
Tyrosine phosphorylation of EGF-R and PDGF-R proteins during acute cutaneous wound healing process...
9
Tyrosine phosphorylation of EGF-R and PDGF-R proteins during acute cutaneous wound healing process in mice Mohan L. Gope, PhD 1 ; Rajalakshmi Gope, PhD 2 1. Department of Biotechnology, City College, Bangalore, India, and 2. Department of Human Genetics, National Institute of Mental Health and Neurosciences (NIMHANS), Bangalore, India Reprint requests: Dr. Rajalakshmi Gope, PhD, Department of Human Genetics, National Institute of Mental Health and Neurosciences (NIMHANS), Bangalore 560 029, India. Tel: 191 80 2699 5125; Fax: 191 80 2656 4830; Email: [email protected] Manuscript received: May 14, 2008 Accepted in final form: August 29, 2008 DOI:10.1111/j.1524-475X.2008.00443.x ABSTRACT The effect of topical application of epidermal growth factor (EGF) and platelet- derived growth factors (PDGFs) on the levels of EGF-R and PDGF-R proteins and their tyrosine phosphorylation were analyzed during an acute cutaneous wound healing process in mice. The growth factor-treated wounds had optimum levels of receptor proteins as early as day 1 compared with the control, which had only a basal level. Analysis of the tyrosine phosphorylation of the receptor pro- teins in control and growth factor-treated wounds indicated that they were phos- phorylated until day 5 after wounding. Only the mature forms of a-PDGF-R and b-PDGF-R proteins were phosphorylated and not their precursors. Our results show that rapid attainment of maximum levels of growth factor receptor proteins and their tyrosine phosphorylation as early as day 1 and the maintenance of the same until day 3 appear to aid faster and better wound healing. Topical applica- tion of PDGF-AA alone did not facilitate the wound healing process and it also antagonized the EGF-medicated wound healing when applied premixed with EGF or within 30 minutes after EGF application. Under these conditions, the receptor proteins were not phosphorylated. Thus, an increased and sustained level of EGF-R and PDGF-R proteins and their tyrosine phosphorylation ap- pear to accelerate the wound healing process. Cutaneous wound healing is a complex, remarkably struc- tured biological process that involves several methodical steps such as granulation, wound closure, and re- epithelialization that are mediated by several endoge- nously released growth factors. 1,2 Many growth factors such as epidermal growth factor (EGF), platelet-derived growth factors (PDGFs), fibroblast growth factor (FGF), and transforming growth factor-b (TGF-b) are released at the wound site, which are absent in the normal skin. Chronic wounds were reported to contain decreased levels of PDGFs and EGF compared with acute wounds. Such deficiencies were suggested to be at least partly responsible for impaired wound healing. 3–8 Therefore, the effects of various growth factors either alone or in combination have been tested in cell culture and animal models and their positive effects on wound healing have been established beyond doubt. Some of these growth factors have been implicated in acceleration of the wound healing process in various tissues including the skin. Among these growth factors, EGF and PDGFs are reported to be important as they promote cell proliferation and differentiation at the wound site. 3–8 EGF has been shown to be necessary for reepithelialization during the wound healing process. 3–5 Reduced expressions of PDGFs and PDGF-R were observed during the early phases of impaired wound healing. 9–12 In addition, topical application of PDGF is reported to enhance wound repair, especially in animals with wound healing defects. 9–14 Previous studied have shown that the growth factor- treated wound healed faster compared with the untreated controls. 15–21 We have shown that EGF and PDGF- treated acute cutaneous wounds healed in approximately 5–7 days, whereas the control wounds healed in 10–15 days. Application of combination of these two growth fac- tors further accelerated the normal wound healing pro- cess. 22 Topical application of PDGF-AA alone had a negative effect and it also antagonized the positive effects of EGF during the cutaneous wound healing process. 22 We have also shown that the topical application of the op- timum concentration of EGF, PDGF-AB, and PDGF-BB individually or in combination led to a two- to four-fold increase in the coordinated expressions of EGF-R, a-PDGF-R, and b-PDGF-R mRNA levels. However, PDGF-AA, either alone or in combination with EGF, in- hibited the EGF-mediated increase in the receptor mRNA levels. 23 The present study was carried out to determine the effects of EGF and PDGFs on the level of expression and phosphorylation of the EGF-R and PDGF-R proteins during acute cutaneous wound healing process. EGF Epidermal growth factor EGF-R Epidermal growth factor receptor FGF Fibroblast growth factor PDGF Platelet-derived growth factor PDGF-AA Platelet-derived growth factor A chain homodimer PDGF-AB Heterodimer of PDGF A and B chain PDGF-BB Platelet-derived growth factor B chain homodimer PDGF-R Platelet-derived growth factor receptor Wound Rep Reg (2009) 17 71–79 c 2009 by the Wound Healing Society 71 Wound Repair and Regeneration
Transcript of Tyrosine phosphorylation of EGF-R and PDGF-R proteins during acute cutaneous wound healing process...

Tyrosine phosphorylation of EGF-R and PDGF-R proteinsduring acute cutaneous wound healing process in mice
Mohan L. Gope, PhD1; Rajalakshmi Gope, PhD2
1. Department of Biotechnology, City College, Bangalore, India, and
2. Department of Human Genetics, National Institute of Mental Health and Neurosciences (NIMHANS), Bangalore, India
Reprint requests:Dr. Rajalakshmi Gope, PhD, Department of
Human Genetics, National Institute of
Mental Health and Neurosciences
(NIMHANS), Bangalore 560 029, India.
Tel: 191 80 2699 5125;
Fax: 191 80 2656 4830;
Email: [email protected]
Manuscript received: May 14, 2008
Accepted in final form: August 29, 2008
DOI:10.1111/j.1524-475X.2008.00443.x
ABSTRACT
The effect of topical application of epidermal growth factor (EGF) and platelet-derived growth factors (PDGFs) on the levels of EGF-R and PDGF-R proteinsand their tyrosine phosphorylation were analyzed during an acute cutaneouswound healing process in mice. The growth factor-treated wounds had optimumlevels of receptor proteins as early as day 1 compared with the control, which hadonly a basal level. Analysis of the tyrosine phosphorylation of the receptor pro-teins in control and growth factor-treated wounds indicated that they were phos-phorylated until day 5 after wounding. Only the mature forms of a-PDGF-R andb-PDGF-R proteins were phosphorylated and not their precursors. Our resultsshow that rapid attainment of maximum levels of growth factor receptor proteinsand their tyrosine phosphorylation as early as day 1 and the maintenance of thesame until day 3 appear to aid faster and better wound healing. Topical applica-tion of PDGF-AA alone did not facilitate the wound healing process and it alsoantagonized the EGF-medicated wound healing when applied premixed withEGF or within 30 minutes after EGF application. Under these conditions, thereceptor proteins were not phosphorylated. Thus, an increased and sustainedlevel of EGF-R and PDGF-R proteins and their tyrosine phosphorylation ap-pear to accelerate the wound healing process.
