Unique and overlapping expression patterns of Arabidopsis CYP85 genes involved in brassinosteroid...
12
Unique and overlapping expression patterns of Arabidopsis CYP85 genes involved in brassinosteroid C-6 oxidation Julie Castle 1 , Miklos Szekeres 2 , Gwen Jenkins 1 and Gerard J. Bishop 1,3, * 1 Institute of Biological Sciences, University of Wales Aberystwyth, SY23 3DD, UK; 2 Institute of Plant Biology, Biological Research Centre of the Hungarian Academy of Sciences, P.O. Box 521, H-6701 Szeged, Hungary; 3 Department of Agricultural Sciences, Imperial College London, Wye Campus, Kent TN25 5AH, UK (*author for correspondence: e-mail [email protected]) Received 10 September; accepted in revised form 29 November 2004 Key words: brassinosteroids, CYP85, differential expression, reporter genes, steroid hormone synthesis Abstract Brassinosteroids (BRs) are steroid hormones that are essential for plant growth and development. To gain insight into potential sites of BR synthesis, we studied promoter activities of the two Arabidopsis BR C-6 oxidase genes (CYP85A1 and CYP85A2) in transgenic plants carrying promoter fusions with the GUS, GFP or LUC reporter genes. BR-dependent feedback regulation of the GUS reporter constructs indicated that their expression corresponded to those of the native genes. Both the CYP85A1 and CYP85A2 pro- moters showed maximum activity during the first week following germination, particularly in the vascular tissues. Compared to CYP85A2, CYP85A1 expression was weaker and confined to the early stages of seedling development. Stronger CYP85A2 promoter activity was evident in both juvenile and adult plants. Comparison of the 5 0 -UTR and TATA box sequences of CYP85A1 and CYP85A2 revealed high homology, indicating a relatively recent gene duplication. We also found that transgenic Arabidopsis plants har- bouring the tomato DWARF promoter-GUS fusion had similarities in the expression pattern to the Arabidopsis genes suggesting common transcriptional regulation of CYP85 genes in the two species. Abbreviations: BL, brassinolide; BR, brassinosteroid; CYP, cytochrome P450; GA, gibberellin Introduction Brassinosteroids (BRs) are steroidal plant hor- mones that play a significant role in developmental processes. When exogenously applied to plants in nanomolar concentrations they elicit many physi- ological responses, such as the induction of cell elongation, pollen tube growth, leaf bending/ expansion, ethylene biosynthesis, xylem differen- tiation, enhancement of stress resistance and de- layed senescence (Mandava, 1988; Clouse and Sasse, 1998). BR biosynthesis or perception mu- tants display a dramatic dwarf phenotype, dark green leaves, reduced fertility, an increased life span and abnormal skotomorphogenesis (Clouse and Feldman, 1999; Schumacher and Chory, 2000; Bishop and Koncz, 2002). The dwarf phenotype of BR-deficient mutants can be restored to wild type by exogenous application of brassinolide (BL), the most active BR. However, this is not possible in the case of BR-insensitive mutants. Mutants deficient in BR biosynthesis or per- ception have provided important information to substantiate the significance of BRs in plant development (Yokota, 1997; Li and Chory, 1999; Bishop and Yokota, 2001). Analysis of these mu- tants has enabled rapid progress in the under- standing of BR regulation and the elucidation of the biosynthetic pathway leading to the produc- tion of BL. Initially it was proposed that BL was Plant Molecular Biology (2005) 57:129–140 Ó Springer 2005 DOI 10.1007/s11103-004-6851-7
Transcript of Unique and overlapping expression patterns of Arabidopsis CYP85 genes involved in brassinosteroid...

Unique and overlapping expression patterns of Arabidopsis CYP85genes involved in brassinosteroid C-6 oxidation
Julie Castle1, Miklos Szekeres2, Gwen Jenkins1 and Gerard J. Bishop1,3,*1 Institute of Biological Sciences, University of Wales Aberystwyth, SY23 3DD, UK; 2Institute of PlantBiology, Biological Research Centre of the Hungarian Academy of Sciences, P.O. Box 521, H-6701 Szeged,Hungary; 3 Department of Agricultural Sciences, Imperial College London, Wye Campus, Kent TN25 5AH,UK (*author for correspondence: e-mail [email protected])
Received 10 September; accepted in revised form 29 November 2004
Key words: brassinosteroids, CYP85, differential expression, reporter genes, steroid hormone synthesis
Abstract
Brassinosteroids (BRs) are steroid hormones that are essential for plant growth and development. To gaininsight into potential sites of BR synthesis, we studied promoter activities of the two Arabidopsis BR C-6oxidase genes (CYP85A1 and CYP85A2) in transgenic plants carrying promoter fusions with the GUS,GFP or LUC reporter genes. BR-dependent feedback regulation of the GUS reporter constructs indicatedthat their expression corresponded to those of the native genes. Both the CYP85A1 and CYP85A2 pro-moters showed maximum activity during the first week following germination, particularly in the vasculartissues. Compared to CYP85A2, CYP85A1 expression was weaker and confined to the early stages ofseedling development. Stronger CYP85A2 promoter activity was evident in both juvenile and adult plants.Comparison of the 50-UTR and TATA box sequences of CYP85A1 and CYP85A2 revealed high homology,indicating a relatively recent gene duplication. We also found that transgenic Arabidopsis plants har-bouring the tomato DWARF promoter-GUS fusion had similarities in the expression pattern to theArabidopsis genes suggesting common transcriptional regulation of CYP85 genes in the two species.
Abbreviations: BL, brassinolide; BR, brassinosteroid; CYP, cytochrome P450; GA, gibberellin
Introduction
Brassinosteroids (BRs) are steroidal plant hor-mones that play a significant role in developmentalprocesses. When exogenously applied to plants innanomolar concentrations they elicit many physi-ological responses, such as the induction of cellelongation, pollen tube growth, leaf bending/expansion, ethylene biosynthesis, xylem differen-tiation, enhancement of stress resistance and de-layed senescence (Mandava, 1988; Clouse andSasse, 1998). BR biosynthesis or perception mu-tants display a dramatic dwarf phenotype, darkgreen leaves, reduced fertility, an increased lifespan and abnormal skotomorphogenesis (Clouse
and Feldman, 1999; Schumacher and Chory, 2000;Bishop and Koncz, 2002). The dwarf phenotype ofBR-deficient mutants can be restored to wild typeby exogenous application of brassinolide (BL), themost active BR. However, this is not possible inthe case of BR-insensitive mutants.
