
Vaccines against bluetongue in Europe
20
Comparative Immunology, Microbiology & Infectious Diseases 31 (2008) 101–120 Vaccines against bluetongue in Europe Giovanni Savini a , N. James MacLachlan b , Jose-Manuel Sanchez-Vizcaino c , Ste´phan Zientara d, a OIE Reference Laboratory for Bluetongue, Istituto Zooprofilattico Sperimentale dell’Abruzzo e del Molise ‘‘G. Caporale’’, Teramo, Italy b Department of Pathology, Microbiology and Immunology, School of Veterinary Medicine, University of California, Davis, CA 95616, USA c Catedra´tico de Sanidad Animal, Dpto. Sanidad Animal, Universidad Complutense, Madrid, Spain d AFSSA Alfort, UMR 1161, 23 Av du Ge´ne´ral de Gaulle, BP 63, 94703 Maisons Alfort, Ce´dex, France Accepted 12 July 2007 Abstract After the incursion of bluetongue virus (BTV) into European Mediterranean countries in 1998, vaccination was used in an effort to minimize direct economic losses to animal production, reduce virus circulation and allow safe movements of animals from endemic areas. Vaccination strategies in different countries were developed according to their individual policies, the geographic distribution of the incurring serotypes of BTV and the availability of appropriate vaccines. Four monovalent modified live virus (MLV) vaccines were imported from South Africa and subsequently used extensively in both cattle and sheep. MLVs were found to be immunogenic and capable of generating strong protective immunity in vaccinated ruminants. Adverse side effects were principally evident in sheep. Specifically, some vaccinated sheep developed signs of clinical bluetongue with fever, facial oedema and lameness. Lactating sheep that developed fever also had reduced milk production. More severe clinical signs occurred in large numbers of sheep that were vaccinated with vaccine combinations containing the BTV-16 MLV, and the use of the monovalent BTV-16 MLV was discontinued as a consequence. Abortion occurred in o0.5% of vaccinated animals. The length of viraemia in sheep and cattle that received MLVs did not exceed 35 days, with the single notable exception ARTICLE IN PRESS www.elsevier.com/locate/cimid 0147-9571/$ - see front matter r 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.cimid.2007.07.006 Corresponding author. Tel.: +33 1 43 96 72 80. E-mail address: [email protected] (S. Zientara).
Transcript of Vaccines against bluetongue in Europe

ARTICLE IN PRESS
Comparative Immunology, Microbiology
& Infectious Diseases 31 (2008) 101–120
0147-9571/$ -
doi:10.1016/j
�CorrespoE-mail ad
www.elsevier.com/locate/cimid
Vaccines against bluetongue in Europe
Giovanni Savinia, N. James MacLachlanb,Jose-Manuel Sanchez-Vizcainoc, Stephan Zientarad,�
aOIE Reference Laboratory for Bluetongue, Istituto Zooprofilattico Sperimentale dell’Abruzzo e del Molise
‘‘G. Caporale’’, Teramo, ItalybDepartment of Pathology, Microbiology and Immunology, School of Veterinary Medicine,
University of California, Davis, CA 95616, USAcCatedratico de Sanidad Animal, Dpto. Sanidad Animal, Universidad Complutense, Madrid, Spain
dAFSSA Alfort, UMR 1161, 23 Av du General de Gaulle, BP 63, 94703 Maisons Alfort, Cedex, France
Accepted 12 July 2007
Abstract
After the incursion of bluetongue virus (BTV) into European Mediterranean countries in
1998, vaccination was used in an effort to minimize direct economic losses to animal
production, reduce virus circulation and allow safe movements of animals from endemic areas.
Vaccination strategies in different countries were developed according to their individual
policies, the geographic distribution of the incurring serotypes of BTV and the availability of
appropriate vaccines. Four monovalent modified live virus (MLV) vaccines were imported
from South Africa and subsequently used extensively in both cattle and sheep. MLVs were
found to be immunogenic and capable of generating strong protective immunity in vaccinated
ruminants. Adverse side effects were principally evident in sheep. Specifically, some vaccinated
sheep developed signs of clinical bluetongue with fever, facial oedema and lameness. Lactating
sheep that developed fever also had reduced milk production. More severe clinical signs
occurred in large numbers of sheep that were vaccinated with vaccine combinations containing
the BTV-16 MLV, and the use of the monovalent BTV-16 MLV was discontinued as a
consequence. Abortion occurred in o0.5% of vaccinated animals. The length of viraemia in
sheep and cattle that received MLVs did not exceed 35 days, with the single notable exception
see front matter r 2007 Elsevier Ltd. All rights reserved.
.cimid.2007.07.006
nding author. Tel.: +331 43 96 72 80.
dress: [email protected] (S. Zientara).

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120102
of a cow vaccinated with a multivalent BTV-2, -4, -9 and -16 vaccine in which viraemia
persisted at least 78 days. Viraemia of sufficient titre to infect Culicoides insects was observed
transiently in MLV-vaccinated ruminants, and natural transmission of MLV strains has been
confirmed. An inactivated vaccine was first developed against BTV-2 and used in the field. An
inactivated vaccine against BTV-4 as well as a bivalent vaccine against serotypes 2 and 4 were
subsequently developed and used in Corsica, Spain, Portugal and Italy. These inactivated
vaccines were generally safe although on few occasions reactions occurred at the site of
inoculation. Two doses of these BTV inactivated vaccines provided complete, long-lasting
immunity against both clinical signs and viraemia, whereas a single immunization with the
BTV-4 inactivated vaccine gave only partial reduction of viraemia in vaccinated cattle when
challenged with the homologous BTV serotype. Additional BTV inactivated vaccines are
currently under development, as well as new generation vaccines including recombinant
vaccines.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Arbovirus; Orbivirus; Bluetongue; Vaccines
Resume
Apres l’incursion du virus de la fievre catarrhale ovine (FCO ou Bluetongue, BT) dans le
bassin mediterraneen en 1998, les autorites italienne, franc-aise, portugaise et espagnole ont
utilise la vaccination pour reduire les pertes economiques directes dans le domaine des
productions animales et pour reduire la circulation virale. Les strategies de vaccination suivies
dans les differents pays ont ete effectuees selon leurs propres politiques vaccinales, les
serotypes du virus de la FCO introduits et leurs distributions geographiques et les capacites
d’acces aux vaccins. Quatre differents vaccins attenues monovalents ont ete importes
d’Afrique du sud. Plusieurs associations de ces vaccins entre eux ont ete utilisees a large echelle
chez les bovins et les moutons. Les etudes experimentales et les donnees de terrain ont montre
que ces vaccins attenues sont immunogenes et capables d’induire une immunite solide chez les
ruminants vaccines. Des effets secondaires ont toutefois ete mis en evidence chez les ovins. En
particulier, des moutons ont presente de la fievre, des oedemes de la face et des boiteries qui
apparaissaient en general dans la seconde semaine suivant la vaccination et disparaissaient 7 a
10 jours apres. Les brebis qui ont manifeste des syndromes febriles ont presente des chutes de
production laitiere. Des avortements ont ete observes chez moins de 0,5% des animaux
vaccines. Des signes cliniques plus severes ont ete observes chez les moutons vaccines avec des
combinaisons de vaccins qui contenaient le serotype 16; en consequence, l’utilisation du vaccin
monovalent 16 a ete arretee. La duree de la viremie chez les moutons et les bovins qui avaient
rec-u un vaccin vivant n’a pas depasse 35 jours, a l’exception notable d’un bovin vaccine avec
un vaccin polyvalent 2, 4, 9 et 16 chez lequel la viremie depassait au moins 78 jours. Le titre de
ces viremies transitoires etait suffisant pour infecter des vecteurs Culicoides et des
transmissions naturelles des souches vaccinales ont ete rapportees sur le terrain. Un vaccin
inactive contre le serotype 2 puis un vaccin inactive contre le serotype 4 ainsi qu’un vaccin
bivalent 2 et 4 ont ete developpes et utilises en Corse, en Espagne, au Portugal et en Italie. Ces
vaccins inactives ont une innocuite satisfaisante meme si parfois des reactions locales aux sites
d’inoculation peuvent s’observer chez les ruminants vaccines. Deux doses de vaccin inactive
induisent une immunite de longue duree et protegent les ruminants a la fois contre les signes
cliniques et la viremie alors qu’une seule dose avec le vaccin de serotype 4 n’induit qu’une
reduction partielle de la viremie chez les bovins infectes apres vaccination. De nouveaux