Cutaneous wound healing is a complex, remarkably struc-tured biological process that involves several methodicalsteps such as granulation, wound closure, and re-epithelialization that are mediated by several endoge-nously released growth factors.1,2 Many growth factorssuch as epidermal growth factor (EGF), platelet-derivedgrowth factors (PDGFs), fibroblast growth factor (FGF),and transforming growth factor-b (TGF-b) are released atthe wound site, which are absent in the normal skin.Chronic wounds were reported to contain decreased levelsof PDGFs and EGF compared with acute wounds. Suchdeficiencies were suggested to be at least partly responsiblefor impaired wound healing.3–8 Therefore, the effects ofvarious growth factors either alone or in combination havebeen tested in cell culture and animal models and theirpositive effects on wound healing have been establishedbeyond doubt. Some of these growth factors have beenimplicated in acceleration of the wound healing process invarious tissues including the skin. Among these growthfactors, EGF and PDGFs are reported to be important asthey promote cell proliferation and differentiation at thewound site.3–8 EGF has been shown to be necessary forreepithelialization during the wound healing process.3–5
Reduced expressions of PDGFs and PDGF-R wereobserved during the early phases of impaired woundhealing.9–12 In addition, topical application of PDGF isreported to enhance wound repair, especially in animalswith wound healing defects.9–14
Previous studied have shown that the growth factor-treated wound healed faster compared with the untreatedcontrols.15–21 We have shown that EGF and PDGF-
treated acute cutaneous wounds healed in approximately5–7 days, whereas the control wounds healed in 10–15days. Application of combination of these two growth fac-tors further accelerated the normal wound healing pro-cess.22 Topical application of PDGF-AA alone had anegative effect and it also antagonized the positive effectsof EGF during the cutaneous wound healing process.22
We have also shown that the topical application of the op-timum concentration of EGF, PDGF-AB, and PDGF-BBindividually or in combination led to a two- to four-foldincrease in the coordinated expressions of EGF-R,a-PDGF-R, and b-PDGF-R mRNA levels. However,PDGF-AA, either alone or in combination with EGF, in-hibited the EGF-mediated increase in the receptor mRNAlevels.23 The present study was carried out to determine theeffects of EGF and PDGFs on the level of expression andphosphorylation of the EGF-R and PDGF-R proteinsduring acute cutaneous wound healing process.
EGF Epidermal growth factor
EGF-R Epidermal growth factor receptor
FGF Fibroblast growth factor
PDGF Platelet-derived growth factor
PDGF-AA Platelet-derived growth factor A chain homodimer
PDGF-AB Heterodimer of PDGF A and B chain
PDGF-BB Platelet-derived growth factor B chain homodimer
PDGF-R Platelet-derived growth factor receptor
Wound Rep Reg (2009) 17 71–79 c� 2009 by the Wound Healing Society 71
Wound Repair and Regeneration

MATERIALS AND METHODS
Animals
The institutional Animal Ethics Committee cleared the ex-perimental protocol. Random bred Swiss albino mice wereused in this study. Three-week-old mice were obtainedfrom the Central Animal Research Facility, National In-stitute of Mental Health and Neurosciences (NIMHANS),Bangalore, and kept in the experimental room at least foran additional 2–3 weeks before use. This time period as-sured that the animals were free of any diseases. All theanimals were provided pelleted animal food (Godrej IndiaLtd., Mumbai, India) and water ad libitum, and were sub-jected to a normal day–night light cycle.
Wounding
The wounds were created as described previously.22,23 Thedorso-lateral sides of the mice were cleaned with a disin-fectant, and an approximately 5 mm�1.5 cm area wascleared of all hair. Each mouse was individually anesthe-tized with ether. Two full-thickness wounds approximately1 cm long and 0.3–0.4 cm deep were created on the dorso-lateral sides of each mouse.22,23
Number of mice used and sample collection
The experiments were performed as described previouslyand they were repeated thrice to obtain statistically signifi-cant data.22,23 Fifty mice were kept as external controls,where the wounds were left untreated. A second group of50 mice were used as additional control, where one side ofthe wound was treated with the diluent alone and thewound on the other side was left untreated. These two setsof animals were external controls. In the third set of 50mice, the wound on one side was treated with the growthfactor and the wound on the other side was treated with adiluent alone, which acted as an internal control. Thus, agroup of 50 mice was used for each of the following growthfactor treatments: (1) EGF, (2) PDGF-AB, (3) PDGF-BB,(4) PDGF-AA, (5) EGF 1 PDGF-BB, (6) EGF 1 PDGF-AB, (7) EGF 1 PDGF-AA either premixed or the EGFwas applied first, followed by PDGF-AA applicationwithin 30 minutes, and (8) EGF 1 PDGF-AA (300), wherethe EGF was applied first, followed by PDGF-AA appli-cation after 30 minutes. The newly formed granular massesof tissues at the wound sites were collected and pooledfrom approximately eight to 10 mice for each time point,that is, days 1, 3, 5, and 7. After the tissue sample collec-tion, the mice were sacrificed and disposed. The remainingmice were kept under observation for approximately30 days for any abnormal growth or rash. Thus, for eachexperiment, a total of 100 mice were used as external con-trol and 400 mice were used for growth factor application.In total 1,500 mice were used for three separate sets ofexperiments.22,23
Growth factors
Recombinant mouse EGF and recombinant humanPDGF-AB, PDGF-BB, and PDGF-AA were obtainedfrom Sigma-Aldrich (St. Louis, MO) as lyophilized pow-
ders. They were diluted in a solution containing 4mMHCl and 0.1% (w/v) bovine serum albumin (BSA) accord-ing to the manufacturer’s instructions (Sigma-Aldrich).Aliquots of appropriate volumes were stored at �20 1Cand they were thawed only once for each use as describedpreviously.22,23
Application of growth factors
The day of wound creation was counted as day 0 and thenext day as 1. Optimum concentrations of the growth fac-tors (EGF, 10 ng; PDGF-AB, 15 ng; PDGF-BB, 15 ng;and PDGF-AA, 20 ng) were applied directly to the woundseach day starting from days 0 to 7. The growth factorswere applied to the wound with a micropipette as de-scribed previously.22,23 For the studies using a combina-tion of growth factors, EGF was applied first, followed byapplication of PDGF-AB or PDGF-BB or PDGF-AA.Alternatively, the growth factors were premixed and ap-plied to the wounds. In a separate set of experimentsPDGF-AA was applied 30 minutes after the EGF applica-tion.22,23 After wounding and application of growth fac-tors the animals were caged separately.