Mutants deficient in BR biosynthesis or per-ception have provided important information tosubstantiate the significance of BRs in plantdevelopment (Yokota, 1997; Li and Chory, 1999;Bishop and Yokota, 2001). Analysis of these mu-tants has enabled rapid progress in the under-standing of BR regulation and the elucidation ofthe biosynthetic pathway leading to the produc-tion of BL. Initially it was proposed that BL was
Plant Molecular Biology (2005) 57:129–140 � Springer 2005DOI 10.1007/s11103-004-6851-7

synthesized via two parallel routes, namely theearly and late C-6 oxidation pathways. However,more recently it has been suggested that BR bio-synthesis may occur via a metabolic grid ratherthan two independent pathways (Shimada et al.,2001). In tomato only the early C-6 oxidation hasbeen demonstrated (Bishop et al., 1999), whereasin Arabidopsis both pathways are functional(Noguchi et al., 2000).
Arguably, one of the most critical steps in theBR pathway is the conversion of 6-deoxocastas-terone (6-deoxoCS) to castasterone (CS), a pen-ultimate step in the biosynthesis of BL. This isespecially significant in tomato where CS isthought to be the main active BR (Bishop et al.,1999). C-6 oxidation is believed to be a rate lim-iting step because of the high accumulation of6-deoxoCS, thus suggesting a key role of C-6 oxi-dase genes in the regulation of BR levels (Shimadaet al., 2003). Dwarf was the first BR C-6 oxidasegene to be identified and characterized (Bishopet al., 1996, 1999). Dwarf was shown to encode acytochrome P450 (CYP), similar to all other BRbiosynthesis genes identified so far, with theexception of DEETIOLATED 2 (DET2) that en-codes a reductase (Li et al., 1996). Recently furtherC-6 oxidase mutants have been reported in rice,clearly demonstrating the critical role of this en-zyme in plant development. These BR-deficientdwarf (brd-1) mutants display a range of abnor-malities in growth and organ development, par-ticularly in stems and leaves (Hong et al., 2002;Mori et al., 2002). In tomato and rice current dataindicate that the mutations causing CYP85 defi-ciency affect a single locus, whereas in Arabidop-sis two homologous genes to DWARF, namelyCYP85A1 and CYP85A2, have been identified.Conversion assays with yeast-expressed Arabidop-sis CYP85As demonstrated that they both cat-alyzed C-6 oxidation (Shimada et al., 2001, 2003),suggesting that this function is redundant in thisspecies. This may be the reason why there are noreports of Arabidopsis CYP85 mutants with acharacteristic BR-biosynthetic dwarf phenotype,whereas in tomato and rice mutations in this genehave a severe phenotypic effect.
To gain better understanding of the role BRsplay in plant development, it is important to dis-cern sites of BR biosynthesis. As the C-6 oxidasescatalyze a penultimate step in the BR biosyntheticpathway, it seems reasonable to assume that the
sites of expression coincide with those of bioactiveBR synthesis. A corresponding paradigm has beenshown with investigations into giberellin (GA)biosynthesis. GA 20-oxidases and 3b-hydroxylasesare enzymes involved in the latter stages ofGA biosynthesis (Ait-Ali et al., 1999). Promoter:b-glucuronidase (GUS) reporter expression ofthese GA biosynthesis genes during early plantdevelopment has revealed information on the sitesof bioactive GA production (Kaneko et al., 2003).Good correlation has been shown between thesites of bioactive GA synthesis and the sites of GAsignalling. It is interesting to note that both theGA 20-oxidase and 3b-hydroxylase enzymes areencoded by more than one gene, which are differ-entially expressed in different tissues and underdifferent conditions (Yamaguchi et al., 2001;Ashikari et al., 2002).
Little is currently known concerning theexpression of BR biosynthesis genes. To gain in-sight into the transcriptional regulation of Ara-bidopsis C-6 oxidases we have investigated thetemporal and spatial expression patterns ofCYP85A1 and CYP85A2, using promoter:reportergene fusion constructs. To verify the consistency ofdata, we used three different reporter genes,namely GUS, GFP (green fluorescent protein), andLUC (firefly luciferase). This approach was used toenhance our knowledge with respect to the cellularlocalization and relative levels of expression. Ourresults support previous microarray, RT-PCR andreal time PCR data showing differential expres-sion between the two Arabidopsis CYP85 genes(Bancos et al., 2002; Shimada et al., 2003), whichindicates reliability of the data obtained. We alsoshow that the organ-specific activity of CYP85promoters correlates with expected sites of BRsynthesis and activity. Our results, therefore, pro-vide valuable information about the transcrip-tional activity at these loci during various stages ofdevelopment.
Materials and methods
Plant material and growth conditions
Seeds from Arabidopsis thaliana ecotype Columbia(Col-0) and its transgenic derivatives were surfacesterilized and grown on half strength MS medium(Murashige and Skoog, 1962) supplemented with
130

1% sucrose and 0.8% agar and, if required, 1 lMBL (CIDtech Research Inc., Missisauga, Ontario,Canada), in 12-hour-light (60–100 lmol/m2/sec)/12-hour-dark cycles at 20 �C. Alternatively, seedswere sown on Levingtons Universal potting com-post in netted pots and grown to the floweringstage in a shaded greenhouse under 8-hour-light/16-hour-dark regime.