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120 103
vaccins inactives sont en cours de developpement ainsi que des vaccins de nouvelle generation
dont des vaccins recombinants.
r 2007 Elsevier Ltd. All rights reserved.
Mots cles: Arbovirus; Orbivirus; Fievre catarrhale ovine; Vaccins
1. Introduction
Bluetongue (BT) is a non-contagious, insect-transmitted disease of certain breedsof sheep and some species of wild ruminants that is caused by bluetongue virus(BTV) [1]. BTV infection of ruminants occurs throughout much of the temperate andtropical regions of the world, coincident with the distribution of specific species ofCulicoides biting midges that are biological vectors of the virus [2,3]. BT typicallyoccurs when susceptible sheep are introduced into areas where virulent strains ofBTV circulate, or when virulent strains of BTV extend their range into previouslyunexposed populations of ruminants. The global distribution of BTV has historicallybeen between latitudes of approximately 40–501N and 351S. However, during therecent northern European epidemic, the virus spread far beyond its prior knownupper northern limits [4].
BTV is the prototype member of the genus Orbivirus, family Reoviridae [5]. Allreoviruses share distinctive common properties including segmented genomes ofdouble-stranded RNA (dsRNA) and characteristic virion morphology and structure.There are currently 12 genera within the family Reoviridae, which include pathogensof plants, crustaceans, fish, insects, reptiles and mammals including humans. Todate, 24 distinct serotypes of BTV have been described that all share common groupantigens but which are distinguished on the basis of serotype-specific virusneutralization assays (VNTs). Importantly, there is considerable variation amongstfield strains of BTV, even those of the same serotype, which reflects differences in thenucleotide sequence of each of the 10 distinct dsRNA segments of the BTV genome[6,7]. Genetic heterogeneity of field strains of BTV occurs as a consequence both ofgenetic drift and genetic shift, the latter as a result of reassortment of viral genesduring mixed infections of either the vertebrate or invertebrate hosts of the virus [8].Variation in the sequence of individual genes occurs through a complex process ofgenetic drift and founder effect during alternating passage of BTV in its ruminantand insect hosts [9].
Five different BTV serotypes (1, 2, 4, 9 and 16) have recently spread throughoutextensive portions of Mediterranean Europe, and BTV serotype 8 emerged innorthern Europe in 2006 [4]. BT epidemics had been reported, although irregularly,from 1979 to 1999 in Greece, in the Eastern Islands (Lesbos, Leros and Kos) and theDodecanese archipelago. In 1999, BT was reported in mainland Greece andsoutheastern Bulgaria, close to the Turkish and Greek borders. From August toSeptember 2000, BTV-9 spread progressively through the Balkan region to Albania,Bosnia Herzegovina, Croatia, Kosovo, Macedonia Republic, Republic of Serbia andMontenegro [10]. BTV-2 emerged in 2000 in France (Corsica), Italy (Sardinia) and

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120104
Spain (Balearic Islands) [11–13] and BTV-9 appeared later during the same year inCalabria (Italy). In 2001–02, BTV-2 and -9 spread throughout Sicily and southernand central Italy. In 2002, serotypes 4 and 16 were detected in the southern regions ofItaly, and in 2003 serotype 4 emerged in Sardinia, Corsica and Menorca; in 2003,serotype 16 was reported in Sardinia and then in 2004 in Corsica [14]. In the sameyear, a new epizootic of BTV-4 affected the mainland of Spain and neighboringregions of Portugal. In 2005, the infection spread to central Spain but no furtheroutbreaks were reported in 2006 [15], in contrast with Portugal where the epidemiccontinued through 2006. In the same year, BTV serotype 8 emerged veryunexpectedly in the North of Europe involving Belgium, France, Germany,Luxembourg and the Netherlands [4].
The incursion of BTV into Mediterranean Europe is having a considerablenegative economic impact, partly due to direct losses from mortality and reducedproduction in affected livestock but, more importantly, because of the total ban ofruminant trade between BTV-infected and non-infected areas [10]. To limit directlosses and in an effort to minimize the circulation of BTV, as well as to allow the safemovement of animals, the Italian, French, Portuguese and Spanish authorities allundertook vaccination of livestock according to their individual national policies,the geographic distribution of the incurring BTV serotype(s), and the availability ofappropriate vaccines. In France, only sheep were vaccinated whereas in Italy, allsusceptible domestic ruminant species were vaccinated, i.e. sheep, goats, cattle andwater buffalos [16]. In Spain, initially only sheep were vaccinated (2001–2003 inBalearic Islands) but in later outbreaks (southwest Spain) both cattle and sheep weresubjected to vaccination [17]. The Italian vaccination campaign was based on a riskassessment that demonstrated that such a vaccination strategy would prevent directeconomic losses, significantly reduce virus circulation [18] and minimize risks linkedto the movement of animals from infected to free areas.
This paper will describe the different vaccines against BT that have been used orwhich have recently been developed in Europe. These include inactivated whole[killed] virus preparations, virus-like particles (VLPs) produced from recombinantbaculoviruses, live attenuated vaccines (modified live viruses, MLVs) and liverecombinant vaccinia or canarypox virus-vectored vaccines [19,20]. All have inherentpotential advantages and disadvantages, but only MLVs and some inactivatedvaccines are currently available under European Community approved nationaldisease control programs. VLPs are also safe and have been shown to be efficaciousin laboratory trials [23–26], but their efficacy in the field is still under evaluation.
2. Inactivated vaccines
The first inactivated vaccine that was developed and used in the field after theemergence of BT in Europe was the vaccine against BTV-2. Subsequently, amonovalent BTV-4 and a bivalent BTV-2 and -4 vaccines have been developed andused in Corsica, Spain, Portugal and Italy. Other inactivated vaccines have recentlybeen developed or are under development (Table 1).

ARTIC
LEIN
PRES
S
Table 1
Bluetongue vaccination campaigns in European countries
Vaccine Year of vaccination
France (Corsica) Italya Spain Portugal Bulgaria
Sheep Sheep Cattle Sheep Cattle Sheep Cattle Sheep
MLV 1999–2000
BTV-3, -8, -9, -10, -11
BTV-2 2000–02b 2002–06 2002–06 2000–01b (Balearic Island)
BTV-4 2004–06 2005–06
BTV-2&4 2003–04 2004–06 2004–06 2003 (Balearic Island)
BTV-2&9 2002–06 2002–06
BTV-16 2004c
BTV-2, -4, -16 2004c 2004c
BTV-2, -4, -9, -16 2004c 2004c
BTV-2, -4, -9 2005–06 2005–06
BTV-9
Inactivated 2005
BTV-2
BTV-4 2005–06 2005–06 2005–06 2005–06
BTV-2&4 2006 2005–06
aIn Italy various monovalent MLV combinations have been used in different regions.bThe 2000 vaccination campaigns started in November.cThe use of the BTV-16 MLV was discontinued.
G.
Sa
vini
eta
l./
Co
mp
.Im
mu
n.
Micro
bio
l.In
fect.D
is.3
1(
20
08
)1
01
–1
20
105

