
Two-dimensional patterns of soluble proteins including three hydrolytic enzymes of mature pollen of...
12
ORIGINAL ARTICLE Two-dimensional patterns of soluble proteins including three hydrolytic enzymes of mature pollen of tristylous Lythrum salicaria A. Kalinowski A. Bocian A. Kosmala K. Winiarczyk Received: 18 July 2006 / Accepted: 9 December 2006 / Published online: 15 March 2007 Ó Springer-Verlag 2007 Abstract Lythrum salicaria, now a widespread invasive species, exhibits tristyly, a form of heteromorphic selfin- compatibility. In tristyly, each plant exhibits one (and only one) of three morphologically different floral forms. Moreover, each flower produces two types of stamens, and these two exhibit different incompatibility reactions. Dif- ferences between stamens of a single flower must be the result of epigenetic phenomena and for that reason, we performed two-dimensional gel electrophoresis (2-DE) to analyze fractions of soluble proteins derived from the pollen coat and protoplast including three hydrolytic enzymes from the six different stamen types (two from each of three floral forms). There were significant differences in the 2-D protein profiles both between pollen from the same flower and between the same type of pollen from two different flowers, in the pollen coat as well as in the protoplast ex- tracts. In five of the six samples of pollen fractions, char- acteristic peptides were found. Quantitative differences between pollen from the same flower were observed in case of esterases. Furthermore, analysis of proteases and acid phosphatases revealed also qualitative differences between these enzymes in pollen from the same flower. Keywords Lythrum salicaria Á Pollen Á Protein/enzymes Á Two-dimensional gel electrophoresis Introduction Classically, (see Darwin 1877) tristylous populations con- tain approximately equal proportion of three genetically determined floral forms. Individuals of each form are moderately self incompatible and incompatible also with other individuals showing the same form. This kind of species has long been studied because it exhibits a fasci- nating syndrome of characteristics. Plants of each form exhibit one, and only one, of three different lengths of styles. Flowers of the long-styled form contain also two types of stamens: mid- and short length. Flowers of the mid-styled form contain both long- and short stamens, and those of the short-styled form contain mid- and long sta- mens. Long stamens, in every case, produce pollen that is larger than that of the mid-length stamens, pollen of which is larger than that of the short stamens. The inheritance of the three floral forms is determined by two diallelic loci, termed S and M. Long-styled plants are homozygous recessive at both loci (ssmm), mid-styled plants are ssM- (meaning ssMM or ssMm), and any plant carrying the dominant S allele will produce short-styled flowers (that is, S is epistatic to M). It should be noted that the S locus referred to here presumably bears no relation to the S locus known to influence homomorphic selfincompatibility. For more recent references, see Barrett (1992, 2002, 2004), Barrett et al. (2000), de Nettancourt (2001) and references Communicated by Celestina Mariani. A. Kalinowski Á A. Bocian Á A. Kosmala (&) Institute of Plant Genetics, Polish Academy of Sciences, Strzeszyn ´ska 34, 60-479 Poznan ´, Poland e-mail: [email protected] A. Kalinowski e-mail: [email protected] A. Bocian e-mail: [email protected] K. Winiarczyk Department of Anatomy and Cytology of Plants, Marie Curie-Sklodowska University, Akademicka 19, 20-033 Lublin, Poland e-mail: [email protected] 123 Sex Plant Reprod (2007) 20:51–62 DOI 10.1007/s00497-006-0042-4
Transcript of Two-dimensional patterns of soluble proteins including three hydrolytic enzymes of mature pollen of...

ORIGINAL ARTICLE
Two-dimensional patterns of soluble proteins including threehydrolytic enzymes of mature pollen of tristylous Lythrumsalicaria
A. Kalinowski Æ A. Bocian Æ A. Kosmala ÆK. Winiarczyk
Received: 18 July 2006 / Accepted: 9 December 2006 / Published online: 15 March 2007
� Springer-Verlag 2007
Abstract Lythrum salicaria, now a widespread invasive
species, exhibits tristyly, a form of heteromorphic selfin-
compatibility. In tristyly, each plant exhibits one (and only
one) of three morphologically different floral forms.
Moreover, each flower produces two types of stamens, and
these two exhibit different incompatibility reactions. Dif-
ferences between stamens of a single flower must be the
result of epigenetic phenomena and for that reason, we
performed two-dimensional gel electrophoresis (2-DE) to
analyze fractions of soluble proteins derived from the pollen
coat and protoplast including three hydrolytic enzymes
from the six different stamen types (two from each of three
floral forms). There were significant differences in the 2-D
protein profiles both between pollen from the same flower
and between the same type of pollen from two different
flowers, in the pollen coat as well as in the protoplast ex-
tracts. In five of the six samples of pollen fractions, char-
acteristic peptides were found. Quantitative differences
between pollen from the same flower were observed in case
of esterases. Furthermore, analysis of proteases and acid
phosphatases revealed also qualitative differences between
these enzymes in pollen from the same flower.
Keywords Lythrum salicaria � Pollen � Protein/enzymes �Two-dimensional gel electrophoresis
Introduction
Classically, (see Darwin 1877) tristylous populations con-
tain approximately equal proportion of three genetically
determined floral forms. Individuals of each form are
moderately self incompatible and incompatible also with
other individuals showing the same form. This kind of
species has long been studied because it exhibits a fasci-
nating syndrome of characteristics. Plants of each form
exhibit one, and only one, of three different lengths of
styles. Flowers of the long-styled form contain also two
types of stamens: mid- and short length. Flowers of the
mid-styled form contain both long- and short stamens, and
those of the short-styled form contain mid- and long sta-
mens. Long stamens, in every case, produce pollen that is
larger than that of the mid-length stamens, pollen of which
is larger than that of the short stamens. The inheritance of
the three floral forms is determined by two diallelic loci,
termed S and M. Long-styled plants are homozygous
recessive at both loci (ssmm), mid-styled plants are ssM-
(meaning ssMM or ssMm), and any plant carrying the
dominant S allele will produce short-styled flowers (that is,
S is epistatic to M). It should be noted that the S locus
referred to here presumably bears no relation to the S locus
known to influence homomorphic selfincompatibility. For
more recent references, see Barrett (1992, 2002, 2004),
Barrett et al. (2000), de Nettancourt (2001) and references
Communicated by Celestina Mariani.
