
Use of plant water relations to assess forage quality and growth for two cultivars of Napier grass...
12
Use of plant water relations to assess forage quality and growth for two cultivars of Napier grass (Pennisetum purpureum) subjected to different levels of soil water supply and temperature regimes S. W. Mwendia A,C , I. A. M. Yunusa A , R. D. B. Whalley A , B. M. Sindel A , D. Kenney A , and I. W. Kariuki B A School of Environmental and Rural Science, University of New England, Armidale, NSW 2351, Australia. B Kenya Agricultural Research Institute, Muguga South, 30148-00100, Nairobi, Kenya. C Corresponding author. Email: [email protected] Abstract. Napier grass (Pennisetum purpureum Schumach.) is an important fodder and relatively drought-tolerant crop in tropical and subtropical regions, especially in developing countries. For this and other species, tools are needed for identifying drought-tolerant cultivars to aid selection for semi-arid environments. We determined tissue water status, carbon assimilation, biomass yield and forage quality for Napier grass cvv. Bana and Atherton grown in bins and subjected to three soil-water supply levels (100, 50 or 25% of field capacity) in glasshouses set at either low (15258C) or high (25358C) temperature regimes, over three growing cycles. Our aim was to explore whether differences in leaf water potential (LWP) and carbon assimilation rates could be reliable indicators of the relative yield potential and forage quality of the two cultivars in environments prone to water and heat stresses. At the low soil-water supply of 25% and low temperature, Bana had lower (more negative) LWP and relative water content (RWC) than Atherton, while at 50% and 100% soil-water supply, Bana had a higher tissue water status. Under the high temperature regime, Bana had consistently more positive LWP and RWC than Atherton, but the differences were not significant. The two cultivars had a similar CO 2 assimilation rate (A) and there were no significant differences in the total dry matter yields over the three growing cycles. Water-use efficiency for above-ground biomass (kg ha –1 mm –1 ) was similar for both cultivars and was 28.5–35.1 under the low temperature regime and 16.9–22.9 under the high temperature regime. Neutral detergent fibre (NDF) was often higher for Bana at low water supply and low temperature than for Atherton, but the trend was reversed under the high temperature regime. Digestibility was generally improved under water-stressed conditions, and there was a positive correlation between NDF and both LWP and RWC measured at midday, but only under the low temperature regime. We conclude that LWP, RWC and A, alone or together, are inadequate for selecting cultivars for dry and hot environments, because cultivars may differ in other mechanistic responses to water stress and high temperatures. Additional keywords: dry matter yield, leaf area, root : shoot ratio, water-use, water-use efficiency. Received 23 April 2013, accepted 21 November 2013, published online 13 December 2013 Introduction Persistence and productivity of crop and pasture species in dry environments depends on the capacity of plants to maintain favourable tissue water status and sustain carbon assimilation rates. Maintaining tissue turgidity during the day is necessary to keep the stomata open. Leaf water potential (LWP) as a measure of the pressure needed to balance the force with which the water is held in the xylem is a useful tool that is routinely used to analyse tissue hydration, and hence the physiological health of the plant under limiting soil water availability (Niklas and Spats 2012). While physiological functions in plants are known to be close to optimum at LWP of ~–0.30 MPa when soil water availability is near maximum, they tend to decline as LWP falls to the theoretical limit of –1.5 MPa (Lambers et al. 2008). Because of its sensitivity to transient changes in availability of soil water, LWP can be an effective tool for understanding drought tolerance in cultivars and possibly identifying those that have this attribute. Prolonged, limited water supply during droughts makes the maintenance of full tissue hydration with the leaves turgid and stomata opened for transpiration and carbon assimilation increasingly difficult (Volaire and Thomas 1995; Zhou et al. 2009). Prolonged, limited water supply also compromises the capacity of plants to fully rehydrate and restore carbon assimilation to full capacity on rewatering (Norris and Thomas 1982). Generally, plant species avoid a deleterious decline in Journal compilation Ó CSIRO 2013 www.publish.csiro.au/journals/cp CSIRO PUBLISHING Crop & Pasture Science, 2013, 64, 1008–1019 http://dx.doi.org/10.1071/CP13254
Transcript of Use of plant water relations to assess forage quality and growth for two cultivars of Napier grass...

Use of plant water relations to assess forage quality and growthfor two cultivars of Napier grass (Pennisetum purpureum)subjected to different levels of soil water supplyand temperature regimes
S. W. MwendiaA,C, I. A. M. YunusaA, R. D. B. WhalleyA, B. M. SindelA, D. KenneyA,and I. W. KariukiB
ASchool of Environmental and Rural Science, University of New England, Armidale, NSW 2351, Australia.BKenya Agricultural Research Institute, Muguga South, 30148-00100, Nairobi, Kenya.CCorresponding author. Email: [email protected]
Abstract. Napier grass (Pennisetum purpureum Schumach.) is an important fodder and relatively drought-tolerant crop intropical and subtropical regions, especially indevelopingcountries. For this andother species, tools areneeded for identifyingdrought-tolerant cultivars to aid selection for semi-arid environments. We determined tissue water status, carbonassimilation, biomass yield and forage quality for Napier grass cvv. Bana and Atherton grown in bins and subjected tothree soil-water supply levels (100, 50or 25%offield capacity) in glasshouses set at either low (15�258C)or high (25�358C)temperature regimes, over three growing cycles. Our aim was to explore whether differences in leaf water potential (LWP)and carbon assimilation rates could be reliable indicators of the relative yield potential and forage quality of the two cultivarsin environments prone to water and heat stresses.
At the low soil-water supply of 25% and low temperature, Bana had lower (more negative) LWP and relative watercontent (RWC) than Atherton, while at 50% and 100% soil-water supply, Bana had a higher tissue water status. Under thehigh temperature regime, Bana had consistently more positive LWP and RWC than Atherton, but the differences were notsignificant. The two cultivars had a similar CO2 assimilation rate (A) and there were no significant differences in the totaldry matter yields over the three growing cycles.Water-use efficiency for above-ground biomass (kg ha–1 mm–1) was similarfor both cultivars and was 28.5–35.1 under the low temperature regime and 16.9–22.9 under the high temperature regime.Neutral detergent fibre (NDF) was often higher for Bana at low water supply and low temperature than for Atherton, butthe trend was reversed under the high temperature regime. Digestibility was generally improved under water-stressedconditions, and there was a positive correlation between NDF and both LWP and RWC measured at midday, but onlyunder the low temperature regime. We conclude that LWP, RWC and A, alone or together, are inadequate for selectingcultivars for dry and hot environments, because cultivars may differ in other mechanistic responses to water stress andhigh temperatures.
Additional keywords: dry matter yield, leaf area, root : shoot ratio, water-use, water-use efficiency.
Received 23 April 2013, accepted 21 November 2013, published online 13 December 2013
Introduction
Persistence and productivity of crop and pasture species in dryenvironments depends on the capacity of plants to maintainfavourable tissue water status and sustain carbon assimilationrates. Maintaining tissue turgidity during the day is necessary tokeep the stomata open. Leaf water potential (LWP) as a measureof the pressure needed to balance the force with which the wateris held in the xylem is a useful tool that is routinely used toanalyse tissue hydration, and hence the physiological health ofthe plant under limiting soil water availability (Niklas andSpats 2012). While physiological functions in plants areknown to be close to optimum at LWP of ~–0.30MPa whensoil water availability is near maximum, they tend to decline as
LWP falls to the theoretical limit of –1.5MPa (Lambers et al.2008). Because of its sensitivity to transient changes inavailability of soil water, LWP can be an effective tool forunderstanding drought tolerance in cultivars and possiblyidentifying those that have this attribute.
Prolonged, limited water supply during droughts makes themaintenance of full tissue hydration with the leaves turgid andstomata opened for transpiration and carbon assimilationincreasingly difficult (Volaire and Thomas 1995; Zhou et al.2009). Prolonged, limited water supply also compromises thecapacity of plants to fully rehydrate and restore carbonassimilation to full capacity on rewatering (Norris and Thomas1982). Generally, plant species avoid a deleterious decline in
Journal compilation � CSIRO 2013 www.publish.csiro.au/journals/cp
CSIRO PUBLISHING
Crop & Pasture Science, 2013, 64, 1008–1019http://dx.doi.org/10.1071/CP13254

LWP by shutting their stomata as the first safeguard againstpossible desiccation, whereas longer term responses includealterations in leaf area and/or root growth (Yordanov et al.2000).
