
Consequence of salt, sugar and fat content modifications in ...
Upload
independentCategory
view
1download
0
Top-down control of prey increases with drying
disturbance in ponds: a consequence of
non-consumptive interactions?
Hamish S. Greig1*, Scott A. Wissinger1,2 and Angus R. McIntosh1
1School of Biological Sciences, University of Canterbury, Private Bag 4800, Christchurch, New Zealand; and 2Biology
Department, Allegheny College, Meadville, PA 16335, USA
Summary
1. Biotic interactions are often expected to decrease in intensity as abiotic conditions become
more stressful to organisms. However, in many cases, food-web and habitat complexity also
change with abiotic stress or disturbance, potentially altering patterns of species interactions
across environmental gradients.
2. We used a combination of field assays and mesocosm experiments to investigate how dis-
turbance from desiccation moderates top-down control of prey by predators across a gradient
of pond duration in New Zealand.
3. Field manipulations of predator abundance in ponds led to an unexpected decrease in the
top-down control of prey biomass by predatory invertebrates as pond duration increased
(decreasing abiotic stress). Predatory fish, which are restricted to permanent ponds, had negli-
gible effects on prey biomass. Mesocosm experiments further indicated the consumptive
effects of fish are weak; a result that cannot be explained by an increase in physical habitat
refugia in relatively more permanent ponds.
4. Manipulations of invertebrate predator diversity in mesocosms (both substitutive and addi-
tive treatments), and the addition of olfactory fish cues, revealed that strong non-consumptive
effects of fish reduced predation by predatory invertebrates, and these effects overwhelmed
the positive influence of invertebrate predator diversity on prey consumption.
5. These results suggest that decreases in top-down control with increasing pond permanence
are likely a result of non-consumptive effects of fish weakening predation by invertebrate
predators in the more complex food webs of permanent ponds. Therefore, considering non-
consumptive effects of predators in complex food webs will likely improve the understanding
of biotic interactions across environmental gradients.
Key-words: Food-web complexity, interaction modification, intraspecific competition, multi-
ple predator effects, pond permanence, risk reduction, trait-mediated interactions
Introduction
Understanding how the strength of species interactions
changes with abiotic context is essential for predicting the
dynamics of ecological communities (Agrawal et al. 2007),
especially in response to changes along environmental gra-
dients. Several traditional models of community organiza-
tion predict decreases in the importance and intensity of
biotic interactions as abiotic conditions become more
stressful to organisms (Connell 1975; Grime 1977; Peckar-
sky 1983; Menge & Sutherland 1987). For example,
abiotic stress can mediate competitive interactions by
reducing densities of interacting species below resource
carrying capacities (Lubchenco 1980; Crain et al. 2004;
Gerhardt & Collinge 2007), and can weaken predation by
disproportionately influencing the abundance or effective-
ness of predators (Menge & Farrell 1989; Wellborn,
Skelly & Werner 1996).
Several authors have argued, however, that the effects
of abiotic stress on species interactions should vary
depending on environmental and community context*Correspondence author. E-mail: [email protected]
© 2013 The Authors. Journal of Animal Ecology © 2013 British Ecological Society
Journal of Animal Ecology 2013, 82, 598–607 doi: 10.1111/1365-2656.12042

(Chesson & Huntly 1997; Crain 2008). For example, phys-
ically benign habitats often have higher taxa diversity
within and across trophic levels and wider body size vari-
ation than disturbed habitats, and hence food webs with
more complex connections (Winemiller 1990; Jenkins,
Kitching & Pimm 1992; Walters & Post 2008). Omnivory,
intraguild predation (IGP) and non-consumptive effects
characteristic of these complex food webs weaken pair-
wise species interactions (Emmerson & Yearsley 2004;
Finke & Denno 2004); whereas the low diversity and
chain-like resource pathways of simple food webs charac-
teristic of disturbed or stressful habitats can strengthen
interactions (Power, Parker & Wootton 1996; Thompson
& Townsend 1999). Furthermore, organisms inhabiting
physically stressful environments often face life-history
constraints that force individuals to trade-off resistance to
biotic interactions with rapid development or efficient
resource use (Power, Parker & Wootton 1996; Wissinger
et al. 2006; Edwards & Stachowicz 2010). These trade-offs
may strengthen top-down control in disturbed habitats.
Finally, changes in habitat morphology along environ-
mental gradients such as habitat size, complexity and
refugia further complicate the community-wide outcome
of biotic interactions across environmental gradients.
Studies that incorporate these parallel and potentially
confounding community and habitat gradients are likely
to reveal more realistic effects of abiotic stress on species
interactions.
Here, we use a combination of field and mesocosm
experiments to investigate how shifts in food-web com-
plexity and habitat structure affect aquatic predator–prey
interactions across a gradient of pond drying disturbance.
The duration and frequency of drying and refilling vary
among individual ponds in a landscape, resulting in a
well-known gradient of abiotic stress across habitat
patches (Wellborn, Skelly & Werner 1996; Williams 1996).
Predator body size and diversity increases with pond
duration and fish are generally restricted to perennial
ponds (Schneider & Frost 1996; Bilton, Foggo & Rundle
2001). Subsequent increases in predation risk with pond
duration are often implicated in species turnover across
the pond permanence gradient (reviewed in Wellborn,
Skelly & Werner 1996; Wissinger 1999). However, pond
area, the abundance of macrophyte refugia (Urban 2004;
McAbendroth et al. 2005) and food-web complexity
(Wissinger et al. 1999) all increase with pond duration,
which may instead weaken community-wide predator–
prey interactions in more permanent ponds. Therefore, if
some predators are able to exploit temporary ponds
through rapid colonization or senescence, patterns of top-
down control of prey biomass across the habitat perma-
nence gradient may be reversed to be most intense in
short duration ponds (Brendonck et al. 2002).
We experimentally manipulated predator biomass in
natural ponds spanning a gradient of permanence to
investigate the influence of drying stress on the strength
of top-down control of prey biomass. We then conducted
two mesocosm experiments to distinguish the mechanisms
driving observed patterns of top-down control across the
natural pond permanence gradient. We first manipulated
the presence of macrophytes and predatory fish in meso-
cosms to examine the potential role of refugia in mediat-
ing predation on benthic invertebrates in permanent
ponds. A second experiment then used additive and sub-
stitutive manipulations of predator diversity (within and
between trophic levels) to determine the influence of
increasing food-web complexity on the strength of
top-down control.
Materials and methods
study sites and natural history
Lentic habitats in the Waimakariri and Rakaia River catchments,
South Island, New Zealand range in size from 2-m diameter
ephemeral pools to large lakes (up to 179 ha) that lie within
fluvio-glacial depressions (Wissinger, Greig & McIntosh 2009).