A. Kalinowski � A. Bocian � A. Kosmala (&)
Institute of Plant Genetics, Polish Academy of Sciences,
Strzeszynska 34, 60-479 Poznan, Poland
e-mail: [email protected]
A. Kalinowski
e-mail: [email protected]
A. Bocian
e-mail: [email protected]
K. Winiarczyk
Department of Anatomy and Cytology of Plants,
Marie Curie-Skłodowska University,
Akademicka 19, 20-033 Lublin, Poland
e-mail: [email protected]
123
Sex Plant Reprod (2007) 20:51–62
DOI 10.1007/s00497-006-0042-4

therein. The present paper is an investigation into the
differences between genetically identical, but behaviorally
dissimilar, pollen from a single form of Lythrum salicaria
(purple loosestrife) plant.
Diversity in forms and functions of flowers within an-
giosperms is favored by natural selection. The spatial and
temporal diversity systems of plant reproductive organs, in
most cases, are associated with the promotion of cross-
fertilization reducing negative effects of self-fertilization.
Floral heteromorphism, like heterostyly and enathiostyly,
is active promoting of a directional pollen dispersal be-
tween plants, while homomorphic self-incompatibility is
passive in character. In heterostylic plants, different polli-
nation and mating systems, even between closely related
taxa, indicate that the traits related with reproduction are
evolutionarily labile (Ganders 1979). The paper by Hus-
band and Barrett (1993) is a good example presenting the
evolutionary instability of tristylous Eichhornia paniculata.
The 24 enzymatic loci from flower buds (11 enzyme sys-
tems) in 167 populations in north-eastern Brazil were
investigated using the starch gel electrophoresis method.
The authors showed that 118 (70.7%) populations were
composed from trimorphic-flower plants, which were poly-
morphic in 20 enzymatic loci. A total of 42 (21.1%) popu-
lations were composed of long- and mid-styled flowers,
which were polymorphic in 16 loci, and the remaining 7
(4.2%) populations were composed of mid-styled flowers
only. No data regarding the analysis of generative parts of
flowers on the molecular level have been found so far.
Different lengths of stamens and styles were observed in
tristylous L. salicaria in relationship with different stigma
and pollen morphology (Hermann et al. 1999). The num-
bers of papillae are the same for each stigma type; how-
ever, the stigma of the long style possesses relatively
bigger, longer and more scarcely distributed papillae,
compared to the stigma of the short style and intermediate
stigma of the mid style (Hermann et al. 1999). The dif-
ferences in profiles of stigma proteins, generated using
two-dimensional gel electrophoresis, revealed that each of
the three stigma types possesses characteristic peptides
(Kalinowski et al., to be published).
Lythrum salicaria shows also differences both in pollen
morphology and color (Mulcahy and Caporello 1970).
Pollen from the long stamen is green, while that from the
mid- and short stamens is yellow. Differences in pollen
diameters were also observed. The biggest pollen comes
from the long stamen and the smallest one from the short
stamen. The diameters were correlated with the amount of
pollen production in the anthers (Mulcahy and Caporello
1970); however, the other results showed no differences in
the pollen diameters (O’Neil 1992). Mal and Hermann
(2000) obtained similar results as Mulcahy and Caporello
(1970) and they pointed out that in bigger populations the
plants produced relatively bigger pollen compared to plants
from small populations. Thus, both the yield of pollen
production and its diameter are influenced by environ-
mental conditions. No references concerning molecular
markers for different pollen types of L. salicaria have been
published so far.
One of the most important phenomena of heteromorphic
incompatibility is a relation between pollen morphology
and directional pollen flow between different morphs. It is
obvious that compatible pollination occurs between the
stigma and anther located on the same level; incompatible
pollination is not as strongly restricted as homomorphic
self-incompatibility. Mulcahy and Caporello (1970) found a
significant amount of self-pollination within the same
flowers after artificial pollination of emasculated flowers of
L. salicaria; however, this pollination was predominant
between the stamen and styles of the same length. These
authors, just like Ottenbreit and Staniforth (1994), con-
cluded that most events of incompatible pollination were
found in mid-styled flowers. Furthermore, the plants with
this type of flower produced the highest set of seed. In
purple loosestrife, asymmetric pollen flow is based on a
limited deposition of pollen on the short style and its
unlimited deposition on long- and mid styles (O’Neil 1992).
The characteristics of possible S loci in determined
heterostyly are still unknown. Barrett et al. (2000) wrote:
‘‘Protein differences between morphs may have nothing
to do with incompatibility per se but instead may simply
be associated with differential organs growth’’, and:
‘‘...ignorance of the molecular genetics of heterostyly will
continue’’.
Some results regarding organ-specific proteins were
obtained for distylous plants. Using one- and two-dimen-
sional gel electrophoresis methods in Averrhoa caram-
bola, Wong et al. (1994) detected 72 kDa protein in the
pin style and 45 kDa protein in the stamens of the thrum
morph. The specific 70 kDa peptide was also found in
vegetative organs. In the stigmas and styles of three spe-
cies of distylous Turnera three characteristic protein bands
were identified. Two of them were only found in the
pollen of short-styled plants. It was evident that genes
controlling the production of these proteins were closely
linked with the S-alleles. The functions of these proteins
have not been recognized, but it is supposed that small
amounts of these proteins are also present in vegetative
organs (Athanasiou and Shore 1997).
Since no data concerning floral-morph-specific molec-
ular markers are available, particularly for pollen and
pistils in tristylous L. salicaria, we started with separate
analyses of the pollen coat and pollen protoplast proteins.
Tristylous species are particularly valuable for such
investigations because each allows comparison of two
pollen types within a single type of flower. That is,
52 Sex Plant Reprod (2007) 20:51–62
123

comparison of the pollen from long- and short stamens
within a single mid-styled plant should be more informa-
tive than comparing pollen from long- and short stamens of
a distylous species, such as Primula sp. In Primula, the
differences may relate to heterostyly, but more likely they
represent irrelevant differences between two individual
plants. With the tristylous species, the comparison can be
made within a single plant, thus eliminating many extra-
neous differences. Furthermore, proteins common to, for
example, long stamens from mid- and short-styled Lythrum
plants can be selected for further investigation if they are
not found in other pollen types of the same plants.
Different proteins (some of them possess enzymatic
activity), lipid-like substances, carbohydrates and second-
ary chemicals, mainly flavones and carotenes, could be
found both in the pollen coat and in the protoplast. Proteins
in the protoplast and in the inner layers of the intine are
under control of microgametophyte genes. Proteins pro-
duced in the tapetum and deposited with other substances
in cavities and on the surface of exine formed a pollen coat
(Heslop-Harrison et al. 1973; Vithanage and Knox 1979;
Murphy and Ross 1998; Doughty et al. 2000). The pollen
coat may be easily washed off with a water solution of
mannitol (Howlett et al. 1975), cyclohexane (Doughty et al.
