
Trophic polymorphism, habitat and diet segregation in Percichthys trucha (Pisces: Percichthyidae) in...
24
Biological Journal of the Linnean Society (1998), 65: 191–214. With 7 figures Article ID: bj980245 Trophic polymorphism, habitat and diet segregation in Percichthys trucha (Pisces: Percichthyidae) in the Andes DANIEL E. RUZZANTE 1 *, SANDRA J. WALDE 1 , VICTOR E. CUSSAC 2,3 , PATRICIO J. MACCHI 2 AND MARCELO F. ALONSO 2 1 Department of Biology, Dalhousie University, Halifax, NS, Canada B3H 4J1 2 Centro Regional Universitario Bariloche, Universidad Nacional del Comahue, 8400 Bariloche, Rio Negro, Argentina 3 Consejo Nacional de Investigaciones Cientificas y Te ´cnicas (CONICET), Argentina Received 18 January 1997; accepted for publication 23 March 1998 Divergent natural selection affecting specific trait combinations that lead to greater efficiency in resource exploitation is believed to be a major mechanism leading to trophic polymorphism and adaptive radiation. We present evidence of trophic polymorphism involving two benthic morphs within Percichthys trucha, a fish endemic to temperate South America. In a series of lakes located in the southern Andes, we found two morphs of P. trucha that could be distinguished on the basis of gill raker length and five other morphological measures, most of which are likely associated with the use of food resources. The differences were consistent across all lakes examined, and were correlated with habitat use and diet. Individuals with longer gill rakers were more abundant in the littoral zone (littoral morph) while the short gill-raker morph was more abundant at 10 m depth and deeper (deep benthic morph). Both morphs fed primarily on benthic invertebrates, but the littoral morph fed more on larval Anisoptera than did the deep benthic morph. Phenotypic correlations among traits were high for the littoral morph, but low and non-significant for the deep-benthic morph. We suggest that gill raker length may influence the relative efficiency of suction feeding for the two morphs. This is the first evidence of trophic polymorphism in fishes from temperate South America. 1998 The Linnean Society of London ADDITIONAL KEY WORDS:—trophic morphology – gill raker – competitive divergence – habitat use – endemics – Patagonia. CONTENTS Introduction . . . . . . . . . . . . . . . . . . . . . . . 192 The Percichthys complex . . . . . . . . . . . . . . . . . . 193 Geological background of northwestern Patagonian lakes . . . . . . 194 Material and methods . . . . . . . . . . . . . . . . . . . 194 * Correspondence to: Daniel E. Ruzzante. E-mail: [email protected]. Present address: Danish Institute for Fisheries Research, Department of Inland Fisheries, Population Genetics Lab, Vejlsoevej 39, DK-8600, Silkeborg, Denmark. 191 0024–4066/98/100191+24 $30.00/0 1998 The Linnean Society of London
Transcript of Trophic polymorphism, habitat and diet segregation in Percichthys trucha (Pisces: Percichthyidae) in...

Biological Journal of the Linnean Society (1998), 65: 191–214. With 7 figures
Article ID: bj980245
Trophic polymorphism, habitat anddiet segregation in Percichthys trucha(Pisces: Percichthyidae) in the Andes
DANIEL E. RUZZANTE1∗, SANDRA J. WALDE1, VICTOR E. CUSSAC2,3,PATRICIO J. MACCHI2 AND MARCELO F. ALONSO2
1 Department of Biology, Dalhousie University, Halifax, NS, Canada B3H 4J1 2 CentroRegional Universitario Bariloche, Universidad Nacional del Comahue, 8400 Bariloche, RioNegro, Argentina 3 Consejo Nacional de Investigaciones Cientificas y Tecnicas (CONICET),Argentina
Received 18 January 1997; accepted for publication 23 March 1998
Divergent natural selection affecting specific trait combinations that lead to greater efficiencyin resource exploitation is believed to be a major mechanism leading to trophic polymorphismand adaptive radiation. We present evidence of trophic polymorphism involving two benthicmorphs within Percichthys trucha, a fish endemic to temperate South America. In a series oflakes located in the southern Andes, we found two morphs of P. trucha that could bedistinguished on the basis of gill raker length and five other morphological measures, mostof which are likely associated with the use of food resources. The differences were consistentacross all lakes examined, and were correlated with habitat use and diet. Individuals withlonger gill rakers were more abundant in the littoral zone (littoral morph) while the shortgill-raker morph was more abundant at 10 m depth and deeper (deep benthic morph). Bothmorphs fed primarily on benthic invertebrates, but the littoral morph fed more on larvalAnisoptera than did the deep benthic morph. Phenotypic correlations among traits werehigh for the littoral morph, but low and non-significant for the deep-benthic morph. Wesuggest that gill raker length may influence the relative efficiency of suction feeding for thetwo morphs. This is the first evidence of trophic polymorphism in fishes from temperateSouth America.
1998 The Linnean Society of London
ADDITIONAL KEY WORDS:—trophic morphology – gill raker – competitive divergence– habitat use – endemics – Patagonia.
CONTENTS
Introduction . . . . . . . . . . . . . . . . . . . . . . . 192The Percichthys complex . . . . . . . . . . . . . . . . . . 193Geological background of northwestern Patagonian lakes . . . . . . 194
Material and methods . . . . . . . . . . . . . . . . . . . 194
∗Correspondence to: Daniel E. Ruzzante. E-mail: [email protected]. Present address: DanishInstitute for Fisheries Research, Department of Inland Fisheries, Population Genetics Lab, Vejlsoevej39, DK-8600, Silkeborg, Denmark.
1910024–4066/98/100191+24 $30.00/0 1998 The Linnean Society of London

D. E. RUZZANTE ET AL192
Study sites and collection methods . . . . . . . . . . . . . . 194Morphological measurements and statistical analysis . . . . . . . . 195
Results . . . . . . . . . . . . . . . . . . . . . . . . 198Species composition . . . . . . . . . . . . . . . . . . . 198Morphological differences between species of the Percichthys complex . . . 199Morphological types within Percichthys trucha . . . . . . . . . . . 199Morphology and resource use . . . . . . . . . . . . . . . 202
Discussion . . . . . . . . . . . . . . . . . . . . . . . 206Acknowledgements . . . . . . . . . . . . . . . . . . . . 210References . . . . . . . . . . . . . . . . . . . . . . . 211
INTRODUCTION
Understanding the ecological and evolutionary forces responsible for trophicpolymorphism and specialization, and ultimately the mechanisms underlying adaptiveradiation is a fundamental problem in evolutionary biology. Divergent naturalselection in an environment with a heterogeneous distribution of resources can leadto specific combinations of traits that improve the efficiency of resource exploitation.This process of morphological diversification is facilitated by frequency dependentcompetition which, within a single population, is expected to be most intense amongindividuals that are phenotypically similar in ecologically relevant traits (Lack, 1947;Van Valen, 1965; Grant, 1986; Grant & Grant, 1989; Schluter, 1996). If the traitsunder selection are quantitative and mating is assortative with respect to them,divergent natural selection for such traits can gradually transform a population withan initial unimodal distribution into a population with a wider and eventuallybimodal distribution (Doebeli, 1996a,b). The process is also facilitated if there issufficient ‘ecological opportunity’ (Schluter, 1996) or ‘vacant niches’ resulting fromlittle competition from other species.
Trophic polymorphisms have been reported for a number of north-temperatefish inhabiting post-Pleistocene lakes, including sticklebacks (Cresko & Baker, 1996),Arctic charr, Salvelinus alpinus (Skulason, Noakes & Snorrason, 1989; Skulason et al.,1993, 1996; Hindar & Jonsson, 1993; Snorrason et al., 1994; Volpe & Ferguson,1996); bluegill sunfish, Lepomis macrochirus (Ehlinger & Wilson, 1988; Ehlinger, 1990),pumpkinseed sunfish L. gibbosus (Robinson et al., 1993; Robinson, Wilson & Shea,1996; Robinson & Wilson, 1996), rainbow smelt, Osmerus mordax (Taylor & Bentzen,1993), and whitefish (Coregonus sp., Lindsey, 1981; Bernatchez & Dodson, 1991,1994; Vuorinen et al., 1993; Bernatchez et al., 1996). Reports from the tropics arefewer, and are generally restricted to cichlids and other fishes from the Neotropics(Roberts, 1974; Meyer, 1987, 1989; Wimberger, 1991, 1992) and to cichlids fromthe Old World (Fryer & Iles, 1972; Meyer, 1993). In the majority of cases thediversification has occurred in species poor communities and has most often resultedin benthic and pelagic forms. These studies and others of a similar nature (seeRobinson & Wilson, 1994) suggest that trophic polymorphism in lake fish might becommon, and that similar processes may be responsible for the very similar patternsof diversification observed in different groups (Schluter, 1996).
Here we present evidence of a new type of trophic polymorphism involving twobenthic morphs in a fish (Percichthys trucha (Cuvier & Valenciennes 1840) Girard1854) inhabiting temperate lakes in the Andes of southern South America. We findtwo morphs inhabiting each lake, and although both morphs feed on benthos, the

