
Axonal cytoskeletal changes after non-disruptive axonal injury
15
Journal of Neurocytology 26, 207–221 (1997) Axonal cytoskeletal changes after non-disruptive axonal injury S. S. JAFARI 1 , W. L. MAXWELL 1 * , M. NEILSON 1 and D. I. GRAHAM 2 1 Laboratory of Human Anatomy, Institute for Biomedical and Life Sciences, University of Glasgow, Glasgow G12 8QQ and 2 University Department of Neuropathology, Institute of Neurological Sciences, Southern General Hospital, Glasgow G51 4TF, UK Received 22 December 1995; revised 18 July, 22 October and 6 December 1996; accepted 12 December 1996 Summary In animal models of human diffuse axonal injury, axonal swellings leading to secondary axotomy occur between 2 and 6 h after injury. But, analysis of cytoskeletal changes associated with secondary axotomy has not been undertaken. We have carried out a quantitative analysis of cytoskeletal changes in a model of diffuse axonal injury 4 h after stretch-injury to adult guinea-pig optic nerves. The major site of axonal damage was the middle portion of the nerve. There was a statistically significant increase in the proportion of small axons with a diameter of 0.5 lm and smaller in which there was compaction of neurofilaments. Axons with a diameter greater than 2.0 lm demonstrated an increased spacing between cytoskeletal elements throughout the length of the nerve. However, in the middle segment of the nerve these larger axons demonstrated two different types of response. Either, where periaxonal spaces occurred, there was a reduction in axonal calibre, compaction of neurofilaments but no change in their number, and a loss of microtubules. Or, where intramyelinic spaces occurred there was an increased spacing between neurofilaments and microtubules with a significant loss in the number of both. Longitudinal sections showed foci of compaction of neurofilaments interspersed between regions where axonal structure was apparently normal. Neurofilament compaction was correlated with disruption of the axolemma at these foci present some hours after injury. We suggest that the time course of these axonal cytoskeletal changes after stretch-injury to central axons is shorter than those changes documented to occur during Wallerian degeneration. * To whom correspondence should be addressed. Introduction It is now widely accepted (Povlishock, 1992; Maxwell et al., 1997) that in human diffuse axonal injury (DAI) the majority of axons do not undergo immediate dis- ruption at the time of injury (primary axotomy), but rather are exposed to non-disruptive axonal injury (Maxwell et al., 1993) leading, after several hours, to disruption (secondary axotomy). Diffuse axonal injury is the most important type of brain damage occurring as a result of blunt head injury. It is a diffuse type of brain damage initiated at the moment of injury as has been shown in both human beings and experimental animals (Adams et al., 1982; Gennarelli et al., 1982), and is the commonest cause of coma in the absence of any intracranial mass lesion, of severe disability and the vegetative state after head injury (Adams, 1992). It is also becoming clear that there is probably a spectrum of axonal pathology, that has its correlates in clinical practice (Gennarelli, 1993), and in models of human blunt head injury (Povlishock, 1992; Maxwell et al., 1997). One of the earliest sequalae to non-disruptive axonal injury is the occurrence of axonal swellings (Povlishock, 1992) in the internodal region of injured nerve fibres. These axonal swellings are focal enlarge- ments of axonal diameter which are posited to precede secondary axotomy (Povlishock, 1992; Maxwell et al., 1997). The occurrence of axonal swellings has been attributed to a focal disruption of axonal transport (Povlishock, 1992; Christman et al., 1994; Maxwell, 1995; Pettus & Povlishock, 1996) manifested ultra-struc- turally as focal accumulations of neurofilaments and membranous organelles together with loss of micro- tubules (Povlishock et al., 1995; Maxwell et al., 1997). However, there has not been a morphometric analysis of changes in the axonal cytoskeleton associated with secondary axotomy after non-disruptive axonal injury. The purpose of this paper is to describe changes that occur within the axonal cytoskeleton in the internodal region of nerve fibres in non-disruptive (stretch) injury (Gennarelli et al., 1989; Maxwell et al., 1991, 1994, 1995), 0300–4864/97 ( 1997 Chapman and Hall
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Axonal cytoskeletal changes after non-disruptive axonal injury

Journal of Neurocytology 26, 207–221 (1997)
Axonal cytoskeletal changes after non-disruptiveaxonal injuryS. S. JAFARI1, W. L. MAXWELL 1*, M. NEILSON 1 and D. I. GRAHAM 2
1 Laboratory of Human Anatomy, Institute for Biomedical and Life Sciences, University of Glasgow, Glasgow G12 8QQ and2 University Department of Neuropathology, Institute of Neurological Sciences, Southern General Hospital,Glasgow G51 4TF, UK
Received 22 December 1995; revised 18 July, 22 October and 6 December 1996; accepted 12 December 1996
Summary
In animal models of human diffuse axonal injury, axonal swellings leading to secondary axotomy occur between 2 and 6 hafter injury. But, analysis of cytoskeletal changes associated with secondary axotomy has not been undertaken. We havecarried out a quantitative analysis of cytoskeletal changes in a model of diffuse axonal injury 4 h after stretch-injury to adultguinea-pig optic nerves. The major site of axonal damage was the middle portion of the nerve. There was a statisticallysignificant increase in the proportion of small axons with a diameter of 0.5 lm and smaller in which there was compaction ofneurofilaments. Axons with a diameter greater than 2.0 lm demonstrated an increased spacing between cytoskeletal elementsthroughout the length of the nerve. However, in the middle segment of the nerve these larger axons demonstrated twodifferent types of response. Either, where periaxonal spaces occurred, there was a reduction in axonal calibre, compaction ofneurofilaments but no change in their number, and a loss of microtubules. Or, where intramyelinic spaces occurred there wasan increased spacing between neurofilaments and microtubules with a significant loss in the number of both. Longitudinalsections showed foci of compaction of neurofilaments interspersed between regions where axonal structure was apparentlynormal. Neurofilament compaction was correlated with disruption of the axolemma at these foci present some hours afterinjury. We suggest that the time course of these axonal cytoskeletal changes after stretch-injury to central axons is shorter than
those changes documented to occur during Wallerian degeneration.Introduction
It is now widely accepted (Povlishock, 1992; Maxwellet al., 1997) that in human diffuse axonal injury (DAI)the majority of axons do not undergo immediate dis-ruption at the time of injury (primary axotomy), butrather are exposed to non-disruptive axonal injury(Maxwell et al., 1993) leading, after several hours, todisruption (secondary axotomy). Diffuse axonal injuryis the most important type of brain damage occurringas a result of blunt head injury. It is a diffuse type ofbrain damage initiated at the moment of injury as hasbeen shown in both human beings and experimentalanimals (Adams et al., 1982; Gennarelli et al., 1982), andis the commonest cause of coma in the absence of anyintracranial mass lesion, of severe disability and thevegetative state after head injury (Adams, 1992). It isalso becoming clear that there is probably a spectrumof axonal pathology, that has its correlates in clinicalpractice (Gennarelli, 1993), and in models of human
* To whom correspondence should be addressed.
blunt head injury (Povlishock, 1992; Maxwell et al.,1997).
0300–4864/97 ( 1997 Chapman and Hall
One of the earliest sequalae to non-disruptiveaxonal injury is the occurrence of axonal swellings(Povlishock, 1992) in the internodal region of injurednerve fibres. These axonal swellings are focal enlarge-ments of axonal diameter which are posited to precedesecondary axotomy (Povlishock, 1992; Maxwell et al.,1997). The occurrence of axonal swellings has beenattributed to a focal disruption of axonal transport(Povlishock, 1992; Christman et al., 1994; Maxwell, 1995;Pettus & Povlishock, 1996) manifested ultra-struc-turally as focal accumulations of neurofilaments andmembranous organelles together with loss of micro-tubules (Povlishock et al., 1995; Maxwell et al., 1997).However, there has not been a morphometric analysisof changes in the axonal cytoskeleton associated withsecondary axotomy after non-disruptive axonal injury.