Temperature stresses due to elevated ambient temperaturescan exacerbate deleterious effects of deficits in soil-water supply(Hatch 1992). Although C4 plants are adapted to high ambienttemperatures, their physiological processes can be adverselyaffected when temperature rises above a certain threshold. Forinstance, when ambient day/night temperature was raised from28/238C to 40/358C, midday LWP declined from –0.5 to–1.3MPa and relative water content (RWC) from 72 to 60%in sugarcane (Wahid and Close 2007). Furthermore, stomatalconductance in tropical grasses was reportedly curtailed as LWPdropped towards –1.5MPa (Neil et al. 2003). However, severaltropical grasses (Panicum maximum, Heteropogon contortusand Cenchrus ciliaris) have been found to maintain favourabletissue moisture and photosynthesis at extremely negative LWPof –2.4 to –1.9MPa, with the gas exchange ceasing only whenLWP dropped below –3.0MPa, or even lower to –4.0MPa, inP. maximum and H. contortus (Ludlow et al. 1985). When highLWP is maintained through stomatal closure, opportunitiesfor assimilating carbon are constrained and would thereforereduce growth in the long term (Yunusa et al. 2005). Periodsof water stress and high temperatures may also affect foragequality, such as protein content and digestibility in addition to drymatter production of pasture grasses (Anindo and Potter 1994;Nyaata et al. 2000).
Species and cultivars may differ in their response to variablesoil-water supply and ambient temperatures (Wilson 1983;Guenni et al. 2002). Mechanisms involved in these responsesare still poorly understood in many respects (Fernández andReynolds 2000). Increased understanding of these responseswould be of value in breeding and selecting appropriatecultivars of pastures and crops for semi-arid environments.
Napier grass (Pennisetum purpureum Schumach.) has severaladvantages as a fodder in tropical and subtropical environments,including ease of establishment, high dry matter production,persistence and regeneration (Nyambati et al. 2011). It istherefore well suited for tropical and subtropical countriessuch as Kenya, where it supports �70% of dairy farmers whoproduce 80% of the traded milk (Staal et al. 1998). Increaseddemand for dairy products by rapidly rising populations inmany of these countries is driving expansion of smallholderdairies into more marginal areas characterised by low rainfalland high temperatures (Esilaba et al. 2011; Qadir et al. 2013).However, this grass has received little research investment tounderstand its physiological responses to water and temperaturestresses. Such an understanding could be used to identifycultivars for these new, drier and hotter environments relativeto the current major cultivation regions of this grass. Thisglasshouse study was undertaken with two aims: (i) tounderstand how two cultivars of Napier grass respond todifferent levels of soil-water supply and ambient temperatureconditions with respect to their tissue water status, carbonassimilation, yield potential and forage quality; and (ii) toexplore whether the responses in LWP and carbon assimilationare reliable indicators of the forage quality and relative yieldpotential of the two cultivars.
Material and methodsStudy design
The experiment was a complete randomised design with twofactors: (i)Napier grasswith two cultivars, (ii) andwater supply atthree levels; these were set up in two temperature environments(glasshouse bays) and replicated four times. The experimentalunit was a plastic bin 0.39m in diameter and 0.46m deep. Theceilings of the glasshouse bays were ~2.5m above the tops of thebins, allowing substantial growth of the Napier grass plants. Thestudy was conducted at the University of New England, NewSouth Wales, Australia.
Planting
The bins were filled with 50 kg of sandy Alfisol (Klingebiel andMontgomery 1961) soil passed through a 10-mm sieve, collectedfrom 0.1–0.2m depth at Trevenna University farm at theUniversity of New England. Diammonium phosphate fertiliser(NPK, 18 : 20 : 0) was applied at a rate of 13 g per bin, equivalentto 26 kg P ha–1. One Napier grass cane (with two nodes) wasplanted per bin with one node below the soil on 12 May 2011.Alcathene beads (1 kg bin–1) were spread on the soil surfaceto reduce evaporation losses. Bins were labelled and theirplacement randomised. The canes were initially grown over28 days before transplanting.
Cultivars
Two cultivars of Napier grass, Bana and Atherton, werecollected at Atherton in Queensland, Australia (17.26258S,145.47698E). Bana is an infertile, interspecific hybrid betweenPennisetumpurpureum andPennisetumglaucum (L.) R.Br. usedas windbreaks between crops in some tropical and subtropicalparts of Australia. The Atherton cultivar (P. purpureum) wasintroduced into Atherton some years ago but its origin isuncertain. It was designated Atherton for the purpose of thisstudy.
Water-supply treatments
All the bins were watered to field capacity at the start of theexperiment. This was achieved bywatering at 100% as guided by‘recharge’ bins, described below. Thereafter, water supply toeach bin was maintained at 25, 50 or 100% of field capacity.To determine the amount of water required to achieve theselevels, eight ‘recharge’ bins were set up (four in each bay).A recharge bin contained 50 kg of soil, with four holes(25.4-mm-diameter fitted with a sieve) at the base, and wasplaced on a pallet to allow excess water to drain throughovernight into a collecting basin. The basin had a polythenesheet fastened over the top with an inlet at the centre to allowwater to enter while inhibiting evaporation. There were tworeplicates of recharge bins for each Napier grass cultivar ateach temperature regime. Prior to watering the treatment bins,the recharge bins were thoroughly watered and allowed to drainovernight. The amount of full recharge was obtained as thedifference between the amounts of water applied and thatcollected in the plastic basin. The average of this amount forthe four recharge bins in each bay was applied to the treatmentbins with 100%water supply; the amounts of water applied to theother treatments were scaled back accordingly. Watering
Napier performance under variable environments Crop & Pasture Science 1009

treatments were initiated on 10 June 2011 and were undertakenwhen the leaves wilted. Wilting was characterised by a visibleloss of rigidity in the leaves, so that they drooped and were nolonger upright. After the first growth cycle, all of the bins werewatered to field capacity and topdressed with urea (46% N)at a rate of 13 g bin–1, equivalent to 26 kgN ha–1. Subsequentwatering (second and third growth cycles) reverted to therespective water treatments when the tillers wilted. In the firstgrowth cycle, there were three irrigations at low temperature andfour at high temperature, whereas in the second and third cyclesthere were four irrigations each at low temperature and five athigh temperature.
Temperature regimes
The two glasshouse bays were maintained at diurnaltemperatures of either 15/258C or 25/358C. The transitionsbetween the limits in set temperatures occurred over 3-hperiods at 07:00–10 : 00 and 19:00–22 : 00 each day.Temperature and relative humidity were automatically loggedusing CN7510A sensors (Honeywell International Inc.,Morristown, NJ, USA) inbuilt in each bay for the entire trialperiod. The sensors were connected to a computer so that thetemperature could be set to the desired levels. Temperature andrelative humidity were used to compute vapour pressure deficit(VPD) according to Monteith and Unsworth (2008).
Measurements
Leaf water potential measurements were made using a pressurechamber (Scholander et al. 1965). Samples were excised fromthe youngest fully developed leaf and immediately pressurisedin the chamber. A pressure reading was recorded once waterappeared on the excised edge. Measurements were done aroundmidday at weekly intervals, and at predawn (05 : 00) and aroundmidday on the day of watering, which occurred at 18 : 00.Following watering, measurements were made duringthe next day at 05 : 00, 07 : 00, 09 : 00, 12 : 00, 16 : 00 and19 : 00. Relative water content was determined as described byBarrs (1968). Leaf segments were excised at the same times asthose for LWPand placed in vials, capped and put in a cooler box.These were weighed fresh (F) and, then after being floated ondeionised water for 4 h at room temperature and mopped with afilter-paper, were reweighed to obtain the ‘fully turgid’ weight(T). They were then dried for 24 h in an oven at 808C to give thedry weight (D). The RWC was calculated as:
½ðF� DÞ=ðT� DÞ� � 100
Data were collected for three growth cycles. Gas exchangewas monitored at midday on sunny days using a Portable
Photosynthesis System LI-6400XT (LI-COR Inc., Lincoln,NE, USA) and ambient light intensity, but photosyntheticallyactive radiation (PAR) generally ranged from 900 to1260mmolm–2 s–1 (full sunlight is ~1300mmolm–2 s–1).Variables measured in the process were photosynthetic carbonassimilation, stomatal conductance and leaf transpiration.Readings were taken on the youngest fully developed leafat week 4 of growth (first growth cycle), weeks 4 and 7(second growth cycle), and week 4 and 8 (third growth cycle).Photosynthetic carbon assimilation was divided by leaftranspiration to obtain photosynthetic water use efficiency(photosynthetic WUE).
