
Transplantation of cerebellar anlagen to hosts with genetic cerebellocortical atrophy
10
Anat Embryol (1987) 176:145 154 Anatomy and Embryology Springer-Verlag 1987 Original articles Transplantation of cerebellar anlagen to hosts with genetic cerebellocortical atrophy Lazaros C. Triarhou 1, 3,., Walter C. Low 2, 3, and Bernardino Ghetti l, 3 1 Department of Pathology, Division of Neuropathology, 2 Department of Physiology and Biophysics, and 3 Program in Medical Neurobiology, Indiana University School of Medicine, Indianapolis, Indiana, USA Summary. Embryonic cerebellar grafts from genetically nor- mal donors were implanted into the cerebellomedullary cistern of adult 'Purkinje cell degeneration' (pcd) and weav- er mutant mice, which are respectively characterized by the selective loss of Purkinje and granule cells. Grafts placed into both mutant recipients exhibited a layered cellular or- ganization reminiscent of the normal cerebellar cortex. Mo- lecular, Purkinje, and granule cell layers were identifiable. Grafted Purkinje cells displayed characteristic cytological features, such as hypolemmal cisterns in association with mitochondria in the perikaryon, and lamellar structures in their axons. The cytological features of granule cell somata in the grafts appeared similar to those of mature granule cells. Electron microscopic examination of the molecular layer of the grafts revealed the presence of parallel fibers, which were not oriented in a parallel fashion; axon termi- nals of such fibers were often presynaptic to dendritic spines. The number of parallel fibers was markedly reduced in grafts implanted into both mutants compared to the nor- mal cerebellar cortex; however, this phenomenon is com- monly seen in cerebellum in tissue culture and in cerebellar transplants into normal hosts. It is concluded, therefore, that the environment of the mutant hosts does not affect the survival of Purkinje or granule cells and that transplan- tation of solid cerebellar grafts in the neurological mutants studied does not seem to pose any apparent limitations beyond those inherent to the process of cerebellar growth and differentiation outside its normal environment. Key words: Neurological mutant mice 'Purkinje cell de- generation' (pcd) Weaver Neural transplants - Cerebel- lum - Light microscopy - Electron microscopy Introduction Mutant mice which lose specific neuronal populations pro- vide valuable experimental models in neural grafting re- search (Krieger et al. 1982; Wille et al. 1983; Errington and Bliss 1984; Sotelo and Alvarado-Mallart 1986; Triarhou et al. 1986a, b; Low et al. 1987). Mutant genes may affect Offprint requests to. B. Ghetti, Department of Pathology (Neu- ropathology), Indiana University School of Medicine, 635 Barnhill Dr., Indianapolis, IN 46223, USA * Dedicated to Herbert yon Karajan their target cells either intrinsically (Mullen and Herrup 1979) or through an influence of the local or systemic envi- ronment (Sidman and Pearlstein 1965; Salaiin 1982). Sever- al factors may interfere with the growth and differentiation of tissue implanted into mutant mice, such as absence of trophic factors or presence of noxious diffusible humoral substances which might be produced (1)as an expression. of the mutation, (2)during neuronal degeneration, and (3) secondarily to tissue alterations which follow neuronal losses. Studies on the viability of solid grafts in mutant mice may yield meaningful clues, especially in the perspec- tive of applying neural transplantation techniques to repair both '"global" (Olson and Seiger 1972; Bj6rklund et al. 1976) and "point-to-point" neuronal systems (Sotelo and Alvarado-Mallart 1986) in degenerative conditions charac- terized by selective neuronal losses. Solid cerebellar grafts are capable of surviving and growing following orthotopic (Das and Altman 1972; Bj6rklund et al. 1984) or heterotopic (Wells and McAllister 1982; Alvarado-Mallart and Sotelo 1982; Kromer et al. 1983) implantation into the brain of normal hosts. The fate of solid cerebellar grafts has not been studied in mice, whether normal or mutant. Among cerebellar mutants, 'Purkinje cell degeneration' (pcd) and weaver (gene sym- bol wv) mice are respectively characterized by the selective loss of Purkinje (Mullen et al. 1976) and granule (Sidman 1968) cells. Adult pcd and weaver homozygotes were used in the present study as recipients of embryonic cerebellar implants to investigate the following issues: (a) viability of the grafted neuronal populations within the environment of the mutant host, (b) ability of the grafts to reach a ma- ture organization, and (c)ultrastructural appearance of grafted Purkinje and granule neurons, their processes, and their synaptic interconnections. Materials and methods Animals Colonies of pcd and weaver mutant mice are maintained at Indiana University Medical Center, originating from mice heterozygous for the genes pcd (pcd/+) or wv (wv/+) from the Jackson Laboratory (Bar Harbor, ME). The pcdmutants are on the C57BL/6J genetic background. Weaver mutants are on the B6CBA-AW-J/A hybrid back- ground, which is obtained by mating females from inbred
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Transplantation of cerebellar anlagen to hosts with genetic cerebellocortical atrophy

Anat Embryol (1987) 176:145 154 Anatomy and Embryology �9 Springer-Verlag 1987
Original articles
Transplantation of cerebellar anlagen to hosts with genetic cerebellocortical atrophy Lazaros C. Triarhou 1, 3 , . , Walter C. Low 2, 3, and Bernardino Ghetti l, 3 1 Department of Pathology, Division of Neuropathology, 2 Department of Physiology and Biophysics, and 3 Program in Medical Neurobiology, Indiana University School of Medicine, Indianapolis, Indiana, USA
Summary. Embryonic cerebellar grafts from genetically nor- mal donors were implanted into the cerebellomedullary cistern of adult 'Purkinje cell degeneration' (pcd) and weav- er mutant mice, which are respectively characterized by the selective loss of Purkinje and granule cells. Grafts placed into both mutant recipients exhibited a layered cellular or- ganization reminiscent of the normal cerebellar cortex. Mo- lecular, Purkinje, and granule cell layers were identifiable. Grafted Purkinje cells displayed characteristic cytological features, such as hypolemmal cisterns in association with mitochondria in the perikaryon, and lamellar structures in their axons. The cytological features of granule cell somata in the grafts appeared similar to those of mature granule cells. Electron microscopic examination of the molecular layer of the grafts revealed the presence of parallel fibers, which were not oriented in a parallel fashion; axon termi- nals of such fibers were often presynaptic to dendritic spines. The number of parallel fibers was markedly reduced in grafts implanted into both mutants compared to the nor- mal cerebellar cortex; however, this phenomenon is com- monly seen in cerebellum in tissue culture and in cerebellar transplants into normal hosts. It is concluded, therefore, that the environment of the mutant hosts does not affect the survival of Purkinje or granule cells and that transplan- tation of solid cerebellar grafts in the neurological mutants studied does not seem to pose any apparent limitations beyond those inherent to the process of cerebellar growth and differentiation outside its normal environment.
