was large and contained a range of bright colors,spatial frequencies, and contrast borders (so that it could be easilyrecognized despite any blur). This, we hoped, would provide anattractive and interesting target that was intellectually nonde-manding. The 1-Hz alternation of some of the elements was de-signed to gain attention, but the alternating elements made up�10% of the total picture area and remained at stable contrast andspatial frequency. There may have been tiny shifts in fixation dur-ing the alternation, say, between the tip of an element of the “�” tothe edge of the “O” defining the eyes, but these would have beensimilar to those induced by fixation shifts between words of the texttarget in study 1. For the “clown” condition, the target size did notchange and so provided an additional looming clue. We chose notto eliminate the looming cues because we wanted to mimic asmany cues to accommodation and vergence as possible, while se-lectively removing disparity, and to compare responses with themore conventional text target where visual angle was kept constant.
Because the clown face also appeared against a black back-ground, it had a similar mean luminance of 0.50 cd/m2 to the textpatch in study 1.
Procedure
The RHP method was the same as in study 1. The subjects wereasked to “look at the clown” with no stress on keeping it clear. Nosubjects commented on blurring of the target at any target dis-tance. In the interval between studies 1 and 2, it had been suggestedto us that there might be a difference between right and left eyeresponses, so both eyes were occluded in turn and compared. Or-der of binocular, right-occluded, and left-occluded conditions wascounterbalanced across subjects.
As in study 1, the prism cover testing was performed at 2, 0.5,and 0.33 m. After initial analysis of the data from study 1 and theresults of published reports from other studies where extremelypoor correlation was found between PCT lens gradient and het-erophoria methods, we did not look at lens gradient AC/A ratioswith this group. PCT heterophoria stimulus AC/A ratios only werecalculated and were 0.91 MA/D (SD � 0.183) or 5.43�/D (SD �1.09), comparing well with study 1.
RESULTS
Vergence
Mean vergence responses for the clown target are illustrated inFig. 2 and Table 3. Binocular vergence response closely followeddemand up to 3 MA, then showed an increase in lag and variancefor the 4-MA target. In study 1, two subjects had poor binocularconvergence responses despite accommodating well and not com-plaining of diplopia. There were no subjects with inappropriatebinocular convergence in study 2, so the mean gain of the response
TABLE 2.Study 1 (N � 10): mean � SD AC/A ratios in both prism diopters per diopter of accommodation (PD/D) and meter anglesper diopter (MA/D).
AC/A Ratios
PCT Methodsa RHP Method
Near (33 cm)Stimulus Lens
Gradient
2 metersStimulus Lens
GradientbStimulus
HeterophoriacStimulus
HeterophoriaResponse
Heterophoria
MA/D 0.26 � 0.19 0.38 � 0.17 0.94 � 0.25 0.46 � 0.31 0.73 � 0.59PD/D 1.56 � 1.07 2.26 � 0.97 5.58 � 1.51 2.75 � 1.82 4.31 � 3.49
a PCT, prism cover test; RHP, remote haploscopic photorefractor.b Two meters chosen for more distant fixation distance to enable comparison with RHP.c Heterophoria methods involve comparison of responses between target distances.
TABLE 3.Study 2 (N � 14): mean � SD convergence in meter angles (MA) and accommodation in dioptes (D) for target demand.
DemandConvergence (MA) Accommodation (D)
Clown BEa Clown RE Clown LE Clown BE Clown RE Clown LE
0.5 0.33 � 1.61 0.54 � 1.72 0.76 � 1.67 0.33 � 0.34 0.22 � 0.67 0.47 � 0.461 1.01 � 1.58 0.75 � 1.65 0.99 � 1.55 0.43 � 0.37 0.29 � 0.65 0.47 � 0.302 2.06 � 1.56 1.14 � 1.64 1.33 � 1.64 0.87 � 0.72 0.74 � 0.36 0.86 � 0.403 2.83 � 2.06 1.60 � 1.83 1.66 � 1.74 2.16 � 0.77 1.41 � 0.88 1.56 � 0.774 3.31 � 2.32 1.64 � 1.66 1.65 � 1.73 2.09 � 1.18 1.45 � 0.84 1.74 � 0.70a BE, both eyes; RE, right eye; LE, left eye.
Accommodation and Convergence in a Natural Setting—Horwood et al. 797
Optometry and Vision Science, Vol. 78, No. 11, November 2001

was higher than that found in study 1 (study 1, 0.706; study 2,0.835). The group mean vergence after left occlusion was 56.7%(slope, 0.34) of the unoccluded total and 64.8% (slope, 0.24) afterright occlusion, again suggesting that eliminating fusion reducesthe total response by slightly less than half. A two-way analysis ofvariance testing the effects of occlusion and target distance showeda significant main effect of occlusion (F2,12 � 24.4, p � 0.001),target distance, (F4,10 � 32.03, p � 0.001), and a significantinteraction (F8,6 � 8.29, p � 0.009) with greatest effects of occlu-sion at greatest target demands.