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120106
Inactivated whole virus vaccines are very safe if properly produced. They can behighly efficacious [21,22] and although not yet available, strategies for differentiatinginfected from vaccinated animals (DIVA) are theoretically possible with these typeof vaccines. Their inherent potential disadvantages include: (1) their high costs ofproduction, as vaccination requires large amounts of antigen; and (2) the need forbooster immunizations, as inactivated vaccines generally induce a relatively transientimmunity.
2.1. Quality control
For quality control testing, no data were given regarding the control testsperformed during the different stages of production of the inactivated vaccine;however, all companies must follow the current guidelines described in the EUlegislation concerning the manufacture of veterinary vaccines.
2.2. Safety
Several studies have been conducted on sheep to evaluate the safety of thesubcutaneous injection of inactivated prototype vaccines against BTV-2, BTV-4 andBTV-2&4 in either simple, repeated or overdose trials. In all conditions, theinactivated BTV prototype vaccines were very well tolerated as demonstrated by theabsence of systemic reaction (fever, weight loss, reproductive dysfunction, etc.)related to vaccination. Some vaccines induced transient local reactions of variableseverity (mild to moderate) with different frequency (unusual to common). Theseusually disappeared within 3 days but, in a single case, a moderate local reactionpersisted for 2 weeks [27]. Anaphylactic shock was also reported in 0.02% sheepfollowing vaccination with a BTV-4 inactivated vaccine. This event was observedonly in areas where BTV-4 MLV had previously been used.
BTV-4 and BTV-2&4 inactivated vaccines have also been tested for safety in cattle[17,28,29]. The vaccines were very well tolerated, and no side effects or localreactions were observed even when five doses of the BTV-4 inactivated vaccine wereadministered to the same animal [17].
No data documenting systemic or local reactions in vaccinated animals areavailable from the field use of these vaccines, as no complaints were reported fromfarmers. In addition, as no significant antigenic variation was yet documentedamongst the BTV-2 and -4 strains currently circulating in Europe, the strains used toproduce the inactivated vaccine are still suitable for the production of effectivevaccines against these serotypes.
2.3. Efficacy
Most companies producing BTV inactivated vaccines follow the guidelines of theEuropean Pharmacopea and Committee for Veterinary Medicines Products forquality and safety control and efficacy. Assessment of efficacy is based on clinicaland virological data as well as on immunogenicity. Immunogenicity is assessed by

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120 107
the analysis of the antibody response induced by each immunization, as measured byELISA and by titration in a VNT against the same serotype (BTV-2 or BTV-4). Theefficacy of the vaccine is evaluated in vaccinated animals by inoculation of aninfective dose of live virulent BTV. The level of viremia after virus challenge isconsidered the most objective way to assess the efficacy of the vaccine-indu-ced immunity. The level of viremia is analyzed by either a BTV-specific quantita-tive real-time RT-PCR assay [30] or by virus isolation. In addition, clinical signs(fever, general congestion of the skin, edema and lameness) are evaluated afterchallenge.
The inactivated prototype BTV vaccines induced significant titres of neutralisingantibodies after either one or two injections in sheep. A booster effect was observedafter the second immunization [27,31]. In cattle, one dose of BTV-4 or BTV-2&4inactivated vaccines induced a weak humoral response which rapidly declined to beundetectable 21 days following vaccination. However, the second dose of vaccineelicited high and stable titers of neutralizing antibodies [17,28].
Table 2 shows the inactivated vaccines that have been tested to date for efficacy.The number, age and breed of animals, as well as the challenge dose and virus strainvary for the different vaccines. A preliminary study was carried out to establish andstandardize the optimal conditions for BTV vaccine trials on sheep and cattle. In thisstudy, the virus strain chosen for the challenge experiment was the isolate BTV-4/SPA-1/2004. The effect of factors like the number of cell culture passages of the virusinoculum, the virus dose, type of inoculum and route of inoculation have all beenevaluated. The infectivity of the field isolate was confirmed after four passages in cellculture, and the route of inoculation (subcutaneous or intravenous) did not affecteither the occurrence of clinical signs or the duration and titer of viremia.Appropriate titers of viremia were also obtained with different inocula (infectedblood or cell culture-propagated virus) at different doses (2� 107 and2� 106 TCID50 (50% tissue culture infective dose)/dose or threshold cycle (TC):25–27 and TC: 30–33). The inoculation of cell culture-propagated virus-induceddetectable viremia 3 days earlier (day 4) than that of blood from highly viremicsheep.
One or two doses of inactivated BTV-2, BTV-4 and BTV-2&4 vaccine at 3–4 weekintervals gave full and significant protection against clinical signs and viremia insheep that were intradermally challenged with virulent BTV-2 and/or BTV-4 a weekor a month after last vaccination. For BTV-2, it was also shown that a single doseprotected sheep against both clinical signs and viremia for at least 6 months [27,31].For BTV-2&4, two doses fully protect sheep for up to 12 months (Table 2).
In cattle, efficacy studies have been performed on BTV-4 and BTV-2&4 vaccines.Two doses of the inactivated BTV-4 vaccine administered at a 24-day intervalprevented viremia in vaccinated animals challenged with the homologous virulentserotype. Similarly, none of the animals vaccinated with two doses of BTV-2&4inactivated vaccine developed detectable viremia following challenge with virulentfield strains of BTV-2 and/or BTV-4 that were performed up to 1 month afterthe second vaccination [28]. However, although a single dose of BTV-4 inacti-vated vaccine prevented viremia in vaccinated animals challenged 2 weeks after
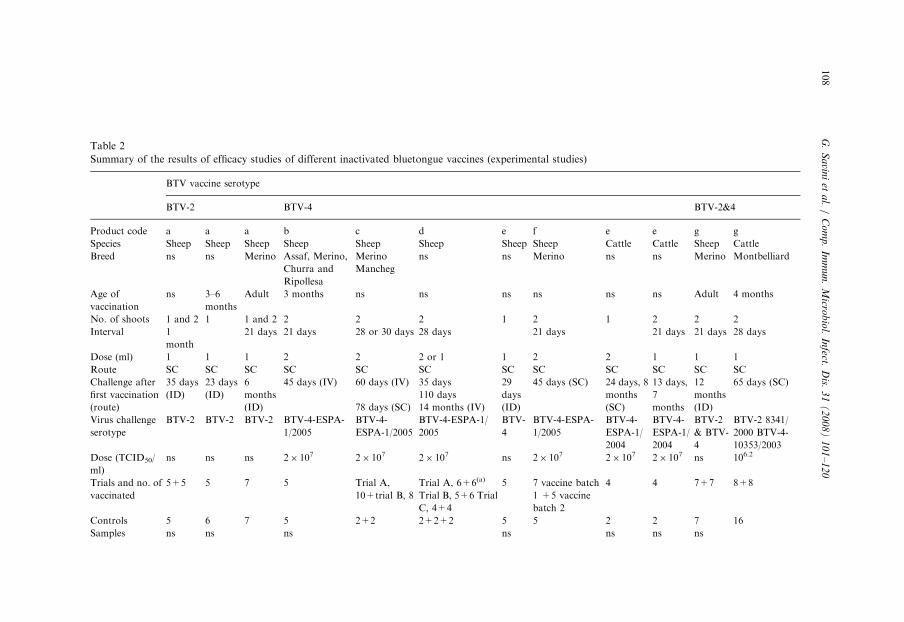
ARTIC
LEIN
PRES
S
Table 2
Summary of the results of efficacy studies of different inactivated bluetongue vaccines (experimental studies)
BTV vaccine serotype
BTV-2 BTV-4 BTV-2&4
Product code a a a b c d e f e e g g
Species Sheep Sheep Sheep Sheep Sheep Sheep Sheep Sheep Cattle Cattle Sheep Cattle
Breed ns ns Merino Assaf, Merino,
Churra and
Ripollesa
Merino
Mancheg
ns ns Merino ns ns Merino Montbelliard
Age of
vaccination
ns 3–6
months
Adult 3 months ns ns ns ns ns ns Adult 4 months
No. of shoots 1 and 2 1 1 and 2 2 2 2 1 2 1 2 2 2
Interval 1
month
21 days 21 days 28 or 30 days 28 days 21 days 21 days 21 days 28 days
Dose (ml) 1 1 1 2 2 2 or 1 1 2 2 1 1 1
Route SC SC SC SC SC SC SC SC SC SC SC SC
Challenge after
first vaccination
(route)
35 days
(ID)
23 days
(ID)
6
months
(ID)
45 days (IV) 60 days (IV) 35 days 29
days
(ID)
45 days (SC) 24 days, 8
months
(SC)
13 days,
7
months
12
months
(ID)
65 days (SC)
110 days
78 days (SC) 14 months (IV)
Virus challenge
serotype
BTV-2 BTV-2 BTV-2 BTV-4-ESPA-
1/2005
BTV-4-
ESPA-1/2005
BTV-4-ESPA-1/
2005
BTV-
4
BTV-4-ESPA-
1/2005
BTV-4-
ESPA-1/
2004
BTV-4-
ESPA-1/
2004
BTV-2
& BTV-
4
BTV-2 8341/
2000 BTV-4-
10353/2003
Dose (TCID50/
ml)
ns ns ns 2� 107 2� 107 2� 107 ns 2� 107 2� 107 2� 107 ns 106.2
Trials and no. of
vaccinated
5+5 5 7 5 Trial A,
10+trial B, 8
Trial A, 6+6(a)
Trial B, 5+6 Trial
C, 4+4
5 7 vaccine batch
1 +5 vaccine
batch 2
4 4 7+7 8+8
Controls 5 6 7 5 2+2 2+2+2 5 5 2 2 7 16
Samples ns ns ns ns ns ns ns
G.
Sa
vini
eta
l./
Co
mp
.Im
mu
n.
Micro
bio
l.In
fect.D
is.3
1(
20
08
)1
01
–1
20
108