TROPHIC POLYMORPHISM IN ANDEAN FISH 193
morphological differences are associated with differences in diet and habitat pref-erence. Our results indicate that the pattern of intraspecific diversification in species-poor lake habitats may be even more general than commonly recognized. Inaddition, the pattern of diversification we describe for P. trucha in these lakes, wherethe species occurs alone, parallels, although is not as extreme as, interspecificdifferences when P. trucha coexists with closely related species in the Percichthyscomplex. Thus within this species complex, as within other fish groups, the patternsof diversification within populations may be at least qualitatively similar to thosemanifesting themselves on macroevolutionary timescales and seen at the interspecificlevel. It may therefore be possible to gain an insight into the process of adaptiveradiation by studying the mechanisms underlying polymorphisms within populations(Robinson et al., 1996; Schluter & McPhail, 1993; Schluter, 1996; Losos, Warheit& Schoener, 1997).
The Percichthys complex
The family Percichthyidae has traditionally been regarded as composed of c. 50freshwater or estuarine species distributed across North and South America, Australiaand Asia (Moyle & Cech, 1982), although more recent studies consider this assemblageas polyphyletic and restrict the family to fresh and brackish water genera of Australiaand South America ( Johnson, 1984, 1993). In South America there are five currentlyrecognized living species within the genus Percichthys (Ringuelet, Aramburu & Alonsode Aramburu, 1967; Arratia, 1982; Arratia, Penafort & Menu Marque, 1983; Lopez,Menni & Miquelarena, 1987; Lopez-Albarello, 1993; Campos & Gavilan, 1996).The Percichthys complex, as is much of the native fish fauna in the Austral bio-geographic region of South America (Ortubay et al., 1994), is endemic to southernArgentina and Chile (Arratia et al., 1983; Almiron et al., 1997), and some of thespecies in the complex are protected within parts of their ranges. Despite theirstatus, they have received relatively little attention. Early studies focused on thedescription of the general and breeding biology of some of the species (GonzalezRegalado, 1945; Mastrarrigo, 1948; Fuster de Plaza & Plaza, 1955; Boschi & Fusterde Plaza, 1957), while more recent efforts have focused on their geographicaldistributions (Arratia, 1982; Arratia et al., 1983), diets (Ferriz, 1989, 1993; Macchi,1991, 1993 and unpublished data), and growth patterns (Guerrero, 1991) in a limitednumber of locations.
Percichthys trucha is the most widely distributed species, both latitudinally andaltitudinally (Arratia et al., 1983) and occurs on both sides of the Andes. One species,P. melanops, occurs exclusively on the western side of the Andes (in Chile), whereasthe remaining three species (P. colhuapiensis, P. vinciguerrai, and P. altispinnins) havesomewhat more restricted geographical distributions and are found in the lakes,streams, and reservoirs of northern, northwestern, and central Patagonia in southernArgentina, a region with otherwise a very sparse native fish fauna (Ringuelet et al.,1967; Lopez et al., 1987; Ortubay et al., 1994). Thus all five species show distributionsthat are partially overlapping. Some of the species have been shown to exhibit amarked degree of intraspecific morphological variation in trophic related charactersboth within and between lakes and some of this variation has tentatively beenattributed to character displacement due to competition for food resources in thepresence of a closely related species (Cussac et al., 1998).

D. E. RUZZANTE ET AL194
Geological background of northwestern Patagonian lakes
The lakes included in this study are developed over a basement of crystallinerocks of Paleozoic age, which were partially covered by sedimentary, volcanic andglacial deposits. The landscape was shaped, in part, by Pleistocene-Holocenevolcanism, but predominantly by glaciation, which is responsible for the deep andwide glacial valleys now occupied by the lakes, and for the coarse gravels and sandsof glacial deposits. The region, now located near the continental divide, with anelevation ranging from 1000 to 2000 m a.s.l., attained in preglacial times, drainstowards the Atlantic Ocean, about 700 km distant, and some of the lakes are isolatedfrom the sea. Lake populations in southwestern Argentina were established after theretreat of Pleistocene glaciers, probably after their last advance between 15 000 and14 500 years b.p. (Mercer, 1976; Lara, 1997). Periodic volcanic activity (mainly inthe form of airborne volcanic ash and mudflows (lahars) causing turbidity) fromvolcanoes located to the west must have stressed the lake populations in the last fewthousand years. Old chronicles tell of intense volcanic activity in the 18th century(Lara, 1997), and there is geomorphological evidence for intense cooling during the‘Little Ice Age’ in the 18th and 19th centuries (Villagran, 1991).
MATERIAL AND METHODS
Study sites and collection methods
We examined the variation in morphology and diet for Percichthys trucha in sixlakes located in the Limay river basin in the Andean forest of southwestern Argentina(Fig. 1 and Table 1). The lakes visited were: Ruca Choroi, Quillen, Falkner-Villarino,Correntoso, Espejo, and Morenito. The two northernmost lakes, Ruca Choroi andQuillen are geographically closer to each other than they are to the other lakes,and drain into the Alumine river, which in turn drains into the larger Limay River.Lakes Villarino and Falkner are interconnected through a short (c. 100 m) river,and thus for the purposes of this study we consider them a single lake. Lake Falkneris downstream of Villarino and drains into the Limay River (Fig. 1). Lakes Correntosoand Espejo are further south and drain into the larger Nahuel Huapi, from whichthe Limay river originates. The southernmost samples were collected in LakeMorenito, the smallest of the six lakes (Table 1), and which also drains directly intoLake Nahuel Huapi (Fig. 1). All samples were collected between January and March1996; Lakes Falkner-Villarino, Correntoso, and Espejo were visited twice duringthis period.
Fish (Percicthys trucha and other species, see Tables 2 and 3) were collected fromtwo sites in each lake. One set of gillnets (stretched mesh sizes ranging from 30 to140 mm) was placed near standing or submerged vegetation (Schoenoplectus californicusMeyer Steud, and Potamogeton linguatus Hangstrom, respectively). In Lakes Correntosoand Espejo, these sites were within enclosed or semi-enclosed bays. The second setwas placed near rocky, exposed coastline with steep bottom gradient and mud, sand,or stone substrate with little or no vegetation. When feasible, gillnets were set atthree depths in each site: in the littoral zone near the surface of the water, and at10 m and 20 m depth in the limnetic zones near the bottom. The nets were put in

TROPHIC POLYMORPHISM IN ANDEAN FISH 195
Argentina
70°
SouthAtlantic
72°
40°
70°
40°
Rio Negro
Piedra delAguila reservoir
Limayriver
Ramos Mexiareservoir
Limayriver
NahuelHuapilake
1
2
34
5
6
N
100 km0 50
Collon Curariver
Neuquen
Alicurareservoir
Alumineriver
Figure 1. Geographic location of lakes. All lakes are situated within the Limay River basin, Provincesof Neuquen and Rio Negro, Argentina. 1: Ruca Choroi. 2: Quillen. 3: Falkner-Villarino. 4: Espejo.5: Correntoso. 6: Moreno. Some major lakes in the area were not sampled. They are shown forcompleteness, but are not labelled for clarity.
T 1. Physical characteristics of six northwestern Patagonian lakes
Lake Latitude Longitude Altitude Surface Area(°S) (°W) (m) (km2)
Ruca Choroi 39°14′ 71°11′ >1000 3.7Quillen 39°25′ 71°15′ 975 33.8Falkner-Villarino 40°27′ 71°32′ >850 17.6Correntoso 40°40′ 71°40′ >800 17.6Espejo 40°38′ 71°45′ >800 19.2Morenito 41°02′ 71°32′ 764 0.08
place before dusk and hauled in after dawn the following morning. All fish wereimmediately removed and placed in 4% formalin.
Morphological measurements and statistical analysis
All measurements were taken from preserved material. For all individuals (n=219) we measured standard length (SL), head length (HL), mouth width (MW),

D. E. RUZZANTE ET AL196
T
2.N
umbe
ran
dle
ngth
(mm
)of
non-
Perc
icht
hyid
aeca
ught
ingi
ll-ne
tsa
mpl
es
Lak
eO
ncor
hync
hus
myk
iss
Sal
velinu
sfo
ntin
atis
Sal
mo
trut
taO
dont
esth
esha
tche
riG
alax
ias
plat
eiD
iplo
mys
tes
vied
men
sis
Ruc
aC
horo
inu
mbe
rof
fish
1611
––
2–
mea
nle
ngth
(SD
)30
1.9(
65.5
)33
6.4(
51.4
)29
7.5
med
ian
leng
th31
034
0ra
nge
ofle
ngth
s18
5–42
525
5–40
027
0–32
5Q
uille
nnu
mbe
rof
fish
205
10–
––
mea
nle
ngth
(SD
)37
6.8
(82.
3)22
9(3
9.7)
493.
5(5
4.8)
med
ian
leng
th39
7.5
240
490
rang
eof
leng
ths
180–
490
180–
270
410–
580
Falk
ner-
Vill
arin
onu
mbe
rof
fish
1317
1–
1–
mea
nle
ngth
(SD
)26
0(1
38.1
)25
4.1
(82.
1)42
020
0m
edia
nle
ngth
185
235
–ra
nge
ofle
ngth
s10
0–47
016
0–42
0–
Cor
rent
oso
num
ber
offis
h12
42
––
–m
ean
leng
th(S
D)
371.
2(1
34.6
)28
1.2
(42.
7)43
2.5
(109
.6)
med
ian
leng
th43
2.5
262.
5ra
nge
ofle
ngth
s10
5–48
525
5–34
535
5–51
0E
spej
onu
mbe
rof
fish
147
–3
1m
ean
leng
th(S
D)
359.
5(8
2.5)
274.
2(9
6.3)
198.
3(7
0.9)
med
ian
leng
th35
029
018
5ra
nge
ofle
ngth
s21
5–46
010
0–37
513
5–27
5M
oren
itonu
mbe
rof
fish
–2
–15
––
mea
nle
ngth
(SD
)26
2.5
(10.
6)26
7.4
(37.
7)m
edia
nle
ngth
268
rang
eof
leng
ths
255–
270
210–
330