Accommodation
The clown stimulus produced a lower accommodative responseboth monocularly and binocularly than the text, especially for thenear target (Fig. 3 and Table 3). The slope for the binocular func-tion was 0.59, and monocularly, right eye 0.40 and left eye 0.41. Atwo-way analysis of variance looking at the effects of occlusion andtarget distance on accommodative response showed significantmain effects of occlusion (F2,10 � 6.81, p � 0.014), a highlysignificant effect of distance (F4,8 � 7.33, p � 0.009), with ahighly significant interaction (F8,4 � 19.01, p � 0.006) as accom-modation was more disrupted by occlusion for the near targets.Examination of individual results showed that three subjects(21%) did not show any pupil crescents for any target on occlu-sion, and two more only produced small crescents of �2 D at the4 D demand positions, indicating almost flat accommodative re-sponses to the clown target. A further subject only produced cres-cents when fixing with one eye. In comparison, there were nosubjects with flat accommodative responses in study 1 when usingthe more accommodatively demanding target.
Accommodation and Convergence Compared
Fig. 5 illustrates the group mean convergence response plottedagainst the mean accommodative response for the clown target inthe unoccluded and both occluded conditions. Individual datapoints represent different target demands. Again, there is a stronglylinear relationship between means of accommodation and conver-gence (unoccluded, r2 � 0.90; right eye fixing, r2 � 0.98; left eyefixing, r2 � 0.90). It is noticeable from this figure how little changein accommodation occurred over the range of target demands forthis target, especially in the occluded conditions. This reflects thepoor accommodative responses to the clown target, which, never-theless, are associated with appropriate convergence responses.
Some subjects commented on difficulty in maintaining fusion at4 D, but none commented on blur for the clown target. Despitethis, the vergence response was more accurate over target distancethan the accommodative response. This suggests that the accom-modative lag was not due to lack of attention, difficulty with fixa-tion, or an artifact of our method.
Mean heterophoria stimulus RHP AC/A in study 2 was 0.40MA/D (SD � 0.25) or 2.41�/D (S.D � 1.52) fixing with theright eye and 0.37 MA/D (SD � 0.38) or 2.19�/D (SD � 2.27)fixing with the left eye. There was no significant difference betweenthese measures across studies 1 and 2 (t � 0.52, p � 0.60), sug-gesting that a similar amount of convergence was produced by bothtargets and when fixing with either eye.
In study 2, the response RHP heterophoria AC/A with theclown target fixing with the right eye was 1.10 MA/D (SD � 2.22)or 6.58�/D (SD � 13.25), and the left eye was 1.00 MA/D (SD �2.40) or 5.95�/D (SD � 14.33). A comparison of the vergenceresponse when viewing the clown vs. the text in Fig. 2 shows thatmean occluded vergence responses were slightly better with theclown target, mainly because of the two subjects who showed poorvergence response to the text target. However, the high responseAC/A ratios found with the clown target were mainly a conse-quence of the group of 5 of the 14 subjects who maintained ap-propriate vergence but had low levels of accommodation: theywere not primarily due to excessive convergence. If the subjectswho produced almost flat accommodative responses on occlusionare removed from this group, the mean response heterophoriaAC/A is right eye 0.72 MA/D (SD � 0.50) or 4.28�/D (SD �3.00) and left eye 0.87 MA/D (SD � 0.52) or 5.16�/D (SD �3.09) (Table 4), similar to the results of study 1.
BETWEEN-SUBJECT COMPARISON ACROSSTARGET TYPE (STUDIES 1 AND 2)
In both studies, there was significant interindividual variabilityin the accommodative vs. vergence response patterns. We at-tempted to categorize the different patterns. The interindividualvariability was found to be more apparent for the accommodativeresponse than for the vergence response (see standard deviations inTables 1 and 3). Three patterns of response were identified in thegroup of subjects tested in these studies, and a further pattern wasfound when experienced subjects were used during pilot trials ofthe study (Fig. 6a to d).
FIGURE 5.Study 2. Mean binocular (left chart) and occluded (right charts) conver-gence (MA) plotted against accommodation (D). Individual points repre-sent different target demands. Accom, accomodation; Conv, convergence;Resp, response.
798 Accommodation and Convergence in a Natural Setting—Horwood et al.
Optometry and Vision Science, Vol. 78, No. 11, November 2001

Subject 1
A 15-year-old female had a response AC/A ratio of 0.5 MA/D andshowed the type of response we expected to find in most subjects,which was the most common response in study 1. She accommodatedappropriately for both the binocular and monocular conditions but
showed a reduced vergence response on occlusion when fusional ver-gence was disrupted. A total of 60% of subjects in study 1 and 43% ofsubjects in study 2 fell into this group.