Generation and analysis of transgenic plants
Upstream regulatory sequences of 1.8 kb includingthe promoter and the complete 50-UTR of bothCYP85A1 and CYP85A2, were PCR amplifiedfrom genomic Arabidopsis DNA. The PCRprimers used were as follows: CYP85A150-GTCCTAGTCGACGATGCCAC and 50-CTCGAACCGTGAACCGCAGCC; CYP85A250-CTACGGATCACGCGATCTCC and 50-CCGCAGCTATGTTGCATGTCC. The PCRproducts, with artificially generated NcoI sites atthe ATG start codons, were cloned into pGEMT-Easy Vector (Promega) and sequenced to makesure that they were free of PCR errors. For GUSfusions the promoter was excised by SpeI andNcoI, then cloned directly into XbaI and NcoI-cleaved pCambia 1301 vector (Centre for theApplication of Molecular Biology to InternationalAgriculture – CAMBIA). GFP5 minus the ERtarget sequence (von Arnim et al., 1998) was fusedto CYP85A1 and CYP85A2 promoters in a similarmanner, i.e. an engineered NcoI site at the startcodon and directly cloned into pCambia 1300(CAMBIA). The firefly luciferase reporterconstruct was generated by fusing the CYP85A2promoter with the firefly luciferase-derivedLUC+ (Promega), and then inserting the chimericgene in the T-DNA-based pPCV812 vector(Koncz et al., 1994) from which GUS had beenremoved. The tomato Dwarf promoter GUS fusionconstruct was generated in a similar manner, withan NcoI site at ATG (further details availableon request). The plant vector constructs weretransferred/conjugated to Agrobacterium tumefic-iens GV3101 and used for flower dip transforma-tion of Arabidopsis (Clough and Bent, 1998).Transgenic plants were selected on MS mediumcontaining 15 mg l)1 hygromycin. GUS, GFP andLUC activity was tested in 10 independent linessegregating 3:1, and seeds from the selfed T2
generation were bulked and used for RNA isola-tion and reporter assays.
Confocal microscopy
Seedlings were grown aseptically to the appropri-ate stage of development. Harvested seedlings wereincubated in 1 lM propidium iodide for 1 h priorto examination under the confocal microscope(BioRad MRC 1024 ES Confocal Laser ScanningMicroscope attached to a Zeiss Axiovert 135).Samples were analysed using an excitation wave-length of 488 nm in combination with a 515 +/)30 nm emission filter. Background levels of fluo-rescence were estimated using untransformed sam-ples and the average background level of expressionwas subtracted from the samples analyzed.
Histochemical and fluorimetric analysisof GUS activity
For GUS histochemical analysis, seedlings weregrown for 2, 4, 7, 11, 15 and 21 days and stainedovernight at 37 �C in 1 mg ml)1 X-gluc in100 mM pH 7.0 sodium phosphate buffer con-taining 5 mM potassium ferrocyanide, 5 mMpotassium ferricyanide and 1 mM EDTA. Stain-ing was examined following the removal of plantpigments by multiple washes with 70% ethanol.For quantitative GUS fluorimetric assays, pooledfresh seedlings were ground in liquid nitrogen andsuspended in GUS extraction buffer: 100 mMsodium phosphate buffer (pH 7.0), containing1 mM EDTA, 0.1% (v/v) Triton X-100 and10 mM b-mercaptoethanol. Spectrofluorimetricassays for GUS activity were performed using4-methylumbelliferyl-b-DD-glucuronide substrate(Sigma), as described in Jefferson et al. (1987). Toobserve the effect of BL treatment, seedlings weregerminated and grown for 7 days in the presenceor absence of 1 lM BL prior to the GUS fluori-metric assays. Wild-type and CPD:GUS trans-genic seedlings (Mathur et al., 1998) were used asnegative and positive controls, respectively. Totalprotein content of the samples was determined byBradford’s method (Bradford, 1976).
Luciferase imaging
Bioluminescence resulting from firefly (Photynuspyralis) luciferase activity was detected by a liquid
131

nitrogen-cooled back-illuminated digital CCDcamera (Princeton Instruments, Trenton, NJ).Plant material was sprayed with a solution of25 mM DD-luciferin (Biosynth A.G., Staad, Swit-zerland) with 0.01% (v/v) Trion X-100 in 5 mMTris-phosphate buffer (pH 8.0), then placed in thedark for 1 h. The distribution of bioluminescencewas detected by 25 min exposures with the CCDcamera using Metamorph imaging software (MetaSeries 4.5, Universal Imaging Co., West Chester,PA).
Transcript analysis
Total RNA was isolated from 100 mg fresh plantmaterial, using Total RNA Isolation Reagentfrom ABgene (Epsom, UK). First strand cDNAwas synthesized using 5 lg total RNA and oligo-dT primer. Semi quantitative RT-PCR assayswere carried out as described in Montoya et al.(2002). The CYP85-specific primers were thoseused by Bancos et al. (2002). GFP primers: 50-CAAGTTGGAATACAACTACAACTC and50-ACTCTAGATAGATCTGTATAGTTCATCC.GUS primers: 50-GTGGATACGTTAGCCGG-GCTG and 50-GGAAATTCGAGCTGGTCAC-CTG. PCR amplification products were analyzedafter 25, 30, 35, 40 and 45 cycles on 2%(w/v) agarosegels with ethidium bromide staining and photo-graphed using Gene-snap (Syngene, CambridgeUK). The optimum cycle number determinedwas 35for both CYP85A1 and CYP85A2. RT-PCR datawere evaluated using Gene Tools manual quantifi-cation (Syngene).
Results
Temporal expression pattern of CYP85 genes
It has been demonstrated that transcript levels ofCYP85A1 and CYP85A2 are differentially accu-mulated (Bancos et al., 2002; Shimada et al.,2003). However, the temporal and spatial expres-sion has not yet been clearly defined. In order toenhance our understanding of the differentialexpression pattern of these two genes duringdevelopment, transgenic Arabidopsis plants weregenerated that contained the respective CYP85promoters fused to reporter genes. Five maingroups of transgenic seedlings were generated,
containing CYP85A1:GUS, CYP85A1:GFP,CYP85A2:GUS, CYP85A2:GFP and CYP85A2:-LUC, respectively.
Histochemical GUS analysis of developingseedlings clearly emphasized the different pro-moter activities of CYP85A1 and CYP85A2 dur-ing the first 21 days of growth. CYP85A1promoter-driven expression was mainly seen in thehypocotyls and roots until day 4 (Figure 1A).Close examination, particularly of 2-day-oldseedlings, showed GUS activity especially in thevascular tissue of the root and hypocotyl. By day 7the expression level was low, with patches of GUSexpression in root tissues and the shoot apicalmeristem. Even in early development, expressionwas not detected in the cotyledons. In contrast,CYP85A2 expression was mainly seen in the cot-yledons, hypocotyl and roots (Figure 1B). GUSactivity remained evident when plants were ana-lyzed after 21 days. However, CYP85A1:GUSactivity was not detectable approximately oneweek after germination. These data indicated thatboth genes were being transcribed in germinatingseeds and young seedlings. This activity reduced asthe seedlings matured and a higher level ofexpression occurred in CYP85A2:GUS lines thanin CYP85A1:GUS lines.
