
Transcriptional synergy mediated by SAF-1 and AP-1: critical role of N-terminal polyalanine and two...
11
Transcriptional Synergy Mediated by SAF-1 and AP-1 CRITICAL ROLE OF N-TERMINAL POLYALANINE AND TWO ZINC FINGER DOMAINS OF SAF-1 * Received for publication, August 14, 2008, and in revised form, November 20, 2008 Published, JBC Papers in Press, November 21, 2008, DOI 10.1074/jbc.M806289200 Deepak Kumar, Alpana Ray 1 , and Bimal K. Ray 2 From the Department of Veterinary Pathobiology, University of Missouri, Columbia, Missouri 65211 Previously we determined that inflammation responsive tran- scription factors AP-1 and SAF-1 synergistically regulate tran- scriptional induction of the MMP-1 gene. The present study investigated the underlying molecular mechanism of cooperat- ivity between these two different groups of transcription factors. We present evidence that knockdown of SAF-1 by small inter- fering RNAs inhibits AP-1-mediated increase of human MMP-1 expression. The two key members of the AP-1 family of proteins, c-Fos and c-Jun, and SAF-1 form a ternary protein complex, which has markedly higher DNA binding activity than either a SAF-1 homodimer or a c-Fos/c-Jun heterodimer. The increased DNA binding activity of the ternary complex is translated into a striking enhancement of their transcriptional activity by which synergistic transcriptional induction of MMP-1 expression is achieved. The SAF-1c-Fosc-Jun ternary complex efficiently promotes transcription from both SAF-1 and AP-1 sites of human MMP-1 promoter. The physical interaction between SAF-1 and AP-1 was demonstrated both in vitro by Far-Western and antibody pulldown assays with recombinant proteins and in vivo by chromatin immunoprecipitation (ChIP), re-ChIP, and co-immunoprecipitation analyses. Two distinct but adjacent domains in SAF-1 are involved in protein-protein contact with c-Fos and c-Jun; one domain resides within two N-terminal polyalanine tracts, and the other is present within the first two zinc finger motifs. Together these findings delineate the mech- anism of synergy and the essential role of SAF-1 and AP-1 in up-regulating human MMP-1 expression under various inflam- matory conditions. A crucial role of matrix metalloproteinase-1 (MMP-1) 3 in the pathogenesis of various diseases associated with extracellular matrix (ECM) degradation is well established (1–3). MMP-1 is a prominent member of more than 26 structurally related matrix MMP proteins that in concert are capable of degrading all components of the ECM at physiological pH (4, 5). Although all MMPs possess some ECM-degrading capacity, the initial cleavage of triple helical collagen can be done only by a small subset of this family of which MMP-1 is one. Normal expres- sion of MMP-1 is thus at a very low level and is highly regulated. Under many pathogenic conditions, MMP-1 protein concen- tration is markedly increased, primarily by transcriptional induction of the MMP-1 gene. Increase of MMP-1 mRNA by many inflammatory stimuli such as cytokines, growth factors, and tumor promoting agents is still a major challenge in suc- cessful inhibition of this molecule for the treatment of all MMP-1-related pathogenicities (6). Several transcription factors have been implicated thus far for transcriptional induction from the MMP-1 gene; these include activator protein 1 (AP-1) (7) and serum amyloid A-ac- tivating factor 1 (SAF-1) (8). AP-1 comprises a group of pro- teins containing a basic leucine zipper DNA-binding domain. AP-1 family members consist of the Fos (c-Fos, Fra-1, Fra-2, FosB) and Jun (c-Jun, JunB, and JunD) proteins (9, 10). Typi- cally, AP-1 protein dimers bind to AP-1-binding elements, and two subunits are formed either by heterodimerization or homodimerization. In cells, the Fos/Jun heterodimer is the most predominant form of AP-1, which has a very high affinity for an AP-1 DNA-binding element, whereas the Jun/Jun homodimer exhibits a very low affinity for the same DNA. AP-1 proteins are transcriptionally induced by the action of various serum and growth factor regulated factors. For example, the c-fos gene is transcriptionally induced by the mitogen-activated protein (MAP) kinase-activated Elk transcription factor, a member of the ternary complex factor (TCF) family, (11, 12) and by protein kinase C during oncogenic Ha-Ras-mediated induction (13). SAF-1 transcription factor is a member of the multiple Cys 2 -His 2 -type zinc finger proteins, which also are activated in response to various inflammatory stimuli, includ- ing cytokines, phorbol 12-myristate 13-acetate, lipopolysac- charide, and oxidized low density lipoproteins (14 –17). Phos- phorylation of SAF-1 via MAP kinase, protein kinase C, or protein kinase A markedly increases its DNA binding and transactivation potential (18 –20). The activation of AP-1 and SAF-1 by similar signal transduc- tion cascades, the MAP kinase and protein kinase C pathways, suggests that these transcription factors may function in a cooperative manner and/or may be dependent on each other to exert their full transactivation potential. A recent study from our laboratory indicates that concurrent participation of AP-1 and SAF-1 is necessary for full transcriptional induction of * The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertise- ment” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. 1 To whom correspondence may be addressed: Dept. of Veterinary Pathobi- ology, University of Missouri, 126 Connaway Hall, Columbia, MO 65211. Tel.: 573-882-6728; Fax: 573-884-5414; E-mail: [email protected]. 2 To whom correspondence may be addressed: Dept. of Veterinary Pathobi- ology, University of Missouri, 124 Connaway Hall, Columbia, MO 65211. Tel.: 573-882-4461; Fax: 573-884-5414; E-mail: [email protected]. 3 The abbreviations used are: MMP, matrix metalloproteinase; AP-1, activator protein 1; ECM, extracellular matrix; SAF, serum amyloid A-activating fac- tor; MAP, mitogen-activated protein; siRNA, small interfering RNA; CAT, chloramphenicol acetyltransferase; ChIP, chromatin immunoprecipita- tion; GST, glutathione S-transferase; RT, reverse transcription; IL, interleukin. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 284, NO. 3, pp. 1853–1862, January 16, 2009 © 2009 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A. JANUARY 16, 2009 • VOLUME 284 • NUMBER 3 JOURNAL OF BIOLOGICAL CHEMISTRY 1853 by guest on June 3, 2016 http://www.jbc.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Transcriptional synergy mediated by SAF-1 and AP-1: critical role of N-terminal polyalanine and two...

Transcriptional Synergy Mediated by SAF-1 and AP-1CRITICAL ROLE OF N-TERMINAL POLYALANINE AND TWO ZINC FINGER DOMAINSOF SAF-1*
Received for publication, August 14, 2008, and in revised form, November 20, 2008 Published, JBC Papers in Press, November 21, 2008, DOI 10.1074/jbc.M806289200
Deepak Kumar, Alpana Ray1, and Bimal K. Ray2
From the Department of Veterinary Pathobiology, University of Missouri, Columbia, Missouri 65211
Previouslywe determined that inflammation responsive tran-scription factors AP-1 and SAF-1 synergistically regulate tran-scriptional induction of the MMP-1 gene. The present studyinvestigated the underlying molecular mechanism of cooperat-ivity between these twodifferent groups of transcription factors.We present evidence that knockdown of SAF-1 by small inter-fering RNAs inhibits AP-1-mediated increase of humanMMP-1expression.The twokeymembers of theAP-1 family of proteins,c-Fos and c-Jun, and SAF-1 form a ternary protein complex,which has markedly higher DNA binding activity than either aSAF-1 homodimer or a c-Fos/c-Jun heterodimer. The increasedDNA binding activity of the ternary complex is translated into astriking enhancement of their transcriptional activity by whichsynergistic transcriptional induction of MMP-1 expression isachieved. The SAF-1�c-Fos�c-Jun ternary complex efficientlypromotes transcription from both SAF-1 and AP-1 sites ofhuman MMP-1 promoter. The physical interaction betweenSAF-1 andAP-1was demonstrated both in vitroby Far-Westernand antibody pulldown assays with recombinant proteins and invivo by chromatin immunoprecipitation (ChIP), re-ChIP, andco-immunoprecipitation analyses. Two distinct but adjacentdomains in SAF-1 are involved in protein-protein contact withc-Fos and c-Jun; one domain resides within two N-terminalpolyalanine tracts, and the other is present within the first twozinc finger motifs. Together these findings delineate the mech-anism of synergy and the essential role of SAF-1 and AP-1 inup-regulating humanMMP-1 expression under various inflam-matory conditions.