Leaf area was measured on a fully developed, second-youngest leaf of one tiller per bin. A single sample leaf washarvested per bin on the day before harvesting the wholeexperiment. The fresh leaves were weighed and their areasmeasured using a CI-202 Portable Laser Leaf Area Meter(CID Bio-Science Inc., Camas, WA, USA). The sample wasthen oven-dried at 608C and dry matter added to the bin drymatter yield. This was done for the three growth cycles. Total leafarea was estimated from the total dry leaf weights. Dry matter(DM) yields were determined at the end of each growth cycle,which lasted 8 weeks. There were three harvests throughout theexperimental period. At each harvest, the plants in each bin werecut off at the level of the alcathene beads using secateurs, andthe fresh weight was determined immediately using an electronicbalance. Harvested plants were taken to the laboratory andmanually separated into leaves and stems, then oven-dried at608C for 48 h. These were weighed separately to compute leaf/stem ratios on a DM basis.
Root biomass was determined after the third growth cycleby weighing all the roots recovered from each bin. The soilcolumn was emptied into an aluminium tray and dividedinto two equal parts, i.e. top and bottom, using a mattock. Soilsamples of ~60 g were taken from the top and bottom halvesseparately weighed, dried at 1058C for 24 h and weighed againfor moisture determination. Roots were clean-washed and blot-dried, weighed, then oven-dried as earlier described. Root dryweights were divided by cumulative aboveground biomass togive root/shoot ratios. Neutral detergent fibre (NDF) wasdetermined on dried samples of stems and leaves (wholeshoot). These were ground to pass through a 1-mm diametersieve andwere then analysed to determineNDF. TheNDF for thefirst harvest was analysed according to procedures of AOAC(1975). Results from this analysis were then used to calibrate anear infrared machine for subsequent NDF analysis (second andthird growth cycles). Nitrogen (N) content was analysed from theground samples using CN-1000 Autoanalyzer (LECOCorporation, St. Joseph, MI, USA).
Table 1. Average temperatures and vapour pressure deficits (VPD) during the three growthcycles in the glasshouse bays set at either low or high temperatures
Values in parentheses denote standard deviation
Growth cycle Temperature regime (8C) VPD (kPa)(dates) Low High Low temp. High temp.
First (08.vi.11–04.viii.11) 20.5 (0.6) 28.5 (0.7) 1.41 (0.16) 2.80 (0.19)Second (05.viii.11–30.ix.11) 20.8 (0.7) 28.6 (0.5) 1.32 (0.17) 2.69 (0.19)Third (01.x.11–25.xi.11) 22.3 (0.9) 29.1 (1.2) 0.99 (0.30) 2.27 (0.43)
1010 Crop & Pasture Science S. W. Mwendia et al.

Water-use efficiency was computed by dividing total DMyield by total water applied over the experimental period. Waterutilised constituted all of the applied water plus soil wateravailable during planting, less water in the soil at end of thethird growth cycle.
Data analyses
Datasets were prepared in an Excel spread sheet and checkedfor entry errors. Analyses of variance were done using Minitab
version 15 (Minitab Inc. 2007). Means were separated by leastsignificant difference. Standard errors were calculated usingthe formula s.e. = (SD/Hn) where SD is the standard deviationand n is the number of observations.
Results
Mean daily temperature during the three growth cycles was20.5, 20.8 and 22.38C under the low temperature regime, and
Table 2. Leaf relative water content andwater potential at predawn andmidday pooled for the three growth periods forNapier grass subjected to variable water supply levels under low and high temperature
For each parameter, either pre-dawn or midday and within temperature regime, means followed by the same letter are notsignificantly different at P= 0.05
Water Low temperature High temperaturesupply Pre-dawn Midday Pre-dawn Midday
Bana Atherton Bana Atherton Bana Atherton Bana Atherton
Relative water content (%)25% 73.5b 84.5ab 63.4c 74.8ab 75.4ab 64.0b 61.4ab 55.1b50% 86.6a 73.6b 77.8ab 63.2c 76.9a 72.8ab 63.5a 59.3ab100% 90.6a 79.9ab 81.1a 67.7bc 81.9a 75.5ab 65.2a 58.3ab
l.s.d. (P= 0.05) 12.4 10.9 11.5 8.0
Leaf water potential (MPa)25% –2.50c –1.73bc –2.88b –2.27b –2.17bc –2.77c –2.85c –2.44bc50% –1.25ab –1.95bc –2.16ab –2.66b –1.97ab –2.31bc –2.27ab –2.68bc100% –0.74a –1.39ab –1.33a –2.03ab –1.28a –1.67abc –1.82a –2.21ab
l.s.d. (P= 0.05) 0.91 0.87 0.79 0.52
80
85
90
95
100
RW
C (
%)
BanaAtherton
(a)
(c)
(b)
(d)
805 7 9 12 16 19 5 7 9 12 16 19
85
90
95
100
2550100
Time of day (hours)
BanaAtherton
2550100
Fig. 1. Mean values (�s.e.) for diurnal trends in leaf relative water content (RWC) for Napier grass cvv.Bana and Atherton averaged for the three growth cycles. The measurements were taken at week 7 (firstcycle) and 5 (second and third cycles) of the growth cycle. (a) Low-temperature glasshouse bay, averagedover all water supply levels; (b) high-temperature glasshouse bay, averaged over all water supply levels;(c) low-temperature glasshouse bay, averaged over both cultivars; (d) high-temperature glasshouse bay,averaged over both cultivars.
Napier performance under variable environments Crop & Pasture Science 1011

28.5, 28.6 and 29.18C under high temperature regime(Table 1). Corresponding daily means for VPD were 1.41,1.32 and 0.99 kPa for the first, second and third growth cycles,respectively, under the low temperature regime, and 2.80, 2.69and 2.27 kPa under the high temperature regime.
Leaf relative water content and water potential
The responses in LWP and RWC were consistent among thetreatments during the three growth cycles; therefore, the datawere pooled and averaged for the whole study period(Table 2). Under low temperature conditions, Bana generallymaintained higher RWC than Atherton in the absence of waterdeficit both at predawn and at midday, whereas Atherton hadhigher RWC at midday when water supply was reduced to 25%.The RWC for both cultivars was lower under high temperatureconditions than low temperature conditions. Bana hadconsistently more positive LWP when fully supplied withwater at both predawn and midday compared with the 25%water treatment, but the LWP of Atherton was notsignificantly affected when water supply was reduced to 25%,under both temperature regimes (Table 2).
Diurnal trends in RWCwere mostly similar for both cultivarsunder both temperature regimes, except for Atherton having ahigher value at predawn under high temperature conditions(Fig. 1b). The RWC declined during the day from predawn to
the lowest values aroundmidday, and the reductionwas larger forplants watered at 25% than either 50% or 100% at lowtemperature, with Bana having the greatest reduction in bothtemperature regimes. Reducing watering to 25% also led to morenegative LWP, especially during the morning hours at hightemperatures (Fig. 2d). The diurnal course in LWP under bothtemperature regimes showed more positive values for Athertonthan for Bana at midday (Fig. 2), at a time that plants becamestressed when water supply was reduced to 25%. Mean LWPdid not become more negative than –1.4MPa for either cultivarunder any conditions (Fig. 2).
Stomatal attributes
Carbon dioxide assimilation rate (A) was higher at full wateringthan at reduced watering, and the rate was halved as wateringwas reduced to 50% under low temperatures (Fig. 3a) and to~75% at high temperatures (Fig. 3b). Both cultivars had similarAat each of the three watering levels under low and hightemperature regimes (Fig. 3a, b).