Subject 2
A 14-year-old female had a RHP response AC/A ratio of 0.21MA/D. This was the most typical response in study 2. Whenbinocular, this subject both accommodated and converged withinthe normal range. There was an unexpected reduction in the slopeof the monocular accommodation response. The vergence re-sponse in this subject when occluded was almost completely ab-sent. A total of 40% of participants in study 1 and 50% in study 2demonstrated a marked reduction in accommodation as well asvergence in this way.
Subject 3 (From Pilot Study)
A 40-year-old female with experience in accommodation andvergence studies showed a surprisingly good accommodative gainfor her age with a response AC/A of 1.29 MA/D. Vergence gainwas �1 for all targets. Accommodation response was appropriatein both the binocular and occluded conditions. Vergence gain washigher than expected when one eye was occluded. All the moreexperienced subjects, whether in their early twenties or approach-ing presbyopia, showed similar lack of degradation of the accom-modative responses and relatively high vergence gain on occlusion.From our pilot trials of the apparatus, this had been the responsewe had expected to find in all subjects.
Subject 4
A 15-year-old female from study 2 was unique in showing al-most no accommodative or convergence response either binocu-larly or monocularly. We initially believed that this subject was notpaying attention and so recalled her a few weeks later for repeattesting, this time using both text and clown targets. On this occa-sion, we questioned her carefully about target clarity during thetesting. She produced identical responses. She claimed that thetarget was not particularly blurred and read the text throughout the
FIGURE 6.Typical types of convergence (MA) and accommodation (D) responsesfound. Charts on the left show binocular responses, and charts on the rightshow monocular responses. Subj, subject; Accom, accomodation; Conv,convergence; Resp, response.
TABLE 4.Study 2 (N � 14): mean � SD AC/A ratios in both prism diopters per diopter of accommodation (PD/D) and meter anglesper diopter (MA/D).
PCTMethoda RHP Methodb
StimulusHeterophoria
StimulusHeterophoria
ResponseHeterophoria
Response Heterophoria Excluding“Flat” Responses
MA/DRE fixing 0.91 � 0.18 0.40 � 0.25 1.10 � 2.22 0.72 � 0.50LE fixing 0.37 � 0.38 1.00 � 2.40 0.87 � 0.52
PD/DRE fixing 5.43 � 1.09 2.41 � 1.52 6.58 � 13.25 4.28 � 3.00LE fixing 2.19 � 2.27 5.95 � 14.33 5.16 � 3.09a PCT, prism cover test; RHP, remote haploscopic photorefractor; RE, right eye; LE, left eye.b RHP AC/A tested fixing both right and left eyes.
Accommodation and Convergence in a Natural Setting—Horwood et al. 799
Optometry and Vision Science, Vol. 78, No. 11, November 2001

test. Subsequent dynamic retinoscopy of the occluded eye obtainedby the retinoscopist lying in the path of the photorefracting camerawhile the subject read the text confirmed that �1D of accommo-dation change across fixation distance had occurred. She wassymptom free and had a small exophoria and good clinical conver-gence to 6 cm. She had a RHP response AC/A ratio of 0.26 but thiswas of little relevance because there was no change in accommo-dation or convergence across target demand.
It is difficult to definitively classify responses in such a smallgroup, and further studies will be carried out to see whether morepatterns of behavior emerge. Some subjects were difficult to clas-sify, and sometimes results were different when fixing with eithereye. Of the subjects across both groups, 20 (83%) continued toaccommodate on occlusion (albeit generally by a noticeably re-duced amount). A total of 16% subjects showed a marked reduc-tion of accommodative response such that no fundal crescents werevisible at any distance. These subjects had shown clear and mea-surable crescents when binocular, so difficulty with crescent inter-pretation was not the reason for absence of a crescent. All exceptone of these subjects also stopped converging at the same time anddemonstrated a conjugate abduction of the occluded eye for thenear targets as the fixing eye adducted. Thus, there was no evidencethat the accommodative or proximal cues remaining were beingused to produce vergence.
Of the 20 subjects who continued to accommodate when oc-cluded, 10 showed some vergence response in the occluded eye.This is the expected response because accommodative and proxi-mal convergence cues are still available. The other 10 subjectsshowed an unexpected conjugate abduction of the occluded eye.This suggests that proximal and accommodative cues were notbeing used to drive vergence in these subjects.
The subjects who stopped accommodating on occlusion with anassociated drop in vergence would have a low ratio on clinicalstimulus AC/A ratio testing. Because accommodation is not mea-sured in this situation, it is assumed that it equals the change indemand across stimulus. Thus, the low convergence response, withsupposedly normal accommodation, would result in an apparentlow clinical AC/A ratio. This is often found in clinical normals.Our method revealed that the response AC/A ratio can be normalin this group, but small vergence changes are associated withequally small accommodative responses.
