
Chromosome evolution in the Salmonidae (Pisces): an update
25
Biol. Rev. (2001), 76, pp.1–25 Printed in the United Kingdom # Cambridge Philosophical Society 1 Chromosome evolution in the Salmonidae (Pisces) : an update RUTH PHILLIPS"* and PETR RA ! B# " Department of Biological Sciences, University of Wisconsin–Milwaukee, Milwaukee, WI 53201, USA (e-mail : rp!csd.uwm.edu) # Laboratory of Fish Genetics, Institute of Animal Physiology and Genetics 277 21 Libechov, Czech Republic (Received 15 February 1999 ; revised 29 June 2000 ; accepted 24 July 2000) ABSTRACT The karyotypes of salmonid fishes including taxa in the three subfamilies Coregoninae, Thymallinae and Salmoninae are described. This review is an update of the (Hartley, 1987) review of the chromosomes of salmonid fishes. As described in the previous review, the karyotypes of salmonid fishes fall into two main categories based on chromosome numbers : the type A karyotypes have diploid numbers close to 80 with approximately 100 chromosome arms (2n fl 80, NF fl 100), and the type B karyotypes have diploid numbers close to 60 with approximately 100 chromosome arms (2n fl 60, NF fl 100). In this paper we have proposed additional sub categories based on variation in the number of chromosome arms : the A« type with NF fl 110–120, the A§ type with NF greater than 140, and the B« type with NF less than 80. Two modes of chromosome evolution are found in the salmonids : in the Coregoninae and the Salmoninae the chromosomes have evolved by centric fusions of the Robertsonian type decreasing chromosome numbers (2n) while retaining chromosome arm numbers (NF) close to that found in the hypothetical tetraploid ancestor so that most extant taxa have either type A or type B karyotypes. In the Thymallinae, the chromosomes have evolved by inversions so that chromosome arm numbers (NF) have increased but chromosome numbers (2n) close to the karyotype of the hypothetical tetraploid ancestor have been retained and all taxa have type A§ karyotypes. Most of the taxa with type B karyotypes in the Coregoninae and Salmoninae are members of the genus Oncorhynchus, although at least one example of type B karyotypes is found in all of the other genera. These taxa either have an anadromous life history or are found in specialized lacustrine environments. Selection for increases or decreases in genetic recombination as proposed by Qumsiyeh, 1994 could have been involved in the evolution of chromosome number in salmonid fishes. Key words : cytogenetics, chromosomes, fish, salmonid, Salmonidae, Pisces, trout, salmon, char, whitefish, grayling. CONTENTS I. Introduction ............................................................................................................................ 2 (1) Salmonid fishes.................................................................................................................. 2 (2) Salmonid karyotypes ......................................................................................................... 2 (3) Qumsiyeh’s hypothesis ...................................................................................................... 3 II. Classification of salmonid chromosomes .................................................................................. 4 (1) Problems in classification .................................................................................................. 4 (2) Categories of karyotypes ................................................................................................... 5 III. Review of salmonid karyotypes ............................................................................................... 5 (1) Coregoninae ...................................................................................................................... 5 (a) Prosopium ..................................................................................................................... 5 (b) Coregonus and Stenodus .................................................................................................. 5 * Author for correspondence.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Chromosome evolution in the Salmonidae (Pisces): an update

Biol. Rev. (2001), 76, pp. 1–25 Printed in the United Kingdom # Cambridge Philosophical Society 1
Chromosome evolution in the Salmonidae
(Pisces) : an update
RUTH PHILLIPS"* and PETR RA! B#
"Department of Biological Sciences, University of Wisconsin–Milwaukee, Milwaukee, WI 53201, USA (e-mail :
rp!csd.uwm.edu)#Laboratory of Fish Genetics, Institute of Animal Physiology and Genetics 277 21 Libechov, Czech Republic
(Received 15 February 1999; revised 29 June 2000; accepted 24 July 2000)
ABSTRACT
The karyotypes of salmonid fishes including taxa in the three subfamilies Coregoninae, Thymallinae andSalmoninae are described. This review is an update of the (Hartley, 1987) review of the chromosomes ofsalmonid fishes. As described in the previous review, the karyotypes of salmonid fishes fall into two maincategories based on chromosome numbers : the type A karyotypes have diploid numbers close to 80 withapproximately 100 chromosome arms (2n¯ 80, NF¯ 100), and the type B karyotypes have diploidnumbers close to 60 with approximately 100 chromosome arms (2n¯ 60, NF¯ 100). In this paper we haveproposed additional sub categories based on variation in the number of chromosome arms: the A« type withNF¯ 110–120, the A§ type with NF greater than 140, and the B« type with NF less than 80. Two modes ofchromosome evolution are found in the salmonids : in the Coregoninae and the Salmoninae thechromosomes have evolved by centric fusions of the Robertsonian type decreasing chromosome numbers (2n)while retaining chromosome arm numbers (NF) close to that found in the hypothetical tetraploid ancestorso that most extant taxa have either type A or type B karyotypes. In the Thymallinae, the chromosomes haveevolved by inversions so that chromosome arm numbers (NF) have increased but chromosome numbers (2n)close to the karyotype of the hypothetical tetraploid ancestor have been retained and all taxa have type A§karyotypes. Most of the taxa with type B karyotypes in the Coregoninae and Salmoninae are members ofthe genus Oncorhynchus, although at least one example of type B karyotypes is found in all of the other genera.These taxa either have an anadromous life history or are found in specialized lacustrine environments.Selection for increases or decreases in genetic recombination as proposed by Qumsiyeh, 1994 could havebeen involved in the evolution of chromosome number in salmonid fishes.
Key words : cytogenetics, chromosomes, fish, salmonid, Salmonidae, Pisces, trout, salmon, char, whitefish,grayling.
CONTENTS
I. Introduction ............................................................................................................................ 2(1) Salmonid fishes.................................................................................................................. 2(2) Salmonid karyotypes ......................................................................................................... 2(3) Qumsiyeh’s hypothesis ...................................................................................................... 3
II. Classification of salmonid chromosomes .................................................................................. 4(1) Problems in classification .................................................................................................. 4(2) Categories of karyotypes ................................................................................................... 5
III. Review of salmonid karyotypes ............................................................................................... 5(1) Coregoninae ...................................................................................................................... 5
(a) Prosopium ..................................................................................................................... 5(b) Coregonus and Stenodus .................................................................................................. 5
* Author for correspondence.

2 Ruth Phillips and Petr Ra! b
(2) Thymallinae ...................................................................................................................... 10(3) Salmoninae........................................................................................................................ 10
(a) Brachymystax and Hucho................................................................................................ 11(b) Salvelinus and Salvethymus ............................................................................................. 11(c) Salmothymus, Acantholingua and Platysalmo..................................................................... 12(d) Salmo ........................................................................................................................... 12(e) Oncorhynchus ................................................................................................................. 12
IV. Types of rearrangements ......................................................................................................... 14(1) Major rearrangements : translocations .............................................................................. 14(2) Minor rearrangements : NORs.......................................................................................... 15(3) Minor rearrangements : heterochromatin additions .......................................................... 15
V. Sex chromosomes..................................................................................................................... 15VI. Chromosome banding.............................................................................................................. 15
VII. Karyotype evolution ................................................................................................................ 16(1) The ancestral salmonid karyotype .................................................................................... 16(2) Two modes of karyotype evolution ................................................................................... 17(3) Parallelism in karyotypes of Coregoninae and Salmoninae .............................................. 17(4) Similarities between taxa with type B karyotypes............................................................. 17(5) Salmonid chromosome evolution and Qumsiyeh’s hypothesis........................................... 17(6) Other aspects of karyotype evolution................................................................................ 19
VIII. Conclusions .............................................................................................................................. 20IX. References................................................................................................................................ 20
I. INTRODUCTION
(1) Salmonid fishes
The ancestrally tetraploid salmonid fishes, familySalmonidae, include the subfamilies Coregoninae(whitefishes, ciscos, inconnu), Thymallinae (gray-lings) and Salmoninae (trouts, salmons and charrs).They have been subjected to intensive study in partbecause many of them are of commercial import-ance. The Salmonidae are found throughout thenorthern hemisphere and are probably of freshwaterorigin. They are especially noted for their extensivephenotypic plasticity, often forming species flocks inrecently glaciated lakes (reviewed in Behnke, 1972;Savvaitova, 1994; Allendorf & Waples, 1996). Thisphenotypic plasticity may be partly a result of theirautotetraploid origin which produced duplicategenes which would be free to develop new functions(reviewed in Ohno, 1970).
There is considerable evidence supporting anautotetraploid origin for the Salmonidae (reviewedin Allendorf & Thorgaard, 1984). The genome sizeis 2.5¬10* base pairs, approximately twice that ofrelated fishes and the number of chromosome arms(NF) which varies from 74 to 170 is also approxi-mately twice that of related fishes (NF¯ 48–60).Meiotic multivalents have been observed in severalspecies as well as residual tetrasomic inheritance forsome loci. Many duplicate loci have been identifiedand duplicated genes that have been sequenced in avariety of species fall into two clades. For example,
two duplicate growth hormone genes, GH1 andGH2 have been studied in the Salmoninae. TheGH1 genes in the genera Brachymystax, Hucho,Salvelinus, Salmo, and Oncorhynchus form a clade, andthe GH2 genes for these same taxa form anotherclade. These results suggest that GH1 and GH2 lociwere produced by ancestral tetraploidizationwhich has been followed by divergence (reviewed inDevlin, 1993; Phillips & Oakley, 1997; Oakley &Phillips, 1999).
(2) Salmonid karyotypes
Salmonids are the best karyologically studied fishgroup in terms of the number of species, populations,individuals and material (adults and embryos)analysed. Since the last karyological review ofsalmonid fishes (Hartley, 1987), new taxa have beendescribed (Chereshnev & Skopets, 1990), karyotypesof some taxa have been described for the first time(Makoedov, 1987; Frolov, 1992a, 1993), and karyo-types of other taxa have been revised (e.g. Phillips,Reed & Ra! b, 1996) or reviewed (Cavender &Kimura, 1989; Ra! b & Jankun, 1992; Ra! b, S) lechta& Flajs)hans, 1994). New information on salmonidphylogenetic relationships based on molecular (re-viewed in Phillips & Oakley, 1997, Oakley & Phillips1999), and osteological data (Sanford, 1990; Smith& Todd 1992; Smith & Stearley, 1993; Glubokovsky& Frolov, 1994) have been obtained. The salmonidsare the only fish group for which all available

3Salmonid chromosome evolution
X Y
Fig. 1. A type A karyotype from a male lake trout, Salvelinus namaycush, with C banding. The second chromosome pairis the sex chromosome pair.
chromosome banding techniques have been appliedincluding physical mapping of cloned genes usingin situ hybridization (Penda! s, Moran & Garcı!a-Vasquez, 1993, 1994; reviewed in Phillips & Reed,1996), molecular analysis of chromosomal rearrange-ments (Phillips & Reed, 1995b) and chromosomemicrodissection (Reed, Bohlander & Phillips, 1995).
A majority of salmonid karyotypes can be dividedinto two main categories (A and B) on the basis ofchromosome number, chromosome arm number andthe distribution of subtelocentric}acrocentric andmetacentric chromosomes (Hartley, 1987). The Akaryotypes (Fig. 1) contain approximately 80chromosomes, 100 chromosome arms and moresubtelocentric than metacentric chromosomes,while the B karyotypes (Fig. 2) contain approxi-mately 60 chromosomes, 104 chromosome arms andmore metacentric than subtelocentric chromosomes.
The recent discovery of the karyotype of Salvethy-mus svetovidovi, an endemic species of char in LakeElgygytgyn from Central Chukotka Peninsula, Rus-sia, with a low diploid chromosome number (2n¯56) (Frolov, 1993) has confirmed the hypothesis thateach of the salmonid lineages, i.e. coregonines,huchonins, chars, Pacific trouts and salmons, andEurasian trouts and salmons, has species with bothhigh (type A) and low (type B) chromosomenumbers. This finding raises a number of questions
about chromosomal evolution in salmonid fishes.Why do each of the salmonid lineages possess sisterspecies with high and low chromosome numbers?Was chromosome evolution after the tetraploidiz-ation event dominated by fusions or fissions or both?Was the chromosomal evolution unidirectional orare there examples of reversals in fusions and fissions?What accounts for the apparent parallel evolution inchromosome number in the salmonid lineages? Isthere a correlation between chromosome numberand life-history traits such as migratory behaviourand}or intralacustrine sympatric speciation?
(3) Qumsiyeh’s hypothesis
Because of the wide variation in diploid chromosomenumber (2n¯ 52–102) and varied life histories insalmonid fishes, they are an excellent vertebrategroup in which to examine the possible adaptivesignificance of variation in chromosome numbers.Recently, Qumsiyeh (1994) has suggested thatselection for increases or decreases in genetic re-combination may explain some aspects of mam-malian chromosome evolution. He has proposed twocompeting strategies for differentiation of mam-malian karyotypes. The first strategy is characterizedby chromosomal rearrangements leading to in-creased recombination, e.g. increase in 2n or NF,

