
Variable Patterns of Axonal Projections of Sensory Neurons in the Mouse Vomeronasal System
10
Cell, Vol. 97, 199–208, April 16, 1999, Copyright 1999 by Cell Press Variable Patterns of Axonal Projections of Sensory Neurons in the Mouse Vomeronasal System The functional and anatomical dichotomy between the main and vomeronasal (or accessory) olfactory sys- tems is further reflected at the level of the molecules Ivan Rodriguez, Paul Feinstein, and Peter Mombaerts* The Rockefeller University New York, New York 10021 that serve as receptors, or putative receptors, for their respective sensory stimuli. In the main olfactory system, odorant receptor (OR) genes encode seven-transmem- brane proteins and are members of a multigene family Summary that may comprise as many as 1000 genes in rat and mouse (Buck and Axel, 1991; for review, Mombaerts, The vomeronasal system mediates pheromonal effects 1999). Recently, the first ligand–receptor relationships in mammals. We have employed gene targeting tech- have been described for vertebrate ORs (Krautwurst et nology to introduce mutations in a putative pheromone al., 1998; Zhao et al., 1998). In the accessory olfactory receptor gene, VR i 2, in the germline of mice. By gener- system, two families of genes encoding seven-trans- ating alleles differentially tagged with the histological membrane proteins have been proposed to encode markers taulacZ and tauGFP, we show that VR i 2 is pheromone receptors. The first family of vomeronasal monoallelically expressed in a given neuron. Axons receptor (VR) genes consists of 30–100 genes that are of VR i 2-expressing neurons converge onto numerous expressed selectively in VSNs of the apical zone of the glomeruli in the accessory olfactory bulb. The pattern epithelium of the VNO (Dulac and Axel, 1995; Saito et of axonal projections is complex and variable. This al., 1998). The second family of VR genes comprises wiring diagram is substantially different from that of 30–140 genes that are expressed in VSNs of the basal the main olfactory system. The projection pattern is zone (Herrada and Dulac, 1997; Matsunami and Buck, disrupted by deleting the coding region of VR i 2, but an 1997; Ryba and Tirindelli, 1997). There are no conserved unrelated seven-transmembrane protein, the odorant motifs between the two families of VRs, and VRs have receptor M71, can partially substitute for VR i 2. no sequence homology with ORs. Neurons in the apical and basal zones express G protein subunits, respec- Introduction tively, G ai2 and G ao (Halpern et al., 1995; Berghard and Buck, 1996), and project their axons to distinct regions The olfactory system provides sensory information about in the mouse AOB (Jia and Halpern, 1996). It is not the chemical composition of the external world. Olfac- known whether these G proteins are involved in signal tory chemoreception initiates in mammals at the level transduction of pheromonal stimuli; they serve as useful of sensory neurons that are located in the main olfactory molecular markers whose expression tends to correlate epithelium (MOE) and the epithelium of the vomeronasal with that of the two families of VR genes. The VR genes organ (VNO) (Farbman, 1992). The MOE (Buck, 1996) encode putative pheromone receptors; there is no evi- mediates the detection of volatile odorants. The VNO dence that any of these molecules is a receptor for a pheromone. Because few mammalian pheromones have (Halpern, 1987) mediates mainly the detection of nonvol- been identified at the molecular level (Sorensen, 1996), atile odorants, such as pheromones. These are chemical ligand–receptor interactions are difficult to define. signals that provide information about gender, domi- For the mouse OR genes I7 and I54, expression has nance, and reproductive status between individuals of been shown to derive mainly, but perhaps not exclu- the same species (Karlson and Lu ¨ scher, 1959; Wilson, sively, from one allele (Chess et al., 1994). The generality, 1963; Keverne, 1983; Sorensen, 1996). They elicit in the mechanisms, and significance of monoallelic expres- recipients innate and stereotyped reproductive and so- sion of OR genes have remained unclear (Chess, 1998a). cial behaviors, along with profound neuroendocrine and In rat and mouse, OSNs expressing a given OR gene are physiological changes. dispersed within one of four zones in the MOE (Ressler et Each olfactory sensory neuron (OSN) projects a single al., 1993; Vassar et al., 1993). Within a zone, a punctate axon to the main olfactory bulb (MOB) into globose expression pattern is observed, without apparent orga- structures termed glomeruli. Axons of OSNs synapse nization. VSNs expressing a given VR gene are also there with the dendrites of second-order neurons, which dispersed in a punctate fashion, within either the G ai2 or project their axons to the olfactory cortex and other the G ao zone of the vomeronasal epithelium (Tirindelli et regions of the brain. There are approximately 1800 glo- al., 1998). meruli per mouse MOB (Royet et al., 1988). Vomeronasal Molecular studies relying on ultrasensitive in situ hy- sensory neurons (VSNs) project their axons to the acces- bridization studies provided the first but indirect evi- sory olfactory bulb (AOB). Axodendritic synapses are dence that glomeruli may serve as common targets for also made within glomeruli. The boundaries of these some OSNs that express a given OR (Ressler et al., glomeruli are generally difficult to define, precluding pre- 1994; Vassar et al., 1994). Subsequent genetic studies cise counting (Meisami and Bhatnagar, 1998). The out- demonstrated that axons of all OSNs expressing a given put neurons of the AOB relay information to the amyg- OR converge onto defined glomeruli (Mombaerts et al., dala and hypothalamus, thus bypassing the cortex. 1996a). Our genetic approach was designed to image axons of individual neurons expressing a particular OR * To whom correspondence should be addressed (e-mail: pm@ rockvax.rockefeller.edu). (Mombaerts, 1996). The method was based on gene
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Variable Patterns of Axonal Projections of Sensory Neurons in the Mouse Vomeronasal System

Cell, Vol. 97, 199–208, April 16, 1999, Copyright 1999 by Cell Press
Variable Patterns of Axonal Projectionsof Sensory Neurons in the Mouse Vomeronasal System
The functional and anatomical dichotomy betweenthe main and vomeronasal (or accessory) olfactory sys-tems is further reflected at the level of the molecules
Ivan Rodriguez, Paul Feinstein,and Peter Mombaerts*The Rockefeller UniversityNew York, New York 10021 that serve as receptors, or putative receptors, for their
respective sensory stimuli. In the main olfactory system,odorant receptor (OR) genes encode seven-transmem-brane proteins and are members of a multigene familySummarythat may comprise as many as 1000 genes in rat andmouse (Buck and Axel, 1991; for review, Mombaerts,The vomeronasal system mediates pheromonal effects1999). Recently, the first ligand–receptor relationshipsin mammals. We have employed gene targeting tech-have been described for vertebrate ORs (Krautwurst etnology to introduce mutations in a putative pheromoneal., 1998; Zhao et al., 1998). In the accessory olfactoryreceptor gene, VRi2, in the germline of mice. By gener-system, two families of genes encoding seven-trans-ating alleles differentially tagged with the histologicalmembrane proteins have been proposed to encodemarkers taulacZ and tauGFP, we show that VRi2 ispheromone receptors. The first family of vomeronasalmonoallelically expressed in a given neuron. Axonsreceptor (VR) genes consists of 30–100 genes that areof VRi2-expressing neurons converge onto numerousexpressed selectively in VSNs of the apical zone of theglomeruli in the accessory olfactory bulb. The patternepithelium of the VNO (Dulac and Axel, 1995; Saito etof axonal projections is complex and variable. Thisal., 1998). The second family of VR genes compriseswiring diagram is substantially different from that of30–140 genes that are expressed in VSNs of the basalthe main olfactory system. The projection pattern iszone (Herrada and Dulac, 1997; Matsunami and Buck,disrupted by deleting the coding region of VRi2, but an1997; Ryba and Tirindelli, 1997). There are no conservedunrelated seven-transmembrane protein, the odorantmotifs between the two families of VRs, and VRs havereceptor M71, can partially substitute for VRi2.no sequence homology with ORs. Neurons in the apicaland basal zones express G protein subunits, respec-