1993) and with diethyl ether (Suen et al. 2003). We pre-
ferred to use an isotonic solution of sucrose with dithio-
treitol and other chemicals—it allowed us to analyze
soluble proteins including enzymes. The use of the solution
without dithiotreitol allowed us to perform in vivo exper-
iments (Kalinowski et al. 2002b).
The current work presents the results of the qualitative
analyses of sporophytically and gametophytically ex-
pressed pollen proteins of L. salicaria together with the
analysis of pollen enzymes without separation into pollen
coat and protoplast fractions, with the use of two-dimen-
sional gel electrophoresis with SDS. The aim of the study
was to find: (1) differences between pollen from the same
morph, (2) differences between the same type of pollen
from different morphs, (3) pollen coat and protoplast
characteristic peptides for each type of pollen.
Materials and methods
Three plants—one with long-, one with mid- and one with
short-styled flowers—were transplanted from their natural
habitat to big flowerpots and then were kept in the garden
to provide the same conditions.
Early morning, before opening, the flowers were col-
lected and kept in Petri dishes with wet filter paper at 4�C
to prevent the dehiscence of the anthers. Indehiscent an-
thers were transferred to sieves. Flowers with opening
anthers were discarded. After 20–30 min. under bubble
light, the anthers were broken and then pollen was sifted
using a little stiff-haired brush. Mature pollen was kept in
Eppendorf tubes at –70�C.
Six samples of pollen were taken for analysis. The
samples were designated as follows: (L, M and S—long-,
mid- and short style, respectively; l, m, and s—long-, mid-
and short stamen, respectively) according to Mulcahy and
Caporello (1970). Thus, Lm indicates a sample from a
long-styled flower, mid-length stamens; Ls—long-styled
flower, short stamens, etc. Samples were as follows: (1)
Lm, (2) Ls, (3) Ml, (4) Ms, (5) Sl and (6) Sm. For con-
venience the following marks are used: pc—pollen coat,
pt—pollen protoplast.
Soluble proteins: pollen coat elution, extraction
of proteins and two-dimensional gel electrophoresis
Pollen coat elution and protein extraction were performed
as in Kalinowski et al. (2002b). Samples of 10 mg of each
type of pollen were shaken gently at room temperature for
three times with 300 ll of isotonic extraction buffer (0.7 M
sucrose, 0.5 M Tris, 30 mM HCl, 0.1 M KCl) according to
Hurkman and Tanaka (1986). During the first wash, pollen
samples were eluted for 20 min. and during the other two
washes for 5 min. Each of the three fractions was then
shaken successively with phenol dissolved in water
(400 ll) to ensure the concentration of proteins from three
eluates in one volume of phenol solution. Pollen devoid of
pollen coat proteins was homogenized with 400 ll of the
extraction buffer and then shaken with phenol. Proteins
were precipitated from the phenol phase by a solution of
0.1 M ammonium acetate in methanol. After drying, the
proteins were dissolved in an appropriate volume of
resolving buffer (9 M urea, 4% Nonidet NP-40, 2% 2-
mercaptoethanol, 2% ampholytes pH 2–4)—20 ll in case
of pollen coat and 30 ll in case of protoplast fraction,
according to Hurkman and Tanaka (1986). On the column
gels, 15 ll of pollen coat and 10 ll of pollen protoplast
extracts were applied.
Two-dimensional gel electrophoresis was performed
according to Hochstrasser et al. (1988). In the first
dimension, isoelectrofocusing (IEF), a 0.8 · 150 mm
capillary was used for polymerization of polyacrylamide
column gel with Servalyte pH range 3–10. In the second
dimension (SDS-PAGE), the proteins were separated in
13% polyacrylamide slabs (1 · 140 · 140 mm). After
electrophoresis, the peptides were stained with silver ni-
trate according to Heukeshoven and Dernick (1985).
Enzymes: extraction of proteins and 2-DE conditions
The 20 mg of each type of pollen (six samples) was
homogenized with 80 ll solution of 15% sucrose with 2%
Sex Plant Reprod (2007) 20:51–62 53
123

dithiotreitol (DTT) and then centrifuged at 14,000 rpm
(Hettich—Universal 32R; 1620A) at 4�C. The volume of
20 ll of extract was applied on column gels for separation
in the first dimension (IEF).
Analyses of esterases (EST) and acid phosphatases
(ACPH) using two-dimensional electrophoresis in dena-
turing conditions (with SDS) and the renaturing of their
activities were conducted according to a previously
developed methodology (Kalinowski et al. 2002a). After
the separation of proteases (PRO) in the first dimension
(IEF), the column gels were transferred into the same
buffer as used for EST and ACPH, containing 0.1% SDS.
In the second dimension, the proteins were separated in 7%
polyacrylamide slabs containing 0.4% SDS and 0.4% gel-
atin from porcine skin as a substrate for protease activity.
Undigested gelatin in the gel was stained with Coomasie
Brillant Blue G-250 (Michaud et al. 1993).
Gel analysis
Each extraction procedure and electrophoretic separation
was performed twice. Total separated protein/enzyme
spots on the gels were scanned (SHARP JX-330) and
subjected to LabScan program processing. The data were
further processed with Image Master 2-D Elite LKB
software. Enzymes were compared also quantitatively on
the basis of light absorption of their particular isoforms
revealed after electrophoresis (using the densitometry
option in Image Master 2-D Elite software). The obtained
data were then used to estimate approximate enzymatic
activities.
The Pharmacia 2-DE calibration kit (17-0582) and
Sigma silver stain SDS molecular weight mixture (M-
6539) were used to standardize the gels and to determine
the molecular weights (MW) and isoelectric points (iP) of
the analyzed proteins.
Results
All the 2-D patterns were highly repeatable. The 2-D
profiles of proteins from the pollen coat were different
from those of the protoplast. Furthermore, differences be-
tween two types of pollen from the same flower and also
between the same type of pollen produced in two morphs
were found. ‘‘Characteristic peptides’’ refer to the peptides
present only in one of the six samples of pollen coat
fractions or pollen protoplast fractions, or the isozymes
present only in one of the six types of pollen. Excluding the
pollen coat of the Sl, characteristic peptides were observed
for the remaining samples both from the pollen coat and
from the protoplast.