4 Ruth Phillips and Petr Ra! b
1 2 3 4 5 6 7
8 9 10 11 12 13 14
15 16 17 18 19 20 21
23 24 25 26 27 28 29
22
30
Fig. 2. A type B karyotype from a female rainbow trout, Oncorhynchus mykiss. This is a reversed image of chromosomesstained with DAPI. Some of the chromosome pairs have DAPI -bright centromeres which show up as dark in thereversed image. The sex chromosome pair is number 24.
increased variation and adaptation to more pliablehabitats, and a decrease in the probability of fixationof new mutations (both chromosomal and genic).The second strategy is characterized by chromosomerearrangements leading to decreased recombination,e.g. decrease in 2n or NF, decreased variation andthus adaptation to a specialized or constant habitat,and an increase in the probability of fixation of anynew mutations (both chromosomal and genic).
It should be possible to test the proposed cor-relation between diploid chromosome number andamount of genetic recombination in salmonidswithin a few years because genome mapping projectshave been initiated in rainbow trout, Oncorhynchus
mykiss, (2n¯ 58–64) (Young et al., 1998), pinksalmon O. gorbuscha (2n¯ 52) (F. W. Allendorf,personal communication), Atlantic salmon, Salmo
salar (2n¯ 56–58) and brown trout, Salmo trutta,(2n¯ 80–82) (Hoyheim et al., 1998). Brown troutand Atlantic salmon are very closely related sistertaxa with a wide variation in chromosome number,so would be ideal for examining this hypothesis. Theinformation from the genome projects should alsoallow determination of homology between chromo-some arms and thus produce answers to questionsregarding the types and directions of fusions andfissions.
Here, we review data published after and}or not
covered by Hartley’s (1987) review, and also datascattered in inaccessible sources. We examine thekaryological data in a phylogenetic context anddiscuss the implications for chromosomal evolutionin the Salmonidae. We also examine the evidence forQumsiyeh’s (1994) proposed correlation betweendiploid chromosome number and habitat.
II. CLASSIFICATION OF SALMONID
CHROMOSOMES
Table 1 summarizes the current karyotype infor-mation on the Salmonidae. Some subspecies whichhave the same karyotypes are not listed (e.g. in theCoregonus laveretus complex and in the Salvelinus
malma}alpinus complex). For additional references,see Klinkhardt, Tesche & Greven (1995).
(1) Problems in classification
The majority of authors classify uni-armed and bi-armed chromosomes according to the guidelines ofLevan, Fredga & Sandberg (1964). Where differ-ences in the number of chromosome arms have beenreported for the same species, this is usually the resultof a difference in the scoring of the subtelocentricchromosomes by different authors. The NF value

5Salmonid chromosome evolution
was originally designed to quantify the centrictranslocations or fissions of the Robertsonian type(Matthey, 1945), but some authors scored subtelo-centrics as bi-armed. Robertsonian rearrangementsare very common in the Salmonidae, so that many ofthe species have a NF number of approximately 100.However, minor inversions around the centromerecan convert telocentric chromosomes into subtelo-centrics. To make trends in karyotype evolutionmore obvious, we have scored all of the chromosomesincluding metacentric and submetacentric chromo-somes as bi-armed and telocentric and subtelocentricas uni-armed. In other words, we consider meta-centric and submetacentric chromosomes to be aproduct of fusions while subtelocentric chromosomesare taken to be a product of another rearrangementalthough we are aware that the situation might bemore complex. Because the intraspecific variation inthe locations and sizes of the chromosomal nucleolarorganizer regions (NORs) have been frequentlydocumented, chromosomes in which the NOR takesup most of the short arm were also scored astelocentric}subtelocentric. The NF number given inTable 1 is the one we calculated using the criteriaexplained above.
(2) Categories of karyotypes
In the earlier review (Hartley, 1987), two majorcategories of salmonid karyotypes were described:type A with a diploid chromosome number close to80 and approximately 100 arms and type B with adiploid number close to 60 and approximately 104arms. It was pointed out that the majority of thespecies with B karyotypes are found in the mostderived genera of Oncorhynchus and Salmo. Recently,all of the western American and East Asian species ofsalmon with B karyotypes have been reclassified intothe genus Oncorhynchus (Smith & Stearley, 1989), asolution suggested much earlier by Regan (1914).Since Hartley (1987), two additional cases of Bkaryotypes have been discovered in other salmonidlineages, but the generalization that the vast ma-jority of such karyotypes are found in Oncorhynchus isstill valid. Another type of A karyotype (AA or A«)has been proposed for huchonine salmonids (Ra! b,S) lechta & Flajs)hans, 1994) which have NF numbersof 112 and above and two additional subtypes (A§and B«) based on differences in NF numbers will beproposed in this review. In Section III we willreview current knowledge about the distribution ofdifferent karyotypes in extant salmonid lineages.
REVIEW OF SALMONID KARYOTYPES
(1) Coregoninae
Coregonine salmonids include two lineages, roundwhitefishes, genus Prosopium and whitefishes, ciscoesand innconu, genera Coregonus and Stenodus. Karyo-types have been described for most of the coregoninespecies (Table 1) and karyological information forthis family has been reviewed recently (Ra! b &Jankun, 1992; Frolov, 1992aa).
(a) Prosopium
Karyotypes have been described for six species ofProsopium from blastula material by Booke (1968,1974, 1975) (Table 1). The widespread species,Prosopium cylindraceum has a typical type A karyotypewith 2n¯ 78 and NF¯ 100. Among the Bear Lakeendemic species, the two whitefish species, P.spilonotus (Bonneville whitefish) and P. abyssicola
(Bear Lake whitefish) have 2n¯ 74, NF¯ 100 and2n¯ 72, NF¯ 100, respectively, which representkaryotypes intermediate (I) between type A andtype B in diploid chromosome number. However,the morphologically diverged Bonneville cisco P.gemmiferum in the same lake has 2n¯ 64, NF¯ 100which is a typical B karyotype.
(b) Coregonus and Stenodus
Karyotypes have been described for the majority ofspecies of the genus Coregonus and the only species ofStenodus (Table 1). Stenodus is included with Coregonus
here because recent evidence from allozymes (Bodalyet al., 1991), mitochondrial DNA (Bernatchez,Colombani & Dodson, 1991; Reist et al., 1996), andribosomal DNA (Sajdak & Phillips, 1997) supportsinclusion of Stenodus in Coregonus. Since the lastreview, revised karyotypes from in vivo or in vitro
preparations have been obtained for a majority ofthese species including those for previously unknownNorth American coregonids (Phillips, Reed & Ra! b,1996). The chromosome number of the all of theNorth American whitefish and ciscos and themajority of the European whitefish and ciscos is2n¯ 80. Revised karyotypes show that the majorityof these species also have very similar NF numbers. S.leucichthys has a karyotype with 2n¯ 74–76 (Booke,1975; Frolov, 1991a ; Makoedov, 1992) with NF¯98–108, which would be an intermediate (I) karyo-type. There are a number of subspecies in the C.lavaretus}C. clupeaformis complex, but all of them have

6R
uth
Phillip
sand
Petr
Ra! b
Table 1. Karyotype data on salmonid fishes
Species Common name 2n NF NOR Type Reference
CoregoninaeCoregonus
albula
Vendace 80}81 96 M A Nygren et al. (1971a)Jankun et al. (1991, 1995)
artedii Lake herring 80}81 94 M A Shields et al. (1990);Booke, (1968)*Phillips et al. (1996)
autumnalis Arctic cisco 78–80 96–98 A Jachenko et al. (1979)chadary Amur cisco 80–84 98–100 A Ra! b, unpublished dataclupeaformis Lake
whitefish80 94 M A Booke (1968)*
Phillips et al. (1996)hoyi Bloater 80 94 S A Booke (1968)*;
Phillips et al. (1996)muksun Muksun 78 100 A Kaidanova (1978)*;
Andriyasheva et al. (1982)*lavaretus Whitefish 80}81 94 M A Nygren et al. (1971b) ;
Sola et al. (1989)nasus Broad
whitefish58–60 92–96 B Andriyasheva et al. (1982)*;
Viktorovsky & Ermolenko (1982)nigripinnis Black-fin cisco 80 94 M A Phillips et al. (1996)peled Peled 74 92, 98 I Nygren et al. (1971);
Viktorovsky (1964)reighardi Shortnose
cisco80 100 A Booke (1968)*
sardinella Least cisco 80}81 96}97 A Frolov (1986, 1990b)tungun Tungen 86–88 106 A Viktorovsky et al. (1983)ussuriensis Amur
whitefish80 100 A Viktorovsky & Maksimova (1978)
zenithicus Shortjawcisco
80 94 S A Booke (1968)*;Phillips et al. (1996)
Stenodus
leucichthys
Inconnu 74–76 108}98 I Booke (1975)*; Frolov (1991a) ;Makoedov, (1992)
Prosopium
abyssicola
Bear Lakewhitefish
72 100 I Booke (1975)*
coulteri Pygmywhitefish
82 100 A Booke (1974)*
cylindraceum Roundwhitefish
78 100 A Booke (1974)*

7Salm
onidchrom
osome
evolutiongemmiferum Bonneville
cisco64 100 B Booke (1975)*
spilonotus Bonnevillewhitefish
74 100 I Booke (1975)*
williamsoni Mountainwhitefish
78 100 A Booke (1975)*
ThymallinaeThymallus
thymallus
Europeangrayling
102 170 S A§ Nygren et al. (1971a) ;Severin (1979);Kalat et al. (1989)
arcticus pallasi Siberiangrayling
98 146 A§ Makoedov (1982b) ;Severin & Zinovyev (1982)
arcticus
arcticus
Arcticgrayling
98–102 160–168 A§ Severin & Zinovyev (1982)
grubei Amurgrayling
98–100 148 A§ Makoedov (1987)
SalmoninaeBrachymystax
lenok Lenok 90–92 110–116 A« Kang & Park (1973);Viktorovsky et al. (1985)
Hucho
hucho hucho Danube salmon 82 112–114 S A« Ra! b & Liehman (1982)hucho taimen Taimen 82–84 112–116 M A« Dorofeeva (1977);
Viktorovsky et al. (1985)Frolov & Frolova (2000)
Parahucho
perryi
Sakhalin taimen 62 100 B Anbinder et al. 1982;Vasilev (1983) Cavender &Kimura (1989)
Salmothymus
obtusirostris
Dalmatiantrout
82 94 A Berberovic et al. (1970)*
Salmo carpio Lake Gardatrout
80 98 A Merlo (1957)
ischan Sevan trout 80–82 96–100 A Ruhkian, (1982, 1984)letnica Lake Ohrid
trout80 104 A Dimovska (1959)
marmoratus Marble trout 80 102 A Al-Sabti (1985)salar Atlantic
salmon54–58 72–74 S B« Roberts (1970); Hartley &
Horne (1984a) ;Barsiene (1981)
trutta Brown trout 78–84 98–102 M A Hartley & Horne (1984b) ;Zenzes & Voiculescu (1975)

8R
uth
Phillip
sand
Petr
Ra! b
Table 1 (cont.)
Species Common name 2n NF NOR Type Reference
Salmoninae (cont.)Salvelinus
confluentus Bull trout 78 100 S A Cavender (1984); Cavender &Kimura (1989); Phillips et al. (1989)
fontinalis Brook trout 84 100 M A Lee & Wright (1981);Phillips et al. (1989)
namaycush Lake trout 84 100 M A Phillips & Zajicek (1982);Phillips et al. (1989)
leucomaenis Potted char 84 100 S A Ueda & Ojima (1983a, b) ;Cavender & Kimura (1989);Frolov & Miller (1992)
pluvius Japanese char 84–86 100 A Ueda & Ojima (1983b) ;Viktorovsky (1975b)
alpinus}malma
complex DollyVardenchar
82 98 S (sm) A Cavender (1984); Cavender &Kimura (1989); Phillips et al.(1989)
albus White char 78–80 98 M A Viktorovsky (1975a)alpinus Arctic char 78 98 M A Cavender (1984); Phillips et al.
(1988); Hartley (1989)elgyticus Small mouthed
char76–78 98 M A Frolov (1992a, 1995)
boganidae Boganid char 76–78 98 M A Frolov (1994a, 1995)kronicus Stone char 78–82 100 M A Viktorovsky (1975a)taranetzi Eastern Arctic
char76–78 98–100 M A Frolov (1997)
levanidovi
malma
lordi
Levanidovi char 78–80 98 M A Frolov & Frolova (1997, 1999)
m. malma DollyVardenchar
78 98 M A Viktorovsky (1975a, b) ;Frolov (1995); Phillips et al.(1999)
m. kraschennkovi DollyVardenchar
82 98 S(st) A Ueda & Ojima (1983a) ;Cavender & Kimura (1989);Frolov et al. (1997)
Salvethymus
svetovidovi Longfinned char 56 98 M B Frolov (1993, 1995)
9Salm
onidchrom
osome
evolution
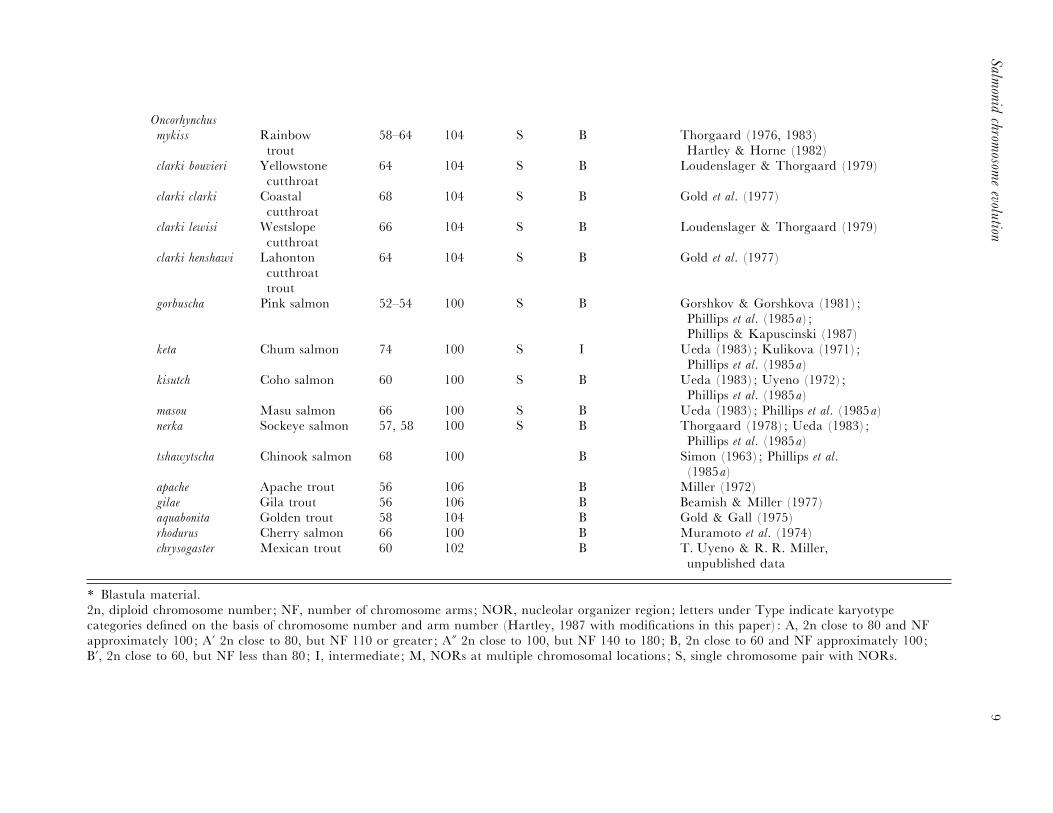
Oncorhynchus
mykiss Rainbowtrout
58–64 104 S B Thorgaard (1976, 1983)Hartley & Horne (1982)
clarki bouvieri Yellowstonecutthroat
64 104 S B Loudenslager & Thorgaard (1979)
clarki clarki Coastalcutthroat
68 104 S B Gold et al. (1977)
clarki lewisi Westslopecutthroat
66 104 S B Loudenslager & Thorgaard (1979)
clarki henshawi Lahontoncutthroattrout
64 104 S B Gold et al. (1977)
gorbuscha Pink salmon 52–54 100 S B Gorshkov & Gorshkova (1981);Phillips et al. (1985a) ;Phillips & Kapuscinski (1987)
keta Chum salmon 74 100 S I Ueda (1983); Kulikova (1971);Phillips et al. (1985a)
kisutch Coho salmon 60 100 S B Ueda (1983); Uyeno (1972);Phillips et al. (1985a)
masou Masu salmon 66 100 S B Ueda (1983); Phillips et al. (1985a)nerka Sockeye salmon 57, 58 100 S B Thorgaard (1978); Ueda (1983);
Phillips et al. (1985a)tshawytscha Chinook salmon 68 100 B Simon (1963); Phillips et al.
(1985a)apache Apache trout 56 106 B Miller (1972)gilae Gila trout 56 106 B Beamish & Miller (1977)aquabonita Golden trout 58 104 B Gold & Gall (1975)rhodurus Cherry salmon 66 100 B Muramoto et al. (1974)chrysogaster Mexican trout 60 102 B T. Uyeno & R. R. Miller,
unpublished data
* Blastula material.2n, diploid chromosome number; NF, number of chromosome arms; NOR, nucleolar organizer region; letters under Type indicate karyotypecategories defined on the basis of chromosome number and arm number (Hartley, 1987 with modifications in this paper) : A, 2n close to 80 and NFapproximately 100; A« 2n close to 80, but NF 110 or greater ; A§ 2n close to 100, but NF 140 to 180; B, 2n close to 60 and NF approximately 100;B«, 2n close to 60, but NF less than 80; I, intermediate ; M, NORs at multiple chromosomal locations ; S, single chromosome pair with NORs.