Introduction tively, Gai2 and Gao (Halpern et al., 1995; Berghard andBuck, 1996), and project their axons to distinct regions
The olfactory system provides sensory information about in the mouse AOB (Jia and Halpern, 1996). It is notthe chemical composition of the external world. Olfac- known whether these G proteins are involved in signaltory chemoreception initiates in mammals at the level transduction of pheromonal stimuli; they serve as usefulof sensory neurons that are located in the main olfactory molecular markers whose expression tends to correlateepithelium (MOE) and the epithelium of the vomeronasal with that of the two families of VR genes. The VR genesorgan (VNO) (Farbman, 1992). The MOE (Buck, 1996) encode putative pheromone receptors; there is no evi-mediates the detection of volatile odorants. The VNO dence that any of these molecules is a receptor for a
pheromone. Because few mammalian pheromones have(Halpern, 1987) mediates mainly the detection of nonvol-been identified at the molecular level (Sorensen, 1996),atile odorants, such as pheromones. These are chemicalligand–receptor interactions are difficult to define.signals that provide information about gender, domi-
For the mouse OR genes I7 and I54, expression hasnance, and reproductive status between individuals ofbeen shown to derive mainly, but perhaps not exclu-the same species (Karlson and Luscher, 1959; Wilson,sively, from one allele (Chess et al., 1994). The generality,1963; Keverne, 1983; Sorensen, 1996). They elicit in themechanisms, and significance of monoallelic expres-recipients innate and stereotyped reproductive and so-sion of OR genes have remained unclear (Chess, 1998a).cial behaviors, along with profound neuroendocrine andIn rat and mouse, OSNs expressing a given OR gene arephysiological changes.dispersed within one of four zones in the MOE (Ressler etEach olfactory sensory neuron (OSN) projects a singleal., 1993; Vassar et al., 1993). Within a zone, a punctateaxon to the main olfactory bulb (MOB) into globoseexpression pattern is observed, without apparent orga-structures termed glomeruli. Axons of OSNs synapsenization. VSNs expressing a given VR gene are alsothere with the dendrites of second-order neurons, whichdispersed in a punctate fashion, within either the Gai2 orproject their axons to the olfactory cortex and otherthe Gao zone of the vomeronasal epithelium (Tirindelli etregions of the brain. There are approximately 1800 glo-al., 1998).meruli per mouse MOB (Royet et al., 1988). Vomeronasal
Molecular studies relying on ultrasensitive in situ hy-sensory neurons (VSNs) project their axons to the acces-bridization studies provided the first but indirect evi-sory olfactory bulb (AOB). Axodendritic synapses aredence that glomeruli may serve as common targets foralso made within glomeruli. The boundaries of thesesome OSNs that express a given OR (Ressler et al.,glomeruli are generally difficult to define, precluding pre-1994; Vassar et al., 1994). Subsequent genetic studiescise counting (Meisami and Bhatnagar, 1998). The out-demonstrated that axons of all OSNs expressing a givenput neurons of the AOB relay information to the amyg-OR converge onto defined glomeruli (Mombaerts et al.,dala and hypothalamus, thus bypassing the cortex.1996a). Our genetic approach was designed to imageaxons of individual neurons expressing a particular OR* To whom correspondence should be addressed (e-mail: pm@
rockvax.rockefeller.edu). (Mombaerts, 1996). The method was based on gene

Cell200
targeting in embryonic stem cells and the histochemicalmarker taulacZ (Callahan and Thomas, 1994). We modi-fied the OR gene P2 in the germline of mice such thatP2-expressing OSNs can be stained blue along theirentire length by virtue of their expression of taulacZ. Inthese mice, labeled axons converge to a pair of glomeruliin the MOB, which reside at stereotyped locations. Addi-tional genetic experiments have fueled the notion thatthe ORs are involved in the wiring of the main olfactorysystem, but they cannot be the only determinant (Mom-baerts et al., 1996a; Wang et al., 1998). The visualizationof this wiring diagram has confirmed and extended thenotion that odorant quality is encoded by a specificcombination of activated glomeruli. Likewise, our under-standing of the logic of pheromone encoding shouldbenefit from a precise description of the wiring diagram Figure 1. Targeted Mutagenesis of the VRi2 Locusof the vomeronasal system. (a) Wild-type VRi2 locus. The black boxes represent the noncoding
Here, we have visualized the topography of sensory regions. The coding region is in yellow. Restriction sites are indi-cated for BamHI (B) and HindIII (H). “ATG” is the start codon; “TGA”projections in the vomeronasal system by means of ais the stop codon.similar, but technically improved, genetic analysis. We(b) VRi2-IRES-taulacZ-LNL targeting vector. The white box labeledhave cloned VRi2, a mouse VR gene expressed in the“i” represents the IRES sequence, the blue box represents the cod-
apical, Gai2-positive zone, and created four alleles by ing sequence of taulacZ, and the gray box labeled “neo” representstargeted mutagenesis. Development of a second histo- the neo-selectable marker LNL flanked by loxP sites (indicated bylogical marker, a fusion between tau and the green fluo- red triangles).
(c) VRi2 locus after homologous recombination with the VRi2-IRES-rescent protein (GFP), greatly enhanced our ability totaulacZ-LNL targeting vector. The probe used to detect homologousimage gene expression and axonal projections.recombination in Southern blots is represented as a horizontal baron the right.
Results (d) VRi2 locus after Cre-mediated excision of the neo cassette, whichleaves a single loxP site behind.(e) VRi2-IRES-tauGFP mutation, after Cre recombination.The Vomeronasal Receptor Gene VRi2(f) (DVRi2) GFP-IRES-taulacZ mutation, after Cre recombination.We isolated several mouse VR genes of the Gai2 type by(g) M71→VRi2-IRES-taulacZ mutation, after Cre recombination.cross-hybridization to probes derived from the coding
region of rat VR genes (Dulac and Axel, 1995). We fo-cused on a particular VR gene that we term VRi2. The developed another histological axonal marker, tauGFPdeduced amino acid sequence of VRi2 (GenBank acces- (see also Brand, 1995). This marker combines the intrin-sion number AF132114) has 77% identity with that of sic green fluorescent properties of GFP (Chalfie et al.,rat VN5. Southern hybridization of mouse genomic DNA 1994; Tsien, 1998) with the microtubule-binding proper-with a VRi2 probe revealed six cross-hybridizing bands ties of the tau protein (Butner and Kirschner, 1991), as-at high stringency, indicating that VRi2 is a member suring that the fluorescent label will be exported downof a small subfamily (data not shown); similar to the the axons to the axon terminals of the VSNs that expressorganization of the OR gene superfamily (Buck and Axel, it. The design of the VRi2-IRES-tauGFP mutation (VG)1991), VR genes can be grouped in many small subfami- mirrors that of the VL mutation, except for the replace-lies that are operationally defined by cross-hybridization ment of the lacZ coding sequence with that of GFP(Dulac and Axel, 1995). In situ hybridization of sections (Figure 1e). Third, to examine whether expression of VRi2through the VNO with a riboprobe for VRi2 revealed a is a determinant of the pattern of axonal projections, wepunctate expression pattern within the apical zone of engineered a deletion allele of the VRi2 gene, termed thethe epithelium (data not shown), demonstrating that VRi2 (DVRi2) GFP-IRES-taulacZ mutation (DV). The intronlessor members of its subfamily are expressed genes. The coding region is excised from start to stop codon andexpression of the VRi2 gene specifically was shown by replaced with a GFP-IRES-taulacZ cassette (Figure 1f).59 RACE and 39 RACE on mRNA isolated from VNO. Inclusion of the GFP coding sequence 59 to the IRESGenomic sequencing revealed that the gene consists of allows for direct comparison with the VL mutation, intwo exons, with the coding region entirely included in which the VRi2 coding sequence precedes the IRES.the second exon (Figure 1a). The VRi2 gene was mapped Fourth, to determine whether other seven-transmem-with a mouse radiation hybrid panel next to marker brane proteins can substitute for VRi2, we replacedD6Mit261 on chromosome 6. the coding region of VRi2 with that of the OR gene