DISCUSSION
The results of this study suggest that the RHP method can be usedfor measuring accommodation, vergence, and AC/A ratios in adults.The mean AC/A ratio measured using RHP was comparable to thosemeasured using either heterophoria or gradient PCT methods, al-though the mean data did not reflect the wide variability found. Asfound in other studies,2, 10, 33, 35, 38 the clinical (PCT) methods ofmeasuring stimulus AC/A ratios correlated poorly with each other andshowed wide individual variation, even when means of more than onelens or fixation distance were used. Ogle and Martens2 found thatheterophoria AC/A ratios are higher than lens gradient ratios and ourresults support this. Our mean clinical values also compared well withclinical studies using PCT methods. Franceschetti and Burian7 foundthat the range of normal lens gradient values could be as low as
0.56�:1 D, and Plenty8 considers a clinical AC/A of under 5�:1 D,with no lower limit, to be normal.
In our study, there was a poor correlation between the clinicalmethods of measuring the AC/A and the RHP method. We failed tofind a correlation either within the different PCT methods or betweenthem and the stimulus RHP method in this normal group. This wasmainly due to lack of any clinically detectable change in angle whenperforming the prism cover test at different fixation distances or withdifferent lenses. The prism cover test produced heterophoria meterangle AC/A ratios close to one and lens gradient AC/A ratios close tozero in many subjects, whereas the RHP produced more intermediatevalues. This reflects greater problems with prism cover test methodsthan the RHP method. Although the dissociation of the cover testshould theoretically remove fusion and thus reduce total convergencefor near, in this group of normal subjects with small heterophorias, itfailed to do so. In a separate smaller pilot study using a clinical groupwith larger, pathological heterophorias, a correlation of r2 � 0.60 (p �0.038) was found between the RHP response AC/A and PCT hetero-phoria stimulus AC/A.
Although we found measurable and, on average, normal AC/Aratios in our groups of subjects, the individual variations evenamong these normal subjects make the practical use of the meanvalues we found limited. It also brings even further into questionwhether any stimulus AC/A ratio is useful as a clinical measurewhen normal responses are so variable and accurate accommoda-tion cannot be assumed (especially in children). Thus, some sub-jects had high response AC/A ratios (up to 5.3 MA/D) because theyhad a reduced accommodative response but an appropriate ver-gence response. The small amount of accommodation producedwas, therefore, associated with a relatively large amount of conver-gence. It is clear from these subjects that even a response AC/Aratio in such near/distance comparisons can only partly explainhow an individual functions.
One possible reason for the individual difference in AC/A ratiosis individual differences in the Hirschberg ratio, i.e., a high AC/Aratio might be due to an overestimation of the Hirschberg ratioleading to an overestimation of the amount of convergence pro-duced. It would have been ideal to have based our calculations onindividual, not mean, Hirschberg ratios, but these data were notavailable. We tested the validity of our AC/A mean ratios by recal-culating the AC/A ratio for a group of 12 subjects for whom indi-vidual Hirschberg ratios were available. The mean unadjustedAC/A ratio for these subjects was 0.81 MA/D (SD � 0.84 MA/D),and the adjusted AC/A ratio was 0.77 MA/D (SD � 0.67 MA/D).The adjusted and unadjusted AC/A ratios were not significantlydifferent from each other (t � 0.109, p � 0.91), although variancewas reduced. Despite this, it is still possible that the subjects withthe high AC/A ratios (those producing large vergence responses incomparison to their accommodation) still had different Hirsch-berg ratios. Of the three subjects with the highest AC/A ratios(1.91, 3.18, and 1.33), the Hirschberg ratios were measured at9.99, 11.05, and 14.85°/mm, respectively, representing the ex-treme highest and lowest Hirschberg values found within thewhole group. Thus, there is no systematic overestimation of theHirschberg ratio in these subjects. A low AC/A ratio is usually theresult of a small vergence response in relation to accommodation.It would require a large individual difference in the Hirschberg
800 Accommodation and Convergence in a Natural Setting—Horwood et al.
Optometry and Vision Science, Vol. 78, No. 11, November 2001

ratio to alter the AC/A ratio in these cases. Such large individualdifferences were not found in the subjects with low AC/A ratios.
Having demonstrated that RHP can be used to measure accom-modation and vergence, the main finding of this paper is that whenaccommodation is not strictly enforced or required, disparity cuesmay play a greater part in driving accommodation than occurswhen accommodation is accurate. When the text target, whichrequires accurate accommodation to read the text, was used, occlu-sion did not significantly reduce the mean accommodative re-sponse. However, when using a target that can be recognized with-out accurate accommodation (the clown), occlusion resulted in areduction in accommodation.
The clown target was designed to be attractive and interesting toinfants, containing a range of colors, spatial frequencies, and contrastand incorporating alternating elements. At the outset of this study, wewere interested in ascertaining the adult response to this target in ourapparatus so that our, and other, photorefractive infant studies ofaccommodation and AC/A development could be compared. Thefindings of this study suggest that the nature of the targets and testingregimes necessary for infant studies make comparisons with highlycontrolled laboratory studies problematic. They also further point tothe necessity for more work in the area of responses to more non-optimal and naturalistic targets in naïve subjects.
