
Translation 3185
279
286 how is this relationship expressed in both ecological groups, the aquatic and semi-aquatic mammals? In this scheme we have attempted to examine simul:taneously the structure of a typical rete mirabili of the dolphin, taking into account the possible pathology, the structure of the mesenteric artery and of some of the lymphatic nodes, representing in this case portions of the lymphatic system. MATERIALS AND . METHODS We had four dolphin specimens available to us: three common porpoises, Phocaena phocaena, and one bottlenose dol- phin, Tursiops truncatus, from the Black Sea, and also four fur seals, Callorhynus ursinus, two Steller's sea lions, Eume- t0DiUS jubatus, and two ha:I:Lour seals, Phoca vitulina, obteined on Robben Island in 1968. Two of the investigated dolphins (one common porpoise and the bottlenose dolphin) died from different diseases, while the remaining two were dissected after an acute physio- logical experiment. Morphological and histological methods were used to study sections of the intercostal, supraclavicular, axillary, lumbar and iliac Portions of the retia mirabilia, sections of the mesenteric artery, the mesenteric, axillary, pancreatic, cervical and other lymph nodes. The sections were stained with haematoxylin and eosin and resorcin-fuchsin. The lymph nodes were measured in the fresh state; some of them were weighed.
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Translation 3185

286
how is this relationship expressed in both ecological groups,
the aquatic and semi-aquatic mammals? In this scheme we have
attempted to examine simul:taneously the structure of a typical
rete mirabili of the dolphin, taking into account the possible
pathology, the structure of the mesenteric artery and of some
of the lymphatic nodes, representing in this case portions of
the lymphatic system.
MATERIALS AND . METHODS
We had four dolphin specimens available to us: three
common porpoises, Phocaena phocaena, and one bottlenose dol-
phin, Tursiops truncatus, from the Black Sea, and also four
fur seals, Callorhynus ursinus, two Steller's sea lions, Eume-
t0DiUS jubatus, and two ha:I:Lour seals, Phoca vitulina, obteined
on Robben Island in 1968.
Two of the investigated dolphins (one common porpoise
and the bottlenose dolphin) died from different diseases,
while the remaining two were dissected after an acute physio-
logical experiment.
Morphological and histological methods were used to
study sections of the intercostal, supraclavicular, axillary,
lumbar and iliac Portions of the retia mirabilia, sections of
the mesenteric artery, the mesenteric, axillary, pancreatic,
cervical and other lymph nodes. The sections were stained
with haematoxylin and eosin and resorcin-fuchsin.
The lymph nodes were measured in the fresh state; some
of them were weighed.

287 • 1- a
THE RETIA.MIRABILIA OF DOLPHINS
Common porpoises No. 2 and No. 3. The arterial net-
work in the intercostal region was investigated. The animals
were dissected after narcotization. The intercostal vascular
network is represented by a spongy tissue, composed of extreme-
ly coiled hollow vessels - the arterioles. Variations in the
diameter of these vessels are not great, from 200 to 1000 IL.
The lumen is gaping and filled with blood. The structure of
the walls of the vessels is typical of vessels of this category
(Figure 1). The internal layer is well developed, after this
comes a distinct thick-layered elastic membrane. In the tunica
media the circular smooth musculature is very dense and abun-
dant. It is bounded on the outside by a second elastic memb-
rane, which is also of considerable thickness and uninterrupted.
The tunica externa of the vessel is maximally reduced, it does 181
not contain collagen and is not firmly accreted to the surroun-
ding tissue. In this manner, the vessel is composed as it
were of a two-layered elastic tube, capable of withstanding
high pressure both from the inside as well as from the outside.
The tissue surrounding the retia mirabilia contains
lymphatic vess'els, sometimes covering the arterioles as it were
with a sheath. The veins form sometimes small sometimes wide,
very thin-walled sinuses. In places extravasations are evi-
dent in the dilated lymphatic capillaries and in the connective
tissue.
The erythrocytes which fill the arterioles are bi-
concave and sometimes crescent-shaped, which may indicate an-
aemia. In places arteritis obliterans was diagnosed, but no
infiltrates in the walls of the vessels were found.

288
p}!c. 1. RllyAectMa ceT6N 113 HaAx.110v11Tloif o6.nac•rf1 MopcKoi ► CIIIII[bil M 2
1-- etl^rpcttltstn 3.iacTUVCCI.aR oGoaolu<a; 2=-11apYacuast 3.7aCTH4CCFaA o1107o4Ha:.
, • :f - cpCAiIRA oûo, o,lha cocyAa; d- m1a+l( 14JTIMCCK11C cocyluIt.X8
Figure 1. Rete mirabile from the supraclavicular region.
of porpoise No. 2.
1 - inner elastic nnEmbrane;2 - outer elastic membrane;3 - tunica media of vessel;4. - lymphàtic vessels. X 8.
Next was studied the arterial "network" from the lum-
bar and iliac aorta (ventrally to this is situated the venous
"rete mirabile"). The general structure of the "network" is
similar to that in the thoracic cavity. All of the layers
are found in the walls of the arterioles. It seems, however,
that the elastic portions are here less strongly developed
and that the vessels are in a somewhat more collapsed state.
The cause of this is not cl.ear: of significance in this res-
pect may be the lowered pressure in the abdominal cavity, in
comparison with the thoracic cavity, the effect of the narcosis
on the autonomic centers, leading to spasms of the vessels,
or the proximity of nidi of helminth infestation.

289
In the lumbar "network" there appear sections, where
the walls of the vessels are • asophilic. From the tunica
interna, in the composition of which there are present muscle
fibres, cells are desquamated and fill the lumen of the vessel.
However alongside the described section there lie normal full-
value arterioles, typical of the retia mirabilia.
Common porpoise No. 1. The animal was dissected after
death, which occurred from an unknown cause. The thoracic
section of the retia mirabilia was traced from the intercostal
artery. The artery was not very elastic, the walls were friable
and the aorta itself was deformed. The rete mirabile starts
from the intercostal artery. Both of the elastic membranes
in the vessels of the network were damaged and the picture
resembled that of typical sclerosis. In the tunica media of
the arterioles there is clearly evident the large-mésh elastic
basis, while the continuity of the circular musculature is
infringed in places. Basophily of the walls and the occurrence
of collagen fibres in the tunica externa was noted.
Around the arterioles of the network there are clearly
expressed and very dilated lymphatic vessels. The veins here
are small and thin-walled. Within the thickness of the struc-
ture are encountered lymphoid follicles.
The bottlenose dolphin. This dolphin died of pneumo-
nia, which was confirmed by the dissection of the lungs. In
the arterial retia mirabilia the vessels lie in a very dense
mass. The diameter of the arterioles ranges within the same
limits as in the common porpoise. In their walls are represen-
ted all of the elements, including both of the elastic membranes.

290 • .1 •
Around the artery there are lymPhatic vessels, though these
are very narrow, and much fat tissue. The circular muscula-
ture in the tunica media of the arterioles is dense, abundant
and stains intensively with eosin. Collagen fibres were not
evident and, consequently, the vessel was in a state of full
tension and retained its elasticity. Apparently, the above-
described picture of arteriosclerosis was created by some other
pathological conditions.
The venous rete mirabile of dolphins. The venous net-
work is situated in the lumbar region ventrally to the aorta,
between the suprarenals. This structure is composed of small
calibre veins and venules. While the arterial network has the
form of a cushion of twisted vessels, the venous network is
composed of numerous anastomoses, braided into a lattice. The
vessels are sinuous, with compact walls. The normal layering
of the venous wall is here indistinctly expressed. Around the
veins are situated lymphatic vessels.
THE MESENTERIC ARTERY
It is known that the mesenteric artery of man has a
mainly circular arrangement of the musculature in its tunica
media. In the mammals there are also found bundles of longi-
tudinal smooth muscles. The longitudinal musculature in the
wall of the vessel provides for its peristaltic contractions,
giving rise to the so-called "peripheral hearts" (Zavarzin,
1939). In the animals studied by us the walls of this were
not uniformly constructed (Table 1, Figures 2-•).

*OTaeablIble peRKIIC no- :mum
480 •
32 40 30
162 280
400 720
foo 240 100 400
140
60
100
340
•180 180
Table 1. Thickness of the layers of the wall of the mesenteric artery, m.
Ta 6 auna 1
Tomnuna canea CTCHKII 6phiaceelmoil apTcpull,
291
Elements of treVer
'Harbour Stellerl I el cilnyq d .
sea-lion
Yur. Bottlenose mopcuow mwiuna
sel dolphin
A Burrpeminfi ujori .6e3 mycKyanyint
"Ilpopuribnan mycxyairrypa • B cpeRneti 0607otwe
C 11,upKyanpu an mycEya a- Typa 11 cpeaudi o6o-.710 , 1KC
D .1Iapywrian o6oacp.wa: . e naoTuan f pleaam
A - Inner layer without musculature
B - Longitudinal musculature in tunica media *Individual sparse fibres •
C - Circular musculature in tunica media D - Tunica externa:
e - dense f - loose
Bottlenose dolphin. The wall of the mesenteric artery
is compact. Its thickness in the bottlenose dolphin is about
620 f, which is about 6.5 times smaller than the outer diameter
of the artery. The tunica media is composed of circular mus-
culature, while the longitudinal musculature is very sparsely
interspersed in the form of individual fibres or bundles (Fi- 183
gure 2). The tunica externa has a thickness of 140m- in all.
There are few collagen fibres; the elastic part is clearly
expressed.

29 2
Puc, 2._CTeinca 6pbuKeet11lori apTepuu mopcKoil cmiltbut (Ns ,i 3):
Burrpotnyin actaouri; cpentinn o6otiogica .; 3 — tiapyxuiaii o6oaogKa.x8 .
Figure 2. Wall of the mesenteric artery of a common porpoise (No; 3)1
1 - tunica interna 2 - tunica media 3 -.tunica externa. X 8.
Seals. The overall thickness of the wall of the artery
is within the same limits as in the bottlenose dolphin; in
the fur seal and the harbour seal it is one sixth, and in
Steller's sea lion one fifth of the diameter of the artery.
Longitudinal musculature in these animals is found in all of
the layers of the wall of the vessel. This musculature may
lie in separate bundles in the center of the circular muscu-
lature (Figure 3), in a continuous layer along the periphery

Z93
4
^w.^^:^^1:::....r^a_.<à,^.w_.,... .^._....^.^.:.^^:.:,,.ks.;-.:.s^,.;M•.. . ^.x^^:;^=u:^
Pt+c. 3. CTCU!ca Gphl^+:ce'tnoii ap•rep!nt atopcr.oro xoin!:a (M, 3):
1- tutpsy.inpnaa ^q'cr.y^arypa cpeAnetl o6)o, ïo4^:n; 2 - npo;toat.ttanxycxy.nat)'ya cpcAned oGoao4xn; 3 - na wnan oGono4h3.?t8
Figure 3. Wall of the mesenteric artery of a fur seal (No. 3):
1- circular musculatùre.of tunica media,-2 -,longitudinal musculature of tunica media;3 - tunica externa. X 8.
of this or in both sections simultaneously, as it were "squee-
zing" the circular musculature and elastic base (Figure 4).
In those places where the longitudinal musculature is stronger
the elastic base is weak and barely evident.
The thickest layer of longitudinal musculature was
found in the Stelier's sea-lion, in which the ratio of its
thickness to the thickness of the circular musculature was
1811.

294
)'•
1„.
'''' • ''' '
Pitc. 4. C-reliKa 61) 1) clœytta: / —Bityrpeutinsi amogKa; 2— 3.rt acT imeci( a n NICNI Cipali a; 3— npop,om..- 11BSI MYCKYJ1TYP ii cpepmeit o6o.logEe; —Itirpxyosiptian mycKyazrry-
pa n cpeiweil o6o.runKe.X8
Figure 4 • Wall of the mesenteric artery of Steller's sea-lion:
1 - tunica interna; 2 - elastic membrane; • 3 - longitudinal musculature in tunica media; 4 - circular musculature in tunica media. X 8.
7 110. In the harbour seal this ratio was 3 110, while in the
fur seal it was 2 :10 (see Table 1).
it is possible that the tunica externa of the vessel
is also dependent on the strength of the contraction of the
longitudinal musculature. The thickness of the tunica externa
is'greatest in the Steller's sea-lion (only one tenth that of
the diameter of the artery, while in the harbour seal it is

295
one thirtieth the diameter, and in the fur seal - one twenty-
seventh). However, in the last two seals the tunica externa
is compact and is closely accreted to the tunica media. In
the Steller's sea-lion, however, the dense layer of the tunica 185
externa is not thick, in all being about 100 'A. in thickness;
over this there lies freely a thick and loose collagen layer,
which in some sections is 4 times as thick as the dense layer.
Probably, within this loose layer there may be accomplished
contractions of the vessel in the longitudinal direction.
LYMPHATIC NODES
The lymphatic nodes as a constituent part of the vas-
cular system are not homogeneous in the animals of the two
groups of animais under comparison:. A marked difference is
evident in the dimensions of the nodes of the dolphins and of 186
the seal (Table 2). For example, the packet of mesenteric
nodes of the dolphin is considerably smaller than that of any
of the seals. The weight of such a node in the dolphin is
170 - 200 gm, While in Steller's sea-lion it is 750 gm. In
the dolphin the nodes are dense and lie in the mesentry; in
the seals they are loose and enclosed in a sac, similarly to
the kidneys, ovaries and other organs.
Common porpoise No. 1. The cervical lymphatic nodes
were studied. In the nodes there were found abundant extra-
neous inclusions (possibly the eggs of helminths) about 12 r_ in diameter. The nodes were hyperemic. Plasmocytes were evi-
dent in the parenchyma, while in the sinuses there could be
seen typical reticular cells with processes, histiocytes with

14,3
15,8
• 6,1
11
13,8
10,6
7,8
139
200
147
158
71
107
103
203
195
120
163,5
114
12,0
I1x5
28x10
28x 9
I9x 8
10x 5
9x 3
700
500
170 — 200
2
12
6
Table 2.
Comparative characteristics of the mesenteric lymphatic nodes.
• T a6.neu.a 2
Cpasinnenbuan xapawrepic-nima me3enTernia.TIbHUX oP.AtcparviecKiix y3n0u
A .nniceie y3non e.11 ■ Iia
BHA rem .
Species
29 6
d % AJ11111C Keluutile C At TCAO
Bee ,riaKera.,
G 06wee 411C/10 yuou
1 rpeumalukciaii -TIO.ICIlb
2 •à.1çpcKoii . HOTIIK .bis2 3
3 MopcKoit 1(01I1K Ne 5
Cinirt •Nt, 1
.5 Jlapra .N'g I
6 .A(panima •
7 ..MapcKan c111111b11
A - Body length, cm B - Greatest leng:th of "packet" of nodes
C — cm d - % of body length
.
e - % of length of kidney
F - Weight of "packet", gin
G - Total number of nodes
1 - Harp seal • 2 - Fur seal No. 3 . 5 — Fur seal No. 5 4 - Steller's sea-lion No. 1 5 — Harbour seal No. 1 6 — Bottlenose dolphin 7 — Common porpoise .
large nuclei and cells with elongate rod-like nuclei. In the
trabeculae, infiltrated with cells, there were present collagen
and elastic fibres and musculature, which here was somewhat
weaker than in the other nodes.

297
Common porpoise No. 2. The lumbar lymphatic node
was studied. The structure of the node was not typical of
the mammals: the cortical substance was arranged along the
peri.phery of the node, however the follicles did not have a
spherical form but were rather thickened zig-zag shaped cords.
The capsule of the node is dense, over this is the muscular
mesenteric membrane, in which the node lies. The trabeculae
in the nodes are not wide,but they are dense and are composed
maxnly of muscle and elastic fibres. The sinuses of the node
are compzressed by the densely branching trabeculae, they are
completely choked up with histiocytes.. Possibly, these are 18?
modified reticular cells which have' rounded up and lost their
processes. In.this organ phagocytic properties of these cells
were not noted.
The nuclei of such cells of the node were light coloured.
The chromatin was disposed in a reticulum and fine clusters.
There were many large and medium-sized lymphocytes. There were
no reactive centers. In the sinuses there were found free
erythrocytes and eosinophilic leucocytes.
The mesenteric lymphatic nodes were studied. The whole
"packet" of nodes was 9 cm in length and 3 cm in width. The
nodes were irregular in form: in places the medullary layer
was found at the surface and was pierced'by a mass of blood-
carrying vessels. Spherical secondary nodules.were also not
found in these nodes. Trabeculation was strikingly expressed
in these nodes. The trabeculae branched so densely and so
finely that they occupied all of the free spaces in the node.
Thus, the sinuses of the node here also were not free, but were

298
CB111-11,11 J\re.). 2 Pm. 5. Me3eirrepnambnbtil enimtpanmecKiiit peg mopcnii LI,eirrpa.nbindi citnyc
I— cmiye; 2 — MHIZOTIILIC uillypbt.x8
Figure 5. Mesenteric lymph node of common porpoise No. 2. Central Sinus
- sinus; 2 - medullary cords. X 8.
filled with histiocytes and and a fibrous reticulum (Figure 5),
not at all similar to that which is formed by the reticular
syncitium in other animais. In rare places there could be
seen portions of the central sinus with an area of no more
than 70X 40}1. The histiocytes have a frothy cytoplasm and
extraneous inclusions. There were few true lymphocytes but
many eosinophils with strongly segmented nuclei.
Common porpoise No. 3. This dolphin, like No. 2 also,
was the freshest of our material and therefore the structure
of the nodes should be the most typical that can be obtained
under conditions of captivity.

299
The mesenteric lymphatic nodes were studied. The
cortical and medullary layers had the normal Proper relation-
ship, but spherical follicles were not found. The trabeculae
were dense and thick; in these there were found more smooth
muscles and elastic fibres, than were found in the preceding
animals. The sinuses were filled with a fibrous mass and
cells, which were similar to endothelial cells. All of the
arteries and veins are of the same caliber. Arteries witbout
a lumen, with a closed cavity. They resemble the arterial
sleeves of the spleen, which end blindly.
The pancreatic lymphatic node is complex: it is com-
posed of several nodes of regular form. Secondary nodules of
zig-zag form are present, but there are also spherical nodules
in which proliferation centers are evident.
The bottlenose dolphin. The overall structure of the
nodes is similar to that described above fvr the common por-
poise. A distinction is found in the finer branching of the
trabeculae and the richness of the musculo-elastic base in
these. The trabeculae surround the secondary nodules and cel-
lular cords. In cross-section the trabecula is composed of
several concentric layers of connective and musculo-elastic
tissue.
PinniPeds. The harbour seal. The meSenteric lymph
nodes form a "packet". The form of eàch node Is regular: the
spherical secondary nodules may be arranged in a crowded man-
ner in the surface portions of the node; in the center is an
extensive portion of medullary substance, The capsule of the
node is relatively thin. The trabeculae do not branch exten-
sively, they are thin, composed of connective and muscle tissue,

300
- with a moderate content of collagen fibres and a very
poor elasticity. Both the central sinus of the node as well
as all of the other sinuses are free of any fibrous elements.
The reticular cells form a weak syncitium within these sinuses.
There are many monocytes, as well as myelocytes and typical
plasma cells with a "cap-like" nucleus and strongly basophilic
cytoplasm. In some sinuses there is a mass of free erythro-
cytes.
The pancreatic lymph node was studied. The parietal
sheet of the capsule of the node is abundantly furnished with
vessels. This comprises the branched venous "rete mirabile"
of the seal, described earlier on other. organs (Sokolov, 1959;
Harrison and Tomlinson, 1956).
In the secondary nodules there can be seen the proli-
feration centers. In the medUllary cords there are many medium-
sized and large lymphocytes. Every secondary nodule on some
sections is enclosed by a trabecular membrane. Within the
sinuses there are found erythrocytes; - There are also cells
phagocytizing the erythrocytes. The latter fill the cytoplasm
of the cells.
Steller's sea-lion. The axillary lymphatic node is
of the regular form, but the secondary nodules may be somewhat
indistinct. The capsule of the node is very thick, in it, as
well as in the trabeculae, there are very many large collagen
fibres. In some places the collagen acquires basophilic pro-
perties. Individual collagen fibres may be encountered also
in the sinuses of the node. Elastic fibres are:distinCti.ni'. ,
the walls of the trabecular arteries.

301
Pile. 6. M.c3e1tTeponabilmit zumcparmecKnet y3e1 cnnytia Ueirrpaablibiri cuilyc
I — cimyc; 2 — mfaannbte tutlypbt; 3 —TpaGerzyma.X8
Figure 6. Mesenteric lymph node of Steller's sea-lion. Central sinus
1 - sinus; 2 - medullary cords; 3 - trabeculae. x8.
The thick and coarse-fibred trabeculae of the node
form large loops (Figure 6), as a result of which the Sinuses
are free for the lymph, both in the middle of the node as well
as in the region of the hilus. The cellular composition of
the sinuses is the saine.
The fur seal (adult male No. 3). The mesenteric lymph
nodes are of the regular form: the follicles lie around the
periphery. The form is spherical. The capsule is very thick,
but the trabeculae are not as thick as in the Steller's sea-
lion. They do not permeate the whole node and do not surround
the follicles. The collagen content in the trabeculae is more
moderate than in the trabeculae of the nodes of the Steller's

302
sea-lion. In the capsule of the node there were many sclero-
tized vessels. Fatty sections were found in the nodes. The
sinuses of the node are wide and free. Free lymphocytes and
eosinophilic leucocytes were noted.
CONCLUSIONS
To account for the structure of the retia mirabilia
E. Slijper (1962) put forward the proposal that in the thora-
cic cavity of the dolphin there is developed a high pressure,
which is considerably higher than that in the abdominal cavity
and other parts of the body. In addition, the arterial pres-
sure on the walls of the arterioles of the network from the
side of the heart is also great (Nagel, Morgane, McFarland and
Galliano, 1968). In such-a case the presence of the double
elastic membrane in the walls of the arterioles would be un-
derstandable. The poorly developed tunica externa of these
vessels and the exceptional paucity of collagen are, in all
probability, conditioned by the extreme lability of this struc-
ture.
The arterioles of the rete are surrounded by lymphatic
vessels; in the normal organ they are narrow, collapsed, while
in edema they are dilated. A similar picture is found in the
lungs of the dolphin, where the lymphatic vessels enclose, as
it were with a sheath, the pulmonary veins (Baudrimont, 1955),
which have a great capacity for contraction, since they carry
arterial blood and, possibly, regulate its flow. In this case
a heavy tunica externa of the vessel*is superfluous, while the
surrounding lymphatic vessels facilitate movement, playing the

mirabilia is, most probably,
animals are held in close and
in the end, to a deformatiun
303
role of a hydraulic shock-absorber. This shock-absorber may
also Protect the rete mirabile from the aspect of the intra-
thoracic pressure.
The fact that in one and the same animal the walls of
the vessels of the rete mirabile in the abdominal cavity are
less elastic than those of the thoracic cavity may, to a cer-
tain extent, demonstrate a difference in Pressure in the men-
tioned parts of the body. Besides the natural pathology
(pneumonia, arteritis, helminthoses etc.), the loss of elasti-
city by the vessels of the retia
a common phenomenon when aquatic
shallow enclosures. This leads,
of the vessels and to death.
The vessels of the venous rete mirabile, taken by us
from the lumbar region, in all probability are not typical.
They should be studied on more extensive material.
In seals under water the activity of the heart is
slowed down (Harrison and Tomlinson, 1962; Elsner, Franklin,
Citters and Kenney, 1966). By vasoconstriction and peristal-
tic contractions the peripheral vessels may set up an extra
pressure in these conditions. Among such vessels may be in-
cluded also the mesenteric artery, which possibly functions
even more intensively in this respect in the seals, which is
indicated by the abundant longitudinal musculature in its wall.
The active operation of this artery as a "peripheral heart"
organ (Zavarzin, 1939) should be expressed more clearly in
Steller's sea-lion than in the other seals and dolphins. This
morphological feature is probably connected with the peculia-
rities of the ecology of this species of seal.

304
• The mesenteric artery supplies both the intestine and
the lymph nodes which lie in the mesentry of the small intes-
tine. Undoubtedly there exists some sort of relationship bet-
ween the structure of the vessel and the organs which that
vessel supplies..
Slijper (1962) wrote that whales have an exceptionally
large number of very large lymph nodes. Rakhimov (1968) des-
cribed 7 groups of nodes in the dolphin. Not long ago a de- 191
tailed study was carried out of five lymph nodes in Black Sea
dolphins (Moskov, Schiwatschewa and Bonew, 1969).
In the Caspian seal Kurdyumov . (1962) noted a small
number of large lymph nodes. We found 18 groups of lymph nodes
in seals and a somewhat smaller number in dolphins. There
is no doubt however as to the difference in the size of the
nodes in the two groups of mammals (Table 2). On account of
what structural characteristics and tissues are the nodes of
seals enlarged in comparison to the nodes of dolphins? In
the first place, apparently, this is due to the free dilated
sinuses and lymphatic vessels, which is not found in the dol-
phins, where the sinuses are narrow and filled with densely-
fibrous tissue of the musculo-elastic type. In addition, the
nodes of the seals, and especially of Steller's sea-lion, are
exceptionally rich in connective tissue (in the form of large
loose collagen fibres), which is scarcely noted at all in the
dolphins. A third cause of the differences is the non-uniform
disposition and composition of the lymphoid tissue and its
disposition in the nodes. In the dolphins the nodes are most
frequently of an irregular structure, the cortical substance

305
does not form the usual spherical follicles but is concentra-
ted in the zig-zag shaped sections, which occupy various po-
sitions in the nodes, including also a central position. The
medullary portion of the nodes is considerably reduced.
There is little.typical lymphoid tissue in the form
of small lymphocytes within the nodes of dolphins, it is fre-
quently replaced by plasmocytes and eosinophils. Such a pic-
ture is observed both in dead animalsas well as in those that
have been specially narcotized. Histiocytes predominate in
the sinuses. Our observations are also confirmed by other
data ( Moskov, Schiwatschewa and Bonew, 1969).
The nodes of seals are richer in actual lymphoid tis-
sue, but here also there is a completely different picture from
that found in the nodes of the majority of terrestrzal mammals.
Within the sinuses of the lymph nodes of both dolphins
and seals there are frequently present free erythrocytes and
eosinophilic leucocytes. In the seals here there are found
macrophages with erythrocytes in their cytoplasm. Slijper
(1962) observed, under a microscope., the process of the break-
ing down of the erythrocytes in the lymph nodes.of the dolphin,
and in connection with this he proposed that the nodes partial-
ly take on the role of the spleen,in this process. However
the size of the spleen, according to our data, is larger 4n
those animals with larger lymph nodes. For example, the length
of the spleen in fur seal No. 3 was 4.3 cm, in the harbour seal
- 23 cm, in the bottlenose dolphin - 7.5 cm and only 2 cm in
the common porpoise.

306
The role of the mesenteric nodes in the deposition of
lymph has been mentioned more than once in the literature
(Bouviere and Valette, 1933; Horstmann, 1952; ghdanov, 1952;
Rusn'yak, Fel'di and Sabo, 1958). It is possible that in the
seals a large part of the lymph is deposited in the lymphatic
nodes, while the lymph nodes of dolphins, which have a great
contractine4 capacity in their trabecular base, may regulate
the movement of the lymph.
REFERENCES
1. Zhdanov D. A. The general anatomy and physiology of the
lymphatic system. Leningrad, Publ. Medgiz (State
Publishing House of Medical Literature), 1952.
2. Zavarzin A. A. A course in histology and microscopic
anatomy. 5th edition. Leningrad, 1939.
3. Ivanova E. I. The morphology of the lymphatic system of 192
the Russian desman. Trudy Khoperskogo gosudarstvennogo
zapovednika (Transactions of the Khoperskii State
Reservation), issue 4, 1970.
k. Kurdyumov N. A., The forms of the efferent lymphatic
pathways of certain animals in a comparative-anatomical
interpretation. Trudy Odesskogo meditsinskogo instituta
(Transactions of the Odessa Medical Institute), vol. 16,
1962.
5. Rakhimov Ya. A. The thoracic duct of mammals. Trudy
Tadzhikskogo meditsinskogo instituta (Transactions of
the Tadzik Medical Institute), vol. 94, 1968.

30?
6. Rusn'yak I., Fel'di M. and Sabo D. The physiology and
pathology of the lymphatic circulation. Budapest,
1957 (translation).
7. Sokolov A. S. Characteristics of the size and disposition
of some of the blood vessels in aquatic (seal) and
terrestrial (dog) mammals. Arkhiv anatomii, gistologii
émbriologii (Archives of Anatomy, Histology and Em-
bryology), 37, No. 12, 1959.
9. Baurdimont A. The structure of the pulmonary veins and
functional circulation of the lung of the common dol-
phin, Delphinus delphinus L.
13. Horstmann E. The functional structure of the mesenteric
lymphatic vessels.
15. Monro A. Comparison of the structure and physiology of
fish. Translated and annotated by J. G. Schneider.
Leipzig, 1787.
16. Moskov M., Schiwatschewa T. and Bonev St. Comparative
histological studies on the lymphatic nodes of mammals.
The lymphatic nodes of the dolphin.
19. Rouviere H and Valette G. The role of the lymph nodes
in the circulation of the lymph.
20. Slijper E. F. Comparative anatomy and systematics of
the Cetacea.

, 3 0 8
- JIHTEPATYPA
1 }Kaatioa ,IL. A. 06utan aturromuu H dnisno,norns: minulumotecuoil enure:Au. II., Mearna, 1952.
2 Sana patin A. A. Kypc rucToaoruit u muKpocuornmecxort auaTomiln. 113/t. 5-e. JI., 1939. . - • . 3 Ileanoaa E. PI. K mop(I)onorini allM(1)HTIPICCIZOÙ ClICTCNIbl pyecKori abtxyxc,in. .. • • lpy,i1A honepcicoro row,. aanoneAli., HUH. 4, 1970.
1.1.1(ypihomoa IL A. (1)0pm:A OE:1301511101x .7iiimdm -rmiecmix lire neKoTopmx >titiBoTimix u cpannuTeabilo-ana -rominiecKom ocoemeitint. Tpy,am OHCCCHCWO me,u.. lin- Ta , T. 16, 1962.
5 Paxu.moa 51. A. rpya,noil upoToK maeizonirraionnix. Tpy.alit Taa.uniNcKoro Nteil. WI-Ta, T. 94, 1968.
0 Perlb.r/K. H., cheizbau M.,.Ccr5o ,r7. (1)113nozonin Ii naToaormtemulioo6paine- muf. ByAmicwT, 1957 (iiepenoR). •
7 CoKome A. C. Oco6einiocTit Kaill6pa ii pacnpeaeâeium ItexoTopmx uponenoc-:lux cocyp,on y . no•rnimx (Tioaenb) il na3eminAx (co6amt) mneNoutrraiounix. Apxlin one.. i'lleT0.1. II Dm6pitomr., 37, ,N2 12, 1959.
• bi Bran K. .1■ I., Murdayglt H. V., Millen Jr. J. E., Robin E. D. Arterial constrictor res panse in a divin,,(Y Mammal. Science, vol. 152, no 3721, 1966.
9 Baurdinzotzt A. Structure des veines pulmonaires - et -circulation functionele du poumon du dauphin Common. Delphinus delphinus L. Bull. de friicrosccipie Apoliquce. 2 Serie. T. 5, no 5 et 6. 1955. . 1 0 Elsner R., Franklin D. E., Van Citters R, L, Kenney D, W, Cardiovascular Defense against Asphyxia. Science, Vol. 153, no 3739, 1966. 11 Harrison R. J., Tomlinson I. D. W. Observatibris on the venous system in . scitain Pinnipedia and Cetacea. Proc. Zool, Ces Soc. London, vol 126, 205, 1956. 12 Harrison R. J. and Tomlinson J. D. W. Anatomical .and physiological adap-Ohms in diving matnrnals. Viewpots in biology, 2..1963.
,
13 Horstmaruz E. Llber die funktionelle Struktur der mesenterialen Lymphgela–iCe.. .,, lorph. Jahrb. 91. 483, 1952. . . 1 e Irving L. Scholander P. F., Grinnel S, W, The. regulation of arterial blood pressuve in the seal during diving. Amer. Physiol. 135. 557, 1942,
, 1illonro A. Vergleichung des liaues und der Physiologie der Fische. Uebersegt FillC mit Bernerkungen versNien von Schneider, J. G. Leipzig. 1787. ,.....
, 10 Aloskort M., Schiwalschezea T., Bonev St. Vergleichshistologische Untersu- chimg der Lymphlmoten der Seg"er. Die Lymphlmoten des Delphins, Anat. Anzg.
' U.)j., N 1, 1969. . J. 7 Nagel E. Z., Morgane P. Z., McFarland W. L. Galliano R. E. Rete mirabile of Dolphin. Its pressure—damping effect on cerebral circulation. Sciense, vol. ,16k no 3844, 1968. lbh - Onzinaney F. D. The vascular Networks (Retia mirabilia) of the Fin—Whale (Balaenoptera physalus). Discovery Reports, 5, 1932. 19 Rouvicre H., Valette G. Role des ganglions lymphatiques dans la circulati- e de la lymphe. 13011. Acad. Med. 110. 189. 1933. '
, ZOSlijper E. F. Die Cetaceen vergleiehend-anatomisch und sYsternatisch. Ca-el, zoologica, VII. 1936. .
lijper E. F. Whaies, Hunchinson of London. 1962. . .. 22 Tyson E. Phocaena or the Anatomy of a Porpess, .dissected at Gresham Colledge. London. 1680. , . . .

309
UDC 599.5 A. S. Leontyuk
PECULIARITIES OF THE DEVELOPMENT OF THE BLOOD-VASCULAR 193
SYSTEM DURING THE EARLY EMBRYOGENESIS OF CETACEANS-
The aquatic mammals differ from the terrestrial mam-
mals by several characteristics, associated with the adapta-
tions to the aquatic medium and, above all, with the periodic
arrhythmic breathing and with the blood circulation. In the
blood-vascular system of the cetaceans there are known several
adaptations, which serve for providing the tissues with an
uninterrupted supply of oxygen. Many authors have noted the
considerable development and peculiarity of the vascular ple-
xuses in cetaceans, that are disposed from the base of the
skull along the thorax and form the so called "retia mirabilia".
Also characterized by having a considerable development in the
cetaceans is the subcutaneous network of blood vessels (B. A.
Zenkovich, 1938; A. G. Tomilin, 1946, 1951; V. E. Sokolov,
1958, 1963; V. M. Berkovich, 1961; Utrecht, 1958, and others),
while in the refion of the flippers there have been described
specific complex vessels (A. G. Tomilin, 1946-1962). To the
latter is alloted an important role in the processes of thermo-
regulation.
The adaptations of the cetacean embryos to supplying
the tissues with oxygen, in connection with the peculiarities
of the respiration and blood circulation of the mother, as was
noted by P. A. Korzhuev and N. N. Bulatova (1951), present an
"even greater puzzle". According to the data of these authors,

310
in cetacean embryos there is noted a high oxygen capacity
of the blood and a high concentration of haemoglobin, repre-
senting an adaptation to unfavourable conditions for embryo-
nic development. It is natural to assume that such adapta-
tions are not the only ones,and are reflected in se-
veral morphological charac -Éeristics of cetacean embryos. In
this connection we conducted studies of the vascular system
in cetacean embryos and directed our attention to adaptations
for ensuring a regular supply of oxygen to the tissues.
Serving as materials for these studies were complete
series of sagittal and transverse sections of cetacean embryos:
of the sperm whale-from 8.5 to 105 mm in length, of the
humpback whale - from 11.5 to 125 mm, and of the fin whale -
from 54 to 73 mm, stained by the Bielschowsky-Buke method, the
Nissl method and with haematoxylin-eosin. The value of silver
staining methods for revealing vascular networks has been con-
firmed on more than one occasion in the laboratories of V. V.
Kupriyanov, D. M. Golub, D. A. Zhdanov and others. The mate-
rial for the study was obtained thanks to the cooperation of
V. A. Zemskii, G. A. Budylenko and D. D. Tormosov, staff mem-
bers of the Atlantic Research Institute of Marine Fisheries
and Oceanography.
The results of the study show that the formation of
the vascular plexuses in cetaceans already occurs during very
early stages of embryonic development. At the same time the
vascular plexuses are formed in their most clearly expressed
form around the brain and spinal cord and around the spinal
ganglia. In a sperm whale embryo 14.5 mm in length, along the
periphery of the spinal cord there was developed a network of

311
•
01
thin-walled capillaries, filled with the formed elements of
the blood. Spreading out cranially, this network surrounds
the developing brain and initially is more clearly expressed
in the region of the posterior cranial fossa at the base of
the brain. The enrichment of the vascular networks proceeds
very intensively, and in the sperm whale embryo 16.2 mm in
length there appear at the base of the skull a considerable
number of large thin-walled veins, surrounded by a network of
finer vessels (Figure 1). Then the vascular system in the
cavity of the skull is constantly represented by very exten-
sive plexuses. At the same time there occurs an enlargement
of the diameter of the large venous trunks an an increase in
the number of small vessels in the network. In the older
group of sperm whale embryos (95 - 105 mm) there are found
extensive vascular plexuses in the cavity of the skull, that
are formed of both the large main veins as well as by the net-
work of small vessels of various diameters (Figure 2). The
vascular plexuses attain a considerable development in the
region of the base of the hypophysis, where they form the basis
of the sinus cavernosus. An intensive development of the vas-
cular networks in the region of the brain and spinal cord is
also characteristic of the embryos of the baleen whales. Thus,.
in a humpback whale embryo 95 mm in length the vascular plexus
around the brain was formed of thin-walled veins and arteries, 195-
located in all parts of the skull and and also extending out
onto the outer base of the skull through its openings.
A second zone of the extremely strong development of
the vascular plexuses is the region where the intervertebral
nodes and the initial sections of the spinal nerves are located.

312
pile. I. Cocy.ji+c'roe CII.Iereune 13 acnouauF nt 11epC(la 3a})o1LtIIla Kj•uiano-ra 16,2 srst ;iauuoil. i%l[ispocj>oro. 06- 2. or. 7
Figure 1. Vascular plexus at the base of the skull in a
sperm whale embryo 16.2 mm in length.
Photomicrogaph. Objective 2, acular 7.
Ÿ`.«^na.v..w.vv.«+..Fv.^'^:ii^+:'.l..t... .^. 7^ t•.'.L^:{ F^: ivi.. .:.117^:f13^^.i:.'.:'..^^t.:.:i: àF::i4r.J..:i.t.:.^
^^^ . . ^ ^" - ._. ...... ^.^ `.. . ^^'._ . . . ..... ^
Here there are formed extensive venous plexuses, surrounding
the spinal ganglia. These plexuses, which connect up with
the internal plexuses of the vertebral canal, are considerably
developed in all of the cetaceans (in both the toothed and the
baleen whales), and.are characteristic also of other mammals
and man. Thus, in a humpback whale embryo 95 mm in length
the venous plexuses are formed of blood vessels.of various
diameter, the branches of which penetrate even right into the
ganglia (Figure 3). In the sperm whale such plexuses in the
region of the spinal ganglia are found in an embryo 23 mm in
length, while later they become more and more extensive. A
characteristic feature of the blood vessels which form the
networks and plexuses in the region of the central nervous
system is the very small thickness of the vascular wall of
both thè arteries as well as the veins. This phenomenon,
which was noted in a dolphin embryo in the region of the

Pile. 2. COCYJUICTOC cnoe-re.ime B 0C110-
B8111111 neperia aapo.abuna KaWaJlOTa 97 AIM NI111101 .1, MinipmpoTo. 06. 2, OK. 5 V..
Figure 2. Vascular plexus at the base of the skull in a
. sperm whale embryo 97 mm in length. Photomicrograph.. Objective 2, ocular 5.
31 3
branching of the internal carotid and vertebral arteries by
E. S. Yakovleva (1951), is probably common to all of the
cetaceans and leads to a convergence of the conditions of
gaseous interchange in the arteries and veins.
Vascular networks and plexuses also develop along the 196
course of the large trunks of the cranial and spinal nerves,
where they are considerably expressed. The vascular plexuses
which accompany the spinal nerves are continuations of the
vascular plexuses of the vertebral canal. They are revealed,
for example, in a sperm whale embryo 65 mm in length, in the
form of a dense network surrounding the intercostal nerves.
A similar picture is found in a fin whale embryo 55 mm in
length. in the region of the trigeminal nerve and its branches

314
Par. 3, Cocy.aucToe cuaereane 13 n03a0•
110 ,1/10111 Eatia.ue II ao nepaci)epail rpy1-
111,1X cilainiomoaroabtx yanoe aapownia rop6aToro Rirra 95 .ADI Rau MIII:po-
4/0T0 . 06. 2, olz. 5
Figure 3. Vascular plexus in the spinal canal and along the
periphery of the thoracic spinal ganglia in a humpback whale embryo 95 mm in length. Photomicrograph. Objective 2, ocular 5.
in sperm whale embryos 97 and 105 mm in length therb is formed
an extensive network of vessels, entwining the trunks of the
nerves and giving rise to branches which penetrate into the
trunks. An extensive vascular network is developed also along
the course and within the trunk of the vagal nerves. The vas-
cular networks within the trunks of the vagal nerves are also
manifested in the embryos of the baleen whales: thus, in a
fin whale embryo 55 mm in length they were traced along the
course of the nerves over a considerable distance. The sympa-
thetic trunks are surrounded by developed vascular plexuses,
connected with the plexuses of the vertebral canal. In the
55 mm long fin whale embryo there are here formed wide-looped

• 315 à '
•
networks. The circum-sympathetic vascular plexuses are very
well expressed in the sperm whale embryos. Already in the
early stages of development of the sperm whale (in embryos
14.5 mm in length) along the course of the sympathetic trunk
there is found a network of blood vessels, having thin walls,
which surrounds the sympathetic trunk and its branches. Sub-
sequently, for example, in a 55 mm long sperm whale embryo
there develops around the sympathetic trunk an extensive net-
work of venous and arterial vessels. A similar vascular net-
work is also retained in the adult cetaceans. In the Atlantic
white-sided dolphin along the vertebral trunk there extend two
strands of vascular plexuses, within which are situated the
sympathetic trunks. In their structure these vessels resemble
the structure of vessels with endarteritis obliterans, when
the flow of blood is very considerably hindered (A. B. Khodos,
1967).
Thus, during the embryogenesis of cetaceans (of both
the toothed and the baleen whales) there is manifested in a
regular fashion the formation of the very strongly developed
and widely distributed vascular plexuses, formed by arterial
and venous vessels in the region of the brain and spinal cord,
of the cranial and spinal nerves, and of the sympathetic trunk.
We believe that such a localization of the vascular plexuses
is not fortuitous but is connected with the necessity for en-
suring the supply, in the first place - of oxygen, to the
centers of the nervous system and nerve trunks.
As has been noted by several investigators, in the
cetaceans the vascular networks and plexuses are also consider-
ably developed in other parts of the body. We directed

316
Pile. 4. Tonorpmpitsi DKerpaRapiLiianbiloro cocyaucToro cum:- realm y 3apo2t.butta Eatila.10Ta 55 .A1:11 ,/(,11111Ort:
/ cocyglictoc cluterelime: 2 — ani(Jaitiza JierKoro: 3-- 3a1uIEGINa Ka; 4-- gpoeemmilite cocyRbs 11 liPII7pa.141106 trverwe Tyaoainusa.
pegeutil cpe3. MeKpoilio -ro. 06. 2., OK. 5
Figure 4. ToPography of the extracardial vascular plexus in a sperm whale embryo 55 mm in lengths
1 - vascular plexus; 2 - lung rudiment; 3 - heart rudiment; 4 - blood vessels in ventral wall of body.
Transverse section. Photomicrograph. Objective 2, ocular 5.
cepn-floue-
attention to the richness of the vascular system, especially
of the venous part, primarily in the sub-pleural space of the
.thoracic cavity. L. B. Slavochinskaya (1965) considers that
the abundance of veins is explained by the level of metabolism
in the pleurae: the veins in the pleura participate in the
regulation of the metabolic processes in the thoracic cavity.
Under the parietal pleura in the region of ribs I - III and
the ventral part of the mediastinal pleura, commencing from
comparatively early stages (sperm whale embryos 34-35 mm in
length), there is formed a network of blood vessels, which
takes the form of an extensive plexus. In the region of the
dome of the pleura this plexus passes over onto the mediastinal

Pue. 5. Tonorpacimm 31:erpaRapirmanbito -
ro cocyRucToro cnacTeulta y aapoRbuua i:inuaaoTa 97 01,1(
3a1eam:a cepRu,a; 2— CT I30.1 Ariad)par-maabtioro neprui; 3— cocyttra
naenpm; 4 — cocyAbt, Ha rpyuoil noBep:otucTif Julad,parmbi. CarirrTam.nEtil cpe3
Mutcpoci)o-ro. 06. 2, OK. 5
317
Figure 5. Topography of the extracardial vàscular plexus
in a sperm whale embi'yo 97 mm in length:
1 - leart rudiment; 2 - trunk of diaphragmatic nerve; 3 - vessels of mediastinal pleura; L. - vessels on thoracic surface of diaphragm.
Sagittal section. Photomicrograph. Objective 2, ocular 5.
pleura and extends ventrally to the root of the lung to the
ventral wall of the body, connecting up ventrally with the
plexuses which are situated under the costal pleura (Figure 4).
Caudally a similar plexus extend down onto the diaphragm and
is distributed on its thoracic surface (Figure 5). As a re-
sult of this the ventral section of the thoracic cavity, in
which is situated the heart in the pericardial sac, is found
to be surrounded by the practiâally continuous extracardial
'vascular plexus, which is situated on the surface of the peri-
cardium. In its structure this network is composed of vessels
of two types: smaller iressels with a gradually forming wall

318
(of the arterial or venous type) which anastomose extensively
among themselves; the second type of vascular structures has
the form of thin-walled lacunae, characteristic of micro-
circulatory adaptations for increasing the capacity of the
vascular stream (V. V. Kupriyanov, 1969). In our preparations 198
these were found to be empty or only partially filled with
blood. Venous sinusoids of this type are considered in the
literature as a part of the cavernous-like formations (A. N. 199
Tikhomirov, 1967). Both types of vessels appear comparatively
early and can be distinctly found in embryos 55 and 65 mm in
length and larger. They are more markedly developed in embryos
97 - 105 mm in length (Figure 6). It should be noted that
the vascular network in the region of the parietal pleura is
distinguishable by its content of vessels of the arterial and venous
type. Characteristic of the mediastinal region, on the other
hand, is the presence of a large number of venous lacunae and
cavernous-like formations alongside the rich network of arterial
and venous vessels. It is interesting that this region in other
mammals is also distinguished by having a characteristic struc-
ture of the vascular system, which has been noted by several
authors (V. V. Kupriyanov, 1969; R. M. Petrova, 1967; L. B.
Slavochinskaya, 1967). Well expressed and widespread extra-
organic vascular plexuses are found in the region of the heart
and especially along the course of the large vessels which run
out of the heart, and further along the course of the aorta
and its branches (Figure 7). These pericardial and periarterial
venous plexuses anastomose extensively with the extracardial
vascular network in many sectors.

319
•
46
fin whale. IE
55 m771 in length we also find a fairly considerable network of 200
blood vessels, though this is more localized, primarily in the
dorso-cranial sectors of the thorax. This network is formed
mainly of arteries and veins. In the mediastinal pleura the
vascular plexus is considerably poorer and is represented by
a considerably less well expressed network of arteries and veins.
Pac, G. CTpyhTypa si.c•rpaxapaEEa.qi,uoïl cocY.aucToii ceriE sapoabi-wa KaA1aOTa 105 Arar ;1,-iuiEOïr.
I- cocy,ai+crbie nahyma; 2- CCTb TOIfKOCTC111i1.IX cocY;Zos. 91Eir.p04)0T0.06. 2, os. 5
Figure 6. The structure of the extracardial vascular net-.
work of a sperm whale embryo 105 mm in length:
1 - vascular lacunae;2 - network of thin-walled vessels.
Photorricrograph. Ubjective 2, ocular 5.
Thus, the disposition of the vascular plexuses in the
thoracic cavity of the sperm whale embryo is distinguished by
the marked peculiarity of the topography and the character of
the blood vessels. Within the thorax of the embryos of the
baleen whales (humpback whale, fin whale) the picture of the
disposition of the vascular networks differs markedly from
that described above. In the subpleural layer of a

I.
S)
Pic. 7. CocyglicToe clue:twine B o6Jtac - , Tit jyrn aopTbt 3apoluatia uall.laaoTa
97 it creitKa aoprm• 2 — cocymicroe cimere7-
Hue. Mulzpo(loTo. 06. 2, ow. 5 •
32 0
Figure 7. Vascular plexus in the region of the arch of the
aorta of a sperm whale embryo 97 mm in length: • 1 - wall of aorta;
2 - vascular plexus.
Photomicrograph. Objective 2, ocular 5.
Among the ecological characteristics of both the baleen
as well as the toothed whales, which have been noted by many
authors (A. G. Tomilin, S.E. Kleinenberg, V. M. Bel'kovich,
V. E. Sokolov and others), belongs the considerable develop-
ment of the network of subcutaneous vessels. The vascular
pattern can be seen distinctly through the integument of the
embryo, commencing from the earliest stages of development.
The architecture of the vascular pattern is especially well
marked out in the region of the head, the base of the flippers,
the base of the tail and the trunk. Only the development of
the pigment layer of the skin gradually conceals this vascular
pattern. With histological studies the development of vascular
networks is found at the base of the flippers and around the

• • 32 1
Pile. 8. lloRimmulan cocyRucTan cen, B 06.1111CTIL GOKOBal c-remui Ty.nounuut aapubmia KannknoTa 105 .11.11
MilKp04)0TO. 06. 2, ()K.' 5.
Figure 8. :Subcutaneous vascular network in the region of
the lateral wall of the trunk of a sperm whale
embryo 105 mm in length.
' Photomicrograph. Objective 2, ocular 5.
phalanges of the fingers, along the course of the bundles of
muscle fibres and onder the skin of the flippers (sperm whale
embryos 72 - 105 mm in length). Extensive vascular networks
are formed under the skin also in the muscle layers of the
trunk (Figure 8).
The data from our studies demonstrate the considerable
richness of the vascular system in cetacean embryos, especial- 201
ly of the toothed whales (sperm whale), manifested in the de-
velopment of vascular plexuses in various parts of the body,
mainly in the zones where vitally important organs are situated:
the central and peripheral nervous systems and the heart. In
this connection, naturally, the question arises as to the bio-
logical significance of such adaptations, their role in the
development of the embryo and, subsequently, in the functions

322
of the adult animals. The concept exists in the literature
that the vascular networks and plexuses in the body of ceta-
ceans fulfil an important, if not fundamental, role in supply-
ing the tissues with blood during diving, functioning as a
store of blood that is rich in oxygen and shortening the path-
way of the blood to the tissues (B. Howell, 1930; F. Ommaney,
1932; A. Krog, 1930; P. Schollander, 1940 - cited by S. E.
Kleinenberg, 1946).
Sharing this point of view, S. E. Kleinenberg empha-
sizes that during diving there is a decrease in the number and
strength of the cardiac contractions and, consequently, the
vascular store permits the blood reserve of oxygen to be more
economically expended and facilitates the work of the heart
during prolonged apnea. Our data permit one to consider that
such adaptations to a prolonged shut-down of external breathing
and the creation of stores of oxygen in the blood are formed
already during the early stages of embryonic development. It
should be borne in mind, however, that the marked development
of vascular plexuses in the region where the centers of nervous
regulation are situated is characteristic not only of the ce-
taceans but also of other mammals, including also man (A. S.
Ieontyuk, 1969), which are not characterized by having exten-
ded periods of apnea. Therefore it is possible to somewhat
expand the concept of the role of the vascular plexuses, which
are formed already in the embryos of the cetaceans and are
retained in the adult animals.
The character of the extra-organic, mainly venous,
plexuses in the region of the brain, spinal cord and peripheral

323
nerves is similar to those in tissues with an increased ex-
change (A. B. Miodos, 1967), since for obtaining the optimal
exchange the rate of the blood-flow should be reduced to a
minimum. It is probable that the developing vascular plexuses
are adapted in their structure primarily to the relative
hypoxia and moderate metabolic acidosis that is so character-
istic of developing embryos (I. A. Arshavskii, 1960; L. S.
Persianinov, 1967, and others). That this is actually the
case_is convincingly indicated by the data of several.authors
on the effect of hypoxia on the structure of the vascular
system (V,. I. Voitkevich, 1958'; E. N. Domontovich, 1958; E.
Van-Lir and K1. Stiknei, 1967; V. V. Kupriyanov, 1969, and
others ), which is manifested in an increased vascularization
of the tissues, a hypertrophy of existing vessels and a growth
of newly formed vessels. The resulting hyperemia leads to an
increased diffusion of oxygen, food substances and metabolites.
The changing conditions of the regional circulation of blood
favour a more complete assimilation of the oxygen in the blood
and the excretion of metabolic products, since the diffusion
of the oxygen from the blood is related to the area of the
diffusion and the partial pressure of oxygen in the venous
part of the vascular flow (A. I. Shik, 1966). V. V. Kupriyanov
(1969) emphasizes that the length of the venous pathways,,_the
diversity of their directions and the extensive roundabout
connections predetermine the pattern of the microcirculatory
system and, in a decisive manner, influence the hemodynamics
in the organs. Convincing data on the considerable reconstruc-
tion of the intra-organic vascular stream in cases of hypoxia

324.
under high mountain conditions were presented by Ya. A. Rakhi-
mov and L. E. Etingen and co-authots (1968).
Thus, the extra-organic network of blood vessels, main-
ly of venous•vessels with a retarded blood flow, which appears
in the early stages of development of cetaceans may be consi-
dered as an adaptation for supplying blood to the tissues in
the specific conditions of the intra-uterine development of
the foetus. In this connection the development of the special
vascular structures during the embryogenesis of the cetaceans
becomes understandable, and these may be considered not only
as storage structures in places with a maximal requirement for
oxygen but also as adaptations for the more complete assimila-
tion of the oxygen in the blood, and for the excretion of meta-
bolites in the zone of the disposition of the vitally impor-
tant organs and systems - the nervous system and the heart.
Taking into consideration the conditions of the life of the
toothed whales and the baleen whales, the more clearly expres-
sed development of the vascular plexuses in the toothed whales,
as compared to the baleen whales, is understandable as corres-
ponding to their ecology. It is evident that the adaptive
characteristics of the vascular system, which are set down in 203
early embryogenesis, retain their significance and also, pro-
bably, acquire new properties in the adult animals.

