
Transmission characteristics for the 1:1 linkage between slowly adapting type II fibers and their...
9
Exp Brain Res (1995) 105:67-75 Springer-Verlag 1995 B. D. Gynther - R. M. Vickery M. J. Rowe Transmission characteristics for the 1:1 linkage between slowly adapting type II fibers and their cuneate target neurons in cat Received: 2 November 1994 / Accepted: 23 February 1995 Abstract Transmission from single, identified, slowly adapting type II (SAIl) tactile fibers to their target neu- rons in the cuneate nucleus was examined in anesthe- tized cats. Simultaneous recordings were made from cu- neate neurons and from fine, intact fascicles of the super- ficial radial nerve in which it was possible to identify and monitor the activity of each group II fiber. Selective activation of individual SAIl fibers was achieved by means of skin stimulation with fine probes, in conjunc- tion with extensive forelimb denervation. Responses were studied for seven SAII-driven cuneate neurons. For three there was unequivocal monitoring of the identified SAII input fiber. However, in six of the seven there was evidence that just one SAIl fiber provided suprathreshold input to the cuneate neuron, and neither temporal nor spatial summation was required for reliable transmission. Cuneate impulse rates, in response to SAIl inputs lasting 1 s, were less than 250 impulses per second, even though the SAIl impulse rates could be 500 s-1. Responses to in- dividual SAII impulses consisted of a burst of 2-3 im- pulses at low SAIl input rates, but burst responses disap- peared at high SAII rates. In all three SAII-cuneate pairs studied, the transmission security (the percentage of SAII impulses that evoked cuneate spike output) exceed- ed 80% in response to static skin displacement and in re- sponse to certain frequencies of skin vibration, in partic- ular, at 100-200 Hz, exceeded 98% when the SAII fiber responded near the 1:1 level (one impulse per vibration cycle). Transmission characteristics for the SAII-cuneate linkage resulted in the cuneate neuron showing tight phaselocking of responses to high-frequency (>100 Hz) vibrotactile stimuli and higher impulse rates than its SAII input (up to input rates of -50 impulses s 1). Secu- rity of transmission across the SAII-cuneate synapse is similar to that demonstrated previously for tactile fibers of the SAI and Pacinian corpuscle (PC)-related classes, B. D. Gynther R. M. Vickery. M. J. Rowe ( ~ ) School of Physiology and Pharmacology, University of New South Wales, Sydney, NSW 2052, Australia; Fax no.: 61-2-385-1059 which suggests that there is no marked differential spe- cialization in transmission characteristics for dorsal col- umn nuclei neurons that receive input from different tac- tile fiber classes. Furthermore, it means that the reported failure of individual SAIl fiber inputs to generate con- scious sensation in man following intraneura] microstim- ulation is not related to transmission failure at the first central relay. Key words Slowly adapting type II Cuneate neuron Synaptic transmission Tactile afferent fiber - Somatosensory system Cat Introduction The slowly adapting type II (SAII) afferent fibers in cat hairy skin each innervate only a single Ruffini ending and therefore have a mechanosensitive receptive field with a single focus of peak sensitivity unrelated to the specialized touch domes that characterize the receptive fields of the SAI afferent fibers (Chambers et al. 1972). Extracellular recording from lemniscal axons or cuneate neurons reveals certain slowly adapting neurons that also have receptive fields with a single punctate focus and re- sponse properties consistent with input from SAII fibers (Brown et al. 1974; Golovchinsky 1980). In view of the sparse distribution of SAIl fibers in the hairy skin (Bur- gess eta1. 1968), it is probable that only single SAII fi- bers provide suprathreshold input to these cuneate neu- rons. Evidence has been deduced from mathematical models by Amassian and Giblin (1974) that single im- pulses of possible SAII origin may be capable of high- transmission security within the cuneate nucleus. How- ever, this has not yet been demonstrated directly for clearly identified SAIl fibers as has been done in our ear- lier studies for single afferent fibers associated with Pa- cinian corpuscles (PC) and for single SAI afferents (Ferrington et al. 1986, 1987a,b; Vickery et al. 1994). As intraneural microstimulation of single SAIl fibers in con- scious human subjects failed in almost all cases to evoke
Transcript of Transmission characteristics for the 1:1 linkage between slowly adapting type II fibers and their...

Exp Brain Res (1995) 105:67-75 �9 Springer-Verlag 1995
B. D. Gynther - R. M. Vickery �9 M. J. Rowe
Transmission characteristics for the 1:1 linkage between slowly adapting type II fibers and their cuneate target neurons in cat
Received: 2 November 1994 / Accepted: 23 February 1995
Abstract Transmission from single, identified, slowly adapting type II (SAIl) tactile fibers to their target neu- rons in the cuneate nucleus was examined in anesthe- tized cats. Simultaneous recordings were made from cu- neate neurons and from fine, intact fascicles of the super- ficial radial nerve in which it was possible to identify and monitor the activity of each group II fiber. Selective activation of individual SAIl fibers was achieved by means of skin stimulation with fine probes, in conjunc- tion with extensive forelimb denervation. Responses were studied for seven SAII-driven cuneate neurons. For three there was unequivocal monitoring of the identified SAII input fiber. However, in six of the seven there was evidence that just one SAIl fiber provided suprathreshold input to the cuneate neuron, and neither temporal nor spatial summation was required for reliable transmission. Cuneate impulse rates, in response to SAIl inputs lasting 1 s, were less than 250 impulses per second, even though the SAIl impulse rates could be 500 s -1. Responses to in- dividual SAII impulses consisted of a burst of 2-3 im- pulses at low SAIl input rates, but burst responses disap- peared at high SAII rates. In all three SAII-cuneate pairs studied, the transmission security (the percentage of SAII impulses that evoked cuneate spike output) exceed- ed 80% in response to static skin displacement and in re- sponse to certain frequencies of skin vibration, in partic- ular, at 100-200 Hz, exceeded 98% when the SAII fiber responded near the 1:1 level (one impulse per vibration cycle). Transmission characteristics for the SAII-cuneate linkage resulted in the cuneate neuron showing tight phaselocking of responses to high-frequency (>100 Hz) vibrotactile stimuli and higher impulse rates than its SAII input (up to input rates of -50 impulses s 1). Secu- rity of transmission across the SAII-cuneate synapse is similar to that demonstrated previously for tactile fibers of the SAI and Pacinian corpuscle (PC)-related classes,
B. D. Gynther �9 R. M. Vickery. M. J. Rowe (~) School of Physiology and Pharmacology, University of New South Wales, Sydney, NSW 2052, Australia; Fax no.: 61-2-385-1059
which suggests that there is no marked differential spe- cialization in transmission characteristics for dorsal col- umn nuclei neurons that receive input from different tac- tile fiber classes. Furthermore, it means that the reported failure of individual SAIl fiber inputs to generate con- scious sensation in man following intraneura] microstim- ulation is not related to transmission failure at the first central relay.