Monitoring
The time taken for wound repair was scored visually andphotographically using the following parameters: (a)wound closure and (b) scar tissue formation, either fine orraised, as described previously.22,23 The newly formed ir-regular masses of granular tissues were collected from thewound site of control and experimental mice. These tissueswere pooled from eight to 10 mice and stored at �70 1Cuntil they were processed.22,23
Preparation of total cell lysate
The granular wound tissues were suspended in approxi-mately 100–150mL of lysis buffer containing 10 mM Tris-HCl, pH 7.0, 150 mM NaCl, 5 mM EDTA, 1.2% TritonX-100, 2 mM phenyl methyl sulfonyl fluoride (PMSF),0.15 U/mL aprotinin, and 10mg/mL leupeptin (Sigma-Aldrich). The tissues were homogenized on ice using polyt-ron, 30 seconds �5, with a 1-minute cooling period in be-tween each mixing cycle. The lysates were clarified bycentrifugation at 8,000�g for 10 minutes. The clear super-natants were transferred to fresh tubes, and the protein con-centrations were measured using a spectrophotometer.24,25
Protein tyrosine phosphatase treatment of the
tissue lysates
The activity of the leukocyte antigen-related protein(LAR-D1 protein) tyrosine phosphatase (Sigma-Aldrich)was first confirmed using the commercially available pro-tein tyrosine phosphatase substrate (Sigma-Aldrich) ac-cording to the manufacturer’s protocol. One hundred andfifty micrograms of the total cell lysate was treated with1–2U of LAR-D1 protein tyrosine phosphatase or serine/threonine phosphatase (Sigma-Aldrich) at 30 1C for1 hour. The reaction was stopped by the addition of5 mL of loading dye containing 12% sodium dodecyl sul-fate, 0.6 mM DTT, 30 mM Tris-HCl, pH 6.8, 0.6%
Wound Rep Reg (2009) 17 71–79 c� 2009 by the Wound Healing Society72
Protein phosphorylation during wound healing Gope et al.

bromophenol blue, and 60% glycerol, denatured by boil-ing for 2 minutes, and loaded onto 7.5% sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE)gels.24,25
In separate reactions, 2mL of a 10-fold diluted phos-phatase inhibitor cocktail 1, which inhibits serine/threo-nine protein phosphatase or cocktail 2, which inhibitstyrosine phosphatase (Sigma-Aldrich) was first added tothe 20mL tissue lysate before the addition of thecorresponding phosphatase. This ensured that the fastermobility of the growth factor receptor bands from thetyrosine phosphatase-treated tissue lysates was due todephosphorylation of the tyrosine residue in the receptorproteins.
Western blot analysis
Western blot analysis was carried out essentially as de-scribed previously.24,25 One hundred and fifty microgramof each lysate was denatured, separated on to a 7.5% SDS-PAGE gel, and transferred to a PVDF membrane (Sigma-Aldrich) using a semidry blotter (Bio-Rad, Hercules, CA).The nonspecific sites on the membranes were blocked witha buffer containing BSA. The membranes were incubatedwith 2–5mg of polyclonal primary antibody to EGF-R ora-PDGF-R or b-PDGF-R for 2 hours. The EGF-R anti-body, Ab-6, was obtained from CN Biosciences Inc. (SanDiego, CA). The a-PDGF-R and b-PDGF-R antibodiesraised against the kinase domain were purchased from BDPharmingen (Singapore). All the primary antibodies usedin this study recongnized mouse as well as human receptorproteins. The primary antibody to the a-PDGF-R isknown not to cross react with the b-PDGF-R and viceversa. The secondary antibody, AP-conjugated protein A(Calbiochem, Nottingham, UK), was diluted 1 : 1,000 andthe membranes were incubated with the diluted secondaryantibody for an additional 1 hour at room temperaturewith shaking. The membranes were washed and the colorwas developed with 1-Step NBT/BCIP (Pierce, Rockford,IL) according to the manufacturer’s protocol. The bandswere photographed with the Bio-Rad Gel-Doc system,and the band intensities were measured using the Bio-Radquantity one software. The wide-range color marker(Sigma-Aldrich) was used to determine the molecularweight of the receptor protein bands. Separate sets of gelscontaining the electrophoretically separated tissue lysateswere hybridized with the b-tubulin primary antibody (BDPharmingen) at a concentration of 2–5mg/mL and second-ary antibody, followed by color development as described.The b-tubulin band intensities were measured and thesevalues were used as internal sample loading controls foreach lane.
Data analysis
The entire experiments, including wounding, applicationof growth factors, collection of tissues, and Western blots,were repeated three times and the mean values were calcu-lated. Only one representative blot from each experimentis presented here. The Bio-Rad quantity one softwarewas used to measure the band intensities from Westernblots.24,25 The value on a blank lane was assigned a valueof zero. The receptor protein band intensities from
the controls and the growth factor-treated wounds weremeasured and the blank value was subtracted. Further,these values were normalized to the band intensity of thecorresponding b-tubulin band intensity, which was used asthe loading control. Thus, the actual band intensities forthe control and the diluent-treated wounds were calcu-lated, which acted as the basal level. The untreated and thediluent-treated controls showed similar values in all thethree separate experiments. Similarly, the band intensitiesof the experimental wounds were calculated and reportedhere as a fold increase as compared with the basallevels obtained for the controls. The a-PDGF-R and theb-PDGF-R antibodies recognized the known two bandseach, and the intensities of both the bands were added andreported here as fold increase for these receptor proteins.
Statistical analysis was carried out with SPSS softwareversion 15.0. Repeated measures and analysis of variantswere performed, followed by post hoc tests for comparisonacross the groups with Bonferroni’s corrections. The meanvalue and standard deviations from three separate exper-iments were calculated and are reported in the Table 1.Statistical significance was fixed at 0.05 levels.
RESULTS
Effects of EGF and PDGFs on the levels of EGF-R and
PDGF-R proteins
Topical application of wounds with EGF and PDGFseither individually or in combination resulted in acoordinated up-regulation of EGF-R, a-PDGF-R, andb-PDGF-R proteins. Consequently, topical application ofEGF resulted in the up-regulation of a-PDGF-R andb-PDGF-R proteins in addition to EGF-R protein. Simi-larly, the topical application of PDGF-AB and PDGF-BBresulted in the up-regulation of EGF-R protein in additionto a-PDGF-R and b-PDGF-R proteins. This up-regula-tion occurred as early as day 1 only in the growth factor-treated wounds and not in the control (Figures 1 and 2;Table 1).
The EGF-R antibody recognized a single band of185 kDa protein. Application of EGF or PDGF-AB orPDGF-BB resulted in a 3.5–4.0-fold increase in the level ofthe EGF-R protein as compared with the control (Figure1; Table 1). EGF and PDGF-AB applications produced apeak level of EGF-R protein on day 1 followed by a grad-ual decline reaching the control level on day 7. Applicationof PDGF-BB produced a 2.5-fold increase of EGF-Ron day 1; however, the peak level of 4-fold was reachedonly on day 3 in all the three separate experiments con-ducted (Figure 1; Table 1).
The a-PDGF-R antibody recognized a mature receptorprotein of 170 kDa and a precursor of 160 kDa (Figure 2).Application of EGF or PDGF-AB or PDGF-BB resultedin a 2.5–3.5-fold increase in the level of a-PDGF-R proteinon day 1. However, the maximum level, which was slightlymore than the initial increase, was observed on days 3, 5,and 1, respectively, for these growth factors. On day 7, thelevels were similar to those of the control for all threegrowth factors (Figure 2; Table 1).