J111TEPATYPA a 1 Apumacutiii H. A. Onsno.Tiorun RpoBoo6pamemin BO nnyTpuprpotillom nepli- .
0A,e. M., 1960. . Beizbuoauct 13. M, 0 (1)11311 11CCKOrl Tepioperyantuni y 6e.aymi. Tp. conem. lixTit-
o onniecxoil KONHICCIIII AH CCCP, inn 12, 1961. •
Ban-Jl up 3., Cruuneii /(4. FlinoNcuil. M., 1967. • . Botiruearm B. H. 13.rinsinue xponinecKoro uncoopo,altoro r000.aantio na Epo-
Bocna6:Kenno mosra. 13 iiii.: eclmaiionorint ii.naTo.norun abixamin, rimontin II OK-
curenoTep arum». KIM, 1953. 5 flo.uonroami E. H. HoKoTophte cIntsuo.noriniecmte mexannambl nplicnoco6.roului
opraiinsma K 111C.10POD,110à 11C;I,OCT11T0 1 1110CT11. B mi.: «.(1mi3no.norim ii narolorlin „a‘pixailitii, rnimmin H oacitronoTepaniut». Kum 1958. b 3enuoomi E. A. TemnepaTypa Te.na KIITOD. ,IJ,o1(.riaa.b1 AH CCCP, T. 18, Bun. 9, 1938. • • 7 i'l eat lOaa E. II. Attain-mu:le oco6ennocTil ' xponenocnoil 11 Rb1X11. 1. 0:11,110ii Cl ic -
-rem pycciwii nbixyxo.nit. B vii.: «1op4loaorittiecuile oco6ounocTii BOP,111,1X mneHoini-Tammilx». M., «Haymi», 1964. • 8 Haanoaa E. II. Honoe e COCT051111111 «npecuoii CCTII» ,u,epuBwroplibix mexamis-
MOB y nenoTopmx rionyBoailux H■ 11130THIJIX. LI,olulaa..1,1 AH CCCP, T. 172, D1,111. 2, 1967. . . •
%. Ii &MOO(' E. II. OCO6e1/110CTII COCY,IIICT0ii CHCTC:111>1 131U1: 11/1X 14.1C1:01111T111011111X.
I Bcecomslioe cc-mental-me 110 liaplenum mopcmtx M.1C101111TD10111.11X. M., 1969.
1 9 Kneiinen6è pe C. E. M•neKonwraimune Lleplioro H 1-‘30eCKOTO Itropeii. M., 1956. 1 Kopmyea H. A., Bynaroaa H. H. Oco6ennocTil HbrikITC.11b110r1 (llyilmilin Bponit
rem6puolion ituitalunia. 2.1,0;1. AH CCCP, T. 81,.131,111. 1, 1951. . 12 Kynpunnoa B. B. rlyrn mithpouitpxyonium. Kilumnen, 1969. 13 J1 COHTION ii. C. Pa31111THC nponenocin,ix cocyaon rpy,aitoro oTiteaa crmintoro
•Nicara II cnitimomosronidx Y:30011 D 9M61/110TCHC3C. B ii i.: <1P1111111C1/11:111.119 11 penac- By.impitaanitn nnyTpenitux opranon meToRom opralionencini». Muncx. 1969. • •
14: Ilepcum".,,... JI. C. Actl,,‘,iin ruioa,r; mi nonopozz -..leinioro. -M.. 1967.
15 Reptant! C. B., COK0.108 A. C., Tomumin A. r. Mopilm-cliyumutolia.-ibilbie oco-CiennocTu apTepliii it Ben Ima131111KOB .H.eabiliinion B C1151311 C .101011101111C11. 1V BCCCO-
103110C C011ellt HO nayii. mopcmix 1\1.1/e1:01111THIO111,11X. M., 1969. . 16 Herpona P. M. K nonpocy o TonorpatImil II minepBaumi me.anacTintaabunii Tuienphi. MaTepliambi OrleT11. liompop. Munemoro me.a,uncTwryTa aa 1963 r. MIII1CK,
1967. .
17 Paxumoa 57. A., .9TUFIZeti JI. E., Benuun B. III., Yemanoa M. Y. Mopqm.aoriin snyTpeinnix oprairon D ycsnorninx nbicoKoropbri. Ryinan6e, 1968.
18 cAaaotimicKast .11. B. HCKOTOplaIC 13031M1CTIlble oco6entiocTil mitupotuipBymiTop-ublx erpyKTyp riplicTeito ■ mon rocupbt ttemneBa. B KII.: 4:Mop(1.fflorin-tecmie oco6eil-11 oon minwoulipBy.nmuni». M., 1967. • 19 Coicoma A. C. Ocori -ennocTii Kaoli6pa II pacime,rtenemin IICKOTOpbIX Bponeitoc- MAX COCYJI.OB y D0111,1X (T10.11C.111.1) H nasemnbix (CO6D1:11) NI.IHKOMITH101.11.11X. Apxnn anaTomun, rucTo.nornu u 3m6piumormi, T. 37, II1,111. 12, 1939. .
20 co/CO400 B. E. Mexamism Teryworaatut y mopcmix maelkonuraioutux. Buria. MOI,M, T. 63, Bun. 4, 1958. •. . . 21 COK0.408 B. E. FucTorenea KOH:II 1111L18.110TH H cluntaana. BCCTHIIIL MFY, 1313111.
5, 1963. . 22 TI1X0.4111poo A. H. K Bonpocy o BnyTpuoprannom cocyancTom nvc•ie nottemlofi ..noxamm. B un.: KM0p(1/0.10C11 11,CCKlIC OCIIOBIA muhpowipt;y:inium». M., 1967.
_ - . 2 3 Tommuti il. F. KrituaooT Kamnarcicoro Mopu. 3oo.n. 1,Kypir., T. 15, nntri, 3,1936, 2 . Tomnun A. r. Topmopery.innuin H reorpaduniecktie pact-A BuToo6pa3imix. 4c . AH CCCP, T. 35, limn. 5, 1946. .. . 4 ToAtimuti A. F. 0 Tepmopery.armitll y ICHT006P113IIIIIX. «HpupoRa», i\i'm 6, 1951. 2o TWIllAtili A. F. KuToo6pa3iib1e CCCP 11 nputaexcainux cTpall, T. IX, - ii3A-130
AU. c,CC1, 1957. . 7 4 amuntin A. F. kiiT006 1m3ilide (1/11Y111.4 111011CrI CCCP. 113n-no AH CCCP, 1962.
2 xoaoc A. B. Minwocxoniniecmiii 111111:1113 113111CIICHIHI B COCYJUICTOM pycle (bac-ulna rollout!. B un.: «Mopclmoortniecxne oco6onnocTil muBpounpxy.annun». M., 1967. 29 Nun: JI. JI. Kacoopo,:uthiii aaupoc u Eposooilpamenue. B KII. C.KIIC.10p0,11,111e1
ReÀKiini opramisma n ero peryituponanne». KIICH, 1966. ..)u Huomneaa E. C. 13 1I3nwnie 11 CTe0e1Ille 1 1V2WC1106 CCTII OCHODHIHISI 'neperia ine-gginiTaiomitx.113necTun H AH PC<DCP, Belli. -35, 1951. „II Barnett C. 11., Harrison R. J., Tomlinson D, W, Variations in the venous uktem of mammals. Biol. Rev., vol. 33,4. 1958. •
iu " Bulrimont A. Sur la signification du double roseau capillaire respiratorice de la elosion interalveolaire du pournon (des niaminiferos maritius et . Fevolution de ces animax. Arch. ana t. histol, et embruol, v, 42, N I--4. 1959. j3 Harrison R. J., Tomlinson D. W, Anatomical arid physiological adaptations. in,divigg mammals. In Vienuoints in 13iology. vol. 2, London. 1063-
325

326
3 14 M .Orgaile P. J., Nage! E. L., Galliano R. Z„ Viamonte M, Angiographie and anatomical study of the blood supply to the brain of the dolphin, Tursiops trutt-psalus. Anat. Record. vo!: 157, 1967- . . . j) °mummer' F. D. The vascular networccs (retia mirabile) of the fin whale
. lgiesol/tieurperphysaltts). Discovery Repts. vol. 5, 1932. cistoa .. P F., Schev i ill W. E. Counter current vascular heat exchange n
he,,fins of whales. J. Appt. Physiol. vat, 8, no 3, 1955. , y Slijper E. I.. On the vascular system of cetacea, Proc, Intern, Congr, Zoo!,
J.Ïb-1.11, Sect. III. 1958. . ..10 Suckline I. A„Stickling E. E., Walker .4.. Suggested function of vascular bundles in the limbs of Periodicticus potto. Nature. Engl. 0 vol. 221, no 5, 178.
• Se . Vianionie M., Morgane P. I„ Galliano R. E., Nage! E. L. McFarland. Angio-graphy in the living dolphin and observations on bladdy supply to the brain. ftn.lerican Journ. of Physiology. Vol. 214, no G. 1968.. • L'f'u Wain:skit R. Some observations on. the vascular systems of a female fetal fjeack. Camegie inst. -Wash. Pub!. 496. 1938 , 14.1 Wilson Il. S. The relu mirabile of the narwhal. Journ. Anal. Physiol, Vol, 14_
• c'( • ' Utrecht W. L. Temperatarregulierende Gerasysteme anderceepider-
maleAtrukturen bei Cetace.e.n. Zool. Anal, vol. 161, N 4. 1958.
REFERENCES
1. Arshavskii I. A. The physiology of the blood circulation
during the intra-uterine period. Moscow, 1960.
2. Bel'kovich V. M. Physical thermoregulation in the beluga.
Trudy soveshchaniya Ikhtiologicheskoi komissii AN SSSR
(Transactions of the Conference of the Ichthyological
Commission of the USSR Academy of Sciences), issue 12,
1961.
3. Van-Lir E. and Stiknei Ml. Hypoxia. Moscow, 1967.
4 , Voitkevich V. I. The effect of chronic oxygen starvation
on the blood supply to the brain. In the books "The
physiology and pathology of respiration, hypoxia and
oxygen therapy". Kiev, 1958.
5. Domontovich E. N. Some physiological mechanisms of the
adaptation of an organism to to an oxygen deficiency.
In the books "The physiology and pathology of respira-
tion, hypoxia and oxygen therapy". Kiev, 1958.

327
16 6. Zenkovich B. A. The body temperature of whales. Doklady
AN SSSR (Reports of the USSR Academy of Sciences),
vol. 18, no. 9, 1938-
7. Ivanova E. I. Adaptive features of the circulatory and
respiratory systems of the Russian desman. In the
book: "Pûorphological characteristics of aquatic mammals",
n,oscow, Publ. "Nauka", 1964.
8. Ivanova E. T. New findings on the state of the retia
mirabilia and derived mechanisms in some semi-aquatic
animals. Doklady AN SSSR, vol. 172,- no. 29. 196?.
Ivanova E. S. Characteristics of the vascular system of
aquatic mammals. 4-th All-Union Conference on the Study
of Marine Mammals. Moscow, 1969.
46 10. Kleinenberg S. E. Mammals of the- Black Sea and the Sea
of Azov. Moscow, 1956.
11. Korzhuev P. A. and Bulatova.i1. N. Characteristics of the
respiratory function of the blood of dolphin embryos.
Doklady AN SSSR,'. vol. 81, no. 1, 1951-
12. Kupriyanov V. V. Idcrocirculatory pathways. Ki.shinev,
1969.
13. Leontyuk A. S. The development of the blood vessels in
the thoracic section of the spinal cord and spinal
ganglia during embryôgenesis. In the book: "The re-
innervation and revascularization of internal organs
by the method of organopexy"'. .Minsk, 1969.
14. Persianikov L. S. Asphyxia of the foetus and newborn.
Moscow, 1967.

328
15. Pershin S. V., Sokolov A. S. and Tomilin A. G. The
morpho-functional characteristics of the arteries and
veins of the fins of dolphins in relation to locomotion.
4th All-Union Conference on the Study of Marine Mammalu
Moscow, 1969.
16. Petrova R. M. The question of the topography and inner-
vation of the mediastinal pleura. Materialy otchetnoi
konferentsii Minskogo medinstituta za 1965 (Papers
presented at the 1965 annual reports conference of the
Minsk Medical Institute). Minsk, 1967.
17. Rakhimov Ya. A., Étingen L. E., Belkin V. Sh. and Usmanov M. U.
The morphology of the internal organs under high mountain
conditions. Dushanbe, 1968.
18. Slavochinskaya I. B. Some age-class characteristics of
the microcirculatory structures of the parietal pleura
of man. In the book: "The morphological characteris-
tics of the microcirculation". Moscow, 1967.
19. Sokolov A. S. Characteristics of the size and disposition
of some of the blood vessels in aquatic (seal) and ter-
restrial (dog) mammals. Arkhiv anatomii, gistologii
émbriologii (Archives of Anatomy, Histology and Embryo-
logy), vol. 37, no. 12, 1959.
20. Sokolov V. E. The mechanism of heat transfer in marine
mammals. Byulletin Moskovskogo obshchestva ispytatelei
prirody (Bulletin of the Moscow Society of Naturalists),
vol. 63, no. 4, 1958.
21. Sokolov V. E. Histogenesis of the skin of the sperm whale
and fin whale. Vestnik Moskovskogo Gosudarstvennogo
Universiteta (Reports of Moscow State University), no. 5,
1963.

329
22. Tikhomirov A. N. The question of the intra-organic
vascular flow of the renal pelvis. In the book: "The
morphological characteristics of the microcirculation".
Moscow, 1967.
23. Tomi.lin A. G. The sperm whale of the Ka.mchatka Sea. 204.
Goologicheskii zhurnal ( Zoological Journal), vol. 15,
no. 3, 1936.
24. Tomilin A. G. Thermoregulation and geographical races
of cetaceans. Doklady AN SSSR ( Reports of the USSR
Academy of Sciences ) , vol. 35, no. 5, 194•6.
25. Tomilin A. G. Thermoregulation in cetaceans. "Priroda"
("Nature" ), No. 6, 1951-
26. Tomilin A. G. Cetaceans of the USSR and neighbouring
countries. vol. IX, Publ. USSR Academy of Sciences,
1957-
27. Tomilin A. G. Cetaceans in the fauna of the seas of the
USSR. Publ. USSR Acad. Sci., 1962.
28. Khodos A. B. Microscopic analysis of changes in the vas-
cular flow of the tibial fascia. In the book: "The
morphological characteristics of the microcirculation".
Moscow, 1967-
29. Shik L. L. Oxygen requirements and the blood circulation.
In the book: "The oxygen regime of the organism and
its regulation". Kiev, 1966.
30. Yakovleva F. S. The development and structure of the
retia mirabilia at the base of the skull of mammals.
Izvestiya APN RSFSR ( Proceedings of the Academy of
Pedagogical Sciences of the RSFSR), issue 35, 1951•

4. 330
32. Baudrimont A. The significance of the double network of
respiratory capillaries in the interalveolar septa of
the lung of marine mammals and the evolution of these
animals.
42. Utrecht W. L. Temperature regulating vascular systems
and other epidermal structures in the Cetacea.

■ ••
I 331
UDC 599.5 599.745
P. A. Korzhuev
THE PROBLEM OF THE OXYGEN RESERVES IN THE ORGANISM 205
OF AQUATIC MAMMALS
One of the most weakly elaborated problems of the bio-
logy of aquatic mammals is the problem of respiration, or more
precisely, the problem of the shutting-off of the external res-
piration during the period of diving. Up to the present time
in the literature there are present contradictory data on the
time spent by the animals under water. While the time of the
shutting-off of external respiration during diving in the diving
birds is measured in minutes, then for the aquatic mammals this
period is measured in tens of minutes, and for the large whales
- even in hours (Korzhuev, 196 )4). According to Pastukhov's
data (1969), the Baikal seal may spend more than an hour under
the water. It is true that this record duration of stay under
water, equaling one hour and eight minutes for one of the test
seals, was observed by the author only once, and later these
seals shut-off their breathing for only 5 - 8 minutes.
The author of the present paper haopened to be a wit-
ness to an experiment, conducted on a young seal in the Northern
Caspian in September of 1965, when after a forced stay under
water of 7 minutes the seal had a very lamentable appearance.
Experiments on the shutting-off of breathing in the Black Sea
dolphins, the white-sided dolphin and the nyalix±hm bottlenose
dolphin (Korzhuev and Bulatova, 1952), when the animals were
on dry land, showed that the white-sided dolphin calmly endured
such an experiment for only two minutes, while the bottlenose

332
dolphin endured this for four minutes. After the expiration
of this period there commenced a very violent reaction, accom-
panied by vomiting, which was.indicative of a very obvious
disturbance of the organism. It may be assumed that, if these
experiments had been conducted in water, the duration of the
experiment may possibly have been increased by a factor of two
or three, but at any rate for the white-sided dolphin this
would not be more than 6 minutes, while for the bottlenose
dolphin it would not exceed 12 minutes. Visual observations
indicate periods of about this order.
Apparently the periods of shutting-off breathing for
the pinnipeds are more prolonged than for the small cetaceans,
though these also measure 15 - 20 minutes (Mordvinov, 1969).
Set somewhat apart is one of the Antarctic species of seals,
the Weddell's seal, which in one experiment dived to a depth
of 600 m and stayed under water for 43 minutes and 20 seconds.
As to the representatives of the large whales, the du-
ration of the stay under water of which is measured in hours
(sperm whale - up to 75 minutes, bottlenose whale - up to two
hours, Irving, 1939), these data are purely visual and there-
fore require confirmation. Apparently, the problem of the
prolonged diving of aquatic mammals will obtain an effective
resolution only when a thorough quantitative determination is
made of those reserves of oxygen which the organism of these
animals has at its disposal.
It is scarcely possible to consider as convincing the
arguments of those investigators who consider that the duration
of diving is related to the capability of the animals to change
over to the anaerobic type of respiration during the period of

333
i diving. It seems to us that the basic condition affecting the
duration of diving and, by the same token also, the cutting-off
of breathing for some period or other is the presence of oxy-
gen reserves and their rational utilization.
However the problem of the oxygen reserves in aquatic
mammals has been very weakly elaborated. On this subject there
are more assumptions than precise facts. As an illustration
of this, one may point to the studies of Scholander (194•0) and
Robinson (1939)s who determined the oxygen reserves in cetaceans
and pinnipeds, for which determinations they assumed that the
probable amount of blood in the organism of the dolphin, for
example, equalled 15.0% of the body weight or that the quanti-
tative characteristics of the body musculature could be taken
as being equal to 35.0% of the body weight, although these
values were not precisely determined.
In this connection it-may be noted only that for the
terrestrial animals also the problem of the oxygen reserves is
one that has not been worked out at all. Only the attitude that
in these animals these reserves are negigible may.be considered
as indisputable.
This fact is often obscured, since during the process
of the evàlution of life on our planet the problem of the re-
serves of various vitally important substances has been resolved
very successfully. In different stages of the ontogeny of
various groups of animals there are formed reserves of proteins,
fats, carbohydrates and vitamins: it is known that the egg
cells of various animals (fishes, amphibians, reptiles, birds,
insects) contain the entire assortment of substances necessary
for normal development; for the period of hibernation

L
3314 •
heterothermal animals lay down reserves of fat, attaining up
to 35.0% of the body weight, while in the aquatic mammals the
fat reserves may be as high as half of the body weight.
However oxygen, which is one of the most important
elements, without which the organism cannot manage to exist,
is practically not stored at all by the organism. Terrestrial
mammals, including man, may cut-off their breathing for only
one to two minutes. This circumstance is quite incomprehensible
since oxygen reserves could protect the organism from many un-
desirable and sometimes also catastrophic consequences.
Apparently there exist some very weighty reasons which
prevent the setting up of considerable oxygen reserves within
the animal organism; the small 'group of aquatic mammals and
birds forms a certain exception to this rule.
To decypher the specific mechanisms, determining the
possibilities by which animals may shut-off their breathing
for some period or other, is a problem which still awaits its
solution. All the same, with respect to the terrestrial verte-
brate animals it may be said that one of the most fundamental
causes, preventing the setting up of oxygen reserves, is to be
found in the properties of oxygen itself.
The fact is, as was shown by Professor A. L. Chizhev-
skii (1960), that it is not a matter of indifference as to
what air we breath. Up to the present time it has been con-
sidered that for respiration it is important that the air should
have in its composition a certain amount of oxygen, nitrogen
and carbon dioxide; however not only the chemical composition
of the air turns out to be important but also the physical

335
state of its molecules and, in the first place, of the oxygen.
Among the factors determining this physical state, the first
place belongs to electricity: the saturation of the air with
electrical charges and the degree of its electrostatic charging,
as a consequence of which the oxygen molecules become ionized.
Particularly important for respiration are molecules of oxygen
having a negative electric charge. If the molecules of the air
are not ionized, then animals rapidly die in such an atmosphere,
in spite of the presence of oxygen molecules in the respired
air. These investigations of A. L. Chizhevskii shed light on
the causes which determine the absence of any considerable re-
serves of oxygen within the organism of animals.
A characteristic feature of the ionized oxygen mole-
cules is the fact that they •exist for a very short period of
time, of the order of ten minutes or a little more, after which
the charge is lost and the oxygen reverts into the inactive
form. This circumstance, apparently, is the fundamental cause
which deprives the animal organism of the possibility of storing
the ionized oxygen, which has a negative charge. The negative-
ly charged oxygen ion possesses several characteristics, such
as, for example, the presence of nine electrons (as in fluorine)
and a changed spin of its nucleus. 4parently these physical
changes, occurring at the level of the electrons and of the
nucleus, play a decisive role in the evolution of the animal
world, as a consequence of which also there is found the absence
of considerable reserves of oxygen even in the higher represen-
tatives of the vertebrate animals.

336
This is a very fundamental circumstance, which has not
been adequately considered by biologists. Nevertheless the
properties of the basic chemical elements, which play an im-
portant role in the life of animals, determine the character
of their evolution. Practically all of the characteristic
features of the organs of aeration and of the haemodynamics
are determined by the properties of oxygen.
As to the aquatic mammals, it seems that here there
occurs a deviation from this principle. It cannot be ruled out
that in the case of the marine mammals there may be some kind
of supplementary adaptations, directed towards overcoming these
properties of oxygen, as a consequence of which there may be
set up within the organism oxygen reserves that are more power-
ful than is characteristic of the terrestrial mammals. However,
until precise data are obtained on the actual periods of the
cutting-off of breathing in the large cetaceans and pinniPeds
in their natural habitats, there are no groundsfor speaking
of deviations from this principle. •
The experimental data that are available at the pres-
ent time permit one to say only that a real- basis for the pro-
longed cutting-off of breathing is the presence of certain oxy-
gen reserves, i.e. the retention of the aerobic type of respi-
ration. And this means that without a precise knowledge of the
oxygen reserves it is impossible to defenitively resolve the
question of the dtiratIon of diving.
UnÉortunately there are very few precise data available
on the amount of oxygen that is found within the organism of
animals. In Table 1 are presented data an the oxygen reserves 1W)

337
Table 1. Oxygen reserveè in the organism of man, certain
mammals and birds, cm3 (Robinson, 1939; Scholander, 1940; Korzhuev, 1964)
'Ta6onua 1
Pe3epnbi nitcnopoAa n oprannme qemonena, IICKOTOpbtX. wienounTaimunx M I1TMU, Cat 3
(PoGlincou 1939; Ilionann,ep, 1940; Kopniyen, 1964)
1 2 3
5 6 7
Kucao-
Beer° pona
"Tea- mtcao- Ha 1 KZ
Blut 13cc, K2 JlerKite Kponb albunnia ttenatt p„,a. Ben
Species We igit Lungs Blood Muscles *"a""h
' kg A. B. U.'
linunam 70000 300000 1400000 1400000 250000 3350 48,0
ByThIJJK0110C 1400 6000 45000 54000 4000 109 78,0
Reni.,(Inui 19 250 600 180 60 1,09 57,0
Moment, . 29 500 1100 '270 100 1,52 52,4
TIOJICIlb 70 . 545 2055 2530 245 5,37 77,0
1-1enoneic 70 900 1160 335 245 2,64 38,0
flinirnun 5,6 120 90 40 20 0,27 48,0
A. - Inter-tissue fluid B. - Total oxygen & 1 C. - Oxygen per 1 kg -body weight, cm3
1 - Fin .whale 2 - Bottlenose whale . 3 - Dolphin 4 - Seal 5 - Seal 6 - Man
Penguin
in the organism of certain representatives of the mammals and
birds. For these the structures of the organism that contain
oxygen are the organs of rspiration - the lungs; the blood,
as a liquid tissue, contains physically dissolved oxygen, as
does also the inter-tissue fluid. However these structures
have a relatively small part of the oxygen. The main portion 209
of this is contained in the form of chemically bound oxygen
jal the respiratory pigments of the blood and muscles in the

338
form of oxyhaemoglobin of the blood and muscles. At least,
in the large cetaceans the portion of oxygen contained in the
organs of aeration and tissue fluids in the form of physically
dissolved oxygen comes to about 10 - 15%, while the portion
of oxygen bound by the haemoglobin of the blood and muscles
comprises about 84% in the fin whale, about 90% in the bottle-
nose whale, from 85 to 90% in seals, and about 70% in the
dolphin. In man the respiratory pigments bind about 55% of
the total oxygen that is present in the organism. It should
be borne in mind that, in spite of the relatively large amount
of oxygen that is bound by the haemoglobin of the blood and
the muscles, in man, for example, the tissues utilize a com-
paratively small amount of this., since the venous blood of man
contains much unutilized oxygen (about 12 - 14 percent by vo-
lume). It is true that such a character of the utilization
of oxygen has its advantages, since the pressure gradient of
oxygen in the capillaries of the body is very high, which
favours a more effective supply of this to the tissues.
In this respect the cetaceans and pinnipeds occupy a
special place, since they utilize the oxygen of the arterial
blood more completely, so that in the venous blood there re-
mains a very small amount of this, of the order of one to two
percent.
Thus, a characteristic feature of the adaptation of
the cetaceans and the pinnipeds is their higher capability
for removing practically all of the oxygen from the blood,
in contrast to the terrestrial mammals.

339
0 If it is considered that the aquatic mammals find
themselves under conditions that approximate to a state of
weightlessness and, consequently, expend practically no energy
for supporting the weight of their own body, then it becomes
understandable that the energy losses in these are considerably
smaller than in their terrestrial relatives. It is very
strange that this circumstance has been completely ignored
in comparisons of the characteristic features of the biology
of aquatic and terrestrial mammals, since in actuality it is
very essential.
Some investigators turn their attention only to the
fact that aquatic mammals may shut-off their breathing for
more prolonged periods, as compared to the terrestrial mammals;
however the rest is accepted as being quite adequate to ter-
restrial conditions. It is considered that the energetics of
the organism are completely the same in both terrestrial as
well as aquatic conditions. However it is precisely in this
that there is a basic difference between the aquatic and ter-
restrial mammals.
Of course, a very important form of.adaptation is the
lowering of sensitivity of the respiratory center to the accu-
mulation of carbon dioxide in the blood. The behaviour of 210
the vascular system in the aquatic mammals at the time of
diving (the redistribution of the blood supply.to various
parts of the body) is completely different from that in the
terrestrial animals. It seems to us, however, that the cause
of this is not the necessity of ensuring a supply of oxygen
only to the vitally important organs (heart, brain) at the

34•0
expense of other parts of the body, which might seemingly be.
deprived of the necessary amounts of oxygen, at the same time
changing to an anaerobic type of respiration, but something
quite different.
From our point of view, the crux of.the matter lies
in the fact that this redistribution of the blood supply
during the period of diving occurs only because in the aquatic
conditions the requirement for oxygen is smaller than under
terrestrial conditions.
The correctness of this assumption finds confirmation
in the fact that the aquatic mammals, in'contrast to the ter-
restrial mammals, possess many times greater reserves of haemo-
globi.n, localized both in the blood and in the muscles of the
body. It is quite obvious that the large amounts of haemo-
globin are the main condition of the creation of the oxygen
reserves during the period of diving. Evidently the expendi-
ture of these reserves is more economical than is the case
under terrestrial conditions, where energy is expended not
only for locomotion but also for supporting the weight of the
body itself.
In other words, life under terrestrial conditions is
linked with the necessity of surmounting the forces of the
earth's attraction, or the forces of.gravitation, and this
surmounting presupposes losses of considerable amounts of ener-
gy, while under aquatic conditions, as a consequence of the
high density of the wate.r, the effect of the gravitational
forces is practically eliminated, although the gravitational
field is retained in the water, as on dry land. The data pre-
sented in Table 1 confirm the stated positions.

341
It may be thought that, if the cetaceans and pinnipeds,
in the process of their adaptation to the aquatic conditions
of existence, had changed over to a gill-type of respiration,
i.e. if they could obtain the oxygen directly from their sur-
rounding medium, as is the case in the fishes, then they would
not have such large reserves of haemoglobin and, by the same
token, of oxygen.
The fishes, as primarily aquatic animals, are charac-
terized by having a very small amount of blood in the organism
and a small amount of haemoglobin per unit weight of the body.
Thus, for example, in fishes on average there are about two
grams of haemoglobin per kilogram of body weight, while in dia-
dromous fishes, which accomplish extensive migrations to their
spawning sites, generally against currents, the degree of pro-
visioning of the organism with haemoglobin comprises about 3
grams per kilogram of body weight. It is true that in addition
to this there is present in these a small amount of physically
dissolved oxygen, contained in the blood and perivisceral
fluids, while in some fish species there is present a small
amount of oxygen that is bound by the muscle haemoglobin.
However, these additional amounts scarcely change the total
figures named above.
If one compares with these figures the degree of pro-
visioning of the organism with haemoglobin in the dolphins
and seals, then it is found that in the white-sided dolphin
this value is approximately equal to 25 g of haemoglobin per
kilogram of body weight, while in the seal it attains a level
of 40 g. This is almost 15 times greater than in fishes.

34,2
o These huge reserves of haemoglobin, required by cetaceans and
pinnipeds for binding oxygen for the periods of diving, would
be superfluous with the gill-type of respiration.
However, since in nature there is found the principle
of the irreversibility of evolution, first discovered and
formulated by the French paleontologist Louis Dollo in 1893,
the change to a gill-type of respiration is impossible. The
cetaceans and pinnipeds are forced to employ the lung-type
of respiration, with all of the consequences arising from
this. This means that the entire evolution of the aquatic
mammals was directed, in the first place, to providing for
one of the most basic functions, th function of respiration.
From this derive: the change in the rhythm of respiration,
the lowering of the sensitivity of the respiratory center to
the accumulation of carbon dioxide in the blood, the redistri-
bution of the blood-supply to the organs during the Period of
diving, and the marked stimulation of the synthesis of hà.emo-
globin not only in the bone marrow but also in the muscles of
the body. And all of this occurred so that it would be pos-
sible to do without breathing during the period of diving.
The aerial type of respiration turned out to be the only
thread which ties in the cetaceans, the group which has moved
furthest in its evolution to the aquatic way of life, with the
terrestrial animals.

34'3
JII'ITE.PATJ'PA
1 Koporcyeo 17. A. remor7o611n. CpaB11nTC7bnan 6noXiotilfl n 4p113110:lorltR.
•allayxa», 1964.2 Kop.mycc+ 17. A., Eqnaroea H, H. jlblaaTCAbnan fiynxunn xpoBn Ae.ab^nnoB
7py7b1 f1nccruryTa mop(Ilo;lorlut :i<liBOTnbix n;,t. Ceuepuoua AH CCCP, Bbin. 6,1952.3 hlopoaur{o(i 10. E. )JA11Ti':1bilOCTb npc6binannn uop, nolIoi! xacnlli'ICxnx Ttoae-
'ueïl. TC311cbt Aor,7a;tOB 4-ro IiceColo3noro coBentannfi no Diol)cxnSr nLlexonllTalO-7I nAt , Ka•aninlnrpan, 1969.
^^ 17(lC7IfX06 B. /^. Hei:OTOpblc Pe3y7hTaTbi na6.nto;tennH nap. 6a}(xaJ1bCt<oÏ1 nep-
n01i B yC.70BI15i\ 31<Cliepll\1e11Ta. Te3ncbl Aoi:.,laltos 4-ro I3Uc0i03110ro co8enjaili(llmo niopci<nm maicl<onirraioil.tu>t. 1Kaaltnnlirpaa„ 1969.5 yra;ccacs:uü A. Jl. Aapouonnc)nxaunsi B napo;tnoM xo3Ni'tcTBe. Ai., I'ocn.naiina-
A T, 1960;^ Harrison S. Experiments with diving Seals Nature. 188, 1960.`z Irving Z. Respiration in diving A'lammals Physiol, Rev., 19, 1939.tj Robinson D. 'Hie muscle hemoglobin of Seals as an Otygen Store in diving.
:Science, 90, 1939.g• Scholander P. Experimental Investigation. on the respiratory function in di-
-ving niaimnals and Birds. Hvalradets Skrifter; no 22, Oslo. 1940.
REFERENCES
1. Korzhuev P. A. Haemoglobin. Comparative biochemistry
and physiology. Moscow, Publ. "Nauka", 1964.
2. Korzhuev P. A. and BulatovaN. N. The respiratory function
of the blo.o.d of dolphins. Trudy Instituta morf ologii
zhivotnykh im. Severtsova AN SSSR (Transactions of the
Severtsov Institute of Animal Priorphology of the USSR
Academy of Sciences), no. 6., 1952.
3. Mordvinov Yu. E<. The duration of the stay under water of
the Caspian seals. Reports of Proceedings of the 4-th
All-Union Conference on marine Mammals, ' Kaliningrad,
1969.
4. Pastukhov V. D. Some results of observations on the Baikal
seal under experimental conditions. Reports of Pro-
ceedings of the II•th All-Union Conference on Marine
yammals, Kaliningrad, 1969.
5. Chizhevskii A. L. Aeroionification in the national economy.
T,loscow, Gosplanizdat (State Publishing house for Litera-
ture on Planning), 1960.

UDC 599.537
T. N. Glazova
344
QUANTITATIVE EVALUATION OF THE HAEMATOPOIETIC FUNCTION 212
OF THE SKELETON OF THE BLACK SEA DOLPHINS
It is considered that in the terrestrial vertebrates
the function of the skeleton consists of supporting the body
in a particular position, and also ofstrengthening and retain-
ing the musculature. In the aquatic vertebrates, according
to the opinion of E. Slijper (1962), the function of the ske-
leton that consists of supporting the body does not have a
great significance, since in the water the body is acted on
by an upward pushing force, which alleviates this function.
gle)
The importance of the latter circumstance is also emphasized
by P. A. Korzhuev (1963), who notes that the organism in the
water is as it were suspended and therefore its energy is ex-
pended only for locomotion, while under terrestrial conditions
the animal must expend considerable energy not only for loco-
motion but also for supporting the.weight of its body.
In both the aquatic and terrestrial conditions one of
the basic functions of the skeleton is the haematopoietic
function. From this point of view there is a special interest
in the study of the weight characteristics of the skeleton and
bone marrow of adult dolphins, as animals which shut-off their
breathing for prolonged periods, and also in the study of dol-
phin embryos, which are subjected to difficulties in acquiring
oxygen not only in particular stages of their development but
also at the time of the shutting-off of breathing by the mother.

345
. In the literature there exists a fairly considerable
amount of data on the weight of the skeleton of the large
cetaceans (Khar'kov, 19•0; lérochkov, 1953; sleptsov, 1961;
Bjarnason, 1954). These data are not precise and at times
are greatly exaggerated in connection with a lack of a thor-
ough cleaning of the skeleton. Probably, more precise data
on the weight of the skeleton of the large cetaceans are to
be found in E. Slijper's (1962) study. Characteristics of
the weight of the skeleton of representatives of the small
cetaceans have been presented in the studies of P. A. Korzhuev
and co-authors (1964, 1967). In these studies there are data
on the weight of the bone marrow in the adult cetaceans and
their foetuses, and also on the. weight of the skeleton of the
latter.
The aim of the present study was to show up the weight
relationships of the skeleton and bone marrow of the developing
embryos and sexually mature Black Sea dolphins.
There were studied embryos of different ages and adult
individuals of the bottlenose dolphin (Tursiops truncatus Mouf),
of the common dolPhin (DelPhinus delphis L.) and of the common
porpoise (Phocaena phocaena L.). The skeleton and bone marrow
was examined in 7 embryos of the . bottlenose dolphin, with a
weight of from 184 g to 16.5 kg and an age of from 4-5 to
11-12 months*; 4 adult individuals e . weighing from 30 to 67 kg,
and 10 embryos of the common dolphin weighing from 2.65 to 7.00
kg and aged 8-10 months; 11 adult individuals of the common
porpoise weighing from 13 to 47 kg and 11 embryos weighing from
* The age of the embryos was determined with an accuracy of to 0.5-1 months on the basis of the length of the body.

346
884 g to 1.996 kg and an age of 5 - 8 months, and also one
embryo weighing 3.07 kg aged 9-10 months.
For obtaining the weight characteristics of the ske-
leton, after the weighing of the whole body there were removed
the muscles, tendons, brain and spinal cord, and the cleaned
skeleton was weighed in sections. The volume of the bone mar-
row was determined by a method based on Archimedes' principle
(Petrov, 1952). For calculating the weight of the bone marrow
there was employed the specific weight of this in only the
sexually mature dolphins, in which it was possible to find
individual sections of cylindrical form in the ribs.
Knowing the specific weight and volume of the bone
marrow, its weight was calculated as a percentage of the body
weight, and the percentage relationships in the different
sections of the skeleton were also calculated.
According to the literature data (E. Slijper, 1962),
in the large crustaceans, such as the sel whale, fin whale and
blue whale, the weight of the skeleton comprises corres .ponding-
ly 13, 16 and 17% of the body weight. In the Black Sea dol-
phins, according to the data of P. A. Korzhuev et al. (1964),
the weight of the skeleton was half as great.
On thorough treatment of the skeleton it was found
that its weight relative to the weight of the body comprised,
on average, 6.5 in the common dolphin and 5.2% in the common
porpoise (Tables 1 and 2). In the Quantitative characteriza-
tion of the skeleton of the two species of dolphins there were
not found any statistically significant sex differences.

347
The axial skeleton takes up the main part of the ske-
leton of the dolphins. Thus, this comprises about 90% of the
weight in the skeleton of the common dolphin, while the ske-
leton of the limbs comprises only 10%. The relationship of
the parts of th skeleton does not remain constant: with in-
creasing weight and length of the dolphin (for example, of the
common porpoise) there is found a decrease in the relative
weight of the skull of from 32.6 to 24.0% of the total weight
of the skeleton, and an increase in the weight of the verteb-
ral column of from 46.0 to 53.7A (Table 2). It la interesting
that the relative weight of the ribs and sternum remains at .
almost the saine level.
While the fraction taken up by the anterior girdle and
free fore limbs in the common porpoise and the common dolphin
is 9 - 10, rin-i2ze the fraction falling to the posterior limbs,
which in dolphins are reduced to two short narrow bones lying
within the muscles, is measured in tenths of a percentage point.
The bones of cetaceans are filled with bone marrow. •
E. Slijper (1962) notes that the skeleton of young animals
contains red bone marrow but that with age there occurs a
replacement of this fraction by the yellow bone marrow.
According to our data, all of the studied dolphin in- 216
dividuals, including also the sexually mature individuals,
possessed only the red fraction of the bone marrow, i.e. the
haematopoietic fraction. The weight of the bone marrow in
the adult dolphins comprises about 2A of the body weight
(Tables 3 and 4). No sex differences were found in the weight
of the bone marrow in the dolphins. The bone marrow is loca-
lized identically in both species of dolphins. The greatest

iength Te2a,
noa •
Sexi cm oo fe" ut. r o
COI.umn
ljeee4 hind
rrepeliniii I 3azilux rore gepena skull
rleepand ,rpyzintst
sternum
44,8
44,1
48,7
eJ
46,4±1,2
10,7
9,8
10.5
10,5
10,4±0,2
6,9 6.9
6,2
6.7
6,5 ± 0,2
32,1
28.9
26,5
27.7
28,8±1,2
12,3
15.6
11,5
13,5
13,2 -±0,9
0,1
0,2
0,1
0,3
0.1-±-0,05
Table 1. The relative weight of sections of the skeleton of adult
, IsD common dolphins, as % of the total weight of the skeleton.
• Ta6au u, a 1
OTHOCHTeJ11,11b1ii BCC OT,Ile.10B CKCJ1CTa B2, p0C.11bIX 06bIKHOBC1111b1X Ae.11:411110B,, % OT e5tuero BCCa cliezeTa 1
TO jULliqadY re.:a, K
W e ght , kg
A • Bc CMC.1 CT2,
Cé CT Beca Ter: a
e"-Ig ul Dec o -r.z.c...:oa cxezeTa.
30,2
41,8
58,3
65,3
7-1-S7C 48,9 ± 7,95
camitaf 128
cameuM 157
camxaf 167.5
cameuM 174
camKa 155,0±10,2 11 CaMeLt f and m.
f - female m - male
A. - Weight of skeleton,:as % of body weight.
1 Translator's not. Sic. Should:be %, rather than kg. t, ;
c5D

6 .1 a 2
• 1: t, , Kg 71-regs--- 1(01! eq:10CTS: •
B .
Bee xpsuna. Ca Ben
body kelet
gepon. skull
Pe6q, ribs stenue
noni cro.163
column fore
nepea.. 'hind.
31.11111X
%04‘, ody
06Temi nec
wjUht, kg
Body length
/Limn rem,
CM
A. Bee cue- ;ICU. !'4 Or Ben
Tela
no.1
Sex
6.3
5,1
• 5,4
• 4,9
• 7,9
5,9 .
5,0
4,9.
4,6
5,2±0,2
0,5
0,3
0,3
0.2
0.8
0,2
0,1 0,2
0,2
0.2
6,2±0,05
7,7
5,5
6,3
3,5
10,5
4,1 9 ,5
3,5
3,8
3,4
4,2
31,2
.31,0
28,9
24,7
32,6
29,8 97,8
27,5
24,2
25,9
24,0
27,5±0,9
11,5
12,1
10,5
11,7
12,0
12,6 13,1 13,7 13,1
12,8
•12,2
12,4±0,3
re . 7,
Table 2. _
I • The relative weight of sections of the skeleton of adult
porpoises, as % of the total weight of the skeleton. common 0.:nocwreobt-rmil sec OT,Ze.,103 cxeneTa e3pocJ1bm mopcmux cBHHe, % OT eGulero Zeca cue-ieTà.
22,00
23,00
26,40
32,00
13,10*
37,45**
38,20"
40,00**
40,00 • •
40,00**
47,00**
xfSx_
34,61±2,54
canut male
» » '
emua . ema_
,»
canieül 129±3
0,9 46,8 9,3 •-.. 0.2
1,2 46,3 8,9 0,5
1.0 49, 9 9,5 - 0.8
1,2 52.9 8,8 0,8
1,2 46,0 ' 8.0 0,1
47.8 8,2 0,2
1,5 49.3 8,1 0,2
1,7 48,3 8,5 0,3
1,6 52,7 8,3 0.2
1,6 50,4 9,0 « 0,3
1,6 53,7 8,6 -
1,3±0,1 1 49,8±0,81 8,7±0,2
114
116
120
127
92 e 133
129
136
136
• 137
138
•
H • , I '
.camica • . m and f • 1lenonono3pezae ocoCh nim cravierinecgoil o6pa60-1- Ne ne
"' Camiza Gepemenue. ymIlThinaeach. •
0,4±-0,1
* Sexually immature individual, not taken into account in.the statistical treatment.
** Pregnant female
A. - Weight of skeleton, as % of body weight.
B. - Weight of cartilage, as % of weightof:
1 Translator's note. Sic. Should be %, rather than kg.

350
amount.ofi this is concentrated in the vertebral columnr ave-
raging 56.7 and 61.6;-) of the total weight of the bone marrow.
The minimal amount of bone marrow is found in the pos-
terior limbs of sexually immature animals. In a young common
porpoise weighing 13.1 kg the pelvic bones contained 0.2%
(Table 1I.). In the rudiments of the posterior limbs of the
sexually mature forms there was no bone marrow. In the skull
it is mainly localized in the occipital bone and is completely
absent in the lower jaw. The average relative weight of the
bone marrow in the skull comprises 20.6 and 15.5%. in both
species of dolphins about 12% is contained in the ribs and
sternum, and also in the bones of the fore limbs.
Before passing on to the description of the weight
characteristics of the skeleton and bone marrow in the embryos
of the Black Sea dolphins, in comparison with the adult forms,
we will give a short account of the regularities revealed for
each species of a particular age.
The greatest period of pre-natal development was co-
vered in the bottlenose dolphin, the embryos of which were re-
presented, on the one hand, by 3-4 month old individuals and,
on the other, by 11-12 month old individuals, i.e. just prior
to birth.
The relative weight of the skeleton of the embryos of
the bottlenose increases, in step with their growth, from 6.75f
in the 3--4. month old embryo to 10.4.0% of the body weight in
the foetus just before birth (Table 5). A positive correlation
is marked between the weight of the skeleton, on the one hand,
and the size of the foe-tus, on the otheri the relative weight

e.,
Z.!
weight of lug,gegpl,akeleton, kg1 andvertebral
peuep H noutouonoro Sen sternum cden
iore hina nepexunx 32AHHX
Length ezima Tua,
of fhdy, cm
A • Cennurt Bec HOCTHOTO
meBra. OT BCC2 Te.7a
30,2
41,8
58,3 65,3
x±Sx 48,9 ±7,95
camn f
cameu,
camxa f cameu,-M
camn CaMeg
f and m
128
157
167,5
174
156,6±10,2
2,0
1,7
1,9
2,2
1,9± 0,1
23,0 14,4
22,5
22,6
20,6±2,1
7,5
16,7
8,2
9,3
10,4±2,1
57,6
M,2
55,5
55,4
56,7 ± 0,7
11,8
10,3
PJ 12,3
12,0±0,7
0,1
0,3 0,2
0,4
0,2 ±0,05
Total body 06Lumg sec
Tem,gz weight, kg
Tion Sex
Table 3. The relative weight of the bone marrow.of adults of the common dolphin, as % of total weight of the bone marrow.
T a 6.1 1,111, a 3
OTHOCHTeablIber BeC KOCTHOr0 mo3ra B3poc.nux O6bIKHOBeliniX aeJ114}11013, % OT 0611.1,CTO Beca KOCTII0f0 mo3ra
f - female m - male
A. - Total weight of bone marrow, as % of body weight.
1 Translator's note. Sic. Should be % of total weight of bone marrow.

Table 4.
The relative weight of the bone marrow of adults of the common porpoise
,: in various sections of the skeleton, as % of total weight of bome 1,1ar1; o^r.
OTHOCSiTCRbHSIH BeC K4CTHoI'0 N.03ra B3110C.?6!Y "f0PCKfiY CBHY,C11 B .^.i137K4!Ib!Y OTAC:SaY CKe.i!eTa, clp CT OG::(efo E^.Ccl.
KOCTHOI'0 M03ra
Total ^ l` ^ iese on, icl 1:OCTt:01'0 bl03:'3 D Lne CrC.boGy I, g th 06ut::i! Bec CC
l^Tl }^SOGutafi sec AnHKa xocrnoro ver-tebralTeTa. c:e ^L,Jra, ?^ I•
c
n03no1roRr:orO xOP.e.liucrel!Te7a aB MOI
o^ ^j^d OT BCC2 TC7a ^CDCIIa pp6ea t rp}-ai+:crp1^e I n 1.nweigh^kg ;jex cm Y1 skull 1 _.^-bs sternum nepC;lc:, 321:131s
22,00
23,00
26,40
32,00
13,10*
37,45
38,20
40,00
40,00
40,00
47,00 **
x±Sx34,61-!-2,64
caaie!1male»
»
ii
casseu HcamKamale
114
116
120
127
92
133
1129
136
136
137
138
129.±2,9 2,0±0,1
and îenl le
13,8
16,6
17,9
13.6
•18,3
18,4
17,7
15,1
13,8
14,4
14,0
15.5.t:0,6
7,5
10,6
8.0
8,1.
8,3
12.3
10,0
8,811,09.2
9,5 ±0,5
1,2
1,9
1,2
1,7
1,3
1,9
2,3
2,8
2,5
2,5
2,0
65,5
59,1
61.0
65,5
62,3
56,7
55,9
60,3
65,3
G0,7
62,8
12,0
11,9
12,0
11,1
9,7
10,8
I1,1
11,6
9,6
11,5
12,0
2,0-?-0,2- 61,6±1,0 1 11,3--+-0,2
0,,8
" Heronoeo3peRas oc06b npit cTaracTKUCCxoi oGpaGoTne R O yti11Tai83naca.Camxa Gepevexaa.
^ Sexually immature individ1.zal,. not considered in the statistical
1
treatment.
Pregnant female
Translator's note. aJc.. Should be ô e
t;

353
of the skeleton increases with the increase in the weight and
length of the embryo. With the growth of the embryo, the re-
lative weight of the skull, which comprises a considerable
fraction of the total weight of the skeleton of the embryos
of the bottlenose dolphins, decreases from 44% in the 4-5 month
foetus to 31% in the 11-12 month foetus, i.e. just before birth.
The largest part of the skeleton consists of the vertebral
column, the relative weight of which increases from 38 to 53%.
The data characterizing the relative weight of the ribs with
the sternum, and also of the girdle and free anterior limbs,
did not display a change with age. It was found that these
parts of the skeleton were about the same in the bottlenose
dolphin embryos of different ages.
The relative amount of *cartilage decreases with the
growth of the embryo. This decrease is manifested most strik-
ingly when the weight of the cartilage is expressed as a per-
centage of the weight of the skeleton. While in the 4-5 em-
bryo weighing 4 kg 55% of the skeleton consists of cartilage,
just before birth in an embryo weighing 15.0 kg the cartilage
fraction comprises only 29%, which values are correspondingly 219
equivalent to 4.2 and 2.8% of the body weight (Table 5).
The bone marrow of the embryos of the bottlenose dol-
phin consists of the active red fraction. In step with the
growth, the total weight of the bone marrow in the skeleton of
the embryos of the bottlenose dolphiruincreases in both abso-
lute and relative terms: the weight of the bone marrow of the
4-5 month embryo comprises 1.7%, while in the embryo in the
last stages of intrauterine development it comprises 4.6% of
the body weight (Table 6).