ARTIC
LEIN
PRES
S
5, 7, 9,
12,
14 dpc
3, 5, 7, 10, 18,
21, 24 and
27 dpc
3, 5, 7, 10, 18, 21,
24 and 27 dpc
3, 5, 7, 10, 18,
21, 24 and
27 dpc
Three times a
week for 6
weeks
Viremia results
in vaccinated
group
0/5 0/5 0/7 ns 0/10 A: 0/6+6 ns Batch 1: 0/7 0/4 0/4 0/7 0/8 0/8
0/5 B: 0/5+6 Batch 2: 0/5 4/4 0/4 0/7
C: 4/4+4
Viremia results
in control group
5/5 6/6 7/7 ns 2/2 A: 2/2 ns 5/5 2/2 2/2 6/7 BTV-2: 8/8
B:2/2 BTV-4: 8/8
C: 2/2
Method used Real
time
RT-
PCR
Real
time
RT-
PCR
Real
time
RT-
PCR
ns Real time
RT-PCR
Real time RT-
PCR
ns Real time RT-
PCR
Real time
RT-PCR
Real
time
RT-
PCR
Real
time
RT-
PCR
Virus isolation
Trial C: two or four animals vaccinated with half-dose had slight viremia when challenged after 14 months post-vaccination.
ID, intradermally; SC, subcutaneously; IV, intravenously; ns: data not provided.
Hamers et al. [27,31]; Savini et al. [28]; MAPA [17].
Real time RT-PCR: Jimenez-Clavero et al. [30]; Toussaint et al. [4].
G.
Sa
vini
eta
l./
Co
mp
.Im
mu
n.
Micro
bio
l.In
fect.D
is.3
1(
20
08
)1
01
–1
20
109

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120110
vaccination, a single vaccination did not fully prevent viremia in animals challenged7 months after vaccination [17].
The efficacy of the inactivated BTV vaccine was indirectly confirmed in the fieldwhen all but 2 of more than 40,000 seasonally migrating vaccinated Spanish cattleremained negative for BTV by RT-PCR after staying in a restricted area in thepresence of BTV circulation [30].
3. Modified live virus (MLV) vaccines
Outside of the European Union (EU), including the USA, Turkey, Republic ofSouth Africa and India, BTV MLVs are available for many BTV serotypes. MLVsare produced by adapting BTV field isolates to growth in vitro through serialpassages in tissue culture or in embryonated chicken eggs. Stimulation of a strongantibody response by these vaccines directly is correlated with their ability toreplicate in the vaccinated host. MLVs are cheap to produce in large quantities, theygenerate protective immunity after a single inoculation and have proven effective inpreventing clinical BT disease in the areas where they are used [16,32,33]. However,BTV MLVs suffer from a variety of documented or potential drawbacks includingunder-attenuation, whose impact may vary with sheep of different breeds. Potentialadverse consequences are depressed milk production in lactating sheep, andabortion/embryonic death and teratogenesis in offspring when used on pregnantfemales [34–39,44]. Another risk associated with the use of MLVs is that of theirpotential for spread by vectors, with eventual reversion to virulence and/orreassortment of MLV genes with those of wild-type virus strains. The frequencyand significance of these events remain poorly defined but natural and localdissemination of BTV-2 and -16 MLV vaccine strains has already been documentedin Europe. Natural dissemination of MLV strains of BTV likely also is responsiblefor the sporadic incidence of teratogenic defects in unvaccinated cattle in SouthAfrica and North America. Finally, the intrinsic inability to serologically distinguishnaturally infected from MLV vaccinated animals precludes the possibility ofdeveloping a DIVA strategy with the MLV vaccines.
After the incursion of BTV into Mediterranean Europe, the Spanish, French,Italian and Portuguese authorities have all carried out compulsory vaccinationcampaigns since 2000 using MLVs produced by Onderstepoort Biological Productsin an attempt to reduce direct losses due to disease and indirect losses due to tradeembargoes caused by the presence of BTV (Table 1). At that time, these were theonly commercially available BTV vaccines. Based on the serotype(s) present in agiven country/area, various MLV monovalent serotype formulations have beenused.
3.1. Quality control
Four monovalent MLV vaccines have been imported from South Africa and usedin the EU (Table 3). Before use, these vaccines were confirmed by the various EU

ARTICLE IN PRESS
Table 3
Characteristics of the bluetongue virus vaccine strains
Modified
live vaccine
Strain Date, place of
isolation
History
BTV-2 Vryheid/
5036
1958, Republic of
South Africa
Virus passaged 50 times in embryonated chicken eggs and
plaque selected three times and passaged twice in BHK21 cells.
BTV-4 Theiler/
79043
1900, Republic of
South Africa
Virus passaged 60 times in embryonated chicken eggs,
followed by three small plaque selections and nine passages in
BHK21 cells.
BTV-9 1900, Republic of
South Africa
Virus passaged 70 times in embryonated chicken eggs,
followed by three small plaque selections and six further
passages in BHK21 cells.
BTV-16 Pakistan/
7766
Pakistan Virus passaged 37 times in eggs, followed by three large plaque
selections, two passages in BHK21 cells and one in VERO
cells.
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120 111
National Laboratories to be free of bacterial, fungal and viral contaminants. Titresand serotype of the MLV batches also have been verified with no major discrepancieswith information provided by the manufacturer. For the BTV-2 MLV, it wasobserved that the titer strongly decreased at temperatures above 35 1C [41], butthat it was retained for at least 36–48 h if sterility conditions were ensured duringrehydration and the reconstituted vaccine was stored at a temperature below19 1C [42].
3.2. Safety
Modified live viruses have different potential adverse impacts according to thespecific formulation used, the specific serotypes and the number of serotypesincluded in the vaccine.
3.2.1. Fever and sickness
Apart from a study performed by the Institute of Animal Health at Pirbright usingeither BTV-2 or BTV-9 and Polled Dorset sheep, in which moderate to severe clinicalsigns of BT, albeit short lived, were observed following vaccination [40], only mildsymptoms were observed in most experimental MLV vaccination studies. These werecharacterized by transient fever starting from the fifth day after vaccination (p.v.)and mild hyperemia of the oral cavity during the second week.
Reports of adverse events in the field greatly vary with the strain of BT MLV usedfor vaccination of the animals. The monovalent BTV-2 MLV vaccine was used inCorsica (from 2001 to 2004) and Italy (from 2002) on approximately 130,000 and4,000,000 sheep and goats, respectively. In both locations, there were no or negligibleadverse reactions reported after vaccination [14,43–45]. However, when the samevaccine was used in 2000–01 in Menorca and Mallorca on 320,000 sheep, adverseevents were observed in 0.13% and abortion in 0.16% of the vaccinated animals [17].