Acrucial role ofmatrixmetalloproteinase-1 (MMP-1)3 in thepathogenesis of various diseases associated with extracellularmatrix (ECM) degradation is well established (1–3). MMP-1 isa prominent member of more than 26 structurally related
matrix MMP proteins that in concert are capable of degradingall components of the ECMat physiological pH (4, 5). Althoughall MMPs possess some ECM-degrading capacity, the initialcleavage of triple helical collagen can be done only by a smallsubset of this family of which MMP-1 is one. Normal expres-sion ofMMP-1 is thus at a very low level and is highly regulated.Under many pathogenic conditions, MMP-1 protein concen-tration is markedly increased, primarily by transcriptionalinduction of the MMP-1 gene. Increase of MMP-1 mRNA bymany inflammatory stimuli such as cytokines, growth factors,and tumor promoting agents is still a major challenge in suc-cessful inhibition of this molecule for the treatment of allMMP-1-related pathogenicities (6).Several transcription factors have been implicated thus far
for transcriptional induction from the MMP-1 gene; theseinclude activator protein 1 (AP-1) (7) and serum amyloid A-ac-tivating factor 1 (SAF-1) (8). AP-1 comprises a group of pro-teins containing a basic leucine zipper DNA-binding domain.AP-1 family members consist of the Fos (c-Fos, Fra-1, Fra-2,FosB) and Jun (c-Jun, JunB, and JunD) proteins (9, 10). Typi-cally, AP-1 protein dimers bind to AP-1-binding elements, andtwo subunits are formed either by heterodimerization orhomodimerization. In cells, the Fos/Jun heterodimer is themost predominant form of AP-1, which has a very high affinityfor an AP-1 DNA-binding element, whereas the Jun/Junhomodimer exhibits a very low affinity for the sameDNA.AP-1proteins are transcriptionally induced by the action of variousserum and growth factor regulated factors. For example, thec-fos gene is transcriptionally induced by themitogen-activatedprotein (MAP) kinase-activated Elk transcription factor, amember of the ternary complex factor (TCF) family, (11, 12)and by protein kinase C during oncogenic Ha-Ras-mediatedinduction (13). SAF-1 transcription factor is a member of themultiple Cys2-His2-type zinc finger proteins, which also areactivated in response to various inflammatory stimuli, includ-ing cytokines, phorbol 12-myristate 13-acetate, lipopolysac-charide, and oxidized low density lipoproteins (14–17). Phos-phorylation of SAF-1 via MAP kinase, protein kinase C, orprotein kinase A markedly increases its DNA binding andtransactivation potential (18–20).The activation of AP-1 and SAF-1 by similar signal transduc-
tion cascades, the MAP kinase and protein kinase C pathways,suggests that these transcription factors may function in acooperativemanner and/ormay be dependent on each other toexert their full transactivation potential. A recent study fromour laboratory indicates that concurrent participation of AP-1and SAF-1 is necessary for full transcriptional induction of
* The costs of publication of this article were defrayed in part by the paymentof page charges. This article must therefore be hereby marked “advertise-ment” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1 To whom correspondence may be addressed: Dept. of Veterinary Pathobi-ology, University of Missouri, 126 Connaway Hall, Columbia, MO 65211.Tel.: 573-882-6728; Fax: 573-884-5414; E-mail: [email protected].
2 To whom correspondence may be addressed: Dept. of Veterinary Pathobi-ology, University of Missouri, 124 Connaway Hall, Columbia, MO 65211.Tel.: 573-882-4461; Fax: 573-884-5414; E-mail: [email protected].
3 The abbreviations used are: MMP, matrix metalloproteinase; AP-1, activatorprotein 1; ECM, extracellular matrix; SAF, serum amyloid A-activating fac-tor; MAP, mitogen-activated protein; siRNA, small interfering RNA; CAT,chloramphenicol acetyltransferase; ChIP, chromatin immunoprecipita-tion; GST, glutathione S-transferase; RT, reverse transcription; IL,interleukin.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 284, NO. 3, pp. 1853–1862, January 16, 2009© 2009 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in the U.S.A.
JANUARY 16, 2009 • VOLUME 284 • NUMBER 3 JOURNAL OF BIOLOGICAL CHEMISTRY 1853
by guest on June 3, 2016http://w
ww
.jbc.org/D
ownloaded from

canine MMP-1 (21). The SAF-1 and AP-1 DNA-binding ele-ments arewell conserved in theMMP-1 promoter amongmanyspecies, which further underscore their importance. Functionalcooperation between candidate proteins, in general, is medi-ated by protein-protein interaction, which is most effectivewhen the DNA-binding elements of the involved transcriptionfactors are proximally located. In correlation with this hypoth-esis, several studies are available that demonstrate the ability ofnuclear proteins to bind each other and to mediate cooperativeDNA binding and promoter activation when their respectivebinding elements are juxtaposed (22, 23). However, in humanor canineMMP-1, SAF-1 and AP-1 DNA-binding elements arenot in close proximity but are more than 250 nucleotides apart.To understand the mechanism of cooperation between SAF-1andAP-1, we have extended the study of humanMMP-1, and inthis report we present evidence for a physical interactionbetween these proteins, the formation of a ternary protein com-plex with markedly high DNA binding ability and a synergisticlevel of transactivation potential. Further, this study provides,for the first time, an identification of novel structural motifs inthe SAF-1 transcription factor for protein-protein contact withAP-1 familymembers. These findings extend the paradigm thatcellular machinery utilizes concurrent activation of a networkof transcription factors and their cooperative interaction forefficient and optimal activation of genes.
EXPERIMENTAL PROCEDURES
Cell Culture and Transfection—HTB-94 human chondro-cyte cells derived fromaprimary grade II chondrosarcomawerecultured in Dulbecco’s modified Eagle’s medium containinghigh glucose, 100 units/ml penicillin, and 100 units/ml strepto-mycin supplemented with 7% fetal calf serum. Transfectionassays were performed as described previously (8). The pSV-�galactosidase (Promega) plasmid DNA was used as an internalcontrol and was assayed as described (24). Cells were harvested24 h post-transfection, and CAT activity was measured asdescribed previously (24). All transfection experiments wereperformed at least three times.Plasmids—MMP-1(�518/�63)-CAT reporter was con-
structed by ligating the human MMP-1 genomic DNAsequence from nucleotide position �518 to �63 intopBLCAT3 vector (25). Mutant MMP-1-CAT reporters wereconstructed by ligating the respective mutant DNA fragmentsgenerated by megaprimer PCR into pBLCAT3 vector. MutantSAF-1 and AP-1 oligonucleotide sequences used were5�-CCTTGCTGACACTGCAGAATGTGGAATTACTAAC-3�and 5�-GGATGTTATAAAGCAGATCTTAGACAGCC-3�,respectively. The underlined bases represent altered sequences.The SAF-1CAT (26) orAP-1CAT reporter plasmids containedthree tandem copies of SAF-1 or AP-1 DNA-binding elementsinto pBLCAT2 vector. Various SAF-1 fragments were con-structed by PCR, and wherever it became necessary, initiatorATG and TGA stop codons were added to the codingsequences. The PCR-amplified DNAs were ligated to theappropriate expression vectors, including pcDNA3 (Invitro-gen), pGEX-5X-1 (GE Healthcare), PRSET (Invitrogen), andpAR (27). Primer sets used for synthesizing various SAF-1derivatives, indicated by beginning and ending amino acids in
parentheses, were as follows: SAF-1-(1–477), 5�-ATGTTCCC-CGTGTTCCCTTGCACGCTG-3� and 5�-AGCTCACCAAG-GCTGGGAGGGAAGTGG-3�; SAF-1-(1–90), 5�-ATGTTCC-CCGTGTTCCCTTGCACGCTG-3� and 5�-TCAGAGAACC-GGGAGCAAGTCCAC-3�; SAF-1-(1–192), 5�-ATGTTCCC-CGTGTTCCCTTGCACGCTG-3� and 5�-TCAGGGCCCCT-TGCTCTTTGTCTTCTTC-3�; SAF-1-(90–192), 5�-ATGCC-GGTTCTCGCCGCCGCGCAGGAATC-3� and 5�-TCAGG-GCCCCTTGCTCTTTGTCTTCTTC-3�; SAF-1-(192–360),5�-ATGAAGGGGCCCTACATTTGCGCCCTG-3� and 5�-TCACTCTGTTGAGTGCACTTGTCTGAC-3�; SAF-1-(192-413), 5�-ATGAAGGGGCCCTACATTTGCGCCCTG-3� and5�-TCACTGGCTGTGCACCTTCATGTGG-3�; SAF-1-(90–301), 5�-ATGCCGGTTCTCGCCGCCGCGCAGGAATC-3�and 5�-TCAGTCCGAGTGGGAGAGCTTGTGTCG-3�; SAF-1-(192–301), 5�-ATGAAGGGGCCCTACATTTGCGCC-CTG-3� and 5�-TCAGTCCGAGTGGGAGAGCTTGTG-TCG-3�; SAF-1-(413–477), 5�-ATGGCCAGGGTCCTCACC-ATGTCTGTG-3� and 5�-AGCTCACCAAGGCTGGGAG-GGAAGTGG-3�.Protein Preparation—Bacterially expressed proteins were
purified by affinity chromatography using nickel-agarose(Invitrogen) or glutathione-Sepharose column chromatogra-phy following themanufacturer’s protocol. pET-cJun and pET-cFos plasmid DNAs were obtained from James A. Goodrich,and the c-Jun/c-Fos heterodimer was prepared as described(28).GST Pulldown Assay—GST-tagged full-length SAF-1 or