• 351i-
Table 5 .The relative weight of sections of the skeleton
of embryos of the bottlenose dolphin, as % of
the total weight of the skeleton.
Tafiauua 5
UTIfOCfi7CllbHbiti üCC OrJleJ1OD CKCAC7'a 11a0t,t01f at^aANiib!,
% OT OÛIi(CCO i3CCa CHCJICTa
0,184
4,0
7,0
11,0
12,0
15,0
16,5
B
n o.1
C
AnnuaTe.7a,ru
caNtha^' 21,5
caa+ica^ 65
cahtica^ 95
ca^icqT^l 96
ca^ma 110
camica 120
ca^seü 114
DB03-pacT,MCCR-
1ICü
3--4
4-5
5-6
5-6
11-12
oxoao12
11-12
IiFe Bec X(v^Ita, çdcKC.le- OT BecaTa,
oT d,necÏ. a
a I cxe-TCAa nCTa
6,8
7,3
9,1
9 ,8
10,4
9,8
10,2
4,2
4,3
3,2
3,52,8
3,2
55,2
47,5
31,4
33,8
28,8
31,4
G Bec oTncnon Cr.e^1cr2, ICI,
^YV ^ x nctttcape6ep u utfovauo:co- u rte-ll
yepcna rpy. ru cro- Pcawlx
yllmw n6a Aoue't-ROCTeII
43,7
38,040,036,530,835,0
7,5
6,3
8,9
6,7
8,1
8,7
37,5
49,1
44,2
49,2
52,8
47,5
7,5
6,6
7,0
7,5
8,38,8
A. - Total weight of body, kg LL
B. - Sex
C. - Length of body, cm
D. - Age, months
E. - Weight of skeleton, as % of body weight
F. - Weight of cartilage, as % of weight of:a - bodyb - skeleton
G. - Weight of sections of the skeleton, kg1w - skullx - ribs and sternumy - vertebral columnz - pectoral girdle and fore limbs.
f - femalem - male
1 Translator's note. Sic. Should be %.

• • 355
Table 6,
The relative weight of the bone marrow of embryos
of the bottlenose dolphin in various sections of the skeleton, as % of the total weight of the bone marrow.
Ta6eltua
OTBOCETCJIMIblii uec Koc -ruoro moara nnoRon ati)a,auubt . paaaugubtx ax cxe.neTa, °,10 ov etimero ileca uoc -ruoro moara
A B C D E F Bec ICOCTII0r0 110311 la CIZeileTe. IC2
06tguii 06tuull oec w
BCC R:ulna Bo3pac-r. ROCTI1010
1C1)Y z
lumen H il
. Ten, 110.1 TC./13. ,,,,,,e, moan, ?k; 1103001104- nepeRinix CAt 01 tieca geperia rry _ 110F0 KOI1C.1110C- X2
Tena juintà CT0.1i511 TCii
•
0,184 caMIa 21,5 3-4 _ - - _ -
4,0 CaNIKa 65 4-5 1,7 54,0 9,0 30,3 6,7
7,0 camK‘ 95 5-6 2,4 42,7 4,4 44,3 8,7
11,0 camet 96 5-6 . - - - _ • _
12,0 ca.mx 110 11--12 3,5 31,8 5,7 54,9 7,7
15,5 UMK 120 OKW10 12 3,5 26,4 9,3 - 53,4 - 10,8
16,5 eaiei.fl 114 11-12 4,6 26,1 4,2 58,9 10,8
A. - Total weight of body, kg • B. - Sex
C. - Langth of body, cm
D. - Age, months E. - Total weight of bone marrow, as % of
body weight
F. - Weight of bone marrow in the skeleton, kg i w - skull x - ribs and sternum y - vertebral column z - pectoral girdle and fore limbs
f - female m - male
1 Translator's note. Sic. Should be %.

Bec cKene-Ta, qc
OT ueca Tena
42,7
40,1
38,2
40,0
35,1
37,7
35,1
35,4
28,3
MA
2,65
.3,00
:3,68
• 4,20
4,90
.5,30
.5,60
.6,00
.6,30
7,00
I) BoapacT, uecnoen
13ec xpnuta, 9t, OT neca
al Pce_ Tel a I n„a
(; Bec „tenon cKeneTa,
in pe6ép 110X0. H 1104H0-
`lePena rpy- ro cron- A1111b1 6a
A
lice Teaa,
he
noca II ne-peanux
.xouen-IIOCTeii
non ji.1{I u a Tena,
C.14
8,1
7,1
8,5
7,3
8.6 9,5
9,2
8,2
7,9 I
4,0
3,6
4,6
2,9
4,0
2,8
2,6
3,4
4,5
3,0
43,1
42,9
47,5
33,8
41,4
27,3
26,7
41,2
40,1
34,5
7-8
7-8
7-8
8-9
8-9
oxono 10
9-10
9-10
10
9-10
9,4
8,5
9,6
8,5
9,5
10,2
9,5
8,3
11,2
8,7
5,8
4,7
5,2
5,9 6,0
5,4
5,5
6,4
4,9
7,1
43,5
42,1 .
48,0
46,9. 50,3
47,4
54,4
49,0
58,6
50,0
ca mica ca men ,cameu.
ca men camxa
nmu
c a Wca CO mica camu
camen
66
67
72
79 • 81
84
83
81
90
84
• 356
o Table 7. The relative weight'of sections of the skeleton of embryos of the common dolphin, as % of the total weight of the skeleton
Ta6.-iiin.a 7
OTHOCIITOJIblibla Bee OTACJI011 cKe.nera naogon oeibncoonennoro.p,e,nbcfmna, % or o6inero neca cKeae -ra
A. - Total weight of body, kg
B. - Sex C. - Length of body, cm
D. - Age, months
E. - Weight of skeleton, as % of body weight
F. - Weight of cartilage, as % of weight of: a - body b - skeleton
G. - Wei&ht of sections of skeleton, kg1
w - skull x - ribs and sternum y - vertebral column z pectoral girdle and anterior limbs.
1 Translator's note.. Sic. Should be %.

357
By the 4-5th month of intra-uterine life about 50% of 220
all of the bone marrow of the bottlenose dolphin embryos is
concentrated in the bones of the skull, but towards the end
of intra-uterine development the greater part of the bone mar-
row, comprising about 60%, is localized in the vertebral column.
The weight of the skeleton of the embryos of the corn-
mon dolphin in the second half of their intra-uterine develop-
ment comprises, on average, 9. 14% of the body weight. As the
embryo grows there is found the same pattern of change of the
relative weight of the different parts of the skeleton and of
the cartilage, as occurs in the embryos of the bottlenose dol-
phin. Thus, with the growth of the embryo there can be traced
a tendency towards a decrease in the fraction of the cartilage
in the skeleton: from 4.6 to 2.8% of the body weight or from
47.5 to 26.7% of the weight of the skeleton (Table 7). In a
manner similar to that in the bottlenose dolphin embryos, there
is manifested a pattern of change with age of the growth of
the skull, the weight of which decreases from 46 to 28% of the
weight of the whole seleton with the increase in the size of
the embryo. In contrast to the development of the skull, in
step with the growth of the embryo there occurs an increase
in the weight of the bones of the vertebral column from 42 to
59% of the weight of the skeleton. Age fluctuations of the
remaining bones of the skeleton are insignificant. The weight
of the bOne marrow of the embryos comprises, on average, 3%
of the body weight (Table 8). On examining the data on the
distribution of the bone marrow in the different sections of
the skeleton of the embryos there is found a tendency towards

358
decreasing the amount of this in the bones of the skull (from
55 to 25% of the total weight of the bone marrow) and an in-
crease in the vertebral column (from 32 to 61%). A similar
pattern is found in the embryos of the bottlenose dolphin.
Unfortunately, the absence of material on embryos in the first
months of intra-uterine development, when there occurs the
formation of the bone marrow as a center of the synthesis of
haemoglobin, does not give us the opportunity of characterizing
this. However in the second half of the embryonic development
there is found only a gradual build-up in the mass of the hae-
matopoietic tissue, which to a considerable degree depends on
the individual characteristics of the developing organism.
On the whole, the distribution of the bone marrow in the ske-
leton of the embryos proceeds in such a manner that the largest
amount of this is localized in the vertebral column - about
50%, then in the skull - about 35%, in the bones of the fore
limbs - about 9%, and in the ribs and sternum - about 6% of
the weight of all of the bone marrow.
In the 5 - 8 month embryos of the common porpoise the
average weight of the skeleton, of which half is composed of
cartilage (53% of the weight of the skeleton), is equal to
8.4% of the body weight (Table 9). We note that the weight
of the cartilage in the skeleton of the adult common porpoises
comprises, on average, only 4.4% of the weight of the skeleton.
In the embryos of this age the relative weight of the skull
and of the vertebral column is about the same ( •2 and 44% cor-
respondingly). In the embryo just prior to birth the weight
of the skull skeleton decreases to 31%, while the weight of
the vertebral column, conversely, increases to 54% of the

Table 8.
A B C D E F Bee uocutoro T•1031":1 B cizene-re. et
06uutii 06nut6 BCC w X Y z
BCC RIIIBB licc e.n.r , KOCTUOCO pe6ep notica,
reia, lloa Te.na, ,„'",,,„„ M03 18, 5, 6 ii uo3uoitog- nepen„,,
cm OT ucca gepetta rpy_ Itur0 Kolictuloc-
IC2 re.qa Alum croA6a TCil ,
2,65 cana' 66 7-8 3,2 40,2 5,5 43,1 11,1
3,00 cameun 67 • 7-8 2,2 54,6 3,9 32,3 9,3
3,68 camearl 72 7-8 • 2,9 40,4 6,5 46,5 6,6
4,20 eamen; 72 8-9 2,5 33,5 7,8 54,0 4,6
4,90 calm 81 8-9 2,7 ' 33,9 5,1 50,1 11,0 .I.
5,30 eantKa 84 omviô 10 3,8 28,1 4,7 57,2 10,0
5,60 cMKak 83 9-10 3,9 26,3 6,2 61,2 6,3
(6,00 camxa 81 9--10 2,5 34,4 6,0 , 47,3 12,3
.6,30 camKa 90 10 3,6 27,5 6,2 52,4 13,9
7,00 caMe4 M 9-10 2,7 27,9 7,9 54,3 ' 9,8
`.7
359
The relative weight of the bone marrow of embryos of the common dolphin.in in various sections of the skeleton, as % of the total weight of the bone marrow.
T a 6 a 11, a 8
OTHOCIITC.,11,111:Iii nee nocTnoro mo3ra I1J10)1,0B o6humonennoro fleabcbitila
13 pa31114111AX oT,ncmax cy.eaera R% or o6atero Beca Eocruoro mo3ra
A. - Total weight of body, kg B. - Sex
C. - Length of body, cm
D. - Age, months
E. - Total weirht of bone marrow, as % of body weight
F. - Weight of bone marrow in the skeleton, kg 1 w - skull x - ribs and sternum
• y - vertebral column z - pectoral girdle and fore limbs
f - female m - male
* about 10
■ .
1 Translator's note. Sic. Should be %.

36 0
Table 9.
The relative weight of parts of the skeleton of embryos of the common porpoise, as % of the total weight of the skeleton.
Ta6aaaa 9
OrnocwreabBbtil Bec qacTeii caeaeTa n.noa,oa mopcaoà % OT o6lus ero Beca CKOJICTa
OT BCCa
G Bee oraeAoe cmeaeTa, E?
A 13.- C P03- BeeEeKe- F Bec xpetue, °6
06ani6 Bec " jltaxita pacT, Aera, % ir w x 1.,,
z Ron,' tocren
Tun, IC nOn TC33, NICCA- OT seca a: b ge pena pe6ep . rpyanub: n° ,3,n„,f1 " fore h.inci c.n Ilea re.la
Tula cKe.iera nepeainix samtex
0,884 ca n a 42 5-6 • 8,1 4,3 53,0 46,7 5,2 0,7 41,8 5,6 -
1,290. camen 48 5-6 8,6 4,6 53,5 42,7 7,4 0,6 42,0 7,2 0,1•
1,392 cameui 49 0_7l 4,7 53,2 41,3 5,3 0,9 45,0 7,5 0,1
1,485 camBa 46 . 5-6 '.: . 8,4 ' 50,9 42,8 5,1 0,7 45,0 6,4 -
1,508 camBa . 48 04-.-7. » 8,5 4,5 53,4 37,7 5,7 0,6 49,0 7,0 0,1
1,528 cameul 48 6-7 8,1 4,3 53,7 43,8 5,2 0,5 43,0 6,9 0,2
1,64 7 ." Camaa: 47 6-7 8,4 4,4 52,5 43,7 7,0 0,7 41,1 7,4 0,1
1,652 camel 50 6-7 9,0 4,5 50,4 42,2 5,6 0.7 43,9 7,5 0,1
1,833 ca meal 1 51 6-7 7,8 4,1 52,8 42,2 5,6 0,7 •44,6 6,8 0,1
1,847 ca /qua 1 52 7 7,7 4,9 54,6 40,7 7,6 0,9 42,4 8,4 0,1
1,996 camel!" 57 7-8 9,5 5,1 5,30 37,9 5,9 . 0,4 48,3 7,4 0,1
um . ramm; 70 9-10 9,9 - - 31,4 5.9 0,4 54,5 7,9 -
A. - Total weight of body, kg B. - Sex
G. - Length of body, cm D. - Age, months
E. - Weight of skeleton, % of body weight F. - Weight of cartilage, % of weight of:
a - body b skeleton
G. - Weight of sections of the skeleton, kg v - skull w - ribs X - sternum y - vertebral column • z - limbs
f - female m - male

é,, Table 10.
rioa lu ma
1103- p a cT,
mecm-uen
5-6
5-6
6-7
5-6
3-7 6-7
6-7
6-7
6-7
7
7-8
2:3
2,7
2,9
2,6
2,7 . 2,4
2,6
2:7
.2,4
.2,6
3,2
59,2 •
54,0
52,9
55,9
5-1,8
55,3
54,7
.54,7
56;7
50,4
57,1
8,6
10,8
9,3
4,6
8,1
9,1
10,0
6,9
8,4
11,1
6,7
0;9
0,3
• 0,4
0,2
0,02
0,3 0,5
• 1,4
0,3
0,4
0,3
26,1
26,9
29,6
29;1
31,5-
27,3
26,7
29,4
27,1
28,3
29,0
6,0
7,0
7,7
10,3
7,8
8,0
7,2
7,2
9,7
6,6
361
The relative weight of the red bone . marrow of the embryos of the common porpoise in . various sections of the skeleton, as % of the total meight of the bone marrow.
Ta6ouga M •
Bec xpacooro KOCTI-10r0 mo3ra 11.'1011,0B mopcBori C1311111,11.
Il pamipuou oraeaax cBeacTa, 0/0 OT oGalero Beca Bocruoro m031a
.06utues nee
. re.na, KO
061mill nec 10C 011010
moara,% OT
neca Toila
V •I gepena I peep
HY1110- H04,101-0
cioa6a
uonAnocre
J. 11111: laanunx
•
F Bec Knerioro Moira H cnenere,
X Wan-
caria " 42
cameo. 48
cameo .49
camaa 46
caMlV! 48
cameo 48
ca man 47
camen, 50
CaMen 51
calm 52
cameo 57
A. - Total weight of body, kg B. - Sex
C. - Langth of body, cm D. - Age, months
E. - Total weight of bane marrow, as.% of body weight
F. - Weight of bone marrow In the. skeleton, kg v - skull w - ribs X - sternum y - vertebral column z - limbs
1 - fore, 2 - hind
f - female m - male
0,884
1,290
1,392
1,485
1,508
1,528
1,647
1,652
1,833
1,847
1,996
0,2
0,1
0,1
0,2
0,1 0,3
0,3
0,1
0,3
I■J

„ , 362
weight of the skeleton. The relative weight of-the ribs and
sternum of the S- 8 month embryos comprises about 7% of the
weight of the skeleton, approximately equally divided between 223
the bones of the girdle and of the free fore limbs. The
smallest fraction in the skeleton is taken up by the rudimen-
tary bones of the hind limbs (averaging 0.1% of the weight of
the skeleton).
The amount of the bone marrow in the embryos ranges
from 2.3 to 3.2% of the body weight (Table 10). With the
growth of the embryo the fraction of the bone marrow in the
skeleton becomes gradually greater, probably mainly on account
of the development of haematopoietic tissue in. the vertebral
column, where there occurs a gradual displacement of the car-
tilage by the bone tissue. In the embryos the main part of
the bone marrow is localized in the bones of the skull (about
60%). Half as much of the bone marrow is contained in the
bones of the vertebral column. On average, 8% of the bone
marrow is contained in the ribs; in the sternum, represented
by a cartilaginous plate with a few centres of ossification,
in which there is also found haematopoi.etic tissue, is located
0.4% of the total bone marrow. In the skeleton of the girdle
and free fore limbs of the embryos there is concentrated about
8% of the bone marrow, while in the pelvic bones there is found
about 0.2% of its total weight.
In generalizing the materials on the weight of the
skeleton and of the bone marrow in the various bones of the
dolphins there are manifested several common patterns. During
the process of the intra-uterine development of the embryo

1 363
•
•
the cartilaginous skeleton is reulaced by the bony skeleton.
The process of the replacement of the cartilage by the bone
occurs especially intensively in the second half of the intra-
uterine development. While during the first phase of develop-
ment the skull occupies the largest part of the skeleton of
the embryo, during the following stages it is the bones of the
vertebral column which become dominant. The bones of the skull
are almost definitively formed towards the time of birth. Thus,
the weight of the skull of the embryos and adults of the white-
sided dolPhin is the same (approximately 28% of the weight of
the skeleton). In the vertebral column the cartilage is almost
completely displaced only in the sexually mature individuals.
With the development of the embryo the amount of bone marrow
increases in both absolute and relative terms, and at the same
time its relative amount decreases in the skull, while it in-
creases in the vertebral column.
According to the data of Z. I. Brodovskaya (1964), in
dolphins during the embryonic period there is found an inten-
sive haematogenesis in the sternum and the bones of the flip-
pers, which continues on also in the adult animals. According
to our data, during the urocess of the development of the em-
bryos there does not occur any marked increase in the relative
weight of the bone marrow in the flippers and sternum (see
Tables 6, 8 and 9), although in the adult animals the amount
of bone marrow in these sections of the skeleton is somewhat
increased (see Tables j and 4). The fluctuations which are
present in the weight of the bone marrow of these structures
in the sexually mature individuals probably, in the main, re-
flect individual variability.

364,
The haematopoiotic canabilities of newly born dolphins
may be judged only by indirect data. Thus, in the adult com-
mon porpoise the amount of bone marrow comprises 2.0%, while
in the 6 - 8 month old embryo this is higher - 2.7% of the
body weight. In the adults of the common dolphin the weight
of the bone marrow comprises 1.9%, while in their 9 - 10 month
old embryos (almost just prior to birth) the bone marrow com-
prises 3.0f of the body weight. Proceeding from these data
it may be concluded that in the newly born dolphins the amount
of bome marrow is one and a half to two times greater than in
the adult individuals. Between the weight of the skeleton of
the newly born and adult dolphins there is found this same re-
lationship. The study of the skeleton and bone marrow of
terrestrial mammals showed that the relative weight of the
skeleton and bone marrow of the newly born individuals is also
approximately twice as great as the corresponding values of
the adult animals (P. A. Korzhuev, 1964; A. N. Evstratova,
1965, and others ) .
During the period of intra-uterine development the em-
bryo finds itself in conditions of a relative physiological.,
hypoxia, the reaction to which is an intensification of haemato-
genesis on account of the increase zn the mass of the bone
marrow and also of its productivity. In the aquatic mammals
during the embryonic period the intensification of haemato-
genesis obviously proceeds to a greater degree along the path
of an increase in the productivity of the bone marrow, while
in the terrestrial mammals this occurs along the path of an
increase in its mass.

365
A basic feature of the adult Black Sea dolphins, as
compared to non-diving mammals that lead an active way of life,
is the smaller weight of their skeleton and bone marrow. Thus,
while the weight of the skeleton of the dolphins is as much
as 6.5% and the weight of the bone marrow - 2% of the body
weight, in the dog (P. A. Korzhuev et al., 1968) these values
are correspondingly equal to 10.5 and 3.3%, and in the reindeer,
according to the data of P. A. Korzhuev and I. S. Nikol'skaya
(1960), correspondingly 10.9 and 4.8% of the body weight.
In spite of the decreased weight of the skeleton and
bone marrow of the Black Sea dolphins, their haematopoietic
Productivity is fairly high, which may be judged by the supply
of haemoglobin to the organism. The haemoglobin supply of the
common dolphin (13 g haemoglobin per 1 kg body weight) is al-
most twice as great as the haemoglobin supply - to the organism
of the guinea pig (Cava Porcellus), which, according to our
data, has a bone marrow weight equalling the weight of the
bone marrow of the common dolohin, and a little e:reater than
the haemoglobin supply to the organism of the dog (11.9 g per
1 kg body weight).
The noted features of the quantitative characteristics
of the main haematopoietic organ of the Black Sea dolphins
are related to the shutting-off of the breathing for some
period or other during diving, and also to the life in a high
density medium, in which, in contrast to the terrestrial en-
vironment, the load on the body is evenly distributed.

366
,ili'iT1:I'ATJ'PA
1 Btm•poroua A. M. C)<e.ie•r, i<oc•rln,til aloar n relo11oa3 n Onrorenc3C y cntnlcïl.
jAol<.nattl,I TCXA, nbIn. 100, 1965.2 Ko/n.Icyca 17. A. J.[t,i\aTe:nwnasl (I)ylrt<IU151 xponll 11 cheseT n03n011091IL1X .[CnnoT-
tII,IX. «ycuexil coupenlelntail 611oaorlut», T. 39, nhln. 2, 1955.3 KopqrcYco 1%. A. IÏpOÛACAIa iieliOCo\toCTn C'l'O4IQI 3I)CI1}1A 3CdI1I03I q)I13IfOA0rnI1.
a. A^I., 1963.a^1,n'naüuolntan If troc^tllttecl<asI ^icAwnn[»^I Koparcyclr H. A., Gnaoûnnoc•.cr JT. B., L•c;crponoocz C. H., Moc7epuTOen 3. M.
OnI,IT xo:u111ecTnennoro onvcae.-Ienna rentor.1o6nr111 n t<ocTnoro I,f03ra y o6[,nttlo-nen110ro lICpuoatopCl:oro 1ze:1141tna. C6. «Mopc[<ne nl,nel<onnTatolunC». Al., «Hayi<a»,
1 3 J 4.5 Kop,;cl/ea 17. A., Tnc1306n 7'. H. O I<Onuvecrncnuaïl xapa[<Tepncrni<e hponn _II
hpouernopnt,Ix oprauon vclnlomOpcr.Iix 1eal,(j)iiiioa. «)Kypnaa 3UoalOlUfOnlloÎ[ 6tto-xur1 mt II c;)ust[onorml:>, T. 3, nwu. 2, 1967.
b Kop^u/ecr TT. A., I nasoaa 7'. H., Arnr:puncr:an 0.• 1•I., Kttnorti3apo6a Al. P.KonWIec•rlieunoe If xaaecTneunoe pacnpetenerlne I<ocrnoro mO3ra y 133poC.antx co-6ai<. c,i(ocmnmecr.asl 6noaor.In if nle,awtnna», jx^ 2, 1968.
rf Kopxc//eu 11. A., tlunoAr,cnasl H. C. KO.nïi1IecTno IrocTnoro uo3ra y cenepnoro
oriclln. RAI] CC'CP, T. 131, nl,nt. 1, 1960.8 h4po1Inoa K. A. I3econoïi if xnatnme.cl<uïI cocTan OTae:Ibnl,ix IIaCTCif Teaa B FIC-
xoroPLIX opranax (j)nnna.za. Tpynl>i ,T•41110, nl,tn. 25, 1953.9 CAeIU{oti M. M. PeayA[,Ta•rhI n3neuliluannst xpytnn,tX It ,Ie,Ii<nX HIlT0O6pa3nl,ls,
Ao61,niaenn,ix na Ila.a>>ucu 13ocTOI<e. Tpyttht 1 tAIïI^ All CCCP, nlrr. 34, 1961.
10 anpor nc< If, H. nlaTepna.tl,[ x neconoa)y If xnntityecr.oN)y cocTany xlITOn. Tpya),T-BIIi4PO, r,I,In. 15, 1940.11 Bjarltnsorl T. Some weight measurement of whales. 1\Torsk. Hvalfan;;sf.Tidende, no i, 1954.12 Slijpel• B. J. Whales. 1-flltchinson of London, 1962.
REFERENCES
1. Evstratava A. M. The skeleton, bone marrow and ha.ematio-
poiesis during ontogeny in the pig. Doklady Timirya-
zevskoi sel'skokhozyaistvennoi akademii (Reports of
the Timiryazev Agricultural Academy), No. 100, 1965-
2. Korzhuev P. A. The respiratory function of the blood and
skeleton of vertebrate animals. "Uspekhi sovremennoi
biologii" ("Advances in contemporary biology"), vol.
No. 2, 1955-
3. Korzhuev P. A. The problem of weightlessness from the
point of view of terrestrial•physiology. .°Aviation
and space medicine". Yoscow, 1963•
39s

367
Korzhuev P. A., Balobanova L. V, Evstropova S. N. and
Moderatova Z. M. An attempt at a quantitative de-
termination of the haemoglobin and bone marrow in the
common Black Sea dolphin. Symp. "Marine mammals".
Moscow, Publ. "rfauka", 1964.
5. Korzhuev P. A. and Glazova T. N. The quantitative charac-
teristics of the blood and of the blood-forming organs
of the Black Sea dolphins. "Zhurnal évolyutsionnoi
biokhimii j fiziologii" ("Journal of Evolutionary Bio-
chemistry and physiology"), vol. 3, No. 2, 1967.
6. Korzhuev P. A., 01azova T. N., Alyakrinskaya O. I. and
Kalandarova M. P. The quantitative and qualitative
distribution of the bone marrow in adult dogs.
"Kosmicheskaya biologiya meditsina" ("Space Biology
and Medicine"), No. 2, 1968.
7. Korzhuev P. A. and Nikol'skaya I. S. The quantity of bone
marrow in the reindeer. Doklady AN SSSR (Reports of
the USSR Academy of Sciences), vol. 13•, No. 1, 1960.
8. Mrochkov K. A. The weight and chemical composition of
different parts of the body in certain organs of the
fin whale. Trudy VNIRO (Transactions of the All-Union
Research Institute of Marine Fisheries and Oceanography),
No. 25, 1953.
Sleptsov M. M. Results of weighings of large and small
cetaceans, caught in the Far East. Trudy Instituta
morfologii zhivotnykh AN SSSR (Transactions of the In-
stitute of Animal Morphology of the USSR Academy of
Sciences), No. 34, 1961.

368
10. Khar'kov I. I. Information on the weight and chemical
composition of whales. Trudy VNIRO (Transactions of
the All-Union Research Institute of Marine Fisheries
and Oceanography), No , 15, 1940.

369
UDC 599.537
G. N. Solntseva
THE COMPARATIVE-ANATOMICAL AND HISTOLOGICAL CHARACTERISTICS 226
. OF THE STRUCTURE OF THE EXTERNAL AND INTERNAL EAR OF
CERTAIN DOLPHINS
The specific characteristics of the way of life of the
cetaceans and those data which we possess, concerning the un-
usually wide frequency stectrum utilized by these animals for
echo-location and communication purposes, makes the study of
their organs of hearing a very pressing question.
Up to the present time many experimental étudies have
been conducted on the capabilities of the cetaceans for per-
ceiving sounds, but insufficient attention has been paid to
the fine morphological structure of their organs of hearing.
For the elucidation of the functional possibilities of the or-
gan of hearing of the cetaceans, the detailed examination of
the morphology and histology of this organ presents consider-
able interest.
The organ of hearing of the terrestrial mammals has
been studied in detail by both Soviet as well as foreign in-
vestigators (Vinnikov and Titova, 1961). Some data are avail-
able on the anatomy and histology of the outer, middle and in-
ner ear of cetaceans. However the histological structure of
Corti's organ has not been studied in detail.

370
MATERIALS AND METHODS
The investigations were carried out on 5 white-sided
dolphins and 5 bottlenose dolphins.
The external and internal ear was studied histologi-
cally and anatomically. The material was fixed in 10:, forma-
lin and embedded in celloidin according to the usual method.
The bullae tympani were collected immediately after the death
of the animal and were fixed in Vitmaak's fixative for 3 weeks,
they were then decalcified and embedded in celloidin. Section
thickness 15e. Staining with haematoxylin-eosin.
EXTERNAL AUDITORY ATUS
It is known that many investigators (Hunter, 1787;
Yurie, 1873; Zille, 1915; jamada, 1953; Fraser and Purves,
1960; Reysenbach de Haan, 1957; Purves, 1955; Purves and
Utrecht, 1964), concerned themselves with the study of the
structure of the external auditory meatus in both the Mystaco-
ceti (Balaenoptera acutorostrata, B. musculus, B. physalus,
Megaptera nodosa) as well as in the Odontoceti (JlobicePhalus 227
melas,l'hyseter catodon, Phocaena Phocaena l Tursiops truncatus).
We have not found any studies in the Russian literature on the
structure of the external auditory meatus in the white-sided
dolphin and the bottlenose dolphin.
The external auditory meatus in the cetaceans starts
with a very small opening *behind the eyes. Further on it is
represented by a winding duct. Its inner end, which abuts on-
to the middle ear, is tightly closed by the tympanic membrane-
ligament, which separates the outer car from the middle ear.

' I •.
,5•7;
371
Pitc. 1. Flapywilmil cnyxonofi npoxo,]. (re,maToKenn1I11-3o3mi, 10x7)
Figure 1. The external auditory meatus (haematoxylin-eosin,
10 x 7).
The whole of the external auditory meatus was divided
into 5 sequential sectors (from the periphery to the center),
the preparations of which were serially prepared.
In sector 1 (Figure 1) the external auditory meatus
in the white-sided dolphin and the bottlenose dolphin is rep-
resented in the form of a very narrow slit. In sector 2 the
dimensions and form of the external auditory meatus change
markedly. The thickness of the wall increases considerably,
and at the saine time the auditory meatus itself is completely
obliterated. The lumen of the auditory meatus appears at the
very end of this sector (Figures 2 and 3). In sector 3 the
lumen of the external auditory meatus acquires an oval form
with protrusions (Figure 4). The wall of the auditory duct,

372
te) Plic. 2. II yliacTol: liztpyKlioro cayxotioro npoxcua
2a — coemiiiirresibuo.mannoe aapacTanne npoKcinta.-ibitoil macTit cnyxotsoro npoxoica, 26— I1J1OT1t.ft ilempopm.leiman coefimmt-
. Teabnayt TIUIIJb, 2o— numc:tuible nytint
Figure 2. Sector II of the external auditory meatus. 2a - connective-tissue obliteration of the pro-
ximal part of the auditory duct, 2b - dense unformed connective tissue, 2v - muscle bundles.
in comparison with the preceding sector, becomes thinner. In 228
sector 4 the auditory meatus has an oval form with a consider-
ably enlarged lumen, in comparison with the preceding sector
(Figure 5). The thickness of the walls of the auditory duct
becomes uniform. And, finally, in sector 5 the externai audi-
tory meatus differs but little in its form from the preceding
sector, though its dimensions are enlarged (Figure 6).

373
Yric. 3. Konüciuoil o1;tca lI YIlacrxa napyNCnoro c.,yronororipoxo^a
^2 - rIOflE.'1Ci'.1'C rlnpryCT^ n r^}rXf^Rp}r P.nO?_nnn 30 - CC.
KPCS1iLIC i4CAC;4bir OICj)^'R^2110111llC C7)'\01301Î no.`: (U
Figure 3. Terminal portion of sector II of the external
auditory meatus.
3a - appearance of lumen'in auditory meatus,3b - secretory glands, surrounding auditory duct.
In the preparations of sector 1 the external auditory
meatus is surrounded by numerous dermal papillae, between which,
within the epidermal cells,melanin is located in the form of
grains and granules.
In sectors 2, 3, 11' and 5 the inner wall of the auditory 229
meatus is surrounded in an annular manner by a layer of dense
connective tissue. In sectors 2 and 3 between the bundles of
connective tissue are situatedc•protein glands and individual
fat cells.

374
O
1) 11c. 4. III rtacToK Hapymoro cnyxonoro npoxon,a 4a — onaehmi (Impta cmyxocoro npoxpAa c nambucc3ut-
IIbMfl 13b11151411118111151M11, 46 — ccl<peTitte "Acme3b1
• Figure 4. Sector III of the external auditory meatus.
4a - oval form of auditory duct with finger-like protrusions,
4h - secretory glands.
In the dense connective tissue there are found almost
undifferentiated cells: fibroblasts and histiocytes. States
of mitosis and amitosis of the colis were found. lying adja-
cent to the layer of dense connective tissue are muscle bundles
(m. zygomatico auricularis and occipiti auricularis), which
extend along the auditory duct and have a varying thickness.
Between the individual muscle bundles and also between the
cartilage and the inner wall of the auditory duct is situated
loose connective tissue, including bundles of collagen fibres •
and seDarate elastic fibres.

375
Plic. '5. IV ytiacox napywnoro cayxoisoro npoxma 5a— oBamillast (popma c.uxonoro iipoxma, 56 111)0-
'11,1C EJICTIZI1*
Figure 5. Sector IV of the external auditory meatus.
5a - oval form of the auditory meatus, 5b - fat cells.
lb)
Directly around the auditory duct there are situated
many blood vessels and blood lacunae, filled with blood.
Commencing from sector 3, cartilage adjoins the exter-
na1 auditory meatus. Initially it appears on one side of the
auditory duct. Then it lies adjacent to the two narrower
sides of the auditory duct, in the form of two caps. Further
on there is traced the fusion of the two separate cartilages
of the preceding section, and already in sector 5 the cartilage
acquires a horseshoe-shaped form, surrounding the auditory
duct on 3 sides. The cartilage forms a tube which joins up

376
Piic. G. YttacTOK ü8P}'iKItOCO C.9)'XO6G1'O itj?OXOi(£IGa - Kpoael!octtvtït cocv^, (:ô - Kponel!octtaa n<u<ylla, Gc--
itAOTltitSi IICO(^lOj?1\lAClil!RSI COC;IUIIIITC^b1i8A TIdBlIb, Gi.-9:Ic2CTiI-
LICCKIIFi XpIIll(
`r'igure 6. Sector of external auditory meatus.
6a blood vessel,6b ^ blood lacuna,6v - dense unformed connective tissue,6g - elastic cartilage.
into the bulla tympani. By comparing the dimensions of the
external auditory ducts of the two species of dolphins under
consideration, by individual sectors, it can be ascertained
that these vary only insignificantly.
Thus, the external auditory meatus runs like a pigmen-
ted tube surrounded by fibrous tissue. After passing through
the fat layer the auditory duct enters a cone-shaped mass of
fibrous tissue, which is considered as being connected with

377 • the auricular muscles and the distal end of the auricular
cartilage (Purves and Utrecht, 196 )4 ).
INNER EAR
On investigating the inner ear of the dolphins it was
noted that, macromorphologically, the cochlea of the dolphins
was distinguishable from that of terrestrial mammals (for
example, of the guinea pig).
The cochlea of the dolphins is flat and completes 1.5
spiral turns, while in the guinea pig it is pyramidal and com-
pletes 4.5 turns (Figure 7).
As in the terrestrial mammals (Ya. A. Vinnikov and
L. K. Titova, 1961), the cochlear canal is divided by Reissner's
membrane into the two scalae, the upper one of which passes
to the apex of the cochlea, while the lower scala, running
from the apex, terminates at the round window.
The upper scala is divided by Reissner's membrane into
two unequal cavities, the smaller of which is the scala media.
Within the scala media on the vestibular surface of the basi-
lar membrane is located the organ of Corti. From the inside
closely abutting to the organ of Corti is the spiral lip, which
passes into the internal spiral notch. The spiral lip is formed
of flat epithelium, which in the dolphins, as in terrestrial
mammals, is arranged in Parallel rows. The cells of the epi-
thelium are large, with . large nuclei, and are disPosed along
the radius of the cochlea.
The cells of the spiral notch pass into the supporting
elements for the inner hair cells. At the point of termination
of the internal spiral notch are disposed the border cells,
which form two rows.

o 378
'
7.1".
Pue. 7. Buenumil uu,u, ,n,em41ma-6e:usGotwu _ ---
Figure 7. External appearance of the cochlea of the white-
sided dolphin.
The receptor elements of the organ of Corti are the
outer (NVK) (Figure 8) and inner hair cells (VVK) (Figure 9).
These cells differ in their structure. The inner hair cells
are arranged in a single row and have a jug-shaped form. The
large nucleus is situated at the base of the cell. They are
situated alonp,side the apices of the outer pillar cells.
In contrast to these, the outer hair cells are arranged in 234
three rows and have a cylindrical form. At their base is lo-
cated the round nucleus, which has a granular structure. The
cytoplasm is granulated.

379
8. kopTueu oprall. Ilapymmc BOAOCKOBMC E.-1C7Elf (remaToiccunuu-3031111,90x7 )
Figure 8. The organ of Corti. .Calter hair cells (haema-
toxylin - eosin, 90 x 7).
The supporting cells are represented by the outer and
inner pillar cells of the organ of Corti, which are located
on the basilar membrane. Between the supporting pillar cells
is located the tunnel and space of- Nun.
There can be clearly traced the inner border cells,
beyond which follows one row of the inner phalangeal cells,
which are situated between two inner hair cells, separating
one from another. Between the inner pillar cells and the bor-
der cells, in the cavity, are arranged the inner hair cells.
Lying adjacent to the inner pillar cells are the cells of
Deiters. With their bases they lie on the basilar membrane
and have a polygonal form with a large nucleus, situated in
the basal part of the cell.

; .
1 t.
38 0
Pile. 9. 1(opTucut oprau. Buyipennue no.nocKoable (teMaTOliC11:11111-
3631111, 90x7)
Figure 9. The organ of Corti. Inner hair cells (haema-
toxylin - eosin, 90 x 7).
• Next follow the cells of Hensen, which lie very closely
adjacent to the cells of Deiters. The cells of Hensen are
situated with their basal ends on the basilar membrane. The
apices of the cells are directed towards the outer hair cells.
They are large, with large nuclei. Beyond the cells of Hensen
are situated the cells of Claudius, which are represented by
short polygonal cells with very distinct intercellular borders.
Their cytoplasm contains a nucleus with nucleoli. The basilar
membrane, on which lie the elements of the organ of Corti,
is clearly traceable. Distinctly evident are the parallelly
arranged fibres, which are located at a uniform distance from
one another.

381
Thus, the analysis of these data makes it possible
to assume that, in contrast to the clearly observed macro-
morphological differences, there were not found any notable
histological differences in the structure of the cochlea in
the dolphins, in comparison with terrestrial mammals (guinea
pig).
CONCLUSIONS
In the study of the external auditory meatus we attempted
to elucidate the question of its obliteration in a particular
sector. Some investigators consider that, along its entire
length, the auditory duct in the toothed whales runs in the
form of an open tube (Fraser and Purves, 1960; Reysenbach de
Haan, 1957), while the studies of other authors indicate that
the external auditory meatus is a tube with a lumen that is
obliterated in some sector (Jamada, 1953).
Our data confirm the data of Jamada. The results of
the anatomical and histological studies showed that in the
white-sided dolphin and the bottlenose dolphin at a distance
of two centimeters from the surface of the skin there occurs
an obliteration of the auditory duct with connective tissue,
and this comprises approximately one twentieth of its length
(see Figure 2).
The obliteration of the auditory duct is of important
significance in the treatment of the mechanism of sound con-
duction. The comolete obliteration in the dolphins cannot
worsen the conduction of the signals to the middle ear in the
water medium and serves, mainly, to preVent water entering the
ear.

v.e^e..ea ve°^eaTRANSLATION 3185 5 of 7
F,a^e,°, 5e^ °^ea,==^°a SERIES NO.(S)aean^e^^°e °id.s°N^°° iame,

r
r
382
The f-Luznel--shaped a.uditory duct of the dolphins, which
is completely isolated from the medium, confirms the position
that they make use of another (as compared to the terrestrial
mammals) pathway for conducting the acoustic signals to the
middle ear.
On account of the "acoustà.c transparency" of the fat
layer of the hypodermis, the signals from the water penetrate,
with minimal losses, through the integument and, reaching the
cavity of the auditory duct, they move along this to the middle
ear.
As to the studies of the inner ear, there are all
grounds for assuming that in its anatomical structure the
cochlea of dolphins is well adapted for the perception of
ultra-sounds. This is indicated by the presence of the lower
loop of the cochlea, since, according to the existing electro-
physiological studies conducted on terrestrial mammals (Altukhov,
1911.9= Andreev, 1911.0, Bekesy, 1951H, Davis, 19,47, and others),
the shorter fibres lying at the base of the cochlea perceive
the high frequencies, while the longer fibres located in the
apex of the cochlea perceive the low f.requenciès.'
JJJ xTLPATYPA
• 1/tns•Yaoo l'. B. Yc,nonnopcrj),^c)iTOpllafl ;tes,TC.m>rioçTh co6axrt ripn srccncpuMc;r-Tü.'twnUT\r noupelaeutnl nepncI)epumCC).oro oTijensr c:Tysol;oro aua.a113aTOpa. «IIpUU,T_
tOIrlIr0JL flK)'CTIIKII», T. 1, 1949.2 flxopeea R. A. O q acoropUx uor,t,ix Aanrn>ix, xapar.Tepnsyloutnx ttesiTWhrrocTt,
nnyt;onoro aua.mtsa'ropa. 7-c coueutsulne no npo6.,,i. rs),tcru. nepnn. +ZesIT., nocn. na-1`u)'ru IJsrnnon<r: Tcsncw uor..q<)AOn. M.-,I1., 19,10. -
3 Bwttuu:oa Sl. A., Tlrrocra 31. K. Koprtrco opt'an. 1•13;>,-rso AH CCCP, M.-,r[_
l ir^31.I' Bckest/ V. Some F.Icctro - Mechanical Propertic^s of the organ of Coi ti.
Aun. or Olol., Rhinol. a. L.aryng., vol. 13, no 2, 1954.5 Davis 11. Biophysics- and physiology of the inner car. Physiol. Rcv., vol.
3, _ 1957.

6 Fraser F. C. and Purves P. E. flearing in 'Cetaceans, Bull , of the htitis-li. mus. (nat. hist.), Zoo logi, vol. 7, No I. 1960.
Huntcr J. Observations on the Strukture .and Oe.mnorny of Whales. Phil. Trans. 77. 1787. 8 Jantada. M. Contribution to the Anatomy of the Organ of Hearing of Wha-
les. Sei Re.p. Whales Research Inst. No 8, Japan. 1953. 9 Lilli J. C. Cetacea. Brit Antarctic. <Terra Nova». Expedition 1910. Nat.
IIist. Report. Zool. 1. no 3, 1915. 10 Marie J. On the Organisation of the Cealing Whale Globicephaius melas. Trans. Zoul. Soc., 8, 1873. _
1 1 Purves P. E. The Wax Plug in the external auditory meatus of the Mysti-ceti. Discovery. Reports 7,7, 1955. 12 Parues P. E. and van Utrecht W. Z. The anatomy and function of the ear of the bottlenosed dolphin Tursiops truncatus «Beaufortia» 9, no 111, 1964. 13 Reysettbach de Haan F. W. Hearing in Whales. Acta Oto— Laryngologica, vol. 137, 1957.
383
REFERENCES
1. Altukhov • G. V. Conditioned reflex activity of the dog
with experimental injury to the peripheral section of
the auditory analyzer. "Problemy fiziologicheskoi
akustiki" (Problems of i)hysiological acoustics), vol. 1,
1949 ;
2. Andreev L. A. Some new data, characterizing the activity
of the auditory analyzer. 7-e soveshchanie po problemam
vysshoi nervnoi deyatel'nosti, posvyashchanno pamyati
Pavlova. Tezisy dokladov (7th conference on problems
. of higher nervous activity, dedicated to the memory of
Pavlov. Reports of Proceedings). Moscow - Leningrad,
1940.
3. Vinnikov Ya. A. and Titiva L. K. The organ of Corti.
Publ. USSR Academy of Sciences, Moscow - Leningrad,
1961.

384.
UDC • 99.74.5
G. M. Kosygin and V. N. Gol'tsev
DATA ON THE MORPHOLOGY AND ECOLOGY OF THE HARBOUR SEAL 238
OF THE TATAR STRAIT
The study of the pinnipeds in the Tatar Strait was
initiated by S. V. Dorofeev (1935), who established that the
harbour seal, ringed seal, bearded seal and ribbon seal are
encountered here in the spring. The predominant species in
the Strait is the harbour seal and it comprises about 80% of
the catch of the hunting of the coastal inhabitants. The re-
maining species are not very numerous. The approximate periods
of whelping of the harbour seal, ringed seal and bearded seal,
and the period of moulting in the harbour seal were establtshed.
Later, P. G. Nikulin (1935) and E. A. Tikhomirov (1956)
placed the Tatar Strait in the category of unpromising regions
for commercial hunting.
These conclusions were made at a time when the seals
were caught for the sake of obtaining the fat. Now, however,
considerable attention is given to procuring fur skins. There
-f ore our attitude to the Tatar Strait as a commercially ex-
ploitable region should be re-examined. In an aero-visual
survey of seals, which was conducted by the Pacific ucean
Research Institute of Fisheries and Oceanography (TINRO) in
1968 (Fedoseev and others, 1968), a fairly large accumulation
of harbour seals was noted in the Tatar Strait. The study of
the harbour seal population that is living here, which has not
been affected by ship-based sealing operations, is of both pure-
ly scientific as well as applied interest.

385
Foin March 14 until April 7 of 1969 co-workers of the
TINR0 on the sealing ship "Sanzar" carried out field studies
in the Tatar Strait. Included in the purpose of the cruise
was the study of the distribution and biology of the seals,
and also the elucidation of the feasibility of commercial
exploitation of the animals. The exploratory transects covered
the region of the strait from 48 to 500 lat. N. The collected
materials were treated by the methods accepted at the TINRO
(Smirnov, 1934; Rokitskii, 1961; Chapskii 1963, 1967; Yablo-
kov, 1963, 1966; Tikhomirov and Klevezal', 1964; Sokolov et
al., 1966). Some of the materials on the harbour seal of the
Sea of Okhotsk were passed on by us to A. V. Yablokov. In •
addition to the authors, A. I. Corshkova participated in the
collection and analysis of the materials.
Many authors have devoted studies to the description
of the harbour seal, and yet the overall morphological charac-
terization of this seal is far from being complete. What has
been said applies to the same degree also to the harbour seal
population, living in the Tatar Strait.
Coloration of the hair coat. Earlier
(Kosygin and Tikhomirov, 1969) in describing the harbour seal
or mottled seal (Phoca larp-ha) its coloration was grouped into 239
4 types: light-coloured, slight mottling, very large mottling
and typical.
In the 64 harbour seals that were examined from the
Tatar Strait, the typical coloration predominated (6 0%); stan-
ding in second place was the slight mottling (33); 6 animals
had the very large mottling, and 1 was lightly coloured. The
colouration of the harbour seals is similar to that described

386
grw
by S. P. Naumov and N. A. Smirnov (1935), S. I. Ognev (1935)
and K. K. Chapskii (1963, 1966) on the basis of individuals
from other parts of the species range. Among all of the white-
coats of the harbour seal that were found in the Tatar Strait
only two pups were encountered that had the smoky coloration,
characteristic of the young of this species of seal from the
Bay of Peter the Great.
Length and weight of body. The average
length of the body (z0 ) of the harbour seal, living in the Sea
of Okhotsk, is 165.3 cm in the males and 151.6 cm in the fe-
males (Pikharev, 1941). There are available measurements of.Z0 on
the harbour seal from the Tatar Strait on individual animals
(Gagichko and Surzhin, 1935) from an autumn catch, which have
not been included with our data, because of the small numbers.
According to our data, the seals of this species from the Ta-
tar Strait are larger than those from the Sea of Okhotsk: the
body length (k) of the males and of the females was, corres-
pondingly, 169.0 and 167.7 cm. On the basis of this feature
they differ little from the harbour seal of the Bay of Peter very.
the Great (Sea of Japan) and are slgnificantly larger than the
females of the Bering Sea, Kuril and Sea of Okhotsk populations
(Table 1). There are also significant differences between the
males. The average length of the body of the whitecoats is
101 - 103 cm. On the basis of the total weight of the body .
(Table 2) and the weight of the carcass without the skin and
the fat ( sculp ) the sexually mature harbour seals from
the Tatar Strait are smaller than the individuals from the Bay
of Peter the Great. The weight of the whitecoats is the same

387
as in the Bering Sea. The weight of the sculp of the
harbour seals of the Tatar Strait ranges from 24 to 43 kg,
averaging 36.5 kg, in the adult males, and from 22 - 56 kg,
with an average of 35.1 kg, in the females. The relative .*
weight of the sculp comprised 31.40% (averaging 37%) in
the males and 29-49% (averaging 39.6%) in the females. In
the harbour seals from the Bay of Peter the Great during the
whelping 1-)eriod this equalled 43.3% in the males and 43.2% in
the females.
Meristic features. Many authors (Yablokov,
1963; Yablokov and Klevezal', 1964; Yablokov, 1966; Chapskii,
1967; Sokolov et al., 1968; Kosygin and Tikhomirov, 1969)
employ the number of vibrissae, of tracheal rings and of ribs
as a morphological feature.
The labial vibrissae of the harbour seal of the Tatar
Strait are seated in 7 - 8 rows. The same number of these
were counted in the harbour seal from the Sea of Okhotsk (Yab-
lokov, in litt.). The number of vibrissae in the males ranges
from 37 to 46, with an average of 43.2 ± 0.43; in the females
- from 37 to 50, with an average of 42.7 ± 0.44. In compari-
son with the harbour seal of the Bay of Peter the Great, no
great differences were found in their total numbers; on ave-
rage, there were somewhat fewer vibrissae in both the males
(t = 0.88) as well as the females (t = 0.36) of the former
population.
In two females (a whitecoat and a sexually mature ani- 242
mal) there was found a null row, in which there were from 1 to
vibrissae on each side of the snout. Such an occurrence has
* Tramslator's note. Sic. Presumably this should be 31 - 40%.