10 Ruth Phillips and Petr Ra! b
m
sm
st-a
1–
–33
20–
34–
–49
–19
Fig. 3. A type A« karyotype from Thymallus thymallus with Giemsa staining.
the type A karyotype with a modal 2n ¯ 80. In fact,all of the species in the genus Coregonus have the typeA or I karyotype with 2n numbers varying from74 to 88, except for C. nasus (broad whitefish) with2n¯ 58–60 and NF¯ 92–98 which has a type Bkaryotype.
(2) Thymallinae
Graylings include one monogeneric group, genusThymallus with five extant species. Graylings are apoorly studied salmonid group and karyotypes havebeen described for only four species (Table 1). Theirkaryotypes are unusual for salmonids with 2n¯ 102,NF¯ 170 for European grayling, T. thymallus
(Severin, 1979, 1985a, b) ; 2n¯ 98, NF¯ 170–178for Arctic grayling, T. arcticus arcticus and 2n¯ 98,NF¯ 146 for the Siberian grayling, T. arcticus pallasi
(Makoedov, 1982a, b ; Severin & Zinovyev, 1979,1982; Makoedov, Frolov & Shevshisin, 1983) and2n¯ 98–100, NF¯ 148 for Amur grayling, T. grubei(Makoedov, 1987). We designate this as the A§karyotype because of the high NF number inaddition to the high diploid number (Fig. 3).Examination of these karyotypes reveals that themetacentric chromosomes are much smaller thanthose found in most other salmonid karyotypes.None of them is larger than typical subtelocentric}acrocentric chromosomes found in other karyotypes.
This suggests that most of the metacentrics havebeen formed by pericentric inversions and othercentromeric shifts and that no centric fusions havebeen involved. The only chromosome banding dataavailable are the localization of NORs in thekaryotype of the European T. thymallus (Kalat et al.,1988). Since morphological (Norden, 1961; Smith &Stearley, 1993), and molecular evidence (S. McKay,personal communication, 1998; R. B. Phillips & T.H. Oakley, unpublished data) supports the place-ment of the Thymallinae as intermediate betweenCoregoninae and Salmoninae, these karyotypes areundoubtedly derived. The fact that the only oc-currence of such small metacentrics in the Salmoninae
is in the more basal genera Brachymystax and Hucho isalso consistent with this view.
(3) Salmoninae
The taxonomic organization of this subfamily iscontroversal (reviewed in Phillips & Oakley, 1997).There have been four major hypothesis proposed.Norden (1961) suggested that there are five mon-ophyletic genera branching off separately from themain stem: Brachymystax, Hucho, Salvelinus, Salmo andOncorhynchus. Kendall & Behnke (1984) proposedthat Brachymystax, Hucho & Salvelinus form a mono-phyletic sister group to Oncorhynchus and Salmo.Dorofeeva (1989) proposed a modification of this

11Salmonid chromosome evolution
m
sm
st
1–
16–
15
a
14
22–
–21
–41
Fig. 4. A type A« karyotype from the Danube salmon, Hucho hucho, with C banding. Note four small metacentricchromosome pairs, ‘huchonine’ markers.
hypothesis in which Parahucho is added as a sistergenus to Hucho, and Salmothymus is added as a sistergenus to Salmo. In the tree proposed by Smith andStearley (1993), Brachymystax is basal, followed bySalmothymus, and then two sister groups of Hucho andSalvelinus, and Salmo and Oncorhynchus. Moleculardata (reviewed in Phillips & Oakley, 1997) supportthe basal position of Brachymystax, and a closerelationship between Salmo and Salmothymus but thebranching relationships of the other genera have notyet been resolved.
(a) Brachymystax and Hucho
The cytotaxonomy of this group has been recentlyreviewed (Ra! b, S) lechta & Flajs)hans, 1994). Karyo-types of three species of Hucho (including Parahucho)and the two subspecies of Brachymystax lenok areknown, while karyotypes of H. bleekeri and H.ishikawai are unknown. B. lenok has a karyotype with2n¯ 90 or 92 and NF¯ 110–116, and the twosubspecies of Hucho hucho: H. hucho hucho and H. hucho
taimen have 2n¯ 82–84 and NF¯ 112–116. All ofthese species share four pairs of small metacentricchromosomes, ‘huchonine’ markers. Biochemicaldata support a close relationship between the twomorphotypes of Brachymystax lenok and Hucho
(Osinov, 1991). Their karyotypes would be classifiedas type A, but Ra! b, S) lechta & Flajs)hans (1994)proposed a new category ancestral (AA) (Fig. 4)
because of its apparent archaic ancestry. We use A«for this category in this paper (Table 1). In contrastto the other huchonines, the karyotype of theSakhalin taimen, Parahucho perryi which has beenplaced into a separate subgenus or genus (Holcı!ket al., 1988) has 2n¯ 62, NF¯ 100 which is a Bkaryotype possessing two pairs of ‘huchonine’marker chromosomes. Recent molecular data sup-port the generic status of Parahucho (Phillips, Oakley& Davis, 1995; Oakley & Phillips, 1999).
(b) Salvelinus and Salvethymus
Karyotypes have been described for 10 species ofSalvelinus (Table 1) and its cytotaxonomy has beenextensively reviewed (Viktorovsky, 1976; Cavender,1984; Cavender & Kimura, 1989; Phillips, Pleyte &Ihssen, 1989; Frolov, 1995). All of these species havekaryotypes with 2n¯ 76–86 and NF numbersaround 100 which are the typical A karyotype.Although a large number of species and subspecieshave been formally named in the S. alpinus}S. malma
complex including several species endemic to dif-ferent lakes in eastern Siberia, they all havechromosome numbers ranging from 76 to 82 (S.elgyticus, S. boganidae, S. kronicus, S. albus, S. levanidovi,S. taranetzi) (Frolov, 1991b, 1992b, 1994a, 1995,1997; Frolov & Frolova, 1997, 1998, 1999; Vikto-rovsky, 1975a, b, 1978a–c ; Viktorovsky & Glubo-kovsky, 1977). Recently a new genus was erected for

12 Ruth Phillips and Petr Ra! b
a morphologically diverged char endemic to LakeElgygytgyn in central Chukotka: Salvethymus sveto-vidovi (Chereshev & Skopets, 1990). Subsequentallozyme, craniological and ontogenic studies havesuggested that this fish is probably a morphologicallydiverged form of Salvelinus alpinus (reviewed inGlubokovsky & Frolov, 1994). This species has akaryotype with 2n¯ 56, NF¯ 98, which is a typicaltype B karyotype (Frolov, 1993). The presence oflarge blocks of telomeric heterochromatin is similarto that found in other forms of Salvelinus alpinus
(Frolov, 1994a). The karyotypes of S. elgyticus and S.boganidae from the same lake are very similar : themajority of cells have 2n¯ 77 (Frolov, 1992b, 1994a,b). Both karyotypes of these species are polymorphicfor a fusion translocation that has caused a reductionin chromosome number.
(c) Salmothymus, Acantholingua and Platysalmo
The ‘archaic trouts ’ of the genera Salmothymus,Acantholingua and Platysalmo are the least knownsalmonines. Hadzisce (1960, 1961) distinguished twomonotypic genera Salmothymus and Acantholingua forDalmatian trout S. obtusirostris and endemic speciesof Lake Ohrid S. (Acanthlolingua) ohridana, respec-tively. Behnke (1968) considered Salmothymus, Acan-tholingua, and his described Platysalmo as subgenera ofthe genus Salmo. Svetovidov (1975) also placedAcantholingua in Salmothymus. Recently Stearley andSmith (1993) suggested that all of these species beclassified in the genus Salmothymus. They alsosuggested that Salmo ischan, an endemic species ofLake Sevan basin in Armenia, and Salmo carpio ofLake Garda, Italy might be included in Salmothymus.A close relationship between S. ischan, S. carpio, S.letnica, Salmothymus (Acantholingua) ohridana and S.trutta was indicated by molecular data (Giuffra,Bernatchez & Guyomard, 1994; Bernatchez &Osinov, 1995; Apostolidis et al., 1997; Phillips et al.,2000). Unfortunately, the karyotypes of these ‘ar-chaic trouts ’ are the least known among salmonids.Salmothymus obstusirostris oxyrhynchus has 2n¯ 82, NF¯ 94 (Berberovic et al., 1970) which is a type Akaryotype similar to Salmo trutta, but the analysis wasbased on anaphase chromosomes. The karyotype ofSalmo ischan is well known. The four ecological racesof this endemic species possess distinct chromosomalsets with 2n¯ 80–82 and NF¯ 96–100 (Dorofeeva& Ruhkian, 1982; Ruhkian, 1984, 1989) which isclearly an A karyotype similar to S. trutta. Karyo-types of Salmothymus species and}or related forms: S.o. salnoita, S. o. krkensis and S. o. letnica inhabiting the
southwestern Balkan Peninsula, S. (Acantholingua)ohridana, and S. (Platysalmo) platycephalus are notknown.
(d) Salmo
Karyotypes have been described for all species of thegenus Salmo, although recent findings (Bernatchez,Guyomard & Bonhomme, 1992, Bernatchez &Osinov, 1995) suggest that the biological diversity ofEurasian Salmo is considerably greater than currenttaxonomy would suggest (Kotellat, 1997), a situationsimilar to that present in the genus Oncorhynchus
(Behnke, 1992). The brown trout, S. trutta is one ofthe cytogenetically best analysed salmonid fishes (fornumerous reports see Klinkhardt, Tesche & Greven,1995). The karyotype and its variations have beendescribed for the broad range of species distributionfrom western Europe (e.g. Hartley & Horne, 1984b),Martı!nez et al., 1991) to eastern Europe (Dorofeeva,1965; Dorofeeva & Ruhkian, 1982) and the upperAmu Darya River in Central Asia (Mazik &Toktosunov, l986). The marble trout, S. marmoratus,a species with restricted distribution in the northernand eastern Adriatic basin, is closely related to butdistinct from brown trout with 2n¯ 80 and NF¯102 (Al-Sabti, 1983, 1985). The least known is S.letnica, an endemic species of Lake Ohrid consideredby Karaman (1966) a ‘ semispecies ’ or form of browntrout. The only chromosome study (Dimovska, 1959)reports 2n¯ 80 and NF¯ 104. Six trout species ofthe genus Salmo have 2n¯ 78–84, with 2n¯ 80, NF96–104 being the most common. All of these aretypical A karyotypes. Atlantic salmon, S. salar, has aunique karyotype for salmonids with 2n¯ 54–56,but NF¯ 72–74 (for numerous reports see Klink-hardt, Tesche & Greven, 1995). This NF number ismuch lower than the widely distributed NF¯ 100.The presence of prominent interstitial C bands onthe larger telocentric chromosomes suggests thatthese chromosomes were formed by either tandemfusions or Robertsonian fusions followed by pericen-tric inversions (Ueda & Kobayashi, 1990). If each ofthese chromosomes is counted as biarmed, than theNF number would be approximately 100, suggestingthat the karyotype of Atlantic salmon is a subtype ofB which we here designate B« (Fig. 5).
(e) Oncorhynchus
The Pacific salmons and western trouts were recentlyclassified in the genus Oncorhynchus (Smith &Stearley, 1989, see also references in Behnke, 1992).

