
Peri-Pubertal Emergence of UNC-5 Homologue Expression by Dopamine Neurons in Rodents
Upload
independentCategory
view
0download
0
Animal Reproduction Science 80 (2004) 15–29
Ultrasonographic evaluation of the pre-pubertaldevelopment of the reproductive tract in beef heifers
Ali Honaramooza, Jayaprakash Aravindakshanb,Ramesh K. Chandoliac, Andrew P. Beardd, Pawel M. Bartlewskie,
Roger A. Piersone, Norman C. Rawlingsf,∗a Center for Animal Transgenesis and Germ Cell Research, New Bolton School of Veterinary Medicine,
University of Pennsylvania, Kennett Square, PA, USAb Human Health Research Centre, INRS-Institut Armand Frappier,
University of Quebec, Quebec City, Que., Canadac Department of Animal Reproduction, Gynaecology and Obstetrics,
Haryana Agricultural University, Hissar, Haryana, Indiad Department of Agriculture, University of Newcastle, Newcastle upon Tyne, UK
e Department of Obstetrics, Gynecology and Reproductive Sciences, Reproductive Biology Research Unit,College of Medicine, University of Saskatchewan, Saskatoon, Sask., Canada
f Department of Veterinary Biomedical Sciences, Western College of Veterinary Medicine,University of Saskatchewan, Saskatoon, Sask., Canada
Received 1 November 2002; received in revised form 6 May 2003; accepted 6 May 2003
Abstract
To study the development of the reproductive tract in heifers, the ovaries, uterus, cervix andvagina were examined by transrectal ultrasonography every 2 weeks, from 2 to 60 weeks afterbirth. First ovulation occurred at 63.7 ± 1.1 weeks of age. Ovarian dimensions increased rapidlyfrom 2 to 14 weeks of age, and increased again after 34 weeks of age (P < 0.05). The size ofthe largest ovarian follicles increased from 8 to 14 weeks of age, from 38 to 42 weeks of age, andfinally from 52 to 60 weeks of age (P < 0.05). The number of follicles≥3 mm in diameter tendedto increase from 6 to 14 weeks of age (P < 0.10) and increased significantly from 6 to 60 weeksof age (P < 0.05). Mean numerical pixel values of the ovarian images decreased from 4 to 26weeks of age, and then rose to 44 weeks of age (P < 0.05). Diameter of the uterine body, cervixand vagina increased from 2 to 20–24 weeks of age, and again after 32 weeks of age (P < 0.05).Mean numerical pixel values for the uterus and vagina decreased initially (uterus: 4–8 weeks andvagina: 6–22 weeks of age) and then increased (uterus: 14–42 weeks and vagina: 22–32 weeks ofage;P < 0.05). Pixel heterogeneity showed a consistent peak at 20–22 weeks of age for the uterus,cervix and vagina (P < 0.05). In summary, in the heifer calf, the marked growth of the reproductivetract in the first few months of age, and prior to first ovulation, reflects phases of increased ovarian
∗ Corresponding author. Tel.:+1-306-966-7357; fax:+1-306-966-7376.E-mail address:[email protected] (N.C. Rawlings).
0378-4320/$ – see front matter © 2003 Elsevier B.V. All rights reserved.doi:10.1016/S0378-4320(03)00136-2

16 A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29
follicle (≥3 mm in diameter) numbers and size. Ultrasonographic image analysis revealed patternsof numerical pixel values and heterogeneity that may be useful in determining important stages ofgrowth and differentiation of the reproductive system.© 2003 Elsevier B.V. All rights reserved.
Keywords:Cattle; Ultrasonography; Reproductive tract; Puberty
1. Introduction
In numerous studies, ultrasonography has been used to describe changes that occur in thereproductive tract of cows and heifers throughout the estrous cycle (Pierson and Ginther,1988; Savio et al., 1988; Sirois and Fortune, 1988; Adams, 1999). However, our understand-ing of the pre-pubertal development of the reproductive tract in heifers is limited and deriveslargely from infrequent (i.e. once a month) examinations of tissues collected at slaughter(Erickson, 1966; Desjardins and Hafs, 1969) or more recent ultrasonographic studies thathave focused on ovarian follicular dynamics (Adams et al., 1994; Bergfelt et al., 1994; Evanset al., 1994a,b; Ginther et al., 1996; Ahmad et al., 1997; Melvin et al., 1999). Non-invasiveassessment of the development of the ovaries and non-gonadal reproductive organs in thesame individual animals over the period of sexual development has not been possible todate.
Data obtained from slaughtering different groups of Holstein heifer calves at monthlyintervals suggested that ovarian weight increased from birth to 5 months of age, followedby a plateau to the time of first ovulation (Desjardins and Hafs, 1969). Growth of the uteruswas slow but linear to first ovulation, and growth of the cervix and vagina was slow to 4months of age with a more rapid increase to first ovulation (Desjardins and Hafs, 1969).