The b-PDGF-R antibody recognized a 180 kDa maturereceptor protein and a precursor of 160 kDa (data not
Wound Rep Reg (2009) 17 71–79 c� 2009 by the Wound Healing Society 73
Protein phosphorylation during wound healingGope et al.
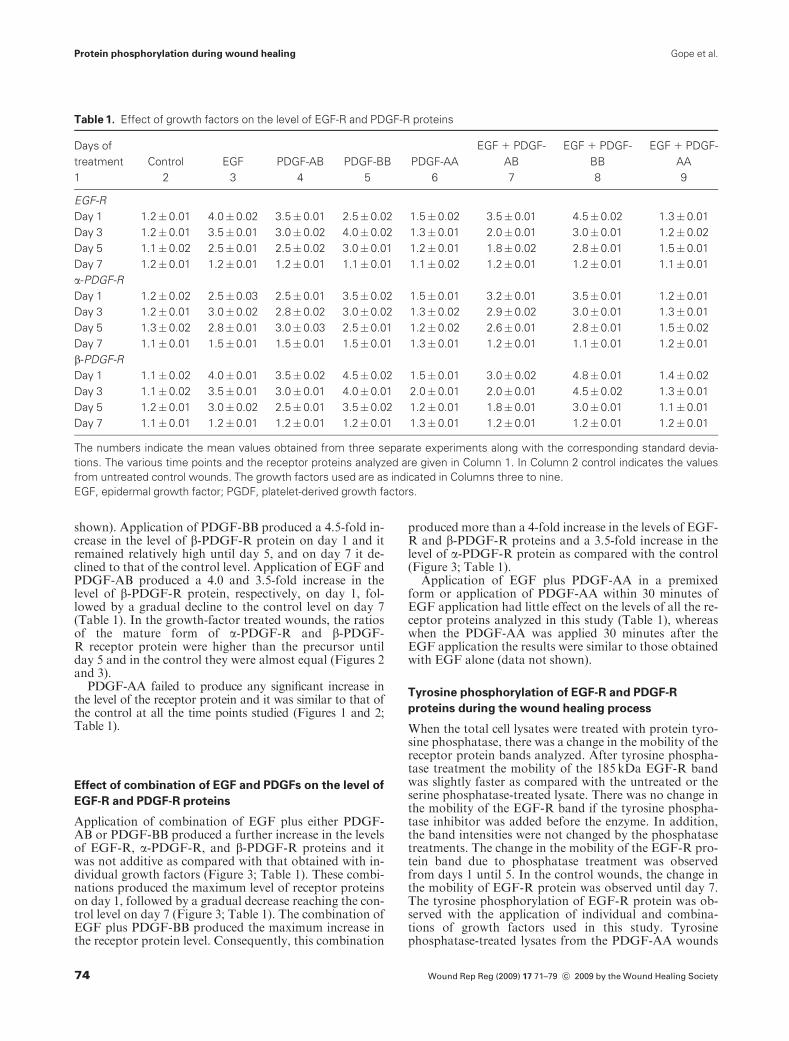
shown). Application of PDGF-BB produced a 4.5-fold in-crease in the level of b-PDGF-R protein on day 1 and itremained relatively high until day 5, and on day 7 it de-clined to that of the control level. Application of EGF andPDGF-AB produced a 4.0 and 3.5-fold increase in thelevel of b-PDGF-R protein, respectively, on day 1, fol-lowed by a gradual decline to the control level on day 7(Table 1). In the growth-factor treated wounds, the ratiosof the mature form of a-PDGF-R and b-PDGF-R receptor protein were higher than the precursor untilday 5 and in the control they were almost equal (Figures 2and 3).
PDGF-AA failed to produce any significant increase inthe level of the receptor protein and it was similar to that ofthe control at all the time points studied (Figures 1 and 2;Table 1).
Effect of combination of EGF and PDGFs on the level of
EGF-R and PDGF-R proteins
Application of combination of EGF plus either PDGF-AB or PDGF-BB produced a further increase in the levelsof EGF-R, a-PDGF-R, and b-PDGF-R proteins and itwas not additive as compared with that obtained with in-dividual growth factors (Figure 3; Table 1). These combi-nations produced the maximum level of receptor proteinson day 1, followed by a gradual decrease reaching the con-trol level on day 7 (Figure 3; Table 1). The combination ofEGF plus PDGF-BB produced the maximum increase inthe receptor protein level. Consequently, this combination
produced more than a 4-fold increase in the levels of EGF-R and b-PDGF-R proteins and a 3.5-fold increase in thelevel of a-PDGF-R protein as compared with the control(Figure 3; Table 1).
Application of EGF plus PDGF-AA in a premixedform or application of PDGF-AA within 30 minutes ofEGF application had little effect on the levels of all the re-ceptor proteins analyzed in this study (Table 1), whereaswhen the PDGF-AA was applied 30 minutes after theEGF application the results were similar to those obtainedwith EGF alone (data not shown).
Tyrosine phosphorylation of EGF-R and PDGF-R
proteins during the wound healing process
When the total cell lysates were treated with protein tyro-sine phosphatase, there was a change in the mobility of thereceptor protein bands analyzed. After tyrosine phospha-tase treatment the mobility of the 185 kDa EGF-R bandwas slightly faster as compared with the untreated or theserine phosphatase-treated lysate. There was no change inthe mobility of the EGF-R band if the tyrosine phospha-tase inhibitor was added before the enzyme. In addition,the band intensities were not changed by the phosphatasetreatments. The change in the mobility of the EGF-R pro-tein band due to phosphatase treatment was observedfrom days 1 until 5. In the control wounds, the change inthe mobility of EGF-R protein was observed until day 7.The tyrosine phosphorylation of EGF-R protein was ob-served with the application of individual and combina-tions of growth factors used in this study. Tyrosinephosphatase-treated lysates from the PDGF-AA wounds
Table 1. Effect of growth factors on the level of EGF-R and PDGF-R proteins
Days of
treatment
1
Control
2
EGF
3
PDGF-AB
4
PDGF-BB
5
PDGF-AA
6
EGF 1 PDGF-
AB
7
EGF 1 PDGF-
BB
8
EGF 1 PDGF-
AA
9
EGF-R
Day 1 1.2� 0.01 4.0� 0.02 3.5� 0.01 2.5� 0.02 1.5� 0.02 3.5� 0.01 4.5� 0.02 1.3� 0.01
Day 3 1.2� 0.01 3.5� 0.01 3.0� 0.02 4.0� 0.02 1.3� 0.01 2.0� 0.01 3.0� 0.01 1.2� 0.02
Day 5 1.1� 0.02 2.5� 0.01 2.5� 0.02 3.0� 0.01 1.2� 0.01 1.8� 0.02 2.8� 0.01 1.5� 0.01
Day 7 1.2� 0.01 1.2� 0.01 1.2� 0.01 1.1� 0.01 1.1� 0.02 1.2� 0.01 1.2� 0.01 1.1� 0.01
a-PDGF-R
Day 1 1.2� 0.02 2.5� 0.03 2.5� 0.01 3.5� 0.02 1.5� 0.01 3.2� 0.01 3.5� 0.01 1.2� 0.01
Day 3 1.2� 0.01 3.0� 0.02 2.8� 0.02 3.0� 0.02 1.3� 0.02 2.9� 0.02 3.0� 0.01 1.3� 0.01
Day 5 1.3� 0.02 2.8� 0.01 3.0� 0.03 2.5� 0.01 1.2� 0.02 2.6� 0.01 2.8� 0.01 1.5� 0.02
Day 7 1.1� 0.01 1.5� 0.01 1.5� 0.01 1.5� 0.01 1.3� 0.01 1.2� 0.01 1.1� 0.01 1.2� 0.01
b-PDGF-R
Day 1 1.1� 0.02 4.0� 0.01 3.5� 0.02 4.5� 0.02 1.5� 0.01 3.0� 0.02 4.8� 0.01 1.4� 0.02
Day 3 1.1� 0.02 3.5� 0.01 3.0� 0.01 4.0� 0.01 2.0� 0.01 2.0� 0.01 4.5� 0.02 1.3� 0.01
Day 5 1.2� 0.01 3.0� 0.02 2.5� 0.01 3.5� 0.02 1.2� 0.01 1.8� 0.01 3.0� 0.01 1.1� 0.01
Day 7 1.1� 0.01 1.2� 0.01 1.2� 0.01 1.2� 0.01 1.3� 0.01 1.2� 0.01 1.2� 0.01 1.2� 0.01
The numbers indicate the mean values obtained from three separate experiments along with the corresponding standard devia-
tions. The various time points and the receptor proteins analyzed are given in Column 1. In Column 2 control indicates the values
from untreated control wounds. The growth factors used are as indicated in Columns three to nine.