Intriguingly, histochemical analysis of GUSactivity revealed that during the reproductivegrowth stage there is also differential regulation ofthe two CYP85 promoters. In transgenic linesharbouring CYP85A1 fusions siliques and em-bryos showed no visible staining even after pro-longed incubation with the GUS substrate(Figure 1A). However, in CYP85A2:GUS linesflowers had high levels of expression in the style,anthers and sepals (Figure 1B). Staining was alsoevident in the siliques where closer analysis re-vealed especially high expression in the funiculi.There was no evidence of GUS activity in theembryos of either GUS transgenic lines until themature embryo stage when the cotyledons ofCYP85A2 lines showed minimal staining afterovernight incubation in GUS substrate. This wasnot seen in any of the CYP85A1 lines analysed.
Spatial patterns of expression
The main difference between the spatial expressionof the CYP85:GUS fusions was identifiable in thecotyledons. In transgenic seedlings harbouring the
132

Figure 1. Temporal expression of CYP85 genes. (A) Tissue-specific distribution of GUS activity in transgenic embryos (i–iv) and
seedlings harbouring CYP85A1 promoter: GUS constructs grown on 0.5· MS medium for 2, 4, 7, 11, 15 and 21 days, (v–x) respec-
tively. Flowers showed no staining in either male or female organs (xi–xii). (B) GUS activity in transgenic embryos (i–iv), funiculus (v–
vi) and seedlings (vii–xii) harbouring CYP85A2 promoter: GUS fusion. Age of the seedlings was the same as above: 2, 4, 7, 11, 15 and
21 days (panels vii–xii, respectively). Flowers show staining in the sepals, anthers and stamen (xiii–xiv).
133
CYP85A1:GUS reporter construct it was evidentfrom histochemical analysis, particularly in youngseedlings, that there was no detectable expressionin the cotyledons, even in very young seedlings.However, in the lines harbouring the CYP85A2:-GUS fusion staining is easily observed in the cot-yledons. Other variations were minimal. Inparticular, staining was intense in vascular tissues,at the root–hypocotyl junction and in the shootapical meristem.
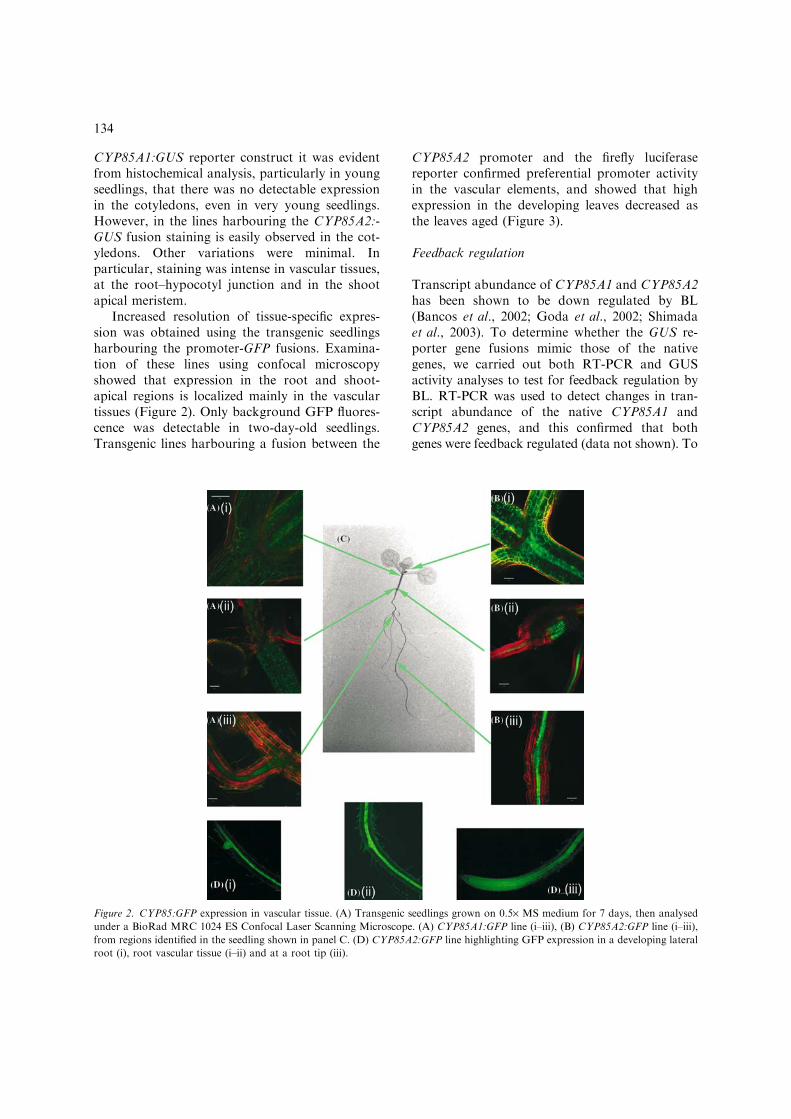
Increased resolution of tissue-specific expres-sion was obtained using the transgenic seedlingsharbouring the promoter-GFP fusions. Examina-tion of these lines using confocal microscopyshowed that expression in the root and shoot-apical regions is localized mainly in the vasculartissues (Figure 2). Only background GFP fluores-cence was detectable in two-day-old seedlings.Transgenic lines harbouring a fusion between the
CYP85A2 promoter and the firefly luciferasereporter confirmed preferential promoter activityin the vascular elements, and showed that highexpression in the developing leaves decreased asthe leaves aged (Figure 3).
Feedback regulation
Transcript abundance of CYP85A1 and CYP85A2has been shown to be down regulated by BL(Bancos et al., 2002; Goda et al., 2002; Shimadaet al., 2003). To determine whether the GUS re-porter gene fusions mimic those of the nativegenes, we carried out both RT-PCR and GUSactivity analyses to test for feedback regulation byBL. RT-PCR was used to detect changes in tran-script abundance of the native CYP85A1 andCYP85A2 genes, and this confirmed that bothgenes were feedback regulated (data not shown). To
Figure 2. CYP85:GFP expression in vascular tissue. (A) Transgenic seedlings grown on 0.5· MS medium for 7 days, then analysed
under a BioRad MRC 1024 ES Confocal Laser Scanning Microscope. (A) CYP85A1:GFP line (i–iii), (B) CYP85A2:GFP line (i–iii),
from regions identified in the seedling shown in panel C. (D) CYP85A2:GFP line highlighting GFP expression in a developing lateral
root (i), root vascular tissue (i–ii) and at a root tip (iii).