M71, thus generating a “swap.” In mice carrying theM71→VRi2-IRES-taulacZ mutation (MV), neurons thatTargeted Mutagenesis of the VRi2 Locus
First, to visualize the topography of VSN axonal projec- transcribe this VR locus instead express an OR (Figure1g). The consequence of this misexpression on the to-tions, we generated the VRi2-IRES-taulacZ mutation (VL)
(Figure 1d). The design should result in coexpression pography of the projections can be examined by visual-izing taulacZ expression.of taulacZ along with VRi2 from bicistronic messages,
according to the same logic of the P2-IRES-taulacZ mu- Germline transmission was obtained for the four tar-geted alleles of the VRi2 locus. We then removed thetation (Mombaerts et al., 1996a). Second, to improve
analysis of gene expression and axonal projections, we neo-selectable marker, which is required in the gene
The Wiring Diagram of the Vomeronasal System201
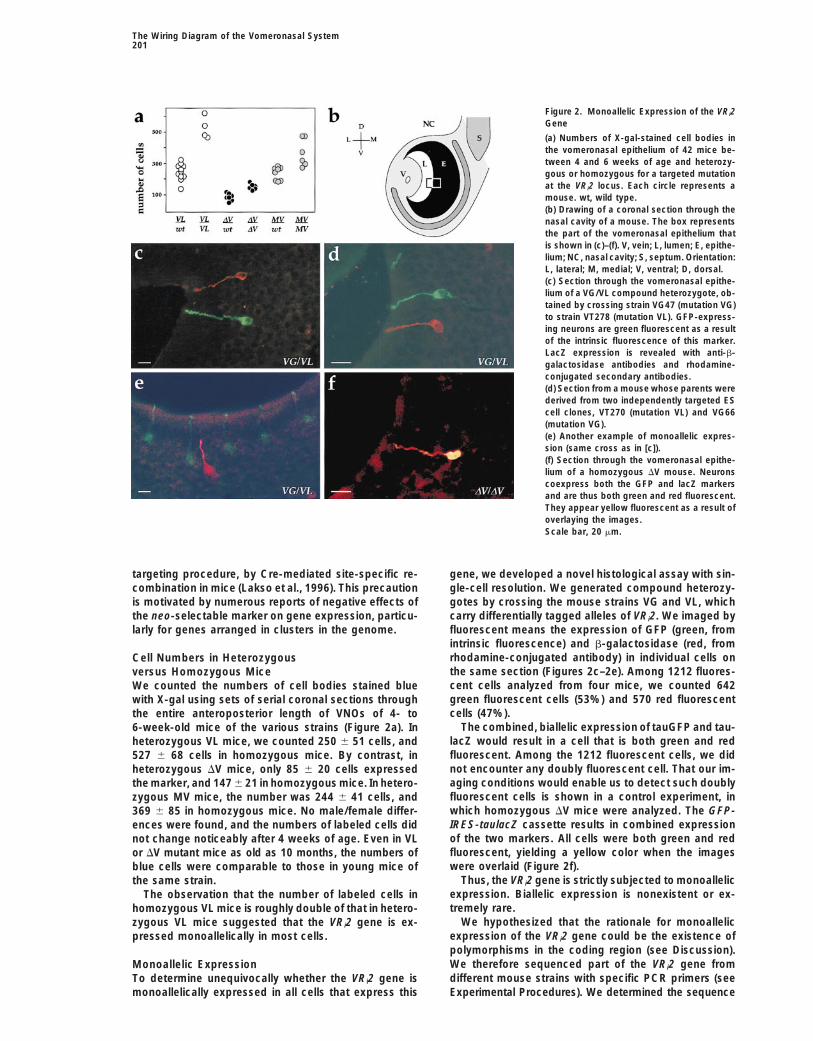
Figure 2. Monoallelic Expression of the VRi2Gene
(a) Numbers of X-gal-stained cell bodies inthe vomeronasal epithelium of 42 mice be-tween 4 and 6 weeks of age and heterozy-gous or homozygous for a targeted mutationat the VRi2 locus. Each circle represents amouse. wt, wild type.(b) Drawing of a coronal section through thenasal cavity of a mouse. The box representsthe part of the vomeronasal epithelium thatis shown in (c)–(f). V, vein; L, lumen; E, epithe-lium; NC, nasal cavity; S, septum. Orientation:L, lateral; M, medial; V, ventral; D, dorsal.(c) Section through the vomeronasal epithe-lium of a VG/VL compound heterozygote, ob-tained by crossing strain VG47 (mutation VG)to strain VT278 (mutation VL). GFP-express-ing neurons are green fluorescent as a resultof the intrinsic fluorescence of this marker.LacZ expression is revealed with anti-b-galactosidase antibodies and rhodamine-conjugated secondary antibodies.(d) Section from a mouse whose parents werederived from two independently targeted EScell clones, VT270 (mutation VL) and VG66(mutation VG).(e) Another example of monoallelic expres-sion (same cross as in [c]).(f) Section through the vomeronasal epithe-lium of a homozygous DV mouse. Neuronscoexpress both the GFP and lacZ markersand are thus both green and red fluorescent.They appear yellow fluorescent as a result ofoverlaying the images.Scale bar, 20 mm.
targeting procedure, by Cre-mediated site-specific re- gene, we developed a novel histological assay with sin-gle-cell resolution. We generated compound heterozy-combination in mice (Lakso et al., 1996). This precaution
is motivated by numerous reports of negative effects of gotes by crossing the mouse strains VG and VL, whichcarry differentially tagged alleles of VRi2. We imaged bythe neo-selectable marker on gene expression, particu-
larly for genes arranged in clusters in the genome. fluorescent means the expression of GFP (green, fromintrinsic fluorescence) and b-galactosidase (red, fromrhodamine-conjugated antibody) in individual cells onCell Numbers in Heterozygousthe same section (Figures 2c–2e). Among 1212 fluores-versus Homozygous Micecent cells analyzed from four mice, we counted 642We counted the numbers of cell bodies stained bluegreen fluorescent cells (53%) and 570 red fluorescentwith X-gal using sets of serial coronal sections throughcells (47%).the entire anteroposterior length of VNOs of 4- to
The combined, biallelic expression of tauGFP and tau-6-week-old mice of the various strains (Figure 2a). InlacZ would result in a cell that is both green and redheterozygous VL mice, we counted 250 6 51 cells, andfluorescent. Among the 1212 fluorescent cells, we did527 6 68 cells in homozygous mice. By contrast, innot encounter any doubly fluorescent cell. That our im-heterozygous DV mice, only 85 6 20 cells expressedaging conditions would enable us to detect such doublythe marker, and 147 6 21 in homozygous mice. In hetero-fluorescent cells is shown in a control experiment, inzygous MV mice, the number was 244 6 41 cells, andwhich homozygous DV mice were analyzed. The GFP-369 6 85 in homozygous mice. No male/female differ-IRES-taulacZ cassette results in combined expressionences were found, and the numbers of labeled cells didof the two markers. All cells were both green and rednot change noticeably after 4 weeks of age. Even in VLfluorescent, yielding a yellow color when the imagesor DV mutant mice as old as 10 months, the numbers ofwere overlaid (Figure 2f).blue cells were comparable to those in young mice of
Thus, the VRi2 gene is strictly subjected to monoallelicthe same strain.expression. Biallelic expression is nonexistent or ex-The observation that the number of labeled cells intremely rare.homozygous VL mice is roughly double of that in hetero-
We hypothesized that the rationale for monoalleliczygous VL mice suggested that the VRi2 gene is ex-expression of the VRi2 gene could be the existence ofpressed monoallelically in most cells.polymorphisms in the coding region (see Discussion).We therefore sequenced part of the VRi2 gene fromMonoallelic Expressiondifferent mouse strains with specific PCR primers (seeTo determine unequivocally whether the VRi2 gene is
monoallelically expressed in all cells that express this Experimental Procedures). We determined the sequence

Cell202
Figure 3. Pattern of VRi2-Expressing Cells inthe VNO
(a) Drawing of a parasagittal section throughthe head of a mouse. VNO, vomeronasal or-gan; S, septum; AOB, accessory olfactorybulb; MOB, main olfactory bulb. The vomero-nasal nerve is schematically represented bythree lines coursing along the septum. Orien-tation: A, anterior; P, posterior; V, ventral; D,dorsal.(b) Whole-mount view of the right half of thehead of a VL mouse (strain VT278), stainedwith X-gal. VRi2-expressing neurons are la-beled intensely blue from the cell bodies (bot-tom left) to the axon terminals (top right). Thearrow indicates the AOB.(c) Lateral whole-mount view of a VNO froma heterozygous OMP-taulacZ mouse.(d) Same whole-mount specimen as in (b) withhigher-power view of the VNO.
of VRi2 in ten inbred strains of the species Mus musculus 1996a), all mature OSNs and VSNs can be stained withX-gal because these cells express the olfactory marker(129/Sv, C57BL/6, BALB/c, CBA, B10, AKR/J, SJL, C3H,
DBA/2, and FVB). No nucleotide polymorphisms were protein (OMP) locus, and thus also the taulacZ reporter.found in the coding region, but these inbred strainsshare common ancestors (Silver, 1995). However, in othersubspecies, which are all inbred strains derived from Patterns in the Accessory Olfactory Bulb
We stained whole-mount specimens of VL mice withwild mice, we found polymorphisms in the coding region(nucleotide [nt], amino acid [aa]): Mus musculus domes- X-gal (Figures 4b–4d). The labeled axons segregate
near the AOB and terminate in multiple targets in theticus/musculus (SKIVE/Ei), 7 nt, 4 aa; Mus musculusdomesticus (WSB/Ei), 1 nt, 0 aa; Mus musculus domes- AOB. In sagittal sections these targets correspond to
glomeruli, which are, as expected, located exclusivelyticus (TIRANO/Ei), 1 nt, 0 aa; Mus musculus castaneus(CASA/Rk), 6 nt, 4 aa; and Mus musculus molossinus in the Gai2-positive region of the AOB (data not shown).