Key words Slowly adapting type II �9 Cuneate neuron �9 Synaptic transmission �9 Tactile afferent fiber - Somatosensory system �9 Cat
Introduction
The slowly adapting type II (SAII) afferent fibers in cat hairy skin each innervate only a single Ruffini ending and therefore have a mechanosensitive receptive field with a single focus of peak sensitivity unrelated to the specialized touch domes that characterize the receptive fields of the SAI afferent fibers (Chambers et al. 1972). Extracellular recording from lemniscal axons or cuneate neurons reveals certain slowly adapting neurons that also have receptive fields with a single punctate focus and re- sponse properties consistent with input from SAII fibers (Brown et al. 1974; Golovchinsky 1980). In view of the sparse distribution of SAIl fibers in the hairy skin (Bur- gess eta1. 1968), it is probable that only single SAII fi- bers provide suprathreshold input to these cuneate neu- rons. Evidence has been deduced from mathematical models by Amassian and Giblin (1974) that single im- pulses of possible SAII origin may be capable of high- transmission security within the cuneate nucleus. How- ever, this has not yet been demonstrated directly for clearly identified SAIl fibers as has been done in our ear- lier studies for single afferent fibers associated with Pa- cinian corpuscles (PC) and for single SAI afferents (Ferrington et al. 1986, 1987a,b; Vickery et al. 1994). As intraneural microstimulation of single SAIl fibers in con- scious human subjects failed in almost all cases to evoke

68
a sensa t ion (Ochoa and TorebjOrk 1983; Val lbo et al. 1984; Mace f i e ld et al. 1990), this may imp ly that S A I l in fo rmat ion is not re l i ab ly t ransmi t ted to h igher levels o f the soma tosenso ry pa thways .
The b road a im of this s tudy was to charac te r ize syn- apt ic t r ansmiss ion f rom s ingle SAI I f ibers to their target neurons in the cuneate nucleus. Spec i f ic a ims o f the s tudy were to examine the t r ansmiss ion character is t ics for the synapt ic l inkage be tween s ingle SAI I f ibers and s ingle cuneate neurons in order to de termine: first, whe ther the input f rom jus t one S A I I f iber is suff icient to act ivate cuneate neurons and, therefore, reveal whether the t r ansmiss ion o f tact i le in fo rmat ion f rom S A I I f ibers can tmke p lace wi thout spat ia l summat ion ; second, whe ther any t empora l summat ion is necessa ry for rel i- ab le t r ansmiss ion o f s ingle S A I I f iber impulses to the cu- nea te neuron; third, whether in format ion about v ibro tac- t i le f requency in the p h a s e l o c k e d impulse pat terns o f the SAI I f ibers can be re ta ined in the course o f t r ansmiss ion through the cuneate nucleus. A p re l imina ry repor t of the da ta has been presen ted in confe rence p roceed ings (Gynthe r et al. 1992b).
Materials and methods
Results came from studies in 24 anesthetized cats used in a con- current study on SAI inputs to the cuneate nucleus (Vickery et al. 1994). Anesthesia was induced and maintained with pentobarbital sodium (40 mg kg -1 i.p.; <2 mg kg -1 la 1 i.v.). Muscle relaxants (gallamine triethiodide, 4 mg kg -1) and artificial ventilation (end- tidal PCO2, 4.0_+0.5%) were employed to improve recording sta- bility in some animals, and a bilateral pneumothorax was occa- sionally performed. In these cases, no effects of the gallamine on neural responsiveness were noted, although no systematic observa- tions were made on this point. In these animals, the depth of anes- thesia was monitored by allowing intermittent recovery from the muscle relaxant and by monitoring blood pressure and pupillary diameter.
Separate fascicles of the lateral branch of the superficial radial nerve were dissected free in continuity to permit recording by means of a platinum hook electrode. Individual slowly adapting fi- bers that innervated the hairy skin on the dorsal side of the fore- arm were identified as belonging to the SAII class and distin- guished from those of the SAI class according to criteria outlined by Burgess et al. (1968), Chambers et al. (1972), and Gynther et al. (1992a). The SAII fibers displayed slowly adapting responses to static indentation of the skin with fine probes (0.25-2 mm di- ameter). They had a single punctate receptive field (based on use of a 0.2-g von Frey hair) of less than 2 mm diameter that was not associated with a touch dome. Furthermore, the SAIl fibers dis- played a vigorous response to skin stretch along one axis, and no response or a reduction of spontaneous activity with stretch along the orthogonal axis. In order to avoid any risk of inputs arising from sources other than the fascicle under study, the ulnar, medi- an, median cutaneous, and musculocutaneous nerves in the fore- arm were all sectioned.
Extracellular recordings were made with tungsten microelec- trodes from single cuneate neurons that received suprathreshold input from individual SAII fibers. The cuneate neurons were all located caudal to the obex. Steady indentation or trains of sinusoi- dal vibration were applied to the skin at the point of maximum sensitivity in the receptive field of the SAII fiber. The feedback- controlled mechanical stimulator was adjusted so that the fiat- tipped probe (0.25, 1, or 2 mm diameter) was perpendicular to the skin but fractionally above it (<100 gin). Step indentations of am- plitude 100-1000 ~tm and duration 1.5 s were repeated every
10-15 s. Sinusoidal vibration (trains of 1 s duration) could be su- perimposed upon the step indentation (usually 100 gm), com- mencing 300 ms after the onset of the step. Data were stored on magnetic tape and in a laboratory computer.
Impulse rates (in impulses per second) were obtained in re- sponse to the 1- to 1.5-s mechanical stimuli and the security of the SAII-cuneate linkage was quantified as the transmission security, by determining the percentage of SAII fiber impulses that generat- ed a response (one or more impulses) in the cuneate neuron. In ad- dition, the extent of phaselocking in responses to vibratory stimuli was examined by constructing cycle histograms, which use a pulse associated with the onset of each successive cycle in a vibration train as a stimulus marker and display the incidence of impulse oc- currences at different times throughout the period of the vibration cycle (Ferrington and Rowe 1980; Ferrington et al. 1987a,b; Vic- kery et al. 1992). Quantitative measures of the tightness of phase- locking in the responses of the SAII fiber and the cuneate neuron to vibration were obtained by calculating an index of dispersion, the minimum standard deviation (SD) in milliseconds, for the dis- tribution of impulse activity within the cycle histogram (Mount- castle et al. 1969; Bennett et al. 1980; Ferrington and Rowe 1980; Greenstein et al. 1987).