truncated SAF-1 proteins and control GST proteins (50 pmoleach) were mixed individually with glutathione-Sepharose 4Bbeads (Amersham Biosciences), incubated for 1 h at 4 °C inbinding buffer (12 mM HEPES, pH 7.9, 4 mM Tris-HCl, pH 7.9,100 mM NaCl, 1 mM EDTA), and washed extensively (fivetimes) with binding buffer. Next, GST-SAF-1, truncated GST-SAF-1, or control GSTprotein immobilized to Sepharose beadswas mixed with bacterially expressed c-Jun/c-Fos heterodimer,incubated for 1 h at 4 °C, and washed extensively with bindingbuffer plus 0.05%Nonidet P-40. The proteins were eluted usingelution buffer (50 mM Tris-HCl, pH 8.0, 100 mM reduced glu-tathione) or boiled in protein sample loading buffer containing1% SDS plus �-mercaptoethanol, analyzed by SDS-PAGE using11% polyacrylamide gel, and visualized by Coomassie Bluestaining.Far-Western Assay—Equal amounts of protein were electro-
phoresed on 11% SDS-PAGE and transferred to a nitrocellulosemembrane. Themembranewas incubated in amodifiedHEPESbinding buffer (25 mM HEPES, pH 7.9, 25 mM NaCl, 5 mMMgCl2, 5 mM �-mercaptoethanol) containing 6 M guanidinehydrochloride for 10 min at room temperature. Membrane-bound denatured proteins were renatured by sequential incu-bation of the membrane for five cycles (10 min each) in buffersthat were diluted each time (1:1) with HEPES binding bufferwithout any guanidine hydrochloride. The membrane receiveda final rinse twice in HEPES binding buffer and then were incu-bated in a blocking solution (50 mM Tris-HCl, pH 7.6, 150 mMNaCl, 5% bovine serum albumin) at 4 °C overnight, furtherincubated in overlay buffer (0.5% bovine serum albumin, 0.25%gelatin, 1% Nonidet P-40, 10 mM NaCl, 1 mM EDTA, 5 mM
Interaction between SAF-1 and AP-1
1854 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 3 • JANUARY 16, 2009
by guest on June 3, 2016http://w
ww
.jbc.org/D
ownloaded from

�-mercaptoethanol, 20 mM HEPES, pH 7.5) containing 32P-labeled pAR-SAF-1 at room temperature for 4 h, washed fourtimes for 10 min each in a washing buffer (50 mM Tris-HCl, pH7.6, 150mMNaCl, 0.5% sarcosyl), dried, and autoradiographed.The pAR-SAF-1 protein was prepared by cloning a full-lengthSAF-1 cDNA in pAR vector (27) that contains a FLAG tag anda heart muscle kinase phosphorylation site, and purified pro-tein was radioactively labeled with [�-32P]ATP and heart mus-cle kinase (Sigma).siRNAExperiments—HTB-94 cells were transfected in dupli-
cate with predesigned human MAZ/SAF-1-specific siRNAs(Dharmacon Research, Lafayette, CO) using Oligofectamine(Invitrogen) according to the manufacturer’s protocol. As neg-ative controls, mock transfection (no siRNA) and nonspecificscrambled siRNAs were used. Forty-eight hours after transfec-tion, cells in each treatment groups were harvested and dividedinto two parts. Also conditioned media were collected formeasuring secreted MMP-1 and MMP-2 proteins. Total RNAwas isolated from one portion of the cells using the guani-dinium thiocyanate method (29), and RT-PCR was performedusing an RT-PCR kit from Invitrogen. The PCR primersequences specific for the genes examined and the predictedproduct sizes, shown in parentheses, were: MAZ/SAF-1 (205bp), 5�-GCCAGGGTCCTCACCATGTCTG-3� (sense) and 5�-CAACTTGGAGCTCACCAGGG-3� (antisense); MMP-1 (234bp), 5�-ATGCTGAAACCCTGAAGGTG-3� (sense) and 5�-CTGCTTGACCCTCAGAGACC-3� (antisense); and GAPDH(178 bp) 5�-TGCACCACCAACTGCTTAG-3� (sense) and 5�-TAGAGGCAGGGATGATGTTC-3� (antisense). The secondset of cells was used forWestern blot analysis using either anti-SAF-1 (prepared as described previously (8)). The conditionedmedia were concentrated by freeze-drying in a lyophilizer, andthe protein contents were measured. Fifty micrograms of pro-teins were subjected toWestern blotting using anti-MMP-1 oranti-MMP-2 antibody (Santa Cruz Biotechnology).For cotransfection analysis, siRNAs were transfected into
HTB-94 cells following the manufacturer’s protocol, and 24 hafter transfection the medium was replaced and further trans-fected with MMP-1-CAT reporter plasmid and pcDc-Fos pluspcDc-Jun plasmids. Cells were harvested 24 h later and used forthe CAT assay.Electrophoretic Mobility Shift Assay—Equal amounts (0.1
�g) of bacterially expressed proteins were used for an electro-phoreticmobility shift assay, whichwas performed as describedpreviously (17). Protein concentrations were measured by theBradford method (30). Radiolabeled probes were prepared bylabeling double-stranded oligonucleotides with [�-32P]dCTP.In some assays, anti-SAF-1 or anti-c-Fos and anti-c-Jun(Santa Cruz Biotechnology) antibodies were added during a30-min preincubation on ice prior to the addition of thelabeled probe. Anti-SAF-1 antibody was prepared asdescribed (8). In some assays, 25 pmol of the following oli-gonucleotides were added in the preincubation mixtures ascompetitor oligonucleotides: SAF-1 oligo, 5�-CCCTTCCT-CTCCACCCACAGCCCCCATGG-3�; AP-1 oligo, 5�-CGC-TTGATGAGTCAGCCGGAA-3�; and nonspecific oligo, 5�-TGTCGAATGCAAATCACTAGAA-3�.
Chromatin Immunoprecipitation (ChIP) and Re-ChIP Assays—ChIP was performed with a chromatin immunoprecipitationassay kit (Upstate Biotechnology, Lake Placid, NY) followingthe manufacturer’s protocol. Briefly, HTB-94 cells (2 � 106each) were stimulated with IL-1� (500 units/ml). Twenty-fourhours later, formaldehyde (1% final concentration) was addeddirectly to the culture medium and incubated at 37 °C for 10min for cross-linking. Cells were lysed in 200 �l of cell lysisbuffer (50 mM Tris-HCl, pH 8.0, 10 mM EDTA, 1% SDS) with aprotease inhibitor mixture and sonicated to generate �500-bplong DNA fragments, and the supernatants were diluted with adilution buffer (20mMTris-HCl, pH8.0, 1.0mMEDTA, 150mMNaCl, 1% Triton X-100, 0.01% SDS, and protease inhibitors).The solutions were precleared with a salmon sperm DNA/pro-tein G-agarose slurry and then incubated with anti-SAF-1 anti-body or a mixture of anti-c-Fos and anti-c-Jun antibodies(Santa Cruz Biotechnology) at 4 °C for 16 h with rotation. Theimmune complexes were precipitated with salmon spermDNA/protein G-agarose. The agarose beads were washedsequentially in a low salt wash buffer (20 mM Tris-HCl, pH 8.0,150mMNaCl, 1 mM EDTA, 1% Triton-X-100, and 0.1% SDS), ahigh salt wash buffer (same as low salt wash buffer except con-taining 500 mM NaCl), LiCl wash buffer (20 mM Tris-HCl, pH8.0, 250 mM LiCl, 0.5% Nonidet P-40, 0.5% deoxycholate, andprotease inhibitors), and 20 mM Tris-HCl, pH 8.0, 1 mM EDTAbuffer. Immune complexes were extracted from the beads withelution buffer (20 mM Tris-HCl, pH 8.0, 1 mM EDTA, 1% SDS,and 50 mM NaHCO3) at 65 °C for 4 h.The Re-ChIP assays were performed as described (31).
Briefly, the eluant of the primary immunocomplex obtainedwith the first antibody was diluted 10-fold with dilution buffer(20mMTris-HCl, pH8.0, 1mMEDTA, 150mMNaCl, 1%TritonX-100, and protease inhibitors) and then further subjected toimmunoprecipitation with the second antibody. Cross-linkingwas reversed, and nucleic acids were isolated from the eluatesby proteinase K and RNase A treatments followed by phenolextraction and ethanol precipitation. Purified DNA was sub-jected to PCR using primers covering the AP-1 or SAF-1 DNA-binding regions in the humanMMP-1 promoter. PCR productswere resolved in a 2% agarose gel and visualized by ethidiumbromide staining. The primers used for amplification of theSAF-1 element in humanMMP-1 (162 bp) were 5�-GAACTT-CAGTCAGTACAGGTGCCGAACAGC-3� (sense) and 5�-TACGGTCAAAGAGTACTCCATGGTC-3� (antisense), andthose for amplification of the AP-1 element in humanMMP-1(132 bp) were 5�-TCTAATGATTGCCTAGTCTATTCAT-AGC-3� (sense) and 5�-CTCCAATATCCCAGCTAGGAAG-CTCCCTC-3� (antisense).Immunoprecipitation and Western Blotting—HTB94 cells,
untreated or treated with IL-1�, were lysed using a lysis buffer(50 mM Tris-HCl, pH 7.6, 150 mM NaCl, 0.1 mM EDTA, 1 mMphenylmethylsulfonyl fluoride, 1% Triton X-100). The proteinswere immunoprecipitated with anti-SAF-1 antibody, anti-c-Jun plus anti-c-Fos antibody, or control nonimmune IgG asindicated. Samples were fractionated in SDS-11% PAGE andelectroblotted onto a nitrocellulose membrane. Immunoblot-ting was performed using anti-c-Fos, anti-c-Jun, or anti-SAF-1antibody as the primary antibody and horseradish peroxidase-
Interaction between SAF-1 and AP-1
JANUARY 16, 2009 • VOLUME 284 • NUMBER 3 JOURNAL OF BIOLOGICAL CHEMISTRY 1855
by guest on June 3, 2016http://w
ww
.jbc.org/D
ownloaded from
conjugated goat anti-rabbit IgG as the secondary antibody.Bands were detected using a chemiluminescence detection sys-tem (Amersham Biosciences).