There were differences between the two cultivars in theirstomatal conductance at 50% watering level (Fig. 3c).Whereas conductance was reduced by up to 50% whenwatering was reduced from 100 to 25% in the low-temperatureregime, the reduction was not significant in the high-temperatureregime. Reduced water supply reduced transpiration at low
–1.6
–1.2
–0.8
–0.4
0.0
BanaAtherton
–1.6
–1.2
–0.8
–0.4
0.0
25
50
100
LWP
(M
Pa)
Time of day (hours)
5 7 9 12 16 19 5 7 9 12 16 19
BanaAtherton
25
50
100
(a)
(c)
(b)
(d )
Fig. 2. Mean values (�s.e.) for diurnal trends for leaf water potential for Napier grass cvv. Bana andAtherton averaged for the three growth cycles. (a) Low-temperature glasshouse bay, averaged over all watersupply levels; (b) high-temperature glasshouse bay, averaged over all water supply levels; (c) low-temperature glasshouse bay, averaged over both cultivars; (d) high-temperature glasshouse bay,averaged over both cultivars.
1012 Crop & Pasture Science S. W. Mwendia et al.

temperatures but not at high temperatures (Fig. 3e, f ).Transpiration at full watering was lower under hightemperature than under low temperature. There was asignificant difference in WUE between the two cultivars at100% under high temperature (Fig. 3h), and generally WUEwas higher under the low than high temperature regime.
Growth variables, water use and water-use efficiency
Leaf area was reduced by reducing water supply under bothtemperature conditions, except for the first growth cycle, wheremost of the differenceswere not statistically significant (P > 0.05)(Table 3).The leaf area increased in both cultivarswith successivegrowth cycles, and both cultivars generally had similar leaf area,except at high temperatures and 100%, watering where Athertonsignificantly outperformed Bana. The leaf area for the thirdgrowth cycle was overall almost three times larger than for thefirst growth cycle.
Generally, more DM was produced under low temperaturethanhighacross thegrowth cycles.The cultivars produced similarDM, except under low temperature during the second growthcycle, where Bana had higher DM than Atherton at 50 and 100%watering (Fig. 4). Over the three growth cycles, DM increased by~25%with each successive growth cycle when water supply waskept at either 50 or 100% field capacity under both temperatureregimes.
Dry matter produced in the shoot and root over the threegrowth cycles was summed and expressed as yield per unit landarea (Table 4). The DM yield was mostly similar between thetwo cultivars and was reduced as water supply was lowered
under the two temperature regimes. The DM fell by 30–50%with each reduction in watering level. The DM produced underhigh temperatures was generally lower by 9–27% than at lowtemperatures. Under the low temperature conditions, the root/shoot ratiowas statistically similar for the two cultivars (P > 0.05)and was reduced by ~30% as water supply was reduced to 25 or50% (Table 4).
0.0
10.0
20.0
30.0
µmol
m–2
S–1
µmol
mm
ol–1
mol
m–2
S–1
mm
olm
–2 S
–1
0.0
0.1
0.2
0.02.04.06.08.0
0.0
25 50 100Ba At Ba At Ba At
25 50 100Ba At Ba At Ba At
2.0
4.0
6.0
Napier cultivar and watering level
(a)
(c)
(e)
(g)
(b)
(d)
(f)
(h)
Fig. 3. Mean values (�s.e.) for leaf processes in Napier grass cvv. Bana (Ba) and Atherton (At). Data are pooled formeasurements made during the three growth cycles at week 4 of growth (first cycle), weeks 4 and 7 (second cycle)and weeks 3 and 8 (third cycle). (a) Low-temperature glasshouse bay, CO2 exchange; (b) high-temperature glasshousebay, CO2 exchange; (c) low-temperature glasshouse bay, stomatal conductance; (d) high-temperature glasshouse bay,stomatal conductance; (e) low-temperature glasshouse bay, transpiration; ( f ) high-temperature glasshouse bay,transpiration; (g) low-temperature glasshouse bay, photosynthetic water-use efficiency; (h) high-temperatureglasshouse bay, photosynthetic water-use efficiency.
Table 3. Leaf area (m2 bin–1) for Napier grass subjected to differentwater supply levels under low and high temperature for three
growth cyclesWithin each temperature regime and growth cycle, means followed by the
same letter are not significantly different at P= 0.05
Water Growth Low temperature High temperaturesupply cycle Bana Atherton Bana Atherton
25% First 0.51ab 0.56ab 0.51a 0.49a50% 0.43b 0.76a 0.73a 0.80a100% 0.60ab 0.50ab 0.71a 0.78a
l.s.d. (P= 0.05) 0.26 0.34
25% Second 0.61c 0.53c 0.51c 0.48c50% 0.94b 0.94b 0.80bc 0.92bc100% 1.50a 1.27a 1.08b 1.72a
l.s.d. (P= 0.05) 0.29 0.47
25% Third 0.87b 0.82b 0.58d 0.59d50% 1.67a 1.48a 1.07c 1.34bc100% 1.54a 1.75a 1.61b 2.11a
l.s.d. (P= 0.05) 0.29 0.36
Napier performance under variable environments Crop & Pasture Science 1013
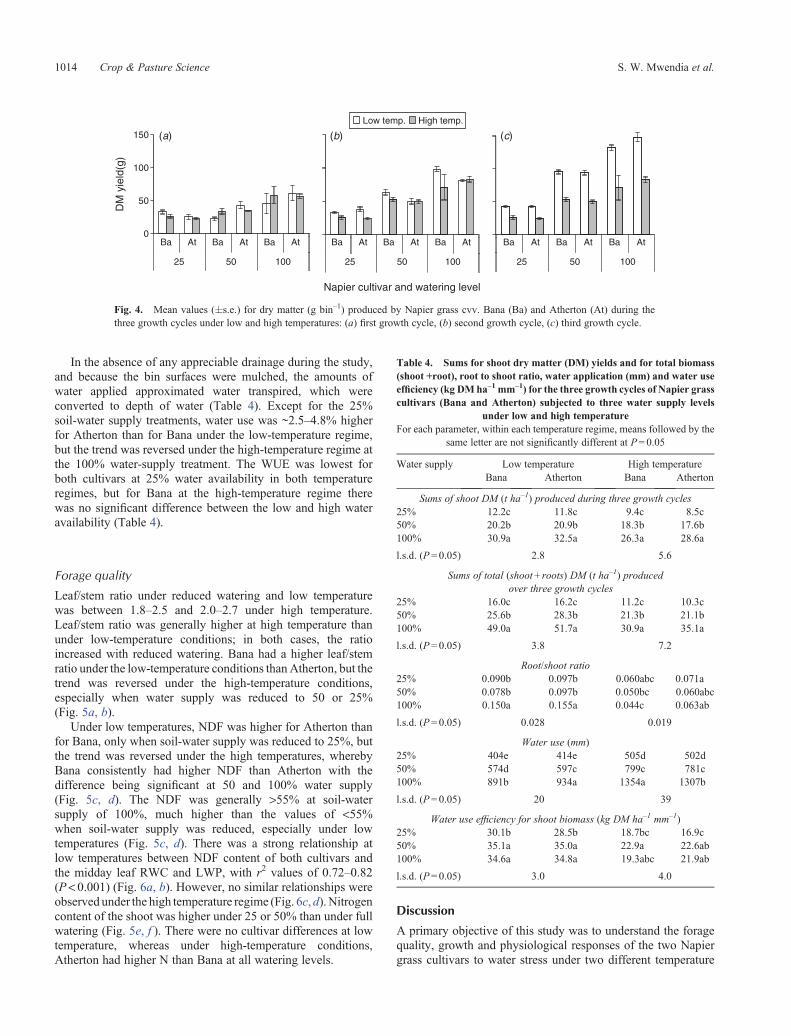
In the absence of any appreciable drainage during the study,and because the bin surfaces were mulched, the amounts ofwater applied approximated water transpired, which wereconverted to depth of water (Table 4). Except for the 25%soil-water supply treatments, water use was ~2.5–4.8% higherfor Atherton than for Bana under the low-temperature regime,but the trend was reversed under the high-temperature regime atthe 100% water-supply treatment. The WUE was lowest forboth cultivars at 25% water availability in both temperatureregimes, but for Bana at the high-temperature regime therewas no significant difference between the low and high wateravailability (Table 4).