Pond hydroperiod (duration of inundation) strongly influences
species richness and community composition in the ponds, and
predator species richness, size and biomass increase with pond
permanence (Wissinger, Greig & McIntosh 2009). Most perma-
nent ponds contain small (< 200 mm) predatory benthic fish (Gal-
axias brevipinnis G€unther, Gobiomorphus breviceps Stokell), and
the larger lakes support low-density populations of longfin eels
(Anguilla dieffenbachia Grey), introduced brown (Salmo trutta L.)
and rainbow trout (Onchorynchus mykiss Walbaum) (Jeppesen
et al. 1997; Wissinger, McIntosh & Greig 2006). The biomass of
predatory invertebrate guilds is dominated by odonates in perma-
nent ponds, and beetles and small Anisops spp. backswimmers in
temporary habitats (Wissinger, Greig & McIntosh 2009).
assays of predator impact in natural ponds
Twelve ponds were selected for predator manipulations based on
their likelihood of retaining water throughout the duration of
experiments, and the presence of submerged vegetation in shallow
water (<60 cm deep). Five of those ponds contained predatory
fish (Table S1, Supporting Information). Visual monitoring of
pond hydrology began March 2005 when temporary ponds were
dry and from 12 October 2005 water depth was recorded hourly
in each pond with stage height data loggers (HT-100; TruTrack
Ltd., Christchurch, New Zealand) placed in the deepest possible
point of the pond basin. Pond permanence was quantified with a
multivariate index derived from principal components analysis of
three aspects of pond hydrology: duration (days) of inundation
following winter refilling, maximum proportion of total depth
lost over the observation period, and number of days inundated
per annum (to account for multiple drying and refilling events;
Table S1, Supporting Information).
In November 2005, we manipulated predator biomass in cages
(1�5 mm mesh on a steel wire frame, 0�25 m2 surface area, 50 cm
height) whose bottom edges were pushed into the sediment within
macrophyte beds and sealed with clay and fine gravel. Three
cages were placed 1 m apart in shallow water (mean depth � SE:
26 � 3 cm) in each pond, with each cage representing one repli-
cate of three different treatments: a ‘all predator reduction’ where
all fish and all predatory invertebrates captured in five sweeps
© 2013 The Authors. Journal of Animal Ecology © 2013 British Ecological Society, Journal of Animal Ecology, 82, 598–607
Disturbance increases top-down control 599

were removed from the cages, a ‘fish removal’, and an open ‘con-
trol’ cage (steel frame only) that allowed access to both predatory
fish and invertebrates. Predators were removed with five succes-
sive sweeps of a 1-mm mesh D-net through the water column
and benthic substrates. The contents of each sweep were trans-
ferred to sorting trays, predatory taxa were removed by hand,
and the remaining invertebrates and detritus were returned to the
cage. This procedure left submergent vegetation intact and
removed all fish biomass and 56 � 10% (mean � SE) of preda-
tory invertebrate biomass when compared with cages without
predator removal. The efficacy of predator removal was not influ-
enced by pond permanence (linear regression on % predator bio-
mass reduction, F1,10 = 1�74, P = 0�24). Predatory invertebrates
were classified as species that obtain the majority of their energy
through the consumption of other macroscopic animals. Small-
bodied predators that could move through the cage mesh (Liodes-
sus spp. beetles, mites and cyclopoid copepods) were not removed
in the manipulation. To control for prey mortality during the
predator manipulations, open-sided cages which represented
ambient levels of predation were covered in a temporary sleeve of
mesh and disturbed using the same procedures as above but with-
out the removal of predators.
After 2 weeks, small-bodied invertebrates (crustaceans, chiron-
omids, worms and molluscs) were subsampled in each cage with
five sweeps of a 500-lm mesh net through a PVC pipe (0�020 m2)
pushed into the substrate. Those subsamples were preserved in
70% ETOH and sorted in the laboratory under 109 magnifica-
tion. The remaining taxa in the cage were sampled with five
sweeps of a 1-mm mesh D-net, sorted on-site and preserved in
70% ETOH. Invertebrates were identified to tribe for Chironomi-
dae, family or genus for Crustacea, and to the lowest possible
taxonomic level (usually species) for remaining taxa. Biomass for
each taxon was calculated using length-dry weight regressions
(Benke et al. 1999; Stoffels, Karbe & Paterson 2003; H. S. Greig
unpublished data) on the body length measurements (ocular
micrometre at 10–209 magnification) of a random subset of 10
individuals from each sample. Dry weights were converted to
ash-free dry mass (AFDM) using taxon-specific estimates of %
ash (Benke et al. 1999, H. S. Greig unpublished data).
Predator impact (PI) on prey biomass was calculated using log
ratio of effect size (Berlow et al. 2004) calculated from the ratio
of prey biomass in unmanipulated predator treatments to prey
biomass in predator reduction treatments (Table 1). Fish PI was
determined by comparing invertebrate biomass in the open con-
trol cages with invertebrate biomass in the fish removal cages.
Predatory invertebrate PI was determined by comparing non-
predatory invertebrate biomass in fish removal cages with that
from the all predator reduction cages. Finally, comparisons of
non-predatory biomass between the open control cage and the all
predator reduction cages produced total PI (Table 1).
The effect of fish on prey biomass in permanent habitats was
analysed by comparing deviations of predator impact from zero
using a one-sample t-test for each of the three prey categories: all
invertebrates, all non-predatory invertebrates and ‘unprotected’
non-predators that did not have cases or shells. The effect of
pond permanence on the strength of predatory invertebrate
impact on all primary consumers and unprotected primary con-
sumers was analysed with linear regression. Finally, we used lin-
ear regression on the biomass of predatory invertebrates in
unmanipulated control cages to assess whether ambient predator
biomass was influenced by pond permanence.
mesocosm experiment 1: manipulation ofpredatory fish and submergent vegetation
Whether habitat refugia could explain the apparent weak con-
sumptive effects of fish in permanent ponds was examined by
manipulating the presence and/or absence of complex vegetation
structure and predatory fish in mesocosms (0�8 m2 oval tanks) in
a 2 9 2 factorial design with four replicates of each treatment
combination. The outdoor mesocosms, which were housed at
University of Canterbury’s Cass Field Station, were filled with
groundwater regulated to 25 cm deep throughout the experiment.
Clumps of Myriophyllum and Carex (surface area: 0�1 m2) were
added to the mud and fine gravel substrate of all tanks, and were
cut to soil level for the no-vegetation treatment. Koaro (Galaxias
brevipinnis) were chosen as predators as they are an abundant
native predatory fish in ponds and lake littoral zones in the study
area and they feed on a large range of macroinvertebrates (Rowe,
Konui & Christie 2002).
The prey community in each mesocosm consisted of 11 macro-
invertebrate species that are common in nearby permanent and
temporary ponds (Fig. S1, Supporting Information). The total
biomass of each prey taxa was approximately equal, and com-
bined biomass of all prey fell within the range observed in natu-
ral ponds (H. S. Greig unpublished data). Prey were added on
the 28th February 2007 10 h prior to the addition of fish (one
koaro per tank, mean fork length: 120 � 5�4 mm). After 2 weeks,
fish were removed from tanks and one pipe sample (0�020 m2)
was taken within the largest clump of Myriophyllum to subsample
small-bodied prey (chironomids and crustaceans) in the same
fashion as the predator assay. Following this procedure, all tank
contents were tipped into a 1-mm mesh net whose contents were
then transferred to a tray for sorting. Soil clumps were also
searched for macroinvertebrates. Invertebrates were preserved
and processed in the same manner as the predator assay.