RESULTS
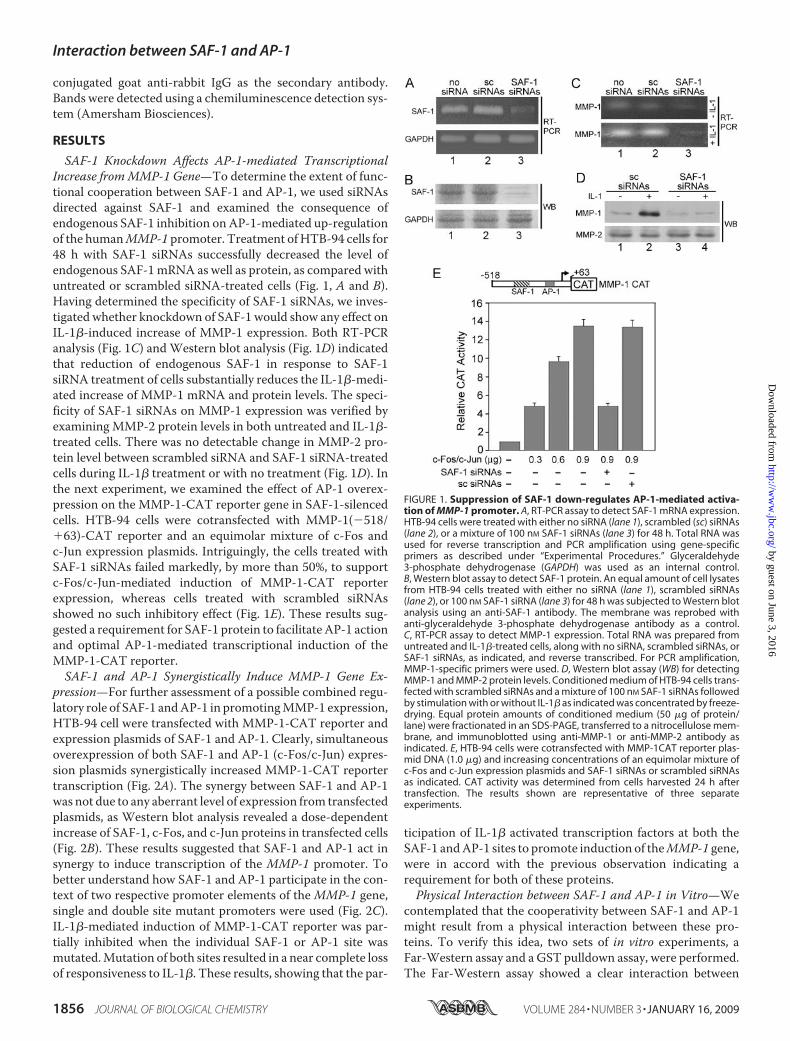
SAF-1 Knockdown Affects AP-1-mediated TranscriptionalIncrease fromMMP-1 Gene—To determine the extent of func-tional cooperation between SAF-1 and AP-1, we used siRNAsdirected against SAF-1 and examined the consequence ofendogenous SAF-1 inhibition onAP-1-mediated up-regulationof the humanMMP-1 promoter. Treatment ofHTB-94 cells for48 h with SAF-1 siRNAs successfully decreased the level ofendogenous SAF-1mRNA as well as protein, as compared withuntreated or scrambled siRNA-treated cells (Fig. 1, A and B).Having determined the specificity of SAF-1 siRNAs, we inves-tigated whether knockdown of SAF-1 would show any effect onIL-1�-induced increase of MMP-1 expression. Both RT-PCRanalysis (Fig. 1C) andWestern blot analysis (Fig. 1D) indicatedthat reduction of endogenous SAF-1 in response to SAF-1siRNA treatment of cells substantially reduces the IL-1�-medi-ated increase of MMP-1 mRNA and protein levels. The speci-ficity of SAF-1 siRNAs on MMP-1 expression was verified byexamining MMP-2 protein levels in both untreated and IL-1�-treated cells. There was no detectable change in MMP-2 pro-tein level between scrambled siRNA and SAF-1 siRNA-treatedcells during IL-1� treatment or with no treatment (Fig. 1D). Inthe next experiment, we examined the effect of AP-1 overex-pression on the MMP-1-CAT reporter gene in SAF-1-silencedcells. HTB-94 cells were cotransfected with MMP-1(�518/�63)-CAT reporter and an equimolar mixture of c-Fos andc-Jun expression plasmids. Intriguingly, the cells treated withSAF-1 siRNAs failed markedly, by more than 50%, to supportc-Fos/c-Jun-mediated induction of MMP-1-CAT reporterexpression, whereas cells treated with scrambled siRNAsshowed no such inhibitory effect (Fig. 1E). These results sug-gested a requirement for SAF-1 protein to facilitateAP-1 actionand optimal AP-1-mediated transcriptional induction of theMMP-1-CAT reporter.SAF-1 and AP-1 Synergistically Induce MMP-1 Gene Ex-
pression—For further assessment of a possible combined regu-latory role of SAF-1 andAP-1 in promotingMMP-1 expression,HTB-94 cell were transfected with MMP-1-CAT reporter andexpression plasmids of SAF-1 and AP-1. Clearly, simultaneousoverexpression of both SAF-1 and AP-1 (c-Fos/c-Jun) expres-sion plasmids synergistically increased MMP-1-CAT reportertranscription (Fig. 2A). The synergy between SAF-1 and AP-1was not due to any aberrant level of expression from transfectedplasmids, as Western blot analysis revealed a dose-dependentincrease of SAF-1, c-Fos, and c-Jun proteins in transfected cells(Fig. 2B). These results suggested that SAF-1 and AP-1 act insynergy to induce transcription of the MMP-1 promoter. Tobetter understand how SAF-1 and AP-1 participate in the con-text of two respective promoter elements of the MMP-1 gene,single and double site mutant promoters were used (Fig. 2C).IL-1�-mediated induction of MMP-1-CAT reporter was par-tially inhibited when the individual SAF-1 or AP-1 site wasmutated.Mutation of both sites resulted in a near complete lossof responsiveness to IL-1�. These results, showing that the par-
ticipation of IL-1� activated transcription factors at both theSAF-1 andAP-1 sites to promote induction of theMMP-1 gene,were in accord with the previous observation indicating arequirement for both of these proteins.Physical Interaction between SAF-1 and AP-1 in Vitro—We
contemplated that the cooperativity between SAF-1 and AP-1might result from a physical interaction between these pro-teins. To verify this idea, two sets of in vitro experiments, aFar-Western assay and a GST pulldown assay, were performed.The Far-Western assay showed a clear interaction between
FIGURE 1. Suppression of SAF-1 down-regulates AP-1-mediated activa-tion of MMP-1 promoter. A, RT-PCR assay to detect SAF-1 mRNA expression.HTB-94 cells were treated with either no siRNA (lane 1), scrambled (sc) siRNAs(lane 2), or a mixture of 100 nM SAF-1 siRNAs (lane 3) for 48 h. Total RNA wasused for reverse transcription and PCR amplification using gene-specificprimers as described under “Experimental Procedures.” Glyceraldehyde3-phosphate dehydrogenase (GAPDH) was used as an internal control.B, Western blot assay to detect SAF-1 protein. An equal amount of cell lysatesfrom HTB-94 cells treated with either no siRNA (lane 1), scrambled siRNAs(lane 2), or 100 nM SAF-1 siRNA (lane 3) for 48 h was subjected to Western blotanalysis using an anti-SAF-1 antibody. The membrane was reprobed withanti-glyceraldehyde 3-phosphate dehydrogenase antibody as a control.C, RT-PCR assay to detect MMP-1 expression. Total RNA was prepared fromuntreated and IL-1�-treated cells, along with no siRNA, scrambled siRNAs, orSAF-1 siRNAs, as indicated, and reverse transcribed. For PCR amplification,MMP-1-specific primers were used. D, Western blot assay (WB) for detectingMMP-1 and MMP-2 protein levels. Conditioned medium of HTB-94 cells trans-fected with scrambled siRNAs and a mixture of 100 nM SAF-1 siRNAs followedby stimulation with or without IL-1� as indicated was concentrated by freeze-drying. Equal protein amounts of conditioned medium (50 �g of protein/lane) were fractionated in an SDS-PAGE, transferred to a nitrocellulose mem-brane, and immunoblotted using anti-MMP-1 or anti-MMP-2 antibody asindicated. E, HTB-94 cells were cotransfected with MMP-1CAT reporter plas-mid DNA (1.0 �g) and increasing concentrations of an equimolar mixture ofc-Fos and c-Jun expression plasmids and SAF-1 siRNAs or scrambled siRNAsas indicated. CAT activity was determined from cells harvested 24 h aftertransfection. The results shown are representative of three separateexperiments.