134

show that the reporter fusion lines behaved simi-larly, down regulation of GUS enzyme activity wasdemonstrated histochemically in both CYP85A1and CYP85A2 fusions (Figure 4). QuantitativeGUS assays verified this observation, and alsoindicated that the CYP85A2 promoter lines havemuch higher activity than those of CYP85A1 lines(Table 1).
Comparison of 50 regulatory sequencesof CYP85 genes
Given the high level of amino acid sequence iden-tity between CYP85A1 and CYP85A2, it is notsurprising that conversion assays with the yeast-expressed proteins have shown similar substratespecificity for both of these P450 enzymes (Shi-mada et al., 2001, 2003). However, it can be clearlyseen that tissue-specific differences exist in theexpression patterns of these genes. Because of thecharacteristic patterns of expression, we compared
the promoters plus 50-UTR sequences of these twogenes using the available data at the TAIR data-base. These regions share short stretches ofhomologous sequence, especially in the 50-UTRsjust upstream of the putative translational startcodons, and at the putative TATA boxes(Figure 5B). The positions of these TATA boxesare 33 and 71 bp upsteam of the nucleotidescorresponding to the longest known cDNAsequences, and are part of the CTATATAAACAmotif (Figure 5B) that is also conserved in thetomato DWARF promoter. Regions further up-stream of the CYP85A1 and CYP85A2 promotersshow only much lower levels of sequence identity(data not shown). On chromosome 5 the CYP85A1promoter may extend to a copia-like element(At5g38975) about 1.8 kb upstream of thecoding sequence. By contrast, on chromosome 3there is a distance of nearly 13 kb between the 50 endof CYP85A2 and the nearest annotated gene(At3g30170, with similarity to MuDR transposase).
Figure 3. Luciferase activity from CYP85A2:LUC fusion in
21-day-old plant. (A) Photograph of an Arabidopsis rosette of a
CYP85A2:LUC transgenic line. (B) Pseudocolour CCD camera
image of the same plant taken after incubation with luciferin
substrate. Maximum luminescence corresponds to white and
red regions.
Figure 4. Feedback regulation of CYP85:GUS fusions by
brassinolide. GUS histochemical staining of seedlings har-
bouring promoter:GUS constructs, grown for 5 days on 0.5·MS medium +/) BL (1 lM), then incubated for 8 h in
X-glucuronide substrate. The seedlings on the left are untreated,
the seedlings on the right show BL inhibition of GUS activity.
(A) CYP85A1:GUS, (B) CYP85A2:GUS.
Table 1. BL feedback regulation of GUS activity in transgenic lines harbouring CYP85A1, CYP85A2 and DWARF promoter-GUSconstructs.
Promoter fusion line GUS enzyme activity
(pmol MUG min)1 mg)1 protein)
Significance T-value
Untreated 1 lM BL
CYP85A1 356 ± 121 137 ± 36 7.34
CYP85A2 48205 ± 5859 38459 ± 3353 3.53
DWARF 4495 ± 483 2282 ± 363 8.97
GUS enzyme activities were calculated on the basis of data obtained from six independent pools of seven-day-old seedlings. Thestandard deviation about the mean is shown. Statistical analysis using a Student’s t-test indicated that the BL effect was significant forall samples tested at P = 0.01.
135

Temporal and spatial expression of the tomatoDWARF promoter-GUS fusions in Arabidopsis
We have shown that the CYP85A1 and CYP85A2genes are differentially regulated in Arabidopsis,even though their products have a high level ofamino acid sequence identity and show the samesubstrate specificity. We wanted to observe tran-scriptional regulation of tomato DWARF inArabidopsis and compare its expression pattern
with those of the Arabidopsis C-6 oxidase genes.Seven-day-old transgenic seedlings harbouring theDWARF:GUS fusion were subjected to histo-chemical analysis and this revealed that DWARFpromoter activity was very similar to that ofCYP85A2. Staining was evident mainly in theroot, root–hypocotyl junction and shoot apicalmeristem. GUS activity was also detected in thehypocotyls, especially in the vascular tissue, beinghighest in the tips of the cotyledons (Figure 6A).
Figure 5. Sequence comparison of Arabidopsis CYP85 promoters. (A) Chromosomal segments surrounding the CYP85 genes.
Arrowheads indicate regions containing coding sequences of the following predicted products: receptor protein kinase-like protein
(At5g38990), unknown protein (At5g38980), Ty1 copia-like element (At5g38975) and MuDR transposase-like protein (At3g30170).
The scheme is based on the most recent annotation available at the TAIR web site: http://www.arabidopsis.org (last accessed on
November 12, 2004). (B) 50 regulatory sequences of the CYP85A1 (At5g38970, accession No. NC_003076) and CYP85A2 (At3g30180,
accession No. NC_003074) genes. Homology is highlighted by shading, predicted TATA boxes and ATG start codons are indicated.
Asterisks mark the most 50 ends of the cDNA sequences associated with each gene (identified with their accession numbers).
136

This mimicked the expression pattern seen intomato (Bishop et al., unpublished data). Theintensity of staining suggested that in the trans-genic lines tested the activity of the DWARF pro-moter was less than that of CYP85A2, but higherthan that of CYP85A1. The expression of theDWARF:GUS fusion was also down regulated byBL, this is shown histochemically (Figure 6B) andfluorometrically by GUS assays (Table 1). Similarregulation of the DWARF promoter in tomato andArabidopsis highlights the conservation of thetranscriptional control mechanisms between thesespecies.
Discussion
One interesting aspect in the study of the BR C-6oxidase genes is the apparent lack of BR-deficientdwarf mutants in these genes in Arabidopsis. Thisis in contrast with tomato and rice where suchmutants have been described (Bishop et al., 1996;Hong et al., 2002; Mori et al., 2002). This is mostprobably due to the functional redundancy of thetwo C-6 oxidase genes in Arabidopsis. In fact,among the SALK T-DNA insertion mutantsdeficient in these P450s only cyp85a2 lines exhibit aweakly dwarfed phenotype, whereas no pheno-typic effect is observable in cyp85a1 plants. Incontrast, the double mutants lacking both CYP85
functions are severe dwarfs (T. Nomura, personalcommunication). This redundancy of CYP85,however, provides a possibility for differentialtranscriptional regulation. Such variation in thecontrol of these oxidases has been supported byprevious studies using RT-PCR and real time PCRto determine mRNA abundance in various organsof Arabidopsis (Bancos et al., 2002; Shimada et al.,2003). The disadvantages in using these methodsare that PCR is invasive, requires multiple RNAextractions and does not allow precise localiza-tion. To overcome these technical shortcomings, weexploited transgenic seedlings harbouring reporterconstructs to identify organ-specific promoteractivities over time, and to correlate these with theexpected sites of BR biosynthesis. Expression of thereporter genes in this study showed good correla-tion with those of the native CYP85 genes. We havedemonstrated that reporter activities are downregulated by BL, as is expression of the nativegene. The feedback effect on CYP85 promoteractivities was clearly observed by GUS histochem-ical staining. However, due to the weaker promoter,this response was found to be more erratic inCYP85A1:GUS transgenic seedlings.