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120112
The BTV-2 MLV vaccine was also used on more than 400,000 cattle during the 2002Italian vaccination campaign with no adverse reactions reported.
The monovalent BTV-4 MLV vaccine has been only used in sheep in Corsica in2004. No adverse reactions were reported [14].
In 2003 a new outbreak of BT occurred in Menorca that was due to BTV-4. Acombined BTV-2 and -4 MLV vaccine was used to vaccinate sheep and goats, and noside effects were observed. The same vaccine combination was used in 2004 inTuscany, and the following year in Tuscany and Sardinia, involving some 4,000,000sheep and goats. Again, no or negligible adverse reactions were reported. Similarly,no adverse side effects were recorded among approximately 400,000 cattle vaccinatedwith the combined BTV-2 and -4 MLVs in 2004 and among a similar number ofanimals in 2005.
A bivalent BTV-2 and -9 MLV vaccine was used to vaccinate sheep and goats insome regions of Italy since 2002. Of the more than 1,700,000 animals vaccinated,only a very small percentage (o0.1%) developed fever and facial edema at 7–14 daysp.v. [45]. In the same vaccination campaign, BTV-2/BTV-9 MLVs were alsoadministered to more than 600,000 cattle and no adverse reactions were reported.Similarly, a trivalent MLV vaccine containing BTV serotypes 2, 4 and 9 was usedsince 2005 on more than 1,000,000 sheep, goat and cattle, and no or negligibleadverse reaction were reported.
In 2004, BTV-16 was isolated in Corsica by the AFSSA Maisons-Alfortlaboratory [46]. Vaccination of sheep with BTV-16 MLV was performed in thesouthern part of the island. Typical signs of BT were reported in vaccinated sheepseveral days after vaccination. The vaccination program was therefore immediatelyterminated. The nucleotide sequence of segments 2, 7, 8, 9 and 10 from the virusisolated from ill animals was determined and found to be exactly identical to that ofthe vaccine MLV strain (Zientara, unpublished).
Also in 2004, a trivalent MLV vaccine containing serotypes 2, 4 and 16 was used inSardinia. However, a few weeks after vaccination, many vaccinated andunvaccinated sheep and goats became ill because of infection with the BTV-16vaccine strain [47]. These incidents were attributed to inadequate attenuation of theBTV-16 MLV that was in the vaccine, and for this reason the use of monovalentBTV-16 MLV vaccine was discontinued [48]. In contrast to what occurred in sheepand goats; however, the BTV-2, -4 and -16 MLV combination did not causesignificant adverse reactions in cattle (approximately 600,000 vaccinated animals).
A polyvalent MLV containing BTV serotypes 2, 4, 9 and 16 was used in southernregions of Italy in 2004, and approximately 1,700,000 sheep and goats werevaccinated. No adverse reactions were reported. The same BTV-2, -4, -9 and -16MLV combination did not cause adverse reactions in cattle (approximately 600,000vaccinated animals).
Finally, the safety of a pentavalent BTV serotypes 1, 2, 4, 9 and 16 MLV vaccinewas evaluated in sheep maintained in isolation facilities at the Afssa SophiaAntipolis laboratory. Significant fever (41–42 1C) was reported in many animals(unpublished data). Based on these observations, it was decided that this vaccineshould not be used in the field.

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120 113
3.2.2. Effect on pregnancy
Experimental infection studies using MLV strains of BTV-2 or BTV-2 and -9were conducted on cattle to evaluate potentially deleterious effects on reproduc-tion. In none of these studies was any adverse effect on pregnancy observed[38,49,50].
Abortions and/or stillbirth have however been reported in the various vaccina-tion campaigns that used BTV MLVs. During the 2000–01 Balearic BTV-2campaign, approximately 0.16% of the 320,000 vaccinated sheep aborted. Similarly,during the Italian BTV-2 vaccination campaign, abortion was reported on 0.42%and 0.18% of vaccinated sheep and cattle, respectively. However the virus wasdetected in only 0.06% and 0.01% of their respective aborted fetuses. In 2002, thecombined BTV-2 and -9 MLV resulted in abortion of 0.53% and 0.14% ofvaccinated sheep and cattle, respectively, although BTV was detected only in a smallpercentage of abortions (0.09% and 0.01% of the sheep and cattle fetuses,respectively) [49].
3.2.3. Effect on semen
The effects of the BTV-2 MLV on the quality of semen were investigated [51] in 23rams vaccinated at a 47-day interval. Although BTV was not detected in any of thesemen samples, a decrease of the semen quality (volume, sperm concentration,motility, abnormal and dead spermatozoa) was demonstrated after the firstvaccination. A decrease in semen quality was also observed after the secondvaccination; however, at day 69, the semen quality of the vaccinated animals was notsignificantly different from those of the 23 controls.
3.2.4. Milk production
Given the economic importance of milk production, numerous studies have beenconducted to determine the effects of several MLV combinations on milk productionin both sheep and cattle.
Vaccination of sheep with either BTV-2 or BTV-2 &4 MLVs did not affect thequantity and quality of milk produced by the vaccinated animals (Zientara,unpublished; and [52–54]).
Quite different were the observations made with BTV-2 and -9, BTV-2 and -16,BTV-2, -4 and -9 or BTV-2, -4, -9 and -16 MLVs. Vaccination with each of thesecombinations had a marked negative impact on total milk production withproduction decreases of 20–30% as compared to normal production levels. Thedecrease, evident in the second week following vaccination, was transient and notaccompanied by significant changes in milk quality (cell count, pH, fat, protein andlactose) [36,53]. It was suggested that the effects of vaccination on milk productionwere primarily due to the transient perturbation of health induced by the vaccine andnot to a direct virus effect on the mammary tissue [36].
In contrast, vaccination of 30 cows with combined BTV-2 and -9 MLV vaccineshad no effect on the production and quality (somatic cell count, pH, milk fat, proteinand lactose content) of their milk [38].

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120114
In the field, data collected between 1999 and 2002 on the quantity and quality ofmilk of 18,000 cows demonstrated that BTV-2 MLV vaccination had no effect onmilk production [54].
3.2.5. Duration and titer of viremia
After immunization with MLVs, the attenuated virus circulates in the blood streamand so potentially can infect competent vectors and be transmitted to other susceptiblehosts. Therefore, MLV vaccination should be performed in the cooler months whenthe Culicoides population and its activity typically are at the lowest level. This will limitthe possibility of transmission of the vaccine strains by biting midges whileimmunizing susceptible animal populations before the next epidemic season.
Transmission of MLV strains of BTV to insects most likely would occur fromviremic animals that are introduced into infection-free areas where competentCulicoides species are present and highly active. In this scenario, the magnitude andduration of viremia in vaccinated animals would be clearly important in determiningwhether or not MLV strains of BTV could be acquired and transmitted by localvectors. Although virus titers in blood less than 103 TCID50/ml have traditionallybeen considered a ‘‘safe’’ threshold, authentic instances of insects acquiring BTVfrom animals with viremic titers less than 103TCID50/ml have been reported. Giventhe complex interaction of BTV, Culicoides vectors and animal hosts in the life cycleof infection, virus titers induced by MLV should be kept to an absolute minimumspecially if field transmission of MLV strains is a concern.
Studies on the duration and titers of viremia have been performed on sheep andcattle following vaccination with different MLV combinations. Viremia followingvaccination with BTV-2, -4, -9 and -16 MLV strains (including multivalentcombinations) was found to persist for up to 24 days in sheep and 78 days incattle [37,38,47,55–59]. Information pertaining to the MLV strains used in Europe ishowever limited; available data suggests that cattle vaccinated with BTV-2 and BTV-9 MLVs can be moved safely 32 days after vaccination [37], whereas sheepvaccinated with the same strains can be moved 28 days following immunization [36].From the viremia data obtained in cattle following BTV-2, -4, -9 and -16 MLVvaccination, it was determined that cattle could be moved safely (risk of infectiono0.01%) at 60 days after vaccination [29]. The latter result, however, is most likelyrelated to the inadequate attenuation of the BTV-16 MLV strain and cannot beextrapolated to MLV vaccines that do not include this serotype.
Apart from some BTV-2 MLV vaccination studies on sheep and cows where virustiters were never found to be higher than 103TCID50/ml [41,49], all other MLVcombinations which have been studied in sheep (BTV-2, BTV-9, BTV-16, BTV-2 and-9, BTV-2 and -4, BTV-2, -4 and -16, BTV-2, -4, -9 and -16) and cattle (BTV-2 and-9, BTV-2, -4, -9 and -16) gave rise, for a brief period of 2–4 days, to viremic titersabove the infecting threshold at least in some of the vaccinated animals[28,29,36,37,40,47,59].
No data have been reported, however, on the duration and titers of viremia inanimals vaccinated with these MLV in the field, but local transmission of BTV-2 andBTV-16 vaccine strains in the field has been demonstrated [39,47].