Within the population tested, there was a wide variability inaccommodative and vergence responses. Individual subjectsshowed categorically different accommodative and vergence be-haviors in response to change in target demand and/or occlusion.This was especially true for the clown target. None of our subjects,however, complained of blur or diplopia for this target, althoughaccommodation and vergence responses were not always accurate.These findings have implications for real-life viewing conditionsbecause the low luminance and undemanding targets that we usedhere are more common in the everyday environment than thedetailed and demanding targets used in most laboratory studies.Because there were differences in color content between the textand clown targets, this might have been responsible for the loweraccommodative response found with the clown as isoluminantcolor contrast has been found to be an insufficient stimulus toaccommodation.39 Our clown target, however, did not containany such isoluminant edges, and all the color edges also containedsignificant brightness contrast.
Another possibility is that we obtained differences in accommo-dative response between the clown and the text targets because theclown target was less detailed. Although accommodation has beenfound to be most efficient at intermediate, rather than high, spatialfrequencies17, 40, 41 and subjective depth of focus does not varywith target size,42 our more detailed text target produced moreaccurate accommodative responses than the clown. It would there-fore seem unlikely that spatial contrast per se is the significantdifference. It is, however, possible that if clarity is not necessary forobject recognition or task performance, the subjects are happy totolerate a blurred image until clarity is required to perform thereading task for the text target.
It could be argued that the low light levels of our apparatus wereresponsible for the poor accommodative response in some subjects.These low light levels were necessary for undilated photorefraction.Johnson43 found that accommodative responses could be reducedat light levels similar to ours, although only one of their four
participants showed significantly reduced accommodation accu-racy. Alpern and David44 also found that accommodation coulddecline at low light levels that were similar to ours. However,Arumi et al.45 and Campbell46 found that lower luminances thanours were necessary to produce markedly reduced responses. It ispossible that the dim lighting conditions of our apparatus mightexaggerate the influence of vergence accommodation,47 especiallyin some subjects, but this is certainly not a universal effect. It isimportant to note that the luminance of both targets was similarlylow, and yet the accommodative response was lower for the clownthan for the text targets. In addition, our subjects who failed toaccommodate when occluded produced better accommodative re-sponses when binocular. In study 1, the target was switched offbetween presentations, whereas in study 2, the target was illumi-nated the whole time, providing looming cues to proximal accom-modation. The additional cue might have been expected to coun-teract the effects of using a less demanding target and of occlusionon the accommodative response, but this was not found.
During the pilot experiment, observers experienced in accommo-dation and vergence experiments, presumably “doing their best to behelpful,” produced different responses from our subsequent naïve sub-jects despite the same instruction set, e.g., Fig. 6c. Accommodationwas generally more accurate in the experienced group and did notreduce on occlusion despite the low luminance levels. Interestingly,vergences were also much less disrupted by occlusion, suggesting thateither accommodative vergence or voluntary effort was more of aninfluence in these experienced observers.
Our naïve subjects’ responses to different targets are in contra-diction to the findings of a study by Ciuffreda et al.,48 who foundrobust accommodative responses to a wide range of targets. Theyalso found consistent over-accommodation to a range of mul-tichromatic targets, which is similar to the responses we found inour pilot group of more experienced subjects. However, their sub-jects were optometry students and staff. In his major review article,Ciuffreda49 emphasized the importance of the instruction set onaccommodative response. Many optometric studies test optometrystudents and researchers: subjects who are likely to have at leastsome subjective awareness of their own response levels, whateverthe instruction set, and may be manipulating their responses viahigher processing, albeit unconsciously. Other studies that aim tostudy “reflex” accommodation in naïve subjects use considerablylonger testing sessions, training periods, and more specific instruc-tion sets than we used here.17, 18 We deliberately chose a minimalinstruction set and completely naïve subjects so that our infantstudies could be compared. Our conditions were as close as possi-ble to “casual” viewing (which is all that is possible in infant re-search) and lasted only a few minutes. We contend that usingexperienced subjects or long practice periods could result in higher,stable, but environmentally unrepresentative responses. This find-ing is particularly important when infant and clinical studies arecompared with carefully controlled laboratory research.
Many orthoptic exercises for convergence insufficiency, hetero-phoria, and intermittent heterotropia teach manipulation of ver-gences first by voluntary manipulation of accommodation.5 Withpractice, this becomes unconscious, and accommodation can even-tually be used normally while vergences remain relatively exerted orrelaxed compared to pretreatment levels. This alone suggests thatlong-term practice effects, which are difficult to quantify, can
Accommodation and Convergence in a Natural Setting—Horwood et al. 801
Optometry and Vision Science, Vol. 78, No. 11, November 2001

change the way that an individual uses cues to achieve a vergenceand accommodative position and may account for differences be-tween naïve and experienced subjects.