Ulltrasonographic examination of the ovaries of beef heifers, for periods of 18 days every6–10 weeks, showed that large antral follicles were present and grew in a wave-like fashionas early as 2 weeks of age (Adams et al., 1994; Evans et al., 1994a,b). The greatest increasein the maximum size and in the number of ovarian antral follicles occurred within the first2 months of life (Bergfelt et al., 1994). There were no major changes in the wave-likepattern of growth and regression of large follicles as heifers approached first ovulation(Evans et al., 1994b), but dominant ovarian follicles were reported to be larger as heifersapproached puberty (Bergfelt et al., 1994). The temporal relationships between pre-pubertalchanges in size and weight of the reproductive organs and ovarian antral follicle numbersin heifer calves have not been studied to date.
In spring-born heifers, there is an early pre-pubertal increase in luteinizing hormone (LH)secretion that is due mainly to a transient increase in LH pulse amplitude between 4 and20 weeks of age (Evans et al., 1992; Honaramooz et al., 1999). It has been proposed thatthis early increase in LH secretion may stimulate the remarkable ovarian follicular changesseen at that time and, therefore, may reflect a critical regulatory stage in the reproductivematuration of heifers (Evans et al., 1994a). LH pulse frequency, however, increases dra-matically during the last few weeks before puberty, resulting in elevated mean circulatingconcentrations of LH (Schams et al., 1981; Day et al., 1984, 1987; Kinder et al., 1987; Dyeret al., 1990; Evans et al., 1992; Chandolia et al., 1997; Honaramooz et al., 1999).

A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29 17
Real time, diagnostic ultrasonography allows for the non-invasive assessment of shape,size and internal structure of organs (Pierson and Adams, 1995). Gray-scale ultrasono-graphic images consist of picture elements (pixels), varying from white to black (Pierson andAdams, 1995). Numerical values ranging from 0 (black) to 255 (white) are assigned to gradepixel brightness. The pixel values depend on the relative density (fluid content, echotex-ture) or the ability of tissues to reflect (echogenic tissues—white) or transmit (non-echogenictissues—black) high frequency acoustic pressure waves, or an ultrasound beam (Pierson andAdams, 1995; Chandolia et al., 1997). For example, the cavities of ovarian antral folliclesappear as black and bones as white on the viewing screen of ultrasound equipment.
In previous studies of reproductive tract development in bull calves, numerical pixelvalues and heterogeneity showed distinct trends that were associated with such events asthe onset and progression of spermatogenesis, and the increase in secretory activity of theglandular accessory structures (Evans et al., 1996). In studies on ovarian follicular growthin adult cattle and humans, pixel values differed between various stages of the lifespan ofthe follicle, and reflected changes in vascularity, steroid synthesis, cell proliferation andfollicle viability (Pierson and Adams, 1995; Singh et al., 1998; Tom et al., 1998). To date,ultrasonographic technique has not been used to see if there are distinct changes in numericalpixel values and heterogeneity for the reproductive organs during sexual maturation in theheifer.
A longitudinal, non-invasive study of the entire reproductive system, in the same individ-ual animals, has not been done before and this information is important to understand thepattern of sexual development and its regulation. Hence, the first objective of this study wasto use transrectal ultrasonographic examination, every 2 weeks, to describe the develop-ment of the entire reproductive system in the heifer, from 2 to 60 weeks of age. The secondobjective was to see if there was a marked temporal pattern to numerical pixel values andpixel heterogeneity of the ultrasound images collected.
2. Materials and methods
2.1. Animals
The age-matched spring-born Hereford heifer calves (n = 5) used in this study werekept at pasture, along with their dams, until weaning at 25 weeks of age. After weaning,they were maintained in corrals and were provided with water, brome-alfalfa hay, mineralsand a ground concentrate ration (Honaramooz et al., 1999). Body weight was recordedevery 2 weeks from birth until puberty. Animals were handled and treated according tothe guidelines of the Canada Council of Animal Care. Data on pre-pubertal patterns ofgonadotropin secretion in these heifers have already been published (Honaramooz et al.,1999).
2.2. Ultrasonography
The internal reproductive tract of the heifer calves was examined every other week frombirth to puberty by transrectal ultrasonography using a B-mode real time echo camera (Aloka

18 A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29
SSD-500, Overseas Monitor Corporation Ltd., Richmond, BC, Canada). The length of theintra-rectal probe used was adjusted as the heifers grew. At 2–10 weeks of age, a 7.5 MHzlinear-array rigid human transrectal probe (35 cm× 1.6 cm) was used. At 12–30 weeksof age, when a longer rigid probe was needed, the cable of a standard I-shaped 7.5 MHzlinear-array probe was stiffened using a 45 cm length of polyethylene tubing (15 mm o.d.,12 mm i.d.), cut along its length and taped around the cable. After 32 weeks of age, theheifers were large enough to allow ultrasound examination by intra-rectal placement of agloved hand holding the probe. The two probes utilized the same transducer head, housedin different configurations. Both ultrasound transducers and imaging methods used in thisstudy were validated for the present applications in heifers (Adams et al., 1994; Evans et al.,1994a). At each observation, the ovaries, uterus, cervix and vagina were examined, and theimages were recorded on high-grade videotapes (Fuji-VHS, ST-120N) using a super-VHSvideocassette recorder (Panasonic, AG-1970, Professional Communications Systems Ltd.,Saskatoon, Sask., Canada) for retrospective analysis.