Key words: Neurological mutant mice 'Purkinje cell de- generation' (pcd) Weaver Neural transplants - Cerebel- lum - Light microscopy - Electron microscopy
Introduction
Mutant mice which lose specific neuronal populations pro- vide valuable experimental models in neural grafting re- search (Krieger et al. 1982; Wille et al. 1983; Errington and Bliss 1984; Sotelo and Alvarado-Mallart 1986; Triarhou et al. 1986a, b; Low et al. 1987). Mutant genes may affect
Offprint requests to. B. Ghetti, Department of Pathology (Neu- ropathology), Indiana University School of Medicine, 635 Barnhill Dr., Indianapolis, IN 46223, USA
* Dedicated to Herbert yon Karajan
their target cells either intrinsically (Mullen and Herrup 1979) or through an influence of the local or systemic envi- ronment (Sidman and Pearlstein 1965; Salaiin 1982). Sever- al factors may interfere with the growth and differentiation of tissue implanted into mutant mice, such as absence of trophic factors or presence of noxious diffusible humoral substances which might be produced (1)as an expression. of the mutation, (2)during neuronal degeneration, and (3) secondarily to tissue alterations which follow neuronal losses. Studies on the viability of solid grafts in mutant mice may yield meaningful clues, especially in the perspec- tive of applying neural transplantation techniques to repair both '"global" (Olson and Seiger 1972; Bj6rklund et al. 1976) and "point- to-point" neuronal systems (Sotelo and Alvarado-Mallart 1986) in degenerative conditions charac- terized by selective neuronal losses.
Solid cerebellar grafts are capable of surviving and growing following orthotopic (Das and Altman 1972; Bj6rklund et al. 1984) or heterotopic (Wells and McAllister 1982; Alvarado-Mallart and Sotelo 1982; Kromer et al. 1983) implantation into the brain of normal hosts. The fate of solid cerebellar grafts has not been studied in mice, whether normal or mutant. Among cerebellar mutants, 'Purkinje cell degeneration' (pcd) and weaver (gene sym- bol wv) mice are respectively characterized by the selective loss of Purkinje (Mullen et al. 1976) and granule (Sidman 1968) cells. Adult pcd and weaver homozygotes were used in the present study as recipients of embryonic cerebellar implants to investigate the following issues: (a) viability of the grafted neuronal populations within the environment of the mutant host, (b) ability of the grafts to reach a ma- ture organization, and (c)ultrastructural appearance of grafted Purkinje and granule neurons, their processes, and their synaptic interconnections.
Materials and methods
Animals
Colonies of pcd and weaver mutant mice are maintained at Indiana University Medical Center, originating from mice heterozygous for the genes pcd (pcd/+) or wv (wv/+) from the Jackson Laboratory (Bar Harbor, ME). The pcdmutants are on the C57BL/6J genetic background. Weaver mutants are on the B6CBA-AW-J/A hybrid back- ground, which is obtained by mating females from inbred

146
strain C57BL/6J-AW-J/A w-J to males from inbred strain CBA/CaGnLe-A/A. Homozygotes are obtained either by crossing pairs of heterozygous parents or by crossing homo- zygous females to heterozygous males. The animals are kept in a 12 h dark-light cycle, and are provided with food pellets and water or wet food ad libitum. For the present study we used 16 pcd and 12 weaver mutants 2-3 months old as tissue recipients. Additional non-grafted normal, pcd and weaver mutant mice were also examined.
Donor tissue
Pairs of normal (+ / + ) C57BL/6J mice (for the pcd experi- ment) and of normal ( + / + ) C57BL/6J-AW-J/A w-J female and normal ( + / + ) CBA/CaGnLe-A/A male mice (for the weaver experiment) were placed in the same cage for a 24 h mating period. Pregnant dams were selected on gestational day 14 15 and anesthetized with equithesin (3.3 ml/kg i.p.). A Caesarean section was performed and the embryos were removed from the uterine horns one at a time. The crown- rump length of the embryos was 11 13 ram. Their brains were dissected out and the cerebellar buds were cut free with the help of fine iridectomy scissors. At this stage of embryonic development the deep nuclear complex has ac- quired a medial position (Goffinet 1983; Altman and Bayer 1985). Thus, segments taken from the dorsolateral part would be expected to contain cerebellar cortical anlage only, while segments that included the medial part of the cerebellar primordium would be expected to contain the deep nuclear complex as well. The grafts were kept intact in lactated Ringer's solution until the time of implantation. Some of the contralateral hemiprimordia were fixed by im- mersion in aldehydes and processed for morphological ex- amination.