13Salmonid chromosome evolution
1 2 3 4 5
6 7 8 9
10 11 12 13 14 15
16 17 18 19 20 21
22 23 24 25 26 27
Fig. 5. The B« karyotype from a male Atlantic salmon, Salmo salar. This is a reversed image of chromosomes stainedwith DAPI. Many of the chromosomes have DAPI-bright bands. The largest acrocentric}subtelocentric chromosomepair appears to be composed of three tandemly fused telocentrics and many of the other large acrocentric}subtelo-centric chromosomes also appear to be the result of tandem fusions.
Karyotypes have been described for all of the majorspecies of Oncorhynchus (Table 1) (see also Hartley,1987 and Klinkhardt, Tesche & Greven, 1995). Thechromosome numbers for all of the species except
Table 2. Revised Oncorhynchus Chromosome data
Speciesa 2n No. M No. small STb No. Ab No. STA NF
O. masou 66 34 24 8 32 100O. tshawytscha 68 32 16 20 36 100O. kisutch 60 40 10 10 20 100O. keta 74 26 6 42 48 100O. nerka 58 42 4 12 16 100O. gorbuscha 52 48 4 0 4 100
aThe taxa are listed in order from the most basal to the most derived among the Pacific salmon (Phillips &Oakley, 1997). Note the decreasing numbers of subtelocentrics (ST) in the more recently evolved fish. This wouldbe expected if acrocentrics are converted to subtelocentrics by inversions over time.bSome intraspecific variation in the number of these two types of chromosomes (ST and A) is seen. This is probablybecause many of the short arms are composed of repetitive DNA which exhibits considerable intraspecific variationin copy number.
chum salmon, O. keta (2n¯ 74, NF¯ 102) varybetween 52 and 68 with NF numbers around 100which is typical of a type B karyotype. O. keta has anintermediate karyotype between type A and type B.

14 Ruth Phillips and Petr Ra! b
If the small subtelocentrics are considered uniarmed,then all of the Pacific salmon and western and EastAsian trouts including O. keta have NF¯ 100 (Table2), suggesting that all the major chromosomerearrangements in this group have consisted ofRobertsonian fusions or fissions.
IV. TYPES OF REARRANGEMENTS
(1) Major rearrangements: translocations
Both type A and type B karyotypes have NF valuesclose to 100, which is consistent with conversion ofone type into the other by Robertsonian trans-locations. The presence of a few species withintermediate karyotypes and the presence of intra-specific Robertsonian variations in many species arealso consistent with this being the major mode ofkaryotype evolution in the Salmonidae. The oc-currence of the lowest chromosome numbers pri-marily in the genus Oncorhynchus also suggests thatfusions that result in a reduction of chromosomenumbers have predominated over fissions. Intra-specific Robertsonian polymorphisms appear to beespecially common in species found over broadgeographical ranges such as Salmo salar, S. trutta,Oncorhynchus mykiss, O. clarki, Brachymystax lenok andCoregonus lavaretus. Typically, lacustrine salmonidassemblages display Robertsonian polymorphisms.Examples include Salmo ischan in Lake Seven,Caucasus (Ruhkian, 1982, 1984, 1989), numerousforms of Salvelinus alpinus complex in lake Azabachye,Kamchatka, (Vasilev, 1975), Lake Nashikinskoe,Kamchatka, (Savvaitova & Vasiliev, 1976; Savvai-tova, 1994), S. malma}S. kronicus complex in LakeKrontskoye, Kamchatka, (Viktorovsky, 1978a), Sal-velinus}Salvethymus complex in Lake Elgygytgyn,Chukotka, (Glubokovsky & Frolov, 1994), possiblyin the Salmo}Salmothymus complex in Lake Ohrid,Balkan Peninsula, (Dimovska, 1959; Berberovic et
al., 1970), Prosopium in Bear Lake, Utah, USA(Booke, 1974), Coregonus lavaretus in Lake Saimaa(Finland) (Jakun et al., 1995b) and Coregonus artedii
in Lake Superior, USA (Phillips, Reed & Ra! b1996). Closely related taxa often differ in one orseveral Robertsonian fusions and or}fissions, e.g.European Hucho hucho hucho with 2n¯ 82 andSiberian H. hucho taimen with 2n¯ 82–84 or thevarious Siberian taxa of chars (2n¯ 76–80). Thegeographical distribution of this interpopulationpolymorphism in widely distributed taxa reflects thedispersal history of particular species and can beirregular or regular. Brown trout, S. trutta has a
uniform karyotype throughout Europe as far as theupper Amu Darya River basin in Central Asia(Mazik & Toktosunov, 1986) except Caucasus andsome rivers in Greece where it is more polymorphic(Ruhkian, 1989; Karakousis, Paschos & Trianta-phyllidis, 1992). Rainbow and cutthroat trouts, O.mykiss and O. clarki have more polymorphic forms inthe southern parts of their ranges. On the otherhand, some species with broad ranges with the Akaryotype such as brook trout, Salvelinus fontinalis,lake trout, Salvelinus namaycush, and bull trout,Salvelinus confluentus do not exhibit Robertsonianpolymorphism although it may occur in restrictedareas (Cavender, 1984; Cavender & Kimura, 1989).
The karyotypes of the Pacific salmon and westerntrouts of the genus Oncorhynchus provide evidence forRobertsonian fissions which increase the diploidchromosome number. In rainbow trout, O. mykiss,the 2n number varies from 58 to 64, but evidencesuggests that the redband trout, an inland form with2n¯ 58 is the most ancestral type (Thorgaard,1983). In the case of O. clarki, the 2n number variesfrom 64 to 68 in different subspecies (Gold, Avise &Gall, 1977, Loudenslager & Thorgaard, 1979). Inthe western trouts (O. apache, O. gilae, O. chrysogaster,O. clarki, O. mykiss), there is evidence that thekaryotype of 2n¯ 64 was achieved independentlythree times, by interior cutthroat trout, coastalrainbow trout in some California populations, andby trouts of the Rio Mayo and Rio Yaqui (Uyeno &Miller, unpublished data, cited in Behnke, 1992).These examples, and the case of O. keta, which has ahigher 2n number than any of the other species inthe genus, but is one of the two most derived species(reviewed in Phillips & Oakley, 1997), suggest thatRobertsonian fissions as well as fusions have oc-curred. Probably some fusions and fissions arereversible, unless inversions occur to convert thechromosomes into submetacentric}metacentric chro-mosomes.
In the future, identification of homologous armsbetween species using in situ hybridization with armspecific paint probes (reviewed in Phillips & Reed,1996) should allow tracing of chromosome re-arrangements and would identify reversible fusions}fissions. It is possible that fusions have involved asubset of chromosomes with the same subclass ofcentric heterochromatin. Isolation and molecularcharacterization of centric heterochromatin has beendone for several salmonids including Arctic char(Hartley & Davidson, 1994) lake trout (Reed &Phillips, 1995b), and rainbow trout (Reed, Dorsch-ner & Phillips, 1997).

15Salmonid chromosome evolution
(2) Minor rearrangements: NORs
Active nucleolar organizer regions, (NORs, sites ofthe rDNA cistrons) stain positively with silver nitrate(Howell & Black, 1980), and chromomycin A3(CMA3) stains NORs regardless of activity in mostfish species (Amemiya & Gold, 1986). The mostaccurate method of identification is in situ hybridiz-ation with a rDNA probe, but this has only beenused in a few species. When all three techniques havebeen applied to the same material (Reed & Phillips,1995a), all silver-positive and CMA3-positive siteshave been confirmed with in situ hybridization withthe rDNA probe. A few additional very small siteswere identified only with the rDNA probe.
Multiple NORs, as revealed by silver and CMA3staining, have been observed among various speciesof Salvelinus including brook trout, S. fontinalis, laketrout, S. namaycush, (Phillips & Ihssen, 1985b ;Phillips et al., 1989), and Arctic char, S. alpinus
(Phillips, Pleyte & Hartley, 1988; Frolov, 1995),Dolly Varden char, S. malma spp. (Frolov & Frolova,1999b), and Salmo trutta (Mayr, Ra! b & Kalat, 1986;Penda! s et al., 1993; Martı!nez et al., 1991, 1993;Castro et al., 1994) and several species of Coregonus
(Phillips, Reed & Ra! b, 1996; Ra! b et al., 1996,Jankun et al., 2000). Although multiple NORs havenot been confirmed in any species of Oncorhynchus,centromeres of several chromosome pairs stainedpositively with CMA3 in O. keta (Phillips, Zajicek &Utter, 1985a). In species with multiple NORs,interindividual variation is common suggesting thattransposition of rDNA genes from one chromosometo another may occur (reviewed in Phillips, Pleyte &Ihssen, 1989); Reed & Phillips, 1995a). Population-specific differences in the number of NORs havebeen found in S. namaycush (Phillips & Ihssen, 1989),S. alpinus (Phillips, Pleyte & Hartley, 1988) and inS. trutta (Castro et al., 1994).
The size and positions of the single rDNA site havebeen shown to vary extensively in the Atlanticsalmon, S. salar (Phillips & Hartley, 1988) where fivedifferent cytotypes were described (Woznicki &Jankun, 1994) including complete deletion of theNOR (Woznicki, Ra! b & Jankun, 1994).
(3) Minor rearrangements: heterochromatinadditions
The heterochromatic short arms of subtelocentricchromosomes in many salmonid species are probablythe result of paracentric inversions of acrocentricchromosomes followed by tandem duplications of
heterochromatin. If we examine the number ofsubtelocentric chromosomes in the genus Oncorhyn-chus (Table 2) we see a trend toward smaller numbersof subtelocentrics in the most derived members of thegenus. This can be explained if these chromosomesare the result of small inversions that accumulatewith time after Robertsonian rearrangements. If weexamine the type B karyotypes found in othersalmonid genera, we find that they usually have asmall number of subtelocentric chromosomes, sug-gesting that these karyotypes have been derivedrelatively recently because they had not time toaccummulate. An especially striking case is that ofthe endemic Salvethymus svetovidovi (2n¯ 56), inwhich there are no subtelocentric chromosomes.
Length variation in specific chromosome pairsresulting from tandem duplications}deletions ofheterochromatin were observed in C. albula ( Jankun,Klinger & Woznicki, 1995), C. lavaretus}clupeaformis
( Jankun et al., 1995; Phillips, Reed & Ra! b, 1996), S.leucomaenis (Ueda & Ojima, 1983a) and S. salar
(Woznicki & Jankun, 1994). Tandem fusions mayhave been involved in the origin of the largestsubmetacentric chromosome found in the karyotypeof the Atlantic salmon, S. salar (Ueda & Kobayashi,1990).
SEX CHROMOSOMES
Males are the heterogametic sex in all of thesalmonids in which this has been examined(Donaldson & Hunter, 1982). Heteromorphic sexchromosomes have been identified in C. sardinella
(Frolov, 1990b), O. mykiss (Thorgaard, 1977; Ueda& Ojima, 1984b ; Frolov, 1989), O. nerka (Thor-gaard, 1978; Ueda & Ojima, 1984a ; Frolov, 1990a),and S. namaycush (Phillips & Ihssen, 1985a). In S.namaycush and O. mykiss there is an XY}XX system,while an XXY system is found in O. nerka and anXYY system in C. sardinella. The formation ofheteromorphic sex chromosomes often involves het-erochromatin addition as in other animals, and thisappears to be the case in S. namaycush and O. mykiss.
VI. CHROMOSOME BANDING
POLYMORPHISMS
In addition to the polymorphisms in size and numberof NORs and heterochromatic short arms of sub-telocentric chromosomes, polymorphisms have also

16 Ruth Phillips and Petr Ra! b
been observed in the presence}absence and stainingproperties of tandem blocks of highly repetitiveDNA at telomeres and centromeres as revealed by Cbanding, Q banding and DAPI banding.
Large blocks of heterochromatin have been foundat the telomeres of many chromosomes in all of thespecies in the genus Salvelinus. These include S.leucomaenis (Abe & Muramoto, 1974; Ueda & Ojima,1983b), S. namaycush (Phillips & Ihssen, 1985b,1986), S. fontinalis (Mayr, Kalat & Ra! b, 1988), S.alpinus (Hartley, 1989), Salvethymus svetovidovi (Frolov,1993), Salvelinus malma (Ueda & Ojima, 1983b), andS. confluentus, R. Phillips unpublished data. Many ofthese blocks stain positively with quinacrine andDAPI in S. namaycush (Phillips & Zajicek, 1982), S.fontinalis (Mayr et al., 1988), S. alpinus (Pleyte,Phillips & Hartley, 1989) and S. leucomaenis (Abe &Muramoto, 1974), and a few of them stain in theother species. These polymorphisms have beenshown to be inherited (Phillips and Ihssen, 1986)and population differences in the number of chromo-somes with these bands have been found in S.leucomaenis (Ueda & Ojima, 1983a) and S. namaycush
(Phillips & Ihssen, 1989).Smaller blocks of heterochromatin have been
found at telomeres in a number of other salmonidspecies including S. trutta (Hartley & Horne, 1984a),S. salar (Hartley & Horne, 1984a, b), O. mykiss
(Thorgaard, 1976; Mayr, Kalat & Ra! b, 1988).Among Oncorhynchus species the largest polymorphicblocks of heterochromatin are found in O. tschawyt-scha in which several chromosomes have largeinterstitial as well as telomeric blocks which stainpositively with quinacrine and DAPI (Phillips,Zajicek & Utter, 1985b).
VII. KARYOTYPE EVOLUTION
(1) The ancestral salmonid karyotype
The polyploid origin of the Salmonidae has beenwell documented (reviewed in Allendorf & Thor-gaard, 1984). It has been suggested that the diploidancestor of salmonids possessed a karyotype with 48telocentric chromosomes. This karyotype has beenproposed as an ancestral teleost karyotype (Ohno,1970) on the basis of shared commonality and thefact that it is widespread in other fish groups(Klinkhardt, Tesche & Greven, 1995). If this iscorrect, the first tetraploid salmonid would have had96 telocentric chromosomes and an NF number of96. The closest known relatives of salmonids arethe osmeriform fishes (sensu Nelson, 1994, i.e. they
include argentinoids, osmeroids and galaxoids) orosmeroid (sensu Johnson & Patterson, 1996, i.e.they include families Osmeridae, Retropinnidae andGalaxiidae), argentinoid and alepocephaloid fishes.Chromosomes of these fish are not well known, butthe available data include diploid chromosomenumbers ranging from 2n¯ 50 in Sprinchus starksi
(Ohno, 1974), to 2n¯ 52 in Hypomesus pretiotus
(Ohno et al., 1969) and 2n¯ 54–56 in Plecoglossus
altivelis (Yamazaki, 1971), Hypomesus olidus (Vasil’ev,1985), H. transpacificus (Kitsada, Tatewaki &Tagawa, 1980) and Osmerus eperplanus (Nygren,Nilsson & Jahnke, 1971a ; Lajus, 1992), all osmeridsand in the salangid Salangichthys microdon (Nogusa,1960). Chromosome arm numbers vary from 60 to78 with a peak at 60–62, suggesting that a fewchromosomal rearrangements have changed orig-inally telocentric chromosomes in their karyotypes.Galaxid fishes, however, experienced a differenttrend of karyotype changes with decreasing diploidchromosome numbers from 2n¯ 44 in Galaxias
brevipinnis, G. fontanus and G. johnstonii, to 2n¯ 38 inBrachygalaxias bullocki, 2n¯ 32 in G. tanycephalus, G.truttaceus, 2n¯ 30 in G. platei, 2n¯ 28 in Paragal-axias dissimilis, P. eleotroides, P. mesotes and P. julianus
to 2n¯ 22 in G. maculatus (Campos, 1972; Merrilees,1975; Johnston, White & Bick, 1981; White, Bick &Webquing, 1987).
It is reasonable to assume that the diploid ancestorof salmonid fishes may have possessed a karyotypeclose to that of the present osmerid and or osmeroidfishes, i.e. around 2n¯ 50 and NF¯ 60. Hartley(1987) proposes two possible alternatives for thesalmonid tetraploid ancestor : (1) 2n¯ 100, NF¯120 with 20 metacentrics and 80 acrocentrics if thediploid ancestor possessed a karyotype close to thatof present osmeroid fishes and (2) 2n¯ 96, NF¯ 96with 96 acrocentrics which would result if a typicalfish karyotype with 2n¯ 48 and NF¯ 48 underwenttetraploidization. We believe the second hypothesisis more likely.
The graylings have karyotypes of 2n¯ 98–102,NF¯ 146–179. According to the first hypothesis ofHartley (1987), this karyotype could result if theancestral salmonid with 2n¯ 100, NF¯ 120 (20m,80a) underwent 50 additional inversions and twofissions. There are two problems with this scenario.First, inspection of the chromosomes of the graylingshows that all of the metacentrics are very smallchromosomes. Thus, it is more likely that they are allderived from acrocentric}subtelocentric chromo-somes as a result of pericentric inversions. Meta-centric chromosomes in other salmonids are always