This apparent variability and flexibility of the accommodativeresponse and AC/A relationship are reflected in clinical practiceand might explain differences found within subjects across testingmethods or between subjects in responses to treatment. One ex-ample is that some patients with esotropias are found to respond toconvex lenses, whereas others do not, despite similar refractiveerror and clinical stimulus AC/A ratio. The possible influence ofluminance does not detract from our argument that accommoda-tion can be used in a varying relationship with vergence dependenton target demand. It is particularly relevant in young childrenwhere larger, nontext targets are the norm for clinical testing. Ourresults also suggest that the effect of dissociation on the angle ofheterophoria may not be just the result of loss of fusional vergence,but in many cases, where accommodation is reduced on occlusion,may also reflect loss of accommodative vergence as well. If this isthe case, and if our findings apply in higher luminances, it raises theimportance of proximal (or voluntary) vergences in the productionof any distance/near change in angle. Kushner and Morton50 dem-onstrated that some intermittent exotropes have strong proximalvergence. It is possible that proximal or voluntary vergences may bemore important for subjects with poor binocular or uniocular ac-commodative gain or abnormally large deviations.
Other researchers have published data showing individuallyvariable responses in much more carefully controlled studies,17 andour minimal control methods might well account for the increasedvariability in response we found here in our completely naïvegroups. Leat and Gargon,34 in a dynamic retinoscopy study of asimilarly naïve population, found wide variations in accommoda-tive response and considered that “real variation between individ-uals is not so much due to a poor technique as real variations inaccommodative response between individuals” (p. 382). Manochaand Brooks33 found that an occluder appears to inhibit accommo-dation. This, sometimes dramatic, reduction in accommodationwhen fusion is disrupted indicates that as Fincham and Walton12
suggested, convergence accommodation may be a major input intothe total accommodative response for many subjects. Wick andCurrie51 also considered that convergence accommodation mayaccount for much of the near accommodative response in manysubjects. Our data support this view, especially with a stimulus thatdoes not require, or produce, accurate accommodation.
The dramatic loss of accommodation response once vergence isdisrupted by occlusion suggests that when subjects are binocular,fusional vergence can be a major drive to accommodation via theCA/C relationship. Although this effect could have been exagger-ated by the low light levels, the subjects were still able to read theprint throughout study 1 and did not complain of an inability tofocus during study 2, despite what must have been considerableoptical blur. In subthreshold conditions, subjects may prefer tooperate at the limits of their depth of field or at a level of blur thatis just sufficient to permit interpretation of the target rather thanuse a clear image at all times. This slight blur may also have theadvantage of enlarging Panum’s Area so that any increased imagedisparity is less noticeable.
Although not directly addressed here, it is possible that theweighting of the use of blur, disparity, and proximal and voluntary
cues may vary between subjects as an idiosyncratic style or withinsubjects depending on task demands and cue availability. Mostexperimental methods control for as much as possible while ma-nipulating one aspect. Our method controlled for little and re-moved one aspect, which may account for our results.
It could be argued that these variable results to non-optimaltargets reflect poor experimental technique, but in other scientificfields, some proponents view variability as vital to the functioningof a dynamic system. Thelen and Smith52 studied the developmentof motor behavior in many domains and noted that when “we lookat adults doing motor or cognitive tasks under controlled labora-tory conditions, in standard environments, we tend to see thestable and predictable features of their behavior. The next step hasoften been to ascribe this stability to executive agents which encodethe details of the action. When subjects are young, unpracticed, orallowed to act more spontaneously and naturally, much of thisstability is no longer apparent, or at least the range of variabilityaround a stable mode is greatly expanded.” In addition, they statethat the “key to biological co-ordination may lie in more looselyassembled patterns, which provide both flexibility and richness toassemble and reassemble the components in adaptive modes.”
This viewpoint could well be an alternative explanation for ourresults. Our results, in the context of this theory, suggest that atleast at low luminance, accommodation and convergence can beinaccurate for much of the time without any subjective awarenesson the part of the subject and may not be related in such fixedAC/A and CA/C relationships as previously thought.
We feel that the extreme variability of responses found shouldnot be dismissed as excessive noise in a poorly controlled experi-mental method, but serve as indication of an inherently flexibleand possibly idiosyncratic system worthy of further investigation inother non-optimal situations. This has particular relevance whencomparing adult with infant studies, where accurate accommoda-tion can never be guaranteed and low light levels are used forphotorefraction.