2.3. Image analysis
The sizes and numbers of ovarian follicles (≥3 mm in diameter), dimensions of theovaries, diameters of the uterus, cervix and vagina were taken from the recorded images.Ovarian dimension measurements excluded the size of superficial follicles≥3 mm in di-ameter, where such follicles distorted the recorded perimeter of the ovary. For the uterus,the greatest diameter of the uterine body was measured; for the cervix, the greatest di-ameter was taken; and for the vagina, the anterior one-third of the vagina was measured.Computer-assisted image intensity (numerical pixel values) analysis was performed as de-scribed previously (Singh et al., 1998). Ultrasonographic images were retrieved from thevideotapes using a PC based image acquisition board (Coreco Inc., St. Laurent, Que.,Canada) at a resolution of 640 pixels×480 pixels and standardized acquisition parameters.Images were transferred to a graphics workstation (SunSparc Station 10, Sun Microsystems,Mt. View, CA). Numerical pixel values comprising the ultrasound images were measuredusing custom developed computer algorithms optimized for ultrasonography (Synergyne1©, R.A. Pierson, University of Saskatchewan, Saskatoon, Sask., Canada). Numerical pixelvalues are described as gray-scale values of individual picture elements ranging from 0(black) to 255 (white). Mean numerical pixel values were determined in four spots (with adiameter of 15 pixels each), placed at random on the image of the parenchyma of the re-productive organs examined. For ovaries, the spots were placed on areas where no obviousfollicles were present. For each spot, the software provides the mean and standard deviationof all encircled pixel units. The average of these means from the four spots in the sametissue in the image was calculated. The average of standard deviations obtained from thefour spots analyzed was calculated to represent the degree of pixel heterogeneity.
2.4. Detection of first ovulation
Starting at 42 weeks of age, single blood samples were collected two or three times aweek from each heifer to measure plasma levels of progesterone, in order to retrospectivelydetermine age at puberty (as described previously for these heifers;Honaramooz et al.,

A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29 19
1999). Age at puberty was defined as the first time at which plasma levels of progesteroneexceeded 1.0 ng/ml, as an indication of first ovulation. Ultrasonographic examination of theovaries was also performed when blood samples were taken to monitor the development ofthe first corpus luteum (CL).
2.5. Statistical analyses
Ultrasonographic examinations of each heifer continued until puberty (first ovulation;range 61–67 weeks of age). As there were no dramatic changes in any of the characteris-tics measured immediately preceding first ovulation, and since growth of CL made ovarianmeasurements unclear, data up to 60 weeks of age were used for statistical analysis. All datawere analyzed for the effect of age using one-way repeated measures analysis of variance(SigmaStat® for Windows, Version 1.0, Jandel Corporation, San Rafael, CA, USA). Theovarian data were also analyzed for the effects of side (left versus right ovary), age and theirinteractions using two-way repeated measures analysis of variance. In all analyses, if themain effects were significant, Student–Newman–Keuls test for least significant differences(P < 0.05) was used as the post-analysis of variance test. For the purpose of presentation,the results of multiple range tests were simplified and are shown on figures by dashed/solidjoints, which illustrate major statistical trends. Major trends delimit periods during whichthere were progressive and significant changes in mean values. Pearson product–momentcorrelation coefficients were calculated amongst some characteristics of ovarian and tubu-lar genitalia (SigmaStat®). All data are presented as mean± standard error of the mean(S.E.M.).
3. Results
3.1. First ovulation
In all heifers, the concurrent presence of a CL was confirmed when plasma levels ofprogesterone rose above 1.0 ng/ml. However, in three heifers, during the week preceding thefirst ovulation, a luteinized follicle was observed and the maximum plasma progesteronevalues at this time in these animals were 0.3, 0.4 and 0.6 ng/ml. As reported previously(Honaramooz et al., 1999), the mean (±S.E.M.) age at puberty in the whole group ofheifers was 63.7 ± 1.1 weeks and ranged between 61 and 67 weeks of age. The mean(±S.E.M.) body weight at puberty was 401± 9 kg, which ranged between 370 and 422 kg.Growth rate of the heifers was 0.90± 0.03 kg per day.
3.2. Ovarian changes
Ovarian dimensions increased with age (P < 0.001) but did not differ between ovariesand there were no interactions (P > 0.05); therefore, data from the left and right ovarieswere pooled. Ovarian length increased from 2 to 14 weeks of age and again from 34 to 60weeks of age (P < 0.05; Fig. 1a). Ovarian diameter increased from 2 to 14 weeks of age(P < 0.05;Fig. 1a) and again from 30 to 44 weeks of age (P < 0.05;Fig. 1a).

20 A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29
Age (wks)
0 4 8 12 16 20 24 28 32 36 40 44 48 52 56 60
Num
eric
al p
ixel
val
ues
10
15
20
25
30
100
120
140
160
Pix
el h
eter
ogen
eity
0 4 8 12 16 20 24 28 32 36 40 44 48 52 56 60
Ova
rian
dim
ensi
ons
(mm
)
0
5
10
15
20
25
30
35LengthDiameter
(a)
(b)
Fig. 1. Ovarian length and diameter (a), numerical pixel values and pixel heterogeneity (b), from 2 to 60 weeksof age in heifers (mean± S.E.M.). Dashed lines show major statistical trends (P < 0.05). Major statistical trendsdelimit the periods during which there were progressive changes in mean values.