Latency histograms (LH) were constructed from the impulse activity in the cuneate neuron in response to impulses in the SAII fiber by treating successively each SAII impulse as time zero (Vic- kery et al. 1994). The LH, like a cross-correlogram, reveals the precise association between individual SAII impulses and the im- pulses generated in the cuneate neuron. However, the LH excludes effects of SAII impulses other than the current time-zero impulse by setting a time window (usually 5 ms) in the response of the cu- neate neuron at a fixed delay, corresponding to the minimum la- tency, after each SAIl impulse. Those SAII impulses at very high instantaneous frequencies (e.g., more than 200 Hz), usually asso- ciated with a burst at the onset of a response, could be excluded from the LH (for further details of the LH see Vickery et al. 1994).
Results
Cunea te neurons with SAI I input: loca t ion and nature of recept ive f ields
Seven cuneate neurons that received input f rom SAII fi- bers were identif ied, six of which had recept ive f ields made up of a s ingle focus o f sensit ivity, s imilar to the re- cept ive f ields of S A I l fibers descr ibed by Gynther et al. (1992a). The remain ing neuron was driven f rom two foci, s t rongly f rom one, and weak ly f rom the other, but both appeared to be re la ted to SAI I fibers. None of these seven neurons showed convergent input f rom other classes o f tact i le fibers, and none d i sp layed evidence o f an inhibi to- ry recept ive field, a l though the extensive deafferenta t ion o f the fore l imb m a y have r emoved some inputs. A l though the sample o f SAII - re l a t ed neurons is small , it should be emphas ized that within the dorsa l co lumn nuclei or at h igher levels o f the pa thway the inc idence o f this class o f neuron appears to be very low. For example , in a sample of 165 lemnisca l axons sampled by Brown et al. (1974), unequivocal ev idence for SAI I input was repor ted for on- ly 3 neurons out of 52 whose recept ive f ields were exam- ined in detail . Fur thermore , within the cuneate nucleus it- s e l l Go lovch insky (1980) found only about 6% of neu- rons with evidence of input f rom hairy skin SAI I fibers.
Re l iab le moni to r ing o f the per iphera l f iber dr iv ing the cuneate neuron was achieved for three of the seven SAII - re la ted neurons. Each SAI I f iber had a s ingle- focus re-

69
I 5 0 m s
.... r ' r filmlfl[17 ] i'[IIL'I]IIIr vr]TrlrrlrTllr 17FTTrIII'[ ' r r r , r I" r ] r r ..... ".-.,.,
2 ........... L/iliiJlllll.I;JJl,JIIll.ll./t,,Ll..ll,l.l.JJtlJ.L.I lllll_i..l,I.],.i,, I _ ", . . . . "r"lllirlllJl II['HllllFl'l l"lTirrrll[[l r l l I"I I l l"[ II"T ? . . . . " - . i
% I "-' ) {
Fig. 1 Secure synaptic linkage between a slowly adapting type II (SAII) peripheral fiber (1 ~ downwards impulses) and a cuneate neuron (2 o). Simultaneously recorded impulse records (700 ms du- ration) are shown with a schematic of the recording sites. The top trace is a replica of the 300-gin-step indentation stimulus. The la- tency histogram (LH) was constructed from activity accumulated over the illustrated 700 ms during six stimulus repetitions. The time window over which cuneate impulses were accumulated in the LH was from 5 to 10 ms after each SAII spike and SAIl in- stantaneous frequencies greater than 200 Hz were not analyzed. Each column in the LH is 0.2 ms in width
ceptive field located on the dorsum of the distal forelimb that corresponded with the receptive field focus of its si- multaneously recorded cuneate target neuron. None of these three SAIl fibers was observed to activate more than the one cuneate neuron in the course of microelec- trode tracking to identify additional cuneate neurons that might have been targets of the monitored SAII fiber. All three target neurons were recorded approximately 1 mm caudal to the obex, at depths of 700-1300 gm below the surface of the medulla, and are therefore likely to be cu- neothalamic projection neurons (Andersen et al. 1964b; Gordon and Jukes 1964).
Secure transmission between SAII peripheral fibers and their cuneate target neurons
In each of the cases where an SAII fiber and cuneate neuron were simultaneously recorded, most impulses in the SAII fiber were effective in generating an impulse in the cuneate neuron at an almost invariant latency. This latency, between recording an SAII fiber impulse periph- erally and a cuneate impulse centrally, was in the range 5.5-5.8 ms for the three pairs. The effectiveness of SAII impulses in activating the cuneate neuron is shown by the simultaneously recorded responses to a 300-gin step indentation of the skin at the receptive field focus (Fig. 1). The record from the fascicle of peripheral nerve (1 ~ contains impulses from two afferent fibers, one of which (Fig. 1, predominantly upward or positive spike) was unresponsive to the tactile stimulus (top trace) but had spontaneous activity and no effect on the cuneate
neuron. The other fiber (Fig. l, impulses predominantly downward or negative in 1 ~ record) was identified as an SAII fiber because it had a regular slowly adapting re- sponse to the step indentation, a punctate receptive field, was excited by skin stretch along one axis of the fore- limb and not along the perpendicular axis, and had spon- taneous activity at a rate of -6 impulses s -1. Almost all impulses in this SAII fiber were followed at a short inter- val by an impulse in the cuneate neuron (Fig. 1). As the SAII impulses were evoked in response to a static dis- placement, and therefore were not synchronized to a stimulus, this indicates that only the monitored SAII fi- ber was responsible for the activity of the cuneate neu- ron. The LH (see Materials and methods) below the im- pulse records confirms the reliable and regular response of this cuneate neuron to the SAII fiber impulses. The modal latency between the recording of an impulse at the two sites was 5.68 ms, with a SD of 0.09 ms.
The transmission security (defined as the percentage of SAII impulses that evoked spike output from the cu- neate neuron) attained values as high as 84%, 90%, and 95% for each of the three pairs, in their responses to static indentation during a 1-s segment that started 300 ms after the onset of a step indentation. Furthermore, these values were obtained with all SAII spikes included, irrespective of instantaneous frequency. Transmission security in the three pairs for the vibration-induced responses (quantified from records such as those in Fig. 2) also attained very high values. For example, when SAII response levels were essentially 1:1 (i.e., one impulse per vibration cycle) in response to vibration frequencies of 100, 100, and 200 Hz, respectively, transmission security values for the three pairs were 98.4%, 99.8%, and 99.9%.