The effects of fish and vegetation structure on total prey bio-
mass were evaluated using 2 9 2 factorial ANOVA with fish and
vegetation as fixed effects and loge-transformed total invertebrate
Table 1. Derivation of predatory impact indices for the field manipulation of predator abundance. Predator impact (PI) was calculated
as the log-ratio effect size PI = ln(Ba/Br) where Ba is the prey biomass in ambient cages and Br is the biomass in predator removal or
reduction cages.
Predator impact type Prey type
Contrasts for PI
Ba Br
Fish PI All invertebrates Open cages Fish exclusion
Predatory invertebrate PI Non-predatory inverts Fish removal All predator reduction
Total PI Non-predatory inverts Open cages All predator reduction
© 2013 The Authors. Journal of Animal Ecology © 2013 British Ecological Society, Journal of Animal Ecology, 82, 598–607
600 H. S Greig, S. A. Wissinger & A. R. McIntosh

AFDM the response variable. Treatment effects on prey commu-
nity composition were assessed with PERMANOVA (PERMANOVA
6; Anderson 2001) on a Bray-Curtis dissimilarity matrix created
from untransformed AFDM of each of the 11 macroinvertebrate
taxa. Significance (P < 0�05) was tested with 999 permutations.
mesocosm experiment 2: multiple predators
The second mesocosm experiment manipulated predator diver-
sity (within and between trophic levels) to determine if increas-
ing food-web complexity dampened top-down control in
permanent ponds. Predator diversity was examined in meso-
cosms using a hybrid design that included both additive and
substitutive manipulations of predator richness (Byrnes &
Stachowicz 2009). This design enabled us to investigate the
changes in intraspecific and interspecific interactions associated
with increases in the density and complexity of the predator
guild with pond permanence (Fig. 1a). We manipulated three
predatory invertebrate taxa including a common beetle, damsel-
fly and dragonfly larva and the non-consumptive effects of a
native and an introduced predatory fish species was also
included in one comparison (Fig. 1b). Treatments were limited
to the subsets of possible combinations that were relevant to
natural predator combinations across the permanence gradient.
The design enabled three models to be tested (indicted by small
cased letters beneath treatments in Fig. 1b): (1) density manipu-
lations (d) of a single widespread predator species, (2) additive
manipulations (a) of multiple predator species in combination
with all possible single species at low density treatments to
enable the calculation of expected predation rates and (3) a sub-
stitutive model (s) at high predator densities in which the effects
of fish chemical cues was also tested. Each of the 11 treatment
combinations (Fig. 1b), including predator-free controls with
and without fish cues, was replicated four times.
Manipulations were conducted in 0�35 m2 white plastic tubs
housed outdoors at the Cass Field Station. Mesocosms were filled
to a depth of 18 cm with groundwater, and three 15-cm high
clumps of plastic aquarium plants, three large cobbles and a layer
of fine sand were added as substrate (Fig. S2, Supporting Infor-
mation). Predator density varied between 4 and 12 individuals
per mesocosm, reflecting the range of predator biomass observed
in natural ponds. The combined cues of two fish species were
added to fish treatments by dripping water (6�2 � 1 L h�1) from
a single tank housing one koaro (145-mm fork length [FL]) and
one rainbow trout (140 mm FL). Fish were fed ad libitum on all
three species of invertebrates used in the experiment to ensure the
complete range of cues from fish feeding was present. A control
drip from an identical tank without fish or invertebrates was
distributed at the same rate into the remaining tanks. Prey were
late instar Chironomus zelandicus Hudson larvae (100 per meso-
cosm), collected from a nearby pond. Chironomus are found in
almost every lentic habitat in the landscape and are an important
component of predator diets. Prey were added at 1600 h on the
31st October 2007 and allowed to acclimate for 24 h before pre-
dators were introduced. Mesocosms were covered with 1 mm
mesh throughout the experiment to contain prey emergence.
After 5 days, adult chironomids that had emerged were hand-
collected, cobbles and artificial plants were washed and
removed, and invertebrates were collected by repeated elutriation
of the tank contents through a 250-lm mesh net followed by an
inspection of the remaining sand. Invertebrates were preserved
in 70% ETOH, enumerated in the laboratory and AFDM
Food
web
com
plex
ity
Pond permanence
Control R
X
R
R R
X
PControl R
R
XX P
R
X
P
d,a da a a d,s s a,s a,s Pre
dato
ry in
verte
brat
ede
nsity
R X
P
R X
P
R XR
(a)
(b)
Fig. 1. (a) Change in the complexity of predator guilds across the gradient of water permanence from temporary to permanent lentic
habitats in Canterbury, New Zealand. Letters denote representative beetle (R, Rhantus suturalis Macleay), damselfly (X, Xanthocnemis
zelandica Mclauchlan) and dragonfly taxa [P, Procordulia grayi (Selys)]. Solid and dashed arrows represent consumptive and non-con-
sumptive effects, respectively. (b) Hybrid experimental design used to investigate multiple predator effects along the permanence gradi-
ent. In (b) each box represents one treatment and box size is proportional to predatory invertebrate density. Capital letters correspond
to species in (a) and shading indicates the presence of chemical cues from fish (Oncorhynchus mykiss and Galaxias brevipinnis). Lower
case letters denote treatment combinations used in the single-species density manipulation (d), and the additive (a) and substitutive (s)
diversity manipulations. Two predator-free controls (with and without fish cues) were also included. Prey density was constant across all
treatments.
© 2013 The Authors. Journal of Animal Ecology © 2013 British Ecological Society, Journal of Animal Ecology, 82, 598–607
Disturbance increases top-down control 601

determined in the manner described earlier. Treatment combina-
tions were grouped according to types of predator manipulation:
(1) Rhantus beetle density treatment, (2) additive diversity
manipulations and (3) substitutive diversity manipulations
(Fig. 1b). Significance of each predator manipulation was tested
using one-way ANOVA with tanks as replicates, followed by
Tukey’s post-hoc comparisons of treatment means. Multiple
predator effects were tested in the additive model by calculating
expected predation rates for the two- and three-species treat-
ments from the low-density single-species treatments using an
additive probability model assuming finite prey resources (Soluk
& Collins 1988). The model was modified for a three species
guild (following Miller 2006:62), whereby expected prey consu-
mption = 100 9 (Pr + Px + Pp � PrPx � PxPp + PrPxPp), where
Pr, Px and Pp are the proportions of initial prey abundance
eaten in the Rhantus, Xanthocnemis and Procordulia single-pred-
ator treatments respectively. The effect of density on intraspe-
cific interactions in Rhantus single-species treatments was also
tested using these models. In this case, expected values were
based on Rhantus low density treatments. Significant differences
between observed predation rates and expected values were
assessed with one-sample t-tests. All analyses for the field and
mesocosm experiments (except PERMANOVA) were conducted in
Statistica 8 (StatSoft 2008).
Results
assays of predator impact in natural ponds
No consistent effects of fish on the biomass of prey were
observed in the in situ assays of predator impact in natu-
ral ponds (Fig. S3, Supporting Information). The biomass
of all invertebrates and primary consumers in cages that
allowed fish access did not differ significantly from cages
that prevented fish access (PI not significantly different
from zero; one-sample t-tests: t4 < 1�14, P > 0�31). The
same pattern was evident when only unprotected prey bio-
mass (species without cases or shells) was considered
(one-sample t-tests: t4 = 1�21, P = 0�29; Fig. S3, Support-ing Information).