A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29 21
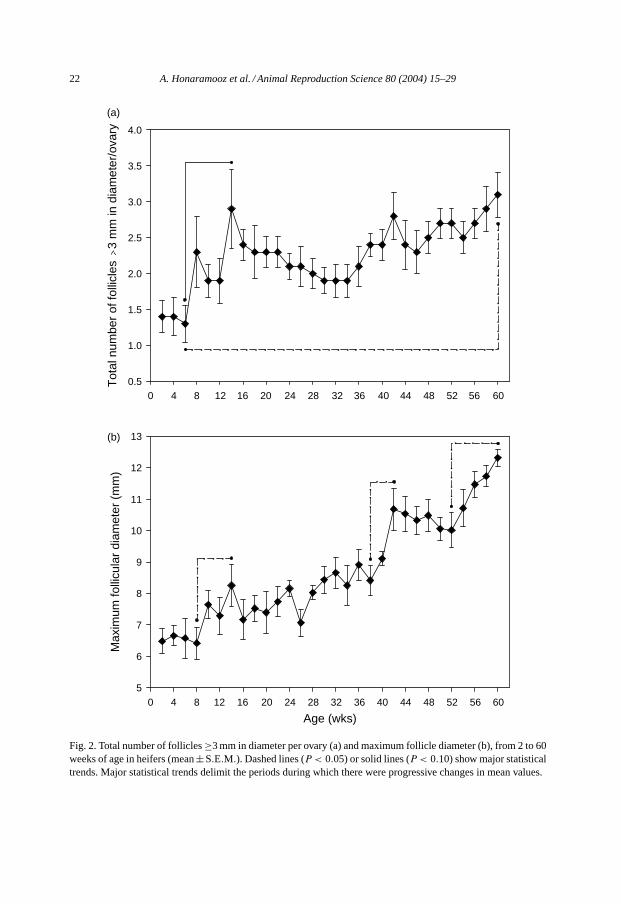
The number of follicles≥3 mm in diameter tended to increase from 6 to 14 weeksof age (P < 0.10; Fig. 2a); however, numbers of follicles≥3 mm in diameter increasedsignificantly from 6 to 60 weeks of age (P < 0.05;Fig. 2a).
The right and left ovaries differed only for the overall size of the largest follicles (P =0.001). The size of the largest follicles on the right ovary was about 1 mm greater than thatof the left ovary (P < 0.05). Although the size of the largest ovarian follicles changed overtime (P < 0.001;Fig. 2b), there were no interactions between age and left or right ovary(P > 0.05), indicating that the pattern of development of the largest ovarian follicle did notdiffer between the right and left ovary. Therefore, data for the left and right ovaries werepooled (Fig. 2). Maximum follicle diameter increased from 8 to 14 weeks, from 38 to 42weeks and finally from 52 to 60 weeks of age (P < 0.05;Fig. 2b).
Mean numerical pixel values measured from ovarian images changed over time for bothovaries (P < 0.01;Fig. 1b). Numerical pixel values decreased from 4 to 26 weeks of age andthen rebounded to 44 weeks of age (P < 0.05; Fig. 1b). The degree of pixel heterogeneityin the ovaries showed no significant changes over time (P > 0.05;Fig. 1b).
3.3. Tubular development
The diameter of the uterine body increased over time (P < 0.001). Growth was rapid to 10weeks of age and then declined to 12 weeks of age, increased from 12 to 24 weeks and againfrom 32 to 60 weeks of age (Fig. 3a). Mean numerical pixel values of the uterine imagesshowed an initial sharp decline from 4 to 8 weeks of age, followed by a gradual increasefrom 14 to 42 weeks of age (P < 0.05; Fig. 3b). The pixel heterogeneity of the uterusalso changed over time (P < 0.001), starting with an abrupt transient increase between 18and 20 weeks of age, followed by a period in which values remained relatively unchanged,except for an apparent increase at 42 weeks of age (P < 0.01;Fig. 3b).