CONCLUSIONS
This study has shown that the remote haploscopic photorefrac-tor can be used to assess binocular and monocular accommodationand convergence in adults, and that stimulus and response AC/Aratios can be obtained that are comparable to previously reportedmeasures in groups of normal adults. However, using dim andundemanding targets and this method of response assessment,there were wide idiosyncratic variations in the amount of accom-modation and vergence actually exerted. In particular, there was adramatic reduction in accommodation exerted by many subjectsunder monocular conditions. It suggests that although a normalAC/A ratio may be demonstrable, many subjects generally performless demanding near tasks under conditions of significant blur,especially if monocular. Because vergences are generally accurate,mean stimulus AC/A ratios do not vary with target type. However,because accommodation can be reduced for stimuli that can berecognized, despite some blur while vergence remains unaffected,the response AC/A ratio may be much higher for less accommoda-tively demanding targets.
For some subjects and targets, accommodative convergence
802 Accommodation and Convergence in a Natural Setting—Horwood et al.
Optometry and Vision Science, Vol. 78, No. 11, November 2001

might play only a minor role in the total near response, particularlywhere light levels are low. Convergence accommodation mightplay a far more important part in driving accommodation, withproximal/voluntary input also being a more important additionalelement, than previously thought. Care should therefore be takenwhen comparing highly controlled laboratory studies on experi-enced observers with naïve and infant studies that generally de-mand less stringent accommodation.
ACKNOWLEDGMENTS
Supported by U.K. Medical Research Council grant G9608874N. We thankJohn Bullinaria for modeling our data, Israel Abramov and Louise Hainlinefor equipment design, and all the participants in the study. We are grateful forthe helpful comments of the anonymous referees on the first version of thispaper, whose suggestions we have included in this version.
Received February 23, 2000; revision received June 15, 2001.
REFERENCES
1. Flom M. On the relationship between accommodation and accom-modative convergence. II: stability. Am J Optom Arch Am AcadOptom 1960;37:517–23.
2. Ogle K, Martens T. On the accommodative convergence and theproximal convergence. Arch Ophthalmol 1957;128:702–15.
3. Sloan L, Sears M, Jablonski M. Convergence-accommodation rela-tionships. Arch Ophthalmol 1960;63:283.
4. Breinin G. Relationship between accommodation and convergence.Trans Am Acad Ophthalmol 1961;61:375–82.
5. Mein J, Trimble R. Diagnosis and Management of Ocular MotilityDisorders, 2nd ed. Oxford: Blackwell Scientific, 1991.
6. Burian HM, von Noorden GK. Binocular Vision and Ocular Motility:Theory and Management of Strabismus. St Louis, MO: Mosby, 1974.
7. Franceschetti AT, Burian HM. Gradient accommodative conver-gence-accommodative ratio in families with and without esotropia.Am J Ophthalmol 1970;70:558–62.
8. Plenty J. A new classification for intermittent exotropia. Br OrthopticJ 1988;45:19–22.
9. Ludwig IH, Parks MM, Getson PR, Kammerman LA. Rate of dete-rioration in accommodative esotropia correlated to the AC/A rela-tionship. J Pediatr Ophthalmol Strabismus 1988;25:8–12.
10. Havertape SA, Cruz OA, Miyazaki EA. Comparison of methods fordetermining the AC/A ratio in accommodative esotropia. J PediatrOphthalmol Strabismus 1999;36:178–83.
11. Hokoda SC, Rosenfield M, Ciuffreda KJ. Proximal vergence and age.Optom Vis Sci 1991;68:168–72.
12. Fincham E, Walton J. The reciprocal actions of accommodation andvergence. J Physiol 1957;137:488–508.
13. McLin LN Jr, Schor CM, Kruger PB. Changing size (looming) as astimulus to accommodation and vergence. Vision Res 1988;28:883–98.
14. Rosenfield M, Ciuffreda KJ, Hung GK, Gilmartin B. Tonicaccommodation: a review. I: basic aspects. Ophthalmic Physiol Opt1993;13:266–84.
15. Currie DC, Manny RE. The development of accommodation. VisionRes 1997;37:1525–33.
16. Judge SJ. How is binocularity maintained during convergence anddivergence? Eye 1996;10:172–6.
17. Mathews S, Kruger PB. Spatiotemporal transfer function of humanaccommodation. Vision Res 1994;34:1965–80.
18. Owens DA. A comparison of accommodative responsiveness and con-trast sensitivity for sinusoidal gratings. Vision Res 1980;20:159–67.
19. Charman WN, Tucker J. Dependence of accommodation response
on the spatial frequency spectrum of the observed object. Vision Res1977;17:129–39.
20. Ciuffreda KJ, Hokoda SC. Effect of instruction and higher levelcontrol on the accommodative response spatial frequency profile.Ophthalmic Physiol Opt 1985;5:221–3.
21. Ciuffreda K. Accommodation and its anomalies. In: Charman WN, ed.Visual Optics and Instrumentation. London: Macmillan, 1991:231–79.
22. Hung GK. Quantitative analysis of the accommodative convergenceto accommodation ratio: linear and nonlinear static models. IEEETrans Biomed Eng 1997;44:306–16.