For comparison, a stained AOB from a heterozygous(MOLD/Rk), 7 nt, 4 aa. No nucleotide deletion or additionwas observed. In a different mouse species, Mus spretus, OMP-taulacZ mouse is shown (Figure 4a).
The pattern of the VRi2 glomeruli is complex and vari-we found 12 polymorphic amino acid residues and, inaddition, two internal deletions of 12 and 4 amino acids. able. Three examples are shown in Figures 4b–4d. Some
labeled glomeruli (3.5 6 1.9, mean 6 SD, n 5 12) arecontained within an area in the rostral part of the Gai2Pattern in the Vomeronasal Organ
The vomeronasal system in the mouse consists of the region and are often superficial, whereas other glomeruli(11.3 6 4.6, mean 6 SD, n 5 12) are located more cau-VNO, the vomeronasal nerve, and the AOB (Figure 3a).
The VNO is a bilateral, tubular, paramedian structure dally and often deeper in the AOB. The central area ofthe Gai2-positive region is consistently devoid of labeledlocated at the base of the nasal cavity. Axons emanate
from the cell bodies of VSNs and join in seven or so glomeruli. It often appears that multiple glomeruli arelinked as in a ganglion-like chain; this point is madefascicles that together form the vomeronasal nerve. This
nerve courses along the nasal septum, penetrates by an image from a VG mutant mouse (Figure 5f). Thepositions of these 15 labeled glomeruli lack the precisionthrough holes in the cribriform plate of the skull, migrates
along the medial surface of the MOB, turns around the and the constancy that have been observed in the mainolfactory system (Mombaerts et al., 1996a). No twomedial–posterior edge of the MOB, and terminates in
the AOB, which is situated dorsally and posteriorly to the stained samples look exactly alike. This variability isbest demonstrated by comparing the left and right AOBsMOB. Axons synapse within loosely defined glomeruli
either in the rostral, Gai2-positive region or the caudal, of VL mutant mice: there is no bilateral symmetry withinan individual (Figure 4d). The average number of labeledGao-positive region.
Figure 3b shows a whole-mount view of a half head glomeruli is the same in heterozygous versus homozy-gous VL mice (data not shown).of a heterozygous VL mouse stained with X-gal. A higher
magnification of the VNO shows stained neurons corre- Serial sections through 8 AOBs from heterozygous VLmice and 14 AOBs from homozygous VL mice weresponding to VRi2-expressing cells (Figure 3d). These
cells are homogeneously scattered throughout the vom- stained with antibodies against b-galactosidase (greenfluorescent color), along with antibodies against the pre-eronasal epithelium and are restricted to the apical part
when analyzed in coronal sections (data not shown). synaptic marker synaptophysin (red fluorescent color)and DAPI (blue fluorescent color). The latter dye labelsAxons of blue VSNs participate in the formation of all
fascicles of the vomeronasal nerve. For comparison, the nuclei, providing an outline of glomerular structures. Thesuperimposition of the green and red fluorescence re-VNO of a heterozygous OMP-taulacZ mouse is shown
in Figure 3c. In mice of this strain (Mombaerts et al., sults in yellow fluorescence. In heterozygous VL mice,
The Wiring Diagram of the Vomeronasal System203
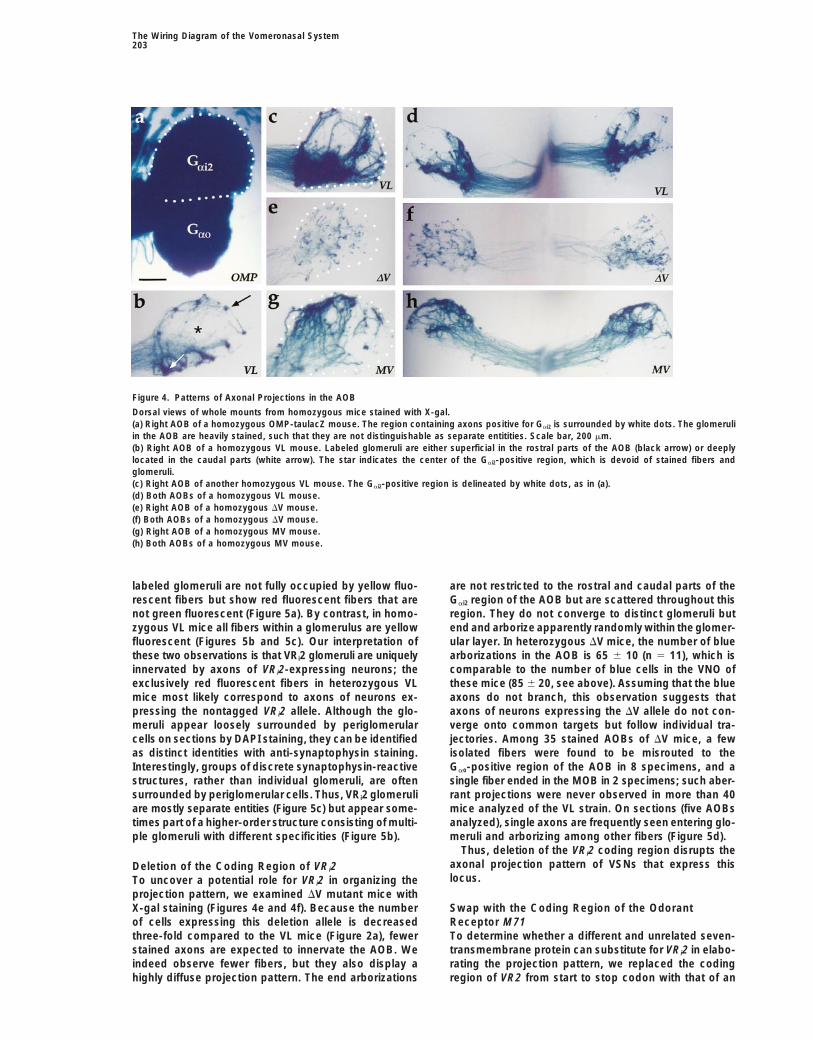
Figure 4. Patterns of Axonal Projections in the AOB
Dorsal views of whole mounts from homozygous mice stained with X-gal.(a) Right AOB of a homozygous OMP-taulacZ mouse. The region containing axons positive for Gai2 is surrounded by white dots. The glomeruliin the AOB are heavily stained, such that they are not distinguishable as separate entitities. Scale bar, 200 mm.(b) Right AOB of a homozygous VL mouse. Labeled glomeruli are either superficial in the rostral parts of the AOB (black arrow) or deeplylocated in the caudal parts (white arrow). The star indicates the center of the Gai2-positive region, which is devoid of stained fibers andglomeruli.(c) Right AOB of another homozygous VL mouse. The Gai2-positive region is delineated by white dots, as in (a).(d) Both AOBs of a homozygous VL mouse.(e) Right AOB of a homozygous DV mouse.(f) Both AOBs of a homozygous DV mouse.(g) Right AOB of a homozygous MV mouse.(h) Both AOBs of a homozygous MV mouse.