Activation of SAII fiber-cuneate neuron pairs by vibrotactile stimulation
Sinusoidal vibrotactile stimuli were effective in activat- ing SAII afferent fibers at rates up to 600 impulses s -1 (Gynther et al. 1992a) and were used to provide con- trolled input to the cuneate neuron. The maximum ira-
70
3 0 Hz
LH 1.0
0
transmission security = 84%
l mean response = 1.9
. ,?L, latency = 5.5 ms
2 0 0 Hz
1 ~
2'
LH 0.20 transmission security = 62%
r~L mean response = 1 .o
o . . . la tency= 6.Oms 0 2.5 5.0 7.5ms
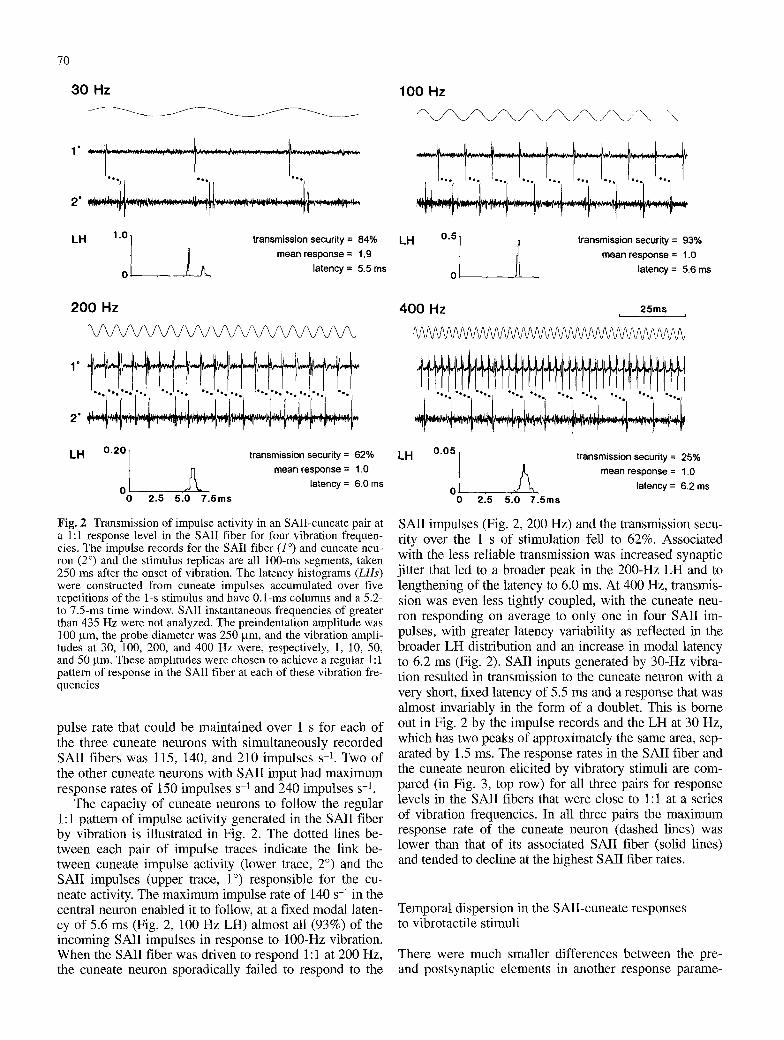
Fig. 2 Transmission of impulse activity in an SAII-cuneate pair at a 1:1 response level in the SAII fiber for four vibration frequen- cies. The impulse records for the SAIl fiber (1 ~ and cuneate neu- ron (2 ~ and the stimulus replicas are all 100-ms segments, taken 250 ms after the onset of vibration. The latency histograms (LHs) were constructed from cuneate impulses accumulated over five repetitions of the 1-s stimulus and have 0.1-ms columns and a 5.2- to 7.5-ms time window. SAIl instantaneous frequencies of greater than 435 Hz were not analyzed. The preindentation amplitude was 100 gin, the probe diameter was 250 l.tm, and the vibration ampli- tudes at 30, 100, 200, and 400 Hz were, respectively, 1, 10, 50, and 50 gm. These amplitudes were chosen to achieve a regular 1:1 pattern of response in the SAIl fiber at each of these vibration fre- quencies
pulse rate that could be maintained over 1 s for each of the three cuneate neurons with simultaneously recorded SAII fibers was 115, 140, and 210 impulses s -1. Two of the other cuneate neurons with SAII input had maximum response rates of 150 impulses s -t and 240 impulses s -t.
The capacity of cuneate neurons to follow the regular 1:1 pattern of impulse activity generated in the SAIl fiber by vibration is illustrated in Fig. 2. The dotted lines be- tween each pair of impulse traces indicate the link be- tween cuneate impulse activity (lower trace, 2 ~ and the SAII impulses (upper trace, 1 ~ responsible for the cu- neate activity. The maximum impulse rate of 140 s -1 in the central neuron enabled it to follow, at a fixed modal laten- cy of 5.6 ms (Fig. 2, 100 Hz LH) almost all (93%) of the incoming SAIl impulses in response to 100-Hz vibration. When the SAII fiber was driven to respond 1:1 at 200 Hz, the cuneate neuron sporadically failed to respond to the
100 Hz
LH ~ t transmission security = 93% mean response = 1.0
0 . latency = 5.6 ms
400 Hz 25ms I l
L H 0 . 0 5 _ _ transmission security = 25%
mean response = 1.0
0 . latency = 6.2 ms 0 2:5 5.0 7.5ms
SAII impulses (Fig. 2, 200 Hz) and the transmission secu- rity over the 1 s of stimulation fell to 62%. Associated with the less reliable transmission was increased synaptic jitter that led to a broader peak in the 200-Hz LH and to lengthening of the latency to 6.0 ms. At 400 Hz, transmis- sion was even less tightly coupled, with the cuneate neu- ron responding on average to only one in four SAII im- pulses, with greater latency variability as reflected in the broader LH distribution and an increase in modal latency to 6.2 ms (Fig. 2). SAII inputs generated by 30-Hz vibra- tion resulted in transmission to the cuneate neuron with a very short, fixed latency of 5.5 ms and a response that was almost invariably in the form of a doublet. This is borne out in Fig. 2 by the impulse records and the LH at 30 Hz, which has two peaks of approximately the same area, sep- arated by 1.5 ms. The response rates in the SAII fiber and the cuneate neuron elicited by vibratory stimuli are com- pared (in Fig. 3, top row) for all three pairs for response levels in the SAII fibers that were close to 1:1 at a series of vibration frequencies. In all three pairs the maximum response rate of the cuneate neuron (dashed lines) was lower than that of its associated SAII fiber (solid lines) and tended to decline at the highest SAII fiber rates.