17Salmonid chromosome evolution
much larger, except for four small pairs found inBrachymystax and Hucho hucho which again are likelyto be the result of pericentric inversions and notfusion events. When subtelomeric chromosomes areconsidered uniarmed in the calculation of NFnumbers, the vast majority of salmonid karyotypeshave an NF number close to 100, suggesting that theancestral karyotype had an NF number close to 50,a value modal for most of their extant diploidrelatives. Second, there is general agreement that thefishes of the subfamily Coregoninae are the mostbasal in the Salmonidae, with the Thymallinaebeing more derived (Kendall & Behnke, 1984;Sanford, 1990, Stearley & Smith, 1993). Howeverthis second objection is based on the premise thatphylogenetically basal taxa will have ancestralkaryotypes. We will not know if this is correct untilchromosome arm homologies can be accuratelydetermined.
(2) Two modes of karyotype evolution
The three subfamilies of salmonid fishes, i.e. core-gonines, thymallines and salmonines exhibit twotypes of karyotypic changes. The chromosomes ofthe Thymallinae have evolved by inversions retaining
chromosome numbers (2n) close to the karyotype of the
hypothetical tetraploid ancestor but increasing substantially
chromosome arm numbers (NF). The chromosomes ofthe Coregoninae and Salmoninae have evolved bycentric fusions of the Robertsonian type decreasing
chromosome numbers (2n) while retaining chromosome arm
numbers (NF) close to the chromosome number (2n) of the
hypothetical tetraploid ancestor. An exception is Atlanticsalmon, S. salar, where NF values were apparentlychanged by subsequent tandem fusions.
(3) Parallelism in karyotypes of Coregoninaeand Salmoninae
There is a conspicuous parallelism in the karyotypesof coregonines and salmonines. Coregonines havepredominantly A karyotypes, with a few I types andone species with the B type, Coregonus nasus. With theexception of the genus Oncorhynchus, salmonines havepredominately A karytoypes. There is only onespecies in each of the other genera with a B or B«karyotype with a substantial reduction in chromo-some number. Species in the genus Oncorhynchus allpossess type B karyotypes except for O. keta whichhas an I type. Examination of close relatives of thesetwo species suggests that fission of already biarmedchromosomes probably resulted in an increase inchromosome numbers.
(4) Similarities among taxa with type Bkaryotypes
What do these species with B karyotypes have incommon? In each case they have a closely related‘ sister ’ species which did not undergo chromo-some rearrangement. In many cases : Salmo salar vs.S. trutta, Hucho}Parahucho perryi vs. H. hucho spp.,Coregonus nasus vs. C. lavaretus, the species with therearranged karyotype is anadromous. Anadromoussalmonids often have small breeding populations(Allendorf & Waples, 1996) which could facilitatefixation of chromosome rearrangements. In at leasttwo cases, the form with the diverged karyotype is anendemic species found in a single lake along with oneor two ‘sister ’ species. These include Salvethymus
svetovidovi (long-finned char) with ‘ sister ’ speciesSalvelinus elgyticus (small-mouthed char) and Sal-velinus boganidae in Lake Elgygytgyn; and Prosopium
gemmiferum (Bonneville cisco) with ‘ sister ’ species P.spilonotus (Bonneville whitefish) and P. abyssicola
(Bear Lake whitefish) found in Bear Lake. Thepossibility exists that these small ’species flocks ’ arethe result of sympatric speciation in which therecould be strong selection for development of re-productive isolating mechanisms.
(5) Salmonid chromosome evolution andQumsiyeh’s hypothesis
Chromosome evolution in the salmonid fishes ap-pears to be a case of ‘karyotype orthoselection’, inwhich there is a series of rearrangements of a specifictype, in this case Robertsonian fusions, which has ledto a general trend towards lower chromosomenumbers in the more derived species and genera.Thus, this group of fishes is an excellent one in whichto examine the forces involved in orthoselection.Two major possibilities exist : first that the rearrange-ments occur without a selective advantage throughrandom drift and population bottlenecks, or second,that the chromosomal rearrangements themselves orin combination have a selective advantage for thespecies in its environment.
If the rearrangements have been acquired withouta selective advantage, then we would expect thatcases in which massive changes have occurred wouldbe associated with very small effective populationsizes. The migratory behaviour exhibited by anadro-mous species may have such an effect because ofprecise homing (Fleming, 1996). Another conse-quence of such behavior is evolution of small, dwarfmales that do not migrate to the sea but reach sexualmaturity in fresh water. With precise homing,

18 Ruth Phillips and Petr Ra! b
brother and sister matings become much more likelyand in some cases would involve dwarf male brothersmating with their sisters which migrated to the sea.In fact anadromy appears to be associated withreduction in chromosome numbers in the salmonids.All of the Pacific salmon in the genus Oncorhynchus
which have B karyotypes are anadromous and wehypothesize that the freshwater species evolved fromlandlocked forms of the migratory ones. In the casesof scattered B karyotypes among sister species of theother genera, Hucho}Parahucho perryi (2n¯ 62) isanadromous, while Hucho hucho spp. (2n¯ 82–84) isnot ; Salmo salar (2n¯ 54–58) is anadromous, whileSalmo trutta (2n¯ 78–84) is generally not, and somepopulations of Coregonus nasus (2n¯ 58–60) areanadromous while Coregonus lavaretus (2n¯ 80–81) isgenerally not. Stenodus leuichthys with the I karyotypeis typically anadromous. The other cases of chromo-some reduction are found in large inland lakes whichhave small ‘ species flocks ’ which could have beenformed as a result of population bottlenecks, againimplying a small effective population size in aspatially limited lacustrine environment.
If the rearrangements have been acquired becauseof a selective advantage of a smaller karyotype in onetype of environment and a larger karyotype inanother type of environment, what explanationcould account for the observed pattern? Onepossibility that has been advanced recently byQumsiyeh (1994) to explain orthoselection in mam-malian chromosome evolution is that chromosomerearrangements may be selected for because of theireffects on genetic recombination. It has been shownin yeast that increasing the number of chromosomesincreases the number of chiasmata and the frequencyof recombination, although there are a number ofother factors which can influence chiasma frequency.Qumsiyeh (1994) suggests that there may be twostrategies. In the first strategy, increasing the diploidnumber or NF number results in increased re-combination and increased variation that is im-portant in adaptation to variable environments. Inthe second strategy, decreasing the chromosomenumber or NF number results in decreased re-combination which allows adaptation to a special-ized more constant environment and increases theprobability of fixation of new mutations. Thus, wewould expect species inhabiting a wide range ofdifferent habitats to have A karyotypes and speciesadapted to a more restricted, constant environmentto have B karyotypes. Studies of genetic variation insalmonid fishes have shown that the amount ofintraspecific genetic variation is much greater in
freshwater species compared to marine or anadro-mous species (reviewed in Allendorf & Waples,1996). For example in one survey, the averagerelative gene diversity between localities was 3.7%for anadromous and 20.4% for freshwater species(Gyllensten, 1985). Therefore in the salmonids wewould expect a correlation between A karyotypesand the more variable freshwater habitats and typeB karyotypes and the more constant ocean en-vironment and anadromous life cycle and}or special-ized lacustrine environment. In fact, this is thepattern that is observed in salmonid fishes. Asexplained above, in the majority of cases in which‘sister ’ species differ widely in chromosome number,the anadromous species has the lower chromosomenumber.
A particularly striking case of closely relatedspecies with A and B karyotypes is the Atlanticsalmon, S. salar (B«) with a low diploid number andlow NF number and brown trout, S. trutta (A) witha high diploid number and NF number. Atlanticsalmon is a specialized anadromous species thatspawns on the east coast of North America and westcoast of Europe, while brown trout is a more widelydistributed species throughout western and easternEurope (reviewed in Behnke, 1965). It has adaptedto lacustrine, fluvial and anadromous life histories.Transplants of Atlantic salmon rarely succeed whilebrown trout is usually successful in waters outsidetheir native range. For example, brown trout hasnaturalised in numerous North American streams.Population genetic studies have shown that much ofthe genetic diversity in brown trout is distributedbetween populations (Ryman, 1983; Ryman &Utter, 1987), while genetic diversity in Atlanticsalmon is mainly within populations (Stahl, 1987).This would support the idea that higher recom-bination rates may have been involved in producinghabitat-specific variation in brown trout. Fortu-nately, both species are the subjects of extensivegenome mapping efforts at the present time, so weshould have data on comparative recombinationrates soon.
There is one case in the Pacific salmon in which asister species (chum salmon) has undergone a seriesof fissions to produce a larger number of chromo-somes. This species is much more widely ranging andthe only one of the Pacific salmon which inhabits theArctic region in northern Alaska and Siberia. Thus,this species may have increased its genetic variabilityto adapt to more variable environments. In the caseof the ‘ species flocks ’ in the inland lakes, the specieswith the decreased number of chromosomes has

19Salmonid chromosome evolution
undergone dramatic morphological changes. Thesehave been recognized by erecting new genera:Salvethymus in one case and Parahucho in another case.These differences could have been the result ofdecreased recombination that increased the prob-ability of fixation of new mutations. This fact alongwith small population sizes and frequent bottlenecksmay account for such morphological derivations.These new mutations may have allowed these speciesto adapt to specialized environments.
The evolution of the A§ karyotype in graylings,genus Thymallus may also be explained in theframework of Qumsiyeh’s (1994) hypothesis. Thegraylings are morphologically very similar to eachother. This fact suggests a decrease in the probabilityof fixation of new mutations, and lower rate ofmorphological differentiation in evolution. They arefreshwater residents, not migratory and occupy verysimilar, and more variable habitats compared withsalmonids with type A karyotypes. We would expectan increase in either 2n or NF, and a substantialincrease in NF is observed.
There are a number of problems with theapplication of Qumsiyeh’s (1994) hypothesis tosalmonids. Several chars, genus Salvelinus, browntrout, Salmo trutta and some coregonines sometimesexhibit migratory behaviour but have A karyotypes.One explanation might be that this behaviour is ofmore recent origin, possibly since the last glaciation,so there has not been enough time to evolvekaryotype differences. This would be consistent withthe hypothesis that migratory behaviour has evolvedseveral times independently (reviewed in Stearley,1992). Additional evidence for this is the apparentlyreversibility of anadromy in the landlocked species ofOncorhynchus. The landlocked and anadromous formshave the same chromosome number in most casesin which it has been determined (R. Phillips, un-published data). Anadromy would virtually alwaysreduce the effective population size, so this might bea better explanation for the fixation of new chromo-some variants. In the case of the coregonid speciesflocks in the Great Lakes, they apparently have avery consistent karyotype of type A in spite ofdifferences in morphology, habitat, and time ofspawning. They practice broadcast spawning, so theeffective population sizes are probably larger thanthose for salmonine fishes. This suggests that geneticdrift may be more important than selection in theproduction of type B karyotypes.
It is clear that there is a correlation between thetype B karyotypes with fewer chromosomes and theanadromous life style, but we do not know whether
genomes with less recombination were at a selectiveadvantage for animals living in the more constantoceanic environment or in specialized (lacustrine)habitats. A correlation between reduction in geneticdiversity and the anadromous life history has beendocumented in these fishes, but it could be relatedprimarily to reduction in breeding population size,which also would favor fixation of chromosomerearrangements. It is possible that selection forreduced recombination was important primarily incases where major genome reorganization occurredin sister species. Increase in probability of fixation ofnew mutations could account for morphologicallymore diverged taxa. We should be able to test thehypothesis that selection for changes in geneticrecombination occurred by determining chiasmataand frequency of recombination in these sister speciespairs. These data are likely to become availablewithin the next decade for the sister species pair ofAtlantic salmon, Salmo salar (2n¯ 56–58) and browntrout, Salmo trutta (2n¯ 78–84) because geneticmapping projects have been initiated for both ofthese species.
(6) Other aspects of karyotype evolution
There are a number of unanswered questionsregarding the pattern of karyotype evolution ob-served in the salmonids. The most frequent karyo-type in both salmonines and coregonines is 2n¯ 80,NF¯ 100. Why has this type been conserved? Wasit achieved independently in each lineage, or does itreflect a common, ancestral character? Preliminaryevidence suggests that fusions and fissions do notinvolve homeologous chromosomes (reviewed inWright et al., 1983), but that rearrangements occurwithin each ‘subgenome’. Is there a pattern to theparticular chromosomes involved in fusions}fissionsand if so, what are the mechanisms involved? Again,the salmonid genome projects should provide thetools for answering some of these questions. Withpreparation of large insert libraries, and chromosomemapping, probes will become available that willallow determination of chromosome arm homologiesand more accurate reconstruction of chromosomeevolution. We will also learn more about theevolution of the two subgenomes found within allsalmonids from the detailed genetic maps that arecurrently being prepared. It is likely that these extragenes are important in providing the raw materialfor ecological plasticity that is so characteristic ofthese fishes.