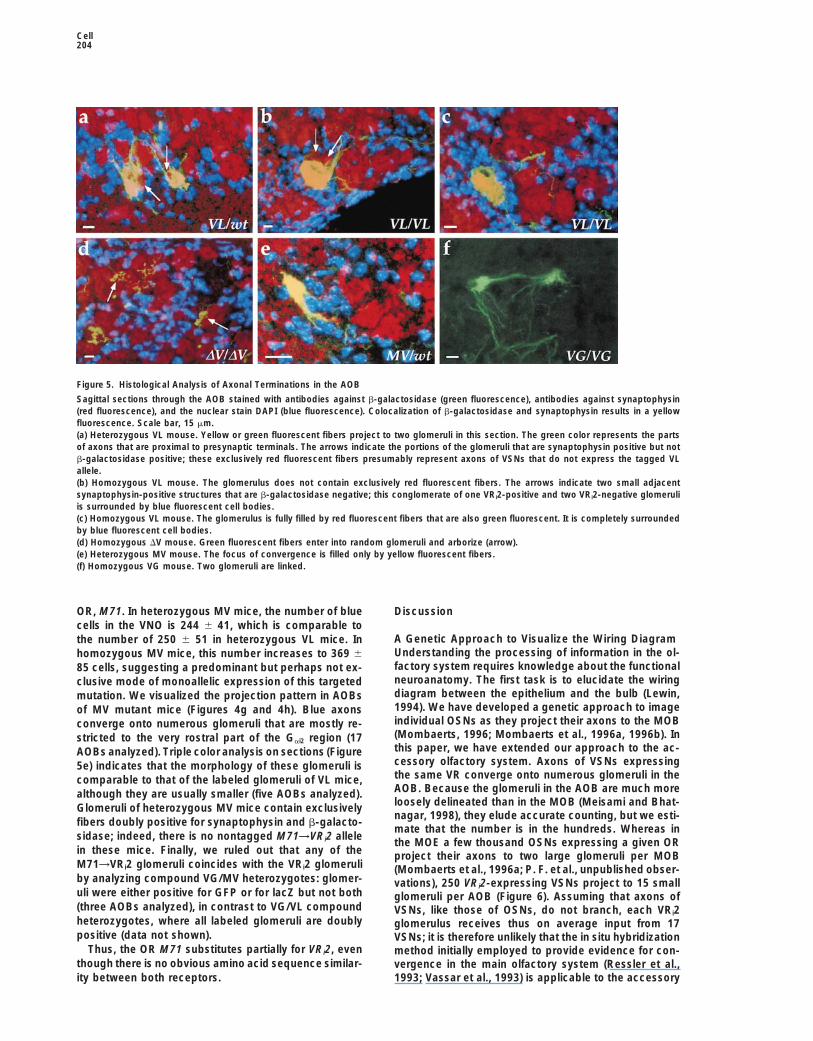
labeled glomeruli are not fully occupied by yellow fluo- are not restricted to the rostral and caudal parts of theGai2 region of the AOB but are scattered throughout thisrescent fibers but show red fluorescent fibers that are
not green fluorescent (Figure 5a). By contrast, in homo- region. They do not converge to distinct glomeruli butend and arborize apparently randomly within the glomer-zygous VL mice all fibers within a glomerulus are yellow
fluorescent (Figures 5b and 5c). Our interpretation of ular layer. In heterozygous DV mice, the number of bluearborizations in the AOB is 65 6 10 (n 5 11), which isthese two observations is that VRi2 glomeruli are uniquely
innervated by axons of VRi2-expressing neurons; the comparable to the number of blue cells in the VNO ofthese mice (85 6 20, see above). Assuming that the blueexclusively red fluorescent fibers in heterozygous VL
mice most likely correspond to axons of neurons ex- axons do not branch, this observation suggests thataxons of neurons expressing the DV allele do not con-pressing the nontagged VRi2 allele. Although the glo-
meruli appear loosely surrounded by periglomerular verge onto common targets but follow individual tra-jectories. Among 35 stained AOBs of DV mice, a fewcells on sections by DAPI staining, they can be identified
as distinct identities with anti-synaptophysin staining. isolated fibers were found to be misrouted to theGao-positive region of the AOB in 8 specimens, and aInterestingly, groups of discrete synaptophysin-reactive
structures, rather than individual glomeruli, are often single fiber ended in the MOB in 2 specimens; such aber-rant projections were never observed in more than 40surrounded by periglomerular cells. Thus, VRi2 glomeruli
are mostly separate entities (Figure 5c) but appear some- mice analyzed of the VL strain. On sections (five AOBsanalyzed), single axons are frequently seen entering glo-times part of a higher-order structure consisting of multi-
ple glomeruli with different specificities (Figure 5b). meruli and arborizing among other fibers (Figure 5d).Thus, deletion of the VRi2 coding region disrupts the
axonal projection pattern of VSNs that express thisDeletion of the Coding Region of VRi2locus.To uncover a potential role for VRi2 in organizing the
projection pattern, we examined DV mutant mice withX-gal staining (Figures 4e and 4f). Because the number Swap with the Coding Region of the Odorant
Receptor M71of cells expressing this deletion allele is decreasedthree-fold compared to the VL mice (Figure 2a), fewer To determine whether a different and unrelated seven-
transmembrane protein can substitute for VRi2 in elabo-stained axons are expected to innervate the AOB. Weindeed observe fewer fibers, but they also display a rating the projection pattern, we replaced the coding
region of VR2 from start to stop codon with that of anhighly diffuse projection pattern. The end arborizations
Cell204
Figure 5. Histological Analysis of Axonal Terminations in the AOB
Sagittal sections through the AOB stained with antibodies against b-galactosidase (green fluorescence), antibodies against synaptophysin(red fluorescence), and the nuclear stain DAPI (blue fluorescence). Colocalization of b-galactosidase and synaptophysin results in a yellowfluorescence. Scale bar, 15 mm.(a) Heterozygous VL mouse. Yellow or green fluorescent fibers project to two glomeruli in this section. The green color represents the partsof axons that are proximal to presynaptic terminals. The arrows indicate the portions of the glomeruli that are synaptophysin positive but notb-galactosidase positive; these exclusively red fluorescent fibers presumably represent axons of VSNs that do not express the tagged VLallele.(b) Homozygous VL mouse. The glomerulus does not contain exclusively red fluorescent fibers. The arrows indicate two small adjacentsynaptophysin-positive structures that are b-galactosidase negative; this conglomerate of one VRi2-positive and two VRi2-negative glomeruliis surrounded by blue fluorescent cell bodies.(c) Homozygous VL mouse. The glomerulus is fully filled by red fluorescent fibers that are also green fluorescent. It is completely surroundedby blue fluorescent cell bodies.(d) Homozygous DV mouse. Green fluorescent fibers enter into random glomeruli and arborize (arrow).(e) Heterozygous MV mouse. The focus of convergence is filled only by yellow fluorescent fibers.(f) Homozygous VG mouse. Two glomeruli are linked.
OR, M71. In heterozygous MV mice, the number of blue Discussioncells in the VNO is 244 6 41, which is comparable to
A Genetic Approach to Visualize the Wiring Diagramthe number of 250 6 51 in heterozygous VL mice. InUnderstanding the processing of information in the ol-homozygous MV mice, this number increases to 369 6factory system requires knowledge about the functional85 cells, suggesting a predominant but perhaps not ex-neuroanatomy. The first task is to elucidate the wiringclusive mode of monoallelic expression of this targeteddiagram between the epithelium and the bulb (Lewin,mutation. We visualized the projection pattern in AOBs1994). We have developed a genetic approach to imageof MV mutant mice (Figures 4g and 4h). Blue axonsindividual OSNs as they project their axons to the MOBconverge onto numerous glomeruli that are mostly re-(Mombaerts, 1996; Mombaerts et al., 1996a, 1996b). Instricted to the very rostral part of the Gai2 region (17this paper, we have extended our approach to the ac-AOBs analyzed). Triple color analysis on sections (Figurecessory olfactory system. Axons of VSNs expressing5e) indicates that the morphology of these glomeruli isthe same VR converge onto numerous glomeruli in thecomparable to that of the labeled glomeruli of VL mice,AOB. Because the glomeruli in the AOB are much morealthough they are usually smaller (five AOBs analyzed).loosely delineated than in the MOB (Meisami and Bhat-Glomeruli of heterozygous MV mice contain exclusivelynagar, 1998), they elude accurate counting, but we esti-
fibers doubly positive for synaptophysin and b-galacto-mate that the number is in the hundreds. Whereas in
sidase; indeed, there is no nontagged M71→VRi2 allele the MOE a few thousand OSNs expressing a given ORin these mice. Finally, we ruled out that any of the project their axons to two large glomeruli per MOBM71→VRi2 glomeruli coincides with the VRi2 glomeruli (Mombaerts et al., 1996a; P. F. et al., unpublished obser-by analyzing compound VG/MV heterozygotes: glomer- vations), 250 VRi2-expressing VSNs project to 15 smalluli were either positive for GFP or for lacZ but not both glomeruli per AOB (Figure 6). Assuming that axons of(three AOBs analyzed), in contrast to VG/VL compound VSNs, like those of OSNs, do not branch, each VRi2heterozygotes, where all labeled glomeruli are doubly glomerulus receives thus on average input from 17positive (data not shown). VSNs; it is therefore unlikely that the in situ hybridization
Thus, the OR M71 substitutes partially for VRi2, even method initially employed to provide evidence for con-though there is no obvious amino acid sequence similar- vergence in the main olfactory system (Ressler et al.,
1993; Vassar et al., 1993) is applicable to the accessoryity between both receptors.