Temporal dispersion in the SAII-cuneate responses to vibrotactile stimuli
There were much smaller differences between the pre- and postsynaptic elements in another response parame-

300.
16o 260 360 460
"g
-~ c- C O
Ct'~ "g l- 0J o i
i 0~
A
400
Q
O "'o-- - -" ~
•O .......
---------n , - n
Id0 200 300 460
71
200.
I00
0 0
. . . . . . . . o . . . .
-4 .
100 200 360 460
0 0
B
r--,300
tn U') C X ~ 200 m m
E 100 L.--.J
C
a '":,,~ " " 0 . . . . . . . . 0 . . . . . . 0
160 260 360 460
Frequency LrHz]
400"
Fig. 3A-C Impulse rates in the responses of three SAII-cuneate pairs to vibratory stimuli that activated the SAII fiber at close to the 1:1 level. The response was taken as the number of impulses generated in the SAII fiber (squares, continuous lines) or cuneate neuron (circles, dashed lines) during the l-s train of vibration. The bottom row plots, as a measure of temporal dispersion, the SD of the cycle histogram distributions constructed from the same data as the upper graphs. For C, the SD values at 30 Hz were off the scale, but were 4.6 ms in the SAII fiber and 3.3 ms in the cuneate neuron. Preindentation amplitudes: A 300 p.m; B 200 gin; C 100 gm. All data were obtained with a 250-gin-diameter probe
300"
200-
tO0-
0 0 160 ..... '" ' 200 '300 460
400-
ter, namely the tightness of phaselocking in responses to a particular vibration frequency (Fig. 3, lower graphs). The extent of phaselocking, or temporal dispersion in the occurrence of impulses during successive vibration cy- cles, was quantified in terms of the standard deviation (SD) of the cycle histogram distributions (see Materials and methods). These SD measures demonstrate the simi- larity in the tightness of phaselocking in the SAII fiber and associated cuneate neuron over the (400 Hz) band- width of vibration frequencies examined, although at the highest frequencies (>200 Hz) the phaselocking ap- peared to be a little poorer in the cuneate neuron than in the paired SAII fiber (Fig. 3, lower graphs). Although there was little change in absolute values of the SDs over the vibration frequency range from 200 to 400 Hz in ei- ther the pre- or postsynaptic elements (Fig. 3), the phase- locking is effectively poorer at the higher frequencies where the cycle period of the vibration is briefer and therefore the SDs become a more substantial fraction of the cycle period.
One of the additional SAII-related cuneate neurons, for which the SAII input fiber could not be adequately
0 0
" '% . . . . o
D
160 260 360 460
discriminated, responded in a tightly phaselocked, essen- tially 1:1 manner to a 1-s train of vibration at 200 Hz (30 gm; SD for the cycle histogram distribution, 0.31 ms). In another unpaired recording, the SAII-related cuneate neuron followed up to 100 Hz at low amplitudes (10 gin) in an essentially 1:1 manner with tight phase- locking (SD 1.08 ms).
Evidence that the monitored SAII fiber was responsible for the cuneate responses
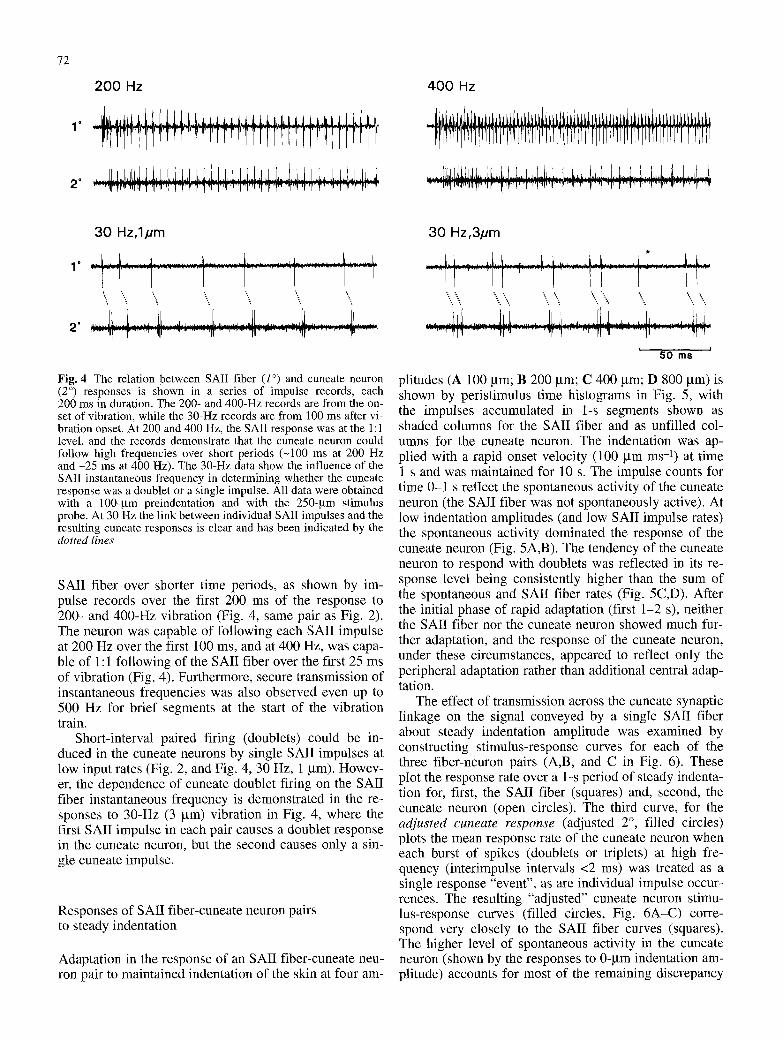
In the three simultaneous paired recordings, the respons- es of the cuneate neurons to vibrotactile stimuli were en- tirely attributable to the input from the SAII fiber under study, as no response was elicited by stimuli outside the receptive field of the fiber (1 mm away from the focus), and impulse failure in the SAII fiber was associated with a correlated failure of response in the cuneate neuron. For example, in Fig. 4 (30 Hz, 3 gm) the SAII fiber's re- sponse to one cycle of the vibration (indicated by the star symbol) is not paired and, correlated with this failure, the corresponding cuneate response lacks the single im- pulse that normally follows the doublet in the response to each vibration cycle.
Capacity of the cuneate neuron to follow high-frequency vibrotactile inputs from the SAIl fiber
Although the cuneate neuron could not sustain impulse rates as high as those in the SAIl fiber over a period of 1 s (Fig. 3), its performance was a better match to the
72
1"
.