Pollen coat proteins
Proteins from six samples of the pollen coat (pc) after two-
dimensional gel electrophoresis separated at a pH range of
4.0–9.8 and their molecular weights were from ~9 to
~80 kDa. In each pc fraction, excluding the Sl, character-
istic peptides were found and their numbers were as fol-
lows: Lm—9, Ls—3, Ml—5, Ms—5 and Sm—19. These
peptides were marked on Figs. 1, 2, and 3.
Taking into account two electrophoretic parameters
(molecular weights and isoelectric points), clear differ-
ences between protein profiles of six pollen coat fractions
were observed. Comparing pollen coat fractions of the two
pollen types found within a single flower morph, we found
that in the pc of the Lm 53 peptides were present, which
were absent in the pc of the Ls. Inversely, 68 peptides were
identified in the pc of the Ls and undetected in the pc of the
Lm. In case of mid-styled flowers, 26 pc proteins charac-
teristic for the Ml were undetected in the pc profiles of the
Ms. Moreover, 85 pc peptides found in the Ms were not
detected in the Ml. The pc of the Sl pollen contained 12
Fig. 1 The 2-D protein patterns
from pollen coat of mid- (Lm)
and short (Ls) stamen of a long-
styled flower. Characteristic
peptides were marked with
arrows. Their molecular
weights in kDa and isoelectric
points at pH (in brackets) were
for the Lm: 11.5 (5.1), 12.0
(5.2), 23.0 (8.8), 27.0 (7.8), 39.0
(5.5 and 5.6) and 56.0 (8.9, 9.2
and 9.4) and for the Ls: 19.0
(5.0), 20.5 (8.7) and 39.0 (9.0)
54 Sex Plant Reprod (2007) 20:51–62
123

peptides, which were absent in the pc of the Sm, and in-
versely, 177 pc peptides from the Sm were not detected in
the pc of the Sl.
Differences were also detected for the same type of
pollen found in different floral morphs. In the pc of the Lm,
34 characteristic peptides were detected, which were absent
in the pc of the Sm. The profile of the pc of the Sm showed
the occurrence of 172 peptides undetected in the profile of
the pc of the Lm. In the coat of the Ls, 13 peptides were
detected, while these were simultaneously undetected in
the pc of the Ms. Inversely, 76 peptides present in the Ms
were absent in the Ls. The comparisons of 2-D protein
profiles within the pc of long-stamen pollen showed that in
the Ml 23 peptides present were undetected in the Sl and 37
peptides were observed in the Sl and simultaneously not
observed in the Ml. Thus, comparing the pollen coat pep-
tide profiles from a single type of pollen from different
forms of flowers, we found that differences were highest
for mid-stamen pollen and lowest for long-stamen pollen.
One of the most interesting results was the identification
of common pollen coat peptides for the same type of pollen
produced in two different types of flowers (the peptides
which were simultaneously absent in the two other pollen
types). For the pc of long-stamen pollen eight (omitting
two weakly stained), for mid stamen—nine and for short
stamen—five such peptides were found. The MWs and iPs
(at pH) of these peptides are given in Table 1.
Fig. 2 The 2-D protein patterns
from pollen coat of long- (Ml)and short (Ms) stamen of a mid-
styled flower. Characteristic
peptides were marked with
arrows. Their molecular
weights in kDa and isoelectric
points at pH (in brackets) were
for the Ml: 11.0 (4.5), 14.2
(4.5), 18.4 (8.5), 29.0 (5.8) and
44.0 (5.9) and for the Ms: 13.0
(7.0), 14.5 (4.4), 26.0 (5.6) 28.5
(7.9) and over 66.0 kDa marker
at pH 6.2
Fig. 3 The 2-D protein patterns
from pollen coat of long- (Sl)and mid (Sm) stamen of a short-
styled flower. Characteristic
peptides were marked with
arrows. Their molecular
weights in kDa and isoelectric
points at pH (in brackets) were
for the Sm as follows: 11.0
(5.9), 11.5 (5.6 and 7.6), 12.0
(5.0, 7.6 and 7.8), 23.0 (7.4),
28.5 (5.4 and 5.5). 32.0 (8.7),
three 33.0–34.0 (5.6–5.7), 36.0
(8.2), 39.0 (4.9), 43.0 (8.3), 46.0
(8.5) and 61.0 (7.4 and 7.6)
Table 1 List of pollen coat and protoplast peptides present only in
one of the long-, mid- or short-stamen pollen
Lm Ls Ml
10.4/4.9 10.5/6.1 11.0/6.8
12.0/7.5 21.5/6.8 18.0/8.7
12.5/5.1 22.0/6.9 21.5/5.9
13.0/5.0 31.0/8.5 23.0/6.9
14.0/4.9 35.0/7.9 23.5/7.1
14.2/4.8 44.0/7.5
15.0/4.7 49.0/5.6
31.0/7.6 62.0/7.3
32.0/7.5
In the first place the MW in kDa and in the second place iP at pH are
given (kDa/pH)
Sex Plant Reprod (2007) 20:51–62 55
123

Pollen protoplast proteins
Proteins from six samples of pollen protoplast (pt) after
two-dimensional gel electrophoresis, as was the case with
pollen coat proteins, were separated at pH 3.5–9.8 range
and their molecular weight differed from ~9 to ~80 kDa. In
each of six pt fractions, characteristic peptides were found
and their numbers were as follows: Lm—7, Ls—6, Ml—8,
Ms—6, Sl—3 and Sm—3. These peptides were marked on
Figs. 4, 5, and 6.
Similar to the 2-D patterns of pollen coat proteins, the 2-
D patterns of protoplast protein fractions between two types
of pollen from the same kind of flower were also different.
In the pt of the Lm, 74 characteristic peptides were present;
simultaneously, these peptides were absent in the pt of the
Ls. Inversely, 47 peptides typical for the pt of the Ls were
undetected in the pt of the Lm. In the case of the mid-style
flowers, 81 proteins identified in the pt of the Ml were not
observed in the pt of the Ms, whereas 39 characteristic
peptides found in the pt of the Ms were not detected in the
Ml. The pt of the Sl pollen contained 39 peptides, which
were absent in the pt of the Sm, while the pt of the Sm
contained 32 peptides not detected in the pt of the Sl.