The Wiring Diagram of the Vomeronasal System205
the accessory olfactory system does not have directconnections to the cerebral cortex.
This difference in wiring diagrams may reflect the dif-ferential functions fulfilled by these systems. The mainolfactory system is designed to detect an immense vari-ety of odorants, often at extremely low concentrations,and to make fine discriminations between structurallyrelated molecules. Convergence to a small number ofglomeruli at fixed positions may increase the sensitivityand specificity of the system. The mammalian accessoryolfactory system, on the other hand, provides informa-tion about chemical signals emanating from individualsof the same species. Pheromonal responses are oftenproduced by “blends” rather than by single compounds(Sorensen, 1996). The accessory olfactory system maythus perceive complex patterns of pheromonal stimulias a “Gestalt” that triggers preprogrammed behavioraland neuroendocrine responses. Furthermore, the ac-cessory olfactory system is more important for the socialand sexual behavior of naive animals than of experi-enced individuals and must relay varying kinds of infor-
Figure 6. Topography of Sensory Projections in the Main and Ac-mation about conspecific signals as the individual ma-cessory Olfactory Systemstures from a newborn to an adult. Having numerous(Top) In the main olfactory system, axons from the MOE convergesmall glomeruli may allow for greater plasticity and re-to a few large glomeruli located at stereotyped positions in the MOB.modeling ability. The vomeronasal system, which under-Mitral cells send their single apical dendrite to a single glomerulus.
Periglomerular cells are represented by blue dots. goes constant neuronal renewal throughout life, may(Bottom) In the accessory olfactory system, axons from the vomero- therefore be more dependent on activity-dependentnasal epithelium (VNE) converge to multiple small glomeruli located mechanisms and be more plastic than the main olfactoryat variable positions in the AOB. Some composite structures are
system, explaining perhaps the variability of the patternsurrounded by cell bodies (blue dots) and contain multiple glomeruliof axonal projections. Because the pattern does notcorresponding to different VRs. Mitral/tufted cells have several api-display bilateral symmetry within one mouse, stochasticcal dendrites that innervate multiple glomeruli. The question marks
reflect the uncertainty about the relationship between glomeruli in- events may also contribute to the generation of the finalnervated by dendrites of a single mitral cell. projection map. Thus, for a given pheromonal stimulus,
multiple wiring diagrams may result in the same re-sponse; the preprogrammed nature of the behavioral orolfactory system. Costaining with DAPI (to outline thephysiological response may not correspond to a fixedrings of surrounding cell bodies) and antibodies againstpattern of connectivity at the first stage of the pathway.synaptophysin (to image presynaptic terminals) sug-
gests that these small glomeruli are exclusively inner-vated by axons of VRi2-expressing VSNs. The distribu- Are VRs Determinants of Axon Guidance?
Wiring the axons of subsets of neurons, which are scat-tion of the VRi2 glomeruli lacks the precision andconstancy observed for MOB glomeruli corresponding tered throughout a zone of the epithelium, to discrete
glomeruli poses a formidable challenge to axon guid-to a given OR. No two mice look alike, even when theleft and right AOBs of a single individual are compared. ance (Lewin, 1994). Genetic studies with the OR gene P2
have led to the hypothesis that the OR itself is intimatelyIt will be important to determine for other VRs whethertheir glomeruli are also exclusively innervated by VSNs involved in the mechanisms that enable axons to navi-
gate to their targets in the MOB. These studies sup-expressing the same VR; perhaps some VSN subsetsexpressing certain VRs coconverge with subsets ex- ported the notion that the OR is a component of the
guidance machinery, but they clearly demonstrate thatpressing other VRs, which may be related in sequence.It has been known for nearly a century that second- it is not the only determinant (Mombaerts et al., 1996a;
Wang et al., 1998). In this paper, we report similar geneticorder neurons in the accessory olfactory system (mitral/tufted cells) project their dendrites to several, often dis- experimentation with the VRi2 gene.
First, when the coding region of the VRi2 gene wastant glomeruli of the AOB (Ramon y Cajal, 1911; Takamiand Graziadei, 1991). By contrast, in the MOB a mitral replaced with a GFP-IRES-taulacZ cassette, the number
of labeled neurons was decreased by a factor of threecell or tufted cell has only a single apical dendrite andprojects this dendrite to a single glomerulus. Perhaps compared to VRi2-taulacZ mutation (in which the coding
region is not altered), suggesting that expression of thea given mitral cell in the AOB sends dendrites to glomer-uli that receive input from neurons expressing different VRi2 gene is required for a normal life span. Alternatively,
the absence of the VRi2 coding region could decreaseVRs (Figure 6). This organization of second-order neu-rons can now be studied with genetic transneuronal the probability that the VRi2 locus is chosen for expres-
sion. Axons of VSNs expressing the deletion allele notracing techniques (Yoshihara et al., 1999). Such an ana-tomical organization would allow for a substantial de- longer converge. At first approximation, this phenotype
would argue for a role of VRi2 in axon guidance, butgree of integration of information within the AOB, beforeit is relayed to other neural areas; it must be noted that we do not yet know whether neurons expressing the

Cell206
deletion allele also express a VR gene. If they do, it is An alternative view is that monoallelic expression ofVRi2 is a side effect of the mechanisms regulating VRpossible that stained VSNs express different VR genes
and project to different glomeruli, perhaps indepen- gene expression but has no apparent biological signifi-cance. It is perhaps the consequence of a DNA re-dently from a direct involvement of the expressed VR(s).
The interpretation of the DV phenotype is therefore am- arrangement event that activates stochastically a singleVR gene, from a single allele, in a given neuron. Asbivalent at this stage. The same caveat precludes sound
conclusions about a similar deletion mutation in P2 a corollary, expression is monoallelic, but in fact, it isprimarily monogenic and also monoallelic. We must(Wang et al., 1998).
Second, when the coding region of the VRi2 gene was stress that there is no evidence for the generation of newORs or VRs sequences through DNA rearrangements,replaced with that of the OR gene M71, the number of
labeled neurons was restored to levels similar to those when cDNA and genomic sequences are compared. Thepurpose would be to restrict expression to a single re-observed in mice carrying the VL mutation. The labeled
axons converged onto multiple glomeruli in the AOB, ceptor type, not to create a new gene sequence. A usefulanalogy would be the regulation of expression of variantwhich are distinct from the VRi2 glomeruli. Because ORs
and VRs have no significant amino acid sequence simi- surface glycoprotein genes in Trypanosomes throughgene conversion.larity, the transduction cascade in VSNs is different from
that in OSNs, and the morphology and location of the Targeted expression of GFP now enables us to iden-tify and isolate live VSNs expressing a given VR. We cansensory neurons are also distinct, it is unlikely that an
M71-expressing VSN is functionally equivalent to an search for DNA rearrangements in the genome of VRi2-expressing cells by PCR on their genomic DNA or byOSN normally expressing M71. Therefore, our results
argue in favor of a role of ORs and VRs in the formation cloning mice with their nuclei (Wakayama et al., 1998).or maintenance of glomeruli that would be distinct fromtheir role in sensory perception. Perhaps axonal projec-
Experimental Procedurestions from neurons expressing the same receptor recog-nize each other in a receptor-dependent fashion, leading
Targeting Vectorsto a coalescence of their termini into foci, which then A portion of the rat VN5 coding region (Dulac and Axel, 1995) wasdevelop into glomeruli. isolated by PCR from rat genomic DNA and used as a probe to
screen a 129/Sv mouse BAC genomic library (Genome Systems). A7 kb HindIII fragment containing VRi2 was subcloned into pBS-SKMonoallelic Expression: Why and How?and sequenced entirely. A PacI site was generated by recombinantIn general, a gene is expressed from both alleles orPCR immediately after the stop codon of VRi2 (plasmid VRi2/Pac).