Pem
Rea:ion nøI
Sex
5,65 9,25
7,11 4, 9 1
7,99 6.43
-0,38
-0,03
+0,18
B3poce c
C2Me/t- camK3 .114 nz Urn
94-111 87-114
84-112 89-102
80-108 78-109
99-108 83-110
Toropc},:trik
3a.niel fleTpa Be./moro
5ep1Ir000 mope
Males CaMIZIpf: tj-3= +2,12
it--4= +2,60 Et-= +1,7O
Females Cammt t t -2,- 0,39
t 1-z +5,81 t_= +5,81
ti_4=±7.67
* Zammie A. H. Benustua. " ,11,anliNe 3. A. Tmomilpona.
* Data of A. N. Belkin ** Data of E. A. Tikhomirov
C CO CO
Table 1.
The body length of sexually mature individuals (z0v ) and
of whitetoats (ze. ) of the harbour seal from different regionsea6xxua 1 crn • ji(amia Te.na no.riosoapembtx oco6eii (Zcv) 6e.nbRo8 (Zc) oaprx paalibm paionon, c.e. •
7
9 7
60 47
4 4
lirn
Tata Strait
Bay df Peter the iGreat
BerinÉ Sea
Darili Islands* KYPubcKtrd ocTpona*
MO-
camen111 cammaf
corneure cammor
. ca eel camof
Ca MC11.-72, comNa.l.
comen.111 camIcaf
• B b X 1-t:
101,2±-1,91 102,7-±3,41
95.8 ± 2.37 M,0±1,61
95,9 ±-1,03 94,1 ±-0,93
103,0 103,7
16 146-173 158,0±-1.68 6,73 35 145-179 157,7-1-13,2 7,81
3 152-168 159,3 10 144-173 155,5±2,78 8,80
12 128-170 148,5±-4,14 14,33 13 123-156 141,2±2.52 9,09
14 134-163 150,0±2,58 9,66 6 135-154 1 45,5±0,89 2,19
19 135-171 153,3±2.18 9,49 16 134-155 142,1 ±-1,52 6,07
t male4 .
cam"a femare
+0,14
-
+1,51
+0,46
+4,10 Sea o'f OkhotgkUol;oe Pe"
m - male f - female

r
I t tLïat^
C3tiC1l-male -Q I Ca)tlSa femalé
Tatar^ Strait
Dây of Petertn_e Great-uril; Islands:<
,B-,^er in Sea
Table 2.Total body weight of the sexually mature individuals and
whitecoats of the harbour seal, caught during the period of ïlt^liiû^i^
in different regions, kg.Q611(L1H IIeC Tc:la iloJI0Q0317CRb1X 0^0Ûei! A^JeJIbISOS :
IaprY.^ J1,O6bITbiX II IIe17I10,^ ll.leHK41 B p23^^^ ^alt OI 1X+ hZ
3a:cIIB 1-leTQâBe:Illxoro
Kypl :zbcKi[eocTpo13a
BepII11rOüo Mope
canlltb 9canlKilf 7
ca1I11i, 4calIr.uf 4
cauub>ni 3Gca>IKII 34
13-30 19.4± 1,7811-31 22,0 =2,7G
8,0-27.0 12,9-_` 1,969,4-25,0 14,7--2,19
15,F-29.5 23,017,0-32.0 27,0
5,8-3:3.8 2 1.9 -!-1,257,3_.3:},7 22,2-!-1,12
5.047,3-1
5,$9 -O,6G 3 IOCi-125 t i3,3
5,80 10 80- 150 11•1,0--6,11
- .- 13 62-113 90,3! 3,99
7,53 - 0,19I 226,51, 16
iIpl1111Ujt HH031)aC9'C OT pomucuusl u 111^ioicl'110 .to oAtloro xecsuta.
Young- from birth to an age ofabou^ one month.
M - malesf - femal.es
t7-147I
55.4 -5,4866,0 - 3,2350-100
males :Ca1fIJ,b1: ES-3= `i' 1,ô'1
ts_,=+2,26
7 60-92 75,2+4,r9
19,3
14,4 +2,4112,7
25,7 +3,0412,9
'i'emales :Cfl1IKH: tI_z=-3,75
fi_a=+2,51t1_.=+5,50
t
,o

390
not -oreviously been noted in the harbour seal, either in the
Bering Sea (Chaoskii, 1967) or in the Bay of Peter the Great
(Kosygin and Tikhomirov, 1969). D)reover, vibrissae were found
in one sexually mature female on the lower jaw. They were
distributed as in the bearded seal (Kosygin, 1968).
In the harbour seals of both the Tatar Strait and the
Bay of Peter the Great there are 1 - 2 nasal vibrissae and 3 - 6
ocular vibrissae.
Between the males of the harbour seal from the Tatar
Straight and from the Bay of Peter the Great, in the average
numbers of labial vibrissae, there is found a significant dif-
ference in the first row (t = 2.5) and a highly significant
differenc (t = 3.33) in the foùrth row. In the remaining
Cases no siîrnificant differences were noted, either in the
total number of vibrissae or in the number of these in the dif-
ferent groups in the populations examined.
The trachea of the harbour seal from the Tatar Strait
has no significant differences in its structure from that which
has already been described by A. Si Sokolov et al. (1968).
In 6 out of 28 individuals (21.4%) the rings situated around
the bifurcation had a left overlap. In the harbour seals
caught in the bay of Peter the Great (26 head), tracheae of a
similar structure were present in 31A of the examined indivi-
duals.
59 - 76 rings were counted in the tracheae: in the
males the average was 67.5 ± 1.17, while in the females this
was 63.7 ± 1.07 rings (Table 3). The differences betweeen
these were significant (t = +2.48). As is evident from Table 3,

l'eeffls ii Um 1 M *m
16
16
02-76
61-71
64-76
61-75 65--73
+2,48
+1,18
+1,33
391
Table 3. Total number of tracheal rings in males and females of the harbour seal from different regions.
Tat-Sauna 3
()Stec nuantiecTuo Ko.nett, Tpaxeu camnors n canon naprif 13 pammix pafiouon
1
Camuu
n I 11m I M-•.m
P.U.LuS Cnmnu p.ca...t..e
Patiu Region
Tatar Strait TaTapcnnii upo- Jilin
Bay of Peter 3a.mil nona t e Great BumEon) Sea of ÛkhOtSPXOTenoe mope (according to (no 516.1oKo-Yablokov, 1. 066PY, 196.6; m in litt, 1969,5 1u' 1969) Kuril Islands' 30ToiTi .p" boepi<'1" e • Bering Sea Bepunr000 m ope
13
10
24
23 8
69,4 ± 0,54
69,5 ±0,74 68,5±:0,96 /1_3=1-1,12
h-4=-1,08 ti-5=-0,66
67,5± 1,17
65,9 ±0,82
98
11 7
«59-71
61-70
60-75
66-75 36-77
71,2±0,83 72,7 ±1,42 t1-2=-0,33 /1_3=-3;18 h-4=-5,73 /1-5=-5,17 •
63,7±1.01
64,1±0,64
68,0 ±0,84
—1,88 —2,46 •
the degree of the differences in this feature between the
harbour seal of the Tatar Strait and the other populations
of this species is not uniform: differences were not found
between the males, though, apparently, they are real between
the females from the Kuril Islands, the Bering Sea and the Sea
of Okhotsk.
A. V. Yablokov (in litt) made a count of the sternal
and and asternal ribs In 16 males and females of the harbour
seal from the Sea of Okhotsk and found a variation in their
number.
In the 23 harbour seals from the Tatar Strait that we
examine d no deviat ions were noted in the .number of ribs : each
had 10 sternal and 5 asternal ribs .
Some data on other internal organs .
The investigation of the internal organs of this spe cies is a

392
continuation of similar studies, which were started on pinni-
peds in 1960 (Scheffer, 1960; Kleinenberg et al., 1965; So-
kolov et al., 1966; Kosygin et al., 1969; Sokolov et al.,
1969).
In a description of the superficial structure of the
lungs of the harbour seal from the Bering Sea, attention was
drawn to the fact that in addition to the so-called typical
form (a not very deep, clearly evident notch on the acute edge)
there are alo found variations, in particular: among the 27
individuals examined in one individual the lungs had a lobed
structure (Sokolov et al., 1966).
A similar phenomenon has also been noted in other po-
pulations. Thus, among 31 specimens which were caught by us
in 1968 in the Bay of Peter the Great, in 8 the lungs had an
atypical form: in one animal there were deep notches and
grooves on the lungs, while in the other 7 there were no notches
of any kind at all.
Of the 80 examined harbour seals from the Tatar Strait,
24 individuals had atypical lungs:. 8 were entire and 16 with
deep notches. Some of the lungs had two notches with deep
grooves running from these, dividing the lungs into almost in-
dependent lobes (Figure 1). •
Either both of the lungs or one of them were atypical. 244
Both the right and left lunes, to an approximately equal deg-
ree, were noted as having a superficially different configura-
tion.
Gagichko and Surzhina (1935) had data on the weight
of the internal organs of the harbour seal from the Tatar Strait

393
Plic. 1. uacTo1•a ncrpevaeNtocrn pas.an11ttoilaropaum acrlnts .iaprn n nonYanunnx:
R- Sepunrona etopsr, 1 ; - 3anuna Ilerpa Bentnco-
100 [
cc ô a S C a e c
A. B % `%
o' UuUlY^
ea.d)opma aernnx:
a- 71muenan; G- a-rtmisynas[ nonblsaran; c- arn-tfanan ücawoasaccnsnsan.
Figure 1. The frequency of occurrence of the different forms
of lungs in harbour seal populations from:
A - Bering Sea; B - Bay of Peter the Great;V - Tatar Straight.
Form of lungs :a - typical; b - atypical lobular; c- atypical
41entire^
but, because of the fragmentary nature of these data, the oc-
casional lack of information on the sex of the animal and also
because this catch was made during the autumn period, these
data, unfortunately, could not be utilized. Our materials in-
dicate that in the adult males, as compared to the females,
the average weightsof the heart, liver, spleen and diaphragm
are somewhat greater and equal, correspondingly, for the former
0.56 ± 0.04; 2.63 ± 0.21; 0.28 ± 0.02; 0.54 ± 0.23 kg, while
for the latter the values are 0.52 ^ 0.006; 1.79 ± 0.07;
0.24• ± 0.02; O.1I-8 ± 0.02 kg. In this case the differences
in the weight of the liver are at a highly significant level
(t = 3.81). The index of this organ is also greater in the
males (Figure 2). In the females there is found a comparatively

394.
60 30 0 30 60C4b/ . Ca^A U,
mmes feriaPile. 2. Bec unyTpetitntx opraxou naprll
906b1TLIX B P8311bIX p8ÎIO11i1X.
060311 a q e fi il n:I- Tarapcrnrt npo.-Iun; 2- lieptrllrono nlopc; 3-3anun 17rrpa Be.vtroro; A- Kypu.il.ctulc orrpona
A- talmevlnlh: 6- tte•tculi; li - nérl<ttc; r- ri<cay;tor,;t( - cepAUe; F. -- ceneaet,ha; )n -- nuachparNla; 3-
noer.a; ff - noa 1;e:ryJw4ltast ;ece.nc3a
Figure 2. The weight of the internal organs of the harbour
seals caught in di.-fferent regions.
Legend
1-Tatar Stra i:t ; 2'Bering Sea; 3 - Bay ofPeter the Great; 4• - Kuril islands.
A - intestine; B - liver; V - lungs;. G - stomach;D - heart; F- spleen; Zh- diaphragm; Z - kid-ney; I - pancreas#
high relative weight of the lungs (22.6 ). Differences bet-
ween males and females in the cardiac and diaphragmal indices
were negligible (in the males and females these were corres-
pondingly equal to 8-5 and 8.6%, and 8.0 and 7.9%).
The fact should be emphasized that the relative weight
of the lungs and heart of the animals from the Tatar Strait
is somewhat I.ower than of those from the Bering Sea. Thus,
in the Bering Sea females the index of the lungs equals 2^a•.8%,
of the heart - 11.1%, of the diaphragm - 8.0%. In general, 21I•5
the relative weight of most organs of the females exceeds that
of the males.

395
The noted characteristics of the internal organs of the har-
bour seal depend on the living conditions, which are specific
to each region (Sokolov et al., 1969).
Craniometric characteristics are pre-
sented in Table 4. The index of the greatest length of the
skull, calculated from the body length (7 ) comprises 13.7
and 13.6%, respectively, in the males and females. Thus, the
sex differences in the leneth of the skull are insignificant.
In this feature the skull of females of the harbour seal from
the Tatar Strait is, on average, somewhat longer than in those
from the Bay of Peter the Great (t = +0.80), the Kuril Islands
(t = +0.73) and the Bering Sea - 207.8 mm (according to the
latest data of Chapskii, 1967).
Notable differences between males and females are pre-
sent only in the rostral width and in the length of the nasal
bones. The absolute length of the nasal bones and the right
upper row of molar teeth in the females from the Tatar Strait
is a little greater than in the females from the Bay of Peter
the Great.
In the skull of the females from the former region the
value of the zygomatic (greatest) width is somewhat smaller
(t = -2.26). This same difference is also noted in the rela-
tive dimensions. Thus the index of this feature, calculated
from the greatest length, on average is equal to 58.0% for the
individuals from the Tatar Strait and 61.4% - for the Bay of
Peter the Great.
The dimensions of the longitudinal diameter of the al-
veoli of the right upper canine show differences that are
close to significant (t = -2.57).

396
For a more complete characterization of the harbour
seal of the Tatar Strait it would be desirable to compare its
craniometric data with those of the neighbouring population,
from the Okhotsk Sea. Unfortunately, we do not have any of
our own skull materials on the Okhotsk Sea harbour seal, while
in the published studies these are either too scarce (Allen,
1902; Ognev, 1935) or have been presented without subdivision
according to sex and region of catch (Naumov and Smirnov, 1935).
Thus, the analysis of this material shows that, on the
basis of the morphological features presented, there are fea-
tures of similarity and noticeable differences in the harbour
seal which lives in the Tatar Strait, in comparison with indi-
viduals from other parts of the species range.
Food objects. Out of 76 harbour seals, aeed
1 to 41 years, that were caught, food remains were contained
in the stomachs of 27 animals. 20 of these animals were caught
on the ice and 7 afloat. In most of the seals the food was
strongly digested; in some stomachs there were retained only
parts of the skeletons and otoliths of fishes, and beaks of
cephalopod molluscs.
The food base of the harbour seal in the Tatar Strait
during March-April consisted of fishes, mainly the Korean cod.
These were found in 25 stomachs (93%). These data confirm
the conclusions of previous authors (Ognev, 1935; Chapskii,
1963; Kenyon, 1965, and others), that fishes play an impor- 247
tant role in the food- Of this seal.
In second place were cephalopod molluscs, found in 10
stomachs (37%), and last were crustaceans, in 3 stomachs*.
* The species affinity of the food objects was determined by L. I. Semenenko.

115
Greatest length
Greatest_zygoma- I1C CIT
Kastoid width
Rostral Width
Length of nasal bones i
Width of nasal bones
Main dimensions of the skull of sexually mature harbour 2 seals from "different regions, mm. . ' Tanza. 4
OcK0eubte pa3Mepb: nepena no.noB03pedimx aapr H3 pa3nb1x parionoti, itAt
Longitudinal dia-mêter of alveo-lus of ilipper can
Length of rostra' portion
Length of upper row of molgr eeth
.1.0.-1jd..I. ›D tel. Ci...L C. May u_L .L-- GUL t,11. U.L c.")..1.,
Tarapcmiet npoatm 3a:tun lima lienmcoro riptt3Harc 110.1 t
Feature sex n i un 1 Al+m 1 a n 1 Uni ' I .41-i:m I e
11aitgonEnuag es.,7u1!a cameo 8 210--225 215.5+1,77 5.01 • 3 205-230 215,9 - - caM!Ç 23 199-226 212,7:4:1,30 6,24 8 196-232 209,4±3,92 11,10 +0,80
Halifianwag criyzoBasi came 8 118-134 126,1 ±1,57 4,45 3 129-141 134,5 - - ampinra camNz 23 116-131 123,5+0,93 4,45 6 123-141 130,0±2,73 6,69 -2,26
Macrounast III npuna caMeLifl 6 117-129 • 123,7±1.70 4,27 3 120-133 125,8 - - ca..+I1 22 113-12G 120, 4 ±- 0,70 3,56 6 117-123 122,3±1,71 4,18 -1.02
PocTparibnam Lunpuna came! 8 37-43 39,4-±-0,65 1.85 3 39-45 42,7 - - cam; 23 32-42 37,7+0,47 2,24 11 35-47 40,4+1,45 3,56 -1,77
11 .1 i 1 f 1 a HOCOBIDIX Komi*/ CI MCTI 8 50-60 5C,6±-1,19 3.38 3 50-59 55,0 - - camK 20 46-59 52,6 ± 0.83 3,73 9 48-58 52,5±0,58 1,75 +0,05
U-iltptilla 1100013b!X KOCTeel camel 8 11-17 14,1 ±0,65 .1,58 3 15-16 15,6 - - CtIME 20 12-15 13.5+0,22 0,97 10 . 13-16 14,3+0,31 1.00 +1,97
/Luna pocTpaohnofi tiac .rn ' ca m e ! 8 103-109 106,3±0.88 2,50 2 101-108 104,5 - - cammz 21 97-115 105,2±0,93 4,28 6 100-119 107,8±2,89 6,85 -0,86
il.inlla Bcpxnero pH,g,a KOpeH- caeu 8 45-48 46.1±0,33 0.96 3 43-49 46.0 - -
111,1X 3y6OB ' • CZIMKt 21 44-50 46,8 ± 0,47 2,17 11 45,9+0,43 1,43 +1,25
11p00abHuii zHameTp aohne- camel 8 10--13 11,8±0.35 0,99 3 11-12 11,6 - - p.ibt Bepxnero Kohnia ca NTE. 21 • 10-13 11,2+0,20 0,92 11 11--14 12,0±0,22 0,72 -2,57 me •
r, camel's- caNika = -1-1j7 ea Nten -CaMxa = +1,97 cameu - cam = +1,79 ca camKa +2,12 male-female
camell - carom +0,88> /7 ca melt - ea mma = +0,86 /9 amen - camiza = +1,21 /9 ea nieu - camKa = +1,50
m male f - female,

398
The cephalopod mulluscs most frequently found Ln the
stomach contents were octoouses, lt is characteristic that
molluscs predominated in the stomachs of sexually mature ani-
mais. This characteristic of the feeding of the harbour seal
has also been observed in the Bering Sea (Gol'tsev, 1969).
Sand and small stones were found only in those stomachs
in which there were contained some benthic animals, which had
been the objects of feeding. It was obvious that these had
been ingested by the harbour seal while hunting for its prey,
as is the case among other species of pinnipeds (Ggnev, 1935;
Kind, 196 )4; Mohr, 1963 and others).
The maximal weight of food found in the stomach (3.27
kg) was discovered in a seven-year old female, containing the
remains of 19 specimens of Korean cod. A weight of food simi-
lar to that found in this seal is not a frequent phenomenon.
S. P. Naumov (1933) reported the finding of 80 specimens of
navaga (saffron cod) in the stomach of a harbour seal caught
on the Shantarskiye Islands. Unfortunately, their weight was
not given.
Distribution. In that part of the Tatar Strait
which was investigated by us, the harbour seal predominated
in encounters of pinnipeds. Females, which had whelped, •with
their young were mainly concentrated 20 - 30 miles from the
continental coast off Sovetskaya Gavan' (Figure 3), and here
there could sometimes be seen up to 77 whitecoats per day from
on board the ship.
Large patches of sexually immature harbour seals were
found in the northernmost part of the region of the studies,
L

399
• not far from the Sakhalin coast. The harbour seals were found
on both flat as well as on hummocky ice.
During the period of observation it was noted that
individual harbour seals whiçh were participating in the repro-
duction were, for the most pà.rt, situated close to the sea
edge of the ice, while the sexually immature and maturing ani-
r;:als kept themselves deep in the ice fields, at times at a con-
szderable distance from the edge.
Some representation of the age structure of the popu-
lation of harbour seals in the Tatar Strait may be obtained
from an analysis of the small series of our samples (76 speci-
mens). Since the hunting was conducted primarily on the whel-
ping patches, in the catch there was a predominance of adult
n.n3.::als and yoinig: of the previous year. There were few sexual-
ly immature animals, aged 1-^^years, and therefore the age
structure of our catch may to some extent characterize only
the part of the herd of bearing age.
1^1•The main part of the catch consisted of seals 5
years old (81%). The remaining 19% comprised animals aged from
15 to 11.1 years. The average age of this part of the herd was
11.8 years.'tTe will note that among the 611. harbour seals that 21I•8
were caught in 1,062 in the Bering Sea, no animals older than
23 years were found (Tikhomzr.ov, 1969). In the Ckhotsk Sea
and Bering Sea populations of harbour seals the average age
is, correspondingly, 11.5 and 9.9 years. The relatively high
average age of the harbour seal population of the Tatar Strait
indicates that it, like the two former populations, has been
little affected by commercial hunting.

4,1 /'i2°
■717 r
U25) IY • 41:,‘ ei3?M7elo r'l Q-ciirc 2
dd •41-9-)./ çt ri
-
A lE CO
r. CO CK
•\- 17 cis\ S *Or S 1
V -
.y Sovetskae Gavadee J rnEro
ç 0[3E1' CK ALI to Ugleh
fiA ri 8v-
5 13 "7r- G
ikoo
Puc. 3. BeTpmemoub Tionenefi n Ta -rapcKom npo:ntne 13
piton C 20 Malrfa no 6 anpeon 1969 r. 060311w-wino-I: A — anpra; B — oaxi.a8; /3 — cuily , t; RphulaT im.
R nun Irreoe — nc -rneveno 110.1011Cii, B 3,IaMelI aTC.1C — Ii TOM qucne 15C.ThKOB. chnrypbt. 1l3o6panienume cunounwil ounncii, — BCTIIC ■10CMOCTt, C 20 no 26 mapTa; npepunucTalt annun — C 27 maivra no 8 anpean; npepbt-
Blic-ran annnsi c TOMBOil C 3 no 6 anpe.in .
Figure 3. The frequency of occurrence of seals in the Tatar
Strait in the period from March 20th to Avril 6th
1969.
Legend: A - harbour seal; B - bearded seal; V - Steller's sea-lion; G - ribbon seal.
In the numerator - seals encountered, in the de-nominator - whitecoats included in above number. Figures, illustrated by a continuous line - en-counters from 1,.arch 20 to 26; interruped line - from iv_arch 27 to Avril 8; interrupted line with dots - from April 3 to 6.

14.01
o
o
249 In the habitats of the harbour seal in the Tatar
Strait there also live other species of pinniPeds. For example,
Steller's sea-lion is encountered here (Naumov, 1933; ügnev,
1935). The former author considered the encounters with the
Steller's sea-lions to be fortuitous.
We have noted considerable accumulations of Steller's
sea-lion. In groups of up to 15 head and singly, these seals
were found near the edge of the ice close to the Sakhalin coast.
Bulls and young animals predominated in these encounters. In
all, we registered 160 Steller's sea-lions. In addition to
these, there were encountered individual,speciplqns of the . .
bearded seal and the ribbon seal.(see
Among the factors determining the concentration of the
harbour seal in particular sections of the Strait, the main
ones, apparently, are the ice conditions and the feeding con-
ditions. According to an oral communication from G. A. Fedo-
seev, A. V. Evzerov and G. P. Kovalev, co-workers from the
T1NRO, who carried out an aerial survey of the seals, on April
1st 1969 in the Tatar Strait the white hummocky ice, suitable
for the whelpinz and resting of the harbour seal, was distri-
buted northwards from the region of our studies for a distance
of 60 miles, to the-latitude of the settlement of Aleksandovsk,
further on up to the narrowest part of the strait there was
slush, and no seals were registered on this. According to our
observations, the southern border of the ice extended not far
from the settlement of Uglegorsk. Here was noted the southern-
most point of encounters with the pinnipeds.

4.02
Under the action of the currents and winds the ice
fields gradually were displaced and extensive stretches of
open water were formed, at times near the continental coast
and at times near the island of Sakhalin. In accordance with
this, the animals were encountered on the edge of the ice
either close to the continent or near the island.
According to V. L. Andreev (1965), in February - April
in the Tatar Strait there occur spawning aggregations of the
Korean cod. These aggregations of the fish shift from place
to place. Probably, following these the harbour seals also
move from one part of the gulf to another.
Periods of' whelping and moulting.
During the period from Llarch 20Y -to April 2 among the encountered
pups there were whitecoats aged 3 - 5 - 10 days. Consequently
the whelping commenced in the second third of 1:.arch, which
once again confirms the conclusions made previously by S. V.
Dorofeev (1935) and G. P. Nikulin (1935).
With the aim of elucidating the conditions under which
the whelPinp.' of the harbour seals occurs, we conducted obser-
vations on the temPerature of the air and of the surface layer
of the water. In the Tatar Strait the temperature of the air
ranges from -2 to -9 ° C, while that of the water ranges from
-1 to 2 °C. It was found that during the period of the whelping
of the harbour seal in various parts of its range (Tatar Strait,
Bay of Peter the Great, Bering Sea) there are no great differ-
ences between the air and water temPeratures.
In many of the harbour seals that were caught in the
first third of April the ovaries already had mature follicles,

1403
which indicates that the animals are ready for ovulation and
the next Pregnancy.
At the end of March - beginning of Aoril the hair coat
of the snotted seals was robust. Among the sexually immature
animals there were found individuals with loosened hair, and
rarely - with portions of bare skin. The first moulting in-
dividuals of the younu harbour seals (ragged-jackets) were
encountered on April 1st.
Recommendations for the commercial
exploitation. It is known that at the present time the
sealing flotilla of the UMRZF* is paying considerable attention
to the procurement of fur pelts of seals. Thus, in the Sea
. of Gkhotsk in recent years the vessels of the UMRZF have been
catching about 6000 harbour seals, among other species of seals.
Because of the severe ice conditions during the period of the
wheloinu of the seals the whitecoats are caught in small num-
bers.
The volume of the procurement of fur pelts may be in-
creased by catches of harbour seals in the Tatar Strait. Ac-
cording to the data from the aerial survey, which was conducted
by the TINRO, about 9000 harbour seals were registered here.
In the past in the Tatar Strait, in the Bay of Tyk there were
caught about 800 head of this species in a year (Gagichko, 1931;
Gagichko ans Surzhin, 1935).
On the basis of the results of the aerial survey, of
the literature sources and the materials obtained during the
cruise of the sealing ship "Sanzar", we recommend the opening
up of a new region for commercial sealing: the Tatar Strait.
*Translator's note. I was unable to find the meaning of this abbreviation.

4o1,.,
Until further information is obtained on the harbour seal,
we propose to establish the quota for its catch for the first
year of the sealing operations at 1000 head.
As has been shown by the experience of the work on
the sealing ship "Sanzar", in years with favourable ice condi-
tions during the period of whelping, in the Tatar Strait it
may be possible to conduct the sealing operations for the
harbour seal both directly from the ship as well as from its
boats. Consequently, whitecoats and beaters may form a con-
siderable portion of the catch. It would obviously be advis able
to commence the sealing at the beginning of Pï.arch.
41J11•ITEPATY PA
1 15esrcux A. H. IIol.bin Bli-Z T IOr7i.!ICi C Kj'pnd!bCK11X oCTponOn -PI o aCinsularis
s^ n. RAt-I CCCP, T. 158, mm. 5, 19G^I.6e.vairt A. l1., Kocbrzurc 1'. il., 17axux. K. H. Hollble nlaTepua:!Lt 110 xapalcre-
I11,CT1!!CC OCTholl!IOrO TIO:IC!!fl.. CG. 4A'IQpCKIio DLRCI<Ol]!1TalO111nC». lIS., 1969.
^-.raau1tno C. !1. Ocetilnlïl lipoN!L1CCn naCTOUOrnX B manne Tbu:. <.PblGnoe xo-
3 tlcTno',Ua:!Lnc!ro I3ocTOxa», Ns l-2, 1931.^-•Fueuw:o C. 1I., Cypxcux C. H. Jlac•ronorue TaTapcsoro npo:111Qa il Amypc!coro
JnI9lana xaK np0\ILIIIS.iC!nloe cLlpbe. TPY;ibl BIiI•iPO, T. 3, 1935.
5^Zopo(1)ee(3 C. 13. A1aTepllaahl x llpO^ulCllosoii 611o1!0ril!t naCT0lfOr!tX B neceu-
lulïl :IC1[ollblü ucpno2 a TaTapexonl npo.nlluC. TpYnbl BIII1PO, T. 3. 1935.6 K,reüxeuüepa C. E., Slonohoa A. B,, K.lceesana r. A., Genbi:vauil B. ,ll.,
3TUx. B. A. Cnpauovuh!e nol<1saTC:m 110 s<;pal(Tel)IICTInCe ne!coTOpb!x ilacrollorl!x
1t KlITOOGpif311b1X. CG. «,^Iopcl:ueaL^er.ouuTa!ou.^ne». \•1., 1^1G5. •DlOpcjlo^or!i^!eCl:Oi{, ^apai:rep!!CT.!niC
^ KOCLL2uH% Ï . M. I'IC1:OTOpL1C DfaTCpna1!LI 110
n,1oRa :IasTal:a. 'fpy1L1 PIIIIPO, T. 65; 113B. "I•kII-IPO, T. 62, 1963.8 Kocnlzurt F. M., Coro loa A. C., Tu.romupoQ 0. A., LUtlcroo A. 17. Iloub!e
j(a11uL1E no nnTE:pbepY aaCT011ornS Cenepll0ïi 9aCTir Tiixoro oliealla. CG. «.^^Iopcr.ue
n! !clmnnTa!ontnc». Al., 1969.9 KOCb121tK r . M., Tuso.rupoa D. A. Jlapra 3an111la IIcTpa Bc 711K01'0. LICTBCpTOe
IiCec0103110C COnelünllnC no 113Y1ICI11!IO 1,101)Cli!IX 19CK01!IfTA1011(IIS. TC311C61^joxna-
Aou. M., 1969.10 !lay.stoa C. 17. Tlo.^!cllu CCCP. 1(oI-i3. i`2.-JL, 1933.

11 Hay.ston C. IL, CIIIIPH08 II. A. AlaTepitaabi no ClICTCMaTIIKU If reorpadmiecoomy pacupeite.neinuo Phocidae celleplion Tiixoro oocana. Tpyina BHPIPO, r. 3, 1935. 17 (knee C. 11..3Bepu CCCP ii npihneffluntx man, T. 3. MAL, 1935. 1.3 Iluxapeu I'. A. Tio.rienii ioru-aanannoii ‘IiICT11 Oxorcooro mopn. 113n. TI111P0,. T. 90, B:10,11.11110CTOK, 1941. 1,
POKIII{KIIII 17. ‘1, . Ocionna napilaimonna CTHTHCTI1KH T(.1151 6110aoron. 1961. -
Cieupnon H. A. 1.1ncTpyounn ,11;111 nomemix pa6or no 6iionoriiii cup:nun it siporpaiamid no inyileinuo npoiabic.nonoii cliaynia ii flp0Mblen0B. t. 1, ) 93l. J. CoK0.400 A. C. BCC0111.151 xapao -repitcruoa mianin opraiion ninuoeinin .macrono-nix. C6. «,\'Iopcolie ffleooriiiraiounie». M., 1969. 17 coso.um A. C. lieltOTOphIC 11T01- 11 H nepcneicrunm i(ciaoro-mopilianorinteconx IlccJIeJWnaliiIii THX0Olie.1111CKIIX Jiacronornx. C6. «Mopcone maeoonirraonnue».
re. C000ion A. C., Kochterm 1'. M., Tuxomupoe D. A. HeooTopide cneuenun o nece nny-rpeumix opranon siacronornx Bepunrona mops'. 143e. T141-1P0, T. 58. B.ria.au-BOCT01:, 1966. 19 C000..con A. C., .Kocbteun T. M., Illycroe A. II. Crpoeinie MerIiIIX 11 rpaxen y 6epiiiironomopconx ;lac -mot- 11x. T. 68. l'1311. T1.11-11)0, T. 62. M., 1968. 2 0 COK0.706 il. C., KOCb1el111 r. M., Ky31i11 A. E., Ilep.non A. C., Taxomupoci D. A., Illycron 17. Cripanotinbie 1101:a311TCAll no xapao-replicTitoe rmyrpeninix °prima Tilmonceancoux aac.roliorirx. C6. «I ■ lopeone ta.rieoonirraimitue». m„, 1969. 21 Tux0,1wpou .9. A. 0 TC1■111X nociipoilanoc.risa cenepo'riix000eznicoux T10:1C11Crl.
C6. «Alopcoini m.leoonirr a onmie». M., 1969. 22 Taxo.unpoa A„ Krietrearmb T. A. Meroulit oripeue.nemin 1303HaCra I101:0TO-
pux :1/ICT01101- 11X. B Ku.: eGiipeaesienue noapacrit nporaliteaonbtx ;lac-11moms: u pa- wionamblIOC IICH0.11,3011anite MOHC1:11>: M.11(.1:01111T11101101x>>. M., 1964. Z3 ganciaiii K. K. Onbrr nepecmoTpa cucTemia li manioc-molt rio.ieiieii noncemefi-cipa Pliouinae. Tp. T. 17, JI., 1955. 2 1 1. LInnentdi K. K. Orpiu Pinnipedia siacronorue. B ou.: «MneooluiTaionnie (Pay-
CCCP».1963. 25 ' ,/anckiili K. K. Mopqm.rioro-raocononintiecoan xapaorepucruoa Jinni
mush Tpy;(1,1 1 -11111P0, T. 21, Mypianco, 1967. • 2 0 /llyeron il. /1., f16.1oKoo A. B. Cpannivremaio-inopqm.imrunecoan xapaorepncrii-oa rpeimanRcooro n nomocaToro rio.rieneiÏ. TpyRta 1-I111IP 0 , 111111. 21, Alypmanco, 1967. 27 )76.fienwe A. 13. Heouropide ripe/inapirremaibie recreplia.nbt K mopilicaoriniecoort xapilKTCplICTIIKe rpeumailienoro T10J1C1111 15C.,10TO H Ppen.rianucooro raopefi. B c6.
liccneRon. pa6or 1962 T. (rpen.rialui,coliii T10,11C111, 11 xox.riati). Apxaureabco, 1963. 28 'M./Iowa A. B. 143mentinnocTb releKolurraionuix. M., 1966. • 29 516.aoKna A. B., Kiieneanizb r. A. Bn6puccia oirroo6pa3max u .nacronornx, ux pacnpe.c.ieii K', crpoenue 11 anal-feline. Mopipo.nornuecone oco6einiocrit nomibtx wieoomiraionnix. M., 1964. 30 Allen 1. A. History of North American Pinnipecls. U. S.- Geogr. S. Z. V, Ferrit. Publ., no .12, 1880. 31 Allen I. A. The Hair Seals (family Pilo' cidae) of the North Pacific ocean and Bering Sea. Bull. Amer. mus. nat.-- History, vol. 16, 1902. 32 Chapskiy K. K. Morphologie Systemaiique Differ'enciation Intra-specifique et Phylogenese du Sous-Genie Phoca sensu stricto Mammalia, t. 24, no 3, 19(30. 33 Doutl K. A review of the genus Phoca. Annals of the Carnegie Museum, vgl. 29, art 4, 1942. 3 14. Kenyon K. W. Food of harbor seals at Amchitka Island, Alacka. I. eMam-mol.», 46, no 1, 1965. 3 .D King 1. E. Seals of the World. Trustes of the British Museum (Natural History), London, 1964.
251
4.05

406
• 36 Mohr E. Ein neuer Westpazifischer Seelitind. Zoolog. Anz. Bd. 133. Nt 3/4. tiarnbur g, 1941.
• 37 -Mohr E. Dic Steine in Robbenrnagen. «Z. Situgetierlcunde», 28, no 3, 1963. . 3bike A4ohr E. Uber Phoea vitulina largha Pallas, 1811 .und Weissgeborene Seeh-
• niy.s. de . «Z. Siiiigetierkunde». Bd. 30. Hamburg. 1965. 3y Sell:4W 1/. B. Seals Sea Lions and Walruses. Stanford, 1958. LFO Scheiter V. B. Weights of organe- and glands in the Nortftprn Fur Seal. hammalia. vol. 24, 3, 1060.
•
REFERENCES
1. Belkin A. P. A new species of seal from the Kuril Islands,
Phoca insularis sp. n. Doklady AN SSSR (Reports of
the USSR Academy of Sciences), vol. 158, no. 5, 1964.
2. Belkin A. N., Kosygin G. M. and Panin K. I. New materials
on the characteristics of the island seal*. Symp.:
"Marine mammals". Moscow, 1969.
3. Gae-iohko S. I. Autumn sealing in the Bay of Tyk. "Rybnoe
khozyaistvo Dal'nego Vostoka" ("The fisheries of the
Far East"), No. 1-2, 1931.
4. Gagichko S. I. and Surzhin S. N. The pinnipeds of the
Tatar Strait and Amur estuary as commercially exploit-
able raw material. Trudy VNIRO (Transactions of the
All-Union Research Institute of Marine Fisheries and
Oceanography), vol. 3, 1935.
5. Dorofeev S. V. Information on the applied biology of
pinniPeds during the sPring ice period in the Tatar
Strait. Trudy VNIRO, vol. 3, 1935.
6. Kleinenberg S. E., Yablokov A. V., Klevezal' G. A., Bel'ko-
vich V. M. and tin V. A. Reference indices for the
characterization of certain pinnipeds and cetaceans.
Symp.: "Marine mammals". bloscow, 1965.
* Translator's note. "ostrovnoi tyulen" - literally "island seal", probably refers to the Kuril seal (Phoca stejnegeri)

407
7. Kosygin G. M. Some data on the morphological character- ,
istics of the foetus of the bearded seal. Trudy VNIRO,
vol. 68; lzvestiya T1NRO (Proceedings of the Pacific
Ocean Research Institute of Fisheries and Oceanography),
vol. 62, 1968.
8. Kosygin G. M., Sokolov A. S., Tikhomirov É. A. and Shustov A. P.
New data on the.interior of pinnipeds of the northern
part of the Pacific Ocean. Symp: "Marine mammals".
Moscow, 1969.
9. Kosygin G. M. and Tikhomirov É. A. The harbour seal in
the Bay of Peter the Great. Chetvertoe Vsesoyuznoe
soveshchanie po izucheniyu morskich mlekopitayushchikh.
Tezisy dokladov (Fourth All-Union Conference on the
Study of Marine Mammals. Reports of Proceedings).
koscow, 1969.
10. Naumov S. P. Seals of the USSR. KOIZ (Vsesoyuznoe koope-
rativnoe izdatel'stvo - All-Union Cooperative Publishing
House). Moscow - Leningrad, 1933.
11. Naumov S. P. and Smirnov N. A. Materials on the systema- 251
tics and geographical distribution of the Phocidae in
the northern part of the Pacific Ocean. Trudy VNIRO,
vol. 3, 1935.
12. Ognev S. I. The animals of the USSR and neighbouring
countries, vol. 3. Moscow - Leningrad, 1935.
13. Pikharev G. A. Seals of the south-western part of the
Sea of Okhotsk. Izvestiya TINRO, vol. 20, Vladivostok,
1941.
1•. Rokitskii P. F. Fundamentals of variational statistics
for biologists. Minsk, 1961.

• 408
15. Smirnov N. A. Instructions for field studies on the
biology of seals. Instruktsiya i programmy po izucheniyu
promyslovoi fauny I promyslov (Instructions and programmes
for the study of exploitable fauna and industries based
on this), No. 1, 4, 1934.
16. Sokolov A. S. The weight characteristics of the muscles
of the locomotory organs of pinnipeds. Symp.: "Marine
mammals". Moscow, 1969.
17. Sokolov A. S. Some results and prospects of ecological
and morphological studies on Pacific pinnipeds. Symp.:
"Marine mammals". Moscow, 1969.
18. Sokolov A. S., Kosygin G. M. and Tikhomirov A. Some
information on the weight of the internal organs of the
pinnipeds of the Bering Sea. Izvestiya TINRO, vol. 58.
Vladivostok, 1966.
19. Sokolov A. S., Kosygin G. M. and Shustov A. P. The struc-
ture of the lungs and tracheae in the Bering Sea pinni-
peds. Vol. 68. Izvestiya TINRU, vol. 62. Moscow, 1968.
20. Sokolov A. S., Kosygin G. M., Kuzin A. A., Perlov A. S.,
Tikhomirov A. and Shustov A. P. Reference indices
for the characterization of the internal organs of the
Pacific pinnipeds. Symp.: "Marine mammals". Moscow,
1969«
21. Tikhomirov El. A. Topics on the reproduction of the seals
of the North Pacific. Symp.: "Marine mammals". Mos-
cow, 1969.

409
22. Tild-iomirov É. A. and Klevezal' G. A. Iiiethods for deter-
mining.the age of certain pinnipeds. In the book:
"Determination of the age of commercial pinnipeds and
the rational exploitation of marine mammals". Moscow,
1964.
23. Chapskii K. K. An attempt at a revision of the systema-
tics and diagnostics of seals of the subfamily Phocinae.
Trudy Zoologicheskogo Instituta '(Transactions of the
Zoological Institute), vol. 17, Leningrad, 1955-
24. Chapskii K. K. The order Pinnipedia, the pinnipeds.
In the book % "A°,ammalian faunas of the USSR". A:oscow -
Leningrad, 1963-
25- Chapskii K. K. Morphological and taxonomical character-
istics of the harbour seal of the Bering Sea. Trudy
PIhlRG (Transactions of the M. Knipovich Polar Research
Institute of D':arine Fisheries and Oceanography), vol.
21, Murmansk, 1967.
26. Shustov A. P. and Yablokov A. V. The comparative morpho-
logical characteristics of-the harp and ribbon seals.
Trudy PIIVRC, issue 21, Murmansk, 1967..
27. Yablokov A. V. Some preliminary data on the morphologi-
cal characteristics of the harp seal of the White and
Greenland Seas. in symp.' of reseach studies 1962
(harp seal and hooded seal). Arkhangelsk, 1963-
28. Yablokov A. V. Variability in mammals. Moscow, 1966.
29. Yablokov A. V. and Klevezal' G. A. The vibrissae of
cetaceans and pinnipeds, their distribution, structure
and significance. Morphological characteristics of
aquat i c mammals. 141o s c ow , 1964.

4.10
32. Chapskiy K. K. The systematic morphology, intra-specific
differentiation and phylogeny of the subgenus Phoca
sensu strictu. Mammalia, vol. 24, no. 3, 1960.
36. Mohr E. A new seal species of the West Pacific.
37. Mohr E. Stones in the stomachs of seals.
38. Mohr E. Phoca vitulina largha Pallas, 1811, and seals
that are white at birth.

4, 11
UDC 599.745.2
V. A. Bychkov
THE SEASONAL VARIATION IN THE DENSITY OF THE FUR IN 253
THE HAIR BUNDLES OF THREE-YEAR-OLD BACHELOR FUR SEAIS
The three-year-old males of the fur seal, as compared
to other age groups, possess the most valuable fur and comprise
the main group that is exploited for commercial purposes.
Therefore, the study of the seasonal variation in the density
of the fur in the hair bundles of this particular age group
of fur seals is of considerable importance for determining
the optimal periods of the kill.
Studies which were carried out on the Pribilof Islands
showed that in all of the fur seal‘males (including also the
three-year-old males) the density of the fur in the bundles
remains almost unchanged during the course of the winter,
spring and summer (i.e. for the greater part of the annual
cycle of their life) and only in the autumn, when the new
hairs grow more rapidly, does the total number of the underfur
hairs in the bundle increase (Scheffer and Johnson, 1963).
Similar studies have not been conducted on the fur
seal population which reproduces on Robben Island. In connec-
tion with this, the present study is intended to elucidate the
characteristics of the variation in the density of the under-
fur hairs in the bundles on the body of the three-year-olds
in relation to the Periods of their kill e -on the basis of
which refinements may be made in the optimal periods for the
sealing operations on the fur seals on Robben Island.

412
The materials for the study of the seasonal variation
in the density of the fur in the bundles of the three-year-olds
were.collected on Robben Island from June to November during
1959 - 1961. There were collected 120 samples of pelt from
24 tagged bachelors of the local population of fur seals. The
samples, which were 2 - 3 sq. cm in size, were taken from the
mane, back, lumbar region, sides and belly (Figure 1), and
were fixed in a 10% solution of formalin. Histological prepa-
rations were prepared from each sample specimen in the form
of series of sections crosswise to the root part of the hair
bundle. The sections were prepared on a freezing microtome,
stained with haematoxylin and eosin, stuck onto a microscope
slide, embedded in gelatine-glycerin and covered with a cover
slip, according to the methods given in the textbook by G. I.
Roskin and L. B. Levinson (1957). The thickness of these
sections was 15 - 30 r . The evaluation of the density of the
fur was carried out under the microscope (with a magnification
of a X7 ocular and X8 objective) in the root portion of the
hair bundle at a depth of 1.0 - 1.5 mm (Figure 2). For each
sample there was determined the density of the fur in 25 homo-
geneous hair bundles. The hairs in each bundle were counted
three times and the average was taken as the "datum" for set-
ting up the variational series. In grouping these "data", the
class interval (i) equalled 5. The arithmetic mean (x), the
standarddeviaticn"cr50' thestandarderr°rofthemean' ) ' (mx
the confidence limits of the mean (5-c.± t 3m) and the confidence
limits of the series in question (51± teR) were calculated, to
one decimal place, by the methods recommended by N. A. Plokhin-
skii (1961).

4,13
rscv; `ls•^ ff':^^'',5;^°
.1^
^^^ ^:.\
^`^ `^.̂1
pile. 1. CxenlaTn1lnoe n3o6paxtellne Tynln xo^QcT»xa
co cnlnlm. (TollhaMll y1:a3al1U1 VtIaCTiQ[ 113flT
414
Pile. 2. 06milii isue, nonocanoro nyrwa na nonepemilom epe- • se iia ray6iiiie 1,0-1,5 .4t.e. boBepxtiocTil izom:11
– -- •
Figure 2. General appearance of a hair bundle in cross
• section at a depth of 1.0 - 1.5 mm from the surface of the skin. •
of fur in the bundles were most frequently found in August -
September. Furthermore, it was established that neither in
October nor in November were there animals among the examined
three-year-olds, in which the numerical values of the modal
classes of the density of the fur in the bundles would exceed
the corresponding indices in June - July.
Even in a comparison of the relative indices of the
density of the fur in the bundles on different parts oe the
body, only in one case was it noted that, in the fur on the

29
95
27
26
28
27
30
23
18
29
20
18
91
26
94
27
25
22
24
16
28
24
15
November
T a 6 el 1111a
Koantiecruo nyxoubtx no:toc n nytuce Ha rime Tpex..nemux xo.nonnicoa n opcnoro 1i:canna
Date of kill .riaTa AO6b14II 511Cp31
of animal
, Modal moe.mei
K111CC
class
Maxi-- Mean Mruzcit- CPung
mym
MUI
Confidence limits JLoiaeiiaJth'I hie
rpaumm
cr7
Mini upi MUM
June 24 moun 1960 r.
24 ». »
29 » »
29 » »
July 2 1110:1fl 1959 r.
10 » »
july 24 1110A51 1960 r.
24 » »
August 10 aurycra 1959 r.
• 11
• 11
• 11
September4ce 6 MrS urn_pst r.
. »
18 » >> •
ShtembeD2 ceirra6pn 1959 r.
Otober
>>
» »
8 non6pn 1959 r.
10 » »
10 » >>
10 » >>
33-37
33-37
38-42
33-37
33-37
38-42
38-42
33-37
28-32
28-32
28-32
28--39
23-27
23-27
33-37
28-32
38-42
33-37
28-32
33-37
33-37
38-42
33--37
28-32
44 36,8 -1- 0,65
46 36,6+1,22
38,4±1,05
49 35,4.10,84
44 36,4+0,81
44 362±0,98
47 37,010,94
45 34,6+0,93
38 28,4:11,02
43 28,6-10,72
43 32,4+1,27
38 30,2 ± 0,96
33 25,2±0,77
37 27,0+0,70
42 33,2-10,79
41 32,4±0,94
47 37,9+0,99
39 32,4:10,80
49 33,0-1.1,09
47 36,2±-0,90
44 35,4-.11,29
48 36,2-11,10
43 36,0±0,89
34 28,6+0,80
31,1-43,5
. 23,2-50,0
27,4-49,4
26,5-44,3
27,9-44,9
26,1-46,3
27,1--46,9
24,3--44,a
17,7-39,1
21,0-36,2
19,0-45,3
20,1-40,3.
15,1-33,3.
19,6-34,4
24,9-41,5.
22,5-42,3.
27,4-48,4
24,0-40,3
21,6-44,4
26,8-45,3
21,8-49,0
24,7-47,T
26,6-45,4
20,2-37,0
21 onrn61)11 1959 r.
23
23
26
256
Table 1.
The numbers of fur hairs in a bundle on the mane of three:7..year7ol.d._ ba.chelors__of... the.. seal.

Da
o
4ne
Jialy
Jiu1ly
ALg.
•
1
Sept.
Oct.
Nov.
1
T a .1 n, a 2
K0,111 1 1CCTLIO nyxonbix »mac U arum Ita mine TpexacTnnx. X0.110CTIIKOIS mopcicoro hOTJIKfl
, ,.;9nIictence ce of kill tini-odalbraxi mean biteptil eamthte Munn- Mo.n.i mud rt Ma melt- ,-Peallsig rpa Itil am
.1-1.a ra A061•1`111 -I Lie P`i mym Kaacc mym :.7÷ III:,:- x -F t1 a— C> animal mura clasi._., mum - x
24 Inomi 1960 r. 27 33-37 45 35,8 ± 0,88 26,6-45,0
24 >> » 26 38-42 47 37,4±1,10 25,8-49,0
29 >> » 31 38-42 47 40,0+0,85 31,1-48,9
29 » » 29 33-37 42 35,8 ± 0,71 25,4-43,2
2 inOnst 1959 r. 32 38-42 47 40,8±0,84 32,0-49,6
10 » » 29 33-37 45 36,4±-1,04 25,4-46,4
24 moan 1960 r. 32 38--42 46 36,8 -±- 0,93 27,1-46,5
24 >> » 24 28-32 46 33,8±0,91 24,4-43,2
10 anrycTa 1959 r. 2-1 33-37 - 43 33,8±1,14 21,9-45,7
11 >> » 27 33-37 49 35,2:9,66 28,3-42,1
11 >> » 20 28-32 45 31,2±1,17 . 18,9-43,5
11 » » 19 28-32 35 29,0-9,89 19,7-38,3
4 ceni Am 1958 r. 20 28-32 38 29,2-2:0,88 20,0-38,4
8 » » 23 28-32 38 30,2±0,87 21,1-39,3
18 >> » 25 . 28-32 41 31,6+0,84 22,8-40,4
22 ceiln 6p g 1959 r. 20 28-32 39 31.0±-1,06 19,9-742,1
21 oKrepn 1959 r. 24 33-37 45 35,6±1,14 24,6-46,4
23 » - >> 26 33-37 46 36,2±0,86 27,2-45,2
23 » » 20 33-37 42 34,4±0,95 24,4-44,4
26 » » 29 38-42 46 . 38,6±0,75 31,0-46,2
8 non6pn 1959 r. 14 33-37 49 3Q,2±1,63 19,0-53,4
10 >> » 20 33-37 43 32,6+1,20 20,0-43,2
10 » » 19 38-42 45 36,61:1,16 24,4-48,8
10 » » 25 28-32 36 31,2+0,66 24,3-38,1
limits
17 3aK. 2354
Table 2.
95T.
The numbers of fur hairs in a bun.dre on
_the_back of ..three-.year-old bachelors, of
the fur seal.