Our observations regarding the temporal andspatial expression patterns of theCYP85 genes wereconfirmed using the microarray data that havebeen made accessible through the GENEVESTI-GATOR database: https://www.geneveastigator.ethz.ch (Zimmermann et al., 2004). Digital North-erns, using all 1434microarray data sets available atthe time of the analysis (November 5, 2004), indi-cated that in most cases CYP85A1 expression wasvery low, within the noise range of the experiments.In the same samples, CYP85A2 expression wassubstantially higher, and feedback regulation ofCYP85A2 expression was seen in response to BLapplication.
Previous studies have indicated that CYP85A1mRNA accumulated primarily in the root (Bancoset al., 2002), but it has also been reported to bemore abundant in the shoot (Shimada et al., 2003).Our data showed that CYP85A1 expression ispredominantly in the root and hypocotyl, andwithin this region primarily at the root–hypocotyljunction, during early development. The apparentdiscrepancy between these data can be attributedto both the low level expression of this gene andthe different separation of root/shoot material,because the bordering root–hypocotyl junction has
Figure 6. Feedback regulation of tomato DWARF:GUS fusion
by brassinolide in transgenic Arabidopsis. Five-day-old
DWARF:GUS lines grown on 0.5· MS medium +/) 1 lM BL.
(A) control, (B) 1 lM BL.
137

high CYP85A1 activity. In seedlings harbouringthe promoter-GFP fusion, fluorescence was de-tected in the shoot apical meristem, root–hypo-cotyl junction and in the roots. However, most ofthe GFP was seen in the root vascular tissue andnot in the peripheral cells or in the root–hypocotyljunction. We found no evidence of CYP85A1promoter activity in the embryo, siliques or inflo-rescence stems. These observations highlight that,in comparison to CYP85A2, CYP85A1 expressionis mainly associated with early developmentalprocesses.
The data obtained from GUS histochemicalanalysis of the CYP85A2 promoter fusion con-struct showed that expression was predominantlyin the cotyledons, although it could also be de-tected at the root–hypocotyl junction. We foundthat CYP85A2 promoter activity was strongerthan that of CYP85A1, as revealed by the analysisof ten representative transgenic lines of each of thetwo promoters. This is in agreement with previousstudies where transcript abundance of CYP85A2 iscomparatively higher than that of CYP85A1(Shimada et al., 2003). Using RT-PCR, Bancoset al. (2002) also showed that in developing seed-lings CYP85A2 expression is stronger in the shootsthan the roots. It is interesting to note that tran-scription from this promoter was evident in thecotyledons of the embryo, suggesting that BRsynthesis takes place in the later stages ofembryogenesis. This highlights a possible link toprevious observations identifying a role of sterolsduring plant embryogenesis (Jang et al., 2000;Schrick et al., 2000). A shared feature of the dataobtained from both CYP85 promoters is thatstrongest expression occurs at an early stage ofdevelopment and is then down regulated overtime. This is consistent with reported RT-PCRdata showing that CYP85A1 and CYP85A2 tran-script levels peaked over the first week of germi-nation (Bancos et al., 2002).
In seedlings carrying GFP fusions with eitherthe CYP85A1 or CYP85A2 promoter, GFPexpression was mainly seen in the vascular tissue.Previous reports that endogenous BRs are in-volved in the promotion of xylogenesis and thatBR dwarf mutants display abnormal xylem aresuggestive that BR biosynthesis would occur in theregion of vascular tissue (Yamamoto et al., 2001).This is particularly interesting because a recentpublication identified a provascular/procambial
cell-specific gene named VASCULAR HIGH-WAY 1 (VH1), which is involved in early vasculardevelopment (Clay and Nelson, 2002). It wasdetermined that the predicted VH1 protein is aleucine-rich repeat receptor kinase, but the sig-nalling molecule which activates the VH1 signallingpathway, inducing differentiation of provascularcells into procambial cells, remains unidentified.The ligand was predicted to be a small peptide orhormone. It was exciting to note that VH1 is closelyrelated to BRASSINOSTEROID INSENSITIVE 1(BRI1), the leucine-rich repeat receptor kinaseinvolved in BR signalling (Li and Chory, 1997). AsCYP85 enzymes are involved in C-6 oxidation ofBRs, it can be assumed that sites of active CYP85transcription coincide with those of BR synthesis,thus implying BL production in the vascular tissues.BR synthesis has previously been shown to increasedramatically during vascular development andBRs were found to be essential in the final stage oftracheary element differentiation (Fukuda, 1997;Yamamoto et al., 2001).
Histochemical data showing intense GUSactivity in the shoot apical meristem, particularlyin CYP85A2:GUS seedlings, support a previousreport in which DWARF (CYP85A) was impli-cated in the earliest stages of leaf development intomato (Pien et al., 2001). Differential displayanalysis, performed to identify asymmetricallyexpressed genes during leaf initiation, showed thatDWARF transcripts accumulated in the region ofthe meristem destined for leaf formation (Pienet al., 2001). Furthermore, Arabidopsis CPD,encoding another P450 enzyme of the BR path-way, was also shown to be highly expressed in theshoot meristematic region (Mathur et al., 1998).These data underline the importance of BRs in leafdevelopment and suggest a role for ArabidopsisCYP85s in this process.