20 Ruth Phillips and Petr Ra! b
VIII. CONCLUSIONS
(1) The diploid salmonid ancestor probably had akaryotype with approximately 100 chromosomesand 100 chromosome arms. (2) There are two modesof karyotype evolution in salmonid fishes. In theCoregoninae and Salmoninae, the chromosomeshave evolved by centric fusions of the Robertsoniantype decreasing chromosome numbers (2n) whileretaining chromosome arm numbers (NF) close tothat found in the hypothetical tetraploid ancestor.In the Thymallinae, the chromosomes have evolvedby inversions so that chromosome arm numbers(NF) have increased but chromosome numbers (2n)close to the karyotype of the hypothetical tetraploidancestor have been retained. (3) In the Coregoninaeand Salmoninae the karyotypes can be divided intotwo main types based on chromosome number, withtype A having approximately 80 chromosomes andtype B having approximately 60 chromosomes.Although most of the taxa with type B karyotypesare found in the genus Oncorhynchus, at least oneexample of type B karyotypes is found in all of theother genera. These taxa either have an anadromouslife history or are found in specialized lacustrineenvironments. (4) Selection for increases or decreasesin genetic recombination (Qumsiyeh, 1994) couldhave been involved in the evolution of chromosomenumber in salmonid fishes.
IX. REFERENCES
A, S. & M, J. (1974). Differential staining of
chromosomes of two salmonid species, Salvelinus leucomaenis
(Pallas) and Salvelinus malma (Waldbaum). Proceedings of the
Japanese Academy 50, 507–511.
A, F. W. & T, G. H. (1984). Tetraploidy
and the evolution of salmonid fishes. Evolutionary Genetics of
Fishes (ed. B. J. Turner), pp. 1–53. Plenum Press, New York.
A, F. W. & W, R. S. (1996). Conservation and
genetics of salmonid fishes. In Conservation Genetics: Case
Histories from Nature (ed. J. C. Avise & J. L. Hamrick), ch. 8.
Chapman and Hall, New York.
A-S, K. (1983). Karyological studies on three Salmonidae
in Slovenia using leukocyte culture technique. Acta Biologica
Iug.-Ichtyologia 15, 41–46.
A-S, K. (1985). Chromosomal studies by blood leukocyte
culture technique on three salmonids from Yugoslavian
waters. Journal of Fish Biology 26, 5–12.
A, C. T. & G, J. R. (1986). Chromomycin A$stains
nucleolus organizer regions of fish chromosomes. Copeia 1986,
226–331.
A, E. M., G, M. K. & P, N. V.
(1982). Karyotype of the Sakhalin trout Hucho perryi. Biologyia
Morya 4, 59–60 (in Russian).
A, M. A., L, A. B. & K, T. I.
(1982). Genetic study of some coregonids. In Abstr. IV
Europ. Ichthyol. Congr. Hamburg, No. 12.
A, A. P., T, C., K, A. &
E, P. S. (1997). Mitochondrial DNA sequence
variation and phylogeography among Salmo trutta L. (Greek
brown trout) populations. Molecular Ecology 6, 531–542.
B, J. V. (1981). Intercellular polymorphism of chromo-
some sets in the Atlantic salmon. Tsitologiya 23, 1052–1059 (in
Russian).
B, R. J. & M, R. R. (1977). Cytotaxonomic study
of gila trout, Salmo gilae. Journal of the Fisheries Research Board of
Canada 34, 1041–1045.
B, R. J. (1965). A systematic study of the family
Salmonidae with special reference to the genus Salmo. Ph.D.
thesis, University of California–Berkeley.
B, R. J. (1968). A new subgenus and species of trout,
Salmo (Platysalmo) platycephalus, from southcentral Turkey,
with comments on the classification of the subfamily Salmo-
ninae. Mitteilungen aus dem Hamburgischen Zoologischen Museum
und Institut 66, 1–15.
B, R. J. (1972). The systematics of salmonid fishes of
recently glaciated lakes, Journal of the Fisheries Research Board of
Canada 29, 639–671.
B, R. J. (1992). Native Trout of Western North America.
American Fisheries Society Monogr. 6. Bethesda, Maryland:
American Fisheries Society. 275 pp.
B, L. J., C, M., H, R. & S-
, A. (1970). Chromosome complement of Salmothymus
obtusirostris oxyrhynchus (Steindachner). Acta Biologica Jug. -
Genetika 2, 55–63 (in Serbo-Croatian).
B, L., C, F. & D, J. J. (1991).
Phylogenetic relationships among the subfamily Coregoninae
as revealed by mitochondrial DNA restriction analysis. Journal
of Fish Biology 39 (Suppl. A), 283–290.
B, L., G, R. & B, P. (1992). DNA
sequence variation of the mitochondrial control region among
geographically and morphologically remote European Salmo
trutta populations. Molecular Ecology 1, 161–173.
B, L. & O, A. G. (1995). Genetic diversity of
brown trout (genus Salmo) from its most eastern native range
based on mitochondrial DNA and nuclear gene variation.
Molecular Ecology 4, 285–297.
B, R. A., V, J., W, R. D., L, M. &
R, J. D. (1991). Genetic comparisons of New and Old
World coregonid fishes. Journal of Fish Biology 38, 37–51.
B, H. E. (1968). Cytotaxonomic studies of the coregonine
fishes of the Great Lakes, USA: DNA and karyotype analysis.
Journal of the Fisheries Research Board of Canada 25, 1667–1687.
B, J. E. (1974). A cytotaxonomic study of roundfishes,
genus Prosopium. Copeia 1974, 115–119.
B, J. E. (1975). Cytotaxonomy of the salmonid fish Stenodus
leucichthys. Journal of the Fisheries Research Board of Canada 32,
295–296.
C, H. H. (1972). Karyology of three galaxiid fishes
Galaxias maculatus, G. platei and Brachygalaxias bullocki. Copeia
1971, 171–174.
C, J., R, S., A, J., S, L. & M,
P. (1994). A population analysis of Robertsonian and AgNOR
polymorphisms in brown trout (Salmo trutta). Theoretical Applied
Genetics 89, 105–111.
C, T. M. (1984). Cytotaxonomy of North American
Salvelinus. In Biology of the Arctic Char. Winnipeg, Manitoba, May

21Salmonid chromosome evolution
1981 (ed. L. Johnson & B. L. Burns), pp. 1–32. Winnipeg,
University of Manitoba Press.
C, T. M. & K, S., (1989). Cytotaxonomy and
interrelationships of Pacific basin Salvelinus. Physiology and
Ecology of Japan. Spec. 1, 49–68.
C, I. A. & S, M. B. (1990). Salvethymus
svetovidovi gen. et sp. nova – new endemic fish of the subfamily
Salmoninae from the Elgygytgyn Lake (central Chukotka).
Voprosy Ikhtiologii 30, 201–213 (in Russian).
D, R. H. (1993). Sequence of sockeye salmon type 1 and 2
growth hormone genes and the relationship of rainbow trout
with Atlantic and Pacific salmon. Canadian Journal of
Fisheries and Aquatic Sciences 50, 1738–1748.
D, A. (1959). Chromosomka garnitura na populacite na
ohridanskom pastrmka (Salmo letnica Karaman). Godisen zb.
Prirodno-matem. Fak. Univ. Skopje 12, 115–135 (in Serbo-
Croatian).
D, E. M. & H, G. A. (1982). Sex control in fish
with particular reference to salmonids. Canadian Journal of
Fisheries and Aquatic Sciences 39, 999–110.
D, E. A. (1965). Karyology and systematic status of
Caspian and Black Sea salmons (Salmo trutta caspius Kessler,
Salmo trutta labrax Pallas). Voprosy Ikhtiologii 5, 38–45.
D, E. A. (1977). Utilization of karyological data for
solving problems of systematics and phylogeny of salmonids.
86–95 In The Principles of the Classification and Phylogeny of
Salmonid Fishes (ed. G. A. Scarlato), 86–95. Zool. Inst.
Leningrad (in Russian).
D, E. A. (1989). The basic principles of classification
and phylogeny of the salmonid fishes (Salmoniformes :
Salmonoidei : Salmonidae). In Biology and Phylogeny of Fishes.
(ed. V. M. Korovinoi, pp. 5–16. USSR Academy of Sciences,
Proceedings of the Zoological Institute 201, St. Petersburg (in
Russian).
D, E. A. & R, R. G. (1982). Differentiation
of Salmo ischan Kessler with respect to karyological and mor-
phological data. Voprosy Ikhtiologii 22, 36–48 (in Russian).
F, I. A. (1996). Reproductive strategies of Atlantic
salmon: ecology and evolution. Reviews in Fish Biology and
Fisheries 6, 379–416.
F, S. V. (1986). Polymorphism and mosaicism in ad-
ditional chromosomes of the Siberian least cisco. Tsitologiya
28, 740–744 (in Russian).
F, S. V. (1989). Differentiation of sex chromosomes in the
Salmonidae. I. Karyotype and sex chromosomes in Parasalmo
mykiss. Tsitologiya (Moscow) 31, 1391–1394 (in Russian).
F, S. V. (1990a). Differentiation of sex chromosomes in
the Salmonidae. II. Karyotype and sex chromosomes in
Oncorhynchus nerka. Tsitologiya (Moscow) 32, 509–514 (in
Russian).
F, S. V. (1990b). Differentiation of sex chromosomes in
the Salmonidae. III. sex chromosomes in Coregonus sardinella.
Tsitologiya (Moscow) 32, 659–663 (in Russian).
F, S. V. (1991a). Karyotype of Stenodus leucichthys nelma
from the Anadyr River. Chromosome Information Service 52,
11–12.
F, S. V. (1991b). Karyotypes of Salvelinus leucomaenis
(Pallas) and white char S. albus Glubokovsky from the
Kamchatka River basin. In Biology of the Chars of the Far East
(ed. I. A. Chereshnev & M. K. Glubokovsky) pp. 103–111
Vladivostok, F. E. B. Russian Academy of Sciences (in
Russian).
F, S. V. (1992a). Some aspects of karyotype evolution in
the Coregoninae. Polish Archives of Hydrobiology 39, 509–515.
F, S. V. (1992b). Karyotype and chromosomal variability
in smallmouth char, Salvelinus elgyticus from Lake Elgygytgyn.
Journal of Ichthyology 32, 61–66.
F, S. V. (1993). Unique karyotype and constitutive
heterochromatin in the endemic salmonine fish Salvethymus
svetovidovi. Journal of Ichthyology 33, 42–49.
F, S. V. (1994a). Variability of the karyotype of the
boganid char (Salvelinus boganidae Berg) from Lake Elgygyt-
gyn. Genetika 30, 262–264 (in Russian).
F, S. V. (1994b). Comparative karyology of the salmonid
fishes of Lake Elgygytgyn. In: Systematics, Biology and Biotech-
nology of Breeding of Salmonid Fishes : Proceedings of the Fifth
Conference, St. Petersburg 203–204 (in Russian).
F, S. V. (1995). Comparative karyology and karyotype
evolution of chars. Journal of Ichthyology 35, 14–23.
F, S. V. (1997). Divergence of Karyotypes of Chukotsk
chars of the genus Salvelinus. Reports of the Academy of Sciences
357 : 703–705 (in Russian).
F, S. V. & F, V. N. (1997). Karyological
differentiation of Levanidov’s char, Salvelinus levanidovi and S.
malma from the Yama River. Biologiya Morya 23, 376–379 (in
Russian).
F, S. V. & F, V. N. (1998). Karyotype of a
Chikchi char from Lake Estiched (East Chukotka. In Biology
and Evolution of Chars of the Northern Hemisphere p. 14. Abstracts
of the ISACF Workshop, 2–10 September 1998. Kamchatka,
Russia.
F, S. V. & F, V. N. (1999). Nucleolar organizer
regions as a phylogenetic marker in karyotypes of the
Dollyvarden, Salvelinus malma malma and the char S. levanidovi.
Biologiya Morya 25 : 407–409 (in Russian).
F, S. V. & F, V. N. (2000). Polymorphism and
karyotype divergence in taimens of the Hucho genus. Genetika
36, 237–240 (in Russian).
F, S. V. & M, I. N. (1992). Karyotypes of Salvelinus
malma and S. leucomaenis from North Primorye. Chromosome
Information Service 52, 12–14.
F, S. V., F, V. N. & M, A. V.
(1997). Karyotype of Salvelinus malma from Yama River and
taxonomical status of the northern and southern S. malma.
Biologiya Morya 5, 309–313 (in Russian).
G, E., B, L. & G, R. (1994).
Mitochondrial control region and protein coding sequence
variation among phenotypic forms of brown trout Salmo trutta
from northern Italy. Molecular Ecology 3, 161–171.
G, M. K. & F, S. V. (1994). Phylogenetic
relations and the systematics of charr’s fishes of Lake
Elgygytgyn. Journal of Ichthyology 34, 128–147.
G, J. R. & G, G. A. E. (1975). Chromosome cytology
and polymorphism in the California high Sierra golden trout
(Salmo aguabonita). Canadian Jounral of Genetics and Cytology 17,
41–53.
G, J. R., A, J. C. & G, G. A. E. (1977). Chromosome
cytology in the cutthroat trout series Salmo clarkii (Sal-
monidae). Cytologia 43, 128–147.
G, S. A. & G, G. V. (1981). Chromosome
polymorphism of the pink salmon Oncorhynchus gorbuscha
(Walbaum) Tsitologiya 23, 954–960 (in Russian).
G, U. (1985). The genetic structure of fish: differences
in the intraspecific distribution of biochemical genetic