2 0 0 Hz
........ II..I.LI.,JtI.,L,I...L.I .I, J .L,I .t,. 1_ LI,LLLI,J~L.LJ,..LI,.L.I~J,.J..,I.L,I.
..... " ] I "WT" IT I iVT"T" I " r r F I " r 'T 7 r r l i l l ~ ' li ~ " ' I " t ] '1 "l'[ 1'f '17" ] r "~ ' t ' r "1 r ' �9 1~ T ~ T .~ IF '~ 11"
4 0 0 Hz
ii,,,,.,(~,,I.lJ.l.t,l J,L, .LL,.i.LL I.ILL.L._L.[.L ],~..L.I,,J,~L~
30 Hz , lpm
.... L,_..~ .. . . . . . . . . . . i . . . . . . . . . . . . . . . . . . .i . . . . . . . . . . . . t . . . . . . . . . . . . . . . . . i . . . . . . . . . . . . . . . L ....... ...L
" . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1 ............. I ,,. - .... - ... .. ,,:
~..,IL..,,[ .............. LL..., .......... . . . . . . . . . . . . . . . . I[ ..................... [L . . . . . . . . . . . . . . J i ........... 2" l r "~ .......... " II ....................... 11" "" ' "71 ................ ~II ............. 11 .........
Fig. 4 The relation between SAII fiber (1 ~ and cuneate neuron (2 ~ responses is shown in a series of impulse records, each 200 ms in duration. The 200- and 400-Hz records are from the on- set of vibration, while the 30-Hz records are from 100 ms after vi- bration onset. At 200 and 400 Hz, the SAII response was at the 1:1 level, and the records demonstrate that the cuneate neuron could follow high frequencies over short periods (-100 ms at 200 Hz and -25 ms at 400 Hz). The 30-Hz data show the influence of the SAII instantaneous frequency in determining whether the cuneate response was a doublet or a single impulse. All data were obtained with a 100-gm preindentation and with the 250-gm stimulus probe. At 30 Hz the link between individual SAII impulses and the resulting cuneate responses is clear and has been indicated by the dotted lines
SAII fiber over shorter time periods, as shown by im- pulse records over the first 200 ms of the response to 200- and 400-Hz vibration (Fig. 4, same pair as Fig. 2). The neuron was capable of following each SAII impulse at 200 Hz over the first 100 ms, and at 400 Hz, was capa- ble of 1:1 following of the SAII fiber over the first 25 ms of vibration (Fig. 4). Furthermore, secure transmission of instantaneous frequencies was also observed even up to 500 Hz for brief segments at the start of the vibration train.
Short-interval paired firing (doublets) could be in- duced in the cuneate neurons by single SAII impulses at low input rates (Fig. 2, and Fig. 4, 30 Hz, 1 gm). Howev- er, the dependence of cuneate doublet firing on the SAII fiber instantaneous frequency is demonstrated in the re- sponses to 30-Hz (3 gin) vibration in Fig. 4, where the first SAII impulse in each pair causes a doublet response in the cuneate neuron, but the second causes only a sin- gle cuneate impulse.
Responses of SAII fiber-cuneate neuron pairs to steady indentation
Adaptation in the response of an SAII fiber-cuneate neu- ron pair to maintained indentation of the skin at four am-
30 Hz,3pm
. . . . . . . . . . . . . ] .................. I I . . . . . . . . . I . . . . . . . . . . . . . . 1 . . . . _ _ �9 : : ; - . - : ". : - ~ - : ; ~,
. . . . . . . . . t ~J~ . . . . . . . . . . . . . 1 ~.iL . . . . . . . . . . . ~ ' . ' l . . . . . . . . . . ] [ / .~ , ,J J . . . . . . . . . lJ .................... JL2 i I
5 0 m s
plitudes (A 100 him; B 200 Ixm; C 400 Ixm; D 800 gm) is shown by peristimulus time histograms in Fig. 5, with the impulses accumulated in 1-s segments shown as shaded columns for the SAII fiber and as unfilled col- umns for the cuneate neuron. The indentation was ap- plied with a rapid onset velocity (100 gm ms -1) at time 1 s and was maintained for 10 s. The impulse counts for time 0-1 s reflect the spontaneous activity of the cuneate neuron (the SAII fiber was not spontaneously active). At low indentation amplitudes (and low SAII impulse rates) the spontaneous activity dominated the response of the cuneate neuron (Fig. 5A,B). The tendency of the cuneate neuron to respond with doublets was reflected in its re- sponse level being consistently higher than the sum of the spontaneous and SAII fiber rates (Fig. 5C,D). After the initial phase of rapid adaptation (first 1-2 s), neither the SAII fiber nor the cuneate neuron showed much fur- ther adaptation, and the response of the cuneate neuron, under these circumstances, appeared to reflect only the peripheral adaptation rather than additional central adap- tation.
The effect of transmission across the cuneate synaptic linkage on the signal conveyed by a single SAII fiber about steady indentation amplitude was examined by constructing stimulus-response curves for each of the three fiber-neuron pairs (A,B, and C in Fig. 6). These plot the response rate over a 1-s period of steady indenta- tion for, first, the SAII fiber (squares) and, second, the cuneate neuron (open circles). The third curve, for the a d j u s t e d c u n e a t e r e s p o n s e (adjusted 2 ~ filled circles) plots the mean response rate of the cuneate neuron when each burst of spikes (doublets or triplets) at high fre- quency (interimpulse intervals <2 ms) was treated as a single response "event", as are individual impulse occur- rences. The resulting "adjusted" cuneate neuron stimu- lus-response curves (filled circles, Fig. 6A-C) corre- spond very closely to the SAII fiber curves (squares). The higher level of spontaneous activity in the cuneate neuron (shown by the responses to 0-gin indentation am- plitude) accounts for most of the remaining discrepancy

A
C
�9 n. .ZI. O. o/]. n. [7. [7 o. n 0 2 4 6 8 10
100
80
?.
I l / l
Ul (U
(:L E
t_..i
4J Ul E 0 I=L Ul
8,
6O
0 . . . . . . . . . . . .
0 2 4 6 8 10
Time Is]
Fig. 5 A - D Peristimulus histograms of the response of an SAII- cuneate pair to a 10-s skin indentation commencing at 1 s. The number of impulses discharged each second is shown by the height of the shaded columns for the SAII fiber and by the unfilled columns for the cuneate neuron. Indentation amplitude was ZOO g m in A, 200 g m in B, 400 g m in C, and 800 p m in D, all de- livered with a 2-mm-diameter probe and with a ramp velocity of 100 gm ms -I. Each column is the mean of five responses
73
B 100
80.
60.
40.