TROPHIC POLYMORPHISM IN ANDEAN FISH 197
MW
HLUJ
DF
SL
CP
Figure 2. Body measurements. SL: standard length. HL: head length. UJ: length of upper jaw. MW:mouth width. DF: height of dorsal spine. CP: caudal peduncle depth. In addition to these externalmorphological measurements, gill raker length and number were also obtained.
length of the upper jaw (UJ), depth of the caudal peduncle (CP), and height of thedorsal fin (DF) (Fig. 2). We also measured the length of the four longest gill rakerson the first left branchial arch (These were usually the rakers located side by sideclosest to the V angle between the two branches of the arch). Gill rakers were firstdrawn under a stereomicroscope and later measured on a digitizing table. Allcharacters are likely related to feeding and/or swimming ability. In addition, headlength, mouth width, upper jaw length, and depth of caudal peduncle are diagnosticvariables for species identification within the Percichthyidae (Ringuelet et al., 1967).
To control for differences among the fish in body size, we standardized allmeasured traits with respect to standard length using the relationship:
Yi=log (Xi) – b [log (SLi) – Mean (log (SL)] (1)
where Yi and Xi are the adjusted and original values for the character in individualsi (i=1, . . . ., N), SLi is the individual standard length, and b is the regressioncoefficient of the logarithm of X on the logarithm of SL. Allometric relationshipsdid not differ between sexes (data not shown), but they differed slightly among lakesfor some of the variables. Variables were therefore standardized with both sexespooled, but separately for each lake.
To determine whether there were consistent morphological types among lakes,we classified individual into two groups using cluster and discriminant analysis ofthe standardized measurements. The analyses were conducted by lake, and we thencompared the morphology of the two groups for each lake.
To further look at the separation of the morphological types, we conductedprincipal component analysis, pooling the measurements (already standardized bylake) from all lakes. Prior to pooling, all standardized measures were brought to a

D. E. RUZZANTE ET AL198
T 3. Number and size of Percichthys trucha of each morphotype collected in six NorthwesternPatagonian lakes
Lake Morphotype Number Length (mm) Proportionof fish females
mean median range (%)
Ruca Choroi littoral 9 254.2 240.0 125–395 44.4deep benthic 6 247 232.5 172–335 50.0
Quillen littoral 51 218.3 220.0 140–355 45.1deep benthic 28 216.9 220.0 130–340 35.7
Falkner-Villarino littoral 15 315.6 325.0 195–370 40.0deep benthic 5 324.6 308.0 290–390 20.0
Correntoso littoral 8 318.1 322.5 280–350 87.5deep benthic 12 332.5 337.5 230–370 25.0
Espejo littoral 16 336.9 357.5 195–395 56.3deep benthic 9 330.0 370 165–420 33.3
Morenito littoral 12 302.1 305 265–340 50.0deep benthic 37 312.2 320 196–390 64.9
common mean using each character- and lake-specific regression coefficient. Con-fidence and prediction (Whitmore, 1986) intervals were calculated for each of theprincipal components.
DietWe obtained information on diet by examining the stomach contents of all
individuals collected from the lakes. Only stomach contents, and not those of theintestine, were included in our analysis to minimize bias arising from differentialpassage rates of prey. For each prey category, i, (i.e. family, genus, or species), wecalculated an index of relative importance, RIi (Pinkas, Oliphant & Iverson 1971):
RIi=(Ni + Vi) (Fi), (2)
where Ni is the percentage of total prey items that were prey of type i, Vi is thepercentage of total prey volume occupied by prey of type i, and Fi is the percentageof non-empty guts that contained at least one prey of type i. Diet overlap betweenpairs of morphs within lakes was determined by comparing the relative importanceindex for each food category.
For lakes Falkner-Villarino, Correntoso, and Espejo, which were visited twicewithin a period of 4 weeks, the data were pooled across dates. In lakes Quillen andRuca Choroi we collected a large number of relatively small fish which weregenerally absent from the samples from other lakes (Table 3). For lake Quillen,where the sample size was relatively large (n=85) we analyzed diet separately forlarge (>250 mm) and small (<250 mm) fish. We were unable to analyze diet separatelyfor large and small fish from lake Ruca Choroi because of limited sample size (n=15) and because the majority were relatively small (Table 3).
RESULTS
Species composition
A number of native and introduced species of fish coexist with the Percichthyidaein these north-western Patagonian lakes, and we had seven such species in our

TROPHIC POLYMORPHISM IN ANDEAN FISH 199
samples, four native and three introduced species (Table 2). Of the introducedspecies, the salmonids, Oncorhynchus mykiss, Salvelinus fontinalis, and Salmo trutta weremost common (Table 2). Overall, the most abundant of these species was O. mykisswith individuals found in all lakes except Lake Morenito. Salvelinus fontinalis weresecond in abundance and were found in all six lakes. Salmo trutta were found inthree lakes (Quillen, Falkner-Villarino, Correntoso). Species composition in LakeMorenito was quite different from that in the other lakes. First, we caught no O.mykiss, and second, the native Odontesthes hatcheri, absent from our samples from theother lakes, were relatively common (Table 2).
Morphological differences between species of the Percichthys complex
A total of 219 ‘percas’ were collected from the six lakes (Table 3). In the foursouthern lakes, most fish were reproductively mature adults, but a significant numberof juveniles were caught in Quillen and Ruca Choroi. Five living species arerecognized within Percichthyidae (Ringuelet et al., 1967; Arratia et al., 1982), howeverwe expected these lake ‘percas’ to be Percichthys trucha since only this species has beenreported for the region containing the study lakes (Ringuelet et al., 1967; Arratia etal., 1982; Bruzone, 1986). To determine if our lake ‘percas’ were indeed P. trucha,we compared their morphology with percas previously collected in two reservoirson the Limay river, approximately 150 km to the east of the lakes (Fig. 1). Threespecies, Percichthys colhuapiensis, P. trucha and P. vinciguerrai are found in one reservoir(Piedra del Aguila), and only two of these, P. trucha and P. vinciguerrai, are found inthe other (Alicura; Cussac et al., 1998). P. colhuapiensis (the ‘large-mouthed’ species)typically have longer heads, longer upper jaws and wider mouths than do P. truchaor P. vinciguerrai, although there can be some overlap. P. vinciguerrai is separated fromP. trucha by its longer dorsal spine. The lake ‘percas’ were clearly not P. vinciguerrai,as their dorsal spines were shorter than the P. vinciguerrai from the reservoirs (Fig.3A). However, for most diagnostic variables the lake ‘percas’ were intermediatebetween the P. colhuapiensis and P. trucha from Piedra del Aguila (Fig. 3B,C,D). Inparticular, upper jaws and heads were distinctively longer (Fig. 3B,C), and mouthswere wider (Fig. 3D) for P. colhuapiensis from Piedra del Aguila, than for most lake‘percas’. The lake ‘percas’ in turn, exhibited upper jaws and heads as well as mouths,that were longer and wider, respectively, than those shown by P. trucha from Piedradel Aguila. This evidence, together with the fact that for the three diagnosticvariables shown in Figure 3, our lake ‘percas’ overlap considerably with P. truchafrom Alicura, where the species exists in allopatry (Cussac et al., 1988) suggest thatour lake ‘percas’ are indeed P. trucha. In addition, preliminary mtDNA evidencesuggests that only one species is present in these lakes (Stewart et al. unpublisheddata). Conclusive identification of the fish as P. trucha awaits further genetic analysis.
Morphological types within Percichthys trucha
Although only one species seems to be present, we found two major morphologicaltypes in each lake. We clustered the fish by lake, using the six morphometricvariables: head length, mouth width, length of upper jaw, peduncle height, dorsalspine length and gill raker length. In each lake, two distinct clusters emerged, where