Forage quality
Leaf/stem ratio under reduced watering and low temperaturewas between 1.8–2.5 and 2.0–2.7 under high temperature.Leaf/stem ratio was generally higher at high temperature thanunder low-temperature conditions; in both cases, the ratioincreased with reduced watering. Bana had a higher leaf/stemratio under the low-temperature conditions thanAtherton, but thetrend was reversed under the high-temperature conditions,especially when water supply was reduced to 50 or 25%(Fig. 5a, b).
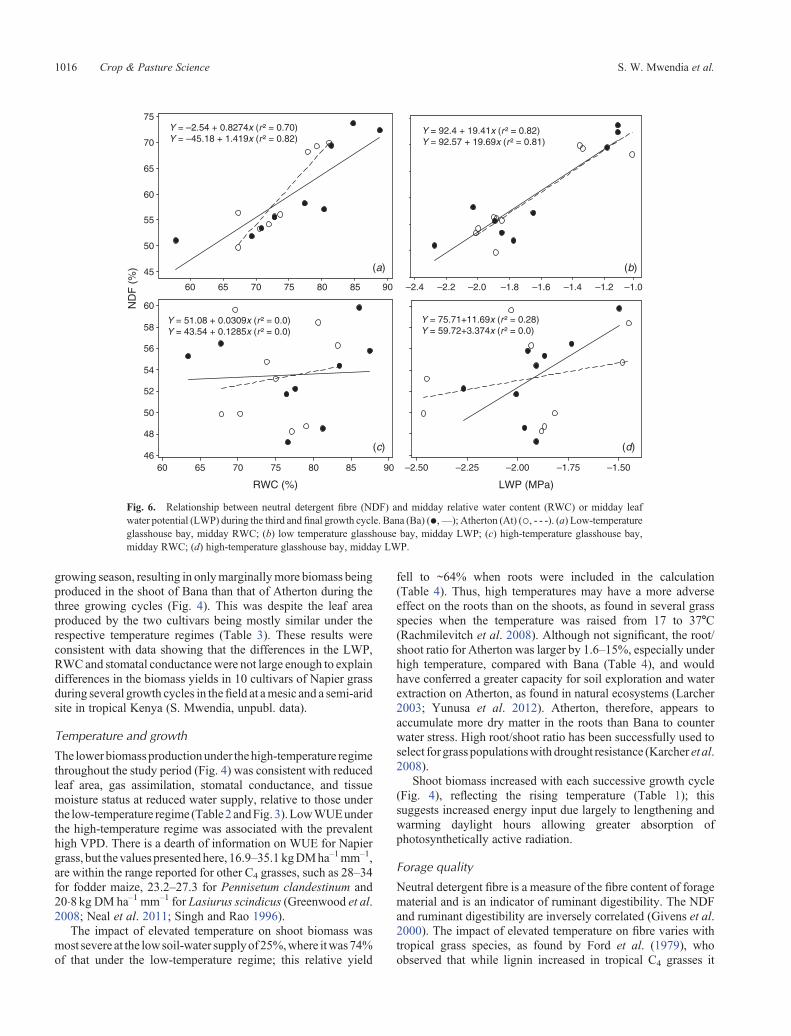
Under low temperatures, NDF was higher for Atherton thanfor Bana, only when soil-water supply was reduced to 25%, butthe trend was reversed under the high temperatures, wherebyBana consistently had higher NDF than Atherton with thedifference being significant at 50 and 100% water supply(Fig. 5c, d). The NDF was generally >55% at soil-watersupply of 100%, much higher than the values of <55%when soil-water supply was reduced, especially under lowtemperatures (Fig. 5c, d). There was a strong relationship atlow temperatures between NDF content of both cultivars andthe midday leaf RWC and LWP, with r2 values of 0.72–0.82(P < 0.001) (Fig. 6a, b). However, no similar relationships wereobserved under the high temperature regime (Fig. 6c,d).Nitrogencontent of the shoot was higher under 25 or 50% than under fullwatering (Fig. 5e, f ). There were no cultivar differences at lowtemperature, whereas under high-temperature conditions,Atherton had higher N than Bana at all watering levels.
Discussion
A primary objective of this study was to understand the foragequality, growth and physiological responses of the two Napiergrass cultivars to water stress under two different temperature
Napier cultivar and watering level
0
50
100
150 (a) (b) (c)
DM
yie
ld(g
)
25 50 100
Ba At Ba At Ba At
25 50 100
Ba At Ba At Ba At
25 50 100
Ba At Ba At Ba At
Low temp. High temp.
Fig. 4. Mean values (�s.e.) for dry matter (g bin–1) produced by Napier grass cvv. Bana (Ba) and Atherton (At) during thethree growth cycles under low and high temperatures: (a) first growth cycle, (b) second growth cycle, (c) third growth cycle.
Table 4. Sums for shoot dry matter (DM) yields and for total biomass(shoot +root), root to shoot ratio, water application (mm) and water useefficiency (kg DMha–1 mm–1) for the three growth cycles of Napier grasscultivars (Bana and Atherton) subjected to three water supply levels
under low and high temperatureFor each parameter, within each temperature regime, means followed by the
same letter are not significantly different at P= 0.05
Water supply Low temperature High temperatureBana Atherton Bana Atherton
Sums of shoot DM (t ha–1) produced during three growth cycles25% 12.2c 11.8c 9.4c 8.5c50% 20.2b 20.9b 18.3b 17.6b100% 30.9a 32.5a 26.3a 28.6a
l.s.d. (P= 0.05) 2.8 5.6
Sums of total (shoot+ roots) DM (t ha–1) producedover three growth cycles
25% 16.0c 16.2c 11.2c 10.3c50% 25.6b 28.3b 21.3b 21.1b100% 49.0a 51.7a 30.9a 35.1a
l.s.d. (P= 0.05) 3.8 7.2
Root/shoot ratio25% 0.090b 0.097b 0.060abc 0.071a50% 0.078b 0.097b 0.050bc 0.060abc100% 0.150a 0.155a 0.044c 0.063ab
l.s.d. (P= 0.05) 0.028 0.019
Water use (mm)25% 404e 414e 505d 502d50% 574d 597c 799c 781c100% 891b 934a 1354a 1307b
l.s.d. (P= 0.05) 20 39
Water use efficiency for shoot biomass (kg DM ha–1 mm–1)25% 30.1b 28.5b 18.7bc 16.9c50% 35.1a 35.0a 22.9a 22.6ab100% 34.6a 34.8a 19.3abc 21.9ab
l.s.d. (P= 0.05) 3.0 4.0
1014 Crop & Pasture Science S. W. Mwendia et al.

regimes. We found that under the low temperature regime at the25% soil-water level Atherton had higher LWP and RWC thanBana, but that trend was reversed at a water supply of either50% or 100% during the study period (Table 2). Thesedifferences in tissue water status did not produce consistentand significant differences in the dry matter yield by the twocultivars (Table 4). Under high temperature, the tissue waterstatus tended to be better for Bana than for Atherton but thedifferences were significant only between Bana at full watersupply and Atherton at 25% water supply. These resultssuggest that the two cultivars differed in their strategies formaintaining favourable tissue water status when soil-watersupply became highly limited and this impacted the quantityand quality of the forage produced, particularly under the lowtemperature regime, as explained in the following sections.
Water stress and growth
Differences in LWP or RWC between the two cultivars were notreflected in the trends in DM yields by the grasses. It is alsonoteworthy that anydecline in carbondioxide assimilation arisingfrom limited soil-water supply was marginal, even as LWPplunged to ~–3.0MPa (Table 5). This suggested that the waterstress was not severe enough even at 25%water supply to impairphotosynthesis. Although there is a lack of literature on Napiergrass in this regard, many tropical grasses can maintain carbonassimilation even at LWP as negative as –4.0MPa (Ludlow et al.