0 2 4 6 8 10
100
80
60
40
20
0 0
D
i
2 4 6 8 10
Time [s I
between the S MI fiber and adjusted cuneate neuron re- sponse rates (Fig. 6). For all three SAII-cuneate pairs, treatment of both the burst of spikes and the individual cuneate impulses as "units" of rate-coded information re- duces the range of response levels of the cuneate neuron, from the unadjusted case (open circles, Fig. 6A-C) to a level almost coinciding with that of the SAII fiber (squares).
Fig. 6 A - C Stimulus-response relations for three SAII-cuneate pairs in response to static indentation. The response was the num- ber of impulses discharged over a 1-s period, commencing 300 ms after the onset of indentation. The data from the SAII fibers are plotted as squares and those from the cuneate neurons as circles. The filled circles plot the response of the cuneate neuron adjusted so that doublet and triplet responses (interimpulse intervals <2 ms) are counted as single impulses. Each data point is the mean of at least five responses and the error bars are SDs. A and B were ob- tained with the 2-mm probe, C with the 250-gm probe
Discussion
Identification of SAII-related cuneate neurons
The seven cuneate neurons were all confidently identi- fied as having SAII input, because the small amount of convergence and the high security of synaptic transmis- sion meant that their response properties and receptive fields were similar to those of SAIl afferent fibers
A 75- 75, 75'
50-
2 5 ~
5o
25
@ 6 260 460 660 g60 IObO
B
6 260 460 660 e60 lobo
Indentation Amplitude [um]
50
25
6 260 4~)o
C
~-" 100 Pm
8O
60
4O
~. 20
0

74
(Chambers et al. 1972; Gynther et al. 1992a). All seven were located behind the obex in the cuneate cluster zone that is known to contain mainly thalamic-projecting neu- rons (Andersen et al. 1964b). The single-focus receptive field observed for six out of the seven SAII-related cu- neate neurons was consistent with the single receptive field focus for the three SAIl lemniscal units studied by Brown et al. (1974) and with Golovchinsky's (1980) findings for his sample of SAil-related cuneate neurons. These observations suggest that SAII input to individual cuneate neurons is almost always limited to a single SAIl fiber. The absence of an inhibitory field for any of the SAII-related cuneate neurons in the present study was also in agreement with findings of Brown et al. (1974) and Golovchinsky (1980).
Security of cuneate transmission for SAII inputs
The small sample of SAII-cuneate pairs obtained reflects the low incidence of central representation of SAIl input (Brown et al. 1974) and the technical difficulties of the paired, simultaneous recording paradigm. Nevertheless, the present study provides the first direct demonstration that the minimal SAII input, a single impulse arriving over an individual fiber, is able to generate spike output from cuneate target neurons. Furthermore, substantial amplification of the SAII input signal may occur as the individual SAIl spike often generates a closely spaced burst of impulses, usually a doublet or triplet, in the cu- neate neuron. This amplification could enhance the prob- ability of detection of weak input signals and could pro- vide for a form of temporal summation to ensure secure transmission at the next synapse in the pathway, at the thalamus.
The results confirm the accuracy of the indirect esti- mates by Amassian and Giblin (1974) of high security for the SAII-cuneate synapse and also establish that the security of synaptic transmission for SAII fibers in the dorsal column nuclei is very similar to that demonstrated in our previous studies for individual fibers of the PC and SAI classes of tactile sensory fibers (Ferrington et al. 1986, 1987a,b; Vickery et al. 1994). Although the early intracellular studies by Eccles' group (Andersen et al. 1964a) had revealed large unitary excitatory postsynaptic potentials (EPSPs) in dorsal column nuclei neurons, the observed EPSPs were, nevertheless, subthreshold. How- ever, our paired recording analysis of transmission for PC, SAI, and now SAIl fibers, confirms that, for each of these sensory fibers, the single-fiber unitary EPSPs must indeed be capable of exceeding threshold. This potent linkage across the dorsal column nuclei may reflect the fact that terminal boutons formed by input fibers (Rozsos 1958; Walberg 1966; Rustioni and Sotelo 1974) are larg- er than, for example, those formed by input fibers to motoneurons, and that individual afferent fibers supply- ing the dorsal column nuclei may give rise to high densi- ties of en passant and terminal boutons (Fyffe et al. 1985).
Signalling of information about static skin displacement and vibrotactile stimuli in the SAII-cuneate linkage
Information about the amplitude of steady skin indenta- tion appeared to be reliably conveyed from the SAII fiber to its cuneate target neuron in terms of an impulse rate that was graded, in both elements of the pair, as a func- tion of indentation amplitude. Furthermore, at most in- dentation amplitudes (Fig. 6), the response level in the cuneate neuron exceeded that in its SAII input fiber on account of the doublet and triplet responses to individual SAII spikes.
The secure transmission in the SAII-cuneate linkage enabled cuneate neurons to replicate, over a 1-s period of response, the 1:1 impulse pattern (1 impulse per vibra- tion cycle) elicited in the afferent fiber by vibration fre- quencies up to a maximum of -200 Hz (Figs. 2-4). Al- though Golovchinsky (1980) reported that some slowly adapting cuneate neurons responded with an entrained, 1:1 pattern up to frequencies of 320-500 Hz, the period over which this pattern of response was sustained was not specified; nor was there any monitoring of the num- ber of active input fibers. As the response traces in our Fig. 4 show, the capacity of the SAII-related cuneate neurons to follow high vibration frequencies (200-400 Hz) in a 1:1 pattern can be sustained only over periods of less than 100-200 ms. However, despite this restriction on the impulse rates of the SAII-related cu- neate neurons, our analysis shows that the cuneate re- sponses remain tightly phaselocked to vibration frequen- cies up to -400 Hz.
It is probable that naturally encountered stimuli would also excite SAII fibers to discharge at the high in- stantaneous frequencies described in the present study. For example, when the skin is brushed against a textured surface this will generate a combination of skin stretch and vibration, conditions that are known to produce high-frequency discharge in SAII fibers in the hairy skin (Gynther et al. 1992a). The present results indicate that the dynamic as well as the static components of informa- tion contained in such natural stimuli are likely to be transmitted with high fidelity to cuneate neurons. It should also be emphasized that in response to long-last- ing static displacement of the skin, of the kind that might arise from skin stretch associated with particular joint positions, the maintained SAIl discharge is reliably transmitted at the SAII-cuneate synapse. This was seen in Fig. 5, where the cuneate neuron maintained its re- sponse to the single SAII input over a 10-s period.