D. E. RUZZANTE ET AL200
100
140
Standard length
Hea
d le
ngt
h
40
ARSR
AL
200 300 400
120
100
60
80
C
100
40
Standard length
Mou
th w
idth
10
AR
SR
AL
200 300 400
30
20
D
150
Spi
ne
len
gth
40
200 300 400
20
30
A
100
40
Upp
er ja
w le
ngt
h
10
AR
SR
AL
200 300 400
30
20
B
250 350
50 50
Figure 3. Relationships between standard length and four morphometric characters for Percichthys truchafrom six Andean lakes, and Percichthys colhuapiensis, P. trucha and P. vinciguerrai from Piedra del Aguilaand Alicura reservoirs on the Limay River in northwestern Patagonia, Argentina. A, (Η) Percichthysvinciguerrai (longer spine) from the reservoir Piedra del Aguila; (Ο) Percichthys trucha from six lakes. Panels(B), (C), and (D) compare the large-mouthed ‘perca’, P. colhuapiensis (Κ), with allopatric P trucha fromthe lakes (allopatric lakes: AL,Ο), with allopatric P. trucha from the Alicura reservoir (allopatric reservoir:AR, +), and with sympatric P. trucha from Piedra del Aguila reservoir (sympatric reservoir: SR, Β).Notice that sympatric P. trucha (small-mouthed ‘perca’) from Piedra del Aguila (SR) are consistentlymore divergent from P. colhuapiensis (large-mouthed ‘perca’) than any of the allopatric P. truchapopulations. (A) spine height vs. standard length (B) upper jaw length vs. standard length, (C) headlength vs. standard length (D) mouth width vs. standard length.
most fish belonged to one of the two categories with >90% certainty (92% and 87%of the cases in morph 1 and 2, respectively (littoral and deep benthic morphs,respectively, see below). These morphological differences are unlikely to be the resultof ontogenetic niche shifts since there are no differences in size between morphs(Ft.201=0.097, P[0.75, see also Table 3). Gill raker length was the single individualvariable that differed most between groups (Table 4), and we call the group withthe longer gill rakers (for a given size) ‘littoral morph’ (see below) and the groupwith the shorter gill rakers ‘deep benthic morph’. The bimodality in the distributionof gill raker size was evident in each lake (Fig. 4). There was no overlap betweenthe two morphs in Lakes Ruca Choroi and Correntoso, and minimal overlap in the
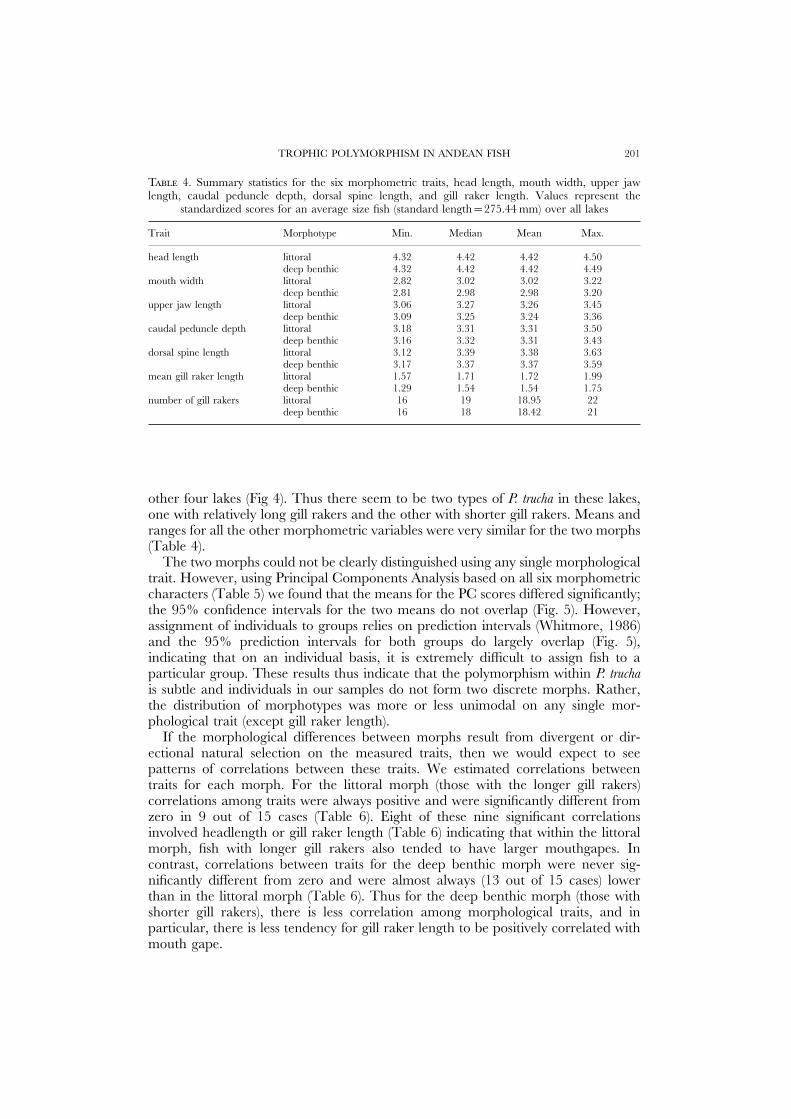
TROPHIC POLYMORPHISM IN ANDEAN FISH 201
T 4. Summary statistics for the six morphometric traits, head length, mouth width, upper jawlength, caudal peduncle depth, dorsal spine length, and gill raker length. Values represent the
standardized scores for an average size fish (standard length=275.44 mm) over all lakes
Trait Morphotype Min. Median Mean Max.
head length littoral 4.32 4.42 4.42 4.50deep benthic 4.32 4.42 4.42 4.49
mouth width littoral 2.82 3.02 3.02 3.22deep benthic 2.81 2.98 2.98 3.20
upper jaw length littoral 3.06 3.27 3.26 3.45deep benthic 3.09 3.25 3.24 3.36
caudal peduncle depth littoral 3.18 3.31 3.31 3.50deep benthic 3.16 3.32 3.31 3.43
dorsal spine length littoral 3.12 3.39 3.38 3.63deep benthic 3.17 3.37 3.37 3.59
mean gill raker length littoral 1.57 1.71 1.72 1.99deep benthic 1.29 1.54 1.54 1.75
number of gill rakers littoral 16 19 18.95 22deep benthic 16 18 18.42 21
other four lakes (Fig 4). Thus there seem to be two types of P. trucha in these lakes,one with relatively long gill rakers and the other with shorter gill rakers. Means andranges for all the other morphometric variables were very similar for the two morphs(Table 4).
The two morphs could not be clearly distinguished using any single morphologicaltrait. However, using Principal Components Analysis based on all six morphometriccharacters (Table 5) we found that the means for the PC scores differed significantly;the 95% confidence intervals for the two means do not overlap (Fig. 5). However,assignment of individuals to groups relies on prediction intervals (Whitmore, 1986)and the 95% prediction intervals for both groups do largely overlap (Fig. 5),indicating that on an individual basis, it is extremely difficult to assign fish to aparticular group. These results thus indicate that the polymorphism within P. truchais subtle and individuals in our samples do not form two discrete morphs. Rather,the distribution of morphotypes was more or less unimodal on any single mor-phological trait (except gill raker length).
If the morphological differences between morphs result from divergent or dir-ectional natural selection on the measured traits, then we would expect to seepatterns of correlations between these traits. We estimated correlations betweentraits for each morph. For the littoral morph (those with the longer gill rakers)correlations among traits were always positive and were significantly different fromzero in 9 out of 15 cases (Table 6). Eight of these nine significant correlationsinvolved headlength or gill raker length (Table 6) indicating that within the littoralmorph, fish with longer gill rakers also tended to have larger mouthgapes. Incontrast, correlations between traits for the deep benthic morph were never sig-nificantly different from zero and were almost always (13 out of 15 cases) lowerthan in the littoral morph (Table 6). Thus for the deep benthic morph (those withshorter gill rakers), there is less correlation among morphological traits, and inparticular, there is less tendency for gill raker length to be positively correlated withmouth gape.

D. E. RUZZANTE ET AL202
10
Mean gill raker length [ln(mm)]1.2
8
6
4
2
01.4 1.6 1.8 2.0
Espejo10
1.2
8
6
4
2
01.4 1.6 1.8 2.0
Morenito
10
Fre
quen
cy
1.2
8
6
4
2
01.4 1.6 1.8 2.0
Falkner & Villarino 10
1.2
8
6
4
2
01.4 1.6 1.8 2.0
Correntoso
10
1.2
5
01.4 1.6 1.8 2.0
Ruca Choroi
10
1.2
8
6
4
2
01.4 1.6 1.8 2.0
Quillen1512
12
LittoralDeep benthic
Figure 4. Empirical probability density (frequency) distributions of mean gill raker length [1n(mm)]for the littoral and deep benthic morphs within Percichthys trucha in six Andean lakes. The area undereach curve is equal to 1.
Morphology and resource use
HabitatAlthough individuals of both morphs were found consistently at all three collection
depths (littoral, 10 m, 20 m), the two morphs differed in their depth distribution(Fisher’s exact test, P=0.011, two sided test): morph 1 (littoral morph) was more

TROPHIC POLYMORPHISM IN ANDEAN FISH 203
T 5. Principal Component Analysis loadings distinguishing two trophic morphs on the basis ofsix morphometric variables (i.e. head length, mouth width, upper jaw length, caudal peduncle height,
dorsal fin spine length, gill raker length)
Trait PC1 PC2 PC3 PC4 PC5 PC6
head length 0.428 −0.095 0.615 0.391 0.299 −0.433mouth width 0.476 −0.299 −0.151 −0.289 −0.652 −0.390upper jaw length 0.518 −0.408 0.047 −0.005 0.113 0.742caudal peduncle 0.252 0.577 0.378 −0.668 0.066 0.103dorsal spine length 0.302 0.615 −0.136 0.560 −0.398 0.200gill raker length 0.409 0.155 −0.660 −0.068 0.557 −0.243% of total variance 32.6 18.3 15.4 13.8 11.4 8.5
2
PC
A 4
–2
3PCA 3
210–2 –1
–1
1
0
2
PC
A 5
–2
3PCA 3
210–2 –1
–1
1
0
2
PC
A 1
–2
3210–2 –1–4
4
0
2
PC
A 2
–2
3210–2 –1
–1
1
0
–3
Figure 5. Scatterplot of Principal Component Analysis (PCA) scores for Percichthys trucha of the littoralmorph (Κ) and of the deep benthic morph (Ο). The symbol + are the group means for the twomorphotypes, and the small ellipses around them are the 95% confidence intervals for the two groupsindicating that on a population wide basis the two morphs differ significantly. The large ellipses arethe 95% prediction intervals and indicate that on an individual basis there is a substantial degree ofoverlap between the two morphs. A, Principal Components 1 vs. 3. B, Principal Components 2 vs. 3.C, Principal Components 4 vs. 3. D, Principal Components 5 vs. 3.

D. E. RUZZANTE ET AL204
T 6. Correlations between standardized morphological variables including gill raker length. Topvalues are for the littoral morph and bottom values are for the deep benthic morph
Morphotype Mouth width Upper jaw Peduncle Spine Gill raker length
head length littoral 0.25∗ 0.45∗∗ 0.23∗ 0.31∗∗ 0.23∗deep benthic 0.10 0.20 0.07 −0.04 0.00
mouth width littoral 0.53∗∗ 0.15 0.14 0.15deep benthic 0.20 0.05 0.02 0.10
upper jaw littoral 0.06 0.11 0.27∗∗deep benthic 0.10 −0.03 0.20
caudal littoral 0.13 0.25∗peduncle deep benthic 0.20 0.04spine length littoral 0.26∗∗
deep benthic 0.20
∗: P<0.05; ∗∗: P<0.01.
70
Collection depth (m)10
Nu
mbe
r of
indi
viu
als
60
50
40
308642
Figure 6. Number of individuals of P. trucha collected in the littoral zone and at depths of 10 m ordeeper in six lakes situated in the Andes of southwestern Argentina. Solid line and symbol: littoralmorph (longer gill rakers). Broken line and open symbol: deep benthic morph (shorter gill rakers).
abundant in the littoral zone than at 10 m depth or deeper, whereas morph 2 (deepbenthic morph) was more abundant in the deeper collections (Fig. 6).
DietAnalysis of stomach contents showed that differences in diet among lakes were
generally minor and in most cases consisted of differences in the relative importanceof prey common to most lakes, rather than of differences in the type of prey (Table7). Depending on availability, ‘percas’ from these lakes fed on aquatic insects(Anisoptera, Zygoptera, Trichoptera, Ephemeroptera, and Chironomidae), as wellas on amphipods, terrestrial insects, crabs (Aegla sp.), and prawns (Samastacus sp.).Regardless of morphotype, larval Anisoptera were always the single most importantprey in all lakes (Table 7). Larval Anisoptera are relatively large and mobileepibenthic prey usually found in the littoral zone in vegetated areas on the substrateor on the vegetation. In five of the six lakes (Ruca Choroi, Quillen, Correntoso,and Falkner-Villarino, and Espejo) Anisoptera tended to be more important in thediet of the littoral morph than in the diet of the deep benthic morph (Wilcoxonpaired test V=15, n=5; P=0.06). In Lake Morenito the observed pattern wasreversed: the littoral morph fed less on Anisoptera than the deep benthic morph