1985). A high stomatal conductance under reduced soil-watersupply can be sustained through other means such as osmoticadjustment to increase in-flow of water into the tissues (Zhanget al. 1999). Such a mechanism has been observed in sorghum,which lowered its osmotic potential by ~0.6MPa for every1.0MPa decline in LWP (Turner et al. 1978). The mostlysimilar RWC for the two cultivars across the three soil-watersupply regimes under the high temperature regime, despite thedifferences in their LWP, suggested that these Napier grasscultivars could have adjusted their osmotic potential tomaintain favourable tissue moisture at low soil-water supply.Thiswas in addition to the parsimony inwater use imposed by thereduced root/shoot ratio at lowwater supply (Table 4), whichwasquite different to the high root/shoot ratio at 100% water supplyand presumably unrestrained extraction and use of water asreported earlier by Snyman (2009).
Differences in the tissue moisture status between the twocultivars occurred only over a few hours around midday(Figs 1, 2), which was when the leaf stomata of Athertonclosed at low soil-water supply. This closurerestrained daytime stomatal conductance and thus reduced theopportunity for carbon assimilation in this cultivar. On theother hand, the stomata of Bana remained open with a higherrate of gas exchange at all soil-water supply levels under the lowtemperature regime and even at low water supply under the hightemperature regime (Fig. 3). This difference in instantaneouscarbon assimilation must have been maintained for much of the
45
50
55
60
65
70
ND
F (
%)
(c) (d)
0.0
1.0
2.0
3.0
4.0(a)
Leaf
: st
em r
atio (b)
0.0
1.0
2.0
3.0
4.0
5.0
Ba At Ba At Ba At
25 50 100
Ba At Ba At Ba At
25 50 100
(e)
N (
%)
Napier cultivar and watering level
(f)
Fig. 5. Mean values (�s.e.) for leaf to stem ratio, neutral detergent fibre (NDF, %) and nitrogen (N, %) inNapier grass cvv. Bana (Ba) and Atherton (At) at the three watering levels under low- and high-temperatureconditions averaged over the three growth cycles. (a) Leaf/stem ratio, low temperature glasshouse bay; (b) leaf/stemratio, high temperature glasshouse bay; (c) NDF, low temperature glasshouse bay; (d) NDF, high temperatureglasshouse bay; (e) N, low temperature glasshouse bay; (f) N, high temperature glasshouse bay.
Napier performance under variable environments Crop & Pasture Science 1015
growing season, resulting in onlymarginallymore biomass beingproduced in the shoot of Bana than that of Atherton during thethree growing cycles (Fig. 4). This was despite the leaf areaproduced by the two cultivars being mostly similar under therespective temperature regimes (Table 3). These results wereconsistent with data showing that the differences in the LWP,RWCand stomatal conductancewere not large enough to explaindifferences in the biomass yields in 10 cultivars of Napier grassduring several growth cycles in thefield at amesic and a semi-aridsite in tropical Kenya (S. Mwendia, unpubl. data).
Temperature and growth
The lowerbiomassproductionunder thehigh-temperature regimethroughout the study period (Fig. 4) was consistent with reducedleaf area, gas assimilation, stomatal conductance, and tissuemoisture status at reduced water supply, relative to those underthe low-temperature regime (Table2 andFig. 3).LowWUEunderthe high-temperature regime was associated with the prevalenthigh VPD. There is a dearth of information on WUE for Napiergrass, but the values presentedhere, 16.9–35.1 kgDMha–1mm–1,are within the range reported for other C4 grasses, such as 28–34for fodder maize, 23.2–27.3 for Pennisetum clandestinum and20�8 kg DM ha–1 mm–1 for Lasiurus scindicus (Greenwood et al.2008; Neal et al. 2011; Singh and Rao 1996).
The impact of elevated temperature on shoot biomass wasmost severe at the lowsoil-water supplyof 25%,where itwas74%of that under the low-temperature regime; this relative yield
fell to ~64% when roots were included in the calculation(Table 4). Thus, high temperatures may have a more adverseeffect on the roots than on the shoots, as found in several grassspecies when the temperature was raised from 17 to 378C(Rachmilevitch et al. 2008). Although not significant, the root/shoot ratio for Atherton was larger by 1.6–15%, especially underhigh temperature, compared with Bana (Table 4), and wouldhave conferred a greater capacity for soil exploration and waterextraction on Atherton, as found in natural ecosystems (Larcher2003; Yunusa et al. 2012). Atherton, therefore, appears toaccumulate more dry matter in the roots than Bana to counterwater stress. High root/shoot ratio has been successfully used toselect for grass populationswith drought resistance (Karcher et al.2008).
Shoot biomass increased with each successive growth cycle(Fig. 4), reflecting the rising temperature (Table 1); thissuggests increased energy input due largely to lengthening andwarming daylight hours allowing greater absorption ofphotosynthetically active radiation.
Forage quality
Neutral detergent fibre is a measure of the fibre content of foragematerial and is an indicator of ruminant digestibility. The NDFand ruminant digestibility are inversely correlated (Givens et al.2000). The impact of elevated temperature on fibre varies withtropical grass species, as found by Ford et al. (1979), whoobserved that while lignin increased in tropical C4 grasses it
90858075706560
75
70
65
60
55
50
45
Y = –2.54 + 0.8274x (r ² = 0.70)Y = –45.18 + 1.419x (r ² = 0.82)
–1.0–1.2–1.4–1.6–1.8–2.0–2.2–2.4
Y = 92.4 + 19.41x (r² = 0.82)Y = 92.57 + 19.69x (r² = 0.81)
90858075706560
60
58
56
54
52
50
48
46
RWC (%)
ND
F (
%)
Y = 51.08 + 0.0309x (r ² = 0.0)Y = 43.54 + 0.1285x (r ² = 0.0)
–1.50–1.75–2.00–2.25–2.50
LWP (MPa)
Y = 75.71+11.69x (r² = 0.28)Y = 59.72+3.374x (r² = 0.0)
(c) (d)
(a) (b)
Fig. 6. Relationship between neutral detergent fibre (NDF) and midday relative water content (RWC) or midday leafwater potential (LWP) during the third and final growth cycle. Bana (Ba) (*,—); Atherton (At) (**, - - -). (a) Low-temperatureglasshouse bay, midday RWC; (b) low temperature glasshouse bay, midday LWP; (c) high-temperature glasshouse bay,midday RWC; (d) high-temperature glasshouse bay, midday LWP.
1016 Crop & Pasture Science S. W. Mwendia et al.

decreased in C3 grasses when daytime temperature was raisedfrom 21 to 328C. In this study, NDF was lower under the high-temperature regime than under the low-temperature regime.Grasses exposed to drought conditions in the field have higherNDF and therefore lower ruminant digestibility (Willard andSchuster 1973). However, in this experiment, NDF was lowerfor plants exposed to reduced water supply averaged overthe three growth cycles under both temperature conditions(Fig. 5c, d). This could be the result of the high leaf/stem ratiowhen watering was reduced to 25%, especially at hightemperature (Fig. 5b). The NDF for the shoots was consistentwith observation in several cultivars of Napier grass by Tudsriet al. (2002).
Differences in NDF between the cultivars (Fig. 5c, d)suggested that, when based on forage quality, Bana would bepreferred because it would be less fibrous than Atherton whenwater supply is limited in mild environments–seasons, as may beexperienced during dry spells in the early or mid-season. Bycontrast, Atherton would provide a superior feed quality in dryand hot conditions, as often experienced in environments withsevere droughts. This is because the lowNDF for Atherton underthe hot conditions means that it accumulates low concentrationsof the mostly complex, poorly digestible polysaccharides and
lignin in the cell wall (Moore and Jung 2001). Despite thesecontrasting responses by the two grass cultivars, digestibility wasgenerally superior for both grasses at reduced soil-water supplyunder both temperature regimes (Fig. 5), because forage cropsreduce concentrations of cell wall fibres in their tissues whenwater-stressed, as found in grasses (Sheaffer et al. 1992; Akinet al. 1994; McCuistion et al. 2010) and legumes (Halim et al.1989).Halim et al. (1989) associated the increased digestibility inlucerne (Medicago sativa) with reduced cellulose concentrationbut increased hemicellulose in the cell wall of water-stressedplants. They also found increased leaf/stem ratiowithwater stressas presented here (Fig. 5). The NDF of both cultivars increasedsignificantly with tissue hydration, implying decreaseddigestibility, but only under the low temperature condition.The absence of significant correlation between NDF and tissuemoisture statusunder thehigh-temperature regime, suchas shownin Fig. 6c, d, suggested other physiological intervention, possiblyosmotic adjustment, in the expression of tissue water statusmentioned earlier.