•
;e of kill Mini ModàlLicx..:- Mean ,11-eZWOW(5-
iiaTa Rowni 3„pg M\1,11,,Itl- M(Ui.a.,:litcll,lii M;I,K,Ce lfl- _q1) . 111,151 rpallIllthl X 1". Mx . Jt' ÷ t a-
- 1 .1.
r: animal mura Class In'15.1M
24 11101151 1960 r. 27 33-37 42 35,4±0,84 26,6-44,2
24 » » 25 33-37 46 34,2± 1,05 23,3-45,1
29 » » 25 38-42 46 37,0±1,28 23,6-50,4
29 » » 25 33-37 40 32,4±0,90 23,0-41,8
2 mo.9 a .1959 r. 31 33-37 43 36,6±068 29,5-43,7
10 » » 29 33-37 44 35,0+0,75 27,2-42,8
24 11102151 1960 r. 23 33-37 40 33,6±0,94 23,7-43,5
24 >> » 26 33-37 45 35,0 -±- 0,94 25,2-44,8
10 aBrycra 1959 r. 19 23-27 35 25,2+0,96 15,1-35,3
11 » » 25. 33-37 42 33,0±0,5 26,1-L-41,9
11 » » 17 28-32 38 30,0 -I= 1,09 18,5-41,5
11 » »' 16 23-27 35 26,2±1,06 15,1-37,3 .
4 ccurepn 1958 r. 26 28--32 41 30,8±0,73 23,0-38,6 -
8 » » 24 28-32 38 31,4±0,72 25.9-38,9
18 » » 24 33-37 42 32,8±0,99 22,4-43,8
22 ccurn6p1 1959 r 19 23-27 40 27,2±1,06 16,0-38,4
21 own 6p a 1959 r. 24 28-32 39 30,4±0,85 21,6-39,2 '
23 » » 24 38-42 43 36,6±1,05 25,6-47,7 .
23. » » 26 33-37 47 37,2±1,05 26,2-48,2
26 » , » 22 33-37 43 33,4 -± 1,1 21,6-45,2. .
8 1Io116pu 1959 r. 12 38-42 47 40,4±1,70 22,5-58,3
10 » » 11 38-42 44 35,4±1,51 19,5-41,3
10 » » 28 33-37 42 36,0+0,69 28,8-43,2
10 » » 23 28-32 35 29,4±0,65 22,6-36,2
limits Da
o
June
July
July
August
Sept.
Sept. Oct.
NOV •
Ta 6 muua 3
KOJ111 ,1CCTII0 uyxonox Bo.noc a urixe im 11051C11111e
TOCXJICT1111X XOTIOCTOKOB MO )cuoro KOTIUt
• 258
Table 3. The numbers of fur hairs in a bundle on
the lumbar region of three-year-old
bachelors of the fur seal.

^^
llate of kill›TA AOGL.^ut sne^a
oi^ animalI
19G0 r.June 24 1110119
»
29 » »
24 » »
Juiy 2 1110nA 19â9 r.
10 » »
1Jua.y24 }nonsl 1900 r.
24 » »
ug . 10 anrycTa 19,-)5) r.
il » »
11 » »
11 » »
1Co7114CCT}t0 nyx06L1X rnaac n}ivuce }la GOKY
TPCXTiCTI}I;X X0:lOCï}:KOC A10[iC1C}1X KOTHKOü
Sept • 4 ceuT}IGps} 1958 r
.1Sept. 22 celmiGpsl 195J r.
21 oxTS}ûpn 1959 r.
23 » »
23 » »
26 » »
Nov • 8 noslGps} 1959 r.
10 » »
10 » »
10 » »
17*
22
1,.odaliGio;ta.it,utatl
kT3cc
class
Pâxi,üat:cu-
mf \(
muni
33-37
33-37
38-42
33-37
38-42
38-42
38-42
33-37
23-27
33-37
28-32
28-32
28-32
28-32
23-27
28-32
33-37
33-37
33-37
38-42
43-47
33-37
38-42
28-32
4G
!^-.e anCIte.lnst st
.r±ni.C
36,0.-L 1,0G
34,0=0,98
37,G-!-1,10
34,2±0,92
37,2 ±0,75
38,8:L0,^02
3G,8z:E 1,09
32,8:h0,89
2G,1± 1,01
35,4±0,G9
31,0-±-1,02
29,1-±- 0,8G
30,4±0,93
30,8 ±0,84
27,0±0,97
31,G± 1,09
33,2±0,9G
37,2±0,70
35,0-1-0,94
38,2±0,93
42,0.±:1,06
30,G:L 1,39
37,2 ±0,7G
31,0 ± 0,75
TaG.-1 nüa 4
Cunfi-d-91rce l imit sAonepnTeat,tiwe
rpcwnma .
.l _fl ..•
24,9-47,1
23,7---44,3
2G,1-49,1
24,6-43,8
29,3-45,1
30,2-47,4.
25,3-48,3
23,4-42,Z'
15,5-37,3
28,2-42,G
20,3-41,7
20,1-38,1
20,7-40,1
22,0-39,G
26,8-37,2
20,1-43,1
23,2-43,2
29,7-44,7
25,1-44,9
28,4-48,0
30,8-53,2
15,9-45,5
29,2-45,2
23,1-38,9
259
Table 4.The numbers of fur hairs'in a bundle on
threé'year-^old bachelors of
the fur seal.
23
20

419 •
24
25
24
22
18
23
21
25
13
19
14
18
18
20
21
18
11
21
24
17
11
G
18
15
;
Sept.
Sept.
T a .1 n 1.t a 5
KOJIIPIeCITO nyxonmx BOJIOC i nruce na 61noxe . TpexaeTnnx. XOJIOCTIII(011 mopcicoro »Tun
Date of kill AaTa ao6lagn »elm
of animal,
• Modal Di .ace mym class mun
Jean ni7
-C-ontidence limits rffl .u. Munn-
mym
MUM
June 24 'Ilona 1960 r.
24 » »
29 » »
29 » »
July 2 mail 1959 r.
10 »
July 24 nio,nst 1960 r.
24 » »
August 11 » »
11 » »
11 » »
4 con -15161m 1958 r.
8
18
22 cenuGpa 1959 r
ÙCtObel^ 21 cuanGpn 1959 r.
23 »
23 >>
26 .»
8 nodipn 159 r.
10 »
10 »
10 .»•
33-37
33-37
33-37
28-32
33-37
28-39
28-32
28-32
18-22
23-27
28-32
23-27
23-27
23-27
28-32
23-27
18-22
33—r
33-37
33-37
33-37
18-22
33-37
18-22
32,3+0,89
31,8±0,89
34,6:1=0,93
30,2+0,99
34,2-±0,84
31,81=0,99
29,6±0,84
32,0 ± 0,80
21,2±0,M
25,8±0,67
27,4 1=0,90
26,6 0,89
24,0+0,69
24,8 ±0,77
27,8±0,81
25,0+0,80
22,0±0,97-
31,6:1:0,89
34,8+0,87
31,4±.-1,28
34,8+1,78
21,8±1,41
34,2±-0,84
20,2+0,72
22,9-41,7
22,4-41,2
24,9-44,3
19,8-40,6
25,4-43,0
21,4-42,2
20,8-38,4
23,6-10,4
12,4-30,0
18,8-32,8
18,0-36;8
17,3-35,9
my-31,3
17,1-32,5
19,0-36,6
16,6-33,4
11,8-32,2
22,3-40,9
25,7-43,9
17,9-44,9
15,9-53,7
6,9-36,7
25,4-43,0
22,6-27,8
39
40
42
39
39
40
38
39
33
3 9
33
33
31
36
37
34
30
38
43
44
40
30
39
26
10 anrycia 1959 r.
NOV
260
Table 5. The numbers of fur hairs in a bundle on
the belly . of three-year-old bachelors of
the fur seal.

Mane
Back
Lumbar region
Si de
Belly
420
4 0 -
f- - -...
30 - 0. c \ , 2 5 - •
40- Cp 1- - -i■ -71- - ... 3 5
- - --. ■̀. '
'., 30- - C
25 - .
40 -
35 - f- - --f. , , 4- - --i 30 -
• ' 1- - -}- tC 0 25 - C
it 0 -
k 35 - I- - -I- r̀) 30- ,0 25 -
0 , 'ç 35 - 4 f- -- i,, , ck 3o—
-f - 1 . z `() 25-
i , . 1 , c ' t) e> V/ V// V/// IX , X X/
>> k U 44 e C e U bf
1. 2 Months 1- parts Pan. 3. Cpe,queB3nemennaa (x:=1:_t atn4 2- amount
ceaonnoro pa3noo6pa3n1 rycToni nyxa B nyincax na 'rule XOSIOCTBKOB 3 OCT
Figure 3. Weighted mean (5I± t3m) of the seasonal variat4n
in density of the fur in the bundles on the body
of 3-year old bachelors.
side of an animal, caught on the 8th of November 1959, the
value of the mode was found to be higher than in animals that
were killed in June - July. The analysis of the individual 261
mean values and confidence limits of the variation in the den:.
sity of the fur in the bundles of the three-year-olds also
indicates the small probability of finding in uctober - November
animals on the body of which there would be found bundles
containing more fur hairs than in June - July.

4.21
The above described characteristics of the change in
the density of the fur in the bundles of three-year-olds is
manifested even more obviously in ,a comparison of the weighted
mean values of this feature (Figure 3). Fromt}ae figure pre-
sented it can be seen that over the entire body-of the bache-
lors the numbers of hairs in August - Septembur are lower
than in the other periods (this•differencé is found to be
statistically significant), while in the comparison of these
same indices in the animals that were killed in June - July
with the animals killed in October - November'only a small 262
difference was detected between these, and moreover this was
not statistically significant.
On the basis of the data obtained, it is not possible
to agr.ee with that interpretation of the seasonal variation,
of the density of the fur in the bundles in bachelors, which
was given by Scheffer and Johnson (1963). The scheme that
was indicated by them is, apparently, atypical. In our opi-
nion, it is more reasonable to propose that the seasonal
changes in the density of the fur in the bundles proceed ac-
cording to the following scheme: as a result of the massive
falling out of the fur hairs of the previous year's generation
their numbers in the bundles on the body of the bachelors de-
creases markedly in August - September. Then, during October
- November, as the fur hairs of the new generation grow up,
the density of the fur in the hair bundles again increases,
but on the average it does not exceed that amount which is
found on the bachelors at the beginning of their appearance
on the hauling grounds in June - July. Taking into account

422
the fact that at the moment of the departure of the bachelors
from the hauling grounds the majority of them have not com-
pletely moulted, the possibility cannot be excluded that the
growth of some portion of the new fur hairs will continue
during the marine period of their life, as a consequence of
which the total number of fur hairs in a bundle will build up
during the winter.
Thus, from the data presented above, it may be conclu-
ded that in August - September the quantity of fur hairs in
the bundles on the body of the three-year-olds is, as a rule,
lower than during other periods of the annual cycle of their
life. However, the violent falling out of the fur from the
bundles in August - September, in its turn, leads to the si-
tuation that the total density of the fur hairs on the skin
of the three-year-olds at this time also decreases. Conse-
quently, under all conditions August - September cannot be.
recommended for the sealing operations on fur seals on Robben
Island.
j1I-ITEPATYPA
. 1 POCKI1H I'. II. .aesancon II. E. Min:pocizontmeexasi Tex a. M., 1957. • 2 1-1.7oxunci;aû il. A. Bname-rplist. 143,-t-uo CO CCCP, Honocemtl)m, 1901. 3 &holier V. B. and Johnson A. AI. MGR in the Northern fin seal. U. S. Dept. Jnter., Fish. Serv., 450, Washington D. C. 1963.
REFEREN CES
1. Roskin G. I. and Levinson L. B. Macroscopic technique.
Moscow. 1957.
2. Plokhinskii N. A. Biometrics. Publ. Siberian Branch
of the USSR Academy of Sciences, Novosibirsk, 1961.

4,23
UDC 599.711•5.2
G. A. Nesterov
THE EFFECT OF THE MUSCLE RELAXANT DITILINE ON THE ORGANTSIvI _ 263.
OF FUR SEALS
In 1963 on the Komandorskiye Islands there were set
up the first tests on the use of the curare-like preparations,
ditiline and diplacâ:n, in the slaughtering of fur seals. Di-
tiline was found to be the most suitable for these purposes.
The effect of the preparation commences after 30-II•0 seconds.
At the moment that the fur seals became completely immobile
they were exsanguinated with a knife.
This method of slaughtering, in comparison with that
existing at the presemt time (killing with a club), has seve-
ral advantages. It is more humane and permits selective
slaughtering to be carried out. The meat and meat products
are suitable for food. It is more economical, since it elimi-
nates bruisings on the pelts. In a slaughter of 10 000 fur
seals the profit may comprise II-5-50 thousand rubels. All of
this permits this method of slaughtering to be recommended for
introduction.
In the selection of methods for the pre-slaughter
stunning of the animals, an important criterion is the possi-
bility of retaining the cardiac activity in the animals, which
is necessary for a better exsanguination. The latter is also
important in the sealing operations on the fur seals, and there-
fore we set ourselves the aim of determining the effect of
ditiline on the organism of the fur seals.

1-12k
From the literature sources it is known that during
the narcotization of animals with non-lethal doses of ditiline
there are not noted any drastically expressed changes in the
blood circulation and respiration (Zhulenko, 19631 Zhulenko
and Korneva, 1965).
Similar studies were carried out by us with fur seals.
As a result of this work there were determined the mean effec-
tive, optimal and lethal doses of ditiline. The effect of the
preparation on the cardiovascular and respiratory systems and
on the body temperature was studied, and at the same time the
consequences of the effect of ditiline on these animals were
revealed.
METHODS OF STUDY
The investigations on the immobilization of the fur
seals with the muscle relaxant were carried out on animals .
which were kept in cages. The animals were first weighed,
after which 5% ttnd 10% solutions of ditiline in various doses
were administered intramuscularly. In the experimental ani-
mals electrocardiograms were taken, the respiration was mea-
sured and the body temperature was measured every five minutes. 264
The electrocardiogram was recorded from 3 standard points of
contact at a current voltage of 1 mv. The changes in the bio-
electrical potentials of the heart under the influence of di-
tiline were registered from the fore and hind flippers on pho-
tographic film with the aid of a portable EKP-60 electrocardio-
graph (Figure 1). The respiration was measured only in animals
which were not held down. The body temperature was measured
rectally.

425
Puc. 1. Dpucoe.nunenne wiewrpo,],on K
Te.ly KoTru.:a npu perucTpanuu cTau- AapTubix celnciummii— I, II, III
Figure 1. The attachment of the electrodes to the body of a fur seal for recording from the standard con-tact points - I, II, III. •
DESCRIPTION AND RESULTS OF EXPERIMENTS
For the purpose of determining the mean effectiveness,
the optimal and lethal doses for the fur seals,there were
tested the following amounts of the preparation: 0.25; 0.5;
0.75; 1.0; 1.25; 1.5; 1.75 and 2.0 mg per 1 kg weight of
the animal. Each dose was tested on 6 male fur seals, aged
2 - 4 years.
The results of the tests indicate that 0.25 mg/kg shows
no effect. A weakly expressed narcotization sets in at doses
of 0.5 and 0.75 mg/kg, within which range was found the mean
effective dose. The optimal immobilizing dose, which brought
about a clearly expressed immobilization of the animal in all

5
ilereTBIle 2111TWOUTA
1.r10 2,00_ Effect of ditiline
He WICITIIWIO Did not ensue Flammwm Ensued' aemumAmmuLethal out corne
0
5
1
9
6
426
of the tests without having a lethal outcome, is'found to be
between 1.0 and 1.25 mg/kg. A dose of 1.5 mg/kg causes a
stoppage of breathing (apnea) in individual animals. Death
always ensued after the administration of 2.0 mg/kg. This is
the absolute lethal dose (Table 1).
Table 1. 265
The effect of the muscle relaxant ditiline on the
organism of fur seals. - -
Ta6.nia 1
Ram-lime mbunequcro peaaucapTa perumpia na opranuam mopcium KOTHKOB
•
The mg/kg
oe 0.50 035
6 4 1
.1triK2
1,00 1,21
0 0
6 6
0 0
o • • 200
• tj
•
s O ()Up - • • e
,r-i too
CI) o
H so
Dos ,a
e me/kg 030`,
£1 75 1,0 1,25 1,5- 2,0
Pitc. 2. linteuenue nyabca y mopci:m Horuzon OT Be:111 ,11M 1 ,1 ;1, 031,1 :11IT 11,11111 a
,,,,, . . • • 0 20 4* /3 .70 0 12 30 0 20 10 30 0 20 43 30 0 40 10 30
apv4sr, A41.0r
Time, min.
Figure 2. Change in the pulse of fur seals depending on
the size of the dose of ditiline.

427
With the aid of the electrocardiograms it was shown
that in optimal doses the °reparation slows down the cardiac
activity, which later returns to the original level (Figure
2). The duration of the intervals in the cardiac cycle is
directly dependent on the dose (Figure 3). The initial part
of the ventricular complex (GPS) is subject to the least
changes. On emergence of the experimental animal from the
state of adynamia, the duration of the intervals in the cardiac
cycle returns to the initial state.
The pulmonary respiration in the narcotized animals
is changed to a greater degree, the greater the amount of the
muscle relaxant reaching the blood. With the administration
of mean effective doses there is an insignificant depression
of respiration, while optimal doses impair gaseous exchange
and may even evoke short-term arrest of respiration (figure 4).
The mean effective dose caused an increase in the body
temperature of the fur seals of 0.6° C; the optimal dose evoked
a temperature increase of 1.0 - 1.5 ° C, while the absolute le-
thal dose evoked a temperature increase of 3 - 4°0 (Figure
With the restoration of the capacity for movement, the body
temperature gradually attains the initial level.
Our investigations showed that the muscle relaxant
affects the fur seals for 15 - 30 minutes, after which the
'animals recover their locomotory function. With the aim of
elucidating the influence of the preparation on the fur seals
on the days following the injection, observations were carried
out on the animals for 1 - 4 days in a cage, after which they
were released onto the rookery. Before release, the fur seals
5 ).

* State of animal Normal
Incomplete immobilization .1••••• ••■•••
Dose mg/kg ,a0 3 a mr/icr
0,25 0,75 1,0 1.5 2,0
428
Dose m5/kg 1. 11* Ilo çja,mr
1.4. 075 fO 1,25 15 j 2.0 j
,14 eft/Mite 1,1 hen rr711110/1
0 01- * Odceioe Hue .xeugammoro ; 1 i
....
–.— tlenomine 0.lu.:ev&vaie i , (1) – C(,` T. ----- if OpMailbmae ■
c•-:---,-- - IlOntioe ocre3gikeyrtdomue i'à i i
.1)
• it i
e02/ 1 . J i f\iN . .1.A\ .. .,.....4 .• 1
E-IÇ:Q eif . • i I I I I i I
P pa.eun ernmem p Fegassi 11/2/5/ P Pcese rerrtus£ piee:greerneee p e7Gias1 fasitgRe 11 fri M epeoebt intervals
-Pm. a. FI31eiteitire npoRomKtrre.ibilocTit unTepnamon 11 cepitetinom unK.ie n 31313IICMIOCTII OT ne.-nnunim no3b1 munanna
11>
Figure .3. The change in the duration of the intervals in the cardiac cycle depending on the quantity of the dose of ditiline.
-1 11•■ •••••■•• # Complete immobilization
V4 •
-P • 70
1`
60 ‘.0 •
0 E 90
0 • te. PO r0 PA •
. 12
"
05 wa 20255101520255 ffl 1520e 5 i0 a 20 e 5 weee epemg 17170X0,YraeH5IR Orkena MUH '
Duration of test, min Pnc. 4. II3menenne •n,bixannn y mopcimx KOT111:00 110/1,
B:111511111CM
Figure I. The change in the respiration of fur seals under the influence of ditiline.
41Ï
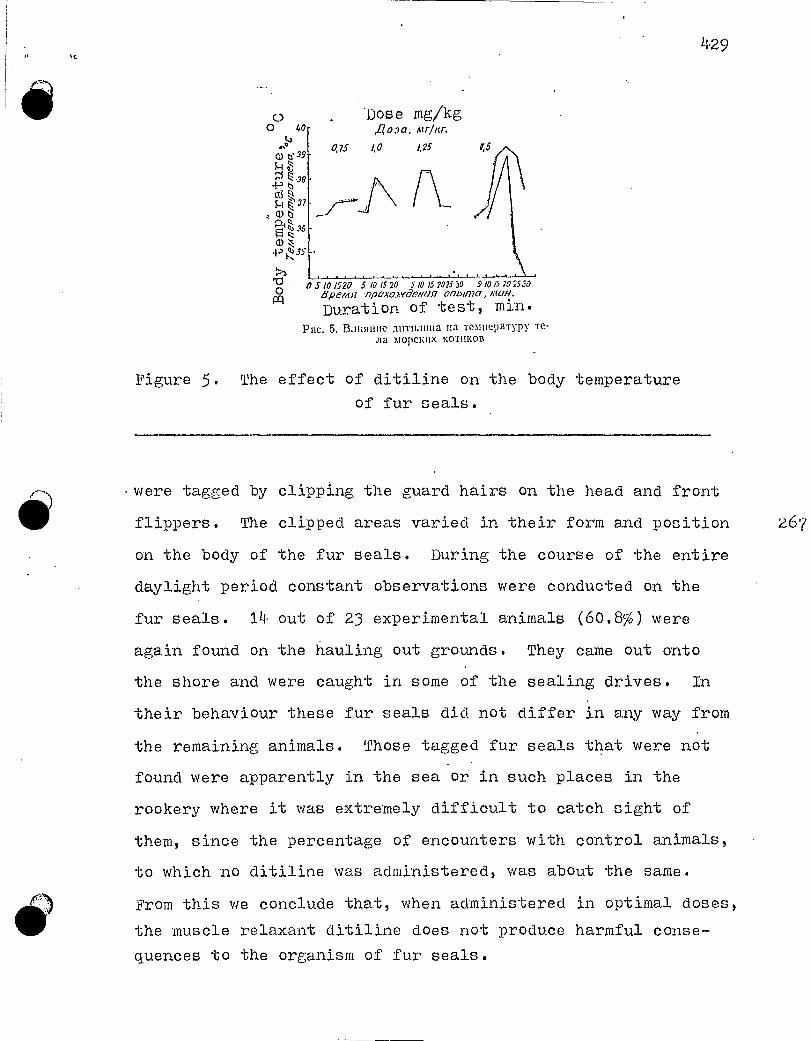
Iiose mg/kg joaa. ht r nr.
0.75 1,0 1.25
429
17 5 10 1520 5 10 15 20 51015 20250 5 10 15 20 2539 gpemn -np0x0.7.yeemag onbuncr,
Duration of test, min. Pile. 5. alliSInue airrnaima Ha Temlieparyp ■,. -re-
mopciaix KOTIIKOB
Figure 5. The effect of ditiline on the body temperature of fur seals.
were tagged by clipping the guard hairs on the head and front
flippers. The clipped areas varied in their form and position 267
on the body of the fur seals. During the course of the entire
daylight period constant observations were conducted on the
fur seals. 14 out of 23 experimental animals (60.8%) were
again found on the ilauling out grounds. They came out onto
the shore and were caught in some of the sealing drives. In
their behaviour these fur seals did not differ in any way from
the remaining animals. Those tagged fur seals that were not
found were apparently in the sea or in such places in the
rookery where it was extremely difficult to catch sight of
them, since the percentage of encounters with control animals,
to which no ditiline was administered, was about the same.
From this we conclude that, when administered in optimal doses,
the muscle relaxant ditiline does not produce harmful conse-
quences to the organism of fur seals.
o 0 0
Wti 39
-P D W% P.37
P-1°-
(1.) -p 3g
1:1 0 - IA

14.30
CONCLUSIONS
The mean effective dose of ditiline for fur seals aged
2 - 4 years lies between 0.5 and 0.75 mg/kg; the optimal dose
- between 1.0 and 1.25 mg/kg, while the absolute lethal dose
is 2.0 mg/kg.
The mean effective and optimal doses, at the start of
the action of the preparation, slow down the cardiac activity
in the fur seals, which later returns to the initial level.
The more ditiline is administered, the more marked is
the dePression of the pulmonary respiration in the narcotized
animals.
In the fur seals the ditiline gave rise to a short-term
increase in body temperature.
No harmful consequences of the effect of ditiline on
the fur seals were observed after the administration of mean
effective and optimal doses of this muscle relaxant to these
animals.

1a-31,d ,L
.rII-rrr:I1,aTS•IIA1%Kymexr:o B. II..IIpe.ay6oiinoe o6cs;LBn;xnuauuc :r.nBO•cuMtX cj)apatar.o;tortt1tc-
ct;uMnt BeutecTaaMut. 1MaTepea.n111 Aox.TIanoB III nay"tnoïr xotujtepenqnn BCTCpnltap-rto•cannT,1.pnoro q)atq•.1r,Te•ra 1\1 11IBLNIII, 196.3.2)I(y,•:exr o B. H., I^oprrecaa B. H. O6c:32Btur:nBarnte Bo.,ncoB n;,tewe;[cü 11,11T11-
nom. ^<I3ercptntap1151:^, \ë 11,'19û5.3 Hecae'poc I'. A. OntAT rtpuNtettenct» ;Urrit.iima na so•rm:oBO.m npoNnAc.-te. I43B.
TI•lI-IPO, r. 58, 1966.
REFER.ENGES
1. Zhulenko V. N. Pre-slaughtering immobilization of animals
with pharmacological substances. Materialy.dokladov
III nauchnoi konferentsii veterinarno-sanitarno fakul-
teta nSTIMMP (Reports of Ilird scientific conference
of the veterinary-sanitary faculty of the IGSoscow Techno-
logical Institute of the ^`Ieat and Dairy Industry), 1063-
2. Zhulenko V. N. and Korneeva V. I. Narcotization of wolves
and bears with ditiline. "Veterinariya" ("Veterinary
Xledi.cine" ), No. 11, 1965-
3. Nesterov G. A. A test of the application of ditiline in
fur sealing operations. Izvestiya TINRO (Proceedings
of the Paci;Cic Ocean Research Institute of Fisheries
and Oceanography), vol. 58, 1966.

4.32
UDC 599.745
V. N. Mineev
QUESTIONS OF CONSERVATION AND REGULATION OF THE STOCKS 269
OF MARINE MAMMALS
In recent times questions concerning the rational ex-
ploitation of the stocks of marine mammals have acquired a
particularly important significance, since these stocks in
different basins have either been undermined or are found in
a stressed state.
The Ministry of Fisheries of the USSR is Paying con-
siderable attention to the rational utilization of the stocks
of marine animals and considerable efforts are being undertaken,
especially in recent years, in this direction.
THE FAR-EASTERN BASIN
The hunting of the sea-otter has been banned everywhere,
which has had a verY favourable influence on the re-establish-
ment of its stocks. The numbers of this species at the present
time in the waters of the Soviet Union comprise 5.5 - 6 thou-
sand head. Ma recent years measures have been undertaken to
strengthen the conservation of the sea-otter; throughout the
course of the whole summer period vessels of the Sakhalin Fi-
sheries Service and the Kamchatka Fisheries Service patrol the
region off the Kuril Islands and Mednyy Island; in 1967 there
were organized two permanent posts for the conservation of sea-
otters on the Kuril Islands, on the islands of Paramushir and
Urup, while in 1968 a third post was set up on the island of
Iturup.

• • 433
o
o
For the further increase in the population of this
very valuable marine animal the ban on its hunting should be
maintained. It is necessary to intensify the work on the
acclimatization of the sea-otter on Bering Island and in other
regions, which used to be its habitats.
Recently commercial organizations are proposing the
introduction of a prophylactic culling of the sea-otters. In
the opinion of Far Eastern scientists, a commercial culling
operation of a so-called "selective" cull afloat is Premature.
At the present time it is sensible to restrict ourselves to
collecting the pelts of sea-otters which have died as a result
of the natural mortality and to carry out further tests on
culling.
Sealing operations on the fur seals are being conducted
on the Komandorskiye Islands and Robben island.
The measures for the conservation and regulation of
the sealing operations on the fur seals on the on-shore rooke-
ries favour the growth in their population numbers.
After the signing in 1957 of the Provisional Convention
on the conservation of fur seals in the northern part of the
Pacific Ocean, the numbers of these in the Far East increased 270
by almost 2.5 times and attained a level of 400 - 420 thousand
head (Komandorskiye Islands - 200; Robben Island - 180 - 200
thousand head).
The dynamics of the catches from 1961 appear as follows:
in 1961 there were killed 12 thousand head, in 1962 - 13.5 thou-
sand, in 1963 - 1•.7 thousand, in 1964 - 18.9 thousand, in
1965 - about 20 thousand, in 1966 - 18.9 thousand, in 1967 -

4•3y., ,.
17.7 thousand, in 1968 - 15.0 thousand and in 1969 - 15.6
thousand head. It should be noted that from 1966 the fur seal
kill began to be reduced in connection with the decrease in
the numbers of the commercially exploited bachelors. The
causes of this decrease still remain unclear, in spite of the
deliberations of a special conference and the organization in
1969 of a trip of a commission to the Kuril Islands ..
It may be supposed that one of the causes of the de-
crease in the numbers of bachelors in the rookeries was the
overestimate of the catch limits, which were established in
previous years, and also the prolongation of the sealing ope-'
rations after August 1st and the frequent driving-off of these
animals from the bachelor hauling grounds.
In recent years several measures have been undertaken
for the further strengthening of the conservation of the fur
sealsi a 30-mile prohibited zone has been established around
the Kuril Islands and Robben Island. It has been resolved to-
kill bachelor males that are no younger than 3-4• years and
of a size not less than 108 and no more than 150 cm. Drives
of fur seals are authorized only from bachelor hauling grounds
with not fewer than 100 head of the commercially exploitable
animals present on the hauling ground.
The population numpers of Steller's sea-lion have
attained a level of 11•0 - 60 thousand head, but these are vir-
tually not being commercially exploited. Thus, "lial'ryba"n°
has cought the following numbers of Steller's sea-lion in dif-
ferent years : in 1964. - 329, in 1965, - 14-6, in 1966 - 40, in
196? - Va., in 1968 -1I•20 head, while about 3000 head could be
caught each year.
i` ":llal'ryba" - Far Fast Trust of Fishing Industry. Enterprises.

4,35
in connection with the reduction in the stocks of the
true seals in the Far East, a limitation of their catches
was initiated from 1967:
In 1967 limits were established for the ringed seal
(11-5-0 thousand head) in the Sea of Okhotsk; the ribbon seal
(5.0 thousand head) in the Bering Sea;
in 1968 for the ringed seal (32.0 thousand head), the
ribbon seal (5.0 thousand head), the harbour seal (11.7 thou-
sand head), the bearded seal - ban in the Sea of Okhotsk; in
the Bering Sea: for the ringed seal (1.0 thousand head), the
ribbon seal (5.0 thousand head), the harbour seal (3.0 thou-
sand head), the bearded seal (1.0 thousand head);
in 1969 for the ringed seal (32.0 thousand head), the
ribbon seql (5.0 thousand head), the harbour seal (11.7 thou-
sand head), the bearded seal - ban in the Sea of Okhotsk; in
the Bering Sea: for the ringed seal (26.0 thousand head), the
ribbon seal (5.0 thousand head), the harbour seal (5.2 thousand
head) and the bearded seal (2.0 thousand head).
From 1970 the sealing has been subdivided into coastal
(sovkhoz*), state shipborne and for the needs of the local
native vovulation.
We consider it advisable.to. switch the hunting opera-
tions for the seals from the spring to the autumn, since during
the autumn sealing all of the seal pelts may be utilized as
high-quality fur raw material. It is necessary that scientific 271
organizations, within a short period of time, give their propo-
sals on the localities and periods for the sealing operations
and on the rational exploitation of the pinniped stocks.
* sovkhoz - state farm.

L36
In connection with the small numbers of the "antur"
of which there are 4.0 thousand head on the Komandorskiye
Islands and about 2.0 thousand head on the Kuril Islands,
there has been a ban on its hunting from 1970.
With the aim of conserving and re-establishing the
stock of the Pacific Ocean walrus, the numbers of which are
estimated at 53 thousand head, the catch limit will be main-
tained at 1.0 thousand head for the needs of the native popu-
lation of Chukotka with a complete ban on shipborne hunting.
In addition, there have been introduced several measures
to strengthen the conservation of the walrus on the hauling
grounds. Every year during the hauling out period of the wal-
ruses an inspector will be present on Wrangel Island.
Commercial whaling for the beluga is not developed in
the waters of the Far East. The numbers of the beluga are not
known. From 1970 the catch of older individuals has been autho-
rized, with the exception of belugas with suckling young.
Scientists should devote their attention to the study
of this animal and should recommend methods of organizing its
exploitation to the industry.
THE NORTHERN COMMERCIAL BASIN
The main regions of the commercial operations are the
White and Barents Seas. From 1965 shipborne sealing for the
harp seal in the White Sea was banned for five years, the
catch of whitecoats did not exceed 20 thousand head.
In 1969 the period of the ban expired but the re-estab-
lishment of the stocks is proceeding extremely slowly and
therefore the limit for 1970 was set at 23 thousand head with a comolete ban on killing the females.

437
o
o
In 1969 for the first time there were carried out ob-
servations on the Norwegian sealing operations in the Barents
Sea. No infringements were found on the part of the Norwe-
gians.
From 1970, in connection with the unsatisfactory state
of the stocks, bans were established agaist sealing for the
bearded seal in the White Sea and the grey seal in the White
and Barents Seas.
At the present time sealing operations for the ringed
seal are not restricted. The SevPINRU (Northern PINRO) and
PINK) (The M. Knipovich Polar Research Institute of Marine
Fisheries and Oceanography) should, in the very near future,
study the state of the stocks of marine mammals in the White,
Barents and Kara Seas, and provide conclusions on their ratio-
nal exploitation.
In recent years the Jan Mayen and Newfoundland stocks
of the harp seal have not been exploited by the Soviet sealing
industry.
CASPIAN BASIN 272
In recent years the hunting for the adult Caspian seal
has been banned, with the exception of the hunting for unmoulted
whitecoats and greycoats during the period from January 25th
to March 15th on the ice in the Northern Caspian within the
limits of the quota. For 1970 the catch limit was set at 60
thousand head.

4.38
THE BAIKAL BASIN
The numbers of the Baikal sea at the present time
comprise 33 thousand head, which permits a yearly catch of
2.5 thousand head. The catch of moulted individuals WO tO 1
year of age ("serka", "kumatkana"*) is authorized during the
period from April 25th to the termination of the sledge sealing
by means of rifles of 5.6 mm caliber with fortified cartridges
and by means of fixed capron** nets under the ice, in the re-
gion to the north of the line which joins the 01'khanskie Vo-
rota to Cape Tonkii, except for the three kilometer zone around
the Ushkan'iye Islands.
In recent years measures have been undertaken to inten-
sify the conservation of the Baikal seal, the staff of inspec- (-)
tors has been increased and additional transportation has been
alloted.
THE BALTIC AND LADOGA BASINS
In the Baltic Sea a limit of 2.5 thousand head has been
set for sealing operations.
In Lake Ladoga a catch of the Ladoga seal in the amount
of 500 head has been authorized.
In accordance with the recommendations of the 3rd All-
Union Conference on Marine Mammals, the "Glavrybvod" (The Main
Directorate for Fish-breeding and Protection of Fishes) devised
Translator's notes. * "serka","kumatkana" - names of older stages of the seal pups. For the harp seal, "serka" would be translated by "beater".
**"capron" - a synthetic fibre

439
the "Provisional regulations for the conservation and exploi-
tation of marine animals for Soviet ships, organizations and
citizens", which were confirmed by the Ministry of Fisheries
of the USSR and came into force from January 1st 1970. After
these have been in force for 1 - 2 years it willbecome clear
as to how good they are and then the question will be decided
on the promulgation of permanent regulations.

UDC 599.7 11.5.2 + 599.7/1,513
D. A. Mikeladze
KEEPING FUR SEALS AND CASPIAN SEAIS IN THE 273
BATUMI AQUARIUM
The study of the way of life, behaviour and many other
questions of the biology of marine mammals is made difficult
by the lack of opportunity for systematic prolonged observa-
tions on individual animals. The keeping of these animals in
an aquarium or oceanarium permits such observations to be made
and, in addition, provides an opportunity for studying such
questions as the dynamics of the growth, the change in weight
and other aspects during particular periods of the year and
in step with the increase in the age of the animal. However,
it should be borne in mind that life in captivity naturally
to some extent changes the instincts and habits of the animals.
In Japan and certain other countries there has already
been accumulated a fairly considerable experience on the keep-
ing in captivity of certain species of pinnipeds, including
also the fur seals. ln the Soviet Union such work has not
been carried out, with the exception of individual short-term
experiments on the purposeful keeping of fur seals in enclosures
and the more prolonged keeping of Caspian seals (Badamshin,
1959). The opportunity for organizing and conducting such
studies in our country has emerged with the bringing into ope-
ration of the marine aquarium in Batumi.
In Uctober of 1966 there were brought by airplane to
the Batumi aquarium three Caspian seals, which had been caught .

441.,,
on Zhemchuzhno Island in the Caspian Sea, but which died in
1`dovember of 1967. Immediately (in this same month) there
were again brought to the aquarium five Caspian seals, that
had been caught on the same island (Table 1). The animals
were transpor.ted by airplane, where they had been placed in
wooden crates without water.
Fur seals were delivered to the Batumi aquarium on
three occasions (Table 2). From Robben Island, located in the
southern part of the Sea of Okhotsk, in August of 1967 there
were brought, in the same way as the Caspian seals, by airplane
in wooden crates six fur seals, including one pup that had
been born this same year. The next batch, amounting to four
fur seals, was brought in September of 1968 also from Robben
Island. Finally, in August of 1969 from Bering Island (the
Isomandorskiye Islands in the Bering Sea) there was brought one
adult female, that was presumably pregnant.
One of the fur seals was an unweaned pup and the attempts
to artificially feed it were not successful. The simultaneous
death of the three Caspian seals and four fur seals in 1967
occurred from food poisoning. U:,0 to December 1969 three Cas- 275
pian seals had survived in the aquarium for 2 years, one fur
seal for more than two years, 3 fur seals for one year and
one female was delivered in August of 1969.
In the northern part of the Pacific Ocean the fur seals
accomplish regular seasonal migrations. Spending the winter
months in the open sea, in the spring, as the waters become
warmer, they move to the north and spend the summer months on
the on-shore rookeries, located on Robben Island and the Kuril,
i;ornandorskiye and Pribilof Islands. The water temperature in

w.n^air.u.r--......r-...r-...-..l.^.ua..iiw.a-L+._`^..1- a..^ .v._......
Table 1.Information on the delivery of -Caspian seals
TaG:i[Iüa ^
CBejkenNH 0 3aCO3C KacnBVlCKnx Tlo!leueïl
►IonSex
Bospacr
AgeCaalert male 2 ro;>,a
yearsCamKa f e Iria.l - 2 ronLa
CaTIKa f I ro;t
Caueültl
Caaleli m
CaNIKa f
Caa1Ka fCaatKa f
Table 2.
2 ro;ta
2 roTLa
I ro;i
1 roi
1 ro:2
Date of!la a,
a.nPuHO3a
devery
OKTS16pb 1966 r.UctoberOKTaGpb 1966 r.
C<KTRGpb 1966 r.
24 IIOAGpfl 1967 r.
24 IIoslGps[ 1967 r.
24 11os16pst 1967 r.
flovember24 11o1Gp5i 1967 r.
24 non6pi 1967 r.
I1puutcuauusiRema.rks
17orn6 8 noslGpa Died,1967 r.
TIorllGna 8 1Io^16psI1967 r. I
IlorltG:[a 8 ]loaGpx1967 r.
I'vov..
of of
it
ilorltG 7 1<Iasl 1969 r.,
I7orI16,na 30 slflBaps[1968 r.
Information on the delivery of fur seals T 11 it a 2
CBeJl,ennH 0 3IIB03e hSOpCK}Ig KOTIIKOB
I'vIe1g-ht Date of Time i^11on 1i03p3C2 Bec, sz Aaru pp1uzo3a 13(: hm nnryrn 1ïPn^^e iai na
Sex lL^e kg dellvery trans ^ y Ke^nar.ks.
Caatetx Il7
Camett M
Carleu 111
Ca;`nca f
Camnta f
Camuca f
Caali:a f
CamKa f
CanlKa fCameu-ill
CaAtKa,f
2 roaa
2 roaa
year:2 roZa
2 ro;la1 ro^L
III
Pup.3 ro;ia
2 ro;ua
I ro:
.1 roa
7 :IeT
year
25
25
23.
21
19
12
26
14
19.5
18,5
42
23 anrYcra ' 1967 r.
23 anrycTa 1967 r.
' august23 anr}•cTa 1967 r.
23 asrycTa 1967 r.
23 aurycra 1967 r.
23 aurycra 1967 r.
9 celn•nGpa 1968 r.
9 cexTSlGpa 1968 r.september
9 cenTStGpst 1968 r.
9 CCIITflGpR 1968 1'.
4 aBr\'CTa 1969 1'.
august
52 ^taca
52 =Iaca
hour52 yaca
52 yaca
52 llaca
52 'Jaca
131uac.
131 llàc.
131 llac.
131 nac.
53 tlaca
17or116 8 1I0aG- Died, iVOT:pH 1967 r.
To >Ice
To Ace
To >Ice
to
W_ayJan.
ifet
n
Ilorit6aa 31 il Sep"CCI1TflG1)H
1967 r.
-çr1lIIorIIGm 21 an " il-
pe,Ia 1969 r.
274

1143
• • -•■ • •
PIIC. 1. 1(0T111:11 13 aKBapityme
Figure 1. Fur seals in the aquarium.
() the regions of the winter habitats of the fur seals keeDs
within the limits of 1-2 to 9-10 oC, occasionally dropping to
0° C. The air temperature in these regions rarely exceeds
12-13°C. With the warming of the water above 14-15 °0 the fur
seals leave the wintering regions and move to the north (Panin
and Panina, 1968; •Arsen'ev, 1964).
The regions of the on-shore rookeries are characterized
by having cloudy weather with drizzling rains and an air tem-
perature ranging from 10 to 15 °0. On occasional sunny days
this may increase to 2000 and more, and then the fur seals
tend to spend the greater part of the day in the sea, where
the temperature of the water generally does not exceed 10-12 00.
In the Batumi aquarium the fur seals encountered com-
pletely different conditions (Figure 1). The water temperature
in the aquarium varied from 6.5 to 25.4°C, while the air tem-
perature varied from 2.5 to 30 0C. The differences in the

CC Ile :flom 1:0>iaa-nopcime wrpo-
BM
6a -rpm KO:Wall:10 1' -
CKIIC OCT pOLIZI
BwryNm
Komait- ',-topewlic ,errrpo-
im Gal-pm
21,2
MA 21,3
94,9
21,5
24,0
6,8
8,2
10,8
11,5
7,8
9,3
6,1
7,7
10,3
11,2
7,2
9,4
Table 3. Comparative data on the water and air temperatures, o C.
, T a 6 au u, a 3
CpanuuTenbume Rauume o TemnepaType nogm H noagyxa, °C
.A. • TeNusepa'Typa B • Temm,parypa ito:./tyxa
d ico.rie6anim von umecm
Year and month
Komatutop- CKIle
f ourpoca
(I KoacCiamist CCpcansm
BaTVMIL
mo June iliolib
July Mon' AUM
Sept . CeltTuCipb
190
June HIM,
July Miom,
18.0-23,5
23,2-2-1,6
22,7-25,6
22,9-25,1
18,0-23,4
23,2-24,6
5,4-8,2
7,6-9,2
9,4-12.6
9,9-13,5
5,8-8,2
6,7-9,8
19 .8 22.4
99,6
21,4
20,9
21,4
14,6-25,6
15,8-28,6
17 ,5-28,7
W1-29,0
17,0-30,6
17,6,-29,3
4,0-7,5
8,5-11,8
9,4-12,6
r)
A. - Water temperature B. - Air temperature
c - Average d - Ranges
e - Batumi f Komandorskiye Islands
4‘)
temperature conditions in the aquarium and in the region of
the Komandorskiye Islands are shown in Table 3.
As can be seen from the table, the hydrometeorological
conditions of the habitats of the fur seals in the region of
the Komandorskiye Islands and in the Batumi aquarium differ
considerably. So abrupt and rapid a change in the conditions 276
of the habitat may lead to certain changes in the organism of
the animais, but the investigation of these questions is a
task for further studie .s.
All that has been said also applies, to a somewhat
lesser degree, to the Caspian seals.

445
With their placement in the aquarium, there also
occurred a considerable change in the food objects of the
animals. In the northern part of the Pacifie Ocean within the
stomachs of the fur seals there have been found the remains
of 45 species of fishes and 8 species of squids. The main
food objects in different regions are the Korean cod, the sand
lance, greenlings, lanternfishes, herring and certain squids
(Panina, 196 )). Under natural conditions the Caspian seals
feed on gobies, "kilka", herring, silversides and gammarids
(Badamshin, 1948, 1959). In the aquarium both the fur seals
and the Caspian seals were fed on the Black Sea scad, and more
rarely on the picarel and the Black Sea anchovy. Predominantly
defrosted fish was provided, more rarely live fish (Figure 2).
Under natural conditions in both the fur seals and the
Caspian seals the weight of the food consumed per day ranged
from 5 to 15; of the weight of the animal itself, though in
this case the regularity of the feeding could not be observed
(Boitsov, 1934; Dorofeev, 1964), since at times the animals
cannot obtain food for themselves over the course of relatively
long periods, while at other times the food is plentiful.
In the aquarium the animals were Provided a regular
feeding twice a day, and in this case, as our investigations
showed, the amount of food consumed ranged within the limits
of 10 - 15% of the body weight. In i:ndividual cases over the
course of two to three days the amount of food consumed was
as much as 20-26% of the weight of the animal, but then the
demand for food dropped markedly. It is possible that with
the higher caloric value food, obtained by the animals in the
aquarium, the feeding rate decreased somewhat.

ena mom Date of delivery
Mm
.S'ex
23 anryna 1967 r. 9 ceuTsiGim 1968 r. ;Se Di 9 ce11r96p1 1968 r. 9 ceuTsulpn 1968 r. 4 anrye -ra. 1968 r. Aug.
31,5 31,5
. 21.5 23,5
- 42,0
Pile. 2. lopm.leuue Kuriu:ors
Figure 2. Feeding the fur seals.
Table 4.
Change in weight of the animals.
T a 6 u a 4
Ilamenenue Lulea ,',,, nOTHMX
4.46
B.Decnaunannyme,ta
13e,nc„,1,1,P," uc,t Aug. 12 ownwpsi j 3 .11IapTa I 12 anrycrn
e2 196S r. 1069 r. 1969 r. A.
AlopcKue Korutai Fur seals Ca meuM 25.0 36,0 28.0 1 Ca mat. 26.0 - 25,0 25,0 Cameum 18.5 18,0 17,0 CamRaf 19,5 19,0 18,5 Ca mizaf 42,0 ' — —
Kacludicraie 710.7efill Caspian seals " 24 11096p9 1967 r. :NOV • Caetti 20,0 I 23.0 22,0 29,0
24 no516ps 1967 r. C9MNaIj 16,0 19,0 19.5 . 25,5 . 24 non6pH 1967 r. Cana1 15,0 !« 1,0 18,5 24,5
3FrIplimegaaue. B nepeol c nena6pn 196S no main. 1939 ro.u. a Asuno-rnbte n ann'apnyme-ne GLUM oi.:c.,cnegenbi z.ocTa -romnum E.0.111 ,1eCTI1ONt KOpM a.
* Note. During the period frOm December 1968 to March 1969 the animals in the aquarium were not provided with an adequate amount of food.
A. - Weight at delivery, kg B. - Weight in aquarium, kg
m - male f - female

F I. I.
44?
o In Table 4 is shown the change in weight of the fur
seals and Caspian seals during the period of their stay in
the aquarium.
A task for further studies is the detailed investiga- 278
tion of the feeding of the animals under the holding conditions
in the aquarium and the influence of this factor on the growth
and development of the organism.
In the Batumi aquarium constant observations are con-
ducted on the behaviour of the animals which are being kept,
and some Preliminary results of these observations may be re-
ported.
The fur seals easily and rapidly become accustomed to
people, especially to those who constantly attend them. They
quickly begin to recognize those who feed them and quietly sub-
mit themselves to these people. Fur seal No. 1, an aborigine
of the Batumi aquarium (the only survivor of the first batch,
who is now already 4 years old) and the adult female do not
• resist when their mouth is opened to examine the state of the
teeth.
The Caspian seals have also become accustomed to people,
they calmly take food from the hand but do not allow themselves
to be touched.
Observations show that the marked change in the living
conditions, and apparently primarily of the temperature regime,
has exerted a considerable influence on the periods of moulting.
Thus, in fur seal No. 1 in 1969 the moult commenced on May 28th,
approximately one month or even earlier than is usual under
the natural conditions. The so-called third stage of the moult.

448
o was noted at the beginning of July, while on the islands this
stage is generally observed in the first half of August (les-
terov, 1968; Bychkov, 1964). In the Caspian seals also the
moult commenced at an earlier period than in the Caspian Sea.
Of great interest is the elucidation of the question
as to the influence of the life under the aquarium conditions
on the quality of the fur pelt of the animals, especially of
the fur seals.
The work with the pinniPeds, which are being kept in
the Batumi aquarium, is only beginning. Included among future
tasks are comprehensive studies on the effect of the stay in
artificial conditions on the vital activity, growth and deve-
lopment of the organism.
011TEPATYPA
1 Apcenbee B. A. 0 cmeninnantitt nom:amuït mopcirttx KOTI1E011. Mopcirrte 10111- lonlamblicro Boctrora. TI1HPO —BHIIPO. M., 1964. 2 Baan.utittut B. Il. 0 nirrounit racintiicKoro Toaenn. Tpybt Boaro-racrinficuoft
cTanuilli, T. 10, ACTPaXallb. 1948. 3 Baaottinin B. II. Onbur co.:tepwatinst racinnicroro Tionenn n neuone. C6opinur
cieeri no 61(o.loritit. Tpy.arr KnenlIFIPO. fliuneupomii3nar, M., 1959. 44. Batioa JI. B. KOTIIIMBOC X0331 fiCTBO. M., 1934.
Bb1lltiO8 B. il. MaTeptianbi no antimite moperux ROTIIIZOB na ocTpoce TIOciellbeM.
opciie KO Bocrora. THI-IPO — BHIIPO, M., 1964.
6 ,z/opooepa C. B. Ceriepirbie mopcirne. ro-rurit (CaIlorllinus ursinus). kt.operile ro-rinat ii,a,unero Bocrora. THHP0— B1-11•IPO. M., 1964 .
7 Ilecrepoo I'. A. MaT.epilaabi o cporax .11111bE.II EOTILKÔB Ha KomatuopcKux ocTpouax. ilacTotiorne ceucpnoii mac-rit Tiixoro oKeatia. M., 1968. •
8 IlaRtma r. K. Flirt- aune No-rin:ori u StnolicRom mope. MopcKire Eo•riikti ,aa.tibue-Boctrinia. T'IWO-B(1'1P°, M., 196-1.
9 Hamm K. II.. [lamina F. K. ',.-/Konorlui ii murpannir OTI1KOB il alimile-necett-unii tiepno,k 1.1 griouch:em ,flacTottot ne ceueptioii tiacTit Tri xoro
1 IIHPO—B1-11•11)0. M., 1968.

449
REFERENCES
1. Arsen'ev V. A. The mixing of populations of fur seals.
Fur seals of the Far East. TINRU - VNIRU (Pacific
Ccean Institute of Fisheries and Gceanography - All-
Union Research Institute of Marine Fisheries and Oceano-
graphy). Moscow, 1964.
2. Badamshin V. I. The feeding of the Caspian seal. Trudy
volgO-kaspiiskoi nauchnoi rybokhozyaistvennoi stantsii
(Transactions of the Volga-Caspian Fisheries Research
Station), vol. 10, Astrakhan'. 1948.
3. Badamshin V. I. An attempt at keeping the Caspian seal
in captivity. Sbornik statei po biologii. Trudy
KaspNIRO (Collection of papers on biology. Transactions
of the Caspian Research institute of Fisheries and
Oceanography). Pishchepromizdat (State Pulishing House
for Literature on the Food Industry), Moscow, 1959.
4, Boitsov L. V. Fur seal_management. Moscow, 1934.
5. Bychkov V. A. 'Information on the moulting of fur seals
on Robben Island. Fur seals of the Far East. TINRU -
VNIRO, Moscow, 1964.
6. Dorofeev S. V. Northern fur seals (Callorhinus ursinus).
Fur seals of the Far East. TINRO VNIRU. Moscow, 1964.
7. Nesterov G. A. Information on the periods of the moulting
of the fur seals on the Komandorskiye Islands. The
pinnipeds of the northern part of the Pacific Ocean.
Moscow, 1968.

8. Panina G. K. The feeding of the fur seals in the Sea of
Japan. Fur seals of the Far-East. TINRU - VNIRG,
Moscow, 1964.
9. Panin K. I. and Panina G. K. The ecology and migrations
of fur seals during the winter and spring period in
the Sea of Japan. The pinnipeds of the northern part
of the Pacific Ocean. TIivRO - VNIRO. Moscow, 1968.