Implantation procedure
Recipient animals were anesthetized with ketamine cocktail (1 ml/kg i.p.) and placed into a stereotaxic apparatus (Da- vid Kopf Instruments). The skin of the dorsal neck was cut and the neck muscles over the occipital bone were re- moved. Donor tissue was placed in the cerebellomedullary cistern, between the area postrema and the uvula vermis, and covered with Gelfoam. The overlying skin was sutured and the animals were allowed to recover from the operation.
Solid grafts were placed ectopically rather than within the cerebellar parenchyma of the hosts in order to distin- guish between the cerebella of the host and graft.
Histological procedure
Mutant pcd recipients were perfused at postoperative days 34 (7), and 39 (8); one animal died postoperatively. Weaver hosts were perfused at postoperative days 20 (2), 27 (2), and 50 (8). The animals were heparinized (500 USP units i.p.), anesthetized with pentobarbital sodium (50 rag/ kg i.p.), and perfused transcardially with a mixture of 1% formaldehyde and 1.25% glutaraldehyde in 0.1 M phos- phate buffer (pH 7.4). Brains were dissected out, postfixed, dehydrated in series of graded ethanols, and embedded in paraffin or Epon. Ten-micrometer thick sections from the paraffin blocks were stained with gallocyanin Nissl stain. Grafts from two pcd hosts and from two weaver hosts were embedded in Epon. One-micrometer-thick sections from the
plastic blocks were stained with toluidine blue for light mi- croscopic examination and orientation. Thin sections were stained with uranyl acetate and lead citrate and examined at 80 kV in a Siemens 101 Elmiskop or at 60 kV in a Phi- lips 300 electron microscope.
Morphometric and semiquantitative analysis
Purkinje cells. Cross-sectional areas of Purkinje cell somata in the grafts and in the normal cerebellum were obtained in 10 pm thick paraffin sections, using the Bioquant image analysis software. Only cells with a clearly visible nucleolus were measured, as the presence of nucleolus may be taken as an indication for the representation of the cell at its full diameter. Neuronal diameters were then calculated from cross-sectional areas, assuming that neuronal somata are spherical in shape.
Parallel fibers. Electron micrographs of molecular layer neuropil in normal, pcd and weaver mutant mice, and in transplants placed into pcdand weaver hosts were taken at a primary magnification of x 5600 and printed at a final magnification of x 14000 (13 micrographs in the case of the normal mouse cerebellum, covering 2786 pm 2, and 15 micrographs covering 3214 gm 2 in each of the other cases). One animal was analyzed for each condition. In normal and mutant cerebella, quantitations were carried out in sagittal sections of the declive vermis. Non-overlap- ping microscopic fields were selected randomly through the entire thickness of the molecular layer. In the photographic fields, axonal and dendritic processes were present, and not neuronal perikarya. Parallel fibers were marked on a plastic overlay and counted. Criteria for the identification of paral- lel fibers were diameter of 70-350 nm, presence of at least two microtubules, and absence of spine apparatus.
Results
Mutant host cerebella
Recipient pcd mutant mice displayed a marked atrophy of the cerebellum. The molecular layer was decreased in thick- ness and did not display the characteristic profiles of Pur- kinje cell dendrites (Fig. 1); the density of parallel fibers was increased in comparison with age-matched controls (Table 1), due to the severe atrophy of the molecular layer. In the Purkinje cell layer, clusters of basket axon terminals were empty; occasionally, debris of degenerated Purkinje cell perikarya was recognized. In the granule cell layer there were scattered degenerated granule cells.
The weaver cerebellum was also atrophic. The molecular layer was thin and contained numerous Purkinje cell den- dritic profiles (Fig. 1). Parallel fibers were remarkably re- duced (Table 1), due to the rarity of granule cells. The few surviving granule cells were mostly localized in the molecu- lar rather than in the granule cell layer. Purkinje cells were not aligned.