EGF, epidermal growth factor; PGDF, platelet-derived growth factors.
Wound Rep Reg (2009) 17 71–79 c� 2009 by the Wound Healing Society74
Protein phosphorylation during wound healing Gope et al.

showed no change in the mobility of EGF-R protein.Treatment of lysates with serine/threonine phosphatasedid not change the mobility of the EGF-R protein band(data not shown).
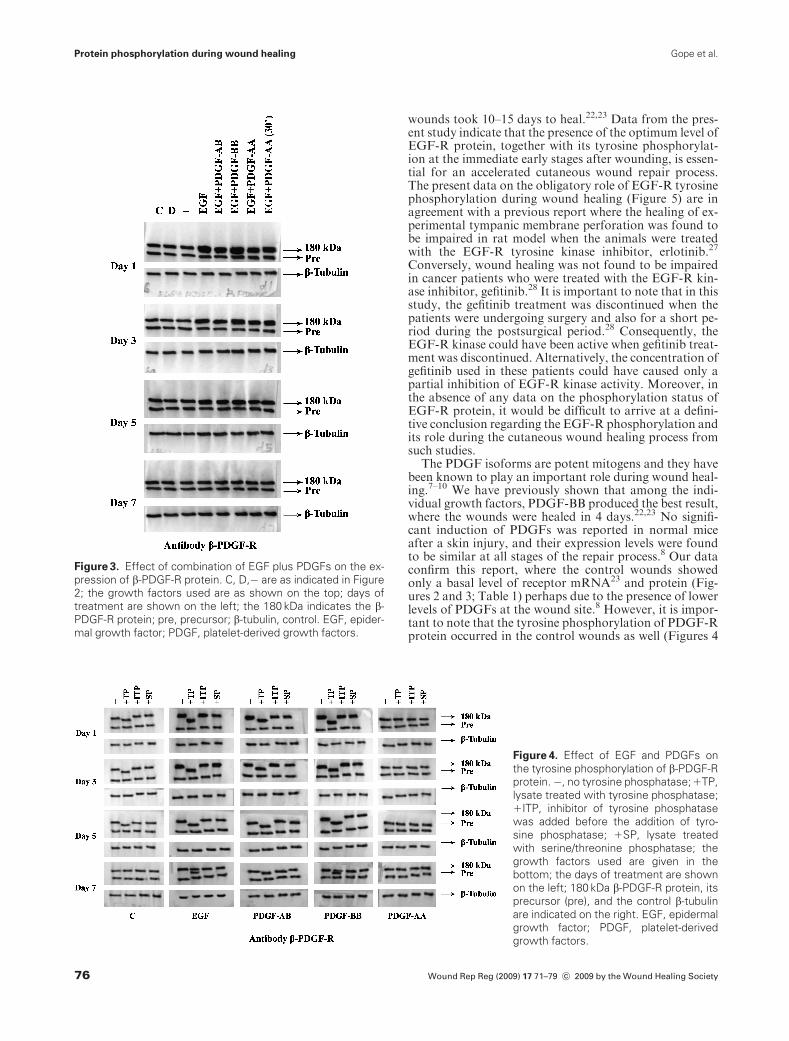
Upon tyrosine phosphatase treatment of the total celllysate, faster mobility was found only for the maturea-PDGF-R and b-PDGF-R protein bands and not for thecorresponding precursor bands, indicating that only the ma-ture forms of these receptor proteins are phosphorylated(Figures 4 and 5). Faster mobility of the mature a-PDGF-Rband was observed from days 1 until 5 in the control as wellas the growth factor-treated wounds. Application of EGFand PDGF-BB individually resulted in phosphorylation of atleast half of themature forms of b-PDGF-R protein on day 7(Figures 4 and 5). The combination of growth factorsshowed tyrosine phosphorylation of all the three receptorproteins until day 5 (Figure 5). Addition of an inhibitor oftyrosine phosphatase before the addition of enzyme or treat-ment with serine/threonine phosphatase did not result in anychange in the mobility of these protein bands (Figures 4 and5). Addition of an inhibitor of serine/threonine phosphatasebefore the addition of the serine/threonine phosphataseenzyme had no effect on the mobility of the a-PDGF-R andb-PDGF-R proteins (Figures 4 and 5).
The PDGF-AA treatment, either individually or incombination with EGF, did not show any significantchange in the mobility of the growth factor receptorbands. However, application of PDGF-AA, 30 minutesafter the EGF application, produced results similarto those obtained with EGF application alone (Figures 4and 5).
DISCUSSION
The positive effects of EGF and PDGF-BB on the exper-imental wounds have been shown by several in vitro andin vivo studies.15–20 EGF binding to its receptor is knownto result in receptor tyrosine phosphorylation, which isknown to play an important role in the downstream sig-naling of EGF-R.26 EGF-R activation has been shown tobe essential for proper epithelial wound healing in vivo.26
Previously, we have shown that daily topical application of10 ng of EGF accelerated acute cutaneous wound healingin a mice model.22,23 A 3.5 to 4-fold increase in the level ofEGF-R protein from days 1 to 3 after wounding and itstyrosine phosphorylation until day 5 (Figure 1; Table 1) isin accordance with our previous report, where we haveshown a similar increase in the level of EGF-R mRNA inthe EGF-treated wounds.23 The untreated and the diluent-treated control wounds had only a basal level of EGF-Rprotein (Figure 1; Table 1), but it is interesting to note thatthey also showed tyrosine phosphorylation (Figures 4 and5). Previously, we have shown that the wounds treatedwith EGF healed in 5 days, whereas the untreated control
Figure 2. Effect of EGF and PDGFs on the expression of
a-PDGF-R protein. C, D, �, as described in Figure 1; 1, growth
factor as indicated on the top; the days of growth factor treat-
ment are shown on the left; 170 kDa indicates the molecular
weight of a-PDGF-R protein band; b-tubulin control is as indi-
cated on the right. Pre, indicates the precursor of a-PDGF-R.