said to be subjected to “biallelic expression.” There are, VRi2-IRES-taulacZ: a PacI fragment containing IRES-taulacZ-LNLhowever, exceptions to this rule (Chess, 1998b): genes was inserted into plasmid VRi2/Pac.on the X chromosome, immunoglobulin and T cell recep- VRi2-IRES-tauGFP: a PacI fragment containing IRES-tauGFP-LNL
was inserted into plasmid VRi2/Pac. The version of GFP employedtor genes, imprinted genes, natural killer cell receptorwas EGFP-1 (Clontech).genes, interleukin-2 and -4 genes, and odorant receptor
(DVRi2) GFP-IRES-taulacZ: the coding sequence of VRi2 was re-genes. We have now shown that the VRi2 gene is ex-moved exactly from the start to the stop codon in plasmid VRi2/
pressed also in a monoallelic fashion, employing a novel Pac, generating plasmid delVRi2/Pac. A PacI fragment containinghistological single-cell assay with differentially tagged the GFP-IRES-taulacZ-LNL cassette was then ligated into plasmidalleles. A difference with the other cases is that the delVRi2/Pac.
M71→VRi2-IRES-taulacZ: the VRi2 coding sequence was replacedfidelity of monoallelism appears to be extremely high:exactly from the start to the stop codon by the M71 coding sequencewe have not encountered a single case of biallellic ex-in the VRi2/Pac plasmid. A PacI fragment containing the IRES-tau-pression out of 1212 cells.lacZ-LNL cassette was then cloned into this plasmid.
It is not obvious why VR genes, or OR genes, should Plasmid requests should be addressed by e-mail to plasmid@be expressed from one allele. We offer the following rockvax.rockefeller.edu.speculation. The functional organization of the olfactorysystem is probably based on the expression of a single
Gene Targetingreceptor type in a given sensory neuron. If polymorphicTargeting vectors were linearized with SalI. Electroporation and cellvariants exist within a species, biallelic expression mayculture of E14 cells (Hooper et al., 1987) were carried out as de-
result in expression, in heterozygous individuals, of two scribed before (Mombaerts et al., 1996a). Genomic DNA from G418-receptors with different specificities in the same neuron resistant ES colonies was digested with HindIII and analyzed by(Lancet, 1994). Indeed, a single amino acid change in the Southern blot hybridization with a 39 probe external to the targeting
vector. Germline transmission was obtained for the VL mutationOR I7 is sufficient to alter the odorous ligand specificity(clones VT270 and VT278), the VG mutation (clones VG47 and VG66),(Krautwurst et al., 1998). In addition, if the receptor isthe DV mutation (DV14), and the MV mutation (clone SV72). Miceimplicated in the choice and formation of a glomerulus,are in a mixed (129 3 C57BL/6) background.
a subtle difference in amino acid sequence could resultin an inappropriate compound code. Therefore, to en-sure that a sensory neuron relays unequivocal informa- Cre Recombination
The neo-selectable marker was removed from the targeted muta-tion, it is may be necessary to restrict expression nottions by crossing mice heterozygous for the LNL allele to EIIa-Creonly to a single receptor gene, but also to one allele oftransgenic mice (Lakso et al., 1996). Transgenic mice used werethat gene. In support of the “polymorphism” hypothesis,backcrossed by us at least four times to the C57BL/6 background.
we have uncovered differences in the deduced amino Intercrossing of loxP-positive mice resulted in loxP heterozygousacid sequence of the VRi2 gene among strains of mice and loxP homozygous mice that were devoid of the Cre transgene.
All analyses were with mice that did not carry the Cre transgene.(see Results).

The Wiring Diagram of the Vomeronasal System207
LacZ Expression Sloan, Basil O’Connor, Guggenheim, Irma T. Hirschl, Klingenstein,McKnight, Rita Allen, and Searle Scholar or Fellow.For whole mounts, tissues were processed as described before
(Mombaerts et al., 1996a), except for the fixation that was carriedout on ice for 8 min. To visualize better the deep glomeruli in the
Received December 31, 1998; revised March 10, 1999.AOB, whole mounts were immersed in methanol for 30 min and thentransferred into a solution of benzyl benzoate and benzyl alcohol(2:1) for 5 min.
References
Immunohistochemistry Berghard, A., and Buck, L.B. (1996). Sensory transduction in vom-Tissues were fixed in 2% formaldehyde, 13.5 mg/ml lysine, 2.1 mg/ eronasal neurons: evidence for Gao, Gai2, and adenlylyl cylcase II asml sodium periodate, and 0.1 M phosphate buffer at pH 7.4 on ice major components of a pheromone signaling cascade. J. Neurosci.for 3 hr and then frozen in OCT. Seven micrometer sections were 16, 909–918.cut. For Gai2 staining, primary antibodies (Wako Pure Chemical In-
Brand, A. (1995). GFP in Drosophila. Trends Genet. 11, 324–325.dustries) were used at a 1:300 dilution followed by a goat anti-rabbitBuck, L.B. (1996). Information coding in the vertebrate olfactoryIgG coupled to rhodamine (ICN Pharmaceuticals). For b-galactosi-system. Annu. Rev. Neurosci. 19, 517–544.dase immunostaining in the vomeronasal epithelium, a rabbit IgG
fraction (Cappel) was used at a 1:100 dilution, followed by a rho- Buck, L., and Axel, R. (1991). A novel multigene family may encodedamine-conjugated anti-rabbit IgG (ICN Pharmaceuticals). For odorant receptors: a molecular basis for odor recognition. Cell 65,b-galactosidase immunostaining in the AOB, a mouse antibody (Pro- 175–187.mega) was used at a concentration of 7 mg/ml, followed by a FITC- Butner, K.A., and Kirschner, M.W. (1991). Tau protein binds to micro-conjugated goat anti-mouse IgG (ICN Pharmaceuticals). A rabbit tubules through a flexible array of distributed weak sites. J. Cell.anti-synaptophysin antibody (DAKO) was used at a 1:80 dilution, Biol. 115, 717–730.followed by a goat anti-rabbit IgG coupled to rhodamine (ICN Phar-
Callahan, C.A., and Thomas, J.B. (1994). Tau-beta-galactosidase,maceuticals). DAPI staining was performed after immunohistochem-
an axon-targeted fusion protein. Proc. Natl. Acad. Sci. USA 91,ical reactions by immersing the slide for 30 s in 0.2 mg/ml DAPI
5972–5976.(Sigma) in PBS and washing them for 1 min in PBS.
Chalfie, M., Tu, Y., Euskirchen, G., Ward, W.W., and Prasher, D.C.(1994). Green fluorescent protein as a marker for gene expression.
Microscopy and Photography Science 263, 802–805.Whole-mount specimens were examined with a Zeiss SV11 stereo-
Chess, A. (1998a). Olfactory receptor gene regulation. Adv. Immunol.microscope; sections were examined with a Zeiss Axioplan 2 fluo-
69, 437–447.rescence microscope. Images were taken with a Canon EOS-DCS
Chess, A. (1998b). Expansion of the allelic exclusion principle? Sci-digital camera and processed with Adobe Photoshop 3.0.ence 279, 2067–2068.