Embryonic donor tissue
A lamination was recognizable (Fig. 2). The following three layers were seen: the external germinal layer dorsally, which was composed of approximately 7-8 layers of cells; some
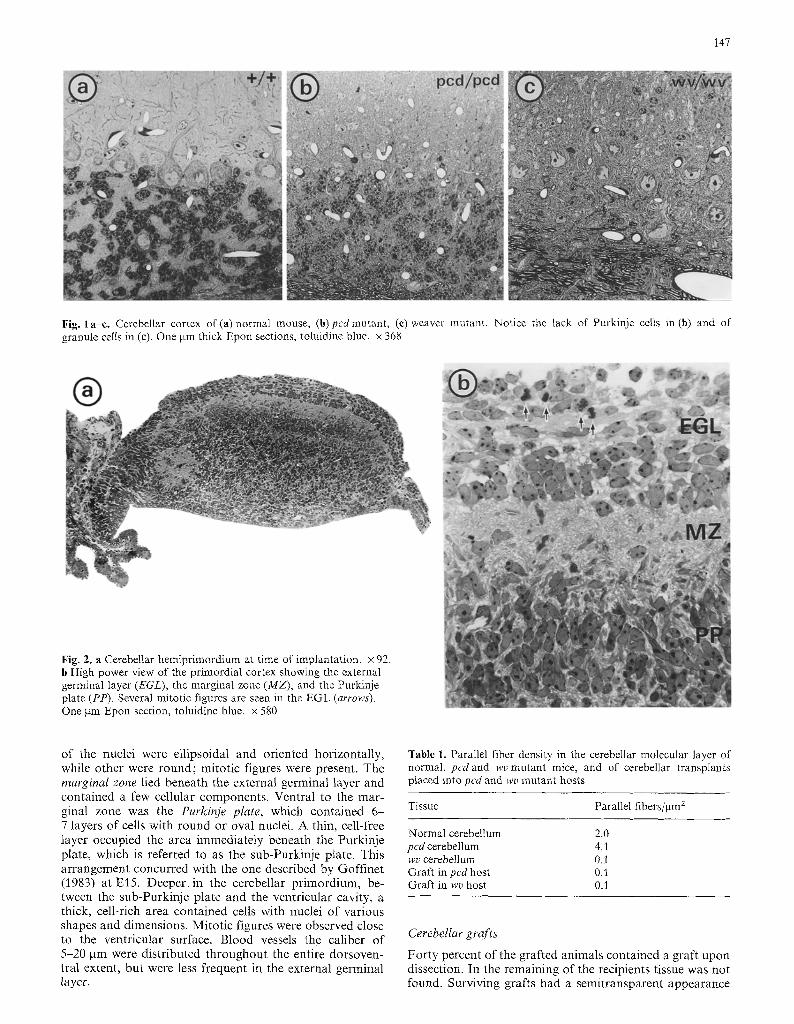
147
Fig. 1 a-c. Cerebellar cortex of (a) normal mouse, (b) pcd mutant, (e) weaver mutant. Notice the lack of Purkinje cells in (b) and of granule cells in (c). One btm thick Epon sections, toluidine blue. • 368
Fig. 2. a Cerebellar hemiprimordium at time of implantation. • 92. b High power view of the primordial cortex showing the external germinal layer (EGL), the marginal zone (MZ), and the Purkinje plate (PP). Several mitotic figures are seen in the EGL (arrows). One gm Epon section, toluidine blue. x 580
of the nuclei were ell ipsoidal and oriented horizontal ly, while other were round; mitot ic figures were present. The marginal zone lied beneath the external germinal layer and contained a few cellular components . Ventral to the mar- ginal zone was the Purkinje plate, which contained 6 - 7 layers of cells with round or oval nuclei. A thin, cell-free layer occupied the area immediately beneath the Purkinje plate, which is referred to as the sub-Purkinje plate. This ar rangement concurred with the one described by Goff inet (1983) at El5 . Deeper. in the cerebellar pr imordium, be- tween the sub-Purkinje plate and the ventricular cavity, a thick, cell-rich area contained cells with nuclei of various shapes and dimensions. Mitot ic figures were observed close to the ventricular surface. Blood vessels the caliber of 5-20 gm were distr ibuted throughout the entire dorsoven- tral extent, but were less frequent in the external germinal layer.
Table 1. Parallel fiber density in the cerebellar molecular layer of normal, pcdand wv mutant mice, and of cerebellar transplants placed into pcd and wv mutant hosts
Tissue Parallel fibers/gm 2
Normal cerebellum 2.0 pcd cerebellum 4.1 wv cerebellum O. I Graft in ped host 0,1 Graft in wv host 0.1
Cerebellar grafts
For ty percent of the grafted animals contained a graft upon dissection. In the remaining of the recipients tissue was not found. Surviving grafts had a semitransparent appearance


149
Fig. 4. Molecular layer of graft in pcdrecipient. Axon terminals are impinging upon dendritic spines (asterisks); d, dendrite. Several spines are unattached, x 19200
and occupied the space between the host cerebellum and the dorsal surface of the brainstem (Fig. 3). Fine, well-per- fused t ransparent b lood vessels connected the grafts with the host brain. These vessels could be clearly seen under the dissecting microscope. Based on measurements in serial paraffin sections stained with gallocyanin, t ransplants placed into pcd mutant hosts were up to 0.6 m m rostrocau- dally, 1.2 mm mediolateral ly, and 1.2 mm dorsoventral ly. Graf ts placed into weaver mutan t hosts were up to 0.7 mm rostrocaudally, 1.0 mm mediolateral ly, and 0.7 mm dorso- ventrally.
On light microscopic examinat ion, grafted cerebella ad- hered to the dorsal surface of the brainstem, beneath the uvula vermis of the host cerebellum (Fig. 3). Graf ts intro- duced into both mutants displayed the characterist ic trila- minar organizat ion of cerebellar cortical tissue. In some of the grafts, a cluster of neurons embedded within the white mat ter apparent ly represented the deep cerebellar nu-
clei. Other grafts were composed entirely of cerebellar cor- tex. Al though the orientat ion of the grafted tissue could not be control led due to the implanta t ion into a cavity with fluid, cerebellar neurons were well organized into sepa- rate molecular, Purkinje, and granule cell layers. The size of the embryonic implant was 1/4 to 1/2 that of the pr imor- dial cerebellum, however, developed grafts were smaller than the size that the same tissue would have reached after normal growth in situ. The thickness of the molecular and granule cell layers was smaller than that in the normal cere- bellum.