EGF, epidermal growth factor; PDGF, platelet-derived growth
factors.
Figure 1. Effect of topical application of growth factors EGF
and PDGFs on the expression of EGF-R. C, control (external
control); D, diluent (external control); �, no growth factor (inter-
nal control); 1, plus growth factor; the days of treatments are
given on the left; 185 kDa indicates the EGF-R protein; the
b-tubulin was used as a control. EGF, epidermal growth factor;
PDGF, platelet-derived growth factors.
Wound Rep Reg (2009) 17 71–79 c� 2009 by the Wound Healing Society 75
Protein phosphorylation during wound healingGope et al.
wounds took 10–15 days to heal.22,23 Data from the pres-ent study indicate that the presence of the optimum level ofEGF-R protein, together with its tyrosine phosphorylat-ion at the immediate early stages after wounding, is essen-tial for an accelerated cutaneous wound repair process.The present data on the obligatory role of EGF-R tyrosinephosphorylation during wound healing (Figure 5) are inagreement with a previous report where the healing of ex-perimental tympanic membrane perforation was found tobe impaired in rat model when the animals were treatedwith the EGF-R tyrosine kinase inhibitor, erlotinib.27
Conversely, wound healing was not found to be impairedin cancer patients who were treated with the EGF-R kin-ase inhibitor, gefitinib.28 It is important to note that in thisstudy, the gefitinib treatment was discontinued when thepatients were undergoing surgery and also for a short pe-riod during the postsurgical period.28 Consequently, theEGF-R kinase could have been active when gefitinib treat-ment was discontinued. Alternatively, the concentration ofgefitinib used in these patients could have caused only apartial inhibition of EGF-R kinase activity. Moreover, inthe absence of any data on the phosphorylation status ofEGF-R protein, it would be difficult to arrive at a defini-tive conclusion regarding the EGF-R phosphorylation andits role during the cutaneous wound healing process fromsuch studies.
The PDGF isoforms are potent mitogens and they havebeen known to play an important role during wound heal-ing.7–10 We have previously shown that among the indi-vidual growth factors, PDGF-BB produced the best result,where the wounds were healed in 4 days.22,23 No signifi-cant induction of PDGFs was reported in normal miceafter a skin injury, and their expression levels were foundto be similar at all stages of the repair process.8 Our dataconfirm this report, where the control wounds showedonly a basal level of receptor mRNA23 and protein (Fig-ures 2 and 3; Table 1) perhaps due to the presence of lowerlevels of PDGFs at the wound site.8 However, it is impor-tant to note that the tyrosine phosphorylation of PDGF-Rprotein occurred in the control wounds as well (Figures 4
Figure 3. Effect of combination of EGF plus PDGFs on the ex-
pression of b-PDGF-R protein. C, D,� are as indicated in Figure
2; the growth factors used are as shown on the top; days of
treatment are shown on the left; the 180 kDa indicates the b-
PDGF-R protein; pre, precursor; b-tubulin, control. EGF, epider-
mal growth factor; PDGF, platelet-derived growth factors.
Figure 4. Effect of EGF and PDGFs on
the tyrosine phosphorylation of b-PDGF-R
protein.�, no tyrosine phosphatase;1TP,
lysate treated with tyrosine phosphatase;
1ITP, inhibitor of tyrosine phosphatase
was added before the addition of tyro-
sine phosphatase; 1SP, lysate treated
with serine/threonine phosphatase; the
growth factors used are given in the
bottom; the days of treatment are shown
on the left; 180 kDa b-PDGF-R protein, its
precursor (pre), and the control b-tubulin
are indicated on the right. EGF, epidermal
growth factor; PDGF, platelet-derived
growth factors.
Wound Rep Reg (2009) 17 71–79 c� 2009 by the Wound Healing Society76
Protein phosphorylation during wound healing Gope et al.

and 5). The only difference between the control and thegrowth factor-treated wound is that the latter had opti-mum levels of receptor proteins as compared with thebasal level observed in the control (Table 1). Therefore, theprolonged time taken for healing of the control wounds(10–15 days) is due to the presence of suboptimal levels ofreceptor mRNAs23 and proteins in these wounds (Table1). Glucocorticoid treatment has been reported to cause asevere defect in wound repair, and it results in decreasedlevels of a-PDGF-R and b-PDGF-R in the early phase ofwound healing.8 Thus, the beneficial effect of topicallyapplied PDGFs observed by others7–10 and by us22,23
could be due to the ability of these growth factors to in-duce and sustain optimum levels of PDGF-R mRNA23
and protein (Table 1) as well as tyrosine phosphorylationof these receptor proteins at the immediate early stages ofthe wound-healing process (Figures 4 and 5).
Previously, it has been suggested that application ofEGF could promote expression of one or more new pro-teins that mediate enhanced cell motility during woundhealing.19 We have reported the pleiotropic effect of EGFon the PDGF-R mRNA and vice versa.23 Data from thepresent study indicate a similar pleiotropic effect of thesegrowth factors on the expression of the receptor protein aswell (Figures 1–3; Table 1). It has been shown that thevarious PDGF isoforms have slightly different signalingpathways.29 In the present study, the maximum levels ofthe receptor proteins and the time of attainment of peaklevels varied slightly depending on the growth factors used(Table 1). This is indicative of the ability of these growthfactors and their receptors to differentially modulate thesignaling pathways at various time points and in turn atdifferent phases during the cutaneous wound healingprocess.