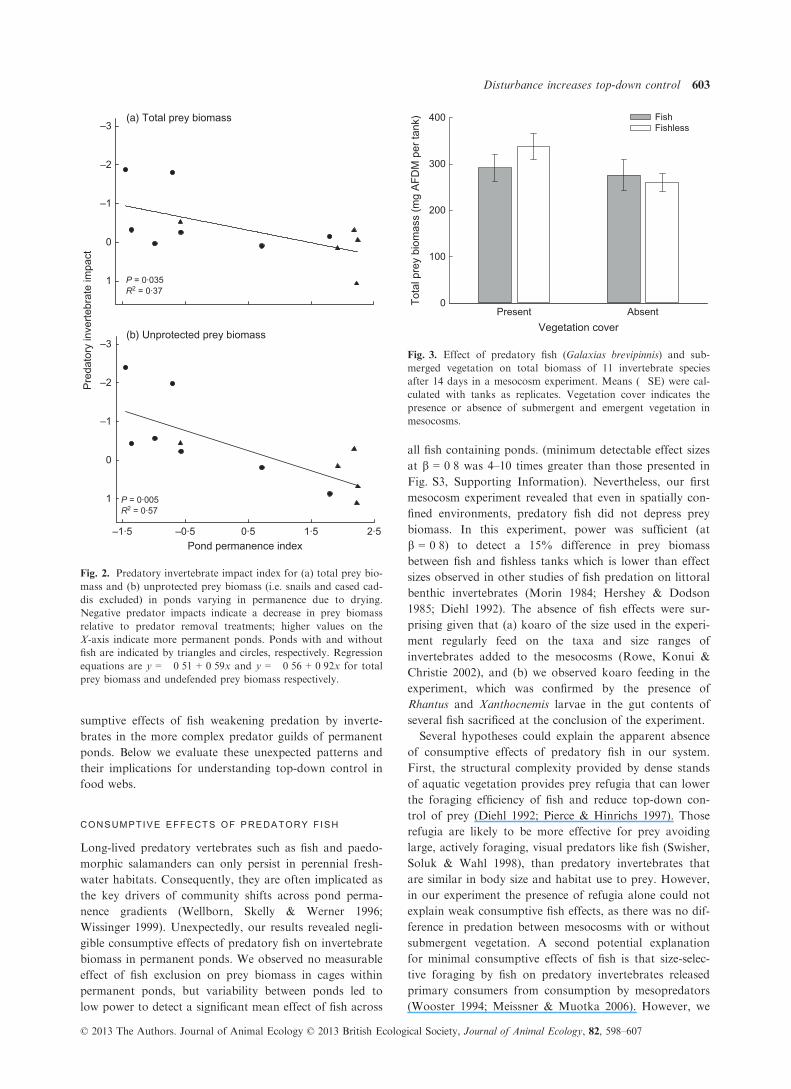
The impact of predatory invertebrates on prey biomass
decreased with increasing pond permanence (Fig. 2a). This
pattern strengthened when only prey without morphologi-
cal defences were considered, owing to an increase in PI in
more temporary ponds (Fig. 2b). The biomass of preda-
tory invertebrates in the unmanipulated cages did not vary
significantly with pond permanence when expressed as
either biomass per cage, or proportion of total invertebrate
biomass (Fig. S4, Supporting Information; P > 0�15).
mesocosm experiment 1: manipulation of fishand aquatic vegetation
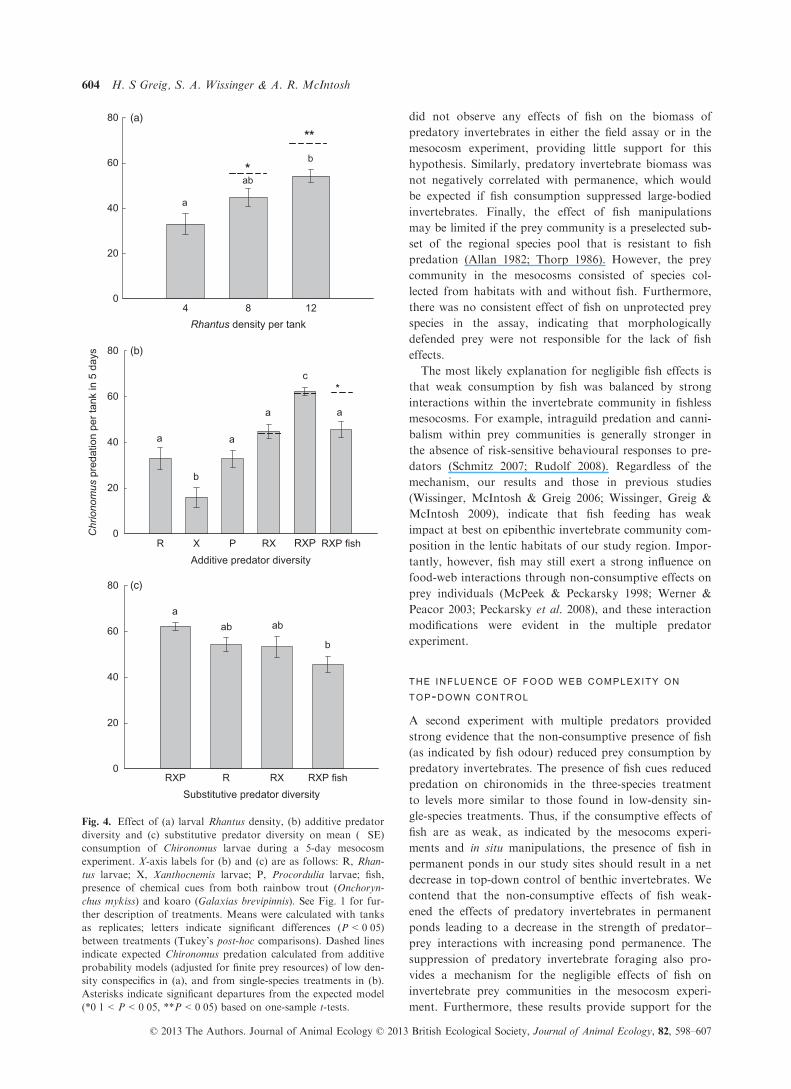
There was no detectable effect of predatory fish on prey
biomass in the mesocosms (Fig. 3). Total prey biomass
was not significantly different between fish and fishless
treatments (F1,12 = 0�29, P = 0�60; Fig. 3), or between
complex and simple vegetation treatments (F1,12 = 2�74,P = 0�12), and there was no fish by vegetation interaction
(F1,12 = 1�03, P = 0�33). Similarly, PERMANOVA indicated
that there were no treatment effects on the composition
of the prey community (fish: F1,12 = 0�84, P = 0�51; vege-tation: F1,12 = 1�95, P = 0�11; interaction: F1,12 = 1�17,P = 0�28). Lastly, fish had no effect on the total biomass
of large-bodied (>4 mg DW) predatory taxa (ANOVA, fish:
F1,12 = 0�52, P = 0�48, interaction: F1,12 = 0�87, P = 0�37).
mesocosm experiment 2: multiple predators
Predation rates of Rhantus beetles as the sole predator
increased significantly with density (one-way ANOVA:
F2,9 = 7�26, P = 0�013; Fig. 4a), but observed predation
rates in high density treatments were significantly lower
than expected based on additive predation at low densities
(Fig. 4a). The additive manipulation of predator diversity
had significant impacts on the consumption of Chirono-
mus (Fig. 4b, One-way ANOVA: F5,18 = 17�9, P < 0�0001).In treatments with single predator species, predation rates
of Rhantus beetle larvae and Procordulia dragonflies were
almost identical, but Xanthocnemis damselflies were a sig-
nificantly weaker predator on Chironomus (Fig. 4b). The
two-predator species treatment resulted in significantly
higher predation rates than by Xanthocnemis alone, but
was not significantly different from rates for Procordulia
or Rhantus alone. In contrast with the single-species den-
sity manipulation, increasing predator density by increas-
ing diversity led to predation rates almost identical to
expected values calculated from the single-species treat-
ments (Fig. 4b). However, the addition of fish chemical
cues significantly decreased predation rates in the three-
species treatment to a level similar to the two-species
treatment and not significantly higher than for Rhantus
and Procordulia alone at low density (Fig. 4a).