Interaction between SAF-1 and AP-1
1856 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 3 • JANUARY 16, 2009
by guest on June 3, 2016http://w
ww
.jbc.org/D
ownloaded from

bacterially expressed His-c-Fos and His-c-Jun proteins withradiolabeled pAR-SAF-1 protein (Fig. 3A, lanes 2 and 3). Radio-active SAF-1 did not interact with the control GST protein (Fig.3A, lane 1). Input of each protein was examined by CoomassieBlue staining (Fig. 3A, lanes 4–6). To gain more evidence, GSTpulldown assays were performed (Fig. 3B). Both bacteriallyexpressedHis-c-Fos andHis-c-Jun proteins interacted strongly
with GST-SAF-1 protein immobi-lized to a glutathione-Sepharosecolumn and were eluted by a molarexcess of glutathione (Fig. 3B, lane5). However when used alone, His-c-Fos or His-c-Jun protein showedno interaction with GST-SAF-1(Fig. 3B, lanes 6 and 7). The AP-1proteins also did not interact withcontrol GST protein (Fig. 3B, lane8). Taken together, these resultsindicated a physical interactionbetween SAF-1 and AP-1 proteinsand further suggested that, in solu-tion, only heterodimers of c-Fos/c-Jun interact efficiently with SAF-1.In Vivo Interaction between SAF-1
and AP-1—To detect the physicalinteraction between SAF-1 andAP-1 in vivo, coimmunoprecipita-tion experiments were performed.The SAF-1 antibody coimmunopre-cipitated c-Fos and c-Jun proteinsalong with SAF-1 in both untreatedand IL-1�-treated cells, albeit at dif-ferent levels (Fig. 4A). In a recipro-cal experiment, SAF-1 protein wasimmunoprecipitated by a mixtureof c-Fos and c-Jun antibodies fromboth untreated and IL-1�-treatedcells, indicating in vivo interactionbetween SAF-1 and members of theAP-1 family. The specificity of thisassay was verified by using controlIgG, which showed no coimmuno-precipitation of c-Fos, c-Jun, orSAF-1. For further verification, weperformed ChIP analysis usinguntreated and IL-1�-treated cells(Fig. 4B). PCR amplification of theAP-1-binding element (Fig. 4B, lane6) and the SAF-1-binding element(Fig. 4B, lane 12) in a cross-linkedchromatin assay of IL-1�-stimu-lated cells indicated active interac-tion of a SAF-1�c-Fos�c-Jun complexwith the MMP-1 promoter at boththe SAF-1 and AP-1 sites duringIL-1� stimulation of cells. Inunstimulated cells, the interactionbetween SAF-1 and AP-1 proteins
seemed to be much less, as the intensity of PCR-amplifiedproducts was low compared with the products obtained fromIL-1�-stimulated cell chromatin DNA (Fig. 4B, comparelanes 3 with 6 and 9 with 12). The specificity of the ChIPassay was verified using control IgG-precipitated chromatin,where no PCR-amplified product was visible (Fig. 4B, lanes2, 5, 8, and 9).
FIGURE 2. Synergistic activation of MMP-1 promoter by SAF-1 and AP-1 proteins. A, HTB-94 cells werecotransfected with MMP-1CAT reporter plasmid, increasing concentrations of pcDSAF-1, and an equimolarmixture of pcDc-Fos and pcDc-Jun expression plasmid DNAs as indicated. CAT activity was determined fromcells harvested 24 h after transfection. The results shown are representative of three separate experiments.B, detection of SAF-1, c-Jun, and c-Fos expression in transfected cells. Equal amount of cell extracts from HTB-94cells transfected with increasing concentrations of pcDSAF-1 (lanes 1–3), pcDc-Jun (lanes 4 – 6), and pcDc-Fos(lanes 7–9) plasmids were immunoblotted with antibodies against SAF-1, c-Jun, and c-Fos as indicated. C, sche-matic showing physical maps of four reporter plasmids containing either wild-type or specific mutation ateither AP-1 or SAF-1 or at both AP-1 and SAF-1 sites as indicated. HTB-94 cells were transfected in duplicatewith these reporter plasmids. Following transfection, one set of cells was incubated with IL-1� (500 units/ml)for an additional 24 h. CAT activity was determined. These results represent an average of three separateexperiments.
FIGURE 3. Physical interaction of SAF-1 with AP-1 in vitro. A, Far-Western assay. Equal amounts (50 pmol) ofpurified bacterially expressed GST (lane 1), His-c-Fos (lane 2), and His-c-Jun (lane 3) proteins were separated electro-phoretically and transferred to a nitrocellulose membrane. The blot was hybridized with a radioactive 32P-labeledpAR-SAF-1 protein and subjected to autoradiography. Lanes 4 – 6 show Coomassie Blue staining of GST (lane 4),His-c-Fos (lane 5), and His-c-Jun (lane 6) proteins. B, GST pulldown assay. GST-SAF-1-(1–477) protein (50 pmol)immobilized on glutathione-Sepharose 4B beads was incubated separately with an equimolar mixture of His-c-Fosand His-c-Jun proteins (lane 5), His-c-Fos protein (lane 7), or His-c-Jun protein (lane 6). GST protein (50 pmol) immo-bilized on glutathione-Sepharose 4B beads was incubated with an equimolar mixture of His-c-Fos and His-c-Junproteins (lane 8). Equal portions of eluted proteins from these columns were analyzed by SDS-PAGE and visualizedby Coomassie Blue staining. Lane 1, 0.5% input GST-SAF-1; lane 2, 0.5% input His-c-Jun; lane 3, 0.5% input His-c-Fos;lane 4, 0.5% input His-c-Fos and His-c-Jun. Samples analyzed in lanes 5– 8 have been described under “ExperimentalProcedures.” His-c-Fos and His-c-Jun protein alone did not interact with GST-SAF-1 (lanes 6 and 7) or together withGST protein (lane 8), whereas His-c-Fos and His-c-Jun proteins together interacted with GST-SAF-1 (lane 5). Theseresults represent the findings from three similar experiments.
Interaction between SAF-1 and AP-1
JANUARY 16, 2009 • VOLUME 284 • NUMBER 3 JOURNAL OF BIOLOGICAL CHEMISTRY 1857
by guest on June 3, 2016http://w
ww
.jbc.org/D
ownloaded from

The interaction of transcription factors to a gene promoter isof a dynamic nature. Therefore, to examine whether SAF-1 andAP-1 proteins are simultaneously localized in theMMP-1 pro-moter, we performed a re-ChIP assay. The cross-linked chro-matin from untreated and IL-1�-treated cells was immunopre-cipitated with the first antibody, and the resultingimmunocomplex was then eluted and further subjected toimmunoprecipitationwith the second antibody or control anti-body. The re-ChIP results indicate that the proximal promoterregion of MMP-1 is occupied by SAF-1 and AP-1 proteins onlyunder conditions of IL-� stimulation (Fig. 4C, lanes 4 and 10).Very little to almost no interaction between SAF-1 and AP-1proteins was detected in unstimulated cells (Fig. 4C, lanes 1 and7). Also, no PCR-amplified product was seen when re-Chipanalysis was performed using control IgG (Fig. 4C, lanes 2, 5, 8,and 11). Taken together, these results indicated the simultane-ous recruitment of SAF-1 and AP-1 to the MMP-1 proximalpromoter region and suggested that they may participate incytokine induction of theMMP-1 gene in vivo.Ternary Complex of SAF-1�c-Fos�c-Jun Shows Markedly
Higher DNA Binding Ability—Protein-protein interactionbetween SAF-1 and c-Fos/c-Jun proteins prompted us to exam-ine the DNA binding activity of the SAF-1�c-Fos�c-Jun ternarycomplex. In the first experiment, the efficacy of the DNA bind-ing activity of bacterially expressed proteins was tested. Alone,bacterially expressed His-c-Fos (Fig. 5B, lane 1) and His-c-Jun(Fig. 5B, lane 2) interacted very poorly with the radioactiveAP-1 element of theMMP-1 promoter, but an equimolar mix-ture of c-Fos/c-Jun proteins showed a considerable level ofDNA-protein interaction with the same probe (Fig. 5B, lane 3).These results are in correlation with other previous observa-tions that reported optimal DNA binding of the AP-1 elementwith only the c-Fos/c-Jun dimer. The c-Fos/c-JunDNA-proteincomplexes were not competed by a nonspecific oligonucleotidebut efficiently competed with a molar excess of nonradioactiveAP-1-binding oligonucleotide (Fig. 5B, lanes 4 and 5). Havingdetermined theDNAbinding efficiency of bacterially expressedproteins, we included an equal amount of bacterially expressedpurified GST-SAF-1 protein in the DNA binding assays. GST-SAF-1 protein did not interact with the radioactive AP-1 ele-ment (Fig. 5B, lane 6) but, together with His-c-Fos and His-c-Jun proteins, formed a very prominent SAF-1�c-Fos�c-JunDNA-protein complex (Fig. 5B, lane 8). The level of SAF-1�c-Fos�c-Jun complex was much higher than the c-Fos�c-Jun com-plex formed with the same AP-1 probe (Fig. 5B, compare lanes7 and 8). The specificity of the ternary complex was verified byusing anti-SAF-1 antibody, which caused a reduction in thelevel of the SAF-1�c-Fos�c-Jun ternary complex (Fig. 5B, lane 9),and by a mixture of c-Fos and c-Jun antibodies, which causedsupershifting of the complex (Fig. 5B, lane 10).In a reciprocal experiment, the extent of SAF-1�c-Fos�c-Jun
ternary complex formation at the SAF-1DNA-binding elementwas examined (Fig. 5E). The interaction of GST-SAF-1 proteinwith the radiolabeled SAF-1 element was found to be modest,and there was no interaction of c-Fos and c-Jun proteins (Fig.5E, lanes 1 and 2). However, when the same SAF-1 protein wasmixed with c-Fos and c-Jun proteins, it interacted with theprobe with much higher affinity and formed two DNA-protein
FIGURE 4. In vivo interaction of SAF-1 with c-Fos and c-Jun. A, HTB-94 cellswere treated with or without IL-1� (500 units/ml) for 24 h. Equal amounts of celllysates were immunoprecipitated (IP) with anti-SAF-1 antibody, normal IgG, or amixture of anti-c-Fos and anti-c-Jun antibodies as indicated. A portion of theinput and immunoprecipitates was then separated by 11% SDS-PAGE, trans-ferred to nitrocellulose membrane, and Western blotted (WB) with anti-c-Jun,anti-c-Fos, or anti-SAF-1 antibody as indicated. These results represent the find-ing from three similar experiments. B, ChIP analysis. HTB-94 cells were treatedwith or without IL-1� (500 units/ml) for 24 h, and the cells were cross-linked with1% formaldehyde. Cell lysates were prepared, and equal amounts of cell lysateswere immunoprecipitated with SAF-1 antibody (Ab; lanes 3 and 6), a mixture ofanti-c-Fos and anti-c-Jun antibodies (lanes 9 and 12), or control IgG (lanes 2, 5, 8,and 11) as indicated. Immunoprecipitated DNA was used for PCR amplification ofthe SAF-1 or AP-1 element in the human MMP-1 gene promoter using specificprimers. Lanes 1, 4, 7, and 8 represent PCR amplification of 10% of input DNA.Amplified products were resolved in 1.5% agarose gel and visualized by ethidiumbromide staining. C, re-ChIP analysis. Chromatin was prepared from HTB-94 cellstreated without (lanes 1, 2, 7, and 8) or with IL-1� (lanes 4, 5, 10, and 11) for 24 h.ChIP was first performed using anti-SAF-1 antibody (lanes 1, 2, 4, and 5) or a mix-tureofanti-c-Fosandanti-c-Junantibodies(lanes 7, 8, 10, and 11)as indicated.Theeluant of each immunocomplex was subjected to further immunoprecipitationusing anti-c-Fos and anti-c-Jun antibodies (lanes 1 and 4), anti-SAF-1 antibody(lanes 7 and 10), or control IgG (lanes 2, 5, 8, and 11) as indicated. The precipitatedchromatin DNA was used for PCR amplification using a pair of primers flankingthe SAF-1 or AP-1 element in human MMP-1 gene promoter. Lanes 3, 6, 9, and 12represent PCR amplification of 10% of input DNA from unstimulated and IL-1-treated cells as indicated. Amplified products were resolved in a 1.5% agarose geland visualized by ethidium bromide staining.