Sequence analysis in Arabidopsis has revealedthe presence of two CYP85 genes encoding ortho-logs of tomato DWARF. CYP85A1 and CYP85A2share 82% amino acid sequence identity, and areabout 70% identical to DWARF (Shimada et al.,2003). It seems likely that in Arabidopsis theredundancy ofCYP85 genes is the result of a recentduplication and, since these sequences are in closeproximity to transposon-like sequences, it is excit-ing to postulate that the duplication may havearisen due to a transposition-based recombinationevent. One consequence of the duplication event
138

would be to enable differential regulation of theresulting genes copies. In accordance with the as-sumed gene duplication, our promoter analysisrevealed that, despite their differential expression,considerable sequence homology can be recognizedbetween the two Arabidopsis C-6 oxidase pro-moters. It was interesting to note that the tran-scriptional regulatory elements of CYP85 genes areapparently well conserved between dicot species,because the DWARF promoter in Arabidopsisconferred a similar expression pattern andBR-responsiveness as in tomato.
It is not unique that P450s of the same sub-family show differential regulation and expression.CYP86A8, a P450 involved in fatty acid modifica-tion, is expressed only in seedling shoots, whereasCYP86A1 of the same subfamily, also involvedin fatty acid modification, is expressed solely inthe roots (Schuler and Werck-Reichhart, 2003).Redundancy of hormone biosynthetic genes, cou-pled with differential expression, is a well-knownphenomenon in the plant kingdom. For instance, inrice two GA 20-oxidase genes were identified. Bothgenes were down regulated by GA, in the samemanner as the BR C-6 oxidase genes are by BL.GA20ox-1 was found to be preferentially expressedin the flowers, whereasGA20ox-2was active in bothleaves and flowers. It was found that in the semi-dwarf variety IR8, mutation of GA20ox-2 resultedin GA-deficiency in leaves, giving rise to the dwarfphenotype (Ashikari et al., 2002).
Although reporter fusions can provide valuableinformation about promoter activities, a crucialproblem is how faithfully these data reflect theexpression of the native gene. In this study theconsistency of results obtained with differentreporters, and their agreement with published dataregarding organ-specificity and feedback regula-tion, clearly indicate that expression of the reporterfusions was representative of that of the endoge-nous CYP85 genes. Our data show that expressionof these highly similar genes is differentially regu-lated during plant development, and suggest thatCYP85A1 and CYP85A2 have roles in differenti-ation, particularly in that of the vascular tissue.
Acknowledgements
We wish to thank those in the Aberystwyth Bot-any Gardens for help with plant care. We are
grateful to S. Bancos and R. Darby for guidanceand advice on GUS assays, as well as T. Nomurafor communicating unpublished results. We thankJ. Jones, Sainsbury Laboratory, UK, for providingplasmids and K. Harrison for help in generatingtransgenic lines. This research was supported byan IBS studentship to J.C. and by the RoyalSociety Joint Project. We also thank the followingfunding agencies that support our research. Bio-technology and Biological Sciences Research Coun-cil (Grant X511786), Human Frontier ScienceProgram (Grant RG 162/2000-M), HungarianScientific Research Fund (Grant T 42639).
References
Ait-Ali, T., Frances, S., Weller, J.L., Reid, J.B., Kendrick, R.E.and Kamiya, Y. 1999. Regulation of gibberellin 20-oxidaseand gibberellin 3b-hydoxylase transcript accumulation dur-ing de-etiolation of pea seedlings. Plant Physiol. 121: 783–791.
von Arnim, A.G., Deng, X.W. and Stacey, M.G. 1998. Cloningvectors for the expression of green fluorescent protein intransgenic plants. Gene 221: 35–43.
Ashikari, M., Sasaki, A., Ueguchi-Tanaka, M., Itoh, H.,Nishimura, A., Datta, S., Ishiyama, K., Saito, T.,Kobayashi, M., Khush, G.S., Kitano, H. and Matsuoka,M. 2002. Loss-of-function of a rice gibberellin biosyntheticgene, GA20 oxidase (GA20ox-2), led to the rice ‘greenrevolution’. Breed. Sci. 52: 143–150.
Bancos, S., Nomura, T., Sato, T., Molnar, G., Bishop, G.J.,Koncz, C., Yokota, T., Nagy, F. and Szekeres, M. 2002.Regulation of transcript levels of the Arabidopsis cyto-chrome P450 genes involved in brassinosteroid biosynthesis.Plant Physiol. 130: 504–513.
Bishop, G.J., Harrison, K. and Jones, J.D.G. 1996. The tomatoDwarf gene isolated by heterologous transposon taggingencodes the first member of a new cytochrome P450 family.Plant Cell 8: 959–969.
Bishop, G.J. and Koncz, C. 2002. Brassinosteroids and plantsteroid hormone signalling. Plant Cell 14: S97–S110.
Bishop, G.J., Nomura, T., Yokota, T., Harrison, K., Noguchi, T.,Fujioka, S., Takatsuto, S., Jones, J.D.G. andKamiya, Y. 1999.The tomato DWARF enzyme catalyses C-6 oxidation inbrassinosteroid biosynthesis. Proc. Natl. Acad. Sci. USA 96:1761–1766.
Bishop, G.J. and Yokota, T. 2001. Plant steroid hormones,brassinosteroids: current highlights of molecular aspects ontheir synthesis/metabolism, transport, perception and re-sponse. Plant Cell Physiol. 42: 114–120.
Bradford, M. 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72: 248–254.
Clay, N.K. and Nelson, T. 2002. VH1, a provascular cell-specific receptor kinase that influences leaf cell patterns inArabidopsis. Plant Cell 14: 2707–2722.
Clough, S.J. and Bent, A.F. 1998. Floral dip: a simplifiedmethod for Agrobacterium-mediated transformation ofArabidopsis thaliana. Plant J. 16: 735–743.
139

Clouse, S.D. and Feldmann, K.A. 1999. Molecular geneticsof brassinosteroid action. In: A. Sakurai, T. Yokota andS.D. Clouse (Eds.), Brassinosteroids Steroidal Plant Hor-mones, Springer, Tokyo, pp. 163–190.
Clouse, S.D. and Sasse, J.M. 1998. Brassinosteroids: essentialregulators of plant growth and development. Annu. Rev.Plant. Physiol. Plant Mol. Biol. 49: 427–451.
Fukuda, H. 1997. Tracheary element differentiation. Plant Cell9: 1147–1156.
Goda, H., Shimada, Y., Asami, T., Fujioka, S. and Yoshida, S.2002. Microarray analysis of brassinosteroid regulated genesin Arabidopsis thaliana. Plant Physiol. 130: 1319–1334.