The purpose of this paper is to describe changes thatoccur within the axonal cytoskeleton in the internodal
region of nerve fibres in non-disruptive (stretch) injury(Gennarelli et al., 1989; Maxwell et al., 1991, 1994, 1995),
J A
a model of human DAI, at 4 h after injury, a time pointwhen axonal swellings are frequent (Povlishock, 1992:Yaghmai & Povlishock, 1992; Povlishock et al., 1995;Maxwell et al., 1997).
Materials and methods
Material was obtained from six adult, male albino DuncanHartley guinea-pigs (Harlan UK) (range 650–750 gm).Animals were deeply anaesthetized with intramuscularketamine (50 mg kg[1) and xylazine (3 mg kg[1 ). The righteyelids were infiltrated with 1% xylocaine containing epi-nephrine. After retracting the lids with 4/0 silk sutures, theconjunctiva was opened through 360° and a sling of sterileumbilical tape placed over the posterior pole of the globe. Inthe three sham operated animals the sling was removed andanimals maintained under anaesthesia until they were killed4 h later. The three experimental animals were placed ina stereotactic frame, the headholder of which had 3° ofmovement to allow alignment of the line of force along theaxis of the optic canal thereby preventing bending of thenerve around the lip of the canal and causing undesirable,localised injury. The sling was then secured to a customdesigned stretch injury apparatus (Gennarelli et al., 1989).The same initial amount of force (30–40 g pre-load) wasplaced on each nerve such that there was no anatomicalslack in the nerve. Displacement of the globe occurred asa pulse generator activated a solenoid piston to provide aforce ranging between 150 and 180 g (plus the pre-load togive a total load of 180–220 g) over periods ranging from 19to 21 ms. A permanent record of the time course of eachexperiment was recorded either on a storage oscilloscopeand the trace photographed using a Polaroid Land camera,or on the hard disc of a PC fitted with a custom built datastorage board. All animals were maintained for 4 h underhalothane anaesthesia. Animals were terminally anaesthe-tised with intraperitoneal barbiturate and killed by trans-cardiac perfusion with 2.5% glutaraldehyde in 0.2 M PIPESbuffer (Baur & Stacey, 1977). The optic nerves were left insitu after perfusion fixation for at least 30 min before dissec-tion and removal of their whole length. Each experimentaland control nerve was cut into three segments, 4–6 mm inlength, the length of the guinea-pig optic nerve being12–20 mm (Gennarelli et al., 1989), segment 1 adjacent to theglobe, segment 2 the midportion and segment 3 the prechias-matic portion of each nerve and left in fixative for 1 h at 22° Cand a further 48 h at 4° C. The middle segment of the leftoptic nerve from each animal was processed as an internalcontrol for the quality of fixation in all of the animals.Segments were washed in 2% sucrose in PIPES buffer andpostfixed in 1% osmium tetroxide in PIPES buffer. Followingdehydration through graded concentrations of ethanol, spec-imens were embedded in Araldite using epoxypropane as anintermediary. Transverse semi-thin sections were obtainedfrom all segments of the nerve. Once the complete transversediameter of the optic nerve had been achieved, semi-thinsections were cut for the purpose of orientation of the block.Ten serial, transverse thin sections were cut from one ran-domly selected end of each segment of both the control and
208
experimental optic nerves, a total of 40 sections per animal.All ultrathin sections were stained with 12.5% methanolic
uranyl acetate and lead citrate. In order to achieve a repre-sentative sample (Williams, 1977) of transverse sections ofthe optic nerves, one randomly selected section out of tensections from each segment of the optic nerve was collectedfrom each animal, that is four sections for each animal. Thesesections were examined in either a Philips 301 or Philips 100TEM. After determination of the spatial extent of the changesin the axonal cytoskeleton from transverse sections, longitu-dinal sections from the two segments of the optic nerve inwhich the greatest degree of axonal damage occurred (seeResults), were examined in TEM. After cutting five longitu-dinal semithin sections to orientate the block, five grids ofserial longitudinal thin sections of segments 2 and 3 from thestretched-injured optic nerves of all experimental animals,a total of 30 grids, were cut. Sections on the third grid in theseries of sections were examined as were areas of particularinterest in the preceding and following sections.
In an attempt to determine whether all fibres in the opticnerve respond in the same way to stretch-injury, axonsand nerve fibres (axons]myelin sheath) were grouped in0.50 lm wide bins and analysed to determine whether therewere changes (1) in the proportion of axons/nerve fibreswithin bins after stretch-injury, and (2) in the spacing ofneurofilaments and microtubules within stretch-injured axons.
For changes in the proportion of axons/fibres within0.50 lm bins, the top left hand corner of each of the ran-domly selected transverse sections was identified. Startingfrom this grid square the top left hand corner of each thirdsuccessive grid square completely overlain by the section,(horizontally from left to the right) throughout the wholetransverse section of the optic nerve was photographedat]3240 magnification. This usually resulted in ten photo-graphs for each section giving a sample for each section ofcirca 700 nerve fibres. The TEM was calibrated using a S104line grating (2160 lines mm[1) (Agar Aids, Berkhamstead,UK) for each cassette of negatives taken. All photographicimages collected from the segments of the control and fourhour survival stretch-injured optic nerves, circa 16 800 fibres,were recorded in a Kontron Vidas image analysis system. Inorder to minimise inter-individual variation, all quantitativedata was collected by one individual (SSJ) who was ‘blinded’as to the particular source of each specimen. The followingparameters were measured for each fibre within the field ofeach negative: axon diameter (defined as the mean trans-verse diameter of the axoplasm), fibre diameter (defined asthe mean transverse diameter of the outer limits of themyelin sheath), the G-ratio (ratio of axonal diameter to fibrediameter; Guy et al., 1989), axonal cross-sectional area, fibrecross-sectional area, and the thickness of the myelin sheath.The results from this analysis (see below) demonstratedthat the greatest degree of change occurred in small axons.However, Gennarelli and colleagues (1989) have alsodocumented changes in the proportion of larger axons/fibres after stretch injury. Therefore we decided to furtherinvestigate changes in two groups of axons, small witha diameter of 0.5 lm or less and larger with an axonaldiameter of 2.0 lm or more.
In order to provide quantitative data to test the hypothesisthat changes in axonal calibre occur as a result of directmechanical alteration of the axonal cytoskeleton an analysisof changes in the spacing of neurofilament/microtubules
F A R I , M A X W E L L , N E IL S ON and GR A H A M
after stretch-injury to small and larger axons was carried out.