23. Lyle TK, Wybar KC. Lyle and Jackson’s Practical Orthoptics in theTreatment of Squint. London: Lewis, 1970.
24. Hainline L, Riddell P, Grose-Fifer J, Abramov I. Development of accom-modation and convergence in infancy. Behav Brain Res 1992;49:33–50.
25. Abramov I, Hainline L, Duckman RH. Screening infant vision withparaxial photorefraction. Optom Vis Sci 1990;67:538–45.
26. Gwiazda J, Grice K, Thorn F. Response AC/A ratios are elevated inmyopic children. Ophthalmic Physiol Opt 1999;19:173–9.
27. Bishop CM. Neural Networks for Pattern Recognition. Oxford: Ox-ford University Press, 1995.
28. Riddell PM, Hainline L, Abramov I. Calibration of the Hirschbergtest in human infants. Invest Ophthalmol Vis Sci 1994;35:538–43.
29. Jones R, Eskridge JB. The Hirschberg test: a re-evaluation. Am JOptom Arch Am Acad Optom 1970;47:105–14.
30. Carter AJ, Roth N. Axial length and the Hirschberg test. Am J OptomPhysiol Opt 1978;55:361–4.
31. Eskridge JB, Perrigin DM, Leach NE. The Hirschberg test: correlationwith corneal radius and axial length. Optom Vis Sci 1990;67:243–7.
32. Paliaga GP. Linear strabismometric methods. Binoc Vis Q 1992;7:139–54.
33. Manocha KM, Brooks SE. Assessment of accommodativeconvergence: accommodation ratio without interrupting fixation:comparison with gradient and heterophoria methods. Invest OphthVis Sci 1995;36:S12.
34. Leat SJ, Gargon JL. Accommodative response in children and youngadults using dynamic retinoscopy. Ophthalmic Physiol Opt 1996;16:375–84.
35. Heyes K, Whittle J. The use of fixation disparity for determiningAC/A ratios. Br Orthoptic J 1998;55:53–6.
36. Owens DA, Wolfe JM. Accommodation for flickering stimuli. Oph-thalmic Physiol Opt 1985;5:291–6.
37. Flitcroft DI. Accommodation and flicker: evidence of a role for tem-poral cues in accommodation control? Ophthalmic Physiol Opt1991;11:81–90.
38. Chin NB, Breinin GM. Ratio of accommodative convergence toaccommodation: accommodative strabismus. Arch Ophthalmol1967;77:752–6.
39. Wolfe JM, Owens DA. Is accommodation colorblind? Focusingchromatic contours. Perception 1981;10:53–62.
40. Owens DA. A comparison of accommodative responsiveness and con-trast sensitivity for sinusoidal gratings. Vision Res 1980;20:159–67.
41. Mathews S. Accommodation and the third spatial harmonic. OptomVis Sci 1998;75:450–8.
42. Atchison DA, Charman WN, Woods RL. Subjective depth-of-focusof the eye. Optom Vis Sci 1997;74:511–20.
43. Johnson CA. Effects of luminance and stimulus distance on accom-modation and visual resolution. J Opt Soc Am 1976;66:138–42.
44. Alpern M, David H. Effects of luminance quantity on accommoda-tion of the eyes. Indus Med Surg 1958;27:551–5.
45. Arumi P, Chauhan K, Charman WN. Accommodation and acuityunder night-driving illumination levels. Ophthalmic Physiol Opt1997;17:291–9.
46. Campbell F. Twilight myopia. J Opt Soc Am 1953;43:925–6.
Accommodation and Convergence in a Natural Setting—Horwood et al. 803
Optometry and Vision Science, Vol. 78, No. 11, November 2001

47. Jiang BC, Gish KW, Leibowitz HW. Effect of luminance on therelation between accommodation and convergence. Optom Vis Sci1991;68:220–5.
48. Ciuffreda KJ, Rosenfield M, Rosen J, Azimi A, Ong E. Accommoda-tive responses to naturalistic stimuli. Ophthalmic Physiol Opt 1990;10:168–74.
49. Ciuffreda KJ. The Glenn A. Fry invited lecture: accommodation togratings and more naturalistic stimuli. Optom Vis Sci 1991;68:243–60.
50. Kushner BJ, Morton GV. Distance/near differences in intermittentexotropia. Arch Ophthalmol 1998;116:478–86.
51. Wick B, Currie D. Convergence accommodation: laboratory andclinical evaluation. Optom Vis Sci 1991;68:226–31.
52. Thelen E, Smith LB. A Dynamic Systems Approach to the Develop-ment of Cognition and Action. Cambridge, MA: MIT Press, 1994.
A. M. HorwoodInfant Vision Laboratory
Psychology DepartmentHarry Pitt Building
University of ReadingPO Box 238
RG6 6ALUnited Kingdom
e-mail: [email protected]
804 Accommodation and Convergence in a Natural Setting—Horwood et al.
Optometry and Vision Science, Vol. 78, No. 11, November 2001




