Comparing pc and pt protein profiles between the same
type of pollen showed that in the pt of the Lm, 156 peptides
were detected—they were absent in the pt of the Sm. In-
versely, 89 peptides in pc of the Sm (undetected in pt of the
Lm) were found. In the pollen coat of Ls, 99 peptides were
detected and simultaneously undetected in the pt of the Ms,
while 73 peptides characteristic for the Ms were not ob-
served in the Ls. Comparisons between the 2-D profiles of
the pt of long-stamen pollen showed that 36 Ml peptides
were undetected in the Sl, while 75 Sl peptides were absent
in the Ml. These comparisons showed that the pollen pro-
toplast fraction profiles were most varied in the case of
mid-stamen pollen and least varied in the case of long-
stamen pollen, similar to the results obtained for pollen
coat protein profiles. We identified also common pollen
protoplast peptides for the same type of pollen produced in
two different types of flowers (the peptides that were
simultaneously absent in two remaining pollen types). Only
two such peptides in the pt of the Ml were found. Their
Fig. 4 The 2-D protein patterns
from pollen protoplast of mid-
(Lm) and short (Ls) stamen of a
long-styled flower.
Characteristic peptides were
marked with arrows. Their
molecular weights in kDa and
isoelectric points at pH (in
brackets) were for the Lm: 20.5
(9.2), 21.0 (8.9), 26.5 (9.2 and
9.3), 29.0 (8.4 and 8.6) and 42.0
(4.6) and for the Ls: 111.5 (9.1),
23.0 (9.1), 24.0 (9.3), 41.0 (9.2),
43.0 (9.1) and 46.0 (9.2)
Fig. 5 The 2-D protein patterns
from pollen protoplast of long-
(Ml) and short (Ms) stamen
of a mid-styled flower.
Characteristic peptides were
marked with arrows. Their
molecular weights in kDa and
isoelectric points at pH (in
brackets) were for the Ml: 12.0
(7.6), 12.5 (7.2), 13.0 (7.7), 15.0
(3.9), 15.5 (8.2), 18.4 (3.7), 40.0
(8.7) and 47.0 (8.3) and for the
Ms: 10.5 (7.8), 15.0 (9.1 and
9.3), 22.5 (4.6), 25.0 (3.9) and
26.0 (3.9)
56 Sex Plant Reprod (2007) 20:51–62
123

electrophoretic parameters (MW and iP) were as follows:
19 kDa at pH 9.6 and 64 kDa at pH 7.8.
Enzymes
Esterases
Four isoesterases were separated using two-dimensional
gel electrophoresis in denaturing conditions (with SDS).
Their MWs ranged from 17.5 to 25 kDa, at pH 5.1–6.3
(Fig. 7). No qualitative differences were observed be-
tween the two types of pollen from the same morph.
Only quantitative differences were found between the Ml
and the Sl, and also between the Lm and the Sm. Dif-
ferences in the activities of some isoesterases from the
same type of pollen from two morphs were observed
after electrophoresis. The isophorm designated as no. 2
from the Lm was twice as active as the isophorm from
the Ls pollen, while three remaining pollen isoesterases
revealed similar, high activities after electrophoresis.
Isophorm no. 4 was significantly less active in the Sl
than in the Sm. Four isoesterases in the Ml and the Ms
pollen were clearly less active compared to the pollen
isoesterases from the L and the S morphs, while these
enzymes from the Ml were slightly more active than
those from the Ms.
Fig. 6 The 2-D protein patterns
from pollen protoplast of long-
(Sl) and mid (Sm) stamen of a
short-styled flower.
Characteristic peptides were
marked with arrows. Their
molecular weights in kDa and
isoelectric points at pH (in
brackets) for Sl were as follows:
38.0 (4.1), 40.0 (3.9) and 45.0
(4.6) and for Sm: 49 (4.3), 50.0
(4.1) and 54.0 (4.0)
Fig. 7 Fragments of gels
showed 2-D profiles of esterases
from six samples of pollen of
purple loosestrife. Activities of
particular isoesterases were
based on light absorption
revealed after electrophoresis
using the densitometry option in
Image Master 2-D Elite
software
Sex Plant Reprod (2007) 20:51–62 57
123
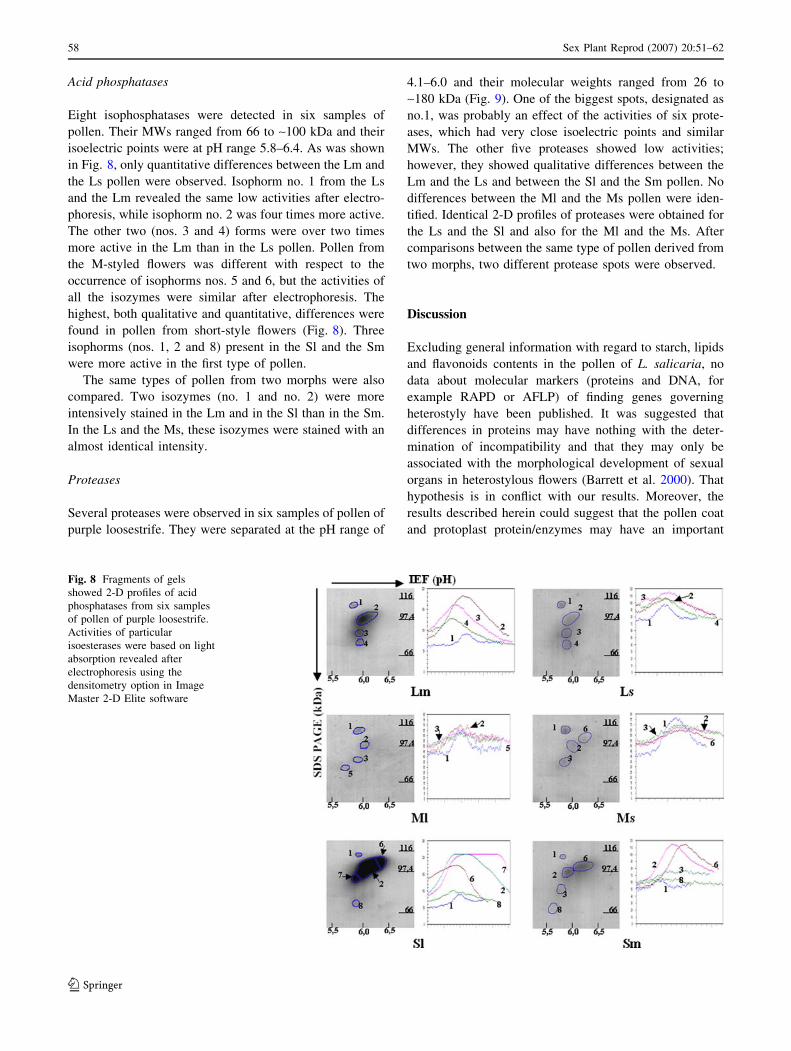
Acid phosphatases
Eight isophosphatases were detected in six samples of
pollen. Their MWs ranged from 66 to ~100 kDa and their
isoelectric points were at pH range 5.8–6.4. As was shown
in Fig. 8, only quantitative differences between the Lm and
the Ls pollen were observed. Isophorm no. 1 from the Ls
and the Lm revealed the same low activities after electro-
phoresis, while isophorm no. 2 was four times more active.