Interaction between SAF-1 and AP-1
1858 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 3 • JANUARY 16, 2009
by guest on June 3, 2016http://w
ww
.jbc.org/D
ownloaded from

complexes, one of which was composed of SAF-1�c-Fos�c-Jun and the other of SAF-1�SAF-1 proteins (Fig. 5E, lane 3).The SAF-1�c-Fos�c-Jun DNA ternary complex was super-shifted by c-Fos/c-Jun antibodies, whereas the SAF-1/SAF-1complex remained unaffected ((Fig. 5E, lane 5). The additionof a molar excess of SAF-1 oligonucleotide (Fig. 5E, lane 6) oranti-SAF-1 antibody (Fig. 5E, lane 7) substantially inhibitedthe formation of both DNA-protein complexes. Theseresults showed that the SAF-1�c-Fos�c-Jun ternary complexcan bind to either a SAF-1 or an AP-1 DNA element with amarkedly higher efficiency.Functional Interaction of SAF-1 with c-Fos/c-Jun Promotes
Synergistic Transcriptional Activation from SAF-1 or AP-1Element—To correlate whether increased DNA binding of theSAF-1�c-Fos�c-Jun ternary complex leads to increased an tran-scriptional function, a series of transfection assays was per-formed. Simultaneous expression from SAF-1, c-Fos, and c-Junplasmids synergistically increased MMP-1(�518/�63)-CAT,AP-1-CAT, or SAF-1-CAT reporter activity (Fig. 6A). Exoge-nous expression of SAF-1, c-Fos, and c-Jun proteins in trans-fected cells was verified by Western blot analysis (Fig. 6B),which showed no discrepancy in the expression pattern oftransfected plasmids. There was a modest increase in theMMP-1 protein level in pcDSAF-1/pcDc-Fos/pcDc-Jun plas-mid-transfected cells (Fig. 6B). These reporter assays clearlyindicated that the synergism between SAF-1 and c-Fos/c-Junproteins can be achieved by the engagement of the ternary com-
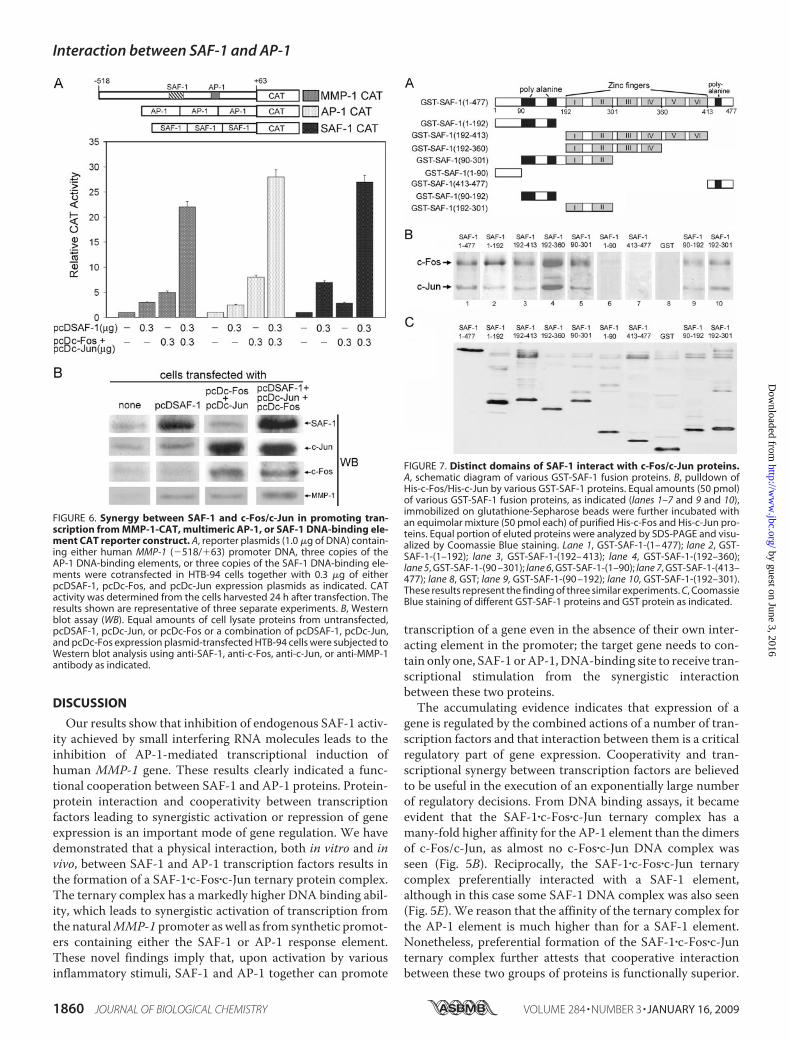
plex at either the AP-1 or SAF-1 element and further suggestedthat synergism between SAF-1 andAP-1 proteinsmay be appli-cable to other genes that contain only one DNA-binding ele-ment, SAF-1 or AP-1, in their promoters.Mapping of the AP-1 Protein-interacting Domains in SAF-1—
To define the domains of SAF-1 involved in interaction withc-Fos/c-Jun proteins, a series ofGST fusion proteins containingdifferent regions of SAF-1 was prepared (Fig. 7A). Among dif-ferent SAF-1 proteins, fragments 1–477, 1–192, 192–413,192–360, and 90–301 showed a good interaction with AP-1proteins (Fig. 7B, lanes 1–5), whereas the two terminal 1–90and 413–477 fragments and negative control GST protein didnot show any interaction with the c-Fos/c-Jun protein mixture(Fig. 7B, lanes 6–8). Further delineation of the domains indi-cated the presence of two independent AP-1-interacting mod-ules in SAF-1 (Fig. 7B, lanes 9 and 10). One domain is centeredwithin the two N-terminal polyalanine tracts (amino acids90–192), and the other domain resides within the first two zincfingers (amino acids 192–301). Coomassie Blue staining wasperformed to verify the input and integrity of different GST-SAF-1 proteins (Fig. 7C). It was interesting to note that theSAF-1-(90–301) fragment containing both interactingdomains displayed a somewhat lower level of interaction withAP-1 proteins than the SAF-1-(192–360) fragment. Nonethe-less, these results clearly mapped the AP-1 protein-interactingdomains of SAF-1.