Both Bana and Atherton used in the current study producedhigh leaf/stem ratios that were mostly >2 (Fig. 5a, b), which iscomparable with values reported by Tudsri et al. (2002) in therange of 1.6–3.1 for their five Napier grass cultivars. The high
Table 5. Minimum leaf relative water content and water potential observed at midday for Napier grass cultivarssubjected to three watering regimes under low and high temperature during three growth cycles
For each parameter, within each temperature regime, means followed by the same letter are not significantly differentat P= 0.05
Water supply Growth cycle and Low temperature High temperatureminimum value date Bana Atherton Bana Atherton
Relative water content (%)25% First, 29 July 2011 68.5b 84.8a 56.2abc 48.4c50% 92.6a 81.2ab 65.0ab 52.8bc100% 90.8a 85.1a 71.3a 57.0abc
l.s.d. (P= 0.05) 14.4 15.5
25% Second, 4 Sept. 2011 62.6bc 89.3a 67.5a 49.8a50% 85.1ab 56.8c 67.6a 63.0a100% 79.3abc 56.0c 62.3a 57.5a
l.s.d. (P= 0.05) 24.8 24.0
25% Third, 29 Oct. 2011 58.7bc 60.2ab 63.2a 64.3a50% 62.7ac 51.8b 58.9ab 61.3a100% 68.5a 52.5b 61.2a 47.9b
l.s.d. (P= 0.05) 8.5 12.9
Leaf water potential (MPa)25% First, 29 July 2011 –2.53b –1.54ab –3.25b –2.13ab50% –0.86a –1.35ab –1.48a –2.12ab100% –0.73a –1.68ab –1.42a –1.95a
l.s.d. (P= 0.05) 1.50 1.16
25% Second, 4 Sept. 2011 –2.28b –0.99a –2.82a –2.82a50% –1.53ab –2.53b –2.77a –2.90a100% –1.55ab –2.50b –1.87a –2.78a
l.s.d. (P= 0.05) 1.19 1.25
25% Third, 29 Oct. 2011 –2.46a –2.67a –2.48ab –2.27a50% –3.17a –2.94a –2.43ab –3.17b100% –2.47a –2.92a –2.03a –2.87b
l.s.d. (P= 0.05) 0.83 0.72
Napier performance under variable environments Crop & Pasture Science 1017

tissue N (2.5–4.0%) for the two cultivars of Napier grasspresented here were much higher than reported (0.9–2.3%) forthis grass species in the literature (Tudsri et al. 2002). Our highvalues were most probably the result of high nutrient availabilitythroughout this study because of topdressing with urea andabsence of leaching from the bins.
As ruminants prefer leaves to stems due to higher nutritionalvalue (Tudsri et al. 2002), the Atherton cultivar would be betterunder high temperature conditions because of its higher leaf/stem ratio,which is also supportedbyhigherNcontent (Fig. 5e, f )and only marginally higher root/shoot ratio.
Summary and conclusion
This study shows that an apparent favourable tissue waterstatus, as shown by Bana over Atherton, does not necessarilyconfer an advantage in forage quality, carbon assimilation andDMaccumulation.Despite its relatively poorer tissuewater statusat low soil water supply, Bana showed a tendency to producemarginally more biomass than Atherton. This was because Bana,unlike Atherton, kept its stomata open to maintain carbonassimilation even when its tissue moisture conditions declinedto well below those of Atherton. As mentioned above, fieldmeasurements also failed to produce significant correlationsbetween tissue moisture status and stomatal conductance, onthe one hand, and biomass yield, on the other hand, for manycultivars tested in a relatively dry year in a tropical environment(S. Mwendia, unpubl. data). However, digestibility declinedwith increased tissue hydration under low temperature assuggested by the increase in NDF. Determining tissue waterstatus may, therefore, not always be an adequate, or evena reliable, predictor of the productivity and quality potential ofa plant under limiting soil-water supply conditions. Its usefulnessfor assessing quality seems dependent on the temperature.Measurements of attributes such as osmotic potential mayhave been beneficial in this study to help understand themechanism in the two cultivars, especially how the Banastomata remain open so that transpiration and photosynthesisrates do not decrease despite the low tissue water status. Therewas evidence of functional convergence between the twocultivars at full or 50% soil water supply, such that despite thedifferences in the leaf area and physiological characteristics(RWC, LWP, stomatal conductance and A) the cultivarsproduced similar DM yields over the three growth cyclesunder the two temperature regimes.
The relative responses of these two cultivars of Napier grassto water stress under different temperature conditions wascomplex with respect to the range of attributes measured inthis glasshouse study. We conclude that simple attributes suchas changes in tissue hydration, specifically LWP and RWC, or A,in response to water temperature stress differences may notalways be reliable indicators of the relative forage quality andyield. This is because, as we show here, gas exchange andbiomass accumulation can be sacrificed in order to maintainfavourable tissue hydration. Further studies, therefore, shouldassess osmotic potential to close the tissue water balance for awider range of cultivars of this grass species to improve ourunderstanding of their physiological responses to environmentalstresses, particularly under natural conditions.
Acknowledgements
This work was supported by funding from the Australian Governmentthrough the Australian Agency for International Development. AssistancefromDr NigelWarick,Michael Faint and all technical staff in the Departmentof Agronomy and Soil Science, University of New England, Australia, ishighly appreciated. We also appreciate the helpful comments from theanonymous reviewers.
References
Akin DE, Kimball BA, Mauney JR, LaMorte RL, Hendrey GR, Lewin K,Nagy J, Gates RN (1994) Influence of enhanced CO2 concentrationand irrigation on sudangrass digestibility. Agricultural and ForestMeteorology 70, 279–287. doi:10.1016/0168-1923(94)90063-9
AnindoDO, PotterHL (1994) Seasonal variation in productivity and nutritivevalue of Napier grass at Muguga, Kenya. East Africa Agriculture andForestry Journal 59, 177–185.
AOAC (1975) ‘Official methods of analysis.’ 12th edn (Association ofOfficial Agricultural Chemists: Washington, DC)
Barrs HD (1968) Determination of water deficits in plant tissue. In ‘Waterdeficit and plant growth’. (Ed. TT Kozlowski) pp. 235–368. (AcademicPress: New York)
Esilaba AO, Muturi GM, Cheruiyot HK, Okoti M, Nyariki DM, Keya GA,Miriti JM, Kigomo JN, Olukoye G, Wekesa L (2011) The desert marginsprogramme approaches in upscaling best-bet technologies in arid andsemi-arid lands inKenya. In ‘Innovations as key to the green revolution inAfrica’. (Eds A Bationo, BWaswa, J Okeyo, FMMaina) pp. 1177–1191.(Springer Science: New York)
Fernández RJ, Reynolds JF (2000) Potential growth and drought toleranceof eight desert grasses: lack of trade off? Oeocologia 123, 90–98.doi:10.1007/s004420050993
Ford CW, Morrison IM, Wilson JR (1979) Temperature effects on lignin,hemicellulose and cellulose in tropical and temperate grasses. AustralianJournal of Agricultural Research 30, 621–633. doi:10.1071/AR9790621
Givens DI, Owen E, Axford RFE, Omed HM (2000) ‘Forage evaluation inruminant nutrition.’ (CABI Publishing: Wallingford, UK)
Greenwood KL, Mundy GN, Kelly KB (2008) On-farm measurement of thewater use and productivity of maize. Australian Journal of ExperimentalAgriculture 48, 274–284. doi:10.1071/EA06094
Guenni O, Marin D, Baruch Z (2002) Responses to drought of fiveBrachiaria species. Biomass production, leaf growth, root distribution,water use and forage quality. Plant and Soil 243, 229–241. doi:10.1023/A:1019956719475
Halim HR, Buxton DR, Hattendorf MJ, Carlson RE (1989) Water-stresseffects on alfalfa forage quality after adjustment for maturity differences.Agronomy Journal 81, 189–194. doi:10.2134/agronj1989.00021962008100020010x
Hatch MD (1992) ‘The making of the C4 pathway.’ (Kluwer AcademicPublishers: Dordrecht, The Netherlands)
Karcher DE, Richard MD, Hignight K, Rush D (2008) Drought tolerance oftall fescue populations selected for high root/shoot ratios and summersurvival. Crop Science 48, 771–777. doi:10.2135/cropsci2007.05.0272
Klingebiel AA, Montgomery PH (1961) ‘Land capability classification.’Agriculture Handbook No. 210. (US Department of Agriculture:Washington, DC)
Lambers H, Chapin FS, Pins TL (2008) ‘Plant physiological ecology.’(Springer Science: New York)
Larcher W (2003) ‘Physiological plant ecology.’ (Springer-Verlag: Berlin,Heidelberg)
Ludlow MM, Fisher MJ, Wilson JR (1985) Stomatal adjustment to waterdeficits in three tropical grasses and a tropical legume in controlledconditions and in the field. Journal of Plant Physiology 12, 131–149.