The other two (nos. 3 and 4) forms were over two times
more active in the Lm than in the Ls pollen. Pollen from
the M-styled flowers was different with respect to the
occurrence of isophorms nos. 5 and 6, but the activities of
all the isozymes were similar after electrophoresis. The
highest, both qualitative and quantitative, differences were
found in pollen from short-style flowers (Fig. 8). Three
isophorms (nos. 1, 2 and 8) present in the Sl and the Sm
were more active in the first type of pollen.
The same types of pollen from two morphs were also
compared. Two isozymes (no. 1 and no. 2) were more
intensively stained in the Lm and in the Sl than in the Sm.
In the Ls and the Ms, these isozymes were stained with an
almost identical intensity.
Proteases
Several proteases were observed in six samples of pollen of
purple loosestrife. They were separated at the pH range of
4.1–6.0 and their molecular weights ranged from 26 to
~180 kDa (Fig. 9). One of the biggest spots, designated as
no.1, was probably an effect of the activities of six prote-
ases, which had very close isoelectric points and similar
MWs. The other five proteases showed low activities;
however, they showed qualitative differences between the
Lm and the Ls and between the Sl and the Sm pollen. No
differences between the Ml and the Ms pollen were iden-
tified. Identical 2-D profiles of proteases were obtained for
the Ls and the Sl and also for the Ml and the Ms. After
comparisons between the same type of pollen derived from
two morphs, two different protease spots were observed.
Discussion
Excluding general information with regard to starch, lipids
and flavonoids contents in the pollen of L. salicaria, no
data about molecular markers (proteins and DNA, for
example RAPD or AFLP) of finding genes governing
heterostyly have been published. It was suggested that
differences in proteins may have nothing with the deter-
mination of incompatibility and that they may only be
associated with the morphological development of sexual
organs in heterostylous flowers (Barrett et al. 2000). That
hypothesis is in conflict with our results. Moreover, the
results described herein could suggest that the pollen coat
and protoplast protein/enzymes may have an important
Fig. 8 Fragments of gels
showed 2-D profiles of acid
phosphatases from six samples
of pollen of purple loosestrife.
Activities of particular
isoesterases were based on light
absorption revealed after
electrophoresis using the
densitometry option in Image
Master 2-D Elite software
58 Sex Plant Reprod (2007) 20:51–62
123

influence on the prezygotic events in L. salicaria. The
investigations presented in this paper represent the first
analysis of L. salicaria pollen on the molecular level and
could be the starting point for wider investigations
regarding the probable role of pollen and style proteins in
compatible and incompatible pollination in purple loose-
strife.
In tristylous Narcisus triandrus, a significantly lower
seed set was observed after self-pollination than after
cross-pollination (Sage et al. 1999). In this species, self-
sterility operates prezygotically as the ovarian self-
incompatibility. Authors hypothesized that the degradation
of the embryo sac was due to the absence of a stimulus.
Similar results have been obtained for L. salicaria by
O’Neil (1994). Pollination induces a cascade of changes in
relation with chemical signals (Du et al. 1996; Cheung and
Wu 1999; Cheung et al. 2000; Lindsey et al. 2002). Thus,
different proteins including probably also some enzymes in
different types of pollen coat may induce different changes
in the style of L. salicaria. Different chemical signals de-
rived from pollen coat chemicals, mainly proteins, induce
changes in the pathway of pollen tube growth that can
inhibit or stimulate this growth. For each of the six pollen
coat samples the characteristic peptides were found. Spe-
cific pollen coat proteins may induce some kinds of
changes in the pistil after incompatible pollination, which
are different from changes started after compatible polli-
nation.
Fig. 9 Fragments of gels
showed 2-D profiles of
proteases from six samples of
pollen of purple loosestrife
Sex Plant Reprod (2007) 20:51–62 59
123

Scribailo and Barrett (1991) showed that in L. salicaria
and L. junceum after incompatible pollination the pollen
tubes growth is inhibited in the stigma or in the upper part
of the style, while in tristylous Pontederia sagittata and P.
cordata that event takes place in the style and in the ovary.
Interactions between specifies are under control of S-genes
present both in the pollen and in the pistil (heteromorphic
incompatibility), (Richards 1986, cited after Scribailo and
Barrett 1991). Thus, it may suggest that some pollen coat
and protoplast characteristic peptides are also present in the
stigmas (Kalinowski, unpublished data) and they may be
involved in this interaction. However, Scribailo and Barrett
(1991) excluded an active rejection by molecular specifies
in P. sagittata. The authors suggested that incompatibility
is rather governed by interactions of heteromorphic traits.
In L. salicaria, except for compatible pollination, the
incompatible pollination was also observed. The lack of
physiological barriers to self-fertilization makes possible
the composition of pollen load on the stigma (Eckert and
Barrett 1994). The authors pollinated stigmas of Decodon
verticillatus using the mixture of self- and out-crossed
pollen and pointed out the possible signification of pollen
competition. The consequences of pollen competition was
seed set and seed germination. One of the requirements for
pollen competition is the existence of pollen polymorphism
(different pollen loads on the stigma); the distance between
the place of pollen arresting to the ovule location and the
number of ovules are also important (Mulcahy 1974;
Mulcahy and Mulcahy 1975; Winsor et al. 2000). Our re-
sults revealed protein polymorphism between six types of
pollen (between two pollen types from each of the three
morphs). Furthermore, characteristic peptides both in the
pollen coat and in the protoplast were found. Additionally,
2-D protein profiles from the stigmas of the three morphs
of L. salicaria (unpublished data) showed significant dif-
ferences.
This paper focuses on the differences in proteins. Two-
dimensional gel electrophoresis of pollen coat and protoplast
proteins as well as enzymes from unwashed pollen showed
differences between pollen from the same morph. Despite
the variations in the degree of incompatibility among pop-
ulations and even among individuals, differences between
pollen of L. salicaria presented in our work, together with
the results obtained by O’Neil (1994), suggest the existence
of preferences for one of the two types of pollen from dif-
ferent morphs in the case of compatible pollinations. These
preferences are probably genetically determined among the
populations because experiments were performed on plants
growing under identical conditions in the greenhouse. The
plants from which pollen was collected for our experiments
were also grown in the same garden conditions.