The substitutive manipulation of predator diversity, in
which total predator density was kept constant, signifi-
cantly influenced predation on Chironomus (Fig. 4c, one-
way ANOVA: F3,12 = 4�29, P = 0�028); however, this was
due to a reduction in predation rates with fish cues in the
three-species treatment (Fig. 4c). Post-hoc tests indicated
no significant difference between the one-, two- and three-
species treatments in the absence of fish cues (P > 0�34).
Discussion
Decreases in the intensity of competition and predation
with increasing abiotic stress or disturbance are central to
several models of community organization across environ-
mental gradients (Menge & Sutherland 1987; Wellborn,
Skelly & Werner 1996; Chesson 2000). Our field assays of
predator–prey interactions across a pond drying gradient
revealed patterns opposite to these predictions. Top-down
control of prey biomass decreased with pond permanence
despite an increase in the size, biomass and diversity of
predators in increasingly permanent ponds (Wissinger,
Greig & McIntosh 2009). Our mesocosm experiment sug-
gested that those patterns could result from the non-con-
© 2013 The Authors. Journal of Animal Ecology © 2013 British Ecological Society, Journal of Animal Ecology, 82, 598–607
602 H. S Greig, S. A. Wissinger & A. R. McIntosh
sumptive effects of fish weakening predation by inverte-
brates in the more complex predator guilds of permanent
ponds. Below we evaluate these unexpected patterns and
their implications for understanding top-down control in
food webs.
consumptive effects of predatory fish
Long-lived predatory vertebrates such as fish and paedo-
morphic salamanders can only persist in perennial fresh-
water habitats. Consequently, they are often implicated as
the key drivers of community shifts across pond perma-
nence gradients (Wellborn, Skelly & Werner 1996;
Wissinger 1999). Unexpectedly, our results revealed negli-
gible consumptive effects of predatory fish on invertebrate
biomass in permanent ponds. We observed no measurable
effect of fish exclusion on prey biomass in cages within
permanent ponds, but variability between ponds led to
low power to detect a significant mean effect of fish across
all fish containing ponds. (minimum detectable effect sizes
at b = 0�8 was 4–10 times greater than those presented in
Fig. S3, Supporting Information). Nevertheless, our first
mesocosm experiment revealed that even in spatially con-
fined environments, predatory fish did not depress prey
biomass. In this experiment, power was sufficient (at
b = 0�8) to detect a 15% difference in prey biomass
between fish and fishless tanks which is lower than effect
sizes observed in other studies of fish predation on littoral
benthic invertebrates (Morin 1984; Hershey & Dodson
1985; Diehl 1992). The absence of fish effects were sur-
prising given that (a) koaro of the size used in the experi-
ment regularly feed on the taxa and size ranges of
invertebrates added to the mesocosms (Rowe, Konui &
Christie 2002), and (b) we observed koaro feeding in the
experiment, which was confirmed by the presence of
Rhantus and Xanthocnemis larvae in the gut contents of
several fish sacrificed at the conclusion of the experiment.
Several hypotheses could explain the apparent absence
of consumptive effects of predatory fish in our system.
First, the structural complexity provided by dense stands
of aquatic vegetation provides prey refugia that can lower
the foraging efficiency of fish and reduce top-down con-
trol of prey (Diehl 1992; Pierce & Hinrichs 1997). Those
refugia are likely to be more effective for prey avoiding
large, actively foraging, visual predators like fish (Swisher,
Soluk & Wahl 1998), than predatory invertebrates that
are similar in body size and habitat use to prey. However,
in our experiment the presence of refugia alone could not
explain weak consumptive fish effects, as there was no dif-
ference in predation between mesocosms with or without
submergent vegetation. A second potential explanation
for minimal consumptive effects of fish is that size-selec-
tive foraging by fish on predatory invertebrates released
primary consumers from consumption by mesopredators
(Wooster 1994; Meissner & Muotka 2006). However, we
Pond permanence index–1·5 –0·5 0·5 1·5 2·5
–3
–2
–1
0
1
–3
–2
–1
0
1
P = 0·005R2 = 0·57
P = 0·035R2 = 0·37
Pre
dato
ry in
verte
brat
e im
pact
(a) Total prey biomass
(b) Unprotected prey biomass
Fig. 2. Predatory invertebrate impact index for (a) total prey bio-
mass and (b) unprotected prey biomass (i.e. snails and cased cad-
dis excluded) in ponds varying in permanence due to drying.
Negative predator impacts indicate a decrease in prey biomass
relative to predator removal treatments; higher values on the
X-axis indicate more permanent ponds. Ponds with and without
fish are indicated by triangles and circles, respectively. Regression
equations are y = �0�51 + 0�59x and y = �0�56 + 0�92x for total
prey biomass and undefended prey biomass respectively.
Vegetation coverPresent Absent
Tota
l pre
y bi
omas
s (m
g A
FDM
per
tank
)
0
100
200
300
400 FishFishless
Fig. 3. Effect of predatory fish (Galaxias brevipinnis) and sub-
merged vegetation on total biomass of 11 invertebrate species
after 14 days in a mesocosm experiment. Means (�SE) were cal-
culated with tanks as replicates. Vegetation cover indicates the
presence or absence of submergent and emergent vegetation in
mesocosms.
© 2013 The Authors. Journal of Animal Ecology © 2013 British Ecological Society, Journal of Animal Ecology, 82, 598–607
Disturbance increases top-down control 603
did not observe any effects of fish on the biomass of
predatory invertebrates in either the field assay or in the
mesocosm experiment, providing little support for this
hypothesis. Similarly, predatory invertebrate biomass was
not negatively correlated with permanence, which would
be expected if fish consumption suppressed large-bodied
invertebrates. Finally, the effect of fish manipulations
may be limited if the prey community is a preselected sub-
set of the regional species pool that is resistant to fish
predation (Allan 1982; Thorp 1986). However, the prey
community in the mesocosms consisted of species col-
lected from habitats with and without fish. Furthermore,
there was no consistent effect of fish on unprotected prey
species in the assay, indicating that morphologically
defended prey were not responsible for the lack of fish
effects.