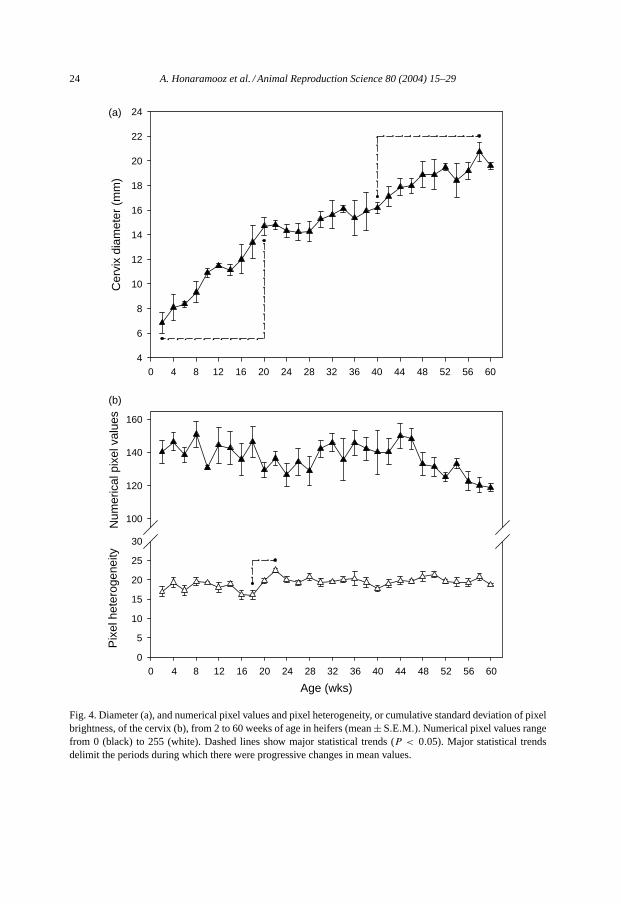
Cervical diameter also showed significant changes over time (P < 0.001). Cervicaldiameter increased from 2 to 20 weeks of age, and again from 40 to 58 weeks of age(P < 0.05; Fig. 4a). Numerical pixel values decreased with time (P < 0.05); however, nosingle time-points differed from another (Fig. 4b). The pixel heterogeneity of the cervicalimages showed a similar pattern to that of the uterine images with a marked peak at 22weeks of age (P < 0.05;Fig. 4b).
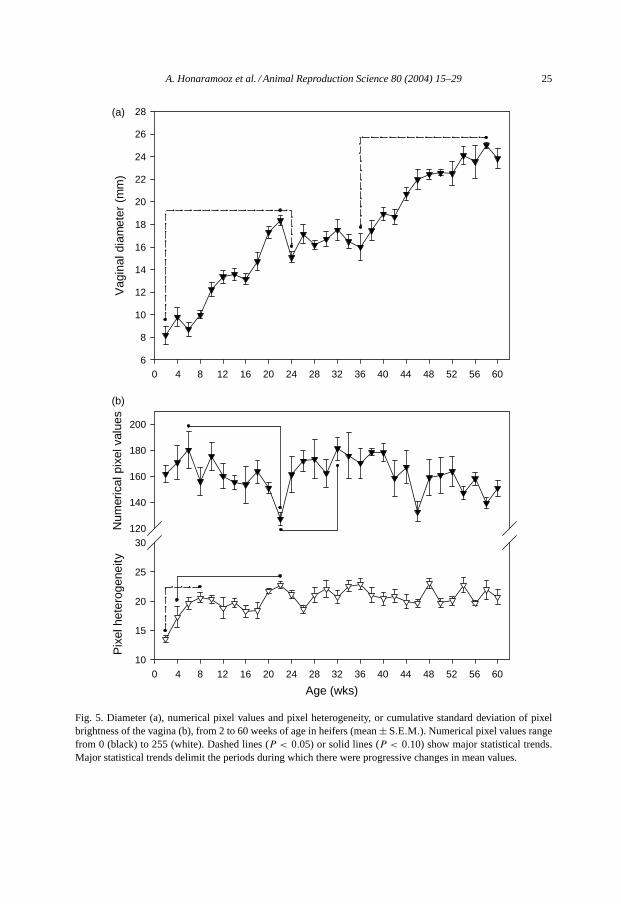
Vaginal diameter increased from 2 to 22 weeks of age, decreased to 24 weeks, plateauedand subsequently increased from 36 to 58 weeks of age (P < 0.05; Fig. 5a). Numericalpixel values of the vaginal images changed significantly over time (P < 0.05; Fig. 5b).However, individual contrasts only approached significance (P < 0.10). Numerical pixelvalues appeared to decline from 6 to 22 weeks of age and increased again from 22 to 32weeks of age (P < 0.05). Vaginal pixel heterogeneity changed over time (P < 0.001) andincreased from 2 to 8 weeks of age (P < 0.05; Fig. 5b). Points at 22, 36, 48 and 54 weeksof age appeared to differ from pixel heterogeneity at 4 weeks of age.
3.4. Correlations between development of the ovary and tubular genitalia
There were strong positive correlations amongst maximum follicle diameter and diameterof each segment of the reproductive tract studied (P < 0.01; Table 1). Numbers of larger
22 A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29
0 4 8 12 16 20 24 28 32 36 40 44 48 52 56 60
Tot
al n
umbe
r of
folli
cles
3
mm
in d
iam
eter
/ova
ry
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
Age (wks)0 4 8 12 16 20 24 28 32 36 40 44 48 52 56 60
Max
imum
folli
cula
r di
amet
er (
mm
)
5
6
7
8
9
10
11
12
13
(a)
(b)
Fig. 2. Total number of follicles≥3 mm in diameter per ovary (a) and maximum follicle diameter (b), from 2 to 60weeks of age in heifers (mean±S.E.M.). Dashed lines (P < 0.05) or solid lines (P < 0.10) show major statisticaltrends. Major statistical trends delimit the periods during which there were progressive changes in mean values.