22 Ruth Phillips and Petr Ra! b
variation between marine, anadromous, and freshwater
species. Journal of Fish Biology 26, 691–699.
H, S. (1960). Zur Kenntnis der Gattung Salmothymus
Berg. Publ. Za Ribarstvo NRM-Skopje 3, 39–56.
H, S. (1961). Zur Kenntnis der Gattung Salmothymus
ohridanus (Steindachner). Verhandlung Internat. Verein Limnologie
14, 785–791.
H, S. E. (1987). The chromosomes of salmonid fishes.
Biological Reviews 62, 197–214.
H, S. E. (1989). Chromosome and constitutive hetero-
chromatin distribution in Arctic charr, Salvelinus alpinus (L.)
(Pisces : Salmonidae). Genetica 79, 161–166.
H, S. E. & D, W. S. (1994). Characterization
and distribution of genomic repeat sequences from Arctic char
(Salvelinus alpinus). In Genetics and Evolution of Aquatic Organisms
(ed. A. R. Beaumont), pp. 271–280. London: Chapman
&Hall.
H, S. E. & H, M. T. (1982). Chromosome poly-
morphism in the rainbow trout (Salmo gairdneri Richardson).
Chromosoma 87, 461–468.
H, S. E. & H, M. T. (1984a). Chromosome
polymorphism and constitutive heterochromatin in Atlantic
salmon, Salmo salar. Chromosoma 89, 377–380.
H, S. E. & H, M. T. (1984b). Chromosome
relationships in the genus Salmo. Chromosoma 90, 229–237.
Hı!, J., H, K., N, J. & S! , L. (1988). The
Eurasian Huchen, Hucho hucho. Perspectives in Vertebrate
Sciences. Dr W. Junk Publishers, Dordrecht, 239 pp.
H, W. M. & B, D. A. (1980). Controlled silver-
staining of nucleolus organizer regions with a protective
colloidal developer: a 1-step method. Experientia 36, 1014–
1015.
H, B., D, R., G, R., H, L.-E.,
P, R. & T, J. Abstract P342. Constructing a
genetic map of salmonid fishes : SALMAP. Plant and Animal
Genome Conference VI, January 1998, San Diego, Cali-
fornia, USA.
J, V. M., M, A. M. & P, E. S.
(1979). Karyotype of the Baikal omul. In Problemy ekologii
pribajkalya (Tez dokl. Ros. Resp. sovetsk.), p. 242. Irkutsk
(in Russian).
J, M., K, M. & W, P., (1995). Chromo-
some variability in European vendace (Coregonus albula L.)
from Poland. Caryologia 48, 165–172.
J, M., R! , P. & V, J. (1991). A karyotype study
of vendace, Coregonus albula (Pisces, Coregoninae). Hereditas
115, 291–294.
J, M., R! , P., V, J. & L, M. (1995).
Chromosomal polymorphism. In Coregonus lavaretus popu-
lations from two locations in Finland and Poland. Archives of
Hydrobiology Special Issues of Advanced Limnology 46, 1–11.
J, M., M P., P, B. G., R! , P., R! ! , M.,
S, L. (2000). rRNA genes map to chromosomes 10,
11 and 12 in European whitefish (Coregonus lavaretus) and to
chromosomes 1, 5, 9 and 10 in vendace (Coregonus albula).
Chromosome Research 8 : 455–456.
J, D. G. & P, C. (1996). Relationships of
Lower Euteleostean fishes. In Interrelationships of Fishes (ed. M.
L. S. Stiassny, L. R. Parenti, G. D. Johnson) pp. 251–332.
Academic Press.
J, R. C. W, R. W. G. & B, Y. A. E. (1981).
Cytotaxonomy of seven species of Galaxias (Pisces : Galaxiidae)
from Tasmania. Genetica 56, 17–21.
K, T. I. (1978). Investigations of the karyotypes of two
kinds of whitefish. Izvestiya GosNIORKh 130, 50–55.
K, M., M, B., R! , P. & S, W. (1988).
Heterochromatin studies and localization of NORs in Euro-
pean grayling (Thymallus thymallus, Pisces, Thymallidae).
Caryologia, 41, 245–249.
K, Y. S. & P, E. H. (1973). Somatic chromosomes of the
Manchurian trout, Brachymystax lenok (Salmonidae). Chromo-
some Information Service 15, 10–11.
K, Y., P, J. & T, C. (1992).
Chromosomal studies in brown trout (Salmo trutta, L.)
populations. Cytobios 72, 117–124.
K, M. (1966). Beitrag zur Kenntnis der Salmoniden
Sudeuropas. Hydrobiologia 28, 1–41.
K, A. W., Jr. & B, R. J. (1984). Salmonidae:
development and relationships. In H. G. Moser (ed.) Ontogeny
and Systematics of Fishes. American Society of Ichthyologists and
Herpetologists Special Publication 1, 142 – 149.
K, J., T, R. & T, M. (1980). Chromo-
somes of the pond smelt Hypomesus transpacificus nipponensis.
Chromosome Information Service 28, 8–9.
K, M., T, M. & G, H. J. (1995). Database
of Fish Chromosomes. Westarp Wissenschaften, Magdeburg, 237
pp.
K, M. (1997). Freshwater fishes of western and central
Europe. Biologia (Bratislava) 52, (Suppl. 5), 1–271.
K, N. I. (1971). Intraspecific variability of karyotypes
of the chum salmon (Oncorhynchus keta) (Walbaum). Journal
of Ichthyology 11, 977–983.
L, D. L. (1992). Chromosome sets of 13 fish species from the
White Sea. In Problems of Study, Rational Exploration and
Conservation of Natural Resources of the White Sea, pp. 254–256.
Petrozavodsk (in Russian).
L, G. M. & W, J. E. (1981). Mitotic and meiotic
analysis of brook trout Salvelinus fontinalis. Journal of Heredity
72, 321–327.
L, A., F, K. & S, A. A. (1964). No-
menclature for centromeric position on chromosomes. Here-
ditas 52, 201–220.
L, E. J. & T, G. H. (1979). Karyotypic
and evolutionary relationships of the Yellowstone (Salmo clarki
bouvieri) and West-Slope (S. c. lewisi) cutthroat trout. Journal
of the Fisheries Research Board of Canada 36, 630–635.
M, A. N., (1982a). Chromosomal polymorphism of
East Siberian grayling and some problems of karyotype
evolution in the subfamily of graylings. Tsitologiya i genetika 16,
53–56 (in Russian).
M, A. N. (1982b). Karyotype of Siberian grayling
Thymallus arcticus (Pallas) from the basins of North-Eastern
Asia. Biology of Freshwater Animals of Far East, 84–86 (in
Russian).
M, A. N. (1987). The problem of stability of karyotype
characters in the grayling fishes. Tsitologiya 29, 490–496 (in
Russian).
M, A. N. (1992). Relationships of coregonid fishes :
karyological aspects. Polskie Archiwum Hydrobiologii 39,
517–522.
M, A. N., F, S. V. & S, A. A., (1983).
Some problems of karyotype of graylings. In Biological Problems
of North, pp. 192–193. 10th All Union Symposium, Magadan
(in Russian).
Mı!, P., V, A., B, C., A, J., A, R, &
S, L. (1991). Cytological characterization of hatchery

23Salmonid chromosome evolution
stocks and natural populations of sea and brown trout from
northwestern Spain. Heredity 66, 9–17.
Mı!, P., V, A., B, C, C, J. & S, L.
(1993). Quantitative analysis of the variability of the NOR
region in Salmo trutta (Pisces, Salmonidae). Genome 36,
1119–1123.
M, R. (1945). L’ evolution de la formule chromosomiale
chez les vertebrees. Experientia 1, 50–56, 78–86.
M, B., K, M. & R! , P. (1988). Heterochromatins and
band karyotypes in three species of salmonids. Theoretical and
Applied Genetics 76, 45–53.
M, B., R! , P. & K, M. (1986). Localisation of NORs
and counterstain-enhanced fluorescence studies in Samo
gairdneri and Salmo trutta (Pisces, Salmonidae). Theoretical and
Applied Genetics 71, 703–707.
M, Y. E. & T, A. T. (1986). Karyotype of the
Amu Darya trout Salmo trutta oxianus (Salmoniformes, Salmo-
nidae). Zoologicheskiy Zhurnal. 65, 1582–1586 (in Russian).
M, S. (1957). Osservazioni cariologiche su Salmo carpio.
Bollettino di Zoologia 24, 253–258 (in Italian).
M, M. H. (1975). Karyotype of Galaxias maculatus from
New Zealand. Copeia 1975, 176–178.
M, R. R. (1972). Classification of native trouts of Arizona
with the description of a new species, Salmo apache. Copeia 3,
401–422.
M, J., A, J. & F, H. (1974). Karyotypes of
nine species of Salmonidae. Chromosome Information Service 17,
20–23.
N, J. S. (1994). Fishes of the World. John Wiley and Sons.
600 pp.
N, S. (1960). A comparative study of chromosomes in
fishes with particular considerations of taxonomy and
evolution. Mem. Hyogo. Univ. Agric. 3, 1–62.
N, C. R. (1961). Comparative osteology of representative
salmonid fishes, with particular reference to the grayling
(Thymallus arcticus) and its phylogeny. Journal of the Fisheries
Research Board of Canada 18, 679–791.
N, A., N, B. & J, M. (1971a). Cytological
studies in Thymallus thymallus and Coregonus albula. Hereditas 67,
269–274.
N, A., L, U., N, B. & J, M. (1971b).
Cytological studies in Coregonus from Sweden. Annales Acade-
miae Regiae Scientiarum Uppsaliensis 15, 5–20.
N, A., N, B. & J, M. (1971c). Cytological
studies in the smelt (Osmerus eperplanus L.) Hereditas 67,
283–286.
O, T. H. & P, R. B. (1999). Phylogeny of
salmonine fish based on growth hormone introns : Atlantic
(Salmo) and Pacific (Oncorhynchus) salmon are not sister taxa.
Molecular Phylogenetics and Evolution 11, 381–393.
O, S. (1970). Evolution by Gene Duplication. Springer-Verlag,
New York.
O, S. (1974). Protochordata, Cyclostomata and Pisces. In
Animal Cytogenetics (ed. B. John), vol. 4. Chordata, I. Born-
traeger, Berlin.
O, S., M, J. K, J. & A, N. B. (1969).
Diploid–tetraploid relationship in clupeoid and salmonid
fishes of the Pacific. Chromosomes Today 2, 139–147.
O, A. G. (1991). Genetic divergence and phylogenetic
relationships between lenoks of genus Brachymystax and
huchens of genera Hucho and Parahucho. Genetika (Moscow) 27,
2127–2135 (in Russian).
P! , A. M., M! , P. & Gı!-V, E. (1993).
Multichromosomal location of ribosomal genes and hetero-
chromatin association in brown trout. Chromosome Research
1, 63–67.
P! , A. M., M! , P. & Gı!-V, E. (1994).
Organization and chromosomal localization of the major
histone cluster in brown trout, Atlantic salmon and rainbow
trout. Chromosoma 103, 147–152.
P, R. B., G, L. I., W, K. M. & DC, F.
(1999). Phylogenetic analysis of the ribosomal ITS1 of the
rDNA supports three subgroups of Dolly Varden char (S.
malma). Canadian Journal of Fisheries and Aquatic Science
56, 1–8.
P, R. B. & H, S. E. (1988). Fluorescent banding
patterns of the chromosomes of the genus Salmo. Genome 30,
193–197.
P, R. B. & I, P. E. (1985a). Identification of sex
chromosomes in lake trout (Salvelinus namaycush). Cytogenetics
and Cell Genetics 39, 14–18.
P, R. B. & I, P. E. (1985b). Chromosome banding
in salmonid fishes : nucleolar organizers in Salmo and Salvelinus.
Canadian Journal of Genetics and Cytology 27, 433–440.
P, R. B. & I, P. E. (1986). Inheritance of Q band
chromosomal polymorphisms in lake trout (Salvelinus namay-
cush). Journal of Heredity 77, 93–97.
P, R. B. & I, P. E. (1989). Population differences in
chromosome banding polymorphisms in lake trout (Salvelinus
namaycush). Transactions of the American Fisheries 118, 64–73.
P, R. B. & K, A. R. (1987). A Robertsonian
polymorphism in pink salmon (Oncorhynchus gorbuscha) in-
volving the NOR region. Cytogenetics and Cell Genetics 44,
148–152.
P, R. B. & K, A. R. (1988). High frequency of
translocation heterozygotes in odd year populations of pink
salmon (Oncorhynchus gorbuscha). Cytogenetics and Cell Genetics 48,
178–182.
P, R. B., M, M. P., K, I. & R, K. M.
(2000). Phylogenetic analysis of nuclear and mitochondrial
sequences supports inclusion of Acantholingua ohridana in Salmo.
Copeia 2000 (2), 546–550.
P, R. B. & O, T. H. (1997). Evolutionary relation-
ships among salmonid fishes inferred from nuclear and
mitochondrial sequences. In Molecular Systematics of Fishes (ed.
T. Kocher & C. Stepien), (pp. 145–162). Academic Press.
P, R. B., O, T. H. & D, E. L. (1995). Support
for the paraphyly of the genus Hucho based on ribosomal DNA
restriction maps. Journal of Fish Biology 47, 956–961.
P, R. B., P, K. A. & H, S. E. (1988). Stock-
specific differences in the number and chromosome positions
of the nucleolar organizer regions in Arctic char (Salvelinus
alpinus). Cytogenetics and Cell Genetics 48, 9–12.
P, R. B., P, K. A. & I, P.E. (1989). Patterns
of chromosomal nucleolar variation in fishes of the genus
Salvelinus. Copeia 1989 (1), 47–53.
P, R. B. & R, K. M. (1995). Molecular cytogenetic
analysis of the double CMA3 chromosome in lake trout,
Salvelinus namaycush. Cytogenetics and Cell Genetics 70, 104–107.
P, R. B. & R, K. M. (1996). Application of fluores-
cence in situ hybridization (FISH) to fish genetics Aquaculture.
140, 197–216.
P, R. B., R, K. M. & R! , P. (1996). Revised
karyotypes of coregonid fishes from the Great Lakes. Canadian
Journal of Zoology 74, 323–329.