TROPHIC POLYMORPHISM IN ANDEAN FISH 205
T 7. Diet composition of Percichthys trucha by lake and morphotype. Values represent the index ofrelative importance (%) which takes into account the number of prey, their volume and their frequencyof appearance (number of stomachs with that prey type): top values are for the littoral morph and
bottom values are for the deep benthic morph
Prey group Ruca Quillen Falkner- Correntoso Espejo MorenitoChoroi Villarino
Anisoptera larvae 0.67 0.82 0.99 0.57 0.84 0.190.49 0.42 0.75 0.30 0.80 0.60
Zygoptera – – <0.01 – – –– – – – – –
Tricoptera larvae – – <0.05 0.01 – 0.06– – <0.01 <0.01 – –
Tricoptera pupae – – <0.01 – – –– – 0.02 – – –
Ephemeroptera 0.12 – – – – –0.22 – – <0.01 – 0.01
Chironomid larvae 0.14 0.10 – – – 0.380.23 0.02 – – – 0.24
Chironomid pupae 0.03 0.03 – – – –– – – – – –
Terrestrial insects – – – – <0.01 –– – – – – –
Aegla sp. 0.05 – – – – –– – – – 0.05 –
Samastacus sp – – <0.01 – 0.05 –– – 0.17 0.25 0.14 <0.01
Amphipoda <0.01 0.05 – 0.37 0.08 –0.06 0.56 – 0.44 0.02 0.03
Quilina sp. – – – – – 0.12– – – <0.01 – 0.11
Galaxias maculatus – 0.01 <0.01 0.06 0.03 0.25– – 0.05 <0.01 – <0.01
Galaxias platei – – <0.01 – – –– – – – – –
EJ
1.0
Lakes
Rel
ativ
e Im
port
ance
In
dex
for
An
isop
tera
RC RC QL QL FKVR CR
n = 150.8
0.6
0.4
0.2
0.0EJ MR MRCRFKVR
n = 7
n = 8
n = 12
n = 16
n = 9
n = 11
n = 7
n = 9n = 6
n = 7
n = 30
Figure 7. Relative importance index for Anisoptera in the diet of Percichthys trucha. [ΕΦ] Littoral morph.(Φ) deep benthic morph. Lakes: RC: Ruca Choroi. QL: Quillen. FKVR: Falkner-Villarino CR:Correntoso. EJ: Espejo. MR: Morenito. n: number of individuals examined.

D. E. RUZZANTE ET AL206
(Fig. 7). However, individuals of the littoral morph in this lake were feeding primarilyon the native fish Galaxias maculatus (RI=25.0%; Table 7), which was rare in thediet of the benthic morph in this lake (RI=0.1%; Table 7), and in the diets of bothmorphs in the other five lakes (RIΖ5.6%; Table 7). Thus, these results suggest thatthere are differences in the diet of the two morphs, with individuals of the littoralmorph generally eating a greater proportion of Anisoptera than those of the deepbenthic morph. When Galaxias maculatus is abundant, the littoral morph may feedmore heavily on this small native fish.
DISCUSSION
Competition for food resources is widespread among vertebrates and is believedto be a major cause of morphological and behavioral polymorphism in fishes. Thedegree of phenotypic variation among morphs showing differential use of resourcescan vary widely: in some species phenotypic variation is unimodal and barelydetectable, while in others it is bimodal and rivals differences between species(Schluter & McPhail, 1992, 1993; Robinson & Wilson, 1994; Skulason & Smith,1995; Smith & Skulason, 1996). The degree of reproductive isolation among morphscan also vary widely, and it has been suggested that intraspecific trophic polymorphismmay constitute an important intermediate stage in the processes of speciation andadaptive radiation (Bush, 1994).
We have provided evidence of a resource based, trophic polymorphism within asingle species of Percichthyidae, Percichthys trucha, inhabiting several lakes within theAndean region of southwestern Argentina. Two morphs were distinguished primarilyon the basis of gill raker length and, to a lesser extent, on the basis of five externalmorphological characters. Three of the external measures (head length, upper jawlength and mouth width) are directly related to mouthgape, and together with gillraker length, are presumed to be associated with the use of food resources. A fourthexternal measure, depth of caudal peduncle, is presumed to affect swimmingmanoeuvrability, and the fifth, dorsal spine length, is probably associated withsusceptibility to predators. The morphological trait that best discriminated betweenthe two groups was gill raker length, with individuals of the littoral morph exhibitinglonger and slightly more numerous gill rakers than individuals of the deep benthicmorph. Phenotypic correlations between morphological traits were relatively highand significant within the littoral morph but not within the deep benthic morph.Although both morphs fed on benthos, the differences in morphology were correlatedwith diet and with habitat use and they were consistent across all lakes examined.
Gill raker morphology, including length and number of rakers, and spacingbetween them has been described as functionally related to food acquisition in avariety of species (Magnuson & Heitz, 1970; Lindsey, 1981; McPhail, 1984, 1993;Schluter & McPhail, 1993; Robinson & Wilson, 1994 and references therein), andgill raker length in particular, is known to affect foraging efficiency in sticklebacks(Gasterosteus aculeatus complex; Bentzen & McPhail, 1984; Lavin & McPhail, 1986;Schluter, 1993; Schluter & McPhail, 1992, 1993). In most of these examples,however, the variation in gill raker architecture involves benthic and limneticmorphs with limnetic, planktivorous individuals generally exhibiting longer andmore numerous gill rakers which are presumed to increase filter feeding efficiency.
The polymorphism within P. trucha differs from most of these examples in one

TROPHIC POLYMORPHISM IN ANDEAN FISH 207
important respect—it involves two benthic morphs which nevertheless differ inhabitat use and diet composition. Individuals of the littoral morph (longer gill rakers)were more abundant in the littoral zone than at 10 m or 20 m depths, whileindividuals of the deep benthic morph were most abundant in bottom collectionsat depths greater than 10 m. Consistent with these differences in habitat distribution,individuals of the littoral morph tended to feed more on Anisoptera larvae, whichtend to be most abundant in the littoral zone (unpublished data), than did individualsof the deep benthic morph. All species within Percichthyidae have protruding jawsand they capture prey by suction feeding. During suction feeding the mouth cavityis rapidly expanded resulting in a negative (suction) pressure inside the oral cavitydrawing water and prey into the mouth (Lauder, 1983; Lauder, 1985a,b). As pressureinside the mouth cavity drops the branchial apparatus forms a dynamic resistanceto water flow from the opercular opening: gillrakers from adjacent gill bars interlockand thus, effectively close off the posterior wall of the mouth (Lauder & Schaffer,1985). The closing of the branchial apparatus prevents the flow of water from theopercular to the mouth cavity and ensures the unidirectionality of water flow duringthe initial phase. The mouth is then closed and water exits posteriorly through thegill bars and out the opercular opening. This method of prey capture is widespreadamong aquatic lower vertebrates (Lauder, 1985a) and has been observed in taxa asdiverse as bluegill sunfish (Lepomis macrochirus, Lauder, 1983) and larval ambystomatidsalamanders (Lauder & Schaffer, 1985, 1993).
P. trucha likely combines suction feeding with a sudden increase in body velocityto overtake prey, and there is probably a large degree of variation among individualsin the extent to which they combine the two feeding modes: some individuals mayremain relatively stationary, specializing in suction feeding, while others may combinesuction feeding with a rapid acceleration from rest. Individuals with shorter gillrakers (the deep benthic morph) may be more efficient at creating sudden negativepressure within the buccal cavity by interlocking gill-rakers from adjacent bars(Lauder, 1983:10). We thus speculate that the deep benthic morph specializes insuction feeding, while the littoral morph feeds predominantly by combining suctionfeeding with sudden body acceleration. (Note that only in the littoral morph wasgill raker length correlated with caudal peduncle). Coupling suction with accelerationmay enable the littoral morph to more successfully capture mobile prey, as suggestedby the tendency of this morph to feed on small fish when available (e.g. LakeMorenito). It is also possible that variation in some of the other traits, such asmouthwidth and upper jaw length may have as much or more functional relevanceas shown by their loadings on PC1, 2, 5, and 6 (Table 5).
The fact that gill raker length and number have consistently been identified asaffecting feeding performance in a wide variety of species suggests these traits andassociated structures may be under strong natural selection in nature (Schluter, 1994for direct experimental results involving changes in gill raker number). We detectedpositive and significant phenotypic correlations among most of the trophic relatedcharacters in individuals of the littoral morph (long gill rakers) but not in individualsof the deep benthic morph. This raises the possibility that directional natural selectionon the suite of trophic related characters may be operating only within the littoralmorph, i.e. the subset of the P. trucha population specializing on capturing activeand mobile prey such as small fish, but not among ‘percas’ in the other group (deepbenthic morph). Such directional selection for the suite of correlated trophiccharacters within the littoral morph would tend to reduce the phenotypic and