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120 115
3.3. Efficacy
An important factor in confirming the efficacy of MLV vaccines is their ability toelicit neutralizing antibodies in vaccinated animals. Neutralizing antibodies play akey role in protecting animals from disease and viremia following infection with thehomologous wild-type BTV. Knowing the duration of the immune status derivedfrom vaccination is of paramount importance for both planning the frequency ofvaccine booster immunizations to adequately protect the animals against disease,and to facilitate the safe movement of vaccinated animals [60].
Experimental challenge studies have demonstrated that vaccination with the BTV-2 MLV strain prevented viremia in at least 90.5% of vaccinated cattle that werechallenged at 7 months after vaccination with a dose of 2� 105.8 TCID50 of virulenthomologous field isolate [57]. In this trial, the control animals had viremias thatpersisted through the 35th day with titers above 105.8 TCID50/ml.
Serological studies performed on cattle and sheep that were vaccinated withseveral MLV combinations have shown that more than 80% of the vaccinatedanimals had specific BTV antibodies [41,43,55,58]. Colostral antibodies were foundin calves born from vaccinated dams until 39 days of age [58].
The efficacy of MLV vaccination has widely been demonstrated in the field.Following the 2000–01 and 2003 BT vaccination campaigns in the Balearic Islands,no outbreaks have been detected since December 2003 in the area.
With regard to the vaccination strategy in Italy, several points warrant attention.First, on the basis of a risk assessment [18] and considering the encouraging resultsof preliminary studies, the Italian Authorities decided to vaccinate all susceptibledomestic ruminant species (i.e. sheep, goats, cattle and water buffalo) in the infectedand at risk areas, with the aim of limiting direct losses and reducing virus BTVcirculation [16]. Mass vaccination of susceptible populations started in January 2002,although the starting dates and the percentages of vaccinated population achievedvaried greatly among regions [16]. In those areas where more than 80% of the targetpopulation was properly vaccinated before the new epidemic peak, clinical disease insheep disappeared almost completely and virus circulation was significantly reduced[16], with substantial benefit to internal animal trade/movement. The resultsobtained in some Italian regions with mass vaccination of all susceptible domesticruminants and the experience gained during the vaccination campaigns contributedto the modifications of BT international standards. Specifically, risk analysis can beused as an alternative to individual testing to assess immunity level in the populationof origin and determine the risk of spreading infection to free areas by movement ofvaccinated animals from infected territories [61]. In particular, the analysisperformed by Giovannini et al. [61] indicates that when more than 80% of thesusceptible population in the territory of origin was vaccinated, the risk associatedwith the movement of vaccinated animals to free areas appeared acceptable andcould further be mitigated by ancillary control measures.
In the absence of effective inactivated vaccines and in an emergency, MLVs stillrepresent a valid option for vaccination, provided that the quality, safety andefficacy of the MLV strains match EU standards. These vaccines can be an

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120116
alternative also in a non-emergency situation when local conditions (e.g. in case alarge amount of animal must be immunized in a very short period of time) indicatetheir use.
4. Recombinant vaccines
Several experimental recombinant vaccines have been described and they clearlyhave numerous inherent potential benefits, including rapid onset of immunity, lackof transmissibility and even a polyvalent strategy.
A recombinant vaccinia virus that expressed both VP2 and VP5 of AustralianBTV serotype 1 induced variable titers of neutralizing antibody in sheep andafforded protection against homologous challenge [62], but this approach has notbeen pursued further.
A recombinant capripoxvirus expressing VP7 was shown to provide partialprotection against heterologous BTV challenge [63], but like the recombinantvaccinia BTV vaccine, its development was not continued.
Finally, a recombinant canarypox virus-VP2/VP5 vaccine was recently describedthat induced highly effective protective immunity in sheep [20]. This vaccine has amajor inherent advantage in that the existing VP7 competitive ELISA assay woulddistinguish vaccinated from naturally infected animals (DIVA), and it utilizes anexpression vector that is incorporated in several vaccines already in use in the EUand elsewhere. The vaccine still is at a development stage.
5. Conclusions
Bluetongue vaccines may be used for different purposes or strategies, dependingon the epidemiological situation of the affected area and strategy desired. The mainpurposes of BT vaccination strategies are (i) to prevent clinical disease, (ii) to limitthe regional extension of BTV infection through reduction of the spread of the virus,(iii) to allow regional or country eradication of the disease based on the reduction ofvirus circulation and (iv) to authorize the safe movement of susceptible animalsbetween affected and free zones.
In BT endemic regions, vaccines have been used to prevent clinical disease anddeath losses in sheep. In these regions, vector Culicoides spp. vectors may be presentyear-round with continuous circulation of different BTV serotypes. This has led tothe design of multivalent vaccines containing different MLV serotypes, as done inSouth Africa, where BTV infection is endemic. The South African MLVs weredeveloped only to control clinical BT disease in sheep, as cattle and other ruminants,although susceptible to BTV infection, usually do not suffer clinical disease.
Since the incursion of BTV into previously non-endemic zones, as in someMediterranean countries, BTV vaccines are used as an aid to prevent furtherextension of the infection to border zones, for local/regional reduction of viruscirculation and for safe movements of animals, which play an important role in the