al
The following procedure was adopted. The top left handcorner of each randomly selected transverse section of anoptic nerve was found and used as an arbitrary startingpoint from which perpendicularly sectioned internodal por-tions of nerve fibres with axonal diameter of both 0.5 lm orless and 2.00 lm and above were photographed at]52 000.This magnification was determined by calibration of theTEM using beef catalase crystals (Agar Aids, Berkhamstead,UK) (Wrigley, 1968) where the last negative in any casetteof negatives was a photograph of a beef catalase crystal.A nerve fibre was selected as being transversely sectionedwhere the circularity factor was 0.8 or greater, and neurofila-ments and microtubules were sectioned perpendicular totheir longitudinal axis. The authors acknowledge thatneurofilaments/microtubules may be visualized in longitu-dinal section. But in transverse sections of axons from theguinea-pig optic nerve it is clear that neurofilaments inparticular are not regularly and uniformly distributedthroughout the axoplasm (Maxwell, 1996, and this paper). Itis therefore essential, in order to reduce labour and errorinduced by examination of serial longitudinal sections, thataxons be examined in transverse section. Axons within thebin sizes defined above were then regularly sampled as oneaxon in every third successive grid square (horizontally fromleft to the right) encountered throughout the whole trans-verse section of the optic nerve. This sampling procedurewas used to generate ten electron micrographs from eachtransverse section. In this way 20 axons (ten small and tenlarger ones) from each segment of control and injured opticnerves (24 thin sections), a total of 480 axons, were sampled.For each neurofilament/microtubule encountered withineach transverse section of an axon the spacing betweena particular neurofilament/microtubule and its nearest fourneighbouring neurofilaments/microtubules was measuredand the mean of these four distances for each neurofila-ment/microtubule was taken as the spacing between neuro-filaments/microtubules. The results for all axons, 20 fromeach segment in each control and in each experimental ani-mal, were used to calculate the mean spacing for each seg-ment in either the control or stretch-injured optic nerve fromeach animal. Results from this analysis demonstrated thatchanges in the spacing of components of the axonal cyto-skeleton occurred first throughout the length of larger axons,and second, within the second and third segments of smallaxons within the optic nerve. Since in small axons compac-tion of neurofilaments occurred only in the middle segmentof stretch-injured optic nerves and the proportion of axonspossessing a so-called periaxonal space (Maxwell et al., 1995)was greatest, we decided, first, to investigate cytoskeletalchanges in detail in that segment. These data provide thebasis for the present communication. Data for nerve fibreswith a periaxonal space was gathered as follows; five axonsfrom each animal, a total of 15 axons with a periaxonal space.The addition of this subgroup of axons provided an overalltotal number of axons analysed, including the right opticnerves from control animals, of 255 axons. Analysis of re-sponses by the axonal cytoskeleton to stretch-injury in thethird segment of the optic nerve will be presented in anothermanuscript (in preparation).
In an attempt to provide evidence to test the hypothesis
Axonal cytoskeletal changes after non-disruptive axon
that changes in spacing occur as a result of either changes indensity of neurofilaments/microtubules within a standard-
ized area of axoplasm (Pettus & Povlishock, 1996), or asa result of a change in their number the current study wasextended to provide data for changes in the density ofneurofilaments/microtubules within the cross-sectional areaof stretch-injured axons. The same micrographs that wereused for spacing measurements in the second segment ofcontrol and stretch-injured axons were used to calculatethe density of neurofilaments and microtubules within astandardised area of axoplasm, a circle with an area of0.0206 lm2, which was randomly superimposed over areasof the axoplasm. Data were collected in an image analysissystem. Neurofilaments and microtubules were digitizedwithin the circle, and their density within the standard areacomputed. A total of 780 areas over the whole sample ofcontrol and experimental animals were analysed. However,neurofilaments/microtubules may also be visualised in lon-gitudinal section. But in transverse sections it is clear that inthe small fibres of the optic nerve, neurofilaments in particu-lar are not regularly and uniformly distributed throughoutthe axoplasm (Maxwell, 1996, and this paper). To determinewhether there was a change in the number of neurofilamentsand/or microtubules in injured axons the total number ofeach in the same micrographs from control and stretch-injured axons in which neurofilaments and microtubuleswere perpendicularly sectioned, were counted. The totalnumber of microtubules and neurofilaments throughout theentire axonal cross-section of ten axons within each controland each experimental group, except for those five axons witha periaxonal space, in each segment from each animal werecounted. The results from all animals (n\6) allowed calcu-lation of the mean and SE of the number of neuro-filamentsand microtubules in 90 control and 30 stretch-injured smallaxons, 90 control and 30 stretch-injured larger axons, anda further 15 axons in which a periaxonal space was present.
Statistical analysis
Changes in axon and fibre parameters were most markedwithin two subgroups of nerve fibres, those with an axonaldiameter of 0.5 lm and less and those with an axonal dia-meter greater than 2.00 lm. Analysis for statistical signifi-cance of changes in the cytoskeleton of axons within thesetwo groups after stretch-injury was carried out in the follow-ing ways. The significance of changes in spacing, density perstandard or unit area of axoplasm and number of neurofila-ments and microtubules in small (\0.5 lm diameter) axonswas determined by means of the Student’s t-test. The signifi-cance of changes in the number of small axons in control andstretch-injured optic nerves was determined by means of thev2 test. Axons ([2.0 lm diameter) of larger nerve fibresdemonstrated two different changes after stretch-injury. Thesignificance of changes in the G-ratio, changes in the spacingand in the number of neurofilaments and microtubules be-tween the control axons and the two subgroups of nervefibres was determined by analysis of variance within 95%confidence intervals. The significance of the changes in theaxonal diameter between larger control fibres and fibreswith a periaxonal space, the G-ratio between control fibresand stretch-injured fibres with periaxonal spaces, was deter-mined by means of the Student’s t-test. The significance ofchanges in the density of neurofilament and microtubules
injury 209
per standard or unit area of axoplasm in larger axons wasdetermined by means of the Student’s t-test.

J
Results
CHANGES IN THE PROPORTIONS OFDIFFERENT SIZES OF AXON AND THESITE OF MAXIMAL AXONAL DAMAGE
Proportions of axons within 0.5 km bins in theoptic nerve
Comparison of the results for all of the axons fallingwithin 0.50 lm wide bins of the internodal axonaldiameter between 0.00 lm and 3.00 lm in each of thethree control and three experimental animals demon-strated a fivefold increase in the number/proportionof axons with a diameter\0.50 lm (Table 1) where thev2 test showed that the differences in probability werehighly significant (p\0.001). There was not, however,a significant change in the proportion of intermediateor larger axons after stretch-injury.
Changes in spacing between neurofilaments/microtubulesthroughout the length of the optic nerve
To provide evidence for diffuse or focal axonal cyto-skeletal responses to stretch-injury optic nerves fromour experimental animals were divided into threeparts, segment 1 adjacent to the globe, segment 2 in themiddle of the nerve and segment 3 in its prechiasmaticportion. There was no significant difference in neuro-filaments/microtubular spacing between all three seg-ments of right control and the left control optic nerves.However, analysis of randomly selected trans-verse sections from all three segments of stretch-in-
210
Change from control (%) ]48.7
(diameter[2.0 lm) there was an increase in spacingbetween (1) one neurofilament and its closest neigh-bouring four neurofilaments, and (2) one microtubuleand its closest neighbouring four microtubulesthroughout the entire length of injured larger axons(Table 2).
Statistical analysis (Student’s t-test) showed thatthe 33.36% increased spacing between neurofilamentswas significantly different from control values in allthree segments of the stretch-injured optic nerves(p\0.05). There was also a statistically significant in-crease in the spacing between microtubules in thesame axons, by 52.79% overall (p\0.05). But it is note-worthy that the increased spacing between micro-tubules was greatest in the second part of the nerve(Table 2). Further evidence in support of the hypothe-sis that the second segment of the optic nerve is thatportion of the length of the nerve where greatestaxonal damage occurred after stretch-injury was pro-vided by calculating the percentage of larger fibreswithin which periaxonal spaces (Maxwell et al., 1995)occurred. Periaxonal spaces occurred in 1.3% of largerfibres in segment 2, but only 0.14% and 0.41% of axonsin segments 1 and 3, respectively.
Changes in the cytoskeleton in small axons afterstretch-injury differed from those in larger axons inthat neurofilament relationships altered but those formicrotubules did not. Further there was not a similarchange throughout the length of these small axons.There was compaction of neurofilaments in the secondsegment but an increase in their spacing in the third
A F A R I , M A X W E L L , N E IL S ON and GR A H A M
Table 2. Mean spacing (nm) and (^SEM) between neurofilaments/microtubules in larger axons with axonal diameter[2.0 lm.