Both of the main neuronal cortical populat ions, i.e., Purkinje and granule cells, had survived in grafts implanted to both mutants. Only a small number of granule cells was present in the most superficial part of the molecular layer. Purkinje cell diameters in grafts placed to pcd mutant hosts were 17 _+ 1.7 gm (mean_+ s tandard deviation, n = 100). Pur- kinje cell diameters in grafts placed to weaver mutan t hosts
Fig. 3a-L Developed cerebellar grafts, a Macroscopic view of the graft in situ (asterisk), lying between the host cerebellum (CER) and brainstem (BS). x 10. b Graft (asterisk) placed into a pcd mutant host, 39 days post-transplantation. IC, inferior colliculus; IV, fourth ventricle, x 25. c Higher power view of the same graft; notice the cerebellar lamination, x 63. d Graft (asterisk) placed in weaver mutant host between the cerebellum (CER) and brainstem (BS), shown at 27 days post-transplantation, x 63. e High power view of the pcd host cortex (H) with the Purkinje cells missing, and of the grafted cortex (G) with both Purkinje and granule cells present, x 250. f High power of the weaver host cortex (H) with granule cells missing, and of the grafted cortex (G) with both Purkinje and granule cells. x 250. Micrographs b through f are taken from t0 gm thick paraffin sections stained with gallocyanin Nissl

150
Fig. 5 a - d . Electron mic roscopy of graf ts in pcdhosts, a A baske t axon te rminal (asterisk) is f o r m i n g an axosoma t i c synapse (arrow) with a Purkinje cell (Pc). x 20000. b H y p o l e m m a l cistern associa ted with a m i t o c h o n d r i o n in a graf ted Purkin je cell. x 32000. c A myel ina ted Purkinje a xon con ta in ing the character is t ic lamellar s t ructure , seen in the granule cell layer o f a graft , x 19200. d Gra f t ed ma tu r e g ranule cells. A dendri te (asterisk) is e m a n a t i n g f rom the cell in the center o f the field, x 8700

151
Fig. 6a--e. Electron microscopy of graft placed into a weaver mutant host. a A cluster of mature granule cells, x 7100. b A bundle of parallel fibers in the molecular layer of the graft, x 30400. c Several parallel fiber terminals are forming adjacent junctional synapses with a dendritic profile (d). x 36800

152
were 16_+ 1.4 pm (n= 100). In the cerebellum of one month old normal mice, Purkinje cell diameters were 17_+ 1.6 gm (n= 100). Purkinje cells were aligned in a single cell layer (Fig. 3). Granule cells formed large aggregates in distinct layers. Granule cell diameters in the normal cerebellum, in grafts placed to pcd mutant hosts, and in grafts placed to weaver mutant hosts ranged from 4 6 gin, with a mean of 5 ~tm. In general, grafts placed to the two strains of mutant recipients did not exhibit any apparent differences.
Under the electron microscope, the molecular layer of the grafts contained unmyelinated fibers 0.1-0.2 gm in di- ameter with a few microtubules and mitochondria. In spite of the lack of orientation and the formation of bundles, these processes were reminiscent of parallel fibers. In the same plane of section, they would be oriented longitudin- ally, transversely or obliquely. Some of these fibers ended with a bulbous terminal which contained numerous round synaptic vesicles. Such endings formed junctional synapses with dendritic spines (Fig. 4). In contrast to the parallel fibers normally seen in the cerebellum of the mouse and of most species, parallel fibers in the grafts were quite sparse and did not form closely packed arrays. The sparsity of parallel fibers was a consistent observation in the grafts. To give a numerical dimension to this observation, a semi- quantitative study of parallel fiber profiles was carried out in the molecular layer of normal, pcd and weaver mutants, and in cerebellar transplants placed into pcd and weaver mutant hosts, which showed a remarkable decrease in the number and density of these profiles in the grafts as com- pared to the normal and pcd cerebellum, and no apparent difference from the density found in the molecular layer of the weaver cerebellum (Table 1). Dendritic shafts were present in the molecular layer of the grafts. They were rich in microtubules, small cisterns of endoplasmic reticulum, and mitochondria. Protrusions in the form of dendritic spines frequently emanated from dendrites. As already men- tioned, such spines were often postsynaptic to parallel fiber endings. Some of the spines were unattached and were enve- loped by electron-lucent astroglial processes (Fig. 4). As ex- pected, climbing fibers were absent. Interneurons of the molecular layer measured 6-12 p.m in diameter and con- tained a large nucleus surrounded by a moderate amount of cytoplasm.
Purkinje cells displayed a relatively large nucleus with the chromatin evenly dispersed throughout the nucleo- plasm. The cytoplasm was abundant and contained a wealth of cytoplasmic organelles, including free ribosomes, rough endoplasmic reticulum, Golgi apparatus, mitochon- dria, microtubules and intermediate filaments. The charac- teristic hypolemmal cisterns (Herndon 1963) were closely associated with mitochondria (Fig. 5). Occasionally, Pur- kinje cells possessed stubby somatic thorns. The polyriboso- mal accumulations which characteristically persist in the Purkinje cells ofpcd mutant mice beyond the 15th postnatal day (Landis and Mullen 1978) were absent from grafted Purkinje cells. Lamellar structures consisting of orderly ar- ranged tubular elements were contained within large den- drites of the molecular layer, within axons in the Purkinje cell layer, and in myelinated axons in the granule cell layer and in the deep nuclear complex (Fig. 5). In the normal cerebellum, such arrays are usually associated with Purkinje cell axons (Uchizono 1969; Mugnaini 1972). Occasionally, Purkinje cell somata were surrounded by glial processes. Axosomatic synapses were formed on Purkinje cells. In
some cases the presynaptic elements contained an abun- dance of synaptic vesicles and intermediate filaments and were interpreted as basket axon terminals (Fig. 5).