Lythrum salicaria produces pollen of different sizes
(Mulcahy and Caporello 1970). Moreover, Mal and
Hermann (2000) suggested differences between three types
of pollen also between populations. While the morphology
of exine and the composition of the pollen coat are under
the control of sporophyte genes, pollen size is under the
control of both sporophyte and microgametophyte genes
(Ottaviano and Mulcahy 1989). Common peptides for the
same type of pollen were mainly found among proteins
derived from the sporophyte; only two peptides were
identified in the protoplast of the Ml pollen. So far, there
are no answers to questions concerning the probable role of
these peptides in the legitimate/illegitimate pollination on
the basis of the presented results.
The 2-DE with SDS analysis of esterases showed no
qualitative differences between two types of pollen in each
of the three morphs. Only quantitative differences between
pollen from the same morph and between the same type of
pollen were observed. In Nicotiana alata, the esterases
were not associated with the expression of self-incompat-
ibility, but they were important in the pollen-pistil
interaction (Blaas and Bredemeijer 1978). Esterases-
hydrolyzing substances present in the stigma and style
facilitated the growth of the pollen tube in Brassica
(Hiscock et al. 2002). The results presented in this paper
may only suggest that pollen esterase activities are corre-
lated with differences in the anatomy of L. salicaria stig-
mas. Thus, pollen esterases could facilitate penetration of
the stigma by the pollen tube in compatible pollination and
this process could be quantitative in character.
The 2-D protein patterns of acid phosphatases showed
both qualitative (with the exception of the Lm and the Ls)
and quantitative differences between pollen from one
morph and between the same type of pollen derived from
different morphs. Roggen and Stanley (1969) stated that
phosphatases, proteinases and a-amylases inhibit pollen
tube growth. Phosphatases through hydrolysis of phosphate
residues act antagonistically to protein kinases, which play
an essential role in signal transductions (Kunz et al. 1996;
Cheung et al. 2000). Thus, the proportion of phosphory-
lation (catalyzed by protein kinases) and dephosphoryla-
tion (catalyzed by phosphatase) of proteins may induce
different signals in the pistil after legitimate pollination
when compared to the signals after illegitimate pollination.
Pollen proteases showed high activities after electro-
phoresis. Protease spot designated as no. 1 was probably an
effect of the activities of six proteases, which had very
close isoelectric points and similar MWs (clearly seen as a
negative picture). Different 2-D profiles of proteases were
observed both between the same type of pollen as well as
pollen from the same morphs, leaving out of account dif-
ferences between types of pollen. Hiscock and Dickinson
(1993) suggested that some proteases caused the death of
the stigma papillary cells. Pollen proteases facilitate pollen
tube growth through the hydrolysis of stigma proteins in
60 Sex Plant Reprod (2007) 20:51–62
123

the transmission tissue of the style (Radłowski et al. 1996).
However, some of the proteases can inhibit the growth of
the pollen tube as well (Roggen and Stanley 1969). Thus,
proteases may play an important role in the pollen–pistil
interaction in tristylous L. salicaria.
The differences between 2-D protein patterns of pollen
within the same morph as well as between the same type of
pollen from two morphs may be partially explained by
post-translational modifications of the observed proteins
(Zannis and Breslow 1981).
Acknowledgment We wish to thank Prof. David L. Mulcahy,
University of Massachusetts, for his inspiration to start the investi-
gation of pollen molecular markers of purple loosestrife, for his
extensive and valuable advice before the performance of the experi-
ments and for his critical reading of the manuscript.
References
Athanasiou A, Shore JS (1997) Morph-specific proteins in pollen
and styles of distylous Turnera (Turneraceae). Genetics
146:669–679
Barrett SCH (1992) Evolution and function of heterostyly In: Barrett
SCH (ed) Springer, Berlin
Barrett SCH (2002) The evolution of plant sexual diversity. Nat Rev
Genet 3:274–284
Barrett SCH (2004) Pontederiaceae (water hyacinth) family. In: Smith
N, Mori SA, Henderson A, Stevenson DW, Heald SV (eds)
Flowering plants of the neotropics. Princeton University Press,
Princeton, pp 474–476
Barrett SCH, Jesson LK, Baker AM (2000) The evolution and
function of stylar polymorphism in flowering plants. Ann Bot
85A:253–265
Blaas J, Bredemeijer GMM (1978) Isoelectric focusing of stylar
esterase isoenzymes in different S-genotypes of Nicotiana alataLink et Otto. Incompat Newsl 10:76–86
Cheung AY, Wu H-M (1999) Arabinogalactan proteins in plant
sexual reproduction. Protoplasma 208:87–98
Cheung AY, Zhan XY, Wong E, Wang H, Wu H-M (2000)
Transcriptional, post-transcriptional and post-translational regu-
lation of Nicotiana stylar transmitting tissue-specific arabinog-
alacta-proteins. In: Clarke A, Bacic A, Nothnagel E (eds) Cell
and developmental biology of arabinogalactan proteins. Kluwer/
Academic/Plenum, New York, pp 133–148
Darwin CR (1877) The different forms of flowers on plants of the
same species. John Murray, London
Doughty J, Hedderson F, McCubbin A, Dickinson H (1993)
Interaction between coating-borne peptides of the Brassicapollen grain and stigmatic S (self-incompatibility)-locus specific
glycoproteins. Proc Natl Acad Sci USA 90:467–471
Doughty J, Wong HY, Dickinson HG (2000) Cysteine-rich pollen
coat proteins (PCPs) and their interactions with stigma S
(incompatibility) and S-related proteins in Brassica putative
roles in SI and pollination. Ann Bot 85A:161–169
Du H, Simpson RJ, Clarke AE, Bacic A (1996) Molecular charac-
terization of a stigma-specific gene encoding an arabinogactan-
protein (AGP) from Nicotiana alata. Plant J 9:313–323
Eckert CE, Barrett SCH (1994) Post-pollination mechanisms and the
maintenance of outcrossing in self-incompatible, tristylous, Dec-odon verticillatus (Lythraceae). Heredity 72:396–411
Ganders FR (1979) The biology of heterostyly. N Z J Bot 17:607–635
Hermann BP, Mal TK, Wiliams RJ, Dollahon NR (1999) Quantitative
evaluation of stigma polymorphism in tristylous weed, Lythrumsalicaria. Am J Bot 86:1121–1129
Heslop-Harrison J, Heslop-Harrison Y, Knox RB, Howlett B (1973)
Pollen wall proteins: ‘‘gametophytic’’ and ‘‘sporophytic’’ frac-
tions in the pollen walls of the Malvaceae. Ann Bot 37:403–412
Heukeshoven J, Dernick R (1985) Simplified method for silver
staining of proteins in polyacrylamide gels and the mechanism of
silver staining. Electrophoresis 6:103–112
Hiscock SJ, Dickinson HG (1993) Unilateral incompatibility within
Brassicaceae: further evidence for the involvement of the self-
incompatibility. Theor Appl Genet 86:744–753
Hiscock SJ, Bown D, Gurr SJ, Dickinson HG (2002) Serine esterases
are requirement for pollen tube penetration of the stigma in
Brassica. Sex Plant Reprod 15:65–74
Hochstrasser DF, Harrington MG, Hochstrasser AC, Miller MJ, Merril
CR (1988) Methods for increasing the resolution of two-
dimensional protein electrophoresis. Anal Biochem 73:424–435
Howlett BJ, Knox RB, Pactons JB, Heslop-Harrison J (1975) Pollen
wall proteins, physiochemical characterization and role in self-
incompatibility in Cosmos bipinatus. Proc Lond Ser B 188:167–
182
Hurkman WJ, Tanaka CK (1986) Solubilization of plant membrane
proteins for analysis by two-dimensional gel electrophoresis.