FIGURE 5. Superior DNA binding ability of the ternary complex of c-Fos, c-Jun, and SAF-1. A, the sequence of the AP-1 DNA-binding element in the humanMMP-1 promoter used as probe in DNA binding assays. B, purified bacterially expressed His-c-Fos, His-c-Jun, and GST-SAF-1 proteins were incubated singly orin combination with 32P-labeled AP-1 oligonucleotide as indicated. In some reactions, a molar excess of unlabeled nonspecific oligonucleotide (lane 4) or ofunlabeled AP-1 oligonucleotide (lane 5), anti-SAF-1 antibody (Ab; lane 9), or a mixture of anti-c-Fos and anti-c-Jun antibodies (lane 10) was included. TheSAF-1�c-Fos�c-Jun ternary complex, c-Fos�c-Jun, c-Jun�c-Jun, and supershifted complexes are indicated. C, Western blot analysis to determine the input ofproteins in the DNA binding assay. Purified bacterially expressed His-c-Fos, His-c-Jun, and GST-SAF-1 proteins, as used in the DNA binding assays, weresubjected to Western blot analysis using anti-c-Fos, anti-c-Jun, and anti-SAF-1 antibodies as indicated. D, the sequence of the SAF-1 DNA-binding element inthe human MMP-1 promoter used as a probe in the DNA binding assays. E, purified bacterially expressed GST-SAF-1, His-c-Fos, and His-c-Jun proteins, singly orin combination, were incubated with 32P-labeled SAF-1 oligonucleotide as indicated. In some reactions, a molar excess of unlabeled nonspecific oligonucleo-tide (lane 4), a mixture of anti-c-Fos and anti-c-Jun antibodies (lane 5), a molar excess of unlabeled SAF-1 oligonucleotide (lane 6), or anti-SAF-1 antibody (lane7) were included. The SAF-1�c-Fos�c-Jun ternary complex, SAF-1�SAF-1 complex, and the supershifted complex are indicated.
Interaction between SAF-1 and AP-1
JANUARY 16, 2009 • VOLUME 284 • NUMBER 3 JOURNAL OF BIOLOGICAL CHEMISTRY 1859
by guest on June 3, 2016http://w
ww
.jbc.org/D
ownloaded from
DISCUSSION
Our results show that inhibition of endogenous SAF-1 activ-ity achieved by small interfering RNA molecules leads to theinhibition of AP-1-mediated transcriptional induction ofhuman MMP-1 gene. These results clearly indicated a func-tional cooperation between SAF-1 and AP-1 proteins. Protein-protein interaction and cooperativity between transcriptionfactors leading to synergistic activation or repression of geneexpression is an important mode of gene regulation. We havedemonstrated that a physical interaction, both in vitro and invivo, between SAF-1 and AP-1 transcription factors results inthe formation of a SAF-1�c-Fos�c-Jun ternary protein complex.The ternary complex has a markedly higher DNA binding abil-ity, which leads to synergistic activation of transcription fromthe naturalMMP-1 promoter as well as from synthetic promot-ers containing either the SAF-1 or AP-1 response element.These novel findings imply that, upon activation by variousinflammatory stimuli, SAF-1 and AP-1 together can promote
transcription of a gene even in the absence of their own inter-acting element in the promoter; the target gene needs to con-tain only one, SAF-1 orAP-1, DNA-binding site to receive tran-scriptional stimulation from the synergistic interactionbetween these two proteins.The accumulating evidence indicates that expression of a
gene is regulated by the combined actions of a number of tran-scription factors and that interaction between them is a criticalregulatory part of gene expression. Cooperativity and tran-scriptional synergy between transcription factors are believedto be useful in the execution of an exponentially large numberof regulatory decisions. From DNA binding assays, it becameevident that the SAF-1�c-Fos�c-Jun ternary complex has amany-fold higher affinity for the AP-1 element than the dimersof c-Fos/c-Jun, as almost no c-Fos�c-Jun DNA complex wasseen (Fig. 5B). Reciprocally, the SAF-1�c-Fos�c-Jun ternarycomplex preferentially interacted with a SAF-1 element,although in this case some SAF-1 DNA complex was also seen(Fig. 5E). We reason that the affinity of the ternary complex forthe AP-1 element is much higher than for a SAF-1 element.Nonetheless, preferential formation of the SAF-1�c-Fos�c-Junternary complex further attests that cooperative interactionbetween these two groups of proteins is functionally superior.
FIGURE 6. Synergy between SAF-1 and c-Fos/c-Jun in promoting tran-scription from MMP-1-CAT, multimeric AP-1, or SAF-1 DNA-binding ele-ment CAT reporter construct. A, reporter plasmids (1.0 �g of DNA) contain-ing either human MMP-1 (�518/�63) promoter DNA, three copies of theAP-1 DNA-binding elements, or three copies of the SAF-1 DNA-binding ele-ments were cotransfected in HTB-94 cells together with 0.3 �g of eitherpcDSAF-1, pcDc-Fos, and pcDc-Jun expression plasmids as indicated. CATactivity was determined from the cells harvested 24 h after transfection. Theresults shown are representative of three separate experiments. B, Westernblot assay (WB). Equal amounts of cell lysate proteins from untransfected,pcDSAF-1, pcDc-Jun, or pcDc-Fos or a combination of pcDSAF-1, pcDc-Jun,and pcDc-Fos expression plasmid-transfected HTB-94 cells were subjected toWestern blot analysis using anti-SAF-1, anti-c-Fos, anti-c-Jun, or anti-MMP-1antibody as indicated.
FIGURE 7. Distinct domains of SAF-1 interact with c-Fos/c-Jun proteins.A, schematic diagram of various GST-SAF-1 fusion proteins. B, pulldown ofHis-c-Fos/His-c-Jun by various GST-SAF-1 proteins. Equal amounts (50 pmol)of various GST-SAF-1 fusion proteins, as indicated (lanes 1–7 and 9 and 10),immobilized on glutathione-Sepharose beads were further incubated withan equimolar mixture (50 pmol each) of purified His-c-Fos and His-c-Jun pro-teins. Equal portion of eluted proteins were analyzed by SDS-PAGE and visu-alized by Coomassie Blue staining. Lane 1, GST-SAF-1-(1– 477); lane 2, GST-SAF-1-(1–192); lane 3, GST-SAF-1-(192– 413); lane 4, GST-SAF-1-(192–360);lane 5, GST-SAF-1-(90 –301); lane 6, GST-SAF-1-(1–90); lane 7, GST-SAF-1-(413–477); lane 8, GST; lane 9, GST-SAF-1-(90 –192); lane 10, GST-SAF-1-(192–301).These results represent the finding of three similar experiments. C, CoomassieBlue staining of different GST-SAF-1 proteins and GST protein as indicated.
Interaction between SAF-1 and AP-1
1860 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 3 • JANUARY 16, 2009
by guest on June 3, 2016http://w
ww
.jbc.org/D
ownloaded from

The AP-1 family of proteins is shown to interact with a varietyof other transcription factors, including NF-�B (32), C/EBP(33), Ets (34), Cbfa1 (35), and STAT-3 (36). In comparison,reports on SAF-1 interacting with other proteins are not soabundant. In yeast two-hybrid assays, SAF-1 was shown tointeract with the catalytic subunit of protein kinase A (37).Another report (15) indicates a functional cooperation betweenSAF-1 and Sp1 leading to synergistic activation of the serumamyloidA gene.However, in this studywe found no evidence ofa physical interaction between SAF-1 and Sp1.The nature of cooperative interaction was further defined
fromvarious transient transfection assays (Figs. 2 and 6). Partialinhibition of cytokine-induced transcription ofmutantMMP-1promoters containing altered AP-1 or SAF-1 element or acti-vation of synthetic promoters driven from multimeric AP-1 orSAF-1 elements clearly showed that a SAF-1�AP-1 complex canexert a biological function through either of these response ele-ments. However, in an earlier study we observed a somewhatdifferent form of cooperativity between SAF-1 and AP-1 pro-teins in regulating humanMMP-9 gene expression (38). In theMMP-9 promoter SAF-1 andAP-1 elements are present in veryclose proximity, placed within 14 nucleotides of each other.Mutation of either SAF-1 or AP-1 element markedly affectscytokine-mediated induction of theMMP-9 promoter and alsocompromises the ability of SAF-1 and AP-1 to activate tran-scription from mutant MMP-9 promoters (38). Such differentresults, perhaps a gene-specific effect, could be explained bytaking into consideration (i) the close spacing between SAF-1and AP-1 that may have created a certain helical arrangementand steric hindrance and/or (ii) the participation of other pro-teins interacting at nearby regions in the human MMP-9 pro-moter. For example, a specific helical phasing is shown to facil-itate the physical interaction between AP-1 with Cbfa1transcription factors in regulating collagenase-3 promoterexpression (39) and between STAT3 and AP-1 in regulating�2-macroglobulin expression (40). However, this mechanismof cross-talk, in which two assembled transcription factors arecapable of contacting the promoter DNA via a single DNA-binding element, is not highly unprecedented. Notable exam-ples include cross-coupling of the NF-�B/p65 and Fos/Jun-po-tentiated biological function via both the �B andAP-1 responseelements (32) and concerted participation of C/EBP andNF-�Bresulting in synergistic transcriptional activation through bothC/EBP and NF-�B sites in the serum amyloid A promoter (41).