Hong, Z., Ueguchi-Tanaka, M., Shimizu-Sato, S., Inukai, Y.,Fujioka, S., Shimada, Y., Takatsuto, S., Agetsuma, M.,Yoshida, S., Watanabe, Y., Uozu, S., Kitano, H., Ashikari, M.and Matsuoka, M. 2002. Loss-of-function of a rice brassi-nosteroid biosynthetic enzyme, C-6 oxidase, prevents theorganised arrangement and polar elongation of cells in theleaves and stem. Plant J. 32: 495–508.
Jang, J.C., Fujioka, S., Tasaka, M., Seto, H., Takatsuto, S.,Ishii, A., Aida, M., Yoshida, S. and Sheen, J. 2000. A criticalrole of sterols in embryonic patterning and meristemprogramming revealed by the fackel mutants of Arabidopsisthaliana. Gene Dev. 14: 1485–1497.
Jefferson, R.A., Kavanagh, T.A. and Bevan, M.W. 1987. GUSfusions: ß-glucuronidase as a sensitive and versatile genefusion marker in higher plants. EMBO J. 6: 3901–3907.
Kaneko, M., Itoh, H., Inukai, Y., Sakamoto, T., Ueguchi-Tanaka, M., Ashikari, M. and Matsuoka, M. 2003. Wheredo gibberellin biosynthesis and gibberellin signaling occur inrice plants? Plant J. 35: 104–115.
Koncz, C.,Martini, N., Szabados, L., Hrouda,M., Brachmair, A.and Schell, J. 1994. Specialized vectors for gene tagging andexpression studies. In: S.B. Gelvin and A.R. Schilperoort(Eds.), Plant Molecular Biology Manual, Vol. B2, KluwerAcademic Publishers, Dordecht, The Netherlands, pp. 1–22.
Li, J.M. and Chory, J. 1997. A putative leucine-rich repeatreceptor kinase involved in brassinosteroid signal transduc-tion. Cell 90: 929–938.
Li, J.M. and Chory, J. 1999. Brassinosteroid action in plants. J.Exp. Bot. 50: 275–282.
Li, J.M., Nagpal, P., Vitart, V., McMorris, T.C. and Chory, J.1996. A role for brassinosteroids in light-dependent devel-opment of Arabidopsis. Science 272: 398–401.
Mandava, B.N. 1988. Plant growth-promoting brassinoster-oids. Annu. Rev. Plant Physiol. Plant Mol. Biol. 39: 23–52.
Mathur, J., Molnar, G., Fujioka, S., Takatsuto, S., Sakurai, A.,Yokota, T., Adam,G., Voigt, B., Nagy, F.,Maas, C., Schell, J.,Koncz, C. and Szekeres, M. 1998. Transcription of theArabidopsis CPD gene, encoding a steroidogenic cytochromeP450, is negatively controlled by brassinosteroids. Plant J. 14:593–602.
Montoya, T., Nomura, T., Farrar, K., Kaneta, T., Yokota, T.and Bishop, G.J. 2002. Cloning the tomato curl3 genehighlights the putative dual role of the leucine-rich repeatreceptor kinase tBRI1/SR160 in plant steroid hormone andpeptide hormone signaling. Plant Cell 14: 3163–3176.
Mori, M., Nomura, T., Ooka, H., Ishizaka, M., Yokota, T.,Sugimoto, K., Okabe, K., Kajiwara, H., Satoh, K.,Yamamoto, K., Hirochika, H. and Kikuchi, S. 2002. Isolationand characterisation of a rice dwarf mutant defective inbrassinosteroid biosynthesis. Plant Physiol. 130: 1152–1161.
Murashige, T. and Skoog, F. 1962. A revised medium for rapidgrowth and bioassays with tobacco tissue cultures. Physiol.Plantarum 15: 473–497.
Noguchi, T., Fujioka, S., Choe, S., Takatsuto, S., Tax, F.E.,Yoshida, S. andFeldmann,K.A. 2000. Biosynthetic pathwaysof brassinolide in Arabidopsis. Plant Physiol. 124: 201–209.
Pien, S., Wyrzykowska, J. and Fleming, A.J. 2001. Novelmarker genes for early leaf development indicate spatialregulation of carbohydrate metabolism within the apicalmeristem. Plant J. 25: 663–674.
Schrick, K., Mayer, U., Horrichs, A., Kuhnt, C., Bellini, C.,Dangl, J., Schmidt, J. and Jurgens, G. 2000. FACKEL is asterol C-14 reductase required for organised cell division andexpansion in Arabidopsis embryogenesis. Gene Dev. 14:1471–1484.
Schuler, M.A. and Werck-Reichhart, D. 2003. Functionalgenomics of P450s. Annu. Rev. Plant Biol. 54: 629–667.
Schumacher, K. and Chory, J. 2000. Brassinosteroid signaltransduction: still casting the actors. Curr. Opin. Plant Biol.3: 79–84.
Shimada,Y., Fujioka, S.,Miyauchi,N.,Kushiro,M., Takatsuto, S.,Nomura, T., Yokota, T., Kamiya, Y., Bishop, G.J. andYoshida, S. 2001. Brassinosteroid-6-oxidases from Arabid-opsis and tomato catalyze multiple C-6 oxidations inbrassinosteroid biosynthesis. Plant Physiol. 126: 770–779.
Shimada, Y., Goda, H., Nakamura, A., Takatsuto, S., Fujioka, S.and Yoshida, S. 2003. Organ-specific expression of brassi-nosteroid-biosynthetic genes and distribution of endogenousbrassinosteroids in Arabidopsis. Plant Physiol. 131: 287–297.
Yamaguchi, S., Kamiya, Y. and Sun, T.P. 2001. Distinct cell-specific expression patterns of early and late gibberellinbiosynthetic genes during Arabidopsis seed germination.Plant J. 28: 443–453.
Yamamoto, R., Fujioka, S., Demura, T., Takatsuto, S.,Yoshida, S. and Fukuda, H. 2001. Brassinosteroid levelsincrease drastically prior to morphogenesis of trachearyelements. Plant Physiol. 125: 556–563.
Yokota, T. 1997. The structure, biosynthesis and function ofbrassinosteroids. Trend. Plant. Sci. 2: 137–143.
Zimmermann, P., Hirsch-Hoffmann, M., Hennig, L., andGruissem, W. 2004. GENEVESTIGATOR: Arabidopsismicroarray database and analysis toolbox. Plant Physiol.136: 2621–2632.
140