Grafted granule cells appeared similar to those in the normal cerebellum (Figs. 5, 6). The nucleus occupied most of the cell body and was surrounded by a thin rim of cyto- plasm. The chromatin was aggregated in large clusters, which were usually located beneath the nuclear envelope. Granule cell perikarya were tightly apposed. In the granule cell layer of the transplants glomerular arrangements con- sisted of mossy fiber-like terminals and small postsynaptic dendritic profiles. Mossy fiber-like terminals contained den- sely packed synaptic vesicles and plentiful mitochondria; their shape was roundish or oval with regular or irregular contours. In grafts placed to both pcd and weaver mutant hosts, granule cell somata had the appearance of normal mature granule cells (Fig. 6). Several bulbous endings of parallel fibers formed almost continuous junctional syn- apses on the same dendritic profile in the molecular layer (Fig. 6).
In some of the preparations, clusters of large neurons were embedded in the white matter at some distance from the granule cell layer. Based on their location, large size, relative paucity of cisterns in the stacks of rough endoplas- mic reficulum, and lack of hypolemmal cisterns in associa- tion with mitochondria, these cells were tentatively identi- fied as deep nuclear neurons. The soma and the initial seg- ment of dendrites of such neurons were synaptically in- vested by axon terminals.
Blood vessels were distributed throughout the trans- planted tissue and endothelial cells lined the walls of the lumen. Astrocytes were distributed throughout all layers of the grafts and were sometimes associated with blood capillaries. Otigodendrocytes were associated with myelin- ated axons of the white matter. Occasionally, macrophages were engaged in the removal of degenerating debris.
Discussion
The findings of the present study show that cerebellar grafts from E15 normal embryos are able to survive and form a structure reminiscent of the cerebellum following implan- tation into adult hosts with inherited Purkinje or granule cell degeneration. Grafts acquire a trilaminar cerebellocorti- cal organization and contain mature forms of both Purkinje and granule cell perikarya. Parallel fibers, however, are markedly reduced in number.
Two groups of parameters which may influence graft survival and growth are discussed. These are related to the mutant host environment and to the separation of the em- bryonic cerebellum from its anatomical context.
The survival of granule cells in grafts implanted into weaver hosts is of interest in view of the hypotheses con- cerning the mechanisms of action of the weaver mutation (Rakic and Sidman 1973c; Sotelo 1980; Goldowitz and Mullen 1982). One view considers the possibility of an effect of the mutation mediated through some toxic substance (Sotelo 1980). This idea is supported by results showing that the viability of granule cells from weaver cerebellum in vitro may be altered by the chemical manipulation of the culture medium (Messer and Smith 1977; Trenkner et al. 1978). The presence of a considerable number of ma- ture granule cell somata forming an internal granular layer in the grafts does not support the view of the action of

153
a putative systemic substance, toxic to these cells in adult weaver mutants.
The pcd mutation is known to act intrinsically within the Purkinje cells (Mullen 1977) and no humoral factors have been associated with the action of the gene. On the other hand, following Purkinje cell loss, astrocytes display several structural and biochemical changes, secondary to the degeneration of these neurons. Glial alterations are ex- pressed as elevation in G m ganglioside (Seyfried et al. 1982; LeVine et al. 1986), decrease in sn-glycerol-3-phosphate de- hydrogenase (Fisher 1984), increase of a nerve growth fac- tor-like protein (Schwartz et al. 1982), and exaggerated cyclic AMP accumulation in vitro (Ghetti et al. 1981). It is not known whether these changes are associated with a modification of the extracellular compartment. In any event, putative humoral changes do not seem to adversely affect the survival of Purkinje cells either when solid grafts are implanted into pcd hosts (present study) or when Pur- kinje cells from cell suspensions are implanted directly into the cerebellar parenchyma ofpcd mutant mice (Sotelo and Alvarado-Mallart 1986).
The 40% rate of graft survival in our study is compara- ble to that found in the case of implantation of solid grafts into normal hosts (50%, Alvarado-Mallart and Sotelo 1982; 45%, Bj6rklund et al. 1984). However, the cerebellar grafts appear rudimentary in that they display a deficient arborization of Purkinje cell dendrites and a paucity of parallel fibers. It is unlikely that these deficits can be attrib- uted to the environment of the mutant host; in fact, defects of this type have previously been reported in grafts im- planted into normal recipients (Alvarado-Mallart and So- telo 1982) and also in cerebellum grown in tissue culture (for review see Seil 1979). The findings of deficient Purkinje cell dendritic arborization and of parallel fiber paucity, which extend previous observations, deserve some com- ments.
The density of parallel fibers was reduced by 96% in the grafts, but small bundles were occasionally observed. The weaver cerebellum is agranular, and therefore, a pau- city of parallel fibers is expected (cf. Table 1); on the other hand, while granule cell somata were present in the trans- plants, there was an extremely low number of parallel fibers. Paucity of parallel fibers is commonly found in tissue cul- tures of mouse cerebellum (Herndon et al. 1981), and in cerebellar transplants implanted in the normal rat (Alvar- ado-Mallart and Sotelo 1982) and in the normal mouse (unpublished results). The paucity of parallel fibers could be due to a loss of granule cell precursors or to an abnor- mality in length and orientation of granule cell axons. Neu- roblasts of the external germinal layer are particularly vul- nerable when removed from their original environment; this has clearly been shown in tissue culture, where a loss of many germinal cells takes place during cerebellar growth (Allerand 1971), leading to a reduction in the number of granule cells. Since these neurons originate postnatally (Miale and Sidman 1961), obtaining the graft at an early stage before granule cell neurogenesis might cause their ger- minal cells to degenerate. Furthermore, the exclusion of extracerebellar mossy fibers from the new environment of the granule cell neuroblasts may deprive the latter of the recognition molecules that may regulate neuronal migration and development. Alternatively, the paucity of parallel fibers in the grafts could relate to an abnormality of granule cell axons either at the horizontal stage, by not growing
both polar processes, or at the migratory and multipolar stages, by retracting one or both of the ah-eady bifurcated branches. These stages in the development of parallel fibers have been described in detail by Ramdn y Cajal (1952) and Larramendi (1969).