Control Experimental Experimental Experimentalaxons segment 1 axons segment 2 axons segment 3
Neurofilaments 75.51^0.51 100.98^5.4 99.96^4.1 101.18^3.8Change from control (%) ]33.73 ]32.37 ]33.99Microtubules 167.4^7.0 249.0^14 267.42^4.1 250.92^5.2
Table 1. Proportion of axons and nerve fibres falling within 0.5 lm wide bins in control (control) andstretch-injured (exp) optic nerves (^SEM).
Bin size (km) Control axon Control fibre Exp axon Exp fibre
0.00–0.50 1.58^0.46 Not applicable 9.48^1.60 Not applicable0.51–1.00 71.60^5.34 34.50^5.60 67.30^3.50 34.53^2.301.01–1.50 22.30^3.60 44.30^1.40 19.48^2.80 41.60^1.201.51–2.00 3.40^0.91 15.47^3.20 2.61^0.78 17.30^1.602.01–2.50 0.96^0.48 3.27^0.67 0.74^0.12 3.57^0.342.51–3.00 0.15^0.02 1.67^0.72 0.30^0.03 1.60^0.353.01–3.50 Not applicable 0.60^0.05 Not applicable 0.48^0.093.51–4.00 Not applicable 0.14^0.03 Not applicable 0.23^0.07
jured nerves demonstrated that in larger axons segment after stretch-injury (Table 3). There was
4 ]59.75 ]49.89

Table 3. Mean spacing (nm) and (^SEM) between neurofilaments/microtubules in small axons with axonal diameter\0.5 lm.
Control Experimental Experimental Experimentalaxons segment 1 axons segment 2 axons segment 3
Neurofilaments 122.89^3.9 160.9^14 57.84^8.4 193.0^14Change from control (%) ]30.9 [52.9 ]57.05Microtubules 102.0^4.3 105.74^5.3 109.9^7.5 104.09^0.14
Axonal cytoskeletal changes after non-disruptive axonal injury 211
Change from control (%) ]3.67 ]7.7 ]2.5
a significant decrease, by 53%, in the second or middlesegment (p\0.05) but a significant increase, by 57%, inneurofilament spacing (p\0.05) in the prechiasmaticportion (Table 3). The increase in neurofilament spac-ing in the first segment was not statistically significant.
MORPHOMETRIC ANALYSES IN THE SECONDSEGMENT OF STRETCH-INJURED OPTIC NERVES
Reduction of axonal diameter and the G-ratio in largerfibres with a periaxonal space
In fibres in which a so-called periaxonal space (Fig. 1a)(Maxwell et al., 1995) occurred there was a significantreduction in axonal diameter (p\0.05) but not in theoverall diameter of the myelin sheath limiting thefibre. This resulted in loss of the intimate relationshipof the axolemma with the internal aspect of the myelinsheath (Fig. 1a, b). The G-ratio in these fibres was re-duced from 0.71 to 0.50, that is a 30% reduction. Analy-sis of variance of the G-ratio of all fibres (control, allfibres in stretched-injured optic nerves and largerfibres with a periaxonal space) showed a statisticallysignificant difference in the G-ratio between thesegroups (F\99.48, df\2]6, p\0.001). In addition,intergroup comparison of 95% confidence intervalsbetween fibres with a periaxonal space and controlsshowed a highly significant difference (p\0.05).
Changes in neurofilament and microtubule spacingand their density
Since there was an increased proportion of axons of\0.50 lm diameter (Table 1) and in some larger fibresthere was a change in the G-ratio that indicateda reduction of axonal calibre in those fibres, wedecided to analyse in more detail changes in thespacing and density/standard or unit area of axo-plasm in control and stretch-injured axons in twogroups of nerve fibre, those with an axon dia-meter\0.50 lm and those with a diameter[2.00 lm(Tables 4 and 5) (Figs 1a, b, 2a, b, 3a, b, 4a, b).
Changes in small axons (\0.5 km in diameter). Quantita-tive analysis of components of the axonal cytoskeletonshowed that in small axons (diameter\0.50 lm) there
was a significant reduction by 52.9% in spacing(p\0.05) or increase by 254.5% in density of neurofila-ments per standard or unit area of axoplasm (p\0.05)(Fig. 2a, b) but no significant change in either spacingor density for microtubules (Tables 4 and 5).
Larger fibres and axons. Larger fibres 4 h after stretch-injury fell into two subclasses. Either those nervefibres in which intramyelinic spaces and myelin intru-sions occurred or those in which periaxonal spacesoccurred (Figs 1a, b, 4a, b). The changes in cytoskeletalrelationships in the axons of these two subclasses offibre differed both from those in small axons andbetween the two groups. In the axons of those nervefibres within which intramyelinic spaces occurredneurofilament and microtubule spacing was increasedby 32.3% and 59.8% respectively (Table 4). This in-creased spacing was correlated with a reduced den-sity/unit area of axoplasm by 58.98% and 56.48%respectively (Table 5). On the other hand, in thoselarger fibres in which periaxonal spaces occurred therewas a reduction by 24.2% in spacing between neuro-filaments (Table 4). Their density/unit area was in-creased by 86% (Table 5). These results provide goodevidence for compaction of neurofilaments (Fig. 1a, b)in larger fibres in which periaxonal spaces occurred.But, changes in respect of microtubules were the oppo-site with a 29.7% increase in spacing (Table 4) ora 52.77% decrease in density/unit area (Table 5). Thus,after stretch-injury, in larger nerve fibres with intra-myelinic spaces there was increased spacing betweenneurofilaments/microtubules. However, there was re-duced spacing between neurofilaments but an in-creased spacing between microtubules in larger nervefibres within which periaxonal spaces occurred.
Analysis of variance for neurofilament spacingwithin the axons of these larger fibres showed thatthere was a highly significant difference between thegroups (F\72.86, df\2]6, p\0.001). Intergroupcomparisons using 95% confidence intervals showedthat filament spacing in stretch-injured larger axons ofoptic nerves was significantly higher than in controls(p\0.05) except in nerve fibres where there was aperiaxonal space. In this case, neurofilament spacingwas significantly reduced from control values
(p\0.05). However, changes within stretch-injured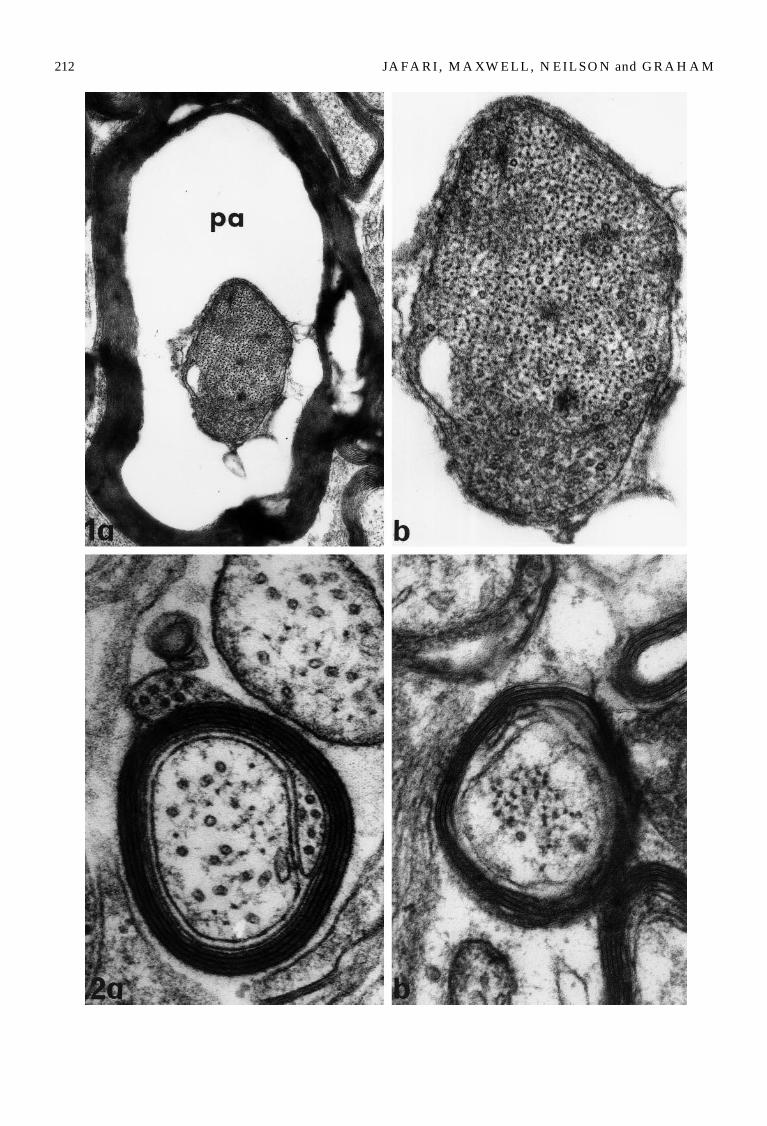
212 M
J A F A R I , M A X W E L L , N E IL S ON and GR A H A
(%)
axons of nerve fibres where intra-myelinic spaces andperiaxonal spaces occurred after stretch-injury.