Other major findings of this study are that (i) only a c-Fos/c-Jun heterodimer can interact with SAF-1, and (ii) two distinctdomains in SAF-1 are involved in forming the protein-proteininteraction with AP-1 proteins (Fig. 7). It is noteworthy thatalthough in a membrane-bound form (Far-Western assay; Fig.3A), c-Fos or c-Jun alone displayed some interaction withSAF-1, only an equimolar mixture of c-Fos and c-Jun proteinswas capable of interacting with SAF-1 protein in solution. Atpresent, it remains to be determined why homodimers of c-Fosand c-Jun failed to interact with SAF-1. The AP-1-interactingdomains in SAF-1 are present within two N-terminal polyala-nine tracts and the first two zinc fingers. Homopolymericamino acid repeats, such as polyalanine, polyglutamine,polyproline tracts are shown to modulate protein-protein
interaction and/or transcriptional regulation (42). A novel sta-ble polyalanine tract expansion in theHOXA13 gene is believedto prevent the protein-protein interaction associated withdominantly inherited hand-foot-genital syndrome disease (43).However, the most interaction between SAF-1 and AP-1 wasnoted with a SAF-1 fragment containing the first four zinc fin-gers (amino acid residues 192–360) or the DNA-bindingdomain of SAF-1. The DNA-binding domains are used forinteracting with the promoter; so, to put it simplistically, ifthese regions are used for protein-protein interaction, thatmayinterfere with DNA-protein interaction and theoreticallyshould lead to a transcriptional repression. It is thereforeintriguing to note that engagement of the SAF-1 DNA-bindingdomain by c-Fos/c-Jun results in a synergistic transcriptionalactivation, which suggests that the ternary complex may con-tain some exposed domains that would allow DNA-proteininteraction to occur. Further studies of the SAF-1 protein struc-ture, such as x-ray crystallography, would be necessary toexplain how the use of a DNA-binding domain for protein-protein interaction could still lead to transcriptional activation.In support of our results, another report indicates protein-pro-tein interaction between the Ets transcription factor, throughits DNA-binding domain, with c-Fos/c-Jun proteins resultingin activation of gene transcription (44).In summary, we have provided a molecular basis for a better
understanding of synergistic gene regulation by SAF-1 andAP-1 transcription factors, which in addition to the MMP-1promoter may be applicable to other genes that contain thesetranscription factor DNA-binding elements. It remains to beseenwhether other factors capable of interacting with SAF-1 orAP-1 also participate in forming multimeric transcriptomecomplexes, thereby allowing sequential enhancement andmax-imal target gene expression in response to different stimuli.
Acknowledgments—We thank Gabriela Aust and James A. Goodrichfor generous gifts of the MMP-1 promoter and pET-cJun/pET-cFosplasmid DNA samples. We also thank Michael Blanar for providingbacterial expression plasmid pAR(�R1)59/60.
REFERENCES1. Ehrlich, M. G. (1985) J. Orthop. Res. 3, 170–1842. Berend, K. R., Toth, A. P., Harrelson, J. M., Layfield, L. J., Hey, L. A., and
Scully, S. P. (1998) J. Bone Jt. Surg. Am. 80, 11–173. Galis, Z. S., Sukhova, G. K., Kranzhofer, R., Clark, S., and Libby, P. (1995)
Proc. Natl. Acad. Sci. U. S. A. 92, 402–4064. Visse, R., and Nagase, H. (2003) Circ. Res. 92, 827–8395. Birkedal-Hansen, H. (1995) Curr. Opin. Cell Biol. 7, 728–7356. Birkedal-Hansen, H., Moore, W. G., Bodden, M. K., Windsor, L. J.,
Birkedal-Hansen, B., DeCarlo, A., and Engler, J. A. (1993) Crit. Rev. OralBiol. Med. 4, 197–250
7. Angel, P., Baumann, I., Stein, B., Delius, H., Rahmsdorf, H. J., andHerrlich,P. (1987)Mol. Cell. Biol. 7, 2256–2266
8. Ray, A., Kuroki, K., Cook, J. L., Bal, B. S., Kenter, K., Aust, G., and Ray, B. K.(2003) Arthritis Rheum. 48, 134–145
9. Shaulian, E., and Karin, M. (2002) Nat. Cell Biol. 4, E131–E13610. Ransone, L. J., and Verma, I. M. (1990) Annu. Rev. Cell Biol. 6, 539–55711. Hipskind, R. A., Rao, V. N., Mueller, C. G., Reddy, E. S., and Nordheim, A.
(1991) Nature 354, 531–53412. Hill, C. S., Marais, R., John, S., Wynne, J., Dalton, S., and Treisman, R.
(1993) Cell 73, 395–406
Interaction between SAF-1 and AP-1
JANUARY 16, 2009 • VOLUME 284 • NUMBER 3 JOURNAL OF BIOLOGICAL CHEMISTRY 1861
by guest on June 3, 2016http://w
ww
.jbc.org/D
ownloaded from

13. Kampfer, S., Hellbert, K., Villunger, A., Doppler, W., Baier, G., Grunicke,H. H., and Uberall, F. (1998) EMBO J. 17, 4046–4055
14. Ray, A., and Ray, B. K. (1998)Mol. Cell. Biol. 18, 7327–733515. Ray, A., Schatten, H., and Ray, B. K. (1999) J. Biol. Chem. 274, 4300–430816. Ray, B. K., Chatterjee, S., and Ray, A. (1999) DNA Cell Biol. 18, 65–7317. Ray, B. K., and Ray, A. (1997) Biochemistry 36, 4662–466818. Ray, A., Yu, G. Y., and Ray, B. K. (2002)Mol. Cell. Biol. 22, 1027–103519. Ray, A., Fields, A. P., and Ray, B. K. (2000) J. Biol. Chem. 275, 39727–3973320. Ray, A., Ray, P., Guthrie, N., Shakya, A., Kumar, D., and Ray, B. K. (2003)
J. Biol. Chem. 278, 22586–2259521. Ray, A., Shakya, A., and Ray, B. K. (2005) Biochim. Biophys. Acta 1732,
53–6122. Gutman, A., and Wasylyk, B. (1991) Trend. Genet. 7, 49–5423. Fry, C. J., and Farnham, P. J. (1999) J. Biol. Chem. 274, 29583–2958624. Sambrook, J., and Russell, D. W. (2001)Molecular Cloning: A Laboratory
Manual, Ed. 3, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY25. Luckow, B., and Schutz, G. (1987) Nucleic Acids Res. 15, 549026. Ray, A., and Ray, B. K. (1996)Mol. Cell. Biol. 16, 1584–159427. Blanar, M. A., and Rutter, W. J. (1992) Science 256, 1014–101828. Ferguson, H. A., and Goodrich, J. A. (2001) Nucleic Acids Res. 29, E9829. Chomczynski, P., and Sacchi, N. (1987) Anal. Biochem. 162, 156–15930. Bradford, M. M. (1976) Anal. Biochem. 72, 248–25431. Kouskouti, A., and Talianidis, I. (2005) EMBO J. 24, 347–357
32. Stein, B., Baldwin, A. S., Jr., Ballard, D. W., Greene, W. C., Angel, P., andHerrlich, P. (1993) EMBO J. 12, 3879–3891
33. Cai, D. H., Wang, D., Keefer, J., Yeamans, C., Hensley, K., and Friedman,A. D. (2008) Oncogene 27, 2772–2779
34. Basuyaux, J. P., Ferreira, E., Stehelin, D., and Buttice, G. (1997) J. Biol.Chem. 272, 26188–26195
35. Porte, D., Tuckermann, J., Becker, M., Baumann, B., Teurich, S., Higgins,T., Owen, M. J., Schorpp-Kistner, M., and Angel, P. (1999) Oncogene 18,667–678
36. Zhang, X., Wrzeszczynska, M. H., Horvath, C. M., and Darnell, J. E., Jr.(1999)Mol. Cell. Biol. 19, 7138–7146
37. Ray, B. K., Chen, J., and Ray, A. (2001) J. Immunol. 167, 2343–234838. Ray, A., Bal, B. S., and Ray, B. K. (2005) J. Immunol. 175, 4039–404839. D’Alonzo, R. C., Selvamurugan, N., Karsenty, G., and Partridge, N. C.
(2002) J. Biol. Chem. 277, 816–82240. Yoo, J. Y., Wang, W., Desiderio, S., and Nathans, D. (2001) J. Biol. Chem.
276, 26421–2642941. Ray, A., Hannink, M., and Ray, B. K. (1995) J. Biol. Chem. 270, 7365–737442. Hancock, J. M., and Simon, M. (2005) Gene 345, 113–11843. Utsch, B., Becker, K., Brock, D., Lentze,M. J., Bidlingmaier, F., and Ludwig,
M. (2002) Hum. Genet. 110, 488–49444. Bassuk, A. G., and Leiden, J. M. (1995) Immunity 3, 223–237
Interaction between SAF-1 and AP-1
1862 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 284 • NUMBER 3 • JANUARY 16, 2009
by guest on June 3, 2016http://w
ww
.jbc.org/D
ownloaded from

Deepak Kumar, Alpana Ray and Bimal K. RaySAF-1
N-TERMINAL POLYALANINE AND TWO ZINC FINGER DOMAINS OF Transcriptional Synergy Mediated by SAF-1 and AP-1: CRITICAL ROLE OF
doi: 10.1074/jbc.M806289200 originally published online November 21, 20082009, 284:1853-1862.J. Biol. Chem.
10.1074/jbc.M806289200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/284/3/1853.full.html#ref-list-1
This article cites 43 references, 22 of which can be accessed free at
by guest on June 3, 2016http://w
ww
.jbc.org/D
ownloaded from




