In turn, the lack of a meaningful number of parallel fibers may affect those features of Purkinje cell dendritic arborization which are controlled by extrinsic factors through interaction with neighbouring cells (Rakic 1975). According to Rakic (1975), " a given neuron may be consid- ered partly as an autonomous active individual unit, and partly as a passive element subordinate to forces that lie outside its boundary". Studies after experimentally-induced (for review see Altman 1982) or genetically-determined (Rakic and Sidman 1973 a, b) agranularity strongly indicate that the initiation and growth of Purkinje cell dendrites and the development of their spines appear to proceed with little influence from parallel fiber input. The site and the final detailed orientation of their dendritic arbor, however, does depend on the parallel fibers (Rakic 1975).
In all, it seems that embryonic tissue explanted from its normal environment and placed into adult mutant hosts may form an elementary cerebellum, similarly to that found in normal hosts. However, in both cases, some of the crucial elements required for normal physiological activity may be deficient. Thus, transplantation in pcd and weaver mutants does not seem to pose any apparent problems beyond those inherent to the process of growth and differentiation of solid cerebellar grafts outside their anatomical context.
Acknowledgements. The authors wish to thank Dr. Shirley A. Bayer of the Laboratory of Developmental Neurobiology, Department of Biology, I.U.P.U.I., for valuable discussion, and Dr. Carleton D. Nordschow of the Department of Pathology and Dr. Marguer- ite Hendrie of the Laboratory Animal Resource Center for support. The skilful technical help of Ms. Constance J. Alyea and Ms. Rose- marie Funkhouser of the Department of Pathology and the photo- graphic assistance of Mr. Joseph Demma of the Department of Medical Illustrations are gratefully acknowledged. This work was supported in part by PHS grants RO1-NS14426 (B.G.), SO7- RR5371 and RO3-AG5575 (W.C.L.), and by grant 85-174 from Eli Lilly and Co. Dr. Triarhou is Postdoctoral Fellow supported by grant RO1-NS14426.
References
Allerand CD (1971) Patterns of neuronal differentiation in develop- ing cultures of neonatal mouse cerebellum : A living and silver impregnation study. J Comp Neurol 142:167-204
Altman J (1982) Morphological development of the rat cerebellum and some of its mechanisms. Exp Brain Res [Suppl] 6:8-49
Altman J, Bayer SA (1985) Embryonic development of the rat cerebellum. III. Regional differences in the time of origin, mi- gration, and settling of Purkinje cells. J Comp Neurol 231:42-65
Alvarado-Mallart RM, Sotelo C (1982) Differentiation of cerebel- lar anlage heterotopically transplanted to adult rat brain: a light and electron microscopic study. J Comp Neurol 212: 247-267
Bj6rklund A, Stenevi U, Svendgaard N-A (1976) Growth of trans- planted monoaminergic neurones into the adult hippocampus along the perforant path. Nature (Lond) 262:787-790
Bj6rklund H, Bickford P, Dahl D, Hoffer B, Olson L (1984) Intra- cranial cerebellar grafts: Intermediate filament immunohisto- chemistry and electrophysiology. Exp Brain Res 55 : 372-385
Das GD, Altman J (1972) Studies on the transplantation of devel- oping neural tissue in the mammalian brain. I. Transplantation

154
of cerebellar slabs into the cerebellum of neonate rats. Brain Res 38 : 233-249
Errington ML, Bliss TVP (1984) Hippocampal transplants in nor- mal and reeler mice. Neurosci Lett 45:291-296
Fisher M (1984) Neuronal influence on glial enzyme expression: Evidence from mutant mouse cerebella. Proc Natl Acad Sci USA 81:4414-4418
Ghetti B, Truex L, Sawyer B, Strada S, Schmidt MJ (1981) Exag- gerated cyclic AMP accumulation and glial cell reaction in the cerebellum during Purkinje cell degeneration in pcdmutant mice. J Neurosci Res 6:789-801
Goffinet AM (1983) The embryonic development of the cerebellum in normal and reeler mutant mice. Anat Embryol 168:73-86
Goldowitz D, Mullen RJ (1982) Granule cell as a site of gene action in the weaver mouse cerebellum: Evidence from hetero- zygous mutant chimeras. J Neurosci 2:1474-1485
Herndon RM (1963) The fine structure of the Purkinje cell. J Cell Biol 18:167-180
Herndon RM, Seil FJ, Seidman C (1981) Synaptogenesis in mouse cerebellum: A comparative in vivo and tissue culture study. Neuroscience 6 : 2587-2598
Krieger DT, Perlow M J, Gibson M J, Davies TF, Zimmerman EA, Ferin M, Charlton HM (1982) Brain grafts reverse hypogonad- ism of gonadotropin-releasing hormone deficiency. Nature (Lond) 298 : 1 3
Kromer LF, Bj6rklund A, Stenevi U (1983) Intraeephalic embryon- ic neural implants in the adult rat brain. I. Growth and mature organization of brainstem, cerebellar, and hippocampal im- plants. J Comp Neurol 218:433-459
Landis SC, Mullen RJ (1978) The development and degeneration of Purkinje cells in pcdmutant mice. J Comp Neurol 177:125-144