Serum factors were known to work in concert with EGFto coordinate cell motility for efficient wound closure. In
addition, EGF was reported to enhance the wound closurein the presence of serum,19 and serum factors that regulatewound healing include PDGF in addition to others.30
Data from our previous studies showed that applicationof combination of EGF plus PDGF-BB produced the bestresult by healing the wound in 3–4 days as compared withthe control wound, which took approximately 10–15 daysto heal.22,23 This combination also produced the maximumlevel of EGF-R and PDGF-R mRNAs on day 1.23 Previ-ously we have suggested that the presence of the maximumlevel of receptor mRNA at the initial stages could be crit-ical for faster and better wound healing.23 Data from thepresent study show that this combination also produced aconcomitant increase in the levels of EGF-R and PDGF-Rproteins (Figure 5; Table 1) and their phosphorylation onday 1 (Figure 5). These results show that the combinationof EGF plus PDGF-BB produced the best pleiotropiceffect and these two growth factors are enough for the ac-celerated cutaneous wound healing process at the molecu-lar level. Statistical analysis of our data showed a directcorrelation between the peak level of receptor proteins andthe time taken for healing as well as the quality of healedwound that we have reported previously.22,23
EGF has been reported to be essential at the early stagesof wound healing,3,4 and the long form of PDGF-AA wasknown to antagonize the EGF-mediated healing.22,23 EGFis known to enhance cell proliferation and motility in bothfibroblasts and epithelial cells during wound repair.5,19
PDGF-AA application has been reported not to elicit di-rected cell migration during microvessel assembly.31
Therefore, it is conceivable that application of PDGF-AAwithin 30 minutes after EGF application could disrupt thecell migration that is necessary for the EGF-mediatedwound repair process. Topical application of PDGF-AAinhibited tyrosine phosphorylation of the EGF-R protein(Figure 5), delayed the cutaneous wound healing process,
Figure 5. Effect of combination of EGF
plus PDGFs on the tyrosine phosphorylat-
ion of receptor proteins. C, control; C1TP,
control lysate treated with tyrosine phos-
phatase; the growth factors used are as
marked above the lanes; 1TP above lanes
after the growth factors indicates the
treatment of lysates with tyrosine phos-
phatase; the last lane in each panel indi-
cates that the PDGF-AA was applied 30
minutes after EGF application. The anti-
bodies used are as indicated at the bot-
tom of each panel; the mature and
precursor (pre) receptor bands and the
b-tubulin bands are as marked on the right
of each panel. EGF, epidermal growth
factor; PDGF, platelet-derived growth
factors.
Wound Rep Reg (2009) 17 71–79 c� 2009 by the Wound Healing Society 77
Protein phosphorylation during wound healingGope et al.

and produced a hypertrophic scar (HS).22,23 PDGF-AAis known to be more potent in stimulating PKC activitythan PDGF-BB.32 PKC activation results in decreasedligand-binding affinity and decreased EGF-R tyrosineautophosphorylation.33,34 The observed tyrosine phos-phorylation of the receptor proteins in the control woundsand the absence of this phosphorylation in PDGF-AAtreated wounds (Figures 4 and 5) are indicative of the in-hibitory effect of PDGF-AA on EGF-R tyrosine phos-phorylation, perhaps via PKC activation,33,34 during anacute cutaneous wound healing process.
The role of PDGF-AA was reported to be differentfrom that of PDGF-BB during mitogenesis.35 Data fromthe present study indicate that PDGF-AA antagonizedthe EGF-mediated wound repair process, perhaps viainhibition of tyrosine phosphorylation of EGF-R protein(Figure 5). It has been suggested that PDGF-AAmight actthrough a different molecular pathway as compared withPDGF-AB and PDGF-BB.35,36 We found a slight increasein the levels of receptor mRNAs23 and proteins(Table 1) with the application of PDGF-AA. However,with the PDGF-AA treatment, the receptor protein wasnot phosphorylated, suggesting that the receptor tyrosinephosphorylation is an essential step during the woundhealing process. Interestingly, the receptor proteins in thecontrol wounds showed tyrosine phosphorylation. It ap-pears that the basal level of receptor proteins found in thecontrol wound is not enough for an accelerated wound re-pair process (Figures 1 and 2; Table 1) even though theproteins were phosphorylated (Figures 4 and 5). It hasbeen shown that the PDGF-BB, PDGF-AA, and fetal bo-vine serum were mitogenic and antiapoptotic, but only thePDGF-BB could induce cell migration.37 Cell migrationhas been reported to be essential for normal wound heal-ing37,38 and it appears that PDGF-AA lacks this ability.PDGFs are known to act through different signaling path-ways that are mediated by various cell types.7,39 Our re-sults show that PDGF-AA could act as an inhibitor ofEGF-R tyrosine phosphorylation during the wound heal-ing process. EGF-R tyrosine phosphorylation appears tobe essential for a normal cutaneous wound repair process.
Further studies are required to understand the specifickinase networks that are involved in the presence of vari-ous growth factors, during an acute cutaneous woundhealing process. Such studies should help us better under-stand wound healing in normal cases and individuals withcancers, which could pave the way toward tackling prob-lems such as impaired wound healing, delayed woundhealing, and HS formation.
ACKNOWLEDGMENTS
The authors thank Dr. J. Suresh Chandra, Mr. R. Rama-chandran, and Mr. K. Manolingam from the CentralAnimal Research Facility (CARF) for their help incarrying out the experiments. The authors thankDr. D.K. Subbakrishna, Department of Biostatistics,NIMHANS, for his help with the statistical analysis ofthe data. The authors also thank the staff of the photogra-phy section, NIMHANS, for their help. This work wasfully supported by a grant from Life Sciences Research
Board (LSRB), DRDO, Government of India, New Delhi,grant number: DBA/48222/c/RD-81.
REFERENCES
1. Martin P. Wound healing—aiming for perfect skin regener-ation. Science 1997; 276: 75–81.
2. Singer AJ, Clark RA. Cutaneous wound healing. N EnglJ Med 1999; 341: 738–46.
3. Brown GL, Nanney LB, Griffen J, Cramer AB, Yancey JM,Curtsinger LJ, Holtzin L, Schultz GS, Jurkiewicz MJ, LynchJB. Enhancement of wound healing by topical treatment withepidermal growth factor. N Engl J Med 1989; 321: 76–9.
4. Nanney LB. Epidermal and dermal effect of epidermal growthfactor during wound repair. J Invest Dermatol 1990; 94: 624–9.
5. Kwon YB, Kim HW, Rob DH, Yoon SY, Baek RM, KimJY, Kweon H, Lee KG, Lee JH. Topical application of epi-dermal growth factor accelerates wound healing by my-ofibroblasts proliferation and collagen synthesis in rat. J VetSci 2006; 7: 105–9.
6. Shah M, Revis D, Herrick S, Baillie R, Thorgeirson S, Fer-guson M, Roberts A. Role of elevated plasma transforminggrowth factor �beta 1 levels in wound healing. Am J Pathol1999; 154: 1115–24.
7. Heldin C-H,Westermark B.Mechanism of action and in vivorole of platelet-derived growth factor. Physiol Rev 1999; 79:1283–316.
8. Antoniades HN, Galanopoulos T, Neville-Golden J, KiritsyCP, Lynch SE. Injury induces in vivo expression of platelet-derived growth factor (PDGF) and PDGF receptor mRNAsin skin epithelial cells and PDGF mRNA in connective tissuefibroblasts. Proc Natl Acad Sci USA 1991; 88: 565–9.
9. Beer HD, Longaker MT, Werner S. Reduced expression ofPDGF and PDGF-receptors during impaired wound heal-ing. J Invest Dermatol 1997; 109: 132–8.
10. Uhl E, Rosken F, Sirsjo A, Messmer K. Influence of platelet-derived growth factor on microcirculation during normaland impaired wound healing. Wound Repair Regen 2003; 11:361–7.
11. Keswani SG, Katz AB, Lim F-Y, Zoltick P, Radu A,Alaee D, Herlyn M, Crombleholme TM. Adenoviral medi-ated gene transfer of PDGF-B enhances wound healingin type I and type II diabetic wounds. Wound Repair Regen2004; 12: 497–504.