45 1
Une 599.745.3
V. D. Pastukhov
THE FEEDING OF THE BAIKAL SEAL 280
UNDER EXPERIMENTAL CONDITIONS
The study of the feeding of the seal under natural
conditions presents certain difficulties. In connection with
the fact that the animals feed mainly during the crepuscular
periods, while, as a rule, they are caught during the day,
the stomachs of the animals are found to be empty. However
in the large (rectal) portion of the intestine there is always
contained a greater or lesser amount of otoliths, which are
very convenient to use for determining the species composition
of the fishes. Moreover, in the sorting of the otolith samples,
removed from the intestine of the seals, a striking feature
was that the otoliths that belong to one and the same fish
species differ markedly from one another in their dimensions.
This suggested to us the idea that the fishes consumed varied
in age, size and weight. From this, naturally, it seemed
possible to establish the data on these fishes on the basis
of their otoliths. However for this there had to be answered
other, no less complex questions: could the otoliths be diges-
ted, how long were they retained and what was the degree of
their state of preservation during their passage through the
gastro-intestinal tract of the seal. All of these points, as
well as a whole series of other questions, could be resolved
only in an experiment.

•-• 452
With these aims I during the period from September -
November 1967 to April - June 1968 there were held in large
tanks (1.2 m3 ; 2.3 m3 ; 7.4 m3 ) three Baikal seals: Ushkan
(aged 1.5 years), Vega (of the same age) and an adult female
Manyunya (more than 10 years old). In the 7.4 m3 tank there
was constant running water, while in the two other tanks the
water was completely changed once a day. The water tempera-
ture in the tanks was close to the mean annual temperature of
the surface waters of Lake Baikal, about 5 0 0.
The animals were fed with fishes, which commonly occur
in their food: the Baikal oil-fish, and pelagic and benthic .
gobies. However, while the Baikal oil-fish predominates in
the natural rations, in the conditions of the experiment the
seals were fed primarily on pelagic gobies. Mainly fresh,
recently caught fish were provided.
For . determining the periods of retention of the undi-
gested remains in the digestive tract, marked food was manu-
factured. Coloured . taas of three different types were employed:
plastic platelets (25 x 6 x 1 mm); finely cut up pieces of
this same Plastic (2 x 2 x 1 mm), which are close in their
dimensions to the otoliths of fishes, and pieces (30 x 10 mn)
of thin rubber (Table 1).
The plastic platelets were retained for a particularly 281
extended period of time. The rubber tags emerged three times
as rapidly as the former. The small pieces of plastic appeared
in the tanks, on average, 1.5 times more rapidly than the rub-
ber tags. Evidently, as a result of the working of the stomach
and the Peristalsis of the intestine, small undigested particles
are expelled more rapidly from the digestive system of animals.

1 • - ■
o 47 8
3. Lilly D.* Man and dolphin. Moscow, "Mysl" ("Thought"),
1965.
L. Rei K.** The flight of three whales. "Amerika" ("America"),
No. 87, 1963.
5. Sokolov V. E. et al., An attempt at studying the charac-
teristics of the swimmïng'of dolphins in a water channel.
Tezisy dokladov 4-go Vsesoyuznogo soveshchaniya -co
izucheniyu morskikh mlekopitayushchikh (Reports of
Proceedings of the kth All-Union Conference on the
Study of 1,arine Mammals). Kaliningrad, 1969.
6. Tomilin A. G. A story of a blind sperm whale. Moscow,
"Iauka" ("Science"), 1965.
Translator's notes. * sic. Should be e. C. Lilly.
** This is a direct transliteration of the Russian uhonetic transliteration. •Probably I... Ray.

4'79L
LDC 599-537
D. A. Yorozov
TRANSPORTATION OF LIVE DOLPHINS 296
The transportation of live dolphins is one of the im-
portant questions in the overall.problem of the maintenance
of these animals in captivity. Dependant on the quality of
the transportation are the health of the animals and the dura-
tion of their life in captivity., and consequently also the
opportunity for conducting variôus studies and experiments.
Within a single experimental station the animals are moved from
the summer sea pens to the winter ;:)ools with warmed water.
Durin`s the course of the cleaning of the pools and in the ?pre-
para.tion of various experiments it is necessary to move the
dolphins from one from one building to another. A mastering
of the technique of transportation of dolphins permits one to
transport experimental animals from one specialized station
to another, to employ trained dolphins in spectacle shows and
to solve problems of an applied character (searches for under-
water objects and securing diving work).
In connection with the expansion of experimental stu-
dies it seems useful to provide a generalization of our expe-
rience concer.ninx?, the transportation of dolphins, which has
been accumulated in conjunction with F. A. Leontovich, V. A.
Kalganov, V. V. T^.',,anukhov, V. V. Belyaev and others.
Cot.ltmencin.!^, this work in 1965 and having available very
limited information on achievements abroad, we were forced to
make and test devices that we had constructed ourselves and

to try out various methods of transportation. 'îdotwithstanding
the diversity of the means of transportation which have been
utilized for this purpose (motor boats, launches, medium Black
Sea seiners, dry-freight transporters, special and passanger
shi.los, several types of motor vehicles, helicopters etc.),
at the present time we may single out only three basic methods:
towing in the sea, placing in a limited volume of water and
transportation without water.
Particularly unfavourable to the dolphins are the con-
ditions of transportation "without water". Under natural. çon-
ditions these animals are found in.a state that is close to
equi_librium and analogous to weightlessness. The force of
buoyancy, evenly applied to the whole body of the dolphin,
.neut,l"ali7.es the force of the weight, and at the same time only
the hydrostatic pressure acts on the internal organs. During
deep dives under the influence of the hydrostatic pressure,
as a consequence of the great motility of the thorax, the vo-
lume of the lungs in the dolphin may decrease tenfold for a
short period (V. V. Babenko and D. A. Prorozov, 1968). During
transportation without water the lungs and other internal or-
gans are compressed for an extended period under the action
of the weight of the animal. Dolphins are unaccustomed to the
drastic limitation of space and mobility, and to the isolation
of animals from one another.
During transportation the following undesirable effects
and traumas may arise.,
1. Compression of the internal organs, which makes
breathing and the work of the heart more difficult.
2Q?

k81
2. Injury to develouinq embryos and premature births.
3. Dislocation or soraining of the joints of the
pectoral flippers.
• . Fechanical injuries by the transporting apparatus:
cuts, bruises, scratches, abrasions and tears to the edges of
the flippers, leading to considerable losses of blood.
5. Temporary loss of motility of the locomotory mus-
culature.
6. Drying of the eyes and Portions of the skin, heat
and sun burns.
7. Overheating of the organism and heat stroke.
8. Entry of water into the blowhole during inhalation.
9. The impossibility of raising the dolphin to the
surface of the water for the normal exchange of air in the
lungs, as a result of entanglement in nets or the dolphin being
under other animals.
10. Stress with a lethal result.
The main inconvenience in the transportation of the
Azov doluhins (common porpoises) is caused by their habit of
being constantly in motion. The active movement of the tail
makes every stage of the transportation of these animals more
difficult. They are inconvenient to carry by hand or in a
stretcher, they frequently push one another down under the water,
they acquire traumas and, conseauently, suffer more from loss
of blood. The specific. difficulties in the transportation of
the bottlenose dolphins arise as a result of the great weiPht
of the adult animals, which in some cases may be as high as
300 kg.

4.82
We will pass on to a description of the procedures
and contrivances which assist in averting the enumerated
troubles. Entanglement of the dolphin in the net may occur
during the catch1ng, when the ourse seine is hauled in.
This is most likely to occur in those places where the net
gathers into folds or is pushed-in by the current into an
overhanging wall. Much more rarely the dolphins will ram
perpendicularly hanging nets. Having gotten caught in the
mesh by its beak and attempting to raise itself to the surface
of the water by the shortest path, the dolphin takes up a
vertical position in the water and in doing this it tangles
its jaws still more deeply in the net. Sometimes at a depth
of only one or two meters from the surface of the sea there
arises a threat of suffocation and drowning of the dolphin.
Therefore, several of those who are participating in the trans-
portation get dressed beforehand in their diving suits, fins
and masks, and, in case of necessity, jump into the water.
Tt is generally possible to free the animal from the net with-
out an aqualung on the diver, but at the saine time watching
the divers from the ship are one or two aqualung divers, who
are ready to quickly come to assist*.
* In working with nets the prototype model "Ukrainy-3" proved itself in an excellent manner; this is a new aqualung by the designer A. I. Gnamm. Special arches (bulges) prevent nets from hooking onto this aqualung. The AVM-1-1',1 aqualung often hooks onto a net with its charging nozzle, minimal Pressuraindi-cator, manometer and shut-off valve. I..ornover, the minimal pressure indicator and other metallic comoonents on the left shoulder strao may injure the delicate skin of the dolphins. For working- in nets one may remove from the AV1.-1-1,-. aqualung the charging nozzle and all of its fittings. A charging Point end-cap cov'érs the ooening of the high pressure nozzle near the shut- off valve. A supplementary arch Prevents the nets from hooking onto the valve.

14.83
NC. 1. Renhcbulibi B «noxrounemiom» Henoese
Figure 1. Dolphins in "submerged" net.
Une of the procedures for protecting the bottlenose
dolphin from injury by the net is the placement of a diver
between the dolphin and the net. The head of the animal is
placed on the shoulder of the man, the net is kept away from
the tail by using one's legs, while the hands protect the
Pectoral flippers from getting tangled in the mesh of the net.
The water suits "Sadko" and "Super-Calypso" without the weighted
belts have a Positive buoyancy and this permits one to lie on
the surface of the water without effort. The animals are
calmed by careful stroking. We did not have a single case of
a dolphin biting while it was being caught, although fairly
frequently it was necessary to grasp them by the jaws with
one:s hands aad even to remove the net from the teeth. The
ideal procedure for protecting the doluhins in a partially

484
hauled in purse seine would be to position divers beside each
dolphin, although under the conditions of mass capture this
is generally not possible (Figure 1).
The ship which transports the dolPhins does not ap-
proach right up to the nurse seine net, and the animals are
towed to it in special cages. The frame of the cage is welded
of duralumin tubing o in such a way that water cannot get into
the internal cavities of the tubes. To increase its buoyancy
and stability, there may be attached to the cage additional
foam-plastic floats. Over the frame of the cage is stretched
fine-mesh capron material, if possible - made of thick threads.
The dolphin is moved out of the purse seine through
the open end side of the net (Figure 2). After one or two
dolphins are placed Ln the cage, it is towed by a row-boat or
small motor boat to the transporting vessel. The Position of
the animals in the cage during the towing is controlled by
swimmers (Figure 3), Particularly restless dolphins are
placed in the cage with a man, whose job it is to support the
head of the animal during the towing, to avoid entanglement
in the net.
The raising of the animal from the water onto the
deck of the transporting ship is.the most difficult and
dangerous, to the animals, operation. Norris (1966) warned
against attempts to raise the dolphins from the water by the
tail, since in this case the blowhole is under the water, the
breathing is impeded and the animals often drown. Pulling out
the animals by hand into the small boat generally leads to
traumatization of the animals against the hard fixtures on the
side of the boat, the ribs and other parts of the boat. The
* Translator's note. sic. However the actual figure 3 is not found in the oa2er.

!".
14.85
Pm. 2. lipzcupoma meril C iteithepincamit
Figure 2. Towing the cage with dolphins.
best method up to the present time remains the method of .
raising the dolphins from the water on a stretcher, usine a
derrick. On special ships the cage with the animal may be
pulled up a slip or through an open ramp.
In Figure /1.*is illustrated the most successful design
for a transporting cage, in which the end walls fold'down on
hinges for the entry of the dolohin into the cage. Other
designs, which do not have the folding-down walls, have to be 300
sunk down on one side in order to, as it were, scoop up the
animal, but in the confinement of the partly hauled in net
this is very inconvenient. The latter design permits one to
take up the dolphin without bringing the cage into the net 301
but placing only the folded-down wall over the top line. The
second foldine wall Permits one to brine the dolphin out of
the cage head first. The convexly arched side wall struts
and the upper connecting tubes have a form which decreases
the -Drobability of the animal beinz injured by them. The skids
*Translator s note. Figure 3.
sic. Description, however, refers to

^^ --^--^--^`C.^ ^-^ ^ ',- ^.,^•^ ^. ^.-^.
^^`^^^^^
Puc. 3. TpaucnopTUan K:ICTb IIOaO}f }iOIICTpYIitlltN
4
figure 3. A tr. ansporting cage of a new design.
^` {1--w ^..+^^•.r ^ ^ ^
Pue. 4. TpattcnopTUpoa:.a Ae1b^titta ma r."t ► ccttpYto-uicPi pame
Figure II.. mrans?porting a dolphin on a hydroplaning frame.
^protect the ;e s roi:l be i-n^^ de.r^a:ed when it i s '?u11ed. u^^ on
shore or u-o a;,li,,) onto the cleck and raise uA), above the Ç1ip-
wa^i or -round, the bottom of the cage on which the dolphin is
1.yinÛ. It is best to make the bottom of rubberized ca^^ron.

4-87
it should have small openings for the drainage of water, so
that when the cage is lifted the excess water is not lifted
with it. The most convenient variant has a removable bottom.
In this case the bottom has a separate framework with 4 - 6
ri 11^ bolts for the attachmient of the slings of the derrick.
The si?e of the botton of the cage, 3.5 x 0.8 m, permits
loading of three-meter Black 'S)ea bottlenose dolphins. The
corresi)ondence of the dimensions of the removable bottom of
the cage with the dimensions of the tubs used for transporta-
tion i)ermi-bs one to place the dolphin into the tub on this
same bottom, avoiding superfluous transfers. A reserve supply
of interchangeable botto,ms considerably exuedites the work of
loading and unloading. A cage of this design may be employed
for the temporary isolation of dolphins in a pen or pool.
For the towing of dol^hins there can be proposed a
hydroplaning frame design (Figure j'`) which was elaborated by
us. The outside dimensions of the frame (3.5 x 0.8 m) permit
it to be placed into a transporting tub. While the removable
bottom of the cage should have a negative buoyancy, the front
and side tubes of this frame should hold it on the surface,
while supporting, a dol-phin. After the securement of the ani-
mal with. straps to the canvas panel the posterior edge of the
frame sinks down. This is achieved by the fact that the
posterior tube fills up with water through its openings and
does not 'serve as a float. Additional .trim at the stern may
be obtained by installing foam-plastic floats in the front
-part of the frame. Gn account of the trim there I s created
the initial angle of attack (see Figure 21,
Translator's notes. * and *,-. sic. * - Figure 1I•. ** - second
f igure in Figure 4.

488
of the towinir, the frame emerges to hydroplane along the sur-
face. This considerably decreases the towing force required.
A distinguishing feature of the towing on the hydroplaning
frame is the securement of the dolphin with straps and the
hydrodynamic lifting of the blowhole above the water. in
securing the dolphin partiCular attention should be Paid to
tiFrhtly fastening down the caudal flukes to the canvas panel,
this prevents the possibility of the animal rolling over onto
its side. The trimming of the frame at the stern helps to
lift the blowhole of the dolphin out of the water even when
the frame is not moving, which is of particular value when it
is installed in the transporting tub. For the greater comfort
of the animal openings are cut In the canvas of the frame for
the Pectoral flippers.
The frame may be utilized as a cradle for ensuring the 302
immobility of a dolphin during the course of a medical exami-
nation of the animal or for experimental purposes.
The described ways of towing may be used in bringing
the dolPhins from the purse seine to the transporting ship,
from the transporting ship to the pen (through a sPecial pas-
sage in the wall of the pen), in moving the animals from one
section of a holding Pen to another or in moving the dolphins
along channels, connecting several pools.
The second method of transporting dolphins, conveyance
in a limited amount of water, was introduced into Practice in
the USSR by F. A. Leontovich*. Aqualung divers were first used
* This information was published by F. A. Leontovich in the re-port "A test of underwater filmirur of dolPhins" at the 3rd ple-nary session of the Section of Underwater Studies of the uceano-graphic Commission of the USSR Academy of Sciences in January of 1966.

489
for removing dolphins from a purse seine on the 9th of July
1965. They held the dolphin and brought up under it in the
water a special stretcher, on which the animals were raised
from the water, transfered and suspended in containers with
water. Used as the containers were the canvas deck tubs on
a metallic frame, of the type that is found aboard the Black
Sea refrigerator ships and seiners. F. A. Leontovich noted
that dolphins suffer considerably from all reloadings and
transfers, and recommended that a large number of stretchers
be employed, so that each animal is only once placed onto and
once removed from the stretcher during the entire journey.
This principle of "straight from the sea onto the suspension
device and to the place of holding without transfers" was set
at the basis of the further imProvement of the Procedures for
the transportation of dolphins.
F. A. Leontovich's observations showed that the dol-
phins which were placed in an individual canvas tub, 2.0 x 0.5
x 0.6 m in size, began to struugle and manifested nervous be-
haviour, while in a transparent aquarium of perspex of the same
size they behaved much more calmly. This was taken into con-
sideration in the design of the sPecial tubs for the transpor-
tation of the dolphins. Before these tubs were made there
were used canvas brine tubs, stretched out on ropes in the hold
of the ship or in the back of a GAZ-69 A truck, and canvas
tubs on wooden end-panel frames. There were also used ply-
wood boxes, lined with porolon*, within which was inserted an
inner lining of polyvinyl chloride film. Lesides these, there
have also been utilized rectangular steel boxes, welded of thin
sheet metal, inflatable rubber boats etc.
* Translator's note. "porolon" - a foam plastic.

4.90
John Lilly (1962) prepared boxes according to the di-
mensions of the animals and attemrted to decrease the volume
of water in these to a minimum. The tubs were sha-.)ed out of
thin plexiszlass in the form of the body of a dolphin, while
the strength and rigidity of the design was ensured by plywood
ribs. In order to lessen -Che spillage of water en route s the 303
box was covered with a lid, with a large slot for the dorsal
fin. In this case there was utilized the principle of simul-
taneous loading of the tub together with the animal and water,
though such a tub was suitable only for a dolphin of a Parti-
cular size.
Cur tubs were designed for the transportation of
various suecies of dolphins. They had a rectangular form,
which permitted, depending on the size of the animals, the
transportation in one tub of one or two bottlenose dolphins,
or 1.11) to five white sided dolphins, or up to nine Azov common
porpoises simultaneously. The most suitable type proved to
be welded tubs of duralumin, which were much lighter than
steel tubs, more durable than plywood tubs, and did not pro-
duce such scratches and abrasions to the delicate skin of the
dolphins as did the canvas tubs. The internal dimensions of
the transporting tubs should be no less than 3.5 x 0.8 x 0.8 m.
In the anterior part of the tub are mounted three rectangular
windows of plexiglass.
In the preparation of the tubs Particular attention
should be Paid to the method of securing the windows. In the
first design the six-sided heads of the bolts which projected
inside the tub and the edges of the tub at the window were the
cause, in several cases, of traumas to the dolphins that were

4•91
being transported. The second design eliminated this defect
by the placement of the bolts on the outside and by the selec-
tion of the bevelling on the plexiglass. ^:;e note that from
the outside the plexiglass should be clamyped down with nuts
through a steel strip which borders the window along its peri--
meter. This ensures a more even distribution of the load on
the fastenings. The hydraulic impacts which arose during
rolling and pitching caused cracks in the windows and loss of
water, if ordinary washers were placed under the nuts instead
of such a strip.
Edging the upper edges of the tub with 7cm diameter
tubing considerably decreases the probability of traumas during
the loading of the dolphin into the tub. The bottom of the
tub was provided with skids as a convenience for moving it,
while on the lower part of the sides of the tub there were
fixed handrails, which could be used for carrÿing the empty
tub by hand or for raising it mechanically. The presence of
drainage plugs considerably facilitates keeping the tub clean
and allows for the possibility of changing the water level in
it. It should be borne in mind that painting the tub with red
lead and the subsequent trans°cortation of an animal in standing
water in such a tub may lead to the poisoning and death of the
animal.
Aboard ship the presence of running water in the tubs
is provided for by making use of sea water from the fire mains.
on motor vehicles, loss of water from the tubs occurs on star-
ting, durin^; turns and. on brakin:; and also when moving over
an uneven road. Strips of porolon, placed on the surface of

1-1.9 2
the water, decrease the splashing. En route, fresh water may
be added: the movement of dolphins into rivers, sometimes
for hundreds of kilometers upstream from the mouth of the ri-
ver, indicates that brief sojourns in fresh water do not harm
them. The susPension of the dolphin in the tub on a stretcher 304
prevents injury of the animal against the walls. In this man-
ner, in 1968 and 1969, there were transported by motor vehicle
four bottlenose dolphins and four white-sided dolphins over a
distance of 250 km. Without a stretcher it is fairly difficult
to restrain an animal on an uneven road § though enthusiasts
were able to deliver dolphins without injury, at the cost of
a five-hour stay in the water together with the animal. The
main thing in this situation is the timely raising, of the del-
Phin by the jaw, so that its blowhole is above the water.
Thanks to the experiments of A. G. Tomilin (1951) it
became known that the loss of heat from the organism of the
dolPhin to the surrounding medium occurs through the flippers*.
If the flippers are not moistened with water for a prolonged
period, then, because of the smaller heat capacity of air,
there may occur overheating of the organism and. death of the
dolphin from heat shock. Drying of the eyes may cause blind-
ness. Some authors recommend wrapping the dolphins in wet
woolen blankets or in sheets. This method has its disadvan-
tages: The fabric does not adhere closely to all parts of the
skin and it hides dried-out portions from the observer. Such
coverings, however, protect the skin from burns by the direct
rays of the sun.
* Translator's note. The Russian word "plavnikin refers not only to the flippers but also to the dorsal fin and tail flukes.

11.93
Spending a prolonged time without movement during trans-
portation may cause a temoorary numbness of the locomotory mus-
culature in dol_ohins. Therefore, when it is let out into the
pen or pool the dolphin sometimes may not be able to raise it-
self to the surface to exchanP - e the air in its lungs. At such
times it is desirable to have an aqualung diver in attendance.
Further studies of the physiology and characteristics
of behaviour of dolphins under experimental conditions will
permit improvements to be made in the methods of transporting
these animals.
JIHTF.PATYPA
1 EaGengo B. B., It lowrion 1.1euoToplite (1)U31ItiecK1Ie 3aKonomepnoe-rn npn - Reaml)nnolu. C6. eMexann3Mht nepetunt:KeHlin H op uieuîa 1H III 1 UROT111,1X».
dlayKorsa Aymt;a», knell, 1968. 2 TO.U11.111H A. F. IleantitiKu oprauta TepNtopery.rnunni laiT006p8311b1X. «PbI6110e
X03grICTDO», 12 u 1950. Tomimivi. A. r. O Tepmoperymmun y Kirr006pa3twx. «Flpnpoaa», N2 6. 1951. I.illi J. C. Man and dolphin. no 4, 1061. 5 whales dolphins and porpoises. Ed. by Norris K. S. Berkeley and Los;
Angeles. Syrnpos. 1965.
REFERENCES
1. Babenko V. V. and ïorozov D. A. Some Physical regulari-
ties in the diving of dolphins. Symp.: "Mechanisms
of locomotion and orientation of animals". Publ.
"Kaukova dumka", Kiev, 1968.
2. Tomilin A. G. The flippers, organs of thermoregulation of
cetaceans. "Rybnoe khozyaistvo" ("Fish L.anagement"),
12, 1950.
3. Tomilin A. G. Thermoreq.ulation in cetaceans. "Priroda"
("nature"), no. 6, 1951.

494
UDC 599.74.5.3
K. K. Chapskii
THE SYSTEMATIC RANK AND SUPRAGENERIC DIFFERENTIATION 305
OF THE 8-INCISOR SEALS
Already in a Paper published ten years ago (Chapskii,
1961) the author argued his views on the subfamily structure
of the ohocids, the main criterion for the systematic subdivi-
sion of which was taken to be the incisor formula. Un this
basis was confirmed the classical suprageneric structure of
the seals in the form of the three subfamilies: the Phocinae,
I,onachinae and Cystophorinae, which had been accepted three
years previously by Scheffer (1958) in his review of the seals
of the world. It may be recalled that it was namely this sys-
tematics of the phocids which were adhered to by the Predomi-
nant majority of mammalogists, commencing with those who laid
the foundations of the systematics of the pinnipeds: Gray,
Gill and others. Many other prominent zoologists of our time
have gone even further and have foilnd it possible to divide
the phocids into four subfamilies (Kellog, 1922; Hay, 1930;
Simpson, 1945; 2rechkop, 1955; Ellerman and Morrison-Scott,
1957, and others).
However the arguments presented for such a division
were, in large Part, very limited and inaccurate.
This "fra=entation" of the family began in the last
quarter of the last century and, aPparently, pertains to Gray
(1874), who, after including the 8-incisor seals in a single
subfamily or tribe for many years, decidedto divide these,

1405
separatinz from the monk seals the southern 8-incisor seals
and including these in the tribe (essentially, from the taxo-
nomic aspect, equated to a subfamily) Jtenorhynchina, equiva-
lent to the Lobodoninae. In the final analysis, even though
this did not improve the systematics as a whole (since in the
process of the rearrangemelit of the genera the monk seal was
placed together with the 10-incisor seals), it focused the
attention of mammalogists on the taxonomie heterogeneity of
the 8-incisor seals and, in particular, on the specific cha-
racteristics of their Antarctic representatives.
THE INCONSISTENCY IN THE TAXONOMIC INTERPRETATION OF THE
8-INCISOR SEALS
The differing views on the systematic position of the
seals that have in common a single incisor formula are, un-•
doubtedly, conditioned by the insufficient clarification of
the elements of similarity and difference (primarily of the
craniological features) between the subtropical and Antarctic
components of this group of phocids.
r'ven in contemoorary reviews it is difficult to find
indications of any distinct osteological differences between
the seals of the genus r._onachus, on the one hand, and the
whole aggregate of the 8-incisor seals from the Southern hemi-
sphere, on the other. This situation may be illustrated by
several sPecific examples.
Almost fifty years after the systematics of the phocids
were enriched, following Gray 's initiative, by the new name
of a distinct subfamily of the southern 8-incisor seals,

o
4.96
Kellog (1922) was forced to state that it was difficult to
find any Precise differences between the Antarctic seals
(Lobodoninae) and the monk seal. Scheffer (ibid.) also aureed
with this, oddityg that, in the Presence of the already existing
subfamily of 8-incisor seals of the Sothern hemisphere, the
seY)aration of a distinct subfamily for the seals of the genus
Uonachus seems to be unsubstantiated.
Not without interest in this connection is the state-
ment by King (1966) that the intergeneric skull differences
in the seals were in general so great that it was difficult
to find any features that were common to the grouP. It is
impossible not to ouree with this statement. Unfortunately,
however, from this King arrived .at extremely different conclu-
sions in another direction.
THE NEWEST ATTEMPT AT A RADICAL RECONSTRUCTION OF THE
SUBFAMILY STRUCTURE OF THE PHOCIDS
In King's opinion, which was expressed in the previous-
ly mentioned paper, the mammalogists who created and perfected
the systematics of the Dhocids had, as it were, "gone along
the line of least resistance" in making use of the number of
incisors as a criterion of the subfamily for precisely the
reason that this is a very convenient.feature, although, from
her point of view, this is by no means sufficiently reliable.
In her own studies, directed towards elucidating the genetic
relationships between the various groups of phocids and, con-
sequently, to the creation of a more rational systematics of

1407
the latter, she completely abandoned the incisor criterion,
considering it as being too variable.
She preferred to place completely new features at the
basis of her reconstituted systematic scheme:
1. The size of the posterior lacerate foramen (fora-
men lacerum).
2. The partially visible, from above, mastoid (os
mastoideum).
3. The ventral part of the petrous bone (os petrosum)
which projects from the lacerate foramen.
It is already known what followed from this. The re-
constructed systematic scheme was found to be reduced to two
subfamilies, which were quite sufficient, as it seemed to King,
to include all of the seals. In one of these, the subfamily
of the northern phocids (Phocinae), she included the 10-incisor
seals and the hooded seal, in the other (Monachinae) - all of
the seals of the Southern hemisphere, including also the ele-
phant seal. King admits that in these subdivisions it is dif-
ficult to clearly Place: firstly, the bearded seal (genus
Erignathus), in which the posterior lacerate foramen is abbre-
viated, and secondly, the monk seal itself (genus Monachus),
since in this two of the basic features (the visibility of the
edge of the mastoid in dorsal view and the distinctly projec-
ting petrous bone in the lacerate foramen) do not correspond
to the requirements of affinity to the subfamily Monachinae
in its new interpretation. Acknowledging these deviations,
King found a way out of a difficult situation by considering
that both of these genera of seals were transitional forms,

k98
ikï
linking the subfamilies of the northern and the southern seals.
Strictly speaking, there are no grounds for disputing this
thesis in general, even if only for the reason that the Antarc-
tic 8-incisor seals undoubtedly belong to one and the same
genetic line with the monk seal, from the ancestral form of
which they also originated. Likewise, there is also no doubt
that the contemporary monk seal (from the ancestor of which
there commenced the divergence which led to the appearance of
the whole group of genera of 8-incisor seals of the Southern
hemisphere) is itself a derivative of the more primitive 10-
incisor seals. •
Probably there is nobody who doubts also the fact of
the relatively more primary and primitive character of the nu-
merous morphological features that are characteristic of the
seals of the genus Erignathus, which aspect has already been
discussed by Winge (1924, 1941), Naumov and Smirnov (1936),
and Chapskii (1955). Nevertheless, it by no means follows
from this that the seals of the Sothern hemisphere could have
originated directly from the predecessor of the contemporary
bearded seal.
The term "transitional form" is a concept from the
phylogenetic arsenal, and by itself is essentially not con-
nected with systematics. Whatever this form is, the compo-
nents which it embraces are taxonomie categories, and these
should have a fully defined position, for which arguments
have been presented in the proper manner. However, in the
system which has been constructed by King on the new morpholo-
gical criteria the position of both the monk seal and of the

11-99
bearded seal is fairly precarious. This is a postulated posi-
tion, rather than one which corresponds to the formal require-
ments of systematics. In actual fact, the motives for inclu-
ding the first of the named species in the subfamily T+'ionachi-
nae (against which, of course, there can be no objections)
clearly will not pass criticism from the formal aspect, since
out of the three criteria of the subfamily, -proposed by King,
two do not fit. At the same time the only and, obviously, in
these circumstances, the decisive feature, the specific cha-
racter of the lacerate foramen, cannot play a similar role
in the second case, in establishing the systematic status of
the bearded seal.
',+hat else can explain these facts, if not the weak 308
aspects of the systematic scheme itself and the unreliability
of the arguments on which it is based? do references to the
transitional character of the species can rectify the situation
here.
The transitional position of any particular species,
genus, subfamily etc. may manifest itself only in those fea-
tures of the structure.which do not belon` to the diagnosis.
To rephrase this postulate, it may be formulated in another
ways the features which are distinguished by an inconstancy
of their manifestation cannot serve as diagnostic features.
Cbservance of these requirements is obligatory for all
of the links of the systematic scheme, particularly for cate-
gories of so hi^^h a rank as the subfamily. Deviations may be
encountered, but only in the form of extremely rare exceptions
to the rule in individual cases. In the system proposed by

500
King, however, the infringement of those few structural fea-
tures to which an exclusively diagnostic significance is as-
cribed seems to be the rule rather than the exceotion! What
then, under these conditions, is the advantage of the new sys-
tematic scheme and why should it be given preference?
SOIE CRITICAL REMARKS CONCERNING THE DIVISION OF THE PHOCIDS
INTO TWO SUBFAMILIES
All that has been said above (the inadequate diagnos-
tic value of the new features which were erected by King to
the position of the primary craniological criteria of the sub-
families, and, as a consequence of this, the presence of the
mentinned "transitional forms") already by itself very much
weakens King's position, which so decisively broke with the
classical systematics of this family of pinnipeds.
There are, however, also other serious grounds for not
agreeing with the new taxonomic structure.
In the first place, there calls attention to itself
the undervaluation of the dental criteria (the incisor formula),
the negative attitude towards which was not argued out in a
proper manner.
The main objections, however, arise from the results
of a special craniological study, undertaken by the author
with the aim of re-examining the taxonomic problem that has
arisen. Having checked.the mentioned new criteria of the sub-
familial integration and, at the same time, having reviewed
the craniological material on the phocids that is available
in the osteological de .00sitory of the Zoological Institute of .

,501
the USSR Academy of Sciences with the aim of revealing more
completely, as far as was possible, the similarities and dif-
ferences between the various groups which were being compared,
the author reached a firm conviction as to the stability of
the previous classical systematic scheme for the phocids
(Chapskii, 1969, in litt.). Without repeating here all of his
arguments, the author nevertheless considers that it would be
useful to present in a very brief form the main motives, on
the basis of which he disputes such radical changes, introduced
by King into the systematics of the phocids.
The objections may be reduced, basically, to the fdl-
lowing points.
1. The significance of the craniological elements of
similarity, which are in fact present in the hooded seal and
the 10-incisor seals, were exaggerated in King's study; at
the same time the cranial differences between these seals
were revealed by her very incompletely and were underrated.
îeanwhile, in fact the number of similar features in these
seals is only about half of the number of the features of
difference: there are about fifteen of the former and at
least twenty five of the latter. In this situation, the cate-
gorical judgement of King on the.inadequate diagnostic value
of the incisor criterion does not appear to be very convincing.
If however one takes into consideration certain additional dia-
,gnostic features, namely: a) the narrowed spindle-shaped bulge
of the mastoid part of the temporal complex (precisely that
portion which is visible beyond the lateral mastoid process
from dorsal view and which in cross section does not exceed

502
half of the length of the tympanicum); h) the direction of
the axis of this bulge, which is inclined markedly downwards
behind the mastoid process; v) the marked swelling of the
maxillary bones immediately in front of the orbits; g) the
well developed anterior palatine foramina, situated in groove-
like depressions, - then, together with the incisor formula,
the subfamily of the ten-incisor seals acquires a fairly good
craniological basis.
2. Speaking convincingly against the systematic bring-
ing together of the elephant seal and the .8-incisor (in-parti-
cular, the Antarctic) seals is the marked predominance of fea-
tures of difference between these over the features of simila-
rity. Strictly speaking, these seals are linked only by the
abbreviated lacerate foramen, while on the basis of almost all
of the remaining elements of the craniology they are differen-
tiated. If, however, there are scrupulously counted all of
the features by which the compared seals are in any way simi-
lar and these are set beside those features which differentiate
them, then, with such an approach, it is found that there are
almost three times as many features of difference as there are
features of similarity. In these conditions it is difficult
to speak of closer genetic ties between the seals which are
being compared.
3. en the other hand, the similarity between the
elephant seal and the hooded seal is far from being exhausted
by those few and, in Ring's opinion, unimportant features which
she mentions. LA actuality, the similarity between them is
much greater. With an attentive examination of the skulls,

é 503
it turns out that there are twice as many elements which bring
these two genera of seals together as there are elements by
which they are differentiated, although there are fairly many
(about 20) of the latter, since these are different genera.
Thus, if the dental formula is accepted as the criterion for
the systematic ,grouping of the genera Cystophora and Mirounga,
i.e. in this case the presence of six incisors (i = 2/1), then
in the seals which are united by this characteristic there are 310
found to be about 40 similar features, and only about 20 dis-
similar features. If however, on the contrary, one is guided
not by the incisor formula but by the relatively little exten-
ded lacerate foramen, and, consequently, one unites the ele-
phant seal with the southern 8-incisor seals, then the total
number of similar elements of the cranial structure in these
will be no greater than five, while at least a dozen features
of difference can be compiled. •
With such numerical ratios, one would think that it
would be difficult to insist on uniting the elephant seal in
one subfamily with the 8-incisor seals. Similarly, it is un-
natural to combine in the second subfamily the hooded seal
with the 10-incisor seals and, as a result, to liquidate the
third subfamily of the 6-incisor seals, the Cystophorinae.
To sum up, the truth in the taxonomic structure of the
phocids is found to be on the side of the classical systematics,
and as the leading criterion of this, at any rate for the con-
temporary members of the family, we may consider the dental
formula*.
* (This footnote is given on the next page of the'translatiOn.)

504
CRANIOLOGICAL SPECIFIC CHARACTERISTICS OF THE 8-INCISOR SEALS
Accepting the incisor formula as being sufficiently
weighty, from the taxonomic aspect, and giving preference to
this classical criterion in_subdividing the phocids into sub-
families, it is impossible not to see also how difficult it
is to find any other craniological features which would in the
same way unite all of the 8-incisor seals and would be speci-
fic only to these. This most important, if not the only, dia-
gnostic element could be expanded by taking into account cer-
tain additional features, namelys
1. The low positioning of the anterio-inferior edge
of the cheekbones (Figure 1).
2. The markedly more developed, than in other seals,
and further backwardly directed zygomatic process of the maxil-
lary bones (Figure 2).
3. The narrowed anterior angle, which is markedly
smaller than a right angle, of the lumen of the orbit in the
horizontal plane (Figure 2).
However all of these additional craniological charac-
teristics are not very expressive, are subject to considerable
variability, are not free of exceptions and may sometimes be
found also in other seals.
Still less diagnostic are several other structural fea-
tures, which, though characteristic of all of the members of
* (From previous page). The author is forced to make the stipulation with respect to
the contemporary phocids at the present time on the grounds that in the upper Tertiary ancestors of the monk seals (genus 1:.ono-therium) there were allegedly present not 2 but ) incisors in each half of the upper jaw (Kellogg, 1922; Boettger, 1951).

505
4
Pttc. 1. KonTypl.[ netsoït ct<ynoneïtI:oCTtt (n^loi^lttnt.) Ttone1[aAFOttI x^I
(8eflxBltït j111C.) il TlD:lettfl y9a^[ ^^I87CthlliCKÿ Rlo'soü(».)oc'fil, Q-
Otio9tte'IC1111S1: a rc noi!^'fe31[If911oC nrRepcrtfe. ee nepei;He-Ila;:fnuil tipafFq
CYCAIIC^RCj1Xl111Ïf uTj1UC70t: i^OC'l'ft,. ^ ^ .-
Figure 1. The contuurs P f the le ft zyg o:matiç boi:e (profile)
of the monk seal (upper figure) and the Weddel
seal (lower figure).
Ilegend: . a - suborbitai foramen, b- body of ^ygo=-matic bone., v-- an^erio-superior proçess of bone,g -'its anterio--inferior edge,
this subfamily without exception, are at the same time inhe-
rent also to the representatives of the subfarriily of the six-
incisor seals. JLmong -these bel-4g; the wide but not niarkedl,y 311
protruding bulge of the mas^oid, whieh is not direeted down-
wards behind the mas.wid proçess; the eonçave outïines of the
projection of the lateral contours of the yre-orb.i.tal part onto
a horizontal plane; the disappearing anterior -Palatine fora-
mina; developed pre-orbital proçesses; a wide inter-ocular
space; abbreviated lacerate foramina.

506
Puc. 2. CxeataTt['tecxoe nsoGpaxtenne ^tacreït vepena• cuepxy:
n-- itwtTypt,t aepcna naprn (npcAcTau11TcAH ABCHTnpC3qOUWX TtoIc-ncit); 6--- t(rparatcuT npmroü cTOporu.t ^tcp, na -1 ro:tcuH-n+onaxa: a- raaa=Hu4nan 4acTb npaEOit croponLt vepena Ttoncnn Y9AACnAa; a- To Ace,
Ttosctts[ Poccaq pcpl,nlncT6tMn JtitlntHa+tt oÜoaitaHctta ;tJnnta (no npoAoJtbftoti oeil 4C-
pena) chy.nonoro oTpocTr;a acascunns:pnoCt hocrn
Figure 2. Schematic depiction of parts of the skull from above s
a - the contours of the skull of the harbour seal(a representative of the 10-incisor seals);
b - fragment of right side of skull of the monk seal;v- orbital part of right side of skull of the
Wed.dell seal;g - the same, of the Ross seal
The interrupted lines indicate the length (alongthe longitudinal axis of the'skull) of the zygoma-tic process of the maxillary bone.
•

5 0 7
o CRANIOLOGICAL GROUNDS FOR THE TAXONOMIC DIFFERENTIATION
OF THE SUBFAMILY OF THE 8-INCISOR SEALS
The inadeouacy of the available characteristics of the
tribes, of their diagnostic features, makes it urgently neces-
sary to have a proper reviSion and more precise definition of
the craniological basis of the established taxonomic subdivi-
sions of the 8-incisor phocids, the P,Ionachinae.
Gray (1874) was probably the first to present certain 312
definite and fairly accurately grasped specific characteristics
of the bones of the skull (in particular, the fusion and elon-
gation of the nasals), which permitted one to more or lesS re-
liably identify the 8-incisor seals of the Southern hemisphere.
Subsequently to these was added one other feature, the elonga-
ted zygomatic processes of the maxillary bones.
AS a result of the most recent examination of these
diagnostic features and all measurements of the craniological
difference of the 8-incisor seals of the Southern hemisphere 313
(Lobodonini) from the subtropical representatives of the
Monachinae, there were found, on the one hand, certain weak
aspects of Gray's diagnosis, while on the other hand there were
revealed several new and more precise elements of difference*.
It was found, in particular, that even such a charac-
teristic feature of the southern 1Lonachinae as the fusion of
the nasal bones is not completely reliable, since divided nasal
bones are not infrequent in Weddell seals (and not only in the
* This work was carried out mainly on the same craniological material in the osteological section of the zoological insti-tute of the USSR Academy of sciences.

508 F
young); it is encountered in the crabeater seal, and a ten-
dency to this is also noted in the leopard seal.
In all of the subtropical monk seals the nasal bones,
of course, are always divided. The second feature, the elon-
gated frontal part ("auex") of the nasal bones, also sometimes
does not give the typical picture in the southern seals, and
at times manifests the opposite development also in the monk
seal. Reference has also been made to the variability in the
length of the zygomatic processes.
Thus, in spite of the characteristic nature of the
craniological features, which one cannot avoid taking into
account, they nevertheless cannot have a primary diagnostic
• sienificance.
Taking into consideration all that has been said, the
cranioloeical part of the expanded diagnoses of both tribes
of seals of the subfamily Monachinae may be represented in
the following form.
TRIBE MONACHINI
Zygomatic bone and profile with a fairly clearly out-
lined anterio-inferior edge, permitting the length of its body
to be measured (without the anteior articular process). The
inferior edee, from the same position, along its whole length
has the form of an arch, which is elevated to the greatest de-
gree in the middle part of the body of the bone (Figure 1).
The end of the anterio-superior process of the zygo-
matic bone extends far anteriorly and terminates almost over
the interior edge of the suborbital foramen. The latter is

509
Pile. 3. Cxehta c -rpeemist mae -rit 'wane erepopm mepena ertepe,mt. rft Ea3HHO cocyroomeulle NICH ■ :(y riestemcennem ripe,D,raa3nwiliere o -reepcTim (ypepeob 'compere e -rmetielt upephusirerell :tumid') H
iiilem eKyaorteii 1:0CTIL (opereem ee pep:mere Hpayi) 060311aulitur. m— TIO.ICI1b-M011ZIX, Il — MO)CI ■Orl neonapjk
Figure 3. Schematic diagram of Part of the right side of the
skull from the front. There is shown the relation-
ship between the position of the pre-orbital fora-
men (the level of which is marked by the double
interrupted line) and the position of the zygoma-
tic bone (the trough of its upper edge).
Legend: M:- monk seal, N - leopard seal.
situated no higher than the level of the greatest deflection
downwards of the upper edge of the body of the zygomatic bone
(Figures 1 and 3).
The nasal bones are not grOwn together with one ano-
ther; the frontal portion of these is generally not longer
than the remaining (anterior) portion (Figures 2b and 4).
The fontanelles in the reeion of the presphenoid are
huge and round. -
The tribe includes the three sPecies of monk seals of
the genus 1\.onachus 21em.

A A
510
Puc. 4. (1)0Inta CJIMAX 11000BIAX 1:0CTal TIOJICIICfi 10)uuoro nouywainut: Tumeun Pocca (A) u mopcuo-
ro Aconapua (5) orpeilliamu ii upephuulcumu Juouumu noicalana
noGnore (*nepuntimi») noconbrx Nocreii. 06038atieinun Jr Kont, 2I —Bepxite!te.moeritari,
n — H000111,10 nocrit _
Figure 4 . The form of the fused nasal bones of the seals of
the Southern hemisphere: the Ross seal (A) and the leopard seal (3).
The arrows and interrupted lines indicate the length of the frontal portion ("apex") of the nasal b-ones.
Legend: fr - frontal bone, m - maxillary bone, n - nasal bones.
TRIBE LOBODONINI*
Profile of the zygomatic bone without any angular bor-
der between the maxillary process•and the body of the bone;
* The 8-incisor seals of the Southern hemisphere were first grouped in a separate tribe, as far as may be judged now on the basis of the available literature sources, by Gray (1871, 1874), thouP:h the author made use of a preoccupied name, Steno-rhynchina, which was subsequently re .placed by another. Since the Presently acceoted name for this tribe is derived from the crenus Lobodon (not Lobodont), then from this it seems that the more correct derivative is namely Lobodini, by which name the tribe is here designated.

511
its inferior edge is either completely without -an arch-sha1ped
curvature, or with a barely expressed, shallow arch only in
the posterior half of the bone. The anterio-suuerior end of
the zygomatic bone reaches far short of the suborbital foramen,
terminating at the level of the latter or below this. The
zygomatic bones in general are set considerably lower and the
greatest bend downwards of their upper edge is situated below
the suborbital foramen (Figures 1 and 3).
The nasal bones, as a rule, are grown together, while
their frontal >?orti.o-n is considerably longer than the remaining
part (Figure 11• ) .
The paired fontanelles in the region of the presphenoid
are in the form of narrow slits, and if they are wider-then
nevertheless they have a distinct tapering in front and behind.
iIMTI•:FATYPA
1 Hatt.lroa C., C.ilupHoa H. MaTepuant>I no cnc•reaa7111;e It reorpacilm[ecr,omy pac-npe,jeneHluo Phocidae cenepxoïi vacTn Tnxoro olceaaa. TpyAtii Bcec01o311oro uay^tn.-ncc:l. III]-Ta aiopcsot•O p1,l611. X03-l;a It o[tea1Iotpa4-un. T. 111, 1936.2 Yancxuit K. K. Onrr nepecaloTpa cucTeiltit it ,Zo-[artlocTIncu Tio.neueit 110;1CedIC31-
crua Phocinac. TpyAbl 3oo:rornvecsoro lui-Ta AI-1 CCCP, T. XVII, 1955.3 yancl:nO K. K. Coupeateatuoe cocTO>:uue If 11pOGae\ibl CIICTC\taTnlU( AaCT0110171IX.
T. corieruanuïi i-lxruo.nornvlccuoü soafliccnli Ali CCCP, nhiu. 12, 1961.
A- yaucr:uù K. K. Bonpo:.bt Taxcouotauvecxoi't crpvlerypbt cenleilcrua n cneTC hpa-niloltorntICCKIiX Ila11u61X. LICTBepTOe BCCC0103IIOe C013enta113ie qo n3Vtie[inlo NtopCliliX
DtTieüOllHTalolllHX. Te311Ch1 j(Oli.7a;1,013. N1., '1969.
5 Boettger S. Notizer zur Verbreitung und iiber Vcrwaridtscha-ftbezichungenctcr nlonchsrobbe (Alonachus albivcntcr I3ocidaert). Zool. Anz. -Leipzig, Bd. 147,Heft 11-12. 1951.6 Gray J. E. Notes on seals (Phocidaè) and the changes in the form of the
lower jaw durinh growth. Ann. k1a5. Nat. Hist. ser. 4.1869.
7 Gray J. E. Supplement to the catalogue of seals and whales in the BritishMuseum. London , Brit. Pylus. 1871.8 Gray J. E. I-land-list of seal'morses, sea-lions, and sea-bears in the British
h•luseuni. London. 1874.9 Eliervuan. and Morrison-Scott. Cheslist of Palaearctic and Indian mammals.
London. 1951.10 Frechkop. Ordre, des P;nnipecies. Traite de Zoologie vol. 17. 1955.
11 Kellogg R. Pinnipeds from Miocene and Pleist, deposits of California. Univ.Calif. Pablic. Gcol. SCi, vol. 13, no .1, 1922.
12 King J. E. The monk seàls (venus Alonachus). Bull. of the British Museum(Nat. 1-list) Zoology. vol. 3, no 5 L. 1956.

51 2
13 King J. E. Relationships of the Hooded and Ekphant se.als (genera Cye-
• tophora and Mirounga) J. of Zoology. Proe. Zoolog. Society of London, vol
.11 18. part 4: 385-398. 1956. •
Sciwf jer V. Seals, sea lions and wali uses. Stanford California. 1958.
1.5 Simpson G. 'Hie principles of classification and a classification of Main- •
Ines. Bull, of the Amer. Mus. of Nat. Ilist. vol . 85. New York. 1915.
10 Weber M. Die Sàugetiere. I3d. 2, Unterordnung: Carnivore Pinnipedia, Jena.
Winge H. Pettedyrsleo.gter. Kobenhavn. 1924 lb Winge il. The interrelationships of the mammalian genera. vol. 11. Koben-
havn. 1941.
REFEREN CES
1. Naumov S. and Smirnov N. Information on the systematics
. and geographical distribution of the Phocidae of the
northern part of the Pacific Ocean. Trudy VNIRO
(Transactions of the All-Union Research Institute of
Marine Fisheries and Oceanography), Vol. III, 1936.
2. Chapskii K. K. An attempt at a revision of the systema-
tics and diagnostics the seals of the subfamily
Phocinae. Trudy Zoologicheskogo instituta AN SSSR
(Transactions of the Zoological Institute of the USSR
Academy of Sciences), 'Vol. XVII, 1955.
3. Chapskii K. K. The contemporary state of and problems in
the systematics of the pinnipeds. Trudy soveshchanii
Ikhtiologicheskoi komissii AN SSSR (Transactions of
conferences of the Ichthyological Commission of the
USSR Academy of Sciences), issue 12, 1961.
F. Chapskii K. K. Questions of the'taxonomic structure of
the family in the light of craniological data. kth
All-Union Conference on the Study of Marine Mammals.
Reports of Proceedings, Moscow, 1969.

513
5. Boettger S. Notes on the propagation and relationships
of the monk seal (Monachus albiventer Boddaert)
10. Èrechkop. The order of the pinnipeds.
16. Weber M. The mammals. * Vol. 2, Ilborder: Carnivora
Pinnipedia. Jena, -1928.
17. Winge H. Mammalian genera. Copenhagen, 1924.
o

•
\ • 514
A. A. Antonyuk
UDC 599 .7115
QUANTITATIVE CHANGES IN THE VERTEBRAE IN THE 3 1 7
VERTEBRAL.COLUMN OF PINNIPEDS
'WO
In considerations of the post-cranial skeleton of main-
mais it is customary to determine the overall vertebral formu-
la. To an equal degree this applies also to the representa-
tives of the order of the pinnipeds. Thus, Flower (1885) and
Turner (1912), on the basis of an examination of about 46 in-
dividuals of various species of the true and eared seals, de-
rived similar formulas for the whole order of the pinnipeds,
namely: C 7, Th 15, L 5, S Cd 8-15 *. An exception was
presented by the walrus, in which these authors noted 14 ver-
tebrae in the thoracic section and 6 in the lumbar section.
Many years later this was still accepted as the vertebral for-
mula and this conclusion was confirmed by King (1956). Two
years later Scheffer (1958) introduced certain corrections,
as a result of which the formula tOok on the form: C 7, Th 15,
L 5, S 3-4, Cd 10-12, with no exceptions being made. Still
later King (1964) corrected this once again: C 7, Th 15, L 5,
S 3, Cs 10-12, an exception to which still remains the walrus
with its 14 thoracic and 6 lumbar vertebrae.
This small review of the literature shows that there
is no agreement in the opinions on the numerical values of the
* C cervical section (pars cervicalis), Th - thoracic section (pars thoracicalis), L - lumbar section (pars lumbaris), S - sacral section (pars sacralis), Gd - caudal section (pars cau-dalis).