in which intramyelinic spaces occurred after stretch-injury and which demonstrated both increased
Fig. 1. (a and b) A transverse thin section of a large myelinated nerve fibre 4 h after stretch-injury demonstrating(a) a periaxonal space (pa) separating the axon from its myelin sheath and (b) the reduced interneurofilament spacing withinthe axoplasm. (a)]41 900, (b)]99 900.
Fig. 2. (a and b) Transverse thin sections from (a) control and (b) stretch-injured small nerve fibres. (a) The control fibreshows a uniform disposition of the myelin sheath, the high relative number of microtubules and the relatively wide spacing ofneurofilaments within the axoplasm. (b) In a small fibre 4 h after stretch-injury the myelin sheath demonstrates a degree of
Table 4. Mean spacing (nm) and (^SEM) between neurofilaments/microtubules and their nearest four neighbours (combinedmeans from ten axons from each segment in each animal in each group, n\3, and from five axons in which a periaxonal spacewas present).
Control Experimental Control Experimental Experimental(small axons) (small axons) (larger axons) (larger axons) axons with\0.50 km \0.50 km [2.00 km [2.00 km periaxonal space
Neurofilaments 122.9^3.9 57.8^4.8 75.5^0.5 99.9^4.1 57.2^1.2Change from control [52.9 ]32.3 [24.2(%)
Microtubules 102.0^4.3 109.9^7.5 167.4^7.0 267.4^4.1 217.1^12Change from control ]7.8 ]59.8 ]29.7(%)
Table 5. Mean density (^SEM) of microtubules and neurofilaments 0.0206 lm[2 axoplasm (combined means from ten axons(five with a periaxonal space) from each segment from each control and experimental animal (n\3 in each group).
Control Experimental Control Experimental Experimental(small axons) (small axons) (larger axons) (larger axons) axons with\0.50 km \0.50 km [2.00 km [2.00 km periaxonal space
Neurofilaments 2.52^0.10 8.43^0.76 5.51^0.18 2.26^0.11 10.26^0.55Change from control ]254.36 [58.98 ]86.20(%)
Microtubules 3.71^0.05 2.73^0.24 1.08^0.05 0.47^0.03 0.51^0.02Change from control [26.41 [56.48 [52.77
Axonal cytoskeletal changes after non-disruptive axonal injury 213
axons did not only result in altered spacing for neuro-filaments. Analysis of variance for spacing betweenadjacent microtubules demonstrated a significant in-crease (F\34.05, df\2]6, p\0.001) both in fibreswith intramyelinic and periaxonal spaces. Thus micro-tubule spacing was increased even in fibres whereneurofilament compaction occurred 4 h after stretch-injury.
The density/unit area of axoplasm of both neuro-filaments and microtubules in axons of larger fibreswith intramyelinic spaces decreased significantly(p\0.05). But in axons with a periaxonal space therewas a significant decrease (p\0.05) only for micro-tubules while the density of neurofilaments increased(p\0.05). There is therefore a differential response in
disrupted structure. There is an almost complete absence of micform a closely packed group within the centre of the axon. (a)
Changes in total number of neurofilaments andmicrotubules in axonsCounts of the means of the total number of neurofila-ments and microtubules in cross-sections of 180 con-trol and 60 stretch-injured axons from the three controland three experimental animals, including 15 axons infibres with a periaxonal space provided the data givenin Table 6.
Small axons. In small axons, after stretch-injury, therewas a significant increase in the number of neurofila-ments (p\0.05) (Table 6). But there was not a signifi-cant change in the number of microtubules (Table 6).
Larger axons with intramyelinic spaces. In larger axons
rotubules from the axoplasm in this axon. The neurofilaments]127 000, (b)]126 150.

214 M
J A F A R I , M A X W E L L , N E IL S ON and GR A H A
(%)
jury, there was a wide variation in axonal diameterover tens of micrometers in the internode (Fig. 7a) tered between regions, about 8–10 lm in length,
Fig. 3. (a and b) A transverse, thin section of a control, large myelinated nerve fibre. (a) At relatively low magnification,the myelin sheath has a close association with the axolemma. (b) A detail of the axoplasm of part of the same fibre from withinthe outlined area in (a) to show the relatively high number of microtubules and the regular spacing of neurofilaments withinthe axoplasm. (a)]30 950, (b)]84 150.
Fig. 4. (a and b) A transverse thin section of a larger, myelinated nerve fibre 4 h after stretch-injury. (a) There is separation ofthe myelin sheath from the axolemma and the axonal profile is irregular. The components of the axonal cytoskeleton are notregularly spaced throughout the axoplasm. (b) A detail of part of the axoplasm from within the area outlined in (a) to show thealmost complete absence of microtubules (one is indicated by the arrow) and the increased spacing between neurofilaments.
Table 6. Mean number (^SEM) of neurofilaments and microtubules in cross-sections of axons in each control and experi-mental group (mean calculated from ten small and ten larger axons within each segment in each animal, a total sample of 240axons, and five axons with a periaxonal space, from each experimental animal; n\3).
Control Experimental Control Experimental Experimental(small axons) (small axons) (larger axons) (larger axons) axons with\0.50 km \0.50 km [2.00 km [2.00 km periaxonal space
Neurofilaments 7.4^1.3 14.8^1.0 246.5^9.0 179.9^4.1 281.9^8.2Change from control ]99.9 [27.0 ]14.4(%)
Microtubules 11.4^2.1 9.6^1.1 74.6^5.5 33.3^1.9 20.7^0.3Change from control [15.6 [55.4 [72.2
Axonal cytoskeletal changes after non-disruptive axonal injury 215
neurofilament and microtubule spacing, analysis ofvariance showed that there was a statistically signifi-cant loss of both neurofilaments (F\48.81, df\2]6,p\0.001) (Table 6) and microtubules (F\69.87,df\2]6, p\0.001).
Larger axons with periaxonal spaces. In larger axons witha periaxonal space in which neurofilament compactionoccurred within the axon there was no significantchange in the number of neurofilaments. However, inthese axons there was a highly significant (p\0.001)loss of axonal microtubules.