24 Ruth Phillips and Petr Ra! b
P, R. B. & Z, K. D. (1982). Q band chromosomal
banding polymorphisms in lake trout (Salvelinus namaycush).
Genetics 101, 222–234.
P, R. B., Z, K. D. & U, F. M. (1985a).
Chromosome banding in salmonid fishes : nucleolar organizer
regions in Oncorhynchus. Canadian Journal of Genetics and Cytology
28, 502–510.
P, R. B., Z, K. D. & U, F. M. (1985b). Q
band chromosomal polymorphisms in chinook salmon
(Oncorhynchus tshawytscha). Copeia 1985 (2), 273–278.
P, K. A., P, R. B. & H, S. E. (1989). Q
band chromosomal polymorphisms in Arctic char (Salvelinus
alpinus). Genome 32, 129–133.
Q, M. B. (1994). Evolution of number and morphology
of mammalian chromosomes. Journal of Heredity 85, 455–465.
R! , P. & J, M. (1992). Chromosome studies of
coregonine fishes : a review. Pol Arch. Hydrobiol. 39, 523–465.
R! , P. & L, P. (1982). Chromosome study of Danube
salmon Hucho hucho (Linnaeus, 1758) (Pisces : Salmonidae).
Folia Zoologica 31, 181–190.
R! , P., R, K. M., J, M. & P, R. B (1996).
Chromosomal NOR phenotypes in several species of Coregonus
as revealed by CMA3, silver staining and FISH with notes on
the ancestral position of NORs in salmonid fishes. VI.
International Symposium on Biology and Management of
Coregonid Fishes, Konstanz, Germany Sept. 23–29, 1996.
R! , P., S, V. & F, M. (1994). Cytogenetics,
cytotaxonomy and biochemical genetics of huchonine sal-
monids. Fol. Zool. 43, 97–107.
R, K. M. & P, R. B. (1995a). Molecular cytogenetic
analysis of the double CMA3 chromosome in lake trout,
Salvelinus namaycush. Cytogenetics and Cell Genetics 70, 104–107.
R, K. M. & P, R. B. (1995b). Molecular charac-
terization and cytogenetic analysis of highly repeated DNAs
of lake trout, Salvelinus namaycush. Chromosoma 104, 242–251.
R, K. M., B, S. K. & P, R. B. (1995).
Microdissection of the Y chromosome and FISH analysis of
the sex chromosomes of lake trout, Salvelinus namaycush).
Chromosome Research 3, 221–226.
R, K. M., D, M. O. & P, R. B. (1997).
Characteristics of two salmonid repetitive DNA families in
rainbow trout (Oncorhynchus mykiss). Cytogenet. Cell Genetics
79, 184–187.
R, C. T. (1914). The systematic arrangement of the fishes
of the family Salmonidae. Annals of the Magazine of Natural
History Vol. XIII (Eighth Series), 405–408.
R, J. D. M, L. D., B, R. A. & C, T. J.
(1996). Variation in D loop sequences of mitochondrial DNA
of new and old-world coregonids. VI International Sym-
posium on Biology and Management of Coregonid Fishes,
Konstanz, Germany, Spet 23–29, 1996.
R, F. L. (1970). Atlantic salmon (Salmo salar) chromo-
somes and speciation. Transactions of the American Fisheries
Society 99, 105–111.
R, R. G. (1982). A comparative analysis of the
karyotypes of the Sevan trout Salmo ischan Kessler. Tsitologiya
(Moskva) 24, 66–77 (in Russian).
R, R. G. (1984). Origin and species identity of
alabalakh trout (genus Salmo, Salmonidae) as revealed by
karyological traits. Voprosy Ikhtiologii 24, 368–373 (in Russian).
R, R. G. (1989). Karyology and origin of the Trans-
caucasian trouts. Acad. Sci. Armenian SSR Press, Yerevan
(in Russian).
R, N. (1983). Patterns of distribution of biochemical
variation in salmonids : differences between species. Aquaculture
33, 1–21.
R, N. & U, F. M. (1987). Population Genetics & Fishery
Management. Seattle : University of Washington Press.
S, S. L. & P, R. B. (1997). Biogeography of North
American coregonids based on ribosomal DNA spacer
sequences. Can. J. Fish. Aquat. Sci. 54, 1494–1503.
S, C. P. J. (1990). The phylogenetic relationships of
salmonid fishes. Bull. Br. Mus. Nat. Hist (Zool) 56, 93–97.
S, K. A. (1994). Patterns of diversity and processes of
speciation in Arctic char. Nordic. J. Freshwat. Res. 71, 81–91.
S, K. A. & V, V. P. (1976). About sympatric
populations in char of the genus Salvelinus (Salmoniformes,
Salmonidae) from the Lake Nachikinskoye (Kamchatka). Sci.
Rep. Higher School, biol. sci. (2), 80–87 (in Russian).
S, S. O. (1979). Chromosome set of European grayling,
Thymallus thymallus, (L.). Voprosy Ichtiologii, 19 (2), 246–250
(in Russian).
S, S. O. (1985a). Evolution and distribution of graylings
(genus Thymallus) : the karyological approach. Abstr. of.
Ph.D. Thesis, Leningrad, 1985, 22 pp. (in Russian).
S, S. O. (1985b). Karyological analysis of three popu-
lations of Thymallus thymallus, (L.) from Kama River basin.
Voprosy Ichtiologii, 25 (6), 899–910 (in Russian).
S, S. O. & Z, E. A. (1979). Karyological
characteristics of graylings in area of species contact (Ural).
p. 80–82 In Structure and function of water biocenosis in the
Ural region: their rational exploration and conservation.
Sverdlovsk (in Russian).
S, S. O. & Z, E. A. (1982). Karyotypes of
isolated populations of Thymallus arcticus (Pallas) in Ob River
basin. Voprosy Ichtiologii, 22 (1), 27–35.
S, B. A., G, K. S. & U, J. C. (1990).
Chromosomal and mitochondrial DNA characterization of a
population of dwarf cisco (Coregonus artedii) in Minnesota.
Canadian Journal of Fisheries and Aquatic Science 1562–1569.
S, R. C. (1963). Chromosome morphology and species
evolution in the five North American species of Pacific salmon
(Oncorhynchus). Journal of Morphology 112, 77–95.
S, G. R. & S, R. F. (1989). The classification and
scientific names of rainbow and cutthroat trouts. Fisheries 14,
4–10.
S, G. R. & S, R. F. (1993). Phylogeny of the
Pacific trouts and salmons (Oncorhynchus) and genera of the
family Salmonidae. Transactions of American Fisheries Society
122, 1–33.
S, G. R. & T, T. N. (1992). Morphological cladistic
study of coregonine fishes. Polskie Archiwum Hydrobiologii 39,
479–490.
S, L., N, G. L. & C, So. (1989). Chromo-
some variability in introduced whitefishes (genus Coregonus)
from 2 lakes in Central Italy. Copeia 1989, 189–192.
S, G. (1987). Genetic population structure of Atlantic
salmon. Chapter 5 In Ryman, N. and Utter, F. (eds.)
Population Genetics and Fishery Management. Seattle :
University of Washington Press.
S, R. F. (1992). Historical ecology of the Salmoninae,
with special reference to Oncorhynchus. In R. L. Mayden, (ed.)
Systematic Historical Ecology and North American Fresh-
water Fishes. Stanford U. Press.
S, R. F. & S, G. R. (1993). Phylogeny of the

25Salmonid chromosome evolution
Pacific trouts and salmons (Oncorhynchus) and genera of the
family Salmonidae. Transactions of American Fisheries Society
122, 1–33.
S, A (1975). A comparative osteological study of the
Balkan endemic genus Salmothymus in relation to its classi-
fication. Zoologicheskii Zhurnal 54, 1174–1190 (in Russian).
T, G. H. (1976). Robertsonian polymorphism and
constitutive heterochromatin distribution in chromosomes of
the rainbow trout. Cytogenet. Cell Genet. 17, 174–184.
T, G. H. (1977). Heteromorphic sex chromosomes in
male rainbow trout. Science 196, 900–902.
T, G. H. (1978). Sex chromosomes in the sockeye
salmon: a Y-autosome fusion. Canadian Journal of Genetic
Cytology 20, 349–354.
T, G. H. (1983). Chromosomal differences among
rainbow trout populations. Copeia 1983, 650–662.
U, T. (1983). Cytogenetical characters of 4 species in the
salmonid fishes. Bulletin of the Faculty of Education, Utsunomiya
University 34, 53–61 (in Japanese).
U, T. & K, J. (1990). Karyotype differentiation of
Atlantic salmon, Salmo salar, especially the sequential karyo-
type change. La. Kromosomo II-58, 1967–1972.
U, T. & O, Y. (1983a). Geographic and chromosomal
polymorphisms in the iwana (Salvelinus leucomaenis). Proceedings
of Japanese Academy 59 (B) 8, 259–262.
U, T. & O, Y. (1983b). Karytoypes with C banding
patterns of two species in the genus Salvelinus of the family
Salmonidae. Proceedings of Japanese Academy 59, Ser B,
343–346.
U, T. & O, Y. (1984a). Sex chromosomes in the
kokanee salmon, Oncorhynchus nerka. Bulletin of Japanese Society
of Fisheries 50, 1495–1498.
U, T. & O, Y. (1984b). Sex chromosomes in the
rainbow trout, Oncorhynchus mykiss. Bulletin of Japanese Society of
Fisheries 50, 1499–1504.
U, T. (1972). Chromosome of offspring resulting from
crossing coho salmon and brook trout. Japanese Journal of
Ichthyology 19, 166–171.
V, V. P. (1975). Karyotypes of some forms of Arctic char
Salvelinus alpinus (L.) from the basins of Kamchatka. Voprosy
Ikhtiologii 15 (3/92), 417–430 (in Russian).
V, V. P. (1983). Some aspects of chromosomal differen-
tiation of fishes. pp. 166–180. In Biological aspects of fisheries :
problems of genetics and selection. Nauka Press. Leningrad
(in Russian).
V, V. P. (1985). Evolutionary karyology of fishes. Nauka
Press, Moscow, 300 pp. (in Russian).
V, R. M. (1964). Chromosome set of peled and
baunt whitefish. Tsitologiya 6, 636–639 (in Russian).
V, R. M. (1975a). Karyotypes of Salvelinus leuco-
maenis (Pallas) and S. malma (Walbaum) (Pisces, Salmoni-
formes, Salmonidae). Zoologicheskii Zhurnal 54 (5), 787–789 (in
Russian).
V, R. M. (1975b). Karyotypes of the endemic charrs
from the Kronotskoe Lake. Tsitologiya 17 (4), 464–466 (in
Russian).
V, R. M. (1976). Comparative karyology, evolution
and speciation in the chars of the genus Salvelinus. pp. 16–19.
In: Ecology and systematics of salmoniform fishes. Leningrad,
Zoologicheskii Institut, Academy of Sciences USSR. 16–19 (in
Russian).
V, R. M. (1978a). Mechanisms of speciation of the
chars of Lake Kronotskoye. Moscow: Nauka (in Russian).
V, R. M. (1978b). The evolution of karyotypes in
chars of the genus Salvelinus. Tsitologiya 20 (7), 833–838 (in
Russian).
V, R. M. (1978 c). Karyotypes of chars from eastern
Asia. Communist Institute of Marine Biology 3, 21–23) (in
Russian).
V, R. M. & E, L. N. (1982). Chromo-
some set of broad whitefish and pidschian and problems with
karyotypic divergence in whitefish. Tsitologiya, 24, 797–801
(in Russian).
V, R. M., E, L. N., M, A. N.,
F, S. V. & S. A. A. (1983). Karyotype
divergence in the genus Coregonus. Tsitologiya 25, 1309–1915
(in Russian).
V, R. M. & G, M. K. (1977). Mechan-
isms and rate of speciation of the chars of the genus Salvelinus
(Salmonidae, Pisces). Doklady USSR Acad. Sci. 235 (4),
946–949) (in Russian).
V, R. M., M, A. N. & S, A. A.
(1985). The chromosomal sets of Brachymystax lenok and Hucho
taimen and the divergency of the salmonid genera. Tstiologiya
27, 703–709 (in Russian).
V, R. M. & M, T. G. (1978). Chromosome
set of Amur whitefish and some problems in the evolution of
the whitefish karyotype. Tsitologiya 20, 967–970.
W, R. W. G., B, Y. A. E. & W, F. (1987).
Karyology of the genus Paragalaxias (Pisces, Salmoniformes,
Galaxiidae). J. Fish Biol. 31, 583–585.
W, P. & J, M. (1994). Chromosome polymorph-
ism of Atlantic salmon (Salmo salar) from Dzwina River, Baltic
Sea Basin: arm length and NOR location variation of the
eighth chromosome. Canadian Journal of Zoology 72, 364–367.
W, P., R! , P. & J, M. (1994). New cytotype of
a highly polymorphic NOR bearing chromosome pair in
Atlantic salmon Salmo salar (L.). Cytobios 79, 59–62.
W J. E., J, K., H, A. & M, B. (1983).
Meiotic models to explain classical linkage, pseudolinkage,
and chromosome pairing in tetraploid derivative salmonid
genomes. In Isozymes : Current Topics in Biological and
Medical Research Vol. 10, Genetics and Evolution 239–260.
Y, F. (1971). A chromosome study of the ayu, a
salmonid fish. Bulletin of Japanese Society of Fisheries 73, 707–710.
Y, W. P. W, P. A. C, B. H., K, P. &
T, G. H. (1998). A detailed linkage map of rainbow
trout produced using doubled haploids. Genetics 148, 1–13.
Z, M. T. & V, I. (1975). C banding patterns in
S. trutta, a species of tetraploid origin. Genetica 45, 531–536.




