51 5
vertebral formula. In the present paper an attempt is made
to investigate the causes of this and, as far as possible, to
clarify the actual situation concerning the vertebral formula
in the pinniped order. With this aim the author again ana-
lyzed the fairly extensive.material which is kept in the col-
lections of the Zoological Institute of the USSR Academy of
Sciences. There were examined two hundred vertebral columns
of almost all of the species of this grouP of animals, of
various sex and and age.
The results are presented in the table, from which
it is evident that, on the basis of the character of the ver-
tebral formulae, there may be established at least seven groups
for the pinnipeds. The first group, which embraces the largest
number of cases (66.5;:; of all of the examined individuals),-is
characterized by the formula: C 7, Th 15, L 5, S 4, Cd 8-16.
In this group are included representatives of all three fami-
lies. In the second, less numerous.group (19 of the examined
individuals), the vertebral formula is: C 7, Th 15, L 5, S 3,
Cd 8-15. In this group the walrus family is completely lacking,
but for the rest the composition is identical to that of the
preceding group. In the third group (6%),with a vertebral
formula of C 7, Th 14, L 6, S 4, Cd 11-12, there are included
in all only two*Udobenus rosmarus and Erignathus barbatus.
The fourth group, which is just as small (5%), includes Histrio-
phoca fasciata, Phoca hisnida lado7ensis and Eumetonias jubatus.
Its vertebral formula is C 7, Th 16, L k, S 4, Cd 8-16. The
* Translator's note. Sic. - Presumably . - "... two species •.."

516
T1G.ttttta
Konliecrno 1103nouhors no oTjtenant no3nonomtitay npeJlcraoure.ncii orpsl,ka nacTOUornx
p11,qu I
Sueciesn I C Th I L I S I Cd
Pt1OCIDA E.Phoca hispida 25 7 15 5 4 12--16
» • 4 7 15 5 3+ 13» 2 7 15 5 3 14-15» 1 7 15 G 4 -
Phoca hispida lado-gensis 5 7 16 4 4 14--16
Phoca sibirica 15 .7 15 5 4 13.-15Phoca caspica . 26 7 15 5 4 13-15Pagophoca ;;roenlsn-
dica 7 7 15 5 4 12-15» 1 7 15 5 4 13
1-listriophoca fasciata 2 7 15 5 4 12» 1 7 15 G 4 --» 2 7 15 . 5 3+ 11» 2 7 16 4 4 12
Phoca Vitulina 18 7 15 5 4 10-12Halichacrus t{rylsus 6 7 15 5 4 11-14 '
» 1 7 15 5 3;- 12Erignathus barbatus 5 7 15 5 4 12-13
» 4 7 14 6 4 11-12» 1 7 16 4 4 15
Cystophora cristata 4 7 15 5 4 10» 2 •1 15 5 3+. 12
» 1 '1 15 5 3 14Monuchus monachus 2 '1 15 5 4 10Leptonychotes wedde- 2 7 15 5 3 --
11t 1 7- ,15 5 4» 2 7 15 5 2 -» 2 7 14 G 3 12-14
Hydrurga leptonyxLobodon carcinopha- 3 7 15 5 3 12-13
ga 2 7 15 5 3 -Mirounga leonina 5 7 15 5 3 11-12Omtnatophoca rossil 2 7 15 5 2 12-13
»
OT.-IRIIDAB8 7 15 5 3+ 8-10
Callorhinus ursinus 9 7 15 5 4 8-92 7 16 4 3+ 9-10
Eumetopias jubatus83
77
1516
44
4,4
8-1412
» 1 7 15 5 3+ -7 7' 15 5 • 3-{- 10-11
Zalophus californianus 4 7 15 5 4 11
OD013G,VIDAEOdobenus râsmarus 7 7 14 6 4 12
» 1 . 7 15 5 4
318
Table
HO d':
CT(!;1
ptits;L:e ^TI)It
xci :;occ>^
chclL 5.IiO
LG,I-qI
ycI-i.nCtul.s`t iT1Sil.'üCiJn
(1'
3t=C1CItciii^s+
M.I'to"
I
The number of vertebrae in different
sections of the -vertebral -column
representatives of the pinniped order.

517
remaining vertebral formulae are scarcely characteristic of
the pinnipeds and each of them is represented by two speci-
mens of some different species, which comprises 1% of the
total number of examined vertebral columns. Thus in a
Weddell seal (Leptonychotes weddelli) the vertebral formula
had the form: C 7, Th 15, L 5, S 2, Cd 11. In a fur seal
(Callorhinus ursinus) there was found a somewhat different
formula: C 7, Th 16, L 4, S 3, Cd 9-10. Finally, the formula
C 7, Th 14, L 6, S 3, Cd 12-14 is charaCteristic of the leo-
pard seal (Hydrurga leutonyx).
In all of the vertebral columns of pinnipeds which
were examined by us and also by other investigators (Allen,
1880; Flower, 1885; King, 1956, 1964, 1969; Scheffer, 1958)
the cervical section was invariably represented by 7 vertebrae.
In the thoracic and lumbar sections there are counted, in to-
tal, 20 trunk vertebrae, which is characteristic also of the
representatives of the order of carnivores. Unly in two cases:
in one ringed seal (Phoca hispida) and one ribbon seal (His-
triouhoca fasciata) were there 21 trunk vertebrae. Such cases
may be considered as atypical deviations from the norm.
In the thoracic section 15 vertebrae were found in 87%
of the examined vertebral columns. This number, evidently,
should be considered as being the most typical for the pinni-
peds. However, as can be seen from the summary table, there
are encountered individuals with 14 and 16 thoracic vertebrae
(6.5% in each case). A thoracic section with 14 vertebrae,
as has already been mentioned above, is typical of the walrus.
The second species in which there are 114 thoracic vertebrae

518
is the bearded seal (Erignathus barbatus) and moreover this
species was also characterized by having 15 and even 16 tho-
racic vertebrae. Such a variation of the vertebrae under con-
sideration possibly conforms, to some extent, with the con-
cepts of certain authors (Winge, 1941; ChaPskii, 1955) on
this species as being little specialized and primitive. It
is interesting to note that in the Ladoga ringed seal (Phoca
hisuida ladogensis) the thoracic section is formed exclusively
of 16 vertebrae. This feature distinguishes this seal not
only from the soecies Phoca hisPida but also from the whole
of the genus Pusa and, possibly, may prove to be a taxonomie
feature. Perhaps the same applies also, though this is less
convincing because of the very small number of individuals
examined, to one other species of true seal - the leopard seal,
the thoracic section of which is composed of 14 vertebrae.
In the eared seals a variation in the number of thora-
oie vertebrae, from 15 to 16, is observed only in the fur seal
(Callorhinus ursinus) and Steller's sea lion (EumetoPias juba-
tus).
The lumbar section in the pinnipeds characteristically 320
has 5 vertebrae, though more rarely there may be encountered
4 or 6. Since, as has been mentioned above, the number of the
trunk vertebrae in the pinnipeds is constant, the frequency
of occurrence of such cases is precisely the same as the nulliber
of encounters of 14 and 16 thoracic vertebrae. In view of the
fact that these values are correlated, all of the arguments
rresented above on the variability in the number of the thora-
oie vertebrae will be valid also for the lumbar vertebrae.

519
As is evident from the literature references, each
author considers that for the oinnipeds that number of sacral
vertebrae is characteristic, which seems to be in accordance
with the results of his own analysis of.the vertebral formula.
In summing up, the authors agree that for the true and eared
seals the typical sacrum is composed of both 4 and 3 vertebrae.
If, however, one again turns to the summary table, then it
can be noted that thm± all of the enumerated investigators
were to some degree right, for in the sacrum there are found
four, three and even two vertebrae. Most often (79%) the
sacrum is comuosed of four fused vertebrae, which, apParently,
may be considered as bein9,- typical of all of the eared and
true seals of the Northern hemiSPhere. Sometimes, however,
in these seals there may be found a sacral bone which is formed
of three fused vertebrae. With a mire detailed examination
of such cases it turns out that the sacrum, even if only mor-
phologically, is nevertheless formed of four vertebrae. Two
facts lead one to this conclusion. Firstly, the caudal epi-
physis of the third sacral vertebra and the cranial euiPhysis,
in this case, of the first caudal vertebra are considerably
flattened, their edges have lost the rounded outlines (figure
1), in contrast to all of the epi -ohyses of the bodies of the
vertebrae in the other sections (Figure 2). Secondly, the
transverse Processes (procesus transversus) of the sacral ver-
tebrae, which here form longitudinal ridges (pars laterales),
in the normal four-vertebral sacrum zradually become narrower
and thinner from the first vertebra to the fourth, merginF
completely with the body of the fourth and last vertebra in

520
Pm. 1. Kayanabilan TpexrIo3B0111:0130r0 kpe- . emit napril ii nepnbtri knocToeoii 110300110K (BIM. C
nekTpambuofi cTopakm, nopuko- Bbie komepa i10 -2B0IIK013 B kpecTu.e)
Figure 1. Caudal portion of three-vertebral sacrum of a
harbour seal and first caudal vertebra (view
from ventral side, the numerals indicate the
ordinal , number of the.vertebrae in the sacrum).
Pue. 2. 4e-rupexao3nokkoublii «TiopmaabkhuI» KpecTeu iaprui (aka c ecin-paibuoii cTopoubl)
Figure 2. Four-vertebral "normal" sacrum of a harbour
seal (view from ventral side). •
its caudal portion (see Figure 1*). In the case, however, of
the three-vertebral sacrum these ridges, on the contrary,
become slightly wider and thicker on the body of the third
di? vertebra, which is clearly not "ended", and only on the first
caudal vertebra do they become thinner and merge with its
body (see 2igure 2*). Such a "morphological incompleteness"
* Translator's note. Sic. Text references to figures obviously

5- 21
i
•n
of some of the three--ve.rtebral sacra considerably reduces,
if it does not practically stop, the freedom of movement of
the J_ast sacral and first caudal vertebrae relative to one
another (such cases are marked in the table by the + sign).
Consequently, the thought arises that such sacra function
like the four-vertebral sacra. Thus, the number of individuals
for which a sacrum composed of four vertebrae is typical be-
comes even 9-:reate-r. There are also encountered cases of a
clearly normal sacrum that is formed out of 'three or even two
vertebrae, representing a morphologi cal ly single sacral bone.
Such a sacrum is bas i ca:lly characteristic of all of the Imtarc-
tic seals, in which there is observed a tendency towards a
reduction in the nurnber of the constituent elements of the
sacrum. In -cne examined series there were also noted two cases,
xkxra* in one individual of the Caspian seal (Phoca caspica)
and one harbour seal (Phoca vitulina largha), in which the
sacrum was composed of five fused vertebrae. This phenomenon
represents a clear deviation from the norm and may be consi-
dered as ateratism s in both cases the first sacral vertebra
was separated from all of the others, while the junctions of
the last sacral vertebra with the first caudal vertebra bear
traces of a distinct ankylosis.
In the caudal section it is generally impossible to
establish any regular pattern of the variation in the number
of vertebrae. in one and the same species the number of ver- 322
tebrae may vary from ô to 12-16. Therefore it is scarcely
possible to utilize the number of vertebrae in this section
as a taxonomic feature of the species, subspecies or popula-
tion rank, at least within the order of the pinnipeds (Yablokov,

522
4i
1963). The only feature which may be noted is the abbrevi-
ated nature of the tail of the eared seals s the tail in
these is formed of a smaller number of vertebrae (from 8 to
].Ii•) than in the true seals, in which the caudal section is
composed of 10 - 16 vertebrae.
Summing up all of the information ,oresented above, the
general vertebral formula of the pinnipeds may be presented
in the following form o C 7, Th 1l1•-16, L 4•--6, S 2--1I•, Cd 8-16.
JI I•ITIà PATS' P A
I ya!u.cai!ï K. K. Onbrr nepecasarpa cisc.reta!,t nRnarnoc•ru!cti Tsoneneit ncsacensers-
c•r!!a Phocinae. TpynL! 3oonor!!vecror(, nneT!rryra AH CCCP, T. XVI1, 1055.2 Sld.ior:oa A. 13. i•ferozopLae npez!iap!rre.^t.!n,!e MaTepna.•!!,! K >sopcpo.nrm!ec!coï!
xapa!nrpt!c•rn!<c rhe!!;!a!!:1c!coro •r!o.aenst lie:!oro n l'pc+nna!tilcxoro mopeil. Ccï•nayt!.-ncc.n. pc16oT CerfllIllPO, Apza!n•c.i!,c!c.'1963.
Allen J. A. 1-iislmy North American Pitinilieds. Washinhton. 1880.^. Flotc^e; IV. 1/. Ost^^oln^y of the mai^^ur!alia. London. I885.
Kinr; J. E. The nsonk seals (hcnus dlonachns). 13u11. Brit. Mus. (Nat.I tit;, Zool. 3: 201--956, M56.
Ring J. G. Seals of the world. London. 196,1.King J. E. Some aspects of the n!!a;tomy 'of the Ross seal, Ommotophoca
r^si (Pinnihedia: Phocidae). British Antarctic Survey, Scientific`reporls, no, 63.London. 1969.8 Scüc(f.er 17. B. Seals, sea lions and walruses. London. 1958.C^ Turner W. 'flic marine mammals in the anatomical rnuscum of the Univer-
s(tv of Hdinburf*. London. 1912.10 ü%in{;r. II. The interrelatiouships of the mammalian l;enera. Vol. 11, Koben-havn, 1Jfl.
REFL,RE:IdCES
1. Chapskii K. K. An attempt at a revision of the systema-
-tics and diagnostics of the seals of the subfamily
Phocinae. Trudy Zoologicheskogo instituta AN SSSR
(Transactions of the Zoolo^ical Institute of the USSR
Academy of Sciences), vol. XVI1:, 1955-

523
2. Yablokov A. Y. Some preliminary data on the morpholo-
gical characteristics of the harp seal of the White
and Greenlad Seas. Sbornik nauchno-issledovatel'skikh
rabot SevPINR0 Uorthern Polar Research and Planning
Institute of Larine Fisheries and ceanogra.)hy),
Archangel, 1963.

524 .
UDC 599 ,7/15 .3
V. A. Zheglov and K. K. Chapskii
TEST OF AN AERIAL SURVEY OF THE RINGED SEAL AND THE GREY SEAL
AND OF THEIR HOLES IN THE GULFS OF THE BALTIC SEA
AND ON IAKE IADOGA
Over the course of many years in the northern and
eastern part of the Baltic the ringed seal (Pusa hisoida gm*)
and the grey seal (Halichoerus grypus F.) have served as
objects of the marine sealing industry; these seals have
been caught here at all periods of the year and without any
limitations. «Furthermore, in Sweden and Finland a bounty is
paid for catching the seals (Curry-Lindahl, 1965).
Every year there are caught by these countries in the
coastal waters about 15-16 thousand seals (Ropelewski, 1952;
Bergman, 1958).
it is natural that such destructive sealing has af-
fected the stocks of the seals.
In the territorial waters of the Soviet Union (in the
Gulf of Riga and the Gulf of Finland) the sealing is carried
out by sealers from fishery kolkhozs in Estonia and Latvia,
the annual catch of which does not exceed 1200 - 1500 head
(according to official data from the Estrybakkolkhozsoyuz**,
in which each 100 luy of oroduction from sealing ooerations is
considered as equivalent to 3 seals).
711rom 1970 a limit has been placed on the catch of Bal-
tic seals in the USSR.
Translator's notes. * Sic. Schreber is the species author. ** Acronym for Estonian fishery collectives union.

525
o
o
Up to the present time the representations of the
numbers of seals in the Baltic remain very indefinite. More-
over, there has been noted a marked reduction in their popu-
lation numbers.
Thus, according to Bergman (1958), "the stocks of the
ringed seal in the coastal waters of Finland have decreased
considerably from the twenties and thirties up to the late
fifties of this century. In the southern part of the Gulf of
Bothnia they are oarticularly small, in the middle part of the
Gulf of Finland and in the central portion of the Gulf of
Bothnia (Salkameri and Melenkuru) they are rather small, in
the shallow waters off the islands of Valgrund and Berge they
are not more than average, and in the eastern part of the
Gulf of Finland they are average, at the level of the 1920s and
1930s. In comparison with the 30s, even in the northern part
of the Gulf of Bothnia the population numbers of the grey seal
have decreased considerably. In the western part of the Gulf
of Finland there were considerably fewer grey seals in 194(7)-
1950 than there were in 1920; there were a few more of these
in the eastern part of the Gulf of Finland. The total number
of grey seals within their territorial waters was, on average,
considerably smaller than the number of ringed seals. Their
ratio, according to Gotberg's data, comprises 1 :3".
According to the tentative estimate of Sundstrem (1962),
the Baltic population of the grey seal at the end of the fif-
ties was about 10 thousand head. A marked lowering of its
population numbers was pointed out in the middle sixties al-
ready by Curry-Lindahl (1965). According to Smith (1966), the
stock of grey seals in the Baltic comprises 5 thousand head.

52 6
The only approximate indication of the population num-
bers of the Baltic ringed seal was found bills in a Finnish
fishermen's journal ("Kalamichen Viesti", no. 1, 1966), where
it was stated that each year after the sealing operations in
Finnish waters there remain about 20 thousand rineed seals.
There are very few operational data of any kind, on the basis
of which judgements can be made on the population numbers of
the two species of seals in the Gulf of Finland and the Gulf
of Riga.
It is even more difficult to judge the population
numbers of the Ladoga ringed seal, of which about 500 head
are cauet each year by hunters and fishermen.
K. K. Chapskii (1932) assumed the possibility that
about 20 thousand seals could be living in Lake Ladoga; in
the opinion of S. P.. Sokorin (1958) the population numbers of
the Ladoga seal were nor high, while A. I. Zubov (1965), re-
ferring to data from the State Research Institute of Lake
Fisheries (GWnIL.RKh), estimates the stocks of the Ladoga
seal at 5 - 10 thousand head.
In connection with the growinz demand of the sealing
industry there has come to a head the necessity for reorga-
nizing this on a scientific basis. A most important condition
for the rational utilization of its resources should be the
knowledrre of the population numbers and other questions re-
lated to this.
2rom 1969 the AtlantnIR() (Atlantic Research institute
of i:.arine Fisheries and ceano,9raphy) has been conductinE work
on the study of the seals in the eastern coastal region of the

527
Baltic Sea and Lake Ladoga, and therefore in arch-April of
1970 there was made the first attempt at an aerial survey of
these seals by workers of the Atlant .HIRu.
MATERIALS AND METHODS
As far back as the forties Dorofeev (1940) wrote about
the advisability of employing aviation for surveying marine
mammals.
af
•
In recent years Tikhomirov (1966), Fedoseev (1966),
Shustov (1969) and other investigators have conducted aerial
surveys and aero-visual counts of seals in the Far East.
In the Gulf of Riga and Gulf of Finland and on Lake
Ladoga no.special aerial surveys of seals were conducted up
to this time.
Conducting an aerial survey of the ringed seal . and the
grey seal in *these regions was accompanied by considerable
difficulties. UD to the present time there has been no metho-
dology worked out fàr the counting of these seals in the zone
of the landfast ice, although we know of the attemnt by Sviri-
dov (1954) and by the far-eastern investigators, who conducted
aero-visual counts of the ringed seal and other seals on broken
drift ice. At the same time, it is known that the ringed seal,
as a rule, does not form concentrated whelping and moulting
patches, similar to the patches of the harp seal, and therefore
we did not employ continuous aerial photo=aphy.
In elaborating the methodolopy for the aerial survey,
we made use of information obtained by Questioning local sealers
and the experience acquired in counting seals by the far-eastern
investigators and other authors.

a M + m C. V.
45 3
Table 1.
Some indices of the periods of retention of tags
in the digestive tract of the experimental animals,
hours.
Ta6auua 1
He KoTopme nourtaaTeas spemess aa,sepiass meTok s nnut,esapinemmom Tpatne no;tonbusbtx :seirtoTtmx, trac
miiTepu la Material
11.TacTmaconb1e MCTIZII 73,5±16,4 73,5 100 i'lastic tags KvottiYieceS oi rubber
59,0 25 ,cm stmatibt 22,64.2,7 13,4
111e,utut Kycotput macTmaccht 14,9±0,8 3,4 22,7 20 Small pieces of
plastic * nPume ■mum.. B CLIN3B C TCM, 4T0 cymcr Beim ,tx pa3.111411rf B 0;1licam1l!bn:
me-rot: cpc.au >1:1moTm ■ IX IIC TZlem Hue 11 imucaeti b! oGoCim,esume ;mumble.
*Note. In view of the fact that significant dif-ferences were not found in the retention times of tags of the same type. between the animals, generalized data are presented in the table.
The retention of the tags and, probably, Of the undi-
gested remains of the food also depends on the amount of food
consumed by the animals and on the characteristics of the
working of the digestive organs. Thus, the plastic platelets
that were given to Ushkan before a prolonged fast were found
after 55 days, while with Vega (in a similar case) they were
found after 20 days. In contrast, when Vega was fed abundantly
during the first third of February (the daily ration exceeded
4 kg) the tags often appeared in the tank 4 - 5 hours after
the ingestion of the food.
However it should be noted that the small pieces of
plastic, just like the otoliths, did not pass through the
gastro-intestinal tract simultaneously: on the average, about
20 t =3,1 AB
t =3,5
2,7 BB

454
80:, of these appeared after 10 hours, about 15 approximately
after one day, and 5 - two days after being given to the
animals. If the working of the digestive system of the ani-
mais under the natural conditions does not differ substantial-
ly from that in the test conditions, it may be assumed that
the otoliths, collected from the intestines of caught animals,
must belong to fishes that were caught by the seals within a
period of 2 - 3 days, although the main mass of the consumed
fishes had been digested less than half a day previously.
In spite of the fact that the animals were held in
small tanks, i.e. in almost immobile conditions, the assimila-
tion rate of the food was found to be very high. The faeces,
which are composed to a considerable degree of undigested or 282
poorly digested remains of fishes (bones, otoliths, crystalline
lenses of the eyes etc.), under a moderate feeding regime
(March - May) on the average had a weight of about 4 - 5% of
the value of the daily ration*. There were found differences
among the animals in the weights of the faeces, expressed as
percentages of the daily ration, though statistically (with
P 0.0) these were insignificant.
Because of an insufficiency of food, the animals did
not always obtain the necessary amount of fish. Moreover,
from October 10th 1967 till January 9th 1968 Vega did not feed
at ail, while Ushkan did not feed at all from January 13th
until March 3rd 1968. During 60 days of fasting Vegas weiet
fell by 10.6 kg (from 38.8 to 28.2 kg), by 29.7% of the initial
weight. (ver 50 days of an enforced fast the weight of Ushkan
dropped from 40 to 29.45 kg or by 26.4%, i.e. he lost 10.55 kg.
The mean daily weight losses comprised: 177 g for Vega and 211 g for Ushkan.
* Urinary wastes were not taken into account. -

455
o The two month fast of the seals in the described con-
ditions should not be considered as the limiting value.
Over 222 days (including the period of fasting) Ushkan
consumed 301.4 kg of fish, i.e. on the average he ate 1358 g
per day. From the initial (39 kg) to the last weighing he lost
3.6 kg or 9.8% of his initial weight. Over 154 days (also in-
cludine the period of fasting) Vega consumed 191.5 kg of fish,
i.e. on the average 1244 g per day. Her weight dropped by
7.95 kg (from 38.8 to 30.85 kg), i.e. by 20.5%.
Ilianyunya obtained food in some amount or other over
the course of almost the whole of the holding period. Never-
theless, the lack of food during the winter period also affected
her. While initially her weight was 65.8 kg, after 184 days
this was equal to 61.9 kg, i.e. she lost 3.9 kg or 6.3% of
the initial weight.
The animals reacted very sensitively to changes in the
size of the ration by changes in their own weight (Figure 1).
With a low daily norm over several days, the weight of the
animals began to fall, while with an increase in the norm the
weight began to increase. At the same time, in the changes
of the body weight with respect to the amount of food consumed
there was observed a certain asynchrony, a semi-antiphase re-
lationship of the maxima and minima. It is quite possible
that this time lag in the changes in weight in relation to the
size of the ration was caused by a differing assimilability
of the food.
In the conditions of the experiment, i.e. with the
markedly restricted movements of the animals, it was found to
be possible to approach the question of estimating the daily

66
621
56 4
2
CIII1 0
pa 401 • 1 36
3 '1 I 28i
L__
456
361
Pic. 1. 1,13menemie peca Teaa ileput MMICIMOCU IDT'IleM1 , 11111U
Cr(VMOrOpMWOlia
I — BCC Tesla (Ke); II— crrognmil palm. on (K2):
a — Dora, 6— Mainoim, o — YulKalt_
Figure 1. The change in body weight of the seals in relation to the size of the daily ration.
- body weight (kg); II - daily ration (kg);
a - Vega, b - lanyunya, v - Ushkan.
ration which, in energetic respects, is close to the value of
the standard (basal) metabolism. With this aim, in March -
May 1968, when the animals were provided with food to an
adequate degree, we attempted to select for each animal just
such a daily ration, i.e. to establish such a moderate feeding 283
regime in which the weight of the seals would remain more or
less stable (Table 2).
The animals were primarily fed with the yellowfin Bai- 284
kal sculpin*. Both species of Baikal oil-fish**, the longfin
Baikal sculpin*** and also the benthic gobies were given in
the form of a small supplement. Even under such conditions,
Translator's notes. * Cottocomephorus comephoroides or C. grewingki ** the "big golomanka" Comephorus baicalensis and the "little. golomanka" - C. dybowskii. *** cottocomeohorus inermis

11•5 .7
Table 2.
Some indices of the daily ration with a stabilized
weight of the experimental animals.
Ta6:inua 2
1-1eNororu:e noha3aTen11 cyTO11110r0 pacqirona npn craGn:11+3nponaHnont eecenoAont,lirthlx xcunoTnblx
[lapaM1terpt,t 26. 111-30. IV 21.11==2.1V
yacupclteicnttü s ï n e^aii^.rF,'^lrE'•ters J'mt<an Ilera
-of distribution Bec eC Body
34,29±0,10
34,0-34,70,24
0,7
6
33,13±0,09
33,2-33,8
0,23
0,7
7
M^E m
lim
IfC. V.
n
^teee c^ô Nozoo par;da^^^j^ ration,
2097-!:49,1
1900-2500282
13,4
36
1872:L 84,2
150-3000
533
28,4
41
12. 11t-14.V
r:dnvûnyaD1autbust
kg.
59,54 ±0,48
59,3-61,8
1,45
2.4
9
g
2559-54,91000-4500
429
16,7
63
when the energy losses for all possible movements were reduced
to a minimum, the animals required a fairly large quantity of
food. In percentages of the body weight the rations compr.ised :
for A:anyunya - 4.2fo, for Vega - 5.6% and for Ushkan - 6.1/.
There is no doubt that under natural conditions the ration is
at least 1.5 times greater than in the conditions of the test.
Just like with other mammals, on a per unit body weight
higherbasis the young animals required a.inaut of energy accumulated
in the food than did the adults.
The food coefficient changes with the age of the ani-
mals. ;t1hile for a body weight gain of 1 kg Vega required 23 kg
of yellowfin i3aikal sculpin, and Ushkan required 26 kg, the
adult female had to consume twice as much of this same fish or
511• kg .
weight,

45 8
The maximal uptake of food at one time comprised 2 kg.
Generally, however, the animals consumed not more than 1 kg
in one meal (within 1 - 5 minutes). When fed "ad libitum"
Manyunya consumed not more than 5.6 kg of pelagic gobies in a
day, Ushkan - 5.3 kg, and Vega - 4.5 kg.
In spite of the fact that the food was usually given
during the day (once every 2 )-i hours), the main mass of the
food was consumed in the evening (creuscular) period.
In March - April, during the period of the optimal
feeding regime of the animals, there were set up experiments
on the digestibility of the otoliths of the big and little
golomankas and also of the yellowfin Baikal sculpin, i.e. the
otoliths of those fishes which comprise the basis of the feed-
ing of the seal under natural conditions. The otoliths were
removed from freshly caught fish, measured under a binocular
along their long and short axes with a precision of to 0.05 mm,
fed to the animals, and later these same otoliths were picked
out of the faeces of the seals and again measured. Subsequent
comparison showed that the average values of the dimensions
of the otoliths of the control and experimental series differed
from one another at a high level of statistical significance
(Table 3). On average, for the three seals, the length of the
otoliths of the big golomanka decreased by 5% after passage
through the gastro-intestinal tract, of the little golomanka -
by 3.4% and of the yellowfin Baikal sculpin - by 4.3%; the
widths decreased, respectively, by 6.9, 4.2 and 5.7% from the
initial dimensions.

M + m C C. V. tAB
A
A
A
A , 13
A
A
11,7
11,5
459
Table 3. Changes in the dimensions of the otoliths of fishes with their passage through the gastro-intestinal tract of the experimental animals, mm.
T a 6 a It us a 3
Ilanicaeuan paudepoa oToasyron pb16' lipli i< ripoxo.:«aellau . . nepea ›Keay,r(oliao-icitaiegablil Tpaar noaonbrrahlx. ›auncrrumx, iLSt
--01011Lhs Parameters—of the di7st-1-1-bution Ilapameind pactipulenetinii
1 Inweimtm c'elm01
chal_g_eL.,g9r ies*
Bo/Lamm coAcmuitea Big golomanka
,annuaL
InpaaaW
RmtnaL
L
• IlInpunziVi
2,246:120,0095 0,288 12.8 919 „
. 2,134±0,0105 0,293 13,7 774
1,799+0,0005 0,197 10,9 919 ,,„ ek
1,675± 0,0077 0,215 12,8 774 '`"-'
MaAaa cOACMIJIlea Little golomanka
1,677:3-0,0036 0,161 9,6 9090 n i
1,620±0,0052 0,179 11,5 1237 ̀.'"
1,279i:0,0031 0,140 10,9 2090
1,224.±0,0015 0,151 12,6 1937 10,1 . . .
XCJITOlepbtAblit 6blitOle. Ye 1101f scu pin
1,883±0,0053 0,145 7,7 748
1,803:120,0044 0,177 9,8 1599
1,096±0,0025 0,069 (3,3 748 1,034:120,0043 › 0,191 18,8 1599
e A — J&O onma, 13 — ontaTa.
* A - before experiment, B - after experiment.
L - length W - width
The degree of digestion of the otoliths was found not
to be uniform in each of the three seals. The otoliths were
found to be somewhat more strongly digested in Manyunya, more
weakly in Vega. lt may be assumed that the intensity of the
digestion of the otoliths is under the control of several fac-
tors, being dependent on the time spent in the gastro-intesti-
nal tract, on the age of the animals, their nutritional state,
motility etc. It is a curious fact that in each of the animals
the otoliths of the big golomanka were most strongly digested, • 285

4•60
less so - of the yellowfin Baikal sculpin, and still less -
of the little golomanka. In this, one cannot but perceive a
species si)ecificity of the otoliths themselves. It is also
interesting that the otoliths decreased in size somewhat more
markedly along their short axis than along the long axis.
Apparently, the digestion of the otoliths depends, among other
factors, also on the surface that comes into.contact with the
gastric juice.
The digestibility of the otoliths, which on the whole
is very similar in the golomankas and in the yellowfin Baikal
sculpin, should nevertheless be evaluated as being insignifi-
cant. The otoliths which.have been in the digestive system
of the seals do not differ superficially in any way from those
which were removed from the fishes. By visual evaluation there
are also no differences in their optical structure: the annual
rings are distinguishable with the same degree of definition
as in the original otoliths.
Thus, it may be stated that the study of the feeding
of the seal on the basis of the otoliths is well-founded.
Lending itself to reconstruction by this method is not o•nly'
the species spectrum of the food but also the size, weight
and age of the fishes consumed by the seals. This last circum-.
stance is of considerable significance for acquiring an under-
standing of the inter-relationships which are established
between the representatives of the two final trophic levels
in the pelagial of Lake Baikal: the golomankas and goby fishes
and the seals.

UDC 599.53r
4.6 1
V. Nt. Hel'kovich and^V. S. Gurevich
QUESTIONS RELATING TO THE CATCHING AND LONG-TERM: HOLDJ:NG
OF DOLPHINS IN CAPTIVITY
The representatives of the suborder of the toothed
cetaceans, the dolphins, attract investigators of the most
diverse specialities. This is connected with the study of the
profound adaptive changes, brought about by the habitat, and
several biological characteristics which are of considerable
interest. Among these, primarily, belong: the methods of
orientation and navigation.in the ocean; the characteristics
of the hydrodynamics; the diving to great depths and for pro-
longed periods; the characteristics of the behaviour and
higher nervous activity.
The necessity for conducting experimental studies to
resolve these problems has forced us to seek ways of trans-
forming dolphins into experimental animals.
In this paper an attempt is made to generalize the
information available to us on the catching, transportation
and medical servicing of experimental dolphins abroad, and
there are presented some of our own studies from the period
1965 - 1969.
THE CATCH
There are no special scientific reports devoted to the
methods and procedure for catching and transporting live dol-
phins. The details of the methods employed are professional
286

4.6 2
secrets of the firms which are engaged in -providing the an!-
mais for aquaria and oceanaria (i'orris, 1966). However, the
scattered framentary data in scientific ouplications.of re
' cent years and our own experience permit us to present a ge-
neral picture. Catching the dolphins may be subdivided into
the catching of individual animals and the catching of groups
of animals.
Individual catching permits one to select the required
specimen from a school and decreases the mortality. A pioneer
of this method was M. Santini, who supplied dolphins to the
Florida "Marineland". In addition to him, such catching is
practiced by F. Brokato, the captain of the ship "Geronimo",
who supplies animais to this same oceanarium, and by Jaques
Ives Cousteau.
Generally the catchers on fast-moving vessels follow
the school until they are able to isolate and catch the marked
out dolphin by means of a throw net or a Special catching de-
vice. M. Santini jumps onto the dolphin in the water and holds
it until the launch approaches.
The catching of groups of dolphins is carried out, as 287
a rule, with the help of nets. In addition, with this method
there are employed vessels or motor boats as beaters, aerial
reconnaissance for finding accumulations of the animals and
also means of transporataion for moving the captured dolphins
to their destination. Such methods of catching were utilized
by K. Rei (1963) for capturing belugas, and by D. Gascin (1966)
for catching bottlenose dolphins.

1 1 ,63
PliC. 1. Re.111.41111b! B • aaomane. CdBlep Bp01131100,1iT no,acynnq
CCT11, a no,nnn pacTnrnnaior ce, trro61,1 ne c.no);;n:m TegenueNt
*Figure 1. Dolphins in "aloman" (purse seine). The seiner takes up the net, while the boats stretch it out so that it is not furled by the current.
R. Busnel (1966) worked out a special method for
catching dolphins with the aid of a powerful acoustic signal
which temporarily stuns the animal.
The existing method for catching dolphins in the Black
Sea was designed for catching groups of animals (Figure 1).
In spite of the fact that it is considered as a commercial
method, the sweep net catch is acknowledged as being the most
effective method up to the present time. The details and tech-
nique of the commercial fishing have been described in detail
in our literature by Tatarinov, Eremaev, Larti, Freiman,
Kleinenberg, Gubenko and others (according to Kleinenberg S. E.
et al., 1964.), and therefore we will not stop to consider this
in detail here.
When we commenced the experimental work with the dol-
phins, utilizing accumulated experience, we somewhat modified

464
the commercial purse seine fishing method, which turned out
to be fully justified. Thus, out of the recently caught dol-
phins (white-sided and bottlenose dolphins) only 24% of the
animals died at the time of the catch. Protective measures
were employed during the catching operations: care was taken
that the current would not furl the purse seine, while during
the hauling in of the net by the boats and the SChS*the net
was kept stretched; during the approach of the transport
vessel to the side of the SChS for the trans-shipping of the
animals between these there were set up spacing struts.
The captured animals were Pulled out of the water with 288
• the aid of stretchers, inflatable rubber mattresses, by suspen-
sion with a soft sling by the tail or simply by hand, depen-
ding on the species and weight of the animal. The main factor
in this operation is speed. The more rapidly the animals are
brought onto deck from the drawn up seine net, the lesser will
be the traumas and the less likely they are to get entangled
in the nets and to choke.
With the contemporary nethods of catching groups of
small dolphins (the common dolphin and the common porpoise)
the best reeults are obtained by the use of a fine-mesh kapron**
Purse seine net (of the scad purse seine type), since with this
the losses of the animals are reduced.
The best method for trans-shipping is the use of a
stretcher with a soft covering and an opening for the Pectoral
fins. The tans-shipping of the small species of dolphins
Translator's notes. * SChS Black Sea semer
** kapron - a synthetic fibre.

465
may be successfully carried out by hand; in transferring large
specimens, as an extreme measure, suspension by the tail in a
soft sling may be tolerated.
TRANSPORTATION
The transportation of dolphins at the Present time is
accomplished by various methods: by ships, airplanes and motor
vehicles.
Transportation of dolphins by sea was first accomplished
in the seventies of the last century, when two belugas were
brought to the Brighton Aquarium in England from Labrador and
Newfoundland in the hold of a ship. Both animals died (Slijper,
1962). In 1907 the New York Aquarium contained in its pools
a group of bottlenose dolphins, while in 1912 in transporting
a new batch of dolphins to this aquarium Taylor first employed
soft litters for their transportation. In recent years the
technique of transporting dolphins by sea has been considerably
improved, which permits them to be transported over fairly
great distances. However, the most successful transportation
is accomplished to those aquaria which are located on the
coast and may receive the dolphins 1 - 2 hours after they are
caueht.
Successful transportations by sea in recent years have
been accomplished by: Bel'kovich (1964), Kritzler (1949), Ray
(1963), Andersen and Dziedzic (1964) and Gascin (1966).
The curator of the ilapier oceanarium in New Zealand
considers that the transportation of dolphins by airplane, in
particular of the bottlenose dolphin, over considerable distances

4.66
in air without water over the course of a period of more than
two days apparently does not cause them any harm (Gascin, 1966).
In 1960 D. Lilly transported two bottlenose dolPhins
by airplane. The dolphins were placed in a special box of
perspex and Plywood, 2.8 x 0.6 m in size. Within the box, at
a distance of 17.5 cm from one another, there were installed
plywood partition-frames, strengthened with perspex. From the
town of St. Augustine (Florida) to the island of St. Thomas 289
the animais were conveyed aboard a cargo plane, the flight of
which was at an altitude of 2000 m and lasted 7 hours. During
the course of the flight the respiratory rhythm of the animals
was measured. The altitude does not have any effect on the
frequency of the respiration, if the dolphins are in water
"in a suspended state". K. Rei (1963) transported three belugas
from the south-western coast of Alaska to the New York ocean-
arium. The animais made the 5900 km journey on stretchers,
boats, a fishing vessel and then on a special B-17 airplane.
The animais were Placed in wooden boxes, partially filled with
water, the Walls and bottom of which were covered with a soft
plastic lining. All three belugas withstood well the 24-hour
non-stop flight. However, immediately after landing at New
York airport one of the belugas died.
Dudok van Heel (1966) successfully transuorted by air-
plane four bottlenose dolphins from the USA to Holland. He
indicated that for the successful transportation by air the
airplane should fly at an altitude of 1500 m. To protect them
from contusions, the animais were suspended in hammocks. The
flight lasted 16 hours and ended successfully.

467
In 1969 there was carried out a successful transporta-
tion of two male common porpoises aboard an IL-14 airplane from
the Crimea to rix)scow; the animals were delivered in a satis-
factory state (Sokolov et al., 1969).
Transportation by motor vehicle is employed mainly
over short distances: from the shore to the oceanarium or
from the oceanarium to the airport. The transportation is
accomplished in open vehicles, employing soft linings and maintain-
ing protective measures to prevent the action of the direct
rays of the sun on the integument of the animals.
It is difficult to say who first employed a truck for
the transportation of dolphins, but as early as 1948 Kritzler
transported two young male pilot whales (Globicephalus macro-
rhynchus), which had been stranded ashore in Florida to Farine-
land.
D. Lilly (1960) delivered by truck, from Marineland
to the airport in St. Augustine, two bottlenose dolphins over
a distance of approimately 30 km. W. Kellog (1966) conveyed
an adult female bottlenose dolphin to his laboratory in an open
truck. Among other investigators, who have used used the me-
thod of conveyina dolphins to the laboratory by motor vehicles,
should be mentioned Schevill and Lawrence (1956), M. Caldwell
and D. Caldwell (1963), Ray (1963), Andersen and Dziedzic
(1964), Heel (1966) and Sokolov (1969).
For the transportation of captured animals we utilized
a refrigerator ship (TKI1S), and moreover we employed several
variants, which may conventionally be divided into two methods:
"semi-aquatic" and "aquatic".

4, 68
The basic factor, necessary for the successful trans- 290
portation of dolphins, is the establishment of optimal condi-
tions for thermoregulation. A prolonged sojourn of the dol-
phins in air, especially under the action of the direct rays
of the sun, leads to a fairly rapid death of the animals. This
is explained not only by the low thermal conductivity of the
air, which causes overheating and heat stroke during the summer
period in literally a few hours, but also by a disturbance in
the activity of the cardio-vascular and respiratory systems.
The latter occurs as a consequence of the infringement of the
"practical weightlessness" on dry land and the compression of
the thorax by the weight of the animal, and also from the
marked shrivelling of the skin,, wrinkling and flaking-off of
fl the euidermis.
11, Thus, the transportation of dolphins out of water brings
to the forefront several requirements, on the correct fulfilment
of which deuends the success of the entire transportation. The
animals must be protected from the action of the direct rays
of the sun, care being taken that the skin is kept moist all
of the time; in addition, to avoid the formation of bed sores,
the dolphins should be placed on thick, fairly soft linings,
which may to some extent compensate the temporary loss of the
state of "suspension" by increasing the area of support of the
body. For this on the deck, shaded by awnings from the rays
of the sun, there is set up a soft bedding; of inflatable
rubber mattresses, boats or thick porolon*. Un these the ani-
mais are Placed individually. A sufficiently thick and soft
beddinP: will somewhat decrease the normal loads in the region
of the thorax and abdomen.
* Translator's note. "Porolon" - presumably an aerated plastic of the styrofoam type.

469
Pnc. 2. Reamjnin B 6opro13or1 II a mi e jia ramane. Ha crinne muna mapaeBan nanne,na
Figure 2. Dolphin on a hammock in a tub aboard ship.
The gauze coverlet is evident on the back of the animal.
The second method of transportation is the "aquatic"
method. For,this there is utilized a tub aboard the trans-
porting vessel, with running water, or special containers, in
which the dolphins are secured in hammocks (Figure 2). The
securing is absolutely necessary,even when there is only a
slight rolling and pitching, in order to minimize the trauma-
tism in general. In addition to this, the individual secure-
ment prevents the possibility of the occurrence of a panic
among the dolphins, during which they fight, ripping the skin
and inflicting serious injury to one another with their power-
ful tails. The securement also prevents, and this is probably
the most important factor, the entry of water into the blowhole.
In one standard shipboard tub, 180 x 250 cm in size,
there are accomodated, depending on their length, 1 - 3 bottle-
nose dolphins or 5 - 6 white-sided dolphins. The containers
are for individuals.

47 0
The elaborated method for transPorting the animals
on the special soft bedding and in the shipboard tubs on the
refrigerator ships (TKhS) were found to be suitable for main-
taining the health of the dolphins for 54 hours and for their
subsequent prolonged holding in captivity.
MEDICAL SERVICING
The problem of the medical servicing of dolphins, kept
in captivity, has attracted the attention of many investigators
(Brown, 1962; Brown and Norris, 1956; Brown and McInture,
1960; Ridgway, 1965; Ridgway and McCormick, 1967; Nagel
et al., 1964, and others).
Dolphins that are kept in captivity most frequently
suffer from skin diseases, which cause real distress and lad,
as a rule, to a lethal outcome (Russel, 1966). In addition
there have been found in them: erysipelatous inflammations,
stomach ulcers, jaundice, insults and infarcts of the myocar-
dium, diabetes, scurvy, inflammation of the lungs, and numerous
external and cavitary parasites (Ridgeway, 1955). One of the
most serious problems is the swallowing by the dolphins of
extraneous objects that fall into the Pools (balls, gloves,
toys). In recent years (Ridgeway, 1955, 1967; Nagel et al.,
1964) there have been elaborated methods for the catherization
and venous puncture of dolphins, of anaesthesia for serious
surgical oPerations, and standards for the application of
antibiotics, tranquilizers, drus and vitamins. In the Tobo
Aquarium (JaPan) there has been established a special hospital
for diseased fishes and dolphins, where they undergo a course

of treatment (Russel, 1.966 ). It has been established that
the medicines which help )eople also help dolphins, in par-
ticular tablets that aid the digestion of food and vitamins,
as a suwlenient to the food ration.
In the work with the dol1?hins we carry out a medico-
biological control on the state of their health (Figure 3).
The importance of this work is dictated among other factors
by the fact that commercial hunting of dolphins in the USSR
is banned and every catch of experimental animals is turned 292
into a complex problem, and therefore we consider that it is
more important to preserve the animals than to catch new ones.
Serving as objective symptoms that are indicative of
the state of health of the animals are : the rate of swimming
and diving; intercourse with other animals; degree of acti-
vity at feeding; character of respiration. A slowing in the
rate of swimialng, long periods spent on the surface of the
water and a reduction in activity at feeding are all patho-
logical symptoms, and in the presence of these the animal
should be examined.
The most important disease symptoms of the animal are:
A disturbance in the rhythm and intensity of the respiratory
act - a prolonged res-piratory act with a sluggish exhalation-
inhalation and the appearance of foaming from the blowhole at
the moment of exhalation; assistance given to any animal on
the part of the others; a yellow or grey colour of the faecal
niasses; a categorical refusal of food. In such cases it is
necessary to immediately examine the dolphin.
lv:ucilagenous discharges from the blowhole (of the spu-
tum type) and a putrescent odour of the expired air are

472
NC. 3. Chcpciutoft àle,locmoTp. CII1TI1e KaplutorpammiA c (1)1111a-eao6nuli
Figure 3. Routine medical examination. Obtaining a cardio-
gram from a white-sided doluhin.
indicative of a flooding of the' respiratory tracts with water
and the presence of inflammatory processes in the lungs. In
this case it is necessary to isolate the diseased animal from
the general group, placing it in a laboratory aquarium for 4 -
5 - days and treating it with antibiotics (penicillin or bicillin*
in the usual doses per 1 kg weight).
Liquid faeces of a yellow or grey colour are indicative
of diseases of the gastro-intestinal tract. In this case it
is necessary that the animal be fed only fresh fish, with which
there may be administered tablets of laevomycetin. Besides
this, use is made of antibiotics (biomycin, synthomycin*1 and 293
vitamins of the "B" group (in the usual doses per 1 kg body
weight). When there is a lack of aupetite and deterioration
of the state, it is advisable to isolate the dolphin for a pro-
longed period from the remaining stocke to avoid the spread of
Translator's notes. * bitsillin or bicillin benzathine penicillin G. ** synthomycin - synthetic streptomycin

473
the infection among the remaining animals, and to carry out
an intensive penicillin therapy.
In the case of the presence on some part of the body
or other of fungal diseases, ulcerations, bedsores and other
skin diseases, there were employed intramuscular injections
of antibiotics and administrations of polyvitamins in the food.
Simultaneously with this there was carried out a treatment of
the infected portions of skin with antiseptic solutions (iodine,
alcohol, potassium permanganate), with a subsequent drying in
air and smearing of the affected sites with sPecial dermato-
logical ointments. A depressive state of the animals may be
revealed only vith systematic observations The superficial
symptoms are: aggressiveness, swimming separately from the
group, loss of appetite and increased caution. All of these
may be evoked by a depressive state, if it proves on examina-
tion that other diseases are absent. In these cases aminazine
was employed.
The methods of medical control which were used in our
work proved themselves to be fully justified. For illustration
we present two examples when the treatment was effective and
operative.
en the 24th of June 1969 a young female bottlenose dol-
phin, which was living its second year in captivity, refused
its food. On the following two days she continued to ignore
the food, swam apart from the group on the surface of the
water, and did not react at all to man and the dolphins. She
was given intramuscular injections of bicillin, 600 thousand
units, and a comolex of the vitamins B 1 , 1-36 . and B121 in 2m1

v
v
aliquots. After 3 - la, hours she becan to move-mors actively
about the pool but continued to refuse the food. Over the
course of the next few c3.ays she made attempts to eat fish,
but without success. On June 30th a thourough medical exami-
nation was made, since the suspicion arose that the dolphin
had swallowed a small rubber ball. In 6 days the animal had
grown considerably thinner, the back was sunk in, the mucôsa
of the mouth cavity was inflazned and a distinct putrescent
odour emanated from this. A strut was placed into the mouth
and an attemnt was made to introduce some machine lubr.icating
oil into the stomach through a rubber tube. This was not
successful. However an inclination to vomiting was noted.
The cavity of the mouth was treated with penicillin and a
solution of potassium permanganate, Intramuscular injections
were macle of bicillin - 1 million 800 thousand units, and a
complex of vitamins (i3i and Bi2 ) in !I ml aliquots. Already
on the following day there were noted attempts, though it is
true that these were still fairly sluggish, by the dolphin to
play with a black rubber ring and with the other dolphins.
On July 3rd repeated injections were made of bicillin
(1 million 200 thousand units) and a complex of vitamins (B1
and B12 ) in 11, nil aliquots, while during the examination the
mouth cavity was aaain irrigated, although the state of the
mucosa was already satisfactory. Towards the evening of this
day the bottlenose dolphin fairly readily and energeticall.y
played with a ball and with rubber rings. Her movements be-
came more confident, energetic and pronounced. She readily
began to eat fish. She began to swim in ai.)air with a young

4.75
bottlenose dolphin, reacting in a lively fashion to the ap-
pearance of a man and making contact with him. During the
course of the following week she was given 3)olyvitamins and
oxytetracycline. This animal soon recovered completely.
In a white-sided dolDhin, which had been caught in May
of 1969, during a routine change on July 16th there was noted
typical loss of coordination of movement, which was expressed
by a rolline over onto the side and bumping into the walls of
the oen. After several convulsive movements of the head she
suddenly sank. We were able to quickly raise her from the
bottom, but she could not maintain herself afloat. It was
necessary to transfer her into a laboratory aquarium and to
secure her on a hammock, so that she would not choke. She was
given an intramuscular injection of bicillin (1 million 200
thousand units) and mellipromin (2m1), after which the water
in the aquarium was disinfected with a solution of potassium
permanganate, while the skin was treated with alcohol and
iodine solutions. The respiration of the animal had a rate
of 31 times in 5 minutes, the heart-beat frequency was 110
beats per minute. On July 19th we repeated the injection of
bicillin (600 thousand units) and of the vitamins B6 and B12
(4 ml of each). The frequency of respiration decreased con-
siderably, to 12 times in 5 minutes. The pulse - to 90 beats.
The irrigation was rel)eated with the disinfectant solutions
and the infected portions of the skin were smeared with un-
decine ointment.
On July 20th, after she had been kept in the laboratory
aquarium for 87 hours without food (refusing the food), she

4.7 6
was let out into the pool 1 here she began to swim actively,
and after 5 minutes to take scad greedily from the hand. Over
the course of a week we continued to give oxytetracycline and
polyvitamins with the food. This white-sided dolphin is still
alive at the present time.
The catching, transportation and relocations, and the
medical servicing during the course of long-term holding in
captivity are important constituent parts, on each of which
is dePendent the overall success of the work with the dolphins.
The tested methods for carrying out the catch and
transportation, taking into account the available practical
possibilities under the conditions in the Black Sea basin,
have shown that they may be successfully utilized in future
work.
The transoortation of the captured animals on a ref-
rigerator ship in containers with running sea water, with the
animais being secured in hammocks, was found to be the most
favourable form of transportation of dolphins by sea. However,
the extension of work with dolphins sets investigators the
task of elaborating and employing method of transportation on
dry land, which is a more complex problem.
The apPlication of therapeutic measures, utilized by
man, deserves by all means to be maintained and to be intro-
duced even more boldly into the practice of the routine work
with dolphins. However, in soite of the assistance rendered,
the mortality among dolphins during long-term holding in cap-
tivity remains high. Therefore there are needed further thor-
ough studies on the causes of the mortality of the animals and
on the elimination of these.