Longitudinal sections
Examination of longitudinal sections again suggesteda differential response by cytoskeletal components instretch-injured small and larger axons. In the former,compaction occurred over lengths of axons in excess of5 lm to result in a central core of neurofilaments witha reduced spacing (Fig. 5). In larger axons, however,cytoskeletal changes occurred throughout a muchgreater length of an axon. There was, however, a widevariation in such changes between fibres or partsof the same fibre. First, perhaps the least amount ofaxonal damage may be seen where axonal calibre wasuniform but where microtubules in particular adopteda coiled or helical disposition within the axoplasm(Fig. 6). Second, with perhaps a greater degree of in-
(a)]21 700, (b)]89 700.
perhaps comparable to the ‘beaded’ appearance ofinjured nerve fibres after ‘light stretch’ (Ochs et al.,1987, 1994, 1996) or accumulation of membranousorganelles after transection (Ballin & Thomas, 1969;Donat & Wisniewski, 1973; Ishise & Rosenbluth, 1986).In the particular example in Fig. 7a, axonal diametervaried from a minimum of 0.8 lm to a maximum of2.03 lm. At the site of minimum axonal diameter (in-dicated by white arrows in Fig. 7a and at higher mag-nification in Fig. 7b) the myelin sheath was grosslydisrupted with separation of myelin lamellae. Theaxoplasm in this region contained a relatively largenumber of longitudinally orientated neurofilamentswhich were closely packed within the axoplasm.Microtubules were largely absent in this region(Fig. 7b). Disorganisation of the axonal cytoskeletonwas greater in regions of increased axonal diameter(area between the dotted lines in Fig. 7a and at highermagnification in Fig. 7c). Although some neuro-filaments still lay parallel to the longitudinal axis ofthe axon, others were now orientated transversely (ar-rows in Fig. 7c) and the spacing between adjacentneurofilaments was greatly increased (Fig. 7c). None-theless, neurofilaments were still recognisable, linearstructures. Third, in examples perhaps indicative ofthe greatest degree of damage in longitudinal sections,although a linear organisation of neurofilaments waspresent, there were multiple foci of neurofilamentcompaction, extending for between 3 and 5 lm, scat-

216 A M
J A F A R I , M A X W E L L , N E IL S ON and GR A H
between neurofilaments and between microtubules. Inaxon and over several hours after stretch-injury. Weprovide additional quantitative data that confirms the
l
Fig. 5. A longitudinal thin section of a small nerve fibre after stretch-injury. The myelin sheath is largely intact. But the axonalcytoskeleton consists of numbers of neurofilaments forming a central core within the axoplasm. ]84 000.
Fig. 6. A longitudinal section of a damaged, larger never fibre in which the components of the axonal cytoskeleton are nolonger longitudinally orientated. Microtubules (arrow) have a helical disposition within the central region of the axoplasm.The peripheral part of the axoplasm is electron lucent. ]32 400.
Fig. 7. (a) A photomontage of a longitudinal section of the internode of a damaged larger nerve fibre. The axon has a variablecalibre. Where the axonal diameter is least there is the greatest disruption of the myelin sheath (arrows). ]22 000. Two parts ofthis fibre are shown at higher magnification. (b) The region of minimum diameter. Here neurofilaments are basicallylongitudinally orientated but with an increased density per unit area of axoplasm. (c) The region of maximum diameter. Herethere is loss of a linearly organised cytoskeleton. Rather neurofilaments have no preferred orientation. Indeed some
where neurofilament spacing was apparently normal(Fig. 8a). At these foci of compaction there was loss ofthe continuity and integrity of the axolemma as re-vealed in both longitudinal and transverse sections(Fig. 8a–c) such that large holes appeared to be presentin the axolemma. In longitudinal sections fragments ofthe membrane extended into the periaxonal space(Fig. 8b). In longitudinal and transverse sections therewas loss of the intimate relationship between theaxolemma, the axonal cytoskeleton and the internalaspect of the myelin sheath together with gross distor-tion of the myelin to form ‘balloon’ like expansions(Fig. 8a–c). At these foci of axolemma disruption therewas compaction of the remaining neurofilamentouscomponent of the axonal cytoskeleton (Fig. 8c).
Discussion
We provide the first quantitative analysis of changesin the axonal cytoskeleton associated with secondaryaxotomy in CNS myelinated fibres after non-disrup-tive axonal injury to those fibres. First, in the stretch-injury model in the guinea-pig optic nerve the majoraxonal response occurs in the mid-region of the lengthof the axon. Second, small and larger axons responddifferently to stretch-injury. In small axons there isposttraumatic compaction of neurofilaments and anincrease in their number, together with an increase inthe proportion of small axons within the optic nerve.We interpret the latter as a reduction in axonal calibrein some axons. There is no statistically significantchange in the number of microtubules in small axons4 h after stretch-injury. But in the internode of largeraxons one of two different responses occur. Whereperiaxonal spaces occur there is focal compaction ofneurofilaments and a reduction in axonal calibre, to-gether with loss of microtubules. But there is no signif-icant change in the number of neurofilaments.Neurofilament compaction is most marked wherethere is loss of the integrity of the axolemma andseparation of the membrane from the axonal cyto-skeleton. In those parts of axons where intramyelinicspaces occur, however, there is an increased spacing
Axonal cytoskeletal changes after non-disruptive axona
neurofilaments run transversely (arrow) through the axoplasm
these axons there is a significant reduction in thenumber of both neurofilaments and microtubules.
Earlier analyses of the site of axonal injury fromthe stretch-injured guinea pig optic nerve suggestedthat the most severe abnormalities occurred in theprechiasmatic portion of the nerve (Gennarelli et al.,1989). But within that study precise details were notgiven of the survivals after stretch-injury at whichdata, morphological or quantitative, were gathered.The major criterion for damaged nerve fibres was anincrease in axoplasmic area. Indeed, Gennarelli andcolleagues (1989) suggested that injured fibres wereenlarged throughout their course. We confirmed thatcytoskeletal changes occurred throughout the lengthof larger axons. However, we extend that study toprovide data that axons also demonstrated reducedcalibre and that these changes were most numerous inthe middle portion of the length of the optic nerve.Since Gennarelli and colleagues (1989) do not givedetails of the survival of animals used for TEM itcannot be stated whether that material was takeneither before or after the time point when secondaryaxotomy is hypothesized to have occurred. It ispossible, therefore, that their morphological andquantitative results reflect changes occurring duringWallerian degeneration where the assumption of anirregular axonal profile is well documented (Ballin& Thomas, 1969; Martinez & Friede, 1970; Donat& Wisniewski, 1973; Schlaepfer, 1974; Hoffman et al.,1985) between 24 and 36 h of axotomy. However, thepurpose of the present study was to determine cyto-skeletal changes prior to secondary axotomy. It is ac-knowledged in the current paper that there are alsostatistically significant changes in the cytoskeleton ofstretch-injured axons in the prechiasmatic segment ofthe guinea pig optic nerve. Investigation of thesechanges is currently underway.
The current study extends recently published find-ings concerning changes in the axonal cytoskeleton atnodes at Ranvier (Maxwell, 1996) after stretch-injury.The present results, therefore demonstrate that cyto-skeletal changes occur over considerable lengths of an
injury 217
. ]44 000.

218 M
J A F A R I , M A X W E L L , N E IL S ON and GR A H A
ferences in detailed changes within the axonal cyto-skeleton in different forms of axonal injury. Earlier present paper extends the latter study to show that
l
Fig. 8. Longitudinal and transverse sections through severely damaged nerve fibres. (a) In this longitudinal section the axonalcytoskeleton still forms a continum but there are a number of foci of compaction of neurofilaments (arrows) separated byregions where the axon has a more normal structure. ]22 000. In this longitudinal (b) and transverse (c) section at a locus ofneurofilament compaction there is disruption of the axolemma such that fragments extend into the periaxonal space
hypothesis (Povlishock & Christman, 1995; Maxwellet al., 1997) that a spectrum of pathological changesoccur after stretch-injury/non-disruptive axonal in-jury. Recent work has shown that compaction ofneurofilaments in the internode of non-disruptivelyinjured large, central nerve fibres occurs within min-utes of injury and lasts for at least 6 h (Pettus & Pov-lishock, 1996). However, our results differed fromthose of other studies (Pettus et al., 1994; Pettus &Povlishock, 1996; Ochs et al., 1994) in that we obtainedevidence for two types of axonal response to the ap-plication of stretch-injury. We confirm the findings ofPettus and colleagues (1994) that there is posttrau-matic compaction of the axonal cytoskeleton in someparts of some axons. But, in addition, we providenovel evidence for an increased spacing or expansionbetween cytoskeletal elements in other parts of stretch-injured axons, in this particular experiment in thesecond segment of the optic nerve. This study alsoprovides novel evidence that small axons are damagedby stretch-injury in that we demonstrate compactionof neurofilaments in those axons. However, the longterm outcome of such cytoskeletal changes has yet tobe determined.