A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29 23
0 4 8 12 16 20 24 28 32 36 40 44 48 52 56 60
Ute
rine
diam
eter
(m
m)
8
10
12
14
16
18
20
22
24
Age (wks)
0 4 8 12 16 20 24 28 32 36 40 44 48 52 56 6010
15
20
25
30
100
120
140
160
Num
eric
al p
ixel
val
ues
Pix
el h
eter
ogen
eity
(a)
(b)
Fig. 3. Diameter (a), and numerical pixel values and pixel heterogeneity, or cumulative standard deviation of pixelbrightness, of the uterine body (b), from 2 to 60 weeks of age in heifers (mean± S.E.M.). Numerical pixel valuesrange from 0 (black) to 255 (white). Dashed lines (P < 0.05) or solid lines (P < 0.10) show major statisticaltrends. Major statistical trends delimit the periods during which there were progressive changes in mean values.
24 A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29
0 4 8 12 16 20 24 28 32 36 40 44 48 52 56 60
Cer
vix
diam
eter
(m
m)
4
6
8
10
12
14
16
18
20
22
24
Age (wks)
0 4 8 12 16 20 24 28 32 36 40 44 48 52 56 600
5
10
15
20
25
30
100
120
140
160
Num
eric
al p
ixel
val
ues
Pix
el h
eter
ogen
eity
(a)
(b)
Fig. 4. Diameter (a), and numerical pixel values and pixel heterogeneity, or cumulative standard deviation of pixelbrightness, of the cervix (b), from 2 to 60 weeks of age in heifers (mean± S.E.M.). Numerical pixel values rangefrom 0 (black) to 255 (white). Dashed lines show major statistical trends (P < 0.05). Major statistical trendsdelimit the periods during which there were progressive changes in mean values.
A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29 25
0 4 8 12 16 20 24 28 32 36 40 44 48 52 56 60
Vag
inal
dia
met
er (
mm
)
6
8
10
12
14
16
18
20
22
24
26
28
Age (wks)0 4 8 12 16 20 24 28 32 36 40 44 48 52 56 60
10
15
20
25
30120
140
160
180
200
Num
eric
al p
ixel
val
ues
Pix
el h
eter
ogen
eity
(a)
(b)
Fig. 5. Diameter (a), numerical pixel values and pixel heterogeneity, or cumulative standard deviation of pixelbrightness of the vagina (b), from 2 to 60 weeks of age in heifers (mean± S.E.M.). Numerical pixel values rangefrom 0 (black) to 255 (white). Dashed lines (P < 0.05) or solid lines (P < 0.10) show major statistical trends.Major statistical trends delimit the periods during which there were progressive changes in mean values.

26 A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29
Table 1Pearson product–moment correlation coefficients (r) between the uterine, cervical and vaginal diameters, and thenumber (allP < 0.10) or maximum diameter (allP < 0.01) of ovarian antral follicles in pre-pubertal heifers
Uterine diameter Cervical diameter Vaginal diameter
Number of follicles≥3 mm in diameter 0.37 0.36 0.33Maximum follicle diameter 0.66 0.64 0.68
antral follicles (≥3 mm in size) were also positively correlated with diameter of the uterus,cervix and vagina, but correlation coefficients were lower (P < 0.10;Table 1).
4. Discussion
In the present study, ovarian dimensions (Fig. 1a) increased rapidly to 14 weeks of agewith a more gradual increase from approximately 34 weeks of age to first ovulation. Earlystudies using slaughter data, showed a rapid increase in ovarian weight to 60–80 days of age(Erickson, 1966) or 5 months of age (Desjardins and Hafs, 1969), but in a marked contrast tothe present study, a plateau through the time of first ovulation (Desjardins and Hafs, 1969).In the heifers of the present study, dimensions collected by ultrasonography excluded largesuperficial follicles that distorted the ovarian profile; therefore, increases with time wouldhave reflected ovarian stromal development and increases in the numbers of and size ofsmall vesicular follicles.
The developmental pattern of numbers of ovarian antral follicles≥3 mm in diameter andmaximum follicle size tended to mirror the pattern for ovarian dimensions, with significantincreases early in life and in the later phase of development prior to first ovulation (Fig. 2aand b). Again, based on monthly observations of post-mortem samples, an early increasein follicle numbers to 60–80 days of age was seen in the study ofErickson (1966), butthe peak occurred later (5 months of age) in the Holstein calves in the study ofDesjardinsand Hafs (1969). In the latter study, and again in contrast to the present results, no furtherincrease in follicle numbers was seen prior to first ovulation. In a previous study, usingultrasonography for 18 days at 2, 8 and 14 weeks of age, a clear early increase in numbersof follicles≥3 mm diameter and maximum follicle diameter was seen (Evans et al., 1994a).This early increase in follicle growth probably reflects the high serum FSH concentrationsand the transient increase in LH secretion, as seen in the present calves between 4 and20 weeks of age (Honaramooz et al., 1999). Interestingly, superovulatory responses togonadotropins increase in young calves over this same time period (Seidel et al., 1971;Armstrong et al., 1992). Prior to first ovulation, a significant increase in size of dominantfollicles was observed in ultrasonographically monitored heifers (Bergfelt et al., 1994). LHpulse frequency increases as heifers approach first ovulation (Day et al., 1984).