The most likely explanation for negligible fish effects is
that weak consumption by fish was balanced by strong
interactions within the invertebrate community in fishless
mesocosms. For example, intraguild predation and canni-
balism within prey communities is generally stronger in
the absence of risk-sensitive behavioural responses to pre-
dators (Schmitz 2007; Rudolf 2008). Regardless of the
mechanism, our results and those in previous studies
(Wissinger, McIntosh & Greig 2006; Wissinger, Greig &
McIntosh 2009), indicate that fish feeding has weak
impact at best on epibenthic invertebrate community com-
position in the lentic habitats of our study region. Impor-
tantly, however, fish may still exert a strong influence on
food-web interactions through non-consumptive effects on
prey individuals (McPeek & Peckarsky 1998; Werner &
Peacor 2003; Peckarsky et al. 2008), and these interaction
modifications were evident in the multiple predator
experiment.
the influence of food web complexity ontop-down control
A second experiment with multiple predators provided
strong evidence that the non-consumptive presence of fish
(as indicated by fish odour) reduced prey consumption by
predatory invertebrates. The presence of fish cues reduced
predation on chironomids in the three-species treatment
to levels more similar to those found in low-density sin-
gle-species treatments. Thus, if the consumptive effects of
fish are as weak, as indicated by the mesocoms experi-
ments and in situ manipulations, the presence of fish in
permanent ponds in our study sites should result in a net
decrease in top-down control of benthic invertebrates. We
contend that the non-consumptive effects of fish weak-
ened the effects of predatory invertebrates in permanent
ponds leading to a decrease in the strength of predator–
prey interactions with increasing pond permanence. The
suppression of predatory invertebrate foraging also pro-
vides a mechanism for the negligible effects of fish on
invertebrate prey communities in the mesocosm experi-
ment. Furthermore, these results provide support for the
Additive predator diversity
0
20
40
60
80
a
b
a
a a
c*
Rhantus density per tank
0
20
40
60
80
a
ab
b
**
*
Substitutive predator diversity
R X P RX RXP RXP fish
4 8 12
RXP R RX RXP fish
Chr
iono
mus
pre
datio
n pe
r tan
k in
5 d
ays
0
20
40
60
80
aabab
b
(c)
(b)
(a)
Fig. 4. Effect of (a) larval Rhantus density, (b) additive predator
diversity and (c) substitutive predator diversity on mean (�SE)
consumption of Chironomus larvae during a 5-day mesocosm
experiment. X-axis labels for (b) and (c) are as follows: R, Rhan-
tus larvae; X, Xanthocnemis larvae; P, Procordulia larvae; fish,
presence of chemical cues from both rainbow trout (Onchoryn-
chus mykiss) and koaro (Galaxias brevipinnis). See Fig. 1 for fur-
ther description of treatments. Means were calculated with tanks
as replicates; letters indicate significant differences (P < 0�05)between treatments (Tukey’s post-hoc comparisons). Dashed lines
indicate expected Chironomus predation calculated from additive
probability models (adjusted for finite prey resources) of low den-
sity conspecifics in (a), and from single-species treatments in (b).
Asterisks indicate significant departures from the expected model
(*0�1 < P < 0�05, **P < 0�05) based on one-sample t-tests.
© 2013 The Authors. Journal of Animal Ecology © 2013 British Ecological Society, Journal of Animal Ecology, 82, 598–607
604 H. S Greig, S. A. Wissinger & A. R. McIntosh

hypothesis that non-consumptive effects of fish in fresh-
water food webs often outweigh their consumptive effects
(e.g. McPeek & Peckarsky 1998).
Our experiments focused on the most commonly
observed predatory invertebrates across the permanence
gradient, and indicate that increased invertebrate predator
diversity in the absence of fish cues led to small positive
effects of diversity on predator impacts, rather than
antagonistic effects among species. The additive model in
particular revealed that increases in predator density by
enhancing diversity may reduce negative intraspecific
interactions apparent in high densities of a single predator
taxa (as in Griffin et al. 2008). This is consistent with the
assertion that intraspecific interactions among predators
are stronger than interspecific interactions (Byrnes & Stac-
howicz 2009). Predation rates of Rhantus beetle larvae at
high densities were significantly lower than the expected
additive predation rate calculated from low density treat-
ments, but predation rates in high density treatments of
multiple predators were almost identical to expected rates.
The Rhantus only treatment is representative of predator
guilds in the most temporary ponds in our study area
(< 1 month inundated), which we were unable to include
in the field experiment because of the logistical constraints
of their short duration. The small positive effects of pred-
atory invertebrate diversity we observed suggest that
predator impact may follow a hump-shaped relationship
with pond permanence when the full range of pond dura-
tion is considered.
Differences we observed in the effects of diversity by
adding trophic levels or adding species within trophic lev-
els reinforce the contention that horizontal and vertical
diversity components of food webs have different but
interactive effects on species interactions (Duffy et al.
2007; Srivastava & Bell 2009). Vertical diversity can
increase top-down control, for example, when top preda-
tors increase spatial overlap of mesopredators and prey
(Grabowski & Kimbro 2005). However, in many cases,
the addition of predator trophic levels should dampen
top-down control. Body size generally increases with tro-
phic level (Woodward et al. 2005), so increasing vertical
diversity in food webs generates size asymmetry between
predator species that promotes intraguild predation and
density- and trait-mediated foraging suppression of meso-
predators. This is analogous to the effects of introducing
size-structure within predator populations (Rudolf 2007).
These negative interactions among trophic levels may
overwhelm positive effects of within-trophic level diver-
sity, as observed in our study when fish cues reduced the
foraging of three species predator guilds to rates similar
to single species guilds.
Several aspects of food web complexity in addition to
diversity may have contributed to the decreased predator
impact with more permanent ponds. Many permanent
pond species have semivoltine life histories, which results
in overlapping cohorts at a given time. Consequently,
size-structured interactions such as cannibalism and intra-
guild predation that can weaken top-down control (Polis
1991; Padeffke & Suhling 2003) are likely to be more pre-
valent in permanent ponds than in temporary ponds
where drying and refilling promotes developmental syn-
chrony within populations. Permanent ponds also often
contain a higher proportion of morphologically defended
prey species (e.g. snails and cased caddisflies) than tempo-
rary ponds, although the negative relationship between
predator impact and permanence actually strengthened
when we excluded morphologically defended prey from
the analysis (Fig. 3b). Finally, prey in resource-limited or
time-constrained habitats such as temporary ponds may
be less likely to exhibit risk-sensitive foraging and other
antipredatory responses, which should strengthen
top-down control.
Conclusions
Interactions between the same sets of species can reverse
in direction or change in strength depending on environ-
mental and consumer context (Crain 2008). Our study
provides evidence that changes in the properties of food
webs along environmental gradients from structurally sim-
ple, chain-like food webs in disturbed habitats, to com-
plex, reticulate food webs in physically benign habitats
are likely to dampen top-down control and generate unex-
pected relationships between abiotic stress and the
strength of species interactions. These results emphasize
that considering food web complexity, especially the non-
consumptive effects of top predators, is essential to under-
stand species interactions across environmental gradients.
Acknowledgements
We thank A. Klemmer, P. Jellyman, E. Isherwood, C. Ross and M.
Galatowitsch for field and laboratory assistance. N. Etheridge and V.
Greig constructed the cages, and M. Fraundorfer, R. Hill, J. Westenra, R.