D. E. RUZZANTE ET AL208
additive genetic variance in these characters within this morph but not within thedeep benthic morph. If this scenario is correct then we would expect the heritabilitiesof these characters to be lower within the littoral morph than within the deepbenthic morph. Significant differences in heritabilities between morphs wouldconstitute circumstancial evidence of assortative mating.
We found the primary difference in diet to be a greater consumption of Anisopteralarvae by the littoral morph than by the deep benthic morph. This pattern wascommon to all lakes except Morenito, where the littoral morph tended to concentrateon small fish. Our data on diet, however are restricted to the summer months. Theabundance and size distribution of aquatic invertebrates in temperate lakes changedramatically throughout the year, the pattern depending on specific life histories. Ifkey resources are scarcer in winter (e.g. see Anon Suarez, 1997 for Chironomidae),the intensity of competition may increase. Diet overlap in trophically polymorphicfishes has been shown to diminish during periods of resource limitation or ecologicalbottlenecks (Meyer, 1989; Malmquist et al., 1992; Snorrason et al., 1994), and thuswe might expect the diet differences between morphs to be more pronounced inthe fall and winter.
Since most of the characters that varied between the two morphs of P. trucha canbe related to food acquisition, competition for food resources would seem to be themost likely mechanism by which these differences arose. At present we do not knowwhether the differences in trophic morphology result from environmental influencesduring ontogeny (i.e. phenotypic plasticity), from genetic differences between morphs,or from a combination of both factors (Robinson & Wilson, 1996). Environmentalone (e.g. diet during ontogeny) can induce trophic polymorphism (Meyer, 1987;Wimberger, 1991, 1992). In addition, the two sources of variation may ultimatelybe confounded given that phenotypic plasticity itself may be genetically determined(Via & Lande, 1985; Via et al., 1995; Robinson & Wilson, 1994). In any case, bothsources of phenotypic variation have thus far been found where both have beenexamined. Genetic differences and phenotypic plasticity both contribute to thetrophic polymorphism of pumpkinseed sunfish (Robinson & Wilson, 1996) and tophenotypic divergence of closely-related stickleback species pairs (Day, Pritchard &Schluter, 1994). The relative importance of each mechanism differs, with most ofthe variation in polymorphic pumpkinseed sunfish due to phenotypic plasticity(Robinson & Wilson, 1996), whereas the phenotypic differences between the twosympatric species of sticklebacks are largely genetically based (McPhail, 1984, 1993,1994; see also Taylor, McPhail & Schluter, 1997) with additive, dominance andepistatic effects all playing significant roles in trophically related characters in thestickleback species complex (Hatfield, 1997).
Regardless of whether the morphological polymorphism within P. trucha is en-vironmentally induced and occurs within each generation (i.e. phenotypic plasticity),or results from genetic differences between morphs, the morphological differencesmay possibly be adaptive, as the two morphs differ in their habitat preferences anddiet. Differences in feeding efficiency among morphs in trophically polymorphic fishspecies have been found whenever they have been studied. For example, feedingperformance on soft and hard prey differs between morphs in the trophicallypolymorphic Neotropic cichlid, Cichlasoma citrinellum (Meyer, 1989). Similarly, thetrophic polymorphism described for bluegill sunfish is associated with habitatpreference and habitat specific feeding efficiency (Ehlinger & Wilson, 1988; Ehlinger,1990). Despite an increasing number of examples of trophic polymorphism on a

TROPHIC POLYMORPHISM IN ANDEAN FISH 209
number of vertebrate groups (see Skulason & Smith, 1995), thorough demonstrationsof functional trade-offs or genotype-environment interactions are available only ina limited number of cases. Two such cases are freshwater sticklebacks, Gasterosteusaculeatus complex (Schluter, 1995) and pumpkinseed sunfish, Lepomis gibbossus (Rob-inson et al., 1996). In both examples, benthic and limnetic forms do more poorly ifraised in the alternative environment, and intermediate forms do worse than extremeforms. Morphology and fitness are thus correlated and disruptive selection canoperate between benthic and limnetic forms.
Morphological differences in trophically polymorphic groups may be large orsmall. The morphological differences between pumpkinseed sunfish from littoraland limnetic habitats rarely exceeded 3% for any single body measure, and thepolymorphism could only be detected using multivariate techniques (Robinson etal., 1996). In contrast, benthic and limnetic sticklebacks are easy to distinguishmorphologically, and are considered separate biological species, which neverthelesscan interbreed in the wild (Schluter, 1996; Hatfield, 1997 and references therein).The morphological polymorphism we have described within P. trucha is similar inmagnitude to that in pumpkinseed sunfish; the differences between morphs arerelatively small, and the two types cannot be reliably distinguished by eye. However,the consistency of the polymorphism found in all 6 lakes, lakes that differedconsiderably in other characteristics (Table 1), suggests that this divergence hasoccurred again and again, and thus is likely of ecological significance.
Intraspecific trophic polymorphism resulting from disruptive selection favoringalternative phenotypes in different habitats may constitute an important intermediatestage in adaptive radiation and in the process of sympatric speciation (Bush, 1994;Schluter, 1996). It is of course uncertain whether or not the polymorphism describedwithin P. trucha represents the initial stages in this process. Some of the importantconditions for resource polymorphism to evolve include underutilized resources withsignificant habitat complexity as well as low interspecific competition (Futuyma,1986; Mayr, 1963; Grant, 1986; Schluter & McPhail, 1992, Robinson & Wilson,1994). It is therefore probably no coincidence that most examples of trophicpolymorphism and adaptive radiation in fishes are often found in recently deglaciatedpost-Pleistocene lakes of the Neartic (Robinson & Wilson, 1994; Skulason & Smith,1995; Schluter, 1996), or in volcanic-, rift-, and crater- East African lakes (Fryer &Iles, 1972; Meyer, 1993). The lakes in our study area, are all recently deglaciatedwater bodies (circa 13 000 yrs ago, see Introduction) and are all situated within thefish-species-poor zoogeographic region of Patagonia in southern Argentina and Chile(Ringuelet et al., 1967). In most lakes only two or three other native species arecommon (Table 2) in gillnet, benthic and littoral collections, and in general theseare much smaller species (e.g. Galaxias sp.) that presumably compete little with P.trucha (and in fact occasionally serve as prey). Thus some of the conditions necessaryfor the process of sympatric speciation or at least character release (Van Valen,1965; Robinson & Wilson, 1994) appear to be fullfilled in these lakes.
We do not yet have information on functional trade-offs for the two morphs, justcorrelations between morphology, habitat use and diet. However, there is a strikingparallel between the trophic polymorphism that we see within P. trucha when it isthe only species of Percichthyidae in a lake, and the differences between P. truchaand another closely related species within the Percichthys complex, the large-mouthed‘perca’, P. colhuapiensis, when they co-occur. In a related study, we found thatmorphology of P. trucha differed when the larger-mouthed P. colhuapiensis was present

D. E. RUZZANTE ET AL210
(Cussac et al., 1998). Most of the characters that differed were related to foodacquisition, the small-mouthed species had a smaller mouthgape in the presence ofthe large-mouthed ‘perca’ than in its absence, suggesting that interspecific competitionfor food resources may influence the trophic morphology of P. trucha when it existsin sympatry with P. colhuapiensis. Thus the two morphs of P. trucha may be movingtoward occupying the niches otherwise occupied by P. trucha and P. colhuapiensis whenthey coexist in a single lake.
This is the first time that a trophic polymorphism has been reported for fish fromtemperate South America. The entire Percichthys complex is endemic to southernArgentina and Chile as indeed are most of the native species in this region (Ringueletet al., 1967; Arratia et al., 1983). Trophic polymorphisms have of course beenreported for a number of northern hemisphere temperate fish (reviewed in Schluter& McPhail, 1992, 1993; Robinson & Wilson, 1994; Skulason & Smith, 1995; Smith& Skulason, 1996, see also Introduction). Our findings suggest that the pattern oftrophic related polymorphism in fishes is even more general and widespread thanpreviously reported, as predicted by Schluter (1996).
Finding trophic polymorphism in the Percichthys complex is also significant for theconservation of native, endemic resources. North temperate salmonids, which havebeen introduced into Patagonian lakes and rivers over the past 100 years (GonzalezRegalado, 1945), are known to have an ecological impact on native Australian andNew Zealand Galaxidae (Tilzey, 1976; McDowall, 1990; Crowl, Townsend &McIntosh, 1992; Glova Sagar & Naslund, 1992; McIntosh, Crowl & Townsend,1994). Introduced salmonids are believed to also be adversely affecting some of thePatagonian native fish likely including Galaxias sp. and Diplomystes viedmensis (Cussacet al. unpublished observations). However, the impact of salmonids on the Patagoniannative fish fauna, and in particular on the various species of Percichthys has receivedlittle attention. Clearly the potential for influencing the nature and intensity of inter-and intraspecific competition within the Percicthyidae exists.
In conclusion, we have identified a new type of trophic polymorphism involvingtwo benthic morphs (littoral versus deep-benthic) in a group of fish endemic tosouthern South America, the Percichthyidae The most important determinant ofmorphological grouping was gill raker length, and although the differences inmorphology were subtle, they were associated with habitat and diet differences.Many of the features of this polymorphism parallel patterns shown by some speciesof sticklebacks and north temperate salmonids, suggesting that the emergence oftrophic polymorphism in fish may be a very general phenomenon. The Percichthysspecies complex offers an opportunity for testing hypotheses concerning the originof morphological polymorphisms, habitat race formation and speciation in a singlebiological system.
ACKNOWLEDGEMENTS
We thank Miguel Battini, Amalia Denegri, and Pablo Vigliano for logisticalsupport while conducting much of this work. We also thank Oscar Jeijo, Claudio‘Bulon’ Knaup, and Valeria Ojeda for much appreciated assistance in the field,David Hamilton for statistical advice, and Marcos Zentilli and Jose Cembrano forassisting with information on the geology of the region. We thank Tom Miyake for