Ovarian numerical pixel values appeared to decrease during the early rapid phase ofovarian development (Fig. 1b). The spots within which pixels were measured were placedrandomly on the ovarian images but care was taken to avoid the antra of obvious follicles.The decrease in numerical pixel may reflect increased growth of small follicles and bloodvessels (angiogenesis). In the present case, the decreases in numerical pixel values most

A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29 27
likely reflect increases in follicle numbers, assuming that numbers of small follicles (≥3 mmin diameter) parallel the trend seen for larger follicles inFig. 2a. This relationship requireshistological verification but, if proven, would further demonstrate the usefulness of ovarianimage analysis in determining the changes in numbers of small antral follicles, or thegrowing follicle pool (Singh et al., 1998; Tom et al., 1998), specifically.
A biphasic pre-pubertal growth pattern, similar to that noted for the ovary, was seen forthe uterus, cervix and vagina (Figs. 3a, 4a and 5a). This is in marked contrast to the find-ings of an earlier study, when weights of the uterus, vagina and cervix were recorded frompost-mortem reproductive tracts collected monthly from Holstein heifers over the first yearof life (Desjardins and Hafs, 1969). In that study, uterine weights increased slowly but lin-early from birth to first ovulation, and vaginal and cervical weights increased slowly at firstwith a somewhat more rapid growth from 4 months of age to first ovulation (Desjardins andHafs, 1969). In the present study, there was a strong correlation between the development ofthe segments of the reproductive tract with the pattern of changes in ovarian maximum fol-licle diameter (Table 1). Larger follicles would be more estrogenic, suggesting that periodsof gonadotropin-driven accelerated follicle growth resulted in estrogen-driven increases ingrowth of the reproductive tract. The reason for the biphasic nature of ovarian and tubu-lar reproductive tract development remains unclear. It has been suggested that increasedgonadotropin secretion (particularly LH) in the early post-natal heifer, may reflect matura-tional changes in the regulation of gonadotropin secretion prior to suppression by estrogen(Evans et al., 1994a,b). Subsequently, reduction in the negative feedback suppression of LHsecretion and another increase in follicle estrogenic capacity may precede and herald thepubertal ovulation in heifers (Day et al., 1984, 1987).
As for the ovary, there was a tendency for numerical pixel values of uterine, cervicaland vaginal images to decrease during phases of rapid growth. However, the most fasci-nating aspect of the ultrasonographic image analysis for the reproductive tract was pixelheterogeneity (Figs. 3b, 4b and 5b). Pixel heterogeneity increased transiently toward theend of the initial post-natal growth of the uterus, cervix and vagina (most notably for theuterus). This could reflect a developmental stage at which marked differentiation occurs,with increased vascularity and/or secretory activity (i.e. changes in numbers and size ofglands). It would be interesting to characterize changes in the tubular genitalia histologi-cally, during pre-determined periods of change in pixel heterogeneity, to see if this aspectof ultrasonographic image analysis could pinpoint particular stages of development anddifferentiation.
5. Conclusions
In conclusion, using repeated ultrasonographic examination and image analysis every 2weeks in the same group of calves, allowed us to create a clear picture of the developmentof the reproductive system in pre-pubertal heifers. There were marked growth and develop-mental changes of the entire reproductive tract in the first 3–4 months of age, followed bya period of plateau or slow growth, and a second phase of accelerated growth prior to firstovulation. Our observations lead us to conclude that two periods of gonadotropin-drivenenhanced follicle growth and, possibly, estrogenicity may stimulate this biphasic pattern

28 A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29
of development of the reproductive organs in pre-pubertal heifers. Ultrasonographic imageanalysis revealed changes in numerical pixel values and heterogeneity that might reflectimportant phases of growth and differentiation of the reproductive system. It would now bejustified and interesting to clearly delineate the basis for these changes with parallel ultra-sonographic and histological studies. Such investigations would be of value for subsequentnon-invasive studies of sexual development and its control.
Acknowledgements
We thank Susan Cook for her excellent technical assistance, Bill Kerr and his staff forcare of the animals, NIDDK and USDA for assay reagents and NSERC for funding thisstudy.
References
Adams, G.P., 1999. Comparative patterns of follicle development and selection in ruminants. J. Reprod. Fertil. 54,17–32.
Adams, G.P., Evans, A.C.O., Rawlings, N.C., 1994. Follicular waves and circulating gonadotrophins in8-month-old prepubertal heifers. J. Reprod. Fertil. 100, 27–33.
Ahmad, N., Townsend, E.C., Dailey, R.A., Inskeep, E.K., 1997. Relationships of hormonal patterns and fertilityto occurrence of two or three waves of ovarian follicles, before and after breeding, in beef cows and heifers.Anim. Reprod. Sci. 49, 13–28.
Armstrong, D.T., Holm, P., Irvine, B., Petersen, B.A., Stubbings, R.B., McLean, D., Stevens, G., Seamark, R.F.,1992. Pregnancies and live birth from in vitro fertilization of calf oocytes collected by laparoscopic follicularaspiration. Theriogenology 38, 667–678.
Bergfelt, E.G.M., Kojima, F.N., Cupp, A.S., Wehrman, M.E., Peters, K.E., Garcia-Winder, M., Kinder, J.E., 1994.Ovarian follicular development in prepubertal heifers is influenced by level of dietary energy intake. Biol.Reprod. 51, 1046–1050.