Chess, A., Simon, I., Cedar, H., and Axel, R. (1994). Allelic inactivationPolymerase Chain Reaction
regulates olfactory receptor gene expression. Cell 78, 823–834.For 59 RACE, reverse transcription and further cycles of amplification
Dulac, C., and Axel, R. (1995). A novel family of genes encodingwere carried out with the SMART kit (Clontech) following the manu-putative pheromone receptors in mammals. Cell 83, 195–206.facturer’s instructions, with reverse nested primers. For 39 RACE,Farbman, A.I. (1992). Cell Biology of Olfaction (New York: Cambridgestandard procedures were followed (Frohman, 1993).University Press).For amplification of the VRi2 sequence from genomic DNA of
different strains, a sense primer located in intron 1 of the VRi2 gene Frohman, M.A. (1993). Rapid amplification of complementary DNAand upstream of the coding region (GTC CAT TTA TCA AGT GAG ends for generation of full-length complementary DNAs: thermalCTG C) and a reverse primer located in the 39 noncoding region of RACE. Methods Enzymol. 218, 340–356.the VRi2 gene (TTT AAG CCA CAG AAT ATT CCC) were used. The Halpern, M. (1987). The organization and function of the vomerona-1389 nt amplification products were cloned and sequenced from sal system. Annu. Rev. Neurosci. 10, 325–362.both strands. A critical control for primer specificity is the absence
Halpern, M., Shapiro, L.S., and Jia, C. (1995). Differential localizationof an amplification product with genomic DNA homozygous for aof G proteins in the opossum vomeronasal system. Brain Res. 677,targeted mutation in the VRi2 gene; the presence of an IRES cassette157–161.precludes efficient PCR because of its length and the unusual se-Herrada, G., and Dulac, C. (1997). A novel family of putative phero-quence of the IRES region. When the amplified sequences weremone receptors in mammals with a topographically organized andcompared to more than 20 other mouse VR sequences of the Gai2
sexually dimorphic distribution. Cell 90, 763–773.type, VRi2 from 129/Sv was unambiguously the closest sequence.Thus, the PCR primers amplify the VRi2 gene in different strains, Hooper, M., Hardy, K., Handyside, A., Hunter, S., and Monk, M.rather than VRi2-like genes. (1987). HPRT-deficient (Lesch-Nyhan) mouse embryos derived from
germline colonization by cultured cells. Nature 326, 292–295.
Jia, C., and Halpern, M. (1996). Subclasses of vomeronasal receptorAcknowledgmentsneurons: differential expression of G proteins (Gia2 and Goa) andsegregated projections to the accessory olfactory bulb. Brain Res.We thank Jenny Yee for technical assistance and the Transgenic719, 117–128.Service of The Rockefeller University for producing chimeric mice.
We acknowledge Clontech for EGFP-1, Chen Zheng for the IRES- Karlson, P., and Luscher, M. (1959). “Pheromones”: a new term fortauGFP cassette, and Charlie Greer for anti-synaptophysin antibod- a class of biologically active substances. Nature 183, 55–56.ies and advice. We thank Stuart Firestein, Charlie Greer, and Pierre Keverne, E.B. (1983). Pheromonal influences on the endocrine regu-Vassalli for comments on the manuscript. We are very grateful to lation of reproduction. Trends Neurosci. 6, 381–384.Heiner Westphal for generously providing EIIa-Cre transgenic mice,
Krautwurst, D., Yau, K.-W., and Reed, R.R. (1998). Identification ofwhich resulted in enormous time savings when generating loxPligands for olfactory receptors by functional expression of a receptorversions of the targeted mutations. I. R. was supported by postdoc-library. Cell 95, 917–926.toral fellowships from the Swiss National Foundation, the EuropeanLakso, M., Pichel, J.G., Gorman, J.R., Sauer, B., Okamoto, Y., Lee,Molecular Biology Organization, and the Human Frontier ScienceE., Alt, F.W., and Westphal, H. (1996). Efficient in vivo manipulationProgram Organization. P. F. was supported by postdoctoral fellow-of mouse genomic sequences at the zygote stage. Proc. Natl. Acad.ships from Bristol-Myers-Squibb and the National Institutes ofSci. USA 93, 5860–5865.Health. P. M. received grant support from the National Institutes of
Health, the Human Frontier Science Program Organization, and the Lancet, D. (1994). Olfaction: exclusive receptors. Nature 372,321–322.March of Dimes Birth Defects Organization. P. M. was an Alfred P.

Cell208
Lewin, B. (1994). On neuronal specificity and the molecular basis of Zhao, H., Ivic, L., Otaki, J.M., Hashimoto, M., Mikoshiba, K., andFirestein, S. (1998). Functional expression of a mammalian odorantperception. Cell 79, 935–943.receptor. Science 279, 237–242.Matsunami, H., and Buck, L.B. (1997). A multigene family encoding
a diverse array of putative pheromone receptors in mammals. Cell90, 775–784.
Meisami, E., and Bhatnagar, K.P. (1998). Structure and diversityin mammalian accessory olfactory bulb. Microsci. Res. Tech. 43,476–499.
Mombaerts, P. (1996). Targeting olfaction. Curr. Opin. Neurobiol. 6,481–486.
Mombaerts, P. (1999). Molecular biology of odorant receptors invertebrates. Annu. Rev. Neurosci. 22, 487–509.
Mombaerts, P., Wang, F., Dulac, C., Chao, S.K., Nemes, A., Mendel-sohn, M., Edmondson, J., and Axel, R. (1996a). Visualizing an olfac-tory sensory map. Cell 87, 675–686.
Mombaerts, P., Wang, F., Dulac, C., Vassar, R., Chao, S.K., Nemes,A., Mendelsohn, M., Edmondson, J., and Axel, R. (1996b). The mo-lecular biology of olfactory perception. Cold Spring Harb. Symp.Quant. Biol. 61, 135–145.
Ramon y Cajal, S. (1911). Appareil olfactif. Muqueuse olfactive etbulbe olfactif ou centre olfactif de premier ordre. In Histologie duSysteme Nerveux de l’Homme et des Vertebres. Vol. 2 (Paris: Ma-loine), pp. 647–674.
Ressler, K.J., Sullivan, S.L., and Buck, L.B. (1993). A zonal organiza-tion of odorant receptor gene expression in the olfactory epithelium.Cell 73, 597–609.
Ressler, K.J., Sullivan, S.L., and Buck, L.B. (1994). Information cod-ing in the olfactory system: evidence for a stereotyped and highlyorganized epitope map in the olfactory bulb. Cell 79, 1245–1255.
Royet, J.P., Souchier, C., Jourdan, F., and Ploye, H. (1988). Morpho-metric study of the glomerular population in the mouse olfactorybulb: numerical density and size distribution along the rostrocaudalaxis. J. Comp. Neurol. 270, 559–568.
Ryba, N.J.P., and Tirindelli, R. (1997). A new multigene family ofputative pheromone receptors. Neuron 19, 371–379.
Saito, H., Mimmack, M.L., Keverne, E.B., Kishimoto, J., and Emson,P.C. (1998). Isolation of mouse vomeronasal receptor genes andtheir co-localization with specific G-protein messenger RNAs. Mol.Brain Res. 60, 215–227.
Silver, L.M. (1995). Mouse Genetics: Concepts and Applications.(New York: Oxford University Press), pp. 3–61.
Sorensen, P.W. (1996). Biological responsiveness to pheromonesprovides fundamental and unique insight into olfactory function.Chem. Sens. 21, 245–256.
Takami, S., and Graziadei, P.P.C. (1991). Light microscopic Golgistudy of mitral/tufted cells in the accessory olfactory bulb of theadult rat. J. Comp. Neurol. 311, 65–83.
Tirindelli, R., Mucignat-Caretta, C., and Ryba, N.J.P. (1998). Molecu-lar aspects of pheromonal communication via the vomeronasal or-gan in mammals. Trends Neurosci. 21, 482–486.
Tsien, R.Y. (1998). The green fluorescent protein. Annu. Rev. Bio-chem. 67, 509–544.
Vassar, R., Ngai, J., and Axel, R. (1993). Spatial segregation of odor-ant receptor expression in the mammalian olfactory epithelium. Cell74, 309–318.
Vassar, R., Chao, S.K., Sitcheran, R., Nunez, J.M., Vosshall, L.B.,and Axel, R. (1994). Topographic organization of sensory projectionsto the olfactory bulb. Cell 79, 981–991.
Wakayama, T., Perry, A.C.F., Zuccotti, M., Johnson, K.R., and Ya-nagimachi, R. (1998). Full-term development of mice from enucle-ated oocytes injected with cumulus cell nuclei. Nature 394, 369–374.
Wang, F., Nemes, A., Mendelsohn, M., and Axel, R. (1998). Odorantreceptors govern the formation of a precise topographic map. Cell93, 47–60.
Wilson, E.O. (1963). Pheromones. Sci. Am. 208, 100–114.
Yoshihara, Y., Mizuno, T., Nakahira, M., Kawasaki, M., Watanabe,Y., Kagamiyama, H., Jishage, K., Ueda, O., Suzuki, H., Tabushi, K.,et al. (1999). A genetic approach to visualization of multisynapticneural pathways using plant lectin transgene. Neuron 22, 33–41.




