McCuistionKC, BeanBW,McCollumFT III (2010)Nutritional compositionresponse to yield differences in brown midrib, non-brown midrib, and
1018 Crop & Pasture Science S. W. Mwendia et al.

photoperiod sensitive forage sorghum cultivars. Forage andGrazinglands, doi:10.1094/FG-2010-0428-01-RS
Minitab Inc (2007) ‘Minitab statistical software, release 15 for windows.’(Minitab Inc.: State College, PA, USA)
Monteith JL, Unsworth MH (2008) ‘Principles of environmental physics.’3rd edn (Elsevier Science: Amsterdam)
Moore KJ, Jung HG (2001) Lignin and fiber digestion. Journal of RangeManagement 54, 420–430. doi:10.2307/4003113
Neal JS, FulkersonWJ, Sutton BG (2011)Differences in water-use efficiencyamong perennial forages used by the dairy industry under optimum anddeficit irrigation. Irrigation Science 29, 213–232.
Neil KL, Tiller RL, Faeth SH (2003) Big sacaton and endophyte-infectedArizona fescue germination under water stress. Journal of RangeManagement 56, 616–622. doi:10.2307/4003936
Niklas KJ, Spats H (2012) Plant water relations. In ‘Plant physics’. (Eds MKoplow, NS Roche) pp. 71–97. (The University of Chicago Press:Chicago, IL)
Norris IB, Thomas H (1982) The effect of droughting on varieties andecotypes of Lolium, Dactylis and Festuca. Journal of Applied Ecology19, 881–889. doi:10.2307/2403290
NyaataOZ,Dorward PT,Keatinge JDH,O’NeillMK (2000)Availability anduse of dry season feed resources on smallholder dairy farms in centralKenya. Agroforestry Systems 50, 315–331. doi:10.1023/A:1006447915074
Nyambati EM, Lusweti CM, Muyekho FN, Mureithi JG (2011) Up-scalingNapier grass (Pennisetum purpureum Schum.) production using“Tumbukiza” method in smallholder farming systems in north westernKenya. Journal of Agricultural Extension and Rural Development 3, 1–7.
Qadir M, Noble AD, Chartres C (2013) Adapting to climate change byimproving water productivity of soils in dry areas. Land Degradation& Development 24, 12–21. doi:10.1002/ldr.1091
Rachmilevitch S, Lambers H, Huang B (2008) Short-term and long-term rootrespiratory acclimation to elevated temperatures associated with rootthermotolerance for two Agrostis grass species. Journal ofExperimental Botany 59, 3803–3809. doi:10.1093/jxb/ern233
Scholander PE, Hammel HT, Bradstreet ED, Hemmingsen EA (1965) Sappressure in vascular plants. Science 148, 339–346. doi:10.1126/science.148.3668.339
Sheaffer CC, Petersen PR, Hall MH, Stordahl JB (1992) Drought effectson yield and quality of perennial grasses in the North Central UnitedStates. Journal of Production Agriculture 5, 556–561. doi:10.2134/jpa1996.0556
Singh KC, Rao AS (1996) Short note on water use and production potentialofLasiurus scindicusHenr. in the Thar desert of Rajasthan, India. Journalof Arid Environments 33, 261–262. doi:10.1006/jare.1996.0062
Snyman HA (2009) Root studies on grass species in a semi-arid South Africaalong a soil-water gradient.Agriculture, Ecosystems&Environment 131,247–254. doi:10.1016/j.agee.2009.01.023
Staal SJ, Chege L, Kenyanjui M, Kimari A, Lukuyu B, Njubi D, OwangoM,Tanner J, Thorpe W, Wambugu M (1998) Characterisation of dairysystems supplying the Nairobi milk market: A pilot survey in Kiambudistrict for the identification of target producers. KARI/MoA/ILRICollaborative Research Project Report. Available at: www.reading.ac.uk/ssc/media/ILRI_2006Nov/Publication/Full%20Text/Staal.pdf(accessed 14 August 2013).
Tudsri S, Jorgensen ST, Riddach P, Pookpakdi A (2002) Effects of cuttingheight and dry season closing date on yield and quality of five Napiergrass cultivars in Thailand. Tropical Grasslands 36, 248–252.
Turner NC, Begg JE, TonnetML (1978) Osmotic adjustment of sorghum andsunflower crops in response towater deficits and its influence on thewaterpotential at which stomata close. Australian Journal of Plant Physiology5, 597–608. doi:10.1071/PP9780597
Volaire F, Thomas H (1995) Effects of drought on water relations, mineraluptake, water-soluble carbohydrate accumulation and survival of twocontrasting populations of cocksfoot (Dactylis glomerata L.). Annals ofBotany 75, 513–524. doi:10.1006/anbo.1995.1053
Wahid A, Close TJ (2007) Expression of dehydrins under heat stress andtheir relationship with water relations of sugarcane leaves. BiologiaPlantarum 51, 104–109. doi:10.1007/s10535-007-0021-0
Willard EE, Schuster JL (1973) Chemical composition of six Southern GreatPlains grasses as related to season and precipitation. Journal of RangeManagement 26, 37–38. doi:10.2307/3896881
Wilson JR (1983)Effects ofwater stress on in vitrodrymatter digestibility andchemical composition of herbage of tropical pasture species. AustralianJournal of Agricultural Research 34, 377–390. doi:10.1071/AR9830377
Yordanov I, Velikova V, Tsonev T (2000) Plant responses to drought,acclimation, and stress tolerance.Photosynthetica38, 171–186. doi:10.1023/A:1007201411474
Yunusa IAM, Thomson SE, Pollock KP, Youwei L, Mead DJ (2005) Waterpotential and gas exchange did not reflect performance of Pinus radiataD. Don in an agroforestry system under conditions of soil-water deficit ina temperate environment. Plant and Soil 275, 195–206. doi:10.1007/s11104-005-1481-7
Yunusa IAM, Zolfaghar S, Zeppel MJB, Li Z, Palmer AR, Eamus D (2012)Fine root biomass and its relationship to evapotranspiration in woody andgrassy vegetation covers for ecological restoration of waste storage andmining landscapes. Ecosystems 15, 113–127. doi:10.1007/s10021-011-9496-9
Zhang J, Nguyen HT, BlumA (1999) Genetic analysis of osmotic adjustmentin crop plants. Journal of Experimental Botany 50, 291–301.
ZhouY, Lambrides C, Kearns R, Ye CR, CaoN, Fukai S (2009) Selecting fordrought tolerance amongAustralian green couch grasses (Cynodon spp.).Crop & Pasture Science 60, 1175–1183. doi:10.1071/CP09172
Napier performance under variable environments Crop & Pasture Science 1019
www.publish.csiro.au/journals/cp


















![Diversity of wild and cultivated pearl millet accessions ( Pennisetum glaucum [L.] R. Br.) in Niger assessed by microsatellite markers](https://static.fdokumen.com/doc/165x107/631521c7fc260b71020fdba0/diversity-of-wild-and-cultivated-pearl-millet-accessions-pennisetum-glaucum-l.jpg)