12. Lee JA, Conejero JA, Mason JM, Parrett BM, Wear-Maggitti KD, Grant RT, Breitbart AS. Lentiviral transfect-ion with the PDGF-B gene improves diabetic wound healing.Plast Reconstr Surg 2005; 116: 532–8.
13. Cheng B, Liu HW, Fu XB, Sun TZ, Sheng ZY. Recombinanthuman platelet-derived growth factor enhanced dermalwound healing by a pathway involving ERK and c-fos in di-abetic rats. J Dermatol Sci 2007; 45: 193–201.
14. Li H, Fu X, Zhang L, Huang Q, Wu Z, Sun T. Research ofPDGF-BB gel on the wound healing of diabetic rats and itspharmacodynamics. J Surg Res 2008; 145: 41–8.
15. Steed DL. Modifying the wound healing response withexogenous growth factors. Clin Plast Surg 1998; 25: 397–405.
16. Babu M, Wells A. Dermal and epidermal communication inwound healing. Wounds 2001; 13: 183–9.
17. Declair V. The importance of growth factors in wound heal-ing. Ostomy Wound Manage 2001; 45: 64–74.
Wound Rep Reg (2009) 17 71–79 c� 2009 by the Wound Healing Society78
Protein phosphorylation during wound healing Gope et al.

18. Heldin C-H, Westermark B. Role of platelet derived growthfactor in vivo. In: Clark RAF, editor. The molecular andcellular biology of wound repair. New York: Plenum, 1996:249–73.
19. Kurten RC, Chowdhury P, Sanders Jr RC, Pittman LM, Ses-sions LW, Chambers TC, Lyle CS, Schnackenberg BJ, JonesSM. Coordinating epidermal growth factor-induced motilitypromotes efficient wound closure. Am J Physiol Cell Physiol2005; 288: C109–21.
20. Lee JH, Lee YJ, Hong SH, Kim JP. Effect of epidermalgrowth factor on full thickness skin defects of the inoperablepatients. J Korean Soc Plast Reconstr Surg 2005; 32: 314–8.
21. Haber M, Cao Z, Panjwani N, Bedenice D, Li WW, ProvostPJ. Effect of growth factors (EGF, PDGF-BB and TGF-beta1) on cultured equine epithelial cells and keratinocytes: im-plications for wound healing. Vet Ophthalmol 2003; 6: 211–7.
22. Gope R. The effect of epidermal growth factor and platelet-derived growth factors on wound healing process. Indian JMed Res 2002; 116: 201–6.
23. Gope ML, Gope R. Topically applied EGF and PDGFsaffect positively the co-ordinate expression of EGF andPDGF receptor genes during acute cutaneous wound healingprocess. Curr Sci 2007; 92: 618–27.
24. Rohini K, Mathivanan J, Antony Herold Prabhu PD, Sub-bakrishna DK, Gope ML, Chandramouli BA, Sampath S,Anandh B, Gope R. Loss of heterozygosity of the p53 geneand deregulated expression of its mRNA and protein inhuman brain tumors. Mol Cell Biochem 2007; 300: 101–11.
25. Mathivanan J, Rohini K, Gope ML, Anandh B, Gope R.Altered structure and deregulated expression of the tumorsuppressor gene retinoblastoma (RB1) in human braintumors.Mol Cell Biochem 2007; 302: 67–77.
26. Schlessinger J. Cell signaling by receptor tyrosine kinases.Cell 2000; 103: 211–25.
27. Kaftan H, Vogelgesang S, Lempas K, Hosemann W, HerzogM. Inhibition of epidermal growth factor receptor byerlotinib: wound healing of experimental tympanic mem-brane perforations. Otol Neurotol 2007; 28: 245–9.
28. Govindan R, Behnken D, Read W, McLeod M. Wound heal-ing is not impaired by the epidermal growth factor receptor-ty-rosine kinase inhibitor gefitinib. Ann Oncol 2003; 14: 1330–1.
29. Diliberto PA, Gordon GW, Yu C-L, Earp HS, Herman B.Platelet-derived growth factor (PDGF) alpha receptoractivation modulates the calcium mobilizing activity of the
PDGF beta receptor in Balb/c3T3 fibroblasts. J Biol Chem1992; 267: 11888–97.
30. Werner S, Grose R. Regulation of wound healing by growthfactors and cytokines. Physiol Rev 2003; 83: 835–70.
31. Zeller PJ, Skalak TC, Ponce AM, Price RJ. In vivo chemo-tactic properties and spatial expression of PDGF in develop-ing mesenteric microvascular networks. Am J Physiol HeartCirc Physiol 2001; 280: H2116–25.
32. Block LH, Emmons LR, Vogt E, Sachinidis A, Vetter W,Hoppe J. Ca21-channel blockers inhibit the action ofrecombinant platelet-derived growth factor in vascularsmooth muscle cells. Proc Natl Acad Sci USA 1989; 86:2388–92.
33. Hunter T, Ling N, Cooper JA. Protein kinase C phosphor-ylation of the EGF receptor at a threonine residue close tothe cytoplasmic face of the plasma membrane. Nature 1984;311: 480–3.
34. Sturani E, Vicentini LM, Zippel R, Toschi L, Pandiella-Alonso A, Comoglio PM, Meldolesi J. PDGF-inducedreceptor phosphorylation and phosphoinositide hydrolysisare unaffected by protein kinase C activation in mouse 3T3and human skin fibroblasts. Biochem Biophys Res Commun1986; 137: 343–50.
35. Pestana IA, Vazquez-Padron RI, Aitouche A, Pham SM.Nicotinic and PDGF-receptor function are essential fornicotine-stimulated mitogenesis in human vascular smoothmuscle cells. J Cell Biochem 2005; 96: 986–95.
36. Yu J, Deuel TF, Kim HR. Platelet-derived growth factor(PDGF) receptor-alpha activates c-Jun NH2-terminal kin-ase-1 and antagonizes PDGF receptor-beta-induced pheno-typic transformation. J Biol Chem 2000; 275: 19076–82.
37. Gao Z, Sasaoka T, Fujimori T, Oya T, Ishii Y, Sabit H,Kawaguchi M, Kurotaki Y, Naito M, Wada T, Ishizawa S,Kobayashi M, Nabeshima Y, Sasahara M. Deletion of thePDGF-R-b gene affects key fibroblast functions importantfor wound healing. J Biol Chem 2005; 280: 9375–89.
38. Li W, Fan J, Chen M, Guan S, Sawcer D, Bokoch GM,Woodley DT. Mechanism of human dermal fibroblastmigration driven by type I collagen and platelet-derivedgrowth factor �BB. Mol Biol Cell 2004; 15: 294–309.
39. Schneider L, Clement CA, Teilmann SC, Pazour GJ, Hoffm-ann EK, Satir P, Christensen ST. PDGFRalphaalpha signal-ing is regulated through the primary cilium in fibroblasts.Curr Biol 2005; 15: 1861–6.
Wound Rep Reg (2009) 17 71–79 c� 2009 by the Wound Healing Society 79
Protein phosphorylation during wound healingGope et al.



