Neurofilaments have been implicated in the main-tenance of axonal calibre (Friede & Samorajski, 1970;Hoffman et al., 1984, 1985; Reles & Friede, 1991) andearly work suggested that this is a simple function oftheir number (Friede & Samorajski, 1970; Lasek et al.,1983). But more recent work has questioned this gen-eral concept since neurofilament density, for example,has been documented to vary between central axons(Szaro et al., 1990) in different tracts of the CNS, oreven along a single, central (Berthold, 1982; Price et al.,1988; de Waegh et al., 1992) or peripheral axon (Reles& Friede, 1991). Microtubules, on the other hand, donot appear to play an important role in regulation ofaxonal diameter but are involved in fast axonal trans-port. Comparison of results obtained in the presentstudy with those cytoskeletal changes documented inother types of axonal injury, for example transectionor crush (Ballin & Thomas, 1969; Martinez & Friede,1970; Donat & Wisniewski, 1973; Hoffman et al., 1984,1985; Ishise & Rosenbluth, 1986; Glass & Griffin, 1991;McHale et al., 1995) or maintained tension/‘lightstretch’ upon nerve fibres during fixation (Ochs &Jersild, 1987; Ochs et al., 1994) allows analysis of dif-
Axonal cytoskeletal changes after non-disruptive axona
(curved arrow) and the neurofilament core is no longer limitedthe myelin sheath to form irregular figures limiting the periph
studies of Wallerian degeneration between 24 and36 h after transection of the rat sural nerve (Ballin& Thomas, 1969), the sciatic nerve in rats, rabbits andcats (Martinez & Friede, 1970; Donat & Wisniewski,1973), the rat saphenous nerve (Schlaepfer, 1974) andcrush injury of the rat sciatic nerve (Hoffman et al.,1985) showed the assumption of an irregular axonalprofile (‘beading’), loss/fragmentation of micro-tubules and neurofilaments and a granular trans-formation of the axoplasm to give a flocculentultrastructure over several centimetres of axons(Donat & Wisniewski, 1973). More recently, however,compaction of neurofilaments to form so-called ‘con-densed filamentous cores’ has been demonstratedwithin 24 h of axonal transection in the lamprey(McHale et al., 1995). There is, therefore, recently ob-tained evidence that neurofilament compaction pos-sibly precedes cytoskeletal loss noted by earlierworkers in Wallerian degeneration. But there is alsoa clear difference in the time course for the develop-ment of an irregular axonal profile or ‘beading’ afterstretch-injury and changes after transection/primaryaxotomy. In animal models of human blunt head in-jury there is rapid loss of axonal microtubules at nodesof Ranvier (Maxwell, 1996) and in the internode oflarge, pontomedullary axons (Pettus & Povlishock,1996). The present paper and the recent literature haveshown that compaction of neurofilaments occurred inboth small axons upto 4 h (this paper) and in largeaxons for at least 6 h after injury (Pettus & Povlishock,1996). In comparison, under conditions where axonsare maintained under mechanical loading during fix-ation, there is compaction of both microtubules andneurofilaments after ‘light stretch’ in the rat (Ochs& Jersild, 1987; Ochs et al., 1994). In the present paperanalysis of changes in the axonal cytoskeleton wasundertaken before the hypothesised time at whichsecondary axotomy might occur (Povlishock, 1992;Maxwell et al., 1997). Therefore we suggest that neuro-filament compaction in stretch-injured central nervefibres precedes the event of axotomy and share theviews expressed by McHale and colleagues (1995) thatafter calcium influx into injured nerve fibres thereis localized collapse of neurofilaments into tightlypacked bundles in the centre of an axon as a result ofeither partial proteloysis of neurofilaments or a combi-nation of partial proteolysis and dephosphorylationof neurofilament sidearms (Hall & Lee, 1995). The
injury 219
by the axolemma (double arrows). There is also disruption ofery of the periaxonal space. (b)]26 000, (c)]58 000.

J
neurofilament compaction after stretch-injury may becorrelated with loss of the integrity of the axolemmaseveral hours after injury.
Acknowledgements
Saeed S. Jafari was supported throughout this studyby a postgraduate studentship from the Ministry ofHealth and Medical Education of the Islamic Republicof Iran. The authors would like to acknowledge thecontribution to this study provided by Professor T. A.
References
ADAMS, J . H. (1992) Head injury. In Greenfield’s Neuro-pathology, 5th Edition (edited by ADAMS, J . H.& DUCHEN, L. W.) pp. 106–52. London: EdwardArnold.
ADAMS, J . H. , GRAHAM, D. I . , MURRAY, L. S. & SCOTT,G. (1982) Diffuse axonal injury due to non-missile headinjury in humans: an analysis of 45 cases. Annals ofNeurology 12, 557–63.
BALLIN, R. H. M. & THOMAS, P. K. (1969) Changes atthe nodes of Ranvier during Wallerian degeneration:an electron microscope study. Acta Neuropathologica 14,237–49.
BAUR, P. S. & STACEY, T. R. (1977) The use of PIPES bufferin the fixation of mammalian and marine tissues forelectron microscopy. Journal of Microscopy 100, 315–27.
BERTHOLD, C. (1982) Some aspects of the ultrastructuralorganization of peripheral myelinated axons in the cat.In Axoplasmic Transport (edited by WEISS, D.) pp.40–54. New York: Springer-Verlag.
CHRISTMAN, C. W.,GRADY, M. S., WALKER, S. A. ,HOLLOWAY, K. L. & POVLISHOCK, J . T. (1994)Ultrastructural studies of diffuse axonal injury in hu-mans. Journal of Neurotrauma 11, 173–86.
DE WAEGH, S. , LEE, V. M.-Y. & BRADY, S. T. (1992) Localmodulation of neurofilament phosphorylation, axonalcaliber, and slow axonal transport by myelinatingSchwann cells. Cell 68, 451–63.
DONAT, J . R . & WISNIEWSKI, H. M. (1973) The spatio-temporal pattern of Wallerian degeneration in mam-malian peripheral nerves. Brain Research 53, 41–53.
FRIEDE, R. L. & SAMORAJSKI, T. (1970) Axon calibrerelates to neurofilaments and microtubules in sciaticnerve fibres of rats and mice. Anatomical Record 167,379–87.
GENNARELLI, T. A. (1993) Cerebral concussion and diffusebrain injuries. In Head Injury (edited by COOPER, P. R.)pp. 137–58. Baltimore: Williams & Wilkins.
GENNARELLI, T. A. , THIBAULT, L. E. , ADAMS, J . H. ,GRAHAM, D. I. , THOMPSON, C. J . & MARCINCIN,R. P. (1982) Diffuse axonal injury and traumatic coma inthe primate. Annals of Neurology 12, 564–74.
GENNARELLI, T. A. , THIBAULT, L. E. , TIPPERMAN, R. ,TOMEI, G. , SERGOT, R. , BROWN, M., MAXWELL,W. L. , GRAHAM, D. I. , ADAMS, J . H. , IRVINE, A.