47?
Gur work does not pretend to provide a complete eluci-
dation of the problems that have been nai.sed. One of its aims
was to prepare the soil for the further development of the
above listed problems, connected with the long-term holding
of dolphins in caotivity.
JIIdTEPt1TYPfl
^ IJen^tiueu^t B. M. tt Op. 3ara;uta ot;cana. n4.. «,^io:to;last raap;ltts3», 1JG5.
2 Knetïxettôcp, C. E. u Op. BeayNa. M., 1964.
3 rlttr,.^tt %I. Lle:to1nex il r^c:n,c^slt1. \^I., «\Itac:tt,», 1965.
,lj• j'aii K. rloacr Tpc\ xtrrots. «>lntcp3n;ar, N, 87. 1963.
5 Cor;naoa B. E. u iJp.Ont,n• nccrte;totlaun;l ocoGewtuc^eü naat:<uulsl Aenli^snttols
L' rltAPos:asta^C. "fe3. Aotc:l. '1-ro licecoloSnoro cotSCnlaliltn Ilo u3)•9ctwto ntohclaix
DtA holtnl'a1o11UfX. Kaan1111111'1>F1;1, 1969.
.^ fjTo.suatttt A. I. Ilcrop,lsl caenoro saluaaoTa. 1\9.. «1Ia^r,a», 1t^G;s.
7 Andersen S., Dziedwic /1 Behavior patterns of cantiv^ Ilarbor porpoise(Paena ll,rocacna), 13u11. Inst. Occano..f^r. Monaco, vol. 63, no 116. 1961,
hro:on D. 1I. »Flic Hcalth problcros of \Valrus colves and retnarl.s on theirgeneral progress in captivity. cln[ernal. 7oo learboolc», 4. 1962.
9 Bro;crt 17. 11., .+Iiclrttttrè R.N. lleallh prublenls of caplive dolphins ,Incl
scals. 4. Amer. Velerin. Mcd. Assoc». vol. 137, no (à, 1960.
ti j.(Y3rowtt D. H.,.i\'orris K. S. Observations of captive and wild Cetacea.
«J. b•1aminal», vol. 37, no 3, 1956.Caldwell D., Brown D., Calcic;:cil M. ]nicr;cncric bchavior by a
iive Pacific Pilot Whalc. «Los i1n;;^elcs Conntv Alns,» no 70. 19G3.
M Gast:Lt D. E.i)olphil, ai 11tc ;^Iapier t^I lri ulancl. «\c\r• "l.ealancl Sei. lic^•.»,
lrurn^l•'M , no 2. 196G.1 D«dok vcrtt Ilcc! W. N. Observations in flizllit re-tctionti of Tursiops
S lil^oilcr ^.u^^^^ ,• ^l,tus (M) with sUllic sUf;'esttonS on 'tli5til planning. «Z.
no 5, 1966.1Il• Keliog Il?. H., Rice C. E. Vistlal Discrimination and Probl^m Solvinl; in aBottlenose Dolphin. «1Vhales, Dolph. a Porp.». 1966.15 Kritzlcr N:
The Pilot \V hale al c^•IarinclancL «1tlt. Ilislor}'», GG, tlo 7, •191J...
jb Nagel E. L., Morgane P. J., :1icl'arland N. L. Anesthesia .for the I3ottlenosellolphin (Tttrsiops irnncatus). «Scicnce», vol. 146, no 3G51. 1961.Nortis N. S. Practical problems. ^\^'halcs, Dolshins <t. Porpoises». 1966.
] Ridgujny S.H. Miclical care of marine Inammttls. «J. Amer. Veterin.:lssos»,
147, no 10, 196,3,19Riilgtonll S. Il, t1lcCorntic J.
G. Ancsthctization of porpoises for major sur-
Hospital. «\emr vol.Scicntisi», 21, no. 6. 19G6.gnvRtsscl>'JI Jal:an's I0iGsil 1
967.
21 Scltevill W. E., Lawrence D. rood-finriin; by a captive porpoise (Tursiopstruncattts). r.I3reviora», no. 63, 1966.22Slijper E. L. l^thales. «I-Itttcilinsun of I-ondon». 1962.
REF FREIN iCES
1. Bel'kovich V. M. et al., A riddle of the ocean. Il+,oscow,
"I^_olodaya. Ûvar• diya" ("The Young Guard"), 1065-
2. !,,Ieinenberg S. E. et al.,
("Science" ), 1965 .
The beluga. TY:oscow, "r'iauka"

52 8
The fairly constant maintenance of the holes by the
ringed seals (Chapskii, 1940) suggested to us the possibility
of counting the number of seals and of their holes as these
were encountered; therefore, in conducting the aerial survey,
we recorded encounters with both seals and their holes, situ-
ated both in direct proximity to a lying animal and also with
no animal present.
It is still difficult to make a completely definitive
judgement as to the number of holes that are made and main-
tained by different individuals of the ringed seal, though
in the opinion of Greve (1896) each ringed seal in the Baltic
Sea has from 5 to 8 breathing holes (according to Ropelewski,
1952).
With respect to the holes of the grey seal Holm (1921)
writes; "There occur cases, when in severe freezing weather
the exit holes of the seals not infrequently freeze over and
many animals are forced to leave these and accumulate at one
hole in numbers of 10 - 15 individuals".
As a result of the studies, conducted on the landfast
ice in the Gulf of Finland in Earch of 1970, by one of the
authors there was thoroughly investigated the region between
Malyi and Bol'shoi Tyuters Islands. In the investigated area
there were found 17 seals and 17 holes of various diameter
(from 3 ).l to 40 cm), visible from above. The ratio of their
upper diameter (on the outer surface of the ice) to the lower
diameter (the edge of the ice that is under the water) com-
prised from 0.57 to 0.63. it is Possible that the size of the
hole is directly deoendent on the size of the animal. The holes
were situated at a distance of from 1 - 1.5 to 3 km from one
another.

529
o
.e4
Besides the 17 separately situated holes, there were
found by us in one place three holes of various sizes situated
next to one another (Figure 1).
Taking into consideration the smaller diameter of two
of these on their upper edge (16 x 15 and 20 x 17 cm), in com-
oarison with the large hole which was found beside these (37 x
40 cm), the thickness of the ice in the place where these holes
were located (about 30 cm) and the tracks of small sized flip-
pers around them (around the large hole there were tracks of
an adult seal), we assume that these two holes were used by
a young animal (possibly born in this same or Ln the previous
year). Such groupines of holes, of 2, 3 or even 4 together,
were observed by us on more than one occasion also from the
airolane.
Further studies of the characteristic features of the
distribution of the seals, and of the disposition and arrange-
ment of their holes will allow us to explain this phenomenon
more specifically.
According to the observations of V. A. Zhelgov every
ringed seal on the landfast ice makes a hole for coming out
onto the surface in the freezim; over patches and cracks
("smorozi") between the ice floes that have been drifting from
the middle of the winter. Later the thickness of the ice on
these "smorozi" increases but it always is less than the thick-
ness of the main frozen together ice floes. In the Gulf of
Finland this difference comprised from 10 to 20 cm. The holes
which are maintained by the ringed seal in the snow drifts
amidst the ice hummocks are also located on "smorozi". As the
thickness of the ice on the "smorozi" increases, the ringed

530
Pile. I. Patnoao)Renue pa3.-uvj- Iforo pa3mepa 13 ()Rawl mec-re. Map .r.
(baueKiiii (Pow A. F. &ammo u B. .4. }Keziwea '
Figure 1. The disposition of holes of various size in ohe
location. March. Gulf of Finland.
Photograph by A. G. Esipenko and V. A. Zheglov.
seal constantly maintains the hole, for which even in the
middle of 1arch it must break through, once or twice a day,
the crust of ice which forms on it (up to 2 cm in a night).
In the course of the investigations on the ice it was
noted that a ringed seal which had been frightened away did
not appear at its hole until the next day. Cbservations in
the regions closest to the hole (in a radius of 3 - 4 km)
showed that the ringed seal did not appear within this distance
on the surface of the ice, even though the weather conditions
for its emer.cecrIce onto the ice were favourable (clear, light
wind, daytime temoerature of +1 to - 2°c),
It is possible that during this period it either made
use of its hiding places (in the hummocks and snow drifts

5 31
with an access hole from the water) or it moved away from this
hole to a distance greater than 3 - 4 km.
Ln the basis of the information presented we assume
that on the landfast ice each ringed seal, up to a certain time
(until the holes are broken up during the break-up of the ice
or due to other factors), makes use of one hole for emerging
onto the surface of the ice and several hiding places with an
access to the water, situated in the ice hummocks, where the
ringed seal lies during bad weather, utilizes these for re-
Plenishing its air reserves during feeding and gives birth to
its younF. The hole which the ringed seal uses for emergence
onto the surface of the ice is clearly visible from an air-
plane. The holes that are located in the hiding places cannot
be counted from an airplane.
During the aerial survey we noted that the break-up
of the ice occurs mainly along the sites of the "smorozi" and
hummockinF. This has also been noted by hydrometeorologists
(Lethodological instructions, 1959). In such cases, both the
holes which are concealed in the hiding places as well as the
open holes, used by the ringed seal for emergence onto the sur-
face of the ice during its break-up, in our opinion, are des-
troyed in anproximately the same ratio. The ringed seals,
whose holes have been destroyed, are apparently forced to make
use of the open patches of water or the holes of their "neigh-
bours".
Taking into consideration the aoproximate periods of
whelpine and moulting of the ringed seal and of the grey seal,
the count of the seals in the Gulfs of Finland and Riga and

532
on Lake Ladoga was conducted in two stages t first - on 1arch
9 - 11, and second - on April 3 - 7. On the basis of informa-
tion obtained from questioning the local sealers and our own
assumutions on the possible distribution of the seals in the
regions under study, the transects for the observation flight
in 1\.arch were set up ahead of time, while in Korn these were
set UP on the basis of the observations made during the first
flights and the direct observations made on the ice in March.
On detection of a region with a more concentrated dis-
tribution of the seals there were "set up" counting areas (see
the schematic maos of the aerial survey). The region was in-
tersected by several counting lines. This permitted us to
estimate more reliably the density of the hauling-out of the
seals in the given region and to determine the borders of their
hauling-out patches.
The flights were made on LI-2 and IL-14 airplanes at
an altitude of from 100 to 150 m, at an average speed of 200
km per hour, from 97.10 a.m. to 3-6 n.m.
The observations were carried out from the right and
left side from the blisters (transparent hemispheres, which
were mounted in place of the portholes in the navigator's com-
partment), which Permitted the observer to see not only to the
side but also to the front and underneath the airplane.
The data on the state of the ice cover was periodical-
ly recorded in the log-book using the generally accepted me-
thod for charting ice mans in accordance with the methodologi-
cal instructions for aerial observations on ice conditions at
sea.

533
Special attention was paid to determining the degree
of hummocking, the snow cover and the closeness of the ice.
Generally on the state of these factors depended the distri-
bution and concentration of the seals and their holes, the
numbers of which and the characteristics of their disPosition
were carefully recorded in the flight logs according to the
time of encounter.
According to our observations, it is best to make thé
simultaneous count of the seals and the holes t from one side,
over a counting strip not more than 150 m in width, although
the seals may be discerned also from the side at a distance
of 300 - 350 m from the airplane. At a greater distance from
the airplane the holes are poorly discernable and it is Pos-
sible to miss them. It is necessary to mention also the par-
ticularly cautious behaviour of the animals themselves in the
Baltic. In the Gulfs of Finland and Riga the ringed seals
escape into their holes 100 - 150 m ahead of the approach of
an LI-2 airplane to it and 200 - 250 m ahead of an IL-14 air-
plane, which forced the observer to look ahead, along the
flight path of the airPlane, from time to time.
The Ladoga seal behaves more calmly in the presence
of the ID-14 airplane, and escapes from it into the water at
a distance of 75-100 m.
It is possible that such differences In the behaviour
of the Ladoga seal and the ringed seal of the Baltic during
the sPring period depend on their physiological state or on
sonie other factors connected with living in bodies of water
with different hydroloe- ical characteristics.

5311-
Ne. 2. llpuuaïl, o6paa0uannLlft 113 upmlocnurn.ac,ta. Anpc:n,. Pnxcit;nit 3:1aua. ( CTpc.Rxa>lu yxa-3a11L1 1I)'1101 K0.7L98T6IX 11Cpn. Ot1nC?ff Iia 1111X
nca:nT ncpna)Asporjioro A. K. 111.voxoru:a
"?ilgure 2. La_nffast ice, formed of ice which has drifted in.
t^-pril. Gulf of Riga. (The arrows indicate holes
of the ringed seal. 13eside one of these lies a.
ses.l ).
Aerial pho-to^ra?)h by A. :•". Shlyundin.
a - hole of sealb - rinEred seal and hole
From the air the ringed seal is similar in form to a
comma (2ed.oseev, 1966). The holes of the seals in the Baltic
are very even dark c i rcles situated beside hummocks. From a
heit:ht of 9.00 - 1.J0 in around the holes there can be clearly
seen the snow that has been crushed down by the seals. The
ed.,es of the holes are not fro..f._: o^c^er., as in the holes of the
haryp seals (Figure 2).
It is almost impossible to de-termine the size of ani-
nials lying singly from an airplane. In April, when groups of
'inirl?a1s were found l^T^.n^ in directdirect ^^ro._ir:i-ty to one another,
t\re observed ?both small as well as lar^,-e seals. it is fairly
cai.ffi ct?l.t to discern awh.itet:oat from an airplane. ',.e no-ted
these only s..oains t ice of a background colour darker than them.

535
The whitecoats are distinzuishable from the adult animals by
their colour and smaller dimensions (about half the length of
the body beside a lying adult animal).
In :arch and J+.nril in the Gulfs of the i3altic Sea we •
noted 2 and 3 whitecoats.
The ,grey seal was visible as a dark-grey comma against 329
the background of the white and grey-white ice. It has a much
lighter colour and is larger than the ringed seal, and lies
more frequently beside cracks. It scarcely reacts at all to
the airplane, and if it does escape into the water, which oc-
curred only in two cases, it does this only right under the
airUane.
• The tentative count of the numbers of the rin,g;ed seal
and the grey seal in the regions under investigation was con-
ducted accordin.q: to the following schemes
1. Drawing the counting transect on a large-scale map
on the basis of the times and orientation points.
2. Transferring from the flight log onto the schema-
tic mao of the transect the information on the sites of finding
of the seals and their holes, and, as changes occurred, the
ice conditions.
3. Gutlining the contours of uniform ice conditions
(according to the types of ice formation), in which we utilized
our own observations and the data from the ice survey by the
hydro-meteorological service, which conducted its work simul-
taneously to us.
4. Calculating the areas of similar ice conditions.
5. Calculating the area of the counting strips for
each type of ice formation (the width of the total counting

5 36
strip from both sides of the airolane was 300 m), determining
the density of the distribution of the seals and their holes
on the counting strips and in the areas with more concentrated
haulinE-out Patches (see the schematic maps of the surveys and
the counting areas on these) for each SQ. km . for each cate-
gory of ice formation.
6, CalculatinP: the approximate numbers of seals and
holes over the area for each type of ice formation (separately
for each Gulf and for Lake Ladoga) by the method of extrapola-
tion of the obtained densities and their distribution.
RESULTS OF SURVEY
As a result of the data obtained by us from the aero-
visual observations, the ice types accepted by other investi-
gators - white, grey-white, grey etc. (Popov, 1966; Shustov,
1969, and other authors), did not permit us to characterize
the conditions and characteristics of the distribution of the
rinp:ed seal and the'exey seal in the Gulfs of the Baltic Sea
and on Lake Ladoga.
For the characterization of the habitat conditions and
the distribution of the seals and their holes we distinguished
several types of ice formations, which were characterized by
a particular density of the distribution. Under the concept
of "density" we imply the average number of seals and of their
holes Per one square kilometer.
1 9 Landfast ice of local origin is
characterized by_a thick ice cover with a thickness of the ice
of more than 30 - 40 cm (from our ice observations), with a

1
537
weak hummocking (up to strength 1), formed by a certain shif-
ting of the ice during the winter; its snow 'cover in the
middle of the winter has an index of up to 3. The grey seal
was not found by us in this type of ice formation; the rineed
seal and its holes are occasionally encou.ntered in hummocks
or beside these .
No seals or their holes were found at a.11 either from
the (airplane or during the observations on the ice in the
Gulfs of the Baltic Sea on landfast ice of local oriein which •
overlies depths of up to 10 - 15 m (see schematic maps of
a.erial survey ). At present it is difficult to explain this.
It is possible that this was due to the absence of the basic
food objects of the seals at small dePths during the winter
period or by the small number of hummoclçs, which are needec3.
'oy the seals for the construction of their hiding places.
Therefore we did not take these areas into• consideration in
calculating the areas of landfast ice of local origin.
2. Landfast ice, formed by the freezing
together of' white ice t.hat has drifted in e
with a degree of hummocking of strength 2 - Li, in our opinion,
i.s the most favourable for the construction of the hiding
places and for the 'maintenance of the holes by the ringed seal
(Figure 3).
At the beginning of January - February 1 during the
hummocking and free 'zing together of the 3_0e le ,thich is drifting
in the ,lfs, the ringed seal, in all probability, is distri-
buted alorrY the o,)e.n oaLtches of water Etmong the ice floes.
Subsequently, when these freeze tor;ether and ice is formed on
the open :.)atches, the seal here makes and mainta.ins its holes,
and in the hummocks - its hidinp_..; -31a.c.:es

- 5 38 • • I
•
. Pile. 3. Tonotinoe mecTo mytioK 1<0.711,‘11iTal
nepnbi an npituae 113 np11110C11010 J11,)1,8. MapT. (1)liocKaii 3a-
. .11113 nepeRuem niane 1:031b,IiITCH"1 nepubl). I'm.° B. A. )Keennea
5. Fir-Jure Typical site for the construction of holes by the
ringed seal on landfast ice derived from drift ice.
r;ulf of Finland. (ln the foreround is a
hole of the ringed seal).
Ehoteraph by V. A. Zheluov
3. Breaking-up landfast ice of frozen
together drift ice is formed mainly with the onset
of warm weather.
In the Gulf of Riga in 1970. this landfast ice began
to be formed from the beginning of Larch; in the Gulf of
2inland and on Lake Ladoga - from the beginning of April
(the closeness of the ice had an index of 8 - 10).
In Larch in the Gulf of Riga.the density of the distri- 331
bution of the seals on the broken up landfast ice (with an
- 9 index of closeness of the ice) attained a level of 0.43
head and the holes had a density of 0.41; in April these
values were only 0.01 head and 0.02 holes. This is probably
connected with the breaking up of the holes and the distribution
of the animals alonu the cracks and on ice formations of other tr e s.

539
In the Gulf of -7'inland on the broken up landfast ice
(closeness of the ice with an index of about 10) in April the
density of the seals was0.15 head, while the density of the
holes was 0. 3 7.
The smaller density of the holesin the Gulf of Riga,
in comparison with the Gulf of loinland, was obviously related
to the greater break-un of the ice, and therefore of the holes,
in this Gulf.
In the Gulf of Finland the opening up of the landfast
ice and its subsequent break-up occurs later than in the Gulf
of Riga, by 10 - 15 days (Atlas of ice conditions in the Bal-
tic Sea, 1960).
4, Ice fields of white and grey-white
ice are formed of frozen together driftine ice floes in
the central and western Part of the Gulf of Finland at the end
of January - beginning of i;'ebruary, and in the middle of the
Gulf of Riga in the middle to the end of February (Atlas of
ice conditions in the Baltic Sea, 1960).
It was noted by us that in . April the ringed seal and
its holes were encountered more frequently (density up to 0.17
head and 0.17 holes) on the ice formations of this type in the
Gulf of Finland than on the landfast ice of local origin.
We consider that the more broken up ice of this type
in the Gulf of Riga also exPlains the smaller density of the
seals and of their holes on this ice (correspondingly 0.02
and 0.13).
In karch Anril, as the ice fields break u), there
occurs a translocation of the seals onto firmer ice, where they

51a-0
distribute themselves along the cracks and oPen :patches of
water, and someti.mes also utilize the holes of the "old resi-
derits". Thus, according to our aerial observa-tions, neither
seals nor holes were found in April on the broken white and
grey-white ice in the Gulf of r'inland and the Gulf of Riga.
At the same time on the ice fields and broken up
landfast ice beside one hole or crack there lay 2 -- 5 seals,
which was noted by us on more than one occasion in April in
the Gulfs of Riga and Finland.
This is indicative of their concentration at the be-
^,.-innin` of A-pril on the firmer ice. In the Gulf of Riga the
fragmentation of the ice occurs in the direction of from south
to north of the -gulf, while in the Gulf of Finland this occurs
from the west towards the north and east (Atlas of ice condi-
tions in the ^,̂ alti.c Sea, 1960). In step with the fragmentation
of the ice the redistribution of the seals apparently occurs
in the same direction.
5. Broke,n-up white and grey-white i.ce,
in our opinion, is unfavourable, in I::arch and in April, for
the hauling out of the ringed seal and the Grey seal. The
areas with this type of ice in the Gulfs of the Baltic Sea
are small (see schematic maps of aerial survey).
In T"arch on ice formations of this type in the region 332
of the Irbenst.i? Strait we noted a relatively high density of
ringed seals, 0.33 head. We explain this as a result of the
dri ftinz of the breakinE--up ice fields through the Irbenskii
^,`),trait into the ouen sea, where the ring,ecl seal does not .)ass
with the ice, but as the broken u-p ice drifts out to the west
the seals concentrate in the :trbenskii Strait.

541
Pitc. 4. Cxema artuaytteTitoro mapulpyra ii aeaonoft o6cration1at I(1)1111éKOM 3a.ritute (mapT 1970 r.)
*. .
Figure 4. Schematic map of the aerial survey transect and ice conditions in the Gulf of Finland (P;arch,'1970).
A. - Counting area.
In Aril, when the edge of the broken-up landfast ice
and of the ice fields moves to the north, and the more broken-
uo grey-white ice is carried out into the open sea, the ringed
seal is no longer encountered here.
6. Light and very light floe grey
ice (area of floes from 40 to 200 sq. m) is formed at the
edge of the ice fields and as it were girdles them with a 5 -
10 km belt: in the Gulf of l'aga from the south, and Ln the
Gulf of Finland from the south-west (see schematic ma) of
aerial survey). The ar:eas of this are small, and in the sprin
it is rapidly broen up. The seals clearly avoid such ice and
were not found by us here.

0 A
®U
00
FED
5 9
®b
EE
it
12
Yc.ionul,te o6o3ua4enlln tc piic. 4-8
1. I]pün<lü ^secTUOro nponc^o.xa^ulsst2. Ilpuuait 113 npuiloalorn cmcp3uicroc5l :n,aa
3. B3.noataunt,tïi npnnai: 113 npniloCnoro ca+epsiue-
rocsi rli>^a4. ^Ielütui,ic iloast Ge-'()ro ti cepo 6e loro .at 1:i5. Iàntl,tïi 6c.1litü n ccpo Geni,li'i 10a6. l^p)'nuo- il nlear.eGnrt+ïi cepi,+ü tc [7. Bo;ta8. I pf111fII[6i 'Cn(IU6 ICdUCLI\ o61)a3oüfln1lÎt il CLIeT-
u[,is naoulaAeït
9. blilpnlp)'T 8nll,lY'IeT110. 13erpeaelnlasl na ',saplupçre xo ibllaT 1 isi nepna
il. IicTpet+euui,lii lui i,\taplnpvTe cep1,tü Tlo.leni,12. L'crpevellnan na asapulp^'Te :nnlca
6",
8 -LT:?`:cnd. to .E''iy^hres I• --
La.nd_C'ast ice of local ori"in.Landfast ice of drifted-in frozen to^,ether ice.Broken-up landfast ice of frozen together drift ice.Ice -.fields of white and Erey-white ice.3rol_en-uu white and gr ey-whi.te ice.I:iEl>.t and very 11.21->.t floe Erey ice.Water.,o_rd.ers of the types of ice formations and counting areas.Transect of aerial sur;Tey. vRinged seal encountered along transect.Grey seal encountered along transect.Hole encountered along transect.
.Un ice formations of other types (ice rind, pancake
ice) the seals were also not found. This is connected with
the fra^^;ilit,y and remoteness of such ice from the ice fields.
The distributions of the listed ice formations, the
encounters with seals and holes alon- the transects, and the
re4dons of their concentrations (counting areas) in the

5 4.3
Pm'. 5. Cxema aDilaytteTnoro maptupyra 06CTa110131:11 Cinalcizom 3amme (aripeab 1970 r.)
Figure 5. Schematic map of the aerial survey transect and
ice conditions in the Gulf of Finland (April, 1970).
investigated bodies of water are presented in the schematic
maps of the aerial survey (Figures 4 - 8).
DYNAMICS OF THE FREQUENCY OF OCCURRENCE OF THE SEALS AND
OF THEIR HOLES IN THE GULF OF FINLAND
In comparing the data on the density of the distribu-
tion of the encountered seals and their holes on the various
types of ice formations, we found:
In March the maximal density of hauling out of the
rinued seal occurred on the landfast ice of drifted-in ice
(up to 0.62 head). A smaller density occurred on the ice
fields of white ice, 0.17 head. The densities of the distri-
bution of the holes were correspondingly expressed as 0.4 and

544
A.- Counting area
Pue. 6. Cxema anuavtieruoro mapuipyra u :Icaonoff (De clauouu B PiGi■CHOM malice (maim. 19/0 r.)
Figure 6. SchematiC mao of the aerial Survey transect and
ice conditions in the Gulf of Riga (iiiarch, 1970).
0.17 holes. There was no broken-uo landfast ice in March.
In summing up, from the calculations, in March there were
about 5000 seals and 4500 holes.
Durine the course of the oeriod between the surveys
profound changes occurred over the whole ice area in the Gulf.
In April there was a decrease in the size of the areas
of nf continuous landfast ice of frozen together ice; these
were replaced by broken-up landfast ice and partially broken-up

A.- Counting area
54.5
•
Pnc. 7. èxema annaytieTnoro-mapniprra n .Icaonort. o6cTa- 110111C11 n PII)liCKOM 3asnme (anpc.lib 1970 r.)
Figure 7. SchematiC map of the aerial survey transect and ice conditions in the Gulf of Riga (April, 1970).
ice fields. The average density . of hauling-out of the ringed
seals on the broken-up landfast ice of frozen together drift
ice in April was 0.15 head, while the density of the holes was
0.37. The ringed seal was not found on the ice fields (the
size of the floes here had decreased to 0.3 - 0.7 sq. km .),
where there were still occasionally found traces, i.e. the holes,
of their prior habitation.

546
The absence of seals on this type of ice formation
is explained, apparently, by their migration to other regions.
According to our calculations, in April in the Gulf
of Finland there were present on the ice about 900 ringed
seals and about 3000 holes.
It is generally known that the ringed seal does not 336
carry out distant migrations, with the exception of slight
seasonal migrations, related to shifts in the edge of the ice
(Bergman, 1958; Chapskii, 1963), and therefore we consider
that the number of ringed seals calculated on the basis of the
encounters in A'uril does not reflect their actual numbers.
The decrease in the frequency of encounters with the
ringed seals on all of the types of ice formations in j1pril
is explained by us as being due to their redistribution onto 337
the edges of the broken up landfast ice, where they form denser
moulting patches, for the detection of which a large counting
area is necessary. Three such patches of ringed seals were
found by us in April in the region of the Islands of ï'ykh ^ya--
Ukhti, Vigrund and 11'ayak. The seals lay here in groups of
2 5 individuals beside one hole or beside an open patch of
water in direct proximity to one another. Groups of ringed
seals were situated at a distance of 0.5 -- 1.5 km from one
another.
The decrease in the total number of holes encountered
in April (according to our calculations there were up to 3000
of these) is related, in our opinion, to the widening of the
"smorozi" and the fragmentation of the hummocked sections on
the ice fields and broken-up landfast ice.
338

5 11.7
o
o Pue. 8. Cxenta annaynernoro maptupyra u sieRonoii o6-
cranonizu n ioclot o3epe (anpe,ib 1970 1.) _ . . . •
2igure 8. Schematic map of the aerial survey transect and
ice conditions on Lake Ladoga (April, 1970).
The data obtained on the distribution of the seals,
on the basis of the calculations for the entire area covered
by ice in the Gulf of Finland, permitted us to make a tenta-
tive estimate of their approximate numbers and to determine
the regions of their spring concentration. It was found that
the main part of the population of the ringed seal in the Gulf
of Finland at this time is concentrated in the eastern part
of the Gulf between the islands of Vaindlo and Seyskari.

548
IMF
o
The approximate number of the ringed seals, present
on the ice, is reckoned by us, on the basis of the encounters
in F.arch, at about 5000 head, with about 4500 holes. The en-
counter with one ig-ey seal in April was a natural occurrence,
since the area investig.ated at this time was 1.6 times lare_er
than in March, and these seals are very rarely encountered on
the ice during this period.
We consider that the number of ringed seals and of
their holes which was calculated by us on the basis of the
density is underestimated. In our case, during the survey
all of the record entries were made by the individual conduc-
ting the count, who had to interrupt his observations to do
this. It has been established experimentally by American in-
vestigators that the error in counting from an airplane, even.
in the case of ungulates on tundra, leads to an underestimate
of the results by up to 20;0 on average. It is therefore re-
commended that those making the count should transmit their
observations, without interrupting these, to a co-worker who
makes all of the record entries (Chervonnyi, 1969).
Takinginto consideration a similar miscalculation in
our work and the approximate percentage of occurrence of the
seals in the water (about 30,%0) duping the daytime in March,
it may be assumed that in the Gulf of Finland during this
period there were oresent about 8 thousand head of the ringed
seal. The calculated number of holes, on the basis of the
findings in b_arch (with a correction for omissions of 20A),
is about 5.3 thousand.

54 9
THE-GULF OF RIGA
The overall character of the distribution of the seals
and their holes according to the types of ice formation is
similar to their distribution in the Gulf of Finland (see
Figure 7).
The number of ringed seals calculated by us on the
basis of the frequency of their encounters'in April is also
smaller (in March - 3.1 thousand head, and in April only 0.12
thousand head). It is probable that the ringed seal in the
Gulf of Riga in April also quits the moving ice and concen-
trates on the edges of the firmer ice (the stationary and
broken-up landfast ice).
In April, in connection with the breaking-up of the
"smorozi" and hummocks, the number of holes decreased consi.L.
derably (according to the calculations there were 3.9 thousand
of these in March, while in April there were only 1.5 thousand
holes).
The number Of ringed seals obtained on the basis of
its encounters in April undoubtedly does not reflect the actual
stocks (as is the case in the Gulf of Finland), and therefore
we calculate the tentative Population numbers of the ringed
seal on the basis of its encounters in March.
In March and April on the breking-up landfast ice in
the Gulf of Riga there were encountered individual grey seals.
In April at the edEe of the ice fields of white and grey-white
ice (with a closeness index of 7-8, and a degree of hummocking
of 2-3) there was found by us a hauline-out patch of grey seals,
where a counting area was set up. The seals here were diffusely

550
distributed, with 1 - 2 beside one hole or edge of an open
uatch of water, at a distance of 0.3 - 1.5 km from one another.
According to our calculations (density 0.89 head), in
this area there were lying about 200 - 250 seals. Besides
this, individual grey seals were also encountered on the
breakine-uu landfast ice.
The finding of a single hauling-out uatch of grey
seals in April evidently cannot serve for evaluating their
population numbers in the Gulf. According to the data of
Sundstrem (1962), adult grey seals are attached to their own
reuions and migrate only in the summer. Therefore we assume
the possible oresence of such hauling-out patches in other
regions of the Gulf.
Un the basis of the results of the aerial survey,
which was carried out by us in D.arch - April, there were reck-
oned to be about 3 thousand ringed seals on the ice in the
Gulf of Riga. Taking into consideration that at the moment
of the count a part of the animals were in the water and an-
other part was omitted while making the record entries, and
introducing the appropriate corrections (tentatively 30% -in
the water, and 20% - for omissions), the auproximate numbers
of ringed seals in the Gulf of Riga were estimated at about
4.5 thousand head.
The calculated number of holes on the basis of the
encounters in 'earch was 3.9 thousand; with the correction
(20%) for omissions the numbers of these come to 4.68 thousand
holes.

551
16 LAKE UDOGA
A tentative picture concerning the places of the dis-
tribution of the Ladoga seal (on the basis of findings of
holes) over the expanse of the lake was provided by the first
test flight over this in T^:ar.ch. holes of the ringed seals
were occasionally found from 5-to 30 km from the eastern and
western shores of the lake.
At the beginning of April up to 20jo of the surface of
the lake remained covered with stationary ice, 60Jo of its re-
maining area was covered with breaking-up landfast ice (Figure
8). The snow cover on the breaking-up landfast ice had an in-
dex ranging from 1 to 2.
At this time on the ice there were found not only holes,
but also ringed seals beside the holes or open patches of water.
The Ladoga seal, like the ringed seal of the Baltic,
prefers landfast ice derived from drifted-in frozen-together
ice. In the northern part of the lake, up to the line of the
Valaam archipelago,.on the landfast ice with very sparse cracks
and hummocks the density of the hauling-out of the ringed
seals in April was 0.028 head, while the density of the holes
was 0.081-1•. The density of the hauling-out of the seals on the
breaking-up landfast ice derived from drifted-in ice was 0.19.
head, while the density of the, holes was 0.36.
The main part of the population of the Ladoga seal
during the syprinf; period was dispersed around the central part 311-0
of the lake, which part is the last to freeze up (in mild win-
ters - in Pebruary, and in severe winters - in January). it
is interesting that, according to the data of Kalesnik (1968),
these places are the basic regions for catching fish.

552
o The number of seals calculated on the basis of their
encounters (2200 head), in our opinion, does not reflect the
actual stocks, since the average yearly catch of these seals
of about 0.5 thousand head has been maintained for several
years without changing. Moreover, the small number of encoun-
ters with these seals during the survey in April may be ex-
plained by the fact that at this time there was cloudy and
rainy weather over Lake Ladoga.
The number of holes of the seals on Lake Ladoga, with
the correction for the inherent error . (20%), is about 5.1 thou-
sand. The partial breking-up of the landfast ice undoubtedly
influenced the numbei's of these, but at the present time we
cannot estimate any correction .factor for this. It can only
be assumed that the numbers of ringed seals in this case should
be greater than the calculated number of holes.
In the analysis of the data obtained in March and
April on the distribution of seals beside the holes and open
patches of water some interesting data were obtained.
In the Gulf of Finland in narch 85% of the seals were
found beside the holes, while in April - only 44%. In the
Gulf of Riga the corresponding value was 48% in Larch, while
in April the rined seals were encountered only beside open
patches of water.
In Lake LFidoga in April 79% Of the ringed seals were
found beside the holes and 21% beside open patches of water.
Un more than one occasion, while conducting the aerial survey,
we noted that on the approach of the airplane the ringed seal
escapes into the water not by way of the polynya but through
its hole, which is sometimes situated fairly close to the

.553
polynya (uo to 3 m). On this basis we judge that there is a
considerable attachment of the seal to its hole. The differing
percentage of the finding of the animals beside the holes or
beside the ooen patches of water in March and in April may be
explained by the availability of the holes, the number of which
decreases as the ice breaks up, or by a differing physiologi-
cal state of the animals, which compels the seals to make
greater use of open patches of water and cracks.
CONCLUSIONS
The comparison of the results of the aerial survey in
March and April showed that:
1. The ringed seal in March and April prefers landfast
ice derived from drift ice and ice fields.
In April it is concentrated on the edges of the breaking-
up landfast ice, where it forms scattered hauling-out patches
(possibly, moulting patches). The number of holes decreases
on the gradually breakin-up ice.
2. On the basis of the présence of whitecoats of the
ringed seal in the Gulfs of Riga and 2inland at the beginning
of March and in April there may be assumed to exist a certain
protraction in the periods of its reproduction.
3. The grey seal (in hauled7out groups and singly)
is found only on broken-up white ice.
I. Taking into consideration that this was the first
attempt to survey the ringed seal and grey seal on the landfast
ice in the conditions of the severe winter of 1969-1970, the
data obtained on the distribution and the numbers of the 'seals

5 54.
o should be considered as tentative , it is possible that the
spring concentrations and numbers of seals in the Gulfs of the
Baltic are not constant and depend on the climatic conditions
each year. In the future, after a thorouFfh study has been
conducted on the ice on the ecology and character of the dis-
tribution of the seals, it would be useful to conduct a repeated
aerial survey of these seals.
REFERENCES
JIHTEPATYPA
1 .AT.,111C .1b,a00 Baal- Um:or° mopst n npltaeratoinitx pailotion. I'n,ipomcreon3..],ar. JI,,,, 1960. .
4 "(op°&Pace C. B. Bo3.aytinibie pa3ne;unt Tio.letteil 13 OXOTCHOM mope. Plan. TI HPO, T. 1, 131.111, 20, 19-10.
Ezopeati A. B. Ba./IT1111CRO2 tope. reorpinpr113, M. 1951. . 3y6oce A. 11. Mepoupturrita no opronmanini upombic:la na Jhukowcizom o3epe
It cp,tincizom 3aanne. «PIAOnce .xo3sn'tcrito», .N": 2, 1965.
g Kanecrite.: C. B. fia,11,0C102 o3epo. FiLapo1e -reon3RaT, IL, 1968. :MeTo:mu-tut:me yKa3anup. .1■i'c 11 (no ,ne.3,onbtm insuattitounbim na6. -uoJtentraNt 112
11311311X), 1:10,pomereoli3;.taT, .11., 1959. . 7 1.1a3aperwo 10. II. Cne,a,entia o ripombtc. -ie CC1321111011 Koithtialoil itepubt. C6.
«ig,opcinte 1.-ieKontrra1outile». e.1-layNa», M., 1969. rso /7onoa .11. A. .11e..tionbiii Rperup 6ito.wron. «Phi()floe X0331r1CTI30», :NI? 10, 1966. y • Cottpuûou II. C. 0111,1T pleTa 3111C.1e11110CT11 pactipeitemenna nepubt na o3epe
Ballica.a c nomoutido comoaera. 113.a. HpKy .rcEoro c/xo3. ini-Ta, nun. 6. 1954. 10 CoKadou A. C. MaTeptiaabi no 6110.1911111 :111,110111C1WrO Ttoaelta Yit. 3an.• nett.' roc., nee.. 1111-Ta. T. 179. JI, 1958. 11 Copmert C. M. 3a1laebl TIO.,1C1111 IS Jla.no:KeKom ofiepe. CO. «Bonpocbt pbrOo-ncocrna it Ro6bniu pulObi»..1-1„ 1958. 12 . c '._,uilpiroce B. C: OnettKa ,1(0CT011epllOCTII rICT11131X ,11111111131X tipti rie.re it:futon-HOCTII )1111130T111 ,1X lia 6oabitnix noonumtstx. CO. «Y‘teut oxern. ;Ktinornbtx na 6o.lb-utitx TeppuToptiax», Ilytutinc-iia- Oie, 1969. 13 - Tuxu.ntipod .9. A. OripeReaeitire Bourn ,o,a:ibitenocTomibtx Tioaelieil c camcae -ra 143 13 . THIWO, T:58
, 1966.
1.11.- Tuxo.itupoo .9.A., Kuucerrep 11. B. ,1:1a.-i1.ileuocr0xitibie .1acT01t0rite. - )._1riabliè- nocy. I:MIMI. 113,1,-110, B.911.:111110CTOK, 1966. 15 •<bedoceeti F. A. K bK0.1011111 pa3mitonceinist Ticmienert cenepitort ttacnt OXOTC110- rotopa. 113u. T 1'1 1-WO, T. 59, 1963. 1 6 Yana:lea K. K. .rlaRoxcicurt Tio.rumb u B031■103EFIOCTb ero npombic.m. 11313.
Hrzuo.nor 1111-Ta, T. XII, 213111. 2, 1932. 17 llancradi K. K. Hernia 3ottaatibix mope ConeTcKoil Apwritint. Tp. ApKr. H1111 nnineenmopnyrit, T. 145, Ji.-M., 1940. 18 gancKua K. K. 0 .rpru1, .lac-rottortix. CO. «MneKontrraiontue CCCP». 4ac -rb 11. 143 1,-no AH CCCP, 1963. 19 gepcoitubtli B. B. .A131layTICT 110111.31111b1X 311 py6eaccm. B c6.: «Y 3 1CTI.31 0X01
-111131611X )1111130111131X na 60.11.untx. Teppirropusix». flyninno-na- Oie, 1969. 20 -Wilma A. 17. Oubur KonintecTnentioro a3ponti3yaabnoro rICTa T10.1C11a 2 cencp0-3anaat1o6 tiacrit Bepintrona i ■topa. CO. «MopcKne noeKonirritiontue», -.ethyl-Ka», M. 1969. . •:. 21' Bergniatz G. «Suomen hyljekannaista». Helsinki. 1958: 2 - Curry-Lindahl K. The plight of the grey seal in the Baltic. «Oryx», vol. 8, - no. 1. 1965.
. .
23 Grave K. Die. geographische Verbreitung der Pinnipetlia Nova Acta K. Lcop. Cai.ol. Akad. Naturf. 1896. ,

555
(.:1■alarnichen Viesti», No 1, Helsinki, 1966, .25'.:illolin 0. De bottniska sjaelarnes lefnadsvanor. Fauna och Flora. Uppeales.
f92 1. •26 . Ropelcu.ski il. Ssaki Baltyku. Nakladew Zakladi Ochrony Przyrody. Kur-
kow. 1952, .27 Smith E. .4..4eviev; of the world's grey seal population. «J. Zoo!». vol.
15eno 4. 1966. ; 28 &Inds/Win: K. Giasiil i norra Ostersjtin. «Sver. nabr. Arsbok, 1962, Aroj. •
53», Sioekhohn. 1962.
1. Atlas of ice conditions in the baltic Sea and adjacent
regions. Gidrometoizdat (State Publishing House of
Hydrometeorolouical Literature). Leningrad, 1960.
2 , Dorofeev S. V. Aerial prospecting for seals in the Sea
of ukhotsk. Izvestiya TINRO (Proceedings of the
Pacific ()cean Research Institute of Fisheries and.
LceanograPhy), vol. 1, issue 20, 1940.
3. Egor'eva A. V. The Baltic Sea. Geografgiz (State Publi-
•shing House of Geographical Literature), Moscow, 1961.
4. Lubov A. 1. Leasures for the organization of the industry
on Lake Ladoga and in the Gulf of Finland. "Rybnoe
khozyaistvo" ("The Fishing industry"), No. 2, 1965.
5. Kalesnik S. V. • Lake Ladoga. Gidrometeoizdat, Leningrad,
1968.
6. Lethodological instructions .No. 14. (for aerial observation
of ice conditions at sea), Gidrometeoizdat, Leningrad,
1959.
7. iazarenko Yu. 1. Information on the industry based on
the northern ringed seal. Sb. "Iorskie mlekopitayushchie"
(Symp. "rarine mammals"), Publ. "i\auka", Moscow, 1969.
8. Popov L. A. i)iolouists adrift on the ice. "Rybnoe
khozyaistvo". ‘Ic). 10, 1966.

556
9. Sviridov à,. S. An attemDt to surveJ- the population
numbers and distribution of seals on Lake ^3aikal with
the aid o^' an airolane . Izv. Irl^.utsko9o sel'sko-
khozyaistvenno`o instituta (Proc. ïrkutsk Agricultural
_i.nst;_-tute ), issue 6, 1954•.
,
40
10. Sokolov A. S. information on the biolog^T of the Ladoga
seal. Uchebnie zapl.ski Leningrad.skon-o gosudarstvenno"o
oedayoüicYiesko`_.o instituta (Scier_tific ilotes of the
Leningrad State ^ed.a^-ogic .institute ), vol . 179 .
Leningrad, 195 8.
11 . Sorokin :^ . T'_. The seal stocks in Lake Ladoga. Sb. •".,top-
r. osv- r^Tbovoc7stva i dobychi r.;*by" "Problems of
f i sh managen.ent and f i sh catching"), Leningrad, 1958-
12. Smirnov V. S. An evaluation of the reliability of count-
in` data in counts of the population numbers of animals
in large areas. ;7b. "Uchety qlcrotnichikh zhivotnykh
na bol"shikh terr,itoriyakh" (S^^r^i_p. "Surveys of hunted
animals over large territories"), Pushchino-na- i;ke,
1;' ; 9 .
13. Ti khomir. ov É. A. The identification of the s-oecies of
_Pri,x-- Eastern seals from an ai rplane . Tzvestilra T1'-i R0,
.vol . 58, 1 966
14 . Ti.lchomirov E. A. and ;•^:izevetter T, V. Far-Eastern uinni-
oeds . Lal'nevostochnoe kni7hnoe izdatel'stvo (:.iar-
East -Doo'•••_ Publishers), Vladivostok, 1966.
c'edoseev À. Contribution to the ecolo^,y of re^?^:,oduc-
tior^ of. the seals of the r_orther.r ?art of the Sea of
;lt;hoiSk. ï?vest.l-cL vol. 59, 1061,.

55?
16. Chapskii K. K. The Ladoga seal and the possibility of its
commercial exploitation. lzvestiya ikhtiologicheskogo
instituta (1-,roceedinf, s of the Ichthyological :institute),
vol. 12, issue 2, 1932.
17. Chapskii K. K. The rinRed seal of the western seas of
the Soviet Arctic. Trudy Arkticheskogo nauchno-
issledovatel'skoiTo instituta Glavsevmorputi (Trans-
actions of the Arctic Research Institute 1Sin Direc-
torate of the lorthern seaway), vol. 14.5, Leningrad -
:..oscow, 1940.
18. Chapskii K. K. The order of the pinni -Jeds. Sb. "Ueko-
pitayushchie SSSR" (Symp. ":,.ammals of the USSR").
Part II. Izd-vo An SSSR (Publ. USSR Academy of Sciences),
1963. •
19. Chervonnyi V. V. Aerial surveying of unulates abroad.
In sym.). "Surveys of hunted animals over large terri-
tories". Pushchino-na-Uke, 1969.
20. Ohustov A. P. An attempt at a .quantitative aero-visual
survey of seals in the north-western part of the ering
Sea. Symp. "Marine mammals", Publ. "Nauka", Moscow,
1969.
21. Bergman G. "finland's seal stocks". Helsinki, 1958.
23. Greve M. The geographical distribution of the Pinnipedia.
24. "2ishermen's re'oort", Ho. 1, Helsinki, 1966.
25. Holm 0. Habits of the seals in the Gulf of Bothnia.
L,launa and 21ora. U3psala, 1921.
26. Ropelewski A. Hamrnals of the ifaltic. Publ. i\ature Pro-
tection Research Center of the Polish Academy of Sciences.
'frak6w, 1952.

2F5. Suncl.str8rii K. The "re,y seals of the northern <ali,.^c .
"Sve r. natur. .jLrsbok, 1962, :lro j 53". Stockholm,
1962.
ri

559
TABLE OF CONTENTS 343
Page Page in in
text trans- lation
Section 1. The ecology and distribution of cetaceans
B. A. Zenkovich. The fate of whales 7 4
V. 1. Krylov and L. P. Medvedev. Ubservations on cetaceans and pinnipeds during the 13th Soviet Antarctic Ext-)edition (SAE) 28 42 .
G. A. Klevezal' and D. D. Tormosov. The separation of local zroupins of sperm whales by the character of the dentine layers of the teeth 35
V. V. Zimushko. information on the reproduction of grey whales 44. 68
V. L. Yukhov. Some data on the feeding of sperm whales in the high latitudes of the Antarc- tic . . . 54 83
L. P. Medvedev. Information on the feeding of the beluga in the Dikson region 60 92
V. 1. Shevchenko. A contribution to the question of the origin of "white scars" on the body of whales 67 105
M. V. Ivashin. Cases of multiple pregnancies and foetal monsters in the baleen whales . . . 75 118
Section 11. The ecology, distribution and commercial exploitation of
seals, fur seals and sea otters
G. A. Fedoseev. The distribution and numbers of seals on whel -oing and moulting patches in the Sea of Ukhotsk 87 135
L. A. Popov. The causes and dimensions of the natural mortality of young harp seals durinsu the lactation period 100 159
• 54

560
*
,
Page Page in in
text trans- lation
V. I. Krylov. The feeding of the Pacific walrus (Ldobaenus rosmarus divergens Ill.) . . . . 110 177
V. D. Pastukhov. Counting the young stock of the Baikal seal 117 190
V. L. Kogai. The state of the stock and the pro- gnosis of the feasible catch of fur seals on Robben Island 125 204
A. V. Zasosov. A method for determining the size of the fur seal oopulation and of the maximal possible stable catch quota 130 211
. Arsen'ev. The local distribution of fur seals in the Sea of Japan 138 226
F. G. Chelnokov. I:ovements of fur seals in the rookeries on Uednyy Island 151 240
V. E. Sokolov and N. S. Stepanov. An experiment on the radio tracking of fur seals 157 250
T. I. Chupakhina. The helminth fauna of the fur seals on Robben island 166 265
A. M. Nikolaev. 1211grations and local regrouoings of the Kuril sea otter 171 274
Section III. The morphology and:physiology of marine mammals and othet questions
E. I. Ivanova. The morphology of the vascular system of cetaceans and pinnipeds 179 285
A. S. Leontyuk. Peculiarities of the development of the blood-vascular system during the early embryogenesis of cetaceans 193 309
— P. A. Korzhuev. The problem of the oxygen réserves in the organism of aouatic mammals . . . . 205
V.
331
T. N. Glazova. Quantitative evaluation of the haematopoietic function of the skeleton of the Black Sea dolphins 212 344
G. N. Solntseva. The comparative-anatomical and histological characteristics of the structure of the external and Internal ear of certain
dolphins 226 369

and Caspian 273 440
Baikal seal
D. A. Morozov. Transportation of live dolphins . 296 479
K. K. Chapskii. generic seals .
The systematic rank and supra-differentiation of the 8-incisor
305 494
561
Page Page in in
text trans lation
G. M. Kosygin and V. N. Gol'tsev. Data on the morphology and ecology of the harbour seal of the Tatar Strait 238 384
V. A. Bychkov. The seasonal variation in the density of the fur in the hair bundles of three-year- old bachelor fur seals 253 411
G. A. Nesterov. The effect of the muscle relaxant ditiline on the organism of fur seals . . . 263 • 423
- V. N. Mineev. Questions of conservation and regula- tion of the stocks of marine mammals . . . 269 432
D. A. Eikeladze. Keeping fur seals seals in the Batumi Aquarium
V. D. Pastukhov. The feeding of the under experimental conditions 280 451
V. E. Bel'kovich and V. S. Gurevich. Questions re- lating to the catching and long-term holding of dolphins in captivity 286 461
A. A. Antonyuk. Quantitative changes in the ver- tebrae in the vertebral column of pinnipeds 317 514
V. A. Zheglov and K. K. ChaPskii. Test of an aerial survey of the ringed seal and of the grey seal and of their holes in the gulfs of the Baltic Sea and on Lake Ladoga . . • 323 524

4 562
RESEARCH ON MARINE MAMMALS
Transactions, issue XXXIX
7i'ditor Ê. S. Mil'chenko
Technical editor T. M. Andreeva
Artistic editor G. P. Burov Proof-reader E. I. Arenko
Delivered for type-setting 10th uctober 1972. Sent to press
19th 1,.ay 1972. Form .Ab uf paper 60 x 90 1/16. Conventional
printers' sheets 21.5. Registered publishers' sheets 21.18.
Edition 1000 copies. KU 04382. Larder 2354. Price 1 rouble, 60 cooecks.
Atlantic Research Institute of Fisheries and Oceanography.
Kaliningrad obl., ul. Dm. Donskogo, 5.
Printing-house of the "Kaliningradskaya pravda" publiShers.
Kaliningrad obi., ul. Karla Marksa, 18.
eee,74.

DUE DATE
1/(1
;
1
201-6503 . printed in USA




