TROPHIC POLYMORPHISM IN ANDEAN FISH 211
help with some of the figures and E. B. Taylor for a stimulating discussion. Thiswork would not have been possible without the cooperation of the ‘DireccionTecnica de Parques Nacionales’ and we especially thank Claudio Chehebar andPablo Martinez for permission to collect native fish within the Parque NacionalNahuel Huapi and Parque Nacional Lanın. SJW was supported by an Organizationof American States fellowship while conducting the fieldwork. Funds for the researchwere provided by an NSERC (Canada) grant to SJW, and by Universidad Nacionaldel Comahue and ANPCYT (Argentina) grants to VEC. We especially thank BerenRobinson and a team of two anonymous referees for comments that greatly enhancedthe quality of this paper. Violeta Ruzzante and Pascal and Sacha Walde Ruzzantecontributed immeasurably (not standardized by size) to our enjoyment of the fieldwork.
REFERENCES
Almiron A, Azpelicueta M, Casciotta J, Lopez Carzola A. 1997. Ichthyogeographic boundarybetween the Brazilean and Austral Subregions in South America, Argentina. Biogeographica 73:23–30.
Anon Suarez DA. 1997. Estructura y dinamica de la taxocenosis Chironomidae (Diptera, Nematocera)de un lago andino. Unpublished Doctoral Thesis. Universidad Nacional de La Plata, Argentina.
Arratia G. 1982. A review of freshwater percoids from South America (Pisces, Osteichthys, Perciformes,Percichthyidae and Perciliidae). Abhandlungen der Senckenbergischen Naturforschenden Ges. 540: 1–52.
Arratia G, Penafort MB, Menu Marque S. 1983. Peces de la region sureste de los Andes y susprobables relaciones biogeograficas actuales. Deserta 7: 48–107.
Bentzen P, McPhail JD. 1984. Ecology and evolution of sympatric sticklebacks (Gasterosteus):specialization for alternative trophic niches in the Enos Lake species pair. Canadian Journal of Zoology52: 2280–2286.
Bernatchez L, Dodson JJ. 1991. Phylogeographic structure in mitochondrial DNA of the lakewhitefish (Coregonus clupeaformis) and its relation to Pleistocene glaciations. Evolution 45: 1016–1035.
Bernatchez L, Dodson JJ. 1994. Phylogenetic relationships among Palearctic and Neartic whitefish(Coregonus sp.) populations as revealed by mitochondrial DNA variation. Canadian Journal of Fisheriesand Aquatic Sciences 51: 240–251.
Bernatchez L, Vuorinen JA, Bodaly RA, Dodson JJ. 1996. Genetic evidence for reproductiveisolation and multiple origins of sympatric trophic ecotypes of whitefish (Coregonus). Evolution 50:624–635.
Boschi EE, Fuster de Plaza ML. 1957. Algunos resultados sobre el mantenimiento de ‘percas’ otruchas criollas en cautividad. Ventajas de su introduccion en nuevos ambientes. Ciencia e Investigacion13: 446–450.
Bruzone JH. 1986. Relevamiento de la fauna icticola de los Parques Nacionales Lanin, NahuelHuapi, Puelo y Los Alerces. Administracion de Parques Nacionales, 20 pp.
Bush GL. 1994. Sympatric speciation in animals: new wine in old bottles. Trends in Ecology and Evolution9: 285–288.
Campos H, Gavilan JF. 1996. Diferenciacion morfologica entre Percichthys trucha y Percichthys melanops(Perciformes: Percichthyidae) entre 36° y 41° L. S. (Chile y Argentina), a traves de analisismultivariados. Gayana Zool 60(2): 99–120.
Cresko WA, Baker JA. 1996. Two morphotypes of lacustrine threespine stickleback, Gasterosteusaculeatus, in Benka Lake, Alaska. Environmental Biology of Fishes 45: 343–350.
Crowl TA, Townsend CR, McIntosh AR. 1992. The impact of introduced brown and rainbowtrout on native fish: the case of Australasia. Reviews in Fish Biology and Fisheries 2: 217–241.
Cussac VE, Ruzzante DE, Walde SJ, Macchi PJ, Ojeda V, Alonso MF, Denegri MA. 1998.Body-shape variation of three species of Percichthys in relation to their coexistence in the Limayriver basin, in Northern Patagonia. Environmental Biology of Fishes 53(2): 143–153.
Day T, Pritchard J, Schluter D. 1994. Ecology and genetics of phenotypic plasticity: a comparisonof two sticklebacks. Evolution 48: 1723–1734.

D. E. RUZZANTE ET AL212
Doebeli M. 1996a. A quantitative genetic competition model for sympatric speciation. Journal ofEvolutionary Biology 9: 893–909.
Doebeli M. 1996b. An explicit genetic model for ecological character displacement. Ecology 72:510–520.
Ehlinger TJ. 1990. Habitat choice and phenotype-limited feeding efficiency in bluegill: individualdifferences and trophic polymorphism. Ecology 71: 886–896.
Ehlinger TJ, Wilson DS. 1988. Complex foraging polymorphism in bluegill sunfish. Proceedings of theNational Academy of Sciences, USA 85: 1878–1882.
Ferriz RA. 1989. Alimentacion de Percichthys colhuapiensis (Mac Donagh, 1955) y P. trucha (Girard,1854) (Osteichthyes, Percichthyidae), en el embalse Ramos Mexia, provincia del Neuquen, Argentina.IHERINGIA, Ser. Zool., Porto Alegre (69): 109–116.
Ferriz RA. 1993. Algunos aspectos de la dieta de cuatro especies ıcticas del Rio Limay (Argentina).Revista de Ictiologia 2/3 (1/2): 1–7.
Fryer G, Iles TD. 1972. The cichlid fishes of the Great Lakes of Africa. Their Biology and Evolution. Edinburgh:Oliver and Boyd.
Fuster de Plaza ML, Plaza JC. 1955. Nuevos ensayos para obtener la reproduccion artificial delas percas o truchas criollas (Percichthys sp.). Publ. Misc. 407. Ministerio de Agricultura y Ganaderıa,47 pp.
Futuyma DJ. 1986. Evolutionary Biology, 2nd ed. Sunderland, Mass: Sinauer.Glova GJ, Sagar PM, Naslund I. 1992. Interaction for food and space between populations of
Galaxias vulgaris Stokell and juvenile Salmo trutta L. in a New Zealand stream. Journal of Fish Biology41: 909–925.
Gonzalez Regalado T. 1945. Los peces de los Parques Nacionales Nahuel Huapi, Lanin, y LosAlerces. Anales del Museo de la Patagonia I: 121–133. Ministerio de Obras Publicas.
Guerrero CA. 1991. Crecimiento de la perca de boca chica (Percichthys trucha) en cuatro ambienteslenticos patagonicos (Argentina). pp. 47–71 En: Trabajos presentados al Segundo Taller In-ternacional sobre Ecologia y Manejo de Peces en Lagos y Embalses, Santiago, Chile. Editado porIrma Vila. FAOCOPESCAL.
Grant PR. 1986. Ecology and Evolution of Darwin’s Finches. Princeton, New Jersey: Princeton UniversityPress.
Grant BR, Grant PR. 1989. Evolutionary dynamics of a natural populations: the large cactus finch of theGalapagos. Chicago: The University of Chicago Press.
Hatfield T. 1997. Genetic divergence in adaptive characters between sympatric species of stickleback.The American Naturalist 149: 1009–1029.
Hindar K, Jonsson B. 1993. Ecological polymorphism in Arctic charr. Biological Journal of the LinneanSociety 48: 63–74.
Johnson GD. 1984. Percoidei: development and relationships. In: Moser HG, Richards WJ, CohenDM, Fahay MP, Kendall, Jr., AW, Richardson SL, eds. Ontogeny and Systematics of Fishes. AmericanSociety Ichthyology and Herpetology, Special Publication, 1.
Johnson GD. 1993. Percomorph phylogeny: progress and problems. Bulletin of Marine Science 52(1):3–28.
Lack D. 1947. Darwin’s finches. Cambridge: Cambridge University Press.Lara L. 1997. Geologia y geoquimica del volcan Lanın (39.5°S–71.5°W), Andes del Sur. IX region,
Chile. Tesis de Magister, Depto. de Geologıa, Universidad de Chile, Santiago. 172 p.Lauder GV. 1983. Prey capture hydrodynamics in fishes: experimental tests of two models. Journal of
Experimental Biology 104: 1–13.Lauder GV. 1985a. Aquatic feeding in lower vertebrates. In: Hildebrand M, Bramble D, Liem KF,
Wake DB, eds. Functional Vertebrate Morphology. Cambridge, MA: Harvard University Press, 210–229.Lauder GV. 1985b. Functional morphology of the feeding mechanisms in lower vertebrates. In:
Duncker H.-R, Fleisher G, eds. Functional morphology in vertebrates. Proceedings of the Ist InternationalSymposium on vertebrate morphology. Stuttgart, 177–188.
Lauder GV, Schaffer HB. 1985. Functional morphology of the feeding mechanism in aquaticAmbystomatid salamanders. Journal of Morphology 185: 297–326.
Lauder GV, Schaffer HB. 1993. Design of feeding systems in aquatic vertebrates: major patternsand their evolutionary interpretations. In: Hanken J, Hall BK, eds. The Skull. Functional and EvolutionaryMechanisms, Vol. 3. Chicago: University of Chicago Press, 113–149.
Lavin PA, McPhail JD. 1986. Adaptative divergence of trophic phenotype among freshwaterpopulations of the Threespine Stickleback (Gasterosteus aculeatus). Canadian Journal of Fisheries andAquatic Sciences 43: 2455–2463.