Larramendi LMH (1969) Analysis of synaptogenesis in the cerebel- lum of the mouse. In: Llinfis R (ed) Neurobiology of cerebellar evolution and development. A M A / E R F Institute for Biomedi- cal Research, Chicago, pp 803 843
LeVine SM, Seyfried TN, Yu RK, Goldman JE (1986) Immunocy- tochemical localization of Gm ganglioside to astrocytes in mur- ine cerebellar mutants. Brain Res 374:260-269
Low WC, Triarhou LC, Ghetti B (1987) Cerebellar transplants into mutant mice with Purkinje and granule cell degeneration. In: Azmitia EC, Bj6rklund A (eds) Cell and tissue transplanta- tion into the adult brain. The New York Academy of Sciences, New York (in press)
Messer A, Smith DM (1977) In vitro behavior of granule cells from staggerer and weaver mutants of mice. Brain Res 130:13 23
Miale IL, Sidman RL (1961) An autoradiographic analysis of histogenesis in the mouse cerebellum. Exp Neurol 4:277-296
Mugnaini E (1972) The histology and cytology of the cerebellar cortex. In: Larsell O, Jansen J (eds) The comparative anatomy and histology of the cerebellum : the human cerebellum, cerebel- lar connections, and cerebellar cortex. The University of Min- nesota Press, Minneapolis, pp 201-264
Mullen RJ (1977) Site ofpcd gene action and Purkinje cell mosaic- ism in cerebella of chimaeric mice. Nature (Lond) 270: 245-247
Mullen RJ, Eicher EM, Sidman RL (1976) Purkinje cell degenera- tion, a new neurological mutation in the mouse. Proc Natl Acad Sci USA 73:208-212
Mullen RJ, Herrup K (1979) Chimeric analysis of mouse cerebellar mutants. In: Breakfield XO (ed) Neurogenetics: Genetic ap- proaches to the nervous system. Elsevier/North-Holland, Am- sterdam, pp 173-196
Olson L, Seiger A (1972) Brain tissue transplanted to the anterior chamber of the eye. 1. Fluorescence histochemistry of immature
catecholamine and 5-hydroxytryptamine neurons reinnervating the rat iris. Z Zellforsch 135:175 194
Rakic P (1975) Role of cell interaction in development of dendritic patterns. Adv Neurol 12:117-134
Rakic P, Sidman RL (1973a) Sequence of developmental abnor- malities leading to granule cell deficit in cerebellar cortex of weaver mutant mice. J Comp Neurol 152:103-132
Rakic P, Sidman RL (1973b) Organization of cerebellar cortex secondary to deficit of granule cells in weaver mutant mice. J Comp Neurol 152:133-162
Rakic P, Sidman RL (1973c) Weaver mutant mouse cerebellum: Defective neuronal migration secondary to abnormality of Bergmann glia. Proc Natl Acad Sci USA 70 : 240-244
Ram6nyCa ja l S (1952) Histologie du syst6me nerveux de 1' homme et des vert6br6s. Consejo Superior de Investigaciones Cientificas, Instituto Ram6n y Cajal, Madrid
Sala/in J (1982) Differentiation of the optic cups from an anoph- thalmic murine strain, in culture and in intrafoetal grafts. J Embryol Exp Morphol 67:71-80
Schwartz JC, Ghetti B, Truex L, Schmidt MJ (1982) Increase of a nerve growth factor-like protein in the cerebellum ofpcd mu- tant mice. J Neurosci Res 8:205-211
Seil FJ (1979) Cerebellum in tissue culture. Rev Neurosci 4:105-177
Seyfried TN, Yu RK, Miyazawa N (1982) Differential cellular en- richment of gangliosides in the mouse cerebellum: analysis us- ing neurological mutants. J Neurochem 38:551-559
Sidman RL (1968) Development of interneuronal connections in brains of mutant mice. In: Carlson FD (ed) Physiological and biochemical aspects of nervous integration. Prentice Hall, Eng- lewood Cliffs, NJ, pp 163 193
Sidman RL, Pearlstein R (1965) Pink-eyed dilution (p) gene in ro- dents: increased pigmentation in tissue culture. Dev Biol 12:93-116
Sotelo C (1980) Mutant mice and the formation of cerebellar cir- cuitry. Trends Neurosci 3 : 33-36
Sotelo C, Alvarado-Mallart RM (1986) Growth and differentiation of cerebellar suspensions transplanted into the adult cerebellum of mice with heredodegenerative ataxia. Proc Natl Acad Sci USA 83:1135-1139
Trenkner E, Hatten ME, Sidman RL (1978) Effect of ether-soluble serum components in vitro on the behavior of immature cere- bellar cells in weaver mutant mice. Neuroscience 3:1093-1100
Triarhou LC, Ghetti B, Low WC (1986a) Purkinje and granule cells survive in cerebellar grafts implanted into hosts with genetically-determined Purkinje or granule cell degeneration. Ann Neurol 20 : 138
Triarhou LC, Low WC, Ghetti B (1986b) Transplantation of ven- tral mesencephalic anlagen to hosts with genetic nigrostriatal dopamine deficiency. Proc Nat1 Acad Sci USA 83 : 8789-8793
Uchizono K (1969) Synaptic organization of the mammalian cere- bellum. In: Llin/~s R (ed) Neurobiology of cerebellar evolution and development. AMA/ERF Institute of Biomedical Re- search, Chicago, pp 549-581
Wells J, McAllister JP (1982) The development of cerebellar pri- mordia transplanted to the neocortex of the rat. Dev Brain Res 4:167-179
Wille W, Goldowitz D, Seiger A, Olson L (1983) The neurological mutation staggerer is expressed in embryonic cerebellar trans- plants matured in the anterior eye chamber of normal mice. Neurosci Lett 42:1-6
Accepted January 16, 1987




