Smith and the Dept. of Conservation granted access to research sites. We
thank B. Peckarsky, R. Didham, B. Sorrell, M. Winterbourn, A. Hildrew
and G. Closs for valuable comments and discussion. The research was
funded by the Miss E.L. Hellaby Indigenous Grassland Research Trust.
HSG was supported by a Top Achiever Doctoral Scholarship and a Foun-
dation for Science, Research and Technology Postdoctoral Fellowship and
SAW was supported by an Erskine Fellowship from the University of
Canterbury.
References
Agrawal, A.A., Ackerly, D.D., Adler, F., Arnold, A.E., Caceres, C.,
Doak, D.F., Post, E., Hudson, P.J., Maron, J., Mooney, K.A., Power,
M., Schemske, D., Stachowicz, J., Strauss, S., Turner, M.G. & Werner,
E. (2007) Filling key gaps in population and community ecology. Fron-
tiers in Ecology and the Environment, 5, 145–152.Allan, J.D. (1982) The effects of reduction in trout density on the inverte-
brate community of a mountain stream. Ecology, 63, 1444–1455.Anderson, M.J. (2001) A new method for non-parametric multivariate
analysis of variance. Austral Ecology, 26, 32–46.Benke, A.C., Huryn, A.D., Smock, L.A. & Wallace, J.B. (1999) Length-
mass relationships for freshwater macroinvertebrates in North America
with particular reference to the southeastern United States. Journal of
the North American Benthological Society, 18, 308–343.Bilton, D.T., Foggo, A. & Rundle, S.D. (2001) Size, permanence and the pro-
portion of predators in ponds. Archiv Fur Hydrobiologie, 151, 451–458.
© 2013 The Authors. Journal of Animal Ecology © 2013 British Ecological Society, Journal of Animal Ecology, 82, 598–607
Disturbance increases top-down control 605

Berlow, E.L., Neutel, A.M., Cohen, J.E., de Ruiter, P.C., Ebenman, B.,
Emmerson, M., Fox, J.W., Jansen, V.A.A., Jones, J.I., Kokkoris, G.D.,
Logofet, D.O., McKane, A.J., Montoya, J.M. & Petchey, O. (2004)
Interaction strengths in food webs: issues and opportunities. Journal of
Animal Ecology, 73, 585–598.Brendonck, L., Michels, E., De Meester, L. & Riddoch, B. (2002) Tempo-
rary pools are not ‘enemy-free’. Hydrobiologia, 486, 147–159.Byrnes, J.E. & Stachowicz, J.J. (2009) The consequences of consumer
diversity loss: different answers from different experimental designs.
Ecology, 90, 2879–2888.Chesson, P. (2000) Mechanisms of maintenance of species diversity.
Annual Review of Ecology and Systematics, 31, 343–366.Chesson, P. & Huntly, N. (1997) The roles of harsh and fluctuating condi-
tions in the dynamics of ecological communities. American Naturalist,
150, 519–553.Connell, J.H. (1975) Some mechanisms producing structure in natural
communities: a model and evidence from field experiments. Ecology and
Evolution of Communities (eds M.L. Cody & J.M. Diamond), pp. 460–490. Bclknap, Harvard, Cambridge, MT.
Crain, C.M. (2008) Interactions between marsh plant species vary in direc-
tion and strength depending on environmental and consumer context.
Journal of Ecology, 96, 166–173.Crain, C.M., Silliman, B.R., Bertness, S.L. & Bertness, M.D. (2004) Physi-
cal and biotic drivers of plant distribution across estuarine salinity gra-
dients. Ecology, 85, 2539–2549.Diehl, S. (1992) Fish predation and benthic community structure: the role
of omnivory and habitat complexity. Ecology, 73, 1646–1661.Duffy, J.E., Cardinale, B.J., France, K.E., McIntyre, P.B., Thebault, E. &
Loreau, M. (2007) The functional role of biodiversity in ecosystems:
incorporating trophic complexity. Ecology Letters, 10, 522–538.Edwards, K.F. & Stachowicz, J.J. (2010) Multivariate trade-offs, succes-
sion, and phenological differentiation in a guild of colonial inverte-
brates. Ecology, 91, 3146–3152.Emmerson, M. & Yearsley, J.M. (2004) Weak interactions, omnivory and
emergent food-web properties. Proceedings of the Royal Society of
London Series B-Biological Sciences, 271, 397–405.Finke, D.L. & Denno, R.F. (2004) Predator diversity dampens trophic cas-
cades. Nature, 429, 407–410.Gerhardt, F. & Collinge, S.K. (2007) Abiotic constraints eclipse biotic
resistance in determining invasibility along experimental vernal pool
gradients. Ecological Applications, 17, 922–933.Grabowski, J.H. & Kimbro, D.L. (2005) Predator-avoidance behavior
extends trophic cascades to refuge habitats. Ecology, 86, 1312–1319.Griffin, J.N., De la Haye, K.L., Hawkins, S.J., Thompson, R.C. &
Jenkins, S.R. (2008) Predator diversity and ecosystem functioning:
density modifies the effect of resource partitioning. Ecology, 89, 298–305.
Grime, J.P. (1977) Evidence for existence of 3 primary strategies in plants
and its relevance to ecological and evolutionary theory. American Natu-
ralist, 111, 1169–1194.Hershey, A.E. & Dodson, S.I. (1985) Selective predation by a sculpin and
a stonefly on 2 chironomids in laboratory feeding trials. Hydrobiologia,
124, 269–273.Jenkins, B., Kitching, R.L. & Pimm, S.L. (1992) Productivity, disturbance
and food web structure at a local spatial scale in experimental container
habitats. Oikos, 65, 249–255.Jeppesen, E., Lauridsen, T., Mitchell, S.F. & Burns, C.W. (1997) Do
planktivorous fish structure the zooplankton communities in New Zea-
land lakes? New Zealand Journal of Marine and Freshwater Research,
31, 163–173.Lubchenco, J. (1980) Algal zonation in the New England rocky intertidal
community: an experimental analysis. Ecology, 61, 333–344.McAbendroth, L., Foggo, A., Rundle, S.D. & Bilton, D.T. (2005) Unrav-
elling nestedness and spatial pattern in pond assemblages. Journal of
Animal Ecology, 74, 41–49.McPeek, M.A. & Peckarsky, B.L. (1998) Life histories and the strengths
of species interactions: combining mortality, growth, and fecundity
effects. Ecology, 79, 867–879.Meissner, K. & Muotka, T. (2006) The role of trout in stream food webs:
integrating evidence from field surveys and experiments. Journal of Ani-
mal Ecology, 75, 421–433.Menge, B.A. & Farrell, T.M. (1989) Community structure and interac-
tion webs in shallow marine hard-bottom communities: tests of an
environmental-stress model. Advances in Ecological Research, 19, 189–262.
Menge, B.A. & Sutherland, J.P. (1987) Community regulation: variation
in disturbance, competition, and predation in relation to environmental-
stress and recruitment. American Naturalist, 130, 730–757.Miller, G.K. (2006) Probability: Modelling and Applications to Random
Processes. Willey, Hoboken, USA.