TROPHIC POLYMORPHISM IN ANDEAN FISH 213
Lindsey CC. 1981. Stocks are chamaleons: plasticity in gill rakers of coregonid fishes. Canadian Journalof Fisheries and Aquatic Sciences 38: 1497–1506.
Lopez Albarello A. 1993. Analisis morfometrico de las especies argentinas de la familia Percichthyidae(Osteichthyies, Perciformes). Seminario de Licenciatura, Universidad Nacional de Buenos Aires,Facultad de Ciencias Exactas y Naturales, 114 pp.
Lopez HL, Menni RC, Miquelarena AM. 1987. Lista de los peces de agua dulce de la Argentina.Biologia Acuatica 12: 1–50.
Losos JB, Warheit KI, Schoener TW. 1997. Adaptive differentiation following experimental islandcolonization in Anolis lizards. Nature 387: 70–72.
Macchi PJ. 1991. Ecologıa trofica de los peces autoctonos del Lago Moreno (Brazo Morenito). Tesisde Licenciatura, Centro Regional Universitario, Universidad Nacional del Comahue. 106 pp.
Macchi PJ. 1993. Patrones de actividad trofica y eficiencia de la alimentacion de los peces autoctonosdel Lago Moreno. Informe Final. Centro Regional Universitario Bariloche – Universidad Nacionaldel Comahue. 80 pp.
Magnuson JJ, Heitz JG. 1971. Gill raker apparatus and food selectivity among mackerels, tunasand dolphins. Fishery Bulletin 69: 361–370.
Malmquist HJ, Snorrason SS, Skulason S, Jonsson B, Sandlund OT, Jonasson PM. 1992.Diet differentiation in polymorphic Arctic charr in Thingvallavatn, Iceland. Journal of Animal Ecology61: 21–35.
Mastrarrigo V. 1948. Reproduccion artificial de la perca o trucha criolla (Percichthys sp). Separatadel Almanaque Ministerio de Agricultura y Ganaderia. 7 pp.
Mayr E. 1963. Animal species and evolution. Cambridge, Massachusetts: Harvard University Press.McDowall RM. 1990. When galaxiid and salmonid fishes meet – a family reunion in New Zealand.
Journal of Fish Biology 37 (Supplement A): 35–43.McIntosh AR, Crowl TA, Townsend CR. 1994. Size related impacts of introduced brown trout
on the distribution of native common river galaxias. New Zealand Journal of Marine and FreshwaterResearch 28: 135–144.
McPhail JD. 1984. Ecology and evolution of sympatric sticklebacks (Gasterosteus): morphological andgenetic evidence for a species pair in Enos Lake, British Columbia. Canadian Journal of Zoology 62:1402–1408.
McPhail JD. 1993. Ecology and evolution of sympatric sticklebacks (Gasterosteus): origin of the speciespairs. Canadian Journal of Zoology 71: 515–523.
Mercer JH. 1976. Glacial history of southernmost South America. Quaternary Research 6: 125–166.Meyer A. 1987. Phenotypic plasticity and heterochrony in Cichlasoma managuense and their implications
for speciation in cichlid fishes. Evolution 41: 1357–1369.Meyer A. 1989. Cost of morphological specialization: feeding performance of the two morphs in the
trophically polymorphic cichlid fish, Cichlasoma citrinellum. Oecologia 80: 431–436.Meyer A. 1993. Phylogenetic relationships and evolutionary processes in East African cichlid fishes.
Trends in Ecology and Evolution 8: 279–284.Moyle PB, Cech JJ. 1982. Fishes. An introduction to ichthyology. Englewood Cliffs, NJ: Prentice-Hall.Ortubay SG, Semenas LG, Ubeda CA, Quaggiotto AE, Viozzi GP. 1994. Catalogo de peces
dulceacuicolas de la Patagonia Argentina y sus parsitos metazoos. Direccion de Pesca, Subsecretaria deRecursos Naturales, Provincia de Rio Negro, Argentina. S.C. Bariloche.
Pinkas L, Oliphant MS, Iverson ZL. 1971. Food habits of albacore bluefin tuna and bonito inCalifornia water. Dept. of Fish and Game Fisheries Bulletin 152: 1–105.
Ringuelet RA, Aramburu RH, Alonso de Aramburu A. 1967. Los Peces Argentinos de Agua Dulce.Provincia de Buenos Aires, Gobernacion: Comision de Investigacion Cientifica.
Roberts, TR. 1974. Dental polymorphism and systematics in Saccodon, a neotropical genus offreshwater fishes (Parodontidae, Characoidei). Journal Zoology, Lond. 173: 303–321.
Robinson BW, Wilson DS. 1994. Character release and displacement in fishes: a neglected literature.The American Naturalist 144: 596–627.
Robinson BW, Wilson DS. 1996. Genetic variation and phenotypic plasticity in a trophicallypolymorphic population of pumpkinseed sunfish (Lepomis gibbosus). Evolution Ecology 10: 631–652.
Robinson BW, Wilson DS, Margosian AS, Lotito PT. 1993. Ecological and morphologicaldifferentiation of pumpkinseed sunfish in lakes without bluegill sunfish. Evolution Ecology 7: 451–464.
Robinson BW, Wilson DS, Shea GO. 1996. Trade-offs of ecological specialization: an intraspecificcomparison of pumpkinseed sunfish phenotypes. Ecology 77: 170–178.
Schluter D. 1993. Adaptive radiation in sticklebacks: size, shape and habitat use efficiency. Ecology74: 699–709.

D. E. RUZZANTE ET AL214
Schluter D. 1994. Experimental evidence that competition promotes divergence in adaptive radiation.Science 266: 798–801.
Schluter D. 1995. Adaptive radiation in sticklebacks: trade-offs in feeding performance and growth.Ecology 76: 82–90.
Schluter D. 1996. Ecological causes of adaptive radiation. The American Naturalist 148: S40–S64.Schluter D, McPhail JD. 1992. Ecological character displacement and speciation in sticklebacks.
The American Naturalist 140: 85–108.Schluter D, McPhail JD. 1993. Character displacement and replicate adaptive radiation. Trends in
Ecology and Evolution 8: 197–200.Skulason S, Smith TB. 1995. Resource polymorphisms in vertebrates. Trends in Ecology and Evolution
10: 366–370.Skulason S, Noakes DLG, Snorrason SS. 1989. Ontogeny of trophic morphology in four sympatric
morphs of Arctic charr, Salvelinus alpinus, in Thingvallavatn, Iceland. Biological Journal of the LinneanSociety 38: 218–301.
Skulason S, Snorrason SS, Ota D, Noakes DLG. 1993. Genetically based differences in foragingbehaviour among sympatric morphs of arctic charr (Pisces: Salmonidae). Animal Behavior 45:1179–1192.
Skulason S, Snorrason SS, Noakes DLG, Ferguson MM. 1996. Genetic basis of life historyvariations among sympatric morphs of Arctic charr, Salvelinus alpinus. Canadian Journal of Fisheries andAquatic Sciences 53: 1807–1813.
Smith TB, Skulason S. 1996. Evolutionary significance of resource polymorphism in fishes,amphibians, and birds. Annual Review of Ecology and Systematics 27: 111–133.
Snorrason SS, Skulason S, Jonsson B, Malmquist HL, Jonasson PM, Sandlund OT, LindemT. 1994. Trophic specialization in Arctic charr Salvelinus alpinus (Pisces, Salmonidae): morphologicaldivergence and ontogenetic niche shifts. Biological Journal of the Linnean Society 52: 1–18.
Svardson G. 1961. Young sibling fish species in Northwestern Europe. In: Blair WF, ed. VertebrateSpeciation), Austin, Texas: University of Texas, 498–513.
Taylor EB, Bentzen P. 1993. Evidence of multiple origins and sympatric divergence of trophicecotypes of smelt (Osmerus) in northeastern North America. Evolution 47: 813–832.
Taylor EB, McPhail JD, Schluter D. 1997. History of ecological selection in sticklebacks: unitingexperimental and phylogenetic approaches. In: Givnish TJ, Sytsma KJ, eds. Molecular evolution andadaptive radiation. Cambridge, UK: Cambridge University Press, 511–534.
Tilzey RDJ. 1976. Observations on interactions between indigenous Galaxidae and introducedSalmonidae in the Lake Eucumbene catchment, New South Wales. Australian Journal of Marine andFreshwater Rev 27: 551–564.
Van Valen L. 1965. Morphological variation and width of ecological niche. The American Naturalist99: 377–390.
Via S, Lande R. 1985. Genotype-environment interaction and the evolution of phenotypic plasticity.Evolution 39: 505–522.
Via S, Gomulkiewicz R, De Jong G, Scheiner SM, Schlichting CD, Van Tienderen PH.1995. Adaptive phenotypic plasticity: consensus and controversy. Trends in Ecology and Evolution 10:212–217.
Villagran C. 1991. El cuaternario en Chile: evidencias de cambio climatico. Simposio geoecologiade montana de los Andes del Sur. Actas, 28 pp.
Volpe JP, Ferguson MM. 1996. Molecular genetic examination of the polymorphic Arctic charrSalvelinus alpinus of Thingvallavatn, Iceland. Molecular Ecology 5: 763–772.
Vuorinen JA, Bodaly RA, Reist JD, Bernatchez L, Dodson JJ. 1993. Genetic and morphologicaldifferentiation between dwarf and normal size forms of lake whitefish (Coregonus clupeaformis) inComo Lake, Ontario. Canadian Journal of Fisheries and Aquatic Sciences 50: 210–216.
Whitmore GA. 1986. Prediction limits for a univariate normal observation. American Statistician 40:141–143.
Wimberger PH. 1991. Plasticity of jaw and skull morphology in the neotropical cichlids Geophagusbrasiliensis and G. steindachneri. Evolution 45: 1545–1563.
Wimberger PH. 1992. Plasticity of fish body shape. The effects of diet, development, family andage in two species of Geophagus (Pisces: Cichlidae). Biological Journal of the Linnean Society 45: 197–218.




