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120 117
European livestock industry. Depending on their availability, MLV or inactivatedserotype-specific vaccines can be used. Climatic and geographic factors as well asabundance of suitable BTV insect vectors are probably all important for the outcomeand persistence (reemergence) of BTV infection in an area. It is therefore commonlyaccepted that vaccines can help limit the spread of the disease. Ideally, for thepurpose of eradication, a successful vaccination campaign should cover allsusceptible ruminant species, attain a high degree of herd immunity and encompassextensive areas surrounding any active BT outbreak. Successful control also requiresrestriction of animal movements between BT-affected and BT-free zones.
Both MLV (outbreaks in Balearic Islands and Corsica in 2000 and 2001 and inItaly in 2003–04) and inactivated vaccines (outbreak in southwest Spain in 2004 and2005) have been successfully used after emergence of BTV in non-endemic areas.Although a risk model integrating factors determinant for BTV persistence(including vaccinations) needs to be developed, it is likely that eradication of BTVwith vaccines may be achievable only under concurrent favorable geo-climaticconditions. These conditions limit or prevent vector activity, and as a result, preventBTV circulation (e.g. cold winter impeding over wintering, geographical barriers,low vector densities, low probability of BTV incursion from neighboring zones), butthis relies on the assumption that vertical BTV transmission does not occur ininsects.
Acknowledgments
The authors gratefully thank the EFSA (European Food Safety Authority)working group on Bluetongue and in particular Dr Philippe Vannier, head of thisgroup. The majority of this paper will be included in an EFSA report on Bluetongue.
References
[1] Verwoerd D, Erasmus BJ. Bluetongue. In: Coetzer JA, Tustin RC, editors. Infectious diseases of
livestock. 2nd ed. Cape Town: Oxford University Press; 2004. p. 1201–20.
[2] Gibbs EP, Greiner EC. The epidemiology of bluetongue. Comp Immunol Microbiol Infect Dis
1994;17:197–206.
[3] Tabachnick WJ. Culicoides and the global epidemiology of bluetongue virus infection. Vet Ital
2004;40:145–50.
[4] Toussaint JF, Vandenbussche F, Mast J, De Meester L, Goris N, Van Dessel W, et al. Bluetongue in
northern Europe. Vet Rec 2006;2:327.
[5] Mertens PP, Diprose J, Maan S, Singh KP, Attoui H, Samuel AR. Bluetongue virus replication,
molecular and structural biology. Vet Ital 2004;40:426–37.
[6] Bonneau KR, Zhang N, Zhu J, Zhang F, Li Z, Zhang K, et al. Sequence comparison of the L2 and
S10 genes of bluetongue viruses from the United States and the People’s Republic of China. Virus Res
1999;61:153–60.
[7] Pritchard LI, Sendow I, Lunt R, Hassan SH, Kattenbelt J, Gould AR, et al. Genetic diversity of
bluetongue viruses in south east Asia. Virus Res 2004;101:193–201.
[8] Bonneau KR, MacLachlan NJ. Genetic diversification of field strains for bluetongue virus. Vet Ital
2004;40:446–7.

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120118
[9] Bonneau KR, Mullens BA, MacLachlan NJ. Occurrence of genetic drift and founder effect during
quasispecies evolution of the VP2 and NS3/3A genes of bluetongue virus upon passage between sheep
and cattle. J Virol 2001;75:8298–305.
[10] Calistri P, Giovannini A, Conte A, Nannini D, Santucci U, Patta C, et al. Bluetongue in Italy: part I.
Vet Ital 2004;40(3):243–51.
[11] OIE World Organisation for Animal Health. Bluetongue in Italy. Dis Inf 2000;13:150.
[12] OIE World Organisation for Animal Health. Bluetongue in Spain: in the Balearic Islands. Dis Inf
2000;13:181.
[13] Zientara S, Sailleau C, Dauphin G, Roquier C, Remond M, Lebreton F, et al. Identification of
bluetongue virus serotype 2 (Corsican strain) by reverse-transcriptase PCR reaction analysis of
segment 2 of the genome. Vet Rec 2002;150(19):598–601.
[14] Breard E, Hamblin C, Hammoumi S, Sailleau C, Dauphin G, Zientara S. The epidemiology and
diagnosis of bluetongue with particular reference to Corsica. Res Vet Sci 2004;77:1–8.
[15] MAPA. Review of the situation of bluetongue in Spain since the outbreak of BT 2 in the Balearic
Island. MAPA report, 2006.
[16] Patta C, Giovannini A, Rolesu S, Nannini D, Savini G, Calistri P, et al. Bluetongue vaccination in
Europe: the Italian experience. Vet Ital 2004;40(4):601–10.
[17] MAPA. Report of the efficacy test performed in Spain with different commercially available vaccine
against BTV4 in cattle and sheep. MAPA report, 2006.
[18] Giovannini A, MacDiarmid S, Calistri P, Conte A, Savini L, Nannini D, et al. The use of risk
assessment to decide the control strategy for bluetongue in Italian ruminant populations. J Risk Anal
2003;24(6):1737–53.
[19] Murray PK, Eaton BT. Vaccine for bluetongue. Aust Vet J 1996;73:207–10.
[20] Boone JD, Balasuriya UB, Karaca K, Audonnet JC, Yao J, He L, et al. Recombinant canarypox
virus vaccine co-expressing genes encoding the VP2 and VP5 outer capsid proteins of bluetongue
virus induces high level protection in sheep. Vaccine 2007;25:672–8.
[21] Stott JL, Barber TL, Osburn BI. Immunologic response of sheep to inactivated and virulent
bluetongue virus. Am J Vet Res 1985;46:1043–9.
[22] Di Emidio B, Nicolussi P, Patta C, Ronchi GF, Monaco F, Savini G, et al. Efficacy and safety studies
on an inactivated vaccine against bluetongue virus serotype 2. Vet Ital 2004;40:640–4.
[23] Roy P, Urakawa T, Van Dijk AA, Erasmus BJ. Recombinant virus vaccine for bluetongue disease in
sheep. J Virol 1990;64:1998–2003.
[24] Roy P, French T, Erasmus BJ. Protective efficacy of virus-like particles for bluetongue disease.
Vaccine 1992;10:28–32.
[25] Roy P, Bishop DH, LeBlois H, Erasmus BJ. Long-lasting protection of sheep against bluetongue
challenge after vaccination with like-like particles: evidence for homologous and partial heterologous
protection. Vaccine 1994;12:805–11.
[26] Roy P. Genetically engineered structure-based vaccine for bluetongue disease. Vet Ital 2004;40:
594–600.
[27] Hamers C, Werle-Lapostolle B, Rehbein S, Blanchet M, Mure-Ravaud K, Schumacher C, Hudelet P.
Six month efficacy of an inactivated BTV-2 industrial vaccine against a virulent BTV-2 challenge in
sheep. In: Proceedings of the ninth international symposium on double-stranded RNA viruses, Cape
Town, South Africa, 21–26 October 2006.
[28] Savini G, Hamers C, Migliaccio P, Leone A, Hudelet P, Schumacher C, Caporale V. Assessment
of efficacy of a bivalent inactivated vaccine against bluetongue virus serotypes 2 and 4 in cattle.
In: Proceedings of the 24th world buiatrics congress, Nice, France, 15–19 October 2006.
[29] Savini G, Conte A, Di Gennaro A, Leone A, Migliaccio P, Bonfini B, Di Ventura M, Monaco F.
Virological and serological responses in cattle following vaccination with modified-live vaccine
against bluetongue virus serotypes 2, 4, 9 and 16. In: Proceedings of the ninth international
symposium on ds-RNA viruses, Cape Town, South Africa, 21–26 October 2006.
[30] Jimenez-Clavero MA, Aguero M, San Miguel E, Mayoral T, Cruz Lopez M, Ruano MJ, et al. High
throughput detection of bluetongue virus by a new real-time on clinical samples from current
mediterranean outbreaks. J Vet Diagn Invest 2006;18:7–17.