Plant Physiol 81:802–806
Husband BC, Barrett SCH (1993) Multiple origin of self-fertilization
in tristylous Eichhornia paniculata (Pontederiaceae): inferences
from style morph and isozyme variation. J Evol Biol 6:591–608
Kalinowski A, Radłowski M, Bartkowiak S (2002a) Maize pollen
enzymes after two-dimensional polyacrylamide gel electropho-
resis in the presence or absence sodium dodecyl sulfate.
Electrophoresis 23:138–143
Kalinowski A, Winiarczyk K, Radłowski M (2002b) Pollen coat
proteins after two-dimensional gel electrophoresis and pollen
wall ultrastructure of Secale cereale and Festuca pratensis. Sex
Plant Reprod 15:75–83
Kunz C, Chang A, Faure JD, Clarke AE, Poly GM, Anderson MA
(1996) Phosphorylation of style S-Rnases by Ca+2-dependent
protein kinase from pollen tubes. Sex Plan Reprod 9:25–34
Lindsey K, Casson S, Chilley P (2002) Peptides: new signalling
molecules in plants. Trends Plant Sci 7:78–93
Mal TK, Hermann BP (2000) Quantitative evaluation of pollen
polymorphism in a tristylous weed, Lythrum salicaria (Lythra-
ceae). Can J Bot 78:1086–1094
Michaud D, Faye L, Yelle S (1993) Electrophoretic analysis of plant
cysteine and serine proteinases using gelatin-containing poly-
acrylamide gels and class-specific proteinase inhibitors. Electro-
phoresis 14:94–98
Mulcahy DL (1974) Correlation between speed of pollen tube growth
and seedling height in Zea mays. Nature 249:491–493
Mulcahy DL, Caporello D (1970) Pollen flow within a tristylous
species: Lythrum salicaria. Am J Bot 57:1027–1030
Mulcahy DL, Mulcahy GB (1975) The influence of gametophytic
competition on sporophytic quality in Dianthus chinensis. Theor
Appl Genet 46:277–280
Murphy DJ, Ross JHE (1998) Biosynthesis, targeting and processing
of oleosin-like proteins, which are major pollen coat components
in Brassica napus. Plant J 13:1–16
de Nettancourt D (2001) Incompatibility and incongruity in wild and
cultivated plants, 2nd edn. Springer, Berlin, p 78
O’Neil P (1992) Variation in male and female reproductive success
among floral morphs in the tristylous plant Lythrum salicaria(Lythraceae). Am J Bot 81:76–84
O’Neil P (1994) Genetic incompatibility and offspring quality in the
tristylous plant Lythrum salicaria (Lythraceae). Am J Bot
79:1024–1030
Sex Plant Reprod (2007) 20:51–62 61
123

Ottaviano E, Mulcahy DL (1989) Genetics of angiosperm pollen. Adv
Genet 26:1–64
Ottenbreit KA, Staniforth RJ (1994) Crossability of naturalized and
cultivated Lythrum taxa. Can J Bot 72:337–341
Radłowski M, Kalinowski A, Adamczyk J, Krolikowski Z, Bart-
kowiak S (1996) Proteolytic activity in the maize pollen wall.
Physiol Plant 98:172–178
Richards AJ (1986) Plant breeding systems. George Allen and Unwin,
London
Roggen HPJR, Stanley RG (1969) Cell-wall-hydrolyzing enzymes in
wall formation as measured by pollen tube extension. Planta
84:295–303
Sage TL, Struma F, Cole WW, Barrett SCH (1999) Differential ovule
development following self- and cross-pollination: the basis of
self-sterility in Narcisus triandrus (Amarylidaceae). Am J Bot
86:855–870
Scribailo R, Barrett SCH (1991) Pollen-pistil interactions in tristylous
Pontederia sagittata (Pontederiaceae). II. Patterns of pollen tube
growth. Am J Bot 78:1662–1682
Suen F, Wu SSH, Wang HC, Dhugga KS, Huang AHC (2003) Cell
wall reactive proteins in the coat and wall of maize pollen.
Potential role in pollen tube growth on the stigma and through
the style. J Biol Chem 278:43672–43681
Vithanage HIM, Knox RB (1979) Pollen development and quantita-
tive cytochemistry of exine and intine enzymes in sunflower,
Helianthus annuus L. Ann Bot 44:95–106
Winsor JA, Peretz S, Stephenson AG (2000) Pollen competition in
natural population of Cucurbita foetidissima (Cucurbidaceae).
Am J Bot 87:527–532
Wong KC, Watanabe M, Hinata K (1994) Protein profiles in pin and
thrum floral organs of distylous Averrhoa carambola L. Sex
Plant Reprod 7:107–115
Zannis VI, Breslow JL (1981) Human very low density lipoprotein
apolipoprotein E, isoprotein polymorphism is explained by
genetic variation and post-translational modification. J Am
Chem Soc 20:1033–1041
62 Sex Plant Reprod (2007) 20:51–62
123




