Chandolia, R.K., Honaramooz, A., Omeke, B.C., Pierson, R., Beard, A.P., Rawlings, N.C., 1997. Assessment ofdevelopment of the reproductive tract by ultrasonography in bull calves and associated endocrine changes.Theriogenelogy 48, 119–132.
Desjardins, C., Hafs, H.D., 1969. Maturation of bovine female genitalia from birth through puberty. J. Anim. Sci.28, 502–507.
Day, M.L., Imakawa, K., Garcia-Winder, M., Zalesky, D.D., Schanbacher, B.D., Kittok, R.J., Kinder, J.E., 1984.Endocrine mechanisms of puberty in heifers: estradiol negative feedback regulation of luteinizing hormonesecretion. Biol. Reprod. 31, 332–341.
Day, M.L., Imakawa, K., Wolf, P.L., Kittok, R.J., Kinder, J.E., 1987. Endocrine mechanisms of puberty in heifers:role of hypothalamo-pituitary estradiol receptors in the negative feedback of estradiol on luteinizing hormonesecretion. Biol. Reprod. 37, 1054–1065.
Dyer, R.M., Bishop, M.D., Day, M.L., 1990. Exogenous estradiol reduces inhibition of luteinizing hormone byoestradiol in prepubertal heifers. Biol. Reprod. 42, 755–761.
Erickson, B.H., 1966. Development and senescence of the postnatal bovine ovary. J. Anim. Sci. 25, 800–805.Evans, A.C.O., Currie, W.D., Rawlings, N.C., 1992. Effects of naloxone on circulating gonadotrophin
concentrations in prepubertal heifers. J. Reprod. Fertil. 96, 847–855.Evans, A.C.O., Adams, G.P., Rawlings, N.C., 1994a. Follicular and hormonal development in prepubertal heifers
from 2 to 36 weeks of age. J. Reprod. Fertil. 102, 463–470.Evans, A.C.O., Adams, G.P., Rawlings, N.C., 1994b. Endocrine and ovarian follicular changes leading up to the
first ovulation in prepubertal heifers. J. Reprod. Fertil. 100, 187–194.

A. Honaramooz et al. / Animal Reproduction Science 80 (2004) 15–29 29
Evans, A.C.O., Pierson, R.A., Garcia, A., McDougall, L.M., Hrudka, F., Rawlings, N.C., 1996. Changes incirculating hormone concentrations, testes histology and testes ultrasonography during sexual maturation inbeef bulls. Theriogenology 46, 345–357.
Ginther, O.J., Wiltbank, M.C., Fricke, P.M., Gibbons, J.R., Kot, K., 1996. Selection of the dominant follicle incattle: minireview. Biol. Reprod. 55, 1187–1194.
Honaramooz, A., Chandolia, R.K., Beard, A.P., Rawlings, N.C., 1999. Effects of season of birth on the prepubertalpattern of gonadotrophin secretion and age at puberty in beef heifers. Theriogenology 52, 67–79.
Kinder, J.E., Day, M.L., Kittok, R.J., 1987. Endocrine regulation of puberty in cows and ewes. J. Reprod. Fertil.Suppl. 34, 167–186.
Melvin, E.J., Lindsey, B.R., Quintal-Franco, J., Zanella, E., Fike, K.E., van Tassell, C.P., Kinder, J.E., 1999.Circulating concentrations of oestradiol, luteinizing hormone, and follicle-stimulating hormone during wavesof ovarian follicular development in prepubertal cattle. Biol. Reprod. 60, 405–412.
Pierson, R.A., Adams, G.P., 1995. Computer-assisted image analysis, diagnostic ultrasonography and ovulationinduction: strange bedfellows. Theriogenology 43, 105–112.
Pierson, R.A., Ginther, O.J., 1988. Ultrasonographic imaging of the ovaries and uterus in cattle. Theriogenology29, 21–38.
Savio, J.D., Keenan, L., Boland, M.P., Roche, J.F., 1988. Pattern of growth of dominant follicles during the oestrouscycle of heifers. J. Reprod. Fertil. 83, 663–671.
Schams, D., Schallenberger, E., Gome, S., Karg, H., 1981. Endogenous patterns associated with puberty in maleand female cattle. J. Reprod. Fertil. Suppl. 30, 103–110.
Seidel, G.E., Larson, L.L., Foote, R.H., 1971. Effects of age and gonadotropin treatment on superovulation in thecalf. J. Anim. Sci. 33, 617–622.
Singh, J., Pierson, R.A., Adams, G.P., 1998. Ultrasound image attributes of the bovine corpus luteum: structuraland functional correlates. J. Reprod. Fertil. 109, 35–44.
Sirois, J., Fortune, J.E., 1988. Ovarian follicular dynamics during the estrous cycle in heifers monitored by real-timeultrasonography. Biol. Reprod. 39, 308–317.
Tom, J.W., Pierson, R.A., Adams, G.P., 1998. Quantitative echotexture analysis of bovine ovarian follicles.Theriogenology 50, 339–346.
Copyright © 2022 FDOKUMEN




