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120 119
[31] Hamers C, Hudelet P, Blanchet M, Werle-Lapostolle B, Sailleau C, Breard E, Zientara S. Efficacy of
an inactivated bivalent BTV-2/BTV-4 industrial vaccine against a BTV-2 or BTV-4 virulent challenge
in sheep. In: Proceedings of the ninth international symposium on double-stranded RNA viruses,
Cape Town, South Africa, 21–26 October 2006.
[32] Caporale V, Giovannini A, Patta C, Calistri P, Nannini D, Santucci U. Vaccination in the control
strategy of bluetongue in Italy. (Developments in biologicals, vol. 119). Control of infectious animal
diseases by vaccination. Basel, Switzerland: S Karger AG; 2004. p. 113–27. [book chapter. conference
paper].
[33] Dungu B, Gerdes T, Smit T. The use of vaccination in the control of bluetongue in southern Africa.
Vet Ital 2004;40:616–22.
[34] MacLachlan NJ, Osburn BI, Stott JL. Ghalib HW orbivirus infection of the bovine fetus. Prog Clin
Biol Res 1985;178:79–84.
[35] Venter GJ, Gerdes GH, Mellor PS, Paweska JT. Transmission potential of South African Culicoides
species for live-attenuated bluetongue virus. Vet Ital 2004;40:198–202.
[36] Savini G, Monaco F, Facchinei A, Pinoni C, Salucci S, Cofini F, et al. Field vaccination of sheep with
bivalent modified-live vaccine against bluetongue virus serotypes 2 and: effect on milk production.
Vet Ital 2004;40(4):627–30.
[37] Monaco F, De Luca N, Spina P, Morelli D, Liberatore I, Citarella R, et al. Virological and
serological response of cattle following field vaccination with bivalent modified-live vaccine against
bluetongue virus serotypes 2 and 9. Vet Ital 2004;40(4):657–60.
[38] Monaco F, De Luca N, Morelli D, Pisciella M, Palmarini S, Di Giandomenico M, et al. Field
vaccination of cattle using a bivalent modified-live vaccine against bluetongue virus serotypes 2 and 9:
effect on milk production. Vet Ital 2004;40(4):661–3.
[39] Ferrari G, De Liberato C, Scavia G, Lorenzetti R, Zini M, Farina F, et al. Active circulation of
bluetongue vaccine virus serotype-2 among unvaccinated cattle in central Italy. Prev Vet Med
2005;68:10–3.
[40] Veronesi E, Hamblin C, Mellor PS. Live attenuated bluetongue vaccine viruses in Dorset Poll sheep,
before and after passage in vector midges (Diptera: Ceratopogonidae). Vaccine 2005;23(48/49):5501–9.
[41] Hammoumi S, Breard E, Sailleau C, Russo P, Grillet C, Cetre-Sossah C, et al. Innocuity and activity
of the monovalent bluetongue serotype 2 vaccine. Specific detection of the genome of the vaccine
strain by RT-PCR. J Vet Med 2003;B50:316–21.
[42] Ronchi G F, Franchi P, Battistini L, Di Emidio B. Stability study of bluetongue attenuated live
vaccine serotype 2 after rehydration. Vet Ital 2003;39(2):11–4.
[43] Gerbier G, Hendrikx, P, Roger F, Zientara S, Biteau-Coroller F, Grillet C, Baldet T, Albina E.
Bluetongue control using vaccines: an experience from the mediterranean islands in Europe. In:
Proceedings of the third OIE bluetongue international symposium, Taormina, Sicily, 26–29 October 2003.
[44] IZSAM, Istituto Zooprofilattico Sperimentale dell’Abruzzo e del Molise ‘G. Caporale’. Safety and
potency testing of bluetongue vaccines. Testing of monovalent bluetongue vaccine (serotype 2) for
innocuity, potency and reversion to virulence. Contract DG Sanco 00/0127. Final report. Teramo,
2001. 19pp. /http://www.east-btnet.izs.it/document_r/research/final_report.pdfS.
[45] IZSAM, Istituto Zooprofilattico Sperimentale dell’Abruzzo e del Molise ‘G. Caporale’. Vaccino e
danni vaccinali, /http://gis2.izs.it:7777/bluetongue/doc_tec/danni_vacc/vaccini.pdfS, 2004.
[46] Zientara S, Breard E, Sailleau C. Bluetongue: characterization of virus types by reverse transcription-
polymerase chain reaction. In: Vannier P, Espeseth D, editors. New diagnostic technology:
applications in animal health and biologics controls. Basel, IABs/Karger series, Developments in
biologicals 2006, vol. 126. p.187–96. ISSN: 1424-6074.
[47] Monaco F, Camma C, Serini S, Savini G. Differentiation between field and vaccine strain of
bluetongue virus serotype 16. Vet Microbiol 2006;116:45–52.
[48] Italian Ministry of Health. Nota del 19 gennaio. Bluetongue—provvedimenti ed impiego del vaccino
sierotipo 16 nella IV campagna di vaccinazione 2004–2005, 2005. n.1720.
[49] Monaco F, Bonfini B, Zaghini M, Antonucci D, Pini A & Savini G. Vaccination of cattle using
monovalent modified-live vaccine against bluetongue virus serotype 2: innocuity, immunogenicity
and effect on pregnancy. Vet Ital 2004;40(4):671–5.

ARTICLE IN PRESS
G. Savini et al. / Comp. Immun. Microbiol. Infect. Dis. 31 (2008) 101–120120
[50] Lucifora G, Rossi P, Calistri P, Giovannini A. Effects of bivalent bluetongue virus serotypes 2 and 9
vaccine on reproductive performance of cattle: a case study in Calabria, Italy. Vet Ital 2004;40:654–6.
[51] Breard E, Pozzi N, Sailleau C, Catinot V, Durand B, Dumont P, et al. Transient effect of the
attenuated bluetongue virus vaccine on the quality of the ram semen. Vet Rec 2007;160(13):431–5.
[52] Caporale V, Di Emidio B, Nicolussi P, Piredda G, Salaris S, De Palmas S, et al. Experimental
vaccination of sheep using a bluetongue attenuated live vaccine: a study of the effects on lactation.
Vet Ital 2003;39(4):5–10.
[53] Cannas EA, Nicolussi P, Bonelli P, Dore S, Denti G, Marongiu E, Migliaccio P, Savini G. Live
modified bluetongue vaccine viruses in Sarda sheep: effects of three different formulations on milk
production. In: Proceedings of the 12th international symposium of the world association of
veterinary laboratory diagnosticians, Montevideo, Uruguay, 16–19 November 2005.
[54] Giovannini A, Conte A, Panichi G, Calistri P, Dessı M, Foddis F, et al. Effects of vaccination against
bluetongue on milk production and quality in cattle vaccinated with live-attenuated monovalent type
2 vaccine. Vet Ital 2004;40(4):648–53.
[55] Savini G, Monaco F, Conte A, Migliaccio P, Casaccia C, Salucci S, et al. Virological and serological
response of sheep following field vaccination with bivalent modified-live vaccine against bluetongue
virus serotypes 2 and 9. Vet Ital 2004;40(4):631–4.
[56] Savini G, Tittarelli M, Bonfini B, Zaghini M, Di Ventura M, Monaco F. Serological response in cattle
and sheep following infection or vaccination with bluetongue virus. Vet Ital 2004;40(4):645–7.
[57] Savini G, Monaco F, Citarella R, Calzetta G, Panichi G, Ruiu A, et al. Monovalent modified-live
vaccine against bluetongue virus serotype 2: immunity studies in cows. Vet Ital 2004;40(4):664–7.
[58] Savini G, Monaco F, Calistri P, Panichi G, Ruiu A, Leone A, et al. Neutralising antibody response in
cattle after vaccination with monovalent modified-live vaccine against bluetongue virus serotype 2.
Vet Ital 2004;40(4):668–70.
[59] Savini G, Cannas EA, Bonelli P, Petruzzi M, Fresi S, Dimauro C, Di Gennaro A, Nicolussi P. Live
modified bluetongue vaccine viruses in sarda sheep: clinical signs, haematology and chemistry. In:
Proceedings of the 12th international symposium of the world association of veterinary laboratory
diagnosticians, Montevideo, Uruguay, 16–19 November 2005.
[60] OIE. Terrestrial animal health code. 2005 ed. World Animal Health Organization.
[61] Giovannini A, Conte A, Calistri P, Di Francesco C, Caporale V. Risk analysis on the introduction
into free territories of vaccinated animals from restricted zones. Vet Ital 2004;40(4):697–702.
[62] Lobato ZI, Coupar BE, Gray CP, Lunt R, AndrewME. Antibody responses and protective immunity
to recombinant vaccinia expressed-expressed bluetongue virus antigens. Vet Immunol Immunopathol
1997;59:293–309.
[63] Wade-Evans AM, Romero CH, Mellor P, Takamatsu H, Anderson J, Thevasagayam J, et al.
Expression of the major core structural protein (VP7) of bluetongue virus, by a recombinant capripox
virus, provides partial protection of sheep against a virulent heterotypic bluetongue virus challenge.
Virology 1996;220:227–31.




