Morin, P.J. (1984) The impact of fish exclusion on the abundance and spe-
cies composition of larval odonates: results of short-term experiments in
a North Carolina farm pond. Ecology, 65, 53–60.Padeffke, T. & Suhling, F. (2003) Temporal priority and intra-guild preda-
tion in temporary waters: an experimental study using Namibian desert
dragonflies. Ecological Entomology, 28, 340–347.Peckarsky, B.L. (1983) Biotic interactions or abiotic limitations? A model
of lotic community structure. Dynamics of Lotic Ecosystems (eds T.D.
Fontaine III & S.M. Bartell), pp. 303–323. Ann Arbor Science Publica-
tions, Ann Arbor, USA.
Peckarsky, B.L., Kerans, B.L., Taylor, B.W. & McIntosh, A.R. (2008)
Predator effects on prey population dynamics in open systems. Oecolo-
gia, 156, 431–440.Pierce, C.L. & Hinrichs, B.D. (1997) Response of littoral invertebrates to
reduction of fish density: simultaneous experiments in ponds with differ-
ent fish assemblages. Freshwater Biology, 37, 397–408.Polis, G.A. (1991) Complex trophic interactions in deserts: an empirical
critique of food-web theory. American Naturalist, 138, 123–155.Power, M.E., Parker, M.S. & Wootton, J.T. (1996) Disturbance and food
chain length in rivers. Food Webs: Contemporary Perspectives (eds G.A.
Polis & K.O. Winemiller), pp. 286–297. Chapman and Hall, New York,
USA.
Rowe, D.K., Konui, G. & Christie, K.D. (2002) Population structure, dis-
tribution, reproduction, diet, and relative abundance of koaro (Galaxias
brevipinnis) in a New Zealand lake. Journal of the Royal Society of New
Zealand, 32, 275–291.Rudolf, V.H.W. (2007) Consequences of stage-structured predators: canni-
balism, behavioral effects, and trophic cascades. Ecology, 88, 2991–3003.Rudolf, V.H.W. (2008) The impact of cannibalism in the prey on preda-
tor-prey systems. Ecology, 89, 3116–3127.Schmitz, O.J. (2007) Predator diversity and trophic interactions. Ecology,
88, 2415–2426.Schneider, D.W. & Frost, T.M. (1996) Habitat duration and community
structure in temporary ponds. Journal of the North American Benthologi-
cal Society, 15, 64–86.Soluk, D.A. & Collins, N.C. (1988) Balancing risks: responses and non-
responses of mayfly larvae to fish and stonefly predators. Oecologia, 77,
370–374.Srivastava, D.S. & Bell, T. (2009) Reducing horizontal and vertical diver-
sity in a foodweb triggers extinctions and impacts functions. Ecology
Letters, 12, 1016–1028.StatSoft (2008) STATISTICA (data analysis software system), Version 8.
Tulsa, Oklahoma.
Stoffels, R.J., Karbe, S. & Paterson, R.A. (2003) Length-mass models for
some common New Zealand littoral-benthic macroinvertebrates, with a
note on within-taxon variability in parameter values among published
models. New Zealand Journal of Marine and Freshwater Research, 37,
449–460.Swisher, B.J., Soluk, D.A. & Wahl, D.H. (1998) Non-additive predation in
littoral habitats: influences of habitat complexity. Oikos, 81, 30–37.Thompson, R.M. & Townsend, C.R. (1999) The effect of seasonal varia-
tion on the community structure and food-web attributes of two
streams: implications for food-web science. Oikos, 87, 75–88.Thorp, J.H. (1986) Two distinct roles for predators in fresh-water assem-
blages. Oikos, 47, 75–82.Urban, M.C. (2004) Disturbance heterogeneity determines freshwater
metacommunity structure. Ecology, 85, 2971–2978.Walters, A.W. & Post, D.M. (2008) An experimental disturbance alters
fish size structure but not food chain length in streams. Ecology, 89,
3261–3267.Wellborn, G.A., Skelly, D.K. & Werner, E.E. (1996) Mechanisms creating
community structure across a freshwater habitat gradient. Annual
Review of Ecology and Systematics, 27, 337–364.Werner, E.E. & Peacor, S.D. (2003) A review of trait-mediated indirect
interactions in ecological communities. Ecology, 84, 1083–1100.Williams, D.D. (1996) Environmental constraints in temporary fresh
waters and their consequences for the insect fauna. Journal of the North
American Benthological Society, 15, 634–650.Winemiller, K.O. (1990) Spatial and temporal variation in tropical fish tro-
phic networks. Ecological Monographs, 60, 331–367.
© 2013 The Authors. Journal of Animal Ecology © 2013 British Ecological Society, Journal of Animal Ecology, 82, 598–607
606 H. S Greig, S. A. Wissinger & A. R. McIntosh

Wissinger, S.A. (1999) Ecology of wetland invertebrates: synthesis and
applications for conservation and management. Invertebrates in Fresh-
water Wetlands of North America (eds D.P. Batzer, R.B. Rader & S.A.
Wissinger), pp. 1043–1086. Willey, New York.
Wissinger, S.A., Greig, H. & McIntosh, A. (2009) Absence of species
replacements between permanent and temporary lentic communities in
New Zealand. Journal of the North American Benthological Society, 28,
12–23.Wissinger, S.A., McIntosh, A.R. & Greig, H.S. (2006) Impacts of intro-
duced brown and rainbow trout on benthic invertebrate communities in
shallow New Zealand lakes. Freshwater Biology, 51, 2009–2028.Wissinger, S.A., Bohonak, A.J., Whiteman, H.H. & Brown, W.S. (1999)
Habitat permanence, salamander predation and invertebrate communi-
ties. Invertebrates in Freshwater Wetlands of North America: Ecology
and Management (eds D.P. Batzer, R.B. Rader & S.A. Wissinger), pp.
757–790. John Wiley & Sons, Inc, New York, USA.
Wissinger, S.A., Whissel, J.C., Eldermire, C. & Brown, W.S. (2006) Preda-
tor defense along a permanence gradient: roles of case structure, behav-
ior, and developmental phenology in caddisflies. Oecologia, 147, 667–678.
Woodward, G., Ebenman, B., Ernmerson, M., Montoya, J.M., Olesen,
J.M., Valido, A. & Warren, P.H. (2005) Body size in ecological net-
works. Trends in Ecology & Evolution, 20, 402–409.Wooster, D. (1994) Predator impacts on stream benthic prey. Oecologia,
99, 7–15.
Received 30 July 2012; accepted 29 November 2012
Handling Editor: Karl Cottenie
Supporting Information
Additional Supporting Information may be found in the online version
of this article.
Table S1. A list of environmental variables for each pond, includ-
ing hydrological parameters underlying the multivariate pond
permanence index, and the identity of predatory fish.
Figure S1. Photos and description of the mesocosm experiment
manipulating the presence and absence of fish and vegetation.
Figure S2. Photo of the mesocosms used in the predator diversity
manipulation.
Figure S3. The impact of predatory fish on invertebrate biomass in
the in situ cage experiment.
Figure S4. Regression of predatory invertebrate biomass in unma-
nipulated cages against pond permanence for the in situ cage
experiment.
© 2013 The Authors. Journal of Animal Ecology © 2013 British Ecological Society, Journal of Animal Ecology, 82, 598–607
Disturbance increases top-down control 607
Copyright © 2022 FDOKUMEN




