220
GENNARELLI, L. M., DUHAIME, A. C., BOOCK, R. &GREENBERG, J. (1989) Axonal injury in the optic nerve:
Gennarelli by the development of the stretch-injuryapparatus. WLM would like to acknowledge supportby the Wellcome Trust for the purchase of a Philips100 TEM and for travel during this study. The authorswould like to express their deep appreciation to Pro-fessor A. P. Payne of the Laboratory of Human Anat-omy, IBLS, University of Glasgow for his unstintingsupport and invaluable advice concerning statisticaltechniques employed in this study. The authors wouldlike to acknowledge technical assistance provided byEuan W. Milne, Caroline Crossan and Jane Patterson.
a model simulating diffuse axonal injury in the brain.Journal of Neurosurgery 71, 315–25.
GLASS, J . D. & GRIFFIN, J . W. (1991) Neurofilament redis-tribution in transected nerves: evidence for bidirectionaltransport of neurofilaments. Journal of Neuroscience 11,3146–54.
GUY, J . , ELLIS, A. , KELLEY, K. & HOPE, G. M. (1989)Spectra of G ratio, myelin sheath thickness, and axonand fibre diameter in the guinea pig optic nerve. Journalof Comparative Neurology 287, 446–54.
HALL, G. F. & LEE, V. M.-Y. (1995) Neurofilament sidearmproteolysis is a prominent early effect of axotomy inlamprey giant central neurons. Journal of ComparativeNeurology 353, 38–49.
HOFFMAN, P. N., GRIFFIN, J . W. & PRICE, D. C. (1984)Control of axonal calibre by neurofilament transport.Journal of Cell Biology 99, 705–14.
HOFFMAN, P. N. , THOMPSON, G. W., GRIFFIN, J . W.& PRICE, D. L. (1985) Changes in neurofilament trans-port coincide temporally with alterations in the calibreof axons in regenerating motor fibres. Journal of CellBiology 101, 1332–40.
ISHISE, J . & ROSENBLUTH, J . (1986) Nodal and paranodalstructural changes in frog optic nerve during early Wal-lerian degeneration. Journal of Neurocytology 15, 657–70.
LASEK, R. J . , OBLINGER, M. & DRAKE, P. (1983) Molecu-lar biology of neuronal geometry: expression of neuro-filament genes influences axonal diameter. Cold SpringHarbour Symposia on Quantitative Biology 18, 731–44.
MARTINEZ, A. J . & FRIEDE, R. L. (1970) Accumulation ofaxoplasmic organelles in swollen nerve fibres. BrainResearch 19, 183–98.
MAXWELL, W. L. (1995) Microtubular changes in axonsafter stretch-injury. Journal of Neurotrauma 12, 363.(Abstract).
MAXWELL, W. L. (1996) Histopathological changes at cen-tral nodes of Ranvier after stretch-injury. Microscopy andResearch Technique 34, 522–35.
MAXWELL, W. L. , IRVINE, A. , GRAHAM, D. I . , ADAMS,J. H. , GENNARELLI, T. A. , TIPPERMAN, R. &STURATIS, M. (1991) Focal axonal injury: the earlyaxonal response to stretch. Journal of Neurocytology 20,157–64.
MAXWELL, W. L. , WATT, C. , GRAHAM, D. I . & GEN-
A F A R I , M A X W E L L , N E IL S ON and GR A H A M
NARELLI, T. A. (1993) Ultrastructural evidence ofaxonal shearing as a result of lateral acceleration of the

l
head in non-human primates. Acta Neuropathologica 86,136–44.
MAXWELL, W. L., ISLAM, M. N., GRAHAM, D. I .& GENNARELLI, T. A. (1994) A qualitative and quant-itative analysis of the cell soma response to stretchinjury in the optic nerve of the adult guinea pig. Journalof Neurocytology 23, 379–92.
MAXWELL, W. L. , MCCREATH, B. J . , GRAHAM, D. I .& GENNARELLI, T. A. (1995) Cytochemical evidencefor redistribution of membrane pump calcium-ATPaseand ecto-Ca-ATPase activity, and calcium influx in my-elinated nerve fibres of the optic nerve after stretchinjury. Journal of Neurocytology 24, 925–42.
MAXWELL, W. L. , POVLISHOCK, J . T. & GRAHAM, D. I .(1997) A mechanistic analysis of non-disruptive axonalinjury. Journal of Neurotrauma 14, in press.
MCHALE, M. K., HALL, G. F. & COHEN, M. J. (1995) Earlycytoskeletal changes following injury of giant spinalaxons in the lamprey. Journal of Comparative Neurology353, 25–37.
OCHS, S., & JERSILD, R. A (1987) Cytoskeletal organellesand myelin structure of beaded nerve fibres. Neurosci-ence 22, 1041–56.
OCHS, S. , JERSILD, R. A., POURMANDS, R. & POTTER,C. G. (1994) The beaded form of myelinated nervefibres. Neuroscience 61, 361–72.
OCHS, S. , POURMAND, R. & JERSILD, R. A. (1996) Originof beading constrictions at the axolemma: presence inunmyelinated axons and after b,b@-iminodipropionitriledegradation of the cytoskeleton. Neuroscience 70, 1081–96.
PETTUS, E. H. & POVLISHOCK, J . T. (1996) Characteriza-tion of a distinct set of intra-axonal ultrastructuralchanges associated with traumatically induced alter-ation in axolemma permeability. Brain Research 722, 1–11.
PETTUS, E. H., CHRISTMAN, C. W., GIEBEL, M. L. &POVLISHOCK, J . T. (1994) Traumatically inducedaltered membrane permeability: its relationship to
Axonal cytoskeletal changes after non-disruptive axona
traumatically induced reactive axonal change. Journal ofNeurotrauma 11, 507–22.
POVLISHOCK, J . T. (1992) Traumatically induced axonalinjury: pathogenesis and pathobiological implications.Brain Pathology 2, 1–12.
POVLISHOCK, J . T. & CHRISTMAN, C. W. (1995) Thepathobiology of traumatically induced axonal injury inanimals and humans: a review of current thoughts.Journal of Neurotrauma 12, 555–62.
POVLISHOCK, J . T. , GIEBEL, M. L. & PETTUS, E. (1995)Traumatically induced alterations in axolemma per-meability are associated with a distinct subset of intra-axonal cytoskeletal changes. Journal of Neurotrauma 12,417.
PRICE, R. L. , PAGGI, P. , LASEK, R. J . & KATZ, M. J .(1988) Neurofilaments are spaced randomly in theradial dimension of axons. Journal of Neurocytology 17,55–62.
RELES, A. & FRIEDE, R. L. (1991) Axonal cytoskeleton atthe nodes of Ranvier. Journal of Neurocytology 20, 450–8.
SCHLAEPFER, W. W. (1974) Calcium-induced degener-ation of axoplasm in isolated segments of rat peripheralnerve. Brain Research 69, 203–15.
SZARO, B. G. , WHITNALL, M. H. & GAINER, H. (1990)Phosphorylation-dependent epitopes on neurofilamentproteins and neurofilament densities differ in axons inthe corticospinal and primary sensory dorsal columntracts in the rat spinal cord. Journal of ComparativeNeurology 302, 220–35.
WILLIAMS, M. A. (1977) Quantitative methods in biology.In Volume 6: Practical methods in electron microscopy(edited by GLAUERT, A. M.), Amsterdam: NorthHolland Publishing Company.
WRIGLEY, N. G. (1968) The lattice spacing of crystallinecatalase as an internal standard of length in electronmicroscopy. Journal of Ultrastructure Research 24, 454–64.
YAGHMAI, A. & POVLISHOCK, J. T. (1992) Traumaticallyinduced reactive change as visualized through theuse of monoclonal antibodies targeted to neurofilament
injury 221
subunits. Journal of Neuropathology and ExperimentalNeurology 51, 158–76.
.




