Cuneate transmission characteristics for single SAII fibers: relation to perceptual effects of intraneural microstimulation
When individual sensory fibers are selectively stimulated in the peripheral nerves of human subjects by means of the intraneural microstimulation procedure, there are marked differences observed in the capacities of the vari-

ous fiber classes to generate subjective sensory experi- ence (Ochoa and Torebj6rk 1983; Vallbo et al. 1984; Macefield et al. 1990). For example, there is no percep- tual response elicited by stimulation of single SAI l fi- bers, in contrast to the distinct perceptual effects associ- ated with the activation o f single fibers o f the PC, RA, or SAI classes. One of the hypotheses tested in the present study was that the differential capacity of these fiber classes to contribute to perceptual experience may be at- tributable to differences in their cuneate transmission se- curity. However, the present demonstrat ion o f a remark- able synaptic security in the cuneate nucleus for SAI l fi- bers, which resembles that demonstrated in our previous studies for PC and SAI fibers, (Ferrington et al. 1986, 1987a,b; Vickery et al. 1994), shows that this explana- tion cannot apply, unless, of course, there were some marked species difference between cat and man in this respect, Perhaps the failure of individual SAII fibers to generate perceptual responses is related to limited diver- gence and a relatively low incidence o f representation for this class o f input within the central pathways. Further- more, SAII inputs may lack a representation in the cortex that is appropriate for perceptual recognition.
Acknowledgements Dr. S. Ghosh participated in some of the ex- periments. We acknowledge the technical assistance of C. Rior- dan, R Mackie, S. Bray, and E Spicer. This work was supported by the National Health and Medical Research Council of Australia and the Australian Research Council. Richard M. Vickery was a New South Wales Medical Research Scholar.
References
Amassian VE, Giblin D (1974) Periodic components in the steady- state activity of cuneate neurones and their possible role in sensory coding. J Physiol (Lond) 243:353-385
Andersen R Eccles JC, Oshima T, Schmidt RF (1964a) Mecha- nisms of synaptic transmission in the cuneate nucleus. J Neu- rophysiol 27:1096-1116
Andersen R Eccles JC, Schmidt RF, Yokota T (1964b) Identifica- tion of relay cells and interueurons in the cuneate nucleus. J Neurophysiol 27:1080-1095
Bennett RE, Ferrington DG, Rowe MJ (1980) Tactile neuron classes within second somatosensory area (SII) of cat cerebral cortex. J Neurophysiol 43:292-309
Brown AG, Gordon, G, Kay RH (1974) A study of single axons in the cat's medial lemniscus. J Physiol (Lond) 236:225-246
Burgess PR, Petit D, Warren RM (1968) Receptor types in cat hairy skin supplied by myelinated fibers. J Neurophysiol 31: 833-848
Chambers MR, Andres KH, Duering M v, Iggo A (1972) The structure and function of the slowly adapting type IX mechano- receptor in hairy skin. Q J Exp Physiol 57:417-445
Ferrington DG, Rowe MJ (1980) Differential contributions to cod- ing of cutaneous vibratory information by cortical somatosen- sory areas I and IX. J Neurophysiol 43:310-331
75
Ferrington DG, Rowe MJ, Tarvin RPC (1986) High gain transmis- sion of single impulses through dorsal column nuclei of the cat. Neurosci Lett 65:277-282
Ferrington DG, Rowe MJ, Tarvin RPC (1987a) Actions of single sensory fibers on cat dorsal column nuclei neurons: vibratory signalling in a one-to-one linkage. J Physiol (Lond) 386: 293- 309
Ferrington DG, Rowe M J, Tarvin RPC (1987b) Integrative pro- cessing of vibratory information in cat dorsal column nuclei neurons driven by identified sensory fibers. J Physiol (Loud) 386:311-331
Fyffe REW, Cheema SS, Light AR, Rustioni A (1985) The organi- zation of neurons and afferent fibers in the cat cuneate nucle- us. Xn: Rowe MJ, Willis WD (eds) Development, organization, and processing in somatosensory pathways. Liss, New York, pp 163-173
Golovchinsky V (1980) Patterns of responses of neurons in cu- neate nucleus to controlled mechanical stimulation of cutane- ous velocity receptors in cat. J Neurophysiol 43:1673-1699
Greenstein J, Kavanagh E Rowe MJ (1987) Phase coherence in vi- bration-induced responses of tactile fibres associated with pa- cinian corpuscle receptors in the cat. J Physiol (Lond) 386: 263-275
Gordon G, Jukes MGM (1964) Dual organization of the extero- ceptive components of the cat's gracile nucleus. J Physiol (Lond) 173:263-290
Gynther BD, Vickery RM, Rowe MJ (X992a) Responses of slowly adapting type II afferent fibres in cat hairy skin to vibrotactile stimuli. J Physiol (Lond) 458:151-169
Gynther BD, Vickery RM, Rowe MJ (1992b) Transmission char- acteristics across a 1:1 linkage between slowly adapting type II afferents and their cuneate target neurones. Proc Aust Neu- rosci Soc 3:75
Macefield G, Gandevia SC, Burke D (1990) Perceptual responses to microstimulation of single afferents innervating joints, mus- cles and skin of the human hand. J Physiol (Lond) 429:113- 129
Mountcastle VB, Talbot WH, Sakata H, Hyvfirinen J (1969) Corti- cal neuronal mechanisms in flutter vibration studied in un- anesthetized monkeys. Neuronal periodicity and frequency discrimination. J Neurophysiol 32:452-484
Ochoa J, lbrebj6rk E (1983) Sensations evoked by intraneural mi- crostimulation of single mechanoreceptor units innervating the human hand. J Physiol (Lond) 342:633-654
Rozsos I (1958) The synapses of Burdach's nucleus. Acta Morp- hol Hung 8:105-109
Rustioni A, Sotelo C (1974) Synaptic organization of the nucleus gracilis of the cat. Experimental identification of dorsal root fi- bres and cortical afferents. J Comp Neurol 155:441-468
Vallbo AB, Olsson KA, Westberg K-G Clark FJ (1984) Micro- stimulation of single tactile afferents from the human hand. Sensory attributes related to unit type and properties of recep- tive field. Brain 107:727-747
Vickery RM, Gynther BD, Rowe MJ (1992) Vibrotactile sensitivi- ty of slowly adapting type I sensory fibers associated with touch domes in cat hairy skin. J Physiol (Lond) 453:609-626
Vickery RM, Gynther BD, Rowe MJ (1994) Synaptic transmission between single slowly adapting type I fibres and their cuneate target neurones in cat. J Physiol (Lond) 474:379-392
Walberg F (1966) The fine structure of the cuneate nucleus in nor- mal cats and following interruption of afferent fibres. An elec- tron microscopical study with particular reference to findings made in Glees and Nauta sections. Exp Brain Res 2:107-128




















